Post #200 – The Corona 3 tripod<br /> by [[Scott K]] in The Filthy Platen on 2014-12-21<br /> accessed on 2025-12-30T15:29:02
7,363 Matching Annotations
- Dec 2025
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Clarke et al. investigate the role of subjective representations of task-based statistical structure on choice accuracy and reaction times during perceptual decision-making. Subjective representations of objective statistical structure are often overlooked in studies of predictive processing and, consequently, little is known about their role in predictive phenomena. By gauging the subjective experience of stimulus probability, expectedness, and surprise in tasks with fixed cue-stimulus contingencies, the authors aimed to separate subjective and objective (task-induced) contributions to predictive effects on behaviour.
Across three different experiments, subjective and objective contributions to predictions were found to explain unique portions of variance in reaction time data. In addition, choice accuracy was best predicted by subjective representations of statistical structure in isolation. These findings reveal that the subjective experience of statistical regularities may play a key role in the predictive processes that shape perception and cognition.
Strengths:
This study combines careful and thorough behavioral experimentation with an innovative focus on subjective experience in predictive processing. By collecting three independent datasets with different perceptual decision-making paradigms, the authors provide converging evidence that subjective representations of statistical structure explain unique variance in behavior beyond objective task structure. The analysis strategy, which directly contrasts the contributions of subjective and objective predictors, is conceptually rigorous and allows clear insight into how subjective and objective influences shape behavior. The methods are consistently applied across all three datasets and produce coherent results, lending strong support to the authors' conclusions. The study emphasizes the critical role of subjective experience in predictive processing, with implications for understanding learning, perception, and decision-making.
Weaknesses:
Despite these strengths, there are several conceptual and technical issues that should be addressed. In Experiments 2 and 3, the authors use a Rescorla-Wagner (RW) learning model to estimate trialwise expectedness (Experiment 2) and surprise (Experiment 3). While the RW model is a well-established model for explaining learning behaviour, it does not represent the objective 'ground truth' statistical structure of the environment, and treating RW trajectories as such imposes assumptions about learning that may not match participants' actual behavior. This assumption could strongly affect the comparison between subjective and 'objective' predictors. It would strengthen the primary conclusions of the manuscript if other implementations of the objective statistical structure, such as the true task-defined probabilities (i.e., 25% or 75%), were considered to provide a complementary 'ground truth' perspective.
Additionally, because objective statistical structure was predictive of subjective ratings in all three experiments, these predictors are likely collinear in the full model. Collinearity can lead to inflated standard errors and unstable coefficient estimates, even if the models converge. Currently, this potential critical problem of the applied statistical models is not assessed, reported on, or controlled for (e.g., by residualizing predictors). RW trajectories are also not reported in the manuscript, limiting the ability to assess how the model evolves over time and whether it maps onto the task-induced probabilities in a sensible way. This is particularly relevant because participants' subjective estimates of the task-induced probabilities seem to converge to the ground truth after just a few trials, especially for the 75% stimuli (Figure 3C).
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary
This study aims to overcome key limitations of single-cell RNA-seq in C. elegans neurons-especially the under-detection of lowly expressed and non-polyadenylated transcripts and residual contamination-by integrating bulk RNA-seq from FACS-isolated neuron types with an existing scRNA-seq atlas. The authors introduce LittleBites, an iterative, reference-guided decontamination algorithm that uses a single-cell reference together with ground-truth reporter datasets to optimize subtraction of contaminating signal from bulk profiles. They then generate an "Integrated" dataset that combines the sensitivity of bulk data with the specificity of scRNA-seq and use it to call neuron-specific expression for protein-coding genes, "rescued" genes not detected in scRNA-seq, and multiple classes of non-coding RNAs across 53 neuron classes. All data, code, and thresholded matrices are made publicly available to enable community reuse.
Strengths
(1) Conceptual advance and useful resource. The work demonstrates in a concrete way how bulk and single-cell datasets can be combined to overcome the weaknesses of each approach, and delivers a high-resolution transcriptomic resource for a substantial fraction of C. elegans neuron classes . The integrated matrices, thresholded expression calls, and non-coding RNA catalog will be useful both for basic neurobiology and for method developers.
(2) Careful benchmarking and transparency. The revised manuscript includes extensive benchmarking of LittleBites and the Integrated dataset against multiple independent "ground-truth" sets: neuron-specific reporter lines, curated non-neuronal markers, and ubiquitous genes. The authors evaluate AUROCs over a wide range of thresholds, explain ROC/AUROC metrics for non-specialists, and quantify how integration affects both sensitivity and specificity relative to scRNA-seq alone.
(3) Improved methodological clarity. In response to review, the authors now provide a much more intuitive description of the LittleBites algorithm, including a stepwise explanation of (1) contamination estimation via NNLS using single-cell references, (2) weighted subtraction tuned by a learning-rate parameter, and (3) performance optimization based on AUROC against ground-truth genes. this makes the approach accessible to readers who are not computational specialists and will facilitate re-implementation.
(4) Systematic analysis of reference dependence. The authors explicitly address the concern that LittleBites depends on the completeness and accuracy of the scRNA-seq reference. They examine how performance varies with cluster size and by simulated degradation of the reference (e.g., reducing the number of cells per cluster), and show that AUROCs remain robust, but that gene-level assignments are more variable for clusters represented by fewer cells. This is an important and honest characterization of when the method is reliable and when users should be cautious.
(5) Additional biological context. The manuscript now more clearly situates the dataset in the context of previous and ongoing work. In particular, the authors highlight that other groups have already used these bulk data to discover and validate cell-type-specific alternative splicing events, strengthening the case that the data are biologically meaningful beyond the immediate analyses presented here. The expanded analysis of non-coding RNAs and GPCR pseudogenes also adds biological interest.
(6) Improved handling and documentation of "unexpressed" genes. The authors have trimmed the original list of 4,440 genes called "unexpressed" in scRNA-seq to a higher-confidence subset and provide new supplementary tables that include gene identities and tissue annotations. They also use a curated set of non-neuronal markers to estimate residual contamination and show that most such markers are not detected in the integrated data, with only a small number of apparent false positives remaining.
Weaknesses
(1) Novel assignments remain predictive rather than experimentally validated. Although the authors have strengthened their benchmarking and refer to external work that validates some splicing patterns from these data, the large sets of newly assigned lowly expressed genes and non-coding RNAs-particularly those rescued from the "unexpressed" gene pool-are still inferred from computational criteria (thresholding plus correlation-based decontamination) rather than direct orthogonal assays (e.g., smFISH, in situ hybridization, or reporter lines). This is understandable given scale and cost, but it means that many of these calls should be interpreted as well-supported predictions, not definitive expression maps. The revised manuscript acknowledges this, and a dedicated "Limitations of this study" subsection will further clarify this point for readers.
(2) Reduced stability for neuron types with sparse single-cell representation. The authors' new analyses show that while integration improves overall correlation and AUROC across a wide range of neuron types, gene-level assignments are less stable for neuron classes represented by relatively few cells in the scRNA-seq reference. For such neuron types, both false negatives and false positives are more likely, and users should be cautious when interpreting cell-type-specific expression differences based solely on these calls.
(3) Residual contamination and misclassification are not completely eliminated. Despite the careful design of LittleBites and the additional correlation-based decontamination of "unexpressed" genes, the authors' benchmarking against curated non-neuronal markers shows that a small fraction of putative non-neuronal genes remains detectable even at stricter thresholds, and some bona fide neuronal genes are removed as likely contaminants. The new supplementary tables documenting "unexpressed" genes and their tissue annotations, together with explicit statements about residual error rates and the predictive nature of these classifications, help users to judge the reliability of specific genes, but they also underscore that the dataset is not a perfect ground truth.
(4) Scope and coverage remain incomplete. As the authors note, the dataset covers 53 neuron classes and does not fully represent all 302 neurons or all known neuron subtypes. In addition, bulk samples represent pools of neurons, and so the approach cannot resolve within-class heterogeneity or subtype-specific expression within those pools. These are inherent limitations of the current experimental design rather than flaws in the analysis, but they are important for readers to keep in mind when using the resource.
Overall, the revised manuscript presents solid evidence for the main methodological and resource claims, with clearly articulated limitations. The work is likely to have valuable impact on the C. elegans community and provides a template for integrating bulk and single-cell data in other systems.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The manuscript introduces a visual paradigm aimed at studying trans-saccadic memory.
The authors observe how memory of object location is selectively impaired across eye movements, whereas object colour memory is relatively immune to intervening eye movements.<br /> Results are reported for young and elderly healthy controls, as well as PD and AD participants.
A computational model is introduced to account for these results, indicating how early differences in memory encoding and decay (but not trans-saccadic updating per se) can account for the observed differences between healthy controls and clinical groups.
Strengths:
The data presented encompasses healthy and elderly controls, as well as clinical groups.
The authors introduce an interesting modelling strategy, aimed at isolating and identifying the main components behind the observed pattern of results.
Weaknesses:
The models tested differ in terms of the number of parameters. In general, a larger number of parameters leads to a better goodness of fit. It is not clear how the difference in the number of parameters between the models was taken into account.
It is not clear whether the modelling results could be influenced by overfitting (it is not clear how well the model can generalize to new observations).
Results specificity: it is not clear how specific the modelling results are with respect to constructional ability (measured via the Rey-Osterrieth Complex Figure test). As with any cognitive test, performance can also be influenced by general, non-specific abilities that contribute broadly to test success.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This study provides novel insights into how individuals regulate the speed of their movements both alone and in pairs, highlighting consistent differences in movement vigor across people and showing that these differences can adapt in dyadic contexts. The findings are significant because they reveal stable individual patterns of action that are flexible when interacting with others, and they suggest that multiple factors, beyond reward sensitivity, may contribute to these idiosyncrasies. The evidence is generally strong, supported by careful behavioral measurements and appropriate modeling, though clarifying some statistical choices and including additional measures of accuracy and smoothness would further strengthen the support for the conclusions.
Major Comments:
(1) Given the idiosyncrasies in individual vigor, would linear mixed models (LMMs) be more appropriate than ANOVAs in some analyses (e.g., in the section "Solo session"), as they can account for random intercepts and slopes on vigor measures? Some figures (e.g., Figure 2.B and 3.E) indeed seem to show that some aspects of behaviour may present variability in slopes and intercepts across participants. In fact, I now realize that LMMs are used in the "Emergence of dyadic vigor from the partners' individual vigor" section, so could the authors clarify why different statistical approaches were applied depending on the sections?
(2) If I understand correctly, the introduction suggests that idiosyncrasies in movement vigor may be driven by inter-individual differences in reward sensitivity. However, the current task does not involve any explicit rewards, yet the authors still observe idiosyncrasies in vigor, which is interesting. Could this indicate that other factors contribute to these consistent individual differences? For example, could sensitivity to temporal costs or physical effort explain the slow versus fast subgrouping? Specifically, might individuals more sensitive to temporal costs move faster to minimize opportunity costs, and might those less sensitive to effort costs also move faster? Along the same lines, could the two subgroups (slow vs. fast) be characterized in terms of underlying computational "phenotypes," such as their sensitivities to time and effort? If this is not feasible with the current dataset, it would still be valuable to discuss whether these factors could plausibly account for the observed patterns, based on existing literature.
(3) The observation that dyads did not lose accuracy or smoothness despite changes in vigor is interesting and suggests a shift in the speed-accuracy tradeoff. Could the authors include accuracy and smoothness measures in the main figures rather than only in supplementary materials? I think it would make the manuscript more complete.
(4) It is a bit unclear to me whether the variance assumptions for ANOVAs were checked, for instance, in Figure 3H.
-
-
arxiv.org arxiv.org
-
Reviewer #3 (Public review):
Summary
Boccato et al. present an ambitious and thoughtfully developed framework, SynaptoGen, which proposes a differentiable model of synaptogenesis grounded in gene-expression vectors, protein interaction probabilities, and conductance rules. The authors aim to bridge the gap between computational connectomics and synthetic biological intelligence by enabling gradient-based optimization of genetically encoded circuit architectures. They support this goal with mathematical derivations, simulation experiments across several RL benchmarks, and a biologically grounded validation using C. elegans adhesion-molecule co-expression data. The paper is timely and conceptually compelling, offering a unified formulation of synaptic multiplicity and synaptic weight formation that can be integrated directly into learning systems.
Strengths
(1) Well-motivated framework with clear conceptual contributions.
(2) Rigorous mathematical development.
(3) Compelling empirical validation.
(4) Excellent framing and discussion of future impact.
Weaknesses
(1) Overstated claims in the abstract and discussion.
(2) Ambiguity in "first of its kind" assertions.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Guy et al. explored the variation in the pathogenicity of carboxy-terminal frameshift deletions in the X-linked MECP2 gene. Loss-of-function variants in MECP2 are associated with Rett syndrome, a severe neurodevelopmental disorder. Although 100's of pathogenic MECP2 variants have been found in people with Rett syndrome, 8 recurrent point mutations are found in ~65% of disease cases, and frameshift insertions/deletions (indels) variants resulting in production of carboxy-terminal truncated (CTT) MeCP2 protein account for ~10% of cases. Many of these occur in a "deletion prone region" (DPR) between c.1110-1210, with common recurrent deletions c.1157-1197del (CTD1) and c.1164_1207del (CTD2). While two major protein functional domains have been defined in MeCP2, the methyl-binding domain (MBD) and the NCoR interacting domain (NID), the functional role of the carboxy-terminal domain (CTD, beyond the NID, predicted to have a disordered protein structure) has not been identified, and previous work by this group and others demonstrated that a Mecp2 "minigene" lacking the CTD retains MeCP2 function suggesting that the CTD is dispensable. This raises an important question: If the CTD is dispensable, what is the pathogenic basis of the various CTT frameshift variants? Prior work from this group demonstrated that genetically engineered mice expressing the CTD1 variant had decreased expression of Mecp2 RNA and MeCP2 protein and decreased survival, but those expressing the CTD2 variant had normal Mecp2 RNA and protein and survival. However, they noted that differences between the mouse and human coding sequences resulted in different terminal sequences between the two common CTD, with CTD1 ending in -PPX in both mouse and human, but CTD2 ending in -PPC in human but -SPX in mouse, and in the previous paper they demonstrated in humanized mouse ES cells (edited to have the same -PPX termination) containing the CTD2 deletion resulted in decreased Mecp2 RNA and protein levels. This previous work provides the underlying hypotheses that they sought to explore, which is that the pathological basis of disease causing CTD relates to the formation of truncated proteins that end with a specific amino acid sequence (-PPX), which leads to decreased mRNA and protein levels, whereas tolerated, non-pathogenic CTD do not lead to production of truncated proteins ending in this sequence and retain normal mRNA/protein expression.
In this manuscript, they evaluate missense variants, in-frame deletions, and frame shift deletions within the DPR from the aggregated Genome Aggregated Database (gnomAD) and find that the "apparently" normal individuals within gnomAD had numerous tolerated missense variants and in-frame deletions within this region, as well as frameshift deletions (in hemizygous males) in the defined region. All of the gnomAD deletions within this region resulted in terminal amino acid sequences -SPRTX (due to +1 frameshift), whereas nearly all deletion variants in this region from people with Rett syndrome (from the Clinvar copy of the former RettBase database) had a terminal -PPX sequence, due to a +2 frameshift. They hypothesized that terminal proline codons causing ribosomal stalling and "nonsense mediated decay like" degradation of mRNA (with subsequent decreased protein expression) was the basis of the specific pathogenicity of the +2 frameshift variants, and that utilizing adenine base editors (ABE) to convert the termination codon to a tryptophan could correct this issue. They demonstrate this by engineering the change into mouse embryonic stem cell lines and mouse lines containing the CTD1 deletion and show that this change normalized Mecp2 mRNA and protein levels and mouse phenotypes. Finally, they performed an initial proof-of-concept in an inducible HEK cell line and showed the ability of targeted ABE to edit the correct adenine and cause production of the expected larger truncated Mecp2 protein from CTD1 constructs.
The findings of this manuscript provide a level of support for their hypothesis about the pathogenicity versus non-pathogenicity of some MECP2 CTT intragenic deletions and provide preliminary evidence for a novel therapeutic approach for Rett syndrome; however, limitations in their analysis do not fully support the broader conclusions presented.
Strengths:
(1) Utilization of publicly available databases containing aggregated genetic sequencing data from adult cohorts (gnomAD) and people with Rett syndrome (Clinvar copy of RettBase) to compare differences in the composition of the resulting terminal amino acid sequences resulting from deletions presumed to be pathogenic (n+2) versus presumed to be tolerated (n+1).
(2) Evaluation of a unique human pedigree containing an n+1 deletion in this region that was reported as pathogenic, with demonstration of inheritance of this from the unaffected father and presence within other unaffected family members.
(3) Development of a novel engineered mouse model of a previously assumed n+1 pathogenic variant to demonstrate lack of detrimental effect, supporting that this is likely a benign variant and not causative of Rett syndrome.
(4) Creation and evaluation of novel cell lines and mouse models to test the hypothesis that the pathogenicity of the n+2 deletion variants could be altered by a single base change in the frameshifted stop codon.
(5) Initial proof-of-concept experiments demonstrating the potential of ABE to correct the pathogenicity of these n+2 deletion variants.
Weaknesses:
(1) While the use of the large aggregated gnomAD genetic data benefits from the overall size of the data, the presence of genetic variants within this collection does not inherently mean that they are "neutral" or benign. While gnomAD does not include children, it does include aggregated data from a variety of projects targeting neuropsychiatric (and other conditions), so there is information in gnomAD from people with various medical/neuropsychiatric conditions. The authors do make some acknowledgement of this and argue that the presence of intragenic deletion variants in their region of interest in hemizygous males indicates that it is highly likely that these are tolerated, non-pathogenic variants. Broadly, it is likely true that gnomAD MECP2 variants found in hemizygous males are unlikely to cause Rett syndrome in heterozygous females, it does not necessarily mean that these variants have no potential to cause other, milder, neuropsychiatric disorders. As a clear example, within gnomAD, there is a hemizygous male with the rs28934908 C>T variant that results in p.A140V (p.A152V in e1 transcript numbering convention). This pathogenic variant has been found in a number of pedigrees with an X-linked intellectual disability pattern, in which males have a clear neurodevelopmental disorder and heterozygous females have mild intellectual disability (see PMIDs 12325019, 24328834 as representative examples of a large number of publications describing this). Thus, while their claim that hemizygous deletion variants in gnomAD are unlikely to cause Rett syndrome, that cannot make the definitive statement that they are not pathogenic and completely benign, especially when only found in a very small number of individuals in gnomAD.
(2) The authors focus exclusively on deletions within the "DPR", they define as between c.1110-1210 and say that these deletions account for 10% of Rett syndrome cases. However, the published studies that are the basis for this 10% estimate include all genetic variants (frameshift deletions, insertions, complex insertion/deletions, nonsense variants) resulting in truncations beyond the NID. For example, Bebbington 2010 (PMID: 19914908), which includes frameshift indels as early as c.905 and beyond c.1210. Further specific examples from RettBase are described below, but the important point is that their evaluation of only frameshift variants within c.1110-1210 is not truly representative of the totality of genetic variants that collectively are considered CTT and account for 10% of Rett cases.
(3) The authors say that they evaluated the putative pathogenic variants contained within RettBase (which is no longer available, but the data were transferred to Clinvar) for all cases with Classic Rett syndrome and de novo deletion variants within their defined DPR domain. Looking at the data from the Clinvar copy of RettBase, there are a number (n=143) of c-terminal truncating variants (either frameshift or nonsense) present beyond the NID, but the authors only discuss 14 deletion frameshift variants in this manuscript. A number of these variants have molecular features that do not fall into the pathogenic classification proposed by the authors and are not addressed in the manuscript and do not support the generalization of the conclusions presented in this manuscript, especially the conclusion that the determination of pathogenicity of all c-terminal truncating variants can be determined according to their proposed n+2 rule, or that all of the 10% of people with Rett syndrome and c-terminal truncating variants could be treated by using a base editor to correct the -PPX termination codon.
(4) The HEK-based system utilized is convenient for doing the initial experiments testing ABE; however, it represents an artificial system expressing cDNA without splicing. Canonical NMD is dependent on splicing, and while non-canonical "NMD-like" processes are less well understood, a concern is whether the artificial system used can adequately predict efficacy in a native setting that includes introns and splicing.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This paper reports on an association between body size and the occurrence of species in cities, which is quantified using an 'urban score' that can be visualized as a 'Species Urbanness Distribution' for particular taxa. The authors use species records from the Global Biodiversity Information Facility (GBIF) and link the occurrence data to nighttime lighting quantified using satellite data (Visible Infrared Imaging Radiometer Suite-VIIRS). They link the urban score to body size data to find 'heterogeneous relationship between body size and urban tolerance across the tree'. The results are then discussed with reference to potential mechanisms that could possibly produce the observed effects (cf. Figure 1).
Strengths:
The novelty of this study lies in the huge number of species analyzed and the comparison of results among animal taxa, rather than in a thorough analysis of what traits allow species to persist under urban conditions. Such analyses have been done using a much more thorough approach that employs presence-absence data as well as a suite of traits by other studies, for example, in (Hahs et al. 2023, Neate-Clegg et al. 2023). The dataset that the authors produced would also be very valuable if these raw data were published, both the cleaned species records as well as the body sizes.
The paper could strongly add to our understanding of what species occur in cities when the open questions are addressed.
Weaknesses:
I value the approach of the authors, but I think the paper needs to be revised.
In my view, the authors could more carefully validate their approach. Currently, any weakness or biases in the approach are quickly explained away rather than carefully explored. This concerns particularly the use of presence-only data, but also the calculation of the urban score.
The vast majority of data in GBIF is presence-only data. This produces a strong bias in the analysis presented in the paper. For some taxa, it is likely that occurrences within the city are overrepresented, and for other taxa, the opposite is true (cf. Sweet et al. 2022). I think the authors should try to address this problem.
The authors should compare their results to studies focusing on particular taxa where extensive trait-based analyses have already been performed, i.e., plants and birds. In fact, I strongly suggest that the authors should compare their results to previous studies on the relationship between traits, including body size and occurrences along a gradient of urbanisation, to draw conclusions about the validity of the approach used in the current study, which has a number of weaknesses.
They should be be more careful in coming up with post-hoc explanations of why the pattern found in this study makes sense or suggests a particular mechanism. This reviewer considers that there is no way in which the current study can disentangle the different possible mechanisms without further analyses and data, so I would suggest pointing out carefully how the mechanisms could be studied
More details should be given about the methodology. The readers should be able to understand the methods without having to read a number of other papers.
References:
Hahs, A. K., B. Fournier, M. F. Aronson, C. H. Nilon, A. Herrera-Montes, A. B. Salisbury, C. G. Threlfall, C. C. Rega-Brodsky, C. A. Lepczyk, and F. A. La Sorte. 2023. Urbanisation generates multiple trait syndromes for terrestrial animal taxa worldwide. Nature Communications 14:4751.
Neate-Clegg, M. H. C., B. A. Tonelli, C. Youngflesh, J. X. Wu, G. A. Montgomery, Ç. H. Şekercioğlu, and M. W. Tingley. 2023. Traits shaping urban tolerance in birds differ around the world. Current Biology 33:1677-1688.
Sweet, F. S. T., B. Apfelbeck, M. Hanusch, C. Garland Monteagudo, and W. W. Weisser. 2022. Data from public and governmental databases show that a large proportion of the regional animal species pool occur in cities in Germany. Journal of Urban Ecology 8:juac002.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The manuscript by Qui et al. explores the issue of spatial learning in both local (rooms) and global (connected rooms) environments. The authors perform a pointing task, which involves either pressing the right or left button in the scanner to indicate where an object is located relative to another object. Participants are repeatedly exposed to rooms over sessions of learning, with one "pre" and one "post" learning session. The authors report that the hippocampus shifted from lower to higher RSA for the global but not the local environment after learning. RSC and OFC showed higher RSA for global object pointing. Other brain regions also showed effects, including ACC, which seemed to show a similar pattern as the hippocampus, as well as other regions shown in Figure S5. The authors attempt to tie their results in with local vs. global spatial representations.
Strengths:
Extensive testing of subjects before and after learning a spatial environment, with data suggesting that there may be fMRI codes sensitive to both global and local codes. Behavioral data suggest that subjects are performing well at the task and learning both global and local object locations, although see further comments.
Weaknesses:
(1) The authors frame the entire introduction around confirming the presence of the cognitive map either locally or globally. There are some significant issues with this framing. For one, the introduction appears to be confirmatory and not testing specific hypotheses that can be falsified. What exactly are the hypotheses being tested? I believe that this relates to the testing whether neural representations are global and/or local. However, this is not clear. Given that a previous paper (Marchette et al. 2014 Nature Neuro, which bears many similarities) showed only local coding in RSC, this paper needs to be discussed in far more depth in terms of its similarities and differences. This paper looked at both position and direction, while the current paper looks at direction. Even here, direction in the current study is somewhat impoverished: it involves either pointing right or left to an object, and much of this could be categorical or even lucky guesses. From what I could tell, all behavioral inferences are based on reaction time and not accuracy, and therefore, it is difficult to determine if the subject's behavior actually reflects knowledge gained or simply faster reaction time, either due to motor learning or a speed-accuracy trade-off. The pointing task is largely egocentric: it can be solved by remembering a facing direction and an object relative to that. It is not the JRD task as has been used in other studies (e.g., Huffman et al. 2019 Neuron), which is continuous and has an allocentric component. This "version" of the task would be largely egocentric. In this way, the pointing task used does not test the core tenets of the cognitive map during navigation, which is defined as allocentric and Euclidean (please see O'Keefe and Nadel 1978, The Hippocampus as a Cognitive Map). Since neither of these assumptions appears valid, the paper should be reframed to reflect spatial representations more broadly or even egocentric spatial representations.
(2) The fMRI data workup is insufficient. What do the authors mean by "deactivations" in Figure 3b? Does this mean the object task showed more activation than the spatial task in HSC? Given that HSC is one of these regions, this would seem to suggest that the hippocampus is more involved in object than spatial processing, although it is difficult to tell from how things are written. The RSA is more helpful, but now a concern is that the analysis focuses on small clusters that are based on analyses determined previously. This appears to be the case for the correlations shown in Figure 3e as well. The issues here are several-fold. For one, it has been shown in previous work that basing secondary analyses on related first analyses can inflate the risk of false positives (i.e., Kriegeskorte et al. 2009 Nature Neuro). The authors should perform secondary analyses in ways that are unbiased by the first analyses, preferably, selecting cluster centers (if they choose to go this route) from previous papers rather than their own analyses. Another option would be to perform analyses at the level of the entire ROI, meaning that the results would generalize more readily. The authors should also perform permutation tests to ensure that the RSA results are reliable, as these can run the risk of false positives (e.g., Nolan et al. 2018 eNeuro). If these results hold, the authors should perform post-hoc (corrected) t-tests for global vs. local before and after learning to ensure these differences are robust and not simply rely on the interaction effect. The figures were difficult to follow in this regard, and an interaction effect does not necessarily mean the differences that are critical (global higher than local after) are necessarily significant. The end part of the results was hard to follow. If ACC showed a similar effect to HC and RSC, why is it not being considered? Many other areas that seemed to show local vs. global effects were dismissed, but these should instead be discussed in terms of whether they are consistent or inconsistent with the hypotheses.
(3) Concerns about the discussion: there are areas involving reverse inference about brain areas rather than connecting the findings with hypotheses (see Poldrack et al. 2006 Trends in Cognitive Science). The authors also argue for 'transfer" of information (for example, from ACC to OFC), but did not perform any connectivity analyses, so these conclusions are not based on any results. Instead, the authors should carefully compare what can be concluded from the reaction time findings and the fMRI data. What is consistent vs. inconsistent with the hypotheses? The authors should also provide a much more detailed comparison with past work. The Marchette et al. paper comes to different conclusions regarding RSC and involves more detailed analyses than those done here, including position. What is different in the current paper that might explain the differences in results? Another previous paper that came to a different conclusion (hippocampus local, retrosplenial global) and should be carefully considered and compared, as it also involved learning of environments and comparisons at different phases (e.g., Wolbers & Buchel 2005 J Neuro). Other papers that have used the JRD task have demonstrated similar, although not identical, networks (e.g., Huffman et al. 2019 Neuron) and the results here should be more carefully compared, as the current task is largely egocentric while the Huffman et al. paper involves a continuous and allocentric version of the JRD task.
(4) The authors cite rodent papers involving single neuron recordings. These are quite different experiments, however: they involve rodents, the rodents are freely moving, and single neurons are recorded. Here, the study involves humans who are supine and an indirect vascular measure of neural activity. Citations should be to studies of spatial memory and navigation in humans using fMRI: over-reliance on rodent studies should be avoided for the reasons mentioned above.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Overall, this is a well-done study, and the conclusions are largely supported by the data, which will be of interest to the field.
Strengths:
Strengths of this study include experiments with solution NMR that can resolve high-resolution interactions of the highly flexible C-terminal tail of arr2 with clathrin and AP2. Although mainly confirmatory in defining the arr2 CBL 376LIELD380 as the clathrin binding site, the use of the NMR is of high interest (Fig. 1). The 15N-labeled CLTC-NTD experiment with arr2 titrations reveals a span from 39-108 that mediates an arr2 interaction, which corroborates previous crystal data, but does not reveal a second area in CLTC-NTD that in previous crystal structures was observed to interact with arr2.
SEC and NMR data suggest that full-length arr2 (1-418) binding with 2-adaptin subunit of AP2 is enhanced in the presence of CCR5 phospho-peptides (Fig. 3). The pp6 peptide shows the highest degree of arr2 activation, and 2-adaptin binding, compared to less phosphorylated peptide or not phosphorylated at all. It is interesting that the arr2 interaction with CLTC NTD and pp6 cannot be detected using the SEC approach, further suggesting that clathrin binding is not dependent on arrestin activation. Overall, the data suggest that receptor activation promotes arrestin binding to AP2, not clathrin, suggesting the AP2 interaction is necessary for CCR5 endocytosis.
To validate the solid biophysical data, the authors pursue validation experiments in a HeLa cell model by confocal microscopy. This requires transient transfection of tagged receptor (CCR5-Flag) and arr2 (arr2-YFP). CCR5 displays a "class B"-like behavior in that arr2 is rapidly recruited to the receptor at the plasma membrane upon agonist activation, which forms a stable complex that internalizes onto endosomes (Fig. 4). The data suggest that complex internalization is dependent on AP2 binding not clathrin (Fig. 5).
The addition of the antagonist experiment/data adds rigor to the study.
Overall, this is a solid study that will be of interest to the field.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This work presents a novel neural network-based framework for parameterizing individual differences in human behavior. Using two distinct decision-making experiments, the author demonstrates the approach's potential and claims it can predict individual behavior (1) within the same task, (2) across different tasks, and (3) across individuals. While the goal of capturing individual variability is compelling and the potential applications are promising, the claims are weakly supported, and I find that the underlying problem is conceptually ill-defined.
Strengths:
The idea of using neural networks for parameterizing individual differences in human behavior is novel, and the potential applications can be impactful.
Weaknesses:
(1) To demonstrate the effectiveness of the approach, the authors compare a Q-learning cognitive model (for the MDP task) and RTNet (for the MNIST task) against the proposed framework. However, as I understand it, neither the cognitive model nor RTNet is designed to fit or account for individual variability. If that is the case, it is unclear why these models serve as appropriate baselines. Isn't it expected that a model explicitly fitted to individual data would outperform models that do not? If so, does the observed superiority of the proposed framework simply reflect the unsurprising benefit of fitting individual variability? I think the authors should either clarify why these models constitute fair control or validate the proposed approach against stronger and more appropriate baselines.
(2) It's not very clear in the results section what it means by having a shorter within-individual distance than between-individual distances. Related to the comment above, is there any control analysis performed for this? Also, this analysis appears to have nothing to do with predicting individual behavior. Is this evidence toward successfully parameterizing individual differences? Could this be task-dependent, especially since the transfer is evaluated on exceedingly similar tasks in both experiments? I think a bit more discussion of the motivation and implications of these results will help the reader in making sense of this analysis.
(3) The authors have to better define what exactly he meant by transferring across different "tasks" and testing the framework in "more distinctive tasks". All presented evidence, taken at face value, demonstrated transferring across different "conditions" of the same task within the same experiment. It is unclear to me how generalizable the framework will be when applied to different tasks.
(4) Conceptually, it is also unclear to me how plausible it is that the framework could generalize across tasks spanning multiple cognitive domains (if that's what is meant by more distinctive). For instance, how can an individual's task performance on a Posner task predict task performance on the Cambridge face memory test? Which part of the framework could have enabled such a cross-domain prediction of task performance? I think these have to be at least discussed to some extent, since without it the future direction is meaningless.
(5) How is the negative log-likelihood, which seems to be the main metric for comparison, computed? Is this based on trial-by-trial response prediction or probability of responses, as what usually performed in cognitive modelling?
(6) None of the presented evidence is cross-validated. The authors should consider performing K-fold cross-validation on the train, test, and evaluation split of subjects to ensure robustness of the findings.
(7) The authors excluded 25 subjects (20% of the data) for different reasons. This is a substantial proportion, especially by the standards of what is typically observed in behavioral experiments. The authors should provide a clear justification for these exclusion criteria and, if possible, cite relevant studies that support the use of such stringent thresholds.
(8) The authors should do a better job of creating the figures and writing the figure captions. It is unclear which specific claim the authors are addressing with the figure. For example, what is the key message of Figure 2C regarding transfer within and across participants? Why are the stats presentation different between the Cognitive model and the EIDT framework plots? In Figure 3, it's unclear what these dots and clusters represent and how they support the authors' claim that the same individual forms clusters. And isn't this experiment have 98 subjects after exclusion, this plot has way less than 98 dots as far as I can tell. Furthermore, I find Figure 5 particularly confusing, as the underlying claim it is meant to illustrate is unclear. Clearer figures and more informative captions are needed to guide the reader effectively.
(9) I also find the writing somewhat difficult to follow. The subheadings are confusing, and it's often unclear which specific claim the authors are addressing. The presentation of results feels disorganized, making it hard to trace the evidence supporting each claim. Also, the excessive use of acronyms (e.g., SX, SY, CG, EA, ES, DA, DS) makes the text harder to parse. I recommend restructuring the results section to be clearer and significantly reducing the use of unnecessary acronyms.
Comments on revisions:
The authors have addressed my previous comments with great care and detail. I appreciate the additional analyses and edits. I have no further comments.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors use calcium recordings from STN to measure STN activity during spontaneous movement and in a multi-stage avoidance paradigm. They also use optogenetic inhibition and lesion approaches to test the role of STN during the avoidance paradigm. The paper reports a large amount of data and makes many claims, some seem well supported to this Reviewer, others not so much.
Strengths:
Well-supported claims include data showing that during spontaneous movements, especially contraversive ones, STN calcium activity is increased using bulk photometry measurements. Single-cell measures back this claim but also show that it is only a minority of STN cells that respond strongly, with most showing no response during movement, and a similar number showing smaller inhibitions during movement.
Photometry data during cued active avoidance procedures show that STN calcium activity sharply increases in response to auditory cues, and during cued movements to avoid a footshock. Optogenetic and lesion experiments are consistent with an important role for STN in generating cue-evoked avoidance. And a strength of these results is that multiple approaches were used.
Original Weaknesses:
I found the experimental design and presentation convoluted and some of the results over-interpreted.
As presented, I don't understand this idea that delayed movement is necessarily indicative of cautious movements. Is the distribution of responses multi-modal in a way that might support this idea; or do the authors simply take a normal distribution and assert that the slower responses represent 'caution'? Even if responses are multi-modal and clearly distinguished by 'type', why should readers think this that delayed responses imply cautious responding instead of say: habituation or sensitization to cue/shock, variability in attention, motivation, or stress; or merely uncertainty which seems plausible given what I understand of the task design where the same mice are repeatedly tested in changing conditions. This relates to a major claim (i.e., in the title).
Related to the last, I'm struggling to understand the rationale for dividing cells into 'types' based the their physiological responses in some experiments.
In several figures the number of subjects used was not described. This is necessary. Also necessary is some assessment of the variability across subjects. The only measure of error shown in many figures relates trial-to-trial or event variability, which is minimal because in many cases it appears that hundreds of trials may have been averaged per animal, but this doesn't provide a strong view of biological variability (i.e., are results consistent across animals?).
It is not clear if or how spread of expression outside of target STN was evaluated, and if or how or how many mice were excluded due to spread or fiber placements. Inadequate histological validation is presented and neighboring regions that would be difficult to completely avoid, such as paraSTN may be contributing to some of the effects.
Raw example traces are not provided.
The timeline of the spontaneous movement and avoidance sessions were not clear, nor the number of events or sessions per animal and how this was set. It is not clear if there was pre-training or habituation, if many or variable sessions were combined per animal, or what the time gaps between sessions was, or if or how any of these parameters might influence interpretation of the results.
Comments on revised version:
The authors removed the optogenetic stimulation experiments, but then also added a lot of new analyses. Overall the scope of their conclusions are essentially unchanged.
Part of the eLife model is to leave it to the authors discretion how they choose to present their work. But my overall view of it is unchanged. There are elements that I found clear, well executed, and compelling. But other elements that I found difficult to understand and where I could not follow or concur with their conclusions.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This is an impressive paper that offers a much-needed 3D standardized brain atlas for the hackled-orb weaving spider Uloborus diversus, an emerging organism of study in neuroethology. The authors used a detailed immunohistological wholemount staining method that allowed them to localize a wide range of common neurotransmitters and neuropeptides and map them on a common brain atlas. Through this approach, they discovered groups of cells that may form parts of neuropils that had not previously been described, such as the 'tonsillar neuropil', which might be part of a larger insect-like central complex. Further, this work provides unique insights into previously underappreciated complexity of higher-order neuropils in spiders, particularly the arcuate body, and hints at a potentially important role for the mushroom bodies in vibratory processing for web-building spiders.
Strengths:
To understand brain function, data from many experiments on brain structure must be compiled to serve as a reference and foundation for future work. As demonstrated by the overwhelming success in genetically tractable laboratory animals, 3D standardized brain atlases are invaluable tools-especially as increasing amounts of data are obtained at the gross morphological, synaptic, and genetic levels, and as functional data from electrophysiology and imaging are integrated. Among 'non-model' organisms, such approaches have included global silver staining and confocal microscopy, MRI, and more recently, micro-computed tomography (X-ray) scans used to image multiple brains and average them into a composite reference. In this study, the authors used synapsin immunoreactivity to generate an averaged spider brain as a scaffold for mapping immunoreactivity to other neuromodulators. Using this framework, they describe many previously known spider brain structures and also identify some previously undescribed regions. They argue that the arcuate body-a midline neuropil thought to have diverged evolutionarily from the insect central complex-shows structural similarities that may support its role in path integration and navigation.
Having diverged from insects such as the fruit fly Drosophila melanogaster over 400 million years ago, spiders are an important group for study-particularly due to their elegant web-building behavior, which is thought to have contributed to their remarkable evolutionary success. How such exquisitely complex behavior is supported by a relatively small brain remains unclear. A rich tradition of spider neuroanatomy emerged in the previous century through the work of comparative zoologists, who used reduced silver and Golgi stains to reveal remarkable detail about gross neuroanatomy. Yet, these techniques cannot uncover the brain's neurochemical landscape, highlighting the need for more modern approaches-such as those employed in the present study.
A key insight from this study involves two prominent higher-order neuropils of the protocerebrum: the arcuate body and the mushroom bodies. The authors show that the arcuate body has a more complex structure and lamination than previously recognized, suggesting it is insect central complex-like and may support functions such as path integration and navigation, which are critical during web building. They also report strong synapsin immunoreactivity in the mushroom bodies and speculate that these structures contribute to vibratory processing during sensory feedback, particularly in the context of web building and prey localization. These findings align with prior work that noted the complex architecture of both neuropils in spiders and their resemblance (and in some cases greater complexity) compared to their insect counterparts. Additionally, the authors describe previously unrecognized neuropils, such as the 'tonsillar neuropil,' whose function remains unknown but may belong to a larger central complex. The diverse patterns of neuromodulator immunoreactivity further suggest that plasticity plays a substantial role in central circuits.
Weaknesses:
My major concern, however, is some of the authors' neuroanatomical descriptions rely too heavily on inference rather than what is currently resolvable from their immunohistochemistry stains alone.
Comments on revisions:
I thought that the authors did an excellent job responding to the reviews, and I have no further comments.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this study, Ho et al. hypothesised that autoreactive T cells receiving enhanced TCR signals during positive selection in the thymus are primed for generating effector and memory T cells. They used CD5 as a marker for TCR signal strength during their selection at the double positive stage. Supporting their hypothesis, naïve T cells with high CD5 proliferated better and expressed markers of T cell activation compared to naïve T cells with lower levels of CD5. Furthermore, results showed that autoimmune diabetes can be efficiently induced after the transfer of naïve CD5 hi T cells compared to CD5 lo T cells. This provided solid evidence in support of their hypothesis that T cells receiving higher basal TCR signaling are primmed to develop into effector T cells. However, all functional characterisation was done on the cells in the periphery and CD5 hi cells in the peripheral lymphoid compartment can receive tonic TCR signaling. Hence, the function of CD5 hi T cells might not be related to development and programming in the thymus. This is a major hurdle in the interpretation of the results and justifying the title of the study. The evidence that transgenic PTPN22 expression could not regulate T cell activation in CD5 hi TCR transgenic autoreactive T cells was weak. Studying T cell development in TCR transgenic mice and looking at TCR downstream signaling could be misleading due to transgenic expression of TCR at all developmental stages.
Strengths:
(1) Demonstrating that CD5 hi cells in naïve CD8 T cell compartment express markers of T cell activation, proliferation and cytotoxicity at a higher level
(2) Using gene expression analysis, study showed CD5 hi cells among naïve CD8 T cells are transcriptionally poised to develop into effector or memory T cells.
(3) Study showed that CD5 hi cells have higher basal TCR signaling compared to CD5 lo T cells.
(4) Key evidence of pathogenicity of autoreactive CD5 hi T cells was provided by doing the adoptive transfer of CD5 hi and CD5 lo CD8 T cells into NOD Rag1-/- mice and comparing them.
Weaknesses:
(1) Although CD5 can be used as a marker for self-reactivity and T cell signal strength during thymic development, it can also be regulated in the periphery by tonic TCR signaling or when T cells are activated by its cognate antigen. Hence, TCR signals in the periphery could also prime the T cells towards effector/memory differentiation. That's why from the evidence presented here it cannot be concluded that this predisposition of T cells towards effector/memory differentiation is programmed due to higher reactivity towards self-MHC molecules in the thymus, as stated in the title.
(2) Flow cytometry data needs to be revisited for the gating strategy, biological controls and interpretation.
(3) Evidence linking CD5 hi cells to more effector phenotype using gene enrichment scores is very weak.
(4) Experiments done in this study did not address why CD5 hi T cells could be negatively regulated in NOD mice when PTPN22 is overexpressed resulting in protection from diabetes but the same cannot be achieved in NOD8.3 mice.
(5) Experimental evidence provided to show that PTPN22 overexpression does not regulate TCR signaling in NOD8.3 T cells is weak.
(6) TCR sequencing analysis does not conclusively show that CD5 hi population is linked with autoreactive T cells. Doing single-cell RNAseq and TCR seq analysis would have helped address this question.
(7) When analysing data from CD5 hi T cells from the pancreatic lymph node, it is difficult to discriminate if the phenotype is just because of T cells that would have just encountered the cognate antigen in the draining lymph node or if it is truly due to basal TCR signaling.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Marcu et al. demonstrate a gut-neuron axis that is required for the lifespan-shortening effects mediated by gut bacteria. They show that the presence of commensal bacteria-particularly Acetobacter pomorum-promotes Tk expression in the gut, which then binds to neuronal tachykinin receptors to shorten lifespan. Tk has also recently been reported to extend lifespan via adipokinetic hormone (Akh) signaling (Ahrentløv et al., Nat Metab 7, 2025), but the mechanism here appears distinct. The lifespan shortening by Ap via Tk seems to be partially dependent on foxo and independent of both insulin signaling and Akh-mediated lipid mobilization.
Although the detailed mechanistic link to lifespan is not fully resolved, the experiment and its results clearly show the involvement of the molecules tested. This work adds a valuable dimension to our growing understanding of how gut bacteria influence host longevity. However, there are some points that should be addressed.
(1) Tk+ EEC activity should be assessed directly, rather than relying solely on transcript levels. Approaches such as CaLexA or GCaMP could be used.
(2) In Line243, the manuscript states that the reporter activity was not increased in the posterior midgut. However, based on the presented results in Fig4E, there is seemingly not apparent regional specificity. A more detailed explanation is necessary.
(3) If feasible, assessing foxo activation would add mechanistic depth. This could be done by monitoring foxo nuclear localization or measuring the expression levels of downstream target genes.
(4) Fig1C uses Adh for normalization. Given the high variability of the result, the authors should (1) check whether Adh expression levels changed via bacterial association and/or (2) compare the results using different genes as internal standard.
(5) While the difficulty of maintaining lifelong axenic conditions is understandable, it may still be feasible to assess the induction of Tk (i.e.. Tk transcription or EE activity upregulation) by the microbiome on males.
(6) We also had some concerns regarding the wording of the title.<br /> Fig6B and C suggests that TkR86C, in addition to TkR99D, may be involved in the A. pomorum-lifespan interaction. Consider revising the title to refer more generally to the "tachykinin receptor" rather than only TkR99D.<br /> The difference between "aging" and "lifespan" should also be addressed. While the study shows a role for Tk in lifespan, assessment of aging phenotypes (e.g. Climbing assay, ISC proliferation) beyond the smurf assay is required to make conclusions about aging.
(7) The statement in Line 82 that EEs express 14 peptide hormones should be supported with an appropriate reference, if available.
Significance:
General assessment: The main strength of this study is the careful and extensive lifespan analyses, which convincingly demonstrate the role of gut microbiota in regulating longevity. The authors clarify an important aspect of how microbial factors contribute to lifespan control. The main limitation is that the study primarily confirms the involvement of previously reported signaling pathways, without identifying novel molecular players or previously unrecognized mechanisms of lifespan regulation.
Advance: The lifespan-shortening effect of Acetobacter pomorum (Ap) has been reported previously, as has the lifespan-shortening effect of Tachykinin (Tk). However, this study is the first to link these two factors mechanistically, which represents a significant and original contribution to the field. The advance is primarily mechanistic, providing new insight into how microbial cues converge on host signaling pathways to influence ageing.
Audience: This work will be of particular interest to a specialized audience of basic researchers in ageing biology. It will also attract interest from microbiome researchers who are investigating host-microbe interactions and their physiological consequences. The findings will be useful as a conceptual framework for future mechanistic studies in this area.
Field of expertise: Drosophila ageing, lifespan, microbiome, metabolism
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors aimed to establish a long-term voltage imaging platform to investigate how coordinated neuronal activity emerges during spinal cord development in zebrafish embryos. Using the genetically encoded voltage indicator ArcLight, they tracked membrane potential dynamics in motor neurons at population, single-cell, and subcellular levels from 18 to 23 hours post-fertilization (hpf), revealing relationships between firing maturation, waveform characteristics, and axonal outgrowth.
Strengths:
(1) Technical advancement in developmental voltage imaging:
This study demonstrates voltage imaging of motor neurons in the developing vertebrate spinal cord. The approach successfully captures voltage dynamics at multiple spatial scales-neuronal population, single-cell, and subcellular compartments.
(2) Insights into the relationship between morphological and functional maturation:
The work reveals important relationships between voltage dynamics maturation and morphological changes.
(3) Kinetics analysis of membrane potential waveform enabled by voltage imaging:
The characterization of "immature" versus "mature" firing based on quantitative waveform parameters provides insights into functional maturation that are inaccessible by calcium imaging. This analysis reveals a maturation process in the biophysical properties of developing neurons.
(4) Matching of voltage indicator kinetics to biological signal:
The authors' choice of ArcLight, despite its slow kinetics compared to newer GEVIs, proved well-suited to the low-frequency activity patterns in developing spinal neurons (frequency ~0.3 Hz).
Weaknesses:
(1) Insufficient comparison with prior calcium imaging studies:
While the authors state that voltage imaging provides superior temporal resolution compared to calcium imaging (lines 192-196, 301), and this is generally true, the current manuscript does not adequately cite or discuss previous calcium imaging studies. Since neural activity occurs at low frequency in the developing spinal cord, calcium imaging is adequate for characterizing the emergence of coordinated activity patterns in the developing zebrafish spinal cord. Notably, Wan et al. (2019, Cell) performed a comprehensive single-cell reconstruction of emerging population activity in the entire developing zebrafish spinal cord using calcium imaging. This work should be properly acknowledged and compared. The specific advantages of voltage imaging over these prior studies need to be more clearly articulated, e.g. detection of subthreshold events and membrane potential waveform kinetics.
(2) Considerations for generalizability of the ArcLight-based voltage imaging approach:
While this study successfully demonstrates voltage imaging using ArcLight in the developing spinal cord, the generalizability of this approach to later developmental stages and other neural systems warrants discussion. ArcLight exhibits relatively slow kinetics (rise time ~100-200 ms, decay τ ~200-300 ms). In the current study, these kinetics are well-suited to the developmental activity patterns observed (firing frequency ~0.3 Hz), representing appropriate matching of indicator properties to biological timescales. However, the same approach may be less suitable for later developmental stages when neural activity occurs at higher frequencies.
(3) Incomplete methodological descriptions:
As a paper establishing a new imaging approach, several critical details are missing or unclear.
(a) Imaging system specifications: The imaging setup description lacks essential information, including light source specifications, excitation wavelength/filter sets, and light power at the sample. The authors should also clarify whether wide-field optics was used rather than confocal or selective plane imaging.
(b) Long-term imaging protocol: Whether neurons were imaged continuously or with breaks between imaging sessions is not explicitly stated. The current phrasing could be interpreted as a continuous 4.5-hour recording, which would be technically impressive but may not be what was actually done.
(c) Image processing procedures: Denoising and bleach correction procedures are mentioned but not described, which is critical for a methods-focused paper.
(d) The waveform classification (Supplementary Figure S6) shows overlapping kinetics between "immature" and "mature" firing, yet the classification method is not adequately justified.
(e) Given that photostability and toxicity are critical considerations for long-term voltage imaging, these aspects warrant further clarification. While the figures suggest stable ArcLight fluorescence during the experiments, the manuscript lacks quantification of photobleaching, a discussion of potential toxicity concerns associated with the indicator, and information regarding the maximum duration over which the ArcLight signal can faithfully report physiological voltage dynamics.
(4) Incomplete data representation and quantification:
(a) The claim of "reduced variability" in calcium imaging (line 194) is not clearly demonstrated in Supplementary Figure S1.
(b) Amplitude distributions for cell/subcellular compartments are not systematically quantified. Figure S3 shows ~5% changes in some axons versus ~2% in others, but it remains unclear whether these variabilities reflect differences between axonal compartments within the same cell, between individual cells, or between individual fish.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
The current paper addresses an important issue in evidence accumulation models: many modelers implement flat decision boundaries because the collapsing alternatives are hard to reliably estimate. Here, using simulations, the authors demonstrate that parameter recovery can be drastically improved by providing the model with additional data (specifically, an EEG-informed estimate of non-decision time). Moreover, in two empirical datasets, it is shown that those EEG-informed models provide a better fit to the data. The method seems sound and promising and might inform future work on the debate regarding flat vs collapsing choice boundaries. As an evidence-accumulation enthusiast, I am quite excited about this work, although for a broader audience, the immediate applicability of this approach seems limited because it does require EEG data (i.e. limiting widespread use of the method or e.g., answering questions about individual differences that require a very large N).
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors have addressed a major question since the discovery of myristate uptake from AM fungi as a non-symbiotic C source. Myristate has been used to grow some AM fungi axenically, but the biological significance of this saprobic attitude in natural or agronomical environments remained unexplored. The results of this research soundly demonstrate that myristate-derived C is used by AM fungi, leading to improved development of both extraradical and intraradical mycelium (at least under low P conditions). However, this does not lead to obvious advantages for the plant, since symbiotic nutrient exchange (carbon and phosphorus) is reduced upon myristate application. Furthermore, myristate-treated plants quench their defence responses.
Strengths:
The study is extensive, based on a solid experimental setup and methodological approach, combining several state-of-the-art techniques. The conclusions are novel and of high relevance for the scientific community. The writing is fluent and clear.
Weaknesses:
Some of the figures should be improved for clarity. The conclusions do not express a conclusive remark that, in my opinion, emerges clearly from the results: myristate application in agriculture does not seem to be a very promising approach, since it unbalances the symbiosis nutritional equilibrium and may weaken plant immunity. This is a very important point (albeit rather unpleasant for applicative scientists) that should be stressed in the conclusions.
-
-
www.medrxiv.org www.medrxiv.org
-
Reviewer #3 (Public review):
Summary:
This is an extremely important manuscript in the evolution of cerebral perfusion imaging using Arterial Spin Labelling (ASL). The number of subjects that were scanned has provided the authors with a unique opportunity to explore many potential associations between regional cerebral blood flow (CBF) and clinical and demographic variables.
Strengths:
The major strength of the manuscript is the access to an unprecedentedly large cohort of subjects. It demonstrates the sensitivity of regional tissue blood flow in the brain as an important marker of resting brain function. In addition, the authors have demonstrated a thorough analysis methodology and good statistical rigour.
Weaknesses:
This reviewer did not identify any major weaknesses in this work.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors generated knockout mice for Atad2, a conserved bromodomain-containing factor expressed during spermatogenesis. In Atad2 KO mice, HIRA, a chaperone for histone variant H3.3, was upregulated in round spermatids, accompanied by an apparent increase in H3.3 levels. Furthermore, the sequential incorporation and removal of TH2B and PRM1 during spermiogenesis were partially disrupted in the absence of ATAD2, possibly due to delayed histone removal. Despite these abnormalities, Atad2 KO male mice were able to produce offspring normally.
Strengths:
The manuscript addresses the biological role of ATAD2 in spermatogenesis using a knockout mouse model, providing a valuable in vivo framework to study chromatin regulation during male germ cell development. The observed redistribution of H3.3 in round spermatids is clearly presented and suggests a previously unappreciated role of ATAD2 in histone variant dynamics. The authors also document defects in the sequential incorporation and removal of TH2B and PRM1 during spermiogenesis, providing phenotypic insight into chromatin transitions in late spermatogenic stages. Overall, the study presents a solid foundation for further mechanistic investigation into ATAD2 function.
Weaknesses:
While the manuscript reports the gross phenotype of Atad2 KO mice, the findings remain largely superficial and do not convincingly demonstrate how ATAD2 deficiency affects chromatin.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The manuscript "Structural mechanisms of pump assembly and drug transport in the AcrAB-TolC efflux system" by Ge et al. describes the identification of a previously uncharacterized lipoprotein, YbjP, as a novel partner of the well-studied Enterobacterial tripartite efflux pump AcrAB-TolC. The authors present cryo-electron microscopy structures of the TolC-YbjP subcomplex and the complete AcrABZ-TolC-YbjP assembly. While the identification and structural characterization of YbjP are potentially novel, the stated focus of the manuscript-mechanisms of pump assembly and drug transport - is not sufficiently addressed. The manuscript requires reframing to emphasize the principal novelty associated with YbjP and significant development of the other aspects, especially the claimed novelty of the AcrB drug-efflux cycle.
Strengths:
The reported association of YbjP with AcrAB-TolC is novel; however, a recent deposition of a preceding and much more detailed manuscript to the BioRxiv server (Horne et al., https://doi.org/10.1101/2025.03.19.644130) removes much of the immediate novelty.
Weaknesses:
While the identification of YbjP is novel, the authors do not appear to acknowledge the precedence of another work (Horne et al., 2025), and it is not cited within the correct context in the manuscript.
Several results presented in the TolC-YbjP section do not represent new findings regarding TolC structure itself. The structure and gating behaviour of TolC should be more thoroughly introduced in the Introduction, including prior work describing channel opening and conformational transitions. The current manuscript does not discuss the mechanistic role of helices H3/H4 and H7/H8 in channel dilation, despite implying that YbjP binding may influence these features. Only the original closed TolC structure is cited, and the manuscript does not address prior mutational studies involving the D396 region, though this residue is specifically highlighted in the presented structures.
The manuscript provides only a general structural alignment between the closed TolC-YbjP subcomplex and the open TolC observed in the full pump assembly. However, multiple open, closed, and intermediate conformations of AcrAB-TolC have already been reported. Thus, YbjP alone cannot be assumed to account for TolC channel gating. A systematic comparison with existing structures is necessary to determine whether YbjP contributes any distinct allosteric modulation.
The analysis of AcrB peristaltic action is superficial, poorly substantiated and importantly, not novel. Several references to the ATP-synthase cycle have been provided, but this has been widely established already some 20 years ago - e.g. https://www.science.org/doi/10.1126/science.1131542.
The most significant limitation of the study is the absence of functional characterization of YbjP in vivo or in vitro. While the structural association between YbjP and TolC is interesting, the biological role of YbjP remains unclear. Moreover, the manuscript does not examine structural differences between the presented complex and previously solved AcrAB-TolC or MexAB-OprM assemblies that might support a mechanistic model.
-
-
-
Reviewer #3 (Public review):
This paper reveals that the neuronal protein PRRT2, previously known for its association with paroxysmal dyskinesia and infantile seizures, modulates the slow inactivation of voltage-gated sodium ion (Nav) channels, a gating process that limits excitability during prolonged activity. Using electrophysiology, molecular biology, and mouse models, the authors show that PRRT2 accelerates entry of Nav channels into the slow-inactivated state and slows their recovery, effectively dampening excessive excitability. The effect seems evolutionarily conserved, requires the C-terminal region of PRRT2, and is recapitulated in cortical neurons, where PRRT2 deficiency leads to hyper-responsiveness and reduced cortical resilience in vivo. These findings extend the functional repertoire of PRRT2, identifying it as a physiological brake on neuronal excitability. The work provides a mechanistic link between PRRT2 mutations and episodic neurological phenotypes.
Comments:
(1) The precise structural interface and the molecular basis of gating modulation remain inferred rather than demonstrated.
(2) The in vivo phenotype reflects a complex circuit outcome and does not isolate slow-inactivation defects per se.
(3) Expression of PRRT2 in muscle or heart is low, so the cross isoform claims are likely of limited physiological significance.
(4) The mechanistic separation between the trafficking of PRRT2 and its gating effects is not clearly resolved.
(5) Additional studies with Nav1.6 should be carried out.
-
-
papers.ssrn.com papers.ssrn.com
-
Reviewer #3 (Public review):
Summary:
This perspective article by Reichmann et al. highlights the importance of moving beyond the search for a single, unified immune mechanism to explain host-Mtb interactions. Drawing from studies in immune profiling, host and bacterial genetics, the authors emphasize inconsistencies in the literature and argue for broader, more integrative models. Overall, the article is thought-provoking and well-articulated, raising a concept that is worth further exploration in the TB field.
Strengths:
Timely and relevant in the context of the rapidly expanding multi-omics datasets that provide unprecedented insights into host-Mtb interactions.
Weaknesses (Minor):
(1) Clarity on the notion of a "unified mechanism". It remains unclear whether prior studies explicitly proposed a single unifying immunological model. While inconsistencies in findings exist, they do not necessarily demonstrate that earlier work was uniformly "single-minded". Moreover, heterogeneity in TB has been recognized previously (PMIDs: 19855401, 28736436), which the authors could acknowledge.
(2) Evolutionary timeline and industrial-era framing. The evolutionary model is outdated. Ancient DNA studies place the Mtb's most recent common ancestor at ~6,000 years BP (PMIDs: 25141181; 25848958). The Industrial Revolution is cited as a driver of TB expansion, but this remains speculative without bacterial-genomics evidence and should be framed as a hypothesis. Additionally, the claim that Mtb genomes have been conserved only since the Industrial Revolution (lines 165-167) is inaccurate; conservation extends back to the MRCA (PMID: 31448322).
(3) Trained immunity and TB infection. The treatment of trained immunity is incomplete. While BCG vaccination is known to induce trained immunity (ref 59), revaccination does not provide sustained protection (ref 8), and importantly, Mtb infection itself can also impart trained immunity (PMID: 33125891). Including these nuances would strengthen the discussion.
Tags
Annotators
URL
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
In this manuscript, the authors investigate how odor-evoked neural activity is modulated by experience within the olfactory-hippocampal network. The authors perform extracellular recordings in the anterior olfactory nucleus (AON), the anterior piriform (aPCx) and lateral entorhinal cortex (LEC), the hippocampus (CA1) and the subiculum (SUB), in naïve mice and in mice repeatedly exposed to the same odorants. They determine the response properties of individual neurons and use population decoding analyses to assess the effect of experience on odor information coding across these regions.
The authors' findings show that odor identity is represented in all recorded areas, but that the response magnitude and selectivity of neurons are differentially modulated by experience across the olfactory-hippocampal pathway.
Overall, this work represents a valuable multi-region data set of odor-evoked neural activity. However, a few limitations in experimental design and analysis restrict the conclusions that can be drawn from this study.
Main limitations:
The authors use a non-associative learning paradigm - repeated odor exposure - to test how experience modulates odor responses along the olfactory-hippocampal pathway. While repeated odor exposure clearly modulates sampling behavior and odor-evoked neural activity, the relevance of this modulation across different brain areas remains difficult to assess.
The authors discuss the olfactory-hippocampal pathway as a transition from primary sensory (AON, aPCx) to associative areas (LEC, CA1, SUB). While this is reasonable, given the known circuit connectivity, other interpretations are possible. For example, AON, aPCx, and LEC receive direct inputs from the olfactory bulb ('primary cortex'), while CA1 and SUB do not; AON receives direct top-down inputs from CA1 ('associative cortex'), while aPCx does not. In fact, the data presented in this manuscript do not appear to support a consistent transformation from sensory to associative, as implied by the authors.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Thank you for inviting me to review this manuscript entitled "Pupil dilation offers a time-window on prediction error" by Colizoli and colleagues. The study examines prediction errors, information gain (Kullback-Leibler [KL] divergence), and uncertainty (entropy) from an information-theory perspective using two experimental tasks and pupillometry. The authors aim to test a theoretical proposal by Zénon (2019) that the pupil response reflects information gain (KL divergence). The conclusion of this work is that (post-feedback) pupil dilation in response to information gain is context dependent.
Strengths:
Use of an established Bayesian model to compute KL divergence and entropy.
Pupillometry data preprocessing and multiple robustness checks.
Weaknesses:
Operationalization of prediction errors based on frequency, accuracy, and their interaction:
The authors rely on a more model-agnostic definition of the prediction error in terms of stimulus frequency ("unsigned prediction error"), accuracy, and their interaction ("signed prediction error"). While I see the point, I would argue that this approach provides a simple approximation of the prediction error, but that a model-based approach would be more appropriate.
Model validation:
My impression is that the ideal learner model should work well in this case. However, the authors don't directly compare model behavior to participant behavior ("posterior predictive checks") to validate the model. Therefore, it is currently unclear if the model-derived terms like KL divergence and entropy provide reasonable estimates for the participant data.
Lack of a clear conclusion:
The authors conclude that this study shows for the first time that (post-feedback) pupil dilation in response to information gain is context dependent. However, the study does not offer a unifying explanation for such context dependence. The discussion is quite detailed with respect to task-specific effects, but fails to provide an overarching perspective on the context-dependent nature of pupil signatures of information gain. This seems to be partly due to the strong differences between the experimental tasks.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The manuscript of Mayer and colleagues analyzes the function of WIPI proteins in mammalian cells. The authors previously identified CROP as a complex consisting of WIPI1 and the retromer complex, primarily in yeast cells. In mammalian cells, both WIPI1 and WIPI2 exist, whereas retromer has a homologous complex termed retriever. They now find that WIPI2 can form a complex with retriever subunits. They named this complex CROP2. Their data further indicate that CROP2 and CROP1 have distinct substrate specificities as knockdown of CROP2 subunits affects beta1 integrin sorting, whereas knockdown of CROP1 affects EGFR and GLUT1. They further identify a similar sequence (FSSS) in both WIPI1 and WIPI2, which is required for their specific binding to retromer and retriever.
Strengths:
CROP1 and CROP2 seem to use similar features for their formation, and have different substrates, which is convincingly shown.
Weaknesses:
The analysis lacks information that this is a complex as claimed. It can be deduced from the interaction analysis, but was not shown.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Taking advantage of the existence in fish of two genes coding for estrogen synthase, the enzyme aromatase, one mostly expressed in the brain (Cyp19a1b) and the other mostly found in the gonads (Cyp19a1a), this study investigates the role of brain-derived estrogens in the control of sexual and aggressive behavior in male medaka. The constitutive deletion of Cyp19a1b, confirmed by the ablation of its transcript, markedly reduced brain estrogen content. This effect is accompanied by reduced sexual and aggressive behavior and reduced expression of the transcripts coding for androgen receptors (AR), ara and arb, in brain regions involved in social behavior regulation. Both AR expression and aspects of social behaviors were restored by adult treatment with estrogens, providing some support for a role of aromatization. Expression analysis of AR isoforms and behavior of mutants of estrogen receptors (ER) indicates that these effects are likely mediated by the activation of the esr1 and esr2a isoforms. Together, these results provide valuable insights into the role of brain-derived estrogens in social behavior in fish.
Strengths:
This study evaluates the role of brain "specific" Cyp19a1 in the social behavior in male teleost fish, which as a taxon are more abundant and yet proportionally less studied that the most common birds and rodents. Therefore, evaluating the generalizability of results from higher vertebrates is important. The study suggests that, as opposed to mammals, the facilitatory role of brain-derived estrogens on mating and aggression is limited to adulthood.
Results obtained from multiple mutant lines converge to show that estrogens most likely synthesized in the brain drives aspects of male sexual behavior.
The comparative discussion of the age-dependent abundance of brain aromatase in fish vs mammals and its role in organization vs activation is important beyond the study of the targeted species.
Weaknesses:
Most experiments are weakly powered (low sample size).
The variability of the mRNA content for a same target gene between experiments (genotype comparison vs E2 treatment comparison) raises questions about the reproducibility of the data (apparent disappearance of genotype effect).
Conclusions :
Overall, the present study provides convincing evidence for a facilitatory role of estrogens originating from the brain on sexual behavior and aggressive behavior in male medaka. The role of specific estrogen receptor isoforms underlying the expression of androgen receptors is supported by converging evidence from multiple mutant lines.
-
-
-
Reviewer #3 (Public review):
Summary:
Metabolic dysfunction associated liver disease (MASLD) describes a spectrum of progressive liver pathologies linked to life style-associated metabolic alterations (such as increased body weight and elevated blood sugar levels), reaching from steatosis over steatohepatitis to fibrosis and finally end stage complications, such as liver failure and hepatocellular carcinoma. Treatment options for MASLD include diet adjustments, weight loss, and the receptor-β (THR-β) agonist resmetirom, but remain limited at this stage, motivating further studies to elucidate molecular disease mechanisms to identify novel therapeutic targets.
In their present study, the authors aim to identify early molecular changes in MASLD linked to obesity. To this end, they study a cohort of 109 obese individuals with no or early-stage MASLD combining measurements from two anatomic sides: 1. bulk RNA-sequencing and metabolomics of liver biopsies, and 2. metabolomics from patient blood. Their major finding is that GTPase-related genes are transcriptionally altered in livers of individuals with steatosis with fibrosis compared to steatosis without fibrosis.
Comments from the first round of review:
(1) Confounders (such as (pre-)diabetes)
The patient table shows significant differences in non-MASLD vs. MASLD individuals, with the latter suffering more often from diabetes or hypertriglyceridemia. Rather than just stating corrections, subgroup analyses should be performed (accompanied with designated statistical power analyses) to infer the degree to which these conditions contribute to the observations. I.e., major findings stating MASLD-associated changes should hold true in the subgroup of MASLD patients without diabetes/of female sex and so forth (testing for each of the significant differences between groups).
Post-rebuttal update: The authors have performed the requested sub-group analysis and find the gene signatures hold for the non-diabetic sub-cohort, but not the diabetic subgroup. They denote a likely interaction between fibrosis and diabetes, that was not corrected for in the original analysis.
(2) External validation
Additionally, to back up the major GTPase signature findings, it would be desirable to analyze an external dataset of (pre)diabetes patients (other biased groups) for alternations in these genes. It would be important to know if this signature also shows in non-MASLD diabetic patients vs. healthy patients or is a feature specific to MASLD. Also, could the matched metabolic data be used to validate metabolite alterations that would be expected under GTPase-associated protein dysregulation?
Post-rebuttal update: The authors confirm that with the present data, insulin resistance cannot be fully ruled out as a confounder to the GTP-ase related gene signature. They however plan future mouse model experiments to study whether the GTPase-fibrosis signature differs in diabetic vs. non-diabetic conditions.
(3).3D liver spheroid MASH model, Fig. 6D/E
This 3D experiment is technically not an external validation of GTPase-related genes being involved in MASLD, since patient-derived cells may only retain changes that have happened in vivo. To demonstrate that the GTPase expression signature is specifically invoked by fibrosis the LX-2 set up is more convincing, however, the up-regulation of the GTPase-related genes upon fibrosis induction with TGF-beta, in concordance with the patient data, needs to be shown first (qPCR or RNA-seq). Additionally, the description of the 3D model is too uncritical. The maintenance of functional PHHs is a major challenge (PMID: 38750036, PMID: 21953633, PMID: 40240606, PMID: 31023926). It cannot be ruled out that their findings are largely attributable to either 1) the (other present) mesenchymal cells (i.e., mesenchyme-derived cells, such as for example hepatic stellate cells, not to be confused with mesenchymal stem cells, MSCs), or 2) related to potential changes in PHHs in culture, and these limitations need to be stated.
Post-rebuttal update: To address the concern of other cells than hepatocytes contributing to the observed effects in culture, the authors performed TGF-beta treatment in independent mono-cultures (Figure R4): LX-2 and hepatocytes, and the spheroid system. Surprisingly, important genes highlighted in Figure 6E for the spheroid system (RAB6A, ARL4A, RAB27B, DIRAS2) are all absent from this qPCR(?) validation experiment. The authors evaluate instead RAC1, RHOU, VAV1, DOCK2, RAB32. In spheroids, RHOU and RAB32 are down-regulated with TGF-B. In hepatocytes DOCK2 and RAC seemed up-regulated. They find no difference in these genes in LX-2 cells. Surprisingly, ACTA2 expression values are missing for LX-2 cells. Together, it is hard to judge which individual cell type recapitulates the changes observed in patients in this validation experiment, as the major genes called out in Figure 6E are not analyzed.
Unfortunately, the 3D liver spheroid model used (as presented in PMID39605182) lacks important functional validation tests of maintained hepatocyte identity in culture (at the very least Albumin expression and secretion plus CYP3A4 assay). This functional data (acquired at the time point in culture when the RNA expression analysis in 6E was performed) is indispensable prior to stating that mature hepatocytes cause the observed effects.
(4) Novelty / references
Similar studies that also combined liver and blood lipidomics/metabolomics in obese individuals with and without MASLD (e.g. PMID 39731853, 39653777) should be cited. Additionally, it would benefit the quality of the discussion to state how findings in this study add new insights over previous studies, if their findings/insights differ, and if so, why.
Post-rebuttal update: The authors have included the studies into their discussion.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This study investigates the functional differences between barrel and septal columns in the mouse somatosensory cortex, focusing on how local inhibitory dynamics (particularly involving SST⁺ interneurons) may mediate temporal integration of multi-whisker (MW) stimuli in septa. Using a combination of in vivo multi-unit recordings, calcium imaging, and anatomical tracing, the authors propose a model in which Elfn1-dependent synaptic facilitation onto SST⁺ interneurons contributes to the distinct sensory responses to MW input in barrels and septa, enabling functional segregation between these domains.
Strengths:
The study presents a thought-provoking and useful conceptual model for understanding sensory processing in the somatosensory cortex. While barrel columns have been widely studied, septal regions remain relatively understudied in mice. If septa indeed act as selective integrators of distributed sensory input, this would suggest a novel computational role for cortical microcircuits beyond the classical view focused on barrels. Although still hypothetical, the proposed model in which SST⁺ interneurons contribute to domain-specific sensory responses between barrel and septal domains is intriguing and opens new avenues for investigating inhibitory circuit mechanisms.
Weaknesses:
The primary limitation of this study lies in the spatial and cellular specificity of the recording techniques. The physiological data rely predominantly on unsorted multi-unit activity (MUA) recorded with low-channel-count silicon probes. Because MUA aggregates signals from multiple neurons over a radius of approximately 50-100 µm (often wider than the typical septal width in mice), this approach makes it difficult to confidently isolate activity originating strictly from within septal domains. The manuscript would benefit from additional analyses to validate the spatial specificity of these recordings, such as systematically varying spike detection thresholds to test the robustness of domain attribution, as suggested by the reviewer. Furthermore, although the authors now appropriately frame their findings in the Elfn1 knockout mice as indirect evidence, it is worth emphasizing that the study lacks direct in vivo, cell-type-specific recordings and manipulations to more definitively test the proposed mechanism.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
In this study, Camuso et al., make use of a knock-in mouse model expressing endogenously mEos4b-tagged GlyRβ subunits to detect endogenous glycine receptors in mouse brain using single-molecule localization microscopy (SMLM). At synapses in the hippocampus GlyRβ molecules are detected at very low copy numbers. Assuming that each detected GlyRβ molecule is incorporated in a pentameric glycine receptor, it was estimated that while the majority of hippocampal inhibitory synapses do not contain glycine receptors, a small population of inhibitory synapses contain a few (up to 10) glycine receptors. Using dual-color SMLM approaches it is furthermore confirmed that the detected GlyRβ molecules are embedded in the postsynaptic domain marked by gephyrin. In contrast to the hippocampus, at inhibitory synapses in the striatum GlyRβ molecules were detected at considerably higher copy numbers. Interestingly, the observed number of GlyRβ detections was significantly higher in the ventral striatum compared to the dorsal striatum. These findings are corroborated by electrophysiological recordings showing that postsynaptic glycinergic currents can be readily detected in the ventral striatum but are almost absent in the dorsal striatum. Using lentiviral overexpression of recombinant GlyRalpha1, alpha2, and beta subunits in cultured hippocampal neurons, it is shown that GlyR alpha1 subunits are readily detectable at synapses, but overexpressed GlyRalpha2 and beta subunits did not strongly enrich at synapses. This could indicate that GlyRa1 expression is limiting the synaptic expression of GlyRβ-containing glycine receptors in hippocampal neurons.
Comments on revised version:
This revised manuscript is significantly improved. New experimental and quantitative analysis is presented that strengthen the conclusions. Overall, the results presented in this manuscript are based on carefully performed SMLM experiments and are well-presented and described. The knock-in mouse with endogenously tagged GlyRβ molecules is a very strong aspect of this study and provides confidence in the labeling, the combination with SMLM is very strong as it provides high sensitivity and spatial resolution. These results confirm previous studies and will be of interest to a specialised audience interested in glycine receptors, inhibitory synapse biology and super-resolution microscopy.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This work provides an overview of the motor neuron landscape in the male reproductive system. Some work had been done to elucidate the circuits of ejaculation in the spine, as well as, the cord but this work fills a gap of knowledge at the level of the reproductive organs. Using complementary approaches the authors show that there are two types of motor neurons that are mutually exclusive: neurons that co-express octopamine and glutamate and neurons that co-express serotonin and glutamate. They also show evidence that both types of neurons express large dense core vesicles indicating that neuropeptides play a role in male fertility. This paper provides a thorough characterization of expression of the different glutamate, octopamine and serotonin receptors in the different organs and tissues of the male reproductive system. The differential expression in different tissues and organs allows building initial theories on the control of emission and expulsion. Additionally, the authors characterize the expression of synaptic proteins and the neuromuscular junction sites. On a mechanistic level, the authors show that neither octopamine/glutamate neuron transmission nor glutamate transmission in serotonin/glutamate neurons are required for male fertility. This final result is quite surprising and opens up many questions on how ejaculation is coordinated.
Strengths:
This work fills an important gap on characterization of innervation of the male reproductive system by providing an extensive characterization of the motor neurons and the potential receptors of motor neuron release.The authors show convincing evidence of glutamate/monoamine co-release and of mutual exclusivity of serotonin/glutamate and octopamine/glutamate neurons.
Weaknesses:
The experiment looking at peristaltic waves in the male organs is missing labeling of the different regions and quantification of the observed waves.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Primary taste cortex neurons show a variety of dynamic response profiles during taste decision-making tasks, reflecting both sensory and decision variables. In the present study, Lang et al. set out to determine how neurons with distinct response profiles contribute to perceptual decisions about taste stimuli.
The methods, with reference to the behavioral task and electrophysiological recordings/data analysis, are straightforward, solid, and appropriate. The computational model is presented in a clear and conceptually intuitive manner, although the details are outside of my area of expertise.
The experimental design features a simple 2-alternative forced-choice design that yielded clear psychometric curves across a range of stimuli. In vivo recordings were performed using Neuropixels and yielded an appropriate sample of single neuron responses. The strength of the model lies in the fact that it consists of single neurons whose response profiles mimic those recorded in vivo, and allows neuron-selective manipulation.
By virtually lesioning specific subsets of neurons in the network, the authors demonstrate that a relatively small population of neurons with specific tuning profiles was sufficient to produce the observed neural dynamics and behavioral responses. This effect was selective as lesioning other responsive neurons did not affect overall response dynamics or performance.
These findings provide new insight into the relation between the response profiles of single neurons in sensory cortex, their population-level activity dynamics, and the perceptual decisions they inform.
The approach is particularly innovative as it uses computational modeling to target functionally-defined "cell types", which cannot necessarily be targeted by more conventional genetic approaches.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This research focuses on a long-lasting and interesting phenomenon in human plasticity. When humans learn basic perceptual skills such as judging the orientation of a simple line, the learned abilities are often limited to the trained condition but not generalizable to untrained conditions. The authors hypothesized that this learning specificity was related to GABA, an inhibitory neurotransmitter in the brain. Using a novel training method that combines reactivation and a brain stimulation method (tDCS) that hypothetically inactivates GABA, the authors hypothesized that learned visual perceptual skills would show greater transfer.
Strengths:
The authors conducted a list of well-conceived behavior studies to demonstrate the effectiveness of their proposed method in enabling learning transfer in two different visual tasks, and carefully conducted comparison studies to elucidate other possible explanations. The sample size was adequate to convey convincing results, and the analyses were thorough.
Weaknesses:
While the authors built their training paradigm on
(1) the hypothetical role GABA plays in inhibiting learning transfer, and
(2) the hypothetical impact tDCS may have on GABA, there was no direct evidence supporting these hypotheses in the current study.
Further, learning specificity takes many formats from features to locations to tasks; it is not yet clear the scope of the observed transfer with the proposed method.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors aimed to overcome the challenges associated with complex, conventional prokaryotic cell-free protein synthesis (CFPS) systems, which require up to thirty-five components, by developing a streamlined and efficient E. coli CFPS platform to encourage broader adoption. The main objective was to reduce the number of reaction components from thirty-five to seven, while also developing an accessible 'fast lysate' preparation protocol that eliminates time-consuming runoff and dialysis steps. The authors also sought to demonstrate the robustness and translational quality of this streamlined system by efficiently synthesising challenging functional proteins, including the cytotoxic restriction endonuclease BsaI and the self-assembling intermediate filament protein vimentin.
Strengths:
This study presents several key strengths of the optimised E. coli cell-free protein synthesis system in terms of its design, performance and accessibility.
(1) The reaction mixture has been dramatically simplified, with the number of essential core components successfully reduced from up to thirty-five in conventional systems to just seven.
(2) The "fast lysate" protocol is a significant advance in terms of procedure.
(3) The system's ability to synthesise challenging, functional proteins is evidence of its robustness.
Weaknesses:
(1) Title: "A simplified and highly efficient cell-free protein synthesis system for prokaryotes".
(a) This title is misleading since one would expect a simplified and highly efficient cell-free protein synthesis system to yield similar protein levels compared to current cell-free protein synthesis systems. What this study shows is that the composition of cell-free protein synthesis systems can be simplified while maintaining a certain level of protein synthesis. Here, optimisation does not involve maintaining protein synthesis yield while simplifying the cell-free protein synthesis system; rather, it involves developing a simplified cell-free protein synthesis system. As mentioned in my comments below, this study lacks a comparison of protein levels with a typical cell-free protein synthesis system.
(b) What do the authors mean by "highly efficient"? Highly efficient compared to what experimental conditions? If one is interested in the yield of protein synthesis, is this simplified system highly efficient compared to current systems?
(2) Figures 1, 3-5 :
(a) What do relative luciferase units represent? How are these units calculated?
(b) In this system, the level of expression depends mainly on the level of NLuc transcripts and the efficiency of NLuc translation. How did the authors ensure that the chemical composition of the different eCFPS buffers only affected protein translation and not transcript levels? In other words, are luciferase units solely an indicator of protein synthesis efficiency, or do they also depend on transcription efficiency, which could vary depending on the experimental conditions?
(c) How long were the eCFPS reactions allowed to proceed before performing the luciferase activity measurement? Depending on the reaction time, the absence or presence of certain compounds may or may not impact NLuc expression. For example, it can be assumed that tRNA does not significantly affect NLuc levels over a short period of time, and that endogenous tRNA in the lysate is present at sufficient concentrations. However, over a longer period of time, the addition of tRNA could be essential to achieve optimal NLuc levels.
(d) The authors show that tRNA and amino acids are not strictly essential for the expression of NLuc, likely due to residual amounts within the cell lysate. However, are the protein levels achieved without added amino acids and tRNA sufficient for biochemical assays that require a certain amount of protein? It is important to note that the focus here is on optimising the simplicity of the buffer rather than the level of protein expression. In fact, the simplicity of the buffer is prioritised over the amount of protein produced. This should be made clear.
(e) How would the NLuc level compare if all the components were optimised individually and present in an optimised buffer, compared to a buffer optimised for simplicity as described by the authors?
(3) Line 71, Streamlining eCFPS: removal of dispensable components. This title is misleading because it creates the false impression that proteins can be produced in vitro without the addition of certain compounds. While this is true, the level of protein produced may not be sufficient for subsequent biochemical analyses. This should be made clear.
(4) Figure 2: In the legend, "(A) Protein expression levels of the eCFPS system measured at varying concentrations of KGlu and MgGlu2" would be more accurate if changed to "(A) Protein expression levels of the eCFPS system using an Nanoluciferase (NLuc) reporter DNA measured at varying concentrations of KGlu and MgGlu2".
(5) Lanes 302-303: "The thorough optimization of the seven core components was a critical step in achieving high protein expression levels". What are "high expression levels"? Compared to what?
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Here Wengert et al., establish a rodent model of KCNC1 (Kv3.1) epilepsy by introducing the A421V mutation. The authors perform video-EEG, slice electrophysiology, and in vivo 2P imaging of calcium activity to establish a disease mechanisms involving impairment in the excitability of fast spiking parvalbumin (PV) interneurons in the cortex and thalamic PV cells.
Outside out nucleated patch recordings were used to evaluate the biophysical consequence of the A421V mutation on potassium currents and showed a clear reduction in potassium currents. Similarly action potential generation in cortical PV interneurons was severely reduced. Given that both potassium currents and action potential generation was found to be unaffected in excitatory pyramidal cells in the cortex the authors propose that loss of inhibition leads to hyperexcitability and seizure susceptibility in a mechanism similar to that of Dravet Syndrome.
Strengths:
This manuscript establishes a new rodent model of KCNC1-developmental and epileptic encephalopathy. The manuscript provides strong evidence that parvabumin interneurons are impaired by the Kcnc1-A421V mutation and that cortical excitatory neurons are not impaired. Together, these findings support the conclusion that seizure phenotypes associated with Kcnc1-A421V are caused by impaired cortical inhibition.
Weaknesses:
The manuscript identifies a partial mechanism of disease that leaves several aspects unresolved including the possible role of subcortical regions in the seizure mechanism. Similarly, while the authors identify a reduction in potassium currents and a reduction in PV cell surface expression of Kv3.1 why the A421V missense mutation leads to a more severe phenotype than previously reported loss-of-function mutations in Kv3.1is not clear.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
In this paper, Sandkuhler et al. reassessed the role of TANGO2 as a heme chaperone proposed by Sun et al in a recently published paper (https://doi.org/10.1038/s41586-022-05347-z). Overall, Sandkuhler et al. conclude that the heme-related roles of TANGO2 had been overemphasized by Sun et al. especially because the hrg9 gene does not exclusively respond to different regimens of heme synthesis/uptake but is susceptible to a greater extent to, for example, oxidative stress. Impaired heme trafficking is then interpreted as due to general mitochondrial dysfunction. In recent years, the discussion around the heme-related roles of TANGO2 has been tantalizing but is still far from a definitive consensus. Discrepancies between results and their interpretation are testament to how ambitious the understanding of TANGO2 and the phenotypes associated with TANGO2 defects are.
The work presented by Sandkuhler et al. is methodologically sound, and the authors have appropriately addressed my concerns in the first round of review. Overall, this paper challenges the recent developments in the field in relation to heme trafficking and provides a wider perspective on the biological roles of TANGO2.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors of this paper were trying to identify how reproducible, or not, their subfield (Drosophilia immunity) was since its inception over 50 years ago. This required identifying not only the papers, but the specific claims made in the paper, assessing if these claims were followed up in the literature, and if so whether the subsequent papers supported or refuted the original claim. In addition to this large manually curated effort, the authors further investigated some claims that were left unchallenged in the literature by conducting replications themselves. This provided a rich corpus of the subfield that could be investigated into what characteristics influence reproducibility.
Strengths:
A major strength of this study is the focus on a subfield, the detailing of identifying the main, major, and minor claims - which is a very challenging manual task - and then cataloging not only their assessment of if these claims were followed up in the literature, but also what characteristics might be contributing to reproducibility, which also included more manual effort to supplement the data that they were able to extract from the published papers. While this provides a rich dataset for analysis, there is a major weakness with this approach, which is not unique to this study.
Weaknesses:
The main weakness is relying heavily on the published literature as the source for if a claim was determined to be verified or not. There are many documented issues with this stemming from every field of research - such as publication bias, selective reporting, all the way to fraud. It's understandable why the authors took this approach - it is the only way to get at a breadth of the literature - however the flaw with this approach is it takes the literature as a solid ground truth, which it is not. At the same time, it is not reasonable to expect the authors to have conducted independent replications for all of the 400 papers they identified. However, there is a big difference trying to assess the reproducibility of the literature by using the literature as the 'ground truth' vs doing this independently like other large-scale replication projects have attempted to do. This means the interpretation of the data is a bit challenging.
Below are suggestions for the authors and readers to consider:
(1) I understand why the authors prefer to mention claims as their primary means of reporting what they found, but it is nested within paper, and that makes it very hard to understand how to interpret these results at times. I also cannot understand at the high-level the relationship between claims and papers. The methods suggest there are 3-4 major claims per paper, but at 400 papers and 1,006 claims, this averages to ~2.5 claims per paper. Can the authors consider describing this relationship better (e.g., distribution of claims and papers) and/or considering presenting the data two ways (primary figures as claims and complimentary supplementary figures with papers as the unit). This will help the reader interpret the data both ways without confusion. I am also curious how the results look when presented both ways (e.g., does shifting to the paper as the unit of analysis shift the figures and interpretation?). This is especially true since the first and last author analysis shows there is varying distribution of papers and claims by authors (and thus the relationship between these is important for the reader).
(2) As mentioned above, I think the biggest weakness is that the authors are taking the literature at face value when assigning if a claim was validated or challenged vs gathering new independent evidence. This means the paper leans more on papers, making it more like a citation analysis vs an independent effort like other large-scale replication projects. I highly recommend the authors state this in their limitations section.
On top of that, I have questions that I could not figure out (though I acknowledge I did not dig super deep into the data to try). The main comment I have is How was verified (and challenged) determined? It seems from the methods it was determined by "Claims were cross-checked with evidence from previous, contemporary and subsequent publications and assigned a verification category". If this is true, and all claims were done this way - are verified claims double counted then? (e.g., an original claim is found by a future claim to be verified - and thus that future claim is also considered to be verified because of the original claim).
Related, did the authors look at the strength of validation or challenged claims? That is, if there is a relationship mapping the authors did for original claims and follow-up claims, I would imagine some claims have deeper (i.e., more) claims that followed up on them vs others. This might be interested to look at as well.
(3) I recommend the authors add sample sizes when not present (e.g., Fig 4C). I also find that the sample sizes are a bit confusing, and I recommend the authors check them and add more explanation when not complete, like they did for Fig 4A. For example, Fig 7B equals to 178 labs (how did more than 156 labs get determined here?), and yet the total number of claims is 996 (opposed to 1,006). Another example, is why does Fig 8B not have all 156 labs accounted for? (related to Fig 8B, I caution on reporting a p value and drawing strong conclusions from this very small sample size - 22 authors). As a last example, Fig 8C has al 156 labs and 1,006 claims - is that expected? I guess it means authors who published before 1995 (as shown in Figure 8A continued to publish after 1995?) in that case, it's all authors? But the text says when they 'set up their lab' after 1995, but how can that be?
(4) Finally, I think it would help if the authors expanded on the limitations generally and potential alternative explanations and/or driving factors. For example, the line "though likely underestimated' is indicated in the discussion about the low rate of challenged claims, it might be useful to call out how publication bias is likely the driver here and thus it needs to be carefully considered in the interpretation of this. Related, I caution the authors on overinterpreting their suggestive evidence. The abstract for example, states claims of what was found in their analysis, when these are suggestive at best, which the authors acknowledge in the paper. But since most people start with the abstract, I worry this is indicating stronger evidence than what the authors actually have.
The authors should be applauded for the monumental effort they put into this project, which does a wonderful job of having experts within a subfield engage their community to understand the connectiveness of the literature and attempt to understand how reliable specific results are and what factors might contribute to them. This project provides a nice blueprint for others to build from as well as leverage the data generated from this subfield, and thus should have an impact in the broader discussion on reproducibility and reliability of research evidence.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The submission from Cronshagen and colleagues describes the application of a previously described method (selection linked integration) to the systematic study of PfEMP1 trafficking in the human malaria parasite Plasmodium falciparum. PfEMP1 is the primary virulence factor and surface antigen of infected red blood cells and is therefore a major focus of research into malaria pathogenesis. Since the discovery of the var gene family that encodes PfEMP1 in the late 1990s, there have been multiple hypotheses for how the protein is trafficked to the infected cell surface, crossing multiple membranes along the way. One difficulty in studying this process is the large size of the var gene family and the propensity of the parasites to switch which var gene is expressed, thus preventing straightforward gene modification-based strategies for tagging the expressed PfEMP1. Here the authors solve this problem by forcing expression of a targeted var gene by fusing the PfEMP1 coding region with a drug selectable marker separated by a skip peptide. This enabled them to generate relatively homogenous populations of parasites all expressing tagged (or otherwise modified) forms of PfEMP1 suitable for study. They then applied this method to study various aspects of PfEMP1 trafficking.
Strengths:
The study is very thorough, and the data are well presented. The authors used SLI to target multiple var genes, thus demonstrating the robustness of their strategy. They then perform experiments to investigate possible trafficking through PTEX, they knockout proteins thought to be involved in PfEMP1 trafficking and observe defects in cytoadherence, and they perform proximity labeling to further identify proteins potentially involved in PfEMP1 export. These are independent and complimentary approaches that together tell a very compelling story.
Weaknesses:
(1) When the authors targeted IT4var19, they were successful in transcriptionally activating the gene, however they did not initially obtain cytoadherent parasites. To observe binding to ICAM-1 and EPCR, they had to perform selection using panning. This is an interesting observation and potentially provides insights into PfEMP1 surface display, folding, etc. However, it also raises questions about other instances in which cytoadherence was not observed. Would panning of these other lines have successfully selected for cytoadherent infected cells? Did the authors attempt panning of their 3D7 lines? Given that these parasites do export PfEMP1 to the infected cell surface (Figure 1D), it is possible that panning would similarly rescue binding. Likewise, the authors knocked out PTP1, TryThrA and EMPIC3 and detected a loss of cytoadhesion, but they did not attempt panning to see if this could rescue binding. The strong selection that panning exerts on parasite populations could result in selection of compensatory changes that enable cytoadherence, which could be very informative, although the analysis could potentially be quite complicated and beyond the scope of the current paper. Nonetheless, these are important concepts to consider when assessing these phenotypes.
(2) The authors perform a series of trafficking experiments to help discern whether PfEMP1 is trafficked through PTEX. While the results were not entirely definitive, they make a strong case for PTEX in PfEMP1 export. The authors then used BioID to obtain a proxiome for PfEMP1 and identified proteins they suggest are involved in PfEMP1 trafficking. However, it seemed that components of PTEX were missing from the list of interacting proteins. Is this surprising and does this observation shed any additional light on the possibility of PfEMP1 trafficking through PTEX? This warrants a comment or discussion.
Comments on revisions:
The authors have responded thoroughly and constructively to suggestions and comments in the initial review. I have no additional comments. This is a great contribution to the literature.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary
GPR30 responds to bicarbonate and plays a role in regulating cellular pH and ion homeostasis. However, the molecular basis of bicarbonate recognition by GPR30 remains unresolved. This study reports the cryo-EM structure of GPR30 bound to a chimeric mini-Gq in the presence of bicarbonate, revealing mechanistic insights into its G-protein coupling. Nonetheless, the study does not identify the bicarbonate-binding site within GPR30.
Strengths
The work provides strong structural evidence clarifying how GPR30 engages and couples with Gq.
Weaknesses
Several GPR30 mutants exhibited diminished responses to bicarbonate, but their expression levels were also reduced. As a result, the mechanism by which GPR30 recognizes bicarbonate remains uncertain, leaving this aspect of the study incomplete.
-
-
www.youtube.com www.youtube.com
-
boardrooms and parliaments, it's somewhere between 3 to 21%. Now, again, numbers are very disputed
for - stats - psychopathy - 3 to 21% in boardrooms and parliaments - more likely to find psychopath in boardroom and parliament than grocery store - SRG comment - stats- shadow side of leadership - high percentage of leaders have dark triad
-
-
www.medrxiv.org www.medrxiv.org
-
Reviewer #3 (Public review):
This work provides a novel statistical model to identify imported malaria cases, which are an important challenge for elimination, particularly in low-transmission areas. This tool was applied in Plasmodium falciparum populations in Mozambique and determined differences in importation rates in 2 low-transmission districts in the South.
Strengths:
The study has several strengths, mainly the development of a novel Bayesian model that integrates genomic, epidemiological, and travel data to estimate importation probabilities. The results showed insights into malaria transmission dynamics, particularly identifying importation sources and differences in importation rates in Mozambique. Finally, the relevance of the findings is to suggest interventions focusing on the traveler population to support efforts for malaria elimination.
Weaknesses:
The study also has some limitations, although the authors have plans to address them. The sample collection was not representative of some provinces, and not all samples had sufficient metadata for the risk factor analysis. Additionally, the authors used a proxy for transmission intensity and assumed some other conditions to calculate the importation probability for specific scenarios. They plan to conduct a new sample collection and include monthly malaria incidence estimates in the future.
Comments on revisions:
- Delete "We added this text to the discussion" in line 302 (Discussion)<br /> - I recommend adding the plans to address limitations indicated in the Response to Reviewers document in the Discussion. This would really strengthen the limitation section.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this study Hammond et al. investigated the role of Dual-specificity Tyrosine Phosphorylation regulated Kinase 1A (DYRK1) in G1/S transition. By exploiting Dependency Map portal, they identified a previously unexplored protein FAM53C as potential regulator of G1/S transition. Using RNAi, they confirmed that depletion of FAM53C suppressed proliferation of human RPE1 cells and that this phenotype was dependent on the presence protein RB. In addition, they noted increased level of CDKN1A transcript and p21 protein that could explain G1 arrest of FAM53C-depleted cells but surprisingly, they did not observe activation of other p53 target genes. Proteomic analysis identified DYRK1 as one of the main interactors of FAM53C and the interaction was confirmed in vitro. Further, they showed that purified FAM53C blocked the ability of DYRK1 to phosphorylate cyclin D in vitro although the activity of DYRK1 was likely not inhibited (judging from the modification of FAM53C itself). Instead, it seems more likely that FAM53C competes with cyclin D in this assay. Authors claim that the G1 arrest caused by depletion of FAM53C was rescued by inhibition of DYRK1 but this was true only in cells lacking functional p53. This is quite confusing as DYRK1 inhibition reduced the fraction of G1 cells in p53 wild type cells as well as in p53 knock-outs, suggesting that FAM53C may not be required for regulation of DYRK1 function. Instead of focusing on the impact of FAM53C on cell cycle progression, authors moved towards investigating its potential (and perhaps more complex) roles in differentiation of IPSCs into cortical organoids and in mice. They observed a lower level of proliferating cells in the organoids but if that reflects an increased activity of DYRK1 or if it is just an off-target effect of the genetic manipulation remains unclear. Even less clear is the phenotype in FAM53C knock-out mice. Authors did not observe any significant changes in survival nor in organ development but they noted some behavioral differences. Weather and how these are connected to the rate of cellular proliferation was not explored. In the summary, the study identified previously unknown role of FAM53C in proliferation but failed to explain the mechanism and its physiological relevance at the level of tissues and organism. Although some of the data might be of interest, in current form the data is too preliminary to justify publication.
Major comments:
(1) Whole study is based on one siRNA to Fam53C and its specificity was not validated. Level of the knock down was shown only in the first figure and not in the other experiments. The observed phenotypes in the cell cycle progression may be affected by variable knock-down efficiency and/or potential off target effects.
(2) Experiments focusing on the cell cycle progression were done in a single cell line RPE1 that showed a strong sensitivity to FAM53C depletion. In contrast, phenotypes in IPSCs and in mice were only mild suggesting that there might be large differences across various cell types in the expression and function of FAM53C. Therefore, it is important to reproduce the observations in other cell types.
(3) Authors state that FAM53C is a direct inhibitor of DYRK1A kinase activity (Line 203), however this model is not supported by the data in Fig 4A. FAM53C seems to be a good substrate of DYRK1 even at high concentrations when phosphorylations of cyclin D is reduced. It rather suggests that DYRK1 is not inhibited by FAM53C but perhaps FAM53C competes with cyclin D. Further, authors should address if the phosphorylation of cyclin D is responsible for the observed cell cycle phenotype. Is this Cyclin D-Thr286 phosphorylation, or are there other sites involved?
(4) At many places, information on statistical tests is missing and SDs are not shown in the plots. For instance, what statistics was used in Fig 4C? Impact of FAM53C on cyclin D phosphorylation does not seem to be significant. IN the same experiment, does DYRK1 inhibitor prevent modification of cyclin D?
(5) Validation of SM13797 compound in terms of specificity to DYRK1 was not performed.
(6) A fraction of cells in G1 is a very easy readout but it does not measure progression through the G1 phase. Extension of the S phase or G2 delay would indirectly also result in reduction of the G1 fraction. Instead, authors could measure the dynamics of entry to S phase in cells released from a G1 block or from mitotic shake off.
Comments to the revised manuscript:
In the revised version of the manuscript, authors addressed most of the critical points. They now include new data with depletion of FAM53C using single siRNAs that show small but significant enrichment of population of the G1 cells. This G1 arrest is likely caused by a combined effects on induction of p21 expression and decreased levels of cyclin D1. Authors observed that inhibition of DYRK1 rescued cyclin D1 levels in FAM53 depleted cells suggesting that FAM53C may inhibit DYRK1. This possibility is also supported by in vitro experiments. On the other hand, inhibition of DYRK1 did not rescue the G1 arrest upon depletion of FAM53C, suggesting that FAM53C may have also DYRK1-independent role in G1. Functional rescue experiments with cyclin D1 mutants and detection of DYRK1 activity in cells would be necessary to conclusively explain the function of FAM53C in progression through G1 phase but unfortunately these experiments were technically not possible. Knock out of FAM53C in iPSCs and in mice suggest that FAM53C may have additional functions besides the cell cycle control and/or that adaptation may have occurred in these model systems. Overall, the study implicated FAM53C in fine tuning DYRK1 activity in cells that may to some extent influence the progression through G1 phase. In addition, FAM53C may also have DYRK1 and cell cycle independent functions that remain to be addressed by future studies.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In the manuscript of Cotten et al., the authors study the 2-thiolation of tRNA in bacterial antibiotic resistance. The wildtype organism, Yersinia pseudotuberculosis, downregulates 2-thiolation as a response to antibiotics targeting the ribosome. In this manuscript, the authors show that a knockout of tusB causes slower translation. They provide evidence on the mechanisms of the slowing by determining transcription and translation, ribosome profiling and performing codon-usage analysis. They successfully determined that 2 codons are drivers of the translation slowdown, and the data is highly conclusive. Technically, I have nothing to criticize.
Strengths:
All in all, the study is very well made, and the writing is clear and concise. It covers a wide array of state-of-the-art analyses to unravel the interplay of tRNA modifications in translation.
Weaknesses:
The only question that remains to be asked is why the slowed translation leads to a better survival of the bacteria under antibiotic stress. In my opinion, the mechanism itself remains unclear. Thus, the statement that "We expect that this reduction in ribosomal proteins is globally reducing the translational capacity of the cell and is responsible for inducing tolerance to ribosome and RNA polymerase-targeting antibiotics" does not truly emphasize the remaining open question of why slowed translation favors survival. Therefore, I would recommend a minor text revision.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
This important study shows that basement membrane (BM) generation is a key event mediating cell 3D organization in response to microenvironmental cues. Such a mechanism participates in the endothelial cell capacity to organize into a capillary vessel segment through the shift of interactions with the interstitial ECM to interactions with vascular BM. This is particularly important for the developing, sprouting vasculature. The authors conclusively show, using TIRF and atomic force microscopy substantiated by 3D sprouting assays, that the lysyl oxidase Loxl2 plays a key role herein. With respect to translation into clinical practice, the dysregulation of adherens junctions and barrier properties associated with Loxl2 dysfunction mediated defects in BM supports its involvement in the progression of long-term microvascular diseases.
An outstanding question not answered in the current MS is how Loxl2 integrates into the Dll4-Notch mediated control of tip-stalk-phalanx cell differentiation in the developing (embryonic) vasculature. The authors focused a lot on Loxl2 loss of function; however, in a (patho)physiological context, Loxl2 gain of function would be relevant. Loxl2 is a hypoxia target and Loxl2 accumulates in the ECM upon hypoxic stress (as occurs during ischemic CVD, stroke/heart infarct). It would be interesting to know how Loxl2 gain-of-function impacts BM assembly, endothelial behavior, mechanosensing, and vessel angiogenic remodeling.
-
-
-
Reviewer #3 (Public review):
Summary:
This manuscript investigates how mutations in the SARS-CoV-2 nucleocapsid protein (N) alter ribonucleoprotein (RNP) assembly, stability, and viral fitness. The authors focus on mutations such as P13L, G214C, G215C combining biophysical assays (SV-AUC, mass photometry, CD spectroscopy, EM), VLP formation, and reverse genetics. They propose that SARS-CoV-2 exploits "fuzzy complex" principles, where distributed weak interfaces in disordered regions allow both stability and plasticity, with measurable consequences for viral replication.
Strengths:
* The paper demonstrates a comprehensive integration of structural biophysics, peptide/protein assays, VLP systems, and reverse genetics.
* Identification of both de novo (P13L) and stabilizing (G214C/G215C) interfaces provides a mechanistic insight into RNP formation.
* Strong application of the "fuzzy complex" framework to viral assembly, showing how weak/disordered interactions support evolvability, is a significant conceptual advance in viral capsid assembly.
* Overall, the study provides a mechanistic context for mutations that have arisen in major SARS-CoV-2 variants (Omicron, Delta, Lambda) and a mechanistic basis for how mutations influence phenotype via altered biomolecular interactions.
Weaknesses:
The weaknesses are shared via detailed comments to follow.
Comments on revisions:
The authors have addressed the criticisms of the original manuscript satisfactorily.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this manuscript the authors examine the processing stages involved in perceptual decision-making using a new approach to analysing EEG data, combined with a critical stimulus manipulation. This new EEG analysis method enables single-trial estimates of the timing and amplitude of transient changes in EEG time-series recurrent across trials in a behavioural task. The authors find evidence for five events between stimulus onset and the response in a two-spatial-interval visual discrimination task. By analysing the timing and amplitude of these events in relation to behaviour and the stimulus manipulation, the authors interpret these events as related to separable processing stages for stimulus encoding (first two events), attention orientation (second event), motor planning (fourth event) and decision (deliberation, final event). This is largely consistent with previous findings from both event-related potentials (across trials) and single-trial estimates using decoding techniques and neural network approaches. However, by taking a data-driven approach (as opposed to theory-driven decoding analyses) a more nuanced picture emerges: there are several stimulus encoding steps which may contribute differently to behaviour, and decision processes extend beyond the planning of the motor response.
Strengths:
This work is not only important for the conceptual advance, but also in promoting this new analysis technique, which will likely prove useful in future research. For the broader picture, this work is an excellent example of the utility of neural measures for mental chronometry.
Weaknesses:
Though beyond the scope of this manuscript, these results should be considered within the broader decision-making literature, where task or domain-specific processes may not generalise (for example, in value-based decision-making).
-
-
arxiv.org arxiv.org
-
Reviewer #3 (Public review):
This manuscript explores whether high-definition transcranial direct current stimulation (HD-tDCS) of the left DLPFC can reduce real-world procrastination, as predicted by the Temporal Decision Model (TDM). The research question is interesting, and the topic - neuromodulation of self-regulatory behavior - is timely.
However, the study also suffers from a limited sample size, and sometimes it was difficult to follow the statistics.
The preregistration and ecological design (ESM) are commendable, but I was not able the find the preregistration, as reported in the paper.
Overall, the paper requires substantial clarification and tightening.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
The goal of the work is to establish the linkage between the spatial transcription factors (STFs) that function transiently to establish the identities of the individual NBs and the terminal selector genes (typically homeodomain genes) that appear in the newborn post-mitotic neurons. How is the identity of the NB maintained and carried forward after the spatial genes have faded away? Focusing on a single neuroblast (NB 7-1), the authors present evidence that the fork-head transcription factor, fd4, provides a bridge linking the transient spatial cues that initially specified neuroblast identity with the terminal selector genes that establish and maintain the identity of the stem cell's progeny.
The study is systematic, concise, and takes full advantage of 40+ years of work on the molecular players that establish neuronal identities in the Drosophila CNS. In the embryonic VNC, fd4 is expressed only in the NB 7-1 and its lineage. They show that Fd4 appears in the NB while the latter is still expressing the Spatial Transcription Factors and continues after the expression of the latter fades out. Fd4 is maintained through the early life of the neuronal progeny but then declines as the neurons turn on their terminal selector genes. Hence, fd4 expression is compatible with it being a bridging factor between the two sets of genes.
Experimental support for the "bridging" role of Fd4 comes from a set of loss-of-function and gain-of-function manipulations. The loss of function of Fd4, and the partially redundant gene Fd5, from lineage 7-1 does not affect the size of the lineage, but terminal markers of late-born neuronal phenotypes, like Eve and Dbx, are reduced or missing. By contrast, ectopic expression of fd4, but not fd5, results in ectopic expression of the terminal markers eve and Dbx throughout diverse VNC lineages.
A detailed test of fd4's expression was then carried out using lineages 7-3 and 5-6, two well-characterized lineages in Drosophila. Lineage 7-3 is much smaller than 7-1 and continues to be so when subjected to fd4 misexpression. However, under the influence of ectopic Fd4 expression, the lineage 7-3 neurons lost their expected serotonin and corazonin expression and showed Eve expression as well as motoneuron phenotypes that partially mimic the U motoneurons of lineage 7-1.
Ectopic expression of Fd4 also produced changes in the 5-6 lineage. Expression of apterous, a feature of lineage 5-6, was suppressed, and expression of the 7-1 marker, Eve, was evident. Dbx expression was also evident in the transformed 5-6 lineages, but extremely restricted as compared to a normal 7-1 lineage. Considering the partial redundancy of fd4 and fd5, it would have been interesting to express both genes in the 5-6 lineage. The anatomical changes that are exhibited by motoneurons in response to Fd4 expression confirm that these cells do, indeed, show a shift in their cellular identity.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
The study by Yadav et al. describes a new setup to quantify a number of aggression and mating behaviors in Drosophila melanogaster. The investigation of these behaviors requires the analysis of large number of videos to identify each kind of behavior displayed by a fly. Several approaches to automatize this process have been published before, but each of them has their limitations. The authors set out to develop a new setup that includes a very low-cost, easy to acquire hardware and open-source machine-learning classifiers to identify and quantify the behavior.
Strengths:
(1) The study demonstrates that their cheap, simple, and easy to obtain hardware works just as well as custom-made, specialized hardware for analyzing aggression and mating behavior. This enables the setup to be used in a wide range of settings, from research with limited resources to classroom teaching.
(2) The authors used previously published software to train new classifiers for detecting a range of behaviors related to aggression and mating and make them freely available. The classifiers are very positively benchmarked against a manually acquired ground-truth as well as existing algorithms.
(3) The study demonstrates the applicability of the setup (hardware and classifiers) to common methods in the field by confirming a number of expected phenotypes with their setup.
Taken together, this work can greatly facilitate research of aggression and mating in Drosophila. The combination of low-cost, off-the-shelf hardware and open-source, robust software enables researchers with very little funding or technical expertise to contribute to the scientific process, and also allows large-scale experiments, for example, in classroom teaching with many students, or for systematic screenings.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this study, Tang, Yu & colleagues investigate the impact of continuous flash suppression (CFS) on the responses of V1 neurons using 2-photon calcium imaging. The report that CFS substantially suppressed V1 orientation responses. This suppression happens in a graded fashion depending on the binocular preference of the neuron: neurons preferring the eye that was presented with the marker stimuli were most suppressed, while the neurons preferring the eye to which the grating stimuli were presented were least suppressed. The binocular neuron exhibited an intermediate level of suppression.
Strengths:
The imaging techniques are cutting-edge, and the imaging results are convincing and consistent across animals.
Weaknesses:
I am not totally convinced by the conclusions that the authors draw based on their machine learning models.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This manuscript presents a series of experiments that further investigate the roles of the BLA and PRH in sensory preconditioning, with a particular focus on understanding their differential involvement in the association of S1 and S2 with shock.
Strengths:
The motivation for the study is clearly articulated, and the experimental designs are thoughtfully constructed. I especially appreciate the inclusion of Table 1, which makes the designs easy to follow. The results are clearly presented, and the statistical analyses are rigorous.
During the revision, the authors have adequately addressed my minor suggestions from the original version.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This is a very interesting paper that documents how humans use a variety of factors that penalize model complexity and integrate over a possible set of parameters within each model. By comparison, trained neural networks also use these biases, but only on tasks where model selection was part of the reward structure. In the situation where training emphasizes maximum-likelihood decisions, only neural networks, but not humans, were able to adapt their decision-making. Humans continue to use model integration simplicity biases.
Strengths:
This study used a pre-registered plan for analyzing human data, which exceeds the standards compared to other current studies.
The results are technically correct.
Weaknesses:
The presentation of the results could be improved.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This study investigates two main questions:
(1) whether brain activity recorded during immersive virtual reality can differentiate facial expressions and stereoscopic depth, and
(2) whether depth cues modulate facial information processing.
The results show that both expression and depth information can be decoded from multivariate EEG recorded in a head-mounted VR setup. However, the results show that the decoding performance of facial expressions does not benefit from depth information.
Strengths:
The study is technically strong and well executed. EEG data are of high quality despite the challenges of recording inside a head-mounted VR system. The work effectively combines stereoscopic stimulus presentation, eye-tracking to monitor gaze behavior, and time-resolved multivariate decoding techniques. Together, these elements provide an exemplary demonstration of how to collect and analyze high-quality EEG data in immersive VR environments.
Weaknesses:
The major limitation concerns the theoretical question about how stereoscopic depth modulates facial expression processing. While previous work has suggested that stereoscopic depth cues can shape natural face perception and emphasize the importance of binocular information in recognizing facial expressions (lines 95-97), the present study reports a null effect of depth. However, the stimulus configuration they used likely constrained the ability to detect any depth-related effects. All facial stimuli were static, frontal, and presented at a fixed distance. This design leads to near-ceiling behavioral performance and no behavioral effect of depth on expression recognition. It makes the null modulation of depth on expression processing unsurprising and limits the theoretical reach of the study. Adding more subtle or naturalistic features (such as various viewing angles and dynamic expressions) to the stimulus set if the authors aim to advance a strong theoretical claim about the role of binocular disparity. Or reframing the work as a technical validation of EEG decoding in this context.
Another issue relates to the claim that eye movements cannot explain the EEG decoding results. It is a real challenge to remove eye-movement-related artifacts and confounds, as the VR setup tends to encourage viewers to explore the environment freely. However, nearly half of the eye-tracking datasets were lost (usable in only 17 of 33 participants), which substantially weakens the evidence for EEG-gaze dissociation. Moreover, it would be almost impossible to decode facial information from only two-dimensional gaze direction, given that with 60 EEG channels, the decoding accuracy was modest (AUC ≈ 0.60). These two factors together limited the strength of the reported null correlation between neural and eye-data decoding.
The decoding analysis appears to use all 60 EEG channels as input features. I wonder why the authors did not examine using more spatially specific channel subsets. Facial expression and depth cues are known to preferentially engage occipito-temporal regions (e.g., N170-related sites), yet the current approach treats all sensors equally. Including all the channels may add noise and irrelevant signals to facial information decoding. Besides, using a subset of spatial-specific channels would align more directly with the subsequent source reconstruction.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This paper explores how spatial attention affects foveal information processing across different spatial frequencies. The results indicate that exogenously directed attention enhances contrast sensitivity for low- to mid-range spatial frequencies (4-8 CPD), with no significant benefits for higher spatial frequencies (12-20 CPD). However, asymptotic performance increased as a result of spatial attention independently of spatial frequency.
Strengths:
The strengths of this article lie in its methodological approach, which combines a psychophysical experiment with precise control over the information presented in the foveola.
Weaknesses:
The authors acknowledge that they used the standard approach of analyzing observer-averaged data, but recognize that this method has limitations: it ignores the uncertainty associated with parameter estimates and the relationships between different parameters of the psychometric model. This may affect the interpretation of attentional effects. In the future, mixed-effects models at the trial level could overcome these limitations.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
In this study, Wu & Turrigiano investigate an ethologically relevant form of associative learning (conditioned taste aversion - CTA) and its extinction in the Shank3 KO mouse model of ASD. They also examine the underlying circuits in the anterior insular cortex (AIC) simultaneously, using two-photon calcium imaging through a GRIN lens. They report that Shank3 KO mice learn CTA slower and suggest that this is mediated by a reduction in tastant-stimulus activity suppression of AIC neurons and a reduced signal-to-noise ratio due to increased noise correlations in AIC neurons. Interestingly, once Shank3 KO mice acquire CTA, they extinguish the aversive memory more rapidly than wild-type mice. This accelerated extinction is accompanied by a faster loss of neuronal and population-level taste selectivity and coding in the AIC compared to WT mice.
This is an important study that uses in vivo methods to assess circuit dysfunction in a mouse model of ASD, related to sensory perception valence (in this case, taste). The study is well executed, the data are of high quality, and the analytical procedures are detailed. Furthermore, the behavioural paradigm is well thought out, particularly the approach for assessing extinction through repeated retrieval sessions (T1-T5), which effectively tests discrimination between saccharin and water rather than relying solely on lick counts or total consumption as a measure of extinction. Finally, the statistical tests used are appropriate and justified.
There is, however, a missing link between the behavioural findings and the underlying mechanisms. More specifically:
(1) The authors don't make a causal link between the behaviour and AIC neurophysiology, both the percentage of suppressed cells and the coactivity measurements. For the % of suppressed cells, it seems that both WT and KO cells are suppressed in the transition between CST1 and CST2 (Figure 1L), yet only the WT mice exhibit CTA (at least by CST2). For the taste-elicited coactivity measure, it seems that there is an increase in coactivity from CST1 to CST2 in WT (Figure 2C - blue, although not statistically tested?), but persistently higher coactivity in KO. Is this change of coactivity in WT important for the expression of CTA? Plotting behavioral performance (from Figure 1G) against coactivity (from Figure 2C) for each animal would be informative.
(2) Shank3 KO cells already show an increase in baseline coactivity (Figure 2- figure supplement 1), and the authors never examine CS-only responses in the KO group, therefore making it difficult to determine whether elevated coactivity and noise correlations reflect a generalized AIC abnormality in Shank3 KOs (perhaps through impaired PV-mediated inhibition in insular cortex - Gogolla et al, 2014) that is not directly responsible/related to CTA?
(3) How do the authors interpret the large range of lick ratios (Figure 1G) for WT (almost bi-modal distribution)? Is there a within-subject correlation with any of the neurophysiological measurements to suggest a relationship between AIC neurophysiology and behavioural expression of CTA?
(4) Indeed, CTA appears to be successfully achieved for Shank3 KO mice delayed by 1 day, as the level of saccharin aversion during the first retrieval session (T1) is comparable between Shank3 KO and WTs. In this context, not extending the first part of the paradigm to include CST3 seems to be a missed opportunity. Doing so would have allowed for within-cell and within-subject comparison of taste-elicited pairwise correlation across the learning and to investigate the neural mechanism of delayed extinction in KOs more effectively.
(5) How to interpret Figure 5F: Absolute discriminability is lower for T5 for CTA WT and CTA KO compared to CS-only? Why would AIC neurons have less information on taste identity by the end of extinction than during the unconditioned (CS-only) condition? And if that is the case, how is decoding accuracy in Figure 6C higher in T5 for CTA WT vs CS-only?
-
-
www.amazon.com www.amazon.com
-
https://www.amazon.com/MicroSpark-Heavyweight-Checklist-Strategy-Tinplate/dp/B0DHYKG6VT?th=1 MicroSpark 3x5" index card product with holder and tin can
Tags
Annotators
URL
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Central pattern generator (CPG) circuits underly rhythmic motor behaviors. Till date, it is thought that these CPG networks are rather local and multiple CPG circuits are serially connected to allow locomotion across the entire body. Distributed CPG networks that incorporate long-range connections have not been proposed although such connectivity has been experimentally shown for several different spinal populations. In this manuscript, the authors use this existing literature on long-range spinal interneuron connectivity to build a new computational model that reproduces basic features of locomotion like left-right alternation, rostrocaudal propagation and independent control of frequency and amplitude. Interestingly, the authors show that a model solely based on inhibitory neurons can recapitulate these basic locomotor features. Excitatory sources were then added that increased the dynamic range of frequencies generated. Finally, the authors were also able to reproduce experimentally observed consequences of cell-type-specific ablations showing that local and long range, cell-type-specific connectivity could be sufficient for generating locomotion.
Strengths:
This work is novel, providing an interesting alternative of distributed CPGs to the local networks traditionally predicted. It shows cell type-specific network connectivity is as important if not more than intrinsic cell properties for rhythmogenesis and that inhibition plays a crucial role in shaping locomotor features. Given the importance of local CPGs in understanding motor control, this alternative concept will be of broad interest to the larger motor control field including invertebrate and vertebrate species.
Weaknesses:
The main weaknesses were addressed in the revision.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
In this manuscript, Davis and colleagues aimed to identify the molecular sensors and signaling cascade that enable collecting lymphatic vessels to increase their spontaneous contraction frequency in response to intraluminal pressure (pressure-induced chronotropy). They tested whether the process is similar to blood vessel myogenic constriction by relying on cation channels (TRPC6, TRPM4, PKD2, PIEZO1, etc.) or instead require the activation of G-protein-coupled receptors (presumably mechanosensitive GNAQ/GNA11-coupled receptors), using ex vivo pressure myography of mouse popliteal lymphatics, smooth muscle-specific conditional knockouts, quantitative PCR validation, and single-cell RNA sequencing for target prioritization. The authors convincingly demonstrate that pressure-induced chronotropy does not require the cation channels implicated in arterial myogenic tone but is blunted by deletion of GNAQ/GNA11 or IP3 receptor 1, supporting a model of GPCR > IP3 > Ca2+ release > Cl⁻ channel activation > depolarization. The core conclusion is robust. The work redefines lymphatic pacemaking as G-protein-coupled receptor-dependent mechanotransduction, distinct from arterial mechanisms, and provides a genetically validated toolkit that is useful for studying lymphatic function and dysfunction.
Strengths:
(1) The data are of high quality and highly sensitive functional readouts
(2) The systematic genetic targeting is a major strength that overcomes pharmacological artifacts
(3) Careful quantitative analyses of frequency-pressure slopes
Weaknesses:
(1) The use of inguinal-axillary vessels for single-cell RNA sequencing rather than the popliteal segment studied functionally.
(2) No direct testing of the specific G-protein-coupled receptor involved.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors investigate deficiencies in various immune responses, and also the prtA toxin's role in OMV toxicity. Some key interpretations are that the Imd pathway contributes to preventing OMV toxicity, but not Toll, and that Hayan and Eater somehow mediate OMV or PrtA toxicity. This descriptive effort is a solid set of experiments, although some experimental results may require further validation.
Strengths:
The breadth of experiments tests multiple immune parameters, providing a systematic effort that ensures a number of potentially relevant interactions can be recovered. Certain findings, such as the PrtA toxicity to flies, appear solid, and some interesting findings regarding Hayan and eater will be of interest to the fly immunity field.
Weaknesses:
It appears almost all results rely on the use of a single mutant representing the deletion of the gene. It's not clear if the mutations are always in the same genetic background, but this can be clarified. There are a couple of results that are confusing and may be internally contradicting, and should be additionally validated and clarified.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
This study is a part of the ongoing series of rigorous work from this group exploring neural coding deficits in the auditory nerve, and dissociating the effects of cochlear synaptopathy from other age-related deficits. They have previously shown no evidence of phase-locking deficits in the remaining auditory nerve fibers in quiet-aged gerbils. Here, they study the effects of aging on the perception and neural coding of temporal fine structure cues in the same Mongolian gerbil model.
They measure TFS coding in the auditory nerve using the TFS1 task which uses a combination of harmonic and tone-shifted inharmonic tones which differ primarily in their TFS cues (and not the envelope). They then follow this up with a behavioral paradigm using the TFS1 task in these gerbils. They test young normal hearing gerbils, aged gerbils, and young gerbils with cochlear synaptopathy induced using the neurotoxin ouabain to mimic synapse losses seen with age.
In the behavioral paradigm, they find that aging is associated with decreased performance compared to the young gerbils, whereas young gerbils with similar levels of synapse loss do not show these deficits. When looking at the auditory nerve responses, they find no differences in neural coding of TFS cues across any of the groups. However, aged gerbils show an increase in the representation of periodicity envelope cues (around f0) compared to young gerbils or those with induced synapse loss. The authors hence conclude that synapse loss by itself doesn't seem to be important for distinguishing TFS cues, and rather the behavioral deficits with age are likely having to do with the misrepresented envelope cues instead.
The manuscript is well written, and the data presented are robust. Some of the points below will need to be considered while interpreting the results of the study, in its current form. These considerations are addressable if deemed necessary, with some additional analysis in future versions of the manuscript.
Spontaneous rates - Figure S2 shows no differences in median spontaneous rates across groups. But taking the median glosses over some of the nuances there. Ouabain (in the Bourien study) famously affects low spont rates first, and at a higher degree than median or high spont rates. It seems to be the case (qualitatively) in figure S2 as well, with almost no units in the low spont region in the ouabain group, compared to the other groups. Looking at distributions within each spont rate category and comparing differences across the groups might reveal some of the underlying causes for these changes. Given that overall, the study reports that low-SR fibers had a higher ENV/TFS log-z-ratio, the distribution of these fibers across groups may reveal specific effects of TFS coding by group.
[Update: The revised manuscript has addressed these issues]
Threshold shifts - It is unclear from the current version if the older gerbils have changes in hearing thresholds, and whether those changes may be affecting behavioral thresholds. The behavioral stimuli appear to have been presented at a fixed sound level for both young and aged gerbils, similar to the single unit recordings. Hence, age-related differences in behavior may have been due to changes in relative sensation level. Approaches such as using hearing thresholds as covariates in the analysis will help explore if older gerbils still show behavioral deficits.
[Update: The issue of threshold shifts with aging gerbils is still unresolved in my opinion. From the revised manuscript, it appears that aged gerbils have a 36dB shift in thresholds. While the revised manuscript provides convincing evidence that these threshold shifts do not affect the auditory nerve tuning properties, the behavioral paradigm was still presented at the same sound level for young and aged animals. But a potential 36 dB change in sensation level may affect behavioral results. The authors may consider adding thresholds as covariates in analyses or present any evidence that behavioral thresholds are plateaued along that 30dB range].
Task learning in aged gerbils - It is unclear if the aged gerbils really learn the task well in two of the three TFS1 test conditions. The d' of 1 which is usually used as the criterion for learning was not reached in even the easiest condition for aged gerbils in all but one condition for the aged gerbils (Fig. 5H) and in that condition, there doesn't seem to be any age-related deficits in behavioral performance (Fig. 6B). Hence dissociating the inability to learn the task from the inability to perceive TFS 1 cues in those animals becomes challenging.
[Update: The revised manuscript sufficiently addresses these issues, with the caveat of hearing threshold changes affecting behavioral thresholds mentioned above].
Increased representation of periodicity envelope in the AN - the mechanisms for increased representation of periodicity envelope cues is unclear. The authors point to some potential central mechanisms but given that these are recordings from the auditory nerve what central mechanisms these may be is unclear. If the authors are suggesting some form of efferent modulation only at the f0 frequency, no evidence for this is presented. It appears more likely that the enhancement may be due to outer hair cell dysfunction (widened tuning, distorted tonotopy). Given this increased envelope coding, the potential change in sensation level for the behavior (from the comment above), and no change in neural coding of TFS cues across any of the groups, a simpler interpretation may be -TFS coding is not affected in remaining auditory nerve fibers after age-related or ouabain induced synapse loss, but behavioral performance is affected by altered outer hair cell dysfunction with age.
[Update: The revised manuscript has addressed these issues]
Emerging evidence seems to suggest that cochlear synaptopathy and/or TFS encoding abilities might be reflected in listening effort rather than behavioral performance. Measuring some proxy of listening effort in these gerbils (like reaction time) to see if that has changed with synapse loss, especially in the young animals with induced synaptopathy, would make an interesting addition to explore perceptual deficits of TFS coding with synapse loss.
[Update: The revised manuscript has addressed these issues]
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This study investigates how various behavioral features are represented in the medial prefrontal cortex (mPFC) of rats engaged in a naturalistic foraging task. The authors recorded electrophysiological responses of individual neurons as animals transitioned between navigation, reward consumption, avoidance, and escape behaviors. Employing a range of computational and statistical methods, including artificial neural networks, dimensionality reduction, hierarchical clustering, and Bayesian classifiers, the authors sought to predict from neural activity distinct task variables (such as distance from the reward zone and the success or failure of avoidance behavior). The findings suggest that mPFC neurons alternate between at least two distinct functional modes, namely spatial encoding and threat evaluation, contingent on the specific location.
Strengths:
This study attempt to address an important question: understanding the role of mPFC across multiple dynamic behaviors. The authors highlight the diverse roles attributed to mPFC in previous literature and seek to explain this apparent heterogeneity. They designed an ethologically relevant foraging task that facilitated the examination of complex dynamic behavior, collecting comprehensive behavioral and neural data. The analyses conducted are both sound and rigorous.
Weaknesses:
Because the study still lacks experimental manipulation, the findings remain correlational. The authors have appropriately tempered their claims regarding the functional role of the mPFC in the task. The nature of the switch between functional modes encoding distinct task variables (i.e., distance to reward, and threat-avoidance behavior type) is not established. Moreover, the evidence presented to dissociate movement from these task variables is not fully convincing, particularly without single-session video analysis of movement. Specifically, while the new analyses in Figure 7 are informative, they may not fully account for all potential confounding variables arising from changes in context or behavior.
Comments on revisions:
The authors have addressed my previous recommendations.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The study provides an interesting contribution to our understanding of Cryptovaranoides relationships, which is a matter of intensive debate among researchers. The authors have modified the manuscript according to most of my suggestions. My main concerns are about the wording of some statements but the authors have the right to put it as they want in the end. Overall the discussion and data are well prepared. I would recommend to publish the manuscript after very minor revisions.
Strengths:
Detailed analysis of the discussed characters. Illustrations of some comparative materials.
Weaknesses:
Abstract: "Our team challenged this identification and instead suggested †Cryptovaranoides had unclear affinities to living reptiles"
Unfortunately I have to disagree again. "unclear affinities to living reptiles" can mean anything including a crown lizard. First, the 2023 paper clearly rejected the squamate hypothesis and presented some evidence that potentially places Cryptovaranoides among Archosauromorpha. In this context "unclear where it would belong within the latter" does not really matter. Second, we are not discussing here if Cryptovaranoides is a squamate or a stem-squamate. We have many more options on the table, so "unclear affinities" is too imprecise. Please change it to "could be an archosauromorph or an indeterminate neodiapsid" in the abstract to show the scale of conflicting evidence.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Ruppert et al. present a well-designed 2×2 factorial study directly comparing methionine restriction (MetR) and cold exposure (CE) across liver, iBAT, iWAT, and eWAT, integrating physiology with tissue-resolved RNA-seq. This approach allows a rigorous assessment of where dietary and environmental stimuli act additively, synergistically, or antagonistically. Physiologically, MetR progressively increases energy expenditure (EE) at 22{degree sign}C and lowers RER, indicating a lipid utilization bias. By contrast, a 24-hour 4 {degree sign}C challenge elevates EE across all groups and eliminates MetR-Ctrl differences. Notably, changes in food intake and activity do not explain the MetR effect at room temperature.
Strengths:
The data convincingly support the central claim: MetR enhances EE and shifts fuel preference to lipids at thermoneutrality, while CE drives robust EE increases regardless of diet and attenuates MetR-driven differences. Transcriptomic analysis reveals tissue-specific responses, with additive signatures in iWAT and CE-dominant effects in iBAT. The inclusion of explicit diet×temperature interaction modeling and GSEA provides a valuable transcriptomic resource for the field.
Comments on revisions:
The authors have addressed any concerns I had.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors developed an interesting novel paradigm to probe the effects of cerebellar climbing fiber activation on short-term adaptation of somatosensory neocortical activity during repetitive whisker stimulation. Normally, RWS potentiated whisker responses in pyramidal cells and weakly suppressed them in interneruons, lasting for at least 1h. Crusii Optogenetic climbing fiber activation during RWS reduced or inverted these adaptive changes. This effect was generally mimicked or blocked with chemogenetic SST or VIP activation/suppression as predicted based on their "sign" in the circuit.
Strengths:
The central finding about CF modulation of S1 response adaptation is interesting, important, and convincing, and provides a jumping-off point for the field to start to think carefully about cerebellar modulation of neocortical plasticity.
Weaknesses:
The SST and VIP results appeared slightly weaker statistically, but I do not personally think this detracts from the importance of the initial finding (if there are multiple underlying mechanisms, modulating one may reproduce only a fraction of the effect size). I found the suggestion that zona incerta may be responsible for the cerebellar effects on S1 to be a more speculative result (it is not so easy with existing technology to effectively modulate this type of polysynaptic pathway), but this may be an interesting topic for the authors to follow up on in more detail in the future.
Comments on revisions:
The authors have appropriately addressed my comments.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This important study combines in vitro and in vivo recording to determine how the firing of cortical and striatal neurons changes during a fever range temperature rise (37-40 oC). The authors found that certain neurons will start, stop, or maintain firing during these body temperature changes. The authors further suggested that the TRPV3 channel plays a role in maintaining cortical activity during fever.
Strengths:
The topic of how the firing pattern of neurons changes during fever is unique and interesting. The authors carefully used in vitro electrophysiology assays to study this interesting topic.
Weaknesses:
(1) In vivo recording is a strength of this study. However, data from in vivo recording is only shown in Fig 5A,B. This reviewer suggests the authors further expand on the analysis of the in vivo Neuropixels recording. For example, to show single spike waveforms and raster plots to provide more information on the recording. The authors can also separate the recording based on brain regions (cortex vs striatum) using the depth of the probe as a landmark to study the specific firing of cortical neurons and striatal neurons. It is also possible to use published parameters to separate the recording based on spike waveform to identify regular principal neurons vs fast-spiking interneurons. Since the authors studied E/I balance in brain slices, it would be very interesting to see whether the "E/I balance" based on the firing of excitatory neurons vs fast-spiking interneurons might be changed or not in the in vivo condition.
(2) The author should propose a potential mechanism for how TRPV3 helps to maintain cortical activity during fever. Would calcium influx-mediated change of membrane potential be the possible reason? Making a summary figure to put all the findings into perspective and propose a possible mechanism would also be appreciated.
(3) The author studied P7-8, P12-14, and P20-26 mice. How do these ages correspond to the human ages? it would be nice to provide a comparison to help the reader understand the context better.
Comments on revisions:
In this revised version, the authors nicely addressed my critiques. I have no more comments to make.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
The current paper investigates neural correlates of trust development in human-AI interaction, looking at EEG signatures locked to the moment that AI advice is presented. The key finding is that both human-response-locked EEG signatures (the CPP) and post-AI-advice signatures (N2, P3) are modulated by trust ratings. The study is interesting, however, it does have some clear and sometimes problematic weaknesses:
(1) The authors did not include "AI-advice". Instead, a manikin turned green or blue, which was framed as AI advice. It is unclear whether participants viewed this as actual AI advice.
(2) The authors did not include a "non-AI" control condition in their experiment, such that we cannot know how specific all of these effects are to AI, or just generic uncertain feedback processing.
(3) Participants perform the task at chance level. This makes it unclear to what extent they even tried to perform the task or just randomly pressed buttons. These situations likely differ substantially from a real-life scenario where humans perform an actual task (which is not impossible) and receive actual AI advice.
(4) Many of the conclusions in the paper are overstated or very generic.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary and Significance:
In this work, Cary and Hayashi address the important question of when, in evolution, certain mobile genetic elements (Ty3/gypsy-like non-LTR retrotransposons) associated with certain membrane fusion proteins (viral glycoprotein F or B-like proteins), which could allow these mobile genetic elements to be transferred between individual cells of a given host. It is debated in the literature whether the acquisition of membrane fusion proteins by non-LTR retrotransposons is a rather recent phenomenon that separately occurred in the ancestors of certain host species or whether the association with membrane fusion proteins is a much more ancient one, pre-dating the Cambrian explosion. Obviously, this question also touches upon the origin of the retroviruses, which can spread between individuals of a given host but seem restricted to vertebrates. Based on convincing data, Cary and Hayashi argue that an ancient association of non-LTR retrotransposons with membrane fusion proteins is most probable.
Strengths:
The authors take the smart approach to systematically retrieve apparently complete, intact, and recently functional Ty3/gypsy-like non-LTR retrotransposons that, next to their characteristic gag and pol genes, additionally carry sequences that are homologous to viral glycoprotein F (env-F) or viral glycoprotein B (env-B). They then construct and compare phylogenetic trees of the host species and individual encoded proteins and protein domains, where 3D-structure calculations and other features explain and corroborate the clustering within the phylogenetic trees. Congruence of phylogenetic trees and correlation of structural features is then taken as evidence for an infrequent recombination and a long-term co-evolution of the reverse transcriptase (encoded by the pol gene) and its respective putative membrane fusion gene (encoded by env-F or env-B). Importantly, the env-F and env-B containing retrotransposons do not form a monophyletic group among the Ty3/gypsy-like non-LTR retrotransposons, but are scattered throughout, supporting the idea of an originally ancient association followed by a random loss of env-F/env-B in individual branches of the tree (and rather rare re-associations via more recent recombinations).
Overall, this is valuable, stimulating, and important work of general and fundamental interest, but still also somewhat incompletely explored, imprecisely explained, and insufficiently put into context for a more general audience.
Weaknesses:
Some points that might be considered and clarified:
(1) Imprecise explanations, terms, and definitions:
It might help to add a 'definitions box' or similar to precisely explain how the authors decided to use certain terms in this manuscript, and then use these terms consistently and with precision.
a) In particular, these are terms such as 'vertebrate retrovirus' vs 'retrovirus' vs 'endogenized retrovirus' vs 'endogenous retrovirus' vs 'non-LTR retrotransposon' and 'Ty3/gypsi-like retrotransposon' vs 'Ty3/gypsy retrotransposon' vs 'errantivirus'.
b) The comment also applies to the term 'env' used for both 'env-F' and 'env-B', where often it remains unclear which of the two protein types the authors refer to. This is confusing, particularly in the methods, where the search for the respective homologs is described.
c) Other examples are the use of the entire pol gene vs. pol-RT for the definition of the Ty3/gypsy clade and for the generation of phylogenetic trees (Methods and Figure S1), and the names for various portions of pol that appear without prior definition or explanation (e.g., 'pro' in Figure 1A, 'bridge' in Figure S1C, 'the chromodomain' in the text and Figure 7).
d) It is unclear from the main text which portions of pol were chosen to define pol-RT and why. The methods name the 'palm-and-fingers', 'thumb', and 'connections' domains to define RT. In the main text, the 'connection' domain is called 'tether' and is instead defined as part of the 'bridge' region following RT, which is not part of RT.
(2) Insufficient broader context:
a) The introduction does not state what defines Ty3/gypsy non-LTR retrotransposons as compared to their closest relatives (Ty1/copia retrotransposons, BEL/pao retrotransposons, vertebrate retroviruses). This makes it difficult to judge the significance and generality of the findings.
b) The various known compositions of Ty3/gypsi-like retrotransposons are not mentioned and explained in the introduction (open reading frames, (poly-)proteins and protein domains, and their variable arrangement, enzymatic activities, and putative functions), and the distribution of Ty3/gypsi-like retrotransposons among eukaryotes remains unclear. The introduction does not mention that Ty3/gypsi-like retrotransposons apparently are absent from vertebrates, and Figure 7 is not very clear about whether or not it includes sequences from plants ('Chromoviridae').
c) The known association of Ty3/gypsi-like retrotransposons from different metazoan phyla with putative membrane fusion proteins (env-like) genes is mentioned in the introduction, but literature information, whether such associations also occur in the context of other retrotransposons (e.g., Ty1/ copia or BEL/pao), is not provided. The abstract is somewhat misleading in this respect. Finally, the different known types of env-like genes are not mentioned and explained as part of the introduction ('env-f', 'env-B', 'retroviral env', others?)
d) Some key references and reviews might be added:
- Pelisson, A. et al. (1994) https://www.embopress.org/doi/abs/10.1002/j.1460-2075.1994.tb06760.x<br /> (next to Song et al. (1994), for the identification of env in Ty3/gypsy)
- Boeke, J.D. et al. (1999)<br /> In Virus Taxonomy: ICTV VIIth report. (ed. F.A. Murphy),. Springer-Verlag, New York.<br /> (cited by Malik et al. (2000) - for the definition and first use of the term 'errantivirus')
- Eickbush, T.H. and Jamburuthugoda, V.K. (2008) https://doi.org/10.1016/j.virusres.2007.12.010<br /> (on the classification of retrotransposons and their env-like genes)
- Hayward, A. (2017) https://doi.org/10.1016/j.coviro.2017.06.006<br /> (on scenarios of env acquisition)
(3) Incomplete analysis:
a) Mobile genetic elements are sometimes difficult to assemble correctly from short-read sequencing data. Did the authors confirm some of their newly identified elements by e.g., PCR analysis or re-identification in long-read sequencing data?
b) The authors mention somewhat on the side that there are Ty3/gypsy elements with a different arrangement (gag-env-pol instead of gag-pol-env). Why was this important feature apparently not used and correlated in the analysis? How does it map on the RT phylogenetic tree? Which type of env is found with either arrangement? Is there evidence for a loss of env also in the case of gag-env-pol elements?
c) Sankey plots are insufficiently explained. How would inconsistencies between trees (recombinations) show up here? Why is there no Sankey plot for the analysis of env-B in Figure 5?
d) Why are there no trees generated for env-F and env-B like proteins, including closely related homologous sequences that do NOT come from Ty3/gypsy retrotransposons (e.g., from the eukaryotic hosts, from other types of retrotransposons (Ty1/copia or BEL/pao), from viruses such as Herpesvirus and Baculovirus)? It would be informative whether the sequences from Ty3/gypsy cluster together in this case.
e) Did the authors identify any other env-like ORFs (apart from env-F and env-B) among Ty3/gypsy retrotransposons? Did they identify other, non-env-like ORFs that might help in the analysis? It is not quite clear from the methods if the searches for env-F and env-B - containing Ty3/gypsy elements were done separately and consecutively or somehow combined (the authors generally use 'env', and it is not clear which type of protein this refers to).
f) Why was the gag protein apparently not used to support the analysis? Are there different, unrelated types of gag among non-LTR retrotransposons? Does gag follow or break the pattern of co-evolution between RT and env-F/env-B?
g) Data availability. The link given in the paper does not seem to work (https://github.com/RippeiHayashi/errantiviruses_2025/tree/main). It would be useful for the community to have the sequences of the newly identified Ty3/gypsy retrotransposons listed readily available (not just genome coordinates as in table S1), together with the respective annotations of ORFs and features.
-
- Nov 2025
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This study aims to develop and characterize phenylhydrazone-based small molecules that selectively activate the ATF6 arm of the unfolded protein response by covalently modifying a subset of ER-resident PDIs. The authors identify AA263 as a lead scaffold and optimize its structure to generate analogs with improved potency and ATF6 selectivity, notably AA263-20. These compounds are shown to restore proteostasis and functional expression of disease-associated misfolded proteins in cellular models involving both secretory (AAT-Z) and membrane (GABAA receptor) proteins. The findings provide valuable chemical tools for modulating ER proteostasis and may serve as promising leads for therapeutic development targeting protein misfolding diseases.
Strengths:
The study presents a well-defined chemical biology framework integrating proteomics, transcriptomics, and disease-relevant functional assays.
Identification and optimization of a new electrophilic scaffold (AA263) that selectively activates ATF6 represents a valuable advance in UPR-targeted pharmacology.
SAR studies are comprehensive and logically drive the development of more potent and selective analogs such as AA263-20.
Functional rescue is demonstrated in two mechanistically distinct disease models of protein misfolding-one involving a secretory protein and the other a membrane protein-underscoring the translational relevance of the approach.
Weaknesses:
ATF6 activation is primarily inferred from reporter assays and transcriptional profiling; direct biochemical evidence of ATF6 cleavage or nuclear translocation remains missing. However, the authors have added supporting data showing that co-treatment with the ATF6 inhibitor CP7 suppresses target gene induction, which partially strengthens the evidence for ATF6-dependent activity.
Although the proposed mechanism involving PDI modification and ATF6 activation is plausible, it is still not experimentally demonstrated and remains incompletely characterized.
In vivo validation is absent, and thus the pharmacological feasibility, selectivity, and bioavailability of these compounds in physiological systems remain untested.
Comments on revisions:
The authors have generally addressed my comments.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This is an important and well-conceived study that identifies the Bearded-type small protein E(spl)m4 as a physical and genetic interactor of TRAF4 in Drosophila. By combining classical genetics, yeast two-hybrid assays, and AlphaFold in silico modeling, the authors convincingly demonstrate that E(spl)m4 acts as an inhibitor of TRAF4-mediated induction of JNK-driven apoptosis in developing larval imaginal wing discs, while not affecting TRAF4's role in adherence junction remodeling.
Based primarily on modeling, the authors propose that the specificity of E(spl)m4 towards TRAF4-mediated signaling arises from its interference with TRAF4 trimerization, which is likely required for the activation of the JNK signaling arm but not for the maintenance of adherence junctions and stability of E-cadherin/β-catenin complex.
Overall, this study is of broad interest to cell and developmental biologists. It also holds potential biomedical relevance, particularly for strategies aimed at modulating TRAF protein activities to dissect and modulate canonical versus non-canonical signaling functions.
Strengths:
(1) The work identifies the Bearded-type small protein E(spl)m4 as a physical and genetic interactor of TRAF4 in Drosophila, extending the understanding of E(spl)m4 beyond its established functions in Notch signaling.
(2) The study is experimentally solid, well-executed, and written, combining classical genetics with protein-protein interaction assays and modeling to reveal E(spl)m4 as a new regulator of TRAF4 signaling.
(3) The genetic and biochemical data convincingly show the ability of E(spl)m4 overexpression to inhibit TRAF4-induced JNK-dependent apoptosis, while leaving the TRAF4 role in adherens junction remodeling unaffected.
(4) The findings have important implications for the regulation of cell signaling and apoptosis and may guide pharmacological targeting of TRAF proteins.
Weaknesses:
The study is overall strong; however, several aspects could be clarified or expanded to strengthen the proposed mechanism and data presentation:
(1) The proposed mechanism that E(spl)m4 inhibits TRAF4 activation of JNK signaling by affecting TRAF4 trimerization relies mainly on modeling. Experimental evidence would strengthen this claim. For example, a native or non-denaturing SDS-PAGE could be used to assess TRAF4 oligomerization states in the absence or presence of E(spl)m4 overexpression, testing whether E(spl)m4 interferes with high-molecular-weight TRAF4 assemblies.
(2) The study depends largely on E(spl)m4 overexpression, which may not reflect physiological conditions. It would be valuable to test, or at least discuss, whether loss-of-function or knockdown of E(spl)m4 modulates the strength or duration of JNK-mediated signaling, potentially accelerating apoptosis. Such data would reinforce the model that E(spl)m4 acts as a physiological modulator of TRAF4-JNK signaling in vivo.
(3) The authors initially identify both E(spl)m4 and E(spl)m2 as TRAF4 interactions, but subsequently focus on E(spl)m4. It would be helpful to clarify or discuss the rationale for prioritizing E(spl)m4 for detailed functional analysis.
(4) E(spl)m4 overexpression appears to protect RpS3 loser clones (Figure 6H-K), yet caspase-3-positive cells are still visible in mosaic wing discs. Please comment on the nature of these Caspase 3-positive cells, whether they are cell-autonomous to the clone or non-autonomous (Figure 6K)?
(5) This is a clear, well-executed, and conceptually strong study that significantly advances understanding of TRAF4 signaling specificity and its modulation by the Bearded-type protein E(spl)m4.
-
-
www.medrxiv.org www.medrxiv.org
-
Reviewer #3 (Public review):
Summary and strengths:
In this manuscript, Grimes presents an extension of the Ellipse of Insignificant (EOI) and Region of Attainable Redaction (ROAR) metrics to the meta-analysis setting as metrics for fragility and robustness evaluation of meta-analysis. The author applies these metrics to three meta-analyses of Vitamin D and cancer mortality, finding substantial fragility in their conclusions. Overall, I think extension/adaptation is a conceptually valuable addition to meta-analysis evaluation, and the manuscript is generally well-written.
Specific comments:
(1) The manuscript would benefit from a clearer explanation of in what sense EOIMETA is generalizable. The author mentions this several times, but without a clear explanation of what they mean here.
(2) The authors mentioned the proposed tools assume low between-study heterogeneity. Could the author illustrate mathematically in the paper how the between-study heterogeneity would influence the proposed measures? Moreover, the between-study heterogeneity is high in Zhang et al's 2022 study. It would be a good place to comment on the influence of such high heterogeneity on the results, and specifying a practical heterogeneity cutoff would better guide future users.
(3) I think clarifying the concepts of "small effect", "fragile result", and "unreliable result" would be helpful for preventing misinterpretation by future users. I am concerned that the audience may be confusing these concepts. A small effect may be related to a fragile meta-analysis result. A fragile meta-analysis doesn't necessarily mean wrong/untrustworthy results. A fragile but precise estimate can still reflect a true effect, but whether that size of true effect is clinically meaningful is another question. Clarifying the effect magnitude, fragility, and reliability in the discussion would be helpful.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In their manuscript entitled "Ubiquitous predictive processing in the spectral domain of sensory cortex", Sennesh and colleagues perform spectral analysis across multiple layers and areas in the visual system of mice. Their results are timely and interesting as they provide a complement to a study from the same lab focussed on firing rates, instead of oscillations. Together, the present study argues for a hypothesis called predictive routing, which argues that non-predictable stimuli are gated by Gamma oscillations, while alpha/beta oscillations are related to predictions.
Strengths:
(1) The study contains a clear introduction, which provides a clear contrast between a number of relevant theories in the field, including their hypotheses in relation to the present data set.
(2) The study provides a systematic analysis across multiple areas and layers of the visual cortex.
Weaknesses:
(1) It is claimed in the abstract that the present study supports predictive routing over predictive coding; however, this claim is nowhere in the manuscript directly substantiated. Not even the differences are clearly laid out, much less tested explicitly. While this might be obvious to the authors, it remains completely opaque to the reader, e.g., as it is also not part of the different hypotheses addressed. I guess this result is meant in contrast to reference 17, by some of the same authors, which argues against predictive coding, while the present work finds differences in the results, which they relate to spectral vs firing rate analysis (although without direct comparison).
(2) Most of the claims about a direction of propagation of certain frequency-related activities (made in the context of Figures 2-4) are - to the eyes of the reviewer - not supported by actual analysis but glimpsed from the pictures, sometimes, with very little evidence/very small time differences to go on. To keep these claims, proper statistical testing should be performed.
(3) Results from different areas are barely presented. While I can see that presenting them in the same format as Figures 2-4 would be quite lengthy, it might be a good idea to contrast the right columns (difference plots) across areas, rather than just the overall averages.
(4) Statistical testing is treated very generally, which can help to improve the readability of the text; however, in the present case, this is a bit extreme, with even obvious tests not reported or not even performed (in particular in Figure 5).
(5) The description of the analysis in the methods is rather short and, to my eye, was missing one of the key descriptions, i.e., how the CSD plots were baselined (which was hinted at in the results, but, as far as I know, not clearly described in the analysis methods). Maybe the authors could section the methods more to point out where this is discussed.
(6) While I appreciate the efforts of the authors to formulate their hypotheses and test them clearly, the text is quite dense at times. Partly this is due to the compared conditions in this paradigm; however, it would help a lot to show a visualization of what is being compared in Figures 2-4, rather than just showing the results.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The article explores the brain's ability to generalize information, with a specific focus on the entorhinal cortex (EC) and its role in learning and representing structural regularities that define relationships between entities in networks. The research provides empirical support for the longstanding theoretical and computational neuroscience hypothesis that the EC is crucial for structure generalization. It demonstrates that EC codes can generalize across non-spatial tasks that share common structural regularities, regardless of the similarity of sensory stimuli and network size.
Strengths:
At first glance, a potential limitation of this study appears to be its application of analytical methods originally developed for high-resolution animal electrophysiology (Samborska et al., 2022) to the relatively coarse and noisy signals of human fMRI. Rather than sidestepping this issue, however, the authors embrace it as a methodological challenge. They provide compelling empirical evidence and biologically grounded simulations to show that key generalization properties of entorhinal cortex representations can still be robustly detected. This not only validates their approach but also demonstrates how far non-invasive human neuroimaging can be pushed. The use of multiple independent datasets and carefully controlled permutation tests further underscores the reliability of their findings, making a strong case that structural generalization across diverse task environments can be meaningfully studied even in abstract, non-spatial domains that are otherwise difficult to investigate in animal models.
Weaknesses:
While this study provides compelling evidence for structural generalization in the entorhinal cortex (EC), several limitations remain that pave the way for promising future research. One issue is that the generalization effect was statistically robust in only one task condition, with weaker effects observed in the "community" condition. This raises the question of whether the null result genuinely reflects a lack of EC involvement, or whether it might be attributable to other factors such as task complexity, training order, or insufficient exposure possibilities that the authors acknowledge as open questions. Moreover, although the study leverages fMRI to examine EC representations in humans, it does not clarify which specific components of EC coding-such as grid cells versus other spatially tuned but non-grid codes-underlie the observed generalization. While electrophysiological data in animals have begun to address this, the human experiments do not disentangle the contributions of these different coding types. This leaves unresolved the important question of what makes EC representations uniquely suited for generalization, particularly given that similar effects were not observed in other regions known to contain grid cells, such as the medial prefrontal cortex (mPFC) or posterior cingulate cortex (PCC). These limitations point to important future directions for better characterizing the computational role of the EC and its distinctiveness within the broader network supporting learning and decision making based on cognitive maps.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
The authors use high throughput neutralisation data to explore how different summary statistics for population immune responses relate to strain success, as measured by growth rate during the 2023 season. The question of how serological measurements relate to epidemic growth is an important one, and I thought the authors present a thoughtful analysis tackling this question, with some clear figures. In particular, they found that stratifying the population based on the magnitude of their antibody titres correlates more with strain growth than using measurements derived from pooled serum data. The updated manuscript has a stronger motivation, and there is substantial potential to build on this work in future research.
Comments on revisions:
I have no additional recommendations. There are several areas where the work could be further developed, which were not addressed in detail in the responses, but given this is a strong manuscript as it stands, it is fine that these aspects are for consideration only at this point.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The stable production of learned vocalizations like human language and birdsong requires auditory feedback. What happens in the brain areas that generate stable vocalizations as performance deteriorates is not well understood. Using a species of songbird, the current study investigates individual cells within the evolutionarily-conserved brain regions that generate learned vocalizations to describe that the complement of neuropeptide (short proteins) signals may be a key feature of behavioral change. Because neuropeptides are important across species, these findings may help explain diminishing stability in learned behaviors even in humans.
Strengths:
The experiments are solid and follow a strong progression from description through manipulation. The songbird model is appropriate and powerful to inform on generalizable biological mechanisms of precisely learned behaviors, including human speech.
Weaknesses:
While it is always possible to perform more experiments, most of the weaknesses are in the presentation of the project, not in the evidence or analysis, which are leading-edge and appropriate. Generally, the ability to follow the findings and to independently assess rigor would be enhanced with increased explicit mention of the statistical thresholds and subjective descriptions. In addition, two prior pieces of relevant work seem to be omitted, including one performing deafening, gene expression measures, and behavioral assessment in zebra finches, and another describing neuropeptide complements in zebra finch singing nuclei based largely on mass spectrometry. The former in particular should be related to the current findings.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
This manuscript provides novel insights into altered glucose metabolism and KC status during early MASLD. The authors propose that hyperactivated glycolysis drives a spatially patterned KC depletion that is more pronounced than the loss of hepatocytes or hepatic stellate cells. This concept significantly enhances our understanding of early MASLD progression and KC metabolic phenotype.
Through a combination of TUNEL staining and MS-based metabolomic analyses of KCs from HFHC-fed mice, the authors show increased KC apoptosis alongside dysregulation of glycolysis and the pentose phosphate pathway. Using in vitro culture systems and KC-specific ablation of Chil1, a regulator of glycolytic flux, they further show that elevated glycolysis can promote KC apoptosis.
However, it remains unclear whether the observed metabolic dysregulation directly causes KC death or whether secondary factors, such as low-grade inflammation or macrophage activation, also contribute significantly. Nonetheless, the results, particularly those derived from the Chil1-ablated model, point to a new potential target for the early prevention of KC death during MASLD progression.
The manuscript is clearly written and thoughtfully addresses key limitations in the field, especially the focus on glycolytic intermediates rather than fatty acid oxidation. The authors acknowledge the missing mechanistic link between increased glycolysis and KC death. Still, several interpretations require moderation to avoid overstatement, and certain experimental details, particularly those concerning flow cytometry and population gating, need further clarification.
Strengths:
(1) The study presents the novel observation of profound metabolic dysregulation in KCs during early MASLD and identifies these cells as undergoing apoptosis. The finding that Chil1 ablation aggravates this phenotype opens new avenues for exploring therapeutic strategies to mitigate or reverse MASLD progression.
(2) The authors provide a comprehensive metabolic profile of KCs following HFHC diet exposure, including quantification of individual metabolites. They further delineate alterations in glycolysis and the pentose phosphate pathway in Chil1-deficient cells, substantiating enhanced glycolytic flux through 13C-glucose tracing experiments.
(3) The data underscore the critical importance of maintaining balanced glucose metabolism in both in vitro and in vivo contexts to prevent KC apoptosis, emphasizing the high metabolic specialization of these cells.
(4) The observed increase in KC death in Chil1-deficient KCs demonstrates their dependence on tightly regulated glycolysis, particularly under pathological conditions such as early MASLD.
Weaknesses:
(1) The novelty is questionable. The presented work has considerable overlap with a study by the same lab, which is currently under review (citation 17), and it should be considered whether the data should not be presented in one paper.
(2) The authors report that 60% of KCs are TUNEL-positive after 16 weeks of HFHC diet and confirm this by cleaved caspase-3 staining. Given that such marker positivity typically indicates imminent cell death within hours, it is unexpected that more extensive KC depletion or monocyte infiltration is not observed. Since Timd4 expression on monocyte-derived macrophages takes roughly one month to establish, the authors should consider whether these TUNEL-positive KCs persist in a pre-apoptotic state longer than anticipated. Alternatively, fate-mapping experiments could clarify the dynamics of KC death and replacement.
(3) The mechanistic link between elevated glycolytic flux and KC death remains unclear.
(4) The study does not address the polarization or ontogeny of KCs during early MASLD. Given that pro-inflammatory macrophages preferentially utilize glycolysis, such data could provide valuable insight into the reason for increased KC death beyond the presented hyperreliance on glycolysis.
(5) The gating strategy for monocyte-derived macrophages (moMFs) appears suboptimal and may include monocytes. A more rigorous characterization of myeloid populations by including additional markers would strengthen the study's conclusions.
(6) While BMDMs from Chil1 knockout mice are used to demonstrate enhanced glycolytic flux, it remains unclear whether Chil1 deficiency affects macrophage differentiation itself.
(7) The authors use the PDK activator PS48 and the ATP synthase inhibitor oligomycin to argue that increased glycolytic flux at the expense of OXPHOS promotes KC death. However, given the high energy demands of KCs and the fact that OXPHOS yields 15-16 times more ATP per glucose molecule than glycolysis, the increased apoptosis observed in Figure 4C-F could primarily reflect energy deprivation rather than a glycolysis-specific mechanism.
(8) In Figure 1C, KC numbers are significantly reduced after 4 and 16 weeks of HFHC diet in WT male mice, yet no comparable reduction is seen in Clec4Cre control mice, which should theoretically exhibit similar behavior under identical conditions.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors report converging evidence from behavioral studies as well as several brain-imaging techniques that geometric figures, notably quadrilaterals, are processed differently in visual (lower activation) and spatial (greater) areas of the human brain than representative figures. Comparison of mathematical models to fit activity for geometric figures shows the best fit for abstract geometric features like parallelism and symmetry. The brain areas active for geometric figures are also active in processing mathematical concepts even in blind mathematicians, linking geometric shapes to abstract math concepts. The effects are stronger in adults than in 6-year-old Western children. Similar phenomena do not appear in great apes, suggesting that this is uniquely human and developmental.
Strengths:
Multiple converging techniques of brain imaging and testing of mathematical models showing special status of perception of abstract forms. Careful reasoning at every step of research and presentation of research, anticipating and addressing possible reservations. Connecting these findings to other findings, brain, behavior, and historical/anthropological to suggest broad and important fundamental connections between abstract visual-spatial forms and mathematical reasoning.
Weaknesses:
I have reservations of the authors' use of "symbolic." They seem to interpret "symbolic" as relying on "discrete, exact, rule-based features." Words are generally considered to symbolic (that is their major function), yet words do not meet those criteria. Depictions of objects can be regarded as symbolic because they represent real objects, they are not the same as the object (as Magritte observed). If so then perhaps depictions of quadrilaterals are also symbolic but then they do not differ from depictions of objects on that quality. Relatedly, calling abstract or generalized representations of forms a distinct "language of thought" doesn't seem supportable by the current findings. Minimally, a language has elements that are combined more or less according to rules. The authors present evidence for geometric forms as elements but nowhere is there evidence for combining them into meaningful strings.
Further thoughts
Incidentally, there have been many attempts at constructing visual languages from visual elements combined by rules, that is, mapping meaning to depictions. Many written languages like Egyptian hieroglyphics or Mayan or Chinese, began that way; there are current attempts using emoji. Apparently, mapping sound to discrete letters, alphabets, is more efficient and was invented once but spread. That said, for restricted domains like maps, circuit diagrams, networks, chemical interactions, mathematics, and more, visual "languages" work quite well.
The findings are striking and as such invite speculation about their meaning and limitations. The images of real objects seem to be interpreted as representations of 3D objects as they activate the same visual areas as real objects. By contrast, the images of 2D geometric forms are not interpreted as representations of real objects but rather seemingly as 2D abstractions. It would be instructive to investigate stimuli that are on a continuum from representational to geometric, e. g., real objects that have simple geometric forms like table tops or boxes under various projections or balls or buildings that are rectangular or triangular. Objects differ from geometric forms in many ways: 3D rather than 2D, more complicated shapes; internal features as well as outlines. The geometric figures used are flat, 2-D, but much geometry is 3-D (e. g. cubes) with similar abstract features. The feature space of geometry is more than parallelism and symmetry; angles are important for example. Listing and testing features would be fascinating.
Can we say that mathematical thinking began with the regularities of shapes or with counting, or both? External representations of counting go far back into prehistory; tallies are frequent and wide-spread. Infants are sensitive to number across domains as are other primates (and perhaps other species). Finding overlapping brain areas for geometric forms and number is intriguing but doesn't show how they are related.
Categories are established in part by contrast categories; are quadrilaterals and triangles and circles different categories? As for quadrilaterals, the authors say some are "completely irregular." Not really; they are still quadrilaterals, if atypical. See Eleanor Rosch's insightful work on (visual) categories. One wonders about distinguishing squashed quadrilaterals from squashed triangles.
What in human experience but not the experience of close primates would drive the abstraction of these geometric properties? It's easy to make a case for elaborate brain processes for recognizing and distinguishing things in the world, shared by many species, but the case for brain areas sensitive to abstracting geometric figures is harder. The fact that these areas are active in blind mathematicians and that they are parietal areas suggest that what is important is spatial far more than visual. Could these geometric figures and their abstract properties be connected in some way to behavior, perhaps with fabrication, construction or use of objects? Or with other interactions with complex objects and environments where symmetry and parallelism (and angles and curvature--and weight and size) would be important? Manual dexterity and fabrication also distinguish humans from great apes (quantitatively not qualitatively) and action drives both visual and spatial representations of objects and spaces in the brain. I certainly wouldn't expect the authors to add research to this already packed paper, but raising some of the conceptual issues would contribute to the significance of the paper.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This research shows that a-mangostin, a proposed nutraceutical, with cardiovascular protective properties, could act through the activation of large conductance potassium permeable channels (BK). The authors provide convincing electrophysiological evidence that the compound binds to BK channels and induces a potent activation, increasing the magnitude of potassium currents. Since these channels are important modulators of the membrane potential of smooth muscle in vascular tissue, this activation leads to muscle relaxation, possibly explaining cardiovascular protective effects.
Strengths:
The authors present evidence based on several lines of experiments that a-mangostin is a potent activator of BK channels. The quality of the experiments and the analysis is high and represents an appropriate level of analysis. This research is timely and provides a basis to understand the physiological effects of natural compounds with proposed cardio-protective effects.
Weaknesses:
The identification of the binding site is not the strongest point of the manuscript. The authors show that the binding site is probably located in the hydrophobic cavity of the pore and show that point mutations reduce the magnitude of the negative voltage shift of activation produced by a-mangostin. However, these experiments do not demonstrate binding to these sites, and could be explained by allosteric effects on gating induced by the mutations themselves.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this study, Freier et al. demonstrate that 3 distinct metabolic pathways are critical for the synthesis of 1C-THF, a metabolite that is crucial for the growth and virulence of Listeria monocytogenes. Using an elegant suppressor screen, they also demonstrate the hierarchical importance of these metabolic pathways with respect to the biosynthesis of 1C-THF.
Strengths:
This study uses elegant bacterial genetics to confirm that 3 distinct metabolic pathways are critical for 1C-THF synthesis in L. monocytogenes, and the lack of either one of these pathways compromises bacterial growth and virulence. The study uses a combination of in vitro growth assays, macrophage-CFU assays, and murine infection models to demonstrate this.
Weaknesses:
(1) The primary finding of the study is that the perturbation of any of the 3 metabolic pathways important for the synthesis of 1C-THF results in reduced growth and virulence of L. monocytogenes. However, there is no evidence demonstrating the levels of 1C-THF in the various knockouts and suppressor mutants used in this study. It is important to measure the levels of this metabolite (ideally using mass spectrometry) in the various knockouts and suppressor mutants, to provide strong causality.
(2) The story becomes a little hard to follow since macrophage-CFU assays and murine infection model data precede the in vitro growth assays. The manuscript would benefit from a reorganization of Figures 2,3, and 4 for better readability and flow of data.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Ceravolo et al. employed functional magnetic resonance imaging (fMRI) to examine how the temporal voice areas (TVA) in the human brain respond to vocalizations from different nonhuman primate species. Their findings reveal that the human TVA is not only responsible for human vocalizations but also exhibits sensitivity to the vocalizations of other primates, particularly chimpanzee vocalizations sharing acoustic similarities with human voices, which offers compelling evidence for cross-species vocal processing in the human auditory system. Overall, the study presents intellectually stimulating hypotheses and demonstrates methodological originality. However, the current findings are not yet solid enough to fully support the proposed claims, and the presentation could be enhanced for clarity and impact.
Strengths:
The study presents intellectually stimulating hypotheses and demonstrates methodological originality.
Weaknesses:
(1) The analysis of the fMRI data does not account for the participants' behavioral performance, specifically their reaction times (RTs) during the species categorization task.
(2) The figure organization/presentation requires significant revision to avoid confusion and redundancy.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this study, authors utilize biophysical modeling to investigate differences in free energies and nucleosomal configuration probability density of CpG islands and nonmethylated regions in the genome. Toward this goal, they develop and apply the cgNA+ coarse-grained model, an extension of their prior molecular modeling framework.
Strengths:
The study utilizes biophysical modeling to gain mechanistic insight into nucleosomal occupancy differences in CpG and nonmethylated regions in the genome.
Weaknesses:
Although the overall study is interesting, the manuscripts need more clarity in places. Moreover, the rationale and conclusion for some of the analyses are not well described.
Comments on revised version:
The authors have addressed my concerns.
-
-
-
Reviewer #3 (Public review):
Summary:
This study examined how young children with minimal reading instruction process letters, focusing on their familiarity with letter shapes, knowledge of letter names, and visual discrimination of upright versus inverted letters. Across four experiments, kindergarten and Grade 1 children could identify the correct orientation of letters even without knowing their names.
Strengths:
This study addresses an important research gap by examining whether children develop letter familiarity prior to formal literacy instruction and how this skill relates to reading-related cognitive abilities. By emphasizing letter familiarity alongside letter recognition, the study highlights a potentially overlooked yet important component of emergent literacy development.
Weaknesses:
The study's methods and results do not effectively test its stated research goals. Reading ability was not directly measured; instead, the authors inferred its relationship with reading from correlations between letter familiarity and reading-related cognitive measures, which limits the validity of their conclusions. Furthermore, the analytical approach was rather limited, relying primarily on simple and partial correlations without employing more advanced statistical methods that could better capture the underlying relationships.
Major Comments:
(1) Limited Novelty and Unclear Theoretical Contribution:
The authors aim to challenge the view that children acquire letter shape knowledge only through formal literacy instruction, but similar questions regarding letter familiarity have already been explored in previous research. The manuscript does not clearly articulate how the present study advances beyond existing findings or why examining letter familiarity specifically before formal instruction provides new theoretical insight. Moreover, if letter familiarity and letter recognition are treated as distinct constructs, the authors should better justify their differentiation and clarify the theoretical significance of focusing on familiarity as an independent component of emergent literacy.
(2) Overgeneralization to Reading Ability:
Although the study measured several literacy-related cognitive skills and examined correlations with letter familiarity, it did not directly assess children's reading ability, as participants had not yet received formal literacy instruction. Therefore, the conclusion that letter familiarity influences reading skills (e.g., Line 519: "Our results are broadly consistent with previous work that has highlighted print letter knowledge as a strong predictor of future reading skills") is not fully supported and should be clarified or revised. To draw conclusions about the impact on reading ability, a longitudinal study would be more appropriate, assessing the relationship between letter familiarity and reading skills after children have received formal literacy instruction. If a longitudinal study is not feasible, measuring familial risk for dyslexia could provide an alternative approach to infer the potential influence of letter familiarity on later reading development.
(3) Confusing and Limited Analytical Approach with Potential for More Sophisticated Modeling:
The study employs a confusing analytical approach, alternating between simple correlational analyses and group-based comparisons, which may introduce circularity - for example, defining high vs. low familiarity groups partly based on performance differences in upright versus inverted letters and then observing a visual search advantage for upright letters within these groups. Moreover, the analyses are relatively simple: although multiple linear regression is mentioned, the results are not fully reported. These approaches may not fully capture the complex relationships among letter familiarity, recognition, visual search performance, RAN, and other covariates. More sophisticated modeling, such as mixed-effects models to account for repeated measures, structural equation modeling to examine latent constructs, or multivariate approaches jointly modeling familiarity and recognition effects, could provide a clearer understanding of the unique contribution of letter shape familiarity to early literacy outcomes. In addition, a large number of correlations were conducted without correction for multiple comparisons, which may increase the risk of false positives and raise concerns about the reliability of some significant findings.
Tags
Annotators
URL
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Despite the abundance of RNA velocity tools, there are still major limitations, and there is strong skepticism about the results these methods lead to. In this paper, the authors try to address some limitations of current RNA velocity approaches by proposing a unified framework to jointly infer transcriptional and splicing dynamics. The method is then benchmarked on 6 real datasets against the most popular RNA velocity tools.
While the approach has the potential to be of interest for the field, and may present improvements compared to existing approaches, there are some major limitations that should be addressed, particularly concerning the benchmark (see major comment 1).
Major comments:
(1) My main criticism concerns the benchmarking: real data lack a ground truth, and are absolutely not ideal for comparing methods, because one can only speculate what results appear to be more plausible.<br /> A solid and extensive simulation study, which covers various scenarios and possibly distinct data-generating models, is needed for comparing approaches. The authors should check, for example, the simulation studies in the BayVel approach (Section 4, BayVel: A Bayesian Framework for RNA Velocity Estimation in Single-Cell Transcriptomics). Clearly, all methods should be included in the simulation.
(2) Related to the above: since a ground truth is missing, the real data analyses need to be interpreted with caution. I recommend avoiding strong statements, such as "successfully captures the correct gene dynamics", or "accurately infer", in favour of milder statements supported by the data, such as "... aligns with the biological processes described" (as in page 12), or "results are compatible with current biological knowledge", etc...
(3) Many methods perform RNA velocity analyses. While there is a brief description, I think it'd be useful to have a schematic summary (e.g., via a Table) of the main conceptual, mathematical, and computational characteristics of each approach.
(4) Related to the above: I struggled to identify the main conceptual novelty of TSvelo, compared to existing approaches. I recommend explaining this aspect more extensively.
(5) A computational benchmark is missing; I'd appreciate seeing the runtime and memory cost of all methods in a couple of datasets.
(6) I think BayVel (mentioned above) should be added to the list of competing methods (both in the text and in the benchmarks). The package can be found here: https://github.com/elenasabbioni/BayVel_pkgJulia .
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors use fluorescent microscopy and fluorescent markers to investigate the requirement of P-bodies during growth on methanol, a common substrate available on plant leaves, by using a yeast edc3 mutant defective in P-body formation. Growth on methanol upregulates the transcription of methanol metabolic genes, which accumulate in granular structures, as observed by microscopy. Co-localization of P-bodies and granules was quantified and described as dynamically enhanced during oxidative stress. Ultimately, the authors suggest a model where methanol induces the accumulation of methanol-induced mRNAs in cytosolic granules, which dynamically interact with P-bodies, especially during oxidative stress, to protect the mRNAs from degradation. However, this model is not strongly supported by the provided data, as the quantification of the co-localization between different markers (of organelles and between P-body and granules) is not well presented or described in the text.
Considering that there is only a small EDC3-dependent overlap between P-bodies and mimRNA granules, the claim that P-bodies regulate mimRNAs is not fully justified. Rather, EDC3 could also be involved in mimRNA granule formation, independent of P-bodies.
Strengths:
(1) The authors could show convincingly that P-bodies (using a P-body-deficient edc3-KO strain) are important for colonizing the plant phyllosphere and for the regulation of methanol-induced mRNAs (mimRNA).
(2) The visualization of mimRNA granules and P-bodies using fluorescent markers is interesting and was validated by alternative methods, such as FISH staining.
(3) The dynamic formation of mimRNA granules and P-bodies was demonstrated during growth on leaves and in artificial medium during oxidative stress. The mimRNA granules showed a similar dynamic as the abundances of several mimRNAs and their corresponding proteins.
(4) A role of EDC3 in the formation of mimRNA granules was demonstrated. However, the link between P-bodies and mimRNA granules was not clearly shown.
Weaknesses:
(1) The study largely relies on fluorescent microscopy and co-localization measurements. However, the subcellular resolution is not very high; it is unclear how dot-like structures were measured and, importantly, how co-localization was quantified.
(2) The text does not clarify to what degree P-bodies and mimRNA granules are different structures. Based on the images, the size of P-bodies and granules seems to be vastly different, making it unclear whether these structures are fused or separate, even if their markers are reported to overlap.
(3) The evidence that mimRNA granules contain ribosome-free and ribosome-associated RNA is only based on inhibitors and microscopy, without providing further evidence measuring granule content by isolation and sequencing approaches.
(4) Similarly, the co-localization with other organelle markers is not supported by quantitative data.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors propose three types of Gaussian process kernels that extend and generalize standard kernels used for sequence-function prediction tasks, giving rise to the connectedness, Jenga, and general product models. The associated hyperparameters are interpretable and represent epistatic effects of varying complexity. The proposed models significantly outperform the simpler baselines, including the additive model, pairwise interaction model, and Gaussian process with a geometric kernel, in terms of R^2.
Strengths:
(1) The demonstrated performance boost and improved scaling with increasing training data are compelling.
(2) The hyperparameter selection step using the marginal likelihood, as implemented by the authors, seems to yield a reasonable hyperparameter combination that lends itself to biologically plausible interpretations.
(3) The proposed kernels generalize existing kernels in domain-interpretable ways, and can correspond to cases that would not be "physical" in the original models (e.g., $\mu_p>1$ in the original connectedness model that allows modeling of anticorrelated phenotypes).
Weaknesses:
(1) While enabling uncertainty quantification is a key advantage of Gaussian processes, the authors do not present metrics specific to the predicted uncertainties; all metrics seem to concern the mean predictions only. It would be helpful to evaluate coverage metrics and maybe include an application of the uncertainties, such as in active learning or Bayesian optimization.
(2) The more complex models, like the general product model, place a heavier burden on the hyperparameter selection step. Explicitly discussing the optimization routine used here would be helpful to potential users of the method and code.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This study presents a powerful and rigorous approach for characterizing stimulus discriminability throughout a sensory manifold, and is applied to the specific context of predicting color discrimination thresholds across the chromatic plane.
Strengths:
Color discrimination has played a fundamental role in studies of human color vision and for color applications, but as the authors note, it remains poorly characterized. The study leverages the assumption that thresholds should vary smoothly and systematically within the space, and validates this with their own tests and comparisons with previous studies.
Weaknesses:
The paper assumes that threshold variations are due to changes in the level of intrinsic noise at different stimulus levels. However, it's not clear to me why they could not also be explained by nonlinearities in the responses, with fixed noise. Indeed, most accounts of contrast coding (which the study is at least in part measuring because the presentation kept the adapt point close to the gray background chromaticity, and thus measured increment thresholds), assume a nonlinear contrast response function, which can at least as easily explain why the thresholds were higher for colors farther from the gray point. It would be very helpful if a section could be added that explains why noise differences rather than signal differences are assumed and how these could be distinguished. If they cannot, then it would be better to allow for both and refer to the variation in terms of S/N rather than N alone.
Related to this point, the authors note that the thresholds should depend on a number of additional factors, including the spatial and temporal properties and the state of adaptation. However, many of these again seem to be more likely to affect the signal than the noise.
An advantage of the approach is that it makes no assumptions about the underlying mechanisms. However, the choice to sample only within the equiluminant plane is itself a mechanistic assumption, and these could potentially be leveraged for deciding how to sample to improve the characterization and efficiency. For example, given what we know about early color coding, would it be more (or less) efficient to select samples based on a DKL space, etc?
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Solyga, Zelechowski, and Keller present a concise report of an innovative study demonstrating clear visuomotor mismatch responses in ambulating humans, using a mobile EEG setup and virtual reality. Human subjects walked around a virtual corridor while EEGs were recorded. Occasionally, motion and visual flow were uncoupled, and this evoked a mismatch response that was strongest in occipitally placed electrodes and had a considerable signal-to-noise ratio. It was robust across participants and could not be explained by the visual stimulus alone.
Strengths:
This is an important extension of their prior work in mice, and represents an elegant translation of those previous findings to humans, where future work can inform theories of e.g., psychiatric diseases that are believed to involve disordered predictive processing. For the most part, the authors are appropriately circumspect in their interpretations and discussions of the implications. I found the discussion of the polarity differences they found in light of separate positive and negative prediction errors, intriguing.
Weaknesses:
The primary weaknesses rest in how the results are sold and interpreted.
Most notably, the interpretation of the results of the comparison of visuomotor mismatches to the passive auditory oddball induced mismatch responses is inappropriate, as suboptimal electrode choices, unclear matching of trial numbers, and other factors. To clarify, regarding the auditory oddball portion in Figure 5, the data quality is a concern for the auditory ERPs, and the choice of Occipital electrodes is a likely culprit. Typically, auditory evoked responses are maximal at Cz or FCz, although these contacts don't seem to be available with this setup. In general, caution is warranted in comparing ERP peaks between two different sensory modalities - especially if attention is directed elsewhere (to a silent movie) during one recording and not during the other. The authors discuss this as a purely "qualitative" comparison in the text, which is appreciated, and do acknowledge the limitations within the results section, but the figure title and, importantly, the abstract set a different tone. At least, for comparisons between auditory mismatch and visuomotor mismatch, trial numbers need to be equated, as ERP magnitude can be augmented by noise (which reduces with increased numbers of trials in the average). And more generally, the size of the mismatch event at the scalp does not scale one-to-one with the size at the level of the neural tissue. One can imagine a number of variables that impact scalp level magnitudes, which are orthogonal to actual cortex-level activation - the size, spread, and polarity variance of the activated source (which all would diminish amplitude at the scalp due to polyphasic summation/cancelation). The variance of phase to a stimulus across trials (cross trial phase locking) vs magnitude of underlying power - the former, in theory, relates to bottom-up activity and the latter can reflect feedback (which has more variability in time across trials; the distance of the scalp electrode from the activated tissue (which, for the auditory system, would be larger (FCz to superior temporal gyrus) than for the visual system (O1 to V1/2)). None of this precludes the inclusion of the auditory mismatch, which is a strength of the study, but interpretations about this supporting a supremacy of sensory-motor mismatch - regardless of validity - are not warranted. I would recommend changing the way this is presented in the abstract.
Otherwise, the data are of adequate quality to derive most of their conclusions.
The authors claim that the mismatch responses emanate from within the occipital cortex, but I would require denser scalp coverage or a demonstration of consistent impedances across electrodes and across subjects to make conclusions about the underlying cortical sources (especially given the latencies of their peaks). In EEG, the distribution of voltage on the scalp is, of course, related to but not directly reflective of the distribution of the underlying sources. The authors are mostly careful in their discussion of this, but I would strongly recommend changing the work choice of "in occipital cortex" to "over occipital cortex" or even "posteriorly distributed". Even with very dense electrode coverage and co-registration to MRIs for the generation of forward models that constrain solutions, source localization of EEG signals is very challenging and not a simple problem. Given the convoluted and interior nature of human V1, the ability to reliably detect early evoked responses (which show the mismatch in mouse models) at the scalp in ERP peaks is challenging - especially if one is collapsing ERPs across subjects. And - given the latency of the mismatch responses, I'd imagine that many distributed cortical regions contribute to the responses seen at the scalp.
I think that Figure 3C, but as a difference of visual mismatch vs halting flow alone (in the open loop) might be additionally informative, as it clarifies exactly where the pure "mismatch" or prediction error is represented.
As a suggestion, the authors are encouraged to analyse time-frequency power and phase locking for these mismatch responses, as is common in much of the literature (see Roach et al 2008, Schizophrenia Bulletin). This is not to say that doing so will yield insights into oscillations per se, but converting the data to the time-frequency domain provides another perspective that has some advantages. It fosters translations to rodent models, as ERP peaks do not map well between species, but e.g., delta-theta power does (see Lee et al 2018, Neuropsychopharmacology; Javitt et al 2018, Schizophrenia research; Gallimore et al 2023, Cereb Ctx). Further, ERP peaks can be influenced by the actual neuroanatomy of an individual (especially for quantifying V1 responses). Time frequency analyses may aid in interpreting the "early negative deflection with a peak latency of 48 ms " finding as well.
Finally, the sentence in the abstract that this paradigm " can trigger strong prediction error responses and consequently requires shorter recording 20 times would simplify experiments in a clinical setting" is a nice setup to the paper, but the very fact that one third of recordings had to be removed due to movement artifact, and that hairstyle modulates the recording SnR, is reason that this paradigm, using the reported equipment, may have limited clinical utility in its current form. Further, auditory oddball paradigms are of great clinical utility because they do not require explicit attention and can be recorded very quickly with no behavioral involvement of a hospitalized patient. This should be discussed, although it does not detract from the overall scientific importance of the study. The authors should reconsider putting this statement in the abstract.
-
-
www.medrxiv.org www.medrxiv.org
-
Reviewer #3 (Public review):
Summary:
The manuscript by Goicoechea et al. assesses the influence of hippocampal-network targeted TMS to parietal cortex on episodic memory using a meta-analytic approach. This is an important contribution to the literature, as the number of studies using this approach to modulate memory/hippocampal function has clearly increased since the initial publication by Wang et al. 2014. This manuscript makes an important contribution to the literature. In general, the analysis is straightforward and the conclusions are well-supported by the results; I have mostly minor comments/concerns.
Strengths:
(1) A meta-analysis across published work is used to evaluate the influence of hippocampal-network-targeted TMS in parietal cortex on episodic memory. By pooling results across studies, the meta-analytic effects demonstrate an influence of TMS on memory across the diversity of many details in study design (specific tasks, stimuli, TMS protocols, study populations).
(2) Selectivity with regard to episodic memory vs. non-episodic memory tasks is evaluated directly in the meta-analysis.
(3) The investigation into supplemental factors as predictors of TMS's influence on memory was tested. This is helpful given the diversity of study designs in the literature. This analysis helps to shed light on which study designs, e.g., TMS protocols, etc., are most effective in memory modulation.
Weaknesses:
(1) My only significant concern is how studies are categorized in the 'Timing' factor (when stimulation is applied). Currently, protocols in which TMS is administered across days are categorized as 'pre-encoding' in the Timing factor. This has the potential to be misleading and may lead to inaccurate conclusions. When TMS is administered across multiple days, followed by memory encoding and retrieval (often on a subsequent day), it is not possible to attribute the influence of TMS to a specific memory phase (i.e., encoding or retrieval) per se. Thus, labeling multi-day TMS studies as 'pre-encoding' may be misleading to readers, as it may imply that the influence of TMS is due to modulation of encoding mechanisms per se, which cannot be concluded. For example, multi-day TMS protocols could be labeled as 'pre-retrieval' and be similarly accurate. This approach also pools results from TMS protocols with temporal specificity (i.e., those applied immediately during encoding and not on board during memory testing) and without temporal specificity (i.e., the case of multi-day TMS) regarding TMS timing. Given the variety of paradigms employed in the literature, and to maximize the utility/accuracy of this analysis, one suggestion is to modify the categories within the Timing factor, e.g., using labels like 'Temporally-Specific' and 'Temporally Non-specific'. The 'Temporally-Specific' category could be subdivided based on the specific memory process affected: 'encoding', 'retrieval', or 'consolidation' (if possible). I think this would improve the accuracy of the approach and help to reach more meaningful conclusions, given the variety of protocols employed in the literature.
(2) As the scope of the meta-analysis is limited to TMS applied to parietal or superior occipital cortex, it is important to highlight this in the Introduction/Abstract. The 'HITS' terminology suggests a general approach that would not necessarily be restricted to parietal/nearby cortical sites.
Minor:
(1) To reduce the number of study factors tested, data reduction was performed via Lasso regression to remove factors that were not unique predictors of the influence of TMS on memory. This approach is reasonable; however, one limitation is that factors strongly correlated with others (and predict less unique variance) will be dropped. This may result in a misrepresentation, i.e., if readers interpret factors left out of this analysis as not being strongly related to the influence of TMS on memory. I do see and appreciate the paragraph in the Discussion which appropriately addresses this issue. However, it may be worth also considering an alternative analysis approach, if the authors have not already done so, which explicitly captures the correlation structure in the data (i.e., shown in Figure S2) using a tool like PCA or an appropriate factor analysis. Then, this shared covariance amongst factors can be tested as predictors of the influence of TMS - e.g., by testing whether component scores for dominant PCs are indeed predictive of the influence of TMS. This complementary approach would capture rather than obfuscate the extent to which different factors are correlated and assess their joint (rather than independent) influence on memory, potentially resulting in more descriptive conclusions. For example, TMS intensity and protocol may jointly influence memory.
(2) Given the specific focus on TMS applied to parietal cortex to modulate hippocampal and related network function, it would be fruitful if the authors could consider adding discussion/speculation regarding whether this approach may be effectively broadened using other stimulation methods (e.g., tACS, tDCS), how it may compare to other non-invasive brain stimulation methods with depth penetration to target hippocampal function directly (transcranial temporal interference, or transcranial focused ultrasound), and/or how or whether other stimulation sites may or may not be effective.
(3) Studies were only included in the meta-analysis if they contained objective episodic memory tests. How were studies handled that included both objective and subjective memory, or other non-episodic memory measures? For example, Yazar et al. 2014 showed no influence of TMS on objective recall, but an impairment in subjective confidence. I assume confidence was not included in the meta-analysis. Similarly, Webler et al. 2024 report results from both the mnemonic similarity task (presumably included) and a fear conditioning paradigm (presumably excluded). Please clarify in the methods how these distinctions were handled.
(4) The analysis comparing memory to non-memory measures is important, showing the specificity of stimulation. Did the authors consider further categorizing the non-memory tasks into distinct domains (i.e., language, working memory, etc.)? If possible, this could provide a finer detail regarding the selectivity of influences on memory vs. other aspects of cognition. It is likely that other aspects of cognition dependent on hippocampal function may be modulated as well, i.e., tasks with high relational/associative processing demands.
(5) In the analysis of the Intensity factor, how were studies using Active (rather than resting) MT categorized? Only resting MT is mentioned in Table S1. This is important as the original theta-burst TMS protocol from Huang et al. 2005 determines intensity based on Active Motor Threshold.
(6) Is there a reason why the study by Koen et al. 2018 (Cognitive Neuroscience) was not included? TMS was performed during encoding to the left AG, and objective memory was assessed, so it would seemingly meet the inclusion criterion.
(7) It would be helpful to briefly differentiate the current meta-analysis from that performed by Yeh & Rose (How can transcranial magnetic stimulation be used to modulate episodic memory?: A systematic review and meta-analysis, 2019, Frontiers in Psychology) (other than being more current).
(8) For transparency and to facilitate further understanding of the literature and potential data re-use, it would be great if the authors consider sharing a supplementary table or file that describes how individual studies/memory measures were categorized under the factors listed in Table S1.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This article presented a novel computer model to address an important question in the field of brain stimulation, using the magnetic stimulation iTBS protocol as an example, how stimulation parameters, frequency in particular, interfere with the intrinsic brain oscillations via plastic mechanisms. Brain oscillation is a critical feature of functional brains and its alteration signals the onset of many neuropsychiatric diseases or certain brain states. The authors suggested with their model that harmonic and subharmonic stimulations close to the individual alpha frequency achieved strong broadband power suppression.
Strengths:
The authors focused on the cortico-thalamic circuitry and managed to generate alpha oscillations in their four-population model. By adding the non-monotonic calcium-based BCM rule, they have also achieved both homeostasis and plasticity in response to magnetic stimulation. This work combined computer simulations and statistical analysis to demonstrate the changes in network architecture and network dynamics triggered by varied magnetic stimulation parameters. By delivering the iTBS protocol to the cortical excitatory population, the key findings are that harmonic and subharmonic stimulations close to the individual alpha frequency (IAF) achieved strong broadband power suppression. This resulted from increased synaptic weights of the corticothalamic feed-forward inhibitory projections, which were mediated by the calcium dynamics perturbed by iTBS magnetic stimulation. This finding endorsed the importance of applying customized stimulation to patients based on their IAFs and suggested the underlying mechanism at the circuitry level.
Weaknesses:
The drawbacks of this work are also obvious. Model validation and biological feasibility justification should be better addressed. The primary outcome of their model is the broadband power suppression and the optimal effects of (sub)harmonic stimulation frequency, but it lacks immediate empirical support in the literature. To the best of my knowledge, many alpha frequency tACS studies reported to increase but not suppress the power of certain brain oscillations. A review by Wang et al., 2024 (Frontiers in System Neuroscience) suggested hybrid changes to different brain oscillations by magnetic stimulation. Developing a model to fully capture such changes might be out of the scope of the present study and challenging in the entire field, but it undermines the quality of the present work if not extensively discussed and justified. Clarity and reproducibility of the work can be improved. Although it is intriguing to see how the calcium-dependent BCM plasticity mediates such changes, the writing of the methods part is not hard to follow. It was also not clear why only two populations were considered in the thalamus, how the entire network was connected, or how the LTP/LTD threshold alters with calcium dynamics. The figures were unfortunately prepared in a nested manner. The crowded layout and the tiny font sizes reduce the clarity. The third point comes to contextualization and comparison to existing models. It will strengthen the work if the authors could have compared their work to other TMS modeling work with plasticity rules, e.g, Anil et al., 2024. Besides, magnetic stimulation is unique in being supra-threshold and having focality compared to other brain stimulation modalities, e.g., tDCS and tACS, but they may share certain basic neural mechanisms if accounting for certain parameters, e.g., frequency. A solid literature review and discussion on this part may help the field better perceive the value and potential limitations of this work.
-
-
www.youtube.com www.youtube.com
-
In regions where rainfall was just 3 in annually, the temporary bean still produced.
for - stats - agriculture - 3 inches annual rain - bean still produced
-
-
-
Reviewer #3 (Public Review):
In this manuscript, Verma et al. set out to visualize cytoplasmic dynein in living cells and describe their behaviour. They first generated heterozygous CRISPR-Cas9 knock-ins of DHC1 and p50 subunit of dynactin and used spinning disk confocal microscopy and TIRF microscopy to visualize these EGFP-tagged molecules. They describe robust localization and movement of DHC and p50 at the plus tips of MTs, which was abrogated using SiR tubulin to visualize the pool of DHC and p50 on the MTs. These DHC and p50 punctae on the MTs showed similar, highly processive movement on MTs. Based on comparison to inducible EGFP-tagged kinesin-1 intensity in Drosophila S2 cells, the authors concluded that the DHC and p50 punctae visualized represented 1 DHC-EGFP dimer+1 untagged DHC dimer and 1 p50-EGFP+3 untagged p50 molecules.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Deletion of the TMA-sensor TAAR5 results in circadian alterations in the gene expression, particularly in the olfactory bulb; plasma hormones; and neurobehaviors.
Strengths:
Genetic background was rigorously controlled.
Comprehensive characterization.
Impact:
These data add to the growing literature pointing to a role for the TMA/TMAO pathway in olfaction and neurobehavior.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
In this paper, the authors investigate how the RNA-binding protein Ssd1 and calorie restriction (CR) influence yeast replicative lifespan, with a particular focus on age-dependent iron uptake and activation of the iron regulon. For this, they use microfluidics-based single-cell imaging to monitor replicative lifespan, protein localization, and intracellular iron levels across aging cells. They show that both Ssd1 overexpression and CR act through a shared pathway to prevent the nuclear translocation of the iron-regulon regulator Aft1 and the subsequent induction of high-affinity iron transporters. As a result, these interventions block the age-related accumulation of intracellular free iron, which otherwise shortens lifespan. Genetic and chemical epistasis experiments further demonstrate that suppression of iron regulon activation is the key mechanism by which Ssd1 and CR promote replicative longevity.
Overall, the paper is technically rigorous, and the main conclusions are supported by a substantial body of experimental data. The microfluidics-based assays in particular provide compelling single-cell evidence for the dynamics of Ssd1 condensates and iron homeostasis.
My main concern, however, is that the central reasoning of the paper-that Ssd1 overexpression and CR prevent the activation of the iron regulon-appears to be contradicted by previous findings, and the authors may actually be misrepresenting these studies, unless I am mistaken. In the manuscript, the authors state on two occasions:
"Intriguingly, transcripts that had altered abundance in CR vs control media and in SSD1 vs ssd1∆ yeast included the FIT1, FIT2, FIT3, and ARN1 genes of the iron regulon (8)"
"Ssd1 and CR both reduce the levels of mRNAs of genes within the iron regulon: FIT1, FIT2, FIT3 and ARN1 (8)"
However, reference (8) by Kaeberlein et al. actually says the opposite:
"Using RNA derived from three independent experiments, a total of 97 genes were observed to undergo a change in expression >1.5-fold in SSD1-V cells relative to ssd1-d cells (supplemental Table 1 at http://www.genetics.org/supplemental/). Of these 97 genes, only 6 underwent similar transcriptional changes in calorically restricted cells (Table 2). This is only slightly greater than the number of genes expected to overlap between the SSD1-V and CR datasets by chance and is in contrast to the highly significant overlap in transcriptional changes observed between CR and HAP4 overexpression (Lin et al. 2002) or between CR and high external osmolarity (Kaeberlein et al. 2002). Intriguingly, of the 6 genes that show similar transcriptional changes in calorically restricted cells and SSD1-V cells, 4 are involved in iron-siderochrome transport: FIT1, FIT2, FIT3, and ARN1 (supplemental Table 1 at http://www.genetics.org/supplemental/)."
Although the phrasing might be ambiguous at first reading, this interpretation is confirmed upon reviewing Matt Kaeberlein's PhD thesis: https://dspace.mit.edu/handle/1721.1/8318
(page 264 and so on)
Moreover, consistent with this, activation of the iron regulon during calorie restriction (or the diauxic shift) has also been observed in two other articles:
https://doi.org/10.1016/S1016-8478(23)13999-9
https://doi.org/10.1074/jbc.M307447200
Taken together, these contradictory data might blur the proposed model and make it unclear how to reconcile the results.
Comments on revisions:
The authors successfully addressed my requests and concerns
-
-
www.researchgate.net www.researchgate.net
-
Reviewer #3 (Public review):
Summary:
This manuscript presents an ambitious integration of multiple artificial intelligence technologies to examine social learning in naturalistic mother-infant interactions. The authors aimed to quantify how information flows between mothers and infants across different communicative modalities and timescales, using speech analysis (Whisper), pose detection (MMPose), facial expression recognition, and semantic modeling (GPT-2) in a unified analytical framework. Their goal was to provide unprecedented quantitative precision in measuring behavioral coordination and information transfer patterns during social learning, moving beyond traditional observational coding approaches to examine cross-modal coordination patterns and semantic contingencies in real-time across multiple temporal scales.
Strengths:
The integration of multiple AI tools into a coherent analytical framework represents a genuine methodological breakthrough that advances our capabilities for studying complex social phenomena. The authors successfully analyzed naturalistic interactions at a scale and level of detail that was not previously possible, examining 33 5-month-old and 34 15-month-old dyads across multiple modalities simultaneously. This sophisticated analytical pipeline, combining speech analysis, semantic modeling, pose detection, and facial expression recognition, provides new capabilities for studying social interactions that extend far beyond what traditional observational coding could achieve.
The specific findings about hierarchical information flow patterns across different timescales are particularly valuable and would not have been possible without this sophisticated analytical approach. The discovery that mothers reduce low-level sensory input when infants focus on objects, while increases in object naming and information rate associate with sustained attention, provides new empirical insights into how social learning unfolds in naturalistic settings. The temporal dynamics analyses reveal interesting patterns of behavioral coordination that extend our understanding of how caregivers adaptively modify their responses to support infant attention across multiple communicative channels simultaneously.
The scale of data collection and the comprehensive multi-modal approach are impressive, opening up new possibilities for understanding social learning processes. The methodological innovations demonstrate how modern computational tools can be systematically integrated to reveal new quantitative aspects of well-established developmental phenomena. The computational features developed for this study represent innovative applications of information theory and computer vision to developmental research.
Weaknesses:
Several major limitations affect the reliability and interpretability of the findings. The sample sizes of 33-34 dyads per age group are relatively modest for the complexity of analyses performed, which include eight different features examined across various time lags with extensive statistical comparisons. The study lacks adequate power analysis to demonstrate whether these sample sizes are sufficient to detect meaningful effect sizes, which is particularly concerning given the multiple comparison burden inherent in this type of multi-modal, multi-timescale analysis.
The statistical framework presents several concerns that limit confidence in the findings. Inter-rater reliability for gaze coding shows substantial but not excellent agreement (κ = 0.628), with only 22% of the data undergoing double coding. Given that gaze coding forms the foundation for all subsequent analyses of joint attention and information flow, this reliability level may systematically influence findings. The multiple comparison correction strategies vary inconsistently across different analyses, with some using FDR correction and others treating lower-level and higher-level features separately. Additionally, object naming analyses employed one-sided tests (p<0.05) while others used two-sided tests (p<0.025) without clear theoretical or methodological justification for these differences.
The validation of AI tools in the specific context of mother-infant interactions is insufficient and represents a critical limitation. The performance characteristics of Whisper with infant-directed speech, the precision of MMPose for detecting facial landmarks in young children, and the accuracy of facial expression recognition tools in infant contexts are not adequately validated for this population. These sophisticated tools may not perform optimally in the specific context of mother-infant interactions, where speech patterns, facial expressions, and body movements may differ substantially from their training data.
The theoretical positioning requires substantial refinement to better acknowledge the extensive existing literature. The authors are working within a well-established theoretical framework that has long recognized social learning as an active, bidirectional process. The joint attention literature, beginning with foundational work by Bruner (1983) and continuing through contemporary theories of social cognition by researchers like Tomasello (1995), has emphasized the communicative and adaptive nature of attentional processes. The scaffolding literature, including seminal work by Wood, Bruner, and Ross (1976), has demonstrated how parents adjust their support based on children's developing competencies. Moreover, there is a substantial body of micro-analytic research that has employed sophisticated quantitative methods to study social interactions, including work by Stern (1985) on microsecond-level interactions and research using time-series methods to examine dyadic coordination patterns.
The cross-correlation analyses have inherent limitations for causal inference that are not adequately acknowledged. The interpretation of temporal correlation patterns in terms of directional influence requires more cautious consideration, as observational data have fundamental constraints for establishing causality. The ecological validity is also questionable due to the laboratory tabletop interaction paradigm and the sample's demographic homogeneity, consisting primarily of white, highly educated, high-income mothers.
-
-
www.researchsquare.com www.researchsquare.com
-
Reviewer #3 (Public review):
The paper by Maggi et al. builds on earlier work by the team (Paatero et al., 2018) on oriented junction-based lamellipodia (JBL). They validate the role of JBLs in guiding endothelial cell rearrangements and utilise high-resolution time-lapse imaging of novel transgenic strains to visualise the formation of distal junctions and their subsequent fusion with proximal junctions. Through functional analyses of Arp2/3 and actomyosin contractility, the study identifies JBLs as localized mechanical hubs, where protrusive forces drive distal junction formation, and actomyosin contractility brings together the distal and proximal junctions. This forward movement provides a unique directionality which would contribute to proper lumen formation, EC orientation, and vessel stability during these early stages of vessel development.
Time-lapse live imaging of VEC, ZO-1, and actin reveals that VEC and ZO-1 are initially deposited at the distal junction, while actin primarily localizes to the region between the proximal and distal sites. Using a photoconvertible Cdh5-mClav2 transgenic line, the origin of the VEC aggregates was examined. This convincingly shows that VE-cadherin was derived from pools outside the proximal junctions. However, in addition to de novo VEC derived from within the photoconverted cell, could some VEC also be contributed by the neighbouring endothelial cell to which the JBL is connected?
As seen for JAILs in cultured ECs, the study reveals that Arp2/3 is enhanced when JBLs form by live imaging of Arpc1b-Venus in conjunction with ZO-1 and actin. Therefore Arp2/3 likely contributes to the initial formation of the distal junction in the lamellopodium.
Inhibiting Arp2/3 with CK666 prevents JBL formation, and filopodia form instead of lamellopodia. This loss of JBLs leads to impaired EC rearrangements.
Is the effect of CK666 treatment reversible? Since only a short (30 min) treatment is used, the overall effect on the embryo would be minimal, and thus washing out CK666 might lead to JBL formation and normalized rearrangements, which would further support the role of Arp2/3.
From the images in Figure 4d it appears that ZO-1 levels are increased in the ring after CK666 treatment. Has this been investigated, and could this overall stabilization of adhesion proteins further prevent elongation of the ring?
To explore how the distal and proximal junctions merge, imaging of spatiotemporal imaging of Myl9 and VEC is conducted. It indicates that Myl9 is localized at the interjunctional fusion site prior to fusion. This suggests pulling forces are at play to merge the junctions, and indeed Y 27632 treatment reduces or blocks the merging of these junctions.
For this experiment, a truncated version of VEC was use,d which lacks the cytoplasmic domain. Why have the authors chosen to image this line, since lacking the cytoplasmic domain could also impair the efficiency of tension on VEC at both junction sites? This is as described in the discussion (lines 328-332).
Since the time-lapse movies involve high-speed imaging of rather small structures, it is understandable that these are difficult to interpret. Adding labels to indicate certain structures or proteins at essential timepoints in the movies would help the readers understand these.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
The article's main question is how humans handle spurious transitions between object features when learning a predictive model for decision-making. The authors conjecture that humans use semantic knowledge about plausible causal relations as an inductive bias to distinguish true from spurious links.
The authors simulate a successor feature (SF) model, demonstrating its susceptibility to suboptimal learning in the presence of spurious transitions caused by co-occurring but independent causal factors. This effect worsens with an increasing number of planning steps and higher co-occurrence rates. In a preregistered study (N=100), they show that humans are also affected by spurious transitions, but perform somewhat better when true transitions occur between features within the same semantic category. However, no evidence for the benefits of semantic congruency was found in test trials involving novel configurations, and attempts to model these biases within an SF framework remained inconclusive.
Strengths:
(1) The authors tackle an important question.
(2) Their simulations employ a simple yet powerful SF modeling framework, offering computational insights into the problem.
(3) The empirical study is preregistered, and the authors transparently report both positive and null findings.
(4) The behavioral benefit during learning in the congruent vs incongruent condition is interesting
Weaknesses:
(1) A major issue is that approximately one quarter of participants failed to learn, while another quarter appeared to use conjunctive or configural learning strategies. This raises questions about the appropriateness of the proposed feature-based learning framework for this task. Extensive prior research suggests that learning about multi-attribute objects is unlikely to involve independent feature learners (see, e.g., the classic discussion of configural vs. elemental learning in conditioning: Bush & Mosteller, 1951; Estes, 1950).
(2) A second concern is the lack of explicit acknowledgment and specification of the essential role of the co-occurrence of causal factors. With sufficient training, SF models can develop much stronger representations of reliable vs. spurious transitions, and simple mechanisms like forgetting or decay of weaker transitions would amplify this effect. This should be clarified from the outset, and the occurrence rates used in all tasks and simulations need to be clearly stated.
(3) Another problem is that the modeling approach did not adequately capture participant behavior. While the authors demonstrate that the b parameter influences model behavior in anticipated ways, it remains unclear how a model could account for the observed congruency advantage during learning but not at test.
(4) Finally, the conceptualization of semantic biases is somewhat unclear. As I understand it, participants could rely on knowledge such as "the shape of a building robot's head determines the kind of head it will build," while the type of robot arm would not affect the head shape. However, this assumption seems counterintuitive - isn't it plausible that a versatile arm is needed to build certain types of robot heads?
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
This is an intriguing paper that reports a potentially novel mechanism of reversible phosphorylation of AGC kinase activation segments by changes in sodium and potassium ion concentrations. The authors show for a variety of AGC kinases that incubating diverse eukaryotic cell types in 450 and 600 mM NaCl results in dephosphorylation of the activation segment. In contrast, phosphorylation of the activation segment for p38 kinases increases. No dephosphorylation of AGC kinases activation segment occurs with sorbitol, thus dephosphorylation is independent of osmotic pressure. This effect is rapidly reversed when cells are returned to normal media and the AGC kinase is re-phosphorylated. This phenomenon is also observed for eukaryotic cell-free extracts, and is induced by other alkali metal ions but not lithium. Importantly, no dephosphorylation is observed in the E. coli cell extract.
The authors also make the following observations:
(1) Dephosphorylation is dependent on PP2A.
(2) Re-phosphorylation is not dependent on PDK1, ATP, and Mg2+.
(3) The K/Na-dependent dephosphorylation/phosphorylation is observed even for relatively short protein segments that incorporate the activation segment.
(4) The phosphorylation observed occurs in cis, i.e., only the activation segment of the protein that is dephosphorylated becomes phosphorylated on reduced KCl. An activation segment from a different length protein is not phosphorylated.
(5) No evidence for auto(de)phosphorylation.
(6) The authors propose three models to explain the dephosphorylation/phosphorylation mechanism. Their experimental data suggest that an acceptor molecule is responsible for accepting the phosphate group and then transferring it back to the activation segment.
Comments on results and experiments:
(1) Are these results an artefact of their assay? The authors mainly use immunoblotting to assess the phosphorylation status of AGC kinase. However, an assay artefact would not show a difference between control and okadaic-acid-treated cells (Figure 3A). Moreover, the authors show dephosphorylation/phosphorylation using radiolabelling (Figure 6C).
(2) Preferably, the authors would have a control to test dephosphorylation/phosphorylation does not occur in the absence of cell extract. The E. coli extract shows that dephosphorylation/phosphorylation is specific to eukaryotic cell extracts.
(3) The authors should show that dephosphorylation/phosphorylation occurs on the same residue of the activation segment (by mass spec).
(4) Since phosphorylation levels are assessed using immunoblots, the levels of dephosphorylation/phosphorylation are not quantified. What proportion of AGC kinase is phosphorylated initially (before Na/K-induced dephosphorylation)?
(5) The experiment to test autophosphorylation (Figure 4, Figure supplement 1B) is not completely convincing because the authors use a cell line with a PKN1 mutant knock-in. Possibly PKN2 or another AGC kinase could phosphorylate the proteins expressed from the transfection vector - although the authors do test with AGC kinase inhibitors.
(6) What are the two bands in Figure 6C (lanes 'Con' and 'diluted)? Only one band disappears with KCl. There is one band in Figure 6 Supplement 2.
In summary, the results presented in this paper are highly unusual. Generally, the manuscript is well written and the figures are clear. The authors have performed numerous experiments to understand this process. These appear robust, and most of their data lend credence to their model in Figure 6Aiii. The idea that a phosphate group can be transferred by an enzyme onto/between molecule(s) is not unprecedented, i.e., phosphoglycerate mutase catalyses 3-phosphoglycerate isomerisation through a phosphorylenzyme intermediate. It will be important to identify this transfer enzyme. One observation that does not fit easily with their model is the role of PP2A. Since protein dephosphorylation by PP2A does not involve a phosphorylenzyme intermediate, if the initial dephosphorylation reaction is catalysed by PP2A, it is very difficult to envision how the free phosphate is then used to phosphorylate the activation segment.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Ji et al. report a novel and interesting light-induced transcriptional response pathway in the eyeless roundworm Caenorhabditis elegans that involves a cytochrome P450 family protein (CYP-14A5) and functions independently from previously established photosensory mechanisms. Although the exact mechanisms underlying photoactivation of this pathway remain unclear, light-dependent induction of CYP-14A5 requires bZIP transcription factors ZIP-2 and CEBP-2 that have been previously implicated in worm responses to pathogens. The authors then suggest that light-induced CYP-14A5 activity in the C. elegans hypoderm can unexpectedly and cell-non-autonomously contribute to retention of an olfactory memory. Finally, the authors demonstrate the potential for this pathway to enable robust light-induced control of gene expression and behavior, albeit with some restrictions. Overall, the evidence supporting the claims of the authors is convincing, and the authors' work suggests numerous interesting lines of future inquiry.
(1) The authors determine that light, but not several other stressors tested (temperature, hypoxia, and food deprivation), can induce transcription of cyp-15A5. The authors use these experiments to suggest the potential specificity of the induction of CYP-14A5 by light. Given the established relationship between light and oxidative stress and the authors' later identification of ZIP-2, testing the effect of an oxidative stressor or pathogen exposure on transcription of cyp-14A5 would further strengthen the validity of this statement and potentially shed some insight into the underlying mechanisms.
(2) The authors suggest that short-wavelength light more robustly increases transcription of cyp-14A5 compared to equally intense longer wavelengths (Figure 2F and 2G). Here, however, the authors report intensities in lux of wavelengths tested. Measurements of and reporting the specific spectra of the incident lights and their corresponding irradiances (ideally, in some form of mW/mm2 - see Ward et al., 2008, Edwards et al., 2008, Bhatla and Horvitz, 2015, De Magalhaes Filho et al., 2018, Ghosh et al., 2021, among others, for examples) is critical for appropriate comparisons across wavelengths and facilitates cross-checking with previous studies of C. elegans light responses. On a related and more minor note, the authors place an ultraviolet shield in front of a visible light LED to test potential effects of ultraviolet light on transcription of cyp-14A5. A measurement of the spectrum of the visible light LED would help confirm if such an experiment was required. Regardless, the principal conclusions the authors made from these experiments will likely remain unchanged.
(3) The authors report an interesting observation that animals exposed to ambient light (~600 lux) exhibit significantly increased memory retention compared to those maintained in darkness (Figure 4). Furthermore, light deprivation within the first 2-4 hours after learning appears to eliminate the effect of light on memory retention. These processes depend on CYP-14A5, loss of which can be rescued by re-expression of cyp-14A5 in mutant animals using a hypoderm-specific- and non-light-inducible- promoter. Taken together, the authors argue convincingly that hypodermal expression of cyp-14A5 can contribute to the retention of the olfactory memory. More broadly, these experiments suggest that cell-non-autonomous signaling can enhance retention of olfactory memory. How retention of the olfactory memory is enhanced by light generally remains unclear. In addition, the authors' experiments in Figure 1B demonstrate - at least by use of the transcriptional reporter - that light-dependent induction of cyp-14A5 transcription at 500 - 1000 lux is minimal and especially so at short duration exposures. Additional experiments, including verification of light-dependent changes in CYP-14A5 levels in the olfactory memory behavioral setup, would help further interpret these otherwise interesting results.
(4) The experiments in Figure 4 nicely validate the usage of the cyp-14A5 promoter as a potential tool for light-dependent induction of gene expression. Despite the limitations of this tool, including those presented by the authors, it could prove useful for the community.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The article develops a CNN-based metastasis scoring system to distinguish cell subsets with high brain metastatic potential and validates its performance using patient platelet data. The robustness of this approach is further demonstrated across diverse single-cell and spatial datasets from multiple cancers, supported by transcription factor and gene set analyses, as well as novel drug identification pipelines. Together, these findings provide strong evidence that reinforces the central theme of the study.
Strengths:
Development of a CNN-based scoring system to reveal the potential of brain metastasis that is robust across multiple cancer cell types, validated by multiple datasets. Other approaches, including transcription factor analyses, cell-cell communication analysis, and spatial transcriptomic, etc., were included to strengthen the work.
Weaknesses:
The author could identify/validate more signaling pathways beyond the VEGF pathway since it's well known in metastasis.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This manuscript examines how locus coeruleus (LC) activity relates to hippocampal ripple events across behavioral states in freely moving rats. Using multi-site electrophysiological recordings, the authors report that LC activity is suppressed prior to ripple events, with the magnitude of suppression depending on the ripple subtype. Suppression is stronger during wakefulness than during NREM sleep and is least pronounced for ripples coupled to spindles.
Strengths:
The study is technically competent and addresses an important question regarding how LC activity interacts with hippocampal and thalamocortical network events across vigilance states.
Weaknesses:
The results are interesting, but entirely observational. Also, the study in its current form would benefit from optimization of figure labeling and presentation, and more detailed result descriptions to make the findings fully interpretable. Also, it would be beneficial if the authors could formulate the narrative and central hypothesis more clearly to ease the line of reasoning across sections.
Comments:
(1) Stronger evidence that recorded units represent noradrenergic LC neurons would reinforce the conclusions. While direct validation may not be possible, showing absolute firing rates (Hz) across quiet wake, active wake, NREM, and REM, and comparing them to published LC values, would help.
(2) The analyses rely almost exclusively on z-scored LC firing and short baselines (~4-6 s), which limits biological interpretation. The authors should include absolute firing rates alongside normalized values for peri-ripple and peri-spindle analyses and extend pre-event windows to at least 20-30 s to assess tonic firing evolution. This would clarify whether differences across ripple subtypes arise from ceiling or floor effects in LC activity; if ripples require LC silence, the relative drop will appear larger during high-firing wake states. This limitation should be discussed and, if possible, results should be shown based on unnormalized firing rates.
(3) Because spindles often occur in clusters, the timing of ripple occurrence within these clusters could influence LC suppression. Indicate whether this structure was considered or discuss how it might affect interpretation (e.g., first vs. subsequent ripples within a spindle cluster).
(4) While the observational approach is appropriate here, causal tests (e.g., optogenetic or chemogenetic manipulation of LC around ripple events and in memory tasks) would considerably strengthen the mechanistic conclusions. At a minimum, a discussion of how such approaches could address current open questions would improve the manuscript.
(5) Please show how "Synchronization Index" (SI) differs quantitatively across behavioral states (wake, NREM, REM) and discuss whether it could serve as a state classifier. This would strengthen interpretations of the correlations between SI, ripple occurrence, and LC activity.
(6) The current use of SI to denote a delta/gamma power ratio is unconventional, as "SI" typically refers to phase-locking metrics. Consider adopting a more standard term, such as delta/gamma power ratio. Similarly, it would be easier to follow if you use common terminology (AUC) to describe the drop in LC-MUA rather than using "MI" and "sub-MI".
(7) The logic in Figure 3 is difficult to follow. The brain state (delta/gamma ratio) appears unchanged relative to surrogate events (3C), while LC activity that is supposedly negatively correlated to delta/gamma changes markedly (3D-E). Could this discrepancy reflect the low temporal resolution (4-s windows) used to calculate delta/gamma when the changes occur on a shorter time scale?
(8) There are apparent inconsistencies between Figures 4B and 4C-D. In B, it seems that the difference between the 10th and 90th percentile is mostly in higher frequencies, but in C and D, the only significant difference is in the delta band.
(9) Because standard sleep scoring is based on EEG and EMG signals, please include an example of sleep scoring alongside the data used for state classification. It would also be relevant to include the delta/gamma power ratio in such an example plot.
(10) Can variability in modulation index (subMI) across ripple subsets reflect differences in recording quality? Please report and compare mean LC firing rates across subsets to confirm this is not a confounding factor.
(11) Figure 6B: If the brown trace represents LC-MUA activity around random time points, why would there be a coinciding negative peak as relative to real sleep spindles? Or is it the subtracted trace?
(12) On page 8, lines 207-209, the authors write "Importantly, neither the LC-MUA rate nor SIs differed during a 2-sec time window preceding either group of spindles". It is unclear which data they refer to, but the statement seems to contradict Figure 6E as well as the following sentence: "Across sessions, MI values exceeded 95% CI in 17/20 datasets for isoSpindles and only 3/20 for ripSpindles". This should be clarified.
(13) The results in Figures 5C and 6F do not align. It seems surprising that ripple-coupled spindles show a considerably higher LC modulation than spindle-coupled ripples, as these events should overlap. Could the discrepancy be due to Z-score normalization as mentioned above? Please include a discussion of this to help the interpretation of the results.
(14) The text implies that 8 recordings came from one rat and two each from six others. This should be confirmed, and it should be explained how the recordings were balanced and analyzed across animals.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors set out to determine how GABAergic inhibitory premotor circuits contribute to the rhythmic alternation of leg flexion and extension during Drosophila grooming. To do this, they first mapped the ~120 13A and 13B hemilineage inhibitory neurons in the prothoracic segment of the VNC and clustered them by morphology and synaptic partners. They then tested the contribution of these cells to flexion and extension using optogenetic activation and inhibition and kinematic analyses of limb joints. Finally, they produced a computational model representing an abstract version of the circuit to determine how the connectivity identified in EM might relate to functional output. The study makes important contributions to the literature.
The authors have identified an interesting question and use a strong set of complementary tools to address it:
They analysed serial‐section TEM data to obtain reconstructions of every 13A and 13B neuron in the prothoracic segment. They manually proofread over 60 13A neurons and 64 13B neurons, then used automated synapse detection to build detailed connectivity maps and cluster neurons into functional motifs.
They used optogenetic tools with a range of genetic driver lines in freely behaving flies to test the contribution of subsets of 13A and 13B neurons.
They used a connectome-constrained computational model to determine how the mapped connectivity relates to the rhythmic output of the behavior.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this manuscript, Lamothe et al. sought to identify the neural substrates of voice identity in the human brain by correlating fMRI recordings with the latent space of a variational autoencoder (VAE) trained on voice spectrograms. They used encoding and decoding models, and showed that the "voice" latent space (VLS) of the VAE performs, in general, (slightly) better than a linear autoencoder's latent space. Additionally, they showed dissociations in the encoding of voice identity across the temporal voice areas.
Strengths:
The geometry of the neural representations of voice identity has not been studied so far. Previous studies on the content of speech and faces in vision suggest that such geometry could exist. This study demonstrates this point systematically, leveraging a specifically trained variational autoencoder.
The size of the voice dataset and the length of the fMRI recordings ensure that the findings are robust.
Comments on revisions:
The authors addressed my previous recommendations.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #5 (Public review):
Summary:
In the research article, "Functional genomics reveals strain-specific genetic requirements conferring hypoxic growth in Mycobacterium intracellulare" Tateshi et al focussed their research on pulmonary disease caused by Mycobacterium avium-intracellulare complex which has recently become a major health concern. The authors were interested in identifying the genetic requirements necessary for growth/survival within host and used hypoxia and biofilm conditions that partly replicate some of the stress conditions experienced by bacteria in vivo. An important finding of this analysis was the observation that genes involved in gluconeogenesis, type VII secretion system and cysteine desulphurase were crucial for the clinical isolates during standard culture while the same were necessary during hypoxia in the ATCC type strain.
Strength of the study:
Transposon mutagenesis has been a powerful genetic tool to identify essential genes/pathways necessary for bacteria under various in vitro stress conditions and for in vivo survival. The authors extended the TnSeq methodology not only to the ATCC strain but also to the recently clinical isolates to identify the differences between the two categories of bacterial strains. Using this approach they dissected the similarities and differences in the genetic requirement for bacterial survival between ATCC type strains and clinical isolates. They observed that the clinical strains performed much better in terms of growth during hypoxia than the type strain. These in vitro findings were further extended to mouse infection models and similar outcomes were observed in vivo further emphasising the relevance of hypoxic adaptation crucial for the clinical strains which could be explored as potential drug targets.
Weakness:
The authors have performed extensive TnSeq analysis but fail to present the data coherently. The data could have been well presented both in Figures and text. In my view this is one of the major weakness of the study.
Comments on revisions:
There is quite a lot of data and this could have been a really impactful study if the the authors had channelized the Tn mutagenesis by focussing on one pathway or network. It looks scattered. However, from the previous version, the authors have made significant improvements to the manuscript and have provided comments that fairly address my questions.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this manuscript, an inducible degron approach is taken to investigate the function of the CHD4 chromatin remodelling complex. The cell lines and approaches used are well thought out, and the data appear to be of high quality. They show that loss of CHD4 results in rapid changes to chromatin accessibility at thousands of sites. Of these locations at which chromatin accessibility is decreased are strongly bound by CHD4 prior to activation of the degron, and so likely represent primary sites of action. Somewhat surprisingly, while chromatin accessibility is reduced at these sites, transcription factor occupancy is little changed. Following CHD4 degradation, occupancy of the key pluripotency transcription factors NANOG and SOX2 increases at many locations genome-wide wide and at many of these sites, chromatin accessibility increases. These represent important new insights into the function of CHD4 complexes.
Strengths:
The experimental approach is well-suited to providing insight into a complex regulator such as CHD4. The data generated to characterise how cells respond to the loss of CHD4 is of high quality. The study reveals major changes in transcription factor occupancy following CHD4 depletion.
Weaknesses:
The main weakness can be summarised as relating to the fact that authors interpret all rapid changes following CHD4 degradation as being a direct effect of the loss of CHD4 activity. The possibility that rapid indirect effects arise does not appear to have been given sufficient consideration. This is especially pertinent where effects are reported at sites where CHD4 occupancy is initially low.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors aimed at a comprehensive phenotypic characterization of the roles of all Rab proteins expressed in PN neurons in the developing Drosophila olfactory system. Important data are shown for a number of these Rabs with small/no phenotypes (in the Supplements) as well as the main endosomal Rabs, Rab5, 7, and 11 in the main figures.
Strengths:
The mosaic analysis is a great strength, allowing visualization of small clones or single neuron morphologies. This also allows some assessment of the cell autonomy of the observed phenotypes. The impact of the work lies in the comprehensiveness of the experiments. The rescue experiments are a strength.
Weaknesses:
The main weakness is that the experiments do not address the mechanisms that are affected by the loss of these Rab proteins, especially in terms of the most significant cargos. The insights thus do not extend far beyond what is already known from other work in many systems.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The study investigates the control of the subspaces in which sequences propagate, through static external and dynamic self-generated inhibition. For this, it first uses a 1D ring model with an asymmetry in the weights to evoke a drift of its bump. This model is studied in detail, showing and explaining that the trajectories take place in different subspaces due to the inhibition of different sets of contributing neurons. Sequence propagation is preserved, even if large numbers of neurons are silenced. In this regime, trajectories are restricted to near-orthogonal subspaces of neuronal activity space. The last part of the results shows that similar phenomena can be observed in a 2D spiking neural network model.
Strengths:
The results are important and convincing, and the analyses give a good further insight into the phenomena. The interpretation of inhibited networks as near-circulant is very elucidating. The sparsening by dynamically maintained winner-takes-all inhibition and the transfer to a 2D spiking model are particularly nice results.
Weaknesses:
I see no major weaknesses, except that some crucial literature has not yet been mentioned and discussed. Further, Figure 2c might raise doubts whether the sequences are indeed reliable for the largest amount of sparsening inhibition considered, and it is not yet clear whether the dynamical regime of the 2D model is biologically plausible.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Strengths:
The core strength of this study lies in its innovative demonstration that an engineered sACE2-Fc fusion redirects virus-decoy complexes to Fc-mediated phagocytosis and lysosomal clearance in macrophages, revealing a distinct antiviral mechanism beyond traditional neutralization. Its complete prophylactic protection in animal models and precise targeting of airway phagocytes establish a novel therapeutic paradigm against SARS-CoV-2 variants and future respiratory viruses.
Weaknesses:
The study attributes the complete antiviral protection to Fc-mediated phagocytic clearance, a central claim that requires more rigorous experimental validation. The observation that abrogating Fc functions compromises protection could be confounded by potential alterations in the protein's stability, half-life, or overall structure. To firmly establish this mechanism, it is crucial to include a control molecule with a mutated Fc region that lacks FcγR binding while preserving the Fc structure itself. Without this critical control, the conclusion that phagocytic clearance is the primary mechanism remains inadequately supported. The strategy of deliberately targeting virus-decoy complexes to phagocytes via Fc receptors inherently raises the question of Antibody-Dependent Enhancement (ADE) of disease. While the authors demonstrate a lack of productive infection in macrophages, this only addresses one facet of ADE. The risk of Fc-mediated exacerbation of inflammation (ADE) remains a critical concern. The manuscript would be significantly strengthened by a direct discussion of this risk and by including data, such as cytokine profiling from treated macrophages, to more comprehensively address the safety profile of this approach. The exclusive use of the K18-hACE2 mouse model, which exhibits severe disease, limits the generalizability of the findings. The "complete protection" observed may not translate to models with more robust and naturalistic immune responses or to human physiology. Furthermore, the lack of data on circulating SARS-CoV-2 variants is a concern. The concept of sACE2-Fc fusion proteins as decoy receptors is not novel, and numerous similar constructs have been previously reported. The manuscript would benefit from a clearer demonstration of how the optimized B5-D3 mutant represents a significant advance over existing sACE2-Fc designs. A direct comparative analysis with previously published benchmarks, particularly in terms of neutralizing potency, Fc effector function strength, and in vivo efficacy, is necessary to establish the incremental value and novelty of this specific agent.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Zhu et al. set out to elucidate how the moral emotions of guilt and shame emerge from specific cognitive antecedents - harm and responsibility - and how these emotions subsequently drive compensatory behavior. Consistent with their prediction derived from functionalist theories of emotion, their behavioral findings indicate that guilt is more influenced by harm, whereas shame is more influenced by responsibility. In line with previous research, their results also demonstrate that guilt has a stronger facilitating effect on compensatory behavior than shame. Furthermore, computational modeling and neuroimaging results suggest that individuals integrate harm and responsibility information into a composite representation of the individual's share of the harm caused. Brain areas such as the striatum, insula, temporoparietal junction, lateral prefrontal cortex, and cingulate cortex were implicated in distinct stages of the processing of guilt and/or shame. In general, this work makes an important contribution to the field of moral emotions. Its impact could be further enhanced by clarifying methodological details, offering a more nuanced interpretation of the findings, and discussing their potential practical implications in greater depth.
Strengths:
First, this work conceptualizes guilt and shame as processes unfolding across distinct stages (cognitive appraisal, emotional experience, and behavioral response) and investigates the psychological and neural characteristics associated with their transitions from one stage to the next.
Second, the well-designed experiment effectively manipulates harm and responsibility - two critical antecedents of guilt and shame.
Third, the findings deepen our understanding of the mechanisms underlying guilt and shame beyond what has been established in previous research.
Comments on revisions:
The authors have addressed the issues I raised in the previous review. I have no more comments on the manuscript.
-
-
osf.io osf.io
-
Reviewer #3 (Public review):
Summary & Strengths:
This review by Yu-Tung Li sheds new light on the processes involved in leukocyte extravasation, with a focus on the inter between leukocytes and the extracellular matrix. In doing so, it presents a fresh perspective on the topic of leukocyte extravasation, which has been extensively covered in numerous excellent reviews. Notably, the role of the extracellular matrix in leukocyte extravasation has received relatively little attention until recently. This review synthesizes the substantial knowledge accumulated over the past two decades in a novel and compelling manner.
The author discusses the relevant barriers leukocytes face during extravasation, addresses interactions with and transmigrate through endothelial junctions, mechanisms supporting extravasation, and how minimal plasma leakage is achieved during this process. The question whether extravasation affects leukocyte differentiation and properties is original and thought-provoking and has received limited consideration thus far. The consequences leukocytes extracellular matrix interaction, non-linear responses to substrate stiffness and effects on macrophage polarization, efferocytosis and the outcome of inflammation are relevant topics raised. Finally, a unifying descriptive framework MIKA is introduced, which provides a tool for classifying macrophages based on their expression patterns and could inform the development of targeted therapies aimed at modulating macrophage identity and improving outcomes in inflammatory scenarios.
In summary, this review provides a stimulating perspective on leukocyte extravasation in the context of extracellular matrix biology.
Weaknesses:
One potential drawback of this review is that the attempt to integrate a vast amount of information has resulted in complex figures, which may lead to important details being overlooked by readers.
Tags
Annotators
URL
-
-
osf.io osf.io
-
Reviewer #3 (Public review):
Summary:
In this article, Barnett examines a pressing question regarding citing behavior of authors during the peer review process. In particular, the author studies the interaction between reviewers and authors, focusing on the odds of acceptance, and how this may be affected by whether or not the authors cited the reviewers' prior work, whether the reviewer requested such citations be added, and whether the authors complied/how that affected the reviewer decision-making.
Strengths:
The author uses a clever analytical design, examining four journals that use the same open peer review system, in which the identities of the authors and reviewers are both available and linkable to structured data. Categorical information about the approval is also available as structured data. This design allows a large scale investigation of this question.
Weaknesses:
My original concerns have been largely addressed. Much more detail is provided about the number of documents under consideration for each analysis, which clarifies a great deal.
Much of the observed reviewer behavior disappears or has much lower effect sizes depending on whether "Accept with Reservations" is considered an Accept or a Reject. This is acknowledged in the results text. Language has been toned down in the revised version.
The conditional analysis on the 441 reviews (lines 224-228) does support the revised interpretation as presented.
No additional concerns are noted.
Tags
Annotators
URL
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
This manuscript addresses an important biological question regarding the mechanisms of muscle cell fusion during regeneration. The primary strength of this work lies in the clean and convincing experiments, with the major conclusions being well-supported by the data provided.
The authors have satisfactorily addressed my inquiries.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this manuscript by Qiao et al., the authors seek to uncover force and contractility dynamics that drive tissue morphogenesis, using the Ciona atrial siphon primordium as a model. Specifically, the authors perform a detailed examination of epithelial folding dynamics. Generally, the authors' claims were supported by their data, and the conceptual advances may have broader implications for other epithelial morphogenesis processes in other systems.
Strengths:
The strengths of this manuscript include the variety of experimental and theoretical methods, including generally rigorous imaging and quantitative analyses of actomyosin dynamics during this epithelial folding process, and the derivation of a mathematical model based on their empirical data, which they perturb in order to gain novel insights into the process of epithelial morphogenesis.
Weaknesses:
There are concerns related to wording and interpretations of results, as well as some missing descriptions and details regarding experimental methods.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Overall, the work is fine; however, I find it very preliminary. To the best of my understanding, to make any claims for altered Notch signaling from this study that is physiologically relevant remains to be discerned.
Strengths:
This manuscript systematically analyzes cancer-associated mutations in the Negative Regulatory Region (NRR) of Drosophila Notch to reveal diverse regulatory mechanisms with implications for cancer modelling and therapy development. The study introduces cancer-associated mutations equivalent to human NOTCH1 mutations, covering a broad spectrum across the LNR and HD domains. The authors use rigorous phenotypic assays to classify their functional outcomes. By leveraging the S2 cell-based assay platform, the work identifies mechanistic differences between mutations that disrupt the LNR-HD interface, core HD, and LNR surface domains, enhancing understanding of Notch regulation. The discovery that certain HD and LNR-HD interface mutations (e.g., R1626Q and E1705P) in Drosophila mirror the constitutive activation and synergy with PEST deletion seen in mammalian T-ALL is nice and provides a platform for future cancer modelling. Surface-exposed LNR-C mutations were shown to increase Notch protein stability and decrease turnover, suggesting a previously unappreciated regulatory layer distinct from canonical cleavage-exposure mechanisms. By linking mutant-specific mechanistic diversity to differential signaling properties, the work directly informs targeted approaches for modulating Notch activity in cancer cells.
Weaknesses:
While this is indeed an exciting set of observations, the work is entirely cell-line-based, and is the primary reason why this approach dampens the enthusiasm for the study. The analysis is confined to Drosophila S2 cells, which may not fully recapitulate tissue or organism-level regulatory complexity observed in vivo. Some Drosophila HD domain mutants accumulate in the secretory pathway and do not phenocopy human T-ALL mutations. Possibly due to limitations on physiological inputs that S2 cells cannot account for, or species-specific differences such as the absence of S1 cleavage.
Thus, the findings may not translate directly to understanding Notch 1 function in mammalian cancer models. While the manuscript highlights mechanistic variety, the functional significance of these mutations for hematopoietic malignancies or developmental contexts in live animals remains untested. Overall, the work does not yet provide evidence for altered Notch signaling that is physiologically relevant.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors present a variant of a previously described fluorescence lifetime sensor for calcium. Much of the manuscript describes the process of developing appropriate assays for screening sensor variants, and thorough characterization of those variants (inherent fluorescence characteristics, response to calcium and pH, comparisons to other calcium sensors). The final two figures show how the sensor performs in cultured cells and in vivo drosophila brains.
Strengths:
The work is presented clearly and the conclusion (this is a new calcium sensor that could be useful in some circumstances) is supported by the data.
Weaknesses:
There are probably few circumstances where this sensor would facilitate experiments (calcium measurements) that other sensors would prove insufficient.
Comment on revised version:
I think the manuscript has been significantly improved and I concur with the eLife Assessment statement.
[Editors' note: There are no further requests by the reviewers. All of them expressed their approval of the new version of the manuscript.]
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Here, Bykov et al move the bi-genomic split-GFP system they previously established to the genome-wide level in order to obtain a more comprehensive list of mitochondrial matrix and inner membrane proteins. In this very elegant split-GFP system, the longer GFP fragment, GFP1-10, is encoded in the mitochondrial genome and the shorter one, GFP11, is C-terminally attached to every protein encoded in the genome of yeast Saccharomyces cerevisiae. GFP fluorescence can therefore only be reconstituted if the C-terminus of the protein is present in the mitochondrial matrix, either as part of a soluble protein, a peripheral membrane protein or an integral inner membrane protein. The system, combined with high-throughput fluorescence microscopy of yeast cells grown under six different conditions, enabled the authors to visualize ca. 400 mitochondrial proteins, 50 of which were not visualised before and 8 of which were not shown to be mitochondrial before. The system appears to be particularly well suited for analysis of dually localized proteins and could potentially be used to study sorting pathways of mitochondrial inner membrane proteins.
Strengths:
Many fluorescence-based genome-wide screen were previously performed in yeast and were central to revealing the subcellular location of a large fraction of yeast proteome. Nonetheless, these screens also showed that tagging with full-length fluorescent proteins (FP) can affect both the function and targeting of proteins. The strength of the system used in the current manuscript is that the shorter tag is beneficial for detection of a number of proteins whose targeting and/or function is affected by tagging with full length FPs.
Furthermore, the system used here can nicely detect mitochondrial pools of dually localized proteins. It is especially useful when these pools are minor and their signals are therefore easily masked by the strong signals coming from the major, nonmitochondrial pools of the proteins.
Weaknesses:
My only concern is that the biological significance of the screen performed appears limited. The dataset obtained is largely in agreement with several previous proteomic screens but it is, unfortunately, not more comprehensive than them, rather the opposite. For proteins that were identified inside mitochondria for the first time here or were identified in an unexpected location within the organelle, it remains unclear whether these localizations represent some minor, missorted pools of proteins or are indeed functionally important fractions and/or productive translocation intermediates. The authors also allude to several potential applications of the system but do little to explore any of these directions.
Comments on revisions:
The revised version of the manuscript submitted by Bykov et al addresses the comments and concerns raised by the Reviewers. It is a pity that the verification of the newly obtained data and its further biological exploration is apparently more challenging than perhaps anticipated.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
In this manuscript, Wang et al employ a chemical biology approach to investigate the differences between the enzymatic and scaffolding roles of tankyrase during Wnt β-catenin signalling. It was previously established that, in addition to its enzymatic activity, tankyrase 1/2 also plays a scaffolding function within the destruction complex, a property conferred by SAM-domain-dependent polymerization (PMID: 27494558). It is also known that TNKS1/2 is an autoregulated protein and that its enzymatic inhibition leads to accumulation of total TNKS proteins and stabilization of Axin punctae (through the scaffolding function of TNKS1/2), leading to rigidification of the DC and decreased β-catenin turnover. The authors surmised that this could, in part, explain the limited efficacy of TNKS1/2 catalytic inhibition for the treatment of colorectal cancers. To test this hypothesis, they evaluated a series of PROTAC molecules promoting the degradation of TNKS1/2 to block both the catalytic and scaffolding activities. They show that IWR1-POMA (their most active molecule) promotes more efficient suppression of beta-catenin-mediated transcription and is more active in inhibiting colorectal cancer cell and CRC patient-derived organoids growth. Mechanistically, the authors used FRAP to demonstrate that catalytic inhibitors of TNKS led to a reduced dynamic assembly of the DC (rigidification), whereas IWR1-POMA did not affect the dynamics.
Overall, this is an interesting study describing the design and development of a PROTAC for TNKS1/2 that could have increased efficacy where catalytic inhibitors have displayed limited activity. Knowing the importance of the scaffolding role of TNKS1/2 within the destruction complex, targeting both the catalytic and scaffolding roles certainly makes sense. The manuscript contains convincing evidence of the different mechanisms of the PROTAC vs catalytic inhibitors. Some additional efforts to quantify several of the experiments and to indicate the reproducibility and statistical analysis would strengthen the manuscript. Ultimately, it would have been great to evaluate the in vivo efficacy of IWR1-POMA in an in vivo CRC assay (APCmin mice or using PDX models); however, I realize that this is likely beyond the scope of this manuscript.
I have some recommendations listed below for consideration by the authors to strengthen their study:
(1) The title is slightly misleading, as it is already known that the scaffolding function of TNKS is important within the DC. The authors should consider incorporating the PROTAC targeting aspect in the title (e.g., PROTAC-mediated targeting of tankyrase leads to increased inhibition of betacat signaling and CRC growth inhibition).
(2) The authors should comment in the manuscript on the bell-shaped curve obtained with treatment of cells with the PROTACs (Figure S2C). This likely indicates tittering of the targets within a bifunctional molecule with increasing concentration (and likely reveals the auto-inhibition conferred by the catalytic inhibition alone).
(3) The authors comment that using G007-LK as warehead was unsuccessful, but they do not show data. Do the authors know why this was the case?
(4) Throughout the manuscript, the authors need to do a better job at quantifying their results (i.e., the western blots and the IF). For example, the degradation of TNKS1/2 in Figure 1D is not overly convincing. Similarly, the IF data in Figure 3 needs to be quantified in some ways. Along the same lines, the effect of IWR1-POMA treatments on the proliferation of cells and organoids should be quantified using viability assays... There is also no indication of how many times these experiments were performed and whether the blots shown are representative experiments. The quantification should include all experiments.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The manuscript by Shukla and colleagues presents a comprehensive study that addresses a central question in kinesin-1 regulation - how cargo binding to the kinesin light chain (KLC) tetratricopeptide repeat (TPR) domains triggers activation of full-length kinesin-1 (KHC). The authors combine AlphaFold3 modeling, biophysical analysis (fluorescence polarization, hydrogen-deuterium exchange), and electron microscopy to derive a mechanistic model in which the KLC-TPR domains dock onto coiled-coil 1 (CC1) of the KHC to form the "TPR shoulder," stabilizing the autoinhibited (λ-particle) conformation. Binding of a W/Y-acidic cargo motif (KinTag) or deletion of the CC1 docking site (TDS) dislocates this shoulder, liberating the motor domains and enhancing accessibility to cofactors such as MAP7. The results link cargo recognition to allosteric structural transitions and present a unified model of kinesin-1 activation.
Strengths:
(1) The study addresses a fundamental and long-standing question in kinesin-1 regulation using a multidisciplinary approach that combines structural modeling, quantitative biophysics, and electron microscopy.
(2) The mechanistic model linking cargo-induced dislocation of the TPR shoulder to activation of the motor complex is well supported by both structural and biochemical evidence.
(3) The authors employ elegant protein-engineering strategies (e.g., ElbowLock and ΔTDS constructs) that enable direct testing of model predictions, providing clear mechanistic insight rather than purely correlative data.
(4) The data are internally consistent and align well with previous studies on kinesin-1 regulation and MAP7-mediated activation, strengthening the overall conclusion.
Weaknesses:
(1) While the EM and HDX-MS analyses are informative, the conformational heterogeneity of the complex limits structural resolution, making some aspects of the model (e.g., stoichiometry or symmetry of TPR docking) indirect rather than directly visualized.
(2) The dynamics of KLC-TPR docking and undocking remain incompletely defined; it is unclear whether both TPR domains engage CC1 simultaneously or in an alternating fashion.
(3) The interplay between cargo adaptors and MAP7 is discussed but not experimentally explored, leaving open questions about the sequence and exclusivity of their interactions with CC1.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The study is grounded in the observations that mitochondrial DNA (mtDNA) exhibits a degree of resistance to mutagenesis under genotoxic stress. The manuscript focuses on the effects of UVC-induced DNA damage on TFAM-DNA binding in vitro and in cells. The authors demonstrate increased TFAM-DNA compaction following UVC irradiation in vitro based on high-throughput protein-DNA binding and atomic force microscopy (AFM) experiments. They did not observe a similar trend in fluorescence polarization assays. In cells, the authors found that UVC exposure upregulated TFAM, POLG, and POLRMT mRNA levels without affecting the mitochondrial membrane potential. Overexpressing TFAM in cells or varying TFAM concentration in reconstituted nucleoids did not alter the accumulation or disappearance of mtDNA damage. Based on their data, the authors proposed a plausible model that, following UVC-induced DNA damage, TFAM facilitates nucleoid compaction, which may serve to signal damage in the mitochondrial genome.
Strengths:
The presented data are solid, technically rigorous, and consistent with established literature findings. The experiments are well-executed, providing reliable evidence on the change of TFAM-DNA interactions following UVC irradiation. The proposed model may inspire future follow-up studies to further study the role of TFAM in sensing UVC-induced damage.
Weaknesses:
The manuscript could be further improved by refining specific interpretations and ensuring terminology aligns precisely with the data presented.
(1) In line 322, the claim of increased "nucleoid compaction" in cells should be removed, as there is a lack of direct cellular evidence. Given that non-DNA-bound TFAM is subject to protease digestion, it is uncertain to what extent the overexpressed TFAM actually integrates into and compacts mitochondrial nucleoids in the absence of supporting immunofluorescence data.
(2) In lines 405 and 406, the authors should avoid equating TFAM overexpression with compaction in the cellular context unless the compaction is directly visualized or measured.
(3) In lines 304 and 305 (and several other places throughout the manuscript), the authors use the term "removal rates". A "removal rate" requires a direct comparison of accumulated lesion levels over a time course under different conditions. Given the complexity of UV-induced DNA damage-which involves both damage formation and potential removal via multiple pathways-a more accurate term that reflects the net result of these opposing processes is "accumulated DNA damage levels." This terminology better reflects the final state measured and avoids implying a single, active 'removal' pathway without sufficient kinetic data.
(4) In line 357, the authors refer to the decrease in the total DNA damage level as "The removal of damaged mtDNA". The decrease may be simply due to the turnover and resynthesis of non-damaged mtDNA molecules. The term "removal" may mislead the casual reader into interpreting the effect as an active repair/removal process.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Overall, this is a well-done study, and the conclusions are largely supported by the data, which will be of interest to the field.
Strengths:
Strengths of this study include experiments with solution NMR that can resolve high-resolution interactions of the highly flexible C-terminal tail of arr2 with clathrin and AP2. Although mainly confirmatory in defining the arr2 CBL 376LIELD380 as the clathrin binding site, the use of the NMR is of high interest (Fig. 1). The 15N-labeled CLTC-NTD experiment with arr2 titrations reveals a span from 39-108 that mediates an arr2 interaction, which corroborates previous crystal data, but does not reveal a second area in CLTC-NTD that in previous crystal structures was observed to interact with arr2.
SEC and NMR data suggest that full-length arr2 (1-418) binding with 2-adaptin subunit of AP2 is enhanced in the presence of CCR5 phospho-peptides (Fig. 3). The pp6 peptide shows the highest degree of arr2 activation, and 2-adaptin binding, compared to less phosphorylated peptide or not phosphorylated at all. It is interesting that the arr2 interaction with CLTC NTD and pp6 cannot be detected using the SEC approach, further suggesting that clathrin binding is not dependent on arrestin activation. Overall, the data suggest that receptor activation promotes arrestin binding to AP2, not clathrin, suggesting the AP2 interaction is necessary for CCR5 endocytosis.
To validate the solid biophysical data, the authors pursue validation experiments in a HeLa cell model by confocal microscopy. This requires transient transfection of tagged receptor (CCR5-Flag) and arr2 (arr2-YFP). CCR5 displays a "class B"-like behavior in that arr2 is rapidly recruited to the receptor at the plasma membrane upon agonist activation, which forms a stable complex that internalizes onto endosomes (Fig. 4). The data suggest that complex internalization is dependent on AP2 binding not clathrin (Fig. 5).
The addition of the antagonist experiment/data adds rigor to the study.
Overall, this is a solid study that will be of interest to the field.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Shimogawa et al. describe the generation of acetylated aSyn variants by genetic code expansion to elucidate effects on vesicle binding, aggregation, and seeding effects. The authors compared a semi-synthetic approach to obtain acetylated aSyn variants with genetic code expansion and concluded that the latter was more efficient in generating all 12 variants studied here, despite the low yields for some of them. Selected acetylated variants were used in advanced NMR, FCS, and cryo-EM experiments to elucidate structural and functional changes caused by acetylation of aSyn. Finally, site-specific differences in deacetylation by HDAC 8 were identified.
The study is of high scientific quality, andthe results are convincingly supported by the experimental data provided. The challenges the authors report regarding semi-synthetic access to aSyn are somewhat surprising, as this protein has been made by a variety of different semi-synthesis strategies in satisfactory yields and without similar problems being reported.
The role of PTMs such as acetylation in neurodegenerative diseases is of high relevance for the field, and a particular strength of this study is the use of authentic acetylated aSyn instead of acetylation-mimicking mutations. The finding that certain lysine acetylations can slow down aggregation even when present only at 10-25% of total aSyn is exciting and bears some potential for diagnostics and therapeutic intervention.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary
Following recent findings that exposure to natural sounds and anthropogenic noise before hatching affects development and fitness in an altricial songbird, this study attempts to estimate the hearing capacities of zebra finch nestlings and the perception of high frequencies in that species. It also tries to estimate whether airborne sound can make zebra finch eggs vibrate, although this is not relevant to the question.
Strength
That prenatal sounds can affect the development of altricial birds clearly challenges the long-held assumption that altricial avian embryos cannot hear. However, there is currently no data to support that expectation. Investigating the development of hearing in songbirds is therefore important, even though technically challenging. More broadly, there is accumulating evidence that some bird species use sounds beyond their known hearing range (especially towards high frequencies), which also calls for a reassessment of avian auditory perception.
Weaknesses
Rather than following validated protocols, the study presents many experimental flaws and two major methodological mistakes (see below), which invalidate all results on responses to frequency-specific tones in nestlings and those on vibration transmission to eggs, as well as largely underestimating hearing sensitivity. Accordingly, the study fails to detect a response in the majority of individuals tested with tones, including adults, and the results are overall inconsistent with previous studies in songbirds. The text throughout the preprint is also highly inaccurate, often presenting only part of the evidence or misrepresenting previous findings (both qualitatively and quantitatively; some examples are given below), which alters the conclusions.
Conclusion and impact
The conclusion from this study is not supported by the evidence. Even if the experiment had been performed correctly, there are well-recognised limitations and challenges of the method that likely explain the lack of response. The preprint fails to acknowledge that the method is well-known for largely underestimating hearing threshold (by 20-40dB in animals) and that it may not be suitable for a 1-gram hatchling. Unlike what is claimed throughout, including in the title, the failure to detect hearing sensitivity in this study does not invalidate all previous findings documenting the impacts of prenatal sound and noise on songbird development. The limitations of the approach and of this study are a much more parsimonious explanation. The incorrect results and interpretations, and the flawed representation of current knowledge, mean that this preprint regrettably creates more confusion than it advances the field.
Detailed assessment
For brevity, only some references are included below as examples, using, when possible, those cited in the preprint (DOI is provided otherwise). A full review of all the studies supporting the points below is beyond the scope of this assessment.
(A) Hearing experiment
The study uses the Auditory Brainstem Response (ABR), which measures minute electrical signals transmitted to the surface of the skull from the auditory nerve and nuclei in the brainstem. ABR is widely used, especially in humans, because it is non-invasive. However, ABR is also a lot less sensitive than other methods, and requires very specific experimental precautions to reliably detect a response, especially in extremely small animals and with high-frequency sounds, as here.
(1) Results on nestling frequency sensitivity are invalid, for failing to follow correct protocols:
The results on frequency testing in nestlings are invalid, since what might serve as a positive control did not work: in adults, no response was detected in a majority of individuals, at the core of their hearing range, with loud 95dB sounds (Figure S1), when testing frequency sensitivity with "tone burst".
This is mostly because the study used a stimulation duration 5 times larger than the norm. It used 25ms tone bursts, when all published avian studies (in altricial or precocial birds) used stimulation of 5ms or less (when using subdermal electrodes as here; e.g., cited: Brittan-Powell et al 2004; not cited: Brittan-Powell et al 2002 (doi: 10.1121/1.1494807), Henry & Lucas 2008 (doi: 10.1016/j.anbehav.2008.08.003)). Long stimulations do not make sense and are indeed known to interfere with the detection of an ABR response, especially at high frequencies, as, for example, explicitly tested and stated in Lauridsen et al 2021 (cited).
Adult response was then re-tested with a correct 5ms tone duration ("tone-pip"), which showed that, for the few individuals that responded to 25ms tones, thresholds were abnormally high (c.a. by 30dB; Figure 2C).<br /> Yet, no nestlings were retested with a correct protocol. There is therefore no valid data to support any conclusion on nestling frequency hearing. Under these circumstances, the fact that some nestlings showed a response to 25ms tones from day 8 would argue against them having very low sensitivity to sound.
(2) Responses to clicks underestimate hearing onset by several days:
Without any valid nestling responses to tones (see # 1), establishing the onset of hearing is not possible based on responses to clicks only, since responses to clicks occur at least 4 days after responses to tones during development (Saunders et al, 1973). Here, 60% of 4-day-old individuals responding to clicks means most would have responded to tones at and before 2 days post-hatch, had the experiment been done correctly.<br /> Responses to tones are indeed observed in other songbirds at 1day post-hatch (see #6).
In budgerigars, hearing onset occurs before 5 days post hatch, since responses to both clicks and tones were detectable at the first age tested at 5dph (Brittan-Powell et al, 2004).
(3) Experimental parameters chosen lower ABR detectability, specifically in younger birds:
Very fast stimulus repetition rate inhibits the ABR response, especially in young:
(a) The stimulus presentation rate (25 stim/ sec) is 6 times faster than zebra finch heat-calls, and 5 to 25 times faster than most previous studies in young birds (e.g., cited: Saunders et al 1973, 1974: 1 stim/sec or less; Katayama 1985: 3.3 clicks/sec; Brittan-Powell et al 2004: 4 stim/sec). Faster rates saturate the neurons and accordingly are known to decrease ABR amplitude and increase ABR latency, especially in younger animals with an immature nervous system. In birds, this occurs especially in the range from 5 to 30 stim/sec (e.g., cited: Saunder et al 1973, Brittan-Powell et al 2004). Values here with 25 rather than 1-4 stim/min are therefore underestimating true sensitivity.
(b) Averaging over only 400 measures is insufficient to reliably detect weak ABR signals:
The study uses 2 to 3 times fewer measures per stimulation type than the recommended value of 1,000 (e.g., Brittan-Powell et al 2002, 2024; Henry & Lucas 2008). This specifically affects the detection of weak signals, as in small hatchlings with tiny brains (adult zebra finches are 12-14g).
(c) Body temperature is not specified and strongly affects the ABR:
Controlling the body temperature of hatchlings of 1-4 grams (with a temperature probe under a 5mm-wide wing) would be very challenging. Low body temperature entirely eliminates the ABR, and even slight deviance from optimal temperature strongly increases wave latency and decreases wave amplitude (e.g., cited: Katayama 1985).
(d) Other essential information is missing on parameters known to affect the ABR:
This includes i) the weight of the animals, ii) whether and how the response signal was amplified and filtered, iii) how the automatised S/N>2 criteria compared to visual assessment for wave detection, and iv) what measures were taken to allow the correct placement of electrodes on hatchlings less than 5 grams.
(4) Results in adults largely underestimate sensitivity at high frequencies, and are not the correct reference point:
(a) Thresholds measured here at high frequencies for adults (using the correct stimulus duration, only done on adults) are 10-30dB higher than in all 3 other published ABR studies in adult zebra finches (cited: Zevin et al 2004; Amin et al 2007; not cited: Noirot et al 2011 (10.1121/1.3578452)), for both 4 and 6 kHz tone pips.
(b) The underlying assumption used throughout the preprint that hearing must be adult-like to be functional in nestlings does not make sense. Slower and smaller neural responses are characteristic of immature systems, but it does not mean signals are not being perceived.
(5) Failure to account for ABR underestimation leads to false conclusions:
(a) Whether the ABR method is suitable to assess hearing in very small hatchlings is unknown. No previous avian study has used ABR before 5 days post-hatch, and all have used larger bird species than the zebra finch.
(b) Even when performed correctly on large enough animals, the ABR systematically underestimates actual auditory sensitivity by 20-40 dB, especially at high frequencies, compared to behavioural responses (e.g., none cited: Brittan-Powell et al 2002, Henry & Lucas 2008, Noirot et al 2011). Against common practice, the preprint fails to account for this, leading to wrong interpretations. For example, in Figure 1G (comparing to heat call levels), actual hearing thresholds would be 30-40dB below those displayed. In addition, the "heat whistle" level displayed here (from the same authors) is 15dB lower than their second measure that they do not mention, and than measures obtained by others (unpublished data). When these two corrections are made - or even just the first one - the conclusion that heat-call sound levels are below the zebra finch hearing threshold does not hold.
(c) Rather than making appropriate corrections, the preprint uses a reference in humans (L180), where ABR is measured using a much more powerful method (multi-array EEG) than in animals, and from a larger brain. The shift of "10-20dB" obtained in humans is not applicable to animals.
(6) Results are inconsistent with previous findings in developing songbirds:
As expected from all of the above, results and conclusions in the preprint are inconsistent with findings in other songbirds, which, using other methods, show for example, auditory sensitivity in:
(a) zebra finch embryos, in response to song vs silence (not cited: Rivera et al 2018, doi: 10.1097/WNR.0000000000001187)
(b) flycatcher hatchlings at 2-3d post hatch (first age tested), across a wide range of frequencies (0.3 to 5kHz), at low to moderate sound levels (45-65dB) (cited: Aleksandrov and Dmitrieva 1992, not cited: Korneeva et al 2006 (10.1134/S0022093006060056)).
(c) songbird nestlings at 2-6d post hatch, which discriminate and behaviourally respond to relevant parental calls or even complex songs. This level of discrimination requires good hearing across frequencies (e.g., not cited: Korneeva et al 2006; Schroeder & Podos 2023 (doi: 10.1016/j.anbehav.2023.06.015)).
(d) zebra finch nestlings at 13d post-hatch, which show adult-like processing of songs in the auditory cortex (CNM) (Schroeder & Remage‐Healey 2021, doi: 10.1002/dneu.22802).
(e) zebra finch juveniles, which are able to perceive and learn song syllables at 5-7kHz (fundamental frequency) with very similar acoustic properties to heat calls, and also produced during inspiration (Goller & Daley 2001, doi: 10.1098/rspb.2001.1805).
NONE of these results - which contradict results and claims in the preprint - are mentioned. Instead, the preprint focuses on very slow-developing species (parrots and owls), which take 2-4 times longer than songbirds to fledge (cited: Brittan-Powell et al 2004; Köppl & Nickel 2007; Kraemer et al 2017).
(7) Results in figures are misreported in the text, and conclusions in the abstract and headers are not supported by the data:
For example:
(a) The data on Figure 1E shows that at 4 days old, 8 out of 13 nestlings (60%) responded to clicks, but the text says only 5/13 responded (L89). When 60% (4dph) and 90% (6dph) of individuals responded, the correct term would be that "most animals", rather than "some animals" responded (L89). Saying that ABR to loud sound appeared "in the majority only after one week" (L93) is also incorrect, given the data. It follows that the title of the paragraph is also erroneous.
(b) The hearing threshold is underestimated by 40dB at 6 and 8Kz on Fig 2C, not by "10-20dB" as reported in the text (L178).
(B) Egg vibration experiment
(8) Using airborne sound to vibrate eggs is biologically irrelevant:
The measurement of airborne sound levels to vibrate eggs misunderstands bone conduction hearing and is not biologically meaningful: zebra finch parents are in direct contact with the eggs when producing heat calls during incubation, not hovering in front of the nest. This misunderstanding affects all extrapolations from this study to findings in studies on prenatal communication.
(C) Misrepresentation of current knowledge
(9) Values from published papers are misreported, which reverses the conclusions:
Most critical examples:
(a) Preprint: "Zebra finch most sensitive hearing range of 1-to-4 kHz (Amin et al., 2007; Okanoya and Dooling, 1987; Yeh et al., 2023)" (L173).<br /> Actual values in the studies cited are:
1-to-7kHz, in Amin et al 2007 (threshold [=50dB with ABR] is the same at 7kHz and 1KHz).
1-to-6 kHz, in Okanoya and Dooling (the threshold [=30dB with behaviour] is actually lower at 6kHz than at 1KHz).
1-to-7kHz, in Yeh et al (threshold [=35-38dB with behaviour] is the same at 7kHz and 1KHz).
Note that zebra finch nestlings' begging calls peaking at 6kHz (Elie & Theunissen 2015, doi: 10.1007/s10071-015-0933-6), would fall 2kHz above the parents' best hearing range if it were only up to 4kHz.
(b) The preprint incorrectly states throughout (e.g., L139, L163, L248) that heat-calls are 7-10kHz, when the actual value is 6-10kHz in the paper cited (Katsis et al, 2018).
(c) Using the correct values from these studies, and heat-calls at 45 dB SLP (as measured by others (unpublished data), or as measured by the authors themselves, but which is not reported here (Anttonen et a,l 2025), the correct conclusion is that heat calls fall within the known zebra finch hearing range.
(10) Published evidence towards high-frequency hearing, including in early development, is systematically omitted:
(a) Other studies showing birds use high frequencies above the known avian hearing range are ignored. This includes oilbirds (7-23kHz; Brinklov et al 2017; by 1 of the preprint authors, doi: 10.1098/rsos.170255) and hummingbirds (10-20kHz; Duque et al 2020, doi: 10.1126/sciadv.abb9393), and in a lesser extreme, zebra finches' inspiratory song syllables at 5-7kHz (Goller & Dalley, 2001).
(b) The discussion of anatomical development (L228-241) completely omits the well-known fact that the avian basilar papilla develops from high to low frequencies (i.e., base to apex), which - as many have pointed out - is opposite to the low-to-high development of sensitivity (e.g., cited: Cohen & Fermin 1978; Caus Capdevila et al 2021).
(c) High frequency hearing in songbirds at hatching is several orders of magnitude better than in chickens and ducks at the same age, even though songbirds are altricial (e.g., at 4kHz, flycatcher: 47dB, chicken-duck: 90dB; at 5kHz, flycatcher: 65dB, chicken-duck: 115dB; Korneeva et al 2006, Saunders et al 1974). That is because Galliformes are low-frequency specialists, according to both anatomical and ecological evidence, with calls peaking at 0.8 to 1.2kHz rather than 2-6kHz in songbirds. It is incorrect to conclude that altricial embryos cannot perceive high frequencies because low-frequency specialist precocial birds do not (L250;261).
The references used to support the statement on a very high threshold for precocial birds above 6kHz are also wrong (L250). Katayama 1985 did not test embryos, nor frequency tones. Neither of these two references tested ducks.
(11) Incorrect statements do not reflect findings from the references cited
For example:
(a) "in altricial bird species hearing typically starts after hatching" (L12, in abstract), "with little to no functional hearing during embryonic stages (Woolley, 2017)." (L33).
There is no evidence, in any species, to support these statements. This is only a - commonly repeated - assumption, not actually based on any data. On the contrary, the extremely limited evidence to date shows the opposite, with zebra finch embryos showing ZENK activation in the auditory cortex in response to song playback (Rivera et al, 2018, not cited).
The book chapter cited (Woolley 2017) acknowledges this lack of evidence, and, in the context of song learning, provides as only references (prior to 2018), 2 studies showing that songbirds do not develop a normal song if the song tutor is removed before 10d post-hatch. That nestlings cannot memorise (to later reproduce) complex signals heard before d10 does not mean that they are deaf to any sound before day 10.
Studies showing hearing in young songbird nestlings (see point 6 above) also contradict these statements.
(b) "Zebra finch embryos supposedly are epigenetically guided to adapt to high temperatures by their parents high-frequency "heat calls" " (L36 and L135).
This is an extremely vague and meaningless description of these results, which cannot be assessed by readers, even though these results are presented as a major justification for the present study. Rather than giving an interpretation of what "supposedly" may occur, it would be appropriate to simply synthesize the empirical evidence provided in these papers. They showed that embryonic exposure to heat-calls, as opposed to control contact calls, alters a suite of physiological and behavioural traits in nestlings, including how growth and cellular physiology respond to high temperatures. This also leads to carry-over effects on song learning and reproductive fitness in adulthood.
(c) "The acoustic communication in precocial mallard ducks depends specifically on the low-frequency auditory sensitivity of the embryo (Gottlieb, 1975)" (L253)
The study cited (Gottlieb, 1975) demonstrates exactly the opposite of this statement: it shows that duckling embryos, not only perceive high frequency sounds (relative to the species frequency range), but also NEED this exposure to display normal audition and behaviour post-hatch. Specifically, it shows that duckling embryos deprived of exposure to their own high-frequency calls (at 2 kHz), failed to identify maternal calls post-hatch because of their abnormal insensitivity to higher frequencies, which was later confirmed by directly testing their auditory perception of tones (Dimitrieva & Gottlieb, 1994).
(12) Considering all of the mistakes and distortions highlighted above, it would be very premature to conclude, based on these results and statements, that altricial avian embryos are not sensitive to sound. This study provides no actual scientific ground to support this conclusion.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public Review):
The article presents a comprehensive study on the stratification of viral shedding patterns in saliva among COVID-19 patients. The authors analyze longitudinal viral load data from 144 mildly symptomatic patients using a mathematical model, identifying three distinct groups based on the duration of viral shedding. Despite analyzing a wide range of clinical data and micro-RNA expression levels, the study could not find significant predictors for the stratified shedding patterns, highlighting the complexity of SARS-CoV-2 dynamics in saliva. The research underscores the need for identifying biomarkers to improve public health interventions and acknowledges several limitations, including the lack of consideration of recent variants, the sparsity of information before symptom onset, and the focus on symptomatic infections.
The manuscript is well-written, with the potential for enhanced clarity in explaining statistical methodologies. This work could inform public health strategies and diagnostic testing approaches.
Comments on the revised version from the editor:
The authors comprehensively addressed the concerns of all 3 reviewers. We are thankful for their considerable efforts to do so. Certain limitations remain unavoidable such as the lack of immunologic diversity among included study participants and lack of contemporaneous variants of concern.
One remaining issue is the continued use of the target cell limited model which is sufficient in most cases, but misses key datapoints in certain participants. In particular, viral rebound is poorly described by this model. Even if viral rebound does not place these cases in a unique cluster, it is well understood that viral rebound is of clinical significance.
In addition, the use of microRNAs as a potential biomarker is still not fully justified. In other words, are there specific microRNAs that have a pre-existing mechanistic basis for relating to higher or lower viral loads? As written it still feels like microRNA was included in the analysis simply because the data existed.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
This manuscript provides evidence that mice have a fusome, a conserved structure most well studied in Drosophila that is important for oocyte specification. Overall, a myriad of evidence is presented demonstrating the existence of a mouse fusome that the authors term visham. This work is important as it addresses a long-standing question in the field of whether mice have fusomes and sheds light on how oocytes are specified in mammals. Concerns that need to be addressed revolve around several conclusions that are overstated or unclear and are listed below.
(1) Line 86 - the heading for this section is "PGCs contain a Golgi-rich structure known as the EMA granule" but there is nothing in this section that shows it is Golgi-rich. It does show that the structure is asymmetric and has branches.
(2) Line 105-106, how do we know if what's seen by EM corresponds to the EMA1 granule?
(3) Line 106-107-states "Visham co-stained with the Golgi protein Gm130 and the recycling endosomal protein Rab11a1". This is not convincing as there is only one example of each image, and both appear to be distorted.
(4) Line 132-133---while visham formation is disrupted when microtubules are disrupted, I am not convinced that visham moves on microtubules as stated in the heading of this section.
(5) Line 156 - the heading for this section states that Visham associates with polarity and microtubule genes, including pard3, but only evidence for pard3 is presented.
(6) Lines 196-210 - it's strange to say that UPR genes depend on DAZ, as they are upregulated in the mutants. I think there are important observations here, but it's unclear what is being concluded.
(7) Line 257-259---wave 1 and 2 follicles need to be explained in the introduction, and how this fits with the observations here clarified.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors dilute fluorescent HCMV stocks in small steps (df ≈ 1.3-1.5) across 23 points, quantify infections by flow cytometry at 3 dpi, and fit a power-law model to estimate a cooperativity parameter n (n > 1 indicates apparent cooperativity). They compare fibroblasts vs epithelial cells and multiple strains/reporters, and explore alternative mechanisms (clumping, accrued damage, viral compensation) via analytical modeling and stochastic simulations. They discuss implications for titer/MOI estimation and suggest a method for detecting "apparent cooperativity," noting that for viruses showing this behavior, MOI estimation may be biased.
Strengths:
(1) High-resolution titration & rigor: The small-step dilution design (23 serial dilutions; tailored df) improves dose-response resolution beyond conventional 10× series.
(2) Clear quantitative signal: Multiple strain-cell pairs show n > 1, with appropriate model fitting and visualization of the linear regime on log-log axes.
(3) Mechanistic exploration: Side-by-side modeling of clumping vs accrued damage vs compensation frames testable hypotheses for cooperativity.
Weaknesses:
(1) Secondary infection control: The authors argue that 3 dpi largely avoids progeny-mediated secondary infection; this claim should be strengthened (e.g., entry inhibitors/control infections) or add sensitivity checks showing results are robust to a small secondary-infection contribution.
(2) Discriminating mechanisms: At present, simulations cannot distinguish between accrued damage and viral compensation. The authors should propose or add a decisive experiment (e.g., dual-color coinfection to quantify true coinfection rates versus "priming" without coinfection; timed sequential inocula) and outline expected signatures for each mechanism.
(3) Decline at high genomes/cell: Several datasets show a downturn at high input. Hypotheses should be provided (cytotoxicity, receptor depletion, and measurement ceiling) and any supportive controls.
(4) Include experimental data: In Figure 6, please include the experimentally measured titers (IU/mL), if available.
(5) MOI guidance: The practical guidance is important; please add a short "best-practice box" (how to determine titer at multiple genomes/cell and cell densities; when single-hit assumptions fail) for end-users.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This study from Jia et al carried out a variety of analyses of terminating ribosomes, including the development of eRF1-seq to map termination sites, identification of a GA-rich motif that promotes ribosome pausing, characterization of tissue-specific termination dynamics, and elucidation of the regulatory roles of 18S rRNA and RPS26. Overall, the study is thoughtfully designed, and its biological conclusions are well supported by complementary experiments. The tools and datasets generated provide valuable resources for researchers investigating the mechanisms of RNA translation.
Strengths:
(1) The study introduces eRF1-seq, a novel approach for mapping translation termination sites, providing a methodological advance for studying ribosome termination.
(2) Through integrative bioinformatic analyses and complementary MPRA experiments, the authors demonstrate that GA-rich motifs promote ribosome pausing at termination sites and reveal possible regulatory roles of 18S rRNA in this process.
(3) The study characterizes tissue-specific ribosome termination dynamics, showing that the testis exhibits stronger ribosome pausing at stop codons compared to other tissues. Follow-up experiments suggest that RPS26 may contribute to this tissue specificity.
Weaknesses:
The biological significance of ribosome pausing regulation at translation termination sites or of translational readthrough, for example, across different tissue types, remains unclear. Nevertheless, this question lies beyond the primary scope of the current study.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this manuscript, the authors define a new paradigm for the attachment and endocytosis of SARS-CoV-2 in which cell surface heparan sulfate (HS) is the primary receptor, with ACE2 having a downstream role within endocytic vesicles. This has implications for the importance of targeting virion-HS interactions as a therapeutic strategy.
Strengths:
The authors show that viruses are internalized via dynamin-dependent endocytosis and that endocytic internalization is the major pathway for pseudotyped SARS-CoV-2 genome expression. They show that HS-mediated viral attachment is a critical step preceding viral endocytosis and also subsequent genome expression. Further, they show that hACE2 acts downstream of endocytosis to promote viral infection, and may be co-internalised with virions after HS attachment. Pseudotyped virus and authentic SARS-CoV-2 provide similar results. In addition, the authors demonstrate that remarkable clusters of multiple HS chains exist on the cell surface, visualised by a number of elegant microscopy methods, and that these represent the docking sites for virions. These visualisations are an important general contribution in themselves to understanding the nanoscale interactions of HS at the cell surface.
The use of a complementary range of methods, virus constructs, and cell models is a strength, and the results clearly support the conclusions.
Overall, the results convincingly demonstrate a different model to the currently accepted mechanism in which the ACE2 protein is regarded as the cell surface receptor for SARS-CoV-2. Here, the authors provide compelling evidence that cell surface clusters of HS are the primary docking site, with ACE2 interactions occurring later, after endocytosis (whilst still being essential for viral genome expression). This is an exciting and important landmark evidence which supports the view that HS-virion interactions should be viewed as a key site for anti-viral drug targeting, likely in strategies that also target the downstream ACE2-based mechanism of viral entry within endosomes.
Weaknesses:
This reviewer identified only minor points regarding citing and discussing other studies and typos, which can be corrected.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
In this paper, authors aimed to investigate carbamylation effects on the function of Cx43-based hemichannels. Such effects have previously been characterized for other connexins, e.g. for Cx26, which display increased hemichannel (HC) opening and closure of gap junction channels upon exposure to increased CO2 partial pressure (accompanied by increased bicarbonate to keep pH constant). The authors used HeLa cells transiently transfected with Cx43 to investigate CO2-dependent carbamylation effects on Cx43 HC function. In contrast to Cx43-based gap junction channels that are here reported to be insensitive to PCO2 alterations, they provide evidence that Cx43 HC opening is highly dependent on the PCO2 pressure in the bath solution, over a range of 20 up to 70 mmHg encompassing the physiologically normal resting level of around 40 mmHg. They furthermore identified several Cx43 residues involved in Cx43 HC sensitivity to PCO2: K105, K109, K144 & K234; mutation of 2 or more of these AAs is necessary to abolish CO2 sensitivity. The subject is interesting and the results indicate that a fraction of HCs is open at a physiological 40 mmHg PCO2, which differs from the situation under HEPES buffered solutions where HCs are mostly closed under resting conditions. The mechanism of HC opening with CO2 gassing is linked to carbamylation and authors pinpointed several Lys residues involved in this process. Overall, the work is interesting as it shows that Cx43 HCs have a significant open probability under resting conditions of physiological levels of CO2 gassing, probably applicable to/relevant for brain, heart and other Cx43 expressing organs. The paper gives a detailed account on various experiments performed (dye uptake, electrophysiology, ATP release to assess HC function) and results concluded from those. They further consider many candidate carbamylation sites by mutating them to negatively charged Glu residues. The paper finalizes with hippocampal slice work showing evidence for connexin-dependent increases of the EPSP amplitude that could be inhibited by HC inhibition with Gap26 (Fig. 10). Another line of evidence comes from the Cx43-linked ODDD genetic disease whereby L90V as well as the A44V mutations of Cx43 prevented the CO2 induced hemichannel opening response (Fig. 11). Although the paper is interesting, in its present state it suffers from (i) a problematic Fig. 3, precluding interpretation of the data shown, and (ii) the poor use of hemichannel inhibitors that are necessary to strengthen the evidence in the crucial experiment of Fig. 2 and others.
Comments on revisions:
The traces in Fig.2B show that the HC current is inward at 20 mmHg PCO2, while it switches to an outward current at 55mmHg PCO2. HCs are non-selective channels, so their current should switch direction around 0 mV but not around -50 mV. As such, the -50 mV switching point indicates involvement of another channel distinct from non-selective Cx43 hemichannels. In the revised version, this problem has not been solved nor addressed. Additionally, I identified another problem in that the experimental traces shown lack a trace at the baseline condition of PCO2 35mmHg, while the summary graph depicts a data point. Not showing a trace at baseline PCO2 35mmHg renders data interpretation in the summary graph questionable.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this manuscript Pinon et al. describe the development of a 3D model of human vasculature within a microchip to study Neisseria meningitidis (Nm)- host interactions and validate it through its comparison to the current gold-standard model consisting of human skin engrafted onto a mouse. There is a pressing need for robust biomimetic models with which to study Nm-host interactions because Nm is a human-specific pathogen for which research has been primarily limited to simple 2D human cell culture assays. Their investigation relies primarily on data derived from microscopy and its quantitative analysis, which support the authors' goal of validating their Vessel-on-Chip (VOC) as a useful tool for studying vascular infections by Nm, and by extension, other pathogens associated with blood vessels.
Strengths:
• Introduces a novel human in vitro system that promotes control of experimental variables and permits greater quantitative analysis than previous models<br /> • The VOC model is validated by direct comparison to the state-of-the-art human skin graft on mouse model<br /> • The authors make significant efforts to quantify, model, and statistically analyze their data<br /> • The laser ablation approach permits defining custom vascular architecture<br /> • The VOC model permits the addition and/or alteration of cell types and microbes added to the model<br /> • The VOC model permits the establishment of an endothelium developed by shear stress and active infusion of reagents into the system
Weaknesses:
• The VOC model contains one cell type, human umbilical cord vascular endothelial cells (HUVECs), while true vasculature contains a number of other cell types that associate with and affect the endothelium, such as smooth muscle cells, pericytes, and components of the immune system. However, adding such complexity may be a future goal of this VOC model.
Impact:
The VOC model presented by Pinon et al. is an exciting advancement in the set of tools available to study human pathogens interacting with the vasculature. This manuscript focuses on validating the model, and as such sets the foundation for impactful research in the future. Of particular value is the photoablation technique that permits the custom design of vascular architecture without the use of artificial scaffolding structures described in previously published works.
Comments on revised version:
The authors have nicely addressed my (and other reviewers') comments.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Fengwen Huang et al. used multiple neuroscience techniques (transgenetic mouse, immunochemistry, bulk calcium recording, neural sensor, hippocampal-dependent task, optogenetics, chemogenetics, and interfer RNA technique) to elucidate the role of the excitatory cholecystokinin-positive pyramidal neurons in the hippocampus in regulating the hippocampal functions, including navigation and neuroplasticity.
Strengths:
(1) The authors provided the distribution profiles of excitatory cholecystokinin in the dorsal hippocampus via the transgenetic mice (Ai14::CCK Cre mice), immunochemistry, and retrograde AAV.
(2) The authors used the neural sensor and light stimulation to monitor the CCK release from the CA3 area, indicating that CCK can be secreted by activation of the excitatory CCK neurons.
(3) The authors showed that the activity of the excitatory CCK neurons in CA3 is necessary for navigation learning.
(4) The authors demonstrated that inhibition of the excitatory CCK neurons and knockdown of the CCK gene expression in CA3 impaired the navigation learning and the neuroplasticity of CA3-CA1 projections.
Weaknesses:
(1) The causal relationship between navigation learning and CCK secretion?
(2) The effect of overexpression of the CCK gene on hippocampal functions?
(3) What are the functional differences between the excitatory and inhibitory CCK neurons in the hippocampus?
(4) Do CCK sources come from the local CA3 or entorhinal cortex (EC) during the high-frequency electrical stimulation?
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This is a clearly written paper that describes the reanalysis of data from a BXD study of the locomotor response to morphine and naloxone. The authors detect significant loci and an epistatic interaction between two of those loci. Single-cell data from outbred rats is used to investigate the interaction. The authors also use network methods and incorporate human data into their analysis.
Strengths:
One major strength of this work is the use of granular time-series data, enabling the identification of time-point-specific QTL. This allowed for the identification of an additional, distinct QTL (the Fgf12 locus) in this work compared to previously published analysis of these data, as well as the identification of an epistatic effect between Oprm1 (driving early stages of locomotor activation) and Fgf12 (driving later stages).
Weaknesses:
(1) What criteria were used to determine whether the epistatic interaction was significant? How many possible interactions were explored?
(2) Results are presented for males and females separately, but the decision to examine the two sexes separately was never explained or justified. Since it is not standard to perform GWAS broken down by sex, some initial explanation of this decision is needed. Perhaps the discussion could also discuss what (if anything) was learned as a result of the sex-specific analysis. In the end, was it useful?
(3) The confidence intervals for the results were not well described, although I do see them in one of the tables. The authors used a 1.5 support interval, but didn't offer any justification for this decision. Is that a 95% confidence interval? If not, should more consideration have been given to genes outside that interval? For some of the QTLs that are not the focus of this paper, the confidence intervals were very large (>10 Mb). Is that typical for BXDs?
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In the submitted article by Lewis et al., the authors investigate how mechanical stimulation influences organ regeneration using the well-characterized zebrafish caudal fin regeneration model. Using a swim flume and a 30min/day exercise regime, the authors found that exercise during the establishment of the blastema reduced regeneration and led to skeletal deformations. Transcriptional profiling of regenerated caudal fin tissue revealed reduced expression of extracellular matrix-associated genes, which were found to be expressed by blastemal fibroblast and osteoblast lineage cells.
Downregulated genes included hyaluronic acid synthases 1 and 2; accordingly, hyaluronic acid levels were found to be reduced in regenerating fins exposed to exercise. The link between regeneration and HA was further confirmed through HA depletion and HA overexpression experiments, which showed a reduction in blastema size and partial rescue of blastema formation, respectively. The authors further show that HA levels, as well as the extent of mechanical loading correlate with nuclear localization of the mechanotransducer Yap and conclude that biomechanical forces play a significant role during regeneration through regulation of HA levels in the ECM and therewith regulation of YAP downstream signaling.
This work expands our understanding of the biochemical signaling connecting biomechanical forces with tissue regeneration. The conclusions are well supported by the data.
Strengths:
(1) Analysis is performed in multiple replicate experimental groups and shows the robust response to the experimental conditions.
(2) The link of HA levels to blastema formation was confirmed through HA overexpression and two different HA depletion experiments.
(3) The use of a previously established fin regeneration single cell dataset does elegantly show the correlation of changes in gene expression levels and specific tissue types, which was further confirmed by in vivo imaging of cell type-specific transgenic lines.
Weaknesses:
Tissue sections stained with hematoxylin and eosin would be helpful to show the changes in tissue architecture more clearly.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This study identifies a novel energy-sensing circuit in Drosophila and mice that directly regulates sweet taste perception. In flies, hugin+ neurons function as a glucose sensor, activated through Glut1 transport and ATP-sensitive potassium channels. Once activated, hugin neurons release hugin peptide, which stimulates downstream Allatostatin A (AstA)+ neurons via PK2-R1 receptors. AstA+ neurons then inhibit sweet-sensing Gr5a+ gustatory neurons through AstA peptide and its receptor AstA-R1, reducing sweet sensitivity after feeding. Disrupting this pathway enhances sweet taste and increases food intake, while activating the pathway suppresses feeding.
The mammalian homolog of neuromedin U (NMU) was shown to play an analogous role in mice. NMU knockout mice displayed heightened sweet preference, while NMU administration suppressed it. In addition, VMH NMU+ neurons directly sense glucose and project to rNST Calb2+ neurons, dampening sweet taste responses. The authors suggested a conserved hugin/NMU-AstA pathway that couples energy state to taste perception.
Strengths:
Interesting findings that extend from insects to mammals. Very comprehensive.
Weaknesses:
Coupling energy status to taste sensitivity is not a new story. Many pathways appear to be involved, and therefore, it raises a question as to how this hugin-AstA pathway is unique.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors set out to extend their previous mapping of Drosophila head mechanosensory neurons (Eichler et al., 2024) by reconstructing their full second-order connectome. Their aim is to reveal how bristle mechanosensory neurons (BMNs) interface with excitatory and inhibitory partners to generate location-specific grooming movements, and to identify the circuit motifs and developmental lineages that support this transformation.
Strengths:
The strengths of this work are clear. The authors present a comprehensive synaptic-resolution connectome for BMNs, identifying nearly all of their pre- and postsynaptic partners. This dataset reveals important circuit motifs:
(1) BMNs provide feedforward excitation to descending neurons, feedforward inhibition to interneurons, and are themselves strongly regulated by GABAergic presynaptic inhibition.
(2) These motifs together support the idea that BMN activity is locally gated and hierarchically suppressed, fitting well with known behavioural sequences of grooming.
(3) The study also shows that connectivity preserves somatotopy, such that BMNs from neighbouring bristle populations converge onto shared partners, while distant BMNs remain segregated.
(4) A developmental analysis reveals both primary and secondary partners, suggesting a layered scaffold plus adult-specific elaborations.
(5) Finally, the identification of hemilineage 23b (LB23) as a core postsynaptic pathway - incorporating previously described antennal grooming neurons (aBN2) - provides a striking link between developmental lineage, anatomical connectivity, and behavioral output.
(6) Together, the dataset represents a valuable resource for the neuroscience community and a foundation for future functional studies.
Weaknesses:
There are also some weaknesses that mostly only limit clarity.
(1) The writing is dense, with results often presented in a cryptic fashion and the functional implications deferred to the discussion. As a result, the significance of circuit motifs such as BMN→motor or reciprocal inhibitory loops is sometimes buried, rather than highlighted when first described.
(2) Some assumptions require more explanation for non-specialist readers - for example, how bristle identity is inferred in EM in the absence of cuticular structures, or what is meant by "ascending" and "descending" in a dataset that does not include the ventral nerve cord. While some of this comes from the earlier paper, it would help readers of this one to explain this.
(3) Visualization choices also sometimes obscure key conclusions: network graphs can be visually appealing but do not clearly convey somatotopy or BMN-type differences; heatmaps or region-level matrices would make the parallel, block-like organization of the circuit more evident.
(4) The data might also speak to roles beyond grooming (e.g., mechanosensory modulation of posture or feeding), and a brief acknowledgement of this would broaden the impact.
(5) The restriction to one hemisphere should be explicitly acknowledged as a limitation when framing this as a 'comprehensive' connectome.
Overall, the authors achieve their main goal: they convincingly show that BMNs connect into parallel, somatotopically organized pathways, with LB23 providing a key lineage-based link from sensory input to grooming output. The dataset is carefully analyzed, and while the presentation could be streamlined, the connectome will be a valuable resource for researchers studying sensory processing, motor control, and the logic of circuit organization.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this study, the authors establish a human in vitro liver model by co-culturing induced hepatocyte-like cells (iHEPs) with induced macrophages (iMACs). Through flow cytometry-based sorting of cell populations at days 3 and 7 of co-culture, followed by bulk RNA sequencing, they demonstrate that bidirectional interactions between these two cell types drive functional maturation. Specifically, the presence of iMACs accelerates the hepatic maturation program of iHEPs, while contact-dependent cues from iHEPs enhance the acquisition of Kupffer cell identity in iMACs, indicating that direct cell-cell interactions are critical for establishing tissue-resident macrophage characteristics.
Functionally, the authors show that iMAC-derived Kupffer-like cells respond to pathological stimuli by producing interleukin-6 (IL-6), a hallmark cytokine of hepatic immune activation. When exposed to a panel of clinically relevant hepatotoxic drugs, the co-culture system exhibited concentration-dependent modulation of IL-6 secretion consistent with reported drug-induced liver injury (DILI) phenotypes. Notably, this response was absent when hepatocytes were co-cultured with monocyte-derived macrophages from peripheral blood, underscoring the liver-specific phenotype and functional relevance of the iMAC-derived Kupffer-like cells. Collectively, the study proposes this co-culture platform as a more physiologically relevant model for interrogating macrophage-hepatocyte crosstalk and assessing immune-mediated hepatotoxicity in vitro.
Strengths:
A major strength of this study lies in its systematic dissection of cell-cell interactions within the co-culture system. By isolating each cell type following co-culture and performing comprehensive transcriptomic analyses, the authors provide direct evidence of bidirectional crosstalk between iMACs and iHEPs. The comparison with single-culture controls is particularly valuable, as it clearly demonstrates how co-culture enhances functional maturation and lineage-specific gene expression in both cell types. This approach allows for a more mechanistic understanding of how hepatocyte-macrophage interactions contribute to the acquisition of tissue-specific phenotypes.
Weaknesses:
(1) Overreliance on bulk RNA-seq data:
The primary evidence supporting cell maturation is derived from bulk RNA sequencing, which has inherent limitations in resolving heterogeneous cellular states and functional maturation. The conclusions regarding hepatocyte maturation are based largely on increased expression of a subset of CYP genes and decreased AFP levels - markers that, while suggestive, are insufficient on their own to substantiate functional maturation. Additional phenotypic or functional assays (e.g., metabolic activity, protein-level validation) would significantly strengthen these claims.
(2) Insufficient characterization of input cell populations:
The manuscript lacks adequate validation of the cellular identities prior to co-culture. Although the authors reference previously published protocols for generating iHEPs and iMACs, it remains unclear whether the cells used in this study faithfully retain expected lineage characteristics. For example, hepatocyte preparations should be characterized by flow cytometry for ALB and AFP expression, while iMACs should be assessed for canonical macrophage markers such as CD45, CD11b, and CD14 before co-culture. Without these baseline data, it is difficult to interpret the magnitude or significance of any co-culture-induced changes.
(3) Quantitative assessment of IL-6 production is insufficient:
The analysis of drug-induced IL-6 responses is based primarily on relative changes compared to control conditions. However, percentage changes alone are inadequate to capture the biological relevance of these responses. Absolute cytokine production levels - particularly in response to LPS stimulation - should be reported and directly compared to PBMC-derived macrophages to determine whether iMAC-derived Kupffer-like cells exhibit enhanced cytokine output. Moreover, the Methods section should clearly describe how ELISA results were normalized or corrected to account for potential differences in cell number, viability, or culture conditions.
(4) Unclear mechanistic interpretation of IL-6 modulation:
The observed changes in IL-6 production upon drug treatment cannot be interpreted solely as evidence of Kupffer cell-specific functionality. For instance, IL-6 suppression by NSAIDs such as diclofenac is well known to result from altered prostaglandin synthesis due to COX inhibition, while leflunomide's effects are linked to metabolite-induced modulation of immune cell proliferation and broader cytokine networks. These mechanisms are distinct from Kupffer cell identity and may not directly reflect liver-specific macrophage function. Consequently, changes in IL-6 secretion alone - particularly without additional mechanistic evidence or analysis of other cytokines - are insufficient to conclude that co-culture with hepatocytes drives the acquisition of bona fide Kupffer cell maturity.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This manuscript reports the discovery of new compounds that selectively inhibit SMARCA4/SMARCA2 ATPase activity and have pronounced effects on uveal melanoma cell proliferation. They induce apoptosis and suppress tumor growth, with no toxicity in vivo. The report provides biological significance by demonstrating that the drugs alter chromatin accessibility at lineage specific gene enhancer regions and decrease expression of lineage specific genes, including SOX10 and SOX10 target genes.
Strengths:
The study provides compelling evidence for the therapeutic use of these compounds and does a thorough job at elucidating the mechanisms by which the drugs work. The study will likely have a high impact on the chromatin remodeling and cancer fields. The datasets will be highly useful to these communities.
[Editors' note: The authors have addressed all of the outstanding issues.]
-
-
www.medrxiv.org www.medrxiv.org
-
Reviewer #3 (Public review):
The introduction does a very good job of discussing the issue around whether there is ongoing replication in people with HIV on antiretroviral therapy. Sporadic, non-sustained replication likely occurs in many PWH on ART related to adherence, drug-drug interactions and possibly penetration of antivirals into sanctuary areas of replication and as the authors point out proving it does not occur is likely not possible and proving it does occur is likely very dependent on the population studied and the design of the intervention. Whether the consequences of this replication in the absence of evolution toward resistance have clinical significance challenging question to address.
It is important to note that INSTI-based therapy may have a different impact on HIV replication events that results in differences in virus release for specific cell type (those responsible for "second phase" decay) by blocking integration in cells that have completed reverse transcription prior to ART initiation but have yet to be fully activated. In a PI or NNRTI-based regimen, those cells will release virus, whereas with an INSTI-based regimen, they will not.
Given the very small sample size, there is a substantial risk of imbalance between the groups in important baseline measures.
Comments on the revised version from the editor:
I appreciate that the authors thoroughly address the reviewer's concerns in the response letter. Most importantly, they acknowledge that "The absence of a pre-specified statistical endpoint or sample size calculation reflects the exploratory nature of the trial." This is vital because the transient impact on total HIV DNA in the intensified versus standard dose arm raises questions about any sustained or meaningful anti-reservoir effect and was also not hypothesized a priori. The authors explanation that HIV DNA may have rebounded due to clonal expansion is interesting but not assessed directly in the trial.
The greater decrease in intact HIV DNA between days 0 and 84 in the intensified arm are notable but are somewhat limited by small sample size, small effect size and lack of data between these two timepoints.
Unfortunately, the hypothesis generating nature of the conclusions which is outlined nicely in the author's response letter is only acknowledged in the discussion of the revised paper. The abstract and results are only marginally different than the original version and still read as definitive when the evidence is only hypothesis generating. For these reasons, the level of evidence remains incomplete as before.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The manuscript demonstrates that starvation induces persister formation in M. abscesses. They also utilized Tn-Seq for the identification of genes involved in persistence. They identified the role of catalase-peroxidase KatG in preventing death from translation inhibitors Tigecycline and Linezolid. They further demonstrated that a combination of these translation inhibitors leads to the generation of ROS in PBS-starved cells.
Strengths:
The authors used high-throughput genomics-based methods for identification of genes playing a role in persistence.
Weaknesses:
The findings could not be validated in clinical strains.
Comments on revisions: No more comments for the authors.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary
The regulation of NF-κB signaling is complex and central to the differentiation and homeostasis of αβT cells, essential to adaptive immunity. γδ T cells are a distinct population that responds to stress/injury-induced cues by producing inflammatory cytokines, representing an important bridge between innate and adaptive immunity. This study from Islam et al. demonstrates that the IKK complex, a central regulator of NF-κB signaling, plays distinct and essential roles in the differentiation and maintenance of γδ T cells. The authors use elegant murine genetic models to generate clear data that disentangle these requirements in vivo.
Although NF-κB activity was found to be dispensable for specification of γδ T cell progenitors and the generation of adaptive γδ T cells, it was required for both the ontogeny of type 1 γδ T cells and the survival of mature adaptive γδ T cells. Subunit-specific analyses revealed parallels with αβ T cells: RELA was necessary for type 1 γδ T cell development, while maintenance of adaptive γδ T cells relied upon redundancy between REL subunits, with cREL and p50 compensating in the absence of RELA but not vice versa. These findings reflect distinct biological requirements for ontogeny versus maintenance, likely driven by differences in receptor signaling, such as TCR and TNFRSF family members. Moreover, IKK also maintained γδ T cell survival through repression of RIPK1-mediated cell death, echoing its dual role in αβ T cells, where it both prevents TNF-induced apoptosis and provides NF-κB-dependent survival signals.
Strengths:
The multiple, unique murine genetic models employed for detailed analysis of in vivo γδ T cell differentiation and homeostasis are a major strength of this paper. NF-κB signaling processes are devilishly complex. The conditional mutants generated for this study disentangle the requirements for the various IKK-regulated pathways in γδ T cell differentiation, cell survival, and homeostasis. Data are clearly presented and suitably interpreted, with a helpful synthesis provided in the Discussion. These data will provide a definitive account of the requirements for NF-κB signaling in γδ T cells and provide new genetic models for the community to further study the upstream signals.
Weaknesses:
The paper would benefit greatly from a graphical abstract that could summarize the key findings, making the key findings accessible to the general immunology or biochemistry reader. Ideally, this graphic would distinguish the requirements for NF-κB signals sustaining thymic γδ T cell differentiation from peripheral maintenance, taking into account the various subsets and signaling pathways required. In addition, the authors should consider adding further literature comparing the requirements for NF-κB /necroptosis pathways in regulating other non-conventional T cell populations, such as iNKT, MAIT, or FOXP3+ Treg cells. These data might help position the requirements described here for γδ T cells compared to other subsets, with respect to homeostatic cues and transcriptional states.
Last and least, there are multiple grammatical errors throughout the manuscript, and it would benefit from further editing. Likewise, there are some minor errors in figures (e.g., Figure 3A, add percentage for plot from IKKDT.RIPK1D138N mouse; Figure 7, "Adative").
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The manuscript by Kim et al. describes a MuSC subpopulation that loses VCam expression in geriatric muscle and shows reduced ability to contribute to muscle regeneration. They propose that this population underlies the reported decline of MuSCs in aged mice, suggesting that these cells remain present in geriatric muscle but are overlooked due to low or absent VCam expression. The identification of a subpopulation that changes with aging would be compelling and of interest to the field.
Strengths:
The authors employ a wide range of assays, from in vitro to in vivo systems, to characterize Vcam-low/negative cells from geriatric muscle. The loss of Vcam appears strong in geriatric mice. They further identify CD63 and CD200 as potential surface markers that remain stable with age, thereby enabling the isolation of MuSCs across different age groups.
Weaknesses:
Some issues remain before establishing whether this population represents a true functional subset or explains the reported decline in MuSC numbers in aged mice. A stronger fate assessment of Vcam-low/negative cells is needed to assess their propensity for cell death in vitro and in vivo (e.g., engraftment efficiency), and if this plays a role in their conclusions. Comparisons include young, middle-aged, and geriatric mice, but not aged (~24 months) mice, which are needed for direct assessment of previous reports of age-related MuSC decline. The suggestion that the Vcam-low/negative population reflects senescence appears premature, with few consistent markers for this fate, as well as the cells not exhibiting irreversible cell-cycle exit. Finally, validation of CD63 and CD200 as reliable age-independent MuSC markers requires further testing, specifically using the Pax7-YFP tracing model and co-labeling in geriatric mice.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public Review):
The manuscript by Carayon and Strutt addresses the role of cell-scale signaling during the establishment of planar cell polarity (PCP) in the Drosophila pupal wing. The authors induce locally the expression of a tagged core PCP protein, Frizzled, and observe and analyze the de novo establishment of planar cell polarity. Using this system, the authors show that PCP can be established within several hours, that PCP is robust towards variation in core PCP protein levels, that PCP proteins do not orient microtubules, and that PCP is robust towards 'extrinsic' re-polarization. The authors conclude that the polarization at the cell-scale is strongly intrinsic and only weakly affected by the polarity of neighboring cells.
Major comments (first round of review):
The data are clearly presented and the manuscript is well written. The conclusions are well supported by the data.
(1) The authors use a system to de novo establish PCP, which has the advantage of excluding global cues orienting PCP and thus to focus on the cell-intrinsic mechanisms. At the same time, the system has the limitation that it is unclear to what extent de novo PCP establishment reflects 'normal' cell scale PCP establishment, in particular because the Gal4/UAS expression system that is used to induce Fz expression will likely result in much higher Fz levels compared with the endogenous levels. The authors should briefly discuss this limitation.
(2) Fig. 3. The authors use heterozygous mutant backgrounds to test the robustness of de novo PCP establishment towards (partial) depletion in core PCP proteins. The authors conclude that de novo polarization is 'extremely robust to variation in protein level'. Since the authors (presumably) lowered protein levels by 50%, this conclusion appears to be somewhat overstated. The authors should tune down their conclusion.
Significance:
The manuscript contributes to our understanding of how planar cell polarity is established. It extends previous work by the authors (Strutt and Strutt, 2002,2007) that already showed that induction of core PCP pathway activity by itself is sufficient to induce de novo PCP. This manuscript further explores the underlying mechanisms. The authors test whether de novo PCP establishment depends on an 'inhibitory signal', as previously postulated (Meinhardt, 2007), but do not find evidence. They also test whether core PCP proteins help to orient microtubules (which could enhance cell intrinsic polarization of core PCP proteins), but, again, do not find evidence, corroborating previous work (Harumoto et al, 2010). The most significant finding of this manuscript, perhaps, is the observation that local de novo PCP establishment does not propagate far through the tissue. A limitation of the study is that the mechanisms establishing intrinsic cell scale polarity remain unknown. The work will likely be of interest to specialists in the field of PCP.
Summary of comments from the Reviewing Editor on the revised version:
In the introduction, when you refer to Figure 1, the definition of Molecular, cellular, tissue scale is indeed not too clear to outside readers. For example, when you first refer to 'cell scale' you define it 'non-local', but probably it is not clear to many readers 'non-local' means 'the mechanism that cannot be explained by 'molecular scale'. (because 'molecular scale = local' is only inferred).
The 'conclusion paragraph' at the end of the Introduction does not have conclusion (only explained 'which question was tested by which method').
Minor comments that can easily be addressed by textual edits:
– they do not explain why gene dosage affects constitutive but not de novo polarization. It seems to me that one would expect de novo to be at least as sensitive if not more.
– Unconventional nomenclature for tissue axes - mediolateral, horizontal - are frequently used. These are sometimes difficult to parse. Please stick with universally accepted anterior, posterior, proximal and distal.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Kumar et al. examine the H3K115 epigenetic mark located on the lateral surface of the histone core domain and present evidence that it may serve as a marker enriched at transcription start sites (TSSs) of active CpG island promoters and at polycomb-repressed promoters. They also note enrichment of the H3K115ac mark is found on fragile nucleosomes within nucleosome-depleted regions, on active enhancers, and CTCF-bound sites. They propose that these observations suggest that H3K115ac contributes to nucleosome destabilization and so may serve as a marker of functionally important regulatory elements in mammalian genomes.
Strengths:
The authors present novel observations suggesting that acetylation of a histone residue in a core (versus on a histone tail) domain may serve a functional role in promoting transcription, in CPG islands and polycomb-repressed promoters. They present a solid amount of confirmatory in silico data using appropriate methodology that supports the idea that the H3K115ac mark may function to destabilize nucleosomes and contribute to regulating ESC differentiation.
Weaknesses:
Additional experiments to confirm antibody specificity are needed. The authors use synthetic peptides for other markers (e.g., H3K122) to support the claim that the antibody is specific, but ChIP-ChIP assays are performed under cross-linked, non-denatured conditions, which preserve structure and epitope accessibility differently than synthetic peptides used for dot blots. Does the antibody give a single band in western blots of histones, and can the H3K115ac peptide block western and immunofluorescence signals of the antibody? Given that the antibody is a rabbit polyclonal, specificity is not a trivial consideration.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
This study investigates the connection between glycolysis and the biosynthesis of sulfur-containing amino acids in controlling fungal morphogenesis, using Saccharomyces cerevisiae and C. albicans as model organisms. The authors identify a conserved metabolic axis that integrates glycolysis with cysteine/methionine biosynthetic pathways to influence morphological transitions. This work broadens the current understanding of fungal morphogenesis, which has largely focused on gene regulatory networks and cAMP-dependent signaling pathways, by emphasizing the contribution of metabolic control mechanisms. However, despite the novel conceptual framework, the study provides limited mechanistic characterization of how the sulfur metabolism and glycolysis blockade directly drive morphological outcomes. In particular, the rationale for selecting specific gene deletions, such as Met32 (and not Met4), or the Met30 deletion used to probe this pathway, is not clearly explained, making it difficult to assess whether these targets comprehensively represent the metabolic nodes proposed to be critical. Further supportive data and experimental validation would strengthen the claims on connections between glycolysis, sulfur amino acid metabolism, and virulence.
Strengths:
(1) The delineation of how glycolytic flux regulates fungal morphogenesis through a cAMP-independent mechanism is a significant advancement. The coupling of glycolysis with the de novo biosynthesis of sulfur-containing amino acids, a requirement for morphogenesis, introduces a novel and unexpected layer of regulation.
(2) Demonstrating this mechanism in both S. cerevisiae and C. albicans strengthens the argument for its evolutionary conservation and biological importance.
(3) The ability to rescue the morphogenesis defect through exogenous supplementation of sulfur-containing amino acids provides functional validation.
(4) The findings from the murine Pfk1-deficient model underscore the clinical significance of metabolic pathways in fungal infections.
Weaknesses:
(1) While the link between glycolysis and sulfur amino acid biosynthesis is established via transcriptomic and proteomic analysis, the specific regulation connecting these pathways via Met30 remains to be elucidated. For example, what are the expression and protein levels of Met30 in the initial analysis from Figure 2? How specific is this effect on Met30 in anaerobic versus aerobic glycolysis, especially when the pentose phosphate pathway is involved in the growth of the cells when glycolysis is perturbed?
(2) Including detailed metabolite profiling could have strengthened the metabolic connection and provided additional insights into intermediate flux changes, i.e., measuring levels of metabolites to check if cysteine or methionine levels are influenced intracellularly. Also, it is expected to see how Met30 deletion could affect cell growth. Data on Met30 deletion and its effect on growth are not included, especially given that a viable heterozygous Met30 strain has been established. Measuring the cysteine or methionine levels using metabolomic analysis would further strengthen the claims in every section.
(3) In comparison with the previous bioRxiv (doi: https://doi.org/10.1101/2025.05.14.654021) of this article in May 2025 to the recent bioRxiv of this article (doi: https://doi.org/10.1101/2025.05.14.654021), there have been some changes, and Met30 deletion has been recently included, and the chemical perturbation of glycolysis has been added as new data. Although the changes incorporated in the recent version of the article improved the illustration of the hypothesis in Figure 6, which connects glycolysis to Sulfur metabolism, the gene expression and protein levels of all genes involved in the illustrated hypothesis are not consistently shown. For example, in some cases, the Met4 expression is not shown (Figure 4), and the Met30 expression is not shown during profiling (gene expression or protein levels) throughout the manuscript. Lack of consistency in profiling the same set of key genes makes understanding more complicated.
(4) The demonstrated link between glycolysis and sulfur amino acid biosynthesis, along with its implications for virulence in C. albicans, is important for understanding fungal adaptation, as mentioned in the article; however, the Met4 activation was not fully characterized, nor were the data presented when virulence was assessed in Figure 4. Why is Met4 not included in Figure 4D and I? Especially, according to Figure 6, Met4 activation is crucial and guides the differences between glycolysis-active and inactive conditions.
(5) Similarly, the rationale behind selecting Met32 for characterizing sulfur metabolism is unclear. Deletion of Met32 resulted in a significant reduction in pseudohyphal differentiation; why is this attributed only to Met32? What happens if Met4 is deleted? It is not justified why Met32, rather than Met4, was chosen. Figure 6 clearly hypothesizes that Met4 activation is the key to the mechanism.
(6) The comparative RT-qPCR in Figure 5 did not account for sulfur metabolism genes, whereas it was focused only on virulence and hyphal differentiation. Is there data to support the levels of sulfur metabolism genes?
(7) To validate the proposed interlink between sulfur metabolism and virulence, it is recommended that the gene sets (illustrated in Figure 6) be consistently included across all comparative data included throughout the comparisons. Excluding sulfur metabolism genes in Figure 5 prevents the experiment from demonstrating the coordinated role of glycolysis perturbation → sulfur metabolism → virulence. The same is true for other comparisons, where the lack of data on Met30, Met4, etc., makes it hard to connect the hypothesis. It is also recommended to check the gene expression of other genes related to the cAMP pathway and report them to confirm the cAMP-independent mechanism. For example, gap2 deletion was used to confirm the effects of cAMP supplementation, but the expression of this gene was not assessed in the RNA-seq analysis in Figure 2. It would be beneficial to show the expression of cAMP-related genes to completely confirm that they do not play a role in the claims in Figure 2.
(8) Although the NAC supplementation study is included in the new version of the article compared to the previous version in BioRxiv (May 2025), the link to sulfur metabolism is not well characterized in Figure 5 and their related datasets. The main focus of the manuscript is to delineate the role of sulfur metabolism; hence, it is anticipated that Figure 5 will include sulfur-related metabolic genes and their links to pfk1 deletion, using RT-PCR measurements as shown for the virulence genes.
(9) The manuscript would benefit from more information added to the introduction section and literature supports for some of the findings reported earlier, including the role of (i) cAMP-PKA and MAPK pathways, (ii) what is known in the literature that reports about the treatment with 2DG (role of Snf1, HXT1, and HXT3), as well as how gpa2 is involved. Some sentences in the manuscripts are repetitive; it would be beneficial to add more relevant sections to the introduction and discussion to clarify the rationale for gene choices.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Nucleotide modifications are important regulators of biological function, however, until recently, their study has been limited by the availability of appropriate analytical methods. Oxford Nanopore direct RNA sequencing preserves nucleotide modifications, permitting their study, however many different nucleotide modifications lack an available base-caller to accurately identify them. Furthermore, existing tools are computationally intensive, and their results can be difficult to interpret.
Cheng et al. present SegPore, a method designed to improve the segmentation of direct RNA sequencing data and boost the accuracy of modified base detection.
Strengths:
This method is well described and has been benchmarked against a range of publicly available base callers that have been designed to detect modified nucleotides.
Comment from the Reviewing Editor:
The authors have provided responses to the weaknesses highlighted previously and the reviewers were not asked to comment. The authors have now requested a Version of Record.
-
-
-
Reviewer #3 (Public review):
Summary:
Pinho et al., investigated the role of the dorsal VS ventral hippocampus and gender differences in mediated learning. While previous studies already established the engagement of the hippocampus in sensory preconditioning, the authors here took advantages of freely-moving fiber photometry recording and chemogenetics to observe and manipulate sub-regions of the hippocampus (drosal VS ventral) in a cell-specific manner. Importantly, the authors validated the sensory preconditioning procedure in male mice. The authors found no evidence of sensory preconditioning in female mice, but rather a generalization effect, stressing the importance of gender differences in fear learning. After validation of a sensory preconditioning procedure in male mice using light and tone neutral stimuli and a mild foot shock as the unconditioned stimulus, the authors used fiber photometry to record from all neurons VS parvalbumin_positive_only neurons in the dorsal hippocampus or ventral hippocampus of male mice during both preconditioning and conditioning phases. They found an increased activity of all neurons, PV+_only neurons, and CAMKII+ neurons in both sub-regions of the hippocampus during both preconditioning and conditioning phases. Finally, the authors found that chemogenetic inhibition of CaMKII+ neurons (but not PV+_only neurons) in the dorsal (but not ventral) hippocampus specifically prevented the formation of an association between the two neutral stimuli (i.e., light and tone cues). This manipulation had no effect on the direct association between the light cue and the mild foot shock. This set of data (1) validates sensory preconditioning in male mice, and stresses the importance of taking gender effect into account; (2) validates the recruitment of dorsal and ventral hippocampi during preconditioning and conditioning phases; (3) and further establishes the specific role of CaMKII+ neurons in the dorsal hippocampus, but not ventral hippocampus, in the formation of an association between two neutral stimuli, but not between a neutral-stimulus and a mild foot shock.
Strengths:
The authors developed a sensory preconditioning procedure in male mice to investigate mediated learning using light and tone cues as neutral stimuli, and a mild foot shock as the unconditioned stimulus. They provide evidence of a gender effect in the formation of light-cue association. The authors took advantage of fiber-photometry and chemogenetics to target sub-regions of the hippocampus, in a cell-specific manner and investigate their role during different phases of a sensory conditioning procedure, and developed a DeepLabCut-based strategy to assess freezing fear responses.
Weaknesses:
The authors went further than previous studies by investigating the role of sub-regions the hippocampus in mediated learning, however, there are a few weaknesses that should be addressed in future studies:
(1) This study found a generalization effect in female mice only. While the authors attempted to neutralize this effect, the mechanism underlying this gender effect and whether female mice can display evidence for mediated learning has yet to be determined.
(2) One of the main effects from which derives the conclusion of this study (i.e., deficit of mediated learning in male mice when CAMKII+ neurons are inhibited in the dorsal HPC during the preconditioning phase) lies in the absence of a significant difference of the freezing response before and during the tone cue presentation when CAMKII+ are chemogenetically inhibited during the Probe Test Tone phase (cf. Fig. 4 Panel B, DPCd group). The fear response before the tone cue presentation in this group (DPCd) seems higher than in Controls_d and DPTd groups and could have masked a mediated learning effect.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
This study by the Boddy and Otomo laboratories further characterizes the roles of SMC5/6 loader proteins and related factors in SMC5/6-mediated repression of extrachromosomal circular DNA. The work shows that mutations engineered at an AlphaFold-predicted protein-protein interface formed between the loader SLF2/SIMC1 and SMC6 (similar to the interface in the yeast counterparts observed by cryo-EM) prevent co-IP of the respective proteins. The mutations in SLF2 also hinder plasmid DNA silencing when expressed in SLF2-/- cell lines, suggesting that this interface is needed for silencing. SIMC1 is dispensable for recruitment of SMC5/6 to sites of DNA damage, while SLF1 is required, thus separating the functions of the two loader complexes. Preventing SUMOylation (with a chemical inhibitor) increases transcription from plasmids but does not in SLF2-deleted cell lines, indicating the SMC5/6 silences plasmids in a SUMOylation dependent manner. Expression of LT is sufficient for increased expression, and again, not additive or synergistic with SIMC1 or SLF2 deletion, indicating that LT prevents silencing by directly inhibiting 5/6. In contrast, PML bodies appear dispensable for plasmid silencing.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In their manuscript, Armand and colleagues investigate the potential of continuing CDK4/6 inhibitors or combining them with CDK2 inhibitors in the treatment of breast cancer that has developed resistance to initial therapy. Utilizing cellular and animal models, the research examines whether maintaining CDK4/6 inhibition or adding CDK2 inhibitors can effectively control tumor growth after resistance has set in. The key findings from the study indicate that the sustained use of CDK4/6 inhibitors can slow down the proliferation of cancer cells that have become resistant, and the combination of CDK2 inhibitors with CDK4/6 inhibitors can further enhance the suppression of tumor growth. Additionally, the study identifies that high levels of Cyclin E play a significant role in resistance to the combined therapy. These results suggest that continuing CDK4/6 inhibitors along with the strategic use of CDK2 inhibitors could be an effective strategy to overcome treatment resistance in hormone receptor-positive breast cancer. However, several issues need to be addressed before considering its publication.
Strengths:
(1) Continuous CDK4/6 Inhibitor Treatment Significantly Suppresses the Growth of Drug-Resistant HR+ Breast Cancer: The study demonstrates that the continued use of CDK4/6 inhibitors, even after disease progression, can significantly inhibit the growth of drug-resistant breast cancer.
(2) Potential of Combined Use of CDK2 Inhibitors with CDK4/6 Inhibitors: The research highlights the potential of combining CDK2 inhibitors with CDK4/6 inhibitors to effectively suppress CDK2 activity and overcome drug resistance.
(3) Discovery of Cyclin E Overexpression as a Key Driver: The study identifies overexpression of cyclin E as a key driver of resistance to the combination of CDK4/6 and CDK2 inhibitors, providing insights for future cancer treatments.
(4) Consistency of In Vitro and In Vivo Experimental Results: The study obtained supportive results from both in vitro cell experiments and in vivo tumor models, enhancing the reliability of the research.
(5) Validation with Multiple Cell Lines: The research utilized multiple HR+/HER2- breast cancer cell lines (such as MCF-7, T47D, CAMA-1) and triple-negative breast cancer cell lines (such as MDA-MB-231), validating the broad applicability of the results.
Comments on revisions:
The authors made a significant effort to improve the manuscript. My comments were sufficiently addressed.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors of this manuscript have addressed a key concept in T cell development: how early thymus gd T cell subsets are specified and the elements that govern gd T17 versus other gd T cell subsets or ab T cell subsets are specified. They show that the transcriptional regulator HEB/Tcf12 plays a critical role in specifying the gd T17 lineage and, intriguingly, that it upregulates the inhibitor Id3, which is later required for further gd T17 maturation.
Strengths:
The conclusions drawn by the authors are amply supported by a detailed analysis of various stages of T cell maturation in WT and KO mouse strains at the single cell level, both phenotypically, by flow cytometry for various diagnostic surface markers, and transcriptionally, by single cell sequencing. Their conclusions are balanced and well supported by the data and citations of previous literature.
Weaknesses:
I actually found this work to be quite comprehensive. I have a few suggestions for additional analyses the authors could explore that are unrelated to the predominant conclusions of the manuscript, but I failed to find major flaws in the current work.
I note that HEB is expressed in many hematopoietic lineages from the earliest progenitors and throughout T cell development. It is also noteworthy that abortive gamma and delta TCR rearrangements have been observed in early NK cells and ILCs, suggesting that, particularly in early thymic development, specification of these lineages may have lower fidelity. It might prove interesting to see whether their single-cell sequencing or flow data reveal changes in the frequency of these other T-cell-related lineages. Is it possible that HEB is playing a role not only in the fidelity of gdT17 cell specification, but also perhaps in the separation of T cells from NK cells and ILCs or the frequency of DN1, DN2, and DN3 cells? Perhaps their single-cell sequencing data or flow analyses could examine the frequency of these cells? That minor caveat aside, I find this to be an extremely exciting body of work.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The manuscript by Chaya and Syed focuses on understanding the link between cell cycle and temporal patterning in central brain type II neural stem cells (NSCs). To investigate this, the authors perturb the progression of the cell cycle by delaying the entry into M phase and preventing cytokinesis. Their results convincingly show that temporal factor expression requires progression of the cell cycle in both Type 1 and Type 2 NSCs in the Drosophila central brain. Overall, this study establishes an important link between the two timing mechanisms of neurogenesis.
Strengths:
The authors provide solid experimental evidence for the coupling of cell cycle and temporal factor progression in Type 2 NSCs. The quantified phenotype shows an all-or-none effect of cell cycle block on the emergence of subsequent temporal factors in the NSCs, strongly suggesting that both nuclear division and cytokinesis are required for temporal progression. The authors also extend this phenotype to Type 1 NSCs in the central brain, providing a generalizable characterization of the relationship between cell cycle and temporal patterning.
Weaknesses:
One major weakness of the study is that the authors do not explore the mechanistic relationship between the cell cycle and temporal factor expression. Although their results are quite convincing, they do not provide an explanation as to why Cdk1 depletion affects Syp and EcR expression but not the onset of svp. This result suggests that at least a part of the temporal cascade in NSCs is cell-cycle independent, which isn't addressed or sufficiently discussed.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Hensley et al. present an important study into the force-detachment behaviour of kinesin-1, the most well-characterised motor protein. One of the key techniques used to characterise kinesins is in vitro optical trapping of purified proteins, which has provided remarkable insights into the biochemical and mechanical mechanisms of motor proteins under single- and multi-motor conditions. This study presents an adapted (from Urbanska et al.) methodological approach of DNA-tethering kinesin-1 to a bead, both under single- and multi-motor conditions, which is then trapped to characterise the run length, processivity, and stall behaviour under unloaded and loaded (both assisting and hindering) conditions. The new approach reduces the vertical or z-force and thus provides insights into the role of horizontal or x-forces acting on the motor. Based on their method of imposing dominant horizontal forces on the motor and their data, they conclude that kinesin-1 exhibits a higher asymmetry in its force-detachment kinetics, is less slippery, and exhibits slip-bond behaviour, particularly under hindering loads. Under assisting loads, similar slip-bond kinetics ensue, but detachment from the microtubule is far more sensitive. To demonstrate the implications of their method and data, they conduct a multi-motor assay and show that multiple kinesin-1 motors can generate significantly higher forces, almost proportional to motor number. Overall, this is important work, and the data are compelling.
Strengths:
The method of DNA-tethered motor trapping is effective in reducing vertical forces and can be easily optimised for other motors and protein characterisation. The major strength of the paper is characterising kinesin-1 under low z-forces, which is likely to reflect the physiological scenario. They report that kinesin-1 is more robust and less prone to premature detachment. The motors exhibit higher stall rates and times. Under hindering and assisting loads, kinesin-1 detachment is more asymmetric and sensitive, and with low z-force shows that slip-behaviour kinetics prevail. Another achievement of this paper is the demonstration of the multi-motor kinesin-1 assay using their low-z force method, showing that multiple kinesin-1 motors are capable of generating higher forces (up to 15 pN, and nearly proportional to motor number), thus opening an avenue to study multiple motor coordination.
Weaknesses:
The method of DNA-tethered motor trapping to enable low z-force is not entirely novel, but adapted from Urbanska (2021) for use in conventional optical trapping laboratories without reliance on microfluidics. However, I appreciate that they have fully established it here to share with the community. The authors could strengthen their methods section by being transparent about protein weight, protein labelling, and DNA ladders shown in the supplementary information. What organism is the protein from? Presumably human, but this should be specified in the methods. While the figures show beautiful data and exemplary traces, the total number of molecules analysed or events is not consistently reported. Overall, certain methodological details should be made sufficient for reproducibility.
The major limitation the study presents is overarching generalisability, starting with the title. I recommend that the title be specific to kinesin-1. The study uses two constructs: a truncated K560 for conventional high-force assays, and full-length Kif5b for the low z-force method. However, for the multi-motor assay, the authors use K560 with the rationale of preventing autoinhibition due to binding with DNA, but that would also have limited characterisation in the single-molecule assay. Overall, the data generated are clear, high-quality, and exciting in the low z-force conditions. But why have they not compared or validated their findings with the truncated construct K560? This is especially important in the force-feedback experiments and in comparison with Andreasson et al. and Carter et al., who use Drosophila kinesin-1. Could kinesin-1 across organisms exhibit different force-detachment kinetics? It is quite possible. Similarly, the authors test backward slipping of Kif5b and K560 and measure dwell times in multi-motor assays. Why not detail the backward slippage kinetics of Kif5b and any step-size impact under low z-forces? For instance, with the traces they already have, the authors could determine slip times, distances, and frequency in horizontal force experiments. Overall, the manuscript could be strengthened by analysing both constructs more fully.
Appraisal and impact:
This study contributes to important and debated evidence on kinesin-1 force-detachment kinetics. The authors conclude that kinesin-1 exhibits a slip-bond interaction with the microtubule under increasing forces, while other recent studies (Noell et al. and Kuo et al.), which also use low z-force setups, conclude catch-bond behaviour under hindering loads. I find the results not fully aligned with their interpretation. The first comparison of low z-forces in their setup with Noell et al. (2024), based on stall times, does not hold, because it is an apples-to-oranges comparison. Their data show a stall time constant of 2.52 s, which is comparable to the 3 s reported by Noell et al., but the comparison is made with a weighted average of 1.49 s. The authors do report that detachment rates are lower in low z-force conditions under unloaded scenarios. So, to completely rule out catch-bond-like behaviour is unfair. That said, their data quality is good and does show that higher hindering forces lead to higher detachment rates. However, on closer inspection, the range of 0-5 pN shows either a decrease or no change in detachment rate, which suggests that under a hindering force threshold, catch-bond-like or ideal-bond-like behaviour is possible, followed by slip-bond behaviour, which is amazing resolution. Under assisting loads, the slip-bond character is consistent, as expected. Overall, the study contributes to an important discussion in the biophysical community and is needed, but requires cautious framing, particularly without evidence of motor trapping in a high microtubule-affinity state rather than genuine bond strengthening.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
This study makes excellent use of a uniquely large dataset of reaching movements collected over several decades to evaluate the origins of systematic motor biases. The analyses convincingly demonstrate that these biases are not explained by errors in sensed hand position or by biomechanical constraints, but instead arise from a misalignment between eye-centric and body-centric representations of position. By testing multiple computational models across diverse contexts-including different effectors, visible versus occluded start positions-the authors provide strong evidence for their transformation model. My earlier concerns have been addressed, and I find the work to be a significant and timely contribution that will be of broad interest to researchers studying visuomotor control, perception, and sensorimotor integration.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Liff et al. have made considerable effort to improve their manuscript. In their revised manuscript, the authors have substantiated their claims of intergenerationally inherited changes in the olfactory system in response to odor-dependent fear conditioning. Several new experiments and analyses now strengthen this study.
I still find that the statement that the study provides "insight into the heritability of acquired phenotypes" is somewhat misleading. In their response to this initially raised point the authors correctly point out that their "results provide basic knowledge that will accelerate our ability to uncover the mechanisms driving heritable changes." That said, current "insights" are not mechanistic in nature.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
In the study the authors performed longitudinal 1P calcium imaging of mouse mPFC across 8 weeks during learning of an olfactory-guided task, including habituation, training, and sleep periods. The authors' goal was to determine how the mPFC representation of the task changed and what aspects of activity emerged between the learning and the learned conditions of the task. The task had 3 arms. Odor was sampled at the end of the middle arm (named the "Sample" period). The animal then needed to run to one of the two other arms (R or L) based on the odor. The whole period until they reached the end of one of the choice arms was the "Outward" period. The time at the reward end was the "Reward" period. They noted several changes from the learning condition to the learned condition:
(1) They classified cells in a few ways. First each cell was classified as SI (spatially informative) if it had significantly more spatial information than shuffled activity, and ~50% of cells ended up being SI cells. Then among the SI cells they classified a cell as a TC (task cell) if it had statistically similar activity maps for R versus L arms, and a GC (goal arm cell) otherwise. Note that there are 4 kinds of these cells: outer arm TCs and GCs and middle arm TCs and GCs (with middle arm GCs essentially being like "splitter cells" since they are not similarly active in the middle arm for R versus L trials). There was an increase in TCs from the learning to the learned condition sessions. They also note the sources of these TCs (some came from GCs, others from non-SI cells).
(2) They analyze activity sequences across cells. They extracted 500 ms duration bursts (defined as periods of activity > 0.5 standard deviations over what I assume is the mean, which is a permissive threshold encompassing a significant fraction of the activity in non-sleep, non-habituation periods). They first noted that the resulting "Burst rates were significantly larger during behavioral epochs than during sleep and during periods of habituation to the arena" and "Moreover, burst rates during correct trials were significantly lower than during error trials". For the sequence analysis they only considered bursts consisting of at least 5 active cells. A cell's activity within the burst was set to the center of mass calcium activity. Then they took all the sequences from all learned and learning sessions together and hierarchically clustered them based on the Spearman's rank correlation between the order of activity in each pair of sequences (among the cells active in both). The iterative hierarchical clustering process produces groups (clusters) of sequences such that there are multiple repeats of sequences within a cluster. Different sequences are expressed across all the longitudinally recorded sessions. They noted "large differences of sequence activation between learning and learned condition, both in the spatial patterns (example animal in Fig. 4D) and the distribution of the sequences (Fig. 4D,E). Rastermap plots (Fig. 4D) also reveal little similarity of sequence expression between task and habituation or sleep condition." They also note the difference in the sequences between learning and learned condition was larger than the different between correct and error trials within each condition. They conclude that during task learning new representations are established, as measured by the burst sequence content. They do additional analyses of the sequence clusters by assessing the spatial informativeness (SI) of each sequence cluster. Over learning they find an increase in clusters that are spatially informative (clusters that tend to occur in specific locations). Finally, they analyzed the SI clusters in a similar manner as SI cells and classified them as task phase selective sequences (TSs) and goal arm selective sequences (GSs) and did some further analysis. However, they themselves conclude that the frequency of TSs and GSs is limited because most sequence clusters were non-SI. In the discussion they say "In addition to GSs and TSs, we found that most of the recurring sequences are not related to behavior (not SI)".
(3) As an alternative to analyzing individual cells and sequences of individual cells, they then look for trajectory replay using Bayesian population decoding of location during bursts. They analyze TS bursts, GS bursts, and non-SI bursts. They say "we found correlations of decoded position with time bin (within a 500 ms burst) strongly exceeding chance level only during outward and reward phase, for both GSs and TSs (Fig 5H)." Fig5H shows distributions indicating statistically significant bias in the forward direction (using correlations of decoded location versus time bin across 10 bins of 50 ms each within each 500ms burst). They find that the Outward trajectories appear to reflect the actual trajectory during running itself, so are likely not replay. But the sequences at the Reward are replay as they do not reflect the current location. Furthermore, replay at the Reward is in the forward direction (unlike the reverse replay at Reward seen in the hippocampus) and this replay is only seen in the learned and not the learning condition. At the same time, they find that replay is not seen during odor Sampling, from which they conclude there is no evidence of replay used for planning. Instead they say the replay at the Reward could possibly be for evaluation during the Reward phase, though this would only be for the learned condition. They conclude "Together with our finding of strong changes in sequence expression after learning (Fig 4E) these findings suggest that a representation of task develops during learning".
This study provides valuable new information about the evolution of mPFC activity during the learning of a odor-based 2AFC T-maze-like task. They show convincing evidence of changes in single cell tuning, population sequences, and replay events. They also find novel forward replay at the Reward, and find that this is present only after the animal learned the task. In the discussion the authors note "the present study, to our knowledge, identified for the first time fast recurring neural sequence activity from 1-p calcium data, based on correlation analysis". Overall, they find a substantial amount of change among the features they analyzed and according to their methods, though they note a small amount of activity was preserved through learning.
One comment is that the threshold for extracting burst events (0.5 standard deviations, presumably above the mean) seems lower than what one usually sees as a threshold for population burst detection, and the authors show (in Supplementary Fig 1) that this means bursts cover ~20-40% of the data. However, it is potentially a strength of this work that their results are found by using this more permissive threshold.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors are reanalyzing previously published data to test the hypothesis that serotonin neurons encode state value. Here, the authors focus on analyzing the firing rate of serotonin neurons during the inter-trial interval, in which no cues or outcomes are delivered. The goal is to quantify and find neurons whose activity is explained by value encoding, and for those that have this property, determine what the timescale of reward integration is (e.g., a few trials, tens of trials, or the entire session) in individual neurons.
Strengths:
The major strengths are the use of a Bayesian modelling approach to extract value and thirst coding features from individual neurons, and comparison of the time course of adaptation of serotonin neurons with a behavioral output, licking in this case. I also appreciate the use of a separate dataset to establish prior distributions for baseline firing rate to be used in the modelling done here, which is an attempt to deal with the main weakness of this study:
Weaknesses:
The weakness of this study is the small number of neurons available for analysis, resulting in a small number of neurons that unequivocally are modulated by value.
The authors did achieve their aims, but the results show that it is hard to unequivocally separate value-coding neurons with long timescales from thirst-coding neurons, which is acknowledged by the authors.
While the experimental results do not allow for a strong conclusion regarding the distinction of value versus thirst coding in serotonin neurons, the methods employed and the rationale for using them are of great utility to the community and for considerations of behavioral task design and data analysis in future studies. This is a point that the authors could discuss/develop more.
Additional significance of the work:
The comparison between time courses for behavior (anticipatory licking) and serotonin activity (as well as the reference to dopamine activity's time course from a previous study) is of great significance for any researcher studying behavioral control. Mounting evidence suggests that multiple brain circuits contribute to any given action selection. Therefore, expecting a perfect alignment between the time course of neuromodulator activity and behavioral output might be unreasonable. For future studies, modelling behavioral output as a combination of policies determined by multiple brain circuits or neuromodulators might be a promising approach.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Fritz et al. investigate the changes in synaptic connectivity between two different life stages of the Drosophila larva, L1 and L3. They focus on 3 types of nociceptive mechanosensory neurons and their connecting 6 downstream interneurons. Connectomic analysis reveals that connectivity and dendritic density are stable across development; however, axonal density, axodendritic overlap, and the number of synapses increase. Finally, using a modeling approach, they demonstrate that this conservation of most features enables stable output across life stages.
Strengths:
The authors analyse two different connectomes from fly larvae in two different life stages. By now, there are only very few such samples available; thus, this is a novel approach and will be helpful to guide further comparative connectomic studies in the future.
Weaknesses:
The authors analyze only a minimal circuit with 9 different cell types on each hemisphere; thus, their findings might be specialised for this specific nociceptive sensory to interneuron peripheral circuit. Also, more animals might need to be analyzed in different life stages to generalize these findings.
-
-
-
Reviewer #3 (Public review):
Summary:
The study uses the food choice task, a well-established method in eating disorder research, particularly in anorexia nervosa. However, it introduces a novel analytical approach-the diffusion decision model-to deconstruct food choices and assess the influence of negative affect on how and when tastiness and healthiness are considered in decision-making among individuals with bulimia nervosa and healthy controls.
Strengths:
The introduction provides a comprehensive review of the literature, and the study design appears robust. It incorporates separate sessions for neutral and negative affect conditions and counterbalances tastiness and healthiness ratings. The statistical methods are rigorous, employing multiple testing corrections.
A key finding-that negative affect induction biases individuals with bulimia nervosa toward prioritizing tastiness over healthiness-offers an intriguing perspective on how negative affect may drive binge eating behaviors.
Weaknesses:
A notable limitation is the absence of a sample size calculation, which, combined with the relatively small sample, may have contributed to null findings. Additionally, while the affect induction method is validated, it is less effective than alternatives such as image or film-based stimuli (Dana et al., 2020), potentially influencing the results.
Tags
Annotators
URL
-
-
-
Reviewer #3 (Public review):
Summary
This paper investigates how disinformation affects reward learning processes in the context of a two-armed bandit task, where feedback is provided by agents with varying reliability (with lying probability explicitly instructed). They find that people learn more from credible sources, but also deviate systematically from optimal Bayesian learning: They learned from uninformative random feedback and updated too quickly from fully credible feedback (especially following low-credibility feedback). People also appeared to learn more from positive feedback and there is tentative evidence that this bias is exacerbated for less credible feedback.
Overall, this study highlights how misinformation could distort basic reward learning processes, without appeal to higher order social constructs like identity.
Strengths - The experimental design is simple and well-controlled; in particular, it isolates basic learning processes by abstracting away from social context - Modeling and statistics meet or exceed standards of rigor - Limitations are acknowledged where appropriate, especially those regarding external validity and challenges in dissociating positivity bias from perseveration - The comparison model, Bayes with biased credibility estimates, is strong; deviations are much more compelling than e.g. a purely optimal model - The conclusions are of substantial interest from both a theoretical and applied perspective
Weaknesses
The authors have done a great job addressing my concerns with the two previous submission. The one issue that they were not able to truly address is the challenge of dissociating positivity bias from perseveration; this challenge weakens evidence for the conclusion that less credible feedback yields a stronger positivity bias. However, the authors have clearly acknowledged this limitation and tempered their conclusions accordingly. Furthermore, the supplementary analyses on this point are suggestive (if not fully conclusive) and do a better job of at least trying to address the confound than most work on positivity/confirmation bias.
I include my previous review describing the challenge in more detail for reference. I encourage interested readers to see the author response as well. It has convinced me that this weakness is not a reflection of the work, but is instead a fundamental challenge for research on positivity bias.
Absolute or relative positivity bias?
The conclusion of greater positivity bias for lower credible feedback (Fig 5) hinges on the specific way in which positivity bias is defined. Specifically, we only see the effect when normalizing the difference in sensitivity to positive vs. negative feedback by the sum. I appreciate that the authors present both and add the caveat whenever they mention the conclusion. However, without an argument that the relative definition is more appropriate, the fact of the matter is that the evidence is equivocal.
There is also a good reason to think that the absolute definition is more appropriate. As expected, participants learn more from credible feedback. Thus, normalizing by average learning (as in the relative definition) amounts to dividing the absolute difference by increasingly large numbers for more credible feedback. If there is a fixed absolute positivity bias (or something that looks like it), the relative bias will necessarily be lower for more credible feedback. In fact, the authors own results demonstrate this phenomenon (see below). A reduction in relative bias thus provides weak evidence for the claim.
It is interesting that the discovery study shows evidence of a drop in absolute bias. However, for me, this just raises questions. Why is there a difference? Was one just a fluke? If so, which one?
Positivity bias or perseveration?
Positivity bias and perseveration will both predict a stronger relationship between positive (vs. negative) feedback and future choice. They can thus be confused for each other when inferred from choice data. This potentially calls into question all the results on positivity bias.
The authors clearly identify this concern in the text and go to considerable lengths to rule it out. However, the new results (in revision 1) show that a perseveration-only model can in fact account for the qualitative pattern in the human data (the CA parameters). This contradicts the current conclusion:
Critically, however, these analyses also confirmed that perseveration cannot account for our main finding of increased positivity bias, relative to the overall extent of CA, for low-credibility feedback.
Figure 24c shows that the credibility-CA model does in fact show stronger positivity bias for less credible feedback. The model distribution for credibility 1 is visibly lower than for credibilities 0.5 and 0.75.
The authors need to be clear that it is the magnitude of the effect that the perseveration-only model cannot account for. Furthermore, they should additionally clarify that this is true only for models fit to data; it is possible that the credibility-CA model could capture the full size of the effect with different parameters (which could fit best if the model was implemented slightly differently).
The authors could make the new analyses somewhat stronger by using parameters optimized to capture just the pattern in CA parameters (for example by MSE). This would show that the models are in principle incapable of capturing the effect. However, this would be a marginal improvement because the conclusion would still rest on a quantitative difference that depends on specific modeling assumptions.
New simulations clearly demonstrate the confound in relative bias
Figure 24 also speaks to the relative vs. absolute question. The model without positivity bias shows a slightly stronger absolute "positivity bias" for the most credible feedback, but a weaker relative bias. This is exactly in line with the logic laid out above. In standard bandit tasks, perseveration can be quite well-captured by a fixed absolute positivity bias, which is roughly what we see in the simulations (I'm not sure what to make of the slight increase; perhaps a useful lead for the authors). However, when we divide by average credit assignment, we now see a reduction. This clearly demonstrates that a reduction in relative bias can emerge without any true differences in positivity bias.
Given everything above, I think it is unlikely that the present data can provide even "solid" evidence for the claim that positivity bias is greater with less credible feedback. This confound could be quickly ruled out, however, by a study in which feedback is sometimes provided in the absence of a choice. This would empirically isolate positivity bias from choice-related effects, including perseveration.
Comments on revisions:
Great work on this. The new paper is very interesting as well. I'm delighted to see that the excessive amount of time I spent on this review has had a concrete impact.
Tags
Annotators
URL
-
-
arxiv.org arxiv.org
-
Reviewer #3 (Public review):
Summary:
Rong et al., compare EEG image responses from a large-scale dataset to state-of-the-art vision and language models, as well as their fusion. They find that the fusion of models provides the best predictivity, with early contribution from vision models and later predictivity from language models. The paper has several strengths: high temporal resolution data (though at the expense of spatial resolution), detailed comparison of alignment (and differences) between vision and language model embeddings, and comparison of "fusion" of different DNN models.
Despite the paper's strengths, it is not clear what is at stake with these findings or how they advance our knowledge beyond other recent studies showing vision versus language model predictions of visual cortex responses with fMRI.
Strengths:
The authors use a large-scale EEG dataset and a comprehensive modeling approach. The methods are sound and involve multiple model comparisons. In particular, the disentangling of vision and language model features is something that has been largely ignored in prior related studies.
Weaknesses:
(1) The authors state their main hypothesis (lines 48-51) that human neural responses to visual stimulation are better modelled by combining representations from a vision DNN and an LLM than by the representations from either of the two components alone, and that the vision DNN and LLM components would uniquely predict earlier and later stages of visual processing, respectively.
While they confirm this hypothesis in largely compelling ways, it is not clear whether these results tell us something about the brain beyond how to build the most predictive model.
In particular, why do language models offer advantages over vision models, and what does this tell us about human visual processing? In several places, the discussion of advantages for the language model felt somewhat trivial and did not seem to advance our understanding of human vision, e.g., "responses for visual stimulation encode detailed information about objects and their properties" (lines 266-270) and "LLM representations capture detailed visuo-semantic information about the stimulus images" (line 293).
(2) It is not clear what the high temporal resolution EEG data tell us that the whole-brain fMRI data do not. The latency results seem to be largely in line with fMRI findings, where the early visual cortex is better predicted by vision models, and the language model is better in later/more anterior regions. In addition, it would help to discuss whether the EEG signals are likely to be restricted to the visual cortex, or could the LLM predictivity explain downstream processing captured by whole-brain EEG signals?
Relatedly, it would help the authors to expand on the implications of the frequency analysis.
(3) While the authors test many combinations of vision and language models and show their "fusion" advantages are largely robust to these changes, it is still hard to ignore the vast differences between vision and language models, in terms of architecture and how they are trained. Two studies (Wang et al., 2023, and Conwell et al., 2024) have now shown that when properly controlling for architecture and dataset, there is little to no advantage of language alignment in predicting visual cortex responses. It would help for the authors to both discuss this aspect of the prior literature and to try to address the implications for their own findings (related to pt 1 about what, if anything, is "special" about language models).
(4) Model features - it would help to state the dimensionality of the input embeddings for each model and how much variance is explained and preserved after the PCA step? I wonder how sensitive the findings are to this choice of dimensionality reduction, and whether an approach that finds the optimal model layer (in a cross-validated way) would show less of a difference between vision/language models (I realize this is not feasible with models like GPT-3).
(5) To better understand the fusion advantage, it would help to look at the results, look for a pair of vision models and a pair of language models. Can a similar advantage be found by combining models from the same modality?
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #4 (Public review):
Summary:
In this manuscript, authors used a learning paradigm in C. elegans; when worms were fed in a saltless plate, its chemotaxis to salt is greatly reduced. To identify learning-related proteins, authors employed nervous system-specific transcriptome analysis to compare whole proteins in neurons between high-salt-fed animals and saltless-fed animals. Authors identified "learning-specific proteins" which are observed only after saltless feeding. They categorized these proteins by GO analyses, pathway analyses and expression site analyses, and further stepped forward to test mutants in selected genes identified by the proteome analysis. They find several mutants that are defective or hyper-proficient for learning, including acc-1/3 and lgc-46 acetylcholine receptors, F46H5.3 putative arginine kinase, and kin-2, a cAMP pathway gene. These mutants were not previously reported to have abnormality in the learning paradigm.
Concerns:
Upon revision, authors addressed all concerns of this reviewer, and the results are now presented in a way that facilitates objective evaluation. Authors' conclusions are supported by the results presented, and the strength of the proteomics approach is persuasively demonstrated.
Significance:
(1) Total neural proteome analysis has not been conducted before for learning-induced changes, though transcriptome analysis has been performed for odor learning (Lakhina et al., http://dx.doi.org/10.1016/j.neuron.2014.12.029). This warrants the novelty of this manuscript, because for some genes, protein levels may change even though mRNA levels remain the same. Although in a few reports TurboID has been used in C. elegans, this is the first report of a systematic analysis of tissue-specific differential proteomics.
(2) Authors found five mutants that have abnormality in the salt learning. These genes have not been described to have the abnormality, providing novel knowledge to the readers, especially those who work on C. elegans behavioural plasticity. Especially, involvement of acetylcholine neurotransmission has not been addressed before. Although transgenic rescue experiments have not been performed except kin-2, and the site of action (neurons involved) has not been tested in this manuscript, it will open the venue to further determine the way in which acetylcholine receptors, cAMP pathway etc. influences the learning process.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors demonstrate that CRF neurons in the extended amygdala form GABAergic synapses onto cholinergic interneurons and that CRF can excite these neurons. The evidence is strong, however, the authors fail to make a compelling connection showing CRF released from these extended amygdala neurons is mediating any of these effects. Further, they show that acute alcohol appears to modulate this action, although the effect size is not particularly robust.
Strengths:
This is an exciting connection from the extended amygdala to the striatum that provides a new direction for how these regions can modulate behavior. The work is rigorous and well done.
Weaknesses:
While the authors show that opto stim of these neurons can increase firing, this is not shown to be CRFR1 dependent. In addition, the effects of acute ethanol are not particularly robust or rigorously evaluated. Further, the opto stim experiments are conducted in an Ai32 mouse, so it is impossible to determine if that is from CEA and BNST, vs. another population of CRF-containing neurons. This is an important caveat.
-
-
www.sciencedirect.com www.sciencedirect.com
-
RRID:ZFIN_ZDB-ALT-110603-3
DOI: 10.1016/j.celrep.2025.116509
Resource: None
Curator: @scibot
SciCrunch record: RRID:ZFIN_ZDB-ALT-110603-3
-
RRID:ZFIN_ZDB-ALT-120117-3
DOI: 10.1016/j.celrep.2025.116509
Resource: (ZFIN Cat# ZDB-ALT-120117-3,RRID:ZFIN_ZDB-ALT-120117-3)
Curator: @scibot
SciCrunch record: RRID:ZFIN_ZDB-ALT-120117-3
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this study, Philipp et al. investigate how a monkey learns to compensate for a large, chronic biomechanical perturbation - a tendon transfer surgery, swapping the actions of two muscles that flex and extend the fingers. After performing the surgery and confirming that the muscle actions are swapped, the authors follow the monkeys' performance on grasping tasks over several months. There are several main findings:
(1) There is an initial stage of learning (around 60 days), where monkeys simply swap the activation timing of their flexors and extensors during the grasp task to compensate for the two swapped muscles.
(2) This is (seemingly paradoxically) followed by a stage where muscle activation timing returns almost to what it was pre-surgery, suggesting that monkeys suddenly swap to a new strategy that is better than the simple swap.
(3) Muscle synergies seem remarkably stable through the entire learning course, indicating that monkeys do not fractionate their muscle control to swap the activations of only the two transferred muscles.
(4) Muscle synergy activation shows a similar learning course, where the flexion synergy and extension synergy activations are temporarily swapped in the first learning stage and then revert to pre-surgery timing in the second learning stage.
(5) The second phase of learning seems to arise from making new, compensatory movements (supported by other muscle synergies) that get around the problem of swapped tendons.
Strengths:
This study is quite remarkable in scope, studying two monkeys over a period of months after a difficult tendon-transfer surgery. As the authors point out, this kind of perturbation is an excellent testbed for the kind of long-term learning that one might observe in a patient after stroke or injury, and provides unique benefits over more temporary perturbations like visuomotor transformations and studying learning through development. Moreover, while the two-stage learning course makes sense, I found the details to be genuinely surprising--specifically the fact that: (1) muscle synergies continue to be stable for months after the surgery, despite now being maladaptive; and (2) muscle activation timing reverts to pre-surgery levels by the end of the learning course. These two facts together initially make it seem like the monkey simply ignores the new biomechanics by the end of the learning course, but the authors do well to explain that this is mainly because the monkeys develop a new kind of movement to circumvent the surgical manipulation.
I found these results fascinating, especially in comparison to some recent work in motor cortex, showing that a monkey may be able to break correlations between the activities of motor cortical neurons, but only after several sessions of coaching and training (Oby et al. PNAS 2019). Even then, it seemed like the monkey was not fully breaking correlations but rather pushing existing correlations harder to succeed at the virtual task (a brain-computer interface with perturbed control).
Weaknesses:
I found the analysis to be reasonably well considered and relatively thorough. However, I do have a few suggestions that I think may elevate the work, should the authors choose to pursue them.
First, I find myself wondering about the physical healing process from the tendon transfer surgery and how it might contribute to the learning. Specifically, how long does it take for the tendons to heal and bear forces? If this itself takes a few months, it would be nice to see some discussion of this.
Second, I see that there are some changes in the muscle loadings for each synergy over the days, though they are relatively small. The authors mention that the cosine distances are very small for the conserved synergies compared to distances across synergies, but it would be good to get a sense for how variable this measure is within synergy. For example, what is the cosine similarity for a conserved synergy across different pre-surgery days? This might help inform whether the changes post-surgery are within a normal variation or whether they reflect important changes in how the muscles are being used over time.
Last, and maybe most difficult (and possibly out of scope for this work): I would have ideally liked to see some theoretical modeling of the biomechanics so I could more easily understand what the tendon transfer did or how specific synergies affect hand kinematics before and after the surgery. Especially given that the synergies remained consistent, such an analysis could be highly instructive for a reader or to suggest future perturbations to further probe the effects of tendon transfer on long-term learning.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #4 (Public review):
Thank you to the authors for their detailed responses and changes in relation to my questions. They have addressed all my concerns around methodological and inference clarity. I would still recommend against the use of feature/pathway selection techniques where there is no way of applying formal error control. I am pleased to read, however, that the authors are planning to develop this in future work. My edited review reflects these changes:
The authors apply what I gather is a novel methodology titled "Multi-gradient Permutation Survival Analysis" to identify genes that are robustly associated with prognosis ("GEARs") using tumour expression data from 15 cancer types available in the TCGA. The resulting lists of GEARs are then interrogated for biological insights using a range of techniques including connectivity and gene enrichment analysis.
I reviewed this paper primarily from a statistical perspective. Evidently an impressive amount of work has been conducted, concisely summarised, and great effort has been undertaken to add layers of insight to the findings. I am no stranger to what an undertaking this would have been. My primary concern, however, is that the novel statistical procedure proposed, and applied to identify the gene lists, as far as I can tell offers no statistical error control nor quantification. Consequently we have no sense what proportion of the highlighted GEAR genes and networks are likely to just be noise.
Major comments:
The main methodology used to identify the GEAR genes, "Multi-gradient Permutation Survival Analysis" does not formally account for multiple testing and offers no formal error control. Meaning we are left without knowing what the family wise (aka type 1) error rate is among the GEAR lists, nor the false discovery rate. I appreciate the emphasis on reproducibility, but I would generally recommend against the use of any feature selection methodology which does not provide error quantification because otherwise we do not know if we are encouraging our colleagues and/or readers to put resource into lists of genes that contain more noise than not. I am glad though and appreciative that the authors intend to develop this in future work.
The authors make a good point that, despite lack of validation in an external independent dataset, it is still compelling work given the functional characterisation and literature validation. I am pleased though that the authors agree validation in an independent dataset is an important next step, and plan to do so in future work.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In the manuscript " Dynamics of mesoscale brain network during decision-making learning revealed by chronic, large-scale single-unit recording", Wang et al investigated mesoscale network reorganization during visual stimulus discrimination learning in mice using chronic, large-scale single-unit recordings across 10 cortical/subcortical regions. During learning, mice improved task performance mainly by suppressing licking on no-go trials. The authors found that learning induced restructuring of functional connectivity, with visual (V1, V2M) and frontal (OFC, M2) regions forming a task-relevant subnetwork during the acquisition of correct No-Go (CR) trials.
Learning also compressed sequential neural activation and broadened stimulus encoding across regions. In addition, a region's network connectivity rank correlated with its timing of peak visual stimulus encoding.
Optogenetic inhibition of orbitofrontal cortex (OFC) and high order visual cortex (V2M) impaired learning, validating its role in learning. The work highlights how mesoscale networks underwent dynamic structuring during learning.
Strengths:
The use of ultra-flexible microelectrode arrays (uFINE-M) for chronic, large-scale recordings across 10 cortical/subcortical regions in behaving mice represents a significant methodological advancement. The ability to track individual units over weeks across multiple brain areas will provide a rare opportunity to study mesoscale network plasticity.
While limited in scope, optogenetic inhibition of OFC and V2M directly ties connectivity rank changes to behavioral performance, adding causal depth to correlational observations.
Weaknesses:
The weakness is also related to the strength provided by the method. It is demonstrated in the original method that this approach in principle can track individual units for four months (Luan et al, 2017). The authors have not showed chronically tracked neurons across learning. Without demonstrating that and taking advantage of analyzing chronically tracked neurons, this approach is not different from acute recording across multiple days during learning. Many studies have achieved acute recording across learning using similar tasks. These studies have recorded units from a few brain areas or even across brain-wide areas.
Another weakness is that major results are based on analyses of functional connectivity that is calculated using the cross-correlation score of spiking activity (TSPE algorithm). Functional connection strengthen across areas is then ranked 1-10 based on relative strength. Without ground truth data, it is hard to judge the underlying caveats. I'd strongly advise the authors to use complementary methods to verify the functional connectivity and to evaluate the mesoscale change in subnetworks. Perhaps the authors can use one key information of anatomy, i.e. the cortex projects to the striatum, while the striatum does not directly affect other brain structures recorded in this manuscript.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors investigate how sleep loss and circadian disruption affect whole-organism metabolism in Drosophila melanogaster. They used chamber-based flow-through respirometry to measure oxygen consumption and carbon dioxide production in wild-type flies and in mutants with impaired sleep or circadian function. These measurements were then integrated with a previously published metabolomics dataset to explore how respiratory dynamics align with metabolic pathways. The central claim is that wild-type flies display anticipatory coordination of metabolic processes with circadian time, while mutants exhibit reactive shifts in substrate use, redox imbalance, and signs of mitochondrial stress.
Strengths:
The study has several strengths. Continuous high-resolution respirometry in flies is challenging, and its application across multiple genotypes provides good comparative insight. The conceptual framework distinguishing anticipatory from reactive metabolic regulation is interesting. The translational framing helps place the work in a broader context of sleep, circadian biology, and metabolic health.
Weaknesses:
At the same time, the evidence supporting the conclusions is somewhat limited. The metabolomics data were not newly generated but repurposed from prior work, reducing novelty. The biological replication in the respirometry assays is low, with only a small number of chambers per genotype. Importantly, respiratory parameters in flies are strongly influenced by locomotor activity, yet no direct measurements of activity were included, making it difficult to separate intrinsic metabolic changes from behavioral differences in mutants. In addition, repeated claims of "mitochondrial stress" are not directly substantiated by assays of mitochondrial function. The study also excluded female flies entirely, despite well-documented sex differences in metabolism, which narrows the generality of the findings.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The focus of the research is to understand how transcription factors with high expression in neural crest cell-derived cancers (e.g., neuroblastoma) and roles in neural crest cell development function to promote malignancy. The focus is on the transcription factor FOXC1 and using murine cell culture, gain- and loss-of-function approaches, and ChIP profiling, among other techniques, to place PKAC inhibitor ARHGAP36 mechanistically between FOXC1 and another pathway associated with malignancy, Sonic Hedgehog (SHH).
Strengths:
Major strengths are the mechanistic approaches to identify FOXC1 direct targets, definitively showing that FOXC1 transcriptional regulation of ARHGAP36 leads to dysregulation of SHH signaling downstream of ARHGAP36 inhibition of PKC. Starting from a screen of Foxc1 OE to get to ARHGAP36 and then using genetic and pharmacological manipulation to work through the mechanism is very well done. There is data that will be of use to others studying FOXC1 in mesenchymal cell types, in particular, the FOXC1 ChIP-seq.
Weaknesses:
Work is almost all performed in NIH3T3 or similar cells (mouse cells, not patient or mouse-derived cancer cells), so the link to neuroblastoma that forms the major motivation of the work is not clear. The authors look at ARHGAP36 levels in association with the neuroblastoma patient survival; however, the finding, though interesting and quite compelling, is misaligned with what the literature shows about FOXC1 and SHH, their high expression is associated with increased malignancy (also maybe worse outcomes?). Therefore, ARHGAP36 expression may be more complicated in a tumor cell or may be unrelated to FOXC1 or SHH, leaving one to wonder what the work in NIH3T3 cells, though well done, is telling us about the mechanisms of FOXC1 as an oncogene in neuroblastoma cells or in any type of cancer cell. Does it really function as an SHH activator to drive tumor growth? The 'oncogenic relevance' and 'contribution to malignancy' claimed in the last paragraph of the introduction are currently weakly supported by the data as presented. This could be improved by studying some of these mechanisms in patient-derived neuroblastoma cells with high FOXC1 expression. Does inhibiting FOXC1 change SHH and ARHGAP36 and have any effect on cell proliferation or migration? Alternatively, does OE of FOXC1 in NIH3T3 cells increase their migration or stimulate proliferation in some way, and is this dependent on ARHGAP36 or SHH? Application of their mechanistic approaches in cancer cells or looking for hallmarks of cancer phenotypes with FOXC1 OE (and dependent on SHH or ARHGAP36) could help to make a link with cellular phenotypes of malignant cells.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors present an in vitro evaluation of drug-drug interactions between artemisinins and quinoline antimalarials, as an important aspect for screening the current artemisinin-based combination therapies for Plasmodium falciparum. Using a revised pulsing assay, they report antagonism between dihydroartemisinin (DHA) and several quinolines, including chloroquine, piperaquine (PPQ), and amodiaquine. This antagonism is increased in CQ-resistant strains in isobologram analyses. Moreover, CQ co-treatment was found to induce artemisinin resistance even in parasites lacking K13 mutations during the ring-stage survival assay. This implies that drug-drug interactions, not just genetic mutations, can influence resistance phenotypes. By using a chemical probe for reactive heme, the authors demonstrate that quinolines inhibit artemisinin activation by rendering cytosolic heme chemically inert, thereby impairing the cytotoxic effects of DHA. The study also observed negative interactions in triple-drug regimens (e.g., DHA-PPQ-Mefloquine) and in combinations involving OZ439, a next-generation peroxide antimalarial. Taken together, these findings raise significant concerns regarding the compatibility of artemisinin and quinoline combinations, which may promote resistance or reduce efficacy.
Throughout the manuscript, no combinations were synergistic, which necessitates comparing the claims to a synergistic combination as a control. The lack of this positive control makes it difficult to contextualize the observed antagonism. Including a known synergistic pair (e.g., artemisinin + lumefantrine) throughout the study would have provided a useful benchmark to assess the relative impact of the drug interactions described.
Strengths:
This study demonstrates the following strengths:
(1) The use of a pulsed in vitro assay that is more physiologically relevant than the traditional 48h or 72h assays.
(2) Small molecule probes, H-FluNox, and Ac-H-FluNox to detect reactive cytosolic heme, demonstrating that quinolines render heme inert and thereby block DHA activation.
(3) Evaluates not only traditional combinations but also triple-drug combinations and next-generation artemisinins like OZ439. This broad scope increases the study's relevance to current treatment strategies and future drug development.
(4) By using the K13 wild-type parasites, the study suggests that resistance phenotypes can emerge from drug-drug interactions alone, without requiring genetic resistance markers.
Weaknesses:
(1) No combinations are shown as synergistic: it could be valuable to have a combination that shows synergy as a positive control (e.g, artemisinin + lumefantrine) throughout the manuscript. The absence of a synergistic control combination in the experimental design makes it more challenging to evaluate the relative impact of the described drug interactions.
(2) Evaluation of the choice of drug-drug interactions: How generalizable are the findings across a broad range of combinations, especially those with varied modes of action?
(3) The study would also benefit from a characterization of the molecular basis for the observed heme inactivation by quinolines to support this hypothesis - while the probe experiments are valuable, they do not fully elucidate how quinolines specifically alter heme chemistry at the molecular level.
(4) Suggestion of alternative combinations that show synergy could have improved the significance of the work.
(5) All data are derived from in vitro experiments, without accompanying an in vivo validation. While the pulsing assay improves physiological relevance, it still cannot fully capture the complexity of drug pharmacokinetics, host-parasite interactions, or immune responses present in living organisms.
(6) The absence of pharmacokinetic/pharmacodynamic modeling leaves questions about how the observed antagonism would manifest under real-world dosing conditions.
-
- Oct 2025
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors present a clearly written and beautifully presented piece of work demonstrating clear evidence to support the idea that BK channels and Cav1.3 channels can co-assemble prior to their assertion in the plasma membrane.
Strengths:
The experimental records shown back up their hypotheses and the authors are to be congratulated for the large number of control experiments shown in the ms.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary
Kong and coauthors describe and implement a method to correct local deformations due to beam-induced motion in cryo-EM movie frames. This is done by fitting a 3D spline model to a stack of micrograph frames using cross-correlation-based local patch alignment to describe the deformations across the micrograph in each frame, and then computing the value of the deformed micrograph at each pixel by interpolating the undeformed micrograph at the displacement positions given by the spline model. A graphical interface in cisTEM allows the user to visualise the deformations in the sample, and the method has been proven to be successful by showing improvements in 2D template matching (2DTM) results on the corrected micrographs using five in situ samples.
Impact
This method has great potential to further streamline the cryo-EM single particle analysis pipeline by shortening the required processing time as a result of obtaining higher quality particles early in the pipeline, and is applicable to both old and new datasets, therefore being relevant to all cryo-EM users.
Strengths
(1) One key idea of the paper is that local beam induced motion affects frames continuously in space (in the image plane) as well as in time (along the frame stack), so one can obtain improvements in the image quality by correcting such deformations in a continuous way (deformations vary continuously from pixel to pixel and from frame to frame) rather than based on local discrete patches only. 3D splines are used to model the deformations: they are initialised using local patch alignments and further refined using cross-correlation between individual patch frames and the average of the other frames in the same patch stack.
(2) Another strength of the paper is using 2DTM to show that correcting such deformations continuously using the proposed method does indeed lead to improvements. This is shown using five in situ datasets, where local motion is quantified using statistics based on the estimated motions of ribosomes.
Weaknesses
(1) While very interesting, it is not clear how the proposed method using 3D splines for estimating local deformations compares with other existing methods that also aim to correct local beam-induced motion by approximating the deformations throughout the frames using other types of approximation, such as polynomials, as done, for example MotionCor2.
(2) The use of 2DTM is appropriate, and the results of the analysis are enlightening, but one shortcoming is that some relevant technical details are missing. For example, the 2DTM SNR is not defined in the article, and it is not clear how the authors ensured that no false positives were included in the particles counted before and after deformation correction. The Jupyter notebooks where this analysis was performed have not been made publicly available.
(3) It is also not clear how the proposed deformation correction method is affected by CTF defocus in the different samples (are the defocus values used in the different datasets similar or significantly different?) or if there is any effect at all.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In the reviewed manuscript, researchers aimed to overcome the obstacles of high-resolution imaging of intact liver tissue. They report successful modification of the existing CUBIC protocol into Liver-CUBIC, a high-resolution multiplex 3D imaging method that integrates multicolor metallic compound nanoparticle (MCNP) perfusion with optimized liver tissue clearing, significantly reducing clearing time and enabling simultaneous 3D visualization of the portal vein, hepatic artery, bile ducts, and central vein spatial networks in the mouse liver. Using this novel platform, the researchers describe a previously unrecognized perivascular structure they termed Periportal Lamellar Complex (PLC), regularly distributed along the portal vein axis. The PLC originates from the portal vein and is characterized by a unique population of CD34⁺Sca-1⁺ dual-positive endothelial cells. Using available scRNAseq data, the authors assessed the CD34⁺Sca-1⁺ cells' expression profile, highlighting the mRNA presence of genes linked to neurodevelopment, biliary function, and hematopoietic niche potential. Different aspects of this analysis were then addressed by protein staining of selected marker proteins in the mouse liver tissue. Next, the authors addressed how the PLC and biliary system react to CCL4-induced liver fibrosis, implying PLC dynamically extends, acting as a scaffold that guides the migration and expansion of terminal bile ducts and sympathetic nerve fibers into the hepatic parenchyma upon injury.
The work clearly demonstrates the usefulness of the Liver-CUBIC technique and the improvement of both resolution and complexity of the information, gained by simultaneous visualization of multiple vascular and biliary systems of the liver at the same time. The identification of PLC and the interpretation of its function represent an intriguing set of observations that will surely attract the attention of liver biologists as well as hepatologists; however, some claims need more thorough assessment by functional experimental approaches to decipher the functional molecules and the sequence of events before establishing the PLC as the key hub governing the activity of biliary, arterial, and neuronal liver systems. Similarly, the level of detail of the methods section does not appear to be sufficient to exactly recapitulate the performed experiments, which is of concern, given that the new technique is a cornerstone of the manuscript.
Nevertheless, the work does bring a clear new insight into the liver structure and functional units and greatly improves the methodological toolbox to study it even further, and thus fully deserves the attention of readers.
Strengths:
The authors clearly demonstrate an improved technique tailored to the visualization of the liver vasulo-biliary architecture in unprecedented resolution.
This work proposes a new biological framework between the portal vein, hepatic arteries, biliary tree, and intrahepatic innervation, centered at previously underappreciated protrusions of the portal veins - the Periportal Lamellar Complexes (PLCs).
Weaknesses:
Possible overinterpretation of the CD34+Sca1+ findings was built on re-analysis of one scRNAseq dataset.
Lack of detail in the materials and methods section greatly limits the usefulness of the new technique to other researchers.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Recent studies have established that trypanocidal drugs, including pentamidine and melarsoprol, enter the trypanosomes via the glyceroaquaporin AQP2 (TbAQP2). Interestingly, drug resistance in trypanosomes is, at least in part, caused by recombination with the neighbouring gene, AQP3, which is unable to permeate pentamidine or melarsoprol. The effect of the drugs on cells expressing chimeric proteins is significantly reduced. In addition, controversy exists regarding whether TbAQP2 permeates the drugs like an ion channel, or whether it serves as a receptor that triggers downstream processes upon drug binding. In this study the authors set out to achieve these objectives: 1) to understand the molecular interactions between TbAQP2 and glycerol, pentamidine, and melarsoprol, and 2) to determine the mechanism by which mutations that arise from recombination with TbAQP3 result in reduced drug permeation.
The cryo-EM structures provide details of glycerol and drug binding, and show that glycerol and the drugs occupy the same space within the pore. Finally, MD simulations and lysis assays are employed to determine how mutations in TbAQP2 result in reduced permeation of drugs by making entry and exit of the drug relatively more energy-expensive. Overall, the strength of evidence used to support the author's claims is solid.
Strengths:
The cryo-EM portion of the study is strong, and while the overall resolution of the structures is in the 3.5Å range, the local resolution within the core of the protein and the drug binding sites is considerably higher (~2.5Å).<br /> I also appreciated the MD simulations on the TbAQP2 mutants and the mechanistic insights that resulted from this data.
Weaknesses:
(1) The authors do not provide any experimental validation the drug binding sites in TbAQP2 due to lacking resources. However, the claims have been softened in the revised paper.
-
-
www.instructables.com www.instructables.com
-
Making Tab Stops for an LC Smith Typewriter : 3 Steps - Instructables<br /> by [[Instructables]]<br /> accessed on 2025-10-30T11:30:29
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The study consists of extensive computational analyses of their previously released Patch-seq data on single MN1-Ib and MNISN-Is neurons. The authors demonstrate the diversity of A>I editing events at single-cell resolution in two different neuronal cell types, identifying numerous A>I editing events that vary in their proportion, including those that cause missense mutations in conserved amino acids. They also consider "noncanonical" edits, such as C>T and G>A, and integrate publicly available data to support these analyses.
In general, the study contains a valuable resource to assess RNA editing in single neurons and opens several questions regarding the diversity and functional implications of RNA editing at single-cell resolution. The conclusions from the study are generally supported by their data; however, the study is currently based on computational predictions and would therefore benefit from experimentation to support their hypotheses and demonstrate the effects of the editing events identified on neuronal function and phenotype.
Strengths:
The study uses samples that are technically difficult to prepare to assess cell-type-specific RNA editing events in a natural model. The study also uses public data from different developmental stages that demonstrate the importance of considering cell type and developmental stage-specific RNA regulation. These critical factors, particularly that of developmental timing, are often overlooked in mechanistic studies.
Extensive computational analysis, using public pipelines, suitable filtering criteria, and accessible custom code, identifies a number of RNA editing events that have the potential to impact conserved amino acids and have subsequent effects on protein function. These observations are supported through the integration of several public data sets to investigate the occurrence of the edits in other data sets, with many identified across multiple data sets. This approach allowed the identification of a number of novel A>I edits, some of which appear to be specific to this study, suggesting cell/developmental specificity, whilst others are present in the public data sets but went unannotated.
The study also considers the role of Adar in the generation of A>I edits, as would be expected, by assessing the effect of Adar expression on editing rates using public data from adar mutant tissue to demonstrate that the edits conserved between experiments are mainly Adar-sensitive. This would be stronger if the authors also performed Patch-seq experiments in adar mutants to increase confidence in the identified edit sites.
Weaknesses:
Whilst the study makes interesting observations using advanced computational approaches, it does not demonstrate the functional implications of the observed editing events. The functional impact of the edits is inferred from either the nature of the change to the coding sequence and the amino acid conservation, or through integration of other data sets. Although these could indeed imply function, further experimentation would be required to confirm such as using their Alphafold models to predict any changes in structure. This limitation is acknowledged by the authors, but the overall strength of the interpretation of the analysis could be softened to represent this.
The study uses public data from more diverse cellular populations to confirm the role of Adar in introducing the A>I edits. Whilst this is convincing, the ideal comparison to support the mechanism behind the identified edits would be to perform patch-seq experiments on 1b or 1s neurons from adar mutants. However, although this should be considered when interpreting the data, these experiments would be a large amount of work and beyond the scope of the paper.
By focusing on the potential impact of editing events that cause missense mutations in the CDS, the study may overlook the importance of edits in noncoding regions, which may impact miRNA or RNA-binding protein target sites. Further, the statement that noncanonical edits and those that induce silent mutations are likely to be less impactful is very broad and should be reconsidered. This is particularly the case when suggesting that silent mutations may not impact the biology. Given the importance of codon usage in translational fidelity, it is possible that silent mutations induced by either A>I or noncanonical editing in the CDS impact translation efficiency. Indeed, this could have a greater impact on protein production and transcript levels than a single amino acid change alone.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This manuscript aims to determine cultural biases and misconceptions in inclusive sex research and evaluate the efficacy of interventions to improve knowledge and shift perceptions to decrease perceived barriers for including both sexes in basic research.
Overall, this study demonstrates that despite the intention to include both sexes and a general belief in the importance of doing so, relatively few people routinely include both sexes. Further, the perceptions of barriers to doing so are high, including misconceptions surrounding sample size, disaggregation, and variability of females. There was also a substantial number of individuals without the statistical knowledge to appropriately analyze data in studies inclusive of sex. Interventions increased knowledge and decreased perception of barriers.
Strengths:
(1) This manuscript provides evidence for the efficacy of interventions for changing attitudes and perceptions of research.
(2) This manuscript also provides a training manual for expanding this intervention to broader groups of researchers.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Hull et al examine Drosophila mutants for the Gaucher's disease locus GBA1/Gba1b, a locus that, when heterozygous, is a risk factor for Parkinson's. Focusing on the Malpighian tubules and their function, they identify a breakdown of cell junctions, loss of haemolymph filtration, sensitivity to ionic imbalance, water retention, and loss of endocytic function in nephrocytes. There is also an imbalance in ROS levels between the cytoplasm and mitochondria, with reduced glutathione levels, rescue of which could not improve longevity. They observe some of the same phenotypes in mutants of Parkin, but treatment by upregulation of autophagy via rapamycin feeding could only rescue the Gba1b mutant and not the Parkin mutant.
Strengths:
The paper uses a range of cellular, genetic, and physiological analyses and manipulations to fully describe the renal dysfunction in the GBa1b animals. The picture developed has depth and detail; the data appears sound and thorough.
Weaknesses:
The paper relies mostly on the biallelic Gba1b mutant, which may reflect dysfunction in Gaucher's patients, though this has yet to be fully explored. The claims for the heterozygous allele and a role in Parkinson's is a little more tenuous, making assumptions that heterozygosity is a similar but milder phenotype than the full loss-of-function.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors aimed to define the somatic mutational landscape and transcriptomic expression of the ZNF217, ZNF703, and ZNF750 genes in breast cancers from Kenyan women and to investigate associations with clinicopathological features like HER2 status and cancer stage. They employed whole-exome and RNA-sequencing on 23 paired tumor-normal samples to achieve this.
Strengths:
(1) A major strength is the focus on a Kenyan cohort, addressing a critical gap in genomic studies of breast cancer, which are predominantly based on European or Asian populations.
(2) The integration of DNA- and RNA-level data from the same patients provides a comprehensive view, linking genetic alterations to expression changes.
Weaknesses:
(1) The small cohort size (n=23) significantly limits the statistical power to detect associations between genetic features and clinical subgroups (e.g., HER2 status, stage), rendering the negative findings inconclusive.
(2) The study is primarily descriptive. While it effectively catalogs mutations and expression changes, it does not include functional experiments to validate the biological impact of the identified alterations.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this study, the authors investigate how the structural state of the microtubule lattice influences the accessibility of the α-tubulin C-terminal tail (CTT). By developing and applying new biosensors, they reveal that the tyrosinated CTT is largely inaccessible under normal conditions but becomes more accessible upon changes to the tubulin conformational state induced by taxol treatment, MAP expression, or GTP-hydrolysis-deficient tubulin. The combination of live imaging, biochemical assays, and simulations suggests that the lattice conformation regulates the exposure of the CTT, providing a potential mechanism for modulating interactions with microtubule-associated proteins. The work addresses a highly topical question in the microtubule field and proposes a new conceptual link between lattice spacing and tail accessibility for tubulin post-translational modification.
Strengths:
(1) The study targets a highly relevant and emerging topic-the structural plasticity of the microtubule lattice and its regulatory implications.
(2) The biosensor design represents a methodological advance, enabling direct visualization of CTT accessibility in living cells.
(3) Integration of imaging, biochemical assays, and simulations provides a multi-scale perspective on lattice regulation.
(4) The conceptual framework proposed lattice conformation as a determinant of post-translational modification accessibility is novel and potentially impactful for understanding microtubule regulation.
Weaknesses:
There are a number of weaknesses in the paper, many of which can be addressed textually. Some of the supporting evidence is preliminary and would benefit from additional experimental validation and clearer presentation before the conclusions can be considered fully supported.
In particular, the authors should directly test in vitro whether Taxol addition can induce lattice exchange (see comments below).
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The AAA+ protease LON1P is a central component of mitochondrial protein quality control and has crucial functions in diverse processes. Cryo-EM structures of LON1P defined inactive and substrate-processing active states. Here, the authors determined multiple new LON1P structural states by cryo-EM in the presence of diverse substrates. The structures are defined as on-pathway intermediates to LON1P activation. A C3-symmetry state is suggested to function as a checkpoint to scan for LON1P substrates and link correct substrate selection to LON1P activation.
Strengths:
The determination of multiple structures provides relevant information on substrate-triggered activation of LON1P. The authors support structural data by biochemical analysis of structure-based mutants.
Weaknesses:
How substrate selection is achieved remains elusive, also because substrates are not detectable in the diverse structures. It also remains in parts unclear whether mutant phenotypes can be specifically linked to a single structural state (C3). Some mutant phenotypes appear complex and do not seem to be in line with the model proposed.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This article is about the neural circuitry underlying motion vision in the fruit fly. Specifically, it regards the roles of two identified neurons, called C2 and C3, that form columnar connections between neurons in the lamina and medulla, including neurons that are presynaptic to the elementary motion detectors T4 and T5. The approach takes advantage of specific fly lines in which one can disable the synaptic outputs of either or both of the C2/3 cell types. This is combined with optical recording from various neurons in the circuit, and with behavioral measurements of the turning reaction to moving stimuli.
The experiments are planned logically. The effects of silencing the C2/C3 neurons are substantial in size. The dominant effect is to make the responses of downstream neurons more sustained, consistent with a circuit role in feedback or feedforward inhibition. Silencing C2/C3 also makes the motion-sensitive neurons T4/T5 less direction-selective. However, the turning response of the fly is affected only in subtle ways. Detection of motion appears unaffected. But the response fails to discriminate between two motion pulses that happen in close succession. One can conclude that C2/C3 are involved in the motion vision circuit, by sharpening responses in time, though they are not essential for its basic function of motion detection.
Strengths:
The combination of cutting-edge methods available in fruit fly neuroscience. Well-planned experiments carried out to a high standard. Convincing effects documenting the role of these neurons in neural processing and behavior.
Weaknesses:
The report could benefit from a mechanistic argument linking the effects at the level of single neurons, the resulting neural computations in elementary motion detectors, and the altered behavioral response to visual motion.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Donofrio et al. report a new observation that in normal aging mice, anti-calbindin whole-mount staining and coronal immunohistochemistry in the cerebellum often show a sagittally patterned loss of Purkinje cells with age. The authors address a central concern that calbindin antibody staining alone is not sufficient to definitively assess Purkinje cell loss, and corroborate their antibody staining data with transgenic Pcp2-CRE x flox-GFP reporter mice and Neutral Red staining. The authors then investigate whether this patterned Purkinje loss correlates with the known parasagittal expression of zebrin-II, finding a strong but imperfect correlation with zebrin-II antibody staining. They next draw a connection between this age-related Purkinje loss to the age-related decline in motor function in mice, with trending but non-significant statistical association between the severity/patterning of Purkinje loss and motor phenotypes within cohorts of aged mice. Finally, the authors look at post-mortem human cerebellar tissues from deceased healthy donors between 21 and 74 years of age, finding a positive correlation between Purkinje degeneration and age, but with unknown spatial patterning.
The conclusions drawn from this study are well supported by the data provided, with image quantification corroborating visual observations. The authors highlight several examples of parasagittal patterning of Purkinje cell degeneration in disease, and they show that proper methodologies must be used to account for these patterns to avoid highly variable data in the sagittal plane. The authors aptly point out that additional work is needed to investigate the spatial patterns of Purkinje cell loss in the human cerebellum.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this manuscript, Edwards et al. describe hamFISH, a customizable and cost-efficient method for performing targeted spatial transcriptomics. hamFISH utilizes highly amplified multiplexed branched DNA amplification, and the authors extensively describe hamFISH development and its advantages over prior variants of this approach.
The authors then used hamFISH to investigate an important circuit in the mouse brain for social behavior, the medial amygdala (MeA). To develop a hamFISH probe set capable of distinguishing MeA neurons, the authors mined published single cell RNA-sequencing datasets of the MeA, ultimately creating a panel of 32 hamFISH probes that mostly cover the identified MeA cell types. They evaluated over 600,000 MeA cells and classified neurons into 16 inhibitory and 10 excitatory types, many of which are spatially clustered.
The authors combined hamFISH with viral and other circuit tracer injections to determine whether the identified MeA cell populations sent and/or received unique inputs from connected brain regions, finding evidence that several cell types had unique patterns of input and output. Finally, the authors performed hamFISH on the brains of male mice that were placed in behavioral conditions that elicit aggressive, infanticidal, or mating behaviors, finding that some cell populations are selectively activated (as assessed by c-fos mRNA expression) in specific social contexts.
Strengths:
(1) The authors developed an optimized tissue preparation protocol for hamFISH and implemented oligopools instead of individually synthesized oligonucleotides to reduce costs. The branched DNA amplification scheme improved smFISH signal compared to previous methods, and multiple variants provide additional improvements in signal intensity and specificity. Compared to other spatial transcriptomics methods, the pipeline for imaging and analysis is streamlined, and is compatible with other techniques like fluorescence-based circuit tracing. This approach is cost-effective and has several advantages that make it a valuable addition to the list of spatial transcriptomics toolkits.
(2) Using 31 probes, hamFISH was able to detect 16 inhibitory and 10 excitatory neuron types in the MeA subregions, including the vast majority of cell types identified by other transcriptomics approaches. The authors quantified the distributions of these cell types along the anterior-posterior, dorsal-ventral, and medial-lateral axes, finding spatial segregation among some, but not all, MeA excitatory and inhibitory cell types. The authors additionally identified a class of inhibitory neurons expressing Ndnf (and a subset of these that express Chrna7) that project to multiple social chemosensory circuits.
(3) The authors combined hamFISH with MeA input and output mapping, finding cell-type biases in the projections to the MPOA, BNST, and VMHvl, and inputs from multiple regions.
(4) The authors identified excitatory and inhibitory cell types, and patterns of activity across cell types, that were selectively activated during various social behaviors, including aggression, mating, and infanticide, providing new insights and avenues for future research into MeA circuit function.
Weaknesses:
(1) Gene selection for hamFISH is likely to still be a limiting factor, even with the expanded (32-probe) capacity. This may have contributed to the lack of ability to identify sexually dimorphic cell types (Fig. S2B). This is an expected tradeoff for a method that has major advantages in terms of cost and adaptability.
(2) Adaptation of hamFISH, for example, to adapt it to other brain regions or tissues, may require extensive optimization. This does not preclude it from being highly useful for other brain regions with extra effort.
(3) Pairing this method with behavioral experiments is likely to require further optimization, as c-fos mRNA expression is an indirect and incomplete survey of neuronal activity (e.g. not all cell types upregulate c-fos when electrically active). As such, there is a risk of false negative results that limit its utility for understanding circuit function.
(4) The incompatibility of hamFISH with thicker tissue samples and minimal optical sectioning introduce additional technical limitations. For example, it would be difficult to densely sample larger neural circuits using serial 20 micron sections.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors present data demonstrating that optogenetic inhibition of either D1- or D2-MSNs in the NAc Shell attenuates expression of sensory-specific PIT while largely sparing value-based decision on an instrumental task. They also provide evidence that SS-PIT depends on D1-MSN projections from the NAc-Shell to the VP, whereas projections from D2-MSNs to the VP do not contribute to SS-PIT.
Strengths:
This is clearly written. The evidence largely supports the authors' interpretations, and these effects are somewhat novel, so they help advance our understanding of PIT and NAc-Shell function.
Weaknesses:
I think the interpretation of some of the effects (specifically the claim that D1-MSNs do not contribute to value-based decision making) is not fully supported by the data presented.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Jo and colleagues set out to investigate the origins and functions of localized FGF/ERK signaling for the differentiation and spatial patterning of primitive streak fates of human embryonic stem cells in a well-established micropattern system. They demonstrate that endogenous FGF signaling is required for ERK activation in a ring-domain in the micropatterns, and that this localized signaling is directly required for differentiation and spatial patterning of specific cell types. Through high-resolution microscopy and transwell assays, they show that cells receive FGF signals through basally localized receptors. Finally, the authors find that there is a requirement for exogenous FGF2 to initiate primitive streak-like differentiation, but endogenous FGFs, especially FGF4 and FGF17, fully take over at later stages.
Even though some of the authors' findings - such as the localized expression of FGF ligands during gastrulation and the importance of FGF/ERK signaling for cell differentiation in the primitive streak - have been reported in model organisms before, this is one of the first studies to investigate the role of FGF signaling during primitive streak-like differentiation of human cells. In doing so, the paper reports a number of interesting and valuable observations, namely the basal localization of FGF receptors which mirrors that of BMP and Nodal receptors, as well as the existence of a positive feedback loop centered on FGF signaling that drives primitive-streak differentiation. In the revised version of their work, the authors have furthermore dissected the role of different FGFs through knockdown approaches. These experiments reveal discrete functions for different FGF genes in their system, as well as interesting differences between the role of specific FGFs in human compared to model systems.
Comments on revisions:
The authors have appropriately addressed all comments and suggestions from the previous round of review. The only textual change that I would still like to suggest is to write explicitly in the main text corresponding to Fig. 1 that the mTESR1 medium used for these initial experiments already contains FGF. This is something that is probably known to experts in the field, but not necessarily to a broader readership.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #4 (Public review):
Summary:
Mason DE et al. have extended their previous study on continuous migration of cells regulated by a feedback loop that controls gene expression by YAP and TAZ. Time scale of the negative feedback loop is derived from the authors' adhesion-spreading-polarization-migration (ASPM) assay. Involvement of transcription-translation in the negative feedback loop is evidenced by the experiments using Actinomycin D. The time scale of mechanotransduction-dependent feedback demonstrated by cytoskeletal alteration in the actinomycin D-treated endothelial colony forming cells (ECFCs) and that shown in the ECFCs depleted of YAP/TAZ by siRNA. The authors examine the time scale when ECFCs are attached to MeHA matrics (soft, moderate, and stiff substrate) and show the conserved time scale among the conditions they use, although instantaneous migration, cell area, and circularity vary. Finally, they tried to confirm that the time scale of the feedback loop-dependent endothelial migration by the effect of washout of Actinomycin D (inhibition of gene transcription), Puromycin (translational inhibition), and Verteporfin (YAP/TAZ inhibitor) on ISV extension during sprouting angiogenesis. They conclude that endothelial motility required for vascular morphogenesis is regulated by a mechanotransduction-mediated feedback loop that is dependent on YAP/TAZ-dependent transcriptional regulation.
Strengths:
The authors conduct ASPM assay to find the time scale of feedback when ECFCs attach to three different matrics. They observe the common time scale of feedback. Thus, under very specific conditions they use, the reproducibility is validated by their ASPM assay. The feedback loop mediated by inhibition of gene expression by Actinomycin D is similar to that obtained from YAP/TAZ-depleted cells, suggesting the mechanotranduction might be involved in the feedback loop. The time scale representing infection point might be interesting when considering the continuous motility in cultured endothelial cells, although it might not account for the migration of endothelial cells that is controlled by a wide variety of extracellular cues. In vivo, stiffness of extracellular matrix is merely one of the cues.
Weaknesses:
ASPM assay is based on attachment-dependent phenomenon. The time scale, including the inflection point determined by ASPM experiments using cultured cells and the mechanotransduction-based theory, do not seem to fit in vivo ISV elongation. Although it is challenging to find the conserved theory of continuous cell motility of endothelial cells, the data is preliminary and does not support the authors' claim. There is no evidence that mechanotransduction solely determines the feedback loop during elongation of ISVs.
Comments on revisions:
The authors' methods using ASPM assay might suggest the feedback loop by their in vitro culture assay. They still need to confirm the loop in vivo using zebrafish intersegmental vessels. The time course of the feedback loop is supported by the ASPM assay. However, the feedback loop is not confirmed in vivo, although it might be suggested by the phenotypes of the ISV treated with drugs. Thus, in the abstract and in the results section, they had better rewrite the interpretation. They have not yet confirmed the feedback loop in vivo.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Cuentas-Condori et al. generate cell-specific tools for visualizing the endogenous expression of, as well as knocking out, four different classes of neurotransmitter vesicular transporters (glutamatergic, cholinergic, GABAergic, and monoaminergic) in C. elegans. They then use these tools in an intersectional strategy to provide evidence for the co-expression of these transporters in individual neurons, suggesting co-transmission of the associated neurotransmitters.
Strengths:
A major strength of the work is the generation of several endogenous tools that will be of use to the community. Additionally, this adds to accumulating evidence of co-transmission of different classes of neurotransmitters in the nervous system.
Weaknesses:
A weakness of the study is a lack of comparison to previously published single-cell sequencing data. These tools are alternatively described in the manuscript as superior to the sequencing data and as validation of the sequencing data, but neither claim can be assessed without knowing how they compare and contrast to that data. It is thus not clear to what extent the conclusions of this paper are an advance over what could be determined from the sequencing data on its own. Finally, some technical considerations should be discussed as potential caveats to the robustness of their intersectional strategy for concluding that certain genes are indeed co-expressed. Overall, claims about co-transmission should be tempered by the caveats presented in the discussion, suggesting that co-expression of these transporters is not in and of itself sufficient for neurotransmitter release.
-
-
-
Reviewer #3 (Public review):
In this manuscript, the authors introduce Megabouts, a software package designed to standardize the analysis of larval zebrafish locomotion, through clustering the 2D posture time series into canonical behavioral categories. Beyond a first, straightforward segmentation that separates glides from powered movements, Megabouts uses a Transformer neural network to classify the powered movements (bouts). This Transformer network is trained with supervised examples. The authors apply their approach to improve the quantification of sensorimotor transformations and enhance the sensitivity of drug-induced phenotype screening. Megabouts also includes a separate pipeline that employs convolutional sparse coding to analyze the less predictable tail movements in head-restrained fish.
I presume that the software works as the authors intend, and I appreciate the focus on quantitative behavior. My primary concerns reflect an implicit oversimplification of animal behavior. Megabouts is ultimately a clustering technique, categorizing powered locomotion into distinct, labelled states which, while effective for analysis, may confuse the continuous and fluid nature of animal behavior. Certainly, Megabouts could potentially miss or misclassify complex, non-stereotypical movements that do not fit the defined categories. In fact, it appears that exactly this situation led the authors to design a new clustering for head-restrained fish. Can we anticipate even more designs for other behavioral conditions?
Ultimately, I am not yet convinced that Megabouts provides a justifiable picture of behavioral control. And if there was a continuous "control knob", which seems very likely, wouldn't that confuse the clustering process, as many distinct clusters would correspond to, say, different amplitudes of the same control knob?
There has been tremendous recent progress in the measurement and analysis of animal behavior, including both continuous and discrete perspectives. However, the supervised clustering approach described here feels like a throwback to an earlier era. Yes, it's more automatic and quantifiable, and the amount of data is fantastic. But ultimately, the method is conceptually bound to the human eye in conditions where we are already familiar.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this paper, Winchester and colleagues investigated melodic perception in natural music listening. They highlight the central role of attentional processes in identifying one particular stream in polyphonic material, and propose to compare several theoretical accounts, namely (1) divided attention, (2) figure-ground separation, and (3) stream integration. In parallel, the authors compare the relative strength of exogenous attentional effects (i.e., salience) produced by two common traits of melodies: high-pitch (compared to other voices), and attractive statistics. To ensure the generalisability of their results to real-life listening contexts, they developed a new uninstructed listening paradigm in which participants can freely attend to any part of a musical stimulus.
Major strengths and weaknesses of the methods and results:
(1) Winchester and colleagues capitalized on previous attention decoding techniques and proposed an uninstructed listening paradigm. This is an important innovation for the study of music perception in ecological settings, and it is used here to investigate the spontaneous attentional focus during listening. The EEG decoding results obtained are coherent with the behavioral data, suggesting that the paradigm is robust and relevant.
(2) The authors first evaluate the relative importance of high-pitch and statistics in producing an attentional bias (Figure 2). Behavioral results show a clear pattern, in which both effects are present, with a dominance of the high-pitch one. The only weakness inherent to this protocol is that behavioral responses are measured based on a second presentation of short samples, which may induce a different attentional focus than in the first uninstructed listening.
(3) Then, the analyses of EEG data compare the decoding results of each melody (the high or low voice, and with "richer" or "poorer" statistics), and show a similar pattern of results. However, this report leaves open the possibility of a confounding factor. In this analysis, a TRF decoding model is first trained based on the presentation of monophonic samples, and it is later used to decode the envelope of the corresponding melodies in the polyphonic scenario. The fitting scores of the training phase are not reported. If the high-pitch or richer melodies were to produce higher decoding scores during monophonic listening (due to properties of the physiological response, or to perceptual processes), a similar difference could be expected during polyphonic listening. To capture attentional biases specifically, the decoding scores in the polyphonic conditions should be compared to the scores in the monophonic conditions, and attention could be expected to increase the decoding of the attended stream or decrease the unattended one.
(4) Then, Winchester and colleagues investigate the processing of melodic information by evaluating the encoding of melodic surprise and uncertainty (Figure 3). They compare the surprise and uncertainty estimated from a monophonic or a polyphonic model (Anticipatory Music Transformer), and analyse the data with a CCA analysis. The results show a double dissociation, where the processing of melodies with a strong attentional bias (high-pitch, rich statistics) is better approximated with a monophonic model, while a polyphonic model better classifies the other melodies. While this global result is compelling, it remains a preliminary and intriguing finding, and the manuscript does not further investigate it. As it stands, the result appears more like a starting point for further exploration than a definitive finding that can support strong theoretical claims. First, it could be complemented by a comparison of the encoding of individual melodies (e.g., AMmono high-voice vs AMmono low-voice, in PolyOrig and PolyInv conditions) to highlight a more direct correspondence with the previous results (Figure 2) and allow a more precise interpretation. Second, additional analyses or experiments would be needed to unpack this result and provide greater explanatory power. Additionally, the CCA analysis is not described in the method. The statistical testing conducted on this analysis seems to be performed across the 250 repetitions of the evaluation rather than across the 40 participants, which may bias the resulting p-values. Moreover, the choice and working principle of the Anticipatory Music Transformer are not described in the method. Overall, these results seem at first glance solid, but the missing parts of the method do not allow for full evaluation or replication of them.
An appraisal of whether the authors achieved their aims, and whether the results support their conclusions:
(1) Winchester and colleagues aimed at identifying the melodic stream that attracts attention during the listening of natural polyphonic music, and the underlying attentional processes. Their behavioral results confirm that high-pitched and attractive statistics increase melodic salience with a greater effect size of the former, as stated in the discussion. The TRF analyses of EEG data seem to show a similar pattern, but could also be explained by confounding factors. Next, the authors interpret the CCA results as the results of stream segregation when there is a high melodic salience, and stream integration when there are weaker attentional biases. These interpretations seem to be supported by the data, but unfortunately, no additional analyses or experiments have been conducted to further evaluate this hypothesis. The authors also acknowledge that their results do not show whether stream segregation occurs via divided attention or figure-ground separation. However, the lack of information about the music model used (Anticipatory Music Model) and the way it was set up raises some questions about its relevance and limits as a model of cognition (e.g. Is this transformer a "better" model of the listeners' expectations than the well-established IDyOM model, and why ?), and about the validity of those results.
(2) Overall, the authors achieved most of the aims presented in the introduction, although they couldn't give a more precise account of the attentional processes at stake. The interpretations are sound and not overstated, with the exception of potential confounding factors that could compromise the conclusions on the neural tracking of salient melodies (EEG results, Figure 2).
Impact of the work on the field, and the utility of the methods and data to the community:
The new uninstructed listening paradigm introduced in this paper will likely have an important impact on psychologists and neuroscientists working on music perception and auditory attention, enabling them to conduct experiments in more ecological settings. While the attentional biases towards melodies with high-pitch and attractive statistics are already known, showing their relative effect is an important step in building precise models of auditory attention, and allows future paradigms to explore more fine-grained effects. Finally, the stream segregation and integration shown with this paradigm could be important for researchers working on music perception. Future work may be necessary to identify the models (Markov chains, deep learning) and setup (data analysis, stimuli, control variables) that do or do not replicate these results.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this study, Wang et al. investigate how herbivorous insects overcome plant receptor-mediated immunity by targeting plant receptor-like proteins. The authors identify two independently evolved salivary effectors, BtRDP in whiteflies and NlSP694 in brown planthoppers, that promote the degradation of plant RLP4 through the ubiquitin-dependent proteasome pathway. NtRLP4 from tobacco and OsRLP4 from rice are shown to confer resistance against herbivores by activating defense signaling, while BtRDP and NlSP694 suppress these defenses by destabilizing RLP4 proteins.
Strengths:
This work highlights a convergent evolutionary strategy in distinct insect lineages and advances our understanding of insect-plant coevolution at the molecular level.
Weaknesses:
(1) I found the naming of BtRDP and NlSP694 somewhat confusing. The authors defined BtRDP as "B. tabaci RLP-degrading protein," whereas NlSP694 appears to have been named after the last three digits of its GenBank accession number (MF278694, presumably). Is there a standard convention for naming newly identified proteins, for example, based on functional motifs or sequence characteristics? As it stands, the inconsistency makes it difficult for readers to clearly distinguish these proteins from those reported in other studies.
(2) Figure 2 and other figures. Transgenic experiments require at least two independent lines, because results from a single line may be confounded by position effects or unintended genomic alterations, and multiple lines provide stronger evidence for reproducibility and reliability.
(3) Figure 3e. Quantitative analysis of NtRLP4 was required. Additionally, since only one band was observed in oeRLP, were any tags included in the construct?
(4) Figure 4a. The RNAi effect appears to be well rescued in Line 1 but poorly in Line 2. Could the authors clarify the reason for this difference?
(5) ROS accumulation is shown for only a single leaf. A quantitative analysis of ROS accumulation across multiple samples would be necessary to support the conclusion. The same applies to Figure 16f.
(6) Figure 4f: NtRLP4 abundance was significantly reduced in oeBtRDP plants but not in oeBtRDP-SP. Although coexpression analysis suggests that BtRDP promotes NtRLP4 degradation in an ubiquitin-dependent manner, the reduced NtRLP4 levels may not result from a direct interaction between BtRDP and NtRLP4. It is possible that BtRDP influences other factors that indirectly affect NtRLP4 abundance. The authors should discuss this possibility.
(7) The statement in lines 335-336 that 'Overexpression of NtRLP4 or NtSOBIR1 enhances insect feeding, while silencing of either gene exerts the opposite effect' is not supported by the results shown in Figures S16-S19. The authors should revise this description to accurately reflect the data.
(8) BtRDP is reported to attach to the salivary sheath. Does the planthopper NlSP694 exhibit a similar secretion localization (e.g., attachment to the salivary sheath)? The authors should supplement this information or discuss the potential implications of any differences in secretion localization between BtRDP and NlSP694 for their respective modes of action.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors aim to investigate the mechanisms of anxiety. The paper focuses on the supramammillary nucleus (SuM) based on a fos screen and recordings showing that footshock and social defeat stress increase activity in this region. Using activity-dependent tagging, they show that reactivation of stress-activated neurons in SuM has an anxiety-like effect, reducing open-arm exploration in the elevated zero task. They then investigate the ventral subiculum as a potential source of anxiety-related information for SuM. They show that ventral subiculum (vSub) inputs to SuM are more strongly activated than dSub when mice explore the open arms of the elevated zero. Finally, they show that DREADD-mediated inhibition of vSub-SuM projections alleviates stress-enhanced anxiety. Overall, the results provide good evidence that SuM contains a stress-activated neuronal population whose later activity increases anxiety-like behavior. It further provides evidence that vSub projects to SuM are activated by stress, and their inhibition alleviates some effects of stress.
Strengths:
Strengths of this paper include the use of convergent methods (e.g., fos plus electrode recordings, footshock, and social defeat) to demonstrate that the SuM is activated by different forms of stress. The activity-dependent tagging experiment shows that footshock-activated SuM neurons are reactivated by social defeat but not by sucrose is also compelling because it provides evidence that SuM neurons are driven by some integrative aspect of stress rather than by a simple sensory stimulus.
Weaknesses:
The strength of some of the evidence is judged to be incomplete. The paper provides good evidence that SuM contains stress-responsive neurons, and the activity of these neurons increases some measure of anxiety-like behavior. However, the evidence that the vSub-SuM projection "encodes anxiety" and that the SuM is a key regulator of anxiety is judged to be incomplete. The claim that SuM generates an "anxiety engram" is also judged to be incompletely supported by the evidence. Namely, what is unclear is whether these cells/regions encode anxiety per se versus modulate behaviors (like exploration) that tend to correlate with anxiety. Since many brain regions respond to footshock and other stressors, the response of SuM to these stimuli is not strong evidence for a role in anxiety. I am not convinced that the identified SuM cells have a specific anxiety function. As the authors mention in the introduction, SuM regulates exploration and theta activity. Since theta potently regulates hippocampal function, there is the concern that SuM manipulations could have broad effects. As shown in Supplementary Figure 2, stimulating stress-responsive cells in SuM potently reduces general locomotor exploration. This raises concerns that the manipulation could have broader effects that go beyond just changes in anxiety-like behavior. Furthermore, the meaning of an "anxiety engram" is unclear. Would this engram encode stress, the sense of a potential threat, or the behavioral response? A more developed analysis of the behavioral correlates of SuM activity and the behavioral effects of SuM manipulations could give insight into these questions.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Zhang et al. investigated how germline tumors influence the development of neighboring wild-type (WT) germline stem cells (GSC) in the Drosophila ovary. They report that germline tumors inhibit the differentiation of neighboring WT GSCs by arresting them in an undifferentiated state, resulting from reduced expression of the differentiation-promoting factor Bam. They find that these tumor cells produce low levels of the niche-associated signaling molecules Dpp and Gbb, which suppress bam expression and consequently inhibit the differentiation of neighboring WT GSCs non-cell-autonomously. Based on these findings, the authors propose that germline tumors mimic the niche to suppress the differentiation of the neighboring stem cells.
Strengths:
This study addresses an important biological question concerning the interaction between germline tumor cells and WT germline stem cells in the Drosophila ovary. If the findings are substantiated, they could provide valuable insights applicable to other stem cell systems.
Weaknesses:
Previous work from Xie's lab demonstrated that bam and bgcn mutant GSCs can outcompete WT GSCs for niche occupancy. Furthermore, a large body of literature has established that the interactions between escort cells (ECs) and GSC daughters are essential for proper and timely germline differentiation (the differentiation niche). Disruption of these interactions leads to arrest of germline cell differentiation in a status with weak BMP signaling activation and low bam expression, a phenotype virtually identical to what is reported here.
Thus, it remains unclear whether the observed phenotype reflects "direct inhibition by tumor cells" or "arrested differentiation due to the loss of the differentiation niche". Because most data were collected at a very late stage (more than 10 days after clonal induction), when tumor cells already dominate the germarium, this question cannot be solved. To distinguish between these two possibilities, the authors could conduct a time-course analysis to examine the onset of the WT GSC-like single-germ-cell (SGC) phenotype and determine whether early-stage tumor clones with a few tumor cells can suppress the differentiation of neighboring WT GSCs with only a few tumor cells present. If tumor cells indeed produce Dpp and Gbb (as proposed here) to inhibit the differentiation of neighboring germline cells, a small cluster or probably even a single tumor cell generated at an early stage might prevent the differentiation of their neighboring germ cells.
The key evidence supporting the claim that tumor cells produce Gpp and Gbb comes from Figures 5 and 6, which suggest that tumor-derived dpp and gbb are required for this inhibition. However, interpretation of these data requires caution.
In Figure 5, the authors use dpp-lacZ to support the claim that dpp is upregulated in tumor cells (Figure 5A and 5B). However, the background expression in somatic cells (ECs and pre-follicular cells) differs noticeably between these panels. In Figure 5A, dpp-lacZ expression in somatic cells in 5A is clearly higher than in 5B, and the expression level in tumor cells appears comparable to that in somatic cells (dpp-lacZ single channel). Similarly, in Figure 5B, dpp-lacZ expression in germline cells is also comparable to that in somatic cells. Providing clear evidence of upregulated dpp and gbb expression in tumor cells (for example, through single-molecular RNA in situ) would be essential.
Most tumor data present in this study were collected from the bam[86] null allele, whereas the data in Figure 6 were derived from a weaker bam[BG] allele. This bam[BG] allele is not molecularly defined and shows some genetic interaction with dpp mutants. As shown in Figure 6E, removal of dpp from homozygous bam[BG] mutant leads to germline differentiation (evidenced by a branched fusome connecting several cystocytes, located at the right side of the white arrowhead). In Figure 6D, fusome is likely present in some GFP-negative bam[BG]/bam[BG] cells. To strengthen their claim that the tumor produces Dpp and Gbb to inhibit WT germline cell differentiation, the authors should repeat these experiments using the bam[86] null allele.
It is well established that the stem niche provides multiple functional supports for maintaining resident stem cells, including physical anchorage and signaling regulation. In Drosophila, several signaling molecules produced by the niche have been identified, each with a distinct function - some promoting stemness, while others regulate differentiation. Expression of Dpp and Gbb alone does not substantiate the claim that these tumor cells have acquired the niche-like property. To support their assertion that these tumors mimic the niche, the authors should provide additional evidence showing that these tumor cells also express other niche-associated markers. Alternatively, they could revise the manuscript title to more accurately reflect their findings.
In the Method section, the authors need to provide details on how dpp-lacZ expression levels were quantified and normalized.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
In this manuscript, the authors explore dynamic chromosomal mobility and transcriptional bursting events in mammalian cells, particularly focusing on ERα-dependent gene activation. The authors investigate how the physical movement of DNA loci changes during different phases of gene transcription (bursting vs. non-bursting, acute vs. chronic stimulation). Using advanced live-cell imaging techniques, including SMT of ERα and dual DNA/RNA visualization, the study reveals a multi-state model of DNA mobility linked to the formation of transcription factor condensates. The authors conclude that differential DNA kinetics serve as a reliable indicator for detecting condensate formation during gene activation, offering new insights into the mechanisms regulating gene expression within the nucleus.
Strengths:
The authors have done substantial work, and a major strength of the manuscript is being able to image both DNA and RNA from the same gene, as well as the TF that acts on that gene. This multi-pronged approach leads to complementary insights into transcription bursting mechanisms.
Weaknesses:
A major weakness of the manuscript is the lack of appropriate controls that support the specificity of the effects observed. The exclusive focus on condensates as the underlying mechanism to explain their data is also a bit limiting.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
CTF18-RFC is an alternative eukaryotic PCNA sliding clamp loader which is thought to specialize in loading PCNA on the leading strand. Eukaryotic clamp loaders (RFC complexes) have an interchangeable large subunit which is responsible for their specialized functions. The authors show that the CTF18 large subunit has several features responsible for its weaker PCNA loading activity, and that the resulting weakened stability of the complex is compensated by a novel beta hairpin backside hook. The authors show this hook is required for the optimal stability and activity of the complex.
Relevance:
The structural findings are important for understanding RFC enzymology and novel ways that the widespread class of AAA ATPases can be adapted to specialized functions. A better understanding of CTF18-RFC function will also provide clarity into aspects of DNA replication, cohesion establishment and the DNA damage response.
Strengths:
The cryo-EM structures are of high quality enabling accurate modelling of the complex and providing a strong basis for analyzing differences and similarities with other RFC complexes.
Weaknesses:
The manuscript would have benefited from a more detailed biochemical analysis using mutagenesis and assays to tease apart the differences with the canonical RFC complex. Analysis of the FRET assay could be improved.
Overall appraisal:
Overall, the work presented here is solid and important. The data is mostly sufficient to support the stated conclusions.
Comments on revisions:
While the authors addressed my previous specific concerns, they have now added a new experiment which raises new concerns.
The FRET clamp loading experiments (Fig. 6) appear to be overfitted so that the fitted values are unlikely to be robust and it is difficult to know what they mean, and this is not explained in this manuscript. Specifically, the contribution of two exponentials is floated in each experiment. By eye, CTF18-RFC looks much slower than RFC1-RFC (as also shown previously in the literature) but the kinetic constants and text suggest it is faster. This is because the contribution of the fast exponential is substantially decreased, and the rate constants then compensate for this. There is a similar change in contribution of the slow and fast rates between WT CTF18 and the variant (where the data curves look the same) and this has been balanced out by a change in the rate constants, which is then interpreted as a defect. I doubt the data are strong enough to confidently fit all these co-dependent parameters, especially for CTF18, where a fast initial phase is not visible. I would recommend either removing this figure or doing a more careful and thorough analysis.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The manuscript by Troyer et al quantitatively measured the membrane localization and diffusion of RNase E, an essential ribonuclease for mRNA turnover as well as tRNA and rRNA processing in bacteria cells. Using single-molecule tracking in live E. coli cells, the authors investigated the impact of membrane targeting sequence (MTS) and the C-terminal domain (CTD) on the membrane localization and diffusion of RNase E under various perturbations. Finally, the authors tried to correlate the membrane localization of RNase E to its function on co- and post-transcriptional mRNA decay using lacZ mRNA as a model.
The major findings of the manuscripts include:
(1) WT RNase E is mostly membrane localized via MTS, confirming previous results. The diffusion of RNase E is increased upon removal of MTS or CTD, and more significantly increased upon removal of both regions.
(2) By tagging RNase E MTS and different lengths of LacY transmembrane domain (LacY2, LacY6 or LacY12) to mEos3.2, the results demonstrate that short LacY transmembrane sequence (LacY2 and LacY6) can increase the diffusion of mEos3.2 on the membrane compared to MTS, further supported by the molecular dynamics simulation. The similar trend was roughly observed in RNase E mutants with MTS switched to LacY transmembrane domains.
(3) The removal of RNase E MTS significantly increases the co-transcriptional degradation of lacZ mRNA, but has minimal effect on the post-transcriptional degradation of lacZ mRNA. Removal of CTD of RNase E overall decrease the mRNA decay rates, suggesting the synergistic effect of CTD on RNase E activity.
Strengths:
(1) The manuscript is clearly written with very detailed methods description and analysis parameters.
(2) The conclusions are mostly supported by the data and analysis.
(3) Some of the main conclusions are interesting and important for understanding the cellular behavior and function of RNase E.
Weaknesses:
The authors have addressed my previous concerns in the revised manuscript.
Comments on revisions:
I have one additional comment. When interpreting the small increase in the diffusion coefficient of RNase E when treating the cell with rifampicin, the authors rule out the possibility that only a small fraction of RNase E interacts with mRNA and suggest that it is more likely the mRNA-RNase E interaction is transient. However, I am wondering about an alternative possibility that RNase E prefers mRNAs with low ribosome density or even untranslated mRNAs?
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors examine the role of the medial frontal cortex of mice in exploiting statistical structure in tasks. They claim that mice are "proactive": they predict upcoming changes, rather than responding in a "model-free" way to environmental changes. Further, they speculate that the estimation of future change (i.e., prediction of upcoming events, based on learning temporal regularities) might be "a main ... function of dorsal medial frontal cortex (dmFC)." Unfortunately, the current manuscript contains flaws such that the evidence supporting these claims is inadequate.
Strengths:
Understanding the neural mechanisms by which we learn about statistical structure in the world is an important goal. The authors developed an interesting task and used model-based techniques to try to understand the mechanisms by which perturbation of dmFC influenced behavior. They demonstrate that lesions and optogenetic silencing of dmFC influence behavior, showing that this region has a causal influence on the task.
Weaknesses:
I was concerned that the main behavioral effects shown in Figure 1F were a statistical artifact. By requiring the Geometric block length to be preceded by a performance-based block, the authors introduce a dependence that can generate the phenomena they describe as anticipation.
To demonstrate this, I simulated their task with an agent that does not have any anticipation of the change point (Reviewer image 1). The agent repeats the previous action with probability `p(repeat)` (similar to the choice kernel in the author's models). If the agent doesn't repeat then the next choice depends on the previous outcome. If the previous choice was rewarded, it stays with `P(WS)` and chooses randomly with `1-P(WS)`. If the previous choice was unrewarded, it switches with `P(LS)` and chooses randomly with `1-P(LS)`.
Review image 1.
An agent with `P(WS)=P(LS)=P(repeat)=0.85` shows the same phenomena as the mice: a difference in performance before the block switch and "earlier" crossing of the midpoint after the switch. https://imgdrop.io/image/aHn6y. The phenomena go away in the simulations when a fixed block length of 20 trials is followed by a Geometric block length.
The authors did not completely rely on the phenomena of Figure 1F for their conclusions. They did a model comparison to provide evidence that animals are anticipating the switch. Unfortunately, the authors did not use state-of-the-art methods in this section of the paper. In particular, they failed to show that under a range of generative parameters for each model class, the model selection process chooses the correct model class (i.e. a confusion matrix). A more minor point, they used BIC instead of a more robust cross-validated metric for model selection. Finally, instead of comparing their "best" anticipating model to their 2nd best model (without anticipation), they compared their best to their 4th best (Supp Fig 3.5). This seems misleading.
Given all of the the above issues, it is hard to critically evaluate the model-based analysis of the effects of lesions/optogenetics.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public Review):
The manuscript presents an intriguing explanation for why grid cell firing fields do {\em not} lie on a lattice whose axes aligned to the walls of a square arena. This observation, by itself, merits the manuscript's dissemination to the journals audience.
The presentation is quirky (but keep the quirkiness!).
But let me recast the problem presented by the authors as one of combinatorics. Given repeating, spatially separated firing fields across cells, one obtains temporal sequences of grid cells firing. Label these cells by integers from $[n]$. Any two cells firing in succession should uniquely identify one of six directions (from the hexagonal lattice) in which the agent is currently moving.
Now, take the symmetric group $\Sigma$ of cyclic permutations on $n$ elements.<br /> We ask whether there are cyclic permutations of $[n]$ such that
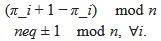
So, for instance, $(4,2,3,1)$ would not be counted as a valid permutation of $(1,2,3,4)$, as $(2,3)$ and $(1,4)$ are adjacent.
Furthermore, given $[n]$, are there two distinct cyclic permutations such that {\em no} adjacencies are preserved when considering any pair of permutations (among the triple of the original ordered sequence and the two permutations)? In other words, if we consider the permutation required to take the first permutation into the second, that permutation should not preserve any adjacencies.
{\bf Key question}: is there any difference between the solution to the combinatorics problem sketched above and the result in the manuscript? Specifically, the text argues that for $n=7$ there is only {\em one} solution.
Ideally, one would strive to obtain a closed-form solution for the number of such permutations as a function of $n$.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
BicD2 is a motor adapter protein that facilitates cellular transport pathways, which are impacted by human disease mutations of BicD2 causing spinal muscular atrophy with lower extremity dominance (SMALED2). The authors provide evidence that some of these mutations result in interactome changes, which may be the underlying cause of the disease. This is supported by proximity biotin ligation screens, immunoprecipitation and cell biology assays. The authors identify several novel BicD2 interactions such as the HOPS complex that participates in the fusion of late endosomes and autophagosomes with lysosomes, which could have important functions. Three BicD2 disease mutants studied had changes in the interactome, which could be an underlying cause for SMALED2. The study extends our understanding of the BicD2 interactome under physiological conditions, as well as of the changes of cellular transport pathways that result in SMALED2. It will be of great interest for the BicD2 and dynein fields.
Strengths:
Extensive interactomes are presented for both WT BicD2 as well as the disease mutants, which will be valuable for the community. The HOPS complex was identified as a novel interactor of BicD2, which is important for fusion of late endosomes and lysosomes, which is of interest, since some of the BicD2 disease mutations result in Golgi-fragmentation phenotypes. The interaction with the HOPS complex is affected by the R747C mutation, which also results in a gain of function interaction with GRAMD1A.
Weaknesses:
The manuscript should be strengthened by further evidence of the BicD2/HOPS complex interaction and the functional implications for spinal muscular atrophy by changes in the interactome through mutations. Which functional implications does the loss of the BicD2/HOPS complex interaction and the gain of function interaction with GRAMD1A have in the context of the R747C mutant?
Major points:
(1) In the biotin proximity ligation assay, a large number of targets were identified, but it is not clear why only the HOPS complex was chosen for further verification. Immunoprecipitation was used for target verification, but due to the very high number of targets identified in the screen, and the fact that the HOPS complex is a membrane protein that could potentially be immunoprecipitated along with lysosomes or dynein, additional experiments to verify the interaction of BicD2 with the HOPS complex (reconstitution of a complex in vitro, GST-pull down of a complex from cell extracts or other approaches) are needed to strengthen the manuscript.<br /> (2) In the biotin proximity ligation assay, a large number of BicD2 interactions were identified that are distinct between the mutant and the WT, but it was not clear why particularly GRAMD1A was chosen as gain of function interaction, and what the functional role of a BicD2/GRAMD1A interaction may be. A Western blot shows a strengthened interaction with the R747C mutant but GRAMD1A also interacts with WT BicD2.<br /> (3) Furthermore, functional implications of changed interactions with HOPS and GRAMD1A in the R747C mutant are unclear. Additional experiments are needed to establish the functional implication of the loss of the BicD2/HOPS interaction in the BicD2/R747C mutant. For the GRAMD1A gain of function interaction, according to the authors a significant amount of the protein localized with BicD2/R747C at the centrosomal region. This changed localization is not very clear from the presented images (no centrosomal or other markers were used, and the changed localization could also be an effect of dynein hyper activation in the mutant). Furthermore, the functional implication of a changed localization of GRAMD1A is unclear from the presented data.
Comments on revisions:
After a major revision, the manuscript is much improved. Additional evidence for the HOPS complex/BicD2 interaction was provided (the interaction was identified in multiple independent screens), and while the authors unfortunately were not able to confirm a direct interaction between BicD2 and the HOPS complex, additional caveats were added in the result section, which clearly state these limitations. The authors also included a very nice discussion of potential physiological roles of the GRAMD1A mislocalization in the disease mutant, which could potentially affect cholesterol transport and homostatis. Limitations of the presented approaches were clearly described as caveats.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors developed an interesting novel paradigm to probe the effects of cerebellar climbing fiber activation on short-term adaptation of somatosensory neocortical activity during repetitive whisker stimulation. Normally, RWS potentiated whisker responses in pyramidal cells and weakly suppressed them in interneurons, lasting for at least 1h. Crusii Optogenetic climbing fiber activation during RWS reduced or inverted these adaptive changes. This effect was generally mimicked or blocked with chemogenetic SST or VIP activation/suppression as predicted based on their "sign" in the circuit.
Strengths:
The central finding about CF modulation of S1 response adaptation is interesting, important, and convincing, and provides a jumping-off point for the field to start to think carefully about cerebellar modulation of neocortical plasticity.
Weaknesses:
The SST and VIP results appeared slightly weaker statistically, but I do not personally think this detracts from the importance of the initial finding (if there are multiple underlying mechanisms, modulating one may reproduce only a fraction of the effect size). I found the suggestion that zona incerta may be responsible for the cerebellar effects on S1 to be a more speculative result (it is not so easy with existing technology to effectively modulate this type of polysynaptic pathway), but this may be an interesting topic for the authors to follow up on in more detail in the future.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The authors have studied a previously published large dataset on the fitness landscape of a 9 base-pair region of the folA gene. The objective of the paper is to understand various aspects of epistasis in this system, which the authors have achieved through detailed and computationally expensive exploration of the landscape. The authors describe epistasis in this system as "fluid", meaning that it depends sensitively on the genetic background, thereby reducing the predictability of evolution at the genetic level. However, the study also finds two robust patterns. The first is the existence of a "pivot point" for a majority of mutations, which is a fixed growth rate at which the effect of mutations switches from beneficial to deleterious (consistent with a previous study on the topic). The second is the observation that the distribution of fitness effects (DFE) of mutations is predicted quite well by the fitness of the genotype, especially for high-fitness genotypes. While the work does not offer a synthesis of the multitude of reported results, the information provided here raises interesting questions for future studies in this field.
Strengths:
A major strength of the study is its detailed and multifaceted approach, which has helped the authors tease out a number of interesting epistatic properties. The study makes a timely contribution by focusing on topical issues like the prevalence of global epistasis, the existence of pivot points, and the dependence of DFE on the background genotype and its fitness. The methodology is presented in a largely transparent manner, which makes it easy to interpret and evaluate the results.
The authors have classified pairwise epistasis into six types and found that the type of epistasis changes depending on background mutations. Switches happen more frequently for mutations at functionally important sites. Interestingly, the authors find that even synonymous mutations in stop codons can alter the epistatic interaction between mutations in other codons. Consistent with these observations of "fluidity", the study reports limited instances of global epistasis (which predicts a simple linear relationship between the size of a mutational effect and the fitness of the genetic background in which it occurs). Overall, the work presents some evidence for the genetic context-dependent nature of epistasis in this system.
Weaknesses:
Despite the wealth of information provided by the study, there are some shortcomings of the paper which must be mentioned.
(1) In the Significance Statement, the authors say that the "fluid" nature of epistasis is a previously unknown property. This is not accurate. What the authors describe as "fluidity" is essentially the prevalence of certain forms of higher-order epistasis (i.e., epistasis beyond pairwise mutational interactions). The existence of higher-order epistasis is a well-known feature of many landscapes. For example, in an early work, (Szendro et. al., J. Stat. Mech., 2013), the presence of a significant degree of higher-order epistasis was reported for a number of empirical fitness landscapes. Likewise, (Weinreich et. al., Curr. Opin. Genet. Dev., 2013) analysed several fitness landscapes and found that higher-order epistatic terms were on average larger than the pairwise term in nearly all cases. They further showed that ignoring higher-order epistasis leads to a significant overestimate of accessible evolutionary paths. The literature on higher-order epistasis has grown substantially since these early works. Any future versions of the present preprint will benefit from a more thorough contextual discussion of the literature on higher-order epistasis.
(2) In the paper, the term 'sign epistasis' is used in a way that is different from its well-established meaning. (Pairwise) sign epistasis, in its standard usage, is said to occur when the effect of a mutation switches from beneficial to deleterious (or vice versa) when a mutation occurs at a different locus. The authors require a stronger condition, namely that the sum of the individual effects of two mutations should have the opposite sign from their joint effect. This is a sufficient condition for sign epistasis, but not a necessary one. The property studied by the authors is important in its own right, but it is not equivalent to sign epistasis.
(3) The authors have looked for global epistasis in all 108 (9x12) mutations, out of which only 16 showed a correlation of R^2 > 0.4. 14 out of these 16 mutations were in the functionally important nucleotide positions. Based on this, the authors conclude that global epistasis is rare in this landscape, and further, that mutations in this landscape can be classified into one of two binary states - those that exhibit global epistasis (a small minority) and those that do not (the majority). I suspect, however, that a biologically significant binary classification based on these data may be premature. Unsurprisingly, mutational effects are stronger at the functional sites as seen in Figure 5 and Figure 2, which means that even if global epistasis is present for all mutations, a statistical signal will be more easily detected for the functionally important sites. Indeed, the authors show that the means of DFEs decrease linearly with background fitness, which hints at the possibility that a weak global epistatic effect may be present (though hard to detect) in the individual mutations. Given the high importance of the phenomenon of global epistasis, it pays to be cautious in interpreting these results.
(4) The study reports that synonymous mutations frequently change the nature of epistasis between mutations in other codons. However, it is unclear whether this should be surprising, because, as the authors have already noted, synonymous mutations can have an impact on cellular functions. The reader may wonder if the synonymous mutations that cause changes in epistatic interactions in a certain background also tend to be non-neutral in that background. Unfortunately, the fitness effect of synonymous mutations has not been reported in the paper.
(5) The authors find that DFEs of high-fitness genotypes tend to depend only on fitness and not on genetic composition. This is an intriguing observation, but unfortunately, the authors do not provide any possible explanation or connect it to theoretical literature. I am reminded of work by (Agarwala and Fisher, Theor. Popul. Biol., 2019) as well as (Reddy and Desai, eLife, 2023) where conditions under which the DFE depends only on the fitness have been derived. Any discussion of possible connections to these works could be a useful addition.
-
-
www.sciencedirect.com www.sciencedirect.com
-
ZFIN: ZDB-ALT-140811-3
DOI: 10.1016/j.celrep.2025.116469
Resource: (ZFIN Cat# ZDB-ALT-140811-3,RRID:ZFIN_ZDB-ALT-140811-3)
Curator: @areedewitt04
SciCrunch record: RRID:ZFIN_ZDB-ALT-140811-3
-
ZFIN: ZDB-ALT-160609-3
DOI: 10.1016/j.celrep.2025.116469
Resource: None
Curator: @areedewitt04
SciCrunch record: RRID:ZFIN_ZDB-ALT-160609-3
-
-
www.thingiverse.com www.thingiverse.com
-
via Aaron Mitchell at https://www.facebook.com/groups/705152958470148/posts/1102482705403836/
Typewriter Typebar Holding Rack by grifter7 - Thingiverse<br /> by [[grifter7]] on [[Thingiverse.com]]<br /> accessed on 2025-10-22T10:26:22
Tool: Typebar Sorting Rack
-