gain
I think this should say GANG not gain. Proof reader missed that.
gain
I think this should say GANG not gain. Proof reader missed that.
https://www.facebook.com/groups/TypewriterCollectors/posts/10163497490649678/
Royal Pica Italic typeface example with P1 as a foundry mark on the slugs.
Several participants talked about the provider transmitting a ‘vibe’ that made them not trust the providers even if theycould not explain the communication cue explicitly in words. PT14 describes the “vibe” of a negative interaction:
Question: I know the participants said they struggled with defining/elaborating but there was a second interviewer there to ask follow up questions and I wish they did. I can't help but wonder about the details of the "weird/awkward vibe". Was it body language, like the physician not making eye contact, not facing or sitting down next to the patient, or too much time on the computer? Was it the tone and if it was paternalistic, too formal and rigid like robotic or reading off a script, or dismissive? I just feel like this doesn't feel very constructive and any concrete detail would help a lot.
hese three aspects may have influenced BIPOC or LGBTQ+ people to be more willing to share theirnegative experiences and how those experiences impacted their life. C
Summary: Health disparities affecting numerous marginalized populations have always been an issue in healthcare. The concurrent tragedies of the COVID-19 pandemic and the well documented violent death of George Floyd during a time of unprecedented ability to vastly disseminate information via the Internet, allowed for often ignored voices and dismissed experiences to take the spotlight. There was a lot of social momentum to promote education and policy changes, and this paper was able to capitalize off that moment.
e did not recruit information about participants’ socio-economic status or insurance coverage.
Connect: I'm actually surprised this information wasn't collected as this paper seems to be quite comprehensive with its emphasis on both BIPOC and LGBTQ+ communities, and covers a large basis of other forms of discrimination as seen in the previous sentence (gave participants a chance to describe the cause of discrimination to be beyond presumed ethnicity/race/sexuality/gender by including religion/physical build/education/income. It's clear to me that the researchers are very well aware of the concept of intersectionality so that is why I am surprised that they didn't inquire about insurance coverage, as it was covered in great detail by multiple videos in the LGBTQIA+ Health Disparities 101 module that a disproportionate amount of queer individuals lack access to healthcare insurance and/or coverage because of factors directly relating to their marginalized identity.
Stop Calling It Memory: The Problem with Every "AI + Obsidian" Tutorial
To state the obvious: your phone is the best place to keep your calendar and inbox and todo list because you always have it with you, but of course that makes it ripe for other intruders. Bundling makes your phone indispensable, but also a menace.
a giant whiteboard that would single-task everything the way i try to single-task my single-monitor
I rarely use paper or physical space this way because it lacks critical conveniences. A huge wall-mounted paper calendar is maybe the best way to plan and visualize a large coordinated effort. (In "making-of" documentaries you find that movie shoots are often planned this way.) Everyone can see it and point to it because it's at human-scale; you can express many dimensions of information simultaneously using shape, color, position, size, and any other physical attribute. But I have a hard time understanding how I'd keep such a calendar up to date. In practice, like almost everyone else, I use a virtual calendar that automatically adds events as I'm invited to them, syncs with other people's calendars, reconciles time zones effortlessly, and interoperates with other programs like email. Could we get the best of both worlds? In other words, shouldn't one goal of rapid technical advancement be some melding of the physical and virtual worlds such that I can sit quietly in an easy chair with pen and pad; or lay cards out on a table to organize my thoughts; or turn a room into the embodiment of a project; and yet have the same flexibility, portability, persistence, and remixability as in the digital versions of these things?
yes! yes!! what of the thinking i can only do on a whiteboard???
Imagine you had the day’s emails to go through. It would be nice if the ones that required a simple decision could be dispatched with a few pen-strokes: I could write down a date that would work for that meeting; check a box to accept that invitation; etc. If an email required me to review a draft, I'd love to mark up a print version on my couch, sans screen, and have those notes scanned and sent off as if I'd done the whole thing on Google Docs. The point is not to give up on virtuality, but just to save the end user from having to interact with it. It's great to be able to send information to anyone in the world instantly; but let me do it without the glaring screen and the thousand distractions. Is such a thing even possible?
i don't like an ipad for this, but why do i not like an ipad for this? what's the equivalent of this little hypothes.is annotation in this world?
https://www.facebook.com/watch/?v=2178175212945906
For stripped screws:<br /> - try manual screwdriver<br /> - locking pliers on head sticking up<br /> - screw extractor with two tips: left hand drill bit to drill hole, then extractor to pull screw out<br /> - hole saw-like extractor (for wood only) and fill with tapered plug afterward
The Trump administration instituted a policy of separating families apprehended at the border and of criminal prosecution for all apprehended crossing the border illegally, including those applying for asylum.
This is not true. This policy was instituted during the Obama administration. Check the sources before writing untruths. https://www.aclu.org/news/smart-justice/president-obama-wants-continue-imprisoning-immigrant-families
eLife Assessment
This important study demonstrates that ocular organoids can generate both retina and lens through a non-canonical, "inside-out" morphogenetic route. The work is supported by convincing data, with well-designed experiments combining imaging, molecular analysis, and transcriptomics to establish that lens formation in organoids follows conserved molecular programs despite an alternative morphogenesis. These findings expand our understanding of self-organization and developmental plasticity, and will be of broad interest to researchers working on eye development, organoids, and tissue engineering.
[Editors' note: this paper was reviewed by Review Commons.]
Reviewer #1 (Public review):
Summary:
The authors focused on medaka retinal organoids to investigate the mechanism underlying the eye cup morphogenesis. The authors succeeded to induce lens formation in fish retinal organoids using 3D suspension culture with minimal growth factor-containing media containing the Hepes. At day 1, retinal precursor cells expressing Rx3:H2B-GFP appear in the surface region of organoids. At day 1.5, Prox1+ cells appear in the interface area between the organoid surface and the core of central cell mass, which develops a spherical-shaped lens later. So, Prox1+ cells covers the surface of the internal lens cell core. At day 2, foxe3:GFP+ cells appear in the Prox1+ area, where early lens fiber marker, LFC, starts to be expressed. In addition, foxe3:GFP+ cells show EdU+ incorporation, indicating that foxe3:GFP+ cells have lens epithelial cell-characters. At day 4, cry:EGFP+ cells differentiate inside the spherical lens core, whose surface area consists of LFC+ and Prox1+ cells. Furthermore, at day 4, the lens core moves towards the surface of retinal organoids to form an eyecup like structure, although this morphogenesis "inside out" mechanism is different from in vivo cellular "outside -in" mechanism of eye cup formation. From these data, the authors conclude that optic cup formation, especially the positioning of the lens, is established in retinal organoids though the different mechanism of in vivo morphogenesis.
In the revised manuscript, the authors have added new data on dissociation and re-aggregation of day one organoids and revealed that differentially adhesive property of lens and retinal precursors cells enables the formation of a spherical lens in the center of the organoid and later movement of lens toward the peripheral region of the organoid for lens evagination. Furthermore, the authors showed that BMP and FGF signaling are required for lens precursor induction and subsequent lens fiber differentiation in the organoid, respectively. In the revised manuscript, they have added new data on target tissue of BMP and FGF signaling pathways by showing phosphorylated Smad1/5/8 and phosphorylated ERK1/2, respectively, and revealed that lens precursor cells formed in the center of day one organoid are target of BMP signaling, whereas lens fiber cells formed in the center of day 1.5 to 2 organoid are targeted by FGF signaling. Finally, the authors conducted bulk RNA-seq analysis of 1-4 dpf embryonic eyes and day 1-4 eye organoids and revealed that lens organoids show a similar temporal profile of gene transcription. These data suggest that, although induction and morphogenesis of lens are differentially regulated between eye organoids and in vivo embryonic eyes, their molecular mechanism seems to be shared.
Significance:
Strength: This study is unique. The authors examined eye cup morphogenesis using fish retinal organoids. Eye cup normally consists of the lens, the neural retina, pigment epithelium and optic stalk. However, retinal organoids seem to be simple and consists of two cell types, lens and retina. Interestingly, a similar optic cup-like structure is achieved in both cases; however, cellular mechanism of lens induction and morphogenesis are different between retinal organoid and in vivo eyes, although their molecular mechanism is conserved.
Limitation: In the revised manuscript, the authors clarified almost obscure points; however, a couple of unclear points are still retained. First, there is one unknown cell-type population located in the interface area between foxe3:GFP+ cells and rx2:H2B-RFP+ cells at day 2 organoid. Second, the authors showed that removal of HEPES from the organoid culture media inhibits lens induction and differentiation. However, the role of HEPES in lens induction and differentiation in the organoid remains to be elucidated.
Advancement: In the revised manuscript, the authors have provided precise description of inductive and morphogenetic process of lens induction and differentiation in retinal organoid as well as their molecular evidence, which impact the research field of cell biology and regenerative medical science using human organoid.
Audience: The target audience of current study are still within ophthalmology and neuroscience community people, maybe translational/clinical rather than basic biology. To beyond specific fields, need to formulate a general principle for cell and developmental biology.
Reviewer #2 (Public review):
Summary:
In this study from Stahl et al., the authors demonstrate that medaka pluripotent embryonic cells can self-organise into eye organoids containing both retina and lens tissues. While these organoids can self-organize into an eye structure that resembles the vertebrate eye, they are built from a fundamentally different morphogenetic process - an "inside-out" mechanism where the lens forms centrally and moves outward, rather than the normal "outside-in" embryonic process. This is a very interesting discovery, both for our understanding of developmental biology and the potential for tissue engineering applications. The study would benefit from some additional experiments and a few clarifications. The authors suggest that the lens cells are the ones that move from the central to a more superficial position. Is this an active movement of lens cells or just the passive consequence of the retina cells acquiring a cup shape? Are the retina cells migrating behind the lens or the lens cells pushing outwards? High-resolution imaging of organoid cup formation, tracking retina cells in combination with membrane labeling of all cells would help elucidate the morphogenetic processes occurring in the organoids. Membrane labeling would also be useful as Prox1 positive lens cells appear elongated in embryos while in the organoids, cell shapes seem less organised, less compact and not elongated (for example as shown in Fig 3f,g).
The organoids could be a useful tool to address how cell fate is linked to cell shape acquisition. In the forming organoids, retinal tissue initially forms on the outside, while non-retinal tissue is located in the centre; this central tissue later expresses lens markers. Do the authors have any insights into why fate acquisition occurs in this pattern? Is there a difference in proliferation rates between the centrally located cells and the external ones? Could it be that highly proliferative cells give rise to neural retina (NR), while lower proliferating cells become lens?
What happens in organoids that do not form lenses? Do these organoids still generate foxe3 positive cells that fail to develop into a proper lens structure? And in the absence of lens formation, does the retina still acquire a cup shape?
The author suggest that lens formation occurs even in the absence of Matrigel. Is the process slower in these conditions? Are the resulting organoids smaller? While there are indeed some LFC expressing cells by day2, these cells are not very well organised and the pattern of expression seems dotty. Moreover, LFC staining seems to localise posterior to the LFC negative, lens-like structure (e.g. Fig.S1 3o'clock).
How do these organoids develop beyond day 4? Do they maintain their structural integrity at later stages?
The role of HEPES in promoting organoid formation is intriguing. Do the authors have any insights into why it is important in this context? Have the authors tried other culture conditions and does culture condition influence the morphogenetic pathways occurring within the organoids?
Significance:
This is a very interesting paper, and it will be important to determine whether this alternative morphogenetic process is specific to medaka or if similar developmental routes can be recapitulated in organoid cultures from other vertebrate species.
Comments on revised version:
The revised manuscript is much improved and addresses all of the points raised by the reviewers.
Reviewer #3 (Public review):
Major Comments on first version:
- The manuscript presents a beautiful set of high-quality images showing expression of lens differentiation markers over time in the organoids. The set of experiments is very robust, with high numbers of organoids analysed and reproducible data. The mechanism by which lens specification is promoted in these organoids is, however, poorly analysed, and the reader does not get a clear understanding of what is different in these experiments, as compared to previous attempts, to support lens differentiation. There is a mention to HEPES supplementation, but no further analysis is provided, and the fact that the process is independent of ECM contradicts, as the authors point out, previous reports. The manuscript would benefit from a more detailed analysis of the mechanisms that lead to lens differentiation in this setting.
- The markers analysed to show onset of lens differentiation in the organoids seem to start being expressed, in vivo, when the lens placode starts invaginating. An analysis of earlier stages is not presented. This would be very informative, allowing to determine whether progenitors differentiate as placode and neuroepithelium first, to subsequently continue differentiating into lens and retina, respectively. Could early placodal and anterior neural plate markers be analysed in the organoids? This would provide a more complete sequence of lens vs retina differentiation in this model.
- The analysis of BMP and Fgf requirement for lens formation and differentiation is suggestive, but the source of these signals is not resolved or mentioned in the manuscript. Are BMP4 and Fgf8 expressed by the organoids? Where are they coming from?
- The fact that the lens becomes specified in the centre of the organoid is striking, but it is for me difficult to visualise how it ends up being extruded from the organoid. Did the authors try to follow this process in movies? I understand that this may be technically challenging, but it would certainly help to understand the process that leads to the final organisation of retinal and lens tissues in the organoid. There is no discussion of why the morphogenetic mechanism is so different from the in vivo situation. The manuscript would benefit from explicitly discussing this.
Significance:
This study describes a reproducible approach to differentiate ocular organoids composed of lens and retinal tissues. The characterisation of lens differentiation in this model is very detailed, and despite the morphogenetic differences, the molecular mechanisms show many similarities to the in vivo situation. The manuscript however does not highlight, in my opinion, why this model may be relevant. Clearly articulating this relevance, particularly in the discussion, will enhance the study and provide more clarity to the readers regarding the significance of the study for the field of organoid research, ocular research and regenerative studies.
Comments on revised version:
The authors presented substantial additional experimental evidence that further strengthens their manuscript and addressed with these experiments and their revised results/discussion in the manuscript the comments and suggestions from the reviewers. I think the manuscript has been greatly improved with the additions presented.
Author response:
The following is the authors’ response to the original reviews
Thank you very much for the positive and constructive feedback on our manuscript. We have revised the manuscript accordingly and have added a substantial number of additional experiments and have extended the data.
Questions of the reviewers were focused mostly on mechanical insight into organoid formation, touching following aspects of lens organoid formation presented in the manuscript:
- Cellular arrangements/re-arrangements during the process of lens formation including potential contribution of differential adhesion-mediated cell sorting to the cellular arrangement in the organoid and characterization of individual contributions of lens- and retina- committed progenitors to this process.
- Activity of BMP and FGF signaling pathways during organoid formation, namely identification of tissue responding to the signaling withing forming organoids.
- Contribution of externally supplemented Matrigel to the differentiation process and cellular arrangements in ocular organoids.
To address those points in detail we included additional experiments that are now presented in revised version of the manuscript, namely in revised Figure 2-figure supplement 1 (addressing contribution of Matrigel); new Figure 4-supplement 1/Video S5 (addressing contribution of differential adhesion-mediated cell sorting); revised Figure 4/Video S6/Video S7 (addressing contribution of lens-committed progenitors); revised Figure 6 (addressing BMP and FGF signaling pathway activities).
Reviewer #1 (Evidence, reproducibility and clarity):
Summary
The authors focused on medaka retinal organoids to investigate the mechanism underlying the eye cup morphogenesis. The authors succeeded to induce lens formation in fish retinal organoids using 3D suspension culture with minimal growth factor-containing media containing the Hepes. At day 1, Rx3:H2B-GFP+ cells appear in the surface region of organoids. At day 1.5, Prox1+cells appear in the interface area between the organoid surface and the core of central cell mass, which develops a spherical-shaped lens later. So, Prox1+ cells covers the surface of the internal lens cell core. At day 2, foxe3:GFP+ cells appear in the Prox1+ area, where early lens fiber marker, LFC, starts to be expressed. In addition, foxe3:GFP+ cells show EdU+ incorporation, indicating that foxe3:GFP+ cells have lens epithelial cell-characters. At day 4, cry:EGFP+ cells differentiate inside the spherical lens core, whose the surface area consists of LFC+ and Prox1+ cells. Furthermore, at day 4, the lens core moves towards the surface of retinal organoids to form an eye-cup like structure, although this morphogenesis "inside out" mechanism is different from in vivo cellular "outside -in" mechanism of eye cup formation. From these data, the authors conclude that optic cup formation, especially the positioning of the lens, is established in retinal organoids though the different mechanism of in vivo morphogenesis.
Overall, manuscript presentation is nice. However, there are still obscure points to understand background mechanism. My comments are shown below.
Major comments
(1) At the initial stage of retinal organoid morphogenesis, a spherical lens is centrally positioned inside the retinal organoids, by covering a central lens core by the outer cell sheet of retinal precursor cells. I wonder if the formation of this structure may be understood by differential cell adhesive activity or mechanical tension between lens core cells and retinal cell sheet, just like the previous study done by Heisenberg lab on the spatial patterning of endoderm, mesoderm and ectoderm (Nat. Cell Biol. 10, 429 - 436 (2008)). Lens core cells may be integrated inside retinal cell mass by cell sorting through the direct interaction between retinal cells and lens cells, or between lens cells and the culture media. After day 1, it is also possible to understand that lens core moves towards the surface of retinal organoids, if adhesive/tensile force states of lens core cells may be change by secretion of extracellular matrix. I wonder if the authors measure physical property, adhesive activity and solidness, of retinal precursor cells and lens core cells. If retinal organoids at day 1 are dissociated and cultured again, do they show the same patterning of internal lens core covering by the outer retinal cell sheet?
The question, whether different adhesive activity is involved in cell sorting and lens formation is indeed very intriguing.
To address this point, we included additional experiments in the revised manuscript. As proposed by the reviewer, we performed dissociation and re-aggregation experiments of day one organoids at the timepoint, when retinal cell fate is already established and first cells with early lens fate (Foxe3::GFP positive) start appearing (see new Figure 4-figure supplement 1).
After dissociation we followed Foxe3::GFP cells over time and observed that initially equally dispersed GFP<sup>+</sup> lens-committed cells gradually sort and establish contact with other GFP<sup>+</sup> cells, ultimately resulting in the formation of a central GFP<sup>+</sup> sphere within a retinal neuroepithelium (AcTub<sup>+</sup>) localized on the surface of the organoid (see new Figure 4-figure supplement 1e and new Video S5). This data show that differential adhesive properties of lens/retinal precursor cells can enable the formation of a spherical lens in the center of the organoid. This is now clearly stated in the revised version of the manuscript.
(2) Optic cup is evaginated from the lateral wall of neuroepithelium of the diencephalon. In zebrafish, cell movement occurs from the pigment epithelium to the neural retina during eye morphogenesis in an FGF-dependent manner. How the medaka optic cup morphogenesis is coordinated? I also wonder if the authors conduct the tracking of cell migration during optic cup morphogenesis to reveal how cell migration and cell division are regulated in lens of the Medaka retinal organoids. It is also interesting to examine how retinal cell movement is coordinated during Medaka retinal organoids.
Looking into the detail of how optic cup-looking tissue arrangement of ocular organoids is achieved on cellular level is of course interesting. Our previous study showed that optic vesicles of medaka retinal organoids do not form optic cups (for details please see Zilova et al., 2021, eLife). We provide evidence that the formation of cup-looking structure of the ocular organoids presented here is mediated by the following processes: establishment of retina and lens domains at specific regions of the organoid – retina on the surface and lens in the center (see Figure 3-figure supplement 1d and Figure 3e, and Figure 4). Further, the dislocation of the centrally formed lens towards the organoid periphery results in the opening of the retina layer, moving the lens to the periphery while retinal cells stay static. We propose that the “cup-like” shape is acquired by an extrusion-like process of the lens from the center of the organoid.
To address the cellular mechanisms involved in this process, we included additional experiments and followed the movements of retinal and lens cells (see new Figure 4c and 4d, new Videos S6, S7 and S8). Retinal cells (tracked as nuclei of the Rx3::H2B-GFP transgenic line) established in the periphery display repeated short distance movements restricted to the retinal epithelium. These movements are characteristic for interkinetic nuclear migration as found in the developing retina. In contrast, Foxe3::GFP lens progenitor cells performed long distance movements from the center to the periphery of the organoid. This movement was accompanied by profound cell shape changes of lens progenitor cells, suggesting an active movement of lens cells to the organoid periphery. These movements are shown in new/extended figures and in new supplementary videos (new Figure 4c and 4d, new Videos S6, S7 and S8) in the revised version of the manuscript.
(3) The authors showed that blockade of FGF signaling affects lens fiber differentiation in day 1-2, whereas lens formation seems to be intact in the presence of FGF receptor inhibitor in day 0-1. I suggest the authors to examine which tissue is a target of FGF signaling in retinal organoids, using markers such as pea3, which is a downstream target of ERK branch of FGF signaling. Since FGF signaling promotes cell proliferation, is the lens core size normal in SU5402-treated organoids from day 0 to day 1?
Assessing the activity of FGF signaling (cross-reference to Reviewer #3) in the organoids is an important point that we have taken care of and included in the revised manuscript.
To address this point, we assessed which tissue/part of the organoid is responding to FGF signaling. To do so we analyzed the presence of phosphorylated ERK (pERK1/2) as FGF signaling target in ocular organoids from day 1 to day 2. At day 1, only low levels of FGF signaling activity were detectable in presumptive retinal or/and lens tissue (see revised Figure 6b). Only half a day later, a significant increase in FGF activity was observed specifically in the central region of the organoids (lens progenitor domain) (at day 1.5), prior to the onset of differentiation of lens fiber cells. This, together with inability of lens progenitor cells to differentiate to lens fiber cells in the presence of FGF inhibitor SU5402 provided during this critical period (day 1 to day 2) demonstrates that FGF signaling activity localized in the lens progenitor cells is required for lens fiber differentiation.
By day 2, FGF activity was detected in both lens and retinal tissue of the organoid. Similar patterns of FGF activity were observed in embryos at 2 days post fertilization (see revised Figure 6b).
The treatment with the FGF signaling inhibitor SU5402 from day 0 to day 1 did have no impact on the core size of organoid the dimension of which were fully comparable to the control (please see Figure 6d).
(4) Fig. 3f and 3g indicate that there is some cell population located between foxe3:GFP+ cells and rx2:H2B-RFP+ cells. What kind of cell-type is occupied in the interface area between foxe3:GFP+ cells and rx2:H2B-RFP+ cells?
That is for sure an interesting question. We are aware of this population of cells. We currently do not have data that clarify the fate of those cells with the required certainty. Rather than speculating, we are currently following up on that question by scRNA sequencing, however we see that beyond the scope of the current manuscript.
(5) Fig. 5e indicates the depth of Rx3 expression at day 1. Is the depth the thickness of Rx3 expressing cell sheet, which covers the central lens core in the organoids? If so, I wonder if total cell number of Rx3 expressing cell sheet may be different in each seeded-cell number, because thickness is the same across each seeded-cell number, but the surface area size may be different depending on underneath the lens core size. Please clarify this point.
The referee is right, figure 5e indicates the thickness of the cell sheet expressing Rx3 positioned at the surface of the organoid. Indeed, the number of Rx3-expressing cells (and lens cells) scales with the size of the organoid as stated in the submitted manuscript. We have taken care to remove ambiguities related to that point in the revised version of the manuscript.
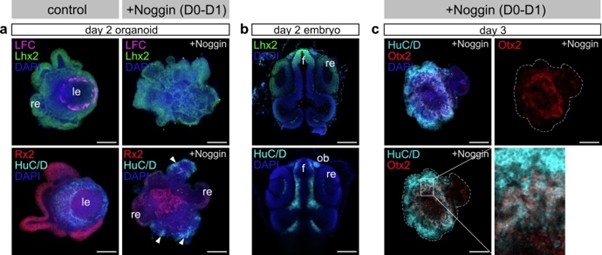
(6) Noggin application inhibits lens formation at day 0-1. BMP signaling regulates formation of lens placode and olfactory placode at the early stage of development. It is interesting to examine whether Noggin-treated organoid expands olfactory placode area. Please check forebrain territory markers.
What tissue differentiates at the expense of the lens in BMP inhibitor-treated organoids is of course an intriguing question.
To address this point, we labeled Noggin treated organoids at day 2 and day 3 with forebrain and olfactory placode markers. We could identify an increase in the domains expressing Lhx2, HuC/D and Otx2 in Noggin-treated organoids, showing a shift of the preferential differentiation of the neurons of anterior forebrain identity (see attached figure for reviewer). However, the available markers Lhx2, HuC/D and Otx2 found in the olfactory placode are in addition also co-expressed in further neuronal cell types of the anterior forebrain. While the speculation is tempting, the shift in expression does not allow to conclusively state the expansion of the olfactory placode.
Author response image 1.
Expression of forebrain and olfactory placode markers.
I have no minor comments
Referees cross-commenting
I agree that all reviewers have similar suggestions, which are reasonable and provided the same estimated time for revision.
Reviewer #1 (Significance):
Strength:
This study is unique. The authors examined eye cup morphogenesis using fish retinal organoids. Eye cup normally consists of the lens, the neural retina, pigment epithelium and optic stalk. However, retinal organoids seem to be simple and consists of two cell types, lens and retina. Interestingly, a similar optic cup-like structure is achieved in both cases; however, underlying mechanism is different. It is interesting to investigate how eye morphogenesis is regulated in retinal organoids,under the unconstrained embryo-free environment.
Limitation:
Description is OK, but analysis is not much profound. It is necessary to apply a bit more molecular and cellular level analysis, such as tracking of cell movement and visualization of FGF signaling in organoid tissues.
Advancement:
The current study is descriptive. Need some conceptual advance, which impact cell biology field or medical science.
Audience:
The target audience of current study are still within ophthalmology and neuroscience community people, maybe translational/clinical rather than basic biology. To beyond specific fields, need to formulate a general principle for cell and developmental biology.
Reviewer #2 (Evidence, reproducibility and clarity):
In this study from Stahl et al., the authors demonstrate that medaka pluripotent embryonic cells can self-organise into eye organoids containing both retina and lens tissues. While these organoids can self-organize into an eye structure that resembles the vertebrate eye, they are built from a fundamentally different morphogenetic process - an "inside-out" mechanism where the lens forms centrally and moves outward, rather than the normal "outside-in" embryonic process. This is a very interesting discovery, both for our understanding of developmental biology and the potential for tissue engineering applications. The study would benefit from some additional experiments and a few clarifications.
The authors suggest that the lens cells are the ones that move from the central to a more superficial position. Is this an active movement of lens cells or just the passive consequence of the retina cells acquiring a cup shape? Are the retina cells migrating behind the lens or the lens cells pushing outwards? High-resolution imaging of organoid cup formation, tracking retina cells in combination with membrane labeling of all cells would help elucidate the morphogenetic processes occurring in the organoids. Membrane labeling would also be useful as Prox1 positive lens cells appear elongated in embryos while in the organoids, cell shapes seem less organised, less compact and not elongated (for example as shown in Fig 3f,g).
Looking into the detail of how the optic cup-like arrangement of ocular organoids is achieved on the cellular level is indeed highly interesting. In the revised manuscript we now provide evidence that the formation of cup-like structure of the ocular organoids presented here is mediated by the following processes: establishment of retina and lens domains at distinct regions of the organoid – retina on the surface and lens in the center (see Figure 3-figure supplement 1d and Figure 3e, and Figure 4). Further, the dislocation of the centrally formed lens towards the organoid periphery results in the opening of the retina layer, moving the lens to the periphery while retinal cells stay static. We propose that the cup-like shape is acquired by an extrusion process of the lens from the center of the organoid.
To address cellular mechanisms involved in this process, we included additional experiments and followed the movements of retinal and lens cells (see new Figure 4c and 4e, new Videos S6, S7 and S8).
Retinal cells (tracked as nuclei of the Rx3::H2B-GFP transgenic line) display repeated short distance movements within the retinal epithelium. These movements are characteristic for interkinetic nuclear migration as found in the developing retina.
In contrast, Foxe3::GFP lens progenitor cells performed long distance movements from the center to the periphery of the organoid. This movement was accompanied by profound cell shape changes of lens progenitor cells, suggesting an active movement of lens cells to the organoid periphery.
These movements are shown in new/extended figures and in new supplementary videos (new Figure 4c and 4e, new Videos S6, S7 and S8) in the revised version of the manuscript.
The organoids could be a useful tool to address how cell fate is linked to cell shape acquisition. In the forming organoids, retinal tissue initially forms on the outside, while non-retinal tissue is located in the centre; this central tissue later expresses lens markers. Do the authors have any insights into why fate acquisition occurs in this pattern? Is there a difference in proliferation rates between the centrally located cells and the external ones? Could it be that highly proliferative cells give rise to neural retina (NR), while lower proliferating cells become lens?
We agree with the reviewer that this is a highly interesting question and in the revised manuscript we followed the advice and dedicated a part of the discussion to this topic. We believe that the arrangement is due to the induction of central lens fates by signal emanating from the retinal epithelium and discuss the role of the diffusion limit and the potential contribution of BMB and FGF signaling to this arrangement. Additional experiments addressing the target tissues of FGF and BMP signaling in the organoid have been provided in response to Reviewer #1. Interfering with FGF signaling that is essential for lens fiber cell differentiation interestingly did not impact on the lens size arguing against an immediate proliferative effect. Although the analysis of the respective proliferation rates at the surface or in the central region of the organoid might show some differences, we do not have any indications, that the proliferation rate itself would be instructive or superior to the cell fate decisions.
What happens in organoids that do not form lenses? Do these organoids still generate foxe3 positive cells that fail to develop into a proper lens structure? And in the absence of lens formation, does the retina still acquire a cup shape?
Lens formation is primarily dependent on the acquisition/specification of Foxe3-expressing lens placode progenitors. In the absence of Foxe3-expression, a lens does not develop. Once Foxe3-expressing progenitors are established, a lens is formed in unperturbed conditions (measured by the presence of expression of crystallin proteins). Organoids that do not have a lens, do not contain Foxe3-expressing cells.
In the absence of a lens, the organoid is composed of retinal neuroepithelium, that does not form an optic cup like shape (for details of such phenotypes please see Zilova et al., 2021, eLIFE). We took care to state that clearly in the revised manuscript.
The author suggest that lens formation occurs even in the absence of Matrigel. Is the process slower in these conditions? Are the resulting organoids smaller? While there are indeed some LFC expressing cells by day2, these cells are not very well organised and the pattern of expression seems dotty. Moreover, LFC staining seems to localise posterior to the LFC negative, lens-like structure (e.g. Fig.S1 3o'clock). How do these organoids develop beyond day 4? Do they maintain their structural integrity at later stages?
The role of HEPES in promoting organoid formation is intriguing. Do the authors have any insights into why it is important in this context? Have the authors tried other culture conditions and does culture condition influence the morphogenetic pathways occurring within the organoids?
We thank the reviewer for pointing this out. In the revised manuscript we made sure to be sufficiently clear in the wording and description of our observation. Indeed, Matrigel is not required for the acquisition of lens fate, which can be demonstrated by the expression of lensspecific markers. However, the presence of Matrigel has a profound impact on structural aspects of organoid formation. Matrigel is essential for organization of retinal-committed cells to form a retinal epithelium (Zilova et al., 2021, eLife). The absence of the structure of the retinal epithelium indeed negatively impacts on the cellular organization and the overall lens structure.
To clarify the contribution of the Matrigel to the organoid organization, we performed additional experiments (see revised Figure 2-figure supplement 1c-f). As mentioned above, the absence of Matrigel impacts on the organization and thickness of retinal neuroepithelium (Rx2<sup>+</sup>, Figure 2-figure supplement 1c). However, measurement of the lens in organoids at day 2 and day 5 showed that size of the lens is not impacted upon in the absence of Matrigel (Figure 3-figure supplement 1d-e). Additionally, taking advantage of the Foxe3::GFP lens reporter line, we measured the onset of lens-specific gene expression in organoids with and without Matrigel. In both conditions, with and without Matrigel supplementation, Foxe3::GFP expression was initiated at 25 hours post aggregation (see revised Figure 4b).
The role of the HEPES in lens formation is indeed very intriguing and currently under investigation. HEPES is mainly used to regulate the pH of the culture media which on its own might have an impact on multiple cellular processes. It will require a significant time investment to address the potential HEPES triggered molecular mechanisms impacting on lens formation (cross reference with Reviewer #3), which goes beyond the scope of the current manuscript.
Referees cross-commenting
Pleased to see that all the other reviewers are positive about the study and raise similar concerns and comments
Reviewer #2 (Significance):
This is a very interesting paper, and it will be important to determine whether this alternative morphogenetic process is specific to medaka or if similar developmental routes can be recapitulated in organoid cultures from other vertebrate species.
Reviewer #3 (Evidence, reproducibility and clarity):
Summary:
The manuscript by Stahl and colleagues reports an approach to generate ocular organoids composed of retinal and lens structures, derived from Medaka blastula cells. The authors present a comprehensive characterisation of the timeline followed by lens and retinal progenitors, showing these have distinct origins, and that they recapitulate the expression of differentiation markers found in vivo. Despite this molecular recapitulation, morphogenesis is strikingly different, with lens progenitors arising at the centre of the organoid, and subsequently translocating to the outside.
Comments:
The manuscript presents a beautiful set of high quality images showing expression of lens differentiation markers over time in the organoids. The set of experiments is very robust, with high numbers of organoids analysed and reproducible data. The mechanism by which lens specification is promoted in these organoids is, however, poorly analysed, and the reader does not get a clear understanding of what is different in these experiments, as compared to previous attempts, to support lens differentiation. There is a mention to HEPES supplementation, but no further analysis is provided, and the fact that the process is independent of ECM contradicts, as the authors point out, previous reports. The manuscript would benefit from a more detailed analysis of the mechanisms that lead to lens differentiation in this setting.
We followed the reviewer’s advice and have included a systematic analysis of the contribution of ECM (Matrigel) to the process of lens formation. In the revised manuscript we made sure to be sufficiently clear in the wording and description of our observation. Indeed, Matrigel is not required for the acquisition of lens fate, which can be demonstrated by the expression of lensspecific markers. However, the presence of Matrigel has a profound impact on structural aspects of organoid formation. Matrigel is essential for organization of retinal-committed cells to form a retinal epithelium (Zilova et al., 2021, eLIFE). The absence of the structure of the retinal epithelium in turn indeed negatively impacts on the cellular organization and the overall lens structure.
To clarify the contribution of the Matrigel to the organoid organization, we performed additional experiments (see revised Figure 2-figure supplement 1c-f). As mentioned above, the absence of Matrigel impacts on the organization and thickness of retinal neuroepithelium (Rx2<sup>+</sup>, Figure 2-figure supplement 1c). However, measurement of the lens in organoids at day 2 and day 5 showed that size of the lens is not impacted upon by the absence of Matrigel (Figure 3-figure supplement 1d-e).
Additionally, taking advantage of the Foxe3::GFP lens reporter line, we measured the onset of lens-specific gene expression in organoids with and without Matrigel. In both conditions (with and without Matrigel supplementation), Foxe3::GFP expression was initiated at 25 hours post aggregation (see revised Figure 4b).
The role of the HEPES in lens formation is indeed intriguing and currently under investigation. HEPES is mainly used to adjust the pH of the culture media, which, on its own might have an impact on multiple cellular processes. It will require a significant time investment to address the potential HEPES triggered molecular mechanisms impacting on lens formation (cross reference with Reviewer #3), which clearly goes beyond the scope of the current manuscript.
The markers analysed to show onset of lens differentiation in the organoids seem to start being expressed, in vivo, when the lens placode starts invaginating. An analysis of earlier stages is not presented. This would be very informative, allowing to determine whether progenitors differentiate as placode and neuroepithelium first, to subsequently continue differentiating into lens and retina, respectively. Could early placodal and anterior neural plate markers be analysed in the organoids? This would provide a more complete sequence of lens vs retina differentiation in this model.
We have taken care to show according stages in embryo and organoid side by side. We provide additional data to highlight the expression of Rx3::H2B-GFP (retina) and Foxe3::GFP (lens and lens placode) markers in earlier developmental stages. For the presumptive eye field within the region of the anterior neural plate (S16, late gastrula) Rx3 represents one of the earliest markers (see revised Figure 3-figure supplement 1). Already before an apparent lens placode is formed (see revised Figure 3d) Foxe3::GFP expression is detected within the presumptive lens ectoderm, demonstrating that Foxe3 is ideally suited as an early marker for placodal progenitors in medaka. The onset of Rx3 and Foxe3-driven reporters is clearly early enough to support the claim about the separate origin of the lens (placodal) and retinal (anterior neuroectoderm) tissues within the ocular organoids now represented in the revised figures.
The analysis of BMP and Fgf requirement for lens formation and differentiation is suggestive, but the source of these signals is not resolved or mentioned in the manuscript. Are BMP4 and Fgf8 expressed by the organoids? Where are they coming from?
Assessing the activity of BMP and FGF signaling (cross-reference to Reviewer #1) in the organoids is an important point that we have taken care of and included in the revised manuscript.
To address this point, we assessed which tissue/part of the organoid is responding to BMP and FGF signaling. To do so we analyzed the presence of phosphorylated SMAD1/5/8 (pSMAD1/5/8) and phosphorylated ERK (pERK1/2) as BMP and FGF signaling target in ocular organoids from day 1 to day 2. BMP signaling activity was detected in the center (region of establishment of lens-committed progenitors (Figure 3e)) of the organoid at day 1 (see revised Figure 6a). At day 1, only low levels of FGF signaling activity were detectable in presumptive retinal or/and lens tissue (see revised Figure S6b). Only half a day later, a significant increase in FGF activity was observed specifically in the central region of the organoids (lens progenitor domain, at day 1.5), prior to the onset of differentiation of lens fiber cells. This, together with inability of lens progenitor cells to differentiate to lens fiber cells in the presence of FGF inhibitor SU5402 provided during this critical period (day 1 to day 2) demonstrates that FGF signaling activity localized in the lens progenitor cells is required for lens fiber differentiation.
By day 2, FGF activity was detected in both lens and retinal tissue of the organoid. Similar patterns of FGF activity were observed in embryos at 2 days post fertilization (see revised Figure S6b).
The treatment with the FGF signaling inhibitor SU5402 from day 0 to day 1 did have no impact on the core size of organoid the dimension of which were fully comparable to the control (please see Figure 6b).
Related to the presence of the corresponding ligands we can state that they are indeed expressed in the organoids at the matching stages based on RNA seq and RT-PCR analyses, however we could not find them specifically localized. This may be due to a widespread, ubiquitous expression or may simply relate to technical problems.
While we can state with confidence that the ligands are present at the relevant time points and trigger the downstream pathways in a localized manner, the question whether the response is due to a localized signal or localized competence remains to be addressed.
The fact that the lens becomes specified in the centre of the organoid is striking, but it is for me difficult to visualise how it ends up being extruded from the organoid. Did the authors try to follow this process in movies? I understand that this may be technically challenging, but it would certainly help to understand the process that leads to the final organisation of retinal and lens tissues in the organoid. There is no discussion of why the morphogenetic mechanism is so different from the in vivo situation. The manuscript would benefit from explicitly discussing this.
Following the shift of the lens in vivo is indeed very relevant suggestion and we have taken care to address this in the revised manuscript.
To clarify this process, we included additional experiments and followed the movements of lens cells (see new Figures 4c, 4d and 4e, new Videos S6 and S7). Foxe3::GFP lens progenitor cells were found to actively move over long distances from center to the organoid periphery. This movement was accompanied by profound cell shape changes of lens progenitor cells with the active extension of lamellipodia and filopodia strongly arguing for an active movement of lens cells to the organoid periphery (cross-reference with Reviewer #1 and Reviewer #2).
Referees cross-commenting
We all seem to have similar comments and concerns. I think overall the suggestions are feasible and realistic for the timeframe provided.
Reviewer #3 (Significance):
This study describes a reproducible approach to differentiate ocular organoids composed of lens and retinal tissues. The characterisation of lens differentiation in this model is very detailed, and despite the morphogenetic differences, the molecular mechanisms show many similarities to the in vivo situation. The manuscript however does not highlight, in my opinion, why this model may be relevant. Clearly articulating this relevance, particularly in the discussion, will enhance the study and provide more clarity to the readers regarding the significance of the study for the field of organoid research, ocular research and regenerative studies.
"History tells a story, one of the earth and time, time before and time now" - Love this!
I like how this page uses poetry and visual art to challenge the Anthropocene as a single linear story by showing that different histories, spaces, and ways of living coexist at the same time.
I never knew that climate change as a literary subject has been avoided in ficiton novels! This definitely challenged my understanding of the Anthropocene, more specifically how people interact with the Anthropocene by showing me that our relationship to it is shaped not only by environmental reality, but by what literature makes visible or leaves out.
They raised funds to hire lawyers or, where appropriate, donated their legal skills freely. Representatives of radical groups acted on negotiation teams in some prison uprisings, working alongside politicians, journalists, and others to represent prisoner interests. Some contributed even more directly: they organized with other prisoners after being incarcerated for their own political activism. While some prisoners became activists out of simple frustration with their conditions, other activists became prisoners. They helped each other, the seasoned activists providing political education while the longtime prisoners providing knowledge of how to navigate or undermine a particular institution.
aided each other, activists went to prison to help inmates get their point of interest
Black Power and New Left radicals. Organizations such as the Black Panther Party (BPP; founded in 1966), the Republic of New Afrika (RNA; founded in 1968), and the Young Lords Party (YLP; a Puerto Rican organization founded in New York in 1969)
groups that aided in the movement
Adaptad el lenguaje al perfil sociocultural de las personas destinatarias. Seleccionad una muestra representativa del territorio o grupo objetivo, considerando variables como género, edad, origen, nivel socioeconómico o localización geográfica. Evitad preguntas sesgadas o que induzcan respuestas. Garantizad la confidencialidad y el anonimato de las respuestas. Incluid preguntas abiertas para captar matices y opiniones no previstas. Asegurad la accesibilidad (lectura fácil, traducción, asistencia si es necesario). Validad el cuestionario con personas del territorio antes de su aplicación masiva.
verbo de la instrucción en infinitivo
Referencias
añdir: Añadir: Batthyány, K., Cabrera, M., Alesina, L., Bertoni, M., Mascheroni, P., Moreira, N., ... & Rojo, V. (2011). Metodología de la investigación para las ciencias sociales: apuntes para un curso inicial. https://repositorio.minedu.gob.pe/handle/20.500.12799/4544
This page is strong because it shows how stories and personal experience can make climate change feel immediate rather than distant. It's also one of my favourite pages because my project also focuses on the importance of personal stories as productive devices in the Anthropocene!
https://www.facebook.com/groups/TypewriterCollectors/posts/10163497483964678
In 9 to 5, Dora Lee seems to be using a Royal (Triumph-Adler) SE1000CD typewriter.
See also trailer at: https://www.youtube.com/watch?v=qni6HOyPNBA
Utilizad metodologías que faciliten los procesos colectivos de toma de decisiones. En este caso, existen multitud de herramientas para ordenar y priorizar las propuestas: pirámide de priorización, técnica MoSCoW, matrices de incidencia estratégica, etc. Las metodologías son únicamente herramientas para facilitar el proceso del taller de priorización. No obstante, su uso debe adaptarse al contexto del grupo y al objetivo del taller. En ocasiones, las herramientas pueden tener alcances limitados a la hora de decidir, por lo que se deberán tener en cuenta otros factores, como la preexistencia de jerarquías dentro del grupo o las divergencias individuales que puedan surgir en el taller. Acompañad el proceso con un equipo técnico que tenga conocimiento sobre las metodologías y propuestas que se llevan a discusión. Distribuid funciones entre las personas participantes para evitar jerarquías informales dentro del grupo.
los verbos de instrucciones en infinitivo
todos los actores involucrados.
todas las personas involucradas.
técnicos
personal técnico
distintos
diferentes
implicado
participantes
agentes
actores
los
quitar
los agentes
quitar
I really like how this page shows that setting is not just a narrative device that initiates the main backdrop and mood for a story. Setting importantly makes climate crisis imaginable to the reader.
This page makes a crucial point that the word "our" can flatten certain groups' responsibility and hide the unequal histories behind this epoch of ecological crisis
I like how this page fames literature as a way of making the Anthropocene feel intellectually and personally graspable/understandable (rather than it being an abstract concept)!
This would change inheritance systems, taxation, and even legal structures. Trade goods would have been replaced with wool and plant-based oils, particularly affecting the mediterranean. Additionally, without cattle, there would be a greater demand for human labor in agriculture.
I like this reimagining of economic structures, especially the shift in labor demands. It might be worth pushing this further—would increased human labor slow down industrialization or fundamentally change class structures?
Essentially, aurochs pioneered the messy, mixed landscapes full of diversity and characteristic of European landscapes. Remove aurochs and the ecosystems become more uniform and less interactive.
I think it's very interesting and impressive how well you've thought out the consequences of removing Aurochs. Again, it emphasizes how much influence cattle have had on our planet over time, something I haven't thought deeply about. I actually recently asked my mom if she knew where cows originated from, so it's funny to be reading a project entirely on this subject.
he absence of cattle from European and North African landscapes would likely reshape early cultures in these regions, snowballing into massive differences in our modern existence. Cattle were not just food, they were engines of agriculture.
It is very interesting to trace back the origin of cattle, because it is such a defining part of our food chain, cultures, and ecosystems. If we want to tackle the issue that factory farming and the domestication of cattle raise, then we have to trace just how far back our relationship with cattle goes, and how deeply we will be affected if we do anything to change this. I haven't thought of it in this way before.
DescriptionDetails
I would have loved to see more images/evidence of how they are cultural symbols in Europe. Maybe even enlarging the image so we can see it better?
image
Did you mean to write "imagine"? Otherwise, I love the idea!
técnicos
personal técnico
Involucrad a la comunidad desde el inicio: una intervención tácticamente exitosa necesita la participación activa y el reconocimiento de actores locales (Lydon, 2012). Sed realista en el alcance: un proyecto pequeño pero bien ejecutado puede generar un impacto mayor que intervenciones complejas sin seguimiento (Mohankumar, 2020). Evaluad el marco regulador: si bien muchas experiencias surgen desde la informalidad o el activismo ciudadano, considerad las condiciones locales para evitar conflictos innecesarios. Proyectad o diseñad la intervención con criterios de inclusividad y perspectiva feminista: intenta eliminar las barreras arquitectónicas, generar recorridos accesibles a pie o en bicicleta, incluid elementos de descanso o reposo e intentad eliminar espacios inseguros o mal iluminados. Documentad todo el proceso: las imágenes, los testimonios y los datos del uso son esenciales para comunicar el valor de la intervención y proponer continuidad (Lydon, 2016). Apostad por la replicabilidad: diseñad con materiales y procesos accesibles que permitan ser replicados en otros puntos del territorio con facilidad.
verbo en infinitivo
¿Cómo?
usar siempre la forma impersonal ej: en lugar de podéis apoyaros poner "se puede apoyarse"
los
eliminar
.
como las recopiladas en este toolkit
encuestas y
eliminar encuesta y poner "la"
el privado
la ciudadanía
efémeras,
efímeras
VERIFY
yes - we us this as a search help for setting up interviews . Match . com - smile every employer has a wish list - different for every job we take the wish list and search candies according. right now each column in a applicants profile can be used as a search filter .
VERIFY
Not exactly how it works . in country is always filled by MTL all process goes though us- we may send it to someone else to process in the end by the paperwork goes though us . A few exertions where an employer does the process _"we have a flag for that in the document status .
VERIFY
all in country are in the same group . MTL just looks at certain details to make sure they match certain employers . That's why we have two visa columns per candidate . one is Visa requested by the candidate : one is MTL assigned visa : This would be sample IN-Counrty H2B + conditions if they apply like : 3yr limit / ISSUE WILL NEED APPROVAL - shorten petition
we have different "flags" set up right now
VERIFY
Foreign or Domestic Foreign - MTL does everything Domestic MTL we don't need visa documents but if hired they might get an offer same as the H2B and if the employer wats we arrange travel curtesy
but for recruitment we need to track them
Declaraciones sobre ética de la investigación (v.g.: consentimiento informado). Fuentes de financiamiento. Considerar el nombre de la institución y el código del financiamiento. Solo es considerado algún tipo de financiamiento monetario.
Averiguar
Dirección postal del autor para correspondencia.
Averiguar
415 TWh of electricity in 2024, re
The energy consumption implications of cellphones and of technology in general makes me think of the digital environmental humanities we spoke about in class. Given that this project is online and because of this it is able to reach a bigger audience faster than something said or written on paper. Do the social and educational benefits of the DEH in some sense outweigh the environmental ramifications of this technology? I'm curious how your project would trouble this and what the way forward would be of considering both or one more than the other.
Facebook alone could hold at least 1.4 billion profiles of deceased users, and potentially far more
I think this page was a perfect mix of statistics and humanities. This page presented the shock factor of the amount of energy consumed from seemingly meaningless actions like using a phone, while also connecting to the images and how images can be damaging not just in judgement and comparison but also on an environmental scale. Tying these two ideas together captures what I believe you are getting at about the Anthropocene; the environmental impacts and the social impacts of humanity in a time of struggle for many.
ingering in clouds and servers I will never see.
This passage really challenged my understanding of the Anthropocene and stuck with me as I continued navigating your Scalar project. I had previously not concretely thought about the relationship with an everyday object, like the phone, and the environmental impacts in places so far from me, that I may never go to. More than that I had not been able to visualize the impacts of data on 'the cloud' until I read this page and thought about the amount of electricity that is being consumed for one photo. I think this is a really interesting way of reaching the local dimensions of the Anthropocene from something as global as phones. It makes you think about your personal impact and how far removed you are from it as we spoke about in class.
The Black Mirror
I love the title of this page. It perfectly plays with the concept of speculative fiction we discussed in class by combining it with a dystopian sense of where the phone that humanity is so attached to is headed in the future.
eLife Assessment
This manuscript reports a valuable modeling study on sequence generation in the hippocampus in a variety of behavioral contexts. The authors model context-depending decision making, and suggest that psychiatric disorders can be interpreted in terms of over or under representation of context information. The presentation is solid, and the work will interest the broad community of researchers studying cortical-hippocampal interactions and sequences.
Reviewer #2 (Public review):
[Editors' note: This version has been assessed by the Reviewing Editor without further input from the original reviewers. The authors have addressed the comments raised in the previous round of review.]
Summary:
Ito and Toyoizumi present a computational model of context-dependent action selection. They propose a "hippocampus" network that learns sequences based on which the agent chooses actions. The hippocampus network receives both stimulus and context information from an attractor network that learns new contexts based on experience. The model is consistent with a variety of experiments both from the rodent and the human literature such as splitter cells, lap cells, the dependence of sequence expression on behavioral statistics. Moreover, the authors suggest that psychiatric disorders can be interpreted in terms of over/under representation of context information.
My general assessment of the work is unchanged, and I still have some questions requesting methodological clarification
Strengths:
This ambitious work links diverse physiological and behavioral findings into a self-organizing neural network framework. All functional aspects of the network arise from plastic synaptic connections: Sequences, contexts, action selection. The model also nicely links ideas from reinforcement learning to a neuronally interpretable mechanisms, e.g. learning a value function from hippocampal activity.
Reviewer #3 (Public review):
Summary:
This paper develops a model to account for flexible and context-dependent behaviors, such as where the same input must generate different responses or representations depending on context. The approach is anchored in the hippocampal place cell literature. The model consists of a module X, which represents context, and a module H (hippocampus), which generates "sequences". X is a binary attractor RNN, and H appears to be a discrete binary network, which is called recurrent but seems to operate primarily in a feedforward mode. H has two types of units (those that are directly activated by context, and transition/sequence units). An input from X drives a winner-take-all activation of a single unit H_context unit, which can trigger a sequence in the H_transition units. When a new/unpredicted context arises, a new stable context in X is generated, which in turn can trigger a new sequence in H. The authors use this model to account for some experimental findings, and on a more speculative note, propose to capture key aspects of contextual processing associated with schizophrenia and autism.
Strengths:
Context-dependency is an important problem. And for this reason, there are many papers that address context-dependency - some of this work is cited. To the best of my knowledge, the approach of using an attractor network to represent and detect changes in context is novel and potentially valuable.
Author response:
The following is the authors’ response to the previous reviews
Public Reviews:
Reviewer #2 (Public review):
Summary:
Ito and Toyoizumi present a computational model of context-dependent action selection. They propose a "hippocampus" network that learns sequences based on which the agent chooses actions. The hippocampus network receives both stimulus and context information from an attractor network that learns new contexts based on experience. The model is consistent with a variety of experiments both from the rodent and the human literature such as splitter cells, lap cells, the dependence of sequence expression on behavioral statistics. Moreover, the authors suggest that psychiatric disorders can be interpreted in terms of over/under representation of context information.
My general assessment of the work is unchanged, and I still have some questions requesting methodological clarification
Strengths:
This ambitious work links diverse physiological and behavioral findings into a self-organizing neural network framework. All functional aspects of the network arise from plastic synaptic connections: Sequences, contexts, action selection. The model also nicely links ideas from reinforcement learning to a neuronally interpretable mechanisms, e.g. learning a value function from hippocampal activity.
Weaknesses:
The presentation, particularly of the methodological aspects, needs to be heavily improved. Judgment of generality and plausibility of the results is severely hampered but is essential, particularly for the conclusions related to psychiatric disorders. In its present form, it is impossible to judge whether the claims and conclusions made are justified. Also, the lack of clarity strongly reduces the impact of the work on the field.
Thank you for pointing this out.
In the revised text, we clarified the definition of “time step” and how hippocampal neurons behaved in each time step (see individual comments below). Also, we clarified the implementation of disorder conditions in our model by indicating the exact neuron numbers of the stimulus domain in H module as below. (Other parameters were common in all conditions.)
“𝑋 consists of two domains: stimulus domain 𝑋 and context domain 𝑋. The neuron ratio in the stimulus domain over the whole neurons dim 𝑋/𝑁 is 16.7% (200 neurons) for the control condition, 2.5% (30 neurons) for the SZ condition, and 50% (600 neurons) for the ASD condition.”
Comments:
The authors have made strong efforts to improve on their description of the methods, however, it is still very hard to understand. As a result of some of their clarifications, new issues appeared that I was not able to extract in the previous version.
(1) Particularly I had problems figuring out how the individual dynamical systems are interrelated (sequences, attractor, action, learning). As I understand it now (and I still might be wrong) there is one discrete time dynamics, where in each time step one action takes place as well as the attractor and sequence dynamics are moved one step forward. Also, synaptic updates happen in every one of those time steps. The authors may verify or correct my interpretations and further improve on their description in the manuscript. It is also confusing that time in the figure panels is given in units of trials, where each trial may consist of (maybe different amounts of) multiple time steps. Are the thin horizontal red ad blue lines time steps?
Thank you for raising the confusing point.
The reviewer’s understanding is correct. In our model, at each time step the agents transition to the next environmental state (which also corresponds to the contextual state). During this step, each processing stage proceeds in order: Context selector performs attractor selection, Sequence composer performs sequence selection, followed by action selection and synaptic updates. As learning progresses and hippocampal sequences begin to predict longer futures, reducing the need for step-by-step planning. However, at least at the beginning of each task, all processes are conducted at each time step (see Fig. 1G).
In all tasks, trials are reset when the agents visit the reward sites (i.e., S4 or S5). n Fig. 2C, for example, one trial consists of three time steps (i.e., three state transitions), and the red and blue shaded regions indicate individual trials. During each time step, two types of hippocampal neurons are activated: a state-coding neuron and a transition-coding neuron. (In contrast, in X, one contextual state is active during one time step). Therefore, in Fig. 2E, two neuronal activities correspond to a single time step.
For clarification, we have revised Fig. 2 and related descriptions in the manuscript as follows.
“Here, we simplified this task by using an environment with five discrete states (S1-S5), i.e., five discrete external stimuli (Figure 2A), where agents transition to the next state at each time step.”
“Figure 2C illustrates an example of both the environmental state transition and the corresponding contextual state transition of an agent, with each trial resetting upon visiting the reward sites (S4 or S5). ”
“At each time step, one state-coding neuron and one transition-coding neuron are active in this order.”
“At each time step, the agents transition between environmental states.”
“The model’s computational dynamics are fundamentally synchronized with the environmental (behavioral) time step, and at each time step, the agents transition to the next environmental state. Upon a state transition, the agents first perform contextual state estimation by Context selector and activate a corresponding hippocampal neuron.”
(2) As a consequence of my new understanding of the model dynamics, I have become doubts about the interpretation of the attractor network as context encoding. Since the X population mainly serves to disambiguate sequence continuation, right before the action has to be taken (active for only two time steps in Figure 1C?) they could also be considered to encode task space (El-Gaby et al. 2024; doi: 10.1038/s41586-024-08145-x).
We thank the reviewer for this insightful comment.
First of all, we would like to clarify that Figure 1C shows the following process: the activity of H at time step t−1 and the external stimulus at time step t jointly provide input to X module, and the activity of X settles into a contextual state at the time step t. As explained in our response to comment (1), the activity of X remains constant during each time step.
The primary function of X module in our model is to disambiguate the environmental states defined by the external stimuli based on the history information. It is true that, in practice, whether an ongoing sequence is maintained or remapped depends on whether the observed stimulus is consistent with the predicted stimulus. However, this is a consequence of the predictive sequence obtained from scratch rather than the primary computational role of X module. In contrast, X module becomes particularly important when past experience does not uniquely determine the next state. In this situation, the agent must infer the contextual state by associating the current situation with previously experienced contexts, rather than relying solely on temporal continuity.
We also add that, in most successful cases, the contextual states learned by the agent often correspond to the hidden states of each task as a result of disambiguation. In this sense, the resulting representation may resemble a “task space” encoding, as suggested by the reviewer. However, an important aspect of our model is that the agent does not assume the existence or number of hidden states a priori. Instead, we considered the situation where the agent initially underestimates the number of contextual states, and through remapping it incrementally increases the number of contextual representations. When the number of contextual states matches the number of hidden task states, the task is typically solved.
(3) Also technically, I wonder why the authors introduce the criterion of 50(!) time steps to allow the attractor to converge, if the state of the attractor network is only relevant in one time step to choose the appropriate continuation of the sequence of actions. Is attractor dynamics important at all? What would happen if just the input and output weights to the X population are kept and the recurrent weights are set 0?
We thank the reviewer for raising this confusing point.
First, we would like to clarify that the “50 iterations” mentioned in the manuscript does not refer to 50 environmental time steps. We implemented multiple iterations of attractor updates (typically until convergence) by Context selector within each behavioral time step.
We clarify this point in the Method section as below.
“After history-based or landmark-based initialization, X iteratively updates its contextual state at the beginning of each time step according to the associative memory dynamics:”
The recurrent connectivity within the X population is essential for attractor updates. If the recurrent weights were removed (i.e., set to zero), the network would lose the ability to retrieve distinct contextual states for the same stimulus. In that case, the model would be unable to solve the context-dependent task as we showed in this manuscript.
(4) Figure 3E: How many time steps are the H cells active (red bars?) Figure 4J: What are the units of the time axis?
Thank you for pointing this out.
In Figure 3E, each time step is indicated in the X-axis ticks (i.e., each environmental state). As we explained in the comment (1), two hippocampal neurons’ activity (red bars) corresponds to each time step.
Similarly, in Figure 4J, each time step is indicated in the X-axis ticks. To better represent the results, we added descriptions of the environmental states in our model to the X-axis tick labels in Figure 4J.
We added the following texts below in Figure captions.
“The x-axis represents each time step (corresponding to environmental states), and the y-axis shows the sorted activity of H module.”
“The x-axis represents each time step (corresponding to environmental states), and the y-axis shows the decoding accuracy of each context based on hippocampal activity.”
I think part of what happened is that many middle- and upper-income households were used to being able to afford low-wage labor on demand - for childcare, for food service, for home health care. Middle- and upper-income households found this frustrating and assumed it was part of the broad story throughout the economy; not realizing that much of this frustration was driven by low-wage workers finally earning a little more bargaining power.
spicy
Michel Kalecki was a Polish economist who argued that full employment would actually be opposed by capitalists despite the positive effects on profits and economic growth.It is true that profits would be higher under a regime of full employment than they are on average under laissez-faire, and even the rise in wage rates resulting from the stronger bargaining power of the workers is less likely to reduce profits than to increase prices, and thus adversely affects only the rentier interests. But “discipline in the factories” and “political stability” are more appreciated than profits by business leaders. Their class instinct tells them that lasting full employment is unsound from their point of view, and that unemployment is an integral part of the “normal” capitalist system.
power structure, not efficiency
But when we focus on the residuals for each group we can see that, in general, the bottom third has the residual closest to zero.
not the most convincing visually
the population that sees the biggest difference between predicted and actual sentiment is the upper- and middle-classes; not the poor.
fun fact…
Before the pandemic, actual and predicted sentiment moved in parallel. But as unemployment and inflation normalized, the predicted sentiment diverged.
tiktok! (joking)
Bolhuis et. al The Cost of Money is Part of the Cost of Living replicated and extended his work, showing that a 1983 change in how inflation was calculated altered the historical relationship between inflation and consumer sentiment. As such, pre-1983 inflation data (which is to say, most of the years that experienced any high inflation at all) is an unreliable guide for predicting contemporary consumer sentiment).
would be interesting to read!
Using the Carter-era “misery index” of unemployment+inflation, right now would be a relatively good (though not necessarily great) economy.
later it puts grains of salt on this
What is the Anthropocene in your experience? Has my exhibition taught you anything new or made you think differently? What are your takeaways? What is your favorite part of living in the Anthropocene? What do you think the world will look like in 100 years?
The Anthropocene, in my experience, is the accumulation of small, everyday choices. It's knowing the systems you rely on are harmful, but participating in them anyway.
The museum perspective made it feel eerie — like our present is already being archived and judged. It's helped my realization that we're not just studying the Anthropocene, but actively shaping it ourselves.
My main takeaway is that there's no outside to these systems. Even the reflections — through AI, media, or this project itself — are part of the same infrastructures it critiques.
My "favourite" part of living in the Anthropocene is the awareness. I think there's something meaningful about recognition, even if it doesn't always lead to resolution.
Unfortunately, I have no idea what the world will look like 100 years from now, and I would rather leave it that way. I don't like thinking too much about things that are out of my control.
he mirror from a site of judgment into a site of intention and love.
I really appreciate this possible alternative of turning from a world of judgment, from the way of looking at oneself with negativity, to the shifting of looking through that same object (the mirror) from a loving perspective. This feels like a wonderful way to repair and rewrite a fundamental question of the self in the Anthropocene.
object/subject relations in the Anthropocene.
The central tenet of this page is the object/subject duality that i had not thought about when creating a visualization of the Anthropocene. The way you articulate its importance is well-founded, and it has challenged my understanding of the Anthropocene. From my perspective as an environmental studies student, I had not thought about the possibility of subject/object dualisms with the self and how the mirror can serve as a vehicle for looking at oneself, but also of staring at oneself at the same time. This relationship is fundamental to the relational aspects of the Anthropocene and how everything is connected, even the self, through different mechanisms. In that exploration, it makes me think of the navigation of worlds between the human and non-human realms of beings and living.
DescriptionDetailsCitationsSource fileNo description available.Scalar URLhttps://scalar.usc.edu/works/a-close-and-personal-guide-to-the-anthropocene/mirror-girl-face-melting (version 1)Source URLhttps://i.pinimg.com/736x/19/40/c4/1940c4c5c92acfedd9a42f4d5ed795f4.jpg (image/JPEG)dcterms:titlemirror girl face meltingView asRDF-XML, RDF-JSON, or HTMLDescriptionDetailsCitationsSource fileNo description available.Scalar URLhttps://scalar.usc.edu/works/a-close-and-personal-guide-to-the-anthropocene/mirror-wiping-makeup (version 1)Source URLhttps://i.pinimg.com/736x/c7/64/d6/c764d61ad99747744ee70846fe8c45d1.jpg (image/JPEG)dcterms:titlemirror wiping makeupView asRDF-XML, RDF-JSON, or HTMLDescriptionDetailsCitationsSource fileNo description available.Scalar URLhttps://scalar.usc.edu/works/a-close-and-personal-guide-to-the-anthropocene/mirror-eye (version 1)Source URLhttps://i.pinimg.com/736x/02/2d/98/022d9879ddc4be6ae46e4f4d021e25fd.jpg (image/JPEG)dcterms:titlemirror EYEView asRDF-XML, RDF-JSON, or HTMLI’ve been thinking a lot about the concept of
The images that you have inserted along with your explanation of the poem, I think fit perfectly with the words you are saying. In one page, you managed to explore three different processes in a seamless manner, through creative writing, poetry and images. I am curious as to whether you created these images and how they play a role in your analysis of the poem as well.
The Looking Glass
I really appreciate how you incorporated poetry into your analysis of the Anthropocene. I think poetry gets at the connection between the human and the more-than-human that radical Western thought tries to remove itself from in the Anthropocene. Your poem is very emotional and thought-provoking and definitely achieves your goal of exploring the subject/object dichotomy.
Thomas and Rachel, our in-house historians, tackle the interview with L.U.M.E.N on this episode of The Curator's Cut to bring some more context to Madigan's environmental contributions. Listen now.
This podcast felt really eerie to me, largely due to the fact that it's AI technology discussing environmental impact. There's something almost oxymoronic about systems that rely on energy-intensive infrastructures reflecting on the damage those same systems contribute to.
On the other hand, streaming, often assumed to be the clean alternative, is not straightforward either. Music streaming's carbon cost is distributed across data centers, network infrastructure, and device manufacturing, collectively significant but invisible to the consumer. Studies suggested that the carbon footprint of streaming an album multiple times could exceed that of producing a physical copy, though this depends heavily on the energy source of the data centers involved.
This section really challenges my idea of "ethical consumption." I always assumed streaming was the more sustainable option, but this complicates that by showing how its environmental cost is just less visible. It feels like there's no clear solution — either you contribute to material production or to invisible digital infrastructures. I'm wondering if that's the point, but then it also leaves me unsure about what action is actually possible.
Mantener en buen estado la infraestructura vial es importante para evitar sobrecostos de operación en el transporte de personas y bienes. Un país que permita el deterioro de su infraestructura vial tendrá sobrecostos de operación vehicular en un rango que se puede ubicar entre 1 y 3% de su producto interno bruto nacional ( Bull, 2003 ). Además, la funcionalidad que debe ofrecer la red de carreteras de un país es crucial para la seguridad y comodidad de los usuarios
CONTEXTO-JUSTI
Un sistema de gestión de pavimentos basado en nuevas tecnologías para países en vía de desarrollo
TITULO
he "free-est" cow I've ever seen have been those that are free to graze in a fenced off pasture, typically close to a road. What does a truly free cow look like? And if all cattle were given the right to freedom, where would they go? We've bred them in such great numbers, is there enough quality habitat area on Earth for them? And if we were to release cows from human dominance, who else would we free consequentially?
This is a really good note to end on; it leaves the reader guessing. Personally, I'm intrigued, and this has me thinking of alternative scenarios regarding our treatment of animals.
What moral obligations do we have to cows?
This is such an interesting perspective. I really enjoy having to stop and think about what we owe to all these species we have destroyed for the sake of "humanity." It's something I haven't thought of before, but you're absolutely right, we do have a responsibility.
3. The Metabolocene
I really like adding your own "-cene", which connects to our class discussion about creating our alternative titles.
The degree to which modern economies are facilitated by simply the existence of the cattle industry cannot be overstated.
I did not realize just how much the cattle industry was involved in the modern economy until I read your speculative fiction about what the world would look like without cattle. It makes you wonder what we can actually change about the industry to ameliorate the planet, when any change could have major consequences.
However, development must be founded on sustainability centered growth, not towards technological prowess and advancement for the purposes of advancement
I completely agree, and, historically speaking, it seems that a big part of the Anthropocene was technological advancement and progress for progress's sake. Technological advancements must center on sustainability if we want to see any change.
took notes of the main lessons and thoughts going through my head as I tried to grapple with these fictional realities.
I also love when you incorporate your own voice, because there was a lot of setting up information, and it felt rewarding to get to the point where it's all connecting, and I'm seeing your thoughts.
Lessons:
This is purely for aesthetic reasons, but including background photos throughout the project would help avoid a grey backdrop. But this is just about playing around with Scalar's features; the actual content is great!
Lessons from Speculative Futures
I really enjoyed all your speculative futures because they helped me truly understand how important cows/cattle are to the modern world and how deeply implicated they are in the Anthropocene. It was a clever way to get me to really think about the information you've provided and then put my brain to work and see "ok, well if they're so important, what would happen if you took them away?" and then you can see how impactful they are on humans, the economy, and the environment.
Rhetoric, instead, can be defined as public discourse that creates meaning and moves communities.
This is a definition of rhetoric NOT the definition of rhetoric
Author context (determines optimal strategy). No institutional affiliation, no career or funding dependency, no active need to establish publication priority against parallel work. Goal 1 (per CLAUDE.md) is patient-directed evidence; Goal 2 is publishable hypothesis paper as a "nice to have." Strategy is calibrated to this profile — specifically NOT optimized for impact factor, career metrics, or priority racing.
Delete
The figure of an all-knowing, all-powerful scientist, whether as a villain or a hero, emphasizes individual and human-dominant solutions in the Anthropocene.
Drawing on our class discussions about masculinity and the danger of the hero trope in the Anthropocene.
create human clones to rip out their organs for money and power
Like the genetically modified pigs in Atwood's "Year of the Flood"!
Because they are all-powerful, they become capable of destroying the planet or humanity.
yes-- this trope marks an interesting shift from religious martyrs/saviors to scientific geniuses who serve similar purposes.
While it is, of course, necessary to rely on scientists to understand, survive and find ecological solutions in our geological epoch, it is also important to avoid mystification that feeds on the presupposed inadequacy of the people
This is why the environmental humanities are so important! They help prevent this mystification and knowledge gap.
A dominant narrative of the Anthropocene casts scientists as hero-like figures
highlighting the Geopower building block.
Amazed by the mountain’s beauty, the viewer wanted to access it, and it is these very movies that made that possible by showing them how to.
Yes this make me think about how we can approach environmental tourism from an ethical perspective...what regulations are necessary? Who has the "right" to visit vs. who does not? Does anyone deserve to more than others?
In other words, consider your audience, and what connects you and your curiosity, interests, and experience to a larger social issue or “motive” for change
love it!
remain open to new perspectives as you research and read, but also recognize and keep track of your stance as you research
it's so easy when your passionate about something and trying to spread valuable information to ignore other perspectives or facts that don't support your argument or research
4. Draft your Inquiry Question using “How,” “Why,” “In what ways,” or “To what extent,” in a way that elicits a conversation in which you will be interested in participating.
to what extent does financial instability limit opportunities for people in the working and lower class, and what can be done to help break those cycles?
3. Once you have your issue, consider your stance. What do you think about this issue? What is your initial opinion? This is your working thesis
my stance is that financial instability has a major impact on people’s opportunities, and it’s not just about individual effort. I think people in the working and lower class face challenges that make it harder to succeed, even if they are capable and motivated. because of this, I believe there should be more support and resources to help people build stability and break out of those cycles. this connects to my experiences and is why I want to be part of helping others improve their financial situations in the future.
2. Once you have a Topic, narrow that down to an issue. An issue is anything that can be debated from multiple sides. For example, starting with the topic of Capitalism, and narrowing that down to the issue of homelessness in San Francisco.
my topic is economic inequality, and I narrowed that down to the issue of how financial instability affects opportunities for people in the working and lower class. this is something that can be debated because some people believe success is based mostly on individual effort, while others argue that systems like income inequality, lack of resources, and limited access to education make it much harder for certain groups to succeed. this issue connects to my own experiences and also to larger systems that affect a lot of people.
1. Start by brainstorming various experiences you can recall related to school, or another large institution. The goal will be to locate your own experiences within larger systems, or be able to relate your own experience to the experiences of others like you.
some experiences I can think of related to school are feeling like I had to figure everything out on my own, especially when it came to assignments and expectations. I’ve also experienced balancing school with other responsibilities, which made it harder to stay consistent. another experience is feeling burnt out or unmotivated even when I knew I needed to do the work. I think a lot of this connects to larger systems because not every student has the same support, time, or resources, so people are coming into school with completely different situations. it also shows how things like financial stress or personal responsibilities can impact how someone performs in school, even if they are capable.
“In what ways does . . .” or “To what extent does . . .,” which open up these more basic questions to a larger audience of opinions and perspectives.
noted.
our goal is to make the conversation interesting enough that readers will want to get involved in it;
I find things that effect a large group people directly tends to be the most impactful, therefore more likely for readers to get involved
especially as you add new perspectives to your schema and your own knowledge and experience around your topic.
new perspectives and personal experiences can add so much impact to a piece of writing.
3. Choose one of these issues to discuss: Why or how would your research of this issue represent an example of “intrinsic motivation” for you?
my research on economic injustice connects to intrinsic motivation because it’s something that directly relates to my life and experiences. I grew up without financial stability, so I understand how much that can affect someone’s stress, opportunities, and overall quality of life. because of that, I’m not just researching it for an assignment, but because I genuinely care about the issue and want to be part of changing it. my goal is to help people in the working and lower class build stability so they don’t have to go through the same struggles or continue that struggle through generations
2. Why are these issues important to you?
I need to help people and children that have lived similar expierences. it's not fair to grow up without the same oppuntities as your peers just because you were born in the wrong place, wrong time, or from the wrong people.
1. What are some personal experiences you have that relate to larger social issues?
I grew up poor, even though my parents worked so incredibly hard to provide for me, it seemed so unfair to see other poeple doing a billion times less in life and get "rewarded" so much for possibly having like one good idea, one time. my parents were robbed of so many great oppurtunites because of societal norms and the economy.
Reading and writing became empowering because they gave these students a written voice for their struggles,
love to hear it, I feel like a lot of like very privledged people, like myself, forgets the importance and impact reading writing truly has and has always had.
reading is more about satisfying your personal curiosities about life’s conundrums
I feel like as an assignment the readings are rarely things I find truly interesting. Not in an egotistical way, but I feel like it's a lot of common sense related readings, or it's something I've already educated myself on so it seem silly to go over it again in a way.
your reading and writing process are integral to meaningful research.
YES, I feel like a lot of people forget how intertwined they are
del espectador
de las personas espectadoras
La seguridad emocional y física del grupo es prioritaria: cuidad el entorno, definid límites claros y estableced protocolos de respeto. Es importante evitar la espectacularización: la performance no busca entretener, sino activar pensamiento y sensibilidad crítica. Escoged espacios con significados o conflictos relevantes para las personas participantes. Podéis combinar este testeo con herramientas como el mapeo colectivo, entrevistas en movimiento o cartografías afectivas. No subestiméis el valor del «juego serio» (Turner, 1982); las acciones lúdicas pueden generar aprendizajes potentes. Documentar el proceso —no solo el resultado— es clave para el análisis posterior.
siempre el verbo en infinitivo
108
es 109
«la intervención urbana sería la acción o conjunto de acciones en el espacio público para tomar parte en un conflicto y convertirse en herramienta para la toma de conciencia».
referencia ( Rosas, 2015, p. 110)
Therefore, does this director actually propose alternative narratives and new ways to film the Anthropocene, or is it only an attempt at it that remains enclosed in mainstream, Western cinematic workings?
Perhaps he is negotiating a compromise between the two...so that he can still have the funds to make inroads in slow cinema.
As spectators, we are constrained to contemplate the "action-deficient scene as a form of violence" (McMenanim 94) against the tree.
I wonder about how effective 'slow cinema' is across age groups...I feel like many Gen Z viewers would get bored, look at their phone, and the technique and consequent message would go over their head...perhaps older generations would engage with it more?
it rejects the idea of one global and blameful humanity by filming local destruction.
Challenging Global Humanity building block through portrayal of an individual's narrative/daily life.
‘Back then, man and beast lived in harmony, but as time went by, most of the great forests were destroyed. Those that remained were guarded by gigantic beasts who owed their allegiance to the great forest spirit. For those were the days of gods, and of demons.’
Wow-- super interesting how the subtitles reframe a message through a more anthropocentric/hierarchical lens.
offer the perspectives of non-human entities.
offering views from below-- through non-human perspectives and imaginative kid perspectives! I also love Ghibli :)
investigadores
academia
los
quitar
soluciones
respuestas
Antes de iniciar el taller, aseguraos de que las personas participantes compartan una comprensión común del problema a abordar. Un diagnóstico previo y compartido alinea expectativas y facilita el trabajo colectivo. Asegurad y promoved los principios fundamentales de los procesos de cocreación: La inteligencia colectiva valora la diversidad de conocimientos y experiencias de todas las personas participantes. La empatía y la ausencia de juicio generan un espacio seguro y respetuoso, donde las personas puedan expresarse con libertad. La colaboración, donde las ideas se construyen colectivamente en lugar de competir. Distribuid papeles facilitadores (como relatoría, dinamización o registro) entre las personas participantes para favorecer el equilibrio en las intervenciones y reforzar el sentido de corresponsabilidad. Fomentad la innovación mediante técnicas que estimulen la creatividad, el diálogo abierto y la escucha activa. Estas condiciones permiten avanzar hacia propuestas transformadoras y conectadas con las realidades de las personas implicadas.
los verbos en infinitivo: asegurar; distribuir, etc
los
las personas
soluciones
respuestas
soluciones
respuestas
solucione
respuestas
a los actores interesados y
quitar
distintos agentes
diferentes actores
de los agentes y personas
de diferentes actores
agentes
actores
Just like these movies, the planet’s story does not have a main plot happening at one precise moment in time, but is constituted of a multitude of times, people and narratives.
Great articulation of how you are disrupting the Global Humanity and History as Stages building blocks through film.
They indeed create an active audience, one that has to think about the story being told. It teaches us to go from being a passive spectator to an intellectually active agent: the films do not resolve themselves without their audience.
Similar to Staurt Hall's theory of active spectatorship!
shake his audience, surprise them, and lead them in the wrong direction. This goal of his has mostly been understood through his shocking display of violence, but here it is also clear in the unusual narrative structure
Great parallel to B + F's Shock of the Anthropocene. It reminds me of our class discussion on the power of the "shock factor" within Anthropocene discourse.
In this sense, places like Pollution Park are not accidental or isolated; they are produced through global systems in which certain environments absorb the material consequences of others.
You do a good job of driving your points home. This sentence is important.
On islands like Caye Caulker, these dynamics become especially visible. Limited waste infrastructure, combine
I really like your analysis and use of theory in this paragraph.
Growth is always ongoing!
In a world that can feel overwhelming, your link to the next section is hopeful, a mindset that is very needed to create lasting change.
I see this page as mainly reading/interpreting because it reshapes how we understand the environment rather than proposing direct action. The idea of “navigating” instead of controlling the environment challenges my assumption that humans can manage ecological systems. This is difficult for me because it replaces certainty with uncertainty and makes environmental responsibility feel less clear.
This page made me reconsider whether environmental education should focus less on “solutions” and more on learning how to exist within uncertainty. That idea is uncomfortable because it suggests we may never fully resolve environmental crises. As we’ve explored in class, Anthropocene thinking often rejects simple “fixes” and instead emphasizes complexity, unpredictability, and ongoing adaptations.
This page challenged my understanding of the Anthropocene by shifting from seeing the environment as something humans manage to something we must navigate inside. I’m so used to thinking of humans as decision-makers acting on nature, but this page suggests that humans are just one part of a larger system. This page tackles a difficult topic but does so adeptly as it destabilizes the idea of human control and responsibility.
Pollution Park into the landscape
This name is witty and ironic. It exemplifies your creativity and points to the deeply sad state of our current environments.
What seems temporary often lasts far longer than expected.
This section does a really good job at showing the sad reality of our world today. The personal concrete examples really drives human activity and its role in contributing to the Anthropocene home.
Both pages (this and Navigating the Environment) focus strongly on reading/interpreting, but The “Local” Species goes further by beginning to hack/undo traditional ideas.
This page made me realize that the Anthropocene collapses the distinction between global and local. Local environments are no longer isolated. They are shaped by global systems like climate change, trade, and urbanization. Anthropocene thinking emphasizes that global processes are experienced through local effects, making scale more complex and harder to separate
Native to everywhere. Leaving nowhere.
A major strength of this page is its originality in questioning something that seems obvious: what counts as “local?” It connects strongly to Anthropocene themes of scale and mobility, showing how global processes reshape local environments.
Referencias
añadir: Añadir: Batthyány, K., Cabrera, M., Alesina, L., Bertoni, M., Mascheroni, P., Moreira, N., ... & Rojo, V. (2011). Metodología de la investigación para las ciencias sociales: apuntes para un curso inicial. https://repositorio.minedu.gob.pe/handle/20.500.12799/4544
Referencias
añadir : Añadir: Batthyány, K., Cabrera, M., Alesina, L., Bertoni, M., Mascheroni, P., Moreira, N., ... & Rojo, V. (2011). Metodología de la investigación para las ciencias sociales: apuntes para un curso inicial. https://repositorio.minedu.gob.pe/handle/20.500.12799/4544
los investigadores
las personas investigadoras
los
quietar "los"
Adecuad la estrategia al contexto: antes de iniciar, delimitad con claridad los objetivos de la observación y adaptad el nivel de inmersión y modalidad (estructurada o no estructurada) al tipo de información que deseáis obtener. Optimizad los tiempos y recursos: programad adecuadamente las sesiones de observación, priorizando los momentos del día o eventos clave que permitan captar con mayor riqueza las dinámicas sociales y espaciales del entorno. Minimizad los sesgos metodológicos: reflexionad sobre vuestra posición como observador u observadora. Reconocer vuestros referentes, expectativas y relaciones con el grupo observado es fundamental para interpretar los datos con mayor objetividad. Mantened una actitud ética y respetuosa: la observación participante implica convivir en entornos reales. Asegurad un comportamiento sensible, sin juicio y respetuoso con las dinámicas culturales, las normas sociales y los ritmos del grupo. Documentad de forma sistemática: utilizad cuadernos de campo, grabadoras, mapas o esquemas para registrar vuestras observaciones. Anotar lo antes posible tras cada sesión os ayudará a evitar la pérdida de matices o detalles relevantes. Contrastad y validad lo observado: siempre que sea posible, compartid vuestras interpretaciones preliminares con otras personas del equipo o incluso con las comunidades participantes, para reforzar la validez del análisis.
el verbo en infinitivo
intermediarios
intermediaciones
observadores
participantes
del observador
de la persona que realiza la observación
a los
quitar "a los"
los
quitar "los"
Referencias
Añadir: Batthyány, K., Cabrera, M., Alesina, L., Bertoni, M., Mascheroni, P., Moreira, N., ... & Rojo, V. (2011). Metodología de la investigación para las ciencias sociales: apuntes para un curso inicial. https://repositorio.minedu.gob.pe/handle/20.500.12799/4544
«Todos aquí pensamos que X, pero difieren en Y»)
Todas las personas participantes pensamos X pero diferimos en Y"
(líderes naturales, participantes reticentes, mediadores espontáneos)
(liderazgos naturales, participaciones reticentes, mediaciones espontáneas)
los
las personas
Elegid una persona que modere con habilidades y competencias para que afloren conversaciones de las personas sin imponer su opinión, con capacidad para fomentar un buen ambiente, intervenir solo cuando sea necesario. Seleccionad y preparad bien a las personas participantes: hay que cerciorarse de que cumplen los perfiles adecuados y que tienen experiencia en los temas que se tratarán en la entrevista. Tamaño del grupo: se recomienda entre seis y diez personas. Para temas sensibles o complejos, los grupos pequeños (3-5) favorecen la participación activa. Para temáticas más generales, pueden incluirse hasta quince personas, teniendo en cuenta los desafíos de moderación. Es útil prever un 20-30 % de participantes adicionales para cubrir posibles ausencias. Elección del espacio: el entorno debe ser cómodo, silencioso, accesible y libre de distracciones (Fàbregues y Paré, 2010). Disponer a las personas en círculo favorece la interacción y la equidad en el uso de la palabra. Es recomendable que el lugar resulte familiar o cercano para las personas participantes y que facilite un ambiente de confianza (Stewart y Shamdasani, 1990). Incentivos y motivación: para fomentar la participación activa y comprometida, puede ser útil ofrecer refrigerios, tarjetas regalo, materiales simbólicos o compartir los resultados del encuentro. Esto ayuda a crear un clima más relajado y colaborativo. Fomentad la participación de las personas, evitando que solo unas intervengan, manejar los conflictos o diferencias de opinión con sensibilidad, reafirmar el valor de todas las opiniones para evitar jerarquías o silenciamientos.
verbo en infinitivo
los
las personas
College research should be driven by open-ended questions that seek to expose real issues or problems occurring in the daily lives of a particular group of people or populations
love love love love love this
Version 3 of this preprint has been peer-reviewed and recommended by Peer Community in Neuroscience.<br /> See the peer reviews and the recommendation.
What it does mean is that we insist on our inherited right to insure our own safety, individually and as a race, to secure the future of our children, to maintain and develop our racial heritage in our own, white, Protestant, American way, without interference. . . .
But that's the exact issue, you don't own that! Yet, when I say this, you'll take it as an attack on your religion now. Which makes it impossible to talk any sense into you! >:O
Protestantism
This would be how you brainwash people. Nothing against the religion or practice itself. But, how the speaker is presenting this is using Protestant as a way to indoctrinate and trap White Americans into only believing in their white supremacy. Do not look into other beliefs as that will go against their religion. :<
The third of the Klan principles is that Protestantism must be supreme; that Rome shall not rule America.
Where the hell did Rome come from???? Like, how are they at all related to this conversation. Rome already fell, why would Americans be afraid of them???????????
This is not pretty and certainly disagrees with the maudlin theories of cosmopolitanism, but it is truth. The world has been so made that each race must fight for its life, must conquer, accept slavery or die. The Klansman believes that the whites will not become slaves, and he does not intend to die before his time. . . .
I see the motivation for why they believe that white must take over the world. They think that this is a race war. That only one race will conquer the world and subjugate everyone in the process and White People MUST take up arms together against this threat.
It reminds me a lot about the Russia and American space race. Where each group were competing with each other, even though they should have been working together... pretty petty if you ask me.
Identificad previamente la situación problemática. Tened claro el objetivo por el que se hace el análisis DAFO. Estableced dimensiones comunes a los cuatro factores. Tened una visión general previa del ámbito objeto de estudio. Utilizad datos de población desagregados por género, edad, etnia, etc. En el DAFO participativo: Asegurad la diversidad de perfiles participantes. Cread un ambiente de confianza y escucha activa. Utilizad dinámicas inclusivas y accesibles. Documentad y devolved los resultados a las personas participantes.
verbo en infinitivo
los
Quirtar los
los
Quitar actores
Manual de metodologías participativas Este manual recopila técnicas y enfoques aplicados en procesos de investigación acción participativa (IAP) en contextos comunitarios, institucionales y territoriales. Aporta herramientas para el análisis relacional y diagnóstico colectivo, incluyendo el sociograma como técnica clave para identificar actores, vínculos, posiciones y conflictos en una comunidad.
quitar este recurso
Diseñad el recorrido y realizadlo previamente a la sesión programada para verificar tiempos, espacios, etc. Adaptad los materiales y las consignas al grupo de trabajo. Clarificad y comunicad los objetivos al grupo desde el comienzo de la sesión. Facilitad y garantizad la horizontalidad en la exposición y reflexión colectiva. Tened en cuenta la accesibilidad y el confort climático según la condición y las necesidades de las personas convocadas.
usar el verbo en infinitivo: Diseñar; Adaptar; Clarificar; etc
Seminal studies in D. melanogasterdemonstrated that shadow enhancers improve theprecision of gene expression and phenotypic robustnessduring animal development, especially under condi-tions of physiological or genetic stress2
Perhaps help with stressful situations?
cis-regulatory elements
CRE's
Play99 is an online platform designed for cricket fans to follow major events such as IPL 2026, T20 World Cup, and other international matches.
The Twilight Saga is a series of romantic vampire fantasy films based on the book series Twilight by Stephenie Meyer. The series has grossed over $3.36 billion worldwide. The first installment, Twilight, was released on November 21, 2008.[1] The second installment, New Moon, followed on November 20, 2009.[2] The third installment, Eclipse, was released on June 30, 2010.[3][4] The fourth installment, Breaking Dawn – Part 1, was released on November 18, 2011, while the fifth and final installment, Breaking Dawn – Part 2, was released on November 16, 2012.[
sadi galli aajjjaaaaaa by ayushmann khurana
hiten is ordering from blinkit (https://www.reddit.com/r/twilight/comments/kr07u4/what_was_it_about_twilight/)
@cookie13
eLife Assessment
This potentially valuable manuscript focuses on the phosphorylation of residue T495 as a mechanism to inactivate HSP70 and disrupt cell cycle progression in response to DNA damage. The evidence supporting this model is solid, but would be significantly strengthened by additional studies defining the extent of T495 phosphorylation induced by DNA damage, identifying the kinase responsible for phosphorylating T495 of HSP70, and further elucidation of the functional implications of T495 phosphorylation in human cells. This work will be of interest to scientists focused on topics including chaperone biology, proteostasis, cell cycle progression, and DNA damage.
Reviewer #1 (Public review):
Summary:
This study identifies a conserved phosphorylation event on Hsp70, at human T495 that is triggered by DNA damage. The authors show that this modification arises in response to MMS and is temporally associated with cell cycle progression through mitosis. Using biochemical analysis, they further argue that the phosphomimetic Hsc70(T495E) adopts an open-like conformation with impaired J protein-stimulated ATP hydrolysis while still retaining client binding. In yeast, both phosphomimetic and phosphonull mutants perturb growth and cell cycle progression, supporting the idea that dynamic regulation of this site helps coordinate DNA damage responses with G1/S control.
Strengths:
A major strength of the paper is that it links prior work on Legionella-mediated Hsp70 phosphorylation to a normal cellular DNA damage response. The study is also commendably multi-level, combining mammalian cell biology, in vitro biochemistry, and yeast genetics to support the central model. Together, the authors provide a coherent story that this Hsp70 site has functional importance in checkpoint-like control rather than being a passive phosphosite, adding to our understanding of the chaperone code.
Minor Weaknesses:
The authors acknowledge that the direct kinases/phosphatases for this site remain unknown. Some conclusions are therefore still somewhat inferential, especially the model that pHsp70 acts as a reversible molecular brake on S-phase entry. These limitations do not undermine the importance of these exciting findings, but they do leave the paper somewhat short of a fully resolved mechanism.
Comments on revisions:
The authors have done a great job in addressing all the previous reviewer concerns. They have provided additional data and refined the text, stating limitations of their proposed model. In doing so, they have produced a much-improved version of the manuscript.
Reviewer #2 (Public review):
The revised manuscript offers little new information and fails to address the critical weaknesses identified in the original submission.
While we can agree that phosphorylation of Thr495 would likely affect Hsp70 function-given the known biochemistry of Hsp70s and the author's previous work on LegK4-the significance of this finding hinges on whether it is a regulated process. If a meaningful fraction of Hsp70 were phosphorylated in a regulated manner triggered by DNA damage or cell cycle progression, it would constitute an important discovery, regardless of its specific impact on fitness in a given context.
However, beyond highlighting the temporal profile of Hsp70 phosphorylation in MMS-treated cells (Figure 4e), the paper fails to rule out the possibility that this correlation is merely an irrelevant side reaction. This "bystander" phosphorylation could simply be caused by the activation of kinases during the experimental MMS treatment and subsequent washout. The authors' claim-that the fraction of phosphorylated Hsp70 increases in a "regulated, cell-cycle dependent manner"-does not sufficiently counter the possibility of it being a non-functional side effect.
This concern could be resolved if the authors had identified the specific kinase, demonstrated its specificity, and manipulated it either genetically or pharmacologically. While I acknowledge this is a "tall order," the lack of such data limits the paper's significance. Furthermore, the current data fails to meet a much lower bar: confirming that a substantial fraction of Hsp70 is actually phosphorylated under the tested conditions. Such a finding would at least suggest the event is capable of impacting the overall Hsp70 pool.
It is surprising that the authors have not provided a ratiometric assay to settle this, such as an immunoblot of total Hsp70 separated on a Phos-tag or IEF gel. Instead, they rely on indirect evidence and data subject to alternative interpretations. Specifically, they argue that the fitness cost of the Thr495Ala mutation (or the phosphomimetic mutation) is due to the loss of regulatory phosphorylation (or deregulated phosphorylation); however, it is equally plausible that the mutations create Hsp70 hypomorphs whose defects are only exposed under stressful experimental conditions.
Reviewer #3 (Public review):
In this manuscript Moss et al. demonstrate that Hsp70 phosphorylation at a conserved threonine residue integrates DNA damage responses with cell-cycle control. The authors present unbiased biochemical, cell-based, and yeast genetic analyses showing that phosphorylation of human Hsp70 at T495 (and the analogous Ssa1 T492 in yeast) is triggered by base-excision-repair intermediates and downstream DDR kinase activity, leading to delayed G1/S progression after DNA damage. They used orthogonal approaches such as ATPase assays, phospho-specific detection, kinase-inhibition studies, synchronization experiments, and phenotypic analyses of phosphomutants. They presented robust data which collectively supported the conclusion that dynamic Hsp70 phosphorylation functions as a conserved "molecular brake" to prevent inappropriate S-phase entry under genotoxic stress.
Comments on revisions:
The authors have addressed all my questions and concerns.
Author response:
The following is the authors’ response to the original reviews
We thank the reviewers for their time and consideration of the manuscript. We have added new data to Figure 5 (Figure 5a) to address concerns regarding the conservation of the Hsp70 phosphorylation in yeast. Additionally, we have changed the title of the manuscript to “Hsp70 is phosphorylated in a conserved response to DNA damage and contributes to cell cycle control” to more accurately represent the conclusions we draw.
Public Reviews:
Reviewer #1 (Public review):
The strength of evidence of the mechanistic and "conserved checkpoint" claims that this site is directly activated by DNA damage is inadequate and fundamentally incorrect.
We respectfully disagree with the reviewer’s characterization of our conclusions. Our data demonstrate that DNA damage induces this phosphorylation in a cell-cycle–dependent manner. We do not claim to have defined the direct kinase or full mechanistic pathway; rather, we establish that site activation is damage-responsive and functionally linked to cell-cycle regulation. Consistent with this, phospho-mutants in yeast exhibit clear cell-cycle defects, supporting a conserved functional role. We address each of the reviewer’s specific concerns below.
Specific comments:
(1) Activation of T495:
The author's premise for the site being activated by DNA damage is Albuquerque et al, where PTMs on MMS treated yeast are analyzed. T492 (the yeast equivalent of human T495) is observed as phosphorylated. However, the authors fail to note that there is no untreated sample analysis in this study, and it is likely that T492 phosphorylation is also present in untreated cells. This is also backed up by later evidence from the same lab (Smolka et al), where they do not identify T492 as being dependent on Mec1/Tel/Rad53 kinases.
We agree with this assessment of the Albuquerque study. Accordingly, we used their data to generate the hypothesis that this site is phosphorylated, and we took it upon ourselves to more rigorously demonstrate phosphorylation with appropriate controls. The validated antibody that we had previously generated[1] to track pHsp70 was the enabling technology to directly track this phosphorylation event. We now directly show phosphorylation of this site (Figure 5a, lines 276-284). Of note, as Reviewer 1 suggested, there is a smaller amount of pHsp70 in the untreated cells, which corresponds with findings from Holt et al 2009 [2]. This could reflect a baseline role of Hsp70 phosphorylation for normal growth that is accentuated upon MMS insult.
(2) The kinase(s) directly responsible for T495 phosphorylation are not identified. Instead, the authors show that knockdown or pharmacological inhibition of DNA-PKcs, ATM, Chk2, and CK1 attenuate pHsp70.
We agree with reviewer 1 that identifying the direct kinase would be an exciting finding, and we believe our manuscript will provide the foundation for future studies to address these questions. While these findings will be impactful, we do not believe their lack detracts from the observations we have made.
(3) ATM siRNA knockdown has no effect, while ATM inhibitors do, which the authors acknowledge but do not resolve. This discrepancy raises concerns about off-target drug effects.
We agree with reviewer 1 that off-target drug effects are always a concern when employing pharmacological inhibitors. To that end, we tested structurally distinct inhibitors of ATM (Figure 3b) to decrease the likelihood of the same off target effect. While complementing this with a genetic knockdown would be ideal, the discrepancies between pharmacological and genetic inhibition of ATM have been well reported (lines 214-216).[3,4] Parallel discrepancies in other kinases have been mechanistically explored by other groups.[5] The preponderance of pharmacological evidence in conjunction with RNAi suggests the most likely interpretation of our data is that ATM is involved in signaling upstream of Hsp70 phosphorylation. Thus, our data compel future work to use more sophisticated genetic methods to more specifically determine how ATM connects with pHsc70.
(4) No in vitro kinase assays, motif analysis, or phosphosite mapping confirming these kinases as direct T495 kinases are presented. Thus, the proposed signaling cascade remains speculative.
We agree that we should carefully circumscribe our conclusions about the potential signaling cascade. To communicate our conclusions more clearly, we rewrote lines 223-226 to highlight that our findings implicate these kinases in upstream signaling rather than direct phosphorylation of Hsp70.
(5) Smolka and many other labs characterized DDR sites as SQ/TQ motifs, and T492 doesn't fit that motif.
We agree, and our response to comment 4 addresses this point. Briefly, we do not claim that Hsp70 is a direct target for DDR. Notably, the SQ/TQ motifs mentioned specifically pertain to ATM and DNA-PK[6], though we would like to note several studies have demonstrated DNA-PK phosphorylation outside of these motifs.[7] Chk2 and CK1 do not prefer SQ/TQ motifs[9]. Additionally, Chk2 is known to phosphorylate non-consensus sequences as well[10].
(6) No genetic tests in yeast (e.g., BER mutants) are used to connect Ssa1 T492 phosphorylation to BER in that system, despite the strong BER-centric model.
We agree that it would be interesting to study BER mutants in yeast, and we believe this will be an exciting prospect for future studies to better establish the signaling cascade. We have included a Western blot (Figure 5a) showing that MMS treatment causes increased Hsp70 phosphorylation in yeast. MMS damage is repaired through BER in S. cerevisiae,[11] and the pathway itself is highly conserved.[12] Our experiments demonstrate that the phosphorylation of Hsp70 occurs as a conserved response to alkylation damage, which is the major conclusion of our paper.
(7) Overexpression of MPG gives only a modest increase in pHsp70, while APE1 overexpression has no effect, and Polβ overexpression does not decrease pHsp70. These mixed results weaken the central claim that Hsp70 phosphorylation is a tuned sensor of BER burden.
We appreciate this incisive question. Though not immediately intuitive, we do not believe these results are necessarily ‘mixed’. The lack of APE1 over-expression having an effect could be attributed to APE1 activity being necessary for the phosphorylation, but not rate-limiting. Regarding Polβ, it is important to note that not its binding, but rather its dRP lyase activity is rate-limiting in base excision repair.[13] As such, if binding sites are already saturated or near saturated, but the lyase activity remains slow, we may not observe a decrease in BER intermediates. While we do claim that phosphorylation of Hsp70 is triggered by BER intermediates (lines 193-194), we do not claim that pHsp70 is a tuned sensor of BER burden.
(8) A major concern is that pHsp70 is only convincingly detected after very high, prolonged MMS (10 mM, 5 h) or 0.5 mM arsenite treatments. Other DNA-damaging agents (bleomycin, camptothecin, hydroxyurea) that robustly activate DDR kinases do not induce pHsp70. This suggests to me that the authors are observing a side effect of proteotoxic stress. This is likely (see Paull et al, PMID: 34116476).
Our data indicate that pHsp70 specifically occurs downstream of base excision repair. Therefore, it is not surprising that drugs that do not activate BER (bleomycin, camptothecin, hydroxyurea) do not elicit the same response. While pHsp70 may arise due to DSBs generated through BER, the fact we do not see phosphorylation after bleomycin treatment could be explained by the cell-cycle dependencies we report (Figure 4e). It is also important to note that MMS-induced pHsp70 occurs primarily in the nucleus, and Western blots of whole cell lysate will contain large amounts of cytosolic Hsp70 that could dilute the signal. Indeed, in our nuclear extraction (Figure 4d), we see faint pHsp70 signal as soon as 1 h after treatment, though it increases in robustness as the time-course progresses. These data are both concordant with a model in which high BER-induced lesion burden in mitosis leads to Hsp70 phosphorylation in late M/G1.
We would like to add that, in the review article cited by Reviewer 1, the authors specifically cite studies implicating a loss-of-function in DDR pathways leading to increased proteotoxic stress (e.g. ATM deficient cells producing higher levels of aggregated proteins compared to WT). However, we find that inhibition of DDR kinases decreases, rather than increases Hsp70 phosphorylation. We thus believe that DNA damage rather than proteotoxic stress is the parsimonious cause of Hsp70 phosphorylation.
(9) A recent study in Nature Communications (Omkar et al., 2025) demonstrates rapid phosphorylation of yeast T492 in a pkc1-dependent manner, diminishing the impact of these findings.
We were excited to see this paper when it was published 3 months after we posted a preprint on bioRxiv, which was released three weeks after our submission to eLife. Rather than diminishing the impact of this paper, we believe that independent lines of evidence from different groups mutually reinforces the impact of the work. We have added a sentence to say that during the review of our work, this group independently observed this phosphorylation event in response to a different stress (lines 421-423). We believe in celebrating the scientific process arriving at consistent results, and the editorial policies of eLife reinforce that philosophy by offering ‘scoop protection.’
We would also like to highlight several differences between the scope of our papers. The phosphorylation reported by Omkar et al. appears highly constrained to yeast as part of the Cell Wall Integrity pathway, whereas ours occurs as a more highly conserved response. Additionally, our paper provides additional biochemical insight into the consequences of this phosphorylation, which is lacking in Omkar et al. If anything, this paper highlights the important regulatory capacity of this residue on Hsp70, and suggests it may serve multiple functions in the cell.
(2) Downstream Effects of T492/T495:
(10) The manuscript's central conceptual advance is that pHsp70 is a cell-cycle-regulated brake on G1/S. Yet in mammalian cells, the authors show only that pHsp70 appears late, after cells have traversed mitosis, and that blocking CDK1 (G2/M) prevents its accumulation.
We would like to clarify the central contribution of this study. Prior work identified this phosphorylation in yeast, but its existence and conservation in human cells had not been established. A primary advance of our study is demonstrating that this site is phosphorylated in mammalian cells and that its accumulation is cell-cycle regulated — coinciding with late M/G1.
We further show that phosphorylation depends on cell-cycle progression, as CDK1 inhibition prevents its accumulation. While these data establish regulation, we agree that they do not by themselves define causality in mammalian cells. To address functional consequences, we leveraged the genetic tractability of S. cerevisiae. Phosphomimetic Ssa1 T492E increases the proportion of G1 cells in the absence of MMS and enforces a stronger G1 arrest following MMS treatment. Together, these findings support a conserved, cell-cycle–linked role for this phosphorylation and provide a foundation for future mechanistic work in mammalian systems.
(11) There is no functional test in human cells: no knockdown/rescue experiments with T495A or T495E, no cell-cycle profiling upon altering Hsp70 phosphorylation state, and no demonstration that pHsp70 actually causes any delay in S-phase entry, rather than simply correlating with late damage responses. The strong conclusion that pT495 "stalls cell cycle progression" (e.g., Figure 6 model) is therefore not supported in the human system.
We agree that we did not directly test the functional consequences of Hsp70 phosphorylation in human cells. Our intent was not to claim that we have demonstrated causality in the mammalian system, but rather to establish that this conserved phosphorylation exists in human cells and is cell-cycle regulated.
We instead used S. cerevisiae to interrogate this due to its increased genetic tractability. In this system, phosphomimetic mutation increases the proportion of G1 cells under basal conditions and enhances G1 arrest following MMS treatment, mirroring the damage-associated phenotype observed in human cells. These findings support a conserved functional role for this modification, although we agree that direct mechanistic testing in mammalian cells will be important for future work.
While we intended the cartoon model to be a speculative illustration of what may be occurring in order to motivate future studies. We now see how this may lead to confusion, so to improve clarity, we have removed Figure 6 from the manuscript.
(12) All functional conclusions rely on T492A/E point mutants at the endogenous SSA1 locus, usually in an ssa2Δ background, in a family of highly redundant Hsp70s. Without showing that this site is actually modified during their MMS treatments, the assignment of phenotypes to loss of a physiological phospho-switch is premature. The authors need to repeat their studies in an Ssa1-4 background, as in https://pubmed.ncbi.nlm.nih.gov/32205407/.
Thank you for this feedback. We have included a Western blot to Figure 5 (Figure 5a) addressing this comment. Briefly, we show that, in yeast, Hsp70 phosphorylation increases upon MMS treatment and is not detectable in the point-mutants in the ssa2∆ background. The latter data suggest that Ssa3-4 modification is negligible in our system.
(13) The authors infer that T495E "locks" Hsc70 in a pseudo-open state based on reduced J-protein-stimulated ATPase activity, unchanged ATP binding, altered trypsin sensitivity, and retained tau binding. However, there is no direct comparison of phosphorylated vs T495E protein (e.g., via in vitro phosphorylation with LegK4 followed by side-by-side biochemical assays, or structural analysis). Thus, it remains unclear to what extent the glutamate substitution mimics a phosphate at this position.
Previously we did show that phosphorylation impacts the ATPase cycle of Hsp70.[1] In this paper, with the phosphomimetic mutant we see an even greater decrease of activity. This is consistent with incomplete phosphorylation yielded by in vitro phosphorylation with LegK4.[1] Due to this incomplete phosphorylation in vitro, we determined that the phosphomimetic mutant would be more useful for the assays we performed, as they rely on bulk readouts.
(14) No client release kinetics, co-chaperone binding assays, or in vivo chaperone function tests are provided, yet the discussion builds a detailed model of a "pseudo-open" state that simultaneously resembles ATP-bound conformation and allows persistent substrate engagement.
We have shown that the conformational cycle of Hsp70 (T495E) is uncoupled from nucleotide state, and that the overall conformation resembles ATP-bound Hsp70. This is consistent with prior studies on AMPylation of the same residue.[14] Additionally, we demonstrate that substrate engagement is similar between WT and T495E. This is consistent with our previously published work showing increased pHsp70 on polysomes,[1] as well as our observations that the phosphomimetic mutant in yeast exerts a phenotype even in the presence of the compensatory isoform SSA2. This dominant-like phenotype is consistent with those seen in mutations locking Hsp70 in a ‘closed’ conformation.[15] We agree that future studies examining client release kinetics and co-chaperone binding would be useful for future structural studies validating and elaborating on our findings.
Reviewer #2 (Public review):
Weaknesses:
The kinase(s) responsible for the phosphorylation have not been identified (and hence remain inaccessible to experimental i.e., genetic or pharmacological manipulation). The mechanistic links to DNA damage repair and the fitness benefits of this proposed adaptation remain obscure. Of greater concern, the data provided in the paper fail to exclude the trivial possibility that the phosphorylation event described (and characterized through biochemical proxies) is biologically neutral, reflecting nothing more than a bystander event in which kinase(s) activated by application of high concentrations of a powerful alkylating agent (MMS) phosphorylate, at meaninglessly low stoichiometry, an abundant protein (Hsp70) on a surface exposed residue. Failure to exclude this (plausible) scenario is this paper's weakness.
We agree that we have not directly quantified the absolute stoichiometry of Hsp70 phosphorylation. However, several lines of evidence argue against the interpretation that this represents a biologically neutral, bystander modification.
First, our pulse-chase experiment (Figure 4e) shows that, after MMS removal, pHsp70 levels increase as cells progress through the cell cycle. Notably, total Hsp70 levels remain constant. This indicates that the fraction of phosphorylated Hsp70 increases in a regulated, cell-cycle dependent manner, rather than through a bystander event during acute stress.
Second, functional perturbation of the homologous site in yeast produces phenotypic consequences. The phosphomimetic Ssa1(T492E) mutant exhibits reduced growth, increased G1 accumulation, and impaired cell-cycle re-entry following MMS treatment (Figure 5). These phenotypes argue that the modification of this residue is functionally consequential.
While the upstream kinase remains to be identified, the genetic and cell-cycle phenotypes observed upon site perturbation argue that this phosphorylation is functionally consequential.
Reviewer #2 (Recommendations for the authors):
(1) The biochemical characterization of the phosphomimetic mutation (T495E) is thorough, relying on ATPase assays and conformational analysis. Figure 1b demonstrates reduced J-protein-stimulated ATPase activity, and Figure 1d shows an ATP-like proteolysis pattern consistent with an open conformation. As the authors are well aware, Hsp70 chaperones act on their substrates via a dynamic cycle that includes binding, ATP hydrolysis, and conformational shifts. One wonders, therefore, at the relevance of the measurement shown in Figure 1f. While it is highly plausible that the T495E mutation mimics the phosphorylation event (BiP T518E mimics key aspects of AMPylation), the lack of a biochemical characterisation of Hsp70 with pThr495 is an important limitation of this paper. Even if such a preparation cannot be accomplished with the endogenous kinase(s) whose identity remains unknown, a characterisation of LegK4-phosphorylated Hsp70 should suffice.
We agree with Reviewer 2 that the rationale for figure 1f does not logically follow the results of 1b and 1d. Rather, this experiment was motivated by the prior findings that phosphorylation of Hsp70 by L.p. lead to an increase occupancy on polysomes[1] (lines 137-139). We sought to better understand the discrepancy between this finding and our own by assaying the capacity of the T495E mutant to bind substrate.
Reviewer 2 raises a valid point in that phosphomimetic proteins do not necessarily behave the same as truly phosphorylated proteins. Previous work from our lab characterized the ATPase activity and in vitro folding capacity of Hsc70 that had been directly phosphorylated by LegK4[1] (lines 114-115). We were motivated to turn to a phosphomimetic mutant as LegK4 only phosphorylates around half of the Hsc70 present in solution[1] (line 116); this mixture of species makes batch analysis difficult. As we had previously published with the in vitro phosphorylated Hsc70, we didn’t believe it necessary to include along with our future analyses.
(2) As noted, the kinase(s) that phosphorylate T495 remain to be identified and is inaccessible genetically. The phenotypic consequences of impaired pThr495 are therefore assessed by a T495A mutation. This most certainly eliminates phosphorylation at that site however, Figure 5C shows quite clearly that the T/A mutation is not neutral. This is expected, given the role of an H-bond network centered upon the homologous residue in the ADP-bound configuration of Hsp70's. Importantly, the biochemical non-neutrality of the T/A mutation also compromises the interpretation of the associated phenotype, as this cannot be attributed solely to a loss of phosphorylation; it may reflect features of the T/A mutations exposed by MMS, but unrelated to the inability of the residue to undergo regulated phosphorylation.
We appreciate this insightful critique. We agree that the alanine substitution may perturb the local H-bond network, and have added a sentence to our discussion to highlight this caveat (lines 379-381). That being said, our conclusions do not solely rely on the T to A mutant. The phenotypes observed in our phosphomimetic mutant overlap with the TA mutant (increased sensitivity to MMS; defects in cell cycle re-entry after MMS treatment) (Figure 5). While the alanine mutation may not represent a purely ‘loss-of-phosphorylation’ state, our findings do implicate the importance of this residue in cell cycle control after DNA damage.
(3) It thus remains formally possible that pThr495 arises as an irrelevant side reaction due to activation of a kinase (with other relevant substrates).
This dismal interpretation of the data would be dispelled somewhat if the stoichiometry of pThr495 were substantial, whereas very low stoichiometry of phosphorylation should leave one wary of the possibility that the surface-exposed Thr495 of ATP-bound Hsc70 is a physiologically irrelevant bystander target of a kinase activated in DNA-damaged cells.
We have included a Western blot in Figure 5 showing pHsp70 in our yeast samples. Here we can see low abundance of Hsp70 phosphorylation in untreated WT yeast, with a clear increase in MMS treated yeast. Additionally, as mentioned in a previous response, Figure 4e shows the accumulation of pHsp70 in human cells even after MMS removal, indicating it is not simply the byproduct of over-activation of the DNA damage response.
Unfortunately, the study does not quantify the stoichiometry of Hsp70 phosphorylation; detection relies on phospho-specific immunoblotting, leaving open the question of whether this modification occurs at physiologically significant levels. This worry is compounded by Figure 2a,f that suggests that phosphorylation occurs only under high-dose MMS or arsenite, raising concerns about physiological relevance.
We agree that we did not quantify absolute phosphorylation stoichiometry. While a precise measurement would be informative, our conclusions are based on regulated dynamics and functional perturbations rather than magnitude alone. Specifically, our pulse-chase (Figure 4e) shows that total Hsp70 levels remain constant while pHsp70 increases in a cell-cycle dependent manner following MMS removal. This indicates a regulated modification rather than a side-effect of kinase over-activation during acute stress. Additionally, perturbation of the homologous site produces cell-cycle phenotypes (Figure 5) in yeast, supporting functional relevance.
However, as mentioned in responses to Comment 3, our pulse-chase assay in Figure 4e indicates the stoichiometry of pHsp70 increases after MMS removal in a cell-cycle dependent manner. Furthermore, as discussed in response to Reviewer 1 Comment 8, Figure 4d highlights a technical limitation with regards to detection of pHsp70 by Western blotting. Namely, as pHsp70 accumulates in the nucleus, signal appears to be diluted by unmodified Hsp70 in the cytosol when whole-cell lysate is probed, thereby reducing detection capacity. It is therefore possible that less stringent doses do lead to phosphorylation, but due to the experiments being run in asynchronous cells and on whole cell lysate we failed to detect it.
Reviewer #3 (Recommendations for the authors):
Major Comments:
(1) Figure 1e - Which antibody was used to probe this blot?
Thank you for catching this omission. This was stained with Coomassie. We have edited the figure legend to reflect this.
(2) Figure 1c- Do the authors have the data of the WT and T495E with DJA2?
The assay was performed with increasing concentrations of DJA2 for both constructs (from 0 µM to 4 µM) (lines 118-119, Figure 1c).
(3) Figure 2- The labeling of the right side of the immunoblots is missing.
We apologize for the confusion. The labeling is on the left. The lines on the right are intended to demarcate blots that came from the same membrane (for easier comparison of loading controls).
(4) Figure 2d- Does MMS treatment lead to a heat shock response?
We have not directly tested this. However, we do not see the massive upregulation of HSPs that would be expected from a heat shock response.
(5) Figure 4c and e - Total protein level of some of the phospho-proteins is missing.
We used housekeeping proteins as loading control. We do not have antibodies for all the non-phospho proteins. For those we have, blots not included in the publication do not show any marked discrepancies between the non-phospho form and the housekeeping proteins.
(6) Figure S1A- Although the authors suggest that the phosphorylation event is reversible, they have not integrated it into the final model in Figure 6.
In line 403 we postulate that dephosphorylation may permit client release. In the interest of clarity, we have now removed the model figure.
(7) Yeast genotype is missing.
We used W303a yeast (line 612).
(8) It is unclear which phosphatase inhibitor was used in their assay (Figure S1A).
We repeated the experiment with both Halt Phosphatase Inhibitor Cocktail (Thermo Scientific 78440) and Roche PhosStop (Roche 04906837001) (lines 524-525).
(9) Please add this most recent and up-to-date reference (PMID: 40976416) related to your study.
We have now added that reference
(10) Can the authors speculate on whether Hsp70- T495E is expected to primarily reside in the nucleus?
We have no data to indicate whether or not phosphorylation at T495 or a phosphomimetic mutation in this site would directly affect nuclear import or export. In cells expressing the Legionella kinase LegK4, pHsp70 exists in the cytoplasm,[1] indicating the phosphorylation in of itself does not force nuclear localization. We thus imagine that the nuclear localization seen in Figure 4d is more likely due to the location of the kinase rather than as a consequence of the phosphorylation. In an over-expression system or in the case of a genomic mutation, we believe the protein is most likely to exist in both the cytoplasm and in the nucleus, though we did not directly test this.
References
(1) Moss, S. M. et al. A Legionella pneumophila Kinase Phosphorylates the Hsp70 Chaperone Family to Inhibit Eukaryotic Protein Synthesis. Cell Host Microbe 25, 454-462.e6 (2019).
(2) Holt, L. J. et al. Global Analysis of Cdk1 Substrate Phosphorylation Sites Provides Insights into Evolution. Science 325, 1682–1686 (2009).
(3) Choi, S., Gamper, A. M., White, J. S. & Bakkenist, C. J. Inhibition of ATM kinase activity does not phenocopy ATM protein disruption. Cell Cycle 9, 4052–4057 (2010).
(4) Menolfi, D. & Zha, S. ATM, ATR and DNA-PKcs kinases—the lessons from the mouse models: inhibition ≠ deletion. Cell Biosci. 10, 8 (2020).
(5) Weiss, W. A., Taylor, S. S. & Shokat, K. M. Recognizing and exploiting differences between RNAi and small-molecule inhibitors. Nat. Chem. Biol. 3, 739–744 (2007).
(6) Kim, S.-T., Lim, D.-S., Canman, C. E. & Kastan, M. B. Substrate Specificities and Identification of Putative Substrates of ATM Kinase Family Members*. J. Biol. Chem. 274, 37538–37543 (1999).
(7) Jette, N. & Lees-Miller, S. P. The DNA-dependent protein kinase: A multifunctional protein kinase with roles in DNA double strand break repair and mitosis. Prog. Biophys. Mol. Biol. 117, 194–205 (2015).
(8) O’Neill, T. et al. Determination of Substrate Motifs for Human Chk1 and hCds1/Chk2 by the Oriented Peptide Library Approach*. J. Biol. Chem. 277, 16102–16115 (2002).
(9) Fulcher, L. J. & Sapkota, G. P. Functions and regulation of the serine/threonine protein kinase CK1 family: moving beyond promiscuity. Biochem. J. 477, 4603–4621 (2020).
(10) Craig, A. et al. Allosteric effects mediate CHK2 phosphorylation of the p53 transactivation domain. EMBO Rep. 4, 787–792 (2003).
(11) Xiao, W., Chow, B. L. & Rathgeber, L. The repair of DNA methylation damage in Saccharomyces cerevisiae. Curr. Genet. 30, 461–468 (1996).
(12) Memisoglu, A. & Samson, L. Base excision repair in yeast and mammals. Mutat. Res.Fundam. Mol. Mech. Mutagen. 451, 39–51 (2000).
(13) Srivastava, D. K. et al. Mammalian Abasic Site Base Excision Repair IDENTIFICATION OF THE REACTION SEQUENCE AND RATE-DETERMINING STEPS*. J. Biol. Chem. 273, 21203–21209 (1998).
(14) Preissler, S., Rato, C., Perera, L. A., Saudek, V. & Ron, D. FICD acts bifunctionally to AMPylate and de-AMPylate the endoplasmic reticulum chaperone BiP. Nat. Struct. Mol. Biol. 24, 23–29 (2017).
(15) Fontaine, S. N. et al. Isoform-selective Genetic Inhibition of Constitutive Cytosolic Hsp70 Activity Promotes Client Tau Degradation Using an Altered Co-chaperone Complement*. J. Biol. Chem. 290, 13115–13127 (2015).
comment from 2026 yo
Ruelle (Not So) Verte
I appreciate this funny play on words and the satirizing notion you are using the critique even just the language we use to describe 'Green Spaces.'
Anthropocene, which often rely on visible, active, and continuous forms of environmental engagement. Montréal’s winter instead reveals that urban nature is uneven not only across space and social groups, but also across time
This is a good connection to our class content, where the Grand Narrative makes big generalizing claims, but even just looking at one city across time and seasons can look so different.
While maps suggest a stable distribution of green space, these representations do not account for the ways in which such spaces are experienced differently across seasons.
This is such an interesting point that I had never thought about before. This raises awareness to the discrepancy between the 'factual' 'scientific' and 'technological' presentations of a city through a map, as opposed to the real lived experiences of people in the city. This makes me think also about the weather app which declares an objective temperature and a temperature that it 'feels like,' but both of these can vary from what the weather truly feels like when you step outside and experience it with your body.
Starvation-induced autophagy occurs independently of the ATG1 complex in Chlamydomonas
Here, you find that autophagic flux proceeds to completion in Chlamydomonas lacking ATG1 complex components, even under acute TOR inhibition, where the canonical mechanism runs directly through ATG1 kinase activity. This is a compelling result worth engaging with carefully.
The gene identifications here rely on sequence similarity and phylogenetic analysis, which establish orthology but not functional equivalence. Have the Chlamydomonas ATG1, ATG11, ATG13, and ATG101 proteins been shown to physically interact and form a kinase-active complex? Without that, another interpretation is available: if the complex has diverged structurally or catalytically in C. reinhardtii, its dispensability may simply reflect that the canonical ATG1 kinase function was already absent. The elevated basal ATG8-PE in ATG1 complex knockouts suggests these proteins are doing something regulatory, but that's consistent with either reading.
Something else must be relaying the TOR signal to the ATG8 lipidation machinery if ATG1 is not. The finding that PI3K remains required suggests it's not a completely orthogonal pathway. Have you considered a systematic approach, such as phosphoproteomics under AZD8055, to identify what TOR is actually targeting in Chlamydomonas when ATG1 is absent? The partial reduction in flux in atg13 and atg11 mutants relative to atg1 also suggests these proteins may have roles outside the canonical complex worth exploring.
This constitution was formed when we were new and unexperienced in the science of government. It was the first too which was formed in the whole United States.
lol
juxtaposes
This is such a creative piece! I think that this screenshot is very effective in juxtaposing these two views, and highlighting how frequently we view both without realizing their differences.
total overview, global and dominating
This is a great connection to class content!
distance and control
I think that acknowledging the flaws of maps is very important, and something that is often overlooked. I use googlemaps all the time without thinking critically about how it influences my view of the world. The psychological consequences of visualizing the world from above (a view from nowhere that has been created through technology) surely erases our personal connection to the world we see at our ground level.
This apparent objectivity, however, is not without its limits.
I think this sentence alone helps illustrate one of the main themes we covered in LIBA 402 this semester – the stereotype of the overly objective nature that science aims to have, compared to the subjective nature of the humanities. We discussed why it is important to acknowledge the flaws of both when studying the environmental humanities and this page lays out that idea.
—where the wheel once worked alongside animals like horses or oxen
Even though connected to animals, the carriages that these animals pull were also designed by humans to transport humans, so maybe they aren't as different as modern forms of transport as we might think?
eLife Assessment
Zandvoort and colleagues have used an innovative approach to study respiration-brain coupling in the context of apnoea in human newborns. This fundamental question is supported with convincing data and analyses. Having addressed all the reviewer comments, there was a general consensus that this work will be of great interest, not only to neonatal clinicians and physiologists, but also broadly to anyone interested in brain-body interactions.
Reviewer #1 (Public review):
Summary:
The authors investigated the extent to which phase-amplitude coupling (PAC) of respiratory and electrophysiological brain activity recordings was related to episodes of life-threatening apnoea in human newborns.
Strengths:
I want to commend the authors for acquiring unique and illuminating data; the difficulty in recording and handling these data has to be appreciated. As far as I can tell, Zandvoort and colleagues are the first to provide robust evidence for respiration-brain coupling in newborns. Their creative use of the phase-slope index for peripheral-central interactions is innovative and credible. If proven to be robust, the authors' findings have important implications well beyond the field of brain-body research.
Comments on revisions:
I would like to thank the authors for a careful revision and additional clarifications; I have no further questions.
Reviewer #2 (Public review):
Summary:
The author's central hypothesis was that the strength of cortico-respiratory coupling in infants is negatively associated with apnoea rate. To prove this, they first investigated the existence of cortico-respiratory coupling in premature and term-born infants, the spatial localisation of the cortical activity and its relationship with the phase of the respiratory cycle, and the directionality of coupling.
Strengths:
The researchers used synchronised EEG and impedance pneumography to detect the phase amplitude coupling.
They have studied a wide range of gestations, from 28 weeks to 42 weeks, including males and females. Their exclusion criteria ensured that healthy babies were studied and potential confounders of impaired respiratory activity were avoided. Their sequential approach in addressing the objectives was appropriate.
Weaknesses:
As a neonatal clinician and neuroscientist, I have commented based on my expertise. I have not commented on signal processing.
There are no major weaknesses to the study. Some minor weaknesses include:
(1) Data relating to the cortical oscillations and the respiratory phase is given. However, whether this would lead to their hypothesis that the strength of cortico-respiratory coupling is negatively associated with apnoea rate is unclear. What preceding data enabled the authors to link the strength of coupling to the rate of apnoea?
(2) If we did not know of data showing the existence of cortico-respiratory coupling in newborn infants, then should it not be the first research question to examine?
(3) What are the characteristics of the infants who contributed data to establish the cortico-respiratory coupling (Figures 2 and 3)?
(4) Although it is the most plausible direction of the relationship, with neural activation driving respiratory muscle contraction, how can the authors prove this with their data? Given that they show coherence between signals, how do we know that the cortical signal precedes the respiratory muscle contraction?
(5) Apgar score is an ordinal variable. The authors should summarise this as median (range).
Comments on revisions:
All the weaknesses are adequately addressed. No more comments
Author response:
The following is the authors’ response to the original reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
The authors investigated the extent to which phase-amplitude coupling (PAC) of respiratory and electrophysiological brain activity recordings was related to episodes of life-threatening apnoea in human newborns.
Strengths:
I want to commend the authors for acquiring unique and illuminating data; the difficulty in recording and handling these data has to be appreciated. As far as I can tell, Zandvoort and colleagues are the first to provide robust evidence for respiration-brain coupling in newborns. Their creative use of the phase-slope index for peripheral-central interactions is innovative and credible. If proven to be robust, the authors' findings have important implications well beyond the field of brain-body research.
Weaknesses:
While the analyses were overall competently conducted and well-justified, I was not entirely convinced by a few methodological choices, specifically i) the computation of PAC surrogates, ii) details of the linear mixed-effects model, and iii) the electrode selection for linking phase-amplitude coupling to apnoea frequency.
Thank you for your kind comments and helpful review of our paper. We have now clarified computation of PAC surrogates, added further details of the linear-mixed effects models and calculated the link between the strength of the cortico-respiratory coupling (phase-amplitude coupling) and apnoea rate with data acquired at all electrodes. We provide further details for each of these in response to your ‘Recommendations for the authors’.
Reviewer #2 (Public review):
Summary:
The author's central hypothesis was that the strength of cortico-respiratory coupling in infants is negatively associated with apnoea rate. To prove this, they first investigated the existence of cortico-respiratory coupling in premature and term-born infants, the spatial localisation of the cortical activity and its relationship with the phase of the respiratory cycle, and the directionality of coupling.
Strengths:
The researchers used synchronised EEG and impedance pneumography to detect the phase amplitude coupling.
They have studied a wide range of gestations, from 28 weeks to 42 weeks, including males and females. Their exclusion criteria ensured that healthy babies were studied and potential confounders of impaired respiratory activity were avoided. Their sequential approach in addressing the objectives was appropriate.
Weaknesses:
As a neonatal clinician and neuroscientist, I have commented based on my expertise. I have not commented on signal processing.
I did not identify any major weaknesses in the study. Some minor weaknesses include:
(1) Data relating to the cortical oscillations and the respiratory phase is given. However, whether this would lead to their hypothesis that the strength of cortico-respiratory coupling is negatively associated with apnoea rate is unclear. What preceding data enabled the authors to link the strength of coupling to the rate of apnoea?
(2) If we did not know of data showing the existence of cortico-respiratory coupling in newborn infants, then should it not be the first research question to examine?
(3) What are the characteristics of the infants who contributed data to establish the cortico-respiratory coupling (Figures 2 and 3)?
(4) Although it is the most plausible direction of the relationship, with neural activation driving respiratory muscle contraction, how can the authors prove this with their data? Given that they show coherence between signals, how do we know that the cortical signal precedes the respiratory muscle contraction?
(5) Apgar score is an ordinal variable. The authors should summarise this as median (range).
Thank you for your useful comments. We have revised the manuscript to address these comments and improve the clarity.
(1) We agree that proceeding data leading to the hypothesis that the strength of cortico-respiratory coupling is negatively associated with apnoea rate is limited. We have clarified in the introduction that adult studies have previously suggested that cortical motor activity may prevent hypoventilation and apnoea seen in patient groups. We have also added further clarification to our hypothesis. In the introduction we now state:
“In adults with congenital central hypoventilation syndrome or obstructive sleep apnoea, a respiratory-linked increase in cortical motor activity suggests that the motor cortex plays an important role in maintaining autonomous respiration, with the authors postulating that cortico-respiratory drive whilst participants are awake may prevent the hypoventilation/apnoea observed in these patients whilst they are asleep.”
And later:
“We hypothesised that cortico-respiratory coupling occurs in newborns and that the strength of cortico-respiratory coupling is negatively associated with apnoea rate (in line with the suggestions made from studies of adults with congenital central hypoventilation syndrome[6] and obstructive sleep apnoea[7]).”
(2) We agree that this was the first research question we examined. We have clarified this in the introduction, now re-writing the hypothesis and aims to state “We hypothesised that cortico-respiratory coupling occurs in newborns and that the strength of cortico-respiratory coupling is negatively associated with apnoea rate (…). To this end, we first examined whether cortico-respiratory coupling exists in both premature and term infants.”
(3) Figures 2 and 3 used the full dataset. We have clarified this in the Figure captions by stating: “For all panels, data included is from 68 infants (28-42 weeks postmenstrual age [PMA] at time of recording) on 104 recording occasions. See Table 1 for further clinical and demographic characteristics.”
(4) We used a cross-frequency version of the phase-slope index to quantify the directionality and strength of information flow between cortical and breathing time series (Figure 3C,D). The phase-slope index investigates phase lags and how these change over narrow frequency ranges by examining the slope of the phase spectrum of their complex coherency. This indicates whether one signal leads or trails another signal (and thus indicating directionality). However, we agree (and as was also noted by Reviewer 3) that this analysis does not ‘prove’ directionality as other factors may influence the analysis. We have added the following to the text to address this point:
“However, caution is needed in the interpretation of these results as signal processing techniques such as the phase-slope index estimate directionality but do not confirm causality. Rather, they show a statistical relationship which can be influenced by a multitude of factors (e.g., signal-to-noise ratio and preprocessing steps). Nevertheless, the results suggest that cortical activity may precede respiration in newborns. Future work is needed to confirm this association by, for example, employing other metrics to estimate directionality, such as the time-lagged cross-correlation and Granger causality and through direct mechanistic studies.”
(5) We have revised Table 1 so that Apgar scores are provided as median and interquartile range.
Reviewer #3 (Public review):
Summary:
This is a strong and important report that presents a framework for understanding cortical contributions to neonatal respiration. Overall, the authors successfully achieved their goal of linking cortical activity to respiratory drive. Despite the correlational nature of this study, it is a crucial step in establishing a foundation for future work to elucidate the interaction between cortical activity and breathing.
Strengths:
(1) The introduction and use of workflows that establish correlational relationships between breathing and brain activity.
(2) The execution of these workflows in human neonates.
Weaknesses:
Interpretations related to causal inference, confounds of sleep and caffeine, and the spatial interpretation of EEG data need to be addressed to ensure that the data appropriately support the conclusions.
Thank you for your useful comments. We have now substantially revised the manuscript in relation to causal inference and our interpretations of the data, in particular adding further detail to the discussion with regards to the limitations of our approach and revising wording that has causal implications. We provide more detail in response to your ‘Recommendations for the authors’.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
I want to elaborate on the three points of methodological criticism, and my apologies in case I have some misconceptions:
(1) It seems like the surrogate distribution to determine PAC significance was computed by shuffling EEG segments and recomputing PAC each time. Surrogate computations are a difficult topic when handling signals as regular as respiration time series. However, random shuffling of data segments is almost always an overly liberal approach (except for trial-based data) since it destroys the temporal autocorrelation of the underlying signal. As the resting-state data in the present study were per sé continuous (and just segmented for analytical purposes), I am not convinced that random shuffling provides an adequate control. Could the authors either a) provide convincing evidence that the temporal autocorrelation of verum and surrogate time series did not differ from one another, or b) conduct additional control analyses based on an alternative approach, e.g., by constructing surrogate respiration phase vectors and recomputing PAC accordingly? We have had good experiences with the IAAFT approach (outlined in Kluger et al., Nat Comms 2023), but others certainly exist.
Thank you for this important comment on the construction of surrogates. We agree that it is essential for any surrogate approach that it destroys the cross-signal coupling whilst preserving the signals’ internal structure (e.g., autocorrelation, spectral profile, and non-stationarities) as much as possible. We apologies for not describing this clearer in the initial manuscript, but we want to clarify that in the surrogate analysis, we did not shuffle time points/segments within EEG trials itself. Instead, we permuted the trial order so that respiration trial T1 was paired with an EEG trial other than T1. This leaves the 4-sec segments used in the PAC analysis unaltered. This surrogate technique preserves the important internal properties of each signal (within-trial autocorrelation, auto-spectra and power distribution of the signals) while destroying the cross-signal alignment across trials, and thus the trial-wise phase locking (e.g., coherence) between respiration and EEG. We have clarified this in the manuscript as follows:
“The surrogate analysis was performed by randomly permuting the trial (4-s segment) order of the EEG amplitude while leaving the respiration trial order unchanged (i.e., respiration segment S1 was paired with an EEG segment Sj, j ≠ 1). Importantly, no temporal samples were shuffled within segments. Thus, the full within-segment temporal structure, including autocorrelation and spectral profile (auto-spectra), was preserved for both signals. This permutation destroys trial-wise cross-signal phase alignment (and therefore coherence) while retaining the intrinsic dynamics of each signal.”
(2) The LMEM approach is very sound, but it seems like ID was the only random effect included in the model. Could the authors clarify whether multiple sessions from individual neonates were considered or whether each ID was only represented once? In case of the former, one possibility would be to include 'session' as an additional random effect; otherwise, the group statistic could be biased. Many thanks in advance for providing insight on this.
Thank you for this important point. Of the 68 infants included in the study, 49 only had a single session. The remaining 19 infants had between 2 – 5 sessions included. Given that most infants only had a single session it is not possible to identify random effects of session reliably and so we have not included session as a random factor. Moreover, postmenstrual age [PMA] (which is related to session order within a subject and is likely a more reliable indicator of variance given that sessions were not at fixed intervals) is already included as a factor in the analysis. Indeed, session ID is not a distinct source of clustering and will be indistinguishable from subject and PMA variance.
In relation to this question, we carefully checked the analysis and realised that we had included infant with a random effect of both slope and intercept. Given that most infants have only one session the random effect of slope cannot be estimated and so we have now removed this from the analysis leading to very minor changes in the results (and no changes in the interpretation). We have clarified in the manuscript that “Infant ID was included as a random effect acting on the intercept.”
(3) It is not entirely clear to me why the authors selected the two electrodes with the strongest overall PAC for the analysis of apnoea frequency. Why not consider all electrodes individually? What is the worry/hypothesis regarding electrodes with low PAC - would one not expect simply to find no relationship with apnoea frequency, and would that information not be instructive? Again, I want to thank the authors in advance for their take on this comment.
We initially included only the two electrodes with the strongest coupling as we would not expect a relationship with apnoea rate at those electrodes without significant coupling (as you say). For completeness, we have now included the relationships with all electrodes individually in Supplementary Figure S4. As expected, the relationship between apnoea rate and coupling (coherence) was not significant for the electrodes without strong coupling.
Reviewer #3 (Recommendations for the authors):
Major Comments:
(1) Causal Language and Overinterpretation are evident throughout the manuscript. The manuscript repeatedly uses language suggesting causality (e.g., "cortical motor activity reduces apnoea"), despite the correlational nature of the findings.
It is recommended that the authors reframe their claims in the abstract and discussion to clarify that the observed associations do not establish causal influence. For example, Abstract: "...revealing novel mechanistic insight....". This correlational observation does not reveal a mechanism but rather supports the concept of mechanistic interactions.
Thank you for this important point. We have now rephrased the manuscript throughout, particularly in the abstract and results/discussion. We have also added the following sentences to the discussion to address the point on causation:
“Nevertheless, it is important to recognise that a limitation of this analysis is that correlation does not imply causation, and future mechanistic studies are required to determine whether and how cortico-respiratory coupling plays a role in reducing apnoea in infants.”
And later:
“The limitations of our study need to be considered, and in particular, directionality of the cortico-respiratory coupling, improved spatial localisation, and a direct mechanistic link between cortico-respiratory coupling and apnoea rate, should be investigated in future work.”
(2) Potential Confounding by Sleep State and Caffeine. Sleep state is a significant determinant of apnoea occurrence and EEG frequency composition, yet no objective sleep-state classification is incorporated. Similarly, caffeine, administered in ~50% of recordings, is a potent respiratory stimulant. A reanalysis of the data, incorporating sleep proxies (e.g., EEG spectral ratios, delta/theta dominance) and caffeine exposure as covariates or stratification factors in the PAC-apnoea models, should be performed.
Sleep state: A limitation of our work is that we did not record sleep state and unfortunately, we do not have anyone trained to annotate sleep states from EEG recordings in our research group. We have added the following to the discussion to address this:
“It is known that most apnoeas in infants occur during active sleep[6][30] and delta- and theta-band frequencies in EEG are strongly related to sleep state[31]. A limitation of our study is that we did not record the sleep state of the infant.”
Caffeine: We agree that caffeine is a respiratory stimulant and, hence, it is important to consider this effect. Moreover, those infants prescribed caffeine are those who are at greatest risk of apnoea and so it is of interest to determine whether the relationship between PAC and apnoea rate occurs in those infants receiving caffeine treatment. We conducted a stratified analysis to address this point, now providing an additional Supplementary Figure.
(3) Directionality Inference from Phase-Slope Index. While PSI suggests a lead-lag relationship, it does not confirm causality and may be influenced by signal-to-noise or preprocessing steps. Validation PSI findings using additional metrics (e.g., time-lagged cross-correlation or Granger causality) or, at a minimum, temper interpretations of cortical "driving" respiration.
We agree that the PSI (and other metrics such as Granger causality) may be influenced by a range of factors. We have therefore changed the wording throughout and also added the following:
“However, caution is needed in the interpretation of these results as signal processing techniques such as the phase-slope index estimate directionality but do not confirm causality. Rather, they show a statistical relationship which can be influenced by a multitude of factors (e.g., signal-to-noise ratio and preprocessing steps). Nevertheless, the results suggest that cortical activity may precede respiration in newborns. Future work is needed to confirm this association by, for example, employing other metrics to estimate directionality, such as the time-lagged cross-correlation and Granger causality and through direct mechanistic studies.”
(4) Limited EEG Spatial Resolution. The attribution of CRC to "cortical motor areas" is overstated, given the use of only 8 EEG electrodes, which provides limited spatial coverage. Avoid overly precise interpretations regarding cortical localization unless source localization or higher-density EEG data are available.
We have added the following to specifically address this limitation.
“It is important to note that the number of electrodes in our montage is limited (with only 8 recording electrodes), and so source localisation was not possible; higher-density recordings are warranted to confirm whether the motor cortex plays a role.”
We have also changed the wording in the summary paragraph and abstract to add this limitation and reworded throughout the manuscript to highlight the limitations of our study.
Minor Comments
(1) Consider color-coding individual points in Figure 4A by PMA or caffeine status to visually disambiguate potential age-related or pharmacological effects.
We agree that this provides additional visual information and have colour-coded the points in Supplementary Figure S6 according to caffeine status.
(2) Clearly define PAC versus CRC. These are used interchangeably. Readers may benefit from a more consistent and precise usage, especially in the abstract.
Thank you for noticing this. We have revised the terms where necessary throughout, and the abstract and introduction to read:
“Using simultaneous electroencephalography (EEG) and impedance pneumography we investigated interactions between cortical and respiratory activity (known as cortico-respiratory coupling) using phase-amplitude coupling.”
“Recently, it was proposed that communication between the cortex and lungs, known as cortico-respiratory coupling, can be identified and quantified through phase-amplitude coupling. This functional coupling arises when the amplitude of cortical activity is modulated by the respiration phase, or vice versa. Phase-amplitude coupling is typically quantified using non-invasive recordings capturing respiratory and neural activity (e.g., magneto- or electroencephalography [EEG]).”
(3) Clarify the overlap with previously published datasets (line 358). Are any EEG-apnoea associations re-analyses of data published in Zandvoort et al., 2024?
We have amended this sentence to explain that the previous study did not investigate respiration/apnoea. We now state:
“Parts of this dataset have been reported earlier in Zandvoort et al. [33] to address a different research question (this study investigated the development of sensory-evoked potentials, which were also recorded in these infants; it did not explore respiration).”
I hate the test
wasssaaaa
eLife Assessment
This important study shows how stochastic and deterministic factors are integrated in Dictyostelium discoideum to reliably drive determination of distinct cell types despite exposure to nearly identical environmental conditions. The authors present convincing evidence that gene expression variability contributes to the robustness of cell fate decisions, which reveals an unexpected role of stochasticity during cell differentiation.
Reviewer #1 (Public review):
Summary:
The authors investigate how stochastic and deterministic factors are integrated in cell fate decisions, using *Dictyostelium discoideum* as a model system. They show that cells in different cell cycle phases (a deterministic factor) are predisposed to different fates, albeit with deviations, when exposed to the same environmental stimulus. However, gene expression variability due to asynchrony in cell cycle phase across cells in the populations and stochasticity of biochemical processes enhances the robustness of cellular responses to environmental cues that disrupt the cell cycle.
Using a simple, tractable mathematical model, the authors characterize the response of cell fate decisions as dependent on a combination of deterministic (cell cycle phase) and stochastic factors (variability in gene expression). They then identify Set1 - a key regulator of gene expression variability - and indicate the mechanism of histone methylation, through which it modulates the variability. Finally, they confirm that gene expression variability contributes to the robustness of cells' response (at the population level) by comparing and contrasting the predictions from the mathematical model versus the outcomes in wild type and set1- mutants.
Strengths:
The authors are careful in their choice of experiments and in measuring gene expression variability, using methods that account for expected trends with average gene expression. The mathematical model chosen is simple to follow intuitively and yet predictive enough (at a qualitative level) of the effects of stochastic-deterministic combination of factors, and burst size/frequency.
Weaknesses:
While the authors show that gene expression variation is a feature of genes associated with fate choice and cell type proportioning, it remains somewhat unclear if this kind of variation, or any amount of it, is always beneficial for robustness or there is some optimum level of it.
Reviewer #2 (Public review):
Summary:
A fundamental problem in developmental biology is how a group of apparently identical cells breaks symmetry and differentiates into, for instance, type A and type B cells in the absence of any external influence such as a gradient of something causing cells at the left side of the group to become type A cells. The authors use the model system Dictyostelium to explore the interplay between a known cell-cycle-dependent musical chairs mechanism (cells are at random phases of the cell cycle, and a signal that hits all the cells causes cells that happen to be in one set of cell cycle phases to become the A cells, and cells that happen to be in other phases become the B cells), and stochastic gene expression. They identified genes whose expression is stochastic (unusually high cell-cell variation). Using a very clever and elegant genetic screen, they then show that these genes often are associated with cell fate choice. The authors then show that the stochastic genes have reduced levels of histone (H3K4) Me3 methylation, and that a histone methylase called Set1 is important for this process. They then bring the work together to show that the cell-cycle-dependent mechanism and stochastic gene expression work in combination to generate the observed differentiation of Dictyostelium cells.
Strengths:
Combination of theory, clever genetic screens, single-cell RNA-seq, and molecular and cell biology to dive into the fundamental problem of cell fate choice.
Results support the conclusions.
Very significant contribution to developmental biology.
Weaknesses:
Because the paper is co-written by people doing theoretical work and people doing experimental work, the theory sections will be difficult for an experimentalist and vice versa, but it is very much worth the effort to read this paper, there is a lot in here. There are no weaknesses of the methods and results.