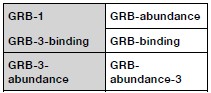
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
This study compares four models - VALOR (dynamic visual-text alignment), CLIP (static visual-text alignment), AlexNet (vision-only), and WordNet (text-only) - in their ability to predict human brain responses using voxel-wise encoding modeling. The results show that VALOR not only achieves the highest accuracy in predicting neural responses but also generalizes more effectively to novel datasets. In addition, VALOR captures meaningful semantic dimensions across the cortical surface and demonstrates impressive predictive power for brain responses elicited by future events.
Strengths:
The study leverages a multimodal machine learning model to investigate how the human brain aligns visual and textual information. Overall, the manuscript is logically organized, clearly written, and easy to follow. The results well support the main conclusions of the paper.
(1) My primary concern is that the performance difference between VALOR and CLIP is not sufficiently explained. Both models are trained using contrastive learning on visual and textual inputs, yet CLIP performs significantly worse. The authors suggest that this may be due to VALOR being trained on dynamic movie data while CLIP is trained on static images. However, this explanation remains speculative. More in-depth discussion is needed on the architectural and inductive biases of the two models, and how these may contribute to their differences in modeling brain responses.
Thank you for this thoughtful comment. We agree that attributing VALOR’s advantage over CLIP solely to ‘dynamic (video) versus static (image) pretraining’ would be incomplete, and that the architectural and inductive biases of the two models are central to understanding the observed performance gap.
Both VALOR and CLIP use contrastive learning to align visual and textual representations, but they differ in several key inductive biases that are particularly relevant for modeling brain responses during continuous movie viewing. First, VALOR is trained to align temporally extended video segments with text, introducing an explicit temporal integration window that aggregates information across consecutive frames. This encourages representations that maintain context, stabilize semantics across time, and encode event-level structure. Second, VALOR’s alignment operates at the level of multi-second narrative units, rather than isolated visual snapshots, biasing the model toward representations that are sensitive to unfolding events and cross-frame consistency.
In contrast, CLIP processes frames independently and aligns single static images with text. As a result, it lacks an intrinsic mechanism for temporal binding, context accumulation, or event-level representation. While CLIP can capture rich visual–semantic associations at the image level, it is less well suited to represent higher-order temporal structure, which is known to strongly drive responses in association cortex during naturalistic narrative perception.
We therefore interpret VALOR’s superior encoding performance as reflecting not only exposure to dynamic audiovisual data, but also inductive biases—temporal integration and event-level alignment—that more closely match how the brain integrates information over time during movie watching. We have revised the Discussion (p. 16) to articulate these architectural and representational differences explicitly, rather than attributing the effect solely to training data modality.
(On page 16) “Additionally, VALOR exceeds the performance of CLIP, a leading static multimodal model, as its training objective aligns multi-second video–text units, enforcing a temporal integration window and event-level semantics that maintain cross-frame consistency and narrative context, whereas CLIP’s image-level alignment provides no intrinsic mechanism for such temporal continuity.”
(2) The methods section lacks clarity regarding which layers of VALOR and CLIP were used to extract features for voxel-wise encoding modeling. A more detailed methodological description is necessary to ensure reproducibility and interpretability. Furthermore, discussion of the inductive biases inherent in these models-and their implications for brain alignment - is crucial.
Thank you for this comment. We agree that reproducibility and interpretability require precise specification of which model representations were used for voxel-wise encoding, as well as clearer discussion of the inductive biases inherent in these models and their implications for brain alignment.
In the revised Methods, we now explicitly specify the feature sources for both models. For CLIP (ViT-B/32), we use the final pooled image embedding after projection into the shared image–text space, extracted frame-by-frame; one representative frame is sampled per TR, and its projected embedding serves as the regressor. For VALOR, we use the final joint video–text projection head, yielding a 512-dimensional embedding computed at the segment/TR level that integrates information across consecutive frames and aligns each multi-second video segment with its associated text. These procedures are now described step-by-step in the Methods (p. 21).
In addition, we expanded the Discussion (p. 16) to explicitly articulate the models’ inductive biases and their relevance for brain alignment. In particular, we contrast CLIP’s image-level, framewise alignment—which lacks intrinsic temporal integration—with VALOR’s event-level, temporally extended video–text alignment, which biases representations toward context maintenance and narrative continuity. This distinction helps explain why the two models differ in their ability to predict neural responses during continuous movie viewing.
(Methods, On page 21)
“(1) Video–text alignment features (VALOR): To extract video-based multimodal features, we used VALOR (VALOR-large checkpoint), an open-source pretrained video–text alignment model24. VALOR combines visual encoders (CLIP and Video Swin Transformer) for extracting visual features and a text encoder (BERT) for extracting textual features 23,51,52. These representations are aligned in a shared embedding space through contrastive learning. We segmented each movie at the TR level and, for each segment, extracted VALOR’s projected video–text embedding from the final projection head of the alignment module to obtain a 512-dimensional feature vector. These embeddings were then time-aligned to the corresponding BOLD responses.
(2) CLIP features: To compare with static image-based multimodal models, we utilized CLIP (ViT-B/32), which aligns visual and textual representations through contrastive learning but processes individual frames independently without capturing temporal context. One video frame was sampled per TR, and the pooled image embedding after CLIP’s projection into the shared image–text space was extracted to obtain a 512-dimensional feature vector. These TR-aligned vectors were used directly as regressors in the voxel-wise encoding models.”
(Discussion, On page 16)
“Additionally, VALOR exceeds the performance of CLIP, a leading static multimodal model, as its training objective aligns multi-second video–text units, enforcing a temporal integration window and event-level semantics that maintain cross-frame consistency and narrative context, whereas CLIP’s image-level alignment provides no intrinsic mechanism for such temporal continuity. More broadly, this difference reflects distinct inductive biases in how the two models represent visual–linguistic information. CLIP is optimized for framewise image–text correspondence, encouraging representations that emphasize instantaneous visual semantics but remain agnostic to temporal structure. In contrast, VALOR is explicitly biased toward aggregating information over multiple consecutive frames and aligning representations at the level of temporally extended events. These inductive biases favor context maintenance, semantic stabilization, and narrative coherence over time, which are known to be critical for driving responses in association cortex during continuous movie perception.”
(3) A broader question remains insufficiently addressed: what is the purpose of visual-text alignment in the human brain? One hypothesis is that it supports the formation of abstract semantic representations that rely on no specific input modality. While VALOR performs well in voxel-wise encoding, it is unclear whether this necessarily indicates the emergence of such abstract semantics. The authors are encouraged to discuss how the computational architecture of VALOR may reflect this alignment mechanism and what implications it has for understanding brain function.
Thank you for this important conceptual question. We agree that improved voxel-wise encoding performance does not, by itself, imply the emergence of fully amodal or modality-independent semantic representations in the brain. In the revision, we therefore avoid framing our findings as evidence for abstract amodal semantics and instead clarify a more constrained interpretation.
Specifically, we suggest that visual–text alignment may support the stabilization and coordination of scene-level meaning across modalities and over time, rather than the formation of modality-free semantic codes. From this perspective, VALOR’s advantage reflects inductive biases that promote (i) integration of visual information over multi-second windows and (ii) alignment of temporally extended visual events with linguistic descriptions, yielding representations that are more temporally stable, context-sensitive, and constrained by language.
We therefore interpret VALOR’s superior encoding performance as identifying cortical regions whose responses are better captured by temporally stabilized, cross-modal representations, rather than as evidence that these regions encode fully abstract semantics independent of input modality. We have expanded the Discussion (p. 16) to articulate this interpretation and to clarify the implications of video–text alignment for understanding how the brain integrates perception and language during naturalistic cognition.
(On page 16) “Together, the relative gains over AlexNet (purely visual), WordNet (manual semantic annotation), and CLIP (static image–text alignment) indicate cortical systems whose responses are best captured by multi-second, multimodal integration, and highlight regions that accumulate and stabilize narrative context over time. At the same time, these findings do not imply that visual–text alignment in the brain gives rise to fully amodal, modality-independent semantic representations. Instead, we suggest that alignment between visual and linguistic signals may serve to stabilize and coordinate scene-level meaning across modalities and over time. From this perspective, VALOR’s architecture—by integrating visual information over multi-second windows and aligning temporally extended video segments with language—provides a computational proxy for how the brain may use linguistic constraints to organize, disambiguate, and maintain coherent representations of unfolding events. The observed encoding gains therefore highlight regions engaged in temporally stabilized, cross-modal integration during naturalistic perception, rather than providing evidence for abstract semantic codes divorced from sensory input.”
(4) The current methods section does not provide enough details about the network architectures, parameter settings, or whether pretrained models were used. If so, please provide links to the pretrained models to facilitate reproducible science.
We appreciate this comment and agree that our original description of model sources and implementation details was not sufficiently explicit. These details are essential for both reproducibility and interpretability. We have now made these specifications explicit in the revised Methods.
In particular, we now state for each model:
VALOR. We use the publicly released pretrained VALOR-large checkpoint. For each movie segment, we extract the joint video–text projection head output (512-D) that encodes the aligned segment-level audiovisual semantics. We report the checkpoint source, the segment duration (in frames/seconds), and how these segment-level embeddings are temporally aligned to TRs for voxel-wise encoding.
CLIP (ViT-B/32). We use the standard pretrained CLIP weights. For each video frame, we extract the final pooled image representation after projection into CLIP’s shared image–text embedding space (512-D). We also clarify that one representative frame is sampled and aligned to each TR, and that these projected embeddings are used as regressors in the encoding model.
AlexNet. We use the ImageNet-pretrained AlexNet. We take activations from conv5, and then apply PCA to reduce them to 512 dimensions before mapping them to the fMRI time series.
For each model, the revised Methods now specify: the pretrained source/checkpoint, the layer or head from which features were taken, output dimensionality, any preprocessing or dimensionality reduction, and the temporal alignment procedure used to generate TR-level regressors. These revisions appear in the updated Methods (page 21).
(On page 21) “(1) Video–text alignment features (VALOR): To extract video-based multimodal features, we used VALOR (VALOR-large checkpoint), an open-source pretrained video–text alignment model24. VALOR combines visual encoders (CLIP and Video Swin Transformer) for extracting visual features and a text encoder (BERT) for extracting textual features 23,51,52. These representations are aligned in a shared embedding space through contrastive learning. We segmented each movie at the TR level and, for each segment, extracted VALOR’s projected video–text embedding from the final projection head of the alignment module to obtain a 512-dimensional feature vector. These embeddings were then time-aligned to the corresponding BOLD responses.
(2) P features: To compare with static image-based multimodal models, we utilized CLIP (ViT-B/32), which aligns visual and textual representations through contrastive learning but processes individual frames independently without capturing temporal context. One video frame was sampled per TR, and the pooled image embedding after CLIP’s projection into the shared image–text space was extracted to obtain a 512-dimensional feature vector. These TR-aligned vectors were used directly as regressors in the voxel-wise encoding models.
(3) AlexNet features: Visual features were extracted by sampling frames at the TR level and processing them with AlexNet, an eight-layer convolutional neural network comprising five convolutional layers followed by three fully connected layers. Features from all five convolutional layers were evaluated in preliminary analyses; the fifth convolutional layer showed the best performance and was used in subsequent analyses. Intra-image z-score normalization was applied to reduce amplitude effects. Principal component analysis (PCA) was used to reduce dimensionality, retaining the top 512 components to match the dimensionality of multimodal feature spaces. This pipeline was implemented using the DNNBrain toolkit 53.
(4) WordNet features: Semantic features were obtained from publicly available WordNet annotations provided with the HCP dataset (7T_movie_resources/WordNetFeatures.hdf5), following the procedure of Huth et al. (2012). Each second of the movie clips was manually annotated with WordNet categories according to predefined guidelines: (a) identifying clear objects and actions in the scene; (b) labeling categories that dominated for more than half of the segment duration; and (c) using specific category labels rather than general ones. A semantic feature matrix was constructed with rows corresponding to time points and columns to semantic categories, with category presence coded as binary values. More specific categories from the WordNet hierarchy were added to each labeled category, yielding a total of 859 semantic features. These features were used directly as regressors. We also evaluated a PCA-reduced 512-dimensional variant (fit within each training fold to avoid leakage); because this version performed slightly worse, we report results from the full 859-dimensional representation in the main text. For the generalization analysis in Study 2, annotations for the SFM dataset were aligned to the same WordNet category space to ensure consistency.”
Reviewer #2 (Public review):
Fu and colleagues have shown that VALOR, a model of multimodal and dynamic stimulus features, better predicts brain responses compared to unimodal or static models such as AlexNet, WordNet, or CLIP. The authors demonstrated the robustness of their findings by generalizing encoding results to an external dataset. They demonstrated the models' practical benefit by showing that semantic mappings were comparable to another model that required labor-intensive manual annotation. Finally, the authors showed that the model reveals predictive coding mechanisms of the brain, which held a meaningful relationship with individuals' fluid intelligence measures.
Strengths:
Recent advances in neural network models that extract visual, linguistic, and semantic features from real-world stimuli have enabled neuroscientists to build encoding models that predict brain responses from these features. Higher prediction accuracy indicates greater explained variance in neural activity, and therefore a better model of brain function. Commonly used models include AlexNet for visual features, WordNet for audio-semantic features, and CLIP for visuo-semantic features; these served as comparison models in the study. Building on this line of work, the authors developed an encoding model using VALOR, which captures the multimodal and dynamic nature of real-world stimuli. VALOR outperformed the comparison models in predicting brain responses. It also recapitulated known semantic mappings and revealed evidence of predictive processing in the brain. These findings support VALOR as a strong candidate model of brain function.
(1) The authors argue that this modeling contributes to a better understanding of how the brain works. However, upon reading, I am less convinced about how VALOR's superior performance over other models tells us more about the brain. VALOR is a better model of the audiovisual stimulus because it processes multimodal and dynamic stimuli compared to other unimodal or static models. If the model better captures real-world stimuli, then I almost feel that it has to better capture brain responses, assuming that the brain is a system that is optimized to process multimodal and dynamic inputs from the real world. The authors could strengthen the manuscript if the significance of their encoding model findings were better explained.
We thank the reviewer for this thoughtful comment and agree with the premise that a model preserving multimodal and temporal structure might a priori be expected to better predict brain responses to naturalistic stimuli. Our intent is not to claim that higher accuracy alone explains brain function, but rather that where and how VALOR improves prediction provides diagnostic insight into cortical processing. We have revised the Discussion to make this distinction explicit.
Specifically, we clarify three ways in which VALOR’s gains are scientifically informative rather than merely unsurprising:
(1) Anatomical specificity of improvement. VALOR’s advantage is not uniform across the cortex; gains are largest in regions implicated in multi-second, cross-modal integration. This spatial pattern constrains where the brain accumulates information over time and stabilizes visual representations using linguistic context.
(2) Model as a computational probe. Beyond prediction accuracy, VALOR’s feature space recovers large-scale semantic organization without manual annotation and enables targeted tests of predictive processing. Features reflecting upcoming content selectively improve fits in specific regions, consistent with anticipatory coding during continuous narrative perception.
(3) Link to individual differences. Individuals whose neural responses are better captured by anticipatory features show higher fluid intelligence, suggesting that VALOR indexes meaningful variability in forward-looking representations rather than merely tracking stimulus complexity.
Accordingly, we have revised the Discussion (p. 16) to frame VALOR as a tool for mapping cortical integration profiles, probing semantic and predictive structure, and linking representational dynamics to cognition, rather than asserting that higher encoding accuracy alone explains brain function.
(On page 16) “Together, the relative gains over AlexNet (purely visual), WordNet (manual semantic annotation), and CLIP (static image–text alignment) indicate cortical systems whose responses are best captured by multi-second, multimodal integration, and highlight regions that accumulate and stabilize narrative context over time.”
(2) In Study 3, the authors show high alignment between WordNet and VALOR feature PCs. Upon reading the method together with Figure 3, I suspect that the alignment almost has to be high, given that the authors projected VALOR features to the Huth et al.'s PC space. Could the authors conduct non-parametric permutation tests, such as shuffling the VALOR features prior to mapping onto Huth et al.'s PC space, and then calculating the Jaccard scores? I imagine that the null distribution would be positively shifted. Still, I would be convinced if the alignment is higher than this shifted null distribution for each PC. If my understanding of this is incorrect, I suggest editing the relevant Method section (line 508) because this analysis was not easy to understand.
Thank you for this helpful comment and for pointing out a potential source of confusion. We apologize that the original Methods description was not sufficiently clear. Importantly, VALOR features were never projected into the Huth et al. PC space, and no optimization or rotation toward the WordNet basis occurred at any stage.
The analysis proceeded as follows:
(1) VALOR PCs. We first fit voxel-wise encoding models using VALOR features on the Huth et al. dataset. We then applied PCA to the resulting cortical weight maps, yielding spatial components (‘VALOR PCs’) that summarize shared patterns of VALOR feature weights across the cortex.
(2) WordNet PCs. We used the semantic principal components reported by Huth et al. (2012) directly as published, with no refitting, projection, or modification using VALOR.
(3) Correspondence analysis. Only after obtaining these two independent sets of cortical maps did we threshold each to their top-loading vertices and compute Jaccard overlap between VALOR PCs and WordNet PCs.
Although a permutation that shuffles VALOR features prior to projection addresses a scenario that does not apply here, we agree that the Methods description should more clearly convey the independence of the two decompositions. We have therefore revised the Methods (p. 24) to describe the procedure step-by-step and explicitly state that no projection, refitting, or optimization toward the WordNet basis was performed.
(On page 24) “We first fit voxel-wise encoding models using VALOR features for each of the five participants in the Huth et al. dataset. For each participant, this yielded a weight map linking each VALOR feature to each voxel. We then stacked these weight maps across participants to form a single voxel-by-feature weight matrix and applied principal component analysis (PCA). The top four principal components from this analysis (“VALOR PCs”) captured shared spatial patterns of VALOR feature weights across cortex. To interpret these components, we projected VALOR feature vectors from >20,000 video segments in the VALOR training set onto each VALOR PC, which revealed dominant semantic axes (e.g., mobility, sociality, civilization). For comparison, we used the semantic principal components reported by Huth et al. (2012) from their WordNet-based encoding model; these “WordNet PCs” were taken directly from the published work and were not refit or reweighted using VALOR.”
(3) In Study 4, the authors show that individuals whose superior parietal gyrus (SPG) exhibited high prediction distance had high fluid cognitive scores (Figure 4C). I had a hard time believing that this was a hypothesis-driven analysis. The authors motivate the analysis that "SPG and PCu have been strongly linked to fluid intelligence (line 304)". Did the authors conduct two analyses only-SPG-fluid intelligence and PCu-fluid intelligence-without relating other brain regions to other individual differences measures? Even if so, the authors should have reported the same r-value and p-value for PCu-fluid intelligence. If SPG-fluid intelligence indeed holds specificity in terms of statistical significance compared to all possible scenarios that were tested, is this rationally an expected result, and could the authors explain the specificity? Also, the authors should explain why they considered fluid intelligence to be the proxy of one's ability to anticipate upcoming scenes during movie watching. I would have understood the rationale better if the authors had at least aggregated predictive scores for all brain regions that held significance into one summary statistic and found a significant correlation with the fluid intelligence measure.
We thank the reviewer for this careful and constructive comment and agree that greater transparency about analytic intent, specificity, and rationale is needed. We have revised the manuscript accordingly.
(1) Analytic scope and a priori restriction. The analysis in Fig. 4C was hypothesis-driven and restricted a priori to two regions — superior parietal gyrus (SPG) and precuneus (PCu) — based on convergent evidence linking frontoparietal and medial parietal systems to fluid reasoning, relational integration, and domain-general cognitive control. Importantly, we did not conduct a whole-brain search across regions or behaviors to identify the strongest correlation post hoc.
(2) Specificity and reporting. In response to the reviewer’s request, we now report the full results for both hypothesized regions. Prediction horizon in SPG showed a statistically reliable association with fluid intelligence, whereas PCu showed a positive but weaker trend that did not survive correction. Reporting both results makes the regional specificity explicit rather than implicit.
(3) Why SPG over PCu? Although both regions are implicated in fluid cognition, SPG has been more consistently linked to active maintenance and manipulation of relational structure and top-down attentional control, whereas PCu is more often associated with internally oriented and mnemonic processes. We therefore interpret the stronger SPG association as consistent with a role for sustained, externally driven predictive processing during continuous perception, rather than as evidence of exclusivity.
(4) Why fluid intelligence? We do not equate fluid intelligence with “anticipation” per se. Rather, we used gF as an a priori proxy for domain-general capacities — maintaining and updating relational context over multi-second windows, integrating multiple constraints, and exerting flexible control — that are plausibly recruited when anticipating upcoming events during naturalistic narratives. The reported relationship is associative and hypothesis-consistent, not causal.
(5) Why not aggregate across regions? We agree that aggregation could reveal more global relationships; however, our goal in this analysis was to test whether predictive timescales in theoretically motivated control regions relate to individual differences, rather than to maximize correlation by pooling heterogeneous regions. We now clarify this rationale in the Results.
These clarifications and additional statistics have been incorporated in the revised Results section (p. 14).
(On page 14) “Finally, we examined whether prediction horizons were linked to individual differences in cognition. We focused on fluid intelligence (gF) because gF is widely taken to index domain-general capacities such as maintaining and updating relational context over several seconds, integrating multiple constraints, and exerting flexible top-down control — functions that should support anticipating what will happen next in a continuous narrative. We targeted two parietal regions, the SPG and the PCu, which have both been repeatedly linked to gF and high-level cognitive control in the individual-differences literature 36,37. For each participant, we correlated fluid cognition scores with that participant’s average prediction horizon in each region. As shown in Fig. 4c, individuals with longer prediction horizons in SPG showed higher fluid cognition scores (SPG: r = 0.172, FDR-corrected p = 0.047). PCu showed a similar positive trend (PCu: r = 0.111, FDR-corrected p = 0.146) but did not reach significance. These associations suggest that the ability to sustain a longer predictive timescale during naturalistic perception co-varies with broader fluid cognitive capacity. No additional brain regions or behavioral measures were examined in this analysis.”
Reviewer #3 (Public review):
In this work, the authors aim to improve neural encoding models for naturalistic video stimuli by integrating temporally aligned multimodal features derived from a deep learning model (VALOR) to predict fMRI responses during movie viewing.
Strengths:
The major strength of the study lies in its systematic comparison across unimodal and multimodal models using large-scale, high-resolution fMRI datasets. The VALOR model demonstrates improved predictive accuracy and cross-dataset generalization. The model also reveals inherent semantic dimensions of cortical organization and can be used to evaluate the integration timescale of predictive coding.
This study demonstrates the utility of modern multimodal pretrained models for improving brain encoding in naturalistic contexts. While not conceptually novel, the application is technically sound, and the data and modeling pipeline may serve as a valuable benchmark for future studies.
(1) Lines 95-96: The authors claim that "cortical areas share a common space," citing references [22-24]. However, these references primarily support the notion that different modalities or representations can be aligned in a common embedding space from a modeling perspective, rather than providing direct evidence that cortical areas themselves are aligned in a shared neural representational space.
We thank the reviewer for this important clarification. We agree that the cited works do not provide direct evidence that cortical areas themselves are aligned in a single neural representational space. Rather, they demonstrate that representations derived from different modalities can be mapped into a shared embedding space from a modeling and computational perspective.
We have therefore revised the text to avoid overstatement and to more precisely reflect what these studies support. In the revised manuscript (p. 4), we now frame the claim in terms of a shared representational framework or feature space used for modeling, rather than implying that cortical areas themselves intrinsically share a unified neural space. This clarification aligns the conceptual claim with the scope of the cited literature.
(On page 4) “As a result, researchers are turning to multimodal deep learning, which learns from visual, linguistic, and auditory streams to model complex brain functions. This trend is supported by neuroscience evidence that cortical responses across regions can be jointly modeled within a common representational space.”
(2) The authors discuss semantic annotation as if it is still a critical component of encoding models. However, recent advances in AI-based encoding methods rely on features derived from large-scale pretrained models (e.g., CLIP, GPT), which automatically capture semantic structure without requiring explicit annotation. While the manuscript does not systematically address this transition, it is important to clarify that the use of such pretrained models is now standard in the field and should not be positioned as an innovation of the present work. Additionally, the citation of Huth et al. (2012, Neuron) to justify the use of WordNet-based annotation omits the important methodological shift in Huth et al. (2016, Nature), which moved away from manual semantic labeling altogether. Since the 2012 dataset is used primarily to enable comparison in study 3, the emphasis should not be placed on reiterating the disadvantages of semantic annotation, which have already been addressed in prior work. Instead, the manuscript's strength lies in its direct comparison between data-driven feature representations and semantic annotation based on WordNet categories. The authors should place greater emphasis on analyzing and discussing the differences revealed by these two approaches, rather than focusing mainly on the general advantage of automated semantic mapping.
Thank you for this thoughtful and constructive comment. We agree with the reviewer that the field has largely transitioned away from manual semantic annotation toward features derived from large-scale pretrained models (e.g., CLIP, GPT-style architectures), and that this shift is now standard rather than a novelty of the present work.
We have revised the manuscript to clarify this positioning. Our goal is not to claim automated semantic extraction as an innovation, but rather to demonstrate how a multimodal, temporally informed video–text model can be used as a direct feature space for voxel-wise encoding of naturalistic movie fMRI data. VALOR is used as a representative example of this broader class of pretrained models, and our emphasis is on the general modeling approach rather than on promoting a specific architecture.
We also agree that our original discussion underemphasized the important methodological shift introduced in Huth et al. (2016, Nature), which moved away from manual semantic labeling in the context of continuous spoken narratives. We now explicitly acknowledge this work and clarify that our use of WordNet-based annotations from Huth et al. (2012) serves a different purpose: it provides an interpretable, historically grounded benchmark for comparison in Study 3, rather than a claim that semantic annotation remains necessary or state-of-the-art.
In response to the reviewer’s suggestion, we have revised the Results (p.10) and Discussion (p.18) to place greater emphasis on what is revealed by directly comparing data-driven multimodal features with category-based semantic annotation under matched conditions. Specifically, we focus on how these two approaches converge at the level of large-scale semantic organization while differing in their flexibility, temporal resolution, and dependence on human-defined categories. These revisions better reflect the current state of the field and sharpen the manuscript’s central contribution as a principled comparison between modeling approaches, rather than a general argument for automated semantic mapping.
(On page 10) “Study 3: Comparing data-driven multimodal representations with category-based semantic annotation
A central question in naturalistic encoding is how data-driven feature representations derived from pretrained models relate to more interpretable, category-based semantic annotations that have historically been used to study cortical semantic organization. Although recent work has shown that pretrained language and vision–language models can capture semantic structure without explicit annotation, category-based approaches such as WordNet remain valuable as interpretable reference frameworks. Here, we leverage the WordNet-based semantic components reported by Huth et al. (2012) 5 not as a state-of-the-art alternative, but as a historically grounded benchmark, allowing a controlled comparison between data-driven multimodal representations and manually defined semantic categories under matched naturalistic movie stimuli.”
(On page 18) “Study 3 demonstrates the utility of video–text alignment models for probing higher-order semantic representations during naturalistic perception. Our comparison between VALOR-derived representations and WordNet-based semantic components highlights an important distinction between data-driven and category-based approaches to modeling meaning in the brain. While multimodal pretrained models offer flexible, high-dimensional representations that capture semantic structure without explicit annotation, category-based frameworks provide interpretability and theoretical anchoring 4,48. Using WordNet-based labeling from prior work as an interpretable reference point, we show that VALOR automatically extracts semantic dimensions—including mobility, sociality, and civilization—that closely mirror those identified using manual semantic categories (Fig. 3). The observed alignment between VALOR PCs and WordNet semantic components suggests that large-scale semantic organization emerges consistently across these approaches, even though they differ in how semantic structure is defined and learned. This convergence supports the use of pretrained multimodal models as practical encoding tools for naturalistic stimuli, while also underscoring the continued value of interpretable semantic benchmarks for understanding which aspects of meaning are represented across cortex. We do not argue that semantic annotation is required for modern encoding models; rather, WordNet-based features serve here as a historically grounded and interpretable reference for contextualizing data-driven multimodal representations.”
(3) The authors use subject-specific encoding models trained on the HCP dataset to predict group-level mean responses in an independent in-house dataset. While this analysis is framed as testing model generalization, it is important to clarify that it is not assessing traditional out-of-distribution (OOD) generalization, where the same subject is tested on novel stimuli, but rather evaluating which encoding model's feature space contains more stimulus-specific and cross-subject-consistent information that can transfer across datasets.
We thank the reviewer for this helpful clarification and agree that the type of generalization tested here should be described more precisely. Our analysis does not assess classical within-subject out-of-distribution (OOD) generalization, in which the same individual is tested on novel stimuli.
Instead, for each HCP participant we train a subject-specific encoding model and transfer it to predict group-mean responses in an independent in-house dataset collected at a different site, with different participants, different movies, and different acquisition conditions. This design evaluates which encoding model’s feature space contains stimulus-locked representations that are consistent across individuals and robust to changes in dataset and experimental context, rather than within-subject stimulus novelty per se.
We have revised the Results (p. 10) and Discussion section (p. 17) to explicitly describe this analysis as a test of cross-subject and cross-dataset transferability of stimulus representations, and to clarify the distinction from traditional OOD generalization.
(On Page 10) “Although this analysis is not a classical within-subject out-of-distribution generalization test, it evaluates the extent to which different feature spaces capture stimulus-locked representations that are consistent across subjects and transferable across datasets, stimuli, and acquisition environments.”
(On Page 17) “By contrast, VALOR exhibited stronger generalization in a cross-cohort, cross-stimulus, and cross-site transfer evaluation.”
(4) Within this setup, the finding that VALOR outperforms CLIP, AlexNet, and WordNet is somewhat expected. VALOR encodes rich spatiotemporal information from videos, making it more aligned with movie-based neural responses. CLIP and AlexNet are static image-based models and thus lack temporal context, while WordNet only provides coarse categorical labels with no stimulus-specific detail. Therefore, the results primarily reflect the advantage of temporally-aware features in capturing shared neural dynamics, rather than revealing surprising model generalization. A direct comparison to pure video-based models, such as Video Swin Transformers or other more recent video models, would help strengthen the argument.
We thank the reviewer for this baseline-focused comment and agree that, in naturalistic movie paradigms, a temporally structured audiovisual model would be expected to outperform static or unimodal feature spaces. Our intent in this comparison is therefore not to claim a surprising advantage, but to isolate which inductive biases matter for cross-dataset transfer of movie-evoked neural responses.
The baseline models were chosen deliberately to span feature spaces that are widely used and interpretable in cognitive neuroscience: AlexNet (vision-only, frame-based), WordNet (human-defined semantic categories without learned visual features), and CLIP (static image–text alignment without temporal context). Comparing VALOR against these established baselines under matched preprocessing, TR alignment, and dimensionality control allows us to attribute performance differences specifically to temporal integration and audiovisual alignment, rather than to generic model capacity.
We agree that a direct comparison with purely visual spatiotemporal encoders (e.g., Video Swin or TimeSformer-style models) would further dissociate the contribution of temporal visual processing from cross-modal video–text alignment. We now explicitly note this as an important direction for future work and frame VALOR as one representative of a broader class of multimodal video models, rather than as a uniquely optimal solution (Discussion, p. 16).
(On page 16) “Second, we did not directly compare VALOR to state-of-the-art video-only spatiotemporal models (e.g., Video Swin Transformer, VideoMAE, and related architectures) that are designed to capture temporal visual structure without language grounding; such comparisons will be important for isolating the specific contributions of temporal visual processing versus cross-modal video–text alignment in naturalistic neural responses.”
(5) Moreover, while WordNet-based encoding models perform reasonably well within-subject in the HCP dataset, their generalization to group-level responses in the Short Fun Movies (SFM) dataset is markedly poorer. This could indicate that these models capture a considerable amount of subject-specific variance, which fails to translate to consistent group-level activity. This observation highlights the importance of distinguishing between encoding models that capture stimulus-driven representations and those that overfit to individual heterogeneities.
Thank you for this thoughtful observation. We agree with the reviewer’s interpretation. In our analyses, WordNet-based models perform reasonably well when fit and evaluated within individual HCP participants, but their performance degrades substantially when transferred to predict group-averaged responses in the independent SFM dataset. This dissociation suggests that, while WordNet annotations capture meaningful variance at the individual level, a larger fraction of that variance may be subject-specific or idiosyncratic, and therefore does not translate into consistent, stimulus-locked responses at the group level.
One motivation for our cross-dataset, cross-subject evaluation is precisely to distinguish encoding models that primarily capture shared stimulus-driven structure from those whose apparent performance depends more strongly on individual heterogeneity. In this context, the reduced transferability of WordNet-based models highlights a potential limitation of category-based semantic features for capturing population-consistent neural dynamics during naturalistic viewing.
We note that this effect likely reflects multiple factors rather than a single failure mode, including differences in annotation schemes, labeling granularity, and semantic coverage across datasets. By contrast, video–text models provide time-aligned linguistic features directly from the stimulus itself, reducing reliance on dataset-specific human annotation and exhibiting stronger transfer across cohorts. We have clarified this interpretation in the revised Discussion (p. 17).
(Page 17) “Together, these findings underscore the importance of distinguishing encoding models that primarily capture shared, stimulus-driven neural structure from those whose performance relies more heavily on subject-specific heterogeneity, particularly when evaluating generalization across participants and datasets.”
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
(1) In the Methods section, please clarify which specific layer of VALOR the 512-dimensional feature vector was extracted from.
Thank you for this suggestion. We have revised the Methods to state explicitly that the 512-dimensional feature vector is extracted from VALOR’s joint video–text projection head, i.e., the final projection layer of the contrastive alignment module that maps video and text representations into a shared embedding space. We also clarify that these 512-D embeddings are computed at the segment/TR level and then time-aligned to the BOLD signal (Methods, p. 21).
(On page 21) “We segmented each movie at the TR level and, for each segment, extracted VALOR’s projected video–text embedding from the final projection head of the alignment module to obtain a 512-dimensional feature vector. These embeddings were then time-aligned to the corresponding BOLD responses.”
(2) It would be helpful to include more detailed descriptions of the network architectures and parameters for all models used.
Thank you for the suggestion. We have revised the Methods to include model-specific subsections for all feature spaces used (VALOR, CLIP, AlexNet, and WordNet). For each model, we now explicitly report (i) the backbone architecture and training objective, (ii) the exact feature source (layer or projection head) and output dimensionality, and (iii) how features were temporally aligned to the BOLD signal. All models were used with their publicly released pretrained parameters, without additional fine-tuning. These additions are intended to improve transparency and reproducibility (Methods, p. 21).
(On page 21) “Movie Feature Extraction
(1) Video–text alignment features (VALOR): To extract video-based multimodal features, we used VALOR (VALOR-large checkpoint), an open-source pretrained video–text alignment model24. VALOR combines visual encoders (CLIP and Video Swin Transformer) for extracting visual features and a text encoder (BERT) for extracting textual features 23,51,52. These representations are aligned in a shared embedding space through contrastive learning. We segmented each movie at the TR level and, for each segment, extracted VALOR’s projected video–text embedding from the final projection head of the alignment module to obtain a 512-dimensional feature vector. These embeddings were then time-aligned to the corresponding BOLD responses.
(2) CLIP features: To compare with static image-based multimodal models, we utilized CLIP (ViT-B/32), which aligns visual and textual representations through contrastive learning but processes individual frames independently without capturing temporal context. One video frame was sampled per TR, and the pooled image embedding after CLIP’s projection into the shared image–text space was extracted to obtain a 512-dimensional feature vector. These TR-aligned vectors were used directly as regressors in the voxel-wise encoding models.
(3) AlexNet features: Visual features were extracted by sampling frames at the TR level and processing them with AlexNet, an eight-layer convolutional neural network comprising five convolutional layers followed by three fully connected layers. Features from all five convolutional layers were evaluated in preliminary analyses; the fifth convolutional layer showed the best performance and was used in subsequent analyses. Intra-image z-score normalization was applied to reduce amplitude effects. Principal component analysis (PCA) was used to reduce dimensionality, retaining the top 512 components to match the dimensionality of multimodal feature spaces. This pipeline was implemented using the DNNBrain toolkit 53.
(4) WordNet features: Semantic features were obtained from publicly available WordNet annotations provided with the HCP dataset (7T_movie_resources/WordNetFeatures.hdf5), following the procedure of Huth et al. (2012). Throughout this manuscript, we use the term “semantic features” to refer to such human-annotated, category-based representations of scene content, and we reserve the term “linguistic features” for continuous language embeddings derived automatically from pretrained language or vision–language models. Each second of the movie clips was manually annotated with WordNet categories according to predefined guidelines: (a) identifying clear objects and actions in the scene; (b) labeling categories that dominated for more than half of the segment duration; and (c) using specific category labels rather than general ones. A semantic feature matrix was constructed with rows corresponding to time points and columns to semantic categories, with category presence coded as binary values. More specific categories from the WordNet hierarchy were added to each labeled category, yielding a total of 859 semantic features. These features were used directly as regressors. We also evaluated a PCA-reduced 512-dimensional variant (fit within each training fold to avoid leakage); because this version performed slightly worse, we report results from the full 859-dimensional representation in the main text. For the generalization analysis in Study 2, annotations for the SFM dataset were aligned to the same WordNet category space to ensure consistency.”
(3) In Figure 3, consider following Huth et al.'s approach by using 3-4 distinct colors to visualize semantic representations across the cortical surface more clearly.
Thank you for this excellent suggestion. We have generated an alternative visualization using a discrete 3–4 color scheme following Huth et al. to display the semantic components on the cortical surface. This version makes the spatial correspondence between components and the boundaries between cortical territories easier to see. We now include this visualization in the Supplement (Fig. S3)
(4) In Figure 2, the brain renderings are too small. Please consider creating a separate, enlarged figure with clearer delineation of relevant ROIs.
We appreciate this suggestion and agree that clear delineation of ROIs is important. We evaluated larger brain renderings; however, within the multi-panel layout of Fig. 2, enlarging them compressed accompanying plots/legends and introduced visual crowding, which reduced overall readability. To preserve a balanced layout and consistent typography across panels, we have kept the current rendering size in the main text and added Fig. S4 with enlarged brain renderings showing clearer ROI boundaries for the same ROIs.
Reviewer #2 (Recommendations for the authors):
(1) From the introduction, I feel like naïve readers would have a hard time understanding what semantic models (e.g., WordNet) are, which the authors write are based on "labor-intensive and subjective manual annotation of semantic content". It would be straightforward to explain the process-how scientists have written descriptions or denoted categories of what's happening within a TR and transformed these into embedding vectors based on language models. This description would explain what the authors mean by "labor-intensive, time-consuming, and subjective". Related to this point, the authors seem to be using the words "semantic model/feature" and "linguistic model/feature" interchangeably, which may exacerbate the confusion.
Thank you for this helpful suggestion. We agree that naïve readers would benefit from a clearer explanation of how “semantic” models such as WordNet are constructed and from a more precise distinction between semantic and linguistic features.
In response, we expanded the Introduction (p. 3) to explicitly describe the process by which semantic features are generated via dense human annotation (i.e., raters label objects, actions, and events within each TR and map these labels onto a predefined ontology to form feature vectors), clarifying why this approach is labor-intensive, time-consuming, and subject to rater variability.
To avoid disrupting the conceptual flow of the Introduction, we placed the explicit terminology clarification in the Methods section (p. 22), where feature extraction is described. There, we now define semantic features as human-annotated, category-based representations of scene content, and linguistic features as continuous language embeddings derived automatically from pretrained language or vision–language models. These revisions are intended to improve clarity and consistency for both expert and non-expert readers.
(On page 3) “Critically, semantic models often rely on dense human annotation. In early naturalistic encoding studies, trained raters watched the stimulus and labeled what was happening within each TR or short time window—for example, identifying objects, actions, or events present in the scene. These labels were then mapped onto a predefined semantic ontology (such as WordNet), yielding high-dimensional categorical feature vectors that served as regressors in encoding models. While this approach provides interpretable semantic features, it is labor-intensive, time-consuming, and inherently subjective, as annotations depend on rater judgment, labeling guidelines, and dataset-specific conventions, limiting scalability and reproducibility.”
(On page 22) “Throughout this manuscript, we use the term “semantic features” to refer to such human-annotated, category-based representations of scene content, and we reserve the term “linguistic features” for continuous language embeddings derived automatically from pretrained language or vision–language models.”
(2) Figure 1A does not look like an accurate schematic of the encoding method. For example, shouldn't the "Train" give rise to weight matrices, and Movies come from moments at Test? I would appreciate it if this schematic figure would explain what the encoding model is to naïve readers.
(3) Figure 1B emphasizes that VALOR is utilizing multimodal features, but does not emphasize that the model is trained on dynamic video. The current figure looks like the model extracted visual and linguistic features from a screenshot of the video, much like the CLIP model.
Thank you for this helpful comment. We agree that the original Fig. 1A did not sufficiently clarify what is learned during training versus what is applied during testing, and that this distinction is particularly important for naïve readers unfamiliar with encoding models. We also agree that the original Fig. 1B did not sufficiently emphasize that VALOR is trained on dynamic video segments, and that the schematic could be misinterpreted as aligning a single video frame with text, similar to CLIP-style image–text models.
We have revised Fig. 1A (p. 6) to make the encoding procedure explicit and pedagogical. Specifically, we now clearly depict that, during the training phase (HCP dataset), voxel-wise encoding models learn feature-to-voxel weight matrices from stimulus features and BOLD responses. These learned weights are explicitly labeled as voxel-wise weight matrices and visually associated with the training stage. In the testing/generalization phase (SFM dataset), we now indicate that these learned weights are held fixed and applied to features extracted from novel movies to generate predicted BOLD responses. Additional labels were added to distinguish “Training (learn weights)” from “Testing/Transfer (apply fixed weights)” and to clarify that the encoding model implements a linear mapping from stimulus features to voxel responses. We have also rewritten the Fig. 1 legend (p. 6) to explicitly explain the encoding workflow in words, including (i) the learning of voxel-specific weights during training, (ii) their reuse during cross-dataset transfer, and (iii) how generalization performance is evaluated. These changes are intended to ensure that Fig. 1A accurately reflects the encoding methodology and is understandable to readers without prior experience with encoding models.
We have revised Fig. 1B (p. 6) to explicitly highlight the temporal nature of the video input used by VALOR. In the updated schematic, the visual stream is depicted as a sequence of consecutive frames spanning multiple seconds, grouped into a video segment, rather than as a single static image. Additional labels indicate that VALOR encodes temporally extended video clips and aligns them with corresponding textual descriptions in a shared embedding space via contrastive learning. We have also updated the figure legend (p. 6) to clarify that VALOR operates on multi-frame video segments and explicitly models temporal structure, distinguishing it from static image–text models such as CLIP. These changes are intended to make clear that VALOR’s advantage derives not only from multimodality, but also from learning representations over time.
(4) Regarding Figure 2, why were paired t-tests conducted in one-sided comparisons? Shouldn't this be two-sided, given that there is no reason to assume one is higher or lower than another?
Thank you for raising this point. We agree that, in the absence of a preregistered directional hypothesis, paired comparisons should be evaluated using two-sided statistical tests.
In response, we have re-run all paired comparisons reported in Figure 2 (p. 9) using two-sided paired t-tests, recomputed the corresponding p-values and false discovery rate (FDR) corrections, and updated the significance markers in the figure and captions accordingly. Importantly, this change does not alter the qualitative pattern of results or the main conclusions reported in the manuscript.
(5) Regarding Study 4, I am curious whether the results are specific to forward-looking representations (predictive coding) or whether the results broadly reveal regions that are sensitive to contexts. For example, if the authors were to incorporate nearby past scenes in the analysis rather than the nearby future scenes, would different brain regions light up?
Thank you for this thoughtful question. We agree that it is important to distinguish forward-looking (predictive) representations from more general sensitivity to temporal context. In Study 4, we deliberately operationalized prediction using future-aligned features, such that only information from upcoming scenes was incorporated into the encoding model. Accordingly, the reported effects should be interpreted as reflecting forward-oriented representations rather than generic context sensitivity.
To make this interpretive scope explicit, we have added a clarifying sentence at the beginning of the Study 4 paragraph in the Discussion (p.18), noting that our analysis incorporates only future-aligned features and that directly contrasting past- and future-aligned features will be an important direction for future work. This clarification is intended to clearly bound our claims while addressing the reviewer’s conceptual distinction..
(On page 18) “In Study 4, we used a video-text alignment model to investigate predictive coding mechanisms. Because our analysis incorporates only future-aligned features, the reported effects should be interpreted as reflecting forward-oriented representations rather than generic sensitivity to temporal context; directly contrasting past- and future-aligned features will be an important direction for future work.”
(6) In the paragraph starting in line 447, were WordNet feature time series also reduced to 512 dimensions like the rest of the model features?
Thank you for the question. In the main analyses, WordNet feature time series were not reduced to 512 dimensions and were instead used at their full dimensionality (859 features).
For comparability with the other feature spaces, we additionally conducted a control analysis in which WordNet features were reduced to 512 dimensions using PCA. The PCA was fit within each training fold to avoid information leakage, and the resulting 512-D features were evaluated using the same encoding pipeline. This PCA-reduced version performed slightly worse than the full 859-D WordNet representation. Accordingly, we report results from the full 859-D WordNet features in the main text. We have clarified this point in the Methods section (p. 22).
(On page 22) “We also evaluated a PCA-reduced 512-dimensional variant (fit within each training fold to avoid leakage); because this version performed slightly worse, we report results from the full 859-dimensional representation in the main text.”
(7) I don't think authors have written what VALOR stands for.
Thank you for the reminder. We now define the VALOR acronym at its first mention in the Abstract and Introduction and use the abbreviation thereafter.
(On page 2) “Using a state-of-the-art deep learning model (VALOR; Vision-Audio-Language Omni-peRception)”
(On page 5) “To answer this, we apply a video-text alignment encoding framework, using VALOR (Vision-Audio-Language Omni-peRception)—a high-performing, open-source model that aligns visual and linguistic features over time—to predict brain responses during movie watching.”
(8) When calculating equation (3), please make sure that the correlation values are Fisher's r-to-z transformed.
Thank you for this reminder. We confirm that all correlation coefficients used in Equation (3) are now Fisher r-to-z transformed prior to any averaging, contrasts, or statistical testing, and this procedure is now explicitly stated in the Methods. We have also updated Fig. 4a (p. 15) to reflect this transformation. Importantly, applying the r-to-z transformation does not change the qualitative pattern of results or their statistical significance.
(9) I wasn't able to check the OSF data/codes because it required permission.
Thank you for flagging this, and we apologize for the inconvenience. We have removed the permission restriction and set the OSF repository to public read-only access, which should resolve the issue.
Reviewer #3 (Recommendations for the authors):
(1) The current approach extracts features from a single "best" layer of each model, which may be suboptimal for predicting neural responses. Prior work has shown that combining features across multiple layers through optimized fusion strategies (e.g., St-Yves et al., 2023) or using model ensembles (e.g., Li et al., 2024) can substantially improve encoding performance. The authors may consider these more comprehensive approaches either as additional baselines or as alternative directions to enhance model accuracy.
Thank you for this constructive suggestion. We agree that combining features across multiple layers or using optimized fusion and ensemble strategies, as demonstrated in recent work (e.g., St-Yves et al., 2023; Li et al., 2024), can substantially improve absolute encoding performance.
In the present study, however, we intentionally evaluated each model using its single best-performing layer within a matched encoding pipeline. This design choice was made to maintain model-agnostic comparability and interpretability, and to ensure that performance differences could be attributed primarily to the type of representation (e.g., temporally informed video–text features versus static or unimodal features), rather than to differences in model complexity, parameter count, or fusion strategy. Importantly, this constraint was applied uniformly across all models and therefore does not favor VALOR over the baselines.
We now explicitly note in the Discussion (p. 19) that multilayer fusion and ensemble approaches represent a natural and promising extension of our framework and are likely to further improve absolute prediction accuracy. Our goal in the current work was to establish the practical utility and generalizability of temporally aligned video–text features for naturalistic movie fMRI under a controlled and comparable evaluation setting..
(On page 19) “Third, for comparability across models we evaluated each model using its single best-performing layer within a matched encoding pipeline rather than using multilayer fusion or ensembling, which allowed us to attribute performance differences to representational format but likely underestimates the absolute performance ceiling.”
(2) Given the naturalistic video-based task, the manuscript would benefit from including state-of-the-art video-only models (e.g., Video Swin Transformer, VideoMAE, and other more recent architectures) as explicit baselines. These models are designed to capture spatiotemporal structure without relying on language input and would provide a more targeted comparison to assess the specific contribution of temporal visual processing.
Thank you for this thoughtful suggestion. We agree that state-of-the-art video-only spatiotemporal models (e.g., Video Swin Transformer, VideoMAE) are highly relevant baselines for naturalistic movie paradigms and would provide a more targeted comparison for isolating the contribution of temporal visual processing independent of language input.
In the present study, our primary goal was not to exhaustively benchmark all possible video architectures, but to evaluate whether temporally informed video–text features can serve as a practical and general-purpose encoding framework that improves upon the models most commonly used in cognitive neuroscience for naturalistic fMRI (e.g., AlexNet for vision, WordNet for semantic annotation, and CLIP for static multimodal alignment). Using these established baselines allowed us to place our results in direct continuity with prior neuroimaging work and to attribute performance differences to representational format under a controlled encoding pipeline.
We agree that incorporating modern video-only spatiotemporal encoders is an important next step, particularly for disentangling the relative contributions of temporal visual structure and cross-modal video–text alignment. We now explicitly note this point in the Discussion (p.19) as a limitation and future direction, and view such comparisons as a natural extension of the current framework within the same TR-aligned encoding setup.
(On page 19) “Second, we did not directly compare VALOR to state-of-the-art video-only spatiotemporal models (e.g., Video Swin Transformer, VideoMAE, and related architectures) that are designed to capture temporal visual structure without language grounding; such comparisons will be important for isolating the specific contributions of temporal visual processing versus cross-modal video–text alignment in naturalistic neural responses.”
(3) An additional consideration is the scale of the AI models used for feature extraction. Previous studies (e.g., Matsuyama et al., 2023) have indicated that model size - particularly the number of parameters - can influence neural prediction performance, independently of architecture. A discussion or analysis of how model size contributes to the observed encoding gains would help clarify whether improvements are due to the representational quality of the model or simply its scale
Thank you for this important point. We agree that model scale—particularly parameter count—can influence neural prediction performance independently of architecture, as noted in prior work (e.g., Matsuyama et al., 2023).
In the present study, our primary goal was to evaluate whether temporally informed video–text representations provide practical advantages over unimodal and static multimodal baselines that are widely used in cognitive neuroscience for naturalistic movie fMRI, under a matched encoding pipeline. We did not perform a systematic scale-controlled analysis in this revision because doing so would require training or evaluating multiple size-matched variants across video-only and video–text architectures, which is beyond the scope of the current work.
We therefore agree that part of the observed performance gains may reflect model capacity in addition to representational format, and we caution against attributing all improvements solely to cross-modal alignment or temporal structure. We now explicitly acknowledge this limitation in the Discussion and note that comparing size-matched video-only and video–text models within the same pipeline is an important next step for disentangling model scale from representational content.
(On page 19) “Finally, part of VALOR’s advantage may reflect model capacity: larger pretrained models often yield higher encoding accuracy, so repeating these analyses with size-matched image-only and image–text models will be critical for disentangling model scale from representational content.”