Houses without roofs, and roofs without houses
The whole town is destroyed and messed up
Houses without roofs, and roofs without houses
The whole town is destroyed and messed up
“She didn’t kill no white man, did she? Well, long as she don’t shoot no white man she kin kill jus’ as many niggers as she please.”
The life of black people don’t matter
So the day of the funeral they came with shame and apology in their faces. They
His friends just wanted to get back for him but they knew wither were wrong and wanted to make things right with Janie.
She was trying to hover him as he closed his teeth in the flesh of her forearm. They came down heavily like that. Janie struggled to a sitting position and pried the dead Tea Cake’s teeth from her arm
If tea cake has rabies, then does that mean he transfer rabies to Jamie when he bit her?
Janie bought him a brand new guitar and put it in his hands. He would be thinking up new songs to play to her when she got there.
After tea cake tried to kill her she still showed love for him at the funeral and showed she truly cared for him and wanted him to get better
After the hurricane teacake gets sick and contracts rabies he gets aggressive and try’s to shoot Janie but she shoots and kills him first
Doctah, Tea Cake ain’t tellin’ yuh everything lak he oughta. We wuz caught in dat hurricane out heah, and Tea Cake over-strained hisself swimmin’ such uh long time and holdin’ me up too, and walkin’ all dem miles in de storm and then befo’ he could git his rest he had tuh come git me out de water agin and fightin’ wid dat big ole dawg and de dawg bitin’ ’im in de face and everything. Ah been ’spectin’ him tuh be sick befo’ now.”
They went to the docter because he’s feeling sick and as well as they think he might have rabbiss so they sent them to the doctor.
Chapter 19 was heartbreaking. Janie had to kill the man she truly loved just to save her own life. The rabies took over Tea Cake and turned him into someone she didn't recognize. It's so sad that after everything they survived together, including the hurricane this is how it ended. But at the same time, Janie finally stood up for herself and survived.
Janie buried Tea Cake in Palm Beach. She knew he loved the ’Glades but it was too low for him to lie with water maybe washing over him with every heavy rain. Anyway, the ’Glades and its waters had killed him. She wanted him out of the way of storms, so she had a strong vault built in the cemetery at West Palm Beach. Janie had wired to Orlando for money to put him away.
I don’t think Janie will ever love another man how she loved Tea Cake.
She was trying to hover him as he closed his teeth in the flesh of her forearm. They came down heavily like that. Janie struggled to a sitting position and pried the dead Tea Cake’s teeth from her arm.
Tea Cake bites Janie and he has rabies so she could probably get rabies too now.
Tea Cake crumpled as his bullet buried itself in the joist over Janie’s head. Janie saw the look on his face and leaped forward as he crashed forward in her arms.
He tried to kill her but she had to defend herself so she ended up shooting him and killing him.
She had to hug him tight for soon he would be gone, and she had to tell him for the last time
janie really loved tea cake, so here she is probably feeling regret that she shot him but it had to be done for her own protection.
This is when Tea Cake started getting crazy from the dog bite
Tea Cake and Janie struggle to survive after the hurricane while Tea Cake unknowingly begins to show signs of rabies after being bitten by a dog, which slowly changes his behavior and creates tension between love and fear.
Janie held his head tightly to her breast and wept and thanked him wordlessly for giving her the chance for loving service. She had to hug him tight for soon he would be gone, and she had to tell him for the last time.
Janie didn’t want to shoot him but had to too protect herself. This shows how much she really cared about him and that he was what she was looking for throughout the whole book.
a collection that started when, in 1978, the proprietor of a Cleveland business machine shop refused to service my mostly plastic typewriter. “A worthless toy!” the man yelled. Yes, yelled. He pointed to shelves full of his refurbished typewriters — already decades old yet all in perfect working order.
The sound of typing is one reason to own a vintage manual typewriter — alas, there are only three reasons, and none of them are ease or speed. In addition to sound, there is the sheer physical pleasure of typing; it feels just as good as it sounds, the muscles in your hands control the volume and cadence of the aural assault so that the room echoes with the staccato beat of your synapses.
You can choose the typewriter to match your sound signature.Remingtons from the 1930s go THICK THICK. Midcentury Royals sound like a voice repeating the word CHALK. CHALK. CHALK CHALK. Even the typewriters made for the dawning jet age (small enough to fit on the fold-down trays of the first 707s), like the Smith Corona Skyriter and the design masterpieces by Olivetti, go FITT FITT FITT like bullets from James Bond’s silenced Walther PPK. Composing on a Groma, exported to the West from a Communist country that no longer exists, is the sound of work, hard work. Close your eyes as you touch-type and you are a blacksmith shaping sentences hot out of the forge of your mind.
Everything you type on a typewriter sounds grand, the words forming in mini-explosions of SHOOK SHOOK SHOOK. A thank-you note resonates with the same heft as a literary masterpiece.
Keeping score at a baseball game with a typewriter is not only possible but is also a much more detailed record of the match. (ORTEGA. Full count! Fouled back three in a row ... OH, THAT BALL’S LANDIN’ WHERE THE FANS ARE STANDIN’!!! Walk. Off. Home. Run. Thanks for your attendance and drive safely.)
Tom Hanks was using typewriters to score baseball games since at least 2013!
the government’s use of data does not and cannot limit itself to focusing solely onindividual needs or rights.
Government data use has to balance individual privacy against collective benefit which means the common good lens is essential here, not just rights.
Ethical frameworks provide helpful guidance aboutwhat you should—and should not—do in relationto data projects
So ethical frameworks are guides, not rulebooks. This matches exactly what the Markkula Framework says there is no ethics algorithm. The judgment call still belongs to the human.
eLife Assessment
This study provides valuable contributions to establish canonical Dhh signaling as a primary mediator in the differentiation of Leydig cells and their steroidogenic capacity. Together, the experimental design using their established stem Leydig cell line alongside relevant genetically mutated models, both derived using the relevant Nile tilapia animal system, provided largely convincing evidence to support their conclusions. The work will be of broad interest to developmental biologists interested in differentiation of steroidogenic or hormone producing cells.
[Editors' note: this paper was reviewed by Review Commons.]
Reviewer #1 (Public review):
[Editors' note: this version has been assessed by the Reviewing Editor without further input from the original reviewers. The authors have addressed the comments raised in the previous round of review.]
Summary:
This manuscript by Zhao et. al investigates the canonical hedgehog pathway in testis development of Nile tilapia. They used complementary approaches with genetically modified tilapia and transfected TSL cells (a clonal stem Leydig cell line) previously derived from 3-mo old tilapia. The approach is innovative and provides a means to investigate DHH and each downstream component from the ptch receptors to the gli and sf1 transcription factors. They concluded that Dhh binds Ptch2 to stimulate Gli1 to promote an increase in Sf1 expression leading to the onset of 11-ketotesterone synthesis heralding the differentiation of Leydig cells in the developing male tilapia.'
Strengths of the methods and results:
- The use of Nile tilapia is important as it is an important aquaculture species, it shares the genetic pathway for sex determination of mammalian species, and molecular differentiation pathways are highly conserved<br /> - The approach is rigorous and incorporates a novel TSL, clonal stem Leydig cell model that they developed that is relatively faithful in following endogenous developmental steps and can produce the appropriate steroid.<br /> - Tilapia are relatively amenable to CRISPR/Cas9 targeting and, with their accelerated developmental time frame, provide an excellent model system to interrogate specific signaling pathways.<br /> - The stepwise analysis from dhh-gli-sf1 is thoughtful and well done.
Achieved Aims: The authors set out to test the hypothesis that the canonical Dhh signaling pathway for Leydig cell differentiation and steroidogenic activity is mediated via ptch2 and gli1 regulation of sf1. The results are strong, there are additional steps needed to verify that redundancy/compensation is not contributing to the outcomes.
This work is important in better understanding of nuanced commonalities and differences in developmental pathways across species. Specific to Leydig cell differentiation and steroidogenesis, their work with tilapia supports conservation of the canonical Dhh pathway; however, there appear to be some differences in downstream mediators compared to mouse. Specifically, they conclude that ptch2/gli1 stimulates sf1 and steroidogenesis in tilapia where gli1 is dispensable in mouse. Instead, Gli3 has recently been shown to play an important role to stimulate Sf1 and support the hedgehog pathway.
Author response:
The following is the authors’ response to the original reviews
Public Reviews:
Reviewer #1 (Public review):
Weaknesses of the methods and results:
- Line 162: need to establish and verify the PKH26-labeled TSL cells were unaffected by the dhh-/- environment. No data to support the claim that they were unaffected.
We thank the reviewer for this important comment. In dhh<sup>-/-</sup> recipient testes, PKH26-labeled TSL cells were observed within the interstitial compartment (Fig. 3C3). Importantly, these PKH26-positive cells could be induced by SAG treatment to differentiate into Cyp11c1-positive steroidogenic cells (Fig. 3E3), indicating that they remained viable in the dhh<sup>-/-</sup> environment.
We have revised the Results section (line 171–173) to “These results suggest that SLC differentiation is inhibited, whereas the survival and engraftment of PKH26-labeled TSL cells were not affected in dhh<sup>-/-</sup> XY tilapia testes.”
- The rescued phenotype caused by the addition of ptch2-/- to the dhh-/- model is a compelling. To further define potential ptch1 contributions, it would be helpful to examine the expression level of ptch1 in the context of the ptch2-/- and ptch2-/-;dhh-/- mutant animals. Any compensatory increase in ptch1 in either case, without obvious phenotype changes, would support the dominant role for ptch2.
We thank the reviewer for this valuable suggestion. We have now performed RT-qPCR analysis of ptch1 expression in XY testes from WT, ptch2<sup>-/-</sup> and dhh<sup>-/-</sup>;ptch2<sup>-/-</sup> fish at 90 dah. As shown in Fig. S8, no significant differences in ptch1 mRNA levels were detected among these genotypes, indicating that loss of ptch2 does not induce compensatory upregulation of ptch1 at the transcriptional level under the conditions examined. We have revised the Discussion section (line 277–290) to “The specificity for Ptch2 in this context might stem from unique co-receptor interactions or expression patterns within the testicular niche. To preliminarily assess potential compensatory regulation, we examined ptch1 expression in XY testes from WT, ptch2<sup>-/-</sup> and dhh<sup>-/-</sup>;ptch2<sup>-/-</sup> fish at 90 dah. No significant differences in ptch1 mRNA levels were detected among these genotypes (Fig. S8), suggesting that loss of ptch2 does not trigger compensatory upregulation of ptch1 at the transcriptional level under the conditions examined. Nonetheless, global ptch2 mutation affects multiple tissues, whereas our mechanistic focus is on SLC differentiation within the testicular niche. Moreover, the early embryonic lethality of global ptch1 mutation in tilapia (Liu et al., 2024) precludes direct assessment of its role in postnatal testis development. Therefore, although our findings strongly support a predominant role for Ptch2 in mediating Dhh signaling in SLCs, definitive resolution of receptor specificity will require future Leydig cell-specific conditional knockout models.”
- Activity of individual gli factors need additional reconciliation. The expression profiles for both alternative gli factors should be quantified in each knockout cell line to establish redundancy and/or compensation.
We agree that quantifying the expression of alternative gli genes might be informative. In the present study, TSL-gli1<sup>-/-</sup> cells completely lose responsiveness to Dhh stimulation in the 8×GLI luciferase assay, whereas TSL-gli2<sup>-/-</sup> and TSL-gli3<sup>-/-</sup> cells retain normal pathway activation (Fig. 5B), which unambiguously suggest that Gli1 is the principal transcriptional effector in tilapia SLCs under our experimental conditions. Redundancy and/or compensation of alternative gli factors need further genetic dissection in the future study.
- Figure 5E: An important control is missing that includes evaluation of HEK293 cells transfected with pcDNA3.1-OnGli1 without the addition of pGL3-sf1.
We don’t think HEK293 cells transfected with pcDNA3.1-OnGli1 without the addition of pGL3-sf1 is an important control in our study. In the dual-luciferase assays, we think pcDNA3.1 + pGL3 (empty reporter) and pcDNA3.1 + pGL3-sf1 controls were sufficient.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
Recommendations for improving the writing and presentation; minor corrections:
- Include Park paper (Endocrinology 2007) somewhere near line 73. Need to acknowledge this paper as it is one of the first to connect Dhh to Sf1.
We have now included the citation of Park et al. (Endocrinology 2007) in the Introduction (now line 81).
- Include Kothandapani paper (PLoS Genetics 2020) somewhere near line 86. Need to acknowledge this paper as it is the only to reconcile the data showing no difference in Gli1 or Gli2 knockouts, but loss of Leydig cell function due to Gli3 activity.
We have now included the citation of Kothandapani et al. (PLoS Genetics 2020) in the Introduction (now line 97).
- Please include sequences of B1 and B2 in sf1 promoter, how conserved are they to the canonical Gli binding sequence?
We have revised the Results section (line 216–218) to “Functional annotation of its promoter region identified two conserved Gli1-binding motifs, B1 (AACCACCCA) and B2 (GAGCCACCCA)”.
- Figure 1 or results text: please clarify that the dhh-/- model used is the delta13bp mutation.
We have clarified in the Results section (line 133) that the dhh<sup>-/-</sup> model corresponds to the 13-bp (CAGGGATGCGGAC) frameshift deletion.
- Figure 5E legend: please clarify that HEK293 cells are used
We have revised the Figure 5E legend to explicitly state that the dual-luciferase reporter assays were performed in HEK293 cells. Revised legend sentence (line 743-746): HEK293 cells were co-transfected with pRL-TK, pGL3, pcDNA3.1, pGL3-sf1, pcDNA3.1-On Gli1, and the indicated cold probe constructs, and luciferase activity was measured 48 hours post-transfection.
- Figure S5E: * indicates the heteroduplex-it seems that there is a heteroduplex highlighted with the asterisk at ~600bp size; based on homozygous and mutant bands, it seems the asterisk should be highlighting the duplex near those sized bands. What are the bands up at ~600bp?
We thank the reviewer for the careful observation. In Figure S5E, the bands observed at approximately ~600 bp represent heteroduplex products formed during the re-annealing of PCR amplicons derived from heterozygous individuals. During denaturation and re-annealing, WT and mutant strands can pair in different configurations, generating distinct heteroduplex conformations that migrate more slowly than homoduplex products in PAGE. As a result, two heteroduplex bands are visible at ~600 bp, reflecting alternative mismatched duplex structures. The homoduplex WT and mutant bands are indicated separately by arrows.
- Figure S7F: dhh-/- data are missing
We thank the reviewer for pointing out this omission. The missing dhh<sup>-/-</sup> dataset has now been added to Figure S7F, and the figure has been updated accordingly.
The manuscript by Tanaka et al. explores the effects of long-acting lenacapavir (LEN) on the structure of the HIV-1 capsid. To understand how LEN incorporation alters capsid architecture and viral infectivity, the authors used cryo-electron microscopy to determine the structure of mature HIV-1 capsid lattice assembled within virus-like particles formed in presence of LEN. The structural analysis suggests that LEN enforces flattened lattice geometry, leading to an altered capsid and larger viral particles. Finally, the authors confirmed the loss of reverse transcriptase activity due to a compromised capsid integrity through biochemical analysis. Overall, the data in this manuscript is well presented and supports the authors' claims. We outline major and minor adjustments to improve clarity in reporting and presentation, as well as providing additional context for a broader audience.
Major comments -While we found the data in Figure 1 (and associated Figures S4 and S7) compelling, we were concerned that when demonstrating the flattening effect of the LEN binding the capsid, all calculated angles for the LEN-negative structures come from models in the electron microscopy data bank (EMDB) rather than your own data. We understand that this may have been done to decrease time and costs. However, to ensure that flattening is not an artifact of the way the capsid was imaged, please consider including a comparison of your own atomic model of capsid without LEN. Alternatively, please elaborate in the discussion why it may not be necessary to include your own LEN(-) 3D atomic structure. -The structures in Figure 1 demonstrate the arrangement of the capsid hexamers in the presence of LEN. However, without a structure of wildtype capsid produced in the absence of LEN, it is difficult for the reader to visualize the structural changes. Please include a wildtype capsid model obtained from your own images or from the EMDB for Figure 1d,e to strengthen your argument that the structure of the hexamers is altered in the presence of LEN. -Figure 3 indicates a loss of reverse transcriptase (RT) activity, but a retention of integrase protein when wildtype capsid is treated with LEN. Given that RT presence was measured indirectly through activity rather than through a western blot or another quantitative protein method, this leaves two possible conclusions; either RT itself is being lost, or the LEN is interacting with RT in some way to reduce its efficacy. It seems unlikely that integrase would be retained with the capsid while reverse transcriptase is lost, especially since reverse transcriptase is a larger protein than integrase. Including a western blot similar to Figure 3b, e (or another quantitative protein analysis method) would allow to investigate changes in RT association with the capsid. We also recommend changing the figure caption to “LEN-containing cores in VLPs exhibit reduced integrity and lose reverse transcriptase activity” since in the current state, the figure does not explore changes in RT protein levels. -In the discussion section: “Recent preprint further reported curvature-dependent destabilization of mature HIV-1 capsids by LEN in post entry cores” references another recent preprint that demonstrates a similar loss of capsid curvature in the presence of LEN (dos Santos et al. bioRxiv 2026). We appreciated referencing a similar preprint, and including such a reference strengthened the findings of your manuscript. We recommend including a few lines in the discussion detailing how your study compares to or differentiates from the dos Santos study. - Please include your rationale for using 293T cells for VLP production and capsid structural analysis since these are not the native cell type HIV-1 infects. Do you believe the capsid structure in the presence of LEN varies if the cell type is produced in 293Ts versus a more native cell type such as MDMs or Jurkats?
Minor comments -Figure 1 a,b,c and Figure 3 i,j,k contains cryoEM and cryoET images, respectively, of VLPs produced in the presence or absence of LEN. Consider grouping or contrasting them in the text to explain the advantage of doing both analyses. -Figure S4 and Figure S7 legends contain description of which structures are LEN (-) and their origin (native virions, VLPs, etc.), but Figure 1h is lacking this information. Please state in the figure what type of structure the LEN (-) map originated from. - On Figure 2b,c,d, showing a color map of the subtraction of the +/-LEN structures would help improve understanding of the structural differences when LEN is bound. -Figure 3h uses a benzonase assay to indirectly measure how open the capsids are; if the capsids are fully closed, the benzonase cannot access the RNA and degrade it, while if the capsids are open the benzonase degrades the RNA. The y axis is titled “relative RNA to benzonase -”, but we did not understand how the RNA levels were normalized between the LEN (-), the LEN (+), and the M66I cores. Please include the raw data without normalization as the M66I virus seems to be less infectious, and the lack of difference upon benzonase-treatment could be due to less RNA overall. - Figure 4a,e demonstrates that the M66I capsid mutant is less infectious than the WT virus, but this is never addressed in the text. Stating in the text that M66I is less infectious and providing a brief explanation or hypothesis as to why, would improve reader comprehension. -The p values and standard deviation bars appear to be missing on Figure 4. Adding these in, along displaying all data points, would strengthen data presentation and improve assessment of data variability. -To increase comprehension in readers not familiar with HIV-1 research, please define Jurkat cells and VSV-G pseudotyped HIV-GFP for Figure 4, as well as a sentence about the effects of RAL and NVP on HIV-1 infection in the discussion of Figure 4. -For the readability of a broader audience please consider including an explanation for the Gag polyprotein in the introduction. -In the discussion, the unfortunate alignment of reference 2 with the pharmacologically relevant concentration of LEN leads to what looks like “5 nM2”, or five nanomolar squared. We recommend shifting the location of the reference in the sentence to prevent any potential misunderstandings. -In the methods for the nuclease sensitivity assay, the MgCl2 has a box in place of the 2.
Camy Guenther and Josy Joseph (Indiana University Bloomington) - not prompted by a journal; this review was written within a Peer Review in Life Sciences graduate course led by Alizée Malnoë with input from group discussion including Carter Collins, Lily Pumphrey and Tahreem Zaheer. None of the authors of this review have extensive expertise in Cryo-EM methodology, and our comments are made with respect to that. We are part of the Dept. of Biology where Adarsh Dharan’s group is located, and Camy Guenther is a graduate student from his group. Adarsh is a collaborator at the Pittsburg Center for HIV Protein Interactions with Barbie Ganser-Pornillos (corresponding author of ref. 42); this interaction did not influence the choice of this preprint for our class.
The corporation itself was ages old, but the actual rightto incorporate had generally been reserved for public works projects orgovernment-sponsored monopolies. After the Civil War, however, thecorporation, using new state incorporation laws passed during the Mar-ket Revolution of the early nineteenth century, became a legal mecha-nism for nearly any enterprise to marshal vast amounts of capital whilelimiting the liability of shareholders. By washing their hands of legal andfinancial obligations while still retaining the right to profit massively,investors flooded corporations with the capital needed to industrialize
Before the civil war, corporations existed but reserved for any gov. use like public projetcs, banks, canals,etc.
After the civil war, new state laws allowed businesses to incorperate. A corporation became a legal business structure that allowed companies to:
raise huge amounts of money from investors expand quickly build factories and railroads “Limiting the liability of shareholders”
This is the key idea.
Shareholders = investors who own stock in a company.
Limited liability means:
if the corporation failed or went into debt, investors usually only lost the money they invested, but their personal property and savings were protected.
Example: If a railroad corporation collapsed:
shareholders might lose their stock investment, but they would not personally owe all the company’s debts.
This made investing much less risky.
and corpo-rate bureaucracies meanwhile allowed for the management of giant newfirms.
This means that as companies became much larger during industrialization, they created corporate bureaucracies to help organize and manage everything.
A bureaucracy is a system with:
many levels of managers departments rules paperwork specialized jobs
Large corporations could no longer be run by just one owner making every decision.
So companies created:
supervisors managers accounting departments human resources sales divisions executive offices
or Taylorism,
What was Taylorism?
Taylor believed work should be studied scientifically to find:
the fastest way to do a task the most efficient movements the best way to organize workers
Managers would:
time workers with stopwatches break jobs into small repetitive tasks set strict rules for productivity
Nabiullina was much praised by the Western technocratic elite and given invitations to Jackson Hole, Basel and the IMF. There, she was fêted for her efforts to defend liberal economic orthodoxy in Russia’s challenging environment as an emerging economy; Christine Lagarde, then managing director of the IMF, said she made ‘central banking sing’.
This explains a lot.
eLife Assessment
This important study provides a comprehensive multi-omics characterization of Leishmania donovani stage differentiation, offering insights into the molecular basis of parasite adaptation across host environments. The authors present convincing evidence that stage transitions are not driven by genomic variation but instead rely on coordinated post-transcriptional regulation, including mRNA turnover, translation, and protein degradation. Although experimental validation of these findings and conclusions remains to be completed, the integration of diverse, high-quality datasets establishes a robust resource that will be of broad utility to researchers investigating Leishmania biology and life-cycle progression.
[Editors' note: this paper was reviewed by Review Commons.]
Reviewer #1 (Public review):
[Editors' note: this version has been assessed by the Reviewing Editor without further input from the original reviewers. The authors have addressed the comments raised in the previous round of review.]
Summary:
The authors describe co-regulated gene modules underlying stage differentiation in Leishmania donovani through a system-level analysis of multiple molecular layers. Using amastigotes isolated from infected hamster spleens and corresponding culture-derived promastigotes, they analyzed genomic variation, transcript abundance, protein levels, phosphorylation states, and metabolite profiles. By combining these, the study identified potential regulatory mechanisms associated with parasite differentiation and generated hypotheses regarding how gene expression is coordinated across different levels.
Strengths:
A major strength of the study is the breadth of the dataset generated. The integration provides an unusually comprehensive view of molecular changes associated with Leishmania differentiation in vitro. Such multi-layer datasets involving bona fide vertebrate host stages remain relatively rare in parasitology and will likely become a valuable resource for the molecular parasitology community. In addition, the use of amastigotes isolated from infected hamsters rather than relying on axenic models provided a biologically relevant framework for the analyses.
The revised manuscript improved several aspects of the original. The RNA-seq analysis is described with a clearer pipeline, and several claims regarding causal regulatory feedback associations have been appropriately toned down. Among the observations reported, the association between parasite differentiation and proteasome-mediated protein degradation is particularly remarkable. The combination of quantitative proteomics with pharmacological inhibition of the proteasome with lactacystin provides support for a role for protein turnover in developmental transitions and paves the way for future mechanistic studies.
Weaknesses:
Most regulatory interpretations remain largely inferential or indirect. The integration identifies correlations between different levels, but direct functional validation is limited/absent. Many of the descriptions should not be interpreted as validated. As highlighted by the authors in this revised version, the mechanistic studies will be part of future work and are beyond the scope of the current work. Of note, the attempt to confirm lactacystin-induced inhibition of proteasomal activity via anti-polyUb immunoblotting did not demonstrate the expected outcome of increase in overall poly-ubiquitylation.Editors' note: this version has been assessed by the Reviewing Editor without further input from the original reviewers. The authors have addressed the comments raised in the previous round of review.]
Reviewer #2 (Public review):
Pescher and colleagues present a revised manuscript detailing the multi-omic characterisation of Leishmania donovani amastigote to promastigote differentiation and integration of this data. The molecular pathways that regulate Leishmania life-stage transitions are still poorly understood, with many approaches exploring single proteins/RNAs etc in a reductionist manner. This paper takes a systems-scale approach and does a good job of integrating the disparate -omics datasets to generate hypotheses about the intersections of regulatory proteins that are associated with life-cycle progression. The differentiation step studied is from amastigote to promastigote using hamster-derived amastigotes which is a major strength. The use of hamsters permits the extraction of parasites that are host adapted and represent "normal", host-adapted Leishmania ploidy; the promastigote experiments are performed at a low passage number. Therefore, this is a strength or the work as it reduces the interference from the biological plasticity of Leishmania when it is cultured outside the host for prolonged periods. The multi-omics datasets presented are robust in their acquisition and analysis and will form an excellent resource for researchers studying the molecular events (particularly proteasomal protein degradation, and phosphorylation) during life-stage progression.
Overall, in the absence of follow up experiments on specific individual examples, some of the claims in the original submission were toned down and reflect a more neutral description of the data now. Significantly, the data still underpin a key role for regulation of the ribosome between the amastigote and promastigote stages (and during the differentiation process). The recursive and reciprocal links between the phosphorylation and ubiquitination systems are interesting and present many opportunities for future investigation.
Reviewer #3 (Public review):
Summary:
The authors proposed to use 5-layer systems level analysis (genomics, transcriptomics, proteomics / protein degradation, metabolomics, phosphoproteomics) to uncover how post-transcriptional mechanisms regulate stage differentiation in Leishmania donovani.<br /> This enabled the identification of several potential regulatory networks, including the regulation of stage-specific gene clusters by RNA stabilisation or decay, proteasomal degradation and protein phosphorylation.
In the new version of this manuscript, the authors have addressed all questions raised by the reviewers.
Strengths:
Although some observations in this study have already been described in the literature, the integrated analysis applied here provides a novel view on how different levels of post-transcriptional networks regulate Leishmania differentiation. This "5-layer system" represents the first analysis of this depth in kinetoplastid parasites.<br /> The revised version with an increased sample number for the RNA-seq now made the authors assumptions adequate to their obtained data.<br /> The use of a proteasomal inhibitor adds an interesting insight in how protein degradation is involved in the parasite differentiation, confirming previous observations in the literature, and help to explain the discrepancies between mRNA and protein expression in the different stages.
Weaknesses:
While this work provides an impressive and foundational dataset, it opens the door for future research to rigorously validate these initial findings and conclusions.
Significance and Impact in the field.
The different datasets generated in this study will be of great interest to the parasitology community, either to be used for hypothesis generation, to validate data from other sources, etc.
The multi-layered analysis performed here identified a series of potential feedback loops and regulatory networks to be further explored in organisms that lack transcriptional control.
Author response:
The following is the authors’ response to the original reviews
Public Reviews:
Reviewer #1 (Public review):
Comments on revised version:
The authors have appropriately addressed my comments and questions from the initial review process. My remaining concern relates to the lack of evidence to confirm proteasomal inhibition by lactacystin in both promastigotes and amastigotes. The immunoblotting experiment newly presented does not reveal a clear increase in the levels of poly-ubiquitylated proteins in treated parasites. In fact, poly-Ub levels were lower at both the 4h and 18h timepoints of treatment. If alternative antibodies or additional immunoblots are not available, the manuscript would benefit from an expanded discussion of this observation and potential explanations. In particular, the interpretation that lactacystin stabilizes ama- and pro-specific degradation would be greatly strengthened by such validation.
Reviewer #2 (Public review):
General comments on the revisions:
My view is that the authors have made significant, satisfactory changes that address the comments and queries I made on the original manuscript (Review Commons).
There are two areas where the authors had to make major changes/justifications where further comment is merited, these were:
RNA-seq.
The most significant issue was the originally underpowered RNA-seq which had only two replicates. This has been repeated with four replicates now. This has not led to changes in the interpretation of the data between the original study and this one. One comment that the authors make in the response to this was : "Given the robustness of the stage-specific transcriptome, and the legal constrains associated with the use of animals, we chose to limit the number of replicates to the necessary". Ensuring that animal experiments are properly powered and that maximum robustness of the data from the minimum sample size is an important part of experimental design for ethical use of animal models. Essentially the replication here could have been avoided if the original study had used 1 more animal. However, the new version of RNA-seq brings appropriate confidence to the interpretation of the data.
Phosphoproteomics.
The authors provide a robust justification of their strategy for the phosphoproteomics and highlight the inclusion criteria for phosphosites: "Phosphosites were only considered if detected with high confidence (identification FDR<1%) and high localisation confidence (localisation probability >0.75) in at least one replicate". The way missing values were dealt with is explained "For statistical analyses, missing values within a given condition were imputed with a well-established algorithm (MLE) only when at least one observed value was present in that condition." This fills in some of the gaps I was missing from the original manuscript, and I am satisfied that the data analysis is entirely appropriate for a discovery/system -based approach such as this one. The authors also edit the manuscript to reflect that "occupancy" or "stoichiometry" might not be the best description of what they were presenting and switched to the terminology of "normalised phosphorylation level" - I think this is an appropriate response.
Overall, in the absence of follow up experiments on specific individual examples, some of the claims in the original submission were toned down and reflect a more neutral description of the data now. Significantly, the data still underpin a key role for regulation of the ribosome between the amastigote and promastigote stages (and during the differentiation process). The recursive and reciprocal links between the phosphorylation and ubiquitination systems are interesting and present many opportunities for future investigation.
Reviewer #3 (Public review):
Summary:
The authors proposed to use 5-layer systems level analysis (genomics, transcriptomics, proteomics / protein degradation, metabolomics, phosphoproteomics) to uncover how post-transcriptional mechanisms regulate stage differentiation in Leishmania donovani.<br /> This enabled the identification of several potential regulatory networks, including the regulation of stage-specific gene clusters by RNA stabilisation or decay, proteasomal degradation and protein phosphorylation.
In the new version of this manuscript, the authors have addressed all questions raised by the reviewers.
Strengths:
Although some observations in this study have already been described in the literature, the integrated analysis applied here provides a novel view on how different levels of post-transcriptional networks regulate Leishmania differentiation. This "5-layer system" represents the first analysis of this depth in kinetoplastid parasites.
The revised version with an increased sample number for the RNA-seq now made the authors assumptions adequate to their obtained data.
The use of a proteasomal inhibitor adds an interesting insight in how protein degradation is involved in the parasite differentiation, confirming previous observations in the literature, and help to explain the discrepancies between mRNA and protein expression in the different stages.
Weaknesses:
While this work provides an impressive and foundational dataset, it opens the door for future research to rigorously validate these initial findings and conclusions.
Significance and Impact in the field.
The different datasets generated in this study will be of great interest to the parasitology community, either to be used for hypothesis generation, to validate data from other sources, etc.
The multi-layered analysis performed here identified a series of potential feedback loops and regulatory networks to be further explored in organisms that lack transcriptional control.
According to the reviewers’ comments, we made the following minor changes:
As suggested by reviewer 1, we have extended the discussion of the results related to the analysis of the ubiquitination pattern by Western blot analysis as follows: “Proteasome inhibition blocked amastigote-to-promastigote differentiation, without inducing rapid global accumulation of ubiquitinated proteins (Figure S7C, upper panel) consistent with a quiescent-like state and low basal ubiquitin–proteasome system activity in amastigotes. After 18 h, ubiquitination levels remained similar to untreated cells, indicating that protein turnover and ubiquitin accumulation are primarily driven by developmental remodeling rather than acute proteasome inhibition. In promastigotes, the lack of detectable change (Fig. S7C, lower panel) may also reflect high basal ubiquitination, engagement of compensatory pathways such as autophagy, and/or only partial proteasome inhibition.”
Recommendations for the authors:
Reviewer #3 (Recommendations for the authors):
Minor comments:
- Supplementary figure 3 is not referenced in the main text.
- The authors removed the "infinite" sign from figures 3 and 4 to better present the data according to their chosen approach to missing values when LFQ=0. However, the sign is still present in the respective figure legends, please adjust.
Supplementary Figure 3 (Figure S3) is now referenced in the main text as requested.
The "infinite" sign has been removed from the legends of Figures 3 and 4 as requested.
eLife Assessment
This study provides valuable insights into mitochondrial cristae organization in Plasmodium falciparum, particularly in the context of its divergent MICOS composition. The authors present convincing evidence, supported by phenotypic and morphological analyses, that cristae junction maintenance can be uncoupled from de novo cristae formation, reinforcing an emerging model of mitochondrial inner membrane organization. Notably, the absence of Mic10 alongside an enlarged and divergent MICOS complex highlights an intriguing evolutionary adaptation, although further characterization of the complex would strengthen the study's overall significance.
[Editors' note: this paper was reviewed by Review Commons.]
Reviewer #1 (Public review):
The manuscript by Tassan-Lugrezin et al. confirms the existence of the MICOS complex in the causative agent of malaria Plasmodium falciparum. Prior to this study, only one of the two core MICOS subunits, Mic60, was found by homology search to be encoded in the apicomplexan parasite's genome. This study demonstrates the absence of the other core subunit, Mic10. It also identifies another MICOS subunit, Mic19, which co-migrates with Mic60 in a very large molecular weight complex upon blue native polyacrylamide gel electrophoresis. The authors then demonstrate that expression of both Mic60 and Mic19 is considerably upregulated during the differentiation of P. falciparum from the pathogenic asexual blood stage (ABS) to gametocytes, which correlates with the activation of oxidative phosphorylation during this process. While gene deletion of Mic19, Mic60 and both simultaneously does not affect this transition, the crista are nevertheless deformed. More significantly, crista junctions are significantly reduced, indicating that MICOS serves the same function in apicomplexans as it does in opisthokonts: maintaining crista junctions. Furthermore, the genetic interaction of mic60 and mic19 observed by augmented crista deformation when both are deleted is further evidence of their biochemical interaction, further supporting their similar complexome profiles. This study represents an important contribution to our understanding of MICOS evolution. Furthermore, the study shows that proper cristae formation is not essential for Plasmodium life cycle progression under in vitro conditions. Moreover, mutant gametocytes are still able to mate in the mosquito vector, albeit with lower efficiency.
Strengths:
The study is a result of a lot of technically challenging work in the model Plamsodium. The technically difficult life cycle progression experiments are well performed as far as I can tell. The electron microscopy is very well done and rigorously analyzed to obtain information about crista parameters. In particular, the authors were able to quantify the occurrence and diameter of crista junctions, which is very challenging in small mitochondria with small cristae. Finally, the authors provide convincing support that Mic60 and the newly discovered Mic19 act to shape crista junctions and MICOS can apparently carry out this function without the core subunit Mic10.
Weaknesses:
In its current form, there are conceptual weaknesses. The authors focus on the development of crista from a highly likely acristate state. This is true. But there can be more insight by considering their result in light of discovering the first functioning MICOS complex without one of its two core proteins, Mic10. The surprisingly large size of is also not really considered by the authors. This brings me the second weakness in my opinion. While I think the study represents a lot of work utilizing appropriate and crucial experiments, it seems the Complexome data was not explored enough. This data revealed Mic19, but what other potential subunits are co-migrating with Mic60 and Mic19 that can explain the large size of Plasmodium MICOS? Also, what is the consequence of the loss of Mic60 and Mic19 on the mitoproteome? Perhaps other MICOS subunits can be identified by their depletion in the knockouts versus the parental cell line.
Comments on latest version:
I am reviewing this manuscript again after reviewing it for Reviewers Commons. I appreciate the author's responses to my comments. The new version is improved but, in my opinion, still needs more work.
These revisions are changes to text, interpretations and obtaining more data from existing data or databases. I do still think one experimental control is necessary to substantiate the authors claim about membrane potential.
Reviewer #2 (Public review):
This manuscript reports the identification of putative orthologues of mitochondrial contact site and cristae organizing system (MICOS) proteins in Plasmodium falciparum - an organism that unusually shows an acristate mitochondrion during the asexual part of its life cycle and then develops cristae as it enters the sexual stage of its life cycle and beyond into the mosquito. The authors identify PfMIC60 and PfMIC19 as putative members and study these in detail. The authors add HA tags to both proteins and look for timing of expression during the parasite life cycle and attempt (unsuccessfully) to localise them within the parasite - lack of signal concluded to be reflect very low expression levels. They also genetically delete both genes singly and in parallel and phenotype the effect on parasite development. They show that both proteins are expressed in gametocytes and not asexuals, suggesting they are present at the same time as cristae development. They also show that the proteins are dispensable for the entire parasite life cycle investigated (asexuals through to sporozoites), however there is some reduction in mosquito transmission. Using mitotracker labelling, the authors observe differences in mitochondrial organisation in gametocytes compared to the transgenic lines. Further investigation at higher resolution using EM techniques, shows data supporting their hypothesis that PfMIC60 and PfMIC19 are important for organising the parasite mitochondrion.
The manuscript is interesting and is an intriguing use of a well-studied organism of medical importance to answer fundamental biological questions. Given the essentiality of mitochondrial respiration for parasite survival in the mosquito, it is surprising that the single and double knock-out transgenics do not give a severe phenotype. However, the authors have been rigorous in characterizing the impact of genetic deletion of both genes throughout the parasite life cycle. Subtle differences in mitochondrial organisation were observed, consistent with their hypothesis that PfMIC60 and PfMIC19 play roles in mitochondrial organisation. Therefore, these data presented give new insights into an organelle that dramatically changes during parasite development and adds to our knowledge of mitochondrial biology in a highly unusual organism.
Comments on revised version:
I previously reviewed this manuscript for Review Commons. This version is greatly improved and the authors should be commended for addressing all comments raised.
Reviewer #3 (Public review):
Summary:
MICOS is a conserved mitochondrial protein complex responsible for organising the mitochondrial inner membrane and the maintenance of cristae junctions. This study sheds first light on the role of two MICOS subunits (Mic60 and the newly annotated Mic19) in the malaria parasite Plasmodium falciparum, which forms cristae de novo during sexual development, as demonstrated by EM of thin section and electron tomography. By generating knockout lines (including a double knockout), the authors demonstrate that knockout of both MICOS subunits leads to defects in cristae morphology and a partial loss of cristae junctions. With a formidable set of parasitological assays, the authors show that despite the metabolically important role of mitochondria for gametocytes, the knockout lines can progress through the life stages and form sporozoites, albeit with diminished infection efficiency.
Major comments (from the previous round of review):
(1) The authors should improve to present their findings in the right context, in particular by:
(i) giving a clearer description in the introduction of what is already known about the role of MICOS. This starts in the introduction, where one main finding is missing: loss of MICOS leads to loss of cristae junctions and the detachment of cristae membranes, which are nevertheless formed, but become membrane vesicles. This needs to be clearly stated in the introduction to allow the reader to understand the consistency of the authors' findings in P. falciparum with previous reports in the literature.
(ii) at the end to the introduction, the motivating hypothesis is formulated ad hoc "conclusive evidence about its involvement in the initial formation of cristae is still lacking" (line 83). If there is evidence in the literature that MICOS is strictly required for cristae formation in any organism, then this should be explained, because the bona fide role of MICOS is maintenance of cristae junctions (the hypothesis is still plausible and its testing important).
(2) Line 96-97: "Interestingly, PfMIC60 is much larger than the human MICOS counterpart, with a large, poorly predicted N-terminal extension." This statement is lacking a reference and presumably refers to annotated ORFs. The authors should clarify if the true N-terminus is definitely known - a 120kDa size is shown for the P. falciparum, but this is not compared to the expected length or the size in S. cerevisiae.
(3) lines 244-245: "Furthermore, our data indicates the effect size increases with simultaneous ablation of both proteins?". The authors should explain which data they are referring to, as some of the data in Figs 3 and 4 look similar and all significance tests relate to the wild type, not between the different mutants, so it is not clear if any overserved differences are significant. The authors repeat this claim in the discussion in lines 368-369 without referring to a specific significance test. This needs to be clarified.
(4) lines 304-306: "Though well established as the cristae organizing system, the role of MICOS in initial formation of cristae remains hidden in model organisms that constitutively display cristae.". This sentence is misleading since even in organisms that display numerous cristae throughout their life cycle, new cristae are being formed as the cells proliferate. Thus, failure to produce cristae in MICOS knockout lines would have been observable but has apparently not been reported in the literature. Thus, the concerted process in P. falciparum makes it a great model organism, but not fundamentally different to what has been studied before in other organisms.
(5) lines 373-378. "where ablation of just MIC60 is sufficient to deplete functionality of the entire MICOS (11, 15),". The authors' claim appears to be contrary to what is actually stated in ref 15, which they cite:
"MICOS subunits have non-redundant functions as the absence of both MICOS subcomplexes results in more severe morphological and respiratory growth defects than deletion of single MICOS subunits or subcomplexes."
This seems in line with what the authors show, rather than "different".
(6) lines 380-385: "... thus suggesting that membrane invaginations still arise but are not properly arranged in these knockout lines. This suggests that MICOS either isn't fully depleted,...". These conclusions are incompatible with findings from ref. 15, which the authors cite. In that study, the authors generated a ∆MICOS line which still forms membrane invaginations, showing that MICOS is not required at all for this process in yeast. Hence the authors' implication that MICOS needs to be fully depleted before membrane invaginations cease to occur is not supported by the literature.
(7) The authors should consider if the first part of their title could be seen as misleading: It suggests that MICOS is "the architect" in cristae formation, but this is not consistent with the literature nor their own findings.
Significance:
The main strength of the study is that it provides the first characterisation of the MICOS complex in P. falciparum, a human parasite in which the mitochondrion has been shown to be a drug target. Mic60 and the newly annotated Mic19 are confirmed to be essential for proper cristae formation and morphology, as well as overall mitochondrial morphology. Furthermore, the mutant lines are characterised for their ability to complete the parasite life cycle and defects in infection effectivity are observed. This work is an important first step for deciphering the role of MICOS in the malaria parasite and the composition and function of this complex in this organism.
The limitation of the study stems from what is already known about MICOS and its subunits in other organisms. MICOS subunit knockouts have been characterised in great detail in yeast and humans with similar findings regarding loss of cristae and cristae defects. The findings of this study do not provide dramatic new insight on MICOS function or go substantially beyond the vast existing literature in terms of the extent of the study, which focuses on parasitological assays and morphological analysis.
Exploring the role of MICOS in an early-divergent organism and human parasite is however important given the divergence found in mitochondrial biology and P. falciparum is a uniquely suited model system. One aspect that would increase the impact of the paper would be if the authors could mechanistically link the observed morphological defects to the decreased infection efficiency, e.g. by probing effects on mitochondrial function. This will likely be challenging as the morphological defects are diverse and the fitness defects appear moderate/mild.
The advance presented in this study is to pioneer the study of MICOS in P. falciparum, thus widening our understanding of the role of this complex to different model organism. This study will likely be mainly of interest for specialised audiences such as basic research parasitologists and mitochondrial biologists. My own field of expertise is mitochondrial biology and structural biology.
Comments on revised version:
The authors have addressed my all of my previous comments in the updated manuscript version.
Author response:
Reviewer #1 (Evidence, reproducibility and clarity):
Summary:
This manuscript reports the identification of putative orthologues of mitochondrial contact site and cristae organizing system (MICOS) proteins in Plasmodium falciparum - an organism that unusually shows an acristate mitochondrion during the asexual part of its life cycle and then this develops cristae as it enters the sexual stage of its life cycle and beyond into the mosquito. The authors identify PfMIC60 and PfMIC19 as putative members and study these in detail. The authors at HA tags to both proteins and look for timing of expression during the parasite life cycle and attempt (unsuccessfully) to localise them within the parasite. They also genetically deleted both gene singly and in parallel and phenotyped the effect on parasite development. They show that both proteins are expressed in gametocytes and not asexuals, suggesting they are present at the same time as cristae development. They also show that the proteins are dispensible for the entire parasite life cycle investigated (asexuals through to sporozoites), however there is some reduction in mosquito transmission. Using EM techniques they show that the morphology of gametocyte mitochondria is abnormal in the knockout lines, although there is great variation.
Major comments:
The manuscript is interesting and is an intriguing use of a well studied organism of medical importance to answer fundamental biological questions. My main comments are that there should be greater detail in areas around methodology and statistical tests used. Also, the mosquito transmission assays (which are notoriously difficult to perform) show substantial variation between replicates and the statistical tests and data presentation are not clear enough to conclude the reduction in transmission that is claimed. Perhaps this could be improved with clearer text?
We would like to thank the reviewer for taking the time to review our manuscript. We are happy to hear the reviewer thinks the manuscript is interesting and thank the reviewer for their constructive feedback.
To clarify the statistical analyses used, we included a new supplementary dataset with all statistical analyses and p-values indicated per graph. Furthermore, figure legends now include the information on the exact statistical test used in each case.
Regarding mosquito experiments, while we indeed reported a reduction in transmission and oocysts numbers, we are aware that this effect might be due to the high variability in mosquito feeding assays. To highlight this point, we deleted the sentence “with the transmission reduction of [numbers]….” and we included the sentence “The high variability encountered in the standard membrane feeding assays, though, partially obstructs a clear conclusion on the biological relevance of the observed reduction in oocyst numbers“
More specific comments to address:
Line 101/Fig1E (and figure legend) - What is this heatmap showing. It would be helpful to have a sentence or two linking it to a specific methodology. I could not find details in the M+M section and "specialized, high molecular mass gels" does not adequately explain what experiments were performed. The reference to Supplementary Information 1 also did not provide information.
We added the information “high molecular mass gels with lower acrylamide percentage” to clarify methodology in the text. Furthermore, we extended the figure legend to include all relevant information. Further experimental details can be found in the study cited in this context, where the dataset originates from (Evers et al., 2021).
Line 115 and Supplementary Figure 2C + D - The main text says that the transgenic parasites contained a mitochondrially localized mScarlet for visualization and localization, but in the supplementary figure 2 it shows mitotracker labelling rather than mScarlet. This is very confusing. The figure legend also mentions both mScarlet and MitoTracker. I assume that mScarlet was used to view in regular IFAs (Fig S2C) and the MitoTracker was used for the expansion microscopy (Fig S2D)?
Please clarify.
We thank the reviewer for pointing this out – this was indeed incorrectly annotated. We used the endogenous mito-mScarlet signal in IFA and mitoTracker in U-ExM. The figure annotation has now been corrected.
Figure 2C - what is the statistical test being used (the methods say "Mean oocysts per midgut and statistical significance were calculated using a generalized linear mixed effect model with a random experiment effect under a negative binomial distribution." but what test is this?)?
The statistic test is now included in the material and method section with the sentence “The fitted model was used to obtain estimated means and contrasts and were evaluated using Wald Statistics”. The test is now also mentioned in the figure legend.
Also the choice of a log10 scale for oocyst intensity is an unusual choice - how are the mosquitoes with 0 oocysts being represented on this graph? It looks like they are being plotted at 10^-1 (which would be 0.1 oocysts in a mosquito which would be impossible).
As the data spans three orders of magnitude with low values being biologically meaningful, we decided that a log scale would best facilitate readability of the graph. As the 0 values are also important to show, we went with a standard approach to handle 0s in log transformed data and substituted the 0s with a small value (0.001). We apologize for not mentioning this transformation in the manuscript. To make this transformation transparent, we added a break at the lower end of the log-scaled y-axis and relabelled the lowest tick as ‘0’. This ensures that mosquitoes with zero oocysts are shown along the x-axis without being assigned an artificial value on the log scale. We would furthermore like to highlight that for statistics we used the true value 0 and not 0.001.
Figure 2D - it is great that the data from all feeding replicates has been shared, however it is difficult to conclude any meaningful impact in transmission with the knock-out lines when there is so much variation and so few mosquitoes dissected for some datapoints (10 mosquitoes are very small sample sizes). For example, Exp1 shows a clear decrease in mic19- transmission, but then Exp2 does not really show as great effect. Similarly, why does the double knock out have better transmission than the single knockouts? Sure there would be a greater effect?
We agree with the reviewer and with the new sentence added, as per major point, we hope we clarified the concept. Note that original Figure 2D has been moved to the supplementary information, as per minor comment of another reviewer.
Figure 3 legend - Please add which statistical test was used and the number of replicates.
Done
Figure 4 legend - Please add which statistical test was used and the number of replicates.
Done. Regarding replicates, note that while we measured over 100 cristae from over 30 mitochondria, these all stem from the same parasite culture.
Figure 5C - the 3D reconstructions are very nice, but what does the red and yellow coloring show?
Indeed, the information was missing. We added it to the figure legend.
Line 352 - "Still, it is striking that, despite the pronounced morphological phenotype, and the possibly high mitochondrial stress levels, the parasites appeared mostly unaffected in life cycle propagation, raising questions about the functional relevance of mitochondria at these stages."
How do the authors reconcile this statement with the proven fact that mitochondria-targeted antimalarials (such as atovaquone) are very potent inhibitors of parasite mosquito transmission?
Our original sentence was reductive. What we wanted to state was related to the functional relevance of crista architecture and overall mitochondrial morphology rather than the general functional relevance of the mitochondria. We changed the sentence accordingly.
Furthermore, even though we do not discuss this in the article, we are aware of mitochondria targeting drugs that are known to block mosquito transmission. We want to point out that it is difficult to discern the disruption of ETC and therefore an impact on energy conversion with the impact on the essential pathway of pyrimidine synthesis, highly relevant in microgamete formation. Still, a recent paper from Sparkes et al. 2024 showed the essentiality of mitochondrial ATP synthesis during gametogenesis so it is very likely that the mitochondrial energy conversion is highly relevant for transmission to the mosquito.
Reviewer #1 (Significance):
This manuscript is a novel approach to studying mitochondrial biology and does open a lot of unanswered questions for further research directions. Currently there are limitations in the use of statistical tests and detail of methodology, but these could be easily be addressed with a bit more analysis/better explanation in the text.
This manuscript could be of interest to readers with a general interest in mitochondrial cell biology and those within the specific field of Plasmodium research.
My expertise is in Plasmodium cell biology.
We thank the reviewer for the praise.
Reviewer #2 (Evidence, reproducibility and clarity):
Major comments:
(1) In my opinion, the authors tend to sensationalize or overinterpret their results. The title of the manuscript is very misleading. While MICOS is certainly important for crista formation, it is not the only factor, as ATP synthase dimer rows make a highly significant contribution to crista morphology. Thus, one can argue with equal validity that ATP synthase should be considered the 'architect', as it's the conformation of the dimers and rows modulate positive curvature. Secondly, while cristae are still formed upon mic60/mic19 gene knockout (KO), they are severely deformed, and likely dysfunctional (see below). Thus, I do not agree with the title that MICOS is dispensable for crista formation, because the authors results show that it clearly is essential. So, the title should be changed.
We thank the reviewer for taking the time to review our manuscript.
Based on the reviewers’ interpretation we conclude the title does not come across as intended. We have changed the title to: “The role of MICOS in organizing mitochondrial cristae in malaria parasites”
The Discussion section starting from line 373 also suffers from overinterpretation as well as being repetitive and hard to understand. The authors infer that MICOS stability is compromised less in the single KOs (sKO) in compared to the mic60/mic19 double KO (dKO). MICOS stability was never directly addressed here and the composition of the MICOS complex is unaddressed, so it does not make sense to speculate by such tenuous connections. The data suggest to me that mic60 and mic19 are equally important for crista formation and crista junction (CJ) stabilization, and the dKO has a more severe phenotype than either KO, further demonstrating neither is epistatic.
We do agree with the reviewer’s notion that we did not address complex stability, and our wording did not make this sufficiently clear. We shortened and rephrased the paragraph in question.
The following paragraphs (line 387 to 422) continues with such unnecessary overinterpretation to the point that it is confusing and contradictory. Line 387 mentions an 'almost complete loss of CJs' and then line 411 mentions an increase in CJ diameter, both upon Mic60 ablation. I do not think this discussion brings any added value to the manuscript and should be shortened. Yes, maybe there are other putative MICOS subunits that may linger in the KOS that are further destabilized in the dKO, or maybe Mic60 remains in the mic19 KO (and vice versa) to somehow salvage more CJs, which is not possible in the dKO. It is impossible to say with confidence how ATP synthase behaves in the KOs with the current data.
We shortened this paragraph.
(2) While the authors went through impressive lengths to detect any effect on lifecycle progression, none was found except for a reduction in oocyte count. However, the authors did not address any direct effect on mitochondria, such as OXPHOS complex assembly, respiration, membrane potential. This seems like a missed opportunity, given the team's previous and very nice work mapping these complexes by complexome profiling. However, I think there are some experiments the authors can still do to address any mitochondrial defects using what they have and not resorting to complexome profiling (although this would be definitive if it is feasible):
i) Quantification of MitoTracker Red staining in WT and KOs. The authors used this dye to visualize mitochondria to assay their gross morphology, but unfortunately not to assay membrane potential in the mutants. The authors can compare relative intensities of the different mitochondria types they categorized in Fig. 3A in 20-30 cells to determine if membrane potential is affected when the cristae are deformed in the mutants. One would predict they are affected.
Interesting suggestion. As our staining and imaging conditions are suitable for such analysis (as demonstrated by Sarazin et al., 2025, https://www.biorxiv.org/content/10.1101/2025.11.27.690934v1), we performed the measurements on the same dataset which we collected for Figure 3. We did, however, not detect any difference in mitotracker intensity between the different lines. The result of this analysis is included in the new version of Supplementary figure S6.
ii) Sporozoites are shown in Fig S5. The authors can use the same set up to track their motion, with the hypothesis that they will be slower in the mutants compared to WT due to less ATP. This assumes that sporozoite mitochondria are active as in gametocytes.
While theoretically plausible and informative, we currently do not know the relevance of mitochondrial energy conversion for general sporozoite biology or specifically features of sporozoite movement. Given the required resources and time to set this experiment up and the uncertainty whether it is a relevant proxy for mitochondrial functioning, we argue it is out of scope for this manuscript.
iii) Shotgun proteomics to compare protein levels in mutants compared to WT, with the hypothesis that OXPHOS complex subunits will be destabilized in the mutants with deformed cristae. This could be indirect evidence that OXPHOS assembly is affected, resulting in destabilized subunits that fail to incorporate into their respective complexes.
While this experiment could potentially further our understanding of the interaction between MICOS and levels of OXPHOS complex subunits we argue that the indirect nature of the evidence does not justify the required investments.
To expedite resubmission, the authors can restrict the cell lines to WT and the dKO, as the latter has a stronger phenotype that the individual KOs and conclusions from this cell line are valid for overall conclusions about Plasmodium MICOS.
I will also conclude that complexome/shotgun proteomics may be a useful tool also for identifying other putative MICOS subunits by determining if proteins sharing the same complexome profile as PfMic60 and Mic19 are affected. This would address the overinterpretation problem of point 1.
(3) I am aware of the authors previous work in which they were not able to detect cristae in ABS, and thus have concluded that these are truly acristate. This can very well be true, or there can be immature cristae forms that evaded detection at the resolution they used in their volumetric EM acquisitions. The mitochondria and gametocyte cristae are pretty small anyway, so it not unreasonable to assume that putative rudimentary cristae in ABS may be even smaller still. Minute levels of sampled complex III and IV plus complex V dimers in ABS that were detected previously by the authors by complexome profiling would argue for the presence of miniscule and/or very few cristae.
I think that authors should hedge their claim that ABS is acristate by briefly stating that there still is a possibility that miniscule cristae may have been overlooked previously.
We acknowledge that we cannot demonstrate the absolute absence of any membrane irregularities along the inner mitochondrial membrane. At the same time, if such structures were present, they would be extremely small and unlikely to contain the full set of proteins characteristic of mature cristae. For this reason, we consider it appropriate to classify ABS mitochondria as acristate. To reflect the reviewer’s point while maintaining clarity for readers, we have slightly adjusted our wording in the manuscript, changing ‘fully acristate’ to ‘acristate’.
This brings me to the claim that Mic19 and Mic60 proteins are not expressed in ABS. This is based on the lack of signal from the epitope tag; a weak signal is detected in gametocytes. Thus, one can counter that Mic19 and Mic60 are also expressed, but below the expression limits of the assay, as the protein exhibits low expression levels when mitochondrial activity is upregulated.
We agree with the reviewer that the absence of a detectable epitope-tag signal does not definitively exclude low-level expression, and we have therefore replaced the term ‘absent’ with ‘undetectable’ throughout the manuscript. In context with previous findings of low-level transcripts of the proteins in a study by Lopez-Berragan et al. and Otto et al., we also added the sentence “The apparent absence could indicate that transcripts are not translated in ABS or that the proteins’ expression was below detection limits of western blot analysis.” to the discussion. At the same time, we would like to clarify that transcript levels for both genes fall within the <25th percentile, suggesting that these low values likely represent background signal rather than biologically meaningful expression. This interpretation is further supported by proteomic datasets in PlasmoDB, which report PfMIC19 and PfMIC60 expression in gametocyte and mosquito stages, but not in asexual blood stages.”
To address this point, the authors should determine of mature mic60 and mic19 mRNAs are detected in ABS in comparison to the dKO, which will lack either transcript. RT-qPCR using polyT primers can be employed to detect these transcripts. If the level of these mRNAs are equivalent to dKO in WT ABS, the authors can make a pretty strong case for the absence of cristae in ABS.
We appreciate the reviewer’s suggestion. As noted in the Discussion, existing transcriptomic datasets already show detectable MIC19 and MIC60 mRNAs in ABS. For this reason, we expect RT-qPCR to reveal low (but not absent) levels of both transcripts, unlike the true loss expected to be observed in the dKO. Because such residual signals have been reported previously and their biological relevance remains uncertain, we do not believe transcript levels alone can serve as a definitive indicator of cristae absence in ABS.
They should highlight the twin CX9C motifs that are a hallmark of Mic19 and other proteins that undergo oxidative folding via the MIA pathway. Interestingly, the Mia40 oxidoreductase that is central to MIA in yeast and animals, is absent in apicomplexans (DOI: 10.1080/19420889.2015.1094593).
Searching for the CX9C motifs is a valuable suggestion. In response to the reviewer´s suggestion we analysed the conservation of the motif in PfMIC19 and included this in a new figure panel (Figure 1 F).
Did the authors try to align Plasmodium Mic19 orthologs with conventional Mic19s? This may reveal some conserved residues within and outside of the CHCH domain.
In response to this comment we made Figure 1 F, where we show conserved residues within the CHCH domains of a broad range of MIC19 annotated sequences across the opisthokonts, and show that the Cx9C motifs are conserved also in PfMIC19. Outside the CHCH domain, we did not find any meaningful conservation, as PfMIC19 heavily diverges from opisthokont MIC19.
(5) Statistical significance. Sometimes my eyes see population differences that are considered insignificant by the statistical methods employed by the authors, eg Fig. 4E, mutants compared to WT, especially the dKO. Have the authors considered using other methods such as student t-test for pairwise comparisons?
The graphs in figures 3, 4 and 5 got a makeover, such that they now are in linear scale and violin plots (also following a suggestion from further down in the reviewer’s comments). We believe that this improves interpretability. ANOVA was kept as statistical testing to assure the correction for multiple comparisons that cannot be performed with standard t-test. A full overview of statistics and exact pvalues can also be found in the newly added supplementary information 2.
Minor comments:
Line 33. Anaerobes (eg Giardia) have mitochondria that do produce ATP, unlike aerobic mitochondria
We acknowledge that producing ATP via OXPHOS is not a characteristic of all mitochondria-like organelles (e.g. mitosomes), which is why these are typically classified separately from canonical mitochondria. When not considering mitochondria-like organelles, energy conversion is the function that the mitochondrion is most well-known for and the one associated with cristae.
Line 56: Unclear what authors mean by "canonical model of mitochondria"
To clarify we changed this to “yeast or human” model of mitochondria.
Lines 75-76: This applies to Mic10 only
We removed the “high degree of conservation in other cristate eukaryotes” statement.
Line 80: Cite DOI: 10.1016/j.cub.2020.02.053
Done
Fig 2D: I find this table difficult to read. If authors keep table format, at least get rid of 'mean' column' as this data is better depicted in 2C. I suggest depicted this data either like in 3B depicting portion of infected vs unaffected flies in all experiments, then move modified Table to supplement. Important to point out experiment 5 appears to be an outlier with reduced infectivity across all cell lines, including WT.
To clarify: the mean reported in the table indicates the mean per replicate while the mean reported in figure 2C is the overall mean for a given genotype that corrects for variability within experiments. We agree that moving the table to the supplementary data is a good idea. We decided to not include a graph for infected and non-infected mosquitoes as this information would be partially misleading, highlighting a phenotype we argue to be influenced by the strong variability.
Fig. 3C-G: I feel like these data repeatedly lead to same conclusions. These are all different ways of showing what is depicted in Fig 2B: mitochondria gross morphology is affected upon ablation of MICOS. I suggest that these graphs be moved to supplement and replaced by the beautiful images.
Thank you for the nice comment on our images. We have now moved part of the graphs to supplementary figure 6 and only kept the Relative Frequency, Sphericity and total mitochondria volume per cell in the main figure.
Line 180: Be more specific with which tubulin isoform is used as a male marker and state why this marker was used in supplemental Fig S6.
We have now specified the exact tubulin isoform used as the male gametocyte marker, both in the main text and in Supplementary Fig. S6. This is a commercial antibody previously known to work as an effective male marker, which is why we selected it for this experiment. This is now clearly stated in the manuscript.
Line 196 and Fig 3C: the word 'intensities' in this context is very ambiguous. Please choose a different term (puncta, elements, parts?). This is related to major point 2i above.
To clarify the biological effect that we can conclude form the measurement, we added an explanation about it in the respective section of the results, and we decided to replace the raw results of the plug-in readout with the deduced relative dispersion.
Line 222: Report male/female crista measurements
We added Supplementary information 2, which contains exact statistical test and outcomes on all presented quantifications as well as a per-sex statistical analysis of the data from figure 4. Correspondingly, we extended supplementary information 2 by a per-sex colour code for the thin section TEM data.
Fig. 4B-E: depict data as violin plots or scatter plots like Fig. 2C to get a better grasp of how the crista coverage is distributed. It seems like the data spread is wider in the double KO. This would also solve the problem with the standard deviation extending beyond 0%.
We changed this accordingly.
Lines 331-333: Please clarify that this applies for some, but not all MICOS subunits. Please also see major point 1 above. Also, the authors should point out that despite their structural divergence, trypanosomal cryptic mitofilins Mic34 and Mic40 are essential for parasite growth, in contrast to their findings with PfMic60 (DOI: https://doi.org/10.1101/2025.01.31.635831).
This has been changed accordingly.
Line 320: incorrect citation. Related to point 1above.
Correct citation is now included in the text.
Lines 333-335. This is related to the above. Again, some subunits appear to affect cell growth under lab conditions, and some do not. This and the previous sentence should be rewritten to reflect this.
This has been changed accordingly.
Line 343-345: The sentence and citation 45 are strange. Regarding the former, it is about CHCHD10, whose status as a bona fide MICOS subunit is very tenuous, so I would omit this. About the phenomenon observed, I think it makes more sense to write that Mic60 ablation results in partially fragmented mitochondria in yeast (Rabl et al., 2009 J Cell Biol. 185: 1047-63). A fragmented mitochondria is often a physiological response to stress. I would just rewrite as not to imply that mitochondrial fission (or fusion) is impaired in these KOs, or at least this could be one of several possibilities.
The sentence has been substituted following the indication of the reviewer. Though we still include the data of the human cells as this has also been shown in Stephens et al. 2020.
Line 373: 'This indicates' is too strong. I would say 'may suggest' as you have no proof that any of the KOs disrupts MICOS. This hypothesis can be tested by other means, but not by penetrance of a phenotype.
Done
Line 376-377; 'deplete functionality' does not make sense, especially in the context of talking about MICOS subunit stability. In my opinion, this paragraph overinterprets the KO effects on MICOS stability. None of the experiments address this phenomenon, and thus the authors should not try to interpret their results in this context. See major point 1.
We removed the sentence. Also, the entire paragraph has been shortened, restructured and wording was changed to address major point 1.
Other suggestions for added value
(1) Does Plasmodium Sam50 co-fractionate with Mic60 and Mic19 in BN PAGE (Fig. 1E)
While we did identify SAMM50 in our BN PAGE, the protein does not co-migrate with the MICOS components but instead comigrates with other components of a putative sorting and assembly machinery (SAM) complex. As SAMM50, the SAM complex and the overarching putative mitochondrial membrane space bridging (MIB) complex are not mentioned in the manuscript, we decided to not include the information in Author response image 1.
Author response image 1.
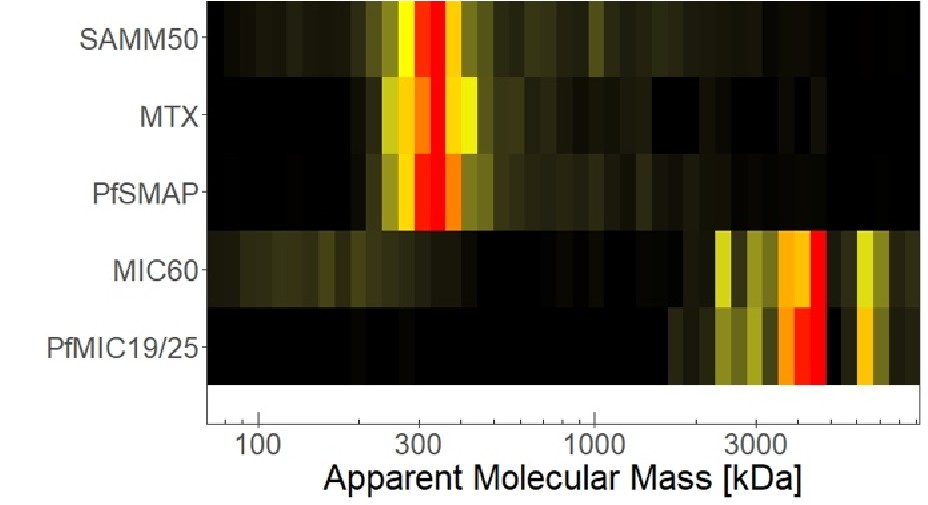
Reviewer #2 (Significance):
The manuscript by Tassan-Lugrezin is predicated on the idea that Plasmodium represents the only system in which de novo crista formation can be studied. They leverage this system to ask the question whether MICOS is essential for this process. They conclude based on their data that the answer is no, which the authors consider unprecedented. But even if their claim is true that ABS is acristate, this supposed advantage does not really bring any meaningful insight into how MICOS works in Plasmodium.
First the positives of this manuscript. As has been the case with this research team, the manuscript is very sophisticated in the experimental approaches that are made. The highlights are the beautiful and often conclusive microscopy performed by the authors. Only the localization of Mic60 and Mic19 was inconclusive due to their very low expression unfortunately.
The examination of the MICOS mutants during in vitro life cycle of Plasmodium falciparum is extremely impressive and yields convincing results. Mitochondrial deformation is tolerated by life cycle stage differentiation, with a modest but significant reduction of oocyte production, being observed.
However, despite the herculean efforts of the authors, the manuscript as it currently stands represents only a minor advance in our understanding of the evolution of MICOS, which from the title and focus of the manuscript, is the main goal of the authors.
In its current form, the manuscript reports some potentially important findings:
(1) Mic60 is verified to play a role in crista formation, as is predicted by its orthology to other characterized Mic60 orthologs.
(2) The discovery of a novel Mic19 analog (since the authors maintain there is no significant sequence homology), which exhibits a similar (or the same?) complexome profile with Mic60. This protein was upregulated in gametocytes like Mic60 and phenocopies Mic60 KO.
(3) Both of these MICOS subunits are essential (not dispensable) for proper crista formation
(4) Surprisingly, neither MICOS subunit is essential for in vitro growth or differentiation from ABS to sexual stages, and from the latter to sporozoites. This says more about the biology of plasmodium itself than anything about the essentiality of Mic60, i.e. plasmodium life cycle progression tolerates defects to mitochondrial morphology. But yes, I agree with the authors that Mic60's apparent insignificance for cell growth in examined conditions does differ with its essentiality in other eukaryotes. But fitness costs were not assayed (e.g. by competition between mutants and WT in infection of mosquitoes)
(5) Decreased fitness of the mutants is implied by a reduction of oocyte formation.
While interesting in their own way, collectively they do not represent a major advance in our understanding of MICOS evolution. Furthermore, the findings bifurcate into categories informing MICOS or Plasmodium biology. Both aspects are somewhat underdeveloped in their current form.
This is unfortunate because there seem to be many missed opportunities in the manuscript that could, with additional experiments, lead to a manuscript with much wider impact. For me, what is remarkable about Plasmodium MICOS that sets it apart from other iterations is the apparent absence of the Mic10 subunit. Purification of plasmodium MICOS via the epitope tagged Mic60 and Mic19 could have verified that MICOS is assembled without this core subunit. Perhaps Mic60 and Mic19 are the vestiges of the complex, and thus operate alone in shaping cristae. Such a reduction may also suggest the declining importance of mitochondria in plasmodium.
Another missed opportunity was to assay the impact of MICOS-depletion of OXPHOS in plasmodium.
This is a salient issue as maybe crista morphology is decoupled from OXPHOS capacity in Plasmodium, which links to the apparent tolerance of mitochondrial morphology in cell growth and differentiation. I suggested in section A experiments to address this deficit.
Finally, the authors could assay fitness costs of MICOS-ablation and associated phenotypes by assaying whether mosquito infectivity is reduced in the mutants when they are directly competing with WT plasmodium. Like the authors, I am also surprised that MICOS mutants can pass population bottlenecks represented by differentiation events. Perhaps the apparent robustness of differentiation may contribute plasmodium's remarkable ability to adapt.
I realize that the authors put a lot of efforts into their study and again, I am very impressed by the sophistication of the methods employed. Nevertheless, I think there is still better ways to increase the impact of the study aside from overinterpreting the conclusions from the data. But this would require more experiments along the lines I suggest in Section A and here.
We thank the reviewer for their extensive analysis of the significance of our findings, including the compliments on our microscopy images and the sophisticated experimental approaches. We hope we have convincingly argued why we could or could not include some of the additional analyses suggested by the reviewer in section 1 above.
With regard to the significance statement, we want to point out that our finding that PfMICOS is not needed for initial formation of cristae (as opposed to organization thereof), is a confirmation of something that has been assumed by the field, without being the actual focus of studies. We argue that the distinction between formation and organization of cristae is important and deserves some attention within the manuscript. The result of MICOS not being involved in the initial formation of cristae, we argue to be relevant in Plasmodium biology and beyond. As for the insights into how MICOS works in Plasmodium we have confirmed that the previously annotated PfMIC60 is indeed involved in the organization of cristae. Furthermore, we have identified and characterized PfMIC19. These findings, we argue, are indeed meaningful insights into PfMICOS.
Reviewer #3 (Evidence, reproducibility and clarity):
Summary:
MICOS is a conserved mitochondrial protein complex responsible for organising the mitochondrial inner membrane and the maintenance of cristae junctions. This study sheds first light on the role of two MICOS subunits (Mic60 and the newly annotated Mic19) in the malaria parasite Plasmodium falciparum, which forms cristae de novo during sexual development, as demonstrated by EM of thin section and electron tomography. By generating knockout lines (including a double knockout), the authors demonstrate that knockout of both MICOS subunits leads to defects in cristae morphology and a partial loss of cristae junctions. With a formidable set of parasitological assays, the authors show that despite the metabolically important role of mitochondria for gametocytes, the knockout lines can progress through the life stages and form sporozoites, albeit with diminished infection efficiency.
We thank the reviewer for their time and compliment.
Major comments:
(1) The authors should improve to present their findings in the right context, in particular by:
i) giving a clearer description in the introduction of what is already known about the role of MICOS. This starts in the introduction, where one main finding is missing: loss of MICOS leads to loss of cristae junctions and the detachment of cristae membranes, which are nevertheless formed, but become membrane vesicles. This needs to be clearly stated in the introduction to allow the reader to understand the consistency of the authors' findings in P. falciparum with previous reports in the literature.
We extended the introduction to include this information.
iii) at the end to the introduction, the motivating hypothesis is formulated ad hoc "conclusive evidence about its involvement in the initial formation of cristae is still lacking" (line 83). If there is evidence in the literature that MICOS is strictly required for cristae formation in any organism, then this should be explained, because the bona fide role of MICOS is maintenance of cristae junctions (the hypothesis is still plausible and its testing important).
To clarify we rephrased the sentence to: “Although MICOS has been described as an organizer of crista junctions, its role during the initial formation of nascent cristae has not been investigated.”
(2) Line 96-97: "Interestingly, PfMIC60 is much larger than the human MICOS counterpart, with a large, poorly predicted N-terminal extension." This statement is lacking a reference and presumably refers to annotated ORFs. The authors should clarify if the true N-terminus is definitely known - a 120kDa size is shown for the P. falciparum but this is not compared to the expected length or the size in S. cerevisiae.
To solve the reference issue, we added the uniprot IDs we compared to see that the annotated ORF is bigger in Plasmodium. We also changed the comparison to yeast instead of human, because we realized it is confusing to compare to yeast all throughout the figure, but then talk about human in this specific sentence.
Regarding whether the true N-terminus is known. Short answer: No, not exactly.
However, we do know that the Pf version is about double the size of the yeast protein.
As the reviewer correctly states, we show the size of 120kDa for the tagged protein in Figure 1G. Considering that we tagged the protein C-terminally, and observed a 120kDa product on western blot, it is safe to conclude that the true N-terminus does not deviate massively from the annotated ORF, and hence, that there is a considerable extension of the protein beyond a 60kDa protein. We do not directly compare to yeast MIC60 on our western blots, however, that comparison can be drawn from literature: Tarasenko et al., 2017 showed that purified MIC60 running at ~60kDa on SDS-PAGE actively bends membranes, suggesting that in its active form, the monomer of yeast MIC60 is indeed 60kDa in size.
To clarify, we now emphasize that we ran the Alphafold prediction on the annotated open reading frame (annotated and sequenced by Bohme et al. and Chapell et al. now cited in the manuscript), and revised the wording to make clear what we are comparing in which sentence.
(3) lines 244-245: "Furthermore, our data indicates the effect size increases with simultaneous ablation of both proteins?". The authors should explain which data they are referring to, as some of the data in Fig 3 and 4 look similar and all significance tests relate to the wild type, not between the different mutants, so it is not clear if any overserved differences are significant. The authors repeat this claim in the discussion in lines 368-369 without referring to a specific significance test. This needs to be clarified.
As a reply to this and other comments from the reviewers we added the multiple testing within all samples. In addition, to clarify statistics used we included a supplementary dataset with all p-values and statistical tests used.
(4) lines 304-306: "Though well established as the cristae organizing system, the role of MICOS in initial formation of cristae remains hidden in model organisms that constitutively display cristae.". This sentence is misleading since even in organisms that display numerous cristae throughout their life cycle, new cristae are being formed as the cells proliferate. Thus, failure to produce cristae in MICOS knockout lines would have been observable but has apparently not been reported in the literature. Thus, the concerted process in P. falciparum makes it a great model organism, but not fundamentally different to what has been studied before in other organisms.
We deleted this statement.
(5) lines 373-378. "where ablation of just MIC60 is sufficient to deplete functionality of the entire MICOS (11, 15),". The authors' claim appears to be contrary to what is actually stated in ref 15, which they cite:
"MICOS subunits have non-redundant functions as the absence of both MICOS subcomplexes results in more severe morphological and respiratory growth defects than deletion of single MICOS subunits or subcomplexes."
This seems in line with what the authors show, rather than "different".
This sentence has been removed.
(6) lines 380-385: "... thus suggesting that membrane invaginations still arise, but are not properly arranged in these knockout lines. This suggests that MICOS either isn't fully depleted,...". These conclusions are incompatible with findings from ref. 15, which the authors cite. In that study, the authors generated a ∆MICOS line which still forms membrane invaginations, showing that MICOS is not required at all for this process in yeast. Hence the authors' implication that MICOS needs to be fully depleted before membrane invaginations cease to occur is not supported by the literature.
This sentence has been deleted in the revised version of the manuscript.
Minor comments:
(1) The authors should consider if the first part of their title could be seen as misleading: It suggests that MICOS is "the architect" in cristae formation, but this is not consistent with the literature nor their own findings.
Title is changed accordingly
- Line 43, of the three seminal papers describing the discovery of MICOS in 2011, the authors only cite two (refs 6 and 7), but miss the third paper, Hoppins et al, PMID: 21987634, which should probably be corrected.
Done, the paper is now cited
- Page 2, line 58: for a more complete picture the authors should also cite the work of others here which shows that although at very low levels, e.g. complex III (a drug target) and ATP synthase do assemble (Nina et al, 2011, JBC).
Done
- Page 3, line 80: "Irrespective of the shape of an organism's cristae, the crista junctions have been described as tubular channels that connect the cristae membrane to the inner boundary membrane (22, 24)." This omits the slit-shaped cristae junctions found in yeast (Davies et al, 2011, PNAS), which the authors should include.
The paper and concept have been added to the manuscript, though the sentence has been moved up in the introduction, when crista junctions are first introduced.
- Line 97: "poorly predicted N-terminal extension", as there is no experimental structure, we don't know if the prediction is poor. Presumably the authors mean either poorly ordered or the absence of secondary structure elements, or the poor confidence score for that region in the prediction? This should be clarified or corrected.
We were referring to the poor confidence score. To address this comment as well as major point 2, we rewrote the respective paragraph. It now clearly states that confidence of the prediction is low, and we mention the tool that was used to identify conserved domains (Topology-based Evolutionary Domains).
- Line 98: "an antiparallel array of ten β-sheets". They are actually two parallel beta-sheets stacked together. The authors could find out the name of this fold, but the confidence of the prediction is marked a low/very low. So, its existence is unknown, not just its "function".
We adapted the domain description to “a stack of two parallel beta-sheets" and replaced the statement on unknown function by the statement “Because this domain is predicted solely from computational analysis, both its actual existence in the native protein and its biological function remain unknown.”
- Fig 1B: The authors show two alphafold predictions of S. cerevisiae and P. falciparum Mic60 structures. There is however an experimental Mic60/19 (fragment) structure from the former organism (PMID: 36044574), which should be included if possible.
We appreciate the reviewer’s suggestion and note that the available structural data indeed provides valuable insight into how MIC60 and MIC19 interact. However, these structures represent fusion constructs of limited protein fragments and therefore capture only a small portion of each protein, specifically the interaction interface. Because our aim in Fig. 1B is to compare the overall domain architecture of the full-length proteins, we believe that including fragment-based structures would be less informative in this context.
- Line: 318-321: "The same trend was observed for PfMIC19 and PfMIC60. Although transcriptomic data suggested that low-level transcripts of PfMIC19 and PfMIC60 are present in ABS (38), we did not detect either of the proteins in ABS by western blot analysis. While this statement is true, the authors should comment on the sensitivity of the respective methods - how well was the antibody working in their hands and how do they interpret the absence of a WB band compared to transcriptomics data?
The HA antibody used in our experiments is a standard commercial reagent that performs reliably in both WB and IFA, although it shows a low background signal in gametocytes. We agree that the sensitivity of the method and the interpretation of weak or absent bands should be addressed explicitly. Transcript levels for both PfMIC19 and PfMIC60 in asexual blood stages fall within the <25 percentile, suggesting that these signals likely represent background. Nevertheless, we acknowledge that low-level protein expression below the detection limit of western blot analysis cannot be excluded. To reflect these considerations, we added the sentence: ‘The apparent absence could indicate that transcripts are not translated in ABS or that the proteins’ expression was below detection limits of western blot analysis.
- Lines 322-323: would the authors not typically have expected an IFA signal given the strength of the band in Western blot? If possible, the authors should comment if the negative fluorescence outcome can indeed be explained with the low abundance or if technical challenges are an equally good explanation.
Considering the nature of the investigated proteins (embedded in the IMM and spread throughout the mitochondria) difficulties in achieving a clear signal in IFA or U-ExM are not very surprizing. While epitopes may remain buried in IFA, U-ExM usually increases accessibility for the antibodies. However, U-ExM comes at the cost of being prone to dotty background signals, therefore potentially hiding low abundance, naturally dotty signals such as the signal of MICOS proteins that localize to distinct foci (at the CJ) along the mitochondrion. Current literature suggests that, in both human and yeast, STED is the preferred method for accurate spatial resolution of MICOS proteins (https://www.ncbi.nlm.nih.gov/pubmed/32567732,https://www.ncbi.nlm.nih.gov/pubmed/3206734 4). Unfortunately, we do not have experience with, nor access to, this particular technique/method.
- Lines 357-365: the authors describe limitations of the applied methods adequately. Perhaps it would be helpful to make a similar statement about the analysis of 3D objects like mitochondria and cristae from 2D sections. E.g. the apparent cristae length depends on whether cristae are straight (e.g. coiled structures do not display long cross sections despite their true length in 3D).
The limitations of other methods are described in the respective results section.
We added a clarifying sentence in the results section of Figure 4:
“Note that such measurements do not indicate the true total length or width of cristae, as the data is two-dimensional. The recorded values are to be considered indicative of possible trends, rather than absolute dimensions of cristae.“
This statement refers to the length/width measurements of cristae.
In the context of Figure 4D we mention the following (see preprint lines 229 – 230): “We expect this effect to translate into the third dimension and thus conclude that the mean crista volume increases with the loss of either PfMIC19, PfMIC60, or both.”
For Figure 5, we included a clarifying statement in the results section of the preprint (lines 269 – 273): “Note that these mitochondrial volumes are not full mitochondria, but large segments thereof. As a result of the incompleteness of the mitochondria within the section, and the tomography specific artefact of the missing wedge, we were unable to confirm whether cristae were in fact fully detached from the boundary membrane, or just too long to fit within the observable z-range.”
- Line 404: perhaps undetected or similar would be a better description than "hidden"?
The sentence does not exist in the revised manuscript.
Reviewer #3 (Significance):
The main strength of the study is that it provides the first characterisation of the MICOS complex in P. falciparum, a human parasite in which the mitochondrion has been shown to be a drug target. Mic60 and the newly annotated Mic19 are confirmed to be essential for proper cristae formation and morphology, as well as overall mitochondrial morphology. Furthermore, the mutant lines are characterised for their ability to complete the parasite life cycle and defects in infection effectivity are observed. This work is an important first step for deciphering the role of MICOS in the malaria parasite and the composition and function of this complex in this organism. The limitation of the study stems from what is already known about MICOS and its subunits in great detail in yeast and humans with similar findings regarding loss of cristae and cristae defects. The findings of this study do not provide dramatic new insight on MICOS function or go substantially beyond the vast existing literature in terms of the extent of the study, which focuses on parasitological assays and morphological analysis. Exploring the role of MICOS in an early-divergent organism and human parasite is however important given the divergence found in mitochondrial biology and P. falciparum is a uniquely suited model system. One aspect that would increase the impact of the paper would be if the authors could mechanistically link the observed morphological defects to the decreased infection efficiency, e.g. by probing effects on mitochondrial function. This will likely be challenging as the morphological defects are diverse and the fitness defects appear moderate/mild.
As suggested by Reviewer 2, we examined mitochondrial membrane potential in gametocytes using MitoTracker staining and did not observe any obvious differences associated with the morphological defects. At present, additional assays to probe mitochondrial function in P. falciparum gametocytes are not sufficiently established, and developing and validating such methods would require substantial work before they could be applied to our mutant lines. For these reasons, a more detailed mechanistic link between the observed morphological changes and the reduced infection efficiency is currently beyond reach.
The advance presented in this study is to pioneer the study of MICOS in P. falciparum, thus widening our understanding of the role of this complex to different model organism. This study will likely be mainly of interest for specialised audiences such as basic research parasitologists and mitochondrial biologists. My own field of expertise is mitochondrial biology and structural biology.
The new cute girl, warm and fun
stolen pictures, all over..
eLife Assessment
This valuable study used genetic and pharmacological manipulations of insulin/IGF signaling to address the role of insulin/IGF axis in the function of renal glomerular podocyte. Solid data are presented to demonstrate that co-inhibition of insulin/IGF signaling in podocytes led to aberrant splicing of mRNAs, which could contribute to the loss of podocytes in vitro and in vivo in mice. In light of the fact that IR/IGF-1R signaling are critically required for normal development and growth in multiple cells and organs, the lack of the assessment of developmental phenotype of podocytes in the mouse model limits the interpretation of the data.
[Editors' note: this paper was reviewed by Review Commons.]
Reviewer #1 (Public review):
Summary:
In this manuscript, the role of the insulin receptor and the insulin growth factor receptor was investigated in podocytes. Mice, where both receptors were deleted, developed glomerular dysfunction and developed proteinuria and glomerulrosclerosis over several months. Because of concerns about incomplete KO, the authors generated and studied podocyte cell lines where both receptors were deleted. Loss of both receptors was highly deleterious with greater than 50% cell death. To elucidate the mechanism of cell death, the authors performed global proteomics and found that spliceosome proteins were downregulated. They confirmed this directly by using long-read sequencing. These results suggest a novel role for insulin and IGF1R signaling in RNA splicing in podocytes.
This is primarily a descriptive study and no technical concerns are raised. The mechanism of how insulin and IGF1 signaling regulates splicing is not directly addressed but implicates potentially the phosphorylation downstream of these receptors. In the revised manuscript, it is shown that the mouse KO is incomplete potentially explaining the slow onset of renal insufficiency. Direct measurement of GFR and serial serum creatinines might also enhance our understanding of progression of disease, proteinuria is a strong sign of renal injury. An attempt to rescue the phenotype by overexpression of SF3B4 would also be useful but may be masked by defects in other spliceosome genes. As insulin and IGF are regulators of metabolism, some assessment of metabolic parameters would be an optional add-on.
Significance:
With the GLP1 agonists providing renal protection, there is great interest in understanding the role of insulin and other incretins in kidney cell biology. It is already known that Insulin and IGFR signaling play important roles in other cells of the kidney. So, there is great interest in understanding these pathways in podocytes. The major advance is that these two pathways appear to have a role in RNA metabolism.
Latest comments:
The new reviewer raised two major points, whether the KO effect on splicing is specific to IGF1 and whether the interpretation could be developmental rather than due to splicing. The reviewer raises some important issues but the evidence to suggest that this is specific is data in the literature that IR/IGF signaling is already known to regulate splicing and that splicing defects were not detected in other models that they have analyzed. I agree with the reviewer (and authors) that the incomplete floxing of the genes is a major complication. The point that there could be a developmental defect with mice being born with fewer podocytes and perhaps the authors should caveat this point. The fact that they mice are born with normal function, that renal function can be maintained with up to 80% loss of podocytes suggest that they are likely born with a good number of podocytes and the dysfunction that occurs at 6 months is due to a process, induced by the loss of IR/IGF signaling that is detrimental to the podocyte.
Reviewer #2 (Public review):
Summary:
In this manuscript, submitted to Review Commons (journal agnostic), Coward and colleagues report on the role of insulin/IGF axis in podocyte gene transcription. They knocked out both the insulin and IGFR1 mice. Dual KO mice manifested a severe phenotype, with albuminuria, glomerulosclerosis, renal failure and death at 4-24 weeks.
Long read RNA sequencing was used to assess splicing events. Podocyte transcripts manifesting intron retention were identified. Dual knock-out podocytes manifested more transcripts with intron retention (18%) compared wild-type controls (18%), with an overlap between experiments of ~30%.
Transcript productivity was also assessed using FLAIR-mark-intron-retention software. Intron retention w seen in 18% of ciDKO podocyte transcripts compared to 14% of wild-type podocyte transcripts (P=0.004), with an overlap between experiments of ~30% (indicating the variability of results with this method). Interestingly, ciDKO podocytes showed downregulation of proteins involved in spliceosome function and RNA processing, as suggested by LC/MS and confirmed by Western blot.
Pladienolide (a spliceosome inhibitor) was cytotoxic to HeLa cells and to mouse podocytes but no toxicity was seen in murine glomerular endothelial cells.
The manuscript is generally clear and well-written. Mouse work was approved in advance. The four figures are generally well-designed, bars/superimposed dot-plots.
Methods are generally well described.
Comments on previous version:
Coward and colleagues have done an excellent job of responding to all the reviewer comments.
Reviewer #4 (Public review):
Summary and background:
This report entitled "The insulin/IGF axis is critically important (for) controlling gene transcription in the podocyte" from Hurcombe et al is based on a mouse double knockdown of the IR and IGF1R and a parallel cultured mouse podocyte model. Insulin/IGF signaling system in mammals evolved as three gene reduplicated peptides (insulin, IGF-1, and IGF-2) and their two receptors IR and IGF1R that cross-react to variable extents with the peptides, are ubiquitously expressed, and signal through parallel pathways. The major downstream effect of insulin is to regulate glucose uptake and metabolism, while that of the IGF pathways is to regulate growth and cell cycling in part through mTORC1. The GH-IGF-1-IGF1R pathway regulates post-natal growth. IGF-2 signaling is thought to play a major role in regulating intrauterine growth and development, although IGF-2 is also present at high levels in post-natal life. Thus, one would anticipate that reducing IR/IGF1R signaling in any cell would slow growth and cell cycling by reducing growth factor and metabolic mTORC1-mediated and other processes including the splicing of RNA for protein synthesis.
Comments on revised version:
The second sentence of the Summary reads "This study sought to elucidate the compound role of the insulin/IGF1 axis in podocytes using transgenic mice and cell culture models deficient in both receptors." The study design and rationale for the proteosome analysis described is predicated on the finding that podocyte-specific knockdown of the IR/IGF-1R in mice is associated with development of proteinuria and reduced eGFR by 20months of life. Since the IR/IGF-1R are critically required for normal development and growth of all cells and organs, the obvious explanation for the observation would be that the model system results in defective podocyte development and deployment (caused by reduced IR/IGF-1) that, in turn, causes subsequent development of proteinuria and glomerulosclerosis (that may be much less dependent on a normal level of IR/IGF-1R expression). Thus, the experimental design does not allow a distinction between podocyte development and steady state function which are different biologic processes. The data provided does not examine podocyte status immediately after birth to confirm that podocyte number and size and structure is normal in mice that subsequently develop proteinuria and glomerulosclerosis. The response to the reviewer suggests that since this would require additional mice it has not been undertaken in order to reduce animal usage. This is not a valid argument, particularly when the investigators have not even used state-of-the-art methods to measure podocyte number, size and density in adult mice, key parameters that would be required to interpret their data. Counting podocyte nuclear number in glomerular cross-sections is simply an inadequate method, even if it is used and reported in other journals, and particularly where the examples given to justify its use can hardly be viewed as representing first rate science.
If the absence of studies that would answer the above questions, the investigators should add a sentence to the Discussion dealing with study limitations as follows. "The study design does not allow us to determine whether the primary effect of reduced IR/IGF-1R expression on the phenotype is during in utero and post-natal podocyte development and deployment, during periods of rapid growth when IGF-1 levels are highest, in steady state adult podocytes, or under all of the above conditions".
Author response:
The following is the authors’ response to the previous reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
In this manuscript, the role of the insulin receptor and the insulin growth factor receptor was investigated in podocytes. Mice, where both receptors were deleted, developed glomerular dysfunction and developed proteinuria and glomerulrosclerosis over several months. Because of concerns about incomplete KO, the authors generated and studied podocyte cell lines where both receptors were deleted. Loss of both receptors was highly deleterious with greater than 50% cell death. To elucidate the mechanism of cell death, the authors performed global proteomics and found that spliceosome proteins were downregulated. They confirmed this directly by using long-read sequencing. These results suggest a novel role for insulin and IGF1R signaling in RNA splicing in podocytes.
This is primarily a descriptive study and no technical concerns are raised. The mechanism of how insulin and IGF1 signaling regulates splicing is not directly addressed but implicates potentially the phosphorylation downstream of these receptors. In the revised manuscript, it is shown that the mouse KO is incomplete potentially explaining the slow onset of renal insufficiency. Direct measurement of GFR and serial serum creatinines might also enhance our understanding of progression of disease, proteinuria is a strong sign of renal injury. An attempt to rescue the phenotype by overexpression of SF3B4 would also be useful but may be masked by defects in other spliceosome genes. As insulin and IGF are regulators of metabolism, some assessment of metabolic parameters would be an optional add-on.
Significance:
With the GLP1 agonists providing renal protection, there is great interest in understanding the role of insulin and other incretins in kidney cell biology. It is already known that Insulin and IGFR signaling play important roles in other cells of the kidney. So, there is great interest in understanding these pathways in podocytes. The major advance is that these two pathways appear to have a role in RNA metabolism.
Comments on revised version:
I'm satisfied with the revised manuscript and the responses to my previous concerns.
Thank you.
Reviewer #2 (Public review):
Summary:
In this manuscript, submitted to Review Commons (journal agnostic), Coward and colleagues report on the role of insulin/IGF axis in podocyte gene transcription. They knocked out both the insulin and IGFR1 mice. Dual KO mice manifested a severe phenotype, with albuminuria, glomerulosclerosis, renal failure and death at 4-24 weeks.
Long read RNA sequencing was used to assess splicing events. Podocyte transcripts manifesting intron retention were identified. Dual knock-out podocytes manifested more transcripts with intron retention (18%) compared wild-type controls (18%), with an overlap between experiments of ~30%.
Transcript productivity was also assessed using FLAIR-mark-intron-retention software. Intron retention w seen in 18% of ciDKO podocyte transcripts compared to 14% of wild-type podocyte transcripts (P=0.004), with an overlap between experiments of ~30% (indicating the variability of results with this method). Interestingly, ciDKO podocytes showed downregulation of proteins involved in spliceosome function and RNA processing, as suggested by LC/MS and confirmed by Western blot.
Pladienolide (a spliceosome inhibitor) was cytotoxic to HeLa cells and to mouse podocytes but no toxicity was seen in murine glomerular endothelial cells.
The manuscript is generally clear and well-written. Mouse work was approved in advance. The four figures are generally well-designed, bars/superimposed dot-plots.
Methods are generally well described.
Comments on revised version:
Coward and colleagues have done an excellent job of responding to all the reviewer comments.
Thank you.
Reviewer #4 (Public review):
Summary and background:
This report entitled "The insulin/IGF axis is critically important (for) controlling gene transcription in the podocyte" from Hurcombe et al is based on a mouse double knockdown of the IR and IGF1R and a parallel cultured mouse podocyte model. Insulin/IGF signaling system in mammals evolved as three gene reduplicated peptides (insulin, IGF-1, and IGF-2) and their two receptors IR and IGF1R that cross-react to variable extents with the peptides, are ubiquitously expressed, and signal through parallel pathways. The major downstream effect of insulin is to regulate glucose uptake and metabolism, while that of the IGF pathways is to regulate growth and cell cycling in part through mTORC1. The GH-IGF-1-IGF1R pathway regulates post-natal growth. IGF-2 signaling is thought to play a major role in regulating intrauterine growth and development, although IGF-2 is also present at high levels in post-natal life. Thus, one would anticipate that reducing IR/IGF1R signaling in any cell would slow growth and cell cycling by reducing growth factor and metabolic mTORC1-mediated and other processes including the splicing of RNA for protein synthesis.
Thank you for this new extra review and assessing our paper with new suggestions (we addressed the previous suggestions to the satisfaction of other reviewers). Of note -regarding this introduction – the podocyte is a terminally differentiated cell and may have unique responses to insulin / IGF as it is accepted it does not generally proliferate (hence we consider understanding the actions of insulin / IGF and their receptors to be of interest). Indeed, we have recently shown a contrasting effect of IGF signalling in the podocyte. Partial suppression of the IGF1 receptor is beneficial in contrast to near complete suppression that results in mitochondrial dysfunction (PMID:38706850).
Mouse IR/IGF1R double knockdown model:
A double knockdown mouse model was generated by interbreeding mice with different genetic backgrounds carrying floxed sites for IR and IGF-1R to produce mixed background offspring with both floxed IR and IGF-1R genes. These mice were crossed so that the podocin promoter driven-Cre (that comes on at about embryonic day 12 bas podocytes are developing) would delete IR and IGF-1R genes. Since podocin is believed to be an absolutely podocyte-specific protein, this podocin promoter this is predicted to specifically knock down the IR and IGF1R genes only in podocytes. The weight and growth of double KO offspring was not different from controls, but some proportion of the double knockdown mice subsequently developed proteinuria by 6 months and 20% died, although no specific data is provided to identify the cause of the deaths since eGFR was not decreased. Surviving mice were evaluated at 6 months of age. The efficacy of knockdown was not demonstrated in the mouse model itself, although a temperature-sensitive cell line developed from these double knockdown mice showed that expression of IR and IGF-1R proteins in the Cre-treated cell line were both reduced by about 50% (no statistical analysis of this result provided).
In the knockout mice, proteinuria was significantly increased by 6 months, but not at earlier time points. Histologic analysis showed proteinaceous casts, glomerulosclerosis and interstitial fibrosis. Podocyte number was stated to be reduced by about 30% in double knockdown mice, although the method by which this was evaluated seems to have been by counting WT1 positive nuclei in glomerular cross-sections, an approach that is well-known not to be a reliable way of assessing true podocyte number. No information is provided about podocyte size, density or glomerular volume.
Comment: If IR/IGF1R deletion plays a significant role in normal podocyte function sufficient to cause proteinuria and glomerulosclerosis then the effect of reduced IR and IGF1R protein expression on podocyte function would have been expected to produce a phenotype before 6 months. A more likely scenario to explain the overall result is that deleting the IR and IGF1R genes at about embryonic day12 impacted podocyte development to a variable extent such that some mice developed fewer podocytes per glomerulus than other mice. As mice grow and their glomeruli and glomerular capillary area increases, those mice with fewer podocytes would not be able to completely cover the filtration surface with foot processes and would develop proteinuria and glomerulosclerosis. If reduced podocyte number per glomerulus is the proximate cause of the observed proteinuria, then modulation of the body and kidney growth rate by calorie restriction to slow growth (lower circulating IGF-1 levels) would be expected to be protective, while a high protein high calorie diet (higher circulating IGF-1 levels) or uni-nephrectomy to increase kidney growth rate would be expected to enhance proteinuria and glomerulosclerosis.
Thank you for these comments. In response to them:
(1) WT1 as a marker of podocyte number. We agree may not be the most accurate way of precisely measuring podocyte number but is widely accepted in the field (PMID:33655004 / PMID:38542564) and we think convincingly shows fewer podocytes at 6-months.
(2) Podocyte size and density was not measured. This was not the focus of the paper and the histology obviously showed a significant phenotype in several mice (Figs 1D-F). Of note we did objectively assess a glomeruloscleorosis index (Fig 1D). We took the approach to understand mechanism through non-biased proteomics and phospho-proteomics of conditionally immortalised podocytes in which we had convincingly knocked down the insulin and IGF1 receptors (Figure 2)
(3) You did not study the mice earlier to ascertain the developmental phenotype. We concede we did not do this but there was no significant proteinuria detected early in the mice so elected not to increase mouse numbers by studying them then (which we consider good practice for reduction, replacement and refinement). We suspect there would have been subtle changes in those mice that had significantly reduced simultaneous IR and IGF1R knockdown. It was precisely because of this that we generated a conditionally immortalised podocyte cell line with robust simultaneous knock-down of both receptors.
(4) You did not show significant insulin and IGF1 receptor knockdown in the conditionally immortalised cell line (reviewer states it was 50%). We clearly knocked both receptors down (insulin and IGF1R) in the podocyte line by >80% which was highly statistically significant (p<0.00001). Figure 2A. We agree this was crucial (and we made the cell line because of the variability in the mouse model).
The model as used may be more representative of a variable degree of podocyte depletion than an effect of impaired IR/IGF1R signaling. Therefore, although the phenotype may be ultimately attributable to the IR/IGF1R gene deletions the proteinuria and glomerulosclerotic phenotype itself was probably a consequence of defective podocyte development. Examining podocyte number, size, density and glomerular volume at earlier time points (4 weeks) would help to answer this question. Therefore, a more appropriate title would be "The insulin/IGF axis is critically important (for) normal podocyte development and deployment". In this context the effect of the knockdowns on splicing would make more sense.
Please see our response (above). We think our final conclusion that in the podocyte the insulin/IGF axis is important for spliceosome activity and control is valid. This is due to our findings (both total and phospho proteomics results) and considering recent other papers showing this axis can rapidly phosphorylate a variety of spliceosome proteins in different cell types (PMID:39939313 / PMID:32888406). All discussed in detail in the manuscript).
Cell culture studies. A cell line was generated using a temperature sensitive SV40 system that has been previously reported from this laboratory. A detailed analysis is provided to show that double knockout cells exhibited abnormal spliceosome activity. This forms the basis for the conclusion that "The insulin/IGF axis is critically important (for) controlling gene transcription in the podocyte". There are several concerns that weaken this conclusion.
(1) In the double knockdown cell culture system about 30% of cells were "lost" by 3 days and about 70% of cells were "lost" by 5days. The studies were done at the 3 day time point. It is not clear whether "lost" cells were in the process of dying, stress-induced detachment, or just growing more slowly than control due to reduced IR and IGF-1R signaling. These processes could have impacted splicing in a non-specific way independent of IR/IGF1R signaling itself.
(2) Can a single cell line derived from the double floxed mice be relied on to provide an unbiased picture of the effect of deleting IR and IGF-1R? Presumably, the transfection and selection process will select for cells that survive thereby including unknown biases, possibly related to spliceosome function. Is a single cell line adequate? These investigators have extensive experience with this type of analysis, but this question is not addressed in the discussion.
(3) To determine whether the effect is specific to reduced IR/IGFR signaling the deletion of IR and IGF-1R could be corrected by transfecting full length IR and IGF-1R cDNAs into the cells to restore normal IR/IGF1R signaling. If transfected cells with intact IR and IGF-1R expression and activity returns spliceosome activity to normal this would be evidence that receptors themselves play some role in spliceosome activity, as opposed to the downstream effect on growth limitation/stress on the cells.
(4) Other ways of testing whether the splicing effect is specifically due to reduced IR/IGF-1R signaling would be to (a) block IR and IGF1R receptors using available inhibitors, (b) remove or reduce insulin, IGF-1 and IGF-2 levels in the culture medium, (c) use low glucose and amino acid culture medium to slow growth rate independent of receptor function, (d) or block intra-cellular signaling via the IR and IGF-1R receptors through mTORC1 inhibition using rapamycin or other signaling targets.
(5) It would be useful to determine whether the cultured cells stressed in other ways (e.g. ischemia, toxins, etc.) also results in the same splicing abnormalities.
Point 1. 70% cell loss was observed at day 7 (not day 5). We found approximately 20% loss at day 3. We opted to go for this early date hypothesising the key detrimental processes would be clear then. This 3 day time point also ensures there has been enough time to allow for the expression of Cre recombinase, receptor gene excision and degradation of existing endogenous IR/IGF1R following lentiviral transduction. Interestingly we did not find a major “death or apoptosis” signal in our data then but agree it should be considered. We think this is a specific pathway as we have examined several other conditionally immortalised detrimental podocyte cell line previously using proteomics with a much more severe phenotype of cell death (E.g. podocyte GSK3 alpha/beta knockdown) and we detected NO spliceosome signal (PMID:30679422). Furthermore, there are now other podocyte proteomics “stress” studies that have been published in which there is proteinuria and significant cell loss / death that also do not show spliceosome dysfunction. These include studying the detailed proteosomal signature of podocytes stressed with Doxorubicin and Lipopolysaccharide endotoxin LPS in mice (PMID:32047005) and bradykinin stimulation of rat podocytes (PMID:32518694).
Point 2. Yes, we think it is valuable and reproducible. We generated a podocyte cell line from insulin receptor and IGF1 receptor homozygous floxed cells. Hence there is no selection bias in the cells when generating the line as both receptors are effectively intact. We then temporally “knocked down” the receptors with extrinsic lentiviral Cre.
Importantly we validated our cell line findings both back in the cells (with Western blotting) and in our transgenic receptor knockdown mice and found evidence of spliceosomal dysregulation (Figure 3E and 3F). Also as discussed above the spliceosome has been identified in other models in the insulin/IGF pathway.
Point 3. We don’t think the experiment of knocking down the receptors and then reconstituting them would prove this hypothesis. This is because if splicing abnormality was due to generalised cell dysfunction (which we do not think is the case in this situation) then putting the receptors back may simply restore cell health and the spliceosomal function (e.g. it does not prove it is via the receptors). Secondly, the process of transduction with multiple lentiviruses may be inherently stressful to the cell and there may be a high level of extrinsic receptor inserted which may also be confounding/detrimental. Finally, as discussed there are now several lines of evidence describing insulin / IGF signalling to spliceosomal proteins which we consider important (discussed in the paper in detail).
Point 4. We think modulating the receptors using the Cre-lox approach is the cleanest approach (with fewer off-target effects) to interrogate the insulin / IGF axis. It allows us to differentiate the cells by thermo-switching (which is crucial for this terminally differentiated cell) and then robustly knocking down both receptors simultaneously to investigate mechanism. We agree these supplementary approaches may give some extra information if their limitations (eg off target effects of inhibitors) are also taken into consideration.
Point 5. They do not. Please see response to point 1 above regarding GSK3, Doxorubicin, LPS and bradykinin challenge.
reply to u/LillieLogang at https://reddit.com/r/typewriters/comments/1tacody/light_touch_jcpenney/
Before you go too deeply here, is the ribbon made of cloth material (nylon, silk, or cotton) or is it a plastic film/carbon type? And if the latter, is it a proprietary cartridge or typewriter spools? Cartridges with carbon can be difficult if not impossible to find.
There's only one Penny's toy typewriter in the database currently which may give you some idea: https://typewriterdatabase.com/1970-jc-penney-j6772-toy-typewriter.11484.typewriter I don't see these pop up often, but there is a related one listed on SGW right now: https://shopgoodwill.com/item/263548866 which may have some unused "ribbon" you might cannibalize if necessary.
It looks like it may have been a rebranded version of some of the PETITE toy typewriters. https://typewriterdatabase.com/no_info.525.typewriter-serial-number-database
Many of these toys are difficult to adjust or fix (if at all), but they tend to be very simple in terms of the mechanics, so you might be able to puzzle out what isn't working and why by looking at it with the hood removed.
A mission should not shy away from a rigorous application of the principle of impartiality for fear of misinterpretation or retaliation.
Peacekeeping operations should be careful to compromise their image of impartiality, but should not be scared of execution and enforcing action. If peacekeeping operations remain focused on their primary goals, they should not have to shy away from taking action due to fear of compromising impartiality.
Universality of consent becomes even less probable in volatile settings
Aren't these settings where peacekeeping is needed most? --> demonstrates the inefficiency of peacekeeping operations.
does not necessarily imply or guarantee that there will also be consent at the local level,
Local citizens could be hostile against the peacekeeping troops if they don't want change or are biased.
a peacekeeping operation risks becoming a party to the conflict
Is this because peacekeeping operations would try to settle peace in an area that doesn't want peace to be settled, thus creating conflict between local parties and peacekeeping operations?
the consent of the main parties to the conflict
Multiple nations often act only in their personal self-interest. This would prevent multiple peacekeeping missions, causing violence or conflict to grow.
They were headed towards the Palm Beach road and kept moving steadily. About an hour later another party appeared and went the same way. Then another just before sundown. This time she asked where they were all going and at last one of the men answered her.
People left Everglades due to warning signs.
great deal of the old crowd were back. But there were lots of new ones too. Some of these men made passes at Janie, and women who didn’t know took out after Tea Cake. Didn’t take them long to be put right, however. Still and all, jealousies arose now and then on both sides. When Mrs. Turner’s brother came and she brought him over to be introduced, Tea Cake had a brainstorm. Before the week was over he had whipped Janie. Not because her behavior justified his jealousy, but it relieved that awful fear inside him.
Tea cake hits Janie cause he is jealous and wants control
great deal of the old crowd were back. But there were lots of new ones too. Some of these men made passes at Janie, and women who didn’t know took out after Tea Cake. Didn’t take them long to be put right, however. Still and all, jealousies arose now and then on both sides. When Mrs. Turner’s brother came and she brought him over to be introduced, Tea Cake had a brainstorm
Teak cake hit Janie to feel like he has control and was jealous
The season closed and people went away like they had come—in droves. Tea Cake and Janie decided to stay since they wanted to make another season on the muck. There was nothing to do, after they had gathered several bushels of dried beans to save over and sell to the planters in the fall. So Janie began to look around and see people and things she hadn’t noticed during the season.
People are starting to relax and enjoy life cause farming season done.
Janie learned what it felt like to be jealous. A little chunky girl took to picking a play out of Tea Cake in the fields and in the quarters
She is feeling jealous because tea cake is hanging with someone else and think they might be up to something.
To Janie’s strange eyes, everything in the Everglades was big and new. Big Lake Okechobee, big beans, big cane, big weeds, big everything. Weeds that did well to grow waist high up the state were eight and often ten feet tall down there. Ground so rich that everything went wild. Volunteer cane just taking the place. Dirt roads so rich and black that a half mile of it would have fertilized a Kansas wheat field. Wild cane on either side of the road hiding the rest of the world. People wild too.
Everything looks weird and diffrent because she’s never explored before
eLife Assessment
This study presents a valuable finding relating to how the state of arousal is represented within the superior colliculus (SC), a principal visuo-oculomotor structure. The main conclusion that the SC's neural representation of arousal is segregated from motor related output appears to have solid support by the data. The work will be of interest to sensory, motor and cognitive neuroscientists.
Reviewer #1 (Public review):
Summary:
Johnston and Smith used linear electrode arrays to record from small populations of neurons in the superior colliculus (SC) of monkeys performing a memory-guided saccade (MGS) task. Dimensionality reduction (PCA) was used to reveal low-dimensional subspaces of population activity reflecting the slow drift of neuronal signals during the delay period across a recording session (similar to what they reported for parts of cortex: Cowley et al., 2020). This SC drift was correlated with a similar slow-drift subspace recorded from the prefrontal cortex, and both slow-drift subspaces tended to be associated with changes in arousal (pupil size). These relationships were driven primarily by neurons in superficial layers of the SC, where saccade sensitivity/selectivity is typically reduced. Accordingly, delay-period modulations of both spiking activity and pupil size were independent of saccade-related activity, which was most prevalent in deeper layers of the SC. The authors suggest that these findings provide evidence of a separation of arousal- and motor-related signals. The analysis techniques expand upon the group's previous work and provides useful insight into the power of large-scale neural recordings paired with dimensionality reduction. This is particularly important with the advent of recording technologies which allow for the measurement of spiking activity across hundreds of neurons simultaneously. Together, these results provide a useful framework for comparing how different populations encode signals related to cognition, arousal, and motor output in potentially different subspaces.
Comments on revised manuscript:
The authors have done a very good job of responding to all of the reviewers' concerns.
Reviewer #2 (Public review):
Summary:
Neurons in motor-related areas have increasingly shown to carry also other, non-motoric signals. This creates a problem of avoidance of interference between the motor and non-motor-related signals. This is a significant problem that likely affects many brain areas. The specific example studied here is interference between saccade-related activity and slow-changing arousal signals in the superior colliculus. The authors identify neuronal activity related to saccades and arousal. Identifying saccade-related activity is straightforward, but arousal-related activity is harder to identify. The authors first identify a potential neuronal correlate of arousal using PCA to identifying a component in the population activity corresponding to slow drift over the recording session. Next, they link this component to arousal by showing that the component is present across different brain areas (SC and PFC), and that it is correlated with pupil size, an external marker of arousal. Having identified an arousal-related component in SC, the authors show next that SC neurons with strong motor-related activity are less strongly affected by this arousal component (both SC and PFC). Lastly, they show that SC population activity pattern related to saccades and pupil size form orthogonal subspaces in the SC population.
Strengths:
A great strength of this research is the clear description of the problem, its relationship with the performed analysis and the interpretation of the results. The paper is very well written and easy to follow.
An additional strength is the use of fairly sophisticated analysis using population activity.
Weaknesses:
(1) The greatest weakness in the present research is the fact that arousal is a functionally less important non-motoric variable. The authors themself introduce the problem with a discussion of attention, which is without any doubt the most important cognitive process that needs to be functionally isolated from oculomotor processes. Given this introduction, one cannot help but wonder, why the authors did not design an experiment, in which spatial attention and oculomotor control are differentiated. Absent such an experiment, the authors should spend more time on explaining the importance of arousal and how it could interfere with oculomotor behavior.
(2) In this context, it is particularly puzzling that one actually would expect effects of arousal on oculomotor behavior. Specifically, saccade reaction time, accuracy, and speed could be influenced by arousal. The authors should include an analysis of such effects. They should also discuss the absence or presence of such effects and how they affect their other results.
(3) The authors use the analysis shown in Figure 6D to argue that across recording sessions the activity components capturing variance in pupil size and saccade tuning are uncorrelated. however, the distribution (green) seems to be non-uniform with a peak at very low and very high correlation, specifically. The authors should test if such an interpretation is correct. If yes, where are the low and high correlations respectively? Are there potentially two functional areas in SC?
Comments on the first revision:
My main concern with the paper is really two-fold. First, I think it is only incremental and adds next to no useful information about the SC. That might not be a fair criticism and certainly is purely subjective, but it affects the standards that eLife has on significance thresholds for papers. As such, this is an issue the editors should talk about.
Second, my main concern with the substance of the paper is that the authors jump immediately into an analysis of the 'arousal-related' effects on SC activity. Before that, I would like to see some behavioral indicators of arousal, such as RT differences, pupil size (the talk about this), or accuracy. The authors first need to describe the objective behavioral indicators of the level of arousal. Using these indices, they need to establish that there are meaningful differences in the level of arousal across the recording session. Having done so, they can proceed to link changes in SC activity with levels of arousal.
Instead, in its current form, the authors find changes in SC activity and describe them immediately as 'arousal-related'. I hope it is clear why that is premature. The 'slow-drift' fluctuations are presumed to be related to arousal, but they could be meaningless random fluctuations, or related to some other cognitive process.
Other than this conceptual issue, I do not have major problems with the analysis per se.
Comments on the latest version:
They have constructively responded to my concerns. I think 'incomplete' should be replaced with 'solidly supported'.
Reviewer #3 (Public review):
Summary:
This study looked at slow changes in neuronal activity (on the order of minutes to hours) in the superior colliculus (SC) and prefrontal cortex (PFC) of two monkeys. They found that SC activity shows slow drift in neuronal activity like in the cortex. They then computed a motor index in SC neurons. By definition, this index is low if the neuron has stronger visual responses than motor response, and it is high if the neuron has weaker visual responses and stronger motor responses. The authors found that the slow drift in neuronal activity was more prevalent in the low motor index SC neurons and less prevalent in the high motor index neurons. In addition, the authors measured pupil diameter and found it to correlate with slow drifts in neuronal activity, but only in the neurons with lower motor index of the SC. They concluded that arousal signals affecting slow drifts in neuronal modulations are brain-wide. They also concluded that these signals are not present in the deepest SC layers, and they interpreted this to mean that this minimizes the impact of arousal on unwanted eye movements.
Strengths:
The paper is clear and well-written.
Showing slow drifts in the SC activity is important to demonstrate that cortical slow drifts could be brain-wide.
Weaknesses:
The authors find that the SC cells with the low motor index are modulated by pupil diameter. However, this could be independent of an "arousal signal". These cells have substantial visual sensitivity. If the pupil diameter changes, then their activity should be influenced since the monkey is watching a luminous display. So, in this regard, the fact that they do not see "an arousal signal" in the most motor neurons (through the pupil diameter analyses) is not evidence that the arousal signal is filtered out from the motor neurons. It could simply be that these neurons simply do not get affected by the pupil diameter because they do not have visual sensitivity.
Comments on revisions:
The authors have given due consideration to the possibility that SC signaling of arousal could be at least in part due to changes in pupil size related responses to ambient light. Discussion of this point in the most recent revision helps to mitigate this concern.
Author response:
The following is the authors’ response to the previous reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
Johnston and Smith used linear electrode arrays to record from small populations of neurons in the superior colliculus (SC) of monkeys performing a memory-guided saccade (MGS) task. Dimensionality reduction (PCA) was used to reveal low-dimensional subspaces of population activity reflecting the slow drift of neuronal signals during the delay period across a recording session (similar to what they reported for parts of cortex: Cowley et al., 2020). This SC drift was correlated with a similar slow-drift subspace recorded from the prefrontal cortex, and both slow-drift subspaces tended to be associated with changes in arousal (pupil size). These relationships were driven primarily by neurons in superficial layers of the SC, where saccade sensitivity/selectivity is typically reduced. Accordingly, delay-period modulations of both spiking activity and pupil size were independent of saccade-related activity, which was most prevalent in deeper layers of the SC. The authors suggest that these findings provide evidence of a separation of arousal- and motor-related signals. The analysis techniques expand upon the group's previous work and provides useful insight into the power of large-scale neural recordings paired with dimensionality reduction. This is particularly important with the advent of recording technologies which allow for the measurement of spiking activity across hundreds of neurons simultaneously. Together, these results provide a useful framework for comparing how different populations encode signals related to cognition, arousal, and motor output in potentially different subspaces.
Comments on revised manuscript:
The authors have done a very good job of responding to all of the reviewers' concerns.
No weaknesses to address.
Reviewer #2 (Public review):
Weaknesses:
(1) The greatest weakness in the present research is the fact that arousal is a functionally less important non-motoric variable. The authors themself introduce the problem with a discussion of attention, which is without any doubt the most important cognitive process that needs to be functionally isolated from oculomotor processes. Given this introduction, one cannot help but wonder, why the authors did not design an experiment, in which spatial attention and oculomotor control are differentiated. Absent such an experiment, the authors should spend more time on explaining the importance of arousal and how it could interfere with oculomotor behavior.
(2) In this context, it is particularly puzzling that one actually would expect effects of arousal on oculomotor behavior. Specifically, saccade reaction time, accuracy, and speed could be influenced by arousal. The authors should include an analysis of such effects. They should also discuss the absence or presence of such effects and how they affect their other results.
(3) The authors use the analysis shown in Figure 6D to argue that across recording sessions the activity components capturing variance in pupil size and saccade tuning are uncorrelated. however, the distribution (green) seems to be non-uniform with a peak at very low and very high correlation specifically. The authors should test if such an interpretation is correct. If yes, where are the low and high correlations respectively? Are there potentially two functional areas in SC?
Comments on revised manuscript:
I remain somewhat concerned that the authors jump immediately into an analysis of the 'arousal-related' effects on SC activity. Before that, I would like to see a more detailed discussion justifying the use pupil size alone (i.e., w/o other indicators such as RT) as indicative of fluctuations in general arousal that are causal to concomitant changes in SC activity. Instead, in its current form, the authors find changes in SC activity and describe them immediately as 'arousal-related'.
Other than this conceptual issue, I do not have major problems with the analysis per se.
We agree with the reviewer that we may have advanced into discussing arousal-related effects in the previous version of the manuscript without providing a thorough explanation for why we think the slow drift axis is associated with changes in the monkey’s arousal levels. Arousal has been linked to the size of the pupil as well as movements of the eyes in numerous previous studies. We have made the following changes in the revised manuscript to address the reviewer’s concern:
(1) When first describing how the spiking responses of SC neurons fluctuate over the course of a recording session (Lines 130-132), we have used the phrase "slow fluctuations in the spiking responses" rather than "arousal-related fluctuations in the spiking responses". Then, when describing these effects in more detail (Lines 136-147), we have explained why we think these fluctuations may be related to arousal. The following text has been added in the revised manuscript for clarification:
“We found that this low-dimensional pattern of activity in the SC was also correlated with pupil size in the present study and with simultaneously recorded data in the prefrontal cortex (PFC), pointing to a link between this brain-wide fluctuation and changes in the monkeys’ arousal levels while performing the task.” (Lines 136-147)
(2) We have changed the subheading in Line 183 of the revised manuscript from "Arousal-related fluctuations are present in the SC and correlated with pupil size and fluctuations in PFC activity" to "Slow fluctuations in SC spiking activity are correlated with pupil size and PFC activity". Given that we have not yet explained the results linking these fluctuations to arousal at this stage of the manuscript, we believe that this revised title is more accurate and avoids jumping too quickly to arousal-related fluctuations without first explaining the link between SC slow drift, pupil size and PFC activity.
(3) We have provided additional justification for using pupil size and PFC activity to assess whether SC slow drift is associated with changes in the monkeys’ arousal levels. In a previous study, we computed an identical slow drift axis for spiking responses in visual cortex (V4) and PFC, and investigated how these low-dimensional neural activity patterns, which were themselves strongly correlated, were associated with various eye-related metrics (e.g., pupil size, microsaccade rate, reaction time, saccade velocity). Results showed that pupil size was the strongest predictor of slow drift in V4 and PFC. Given that the eye metrics were also strongly correlated with each other, we believe that the observed relationship between SC slow drift, pupil size and PFC activity provides sufficient evidence to suggest that the fluctuations observed in the SC are arousal-related. The following text has been added to the Results section of the revised manuscript:
“Moreover, previous work in our laboratory computed a similar slow-drift axis using spiking activity in visual cortex (V4) and PFC, and investigated the relationship between these low-dimensional neural activity patterns and different eye-related metrics (e.g., pupil size, microsaccade rate, reaction time, saccade velocity). In addition to observing a strong correlation between V4 and PFC slow drift, we found that, relative to the other eye-related metrics, pupil size was the strongest predictor of these fluctuations (Johnston et al., 2022a). Thus, to further confirm the link between the SC slow drift axis and changes in the monkeys’ arousal levels while they performed the MGS task, we next sought to explore if projections onto the SC slow drift axis were associated with pupil size.” (Lines 236-344)
Reviewer #3 (Public review):
Summary:
This study looked at slow changes in neuronal activity (on the order of minutes to hours) in the superior colliculus (SC) and prefrontal cortex (PFC) of two monkeys. They found that SC activity shows slow drift in neuronal activity like in the cortex. They then computed a motor index in SC neurons. By definition, this index is low if the neuron has stronger visual responses than motor response, and it is low if the neuron has weaker visual responses and stronger motor responses. The authors found that the slow drift in neuronal activity was more prevalent in the low motor index SC neurons and less prevalent in the high motor index neurons. In addition, the authors measured pupil diameter and found it to correlate with slow drifts in neuronal activity, but only in the neurons with lower motor index of the SC. They concluded that arousal signals affecting slow drifts in neuronal modulations are brain-wide. They also concluded that these signals are not present in the deepest SC layers, and they interpreted this to mean that this minimizes the impact of arousal on unwanted eye movements.
Strengths:
The paper is clear and well-written.
Showing slow drifts in the SC activity is important to demonstrate that cortical slow drifts could be brain-wide.
Weaknesses:
The authors find that the SC cells with the low motor index are modulated by pupil diameter. However, this could be completely independent of an "arousal signal". These cells have substantial visual sensitivity. If the pupil diameter changes, then their activity should be influenced since the monkey is watching a luminous display. So, in this regard, the fact that they do not see "an arousal signal" in the most motor neurons (through the pupil diameter analyses) is not evidence that the arousal signal is filtered out from the motor neurons. It could simply be that these neurons simply do not get affected by the pupil diameter because they do not have visual sensitivity. So, even with the pupil data, it is still a bit tricky for me to interpret that arousal signals are excluded from the "output layers" of the SC.
Of course, the general conclusion is that the motor neurons will not have the arousal signal. It's just the interpretation that is different in the sense that the lack of the arousal signal is due to a lack of visual sensitivity in the motor neurons.
I think that it is important to consider the alternative caveat of different amounts of light entering the system. Changes in light level caused by pupil diameter variations can be quite large. Please also note that I do not mean the luminance transient associated with the target onset. I mean the luminance of the gray display. it is a source of light. if the pupil diameter changes, then the amount of light entering to the visually sensitive neurons also changes.
Comments on revised manuscript:
The authors have addressed my first primary comment. For the light comment, I'm still not sure they addressed it. At the very least, they should explicitly state the possibility that the amount of light entering from the gray background can matter greatly, and it is not resolved by simply changing the analysis interval to the baseline pre-stimulus epoch. I provide more clear details below:
In line 194 of the redlined version of the article (in the Introduction), the citation to Baumann et al., PNAS, 2023 is missing near the citation of Jagadisan and Gandhi, 2022. Besides replicating Jagadisan and Gandhi, 2022, this other study actually showed that the subspaces for the visual and motor epochs are orthogonal to each other
We thank the reviewer for this comment and apologize that the citation to Baumann et al., PNAS, 2023 was missing in the previous version of the manuscript. In addition to including this citation in the revised version, we have provided a much more comprehensive description of all three cited studies and clarified that, in addition to replicating the results of Jagadisan and Gandhi, Baumann et al., PNAS, 2023 showed that the subspaces for the visual and motor epochs are orthogonal to each other. The following lines have been added to the Introduction of the revised manuscript:
“A similar separation has been observed for visual and motor responses in the SC (Jagadisan and Gandhi, 2022; Ayar et al., 2023; Baumann et al., 2023). For example, Jagadisan and Gandhi (2022) used linear microelectrode arrays to investigate why early eye movements are not triggered when neuronal responses to a visual target, presented before a delayed saccade to that target, cross a threshold. They found that population activity in the SC was less stable during the visual epoch of a delayed saccade task, relative to the saccade epoch. Moreover, saccades could be evoked more easily by patterned microstimulation when the temporal structure of the microstimulation was stable across electrodes, providing a potential explanation for how downstream regions differentiate between visual and motor responses. Similar results were reported by Baumann et al. (2023) who found that the strength of SC motor responses during a saccade to a visual image depends on the features of that image (e.g., contrast, orientation). When dimensionality reduction was applied to the spiking responses of neuronal populations in the SC, the population trajectory during the initial visual response to the image was orthogonal to that during the motor response. These findings replicate the separation in temporal population structure reported by Jagadisan and Gandhi (2022) and support the results of Ayar et al. (2023). They found that, although not completely orthogonal, population activity in the SC is distinct for visual and motor responses during the same oculomotor task and across different tasks, which could further facilitate the decoding of signals related to sensation, action and context by downstream regions.” (Lines 110-127)
Line 683 (and around) of the redlined version of the article (in the Results): I'm very confused here. When I mentioned visual modulation by changed pupil diameter, I did not mean the transient changes associated with the brief onset of the cue in the memory-guided saccade task. I meant the gray background of the display itself. This is a strong source of light. If the pupil diameter changes across trials, then the amount of light entering the eye also changes from the gray background. Thus, visually-responsive neurons will have different amount of light driving them. This will also happen in the baseline interval containing only a fixation spot. The arguments made by the authors here do not address this point at all. So, please modify the text to explicitly state the possibility that the global luminance of the display (as filtered by the pupil diameter) alters the amount of light driving the visually-responsive neurons and could contribute to the higher effects seen in the more visual neurons.
We apologize that our analysis did not fully address the reviewer’s concern that the presence of fluctuations in visual neurons and their absence in motor neurons may have arisen indirectly due to changes in the amount of light entering the eye caused by changes in pupil size. As per the reviewer’s suggestion, we have now raised the possibility that visual neurons in the SC may have firing rates that are monotonically related to slow trends in overall luminance induced by pupil size changes, whereas motor neurons do not. Although we believe this to be an unlikely explanation, the paragraph from lines 374-398 has been modified to better describe this possibility, including the following text:
“Given that slow drift is found in traditionally defined visual areas (e.g., area V4) and in regions that show mixed selectivity for multiple task variables (e.g., PFC) (Cowley et al., 2020), it seems unlikely that slow drift is caused by luminance fluctuations alone and more likely that it reflects global changes in arousal. At the same time, these arousal-related fluctuations covary with changes in pupil size (Johnston et al., 2022a), which could modulate the amount of light entering the eye from the display. This might affect visual neurons but not motor neurons due to their lack of visual sensitivity. Because SC neurons exist on a continuum, with visual responses decreasing and motor responses increasing from the intermediate to deep layers (Massot et al., 2019; Heusser et al., 2022) and no clear categorical boundary for motor-only neurons, any readout strategy would still need to avoid corruption of the motor output by slow drift, even if it were caused by changes in the amount of light entering the eye.” (Lines 387-398)
The figures (everywhere, including the responses to reviewers) are very low resolution and all equations in methods are missing.
We thank the reviewer for bringing this to our attention. We believe this issue may have arisen during conversion of the manuscript file for review, as the figures were of sufficient quality and the equations visible in the version that appeared online (https://doi.org/10.7554/eLife.99278.2). In any case, we will ensure that high-resolution figures are submitted with the revised manuscript and apologize that they were low resolution in the previous version.
I'm very confused by Fig. 2 - supplement 2. Panel B shows a firing rate burst aligned to *microsaccade* onset. Does that mean you were in the foveal SC? i.e. how can neurons have a motor burst to the target of the memory-guided saccade and also for microsaccades? And which microsaccade directions caused such a burst? And what does it mean to compute the motor index and spike count for microsaccades in panel C? if you were in the proper SC location for the saccade target, then shouldn't you *not* get any microsaccade-related burst at all? This is very confusing to me and needs to be clarified
We agree that clarification is needed here and thank the reviewer for their comment. The eccentricity of the targets was set to match the endpoints of the evoked saccades, which for some sessions were relatively close to the fovea. The mean eccentricity of the targets across sessions was 4.52° (SD = 2.89°). These values are now reported in the Methods section of the revised manuscript (Line 637). For the neuron shown in Figure 2–figure supplement 2, the eccentricity of the targets was 3°. Previous research has shown that some SC neurons respond during microsaccades as well as slightly larger saccades (see Hafed & Krauzlis, 2012, J. Neurophysiol., Fig. 4B). This likely explains why the neuron shown in Figure 2–figure supplement 2, which had a receptive field at ~3° based on saccades evoked by microstimulation, also responded during microsaccades. We apologize that this was not explained in the previous version and agree that it could have been confusing for the reader. To address this, the legend for this supplementary figure has been edited in the revised version and now reads:
“(B) PSTH for an SC neuron that responded around the time of a microsaccade. Firing rates were computed in 1ms bins, averaged across trials and smoothed using a Gaussian function (σ = 5ms). Note that the targets were set to 3º in this session based on saccades evoked by microstimulation (see Methods). Previous research has shown that some SC neurons respond during microsaccades as well as to slightly larger saccades (Hafed and Krauzlis, 2012). This likely explains why this SC neuron, which had a RF at ~3º based on saccades evoked by microstimulation, also responded around the time of a microsaccade.” (Lines 1026-1031)
eLife Assessment
This study explores how exogenous attention operates at the finest spatial scale of vision, within the foveola - a topic that has not been previously explored but is of interest to visual neuroscientists. The question is important for understanding how attention shapes perception, and how it differs between the periphery and the central regions of highest visual acuity. The evidence indicating that attention near the fovea preferentially enhances low spatial frequencies is compelling, as shown by carefully designed experiments with state-of-the-art eye tracking to monitor attended locations just a few tens of minutes of arc away from the fixation target.
Reviewer #1 (Public review):
[Editors' note: this version has been assessed by the Reviewing Editor without further input from the original reviewers. The authors have addressed the weaknesses noted above, which were raised in the previous round of review.]
Summary:
The manuscript investigates how exogenous attention modulates spatial frequency sensitivity within the foveola. Using high-precision eye-tracking and gaze-contingent stimulus control, the authors show that exogenous attention selectively improves contrast sensitivity for low- to mid-range spatial frequencies (4-8 cycles/degree), but not for higher frequencies (12-20 CPD). In contrast, improvements in asymptotic performance at the highest contrast levels occur across all spatial frequencies. These results suggest that, even within the foveola, exogenous attention operates through a mechanism similar to that observed in peripheral vision, preferentially enhancing lower spatial frequencies.
Strengths:
The study shows strong methodological rigor. Eye position was carefully controlled, and the stimulus generation and calibration were highly precise. The authors also situate their work well within the existing literature, providing a clear rationale for examining the fine-grained effects of exogenous attention within the foveola. The combination of high spatial precision, gaze-contingent presentation, and detailed modeling makes this a valuable technical contribution.
Weaknesses:
The manipulation of attention raises some interpretive concerns. Clarifying this issue, together with additional detail about statistics, participant profiles, other methodological elements, and further discussion in relation to oculomotor control in general, could broaden the impact of the findings.
Reviewer #2 (Public review):
Summary:
This study aims to test whether foveal and non-foveal vision share the same mechanisms for endogenous attention. Specifically, they aim to test whether they can replicate at the foveola previous results regarding the effects of exogenous attention for different spatial frequencies.
Strengths:
Monitoring the exact place where the gaze is located at this scale requires very precise eye-tracking methods and accurate and stable calibration. This study uses state-of-the-art methods to achieve this goal. The study builds on many other studies that show similarities between foveal vision and non-foveal vision, adding more data supporting this parallel.
Weaknesses:
The study lacks a discussion of the strength of the effect and how it relates to previous studies done away from the fovea. It would be valuable to know if not just the range of frequencies, but the size of the effect is also comparable.
Reviewer #3 (Public review):
Summary:
This paper explores how spatial attention affects foveal information processing across different spatial frequencies. The results indicate that exogenously directed attention enhances contrast sensitivity for low- to mid-range spatial frequencies (4-8 CPD), with no significant benefits for higher spatial frequencies (12-20 CPD). However, asymptotic performance increased as a result of spatial attention independently of spatial frequency.
Strengths:
The strengths of this article lie in its methodological approach, which combines a psychophysical experiment with precise control over the information presented in the foveola.
Weaknesses:
The authors acknowledge that they used the standard approach of analyzing observer-averaged data, but recognize that this method has limitations: it ignores the uncertainty associated with parameter estimates and the relationships between different parameters of the psychometric model. This may affect the interpretation of attentional effects. In the future, mixed-effects models at the trial level could overcome these limitations.
Author response:
The following is the authors’ response to the original reviews.
eLife Assessment
This study explores how exogenous attention operates at the finest spatial scale of vision, within the foveola - a topic that has not been previously explored. The question is important for understanding how attention shapes perception, and how it differs between the periphery and the central regions of highest visual acuity. The evidence is compelling, as shown by carefully designed experiments with state-of-the-art eye tracking to monitor attended locations just a few tens of minutes of arc away from the fixation target, but additional clarification regarding analyses and implications for vision and oculomotor control would broaden the impact of the study.
We thank the editors and reviewers for their thorough evaluation of our work. We have carefully revised the manuscript and substantially reworked the Discussion to address all of the points raised, eliminate redundancies, streamline the text, and clarify the implications of our findings for vision and oculomotor control. We have also expanded the documentation of our power analyses and conducted the additional analyses requested by the reviewers. Our point-by-point responses are provided.
Public Reviews:
Reviewer #1 (Public review):
Summary:
The manuscript investigates how exogenous attention modulates spatial frequency sensitivity within the foveola. Using high-precision eye-tracking and gaze-contingent stimulus control, the authors show that exogenous attention selectively improves contrast sensitivity for low- to midrange spatial frequencies (4-8 cycles/degree), but not for higher frequencies (12-20 CPD). In contrast, improvements in asymptotic performance at the highest contrast levels occur across all spatial frequencies. These results suggest that, even within the foveola, exogenous attention operates through a mechanism similar to that observed in peripheral vision, preferentially enhancing lower spatial frequencies.
Strengths:
The study shows strong methodological rigor. Eye position was carefully controlled, and the stimulus generation and calibration were highly precise. The authors also situate their work well within the existing literature, providing a clear rationale for examining the fine-grained effects of exogenous attention within the foveola. The combination of high spatial precision, gazecontingent presentation, and detailed modeling makes this a valuable technical contribution.
Weaknesses:
The manipulation of attention raises some interpretive concerns. Clarifying this issue, together with additional detail about statistics, participant profiles, other methodological elements, and further discussion in relation to oculomotor control in general, could broaden the impact of the findings.
We thank the reviewer for the helpful comments. In the Discussion, we have now considered additional factors that could have contributed to the observed attentional effects. First, the exogenous cue might have functioned as a temporal warning signal. However, the interval between cue and stimulus onset was fixed across trials, meaning that the cue did not provide temporal information beyond what participants could already anticipate. Furthermore, participants completed a large number of trials (≥ 4000), making it highly likely that the temporal relationship between trial onset and target onset was overlearned. These considerations indicate that the observed benefit in the valid condition was predominantly attributable to spatial reorienting induced by the cue, rather than to differences in the temporal predictability of the target across conditions.
Another possibility is that the 100% validity of the exogenous cue could potentially have promoted endogenous attentional engagement. Yet, several characteristics of our task strongly limited the extent to which such endogenous engagement could meaningfully influence performance. Endogenous attentional benefits typically emerge only after ~150-200 ms (Posner & Petersen, 1990; Carrasco, 2011), whereas our cue-target SOA was 100 ms, and the target remained visible for only 50 ms. Under these temporal constraints, any voluntary, slow endogenous enhancement would primarily occur after the stimulus offset. Thus, although endogenous maintenance is theoretically possible given the cue’s validity, it is unlikely to have substantially contributed to the observed attentional benefits in our task.
Regarding the points on statistical reporting and participant details, we followed the reviewer’s suggestions by adding post hoc power analyses and providing more comprehensive reporting of the linear model outputs (see Appendices 1 and 2). We also expanded the description of the training procedures conducted with participants prior to formal data collection in the Methods section.
We appreciate the reviewer for raising the important question of how our findings may relate to oculomotor control. To address this, we analyzed trials excluded from the manuscript due to saccades. This analysis revealed that saccade latencies were shorter in the valid condition than in the neutral condition (see Figure 2 — Supplementary Figure 2). This earlier saccade onset may reflect exogenously triggered preparatory activity in the oculomotor system in response to the salient cue. Future studies are needed to examine whether this preparatory mechanism serves to efficiently guide microsaccades or saccades toward behaviorally relevant stimuli in everyday vision. We have incorporated this point into the Discussion, highlighting a potential mechanistic link between exogenous attention and oculomotor behavior.
Reviewer #2 (Public review):
Summary:
This study aims to test whether foveal and non-foveal vision share the same mechanisms for endogenous attention. Specifically, they aim to test whether they can replicate at the foveola previous results regarding the effects of exogenous attention for different spatial frequencies.
Strengths:
Monitoring the exact place where the gaze is located at this scale requires very precise eyetracking methods and accurate and stable calibration. This study uses state-of-the-art methods to achieve this goal. The study builds on many other studies that show similarities between foveal vision and non-foveal vision, adding more data supporting this parallel.
Weaknesses:
The study lacks a discussion of the strength of the effect and how it relates to previous studies done away from the fovea. It would be valuable to know if not just the range of frequencies, but the size of the effect is also comparable.
We thank the reviewer for raising these important issues. In response, we have expanded the Discussion to link our findings to prior work. First, we included a direct comparison of our effect sizes with those reported in previous studies. This analysis revealed that our effect sizes are highly comparable to those earlier studies (see Figure 3 — Supplementary Figure 4). Second, we contextualized our findings within the popular framework of normalization model of attention in the Discussion. We detected a mixture of contrast and response gain effects, consistent with predictions from the normalization framework given our experimental design. Finally, we extended the Discussion to consider potential underlying neural mechanisms. Specifically, we suggested that differences in attentional modulation, particularly the manifestation in response gain vs. contrast gain between the fovea and extrafovea, may reflect distinct characteristics of foveal neurons relative to those in extrafoveal regions.
Reviewer #3 (Public review):
Summary:
This paper explores how spatial attention affects foveal information processing across different spatial frequencies. The results indicate that exogenously directed attention enhances contrast sensitivity for low- to mid-range spatial frequencies (4-8 CPD), with no significant benefits for higher spatial frequencies (12-20 CPD). However, asymptotic performance increased as a result of spatial attention independently of spatial frequency.
Strengths:
The strengths of this article lie in its methodological approach, which combines a psychophysical experiment with precise control over the information presented in the foveola.
Weaknesses:
The authors acknowledge that they used the standard approach of analyzing observeraveraged data, but recognize that this method has limitations: it ignores the uncertainty associated with parameter estimates and the relationships between different parameters of the psychometric model. This may affect the interpretation of attentional effects. In the future, mixed-effects models at the trial level could overcome these limitations.
We thank the reviewer for this comment. Our Methods section continues to transparently discuss these limitations, as well as the fact that these limitations are shared with most published studies in psychophysics. Additionally, we now include measures of uncertainty for all key effects (see Appendices 1 and 2), and we have reported effect sizes throughout the Results section. Finally, we have added post hoc power analyses to the Methods. Following previous approaches to power calculation for related experiments, we found that our study was sufficiently powered to detect the main effect of attention and had moderate power to detect the interaction between attention and spatial frequency.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
(1) The manipulation of attention raises some interpretive concerns. Since only valid and neutral cue conditions were included, the results might reflect differences in temporal predictability rather than true spatial reorienting of attention. In other words, the valid cue could act mainly as a temporal warning signal that reduces uncertainty about stimulus onset. Without invalid trials or a non-predictive control cue, it remains difficult to separate spatial and temporal contributions to exogenous attention.
We thank the reviewer for raising this point. In this regard, we would like to clarify that there was no temporal uncertainty in stimulus onset: across all conditions and trial types, the stimulus was presented at the same time relative to the start of the trial, i.e., 600 ms after the start. Yet, we acknowledge that the shorter temporal proximity between the cue and stimulus in valid trials could serve as an additional temporal warning signal, potentially conferring an advantage relative to the neutral condition. While we cannot completely rule out a contribution of such temporal cueing within the constraints of the current experimental design, we believe its impact was limited. Specifically, the fixed cue-stimulus interval reduced the cue’s ability to convey additional temporal information. Furthermore, observers completed a large number of trials (≥4000), and the temporal contingency between trial onset and target onset was likely overlearned. Taken together, these considerations indicate that the observed benefit in the valid condition was predominantly attributable to spatial reorienting induced by the cue, rather than to differences in the temporal predictability of the target across conditions. We now mention this in the revised Discussion (lines 309-318).
We recognized that the original Figure 2 illustrating the experimental paradigm may have caused confusion regarding the timing structure of the task. We have therefore updated the figure to more explicitly illustrate the trial timeline in both conditions.
(2) The reported effects seem small, and no power analysis is provided. With only seven participants, the study may not have enough statistical power to confirm that the observed differences are reliable or generalizable. Although the technical precision in gaze and stimulus control is impressive, it cannot offset the limitations of a small sample. The authors should include effect size estimates, confidence intervals, and ideally a post-hoc power analysis.
The statistical results are reported only as χ² values from model comparisons, which do not show the direction or size of the effects. For clarity and transparency, these tests should be accompanied by fixed-effect estimates with their standard errors and confidence intervals, so readers can better assess both the reliability and perceptual relevance of the findings.
The reviewer raised several important points regarding the study's statistical rigor.
In the revised manuscript, we now report effect size estimates (Cohen’s d) in the Results section and Appendices. Effect sizes were in the medium-to-large range, including the effect of attention on contrast sensitivity at 4 and 8 CPD, and the difference in attentional benefit on contrast sensitivity between 4 and 12 CPD and between 8 and 12 CPD. We have also included the full model outputs, including standard errors and confidence intervals, in the Appendices.
The sample size for the current study was determined based on the magnitude of the attentional effects observed in our previous work (Guzhang et al., 2021). The experimental design and dependent measures were highly similar across the two studies, and the prior study revealed a robust effect, which accounted for a substantial proportion of within-observer variance in a tightly controlled repeated-measures design.
We have revised the manuscript, adding bootstrap-based power estimates, following the procedure described by Jigo and Carrasco (2020), using data from Guzhang et al. (2021). Assuming the effect size in our current study would be comparable to the prior one, 2 to 12 observers were randomly sampled with replacement, and a one-way repeated-measures ANOVA with attention as the main factor was used. This procedure was repeated 10,000 times, and power was estimated as the proportion of iterations yielding a significant main effect for each sample size. The results of this analysis indicate that a sample size of five observers would have been sufficient to achieve approximately 80% power to detect the main effect of attention in the prior study. Based on these estimates, the sample size used in the current study (seven observers) is adequately powered.
We also conducted a post hoc power analysis to evaluate the power of our design to detect the main effects and their interaction. It was performed using the R package simr, which estimates statistical power for mixed-effects models through model-based simulation. Specifically, simr generated datasets based on the fixed- and random-effect structure of the fitted model, preserving the observed effect sizes and variance components. For each simulated dataset, the model was refit, and the effect of interest was tested. By repeating this procedure 501 times across different sample sizes, power was estimated as the proportion of simulations in which the effect was statistically significant. Based on these post hoc simulations, we estimated that our study had high power (>95%) to detect the main effects and moderate power (>65%) to detect the interaction. Although the estimated power for the interaction was lower than for the main effects, the observed effect size was substantial (as indexed by Cohen’s d), indicating that the interaction was not trivially small.
We now describe these analyses in lines 501-532 in the Methods section.
(3) The task seems quite demanding, requiring fine spatial discrimination, very small stimuli, and head stabilization with a bite bar. It is not clear whether participants were naïve or experienced observers. If they had prior psychophysical training, practice effects could have influenced the results, particularly given the lack of invalid trials. The manuscript would benefit from clarifying participants' experience level and describing any training or familiarization procedures.
We appreciate the reviewer’s concern regarding potential training effects. All observers had prior experience with similar tasks, but were naïve to the scope of this study. Each participant underwent an initial familiarization phase of approximately 50 trials with the experimental setup of this study. They then completed an additional ~50 trials to estimate their individual contrast thresholds per spatial frequency level before we proceeded with data collection at the five predefined contrast levels.
Based on our experience, we have found that, for experiments similar to the one described here, observers quickly adapt to the setup and are generally able to maintain reliable fixation and stable performance, even during the initial training phase. In addition, each participant completed approximately 400 trials before the data collection started. Even observers who began the session with no prior experience would have become practiced with the setup by the time the actual data-collection phase started, during which ~4000 trials were collected per observer. Therefore, whether an observer participated in previous experiments is unlikely to meaningfully affect the results, as the large number of trials ensures comparable levels of task familiarity across individuals.
Crucially, valid and neutral trials were interleaved throughout the session. Any general learning or practice would therefore influence both conditions equally. Despite this, we still observed clear performance improvements in the valid condition relative to the neutral condition, indicating that the observed benefits cannot be attributed solely to practice and reflect an attentional enhancement. We have added elaboration on the training procedures in Methods (lines 411-429).
Finally, we recognize that the lack of invalid trials may raise concerns given our 100% spatially predictive cue, as noted in Reviewer 3’s first comment. We refer the reader to our response to that point for a more detailed discussion of cue validity and the distinction between exogenous and endogenous influences in our paradigm.
(4) The study would benefit from a clearer connection between the behavioral results and possible underlying neural mechanisms. How might the observed changes in contrast sensitivity relate to known physiological processes at the retinal, thalamic, or cortical level? The discussion could be strengthened by framing the findings within established models of attentional modulation or by referring to known effects of attention in the early visual cortex.
This is an important point, and we agree that framing the findings within established models of attentional modulation can strengthen the discussion. We believe that the normalization model of attention (Reynolds and Heeger, 2009; Herrmann et al., 2010) offers a useful framework for interpreting our behavioral findings, especially the attention-related changes in contrast sensitivity and asymptotic performance observed at the foveal scale. We have now added a more detailed discussion linking our results to this model and considering, explicitly as speculation, how known physiological processes at different stages may contribute to the observed effects in Discussion (lines 264-307).
(5) The ecological relevance of the results is not fully developed. The authors propose that the observed effects may resemble natural attentional shifts triggered by salient events, yet the brief, highly localized flashes used here are somewhat artificial. A more likely interpretation is that these mechanisms relate to oculomotor control within the fovea, perhaps reflecting preparatory activity for microsaccades or fine fixation adjustments. Considering this view could broaden the impact of the findings and link them to current discussions on the relationship between attention and oculomotor control.
We thank the reviewer for raising this important point regarding the ecological relevance of our findings, which we did not sufficiently address in the original manuscript. Although we briefly motivated scenarios that engage exogenous attention at high spatial resolution, such as detecting road signs or traffic lights at a distance while driving, we did not fully elaborate on how such attentional processes may link to downstream visual and oculomotor functions.
In our experiment, observers maintained fixation and avoided saccades throughout the trial. Nevertheless, in a subset of trials (on average 17% ± 3%), observers made saccades after stimuli disappeared and prior to providing a response. Typically, these movements were microsaccades with amplitudes smaller than 0.5°, directed toward the target location, in both valid and neutral trials. These saccades were discarded prior to the analyses performed in the manuscript. Inspired by the reviewer’s feedback, we decided to examine the saccade latency in these trials relative to the onset of the response cue to assess whether exogenous cueing influenced oculomotor timing. Notably, we observed an earlier onset of microsaccades in valid compared to neutral trials (71 ms ± 50 ms faster, P < 0.01). We have now added this observation as Figure 2 — Supplementary Figure 2 in the manuscript. Because the presence of an exogenous pre-cue was the only difference between the two trial types, the earlier microsaccade onset likely reflects exogenously triggered preparatory activity in the oculomotor system in response to the salient pre-cue. Such fine-grained attention may prime potential eye movements toward behaviorally relevant stimuli for further examination. This interpretation is consistent with the reviewer’s suggestion and supports a mechanistic link between exogenous attention and oculomotor behavior, extending the ecological relevance of our findings. This point has been added to the Discussion on lines 329 to 340.
We also conducted analysis to examine ocular drift behavior following the response cue. Although trials included in the manuscript analyses were constrained such that fixation during target presentation remained within a small window (10’ radius) around the fixation marker, we did not assess whether gaze subsequently drifted closer to the target location after the response cue. One possibility is that exogenous attention might bias ocular drift, shifting the preferred locus of fixation closer to the target. To address this, we computed the average Euclidean distance between gaze position and the target location following response cue onset for valid and neutral trials. However, we found no significant difference in gaze-target distance between valid and neutral trials (p = 0.57).
Although the spatial cueing approach has long been used to probe exogenous attention in a controlled manner in psychophysical experiments, we fully recognize the importance of understanding attention under more naturalistic viewing conditions that allow observers to freely move their eyes. Developing paradigms that incorporate more naturalistic, salient stimuli would be an important direction for future work, enabling investigation of exogenous attention in ecologically valid settings and its influence on sequential actions and processes, including oculomotor behavior.
(6) There is no statement about the availability of the data and code used for the experiment.
We have now added the data and code for the analysis pipeline to the Open Science Framework (OSF).
Reviewer #2 (Recommendations for the authors):
(1) The study could discuss the strength of the effect and how it relates to previous studies.
We thank the reviewer for raising this point. To facilitate direct comparison with the study by Jigo and Carrasco (2020), we computed attentional benefit as the ratio of contrast sensitivity between the valid and neutral conditions (now shown in Figure 3 — Supplementary Figure 4). In their data, the attentional benefit at 0° eccentricity peaked just below 4 CPD, with a ratio of approximately 1.2, corresponding to a ~20% increase in contrast sensitivity. This magnitude closely matches the benefit we observed for fine-grained attentional shifts within the foveola at spatial frequencies between 4 and 8 CPD (17% ± 12% and 16% ± 14% for 4 and 8 CPD, respectively). We have added this comparison to the Discussion (lines 246-262).
In addition, we acknowledge that prior studies have reported heterogeneous attentional effects, including pure contrast gain, pure response gain, or a mixture of the two. We now explicitly reference these findings in the Discussion and use the normalization model of attention (Reynolds and Heeger, 2009; Herrmann et al., 2010) to account for how differences in stimulus configuration, attention field size, and eccentricity may account for discrepancies between our findings and prior studies examining attention in the extrafovea or when broadly distributed across the fovea (lines 264-307).
(2) Minor details:
(a) The abstract mentions gaze-contingent-display, but if I understand correctly, the stimulus was not presented in a gaze-contingent manner.
That’s correct. Although stimuli were not presented gaze-contingently, we used a gaze-contingent calibration procedure (see Methods, lines 386-389) to achieve higher precision in localizing the line of sight. This increased accuracy was essential for selecting trials in which stimuli remained at the intended eccentricity relative to the preferred locus of fixation. To avoid potential confusion, however, we have removed this detail from the abstract.
(b) Line 361: What is the manual calibration the authors are referring to? It does not appear to be described.
The text has been updated to explain more explicitly what auto and manual calibrations are.
(c) Line 402: There may be a typo towards the end of the line "t0" should be "to"?
Text has been updated. Thank you.
(d) Line 405. What are the units of 30?
It’s in arcminutes. Text has been updated.
Reviewer #3 (Recommendations for the authors):
I found this paper very interesting, with a solid methodological approach and excellent data analyses. The authors present a well-designed psychophysical study that contributes valuable insights into the mechanisms of attention in the foveola. The methodology is rigorous, and the analyses are thoughtfully conducted and clearly presented.
That said, I would like to offer a few comments and suggestions for clarification and further consideration:
(1) Exogenous attention:
If a 100% spatially predictive cue is compared to a neutral cue, the observed attentional effect should not be described as (purely) exogenous, since the cue fully predicts where the post-cue will request a response. This situation represents a case in which attention is exogenously driven but endogenously maintained (see e.g., Chica et al., 2013, Behavioural Brain Research). I recommend clarifying this distinction in the manuscript (and title) to avoid conceptual ambiguity.
We thank the reviewer for raising this important conceptual point. We agree that because the pre-cue was 100% spatially predictive, the resulting attentional allocation cannot be considered purely exogenous. Although the abrupt, salient onset of the cue obligatorily triggers an exogenous shift of attention, its validity could also promote endogenous maintenance of attention at the cued location. Yet, several characteristics of our task strongly limit the extent to which such endogenous engagement could meaningfully influence performance. Endogenous attentional benefits typically emerge only after ~150-200 ms (Posner & Petersen, 1990; Carrasco, 2011), whereas our cue-target SOA was 100 ms, and the target remained visible for only 50 ms. Under these temporal constraints, any voluntary, slow endogenous enhancement would primarily occur after the stimulus offset. Thus, although endogenous maintenance is theoretically possible given the cue’s validity, it is unlikely to have substantially contributed to perceptual encoding in our task.
We also considered the possibility that our response cue (a retro-cue indicating the target location) might recruit endogenous attention to the internal perceptual representation. Importantly, however, this retro-cue was equally informative in valid and neutral conditions. Any enhancement driven by the retro-cue should therefore benefit both trial types to the same extent. The fact that we still observe a robust advantage in valid trials supports the conclusion that the performance improvements predominantly reflect fast, spatially specific exogenous facilitation rather than slower endogenous processes.
We have revised the manuscript to clarify that although the cue obligatorily triggers an exogenous attentional shift, its 100% validity could allow for endogenous attention maintenance as shown by Chica et al. (2013). We also added an explanation detailing why such endogenous contributions are unlikely to drive our main results, given the rapid cue-target timing in our task in Discussion (lines 319-327). Finally, to further prevent ambiguity, we updated the manuscript title to refer to “exogenously triggered attention,” rather than simply “exogenous attention.”
(2) Interpretation of statistical effects:
The statement "Therefore, asymptotic performance showed only independent, additive effects of frequency and attention, without a systematic influence of spatial frequency on the attentional benefit" seems not to be supported by the data, as the main effect of frequency was not significant.
We thank the reviewer for this helpful observation. We agree that the original phrasing did not accurately reflect the results, as the main effect of spatial frequency was not significant (p = .0545). We have revised the sentence to “Therefore, asymptotic performance reflected an effect of attention alone, with no detectable contribution of spatial frequency or of the interaction between spatial frequency and attention” to avoid implying such an effect (lines 210-211).
If data from two participants were missing in one condition, the authors should consider replacing this data with new participants.
We agree with the reviewer that having two observers with missing data in one condition is not ideal. However, the 20 cpd condition was deliberately positioned near the resolution limit at the tested eccentricity and was therefore extremely demanding. Observers also had to monitor two stimulus locations simultaneously, further increasing task difficulty. This condition was challenging for all observers and, despite testing up to the highest contrast, two of seven observers were unable to perform above chance, indicating that for a non-trivial fraction of observers, this condition was effectively unmeasurable with our paradigm. As noted in the manuscript, the 20 cpd condition also has a statistical limitation: thresholds clustered near the upper bound (approaching 100% contrast), compressing the dynamic range and markedly reducing variance relative to lower spatial frequencies, which violates the homoscedasticity assumption of linear models. For these reasons, we did not pursue additional data collection in this condition. Nevertheless, we report the data that were successfully obtained, as they remain informative about performance near the resolution limit.
We finally note that even when setting aside the 20 CPD condition, our data support this conclusion: comparisons between 4 and 12 CPD, as well as between 8 and 12 CPD, revealed large differences in the magnitude of the attentional benefit (d = 0.65, 95% CI [0.11, 1.18] and d = 0.62, 95% CI [0.08, 1.14], respectively). To further quantify these effects, we have added Cohen’s d to report the effect sizes for these spatial-frequency comparisons across texts in Results as well as in tables in Appendices.
(3) Sample size:
As this is a psychophysical experiment with many trials and few participants, I am curious about how the authors determined the appropriate sample size and the number of trials required to detect the expected effects. Given that many effects were found to be significant, it seems that statistical power was adequate; however, it would be helpful if the authors could explain how this issue was addressed a priori during experimental planning.
We appreciate that the reviewer raised this point. Please see the reply to the second point from Reviewer 1, who raised a related question about statistical power.
(4) Figure 2 clarification:
In Figure 2B, I do not fully understand the "Valid" and "Neutral" representation. Both conditions include a post-cue indicating the right position; however, in the neutral condition, there is a central fixation square, whereas in the valid condition, there is not. Please clarify this aspect of the figure. I think I understood the paradigm, but this part of the figure is misleading.
Precue only exists in valid condition. But there is a mistake where fixation marker is missing in valid condition in panel B.
We thank the reviewer for pointing this out. We have updated Figure 2 to explicitly show the sequence of valid vs. neutral trials. The fixation mark remained on the screen throughout the trial in both the valid and neutral conditions. After a 500 ms fixation period, an exogenous cue was presented for 30 ms in valid trials, followed by a 70 ms interval before stimulus onset. In neutral trials, no cue was presented, and the screen remained blank for 100 ms before the stimuli appeared. In conditions, a response cue would appear 50 ms after stimulus offset.
Es ist das erste MACH-Alliance-zertifizierte Composable ERP für E-Commerce, basierend auf einer Microservices-, API-first-, Cloud-nativen, Headless-Architektur. Wenn diese Begriffe abstrakt klingen, spricht die Kundenliste konkreter: Nintendo of Europe, EDEKA, ALDI Onlineshop, Mondelez und tesa setzen alle auf Actindo.
Add information about sales channels integration with amazon, ebay, Otto and own stores using shopify, salesforce or shopware. Use the word "Vertriebskanal" for "Sales channels".
Afterbuy bietet die breiteste native Integration mit deutschen Marktplätzen (Amazon, eBay, Otto, Kaufland, Hood.de, Willhaben, Kleinanzeigen, ManoMano) und ist damit die erste Wahl für Händler, deren Geschäft auf deutschen Marktplätzen aufgebaut ist.
Change the beginning to "Afterbuy bietet im Hinblick auf deutsche Marktplätze" to emphasise the view on the german market.
Kostenbewusste deutsche Onlinehändler, die einen einzelnen Shop plus ein bis zwei Marktplatzkanäle betreiben, mit On-Premise-Software vertraut sind und einen kostenlosen Einstieg gegenüber Abo-Verpflichtungen bevorzugen.
Change the information to past tense and add, that JTL prices are on the rise
Zusammen mit der Auszeichnung als Leader bei den OMR Reviews in der Kategorie Multichannel Retail für Q1 2026 zeigt das:
Correct the Award from "als Leader bei den OMR Reviews in der Kategorie Multichannel Retail für Q1 2026" to "zum besten Tool für Multichannel und Marketplace Tools bei den E-Commerce Germany Awards" and delete "das" before the colon
alles verbunden mit mehr als 150 Marktplätzen, Shopsystemen, Zahlungsanbietern und Versanddienstleistern.
Replace "Marktplätze, Shopsystem" with "Vertriebskanälen" and add "weltweit" at the end of the sentence
Schnellvergleich
Extend the table in thid chapter and add two more variables. Use information provided from this table: https://www.plentyone.com/de/jtl-alternative#vergleichstabelle
E-Commerce
I am not allowed to send you my email via Upwork. If you like to, we can also handle invoicing outside of Upwork. Upworks fees are killing me and there is also the issue with the invoices that to not abide german tax law (invoices are posted without sales tax).
I already tried to contact you via the contact form on your website and the e-mail provided.
Please let me know if you are interested in working outside of Upwork. I will of course send you a valid invoice for my work.
kontakt@jc-hohaus.de
eLife Assessment
This is a valuable report describing tracheal terminal cells (TTCs) in Drosophila as an immune privileged organ. The authors demonstrated that TTCs lack expression of the membrane-associated peptidoglycan recognition receptor PGRP-LC, which protects these cells from immune pathway activation and JNK-mediated cell death to maintain TTC homeostasis. While the genetic experiments using RNAi and overexpression are convincing and solid, the broader biological significance of this phenomenon requires further investigation. This work will be of interest to researchers in innate immunity across various model systems.
Reviewer #1 (Public review):
Summary:
In their manuscript entitled "Terminal tracheal cells of Drosophila are immune privileged to maintain their Foxo-dependent structural plasticity", Bossen and colleagues determine that the terminal cells of the tracheal system differ from other larval tracheal cells in that they do not typically show an Imd-dependent immune response to fungal and viral infections. Authors reach this conclusion based on the expression of a reporter line, Drs-GFP. The authors speculate that this difference may reflect differential expression of an immune pathway component, as tracheal terminal cells (ttcs) do not respond to forced expression of PRGP-LS. The authors then go on to show that, unlike the other cells of the tracheal system, terminal cells do not express PGRP-LC as reported by a GAL4 enhancer trap. Forced expression of PGRP-LC in terminal cells resulted in reduced branching, cell damage and features of the cell death program. These effects could be suppressed by depletion of AP-1 or Foxo transcription factors. Authors show that Foxo plays a negative role in branching of ttcs, with ectopic branching occurring upon RNAi (or under hypoxic conditions). The authors speculate that immune privilege of the ttcs may have evolved to permit Foxo regulation of ttc branching.
Strengths:
The authors provide compelling genetic data that support their overall conclusions.
Weaknesses:
FC do not appear to express DRS reporter in Figure 1 or elsewhere, raising the question of whether fusion cells are also immune privileged.<br /> Fig 5, TRE_RFP expression, is convincing in wt ttc, but not in ttc o/x PGRP-LCx
Reviewer #2 (Public review):
Summary:
In this study, Bossen et al. looked at the immune status of the tracheal terminal cells (TTCs) in Drosophila larvae. The authors propose that these cells do show PGFP-LCx expression and, hence, lack immune function. Artificial overexpression of the PGRP-LCx in the TTCs causes these cells to undergo apoptosis.
Strengths:
Only a few groups have tried to look at the immune status of the trachea, though we know that AMPs are expressed there after infection. This exciting study attempts to understand the differences in the tracheal cells that do not produce AMPs upon infection.
Weaknesses:
The reason why the TTCs have some immune privilege still needs to be completely clear. Whether the phenotype is cell autonomous or contributes to the cellular immune system is not evaluated. As we know, crystal cells also maintain oxygen levels in larvae; whether in the absence of a terminal trachea, the crystal cells have any role is not explored.
My particular comments on the figures are as follows:
(1) In Figure 2, the PGRP-LCx signal should be quantified as done for Drosomycin GFP, as shown in Figure 1.<br /> - The authors have now done this.
(2) In Fig 2F and G are the larvae infected? If not, what happens to PGRP-LCx expression post Ecc15 infection?<br /> - The authors have answered this question, saying infection has no effect on TTCs' Dr-GFP expression.
(3) Is the effect of overexpression of LCx exaggerated post-infection? In particular, when it comes to the escape phenotype.<br /> - This was not done; the infection experiment was done with PGRP-LE overexpression.
(4) Does overexpression of anti-apoptotic genes in TTC and PGRP-LCx rescue the TTC branching?<br /> - This was not done.
(5) Have the authors tried to rescue the larvae with shallow food?<br /> - This was not done.
(6) Is there any effect on the circulating hemocytes or lymph gland in the PGFRP-LCx overexpressing animals?<br /> - This was not done.
Reviewer #3 (Public review):
Summary:
The authors report that tracheal terminal cells (TTCs) in Drosophila do not activate innate immunity following bacterial infection, and attribute this to the absence of PGRP-LCx expression in these cells. Forced activation of the Imd pathway in TTCs leads to JNK-mediated cell death and reduced tracheal branching. The authors propose that this immune-privileged status preserves Foxo-dependent structural plasticity, which is essential for TTCs to respond to changing environmental conditions such as hypoxia.
Strengths:
The revised manuscript represents a meaningful improvement over the initial submission. The addition of multiple antimicrobial peptide reporters substantially strengthens the key observation that TTCs do not mount a humoral immune response upon infection, moving beyond reliance on the Drs-GFP reporter alone. The mechanistic dissection of the cell death pathway - demonstrating roles for JNK, AP-1, and Foxo downstream of ectopic PGRP-LCx activation - is well-executed and provides solid mechanistic insight. The inclusion of a second, independent UAS-PGRP-LCx line with a milder phenotype adds useful calibration. The hypoxia sensitivity assays provide physiological context, and the discussion of the gradient hypothesis, while based on qualitative observation, is logically reasoned and addresses a legitimate alternative interpretation.
Weaknesses:
The primary remaining concern is that the absence of PGRP-LCx expression in TTCs is supported by a single GAL4 enhancer trap line, without independent validation by complementary methods such as in situ hybridization, antibody staining, or reanalysis of publicly available single-cell transcriptomic data. The authors acknowledge this limitation transparently. While the convergent evidence from infection experiments - in which neither the Drs-GFP reporter nor the PGRP-LCx-Gal4 line shows TTC activation - lends indirect support, orthogonal confirmation would more definitively establish this mechanistic claim.
Additionally, the finding that Dcp-1 cleavage occurs in non-TTC tracheal cells as well suggests that Imd-mediated apoptotic signaling is not uniquely restricted to TTCs, and the Discussion could more explicitly address what distinguishes the TTC response in terms of degree or cellular context.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
Summary:
In their manuscript entitled "Terminal tracheal cells of Drosophila are immune privileged to maintain their Foxo-dependent structural plasticity", Bossen and colleagues determine that the terminal cells of the tracheal system differ from other larval tracheal cells in that they do not typically show an Imd-dependent immune response to fungal and viral infections. The authors reach this conclusion based on the expression of a reporter line, Drs-GFP. The authors speculate that this difference may reflect differential expression of an immune pathway component, as tracheal terminal cells (TTCs) do not respond to forced expression of PRGP-LS. The authors then go on to show that, unlike the other cells of the tracheal system, terminal cells do not express PGRP-LC as reported by a GAL4 enhancer trap. Forced expression of PGRP-LC in terminal cells resulted in reduced branching, cell damage, and features of the cell death program. These effects could be suppressed by the depletion of AP-1 or Foxo transcription factors. The authors show that Foxo plays a negative role in the branching of TTCs, with ectopic branching occurring upon RNAi (or under hypoxic conditions). The authors speculate that the immune privilege of the TTCs may have evolved to permit Foxo regulation of TTC branching.
Strengths:
The authors provide compelling genetic data.
Weaknesses:
(1) The authors state that after infection 34% of larvae were not GFP+ as defined by the detection of Drs-GFP in dorsal branches. The authors should clarify if these larvae are completely without response to infection, with no Drs-GFP in dorsal trunks and or other tracheal branches. If these larvae are entirely unresponsive, could authors indicate why this might be? Also, at this point in the manuscript, the authors are somewhat misleading regarding TTC expression of Drs-GFP - they should state at this point that there are some TTCs that do express Drs-GFP, and also should address their prior study of Drs-GFP induction which does not claim exclusion of TTC Drs-GFP expression.
GFP– indicates the absence of detectable fluorescence in regions proximal to the TTCs (dorsal branch and fusion cells). Our analysis specifically focused on these regions and did not assess fluorescence in other parts of the tracheal system. Therefore, the reported 34% of larvae classified as GFP– does not imply a complete absence of response in these animals; rather, no fluorescence was detected within our defined region of interest. To clarify how fluorescence in TTCs was quantified, we have added a schematic (new Fig. 1F). In addition, new Fig. S1 illustrates that AMP reporter activation frequently occurs in other tissues.
Our observations are consistent with earlier reports. In the original description of the AMP reporter lines, Tzou et al. (2000; https://doi.org/10.1016/S1074-7613(00)00072-8) reported that “only a fraction of the flies or larvae exhibited fluorescence in surface epithelia, and the proportion of GFP-expressing animals was variable from one culture vial to the next. In addition, fluorescence was rarely distributed throughout the whole tissue and was limited to restricted areas of the epithelium,” suggesting that AMP reporter activation can occur locally rather than uniformly across tissues.
In a previous study (https://doi.org/10.1186/1471-2164-9-446), we reported that airway epithelial cells, including the finest tracheal endings on target organs, can activate drosomycin transcription following infection. However, that study focused specifically on infected larvae. Importantly, it did not quantify the frequency of reporter activation or analyze TTC-specific phenotypes. As such, those statements should not be interpreted as implying uniform or ubiquitous reporter activation across all tracheal cells.
(2) The authors describe the terminal cell phenotype as "shrunken" but this implies loss of size or pruning, however, it is not clear whether the defects could equally be due to lack of growth or slower growth.
We omitted the term “shrunken” in the present manuscript to avoid potential misinterpretation.
(3) Figure 1 suggests that GFP+ dorsal branches are not uniform in their expression of Drs-GFP, it seems more patchy. The authors should define the fraction of dorsal branch cells that are Drs-GFP positive. Also, are fusion cells Drs-GFP positive?
We included a schematic illustrating our quantification approach (new Fig. 1F). We also revised the wording to clarify that GFP<sup>+</sup> animals include fluorescence not only in the dorsal branch (DB) but also in fusion cells (FCs), i.e., structures located between the dorsal trunks and the terminal tracheal cells (TTCs). Any structure in proximity to the TTCs that shows GFP expression was scored as GFP<sup>+</sup>. In most cases, GFP expression was observed in the dorsal fusion cells.
(4) Drs-GFP expression is largely absent from terminal cells; however, a still significant # of terminal cells show expression (8%). Authors argue that PRGP-LC expression is absent based on a GAL4 transgenic line. If this line reflects endogenous PRGP-LC expression, should there not be 8% positive TTCs? Or is the 8% Drs-GFP expression independent of the IMD receptor?
We detected PGRP-LE expression in approximately 3% of epithelial tracheal cells that expressed Drs after infection (Fig. 3F,G). This observation suggests that Drs activation can occur through a mechanism independent of PGRP-LCx. We have incorporated this finding into both the Results and Discussion sections.
(5) Figure 2: the authors state that TTCs are negative even with induced PRGP-LE expression - should there not be at least 8% that are positive?
We included infection of the PGRP-LE overexpression and could see Drs-GFP expression in 3 % of the cases, which we did not see without infection.
(6) The authors compare PRGP-LC expression to induction of cell death by expression of reaper and hid. Reaper and Hid had stronger effects and eliminated TTCs. See cleavage of caspase Dpc-1 in PRGP-LC expressing cells. Is caspase cleavage always diagnostic of apoptosis or could the weaker than rpr/hid phenotype imply a different function?
We have included the potential non-apoptotic functions of Dcp-1 in the Discussion. The weaker phenotype observed could therefore be explained by a non-apoptotic role of Dcp-1.
(7) Drs-GFP expression is said to be "completely" absent from tracheal terminal cells when the entire tracheal system is expressing PGRP-LE.
We have revised the wording accordingly.
(8) Figure 5, TRE_RFP expression, is not convincing that it is higher or in terminal cells. https://doi.org/10.7554/eLife.102369.1.sa2
We have revised the wording in line 230.
Reviewer #2 (Public review):
Summary:
In this study, Bossen et al. looked at the immune status of the tracheal terminal cells (TTCs) in Drosophila larvae. The authors propose that these cells do show PGFP-LCx expression and, hence, lack immune function. Artificial overexpression of the PGRP-LCx in the TTCs causes these cells to undergo apoptosis.
Strengths:
Only a few groups have tried to look at the immune status of the trachea, though we know that AMPs are expressed there after infection. This exciting study attempts to understand the differences in the tracheal cells that do not produce AMPs upon infection.
Weaknesses:
The reason why the TTCs have some immune privilege still needs to be completely clear. Whether the phenotype is cell autonomous or contributes to the cellular immune system is not evaluated. As we know, crystal cells also maintain oxygen levels in larvae; whether in the absence of terminal trachea, the crystal cells have any role is not explored. https://doi.org/10.7554/eLife.102369.1.sa1
In addition to the Drs-GFP reporter line, we performed new infection experiments using additional antimicrobial peptide reporters to further support our observations. While these experiments confirm the humoral immune response, they do not address the mechanisms underlying the apparent immune privilege. Our analysis therefore focuses specifically on the humoral immune response and does not allow conclusions regarding potential contributions of the cellular immune system, including crystal cells, to maintaining oxygen levels in animals with impaired TTCs. Notably, complete loss of TTCs is lethal, as demonstrated by TTC ablation using hid;rpr expression (Fig. 4F).
Reviewer #3 (Public review):
Summary:
The authors report that tracheal terminal cells (TTCs) in Drosophila do not activate innate immunity following bacterial infection. They attribute this to the lack of expression of PGRP-LCx in these cells. Forced activation of the Imd pathway in TTCs leads to cell death and a reduction in tracheal branching. The authors propose a mechanism for cell death induction via pathways involving JNK, AP-1, and foxo. They suggest that the suppression of innate immunity in TTCs may serve to maintain their plasticity, preparing them for responses to hypoxic conditions.
Strengths:
(1) The study addresses the understudied area of immune privilege in innate immunity, providing a potentially important example in Drosophila TTCs.
(2) The molecular characterization of the cell death pathway induced by forced Imd activation is well-executed and provides solid mechanistic insights.
(3) The authors draw interesting parallels between Drosophila TTCs and mammalian endothelial cells, suggesting broader implications for their findings.
Weaknesses:
(1) The core premise of the study - that TTCs do not activate innate immunity following bacterial infection - relies heavily on a single readout (Drs reporter). Additional markers of immune activation would strengthen this crucial claim.
We included new experiments using additional antimicrobial peptide reporter genes that show results similar to those obtained with the Drs-GFP reporter (new Fig. 1).
(2) The evidence for the lack of PGRP-LCx expression in TTCs is based on a single GAL4 reporter line. Given the importance of this observation to the authors' model, validation using alternative methods would be beneficial.
Although we were not able to include alternative methods to further confirm our hypothesis, we performed additional infection experiments. Upon bacterial infection, we observed a strong increase in GFP fluorescence throughout the animal and in many other tissues, while still detecting no response in the TTCs. These results further support our hypothesis.
(3) The phenotypes observed upon forced activation of the Imd pathway in TTCs, while intriguing, may be influenced by non-physiological levels of pathway activation. The authors should address this potential caveat and consider examining the effects of more moderate pathway activation. https://doi.org/10.7554/eLife.102369.1.sa0
We used two independent UAS-PGRP-LCx lines located on different chromosomes. One line (III) produced a stronger phenotype than the other (II). We clarified this point in the Results section (Fig. 4C,D) and added supplementary data (new Fig. S2) showing that both lines produce comparable phenotypes when expressed using an alternative tracheal driver. The epithelial thickening observed follows the same pattern as the phenotype detected in TTCs, indicating that even moderate pathway activation leads to similar effects. However, we acknowledge that this represents ectopic pathway activation and therefore likely reflects a non-physiological level of signaling.
Recommendations for the authors:
Reviewer #2 (Recommendations for the authors):
My particular comments on the figures are as follows:
(1) In Figure 2, the PGRP-LCx signal should be quantified as done for Drosomycin GFP, as shown in Figure 1.
We agree and have added a quantification.
(2) In Figure 2F and G are the larvae infected? If not, what happens to PGRP-LCx expression post Ecc15 infection?
We also included infected larvae to test whether infection induces GFP expression in TTCs. However, GFP expression was never observed in TTCs, although overall fluorescence increased in other tissues.
(3) Is the effect of overexpression of LCx exaggerated post-infection? In particular when it comes to the escape phenotype.
We induced mild Imd pathway activation by expressing PGRP-LE using a tracheal driver active in all tracheal cells, including TTCs, for 24 hours. In addition, these larvae were infected and their sensitivity to hypoxia was assessed. Animals expressing PGRP-LE in the trachea showed increased sensitivity to hypoxia, which was further enhanced following infection.
(4) Does overexpression of anti-apoptotic genes in TTC and PGRP-LCx rescue the TTC branching?
This point was not addressed.
(5) Have the authors tried to rescue the larvae with shallow food?
This point was not addressed.
(6) Is there any effect on the circulating hemocytes or lymph glands in the PGFRP-LCx overexpressing animals?
This point was not addressed.
Reviewer #3 (Recommendations for the authors):
The authors present an intriguing model of immune privilege in Drosophila tracheal terminal cells (TTCs). This model is built upon three key pillars: (1) the absence of innate immune activation in TTCs, (2) the lack of PGRP-LCx expression in TTCs, and (3) the induction of cell death when innate immunity is activated in TTCs. However, the experimental evidence supporting each of these critical points requires substantial strengthening. The reviewer recommends the following improvements and additional experiments to address these core issues:
(1) Innate immune activation in TTCs:
Evaluate the expression of additional antimicrobial peptide reporters to provide a more comprehensive assessment of innate immune activation in TTCs.
In addition to the Drs-GFP reporter line, we performed new infection experiments using other antimicrobial peptide reporters to confirm our results.
(2) PGRP-LCx expression in TTCs:
Validate the PGRP-LCx-GAL4 line used in the study to ensure it accurately reflects endogenous PGRP-LCx expression.
Employ complementary techniques such as in situ hybridization and antibody staining to corroborate the absence of PGRP-LCx in TTCs.
We also included infection experiments using PGRP-LCx-Gal4 larvae. Infection did not trigger GFP expression in TTCs. However, the overall PGRP-LCx expression pattern observed in other larval tissues supports that the results reflect endogenous PGRP-LCx expression.
(3) Cell death induction upon immune activation in TTCs:
Address the possibility that the observed cell death is an artifact of strong, forced Imd pathway activation. To do that,
perform control experiments activating the Imd pathway in non-TTC tracheal cells to determine if cell death is specific to TTCs.
Use broader tracheal drivers (e.g., ppk4-GAL4 or btl-GAL4) to activate the Imd pathway and verify if cell death is indeed restricted to TTCs.
We included results from PGRP-LCx overexpression using the tracheal driver ppk4-Gal4 and stained for the apoptosis marker Dcp-1 (new Fig. S3). We observed increased Dcp-1 signal in dorsal trunk cells, indicating that PGRP-LCx-mediated Dcp-1 cleavage is not restricted to TTCs.
Ideally, generate a transgenic line expressing physiological levels of PGRP-LCx in TTCs and demonstrate that bacterial infection induces cell death specifically in TTCs through the proposed pathway. The reviewer acknowledges the complexity of this experiment but believe it would significantly strengthen the authors' conclusions.
We did not generate a new transgenic line but instead used an alternative UAS-PGRP-LCx line (II), which exhibits a milder phenotype. This has now been clarified more prominently in the Results section (Fig. 4C,D). Additionally, we performed further experiments showing an epithelial thickening phenotype whose severity depends on the UAS-PGRP-LCx line used (new Fig. S2).
In addition to the above major points
(4) Quantitative data presentation:
Provide quantitative analyses for the results presented in Figures 2 and 3J-K to allow for a more rigorous evaluation of the data.
We included a quantitative analysis of the results shown in Fig. 2 (now presented in new Fig. 3). In addition, we added quantification of fluorescence in the TTCs of infected larvae.
(5) Alternative hypothesis:
Consider and address an alternative explanation for the lack of innate immune activation in TTCs: the potential gradient of bacterial ligands from proximal trachea to distal TTCs. If this hypothesis is correct, one might expect to see a gradient of Drs expression correlating with the distance from the proximal trachea. Addressing this possibility would strengthen the authors' proposed model.
We now included the following paragraph as part of the discussion section.
“An alternative explanation for the observed lack of an immune response in TTCs could be their maximal distance from the spiracles. In this scenario, a gradient of bacterial inducers along the tracheal system might be expected, resulting in a gradual decrease in immune activation from the spiracles toward the TTCs. However, this is not what we observed. In tracheae that displayed an immune response, the response was largely homogeneous along the entire length of the tracheal system, from the spiracles to the TTCs. Only at the transition to the TTCs did the immune response drop abruptly. This observation argues against the gradient hypothesis and suggests that TTCs are specifically excluded from the immune response.”
The length and physical impossibility of the poem's mapped itinerary reveal that Paris: A Poem is not a literal walk through the 1920s city, but a ghostlike traversal across time. A traversal where past, present, and future coalesce into a single day, and Paris lives on through the generations of artists, revolutionaries, and ordinary inhabitants who haunt its streets.
It's tiring on eyes to read an entire paragraph of boldface type. Can you switch this to regular text and make a transition to invite readers outside the map? Talk to us the way you did at the beginning, e.g.: If you feel a bit dizzy after that tour of Paris, you have begun to realize the length and physical impossibility... You might also productively include some of the calculations you mentioned in class, like how many miles were traversed in navigating the city in this way, and how long it might take to do so.
eLife Assessment
By screening an FDA-approved small-molecule library against a leucine-dependent M. tuberculosis strain, this study identifies semapimod as an inhibitor of Mtb growth that functions by impairing leucine import. The work is useful in linking leucine uptake to cell wall lipid biology in Mtb. However, the mechanistic understanding remains incomplete. Additional experimental evidence is required to clarify how PDIM contributes to or regulates leucine uptake.
Reviewer #3 (Public review):
Agarwal et al identified the small molecule semapimod from a chemical screen of repurposed drugs with specific antimycobacterial activity against a leucine-dependent strain of M. tuberculosis. To better understand the mechanism of action of this repurposed anti-inflammatory drug, the authors used RNA-seq to reveal a leucine-deficient transcriptomic signature from semapimod challenge. The authors then measured a decreased intracellular concentration of leucine after semapimod challenge, suggesting that semapimod disrupts leucine uptake as the primary mechanism of action. Unexpectedly however, resistant mutants raised against semapimod had a mutation in the polyketide synthase gene ppsB that resulted in loss of PDIM synthesis. The authors believe growth inhibition is a consequence of decreased accumulation of leucine as a result of an impaired cell wall and a disrupted, unknown leucine transporter. This study highlights the importance of branched-chain amino acids for M. tuberculosis survival and the chemical genetic interactions between semapimod and ppsB indicate that ppsB is a conditionally essential gene in a medium deplete of leucine.
The conclusions regarding the leucine and PDIM phenotypes are moderately supported by experimental data. The authors do not provide experimental evidence to support a specific link between leucine uptake and impaired PDIM production. Additional work is needed to support these claims and strengthen this mechanism of action.
A mechanistic gap still exists for the model of semapimod antitubercular activity. The basis for semapimod activity is that the leucine auxotroph strain cannot acquire leucine from its environment, and thus the bug ceases to grow. Under normal growth conditions, the leucine auxotroph strain produces PDIM and acquires exogenous leucine through some mechanism (either through a transporter or through PDIM). Semapimod binding to PpsB causes the cell to alter its PDIM profile (lacking experimental for this), and now with the altered PDIM profile the cell cannot acquire enough exogenous leucine to sustain growth (either because the altered PDIM profile interferes with the leucine transporter activity or through PDIM uptake). Acquiring a mutation in ppsB results in cells unable to produce PDIM (some evidence supporting this) but can now acquire enough exogenous leucine to sustain growth. I cannot find the connection between cells that have normal PDIM with normal leucine uptake and cells that are missing PDIM with normal leucine uptake.
(1) The manuscript would benefit from adding additional antibiotic controls to experiments. With the current experimental approaches, it is unclear if these signatures are the result of semapimod specifically or the effect of an antimicrobial agent. Adding additional strains to the 2D TLC experiments could provide more confidence in the absence or modifications of the PDIM band.
(2) The intriguing observation that wild-type H37Rv is resistant to semapimod but the leucine-auxotroph is sensitive should be further explored. If the authors are correct and semapimod does inhibit leucine uptake through a specific transporter or modified PDIM profiles, testing semapimod activity against the leucine-auxotroph in various concentrations of BCAAs could highlight the importance of intracellular leucine. Cells might recover growth in the presence of semapimod treatment if enough leucine is provided in the media and some fraction is able to enter the cell through the impaired PDIM barrier.
Reviewer #4 (Public review):
Summary:
In this study, the authors screened an FDA-approved repurposed library of small-molecule inhibitors against the auxotrophic strain Mtb mc2 6206 and found that semapimod exclusively inhibited its growth. Further studies showed that it inhibits L-leucine uptake by interacting with PpsB, although the exact mechanism remains unknown. Interestingly, semapimod showed antibacterial activity against H37Rv only in vivo, not in vitro, suggesting a dependence on host-derived exogenous leucine during intracellular growth. This work therefore suggests that uptake of host-derived leucine can be targeted as an effective strategy to reduce intracellular survival of Mtb.
Strengths:
The authors have used different approaches to understand the mechanism of L-leucine uptake in Mtb. To start, they conducted an in vitro screen using an FDA-approved library, followed by transcriptomic and metabolic analyses of different Mtb mutants. Through whole-genome sequencing, they identified mutations conferring resistance to semapimod to gain further mechanistic understanding. This led to the analysis of semapimod-PpsB interaction by BLI-Octet and analysis of cell-wall apolar lipid, which explained how PDIM loss resulted in sensitivity to vancomycin. Finally, infection experiments in mice surprisingly showed that semapimod was effective against intracellular Mtb in vivo but not in vitro.
Weakness:
The major weakness of this study is that it is unclear what role PpsB plays in L-leucine uptake. It is also not clear why intracellular Mtb relies on exogenous leucine rather than endogenous leucine. Does intracellular Mtb lose its ability to synthesize leucine, which is why semapimod is active in vivo but not in vitro? Or semapimod has any other effect on host immunity that has not been explored. I have a few minor comments, which are as follows:
(1) Authors state that "The colony forming unit (CFU) estimation further shows a bactericidal activity of this molecule which causes 88% reduction of bacterial viability on day 2 and >99% reduction after 5 days of incubation" (Fig. 1d). However, this is only true when compared to the untreated control. Compared to the Day 0 control, treated bacteria appear to have undergone little or no change, suggesting that the compound is bacteriostatic, not bactericidal. The drug concentration used for Fig 1d is not mentioned. For Fig. 1e, there is no day 0 control, and the comparison is with the untreated control at Day 6, which again does not suggest bactericidal action of Semapimod.
(2) The authors report that "Notably, no cytotoxic effect was observed at this concentration against THP1, thus ruling out the possibility of cell lysis by semapimod," but the data are not shown. Similarly, authors state that "As a control, interaction of semapimod was also analyzed with the purified Ppe60, which fails to exhibit any binding," but the data is not shown.
(3) Line 235: change "promote" to "promoter".
Reviewer #5 (Public review):
Summary:
The authors have extensively characterized the response of the leucine and pantothenate auxotroph Mtb strain H37Rv mc26 206 to an FDA-approved compound library and identified semapimod that is, at best, bacteriostatic in its action against the pathogen. The authors have used transcriptional profiling, metabolite quantification and a screening of genetically-resistant mutants to identify changes in leucine uptake under semapimod exposure. Based on these data, the authors attribute changes in antibiotic susceptibility to differences in environmental leucine availability and bacterial PDIM architecture. While the work presents an interesting avenue of investigation of metabolite uptake and utilization in a comparative fashion between fully virulent and auxotroph Mtb strains, it lacks clear and direct evidence to link the observations with a mechanistic explanation.
Strengths:
The authors used a well-designed screening strategy for FDA-approved compounds against a metabolically defined strain and follow up characterization of semapimod exposure through RNA-seq and pathway analysis, metabolomics and time-course analysis of drug effects. The data has been interestingly interpreted to identify a phenotypic connection between PDIM and altered drug susceptibility.
Weaknesses:
The major gap in the study is the speculative nature of the mechanism underpinning the connection between PDIM architecture and changes in leucine uptake under various bacterial growth conditions.
(1) Despite claims of identifying a "novel leucine uptake mechanism", the authors only provide endpoint metabolite measurements rather than kinetic leucine transport studies.
(2) A clear explanation for the differences in susceptibility between auxotroph and fully virulent Mtb strains through changes in "PDIM architecture" is not supported by any direct evidence such as structural analysis, lipidomics, or direct measurement of PDIM architectural changes.
(3) The figures 1D (lines 110-112, "kills bacteria") and 7c (lines 283-285) are used to infer a bactericidal role of semapimod, which maybe a mischaracterization of drug activity. The trend in CFUs in both cases seems of no bacterial growth rather than a CFU reduction- therefore interpreted as "bacteriostatic" at best. These observations would in fact align with the general antibiotic/stress response signature identified by RNA-seq, where leucine transport related genes only happen to be a small subset of many dysregulated genes. How do the authors disentangle these generic signatures from the leucine transport evidence, other than endpoint metabolite quantification?
(4) Furthermore, the studies with supplementation of leuCD (and not panCD) in rescuing from semapimod susceptibility are not supported by a clear mechanistic link. The complementation of leuCD does not completely rescue growth- does this indicate differences in uptake and metabolism? The authors should test this by monitroing the growth of the strains in minimal medium in presence and absence of exogenous leucine.
(5) It remains unclear if the authors attribute leucine uptake differences to a loss of PDIM or changes in PDIM amount and architecture. No direct evidence is provided for differences in PDIM production in the WT H37Rv strain and the auxotroph mc2 6206 strains used in this study. Mulholland et al (2024) report similar PDIM levels for WT and auxotrophic Mtb (mc2 6206) in their stocks passaged to maintain PDIM. This could change for stocks maintained differently. Since the presence of PDIM has classically been used to explain a penetration barrier for small molecules and the schematic provided by the authors at the end of the manuscript (figure 8c) suggest free leucine penetration in the absence of PDIM, how do the authors explain the increased leucine uptake and sensitivity of a PDIM positive auxotroph to semapimod through direct experimental evidence? Further on the point of PDIM production, the WT auxotroph strain seems to produce limited amounts of PDIM as evidenced by the TLC data in Figure 6b. To solidify this point, the authors should test other point mutants for PDIM production (not attenuated for growth) through TLC and quantify these differences. These data should be compared with PDIM production in the WT Mtb H37Rv strain (used by the authors) under in vitro growth conditions. A comparative lipidomics of cell envelope components might be insightful in explaining these differences. I believe answering this query is crucial and within the scope of the work whose central claim is the identification of a novel leucine uptake mechanism. It would be interesting, in fact, to identify a novel transporter associated with the PDIM layer on the cell envelope.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
Summary:
In this manuscript, the authors used a leucine/pantothenate auxotrophic strain of Mtb to screen a library of FDA-approved compounds for their antimycobacterial activity and found significant antibacterial activity of the inhibitor semapimod. In addition to alterations in pathways, including amino acid and lipid metabolism and transcriptional machinery, the authors demonstrate that semapimod treatment targets leucine uptake in Mtb. The work presents an interesting connection between nutrient uptake and cell wall composition in mycobacteria.
Strengths:
(1a) The link between the leucine uptake pathway and PDIM is interesting but has not been characterized mechanistically. The authors discuss that PDIM presents a barrier to the uptake of nutrients and shows binding of the drug with PpsB. However it is unclear why only the leucine uptake pathway was affected.
We observe interference of L-leucine, but not of pantothenate, uptake in mc2 6206 strain upon semapimod treatment. At present, we do not have any clue whether PDIM presents a barrier exclusively to the uptake of L-leucine. Further studies may shed a light on underlying mechanism(s) by which L-leucine uptake is modulated by this small molecule.
(1b) We still do not know what PpsB actually does for amino acid uptake - is it a transporter?
By BLI-Octet we do not find any interaction between L-leucine and PpsB. Therefore, we doubt that PpsB is a transporter of L-leucine.
(1c) Does semapimod binding affect its activity?
Our study suggests that semapimod treatment alters PDIM architecture which becomes restrictive to L-leucine. However, at present the exact mechanism is not clear. Further studies are required to thoroughly examine the effect of semapimod on Mtb PpsB activity and alterations in PDIM by mass spectrometry.
(1d) Does the auxotrophic Mtb have lower PDIM levels compared to wild-type Mtb?
As per the published report by Mulholland et al, and by vancomycin susceptibility phenotype in our study, both the strains appear to have comparable PDIM levels.
(2) The authors show an interesting result where they observed antibacterial activity of semapimod against H37Rv only in vivo and not in vitro. Why do the authors think this is the basis of this observation? It is possible semapimod has an immunomodulatory effect on the host since leucine is an essential amino acid in mice. The authors could check pro-inflammatory cytokine levels in infected mouse lungs with and without drug treatment.
Semapimod inhibits production of proinflammatory cytokines such as TNF-α, IL-1β, and IL-6, which would indeed help pathogen establish chronic infection. However, a significant reduction in bacterial loads in lungs and spleen upon semapimod treatment despite inhibition of proinflammatory cytokines clearly indicates bacterial dependence on host-derived exogenous leucine during intracellular growth.
(3) The authors show that the semapimod-resistant auxotroph lacks PDIM. The conclusions would be further strengthened by including validations using PDIM mutants, including del-ppsB Mtb and other genes of the PDIM locus, whether in vivo this mutant would be more susceptible (or resistant) to semapimod treatment.
PDIM is a virulence factor, and plays an important role in the intracellular survival of the TB pathogen. Mtb strains lacking PDIM are expected to show attenuated growth during infection, even without semapimod treatment. In such a case, it might be difficult to draw any conclusions about the effect of semapimod against PDIM(-) strains in vivo.
(4) Prolonged subculturing can introduce mutations in PDIM, which can be overcome by supplementing with propionate (Mullholland et al, Nat Microbiol, 2024). Did the authors also supplement their cultures with propionate? It would be interesting to see what mutations would result in Semr strains with propionate supplementation along with prolonged semapimod treatment.
Considering the fact that extensive subculturing may result in loss of PDIM, we avoided prolonged subculturing of bacteria. As presented in Fig. 6b, the WT bacteria retain PDIM. While performing the initial screening of drugs, we did not anticipate such phenotype, and hence bacteria were cultured in regular 7H9-OADS medium without propionate supplementation.
A comprehensive future study would help examining the effect of propionate on generation of semapimod resistant mutants in Mtb mc2 6206.
Weaknesses:
I have summarized the limitations above in my comments. Overall, it would be helpful to provide more mechanistic details to study the connection between leucine uptake and PDIM.
Reviewer #2 (Public review):
Summary
This important study uncovers a novel mechanism for L-leucine uptake by M. tuberculosis and shows that targeting this pathway with 'Semapimod' interferes with bacterial metabolism and virulence. These results identify the leucine uptake pathway as a potential target to design new anti-tubercular therapy.
Strengths
The authors took numerous approaches to prove that L-leucine uptake of M. tuberculosis is an important physiological phenomenon and may be effectively targeted by 'Semapimod'. This study utilizes a series of experiments using a broad set of tools to justify how the leucine uptake pathway of M. tuberculosis may be targeted to design new anti-tubercular therapy.
Weaknesses
(1) The study does not explain how L-leucine is taken up by M. tuberculosis, leaving the mechanism unclear. Even though 'Semapimod' binds to the PpsB protein, the relevant connection between changes in PDIM and amino acid transport remains incomplete.
While Leucine uptake involves specific transporters in other bacteria, such transport system is not known in Mtb. By screening small molecule inhibitors, we came across a molecule, semapimod, which selectively kills the leucine auxotroph (mc2 6206), but not the WT Mtb. To understand the underlying mechanism of differential susceptibility of the WT and auxotrophic strains to this molecule, we evaluated the effect of restoration of leuCD and panCD expression on susceptibility of the auxotrophic strain to semapimod. Interestingly, our results demonstrated that upon endogenous expression of leuCD genes, mc2 6206 strain becomes resistant to killing by semapimod. In contrast, no effect of panCD expression was observed on semapimod susceptibility of mc2 6206. These findings were further substantiated by gene expression analysis of semapimod treated mc2 6206, which exhibits differential regulation of a set of genes that are altered upon leucine depletion in Mtb as well as in other bacteria. Overall results thus provide first evidence of perturbation of L-leucine uptake by semapimod treatment of the leucine auxotroph.
To further gain mechanistic insights into the effect of semapimod on leucine uptake in Mtb, we generated the semapimod resistant strain which exhibits point mutation in 4 genes including ppsB. Interestingly, overexpression of wild-type ppsB, but not of other genes, restored susceptibility of the resistant bacteria to semapimod. Our observations that semapimod interacts with PpsB, and semapimod resistant strain accumulates mutation in PpsB resulting in loss of PDIM together support the involvement of cell-wall PDIM in regulation of L-leucine transport in Mtb.
As mentioned above, we anticipate that semapimod treatment brings about certain modifications in PDIM which becomes more restrictive to L-leucine. A comprehensive future study will be helpful to examine the effect of semapimod on Mtb physiology.
(2) Also, the fact that the drug does not function on WT bacteria makes it a weak candidate to consider its usefulness for a therapeutic option.
We agree that semapimod is not an appropriate drug candidate against TB owing to its inhibitory effect on production of proinflammatory cytokines such as TNF-α, IL-1β, and IL-6 that help pathogen establish chronic infection. However, a significant reduction in bacterial loads in lungs and spleen upon semapimod treatment despite inhibition of proinflammatory cytokines clearly indicates bacterial dependence on host-derived exogenous leucine during intracellular growth. Therefore targeting L-leucine uptake can be a novel therapeutic strategy against TB.
Reviewer #3 (Public review):
(1) Agarwal et al identified the small molecule semapimod from a chemical screen of repurposed drugs with specific antimycobacterial activity against a leucine-dependent strain of M. tuberculosis. To better understand the mechanism of action of this repurposed anti-inflammatory drug, the authors used RNA-seq to reveal a leucine-deficient transcriptomic signature from semapimod challenge. The authors then measured a decreased intracellular concentration of leucine after semapimod challenge, suggesting that semapimod disrupts leucine uptake as the primary mechanism of action. Unexpectedly, however, resistant mutants raised against semapimod had a mutation in the polyketide synthase gene ppsB that resulted in loss of PDIM synthesis. The authors believe growth inhibition is a consequence of decreased accumulation of leucine as a result of an impaired cell wall and a disrupted, unknown leucine transporter. This study highlights the importance of branched-chain amino acids for M. tuberculosis survival, and the chemical genetic interactions between semapimod and ppsB indicate that ppsB is a conditionally essential gene in a medium depleted of leucine.
The conclusions regarding the leucine and PDIM phenotypes are moderately supported by experimental data. The authors do not provide experimental evidence to support a specific link between leucine uptake and impaired PDIM production. Additional work is needed to support these claims and strengthen this mechanism of action.
As mentioned above, overall results from this study provide first evidence of perturbation of L-leucine uptake by semapimod treatment of the leucine auxotroph. Our observations that semapimod interacts with PpsB, and semapimod resistant strain accumulates mutation in PpsB resulting in loss of PDIM together support the involvement of cell-wall PDIM in regulation of L-leucine transport in Mtb.
As hitherto mentioned, it appears that semapimod treatment brings about certain modifications in PDIM which becomes restrictive to L-leucine. Future studies are required to gain detailed mechanistic insights into the effect of semapimod on Mtb physiology.
(2) Since leucine uptake and PDIM synthesis are important concepts of the manuscript, experiments would benefit from exploring other BCAAs to know if the phenotypes observed are specific to leucine, and adding additional strains to the 2D TLC experiments to provide confidence in the absence of the PDIM band.
We thank the peer reviewer for this suggestion. We would be happy to analyse the effect of semapimod on the level of other amino acids including BCAA by mass spectrometry.
(3) The intriguing observation that wild-type H37Rv is resistant to semapimod but the leucine-auxotroph is sensitive should be further explored. If the authors are correct and semapimod does inhibit leucine uptake through a specific transporter or disrupted cell wall (PDIM synthesis), testing semapimod activity against the leucine-auxotroph in various concentrations of BCAAs could highlight the importance of intracellular leucine. H37Rv is still able to synthesize endogenous leucine and is able to circumvent the effect of semapimod.
We thank the peer reviewer for this suggestion. We would explore the possibility of analysing the effect of increasing concentrations of BCAAs on mc2 6206 susceptibility to semapimod.
Recommendations for the authors:
(1A) Intracellular leucine can decrease from:
inhibition of transport/uptake via semapimod as the authors claim or
decreased uptake/requirement of many metabolites due to cells entering static growth arrest from challenge by semapimod
To rule out the growth-inhibitory effect of semapimod on L-leucine uptake, we estimated intracellular L-leucine in Mtb after brief exposure of 24 hours to 50ng/ml semapimod (kindly refer Materials and Methods). We confirmed that 24 hours of treatment with 50ng/ml semapimod does not cause cells entering static growth arrest.
(1B) increased consumption/utilization of leucine for some programmed response to semapimod challenge
Our results show reduced expression of genes involved in leucine catabolism such as accD1, bkdA and bkdB in semapimod-treated cells, and thus the above hypothesis seems unlikely.
(1C) Additional metabolites should be measured to determine the specificity of the semapimod challenge.
As mentioned below, we measured intracellular valine in the semapimod-treated Mtb 6206 by LC-MS/MS, which shows no change in its level. These observations thus corroborate a specific effect of semapimod on L-leucine level in the cell.
(2) The effect of Semapimod on L-leucine uptake is largely based on indirect evidence, without showing reduced transport of the amino acid. Gene expression data is not enough to prove that the amino acid transport is blocked. More compelling evidence is required to confirm this mechanism.
The authors could perform leucine uptake assays to directly confirm the functioning of Semapimod, inhibiting L-leucine transport. Another possibility would be to try out measuring intra-bacterial leucine levels for drug-treated versus untreated M. tuberculosis strains.
Data presented in the Fig. 3b shows lesser intracellular L-leucine upon semapimod treatment; in contrast, Sem<sup>R</sup> strain exhibits ~3-fold more intracellular L-leucine, as estimated by mass spectrometry (kindly refer our response to comment #6 below). Together, these observations indicate an inhibitory effect of semapimod on L-leucine uptake by the auxotroph.
(3) The authors show that the overexpression of leuC-leuD restores Semapimod resistance in the auxotroph (Figs. 3C-3E). Is it possible to examine Semapimod resistance of WT-H37Rv or the complemented mutant grown in leucine-limiting conditions? This sort of evidence will be more direct on the specific drug-target beyond the auxotroph (mc<sup>2</sup> 6206).
Because endogenous L-leucine synthesis pathway is functional in WT-H37Rv, as well as complemented auxotrophic strain, leucine-limiting conditions are unexpected to yield any effect on susceptibility to semapimod.
Author response image 1.

(4) Biolayer Interferometry (BLI) shows Semapimod binds to PpsB (Fig. 6); however, there is no clear evidence that it disrupts PDIM synthesis. More direct evidence would be to study the effect of Semapimod on a ppsB mutant (may be a knock-down). This would prove the specificity of Semapimod for PpsB. Likewise, it would be worth looking into the effect of Semapimod using mutant M. tuberculosis defective for PDIM synthesis.
As recommended by the peer reviewer, we created the ppsB knockdown strain in the Mtb mc2 6206 by CRISPRi and examined its vulnerability to semapimod treatment. As can be seen in the Author response image 1, ppsB KD strain shows lesser susceptibility to semapimod when compared with the pDcas9-control strain which exhibits significant growth inhibition on the 7H11-OADS-PL agar plate containing 200nM semapimod.
(5) Metabolomics experiments would benefit from including other control BCAAs like isoleucine and valine to determine if decreased intracellular levels of leucine are specific to semapimod or a general consequence of growth arrest from an antimicrobial agent.
As suggested by the reviewer, we measured intracellular valine as well as proline levels in the semapimod-treated Mtb 6206 by LC-MS/MS; data presented in the supplimentry figure 5 clearly show no change in their levels upon semapimod treatment.
(5) Figure 3c, pyrazinamide susceptibility assay could be included on the panCD strain to ensure complementation leads to functional panCD. Parent strain would be resistant to PZA, complement strain would be susceptible. (doi: 10.1038/s41467-019-14238-3).
The wild-type Mtb 6206 is unable to grow in the absence of pantothenate. We verified resumption of growth of Mtb 6206 in 7H9-OADS-L-leucine medium lacking pantothenate upon PanCD overexpression, which provides more direct evidence of the expression of functional copies of panCD genes.
(6) does the Sem-R mutant have increased levels of leucine?
As can be seen in the supplimentry figure 7, Sem<sup>R</sup> strain shows ~3.0 fold increase in the intracellular L-leucine level when compared with the WT strain. In contrast, a comparable level of another BCAA– valine, is observed in both the strains
eLife Assessment
This study presents valuable findings on the differential effects of RNA on the phase separation, aggregation dynamics, and bioactivity of PSMα3 and LL-37. The authors provide solid evidence from complementary biophysical and cell-based experiments that RNA influences peptide assembly and associated in vitro activities. The study is of interest for understanding interactions between amyloidogenic peptides and nucleic acids, although the physiological significance and some aspects of the mechanistic interpretation would benefit from further clarification.
Reviewer #1 (Public review):
Summary:
The manuscript by Rayan et al. aims to elucidate the role of RNA as a context-dependent modulator of liquid-liquid phase separation (LLPS), aggregation, and bioactivity of the amyloidogenic peptides PSMα3 and LL-37, motivated by their structural and functional similarities.
Strengths:
The authors combine extensive biophysical characterization with cell-based assays to investigate how RNA differentially regulates peptide aggregation states and associated cytotoxic and antimicrobial functions.
Weaknesses:
While the study addresses an interesting and timely question with potentially broad implications for host-pathogen interactions and amyloid biology, some aspects of the experimental design and data analysis require further clarification and strengthening.
Reviewer #2 (Public review):
In this paper, Rayan et al. report that RNA influences cytotoxic activity of the staphylococcal secreted peptide cytolysin PSMalpha3 versus human cells and E. coli by impacting its aggregation. The authors used sophisticated methods of structural analysis and describe the associated liquid-liquid phase separation. They also compare to the influence of RNA on aggregation and activity of LL-37, which shows differences to that on PSMalpha3.
That RNA impacts PSM cytotoxicity when co-incubated in vitro becomes clear. However, I have two major problems with this study:
(1) The premise, as stated in the introduction and elsewhere, that PSMalpha3 amyloids are biologically functional, is highly debatable and has never been conclusively substantiated. The property that matters most for the present study, cytotoxicity, is generally attributed to PSM monomers, not amyloids. The likely erroneous notion that PSM amyloids are the predominant cytotoxic form is derived from an earlier study by the authors that has described a specific amyloid structure of aggregated PSMalpha3. Other authors have later produced evidence that, quite unsurprisingly, indicated that aggregation into amyloids decreases, rather than increases, PSM cytotoxicity. Unfortunately, yet other groups have in the meantime published in-vitro studies on "functional amyloids" by PSMs without critically challenging the concept of PSM amyloid "functionality". Of note, the authors' own data in the present study that show strongly decreased cytotoxicity of PSMalpha3 after prolonged incubation are in agreement with monomer-associated cytotoxicity as they can be easily explained by the removal of biologically active monomers from the solution.
In their revision and in the rebuttal, the authors have further described their concept regarding what they call "functionality" of PSMalpha3 amyloids. They now admit that monomers are the active cytolytic form, like other researchers have stressed, whereas amyloids are not. This represents a considerable difference to earlier papers in which they ascribed functionality, i.e. cytolytic capacity, to PSMalpha3 amyloids, a claim that has raised considerable controversy. Now, they use the term "functional " to describe that PSMalpha3 amyloids, while not cytolytic, can be reversed to a cytolytic monomeric state, calling them a "dynamic reservoir". There is no evidence that such a reservoir is necessary for the cytolytic activity of the monomers to be established; also, there is no evidence that in a biological system, such an amyloid reservoir exists. To continue calling PSMalpha3 amyloids "functional" based on this - considerably changed - concept of the authors appears inappropriate, given the finally admitted absence of cytolytic activity of the PSM amyloids in addition to the continuing complete lack of evidence of any biological relevance of PSM amyloid formation.
(2) That RNA may interfere with PSM aggregation and influence activity is not very surprising, given that PSM attachment to nucleic acids - while not studied in as much detail as here - has been described. Importantly, it does not become clear whether this effect has biologically significant consequences beyond influencing, again not surprisingly, cytotoxicity in vitro. The authors do show in nice microscopic analyses that labeled PSMalpha3 attaches to nuclei when incubated with HeLa cells. However, given that the cells are killed rapidly by membrane perturbation by the applied PSM concentrations, it remains unclear and untested whether the attachment to nucleic acids in dying cells makes any contribution to PSM-induced cell death or has any other biological significance.
Overall, the findings can be explained in a much more straightforward way with the common concept of cytotoxicity being due to monomeric PSMs, and the impact of nucleic acids on cytotoxicity being due to lowering of the concentration of that active form by RNA attachment. Further limiting the significance of the findings, whether this interaction has any biological significance on the physiology or infectivity of the PSM producer remains largely unexplored.
Further remarks:
• Circumstantial evidence based on the "amyloid inhibitor", EGCG: The results with EGCG, which has been shown to have a moderate amyloid-reducing effect on PSMalpha 1 and PSMalpha4, should not be taken as evidence for amyloid-based cytotoxicity. While increased concentrations of EGCG reduced the cytotoxic effect of PSMalpha3, it is not convincingly shown that this is due to a lower concentration of amyloid vs. monomeric PSM.
• It is appreciated that the authors refrain from presenting the unsubstantiated concept of "functional" PSM amyloids in the discussion. However, wording in that direction must also be removed from other parts of the manuscript (e.g. "bioactive fibrillar polymorphs". "The formation of cross-alpha amyloids has been correlated with toxic activity", etc.), generally refraining from uncritically implying that amyloid formation underlies PSM biological activity, and rather discussing that the much more likely explanation of the findings is a lowering of cytolytically active, monomeric PSM concentration.
• Discussion: "PSM alpha3 interaction with nucleic acids within human cells ...supports a comparable mechanism...". Delete. Unsubstantiated.
• The authors should cite papers that have argued against their hypothesis and not only their own manuscripts.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
Summary:
The manuscript by Rayan et al. aims to elucidate the role of RNA as a context-dependent modulator of liquid-liquid phase separation (LLPS), aggregation, and bioactivity of the amyloidogenic peptides PSMα3 and LL-37, motivated by their structural and functional similarities.
Strengths:
The authors combine extensive biophysical characterization with cell-based assays to investigate how RNA differentially regulates peptide aggregation states and associated cytotoxic and antimicrobial functions.
Weaknesses:
While the study addresses an interesting and timely question with potentially broad implications for host-pathogen interactions and amyloid biology, several aspects of the experimental design and data analysis require further clarification and strengthening.
Major Comments:
(1) In Figure 1A, the author showed "stronger binding affinity" based on shifts at lower peptide concentrations, but no quantitative binding parameters (e.g., apparent Kd, fraction bound, or densitometric analysis) are presented. This claim would be better supported by including: (i) A binding curve with quantification of free vs bound RNA band intensities (ii) Replicates and error estimates (mean {plus minus} SD).
We thank the reviewer for this suggestion. To quantitatively support the binding differences observed in Figure 1A, we have now performed densitometric analysis of the EMSA data and included the results in Figure S1. The analysis showed that the Kd for PSMα3 binding to polyAU and polyA RNA is in the same order of magnitude but lower for the polyAU, indicating a stronger binding. A description was added to the results in lines 137-145 of the revised version.
(2) The authors report droplet formation at low RNA (50 ng/µL) but protein aggregation at high RNA (400 ng/µL) through fluorescence microscopy. However, no intermediate RNA concentrations (e.g., 100-300 ng/µL) are tested or discussed, leaving a critical gap in understanding the full phase diagram and transition mechanisms.
Our initial choice of 50 ng/µL (low RNA) and 400 ng/µL (high RNA) was guided by a broader RNA titration performed by turbidity measurements across 0, 10, 20, 50, 100, 200, and 400 ng/µL (Figure S2 in the revised version). In this screen, turbidity increased up to 50 ng/µL and then decreased dose-dependently from 100–400 ng/µL. We interpret this non-monotonic behavior as consistent with a transition from a droplet rich regime (maximal light scattering at intermediate dense-phase volume) toward conditions where assemblies become larger and/or more compact and sediment out of the optical path. This is described in lines 158-161 of the revised version.
Of note, additional intermediate RNA conditions (100 and 200 ng/µL) are included in Figure S14 (of the revised version). While these experiments were performed under the heat-shock perturbation, they nevertheless support the central point that RNA tunes assembly state across intermediate concentrations rather than producing a binary low/high outcome.
Importantly, we agree with the reviewer that a full phase diagram would be the most rigorous way to define the transition mechanism. However, establishing csat and constructing a complete phase diagram would require systematic measurements of dilute-phase concentrations (e.g., centrifugation/quantification or fluorescence calibration), controlled ionic strength titrations, and time-resolved mapping, which is beyond the scope of the present study. We have therefore revised the text to avoid implying that we provide a complete phase diagram. Instead, we frame our results as a qualitative with multi-assay characterization showing that RNA concentration drives a shift from liquid-like condensates (at low RNA) toward solid-like assemblies (at high RNA), with an intermediate regime suggested by the turbidity transition and supported by additional imaging under stress. Finally, to address the “critical gap” concern directly, we add a sentence (lines 239-241) stating that: “Future work will be required to quantitatively define the phase boundaries and delineate the dominant mechanisms, such as sedimentation, dissolution, or coarsening/aging, across intermediate RNA concentrations”.
(3) Additionally, the behaviour of PSMα3 in the absence of RNA under LLPS conditions is not shown. Without protein-only data, it is difficult to assess if droplets are RNA-induced or if protein has a weak baseline LLPS that RNA tunes. The saturation concentration (csat) for PSMα3 phase separation, either in the absence or presence of RNA, should be reported.
In response to the reviewer’s request, we have added Figure 2F, which shows PSMα3 alone in the absence of RNA under the same conditions. PSMα3 does not form droplets in this condition, indicating that condensate formation is RNA-dependent in the tested conditions. This is referred to in the text in lines 190-193 of the revised version. Please see our response about determining the csat in the response to the previous comment.
(4) For a convincing LLPS claim, it is important to show: Quantitative FRAP curves (mobile fraction and half-time of recovery) rather than only microscopy images and qualitative statements.
We have included quantitative FRAP analysis in Figure S4 of the revised version, showing normalized recovery curves along with extracted mobile fractions and half-times of recovery (t₁/₂). These quantitative measurements support the dynamic nature of the PSMα3–RNA. This is referred to in the text in lines 179-184 of the revised version.
(5) The manuscript highly relies on fluorescence microscopy to show colocalization. However, the colocalization is presented in a qualitative manner only. The manuscript would benefit from the inclusion of quantitative metrics (e.g., Pearson's correlation coefficient, Manders' overlap coefficients, or intensity correlation analysis).
In response, we have added quantitative colocalization analysis to the revised manuscript. Specifically, we now report Pearson’s correlation coefficients and Manders’ overlap coefficients for the dual-channel fluorescence microscopy datasets in Figure S5 of the revised version. These metrics provide an objective measure of co-distribution and complement the qualitative imaging.
The analysis supports that at low RNA concentrations (droplet/condensate conditions), PSMα3 and RNA show strong colocalization, consistent with RNA being incorporated within, or closely associated with, the peptide-rich phase. In contrast, at high RNA concentrations, where the assemblies are more solid-like/amyloid-positive, the quantitative coefficients decrease, consistent with reduced overlap and an apparent spatial demixing in which RNA becomes partially excluded from the peptide-rich structures. This is referred to in the text in lines 194-203 of the revised version.
(6) In Figures 3 B and 3C, the contrast between "no AT630 at 30 min, strong at 2 h" (50 ng/μL) and "strong at 30 min" (400 ng/μL) is compelling, but a simple quantification (e.g., mean fluorescence intensity per area) would greatly increase rigor.
We have included quantitative analysis of AmyTracker630 fluorescence intensity in Figure S6 of the revised version, reporting the mean fluorescence intensity per area for the indicated conditions and time points. This quantification supports the qualitative differences observed in Figures 3B and 3C. This is now referred to in the text in lines 233-236 of the revised version.
(7) In Figure S3 ssCD data, if possible, indicate whether the α-helical signal increases with RNA concentration or shows a non-linear dependence, which might link to the LLPS vs solid aggregate regimes.
The ssCD spectra displayed in Figure S7 in the revised version (corresponding to Figure S3 in the original submission) show that the α-helical signature of PSMα3 is markedly enhanced in the presence of RNA compared to peptide alone, as evidenced by increased signal intensity, deeper minima, and more pronounced spectral features characteristic of α-helical structure. Importantly, this enhancement is more pronounced at 400 ng/µL Poly(AU) RNA than at 50 ng/µL, particularly after 2 hours of coincubation, indicating that RNA concentration influences the stabilization of α-helical assemblies. This is now more specifically detailed in the text in lines 258-263 of the revised version.
We note that solid-state CD does not allow direct quantitative deconvolution of secondary structure content (e.g., % helix) in the same manner as solution CD, due to sample anisotropy, scattering, and orientation effects inherent to dried or aggregated films. Consequently, our interpretation is qualitative rather than strictly quantitative. The ssCD data therefore suggest a non-linear dependence on RNA concentration, rather than a simple linear dose–response. This is also expected considering that phase transition, suggested by the other findings, is intrinsically non-linear.
(8) In Figure 5B, FRAP recovery in dying cells may reflect artifactual mobility rather than biological relevance. Additionally, the absence of quantification data limits interpretation; providing recovery curves would clarify relevance.”
We added quantitative FRAP analysis of the effect on PSMα3 within HeLa cells, shown in Figure S8 of the revised version. Compared to PSMα3 assemblies in vitro, nucleolar PSMα3 exhibits slower fluorescence recovery and a reduced mobile fraction. The nucleolus represents a highly crowded, RNA-rich cellular environment, which is expected to impose additional constraints on molecular mobility and likely contributes to the slower recovery kinetics observed in cells. This is now more specifically detailed in the text in lines 324-333 and discussed in lines 597-607 of the revised version.
(9) The narrative conflates cytotoxicity endpoints (membrane damage, PI staining, aggregates) with localization data (nucleolar foci), creating ambiguity about whether nucleolar targeting drives toxicity or is a consequence of cell death. Separating toxicity assessment from localization analysis, or clearly demonstrating that nucleolar accumulation precedes cytotoxicity, would resolve this ambiguity.
We thank the reviewer for raising this important point. We agree that, in the current dataset, cytotoxicity readouts (membrane damage, PI staining, aggregate formation) and subcellular localization (nucleolar accumulation) are observed in close temporal proximity, which limits our ability to unambiguously assign causality. In the experiments presented here, PSMα3 was applied at concentrations known to induce rapid membrane disruption and cytotoxicity in HeLa cells. Under these conditions, PSMα3 accumulates on cellular membranes and penetrates into the cell and nucleus on very short timescales (seconds to minutes), likely preceding the temporal resolution accessible by standard live-cell fluorescence microscopy. As a result, nucleolar accumulation and cytotoxic endpoints are detected essentially concurrently, precluding a definitive determination of whether nucleolar association actively drives toxicity or occurs as a downstream consequence of membrane permeabilization and cell damage.
We therefore emphasize that, in this study, nucleolar localization is presented as a phenomenological observation consistent with RNA-rich compartment association, rather than as a demonstrated causal mechanism of cytotoxicity. We have revised the Discussion (lines 597-607) to clarify this distinction and to avoid implying that nucleolar targeting is the primary driver of cell death.
We agree that resolving this ambiguity would require systematic time-resolved and concentration-dependent experiments, including analysis at sub-toxic PSMα3 concentrations below the membrane-disruptive threshold, combined with orthogonal imaging approaches. Such experiments are planned for future work but are beyond the scope of the present study.
(10) In Figure 8, to strengthen the LLPS assignment for LL-37, additional evidence, such as FRAP analysis or observation of droplet fusion events, would be valuable. This is particularly relevant given that the heat shock conditions (65 °C for 15 minutes) could potentially induce partial denaturation or nonspecific coacervation.
In response to this comment, we have added FRAP analysis of LL-37 assemblies in the revised manuscript (Figure S12), including representative images and corresponding fluorescence recovery curves. The FRAP measurements show minimal fluorescence recovery over the acquisition window, indicating that the LL-37–RNA assemblies formed under these conditions are largely immobile and solid-like, rather than liquid-like droplets. This is now referred to in the text in lines 458-462 of the revised version.
Reviewer #2 (Public review):
In this paper, Rayan et al. report that RNA influences cytotoxic activity of the staphylococcal secreted peptide cytolysin PSMalpha3 versus human cells and E. coli by impacting its aggregation. The authors used sophisticated methods of structural analysis and described the associated liquid-liquid phase separation. They also compare the influence of RNA on the aggregation and activity of LL-37, which shows differences from that on PSMalpha3.
Strengths:
That RNA impacts PSM cytotoxicity when co-incubated in vitro becomes clear.
Weaknesses:
I have two major and fundamental problems with this study:
(1) The premise, as stated in the introduction and elsewhere, that PSMalpha3 amyloids are biologically functional, is highly debatable and has never been conclusively substantiated. The property that matters most for the present study, cytotoxicity, is generally attributed to PSM monomers, not amyloids. The likely erroneous notion that PSM amyloids are the predominant cytotoxic form is derived from an earlier study by the authors that has described a specific amyloid structure of aggregated PSMalpha3. Other authors have later produced evidence that, quite unsurprisingly, indicated that aggregation into amyloids decreases, rather than increases, PSM cytotoxicity. Unfortunately, yet other groups have, in the meantime, published in-vitro studies on "functional amyloids" by PSMs without critically challenging the concept of PSM amyloid "functionality". Of note, the authors' own data in the present study, which show strongly decreased cytotoxicity of PSMalpha3 after prolonged incubation, are in agreement with monomer-associated cytotoxicity as they can be easily explained by the removal of biologically active monomers from the solution.
We thank the reviewer for this important critique and agree that direct cytotoxicity is most plausibly mediated by soluble PSM species, while extensive fibrillation generally reduces toxicity by depleting these forms, a conclusion supported by our data and by other studies (e.g., Zheng et al 2018 and Yao et al 2019). We do not propose mature amyloid fibrils as the primary toxic entities. Rather, we use the term functional amyloid in a regulatory sense, consistent with other biological amyloids whose fibrillar states modulate activity (e.g., hormone storage amyloids or RNA-binding proteins).
In line with emerging findings, we interpret PSMα3 toxicity as arising from a dynamic assembly process rather than from a single static molecular species. We previously showed that PSMα3 forms cross-α fibrils that are thermodynamically and mechanically less stable than cross-β amyloids and readily disassemble upon heat stress, fully restoring cytotoxic activity (Rayan et al., 2023). This behavior contrasts with PSMα1, which forms highly stable cross-β fibrils that do not recover activity after heat shock, suggesting that the limited thermostability of PSMα3 is an evolved feature enabling reversible switching between inactive (stored) and active states.
Consistent with this view, both PSMα1 and PSMα3 are cytotoxic in their soluble states, yet mutants unable to fibrillate lose activity, indicating that fibrillation is required but not itself the toxic end state (Tayeb-Fligelman et al., 2017, 2020; Malishev et al., 2018). Our other studies further show that cytotoxicity toward human cells correlates with inherent or lipid-induced α-helical assemblies, rather than with inert β-sheet amyloids (RagonisBachar et al., 2022, 2026; Salinas 2020, Bücker 2022). Together, these findings support a model in which membrane-associated, dynamic α-helical assembly, which requires continuous exchange between soluble species and growing fibrils, drives membrane disruption, potentially through lipid recruitment or extraction, analogous to mechanisms proposed for human amyloids such as islet amyloid polypeptide (Sparr et al., 2004).
In the present study, we further show that RNA reshapes this dynamic landscape: while PSMα3 alone progressively loses activity upon incubation, co-incubation with RNA preserves cytotoxicity by stabilizing bioactive polymorphs and condensate-like states, whereas high RNA concentrations promote solid aggregation but nevertheless preserve activity. Thus, aggregation is neither inherently functional nor toxic, but context dependent and environmentally regulated. Taken together, our data support a model in which PSMα3 amyloids act as a dynamic reservoir, enabling S. aureus to tune virulence by reversibly shifting between dormant and active states in response to environmental cues such as heat or RNA.
This is now discussed in lines 56-76 and 523-553 of the revised version.
(2) That RNA may interfere with PSM aggregation and influence activity is not very surprising, given that PSM attachment to nucleic acids - while not studied in as much detail as here - has been described. Importantly, it does not become clear whether this effect has biologically significant consequences beyond influencing, again not surprisingly, cytotoxicity in vitro. The authors do show in nice microscopic analyses that labeled PSMalpha3 attaches to nuclei when incubated with HeLa cells. However, given that the cells are killed rapidly by membrane perturbation by the applied PSM concentrations, it remains unclear and untested whether the attachment to nucleic acids in dying cells makes any contribution to PSM-induced cell death or has any other biological significance.
We thank the reviewer for this important point and agree that PSM–nucleic acid interactions are not unexpected and that our data do not support a direct intracellular role for RNA binding in mediating cytotoxicity. Accordingly, we do not propose nucleolar or nuclear association of PSMα3 as a causal mechanism of cell death. At the concentrations used, PSMα3 induces rapid membrane disruption, and nucleic acid association is observed along with membrane attachment, precluding conclusions about intracellular function. This limitation is now explicitly clarified in the revised manuscript. The biological significance of our findings lies instead in extracellular and environmental contexts, where PSMα3 encounters abundant nucleic acids, such as RNA or DNA released from damaged host cells or present in biofilms as now addressed in lines 622631. Our data show that RNA modulates PSMα3 aggregation trajectories, shifting the balance between liquid-like condensates and solid aggregates, and thereby regulates the persistence and timing of cytotoxic activity. In this framework, RNA acts as a context dependent regulator of virulence, rather than as an intracellular cytotoxic cofactor, an aspect which would be studied in depth in future work. This is now addressed in the text in lines 597-607 of the revised version.
Reviewer #3 (Public review):
Summary:
The manuscript by Rayan et al. aims to investigate the role of RNA in modulating both virulent amyloid and host-defense peptides, with the objective of understanding their self-assembly mechanisms, morphological features, and aggregation pathways.
Strengths:
The overall content is well-structured with a logical flow of ideas that effectively conveys the research objectives.
Weaknesses:
(1) Figure 2 displays representative FRAP images demonstrating fluorescence recovery within seconds. To gain a more comprehensive understanding of how recovery after photobleaching varies under different conditions, it is recommended to supplement these images with corresponding quantitative fluorescence recovery curves for analysis.
In response to this comment, we have supplemented the representative FRAP images with quantitative fluorescence recovery curves, reporting normalized recovery kinetics for the indicated conditions. These data are now provided in Figure S4 of the revised manuscript, allowing direct comparison of recovery behavior across conditions (shown by microscopy in Figure 2). In addition, we have included quantitative FRAP analyses for the cellular imaging shown in Figure 5 (presented in Figure S8) and for LL-37 assemblies formed under heat-shock conditions (Figure S12). Together, these additions provide a quantitative framework for interpreting the FRAP results and strengthen the distinction between liquid-like and solid-like assembly states.
(2) Ostwald ripening typically leads to the shrinkage or even disappearance of smaller droplets, accompanied by the further growth of large droplets. However, the droplet size in Figure 2D decreases significantly after 2 h of incubation. This observation prompts the question, what is the driving force underlying RNA-regulated phase separation and phase transition?”
We thank the reviewer for this observation. Across multiple samples, we consistently observe a coexistence of small droplets and larger aggregates, rather than systematic growth of larger droplets at the expense of smaller ones or a uniform decrease in droplet size. In addition, the timescales examined do not allow us to reliably assess whether diffusion-driven droplet coalescence is fast enough to draw firm conclusions about droplet size evolution. This is now addressed in the text in lines 181-184 of the revised version.
A decrease in droplet size over time is nevertheless observed in some instances and is more consistent with a time-dependent conversion of initially liquid-like condensates into more solid-like assemblies, which would reduce molecular mobility and suppress droplet coalescence. In parallel, progressive fibril formation may act as a sink for soluble peptide, leading to partial dissolution or shrinkage of less mature condensates. Together, these observations are consistent with a non-equilibrium aging process, in which RNAregulated assemblies evolve from dynamic condensates toward more solid structures rather than following equilibrium Ostwald ripening.
(3) The manuscript aims to study the role of RNA in modulating PSMα3 aggregation by using solution-state NMR to obtain residue-specific structural information. The current NMR data, as described in the method and figure captions, were recorded in the absence of RNA. Whether RNA binding induces conformational changes of PSMα3, and how these changes alter the NMR spectra? Also, the sequential NOE walk between neighboring residues can be annotated on the spectrum for clarity.
The solution-state NMR experiments were performed specifically to characterize the potential binding of EGCG to PSMα3. Due to the strong tendency of PSMα3 to undergo rapid aggregation and line broadening upon RNA addition, solution state NMR spectra in the presence of RNA could not be obtained at sufficient quality for residue-specific analysis. As suggested, we have updated and annotated the sequential NOE walk between neighboring residues on the relevant NOESY spectra to improve clarity.
(4) The authors claim that LL-37 shares functional, sequence, and structural similarities with PSMα3. However, no droplet formation was observed of LL-37 in the presence of RNA only. The authors then applied thermal stress to induce phase separation of LL-37. What are the main factors contributing to the different phase behaviors exhibited by LL37 and PSMα3? What are the differences in the conformation of amyloid aggregates and the kinetics of aggregation between the condensation-induced aggregation in the presence of RNA and the conventional nucleation-elongation process in the absence of RNA for these two proteins?
We appreciate this important question and have clarified both the basis of the comparison and the origin of the divergent phase behaviors of LL-37 and PSMα3. While PSMα3 and LL-37 share key properties as short, cationic, amphipathic α-helical peptides that self-assemble and interact with nucleic acids, they differ fundamentally in their assembly architectures. PSMα3 is an amyloidogenic peptide that forms cross-α amyloid fibrils, in which α-helices stack perpendicular to the fibril axis. In contrast, LL-37 can form fibrillar or sheet-like assemblies (observed in cryo grids), but these lack canonical amyloid features without clear cross-α or cross-β amyloid order, as so far observed by crystal structures. This is now clarified in different parts of the text of the revised version. Thus, the comparison between the two peptides is functional and physicochemical rather than implying identical amyloid mechanisms. These structural differences likely underlie their distinct phase behaviors.
Because LL-37 does not follow a classical amyloid nucleation–elongation pathway, and high-resolution structural information (e.g., cryo-EM) is currently lacking, partly due to its sheet-like, non-twisted morphology (unpublished results), it is not possible to directly compare aggregation kinetics or nucleation mechanisms between LL-37 and PSMα3. It is possible that amyloidogenic systems such as PSMα3 exhibit greater flexibility in prefibrillar and fibrillar polymorphism, enabling RNA-regulated phase behavior, whereas non amyloid assemblies such as LL-37 are more prone to stress-induced solid aggregation. We note that this interpretation is necessarily tentative and does not imply a general rule, but rather reflects differences evident in the present system.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
Minor Comments:
(1) In the abstract, replacing the word "overriding" with "counteracting" may provide a scientifically neutral tone.
In the course of revision, the abstract was substantially rewritten to more precisely convey the mechanistic framework and key conclusions of the study. As part of this rewrite, the term "overriding" was removed and the language throughout was revised to adopt a more scientifically neutral tone, consistent with the reviewer's suggestion.
(2) In abstract, the final sentence is ambitious but heavy. It may benefit from being split into two shorter sentences, for example:
"These findings establish RNA as a potent, context-dependent modulator of both virulent amyloids and host-defense peptides. They further reveal phase transitions as tunable regulators of peptide activity and potential therapeutic targets across infectious and neurodegenerative diseases."
As part of the broader abstract revision, the final sentence was restructured and the abstract as a whole was rewritten to improve clarity and readability, in the spirit of the reviewer's recommendation.
(3) In the Introduction section,
The phenol-soluble modulins (PSMs) produced by Staphylococci contain amyloid-forming short peptides which play multiple functional roles...", consider "Staphylococcal phenolsoluble modulins (PSMs) are short, amyloidogenic peptides that perform multiple roles central to pathogenesis....
In accordance with the suggestion, the sentence has been revised.
(4) To improve narrative flow in the final paragraph of the Introduction, a short bridging sentence could be added, such as:
"Given these nucleic acid interactions, we next examined whether RNA can drive phase separation or structural reorganization of these amyloidogenic peptides."
We thank the reviewer for this helpful suggestion. It provided an opportunity to clarify an important distinction between the two peptides studied. While LL-37 can self-assemble into higher-order α-helical structures, it is not amyloidogenic, in contrast to PSMα3. We therefore revised the bridging sentence in the final paragraph of the Introduction to read: “Given their shared cationic, amphipathic α-helical character, but distinct amyloidogenic properties, we sought to examine whether RNA differentially influences the assembly landscapes and bioactivity of PSMα3 and LL-37. “
(5) The rationale for selecting Poly(A) and Poly(AU) would benefit from further clarification. It would be helpful to specify whether these RNAs are intended to model particular host or bacterial RNA species, such as AU-rich elements, rRNA-like sequences, or mRNA-like contexts.
Poly(A) and Poly(AU) RNAs were selected as simplified, well-defined model RNAs to probe general peptide–RNA interactions in an unbiased manner, as no prior information was available regarding whether such interactions occur or which specific RNA species might be involved. This rationale is now clarified in the revised text (lines 128–131).
These RNAs are not intended to represent a single biological transcript, but rather generic RNA features relevant to both host and bacterial contexts, including single-stranded homopolymeric regions and AU-rich elements commonly found in mRNAs and stress srelated RNAs. The use of such reductionist RNA models to study RNA–protein interactions, phase behavior, and RNA-modulated aggregation is well established. We nevertheless agree that RNA sequence and structure may influence peptide assembly and activity, and future studies will address sequence-specific and biologically derived RNAs.
(6) In Figure 1A, essential EMSA controls- RNA alone, peptide alone, and a nonspecific peptide or PSMα3 should be included to distinguish specific complexes from artifacts, even if presented in the supplementary information. In addition, a competition assay using unlabeled RNA would help confirm binding specificity and rule out predominantly nonspecific electrostatic interactions; these data could also be reported in the supplementary figures.
An RNA-alone control is already included in Figure 1A of the revised version. The first lane (“0 µM”) shows free Poly(A) or Poly(AU) RNA in the absence of peptide and serves as the negative control against which PSMα3-induced mobility shifts are evaluated. A peptide-alone EMSA cannot be performed, as PSMα3 is highly cationic and does not migrate into the gel in the absence of RNA; moreover, EMSA in this format reports on RNA mobility rather than peptide migration.
With respect to binding specificity, we compared Poly(A) and Poly(AU) RNAs and observed distinct binding behaviors, which would not be expected for purely nonspecific electrostatic interactions. In addition, the extracted Hill coefficients (>1) are consistent with cooperative binding, further arguing against simple charge-driven association. Finally, the RNA-dependent association of PSMα3 is independently supported by fluorescence microscopy and quantitative colocalization analyses, which corroborate the EMSA results. Together, these orthogonal approaches support the relevance of the observed peptide–RNA interactions.
(7) In Figure 1B, there is a time mismatch between EMSA (30 minutes) and TEM (2 hours). If aggregation progresses over time, the EMSA pattern at 2 hours may differ. This point could be acknowledged or experimentally addressed, as RNA-peptide assemblies may evolve from liquid-like condensates to more solid aggregates.
The EMSA and TEM experiments were intentionally performed at different time points to capture distinct stages of the PSMα3–RNA assembly process. The EMSA assay (30 minutes) was designed to probe early RNA–peptide complex formation and binding interactions, before extensive higher-order aggregation occurs. At this stage, we aim to detect mobility shifts reflecting complex formation rather than mature assemblies. In contrast, TEM was performed after 2 hours to visualize later-stage structural outcomes, including fibrillation and morphological reorganization. As aggregation progresses over time, the assemblies evolve from early RNA–peptide complexes into more ordered fibrillar structures, which are best assessed by electron microscopy at later time points. To improve clarity and avoid potential confusion, we have streamlined Figure 1 to focus on the EMSA data, which specifically addresses early binding events. The TEM data were removed from Figure 1 and are now presented in Figure 3, where later-stage structural transitions and fibrillation are shown more comprehensively and in the appropriate mechanistic context.
(8) In Figure 1B, if feasible, complementing TEM with a confirmatory fibril assay (e.g., ThT kinetics) under the same conditions would strengthen the conclusion that the morphology difference is robust, but it is not mandatory.
We attempted to perform ThT fibrillation kinetics under the same RNA containing conditions; however, these assays were not informative for this system. PSMα3 aggregates extremely rapidly, producing an immediate and steep increase in ThT fluorescence (Fig. S9 in the revised version), which prevents reliable resolution of RNA dependent differences in aggregation kinetics or lag phases. In addition, Poly(AU) RNA interferes with ThT readout through electrostatic interactions between the negatively charged RNA and the cationic dye, as well as through RNA-induced changes in fibril morphology, both of which complicate quantitative interpretation of fluorescence kinetics. Based on these technical constraints and prior experience with RNA–amyloid systems, ThT kinetics under identical RNA conditions would not provide a robust or interpretable confirmation of the morphological differences observed by TEM.
(9) In Figure 1B, PSMα3 alone control is missing in TEM images.
A TEM image of PSMα3 alone is included in Figure 3, where we systematically present fibrillation outcomes across different RNA concentrations alongside the peptide-only control. Figure 1 was streamlined to focus on early RNA– peptide interactions assessed by EMSA, whereas Figure 3 provides a comprehensive TEM analysis of later-stage structural outcomes. This organization was chosen to clearly separate early binding events from subsequent assembly transitions and to avoid redundant presentation of TEM images under similar conditions.
(10) Although it is experimentally practical to focus on Poly(AU), the justification is very one-sided. The Poly(A) condition, which yields amorphous aggregates, may be equally informative for understanding toxicity, LLPS, or nonfibrillar states and could be discussed more explicitly.
We agree that Poly(A)-induced amorphous aggregation is informative for understanding non fibrillar assembly states. However, the primary aim of this study was to dissect RNA-dependent regulation of fibrillar assembly and phase behavior, which is most clearly captured using Poly(AU). Poly(A) was therefore included as a comparative condition rather than as a focus for detailed mechanistic analysis. A more systematic comparison of different RNA classes and their effects on non fibrillar states and toxicity is an important direction for future work but is beyond the scope of the present study.
(11) To improve readability of the manuscript, the main text should follow the order of the figure panels (e.g., A, B, C, D, and E) and numbers (Figure 1, 2...) sequentially, so that readers can easily align with the corresponding images.
We have revised the manuscript to improve alignment between the main text and the figures, adjusting panel ordering and numbering where appropriate so that the text now follows the figure panels and figure numbers more sequentially. These changes were made to enhance readability while maintaining a logical visual flow within the figures.
(12) In the result section of Figure 2, the analogy to Ddx4-like systems is a helpful concept, but should be clearly framed as an analogy, not evidence. It would be more accurate to say that the behavior is "conceptually similar to" those systems, while noting that the molecular context is significantly different.
We have revised the text to explicitly frame the comparison to Ddx4-like systems as a conceptual analogy rather than evidence: lines 158-161 in the revised version.
(13) In Figure 4, inclusion of positive and negative controls to validate assay performance (e.g., untreated bacteria or HeLa cells, lysis buffer, media alone) would strengthen confidence in the bioactivity measurements.
We wish to clarify that appropriate positive and negative controls were included in all bioactivity assays and were used to normalize the data presented in Figure 4. For the HeLa cytotoxicity assay (LDH), untreated cells were used to determine spontaneous LDH release (negative control), and cells treated with the manufacturer supplied lysis buffer were used to determine maximum LDH release (positive control). The percent cytotoxicity shown in Figure 4B was calculated relative to these internal controls, as described in the Methods. For the antibacterial assay (PrestoBlue), wells containing E. coli without peptide served as the positive control for 100% viability, while wells containing sterile LB medium alone were used as blanks. Viability values in Figure 4A were normalized to these controls. We have ensured that the Methods section explicitly describes these controls to reinforce confidence in the bioactivity measurements.
(14) To enhance clarity, consider presenting the RNA concentration and time-dependent effects on PSMα3 bioactivity in a comparison table within the main text or as a supplementary figure.
We appreciate this suggestion and carefully considered presenting the data in tabular form. However, we found that graphical representation more effectively conveys the trends, transitions, and comparative patterns between conditions. A table would not adequately capture these relationships.
Reviewer #2 (Recommendations for the authors):
Further remarks:
(1) Circumstantial evidence based on the "amyloid inhibitor", EGCG: The results with EGCG, which has been shown to have a moderate amyloid-reducing effect on PSMalpha 1 and PSMalpha4, should not be taken as evidence for amyloid-based cytotoxicity. While increased concentrations of EGCG reduced the cytotoxic effect of PSMalpha3, it is not convincingly shown that this is due to a lower concentration of amyloid vs. monomeric PSM.
We agree that the effects of EGCG should not be interpreted as evidence for amyloid fibrils being the cytotoxic species. Our data instead support a mechanism in which EGCG primarily targets soluble PSMα3, thereby redirecting its assembly pathway and depleting bioactive species. Specifically, solution-state NMR (Fig. 7) shows that EGCG binds defined residues of monomeric PSMα3, consistent with sequestration of soluble peptide rather than selective inhibition of fibrils. Complementary light and electron microscopy, together with kinetic measurements, indicate that EGCG does not simply stabilize monomers but instead diverts PSMα3 into amorphous, non-functional aggregates, as visualized by TEM (Fig. 6B) and reflected in altered ThT responses (Fig. S9). Importantly, these EGCG-induced aggregates are non-cytotoxic (Fig. 6A/C) and fail to associate with membranes or cells, in contrast to untreated PSMα3, which forms membrane-associated assemblies and induces disruption (newly added Movies S1-S2). Thus, EGCG potentially reduces cytotoxicity by remodeling the aggregation landscape and depleting active soluble species, rather than by selectively inhibiting specific fibril formation. This clarification is now added to the Discussion in lines 554-564 of the revised version.
(2) It is appreciated that the authors refrain from presenting the unsubstantiated concept of "functional" PSM amyloids in the discussion. However, wording in that direction must also be removed from other parts of the manuscript (e.g. "bioactive fibrillar polymorphs". "The formation of cross-alpha amyloids has been correlated with toxic activity", etc.), generally refraining from uncritically implying that amyloid formation underlies PSM biological activity, and rather discussing that the much more likely explanation of the findings is a lowering of cytolytically active, monomeric PSM concentration.
As detailed in our response to Major Comment #1, we agree that uncritical language implying that amyloid fibrils themselves are the cytotoxic species should be avoided. Accordingly, we have revised the manuscript to consistently frame amyloid formation in regulatory terms. Aggregation, depending on context, modulates activity by altering the availability, persistence, and assembly pathways of these species. Distinct aggregation states are therefore presented as correlated with, but not equivalent to, cytotoxic activity, and as components of a dynamic assembly landscape rather than as direct toxic entities.
(3) Discussion: "PSM alpha3 interaction with nucleic acids within human cells ...supports a comparable mechanism...". Please delete this as it is unsubstantiated.
We agree that the original phrasing overstated the evidence. The sentence was removed and the Discussion was revised to clearly frame nucleolar accumulation as a phenomenological observation reflecting PSMα3's intrinsic nucleic acid–binding capacity, rather than as evidence for a comparable intracellular mechanism. Specifically, the revised Discussion (lines 597–607) states that nucleolar localization is "unlikely to represent a distinct intracellular toxic mechanism" and instead "reflects binding competence within RNA-rich compartments following cellular entry." The biological relevance of this interaction, particularly at sub-cytotoxic concentrations, is noted as an open question requiring further investigation.
(4) The authors should also cite papers that have argued against their central hypothesis of "functional" PSM amyloids.
We thank the reviewer for this suggestion. Accordingly, we have revised the manuscript to explicitly cite and discuss studies that argue against amyloid fibrils as the primary cytotoxic species, and that instead attribute PSM cytotoxicity to soluble or membrane-associated forms. These perspectives are now incorporated in the Discussion to provide a balanced view of the field and to clarify how our findings align with, and differ from, existing models of PSM activity.
I think it would be helpful for the introduction to mention how proximity labeling works and the difference between bioid, turboid, and ultraid.
Mia Sato. YouTube reveals millions of incorrect copyright claims in six months. The Verge, December 2021. URL: https://www.theverge.com/2021/12/6/22820318/youtube-copyright-claims-transparency-report (visited on 2023-12-08).
I chose this source because it shows that copyright moderation can go wrong on a very large scale. If so many claims were later overturned, that suggests legal moderation is not always accurate, even on a platform as big as YouTube. That connects well to this chapter’s point that moderation is often shaped by legal pressure, but the results are not always fair.
Without quality control moderation, the social media site will likely fill up with content that the target users of the site don’t want, and those users will leave.
I thought this part was interesting because it shows that moderation is not only about ethics or safety, but also about making a platform usable at all. A lot of arguments about moderation focus on free speech, but this section reminds me that if everything is allowed, the site can become full of spam, scams, or repetitive content and stop being useful to regular users. That makes moderation feel less like an extra feature and more like part of what makes a platform function in the first place.
has a documented life in the theatre.
Tense and Repetition
eLife Assessment
This important work advances our understanding of the development of the visual system. The data presented is compelling and provides a detailed single-cell atlas of post-natal anterior chamber development in mice, highlighting the trabecular meshwork and Schlemm's canal.
Reviewer #2 (Public review):
Summary:
This study presents a detailed single-cell transcriptomic analysis of the post-natal development of mouse anterior chamber tissues. The dataset is robust, consisting of ~130,000 cells collected across seven time points from early post-natal development to adult. Analysis focused on the development of cells that comprise Schlemm's Canal (SC) and trabecular meshwork (TM).
Comments on revisions:
My critiques have been adequately addressed.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
Summary:
This study presents a comprehensive single-cell atlas of mouse anterior segment development, focusing on the trabecular meshwork and Schlemm's canal. The authors profiled ~130,000 cells across seven postnatal stages, providing detailed and solid characterization of cell types, developmental trajectories, and molecular programs.
Strengths:
The manuscript is well-written, with a clear structure and thorough introduction of previous literature, providing a strong context for the study. The characterization of cell types is detailed and robust, supported by both established and novel marker genes as well as experimental validation. The developmental model proposed is intriguing and well supported by the evidence. The study will serve as a valuable reference for researchers investigating anterior segment developmental mechanisms. Additionally, the discussion effectively situates the findings within the broader field, emphasizing their significance and potential impact for developmental biologists studying the visual system.
Weaknesses:
The weaknesses of the study are minor and addressable. As the study focuses on the mouse anterior segment, a brief discussion of potential human relevance would strengthen the work by relating the findings to human anterior segment cell types, developmental mechanisms, and possible implications for human eye disease. Data availability is currently limited, which restricts immediate use by the community. Similarly, the analysis code is not yet accessible, limiting the ability to reproduce and validate the computational analyses presented in the study.
In the revised version we have added an additional paragraph to the discussion section highlighting the human relevance of our work. Additionally, data is public on single cell portal and GEO, accession numbers have been updated. Codes are available on Github (https://github.com/revathi-balasubramanian/Anterior-segment-development-single-cell-data-analysis).
Reviewer #2 (Public review):
Summary:
This study presents a detailed single-cell transcriptomic analysis of the postnatal development of mouse anterior chamber tissues. Analysis focused on the development of cells that comprise Schlemm's Canal (SC) and trabecular meshwork (TM).
Strengths:
This developmental atlas represents a valuable resource for the research community. The dataset is robust, consisting of ~130,000 cells collected across seven time points from early post-natal development to adulthood. Analyses reveal developmental dynamics of SC and TM populations and describe the developmental expression patterns of genes associated with glaucoma.
Weaknesses:
(1) Throughout the paper, the authors place significant weight on the spatial relationships of UMAP clusters, which can be misleading (See Chari and Patcher, Plos Comb Bio 2023). This is perhaps most evident in the assessment of vascular progenitors (VP) into BEC and SEC types (Figures 4 and 5). In the text, VPs are described as a common progenitor for these types, however, the trajectory analysis in Figure 5 denotes a path of PEC -> BEC -> VP -> SEC. These two findings are incongruous and should be reconciled. The limitations of inferring relationships based on UMAP spatial positions should be noted.
(2) Figure 2d does not include P60. It is also noted that technical variation resulted in fewer TM3 cells at P21; was this due to challenges in isolation? What is the expected proportion of TM3 cells at this stage?
(3) In Figures 3a and b it is difficult to discern the morphological changes described in the text. Could features of the image be quantified or annotated to highlight morphological features?
(4) Given the limited number of markers available to identify SC and TM populations during development, it would be useful to provide a table describing potential new markers identified in this study.
(5) The paper introduces developmental glaucoma (DG), namely Axenfeld-Rieger syndrome and Peters Anomaly, but the expression analysis (Figure S20) does not annotate which genes are associated with DG.
(1) We agree that inferring biological relationships from the spatial arrangement of UMAP clusters has limitations and we have qualified our interpretation accordingly in the text. We have also added clarifying language to the trajectory analysis in Figure 5. The intended developmental trajectory is PEC → VP → BEC and SEC; however, the cluster labels in Figure 5 were applied incorrectly. Specifically, VP, BECs cluster was mislabeled as BECs, which led to the confusion. This cluster contains VPs that transition into BECs as well as VPs that are precursors to SECs.
(2) We recently published the P60 dataset separately (Tolman, Li, Balasubramanian et al., eLife 2025); these data consist of integrated single-nucleus multiome profiles that were subjected to in-depth analysis. Additionally, we found that integrating the P60 dataset with the developmental datasets obscured sub-clustering of mature cell types. In future manuscripts, we will pursue a more detailed analysis of TM development and perform time point–specific clustering, similar to the approach we used for endothelial cells (Figure 4e).
Comparing proportions of cells at different ages and as the eyes grows needs to be done cautiously. Notwithstanding the limitations, the proportions of TM1, TM2, and TM3 clusters are expected to be similar between P14 and P21 as the proportions at P14 and P60 are similar when comparing to the separately analyzed P60 data. Importantly, our dissection strategy changed with age: from P2 to P14, we removed approximately one-third of the cornea, whereas at P21 and P60 we removed most of the cornea to help maximize representation of limbal cells as the eyes grew. This change in dissection likely contributed to the reduced number of TM3 cells observed at P21. TM3 cells are enriched anteriorly (at-least in adult) and so are located closer to the corneal cut during dissection of the P21 eyes (which despite being larger than younger ages are still small and more delicate to accurately dissect than at P60) and are therefore more likely to be lost. Additional details are provided in the Methods section and the caveats surrounding our dissection method have now been included.
(3) For Figure 3a and b, we have now pseudo-colored the spaces and provided a quantification of how both TM volume and intratrabecular spaces change with developing age (Figure 3c).
(4) We have now included a supplemental table of markers for developing and mature TM and SC cell types (Table S3).
(5) We have highlighted DG genes in rectangular boxes in Figure S20.
Vorteile Beste Preise im Premium-Segment 600+ Marken 3 Gratisproben pro Bestellung Transparentes Punktesystem Eigene PV-Anlage am Logistikstandort Gute Naturkosmetik-Kategorie Nachteile Kein eigenständiger Nachhaltigkeitsbericht Kaum Exklusivmarken Proben automatisch statt nach Wahl Weniger Premium-Feeling als NB oder Breuninger Operative Maßnahmen ohne übergeordnete Strategie auf Tochter-Ebene
please check to whcih degree they are operationally and thus sustainability wise included in the mother company douglas? if douglas has a strong sustainability strcuture and certification and ParfumeDreams even influences these douglas calculations and reports, it is possible that they in a way share the same goals and should have stronger sustainabiliity crediential than it seems, soley based on being part of the douglas group. please verify this and adapt accordingly. feel free to add or change the entire section 3 ParfumeDreams to adjust based on your research results
Der Vergleich im Überblick
Maybe we just remove the entire box?
Die 7 besten Online-Shops für Luxus-Beauty 2026: Wo Kenner kaufen und was wirklich nachhaltig ist
Please find a better title that's a bit different than the ones that already went online.
Ingredients
Good recipe with unnecessary steps. My changes: 1. Carmelize the onions in 1 T of the butter 2. Add a large clove of garlice and the thyme (I used a tsp. fresh) 3. Add the mushrooms and another T of butter and saute for a few minutes. 4. Add the final T of butter and the 3 Tbsp of flour; cook for a minute 5. Whisk in the stock, cook for about five minutes. 6. Add the dairy, bring to a brief simmer. 7. Cut the heat and add the sherry (no need for a blender!!)
crime is a breach of the criminal law that requires two elements: a mental element (mens rea—a guilty mind) and a physical element (actus reus—the guilty act). It’s not possible to directly establish mens rea using physical science—this must be inferred from the activities and behaviour of the accused. To commit a crime, the accused must have the intent to do wro
sjdvcshdvckshdvc
community, fueled by a strong desire to re-tain what already exists. Typically, the cases are carefully swaddled inappeals to skill, to being good enough, and to working hard enoughto make it. All these tropes are at the center of any sort of merito-cratic appeal. If the harassed were tough enough to take it, then theywould be able to reap the rewards of success. Systemic harassment setsthe terms on which players engage, giving stark advantage to thosewho are not targeted and retaining power for those who have alreadyclimbed the ladder.
There's this non-homogeneous group of white privilege people that yearn to continue playing these types of games, and that may even see themselves as activists when buying them. These may be big mainstream titles, but much like in cinema and TV, their budgets are also big. They know, and they don't mind, they wish these games be as larger and ambitious as possible, ever bigger, and more complex, and continuously "improving", and "innovating" in this sense. They see defending this kind of consumption as defending their identity, defending who they are, defending dark comedy and freedom of speech... freedom of speech, at which point does it become hate speech? Why should their tone for people that have no skin in the game and who aim to get rid of their identity, of their way of living, without asking? You see how both sides have self-reinforcing narratives, and they may even acknowledge this, and although many left-wingers would love to parse out this radically big titles, instead of talking it out and recognising the current exclusionary and biased present (not perpetuating endless debates), some prominent white privilege people push a zero-sum incompatibility competition narrative where one must survive, and it will be them.
You can't expect a person who's played 5000 hours, to quit Fifa overnight.
Also, notably, this is the argument thrown at women and leftists too, arguing for a sort of inevitability of the system we live in. It's "You got your ass touched? Thank you didn't get raped", or "You cry for those people in Sudan? Why don't you go there?"... the cynicism and instrumentalist value-ridden statements that plague these ideologies... it's a red herring distraction that allows toxic masculinity with its inherited Western binary of mind-body, that separates the psyche and tells boys not to cry, not to share their emotions, to grow a thicker skin, to not make a fuss about it, and on the grounds of "tolerance" and not being a "kill-joy" or a "virtue signaler" this allows machoism, revenge sex, and rape fantasies to remain.
Dropdown for type (PREVIOUS_PERIOD, PREVIOUS_YEAR)
The component already exists within the datepicker from PAX, so we don't need to create a new one. The event change from the picker is the same for changing a date and setting comparison date also.
RETURN_INSIGHTS_M3_FLEXIBLE_DATES
the feature flag should be named DATE_PICKER_M3
You can find more UK travel photography in the NHC Search Engine. Or see the original source.
Read the UK Travel Photography Blog for interesting locations and beautiful photos. Personalised with occasional gaming stories.
When The Sassy Owl isn't showing you great photos from the UK, they may be entertaining you with gaming stories. Such as Kubb Lawn Game.
eLife Assessment
This study provides a useful demonstration that, at least for the systems examined, aspects of the entropic contribution to protein-ligand binding can be inferred directly from crystallographic data. In doing so, it strengthens a view of crystal structures as heterogeneous ensembles that are amenable to statistical-mechanical analysis rather than purely static models. The analytical approaches are carefully developed and transparently discussed, with thoughtful consideration of both successful and less effective methods, lending solid support to the central conclusions. However, because the analysis is based on a relatively small and narrowly sampled set of protein-ligand complexes, the generality of these findings remains speculative and will require broader validation.
Reviewer #1 (Public review):
Summary:
The authors show that if they generate a weighted multi-conformer ensemble of structural models to fit crystallographic electron density data, the application of statistical mechanical methodologies to that ensemble can provide reasonable estimates of configurational entropy terms related to protein-ligand binding.
Strengths:
A fair range of proteins (12) and ligands (70) is included in the study. The analytical methodologies are well described. Both successful and less successful analytical approaches are discussed, and the latter are frequently as insightful as the former.
Weaknesses:
Compared to the universe of protein-ligand complexes, this dataset is inevitably very limited, so the generality of the observations made here remains speculative. Though a fair range of proteins is studied, the dynamic range in the binding affinity data is limited. The practical utility of the approach is never really commented on.
Reviewer #2 (Public review):
The manuscript by Miller and Wankowicz (M&W) develops a crystallographic approach to predict the contribution of protein conformational entropy to the total binding entropy using multi-conformer ensemble models. The approach loosely follows the path developed by Wand using NMR relaxation methods. Their approach is to generate local crystallographic order parameters (analogous to NMR order parameters) to estimate protein conformational entropy and then combine this with statements about water entropy. The static view of the ensemble is perhaps easier to grasp, with respect to entropy, than the NMR-based dynamical view. This approach is potentially ground-breaking and of great importance given the ease, relative to NMR, with which the source data can be obtained. However, the approach has several deficiencies, only some of which are noted by the authors.
Like the initial Wand approach (Frederick et al Nature, 2007), M&W develop a simple counting relationship between members of the ensemble and a statement about conformational entropy. For reasons that are not clear, M&W utilize "per residue" scaling, which was initially introduced by Wand but later discarded for the more physically meaningful "per torsion angle" scaling. As noted in the Nature 2007 paper, this assumes uncorrelated occupancy. The current Wand approach (Caro et al PNAS, 2017) subsumes correlated occupancy and potentially incomplete sampling of the ensemble into an empirically determined scaling parameter (sd). This is likely a major contributor to the mysterious 1/4 scaling factor that is introduced. It is not clear to me how discrete conformational states are counted from the qFit models. Using the B-factor, as opposed to a thermal factor, to account for motion in a rotamer well seems suspect. With some irony, M&W only look at chi-1 rotamers in distinct contrast to the NMR approach, which looks at the end of the side chain, which captures the entire disorder. On the other hand, the crystallographic approach "sees" all side chains, whereas the NMR approach, as currently rendered, looks only at methyl-bearing side chains and requires coupling to neighbors to report on all side chains (see Kasinath JACS 2013 and Wand & Sharp ARB 2018).
Nevertheless, as noted by Nature 2007, the fact that a linear relationship is seen between the apparent conformational entropy and total binding entropy suggests that the former is a major component of the latter. It also reinforces the idea that dSrt is constant for higher affinity complexes, i.e., residual rigid-body motion of protein relative to ligand is limited (a conclusion reached in PNAS 2017) but not mentioned. This is an important result.
The classic hydrophobic effect is potentially a significant component of total binding entropy. Here, the manuscript falls flat by focusing on crystallographically resolved waters. As shown in site-resolved detail (Nucci et al, NSMB 2011 and others), hydration water has a range of residual motion (entropy) that will modulate contributions to water entropy upon displacement from an interface. A very clear example of the potential for large contributions was demonstrated in the wet interface of a barnase-DNA complex (PNAS 2017). The fact that the classic dASA treatment failed, I think, points to problems elsewhere in the approach.
I note that the range of ligand types explored by M&W is quite limited as compared to PNAS 2017, making generalization somewhat difficult (see Wand Cur. Opin. Struct. Biol, 2013 for why this is important). Finally, it is disappointing that the authors chose not to examine systems common to PNAS 2017, making direct comparison to the NMR method impossible.
In summary, this manuscript sets the field in a new direction. It is a first serious look at conformational entropy using crystallographic approaches. If fully validated, this approach would permit an explosion of insight since the crystallography is now straightforward, very fast and capable of approaching larger systems, relative to the NMR approach. However, there are missing quantitative elements represented by a formal relationship that is fitted by the data. I do not think this is a fatal flaw for this manuscript, however. If the supplementary material is improved for clarity and completeness (e.g, include tables of thermodynamic data; conformer analysis; B-factors) such that all figures could be independently reproduced and therefore analyzed in different ways, and the comments made above are addressed, if not resolved, then I think this manuscript could become a keystone for this new direction.
eLife Assessment
This study provides valuable insights into how cells maintain sphingolipid homeostasis through transcriptional control and regulated protein degradation in response to changes in sphingolipid levels. The evidence supporting the conclusions is convincing overall, with solid genetic and biochemical approaches, while some mechanistic aspects remain to be clarified. This work will be of interest to researchers studying lipid metabolism and membrane biology.
Reviewer #1 (Public review):
Matsumoto et al. identify Com2, a C2H2-type zinc finger transcription factor not previously linked to sphingolipid metabolism, as a regulator of this pathway in budding yeast. They show that depletion of sphingolipids by myriocin, an inhibitor of serine palmitoyl transferase, increases Com2 expression. This, in turn, promotes the expression of the protein kinase Ypk1 and enhances TORC2-dependent phosphorylation of Ypk1. The authors identify a Com2-binding site in the YPK1 promoter and provide evidence that Com2 functions upstream of Ypk1 to regulate its<br /> expression. They further report that Com2 abundance is controlled by the ubiquitin-proteasome system: degradation of Com2 is inhibited by myriocin treatment and enhanced by phytosphingosine. Mutational analyses of putative phosphorylation and ubiquitination sites support a role for these modifications in regulating Com2 stability. Based on these findings, the authors propose that Com2 acts as a transcriptional regulator of sphingolipid metabolism that responds to sphingolipid levels and promotes Ypk1 expression.
Strengths:
This study provides a valuable finding on the regulation of sphingolipid synthesis by the transcription factor Com2 in budding yeast. The evidence supporting the authors' claims is solid, although additional evidence clarifying the mechanisms and biological significance of ubiquitin-proteasome-mediated degradation of Com2 would strengthen the work. This work will be of interest to microbiologists studying budding yeast.
Weaknesses:
The biological significance of Com2 degradation is not sufficiently clear, which represents an important limitation of the study. It would also be important to determine whether Com2 is actively degraded under normal growth conditions, such as during logarithmic growth in the absence of drug treatment.
Reviewer #2 (Public review):
Summary:
In this study, Matsumoto and co-workers use budding yeast as a model organism to identify and characterize transcriptional mechanisms that homeostatically regulate sphingolipid metabolism. Through a genetic suppressor screen and a series of genetic, molecular, and biochemical analyses, they identify the transcription factor Com2 as a key regulator that responds to sphingolipid levels and regulates the expression of genes such as YPK1, which in turn controls the activity of several enzymes in the yeast sphingolipid biosynthetic pathway.
Com2 itself is further regulated by the ubiquitin proteasome system in response to sphingolipid levels. High sphingolipid levels promote proteasomal degradation of Com2, whereas low sphingolipid levels stabilize Com2. These findings suggest that Com2 is a central component of a feedback system that helps maintain sphingolipid homeostasis.
Strengths:
The identification of Com2 as an upstream regulator of the TORC2-Ypk1 pathway is supported by multiple orthogonal lines of evidence. The authors also provide mechanistic insight into how Com2 protein levels are dynamically controlled through phosphorylation and ubiquitin-mediated degradation. Stabilization of Com2 in response to sphingolipid depletion appears to be required for the transcriptional upregulation of YPK1 expression.
Weaknesses:
Although several important questions remain unresolved, such as which kinases function upstream of Com2 and which ubiquitin ligase(s) target Com2, this work is nevertheless likely to have a meaningful impact on the field of sphingolipid metabolism. The identification of a regulated transcription factor that responds to sphingolipid levels may also be of broader interest to researchers studying membrane homeostasis.
Reviewer #3 (Public review):
This paper extends the authors' 2022 studies of how the synthesis of membrane sphingolipids is regulated in budding yeast. Here, they hypothesized that overexpression of a protein involved in sphingolipid (SL) biosynthesis would confer resistance of lip1-1 cells, which are Dox-inducibly defective in expression of a ceramide synthase regulatory subunit, to myriocin (Myr), a serine palmitoyltransferase inhibitor that inhibits SL synthesis. To test this idea, they transformed lip1-1 cells with a multi-copy genomic library, selecting for Myr resistance. Apart from LIP1 itself and YPK1, a protein kinase downstream of TORC2, COM2, which encodes the Com2 C2H2-type zinc finger transcription factor, was the most frequent hit in the screen. They went on to show that com2Δ cells exhibited Myr sensitivity, and that Com2 protein expression was induced under conditions that reduced complex sphingolipid synthesis, such as Myr-treatment. Using ypk1-as ypk2Δ cells and the 3-MB-PP1 Ypk1as a selective Ypk1as kinase inhibitor, they showed that Com2 phosphorylation was independent of Ypk1 activity, suggesting that Ypk1 lies downstream of Com2. Consistently, Myr treatment, which reduces SL synthesis, resulted in an increase in both Com2 and Ypk1 proteins. By generating a Ptet-off-GFP-COM2 strain they showed that when Dox was removed to induce GFP-Com2 overexpression, Myr resistance was increased. They went on to show that Com2 binds to a Com2 response element in the YPK1 promoter and drives expression of Ypk1. This was confirmed by showing that expression of a YPK1-driven lacZ reporter gene was also elevated when GFP-Com2 overexpression was induced. CRISPR deletion of the putative Com2-binding site (CBS) from the endogenous YPK1 promoter was used to generate PYPK1-ΔCBS cells, which showed a significant reduction in Ypk1 expression and exhibited intermediate Myr sensitivity, suggesting that Com2 is important for but not the only regulator of Ypk1 expression. Analysis of SL levels showed that they largely paralleled the levels of Ypk1 protein and active pT662 Ypk1. Using deletion analysis of the COM2 gene, they showed that residues 2-190 and the C-terminal DNA binding domain of Com2 were essential for Com2 function in the SL synthesis pathway. Deletion of {greater than or equal to}40 amino acids from the N-terminus increased expression of Com2 protein irrespective of Myr treatment, suggesting that Com2 protein levels are regulated by protein stability. Consistently, they found the high level of Com2 protein induced by Myr was rapidly reversed by treatment with phytosphingosine (PHS), a ceramide precursor that bypasses the Myr-blocked step and restores SL synthesis. The reduction in Com2 protein plus PHS was prevented by MG132 proteasome inhibitor treatment and led to the accumulation of polyUb-Com2 species, consistent with Com2 being negatively regulated by SL-induced UPS-mediated degradation. Based on the use of selective inhibitors of different steps in SL synthesis, they showed that SL biosynthesis up to the level of MIPC (mannnosyldiinositol phosphorylceramide) is required for the SL-mediated degradation response. Based on individual and combined K to R mutagenesis of the three Lys in Com2 1-49, they showed that K23, K35 and K51 in combination are needed for PHS-induced Com2 degradation, and therefore are likely to be the main Com2 Ub sites. Finally, they observed that PHS induced an increase in K3R Com2 phosphorylation, finding that an S/T10A mutant was only weakly phosphorylated and was resistant to PHS-induced degradation, suggesting that phosphorylation of Com2 is required for PHS-dependent degradation.
The paper is clearly written, and the data in Figures 1-6 show convincingly that the Com2 zinc finger protein, by inducing the expression of a set of genes, including YPK1 and LCB1, plays an important role in sphingolipid (SL) homeostasis in yeast under conditions when sphingolipid levels are low. However, the data in Figures 7 and 8, where the authors provide evidence that the Com2 protein was rapidly degraded in a proteasome-dependent manner in response to phytosphingosine (PHS) treatment, dependent on the N-terminal 40 residues of Com2 and a combination of three Lys residues in this region, are intriguing but incomplete. There are a number of issues, including the identity of the Com2 ubiquitylation sites. They showed that the K23/35/51R Com2 mutant was stabilized, but did they provide direct evidence that these three Lys are in fact ubiquitylated (e.g. GG-K peptide enrichment based MS analysis of Ub-Com2 from PHS-treated, MG132-treated cells). They showed that PHS treatment increased Myc13-tagged Com2 ubiquitylation in the presence of MG132, but did not show that the K3R Com2 mutant (or the S/T10A phosphorylation site Com2 mutant) failed to be ubiquitylated. They also found that the WT Com2 and particularly the K3R Com2 mutant protein exhibited hyperphosphorylation in response to PHS treatment, and that mutation of 10 potential pSer sites to Ala abolished this effect, and stabilized the Com2 protein. However, it is unclear whether the K3R mutation led to increased Com2 hyperphosphorylation per se following PHS treatment, or whether this is because there is more K3R protein, as they suggest might be the case. It is also not clear what protein kinase is responsible or how it might be activated when SL levels are high. In addition, the E3 Ub ligase needed for Com2 degradation was not identified, and it is not clear whether Com2 phosphorylation is directly involved in its recognition by a phosphodependent E3 Ub ligase, as they propose in the model shown in Figure 9. Finally, and perhaps most importantly. It is unclear how elevated levels of phytosphingosine or any sphingolipid are sensed by the Com2 pathway in order to switch on the degradation response as a negative feedback event. The model depicted in Figure 9 exposes all of these unknowns. The paper would be significantly strengthened by additional experiments defining how complex SL levels are sensed, how Com2 is phosphorylated in response to SL sensor signals, and how (phospho)Com1 is recognized for ubiquitylation and degradation.
In summary, the finding that the Com2 zinc finger transcription factor is an upstream regulator of the sphingolipid biosynthesis pathway in budding yeast, acting as part of an SL sensor system to maintain sphingolipid homeostasis, is new and potentially important. However, more mechanistic work needs to be done to address the unanswered questions raised by the data in Figures 7 and 8.
eLife Assessment
This study presents important findings on the molecular mechanisms governing how the natural killer cell receptor KIR2DL4 interacts with HLA-G and undergoes internalization. The authors provide solid evidence for an allosteric disulfide-bond switch that regulates receptor activity, using a multifaceted approach that includes mutagenesis, mass spectrometry, and imaging. The work would be further strengthened by validating these mechanisms in primary immune cells and providing direct structural evidence for the proposed ligand-binding interface.
Reviewer #1 (Public review):
Summary:
This paper asks how the NK cell receptor KIR2DL4 binds HLA-G and undergoes endocytosis. The authors propose that an allosteric disulfide-bond switch controls whether the receptor is in a ligand-binding or non-binding state, and they support this model using mutagenesis, imaging, mass spectrometry, and structural prediction.
Strengths:
A major strength is the use of diverse, complementary approaches to validate the central claim. The authors combined unbiased random mutagenesis to identify key residues, confocal microscopy to track cellular localization , and mass spectrometry to quantify the redox states of specific disulfide bonds. These methods consistently support a single model: an allosteric disulfide switch. The transition between a Cys10-Cys28 bond and a Cys28-Cys74 bond serves as a functional switch that controls whether the receptor resides at the plasma membrane to bind ligand or remains inactive in endosomes.
Weaknesses:
The core model is interesting, but some of the strongest mechanistic claims still rely heavily on structure prediction rather than direct structural evidence, especially the proposed HLA-G contact surface in Figure 6.
The paper supports an effect of the disulfide state on trafficking and uptake, but the case for direct KIR2DL4-HLA-G binding still feels somewhat indirect. The manuscript itself notes that direct binding had not been previously shown, and the current explanation partly depends on inference about which disulfide state is present.
Most of the main experiments are done in transfected 293T cells, so it is still not fully clear how strongly this mechanism carries over to the more relevant NK-cell setting discussed in the paper.
The cellular evidence for the PDI story is not specific, since it depends a lot on inhibitor and blocking experiments that could affect the broader extracellular redox environment.
Reviewer #2 (Public review):
Summary:
Rajagopalan et al show how extracellular domain features regulate KIR2DL4 internalization. The trafficking phenotypes of cysteine mutants are logically organized, and well-summarized in a Table. The disulfide mapping and differential alkylation strategy are appropriate and provide strong support for alternative disulfide configurations in D0. The higher accessibility or more selective reduction of Cys10-Cys28 as compared to Cys28-Cys74 by PDI is a key mechanistic anchor.
Strengths:
The identification of a conformational switch in KIR2DL4 is conceptually novel. Experimental elegance, detailed and well-written.
Weaknesses:
Most of the mechanistic work was shown in HEK293. The authors should exhibit relevance using primary NK cells (using primary NK)
Do you love reading about Science, Technology, Engineering, and Mathematics? Then cyprianj's STEM Blog is just right for you. Browse the STEM Blog now for interesting facts, experience and opinions.
As a researcher studying killer toxin, I have been waiting for a structural prediction based on AF2 (or AF3). However, the structure shown is that of the precursor, including the γ-peptide, so I request a structural prediction of the dimer formed after cleavage at the processing site, if possible. Of course, I understand that the precursor structure is also important, as the protein folds as a precursor before undergoing processing. I believe the SMKT in Figure 7 is the structure of the β-chain from the PDB, but I have confirmed that the actual SMKT forms a structure as an α-β dimer and that the individual subunits do not have a structure on their own. I thought this might be inappropriate for use in this figure.
Horizontal carousels may make navigation harder - Potential accessibility issue
This horizontal carousel may create accessibility challenges. Some content is partially hidden off-screen, and users need to scroll sideways or use small arrow controls to view everything. This could make navigation less intuitive for keyboard users, screen reader users, or people with motor impairments. From the Operable principle, content should be easy to navigate without requiring precise or difficult interactions.
Large visual product carousel — Potential accessibility issue
This large visual product carousel may be less accessible for some users. Because the section relies heavily on images and horizontal navigation, users with visual impairments, motor impairments, or those using keyboard navigation may find it more difficult to browse all of the content. This connects to the Operable principle, which emphasizes that web interfaces should be easy to navigate without requiring overly precise or complex interaction.
Clear category structure
Clear category labels, such as “Kitchen & appliances” and “Outdoor products,” make the page easier to understand and navigate. This supports the Understandable principle of accessibility because users can quickly identify where different types of content are located. A clear category structure is also helpful for users who rely on screen readers or who may feel overwhelmed by a visually busy shopping page.
“28% off, save $70"
This is a positive accessibility feature because the sale is communicated through both colour and text, rather than colour alone. The red price helps draw attention visually, but the written discount information, such as “28% off, save $70.00,” ensures that users who are colourblind or using assistive technologies can still understand the meaning. This supports the Perceivable principle.
Clear section headings - “Inspiration for every room”
This is a good example of the Understandable principle of web accessibility. Clear section headings help users quickly understand how the page is organized and what kind of content appears in each area. They also support screen reader navigation, since users can move through a page by headings instead of reading every item in order.
Debugging is also like an experimental science. Once you have an idea what is going wrong, you modify your program and try again. If your hypothesis was correct, then you can predict the result of the modification, and you take a step closer to a working program. If your hypothesis was wrong, you have to come up with a new one. As Sherlock Holmes pointed out, When you have eliminated the impossible, whatever remains, however improbable, must be the truth. (A. Conan Doyle, The Sign of Four)
Never thought about it this way. Guess that's part of why it's called compsci.
We now move from the crime scene to the laboratory and the various stages of recovering, documenting, and analysing evidence. Some of the principles and processes will now be familiar to us as they reflect those applied at crime scenes. The new dimension is the specific application of scientific testing of case items and the range of scientific disciplines involved. In this chapter, we will cover the types of examinations carried out in particular case types and the specific scientific and legal procedures required to meet the standards of criminal law.
sdvasdva
ipfs white paper
Anya Kamenetz. Selfies, Filters, and Snapchat Dysmorphia: How Photo-Editing Harms Body Image. Psychology Today, February 2020. URL: https://www.psychologytoday.com/us/articles/202002/selfies-filters-and-snapchat-dysmorphia-how-photo-editing-harms-body-image (visited on 2023-12-08).
The article talks about how using selfies, filters, and photo-editing apps can mess with how people see their own appearance, sometimes leading to what’s called “Snapchat dysmorphia.” It also points out that constantly seeing edited, “perfect” images on social media can make people feel worse about their own bodies and set unrealistic beauty standards.
When you hear the term “online presence,” what comes to mind?
When I think of online presence, I think in social media, content creation and websites. I also associate it with brands and influencers that make that a product or brand be viral.
brew install --cask --no-quarantine hovancik/stretchly/stretchly
Error: Calling the --[no-]quarantine switch is disabled! There is no replacement.
it didn't work for me
It is typical for services to claim some form of access or control over your words and images, such as the right to share it with 3rd party clients, so be prudent and selective about what you think is reasonable or fair.
I feel a really important aspect of content rights nowadays is the use of built-in AI functionality. Because there is no governing body to regulate it, people of all ages/backgrounds can be easily convinced to create using AI because it's free and easy, unknowingly creating personal and derived training data for the firm to feed back into the system.
Malwarebytes is an free add-on to make sure your device is regularly scanned for viruses.
When I first got my desktop computer, a friend of mine suggested that I download Malwarebytes to serve this purpose and better protect my device. I have always appreciated the extra layer of security, and have performed many scans of my computer over the years using Malwarebytes.
However, I've never looked into how it actually works. When Malwarebytes conducts a scan of a computer, what evidence or clues does it look for to detect a virus? Does it scan files, code, or application data? And if it does discover problematic results, what does it do to remedy the problem? It is interesting to me that the term "virus" has become regular vernacular, yet most people don't actually understand what a virus is, or how they can be resolved (myself included).
The volcanic complexes include ring dykes and central domes together with ignimbrites.
excuse me, together with ignimbrites?!?
For example, in BC, educators can use the Fair Dealings Policy[3] to upload and create copies of up to 10% of a copyright-protected work for their courses.
I wonder if this a helpful tool for educators in BC or if it isnt used very much.
For the Benefit of All BeingsTap to unmute2xFor the Benefit of All BeingsGarchen Buddhist Institute 58,573 views 2 years agoSearchInfoShoppingJohn C. McGinley's Secret To Memorizing Lines: Juggling | CONAN on TBSCopy linkIf playback doesn't begin shortly, try restarting your device.Pull up for precise seekingMute7:54•You're signed outVideos you watch may be added to the TV's watch history and influence TV recommendations. To avoid this, cancel and sign in to YouTube on your computer.CancelConfirmUp nextLiveUpcomingCancelPlay NowShareInclude playlistAn error occurred while retrieving sharing information. Please try again later.0:000:22 / 1:40:41Live•Watch full video••35:34Dharma Chat with H.E. Garchen RinpocheDaniel Aitken7.4K views • 1 month agoLivePlaylist ()Mix (50+)22:08Dudjom Rinpoche ~ Knowing the One Thing ~ DzogchenSamaneri Jayasāra - Wisdom of the Masters34K views • 1 month agoLivePlaylist ()Mix (50+)1:16:33THE YOGIS OF TIBET - Rare Documentary FilmNyishar5.9M views • 6 years agoLivePlaylist ()Mix (50+)34:09This Music Will Protect You & Light Your Path | 7 Sacred Native American ChantsNative Soul Romance29K views • 6 days agoLivePlaylist ()Mix (50+)46:05THE SPIRIT OF TIBET : The Life and World of Dilgo Khyentse RinpocheShechen/Dilgo Khyentse Fellowship248K views • 2 years agoLivePlaylist ()Mix (50+)15:17The Child Prince Who Remembered a Past Life — and the Karmapa ControversyLei's Looking Glass264K views • 5 months agoLivePlaylist ()Mix (50+)1:13:37Searching for the Lotus-Born Master : 8 Manifestations of Quantum Energy. Directed by Laurence BrahmShambhala Studio Films | Laurence Brahm783K views • 6 years agoLivePlaylist ()Mix (50+)1:22:21The Hidden System Running the World and How You Can Escape It with Simon DixonNatalie Brunell182K views • 3 weeks agoLivePlaylist ()Mix (50+)23:27Scientists Reveal Shocking Genetic Origin of HungariansGeneTrace and Dust & Data78K views • 10 days agoLivePlaylist ()Mix (50+)1:44:39This Is The Dark State In Action | Fitts, Dixon, DavisKyle Chasse crypto and 2 more7K views • 9 hours agoLivePlaylist ()Mix (50+)20:10The Samādhi Empowerment台灣噶千佛學會Garchen Dharma Institute14K views • 7 years agoLivePlaylist ()Mix (50+)1:40:42Garchen Rinpoche - For the Benefit of All BeingsDrikung Dharma Surya Center277 views • 5 months agoLivePlaylist ()Mix (50+)1x For the Benefit of All Beings
x
The famous rationalist rug is the Home Depot Ethereal shag rug in cream beige. It comes in seven different sizes, and there are also four other color options if you want to be original.
God bless these people
Observed changes in atmospheric CO₂ concentration during the instrumental period andisotopic signatures back to the Little Ice Age are likely explained by natural biogeochem-ical feedbacks, with no detectable anthropogenic contribution.
We know that, "uptake of atmospheric CO2 should react not to a change in emissions, but to a change in concentrations" (Knorr 2009). So if our current anthropogenic emissions of 4.9 ±0.5 ppmv/yr of fossil CO2 were to suddenly cease, the CO2 trend would be reduced by that same amount: 4.9 ±0.5 ppmv/yr. That means it would be negative instead of positive: initially averaging about -2.4 ±0.6 ppmv/year instead of the current average of +2.5 ±0.1 ppmv/year.
In other words, were it not for ongoing anthropogenic CO2 emissions the atmospheric CO2 concentration would currently be falling, rather than rising.
That obviously means that the current upward trend in atmospheric CO2 concentration is entirely due to anthropogenic emissions.
● Mankind is ADDING CO2 to the atmosphere
● 'Nature' (the net sum of all non-anthropogenic carbon fluxes) is REMOVING CO2 from the atmosphere
● The atmospheric CO2 level is currently rising because mankind is currently adding CO2 faster than nature is removing it.
short CO₂ residence times of 3.5–4 years,
This confuses the 3-5 year short "residence time" of carbon in the atmosphere with the roughly 50 year "adjustment time" for added CO2.
It's the adjustment time, not the residence time, which determines the duration of effect of additions of CO2 to the atmosphere.
The four(!) different "lifetimes" of added CO2 are explained here:
Burton, D.A. (2024). "Comment on Stallinga, P. (2023), Residence Time vs. Adjustment Time of Carbon Dioxide in the Atmosphere." OSF Preprints. https://doi.org/10.31219/osf.io/brdq9 (and supplemental material here https://sealevel.info/Comment-on-Stallinga2023/)
When a CO2 molecule is absorbed by a puddle or raindrop, that's the end of its "residence time." When that puddle or raindrop evaporates, releasing the dissolved CO2 molecule back into the air, that's the start of a new "residence time." But, of course, the amount of CO2 in the atmosphere is unaffected when a CO2 molecule temporarily leaves the atmosphere, and then returns to it.
Similarly, if a carbon atom is exchanged between the air and the terrestrial biosphere or ocean, either immediately or perhaps after a growing season, that exchange "resets the timer" if you're counting "residence times" of carbon atoms, but it does so without affecting the amount of CO2 in the air.
David Berman highlights some key details and when he started to talk about them I started to connect them to our community. The first thing that came to my mind was how he connected sustainability with design and what David Berman expressed is that designers should have all there work be beneficial to both the people and overall the whole community. Next I wanted to also mention that David Berman had also mentioned that design can play a positive impact on the community and overall the society. Overall, this was a great lecture to understand all of the key details that David Berman pointed out.
David Berman on Sustainable Design Thinking StrategyTap to unmute2xDavid Berman on Sustainable Design Thinking Strategydavidbermancom 973 views 11 years agoSearchCopy linkInfoShoppingIf playback doesn't begin shortly, try restarting your device.Pull up for precise seekingMute16:02•You're signed outVideos you watch may be added to the TV's watch history and influence TV recommendations. To avoid this, cancel and sign in to YouTube on your computer.CancelConfirmUp nextLiveUpcomingCancelPlay NowI have this medicine.ShareInclude playlistAn error occurred while retrieving sharing information. Please try again later.3:033:31 / 49:15Live•Watch full video••34:42Design Thinking workshop with Justin Ferrell of Stanford d. School at The Irish TimesJohnny Ryan372K views • 12 years agoLivePlaylist ()Mix (50+)47:20Current Work: Michael BierutThe Architectural League8.6K views • 9 years agoLivePlaylist ()Mix (50+)lofi hip hop radio 📚 beats to relax/study toLofi Girl26K watching • 3 years agoLivePlaylist ()Mix (50+)39:16Michael Bierut on how to think like a designerDesign Indaba379K views • 9 years agoLivePlaylist ()Mix (50+)25:215 steps to designing the life you want | Bill Burnett | TEDxStanfordTEDx Talks11M views • 8 years agoLivePlaylist ()Mix (50+)1:11:37Learn Copywriting in 76 Minutes – Harry DryDavid Perell831K views • 1 year agoLivePlaylist ()Mix (50+)37:35Clara Mattei: capitalism is not natural - it’s enforcedChannel 4 News977K views • 2 months agoLivePlaylist ()Mix (50+)22:17The Shadow Docket: Last Week Tonight with John Oliver (HBO)LastWeekTonight891K views • 13 hours agoLivePlaylist ()Mix (50+)32:32The Strange Math That Predicts (Almost) AnythingVeritasium11M views • 9 months agoLivePlaylist ()Mix (50+)20:50The Power of Vulnerability | Brené Brown | TEDTED24M views • 15 years agoLivePlaylist ()Mix (50+)42:56Seth Godin – Leadership vs. Management - What it means to make a differenceNordic Business Forum2M views • 4 years agoLivePlaylist ()Mix (50+)35:50Understand & Improve Memory Using Science-Based Tools | Huberman Lab EssentialsAndrew Huberman124K views • 3 weeks agoLivePlaylist ()Mix (50+)1x David Berman on Sustainable Design Thinking Strategy
Tripple Bottom line includes Profit (Economic), Planet (Environmental), and People (Social), which a lot of companies use as a measurement of success. This means that today’s Designers must be cognate of how their design impacts each attribute of the Tripple Bottom line. For example, when designing a part for a vehicle, the part might be better for the environment if it’s 100% made from recyclable material. However, if that recycled part costs too much to make, it might affect the profit.
we will need to find or create spaces in our lives where we can "practice" democracy -- beginning with our families (responsibilities shared equitably between parents) and our places of work (increased worker participation)
This is interesting the workplace as a place to practice democracy. Most workplaces are hierarchical by design.
These metrics rely almost exclusively on temperature data from the Argo profiling float array.
I wish that were so! Unfortunately, there were no Argo floats in the 20th century, and NCEI OHC estimates go back to 1955!
https://www.ncei.noaa.gov/access/global-ocean-heat-content/index.html
For 2/3 of the time period for which they claim to know OHC there's almost no measurement data from which it can be determined. It's models, all the way down.
every person in an organization is morally responsible for his or her own behavior, and any efforts to change that behavior should focus on the individual.
This is how most corporate ethics training works for protocols and individual compliance. But if the whole system is broken, individual training alone can't fix it.
.
Painter, C. C. The Condition of Affairs in Indian Territory and California. Philadelphia: Indian Rights Association, 1888.
.
Bauer Jr., William J. California Through Native Eyes: Reclaiming History. Seattle: University of Washington Press, 2016. p. 88
our historical traditions place a high value on individual freedom, on personal rights, and on allowing each person to "do her own thing".
This is why the common good is so hard to sell in Western cultures. Rights language always wins the argument. Makes me wonder if the common good approach needs a different vocabulary to actually gain traction.
re and leaving notes in the card.
test
COURSE LEVEL SCOPE NOTES: This course introduces the major intervention strategies used in generalist social work practice, including problem-solving approaches, crisis intervention principles, and evidence-informed decision-making. Learners examine intervention processes across individual, family, group, organizational, and community contexts. Emphasis is placed on culturally responsive, strength-based, and trauma-informed approaches. Learners develop foundational skills in planning and evaluating interventions appropriate for entry-level practice.
Hey just checking this out.
Individuals should be treated the same, unless they differ in ways that are relevant to the situation in which they are involved.
This is the key line. Equal treatment doesn't mean identical treatment. Connects to workplace equity debates same rules don't always produce fair outcomes
By selecting clones based on criteria such as possessing at least one lineage of interest, for example ILCs, and then clustering these clones according to their lineage outputs, it would be possible to identify progenitor populations with distinct lineage output patterns based on the sharing patterns between ILCs and other lineages.
Is the final stage of ILC specification truly the bifurcation point from the NK lineage?
In addition, it would also be interesting to select clones that retained MPPs and classify them by clustering analysis. Such an approach might provide insight into what types of lineage-output progenitors are present within selfrenewing MPPs.
Filtering based solely on Pgen appears likely to exclude cases where truly unique clones, as well as include cases where non-unique clones are retained. Specifically, in Fig. S3, there are cases with Pgen around 10^2 without high barcode frequency, as well as cases with Pgen greater than 10^4 that lie just above the proportional relationship between BC frequency and Pgen. Because barcode clones detected only once among the n=7 samples contain, on average, at least >85.7% unique clone information, it may be possible to generate a more reliable dataset by combining multiple criteria, such as Pgen, read frequency, and uniqueness across samples.
Relying exclusively on a rights approach to ethics tends to emphasize the individual at the expense of the community.
This feels very relevant to business ethics. Companies that focus only on shareholder rights often ignore community harms.
rights should not be the sole consideration in ethical decision-making
This is interesting even the rights article admits rights alone are not enough. Connects to the utilitarianism reading too. Seems like no single framework is sufficient on its own.
it's often difficult, if not impossible, to measure and compare the values of certain benefits and costs.
This is the biggest practical problem. How do you put a number on human dignity or community harm..? Business decisions get made this way all the time though.
the greatest good for the greatest number."
who counts as part of "the greatest number"? Employees? Shareholders? Future generations?
The greatestEnd of search result challenge in business decision-making is moving beyond the letter of the law to create a culture of ethics
Who is actually responsible for building that culture? leadership, employees, or both?
Menu
The navigation menu uses a dropdown feature to organize information into smaller categories. This supports the understandable and operable principles. However, it is crucial that dropdown menus are keyboard accessible, otherwise they can act as a barrier. This is a good practice if it is implemented properly.
Government of Canada
The use of white text on a darker background may be difficult for some users to interact with and read. Some individuals with low vision or colour blindness may struggle with reading this information. This connects to the perceivable principle as it requires for information to be presented clearly and may be also be seen as a poor practice.
Jobs
Links are used to lead individuals to more descriptive pages instead of providing chunks of information. A small description is provided as well. This helps support the understandable principle and is a good practice for accessibility and readability as information can easily be accessed through links and is condensed.
Canada.ca The official website of the Government of Canada
A high contrast text is used here against a colourful background. There is also a simple font used which makes the text easier to read for individuals who have low vision. This supports the perceivable principle. This is a good practice as it enhances legibility and reduces barriers when it comes to accessing content.
short CO₂ residence times of 3.5–4 years
This confuses the 3-5 year short "residence time" of carbon in the atmosphere with the roughly 50 year "adjustment time" for added CO2.
It's the adjustment time, not the residence time, which determines the duration of effect of additions of CO2 to the atmosphere.
The four(!) different "lifetimes" of added CO2 are explained here:
Burton, D.A. (2024). "Comment on Stallinga, P. (2023), Residence Time vs. Adjustment Time of Carbon Dioxide in the Atmosphere." OSF Preprints. https://doi.org/10.31219/osf.io/brdq9 (and supplemental material here https://sealevel.info/Comment-on-Stallinga2023/)
When a CO2 molecule is absorbed by a puddle or raindrop, that's the end of its "residence time." When that puddle or raindrop evaporates, releasing the dissolved CO2 molecule back into the air, that's the start of a new "residence time." But, of course, the amount of CO2 in the atmosphere is unaffected when a CO2 molecule temporarily leaves the atmosphere, and then returns to it.
Similarly, if a carbon atom is exchanged between the air and the terrestrial biosphere or ocean, either immediately or perhaps after a growing season, that exchange "resets the timer" if you're counting "residence times" of carbon atoms, but it does so without affecting the amount of CO2 in the air.
In summary, we find striking mathematical similarity among animal ‘song’ vocalizations and human musical sounds regarding the stability of CES, along with some well-defined differences across evolutionary distant taxa that evolved singing behavior independently (i.e. anurans, birds, primates)
I don't think you have yet shown enough to make this claim. The paper would greatly benefit from the use of null models. Given that all the data used in the paper share informational structure (i.e., they're way detectable in some way as "songs" to humans), are the relationships you identify actually surprising? It's very hard to gauge this without a null, either empirical or simulated, to compare to.
Similarly, it's impossible to know whether the comparisons are fair without a deeper examination of how parameter choice (e.g., window sampling size) may differentially affect the CES estimation across song types. Even if you find significant similarity between human/animal song times compared to a null, how will you know if this isn't a product of bias in your CES estimation?
I took a look at i4 about the right to privacy and how it protects people from government and company interference with their personal information. One thing that stood out to me was how privacy laws differ so much around the world, especially with countries like the European Union having stronger protections through GDPR compared to the US. I also found it interesting how technology and social media have made privacy more complicated because companies can collect big amounts of personal data online. Reading about "mass surveillance" and data collection made me realize how important privacy rights are now that so many things are digital and how difficult it can be to balance safety, security, and personal freedom.
I read i19 and one part of this article that stood out to me was the idea that employees often have an “illusion of privacy” when using workplace communication tools. I learned that many people assume private messages or deleted chats cant be seen, but companies can actually access emails, Slack messages, and even deleted content. This connects strongly to the topic of digital privacy because it shows how technology can blur the line between personal and professional communication. It also made me think about how important transparency is and why workers should clearly know what information employers can monitor and store.
One source from the bibliography that stood out to me was the Washington Post article h8 “Everything Everywhere All at Once is a deeply Asian American film.” The article explains how the multiverse in the movie represents the immigrant experience and the pressure of balancing different identities. I thought this was interesting because it shows that the movie is not only about action or science fiction, but also about family expectations, cultural identity, and generational trauma. The writer explains that Evelyn feels split between many versions of herself, which was meant to show how many immigrants or children of immigrants can feel caught between cultures. This detail helped me better understand why the film connected so strongly with audiences and why it is considered such an important movie for Asian American representation. This was a good read!
One thing I found interesting in this section was the idea of spurious correlations. Just because two things seem connected in data doesnt mean one actually causes the other. The example of comparing COVID cases and Yankee Candle reviews was funny but also showed how easy it is to misinterpret data online. I think this is important because social media companies and researchers use huge amounts of data to make decisions, so people need to think critically about whether the conclusions are actually reliable.
I thought this section was interesting because it showed how social media platforms could change what people see online by filtering posts with positive or negative sentiment. Even though only showing positive news might help some people feel better, I also think it could create an unrealistic view of the world if users never see important negative news. It made me think about how much control algorithms have over people’s emotions and perspectives online.
What Makes the Hanx Writer Click?<br /> by [[Silvia Killingsworth]] in The New Yorker<br /> accessed on 2026-05-11T14:49:13
One editor recalls that you could tell whether your neighbor was being productive that day just by the typewriter sounds coming out of his office (thus pressuring you to get down to work).
Indeed, I.B.M. really knew how to make a keyboard: there is an entire fan site dedicated to the I.B.M. model M keyboard, complete with the full text of the patent for the “buckling spring snap torsional actuator,” granted in 1978.
What characteristics, actions, and behaviors do you want to capture your identity? Are there any misalignments between who you appear to be and who you want to be? What is the best representation of your desire to show up daily?
Characteristics, actions & Behaviors - Humble, open minded, innovative/imaginative, respectful, considerate, courageous Misalignments - Do more research and exploring of different practices and strategies and skills, network/partner more
Nora Ephron and The New York Observer: A Footnote<br /> by [[Foster Kamer]] in Observer <br /> accessed on 2026-05-11T14:37:21
Rosenbaum, Ron. 1999. “The Last Luddite Gets Wired.” Slate. https://slate.com/news-and-politics/1999/05/the-last-luddite-gets-wired-4.html (May 11, 2026).
I believe a case can be made–indeed a case has been made by others, by historians of Apple and hacker culture–that “Secrets of the Little Blue Box,” a story I wrote for Esquire back in 1971 about “phone phreaks” and the first computer hackers (it was only the second magazine story I’d ever had published) played a crucial role in the careers of the founders of Apple and of a legendary ur-hacker I made famous who went by the name of Captain Crunch.
But there’s something about the sharp staccato sound of my Olympia Report Deluxe–like successive volleys of rifle shots rather than the mouselike scribble-scrabble of the keyboard pad–that I feel reluctant to abandon.
neo-Nazi pinheads who, along with child pornographers and Bill Gates, seem to me to be the only unequivocal beneficiaries of wired culture.
The thing that always used to get to me was the way a certain kind of arrogant but aggressively mediocre writer would harangue me about how I had to get a computer because ever since he got one, “I’ve been able to turn out so much more,” as if this were an unmixed blessing the world would welcome.
For the first 10 years of my life as a writer, I was a deeply agonized, chronically blocked writer, and it was not until I devised a method of writing-by-rewriting on my Olympia Report Deluxe that I was able to become a productive–if still slow and labor-intensive–writer. It’s not just superstition that kept me from switching to a computer; I suppose I could adopt my rewriting-from-the-top method to a computer if I were to print out each successive draft and rewrite the next one on the screen, then print that out and start rewriting from the top again.
Actually, that typewriter, my Olympia Report Deluxe, or rather Deluxes (I have three of them because they’re no longer manufactured, and I need one for backup and one to cannibalize for parts) are the real reason I’ve rejected switching to a computer for so long.
You know that promo slogan for Alien: “In space, no one can hear you scream.” In cyberspace, no one can hear you doubt.
I’d made the mistake of adopting the moniker “Redhead,” and the supposedly philosophical participants were all over me like a cheap suit the moment I logged on, assuming, it finally dawned on me, that I was a different gender of Redhead.)
Blackbirds
explain who the Blackbirds were.
critical work has focused mostly on historical contextualization rather than reading posthumous textual portraits of Mills.
I'm not sure what is the difference between historical contextualization and reading posthumous portraits.
Shuffle Along
put title in Italics. Also have you introduced Shuffle Along and explained her role in it already? Like Du Bois, this important reference needs more contextualizing to explain its significance as the first Broadway musical with an all Black cast (rather than white performers in blackface) and Florence Mills' role in the production. Was it her breakout performance?
Scholars probe the edges of this absence, weaving threads across the gaping void to try to understand.
Why is this text larger?
The story map is rich with archival details, but I'm having a bit of trouble distinguishing your narrative from the primary and quoted sources. Might be more effective to have a consistent rule of your narrative being in regular paragraph form, and quotations from primary sources appearing in those flying boxes?
The fact that no archival footage remains of her voice has only deepened her enigma. Audiences tried to describe it,
I agree about the red curtain and think this text and the screen shots from newspapers would probably be more effective if just delivered, with the sentence above, in regular text/image format
Even in her lifetime, Mills was mythologized.
Very effective animated collage that rebuilds the mythology even as it immerses me in it.
Your Ultimate Domination Awaits
Pictures all over the place. Telegram has four entities, a few marked bots.
developmental refugees
Nixon, Rob. “Megadams, Monumental Modernity, and Developmental Refugees.” Slow Violence and the Environmentalism of the Poor, n.d.
specifically Black women
Dokshina, Darya, Sidonie K. Roque, Sydney Berry, et al. “Racial and Ethnic Differences in Emotional Reactions to the Flint Water Crisis among Michigan Women in Communities Outside of Flint.” Ethnicity & Disease 34, no. 3 (2024): 129–36. https://doi.org/10.18865/EthnDis-2023-58.
If you
David Fasenfest, A Neoliberal Response to an Urban Crisis: Emergency Management in Flint, MI, Critical Sociology 45, no. 1 (2019): 34, https://doi.org/10.1177/0896920517718039.
Water
Fennell, Catherine. “Are We All Flint?” Limn, no. 7 (July 2016): 21–21.
Feature RequestThis is an on-demand feature. Please fill out the form to get this feature activated on your account.
This section to be removed
ConclusionThrough the blending of both the flâneuse's internal thoughts as well as her perceptions of the external world, Mirrlees constructs a multi-sensory experience that invites readers into her interiority, allowing them to engage with the poem, as she does with Paris: in a meaningful and embodied way.
Though clear and coherent, this conclusion feels like a bit of a let down after such subtle, perceptive, and engaging analysis. I think you might say how, after (or as an extension of ) collapsing the distinction between flaneuse and writer, the poem animates the reader in their combined role, inviting us to experience an autonomous, subjective, mutlisensory interiority, in which the self is not objectified nor objectifies others, but experiences its own power and permeability. In this way, the poem reminds me a bit of Walt Whitman's Song of Myself, which concludes by hailing the reader, saying: if you want me, look under your boot soles. We become Whitman, walking the streets of the newly formed American nation in his shoes.
e visualization of the letters are being drawn by Mirrlees as the writer, rather than the wandering flâneuse, as they implicate the page
very intriguing and perceptive distinction.
While digital media cannot materially reproduce scent, Mirrlees’s language enables readers to use their imagination, as the flâneuse does throughout the poem, in order to vicariously experience the sensation, thereby “becoming-with” the poem itself.
Nicely done!
portrait of Paris.
Paris at a particular point in the 1920s, when there were still roosters! You'd be unlikely to hear that today!
