Reviewer #3 (Public Review):
Summary:
This study proposes visual homogeneity as a novel visual property that enables observers perform to several seemingly disparate visual tasks, such as finding an odd item, deciding if two items are same, or judging if an object is symmetric. In Exp 1, the reaction times on several objects were measured in human subjects. In Exp 2, visual homogeneity of each object was calculated based on the reaction time data. The visual homogeneity scores predicted reaction times. This value was also correlated with the BOLD signals in a specific region anterior to LO. Similar methods were used to analyze reaction time and fMRI data in a symmetry detection task. It is concluded that visual homogeneity is an important feature that enables observers to solve these two tasks.
Strengths:
(1) The writing is very clear. The presentation of the study is informative.<br />
(2) This study includes several behavioral and fMRI experiments. I appreciate the scientific rigor of the authors.
Weaknesses:
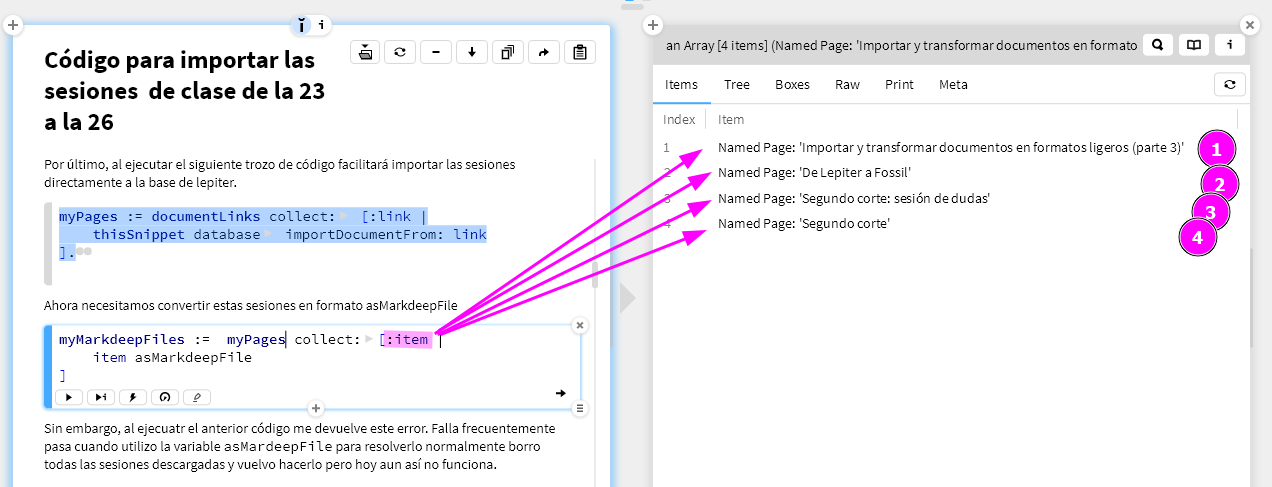
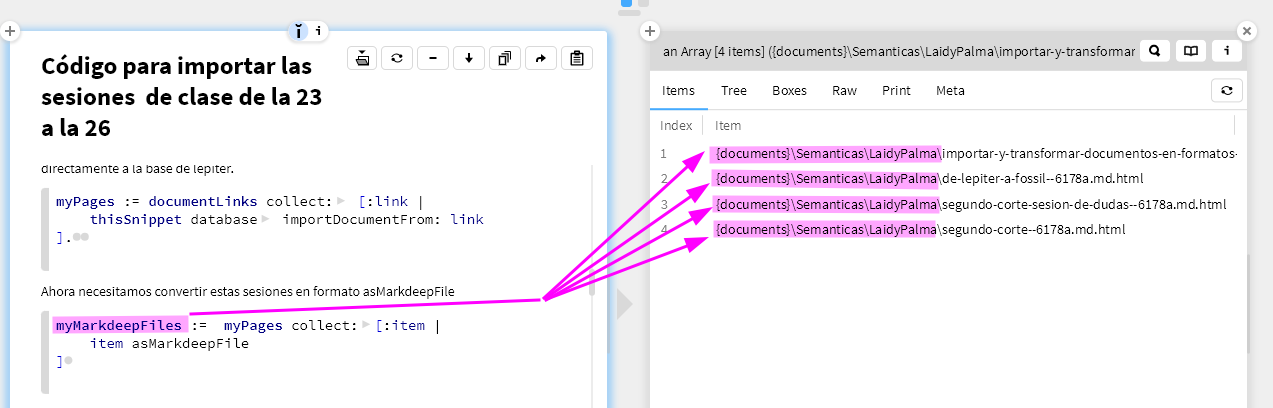
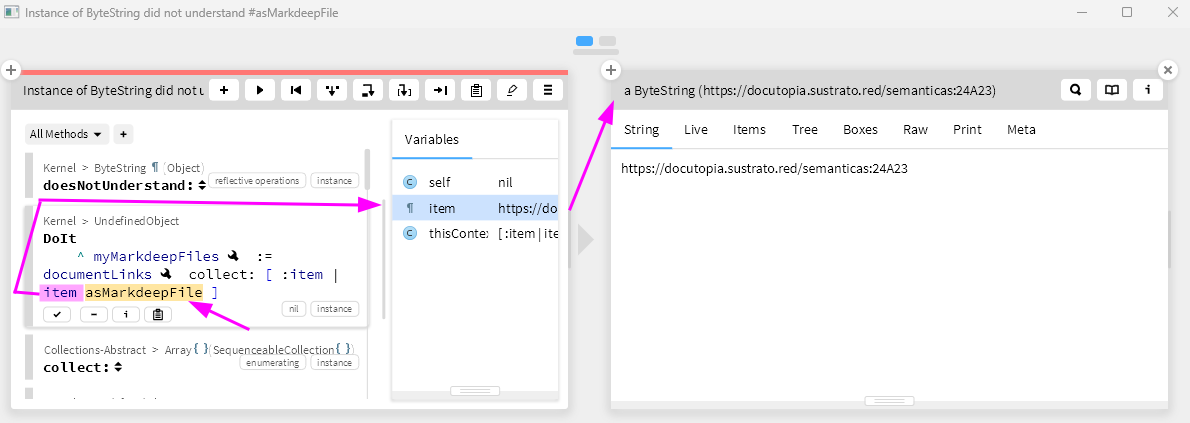
(1) My main concern with this paper is the way visual homogeneity is computed. On page 10, lines 188-192, it says: "we then asked if there is any point in this multidimensional representation such that distances from this point to the target-present and target-absent response vectors can accurately predict the target-present and target-absent response times with a positive and negative correlation respectively (see Methods)". This is also true for the symmetry detection task. If I understand correctly, the reference point in this perceptual space was found by deliberating satisfying the negative and positive correlations in response times. And then on page 10, lines 200-205, it shows that the positive and negative correlations actually exist. This logic is confusing. The positive and negative correlations emerge only because this method is optimized to do so. It seems more reasonable to identify the reference point of this perceptual space independently, without using the reaction time data. Otherwise, the inference process sounds circular. A simple way is to just use the mean point of all objects in Exp 1, without any optimization towards reaction time data.
(2) Visual homogeneity (at least given the current from) is an unnecessary term. It is similar to distractor heterogeneity/distractor variability/distractor statics in literature. However, the authors attempt to claim it as a novel concept. The title is "visual homogeneity computations in the brain enable solving generic visual tasks". The last sentence of the abstract is "a NOVEL IMAGE PROPERTY, visual homogeneity, is encoded in a localized brain region, to solve generic visual tasks". In the significance, it is mentioned that "we show that these tasks can be solved using a simple property WE DEFINE as visual homogeneity". If the authors agree that visual homogeneity is not new, I suggest a complete rewrite of the title, abstract, significance, and introduction.
(3) Also, "solving generic tasks" is another overstatement. The oddball search tasks, same-different tasks, and symmetric tasks are only a small subset of many visual tasks. Can this "quantitative model" solve motion direction judgment tasks, visual working memory tasks? Perhaps so, but at least this manuscript provides no such evidence. On line 291, it says "we have proposed that visual homogeneity can be used to solve any task that requires discriminating between homogeneous and heterogeneous displays". I think this is a good statement. A title that says "XXXX enable solving discrimination tasks with multi-component displays" is more acceptable. The phrase "generic tasks" is certainly an exaggeration.
(4) If I understand it correctly, one of the key findings of this paper is "the response times for target-present searches were positively correlated with visual homogeneity. By contrast, the response times for target-absent searches were negatively correlated with visual homogeneity" (lines 204-207). I think the authors have already acknowledged that the positive correlation is not surprising at all because it reflects the classic target-distractor similarity effect. But the authors claim that the negative correlations in target-absent searches is the true novel finding.
(5) I would like to make it clear that this negative correlation is not new either. The seminal paper by Duncan and Humphreys (1989) has clearly stated that "difficulty increases with increased similarity of targets to nontargets and decreased similarity between nontargets" (the sentence in their abstract). Here, "similarity between nontargets" is the same as the visual homogeneity defined here. Similar effects have been shown in Duncan (1989) and Nagy, Neriani, and Young (2005). See also the inconsistent results in Nagy& Thomas, 2003, Vicent, Baddeley, Troscianko&Gilchrist, 2009.<br />
More recently, Wei Ji Ma has systematically investigated the effects of heterogeneous distractors in visual search. I think the introduction part of Wei Ji Ma's paper (2020) provides a nice summary of this line of research.
I am surprised that these references are not mentioned at all in this manuscript (except Duncan and Humphreys, 1989).
(6) If the key contribution is the quantitative model, the study should be organized in a different way. Although the findings of positive and negative correlations are not novel, it is still good to propose new models to explain classic phenomena. I would like to mention the three studies by Wei Ji Ma (see below). In these studies, Bayesian observer models were established to account for trial-by-trial behavioral responses. These computational models can also account for the set-size effect, behavior in both localization and detection tasks. I see much more scientific rigor in their studies. Going back to the quantitative model in this paper, I am wondering whether the model can provide any qualitative prediction beyond the positive and negative correlations? Can the model make qualitative predictions that differ from those of Wei Ji's model? If not, can the authors show that the model can quantitatively better account for the data than existing Bayesian models? We should evaluate a model either qualitatively or quantitatively.
(7) In my opinion, one of the advantages of this study is the fMRI dataset, which is valuable because previous studies did not collect fMRI data. The key contribution may be the novel brain region associated with display heterogeneity. If this is the case, I would suggest using a more parametric way to measure this region. For example, one can use Gabor stimuli and systematically manipulate the variations of multiple Gabor stimuli, the same logic also applies to motion direction. If this study uses static Gabor, random dot motion, object images that span from low-level to high-level visual stimuli, and consistently shows that the stimulus heterogeneity is encoded in one brain region, I would say this finding is valuable. But this sounds like another experiment. In other words, it is insufficient to claim a new brain region given the current form of the manuscript.
REFERENCES<br />
- Duncan, J., & Humphreys, G. W. (1989). Visual search and stimulus similarity. Psychological Review, 96(3), 433-458. doi: 10.1037/0033-295x.96.3.433<br />
- Duncan, J. (1989). Boundary conditions on parallel processing in human vision. Perception, 18(4), 457-469. doi: 10.1068/p180457<br />
- Nagy, A. L., Neriani, K. E., & Young, T. L. (2005). Effects of target and distractor heterogeneity on search for a color target. Vision Research, 45(14), 1885-1899. doi: 10.1016/j.visres.2005.01.007<br />
- Nagy, A. L., & Thomas, G. (2003). Distractor heterogeneity, attention, and color in visual search. Vision Research, 43(14), 1541-1552. doi: 10.1016/s0042-6989(03)00234-7<br />
- Vincent, B., Baddeley, R., Troscianko, T., & Gilchrist, I. (2009). Optimal feature integration in visual search. Journal of Vision, 9(5), 15-15. doi: 10.1167/9.5.15<br />
- Singh, A., Mihali, A., Chou, W. C., & Ma, W. J. (2023). A Computational Approach to Search in Visual Working Memory.<br />
- Mihali, A., & Ma, W. J. (2020). The psychophysics of visual search with heterogeneous distractors. BioRxiv, 2020-08.<br />
- Calder-Travis, J., & Ma, W. J. (2020). Explaining the effects of distractor statistics in visual search. Journal of Vision, 20(13), 11-11.