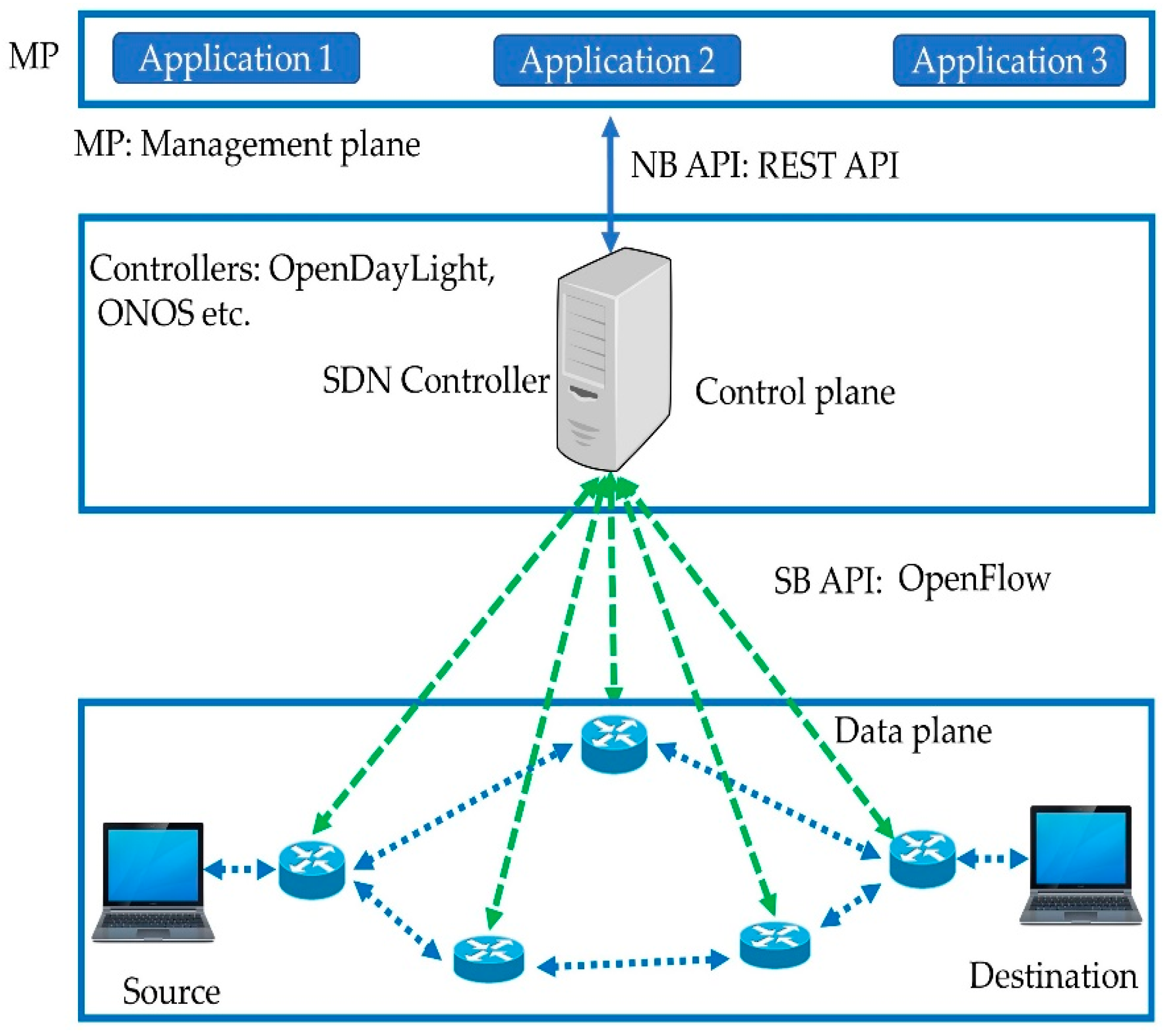
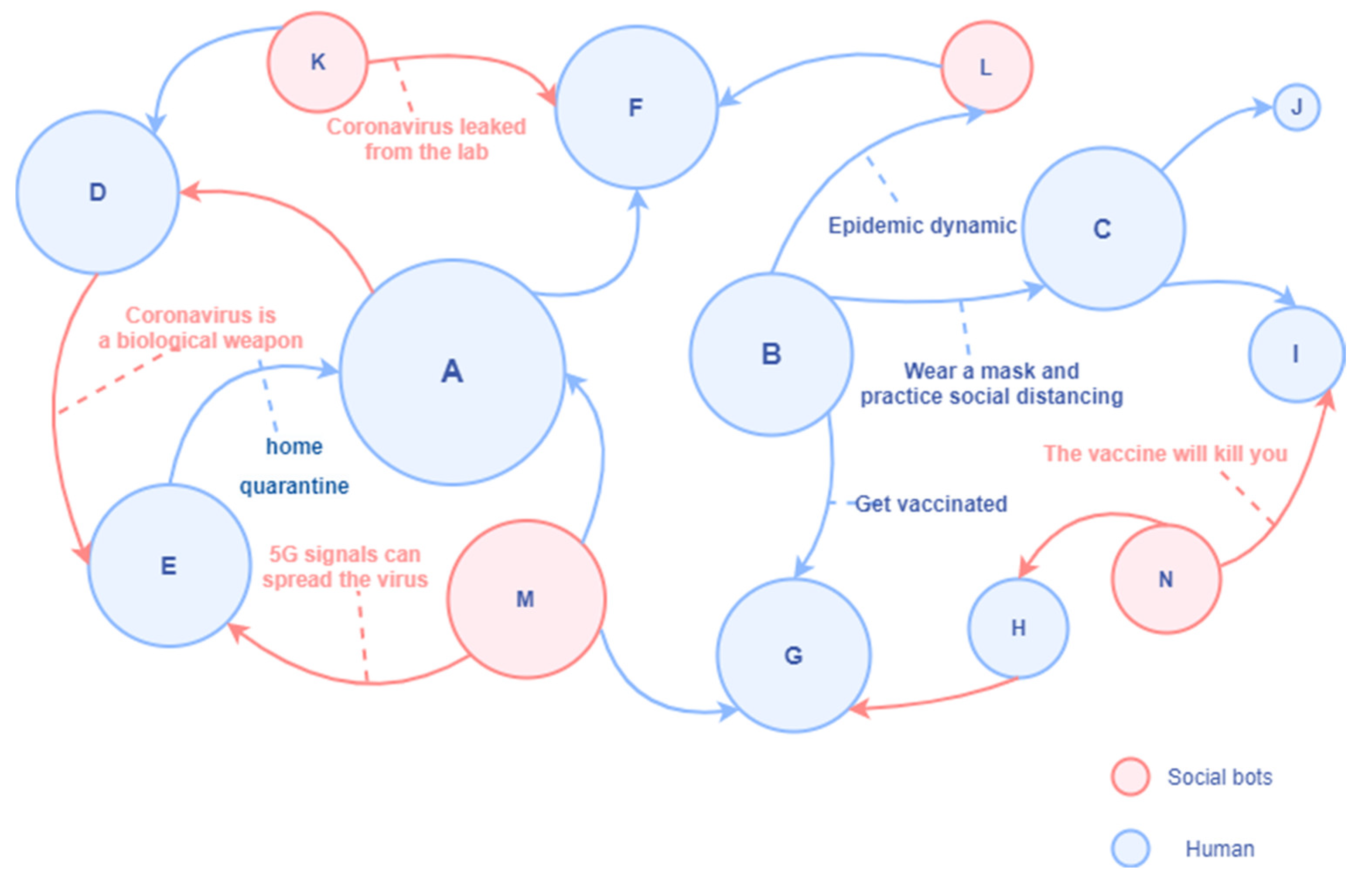
<svg xmlns="http://www.w3.org/2000/svg" viewBox="0 0 800 700" width="800" height="700" preserveAspectRatio="xMidYMid meet" style="width: 100%; height: 100%; transform: translate3d(0px, 0px, 0px);"><defs><clipPath id="__lottie_element_195"><rect width="800" height="700" x="0" y="0"></rect></clipPath><image xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAKwAAAA8CAMAAADMiaGpAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAB1UExURUdwTJNDz4g9zf///5lF0rmI5P///////////6dj2P///////6BI1aRK1pRD0Yk+zoo+zaBI1Z9I1P///////////4Q7zKBI1P///////4Q7zP///5FBz////5RC0P///6FJ1ZJC0JhF0qtO2H04yYo+zoU8zFmAC+wAAAAfdFJOUwBAnt8gECC/oBtbQL5kgNxj3JZwkO/v74DPu7AwMFBA+hIEAAAF60lEQVRo3tWai7aiOBBFQQlh8X6qKBpA8f8/carygKCo0NOOTFavexHxsq2cOlUJbRiTwwqj3W13jI31DxLecNxv9+iydtZ4x1GBFcbGWjOqeeRB3R2PEcLed+F6FbDhCtiFBF5UiNt1UbxmBdyOau7jqMOxRi1wBdxukTmcqnactuORXqMCdHwR2a5ZmRZCoYCNFkLr2CnWpmmi1WjBjIRYTd1t75w0raq04WMd0rWEWHfxKNc4qsg1a4Ow7Qq0QCYUYB55WIcKFkdI2yY/Dq41oYANN9jdyAPCfYsj/D3rgwI46/EhitaG0wY/hOWGpYfQlHXLnEjDBGD3v1OChWGt9FzDhqB71RGkQJv+NLCxnmucVc81UpnDKwKx3T9XM5vS01kcZpRmSwDgozSfeS0oNhor4N7puVb5aFp+H/ugresJgVDGSnFUMkaXwLqMsWLmtegDfZRFT6jlGpHloOlNoKrreiLFMrilxwMFB+4S2Bw+wOzZsJsRq66AYCdIm8EErGlYUjCWK2p7CSxl87/eABsjqq4A88hRozTkJkDeweI9C6mC06KkQVbmLIVFvYZ6YRACSIk0geAtrCd0sFgFHofdLoStUAJ6YZBq5a5KoHr5b2ENoYNs9o01scMgy2AvAGtqfWyjwxrJR9gD10EpppRkFOALh+fcmZ4OIpeoA/5GwOaIt2Uln4ETyKboZyOn1PMoK6T7gQ26Xl4UNCdj2FiDDTnqLg4huQSs/xEWBeDhD/Rbh8mRy5m2pa4dceG2zyo4cqlMTqF8NqjY6V+WnBYWsj1sJ2HjBrttEGsINqBg6w+weKM8E3brsn54k7DqLeOM3y7vrZkOH3T178zYQUT2rmA7BRsBq4/Hi2BRfXJC0Tsd14MpZtk0LD14tioJ+KMYInvKXEeGFiNLDy7+OfoQWQVrggREoQjbHrb+CEvEjNninlt5xpmEPWslgWpnqAxhKf6AI/WhfPHeR7ZTsKDUpnoDew3e1CJHCvE0lN4J2N7dREIWfAb6K8T8i19UrzS3+xMsGqyxPLLCMj1p9Ad5z/ItLBFCUYz9wWEM68q/cH8HCx7QGo+w11ewGNDCeIDdvoWVpSRX7qxgJZ0Oe/4QWQtYE2O2DBB2uwwWI+g4Dh1f8QQrv9O7yAb7oX/RI/sXYR+s6o9h+RJGBVaDvf5N2GKAzRfCNgOs5fPFYb/y/g7seWAVvfufwYpld9r3F8lXYF116Mhe5hNs18lGBipsLGFb3m77Q2+LhpXOh90+wZ4nYR31jlpozIUlau/N4mJt9zoSqOB6WQ574idsZb9PsKWwU0GTffbZTsHy2Y/CON03YwXgohECK1NtLqwj6ahGd3qAJUqq/OgkmyFVDAXsSa9gA6yl2u2xAmDRCGFVgZ0HK0u7LRCI7EPII6zH+jWQrCeqN5DNAMA6RJP0AGtUkURNdJpgX+NQCx5zDiwG5nBWXReeZ9BKFewBNmOqJ+A4tvDdrXNWbRaeLZzzQS18oq6LtM0sgN2P9pJQAfU16SMdAKz5CdYbHMlWHU7fUw+wJyVmwe2+62e5OjZ9LeBTbl4qY6wAGDp+ArDk9apaLsBOo3vYWx2W9LDlsPjyxHvDSmE7XilsufeZYK+vNuClAnzt7RRYX+51ZbRUmzEZ56My6+0Mm0UvExtFsNBy1fVl31/yz6Ib2Gh4ByJhKb4scrkVcQR/ndzRNhOOmug7CT6wXmfuItrjvY5ZOx/CuvqZE9ZFhpkUJvAUXMuXCtBFgajXb+4m0/F+h/O0byZMYPx0g4RCAbrbBvuvs36GFU83Wv3phlSA/6SAa2Iav4XlTze0HkspIHhSwP7bDxrnwIpFNxZZs7qkzwoIhQLSr2/Qz4NVTzfatm5r+OdXui38FwpQjqfDuuWrjelLwklbtKtAtwWOev3lQ5rJIoC4YwUQpYCVPRXHKKY1j2z4oADfNNY4LFm2Ak0B+8BY6wgkbppKsYbEWO9ALWC3XQsFrPp/Hykt/Gd29e+1wCXgB8T4fwzLWh3pPwaHCY6L2yqyAAAAAElFTkSuQmCC"></image><image xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAEIAAABCCAMAAADUivDaAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAzUExURUdwTAICAgEBAaenpwQEBAMDAwICAhkZGQoKCgcHB/Ly8k9PT8nJyeTk5LCwsJKSkvr6+uwWQioAAAAQdFJOUwAzQAETHCYDBgzsXK3Rk3xWRxyEAAACnElEQVRYw8WY63KEIAyFlewuiqi8/9NWbrkIiG5n2rQz7Y/m25PDLekw/EkADPBlIo9nkJylMfyvGXWLMLD06fhioFsQnx4B0zkipkPJ8qeYP/NAir5kRIJIfx3BIZP/i6a3XmQkUDYFYi4cAbQg579ZZEr2pI2IAMr/fMJ3pkSIbgiRhJAdYkw/CZKLaRiJAE8YKQiCxZwZpCEQPhmgfIwqUS4ZHkEaIkCJyJDECLtMrmcqQwCWza7GGbPafckQzoCyjEwIgGVbHQsbIZlRVOLNzD4kwm7cKTxEMviqMBGecEhYXRlmKxjCCiwjEIyrhj0xSAWKiEY0CcSoI6iMNiEzDhl+VZihQCJ8GdZdxH4wsgxmBggR2xXBmQVl8EqkCHOJCKWgDIm4J0LKwEp4HUqtHYTbEuIVKhGIWMfSI7gVK2EIYFZsXYSv5IQIByTtzBt1OMfNwELQzVuIrYMwfYTlS1JBuN8jnqqArxClF/DUzr1mJ9ta9qtFHfgp2/tbq7Y7+RlZzC038aiWJ1X1K9nFhXFC3KpkrV5bwF6A7posWAe/tdCMd+/2xUsrbSxSQcvqZWzdO6t6/XIZF46ahTmBJ4QqmUNvc8lYFN56fkXZiwjMjQuGEQTZHpx7g9rDLp72uXiWsckhhrInyLrLBsM3NY0+CRmK9SjGZkCrRSkYCaL23Vq7hTZJNFvVdk1rxoitzljr1hJhKpvG9LwzRqKMqWVEwKvZ/0JiTKL5HT/UuTLC1OjCAf2QHXTZP2vdROisI84R73dzFGiMaQAF5MXS2RgQrITWXKcJMpcTTVTQmkZoukyz2VwEzWa9CZONh5jMhju5rRuzNY6p5wlT3x52i2EZB+YB4OnMHSfbNN/eHrcr/zKAR5/+X/ED5vZsdcVjAFYAAAAASUVORK5CYII="></image><image xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAASgAAADVCAMAAADTop1YAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAADnUExURUdwTA0hVQoeThAjVEZRfRwoWgwhUUZIgEFKekRIfQ8kWBEfVhUlViEaWxQbUxgaVRgnWg8dUhUeWBscWRMmWR0ZVyUdXykgYywkZxgtYC8mbDQqcDkudiIqZT80fiAkYCo2YtHU3lBbgP+JTf9qTfvpTT5Kd/lN6//KTf+qTWz/UalQ57T3TY/7Tf9NqNnyTcJP6d1O7E348/9Nf03Z+HZd9PpQVf9Nz02/+pJU6l//db3Bz02Y/E10/1H/1lhl/WdwkYmRqlv/kk2r+02G/lj/p1T/vf9obaWrvk1tUn5DVCdjiWcvhV5pCsgAAAAKdFJOUwD///+r//9/P9Pxgf6aAAAgAElEQVR42uSby3LjOBJFuwATttlYoDroBRcKUVzwJ7SRdvr/LxrkA8hMkNSjyl0zEQPZkiXTqtCpmxcXCfKvv1bj4/3tc/7/HZ9vb+8ffz0c72/52GVapv+1MfIdfm3eRvzOo8s3vMMHesRvHhPd0Rvxl4xlWYDWA0wAqfvlcVT39lX9bY+mkeieR6SBj758Re/rUx4ebvwF3wF+F+gpj+DNwL/Ch74f+iGPfuyHcexHM6bM6u3jLqbuz49Et8yny7eYHxNA6uLO8OXOF1axoAlMJtSfQ31ixuB9H4c6+jHfpZbVnqre/iuYEBQhQkop8rMY91ixvISP0VCGUumEQM/akf+yj35gUQ0jfI1jwntBNX9uierjc57+MKFIdzHhfUdqilx4AikpIXHx+VJ5nmoRP3wQRJ4Q8U+BXxdiPWhqGJSmiNdoVbXM72tORU5x72N9fT+kep+o6FhMVHbdhpKUYUX2HF1WkSsucP0FhiS6CurIPkvKK0j5brSaGtflVzjFlz/rL2OiP09ACVAhoq7y6XZKz4stsS/DLUTwceQTpOhCYFqeRWWqcEBRZVi9wGo45TF/butJM/j6dhm1gBlSRBWliHZeDarFpPhUPRVXioEBFQEVeypf6ifB1COqXiSFmCyqyWrqc1mVWNwsuq/v9afUdVJtBky3N9VJIogehUT3IdbKKvUWpPYqtnbu6wdvPGpcaWrSPvU2f59KnnemCJSKhgqrLqr6U2Kqrl3KDfBguUXiUEEEMafCyYnEavUNQInKDyH14zCsMaFPfUjh7Qkm/gokTUO5UTSHYNUlnOrYvbv95KQnu2JKpeRYURKgNCTzg+Y00De6OeVOSlPjKnsqm/r4XP54KKAY3pXYxLXWPU5OHJ6KfYM5xeLURTXRNcUnT0pOCFh3UHgmS2E+WImqFN/7fFCppmt8PH5/UohUeCkymxTb6c4mgvUc5z27k0z5dbbTMrKDcXklq95rk6KIsNLUMrOTT/er7etJPvFutUYqOElOpKfKqNua50ROkUOTCuCR7CmKTGTSU5WHDlXnQO3oPUapYaiaAqNK43rpR5L6mC2U+Ku58WlrIxFJbNqOA3rdy17uWU+cxJFULEkycNTcEZOoigWF6RxJ+aEXTW3YOUvqbfkjE171dXxQkanbZ6SXvnpNRx+3SilIwAQWaFEyXANMqjUrKvaG0lY+wIgAEx8v8b7uL1u+XgAX9zhGaRcU467m1G02CaqRe0kJ4k1l8WvMWmNxW6Liqa/nJOX7aui1ALdq72P+LkL7MosFX+q45lKJBHeqjhOBb90pqPVdyVD44V1hAZpyRlSGUqm+vidMXlMaNjxqXHJCeF++aU57pgRTJEWVouOV8F7ZebUExpDpuU1Q2k2l4WQF42rNubb26iJH+blEBGxLbSQpNKlNi4rfnpvImXDBkmgJ/KAzV9cr0Xabioi8aqM0TqWU5ISXURTfo6T6gUnVbN6vWGWT+nyhC/X1m/XH7oQ6YhOXBlS0YYBzuKzsgkQC5eghyEpuS0fIiu8bVsql+ro2HvotRQGo2q776v6V2S/KMgaSU1cbvarZ21lKNWBG08VU4bLIIshk5yQwMS7n1qUntk66BJuSXgvkKP5eh/MHfc2v7zEo7M1lRrWfst1IkZLzaqZrGklB+k26R+C0niodp7QlpNQ7NqlzYEr9q6B+U1MSximKR942SNvLOio4XqnE2t8uvcugoqXhZWOTc6Xo+CfnGkPnBU/vudGpKQ0bFjUuDah/o09HjSesuRRNF5M0ldp1XVnVmdZcSQVqtRKEkxfRKEEhKFV9rl3z+R9oVICKd6+QUZs4D2tQltfX98gspY4XdqnsQ3XrWa+oKMp+QZTcpDNTned82IrgrCPHPuWCU+IyoZNg9dhGYFHBJl+/1RF+vxvMf78Uad8AeipkTnu98Nrj9d63eop1oy6UNsH2mk5NeCwmJlR9fbWY6cmlBornI7ak1knqsKeo70GkNzeRjXTFt/cNOGXarbogfQKvNgrqisX5lW2zfogQ+pOSlBNHD2hTyAn0VGe+tHbz+V8CpbbJY+q4O5fSnU3NumCJZjtTpXAVqpUjZVCuwRQcl5wTXbkaqCQfmH5L2V/nzNk/MPPdMc2vA00FEZ1PwE4+Te12OVn4tCxR6q6uenXdya4KfTvl4EZMVUtSeM7ZiC59Kao8L7sxKbWFd7ijqDjf1Jjm0+nyYuVFIsURk3LCcj2fTqfbdVoL6no6t72UaAKPKroCyG+s5lx1cE2rPGnsvKAye1Zjs1/8QFGXkxovgqrdlBIw2Ziu9Q0v7T6UJ1C1+srGgVrTeZXF8cGF7aoLTiCJqNqFDLzPj7LeM7tWyUx6B/jOoPZO85nmZZ4zrcucf+heAMUb5OjgHWUCtvDM6XyZlwV4zW3tZVDVn4p320xeP5/TycC1sancFK6irrC35Bv6ui4e173gwzg98qgMin69vKKoVE9QSZFTAXzlN7tNWIb53c5jtGc6AajmzCa9/G36ck735FxQoUDpyaiq8fOiJwxSvd0xbtPmYzOPFRQpalLymybdnynP6JFWvykfntT5KGeiQ/IpksrHUy4gUHnk/75OK2nqhJANT0hLZv3qTS7o6gt6+jOlh8mco7k6U2OD1eEJRS1VUfMNjJjBwc+nKzyZz7fucoaq6iZ8vGJrLsHP+cXaKJiVM03zJc9xccRjbgt7FGJBu79OQOqW3zIfMflgTySovuSUfTtVbiIlrShXzYqY/wilca765rANo/f2DtWj7oH6RyvqRjZ8ntjpz2d6cqm/uvDjFSoPjAgxjBSeQEWTnPIUANe5WHsBFZby0pw/w/l0hXecbIMAlGS6A7VBt0PJ1p9udUo80KQMJqWowzMeNeP/fDddqQTxSdbS6UbQsrTmMx+SP9uS0gVeTFC7FzX/2xie/+IyRmCzMKiM7jx3Pr/XeQoZVP4HrpcuNKXnKxiuvGaGs7Oe/E4XX7XAXqWDYVyZ1IFo3Zn1jEdB3ZzxJWRz4yf51RlAXRlqFlhKM0gqf+Ir2vgVjAnN/KZBZU5w4MWTsG4eQYUrSin6XOlXUNTpEtRmuW0QmDAQ9oVkIOrISRaFMwibVMK6s/nggLenZ70aD67AKD+Z8AxrYHQBWh3KDGsuAaP8SWc8GRlYJqsor+Y5/s1pJEXdABmMTDWEM1ajiU6rdkqVCrv2fVSmh1DiZl7jlXMU02Bz+aGQemLWW4qiBNSkoqgBdcFTVACUjquXWHHoJmaW2I3CZX7zhUChkPJw+eAug7rZMM4VV1nVCe6xnlw77XmxqL6PPWMas6j0Oa8Hqr3DrykKLOrK41JnRgAFERxAYWYqhywECnCIPxVQIHwBdSZQwStQW92UYBYp4RlIW80WJhWrRyU6hzopKydUr3iUAXUxx8wCKpGiiIrZswMrKk0C+BsqPY81WEoP3QoUxKV3K9sAWw06RSg0j3dwmS6LaR0MIKVEsNJ4FIN6RVGL8ahq5tc83RlQ0EoBRYGZUxy/1vAkMyDkggloXTA6VTMHIS3wKcDvQqsotf43afupoUu2bMMUQdWzqBM7VNIzXgG1vJDMC6iZgifM4uRjoKQJbBvcHBTVMZWLTpkZz3UZKWZmRB1kAe+XG051Eg+Ch8QwiaLWvd5gnDs8icqUnt7YiXU5nIBV0rX3Ox61CpygqESK6sijIsSpMxxyq+Xna8AEz84lBzzgEBQWJfP5VF4KYe1R4k7VpfZS00MvLzEKL4uJg4oHWHdHUdNLirKl1+GahWRFM2NKpfRIURnOlZYwo+4SzIQqL1pw9TthlAdZ+bqEudEB0CQQUE5n8Borw7NikuUfjp8ql+O1HjUdJKq+NkUdHoHaau2WsZSeJ54DzedjknoSX9KyLLnQmtYvXOXVSXuuW5apuRAqHzGtuuAWUghNDndPznvlrX7Wf432WSuoMZGPJ5LUQQXOX77sDHYx6zmrpptZJrtu+3wne1mYV1dD7bUJTBNTFna63/S46myDUxtUFD2RQQ1lztOKeqYXfrzfxUx0Cp1cfLB/XpiXvfJyUpjuYZqTeze3o6TqKqvwfPWFWnvqv4odCiANY9XU8WmP4usQj+aV46oWE+8h8Bk9ZQ/B7iCUky/86iQeVXY7ePQOgguqhamYPVl6BZNWFLi5EtSQpGNe5PTQo453RJXsZgtttNTLEu0FZOZqTUsrWH/aY+VET+GFRNA0DuD2k1HJPkbk2ktcdzTnHUvfgEjtgTq+cN0mIeITngjTxll09prWDT15s8W5O2zP8oUIJVPeT3RzuOXx99+ZU6TlMJw6fRyOJCkqPBhw/4yZcwkeWj3FIqVYzitIdeO8PGyg2iAVvN6RWnk5pyfZKZf0+IKoeBoATv7nT32NcQ3lWHygqSOZVIGlS+/4ip5SrbwkVwOnchZGjCs9/Yeza+1tG1eisAXpgy3LlK3EVnpVBUiKNE1v2jSp2xR9AE1b4Pr2//+e5TxIDh+SnR3KdpJdLLqnZ86cGVFMlpXhPuhIn5wRTGZdUTzLhocw1dO6EMv83czLOSCFlFIu9zj1GKSIUf1z0o/ZlDNEytQ9t0ssepQlmXD2dp1jVLIHnkYu6jlRI6cILQRKU2pOQAFSq7LRf27CqtcaZcveoJj3IUJd/JS+crdZcoMVPTatUmyynMqycqDguSnmAJvE9oFUf1ccAqk29/QApqywiTefA06rkhlFNqr3KUVCJYCS8JwPcKrP/YzzohRppyJnkFJw+ajvgIRPzVz8uZ1daKBqHYhTTTo+JyGn3FvR37OmU6u40eudhzq+henCwxwMoaQvQAlXaUqZbfWxNLntKcMeSu6bez5KlWEVBSo51zvLqGbFedf0ivLOS7zzw0D1gd8UzskdeyHoNJB2ccqJniXL5CbDocalCB350SBplGCBQmlOEZ+g7BFKiJNOPavlrE+YeybxJFCdQyI4GqSPt/M4pErpCAaeQkh4giLL5CPRgXua+gAV07EbdkNNX40wWahqVPJsSoSqAaVsTslXrhAjzaoeGBWkXidTbwiWjtUcUWxNrSNO5ZJNxmeOPUiWLHlyn2E25Avc/sI0XOFuA5ZvwypMuSm/QKMqzSdQKVPzVLNCRvVyHNV3naBUMvW6RFvnNXeekpuzVNRwvUvbcfcI8KgVt21euOciQSiJUW1IZQjFDkrjpAlVo4XimqdL3soqlIYIL5InzjwJVJeobvqzjT050cmdVVCOKlSZHehaBoByXd2B3EsAWFtGVbgg60wUVVFlWaVBqhCplf7DNyU5KHpB/8JIpYDy8o6waZP7DR2RciHnKT8uhgZZSqcKcURBEqqp3FBQDGwmkIapIIxqA5FhF3AKkhBRqsu6nmuoKjIHK1ioUUCpts+RUhormXtRC9MFlsBHTLlZQW4USiUmBbEzGEi60elKhE3SShWhAyekap9RgBOgVIBC1RlKeVnNV+g2S4QJlFz1us6bktdaQp0boLo8CVakT0rZTWI4LVCy4JXUNR3sXIQ0ZdmYNKUhKUb8eM2UqixI1h2g3dRI1VWGC/hk3KYiiVKIlGKkOpDz2B50Xr/SCbjaZObR9kwVKFQTaXj4aHkmjypKjQrc7aji4KC3kJueiE7mVZkvTamr64o5hThVgNJ8hZRSzChAie1B7wlUoFGdFKmWftBKNy7PdLBDlRCjJngSOD1VKUa8+NS/DVwM3R6fJqwTLWRSPWU/QNUPvgHRKphRmYYK0w77F82oNem5KXtd70GVrHqST70cZBqgrNksS+XPCsyBevlzjvnLadloS3ly3zMC/hfj2OgLYgsrGVrLVwSSzjt9odv0ofI0yiHUDZW7XFQ5pZQ4QKzMz/e7S4qLywsb1xfXFK85Xpl4R/GW478Ubzi+QPz8+fP3799//vz5ruMDxwPEZ4inp6evOn79+vVNxw+IjxCPGDcY72/eU9xRXHF8wrjFuL/f/yU+5SDmALf2B52MhOFsDWat+VopfzRuks90weDYynODEsCkgbo0MBFQr2EBTLAQJlgaJlgIEyL1Bhbi9IZwioB68HF6QpwsUA6nmxsPqLs7ROoKlsWJYdJA6dgzUmwOwB4kNKr1RLv10q4V/Z2AirnEmrS/vNwxTIxUQKnXTKlXr31GvWVKEVAGp5BRDqkHhOqzhupJMIqR+ghIPRqoHhOMurrzGPVJQLVHQqmeE7hLMco0LMaG962FSIX3EZzfhJlgQ1jtdpeSURIkTj0GyuLEjILUe+dwskDFjDKJ92GAUT9CRj16jHpvKYUwXUWMur/fbXOScmcO4tRrXQvcce61wbRABZmHjEJC5XsEykmUACtIPQmVxyhLqQOMYonyGfXN5B5TyiXfe0uplEbdfnJI7e53u03LjAJbLjPvTIq5MOGddxshNc5U0jXtQ5gMTteeSCUZxUAFGoWESmnUw4dIo76GGvUxxaikRsnMA6T2eW8YRWoe24Mul/0vSFZPnyo180WgDE55vgsIlap6hlAho5xIxalnGfVdI/VBIPX5IdaoHwapx7GqZzQKcy8CaveCcq/rcg1OHzGqF95A94RywML3D/JIn6jWwSr3RqF2IVLXHk7XCUYJLX8biXmy6nka9eRpOWUeIXXjazlrVKDlUqQAp92u6ZlQXVqjpHsysq5f1o9HeQcqbmi1w/BhuhyqegcYZXF6M6pRgNRT4KO+JX0UQ3VnNEoyylLq3lJqY0yrr+Zn/ijYc5pDroDzDgNv258TSLbqydy7vhipej5MgY/6Ml71HpJV76MlVMgoU/SuYo26J0oRo/Zc84BQjNSZ58w9U26SL8w5kXq0Kxuzb490EowKDMJF0nAmoPJSb4BREqfYRx3PKEmoW0coBApqXu8plGSU3xNLC+UH3ihsSJ7IRO0x8S59Z26MeSTmBqd3vj0INerLkEbFVe/XVz/zRhllcbqSdlOIlOqN3exjjerSg+CITbCFqAQmEZUQqHJhGEVQXVwGLmoAqBCpgxr1wfMHCWf+4xhGScN562sUAUUNjGv1zugCoGxH7Pq71k3qEtGAlJNAcdELGZXUcpN6rtkb06g3/6rqQdkzjEJKBYxy9uBTyCjCaTfrOfn83LMtTBdnX6jjdksaAcUwlWufUUGzdz3Y68W5N9LrfT9Q9X7FuTfKKF+kRNXb0+SAQdpaUvljFtEdt2pAylnFS/MyNippOEc0KtDyY3s9IeZH9XoDGvUp6mBE1ZNqvsWiR4zq7K1Orna9ygezDnnVkJzzRDP3bFSQewKn6wMadVyv53qYhI/ypwc3QVcc5d5t7Dhf8CzKM1E+o7rw1l2KTtZDlaWFqtx5Rkrag2vPHwRTljGN+vLvnHk8jxLOfKTqOUpxzSOoNpB7Z1KjROL12OAxWqEpoLZF3mppyll5PsKo4V4vXfXSvZ6oesnpge31xPTg5jHFKNnD3Ma5ZzNPsElUvb61eddao5lwB7jF2KcTN3tOpC4Gjfl1am435KOOZNTIhJOHB0HmeR1MnHmCTppPAirWqF4MNlVqAMU4md97weYAUSqbLPcZ5c8Phnu9V/486qhe72GQUd/c9EAyys+9K1/OQ0ZtvcwDKd/6GsX3hXuqecpzBqX1BqThpsvDW1LwVTab5btdYnYgGXVIy5/b68Ua9cNND2Tm+RqVwskChUpucm+DjIKiB8s5c2aUErsw/GJnnk1mUtm3rJlpxPIhRoW93iF/kO71vqcmnJ8TE06vh/EmnBapeB5lxPyFhanfOD5R+r1ke6Btu9igqRI9nmqCIMMJJ+3i71fZpxiFZe+46UEo5oc16skw6qtw5pJRj0ktvxtw5vtNv+iFLd8YG3Wmr3MWc9EHq7QrQGPQWJkilBp63g2PN6m823qXg/f1DiJ11Mw8vK+XdJwy9UJGhdOD/X963J9Bvhxh2pI5wCnLlsUcbYHdhhHt+W1s7jVWwjFmJRzXYQ8nzs/3XvyPL7H4Oib+T+tQvIQFVyL+mjUW+79/t+v+dOHuLANMG9PBkEgJH9UKmJRSQ5lXuhlLY89On5ntGLTpYpLZg9Fge0ld200Gpaaefq/whL9Kf117R9LRe02PGoj9hfhfxYNqZ/PZZD6vKx3Laqmjgt2Yk3ldV3Wt3/U3K4jlyVJf82W1qvQ3K/1v4D9Zw1qsFxBrHSf5ycn65CTv132+6E+tRhm3acT8jDWK7uNpmWpT9kk1gUBRyWvo98/M5P6LyQRe5q2YTApY0Q6Mwp6pUqRPs8fNFf7pdZnZLW6jwst8qcGreUHQux9LwFbDti5x5Xqt80UOrwVn3ikwamNr3pbnLGw4W2yCzZ6VgdGKZ6L4t89kaA4kVhOCa5JpgOADYTJQTZKP37kToGjfPey2pO3zABb8pIKfTH2MqoqRImjqqmKMaoGS+VzyO5FwtVwv1xSA1FqL+MISqrNukwmFpEKNas1mzdbcc0nknRJDcnrIpkGI8MAOQSeEamKiiBgVnC06Fdt/62JqGVO4TZewS66qp5XAiL5hQnk8SsLk+FRpkMrlWiO1WmtKIVKaTeu1KXin/Wm3QU5pgDYi9XxGJZ/cYJRKZRmFMOHBcASSTL5JBNWkSEAVpFxtoOKHoIg/TCPY3GQ2QbtEqyWjDKlqi840mXbEqPUSKXWCqQeMgrWQGsUNzHbLlNqiRpEd7xVSijbvJ90mj8ixw2MmzczJZ/jFzOSdTjzMvAJhGtgOPahNRS338OKzKzLl7CdSi2DyYlql5Mkmnk49k3WoUAuAyTFq02+BUb1tX2ixmCsc1B2h41zzgE7abM4yq08z5hPDRPKEuZcl9rAmeIXP8xRTkXpGlyyb+LOyKs5UsqmXhAjZtLRVclkCm/ANsVoQVhqsU8Mo9lCI1BZxIkbZ2y3tqDw5N85aDgJFhwpZajFWGYk4ClSUedNpKg2nhlCGVO6jKkiUMpKkgrZjOk2qqwOxJKwAqJJSb32yzunSdDKMWpCJ2nTAp44yjy3ndvuSnXnLs98YpX8Yu5YlV3ElGLKiWUg2tDDgVzvcq/v/n3hVT5WE6DPYxpyeWWVkZmWVBG45FYBKlKICuzmB5IhPak/DfzxOYlLJSi4VyQFOiE6+UkLZLJAsXKcOTKQ6ItQV/WmiEJUJhThdLgITEupzv1GMIkbdn080c2Pijz/5JKyin1qjcsePuZaKx2iBOTn/JzSGTaS6YWhwEphYbgOexiP3/oNQnEwFKKh76E/rBWJmFt6lKI9C1PsGON3JzE3gfBwF8mVPp5DVR7/JGrwgVaIm8eo4FRzjZqDBO1YG5pLn+3wEMG8IlA5r3E53iFIW3gofxGmlcHClGIWM2sjGYW+seBRb1Js8qjdXodvbdbaiXg4PhAu0eTxYW6IsTlVvEOU1AjwdQJUKTr6ilZeopFFABedLAk9/+pNojkveyuLLBgW9TA4FWXsAlTLqQ7q73SkegPhIeu/vgwBlRis0+yV/CtAKE0aaNUV0nsPT8QbyLlqJ0zjfd5j0w4QCPqkGuV85bFRaPjFCk1gUePkX2HluXsDLL+jlGzMKu5fb/X4jSj1pzoJmzrOVx39wp4Wfmrew6DQbOJMyPXctXPcOScQWLkbeHH608ZKo5LFvga8R/5UOsZHoZHMm4bSC8lB90BujQdHLeNSH+ASMYi8HRv0+LJceNmbuRgb8i1hIJByv5IwZND0RVIyRgcg1ACX6SypPtkhpsLHANr0jWJUXIxf/Tn9aEp85OllGrWjkCBM2xAATFr2PWhR1xIjTU1sYZBTGp0dV8zp0AjZhNA/YvgSJmQia0AkNyldWLhcnx6pzJ8WIQKuyQOVPaOF+TCYVVJXOH0A1FTaZQI44rezl6zXXvAvqDumU3xtCBQWPsbpzOJB48CCEBKqlH8rP8shvfmasD8ahvKETe/meUCf8ryf9QxLATLeC8IiJj2aQMtbdb5dTBRsLExv5SBOqDBO+MpngAyhdL7aBQUpxNGCoOJn/vvuZoFpyWXCmsvDDh3W0gl/OVc3wQPmgHarAP0/EppMTRjlKAXiPfa06jw7uK4DEnvwhnyZRHI0KZLbHfEI6TQjTyiZ1FfFlh9poFkVmjoy647z8CZRSoB7tjpWO7nS0shgnz9AE6fJ8k8kLXAARiA9plapU0MQBKHwME1PKjxWZ/EG1U70xgdifVHcr8ulMSIHowMxxaHAhnDKjbtuNGXW7Y83Lbo7KE4/qLwgvZU53Djg04N9YC9TfcZcXvE41PdU8Fp7O605OGKV2TpKT9+DbGQpwydPYEqFiNhFA52MTn2x0sj6+ElYw+b0im648Es5WvmE+2Fh5N9UeTVmQVVj1evtVyqowJXJcmVqQSwoRMSmoRxFMNpfrVWZU5pRD8z4BTJEDQsfB8XZ7yUoSmeTcUuls49LY2lJ9rOJRMCWHaICTA4Bqw3SQgdpgypI7PY5RaOQgvicz6vGPBMVo4c/Q4bsMDRwzymPK9IPbDzLRmgbj45idnNS6QTDyXOH4TXhp+PbtANNm75EGKSq9cYcQvr5gJeGKylsvTKiNhbeR8rDs6Y19nKL2jFpKfjq3OAX62SKZ/4bSw8ggSlOUG6qwydHAAUKOG2DAyeEZFk+8bYIxGKSxGfKqfc+FS1Nx8B6FGt2h9K6kPTSpDQzqupFHUc17g0nhHZDv0r/8KqNq3fUyFP5MptCJrUlqn1PV+U4YJ4dCX8o4Jodkci6lju5Ic2PiW6QzRFFa4LgvdZPySWYEY+tMDafWleIBsYktShh1I06RlaNHoZc/368doxbdemi7FsXpzD8CqYMoF5yQaZC5pjDKIUQn/KIIlcCdsOg5xsgxpzzRig5YnMt4ZYQYm1jZEXNq1rxk11cEpTqQr0Z84OZq5GRUG7zIy5FO5Z5aMPJfkh7Fg0eBqbfLAFHyhUy8ppCBKlMoTpmmxXMmZQ7S/SVMDzzpdS2lZBkTVjhj+kfT+x8UZ8udoESRPAsOAhSghEsvpCqpL8QAABaZSURBVLuNSh5ZObJJLOrF0lse1TpCz8zhgfs0OwjVIErBogTltNQRpRx/J7hOxKfkCkiu+BPBhNQ6hGg6AzqzgjTamdw0NbitUukm0R2uC2ewLpf1g5T6IKOQUAAT3qZ953DwJocCnH41cPJDz/sWhdicQygzzSD1zvEPPJS+Rb5OJRc4yk+QyAWnqFbeZnIdocQitL36dNQ0WpmNNY/GtVYeHpfVWBTmAspQ2Z+g6FHJu5FFPTEc/L6QUa1HdQcsgdUnysNIQNJzgtMgK55MIvrgGcFCbxqc64zFhVDApngousm2KaNgtM+WghGCtVYocdWjCcv2xRa1sUVtcts/JU3MUBmk4lEcpJbv72XvUCGYaidRHHw8SHfHc3LbCsPVibo7R1UO1OYAJXbw2LcoASgeDiyl0O2sqaZTSZiIkoD1lRnF9e5yISNnqBAmDpvKJ1be8/U/jQeLpKgOUougVOCSxkWN3OjOlbUqItXA9Y7+7MSc8ieSJ0UjPcQoHnr4bBo6i1EF26hymxpCwfGVsfoimC4XzZqbJRQiJRkKkBLpwcNfv7sebshU2hav0jMr5/WKApkTUcpRLBBCeYxRJLNYoIp8igBTOTV0OtNi7zge1ri15Esx8WktUAGj8PhCKtGbGSXLCsinN7MJ1UdAPQyfvjvK00Y4lFl50MUEWaGSFKAvWlIncyJ3csaiIsYA0VyKghLAA2DFys4BGvxu1+kO4GK5dWACpFR62BEDTsAngonY9C7KA5xe4lFLJ2wyn86hIlTITAJoqOyVqOm13En7AgZOdo5IoVEBWCk6Dkt45k+iE+4LM8GSTjNlzNEw6RAprXSrWLlGAyFUPiGdPkon0Z2GTWLUrzDKVr1uyVPdmYkBN8O6y8AiZFjFtDq5gZsWUB59iEWqu8jsiggSWRRfKFZAopkDwTTDdcfEd00wolP5E+FkvFyOj4RyptT9gFFLd2vdUmycHFw3Q9FY05n9PV6ZVPpidKahJAPBSbRGYEVGKCVBiQxqjLNS6zzJ0NLiMx4iZIpd5eVZdexSm+JEfOKidxeLEpB+X3AUM5edBq30llZ2NNx0duNKpbim7JkD+JRl54hEPtKWS1FeMr4UMVoSm2a1ceRTptJZdlv0RMeXe81ZjypAfQir22ejTH67d7IBS+/9bU1q2WFVdmPg4cDIfYVAIRDTiC+c5NBEmFEmcAgMVbkiuSRiYzqJLRX5tRbex2otHFKkphamL8lQm6FUptOtwYlAegqjFh2vfPfSgcQnVJ6QKLgeVIMu6hnnQrXRiY4oICWlUkrRDAsi8YgzATiS7CE4E1LnhkBV71v6umlnUOulxAMV3rZxj/e5y3iFkPp9MVQKlM4MGqxCrT7kFMNUhygzq+NU6XayS0lqHpe86hip2pW0OZd5ylhVOdleqOe1zuE2YU592VUWRS2ezOt2unsZj1r2K1RNKufxEyaCsENhcDaaV/bEAlQ+eSdCK7pLxKdS8VB5I1EJPxNvVN0L7lqBxN9NtZu6NW8rOAlWCBTT6bdSngBV3Rb0XcHkTdkD4QXmUxAgmgkLXxQm8ZSAgCrRqRCKuDUSpWLxpFkYZXZfmr0WvJOgmcsJVHs+7RglwYCNHJu8G4VN41BPw6iCzZFHlcGB0VwBy65PFaC8UR1dxFz0XIEolTPBhHGAfDy/IF3OOH2Cy2nuEKkQSiGzJr52DWptcOLVvIxSbeUvozwbD3qzuiVoMid/go9HiGzlK6udUuYKQmLjiFJkoLjeiQS5aRlZcnSa4oyePukyCxv5yku+63Xaz5smNacGJ+nyNEOp7G63qhluepfGo8SmvtsUhYw6l3zgnCWLRigniZz+oPaUPHp4RscxLFjzLJkiu5PqjkQ3oTnNKrd5Kv9Ym7U6O8U8PqzyNnYoHBrwIOq+J5QcP8KonuaK7phPkDZRewF119yfwIyyBS/yFwDiRHMVRCnGMiyYy1IUc2nsdClX0tsVlsZNYqosfOrjJHwyCYqFV3CiMflv41CvHzFzw6dvEw1C1RA7Z4peafP2mnPO/D8qOVPuTDoY1cYlFXD/i740T5zGx8ma1Dq1Nv5HKthFqM0gdWOc3rcyhto51I+tegxV6HZ5BJVjWu1i1KAarHCKyiq6CyzFBqym2mUzJ4sCbOa2j5t1RwoSCx3KmlH56sJEQwMteBeh01atUEnUrIz8pzCq6+OLndnJHunAO3xqnzK5s+JT4oAQ7SQlVtmA/GkWHzf9CtoUVLxpqksecOm6c6VpPRKdKG+9VMK7Gd3dCqHu+2zww0DhCszS2XlvhcfbWGCTj+PRgatDpWvFhzApHqkxKIYoAkKRvsia5nFuJuKzTQVAJXgrd6bpL2syEHUMiqV3L9FAUSq6+xEzZ4weYuJLs5ywcDOM/bDnfs9XbJI81R4RZedjbHsWHT7NUvUIIATL6K6UPcpMa71Cvv6LUZd2ANzi1GaD+3NX8yyj6mAQStZcmE6wqzVUzXCVyF3ds1RlL9aTubq9A9HNpeQhp+ju1rkQKV9cr7MkctgpDuuYK7Hqn8elOuqKJzDtpwYv6l1+KkYJnapsYBddMGkKUOH/nF3tchs5DrQqqY3DOVo6cj5kSZek7nn2/Z/nNMQ3CI68pzheZ392NRqNBmZ88kIeUAkVHL5sn9OfAmwqyCP8W4quuYJkqmANGjz7Wcr5CzDlPjLQllygir3mA/n0eDBQo9klwSEiAZROnlHS9jp/ME3Y7igd94QqZSqMUUPrXD5Au516l0pxQbsSX76GEXnMxSgUMsrySUOlHfnjAV8I1Ge4VkBGnb4xrWwSJe7pL/VvhRLCNPigkO9gYcG1kjtD3Z0L/1GZCgzDy9c+eenIZDIDazWjhvdAc/BQGtXqL5FAUcebKTWA/VQ3wAQ8EqReobQTiiD6ILTOKORawqs8QAacgmOLr5SdoZOZ8Tal5NzyrjaFAoFSjJrTJ+d0c+LWp/OVE5VfCkAhVv1b69OQT4W+f0ykS5iOC5eo/mrlsQWDFbwFw0OCIzYtxhes2pFv3X6qk3IFlGjUBXmU3BWi4JROX/8AQuPKQ4e5Q4USToTaUWsoVRnsQMhrVRlUxWOL415nYpWu44nPZK/Jw/BNKg8JpTUqDlnIQXHlpVclB6bAg0MmsyhePeXpA957gXXHEgWvcbDpU6XlAQBUa/VeyXQ6V3Qm+rWm/Hp1U57lkzDqolOVPjgY8+mvVzidRkX3hAR+EoUCVuHoghKF73HAh8faP85d0WWFVZYf1Gyn7lbU8AKW3DS8q5lcHqDjDNR/PvtdnpEoMgb/qPrGDa/Zy2/c7YRRykI1VlW2TlR5bVoRJtXOLsnI0oG0cgYlk0u0Gb4biUKYrI9KvE2whEK7+XWopsOeVxqjqOTK9EEAFTQDhccWflCayHUg3FnNdVh2izHksj0/kPL7zc14nlGfJOS4diF3cAKA0j/h03Rioxl3vP2Xa5EbJ06d8a/8o9IHQpXavjsFhwrTwsTuySxckFG642Hh+RjqRmsXkae+9FIYQ6lQ86uMOnSaJfg0HScqNR3neqNn7/EzSAeYVnZg8TEdZr+bz8klh7obDyWVR0Cl6NxAuyizfXnJp7F72quuYUXOQJHprOoPuEQgnduTiPh8hu112ZhwPwLjWd2qIk1lDbpbA6m8h8HJMCrxnznQKOU4T33ii0HeNDLlReRpIvP0wegQPPjT/ioeYhQSamGglIRnXW7ZxL1Qk9LrtDxpX7CxIb+yNbh5r7nD1ICy9iBFxfdSySEd//btoOZAxP3nXNCQF4qeiryQ56wq7kDKs+p1ru7EZkazi/qdXy6FMnT6pUrP5it20AuM1LdRtztqdhN7cRzxuN2pmsPsoCplOqN56r04uSUkUYDSqqtOyTjBtPl9gs7JXelhZJ7EH3TWwCcs/8cQjDNwwCiuvWLDFbKY+FR51b6papEiSrlWx25cDJRLV7pYkzqeF6gnoTyj0pxSJ+YnPjcgbqUYp2OwCjNK113Z39BHdIKGV9GDk46TREWRXOZOl0MdX7VC7fjo9HfTMN1tDqVhsqWX5qFGMTRBBaYxn0r/T6g6hdL+GkMU8tq+zvw2HoBoUbZA4KJZJasgxXnMVRGKj1bUNfl27RyUiuvEGEQaRU/lzYGWK6zSgE4vFApGPEozNaWQUKJLZ+OdegclfsmMK4EVl286MpCed7vKE1T3PjTAlteA+ry4LXpgo/ogWBvQL1Rd6euuMKN2O9Bep9YSg85jVmIWq1TO2h9kn6VYNmklN2mdcZpeyh/688uXHiV3vZSTP7AOHc/vEKWw/oogVUI1R2I1uAIeCVALOynyTDnrld3Sabi/FVtNpinHBhSUGynXdAIxl643qDvxUckSCv/HIFJBF441N4AICUVZig4LuNvBV5CpcJtbegW38iT2yQ/DN7N5iZ0Bld6nv9ec++RAY+QnmlHZFVKmAUjQ7lrZFbDh0ucO9YlFKh80O193u4Hq9523q0Kp9+TEJ9GoZMaYYNSzK5ikURsKVEnIpWkEU8NoLzlsf1UJ1KJMOfiD6iI6c2roYRqrk3KaCqr77R6EBsQnwyimVBggBFp+kBUU7Q5MTPDRFV4pmkfnXp6IUtmkKdllvaHDlH6nTDlZAyvkwUaBGIWGM1zmWWcwMgfTs/BiPsGkclB2jU1gniRO6frdUu2qJZtcJR/OdZm8E+oTy9NmnoXtYNIW6oF8EjEnEe9mmBOlB6k35QnZlACcJDD9qzCTxlC1qkN7wO7p3LMKtTy7CY9UfPHSZCOozYdQMriI11RxnVLyX7+6rpfUpc88HvWS91HhUqqQPM1jlECdoOoGrmDR1iBXiVEG1gnm37yaqcXNwlfd8ChfMTF51PGQUVx0cwrdwWm81ms0Sqd+z1KO2x1pVFX5XBmY8oxYZTvaRXFK9hht/lostJo3s3mxjnyH6aGAsq8VcYTySo5WfXeYaRQTTNMRm8iOF7ZRxc3B2hk8seK0dxA4Sal5sIhOsnVxi2Ev5ErJH+IOoPRiV6C8Zjo5Kcdi3Pk0wbdgcxdL1FlaHrHoxWd/rLyKFegeTxRp6j2mTaDsic81ukHkjeeDyaRKL5mrqM4XmG6Hws4TSxo4gzGdyGtWY82jT64YGeSsI19tCla/tVuj5NfdrZhjKI3T3aYrKOem9PwD+8H8IoWYTlB3KQFUCfAqOLWUQ32qCqIvfBomNdsAKpAnu20J5ekq5+RqxrM7dCFUx6jLTDLe1V802uE1JzS7NJxeXsj4V1CCl/UBo1Tu63dR1oavZgze1FaYFWq76bhO4pV+HH4IVMQoBdJsrxElamkKnnAT3OiE6pReLxH8iEcGKgZsMfnKIsrk1lG4iuorb+vPo8mQb/G+s/PkquNZwxlPefZuM8EFRmMUK1Tyae8rOhU1ubzi1ILF59cILk5xVadgUpGmkihZeIZTHjY8VXmNUUlGvNnct5Irb2ilXZhS4xQcRZ8EoaSAGkV0jNFZe6gBPnUBA4XOQBvMJfdr4LwGHtPIOEcrG6F0IFBGxX3ppaP9C8pVAyjR8IJsSn5/UA7TJ5ZzrLoS1l2myIDYFOxYKMvsO94mFysep51N2y18GvZ+t6u8h8GqAZXYmncLGDlxZTopXTpBy0uyCg5DlXrB7InAOtbyrNWcwzkTGcjjroGSb8KoVSeanTO4q80wFd5dCZRnVBrxicbhhKya2JE3kHasUoHvFNQ9h7znf+eGyewiuq+3vAzyRB5K4aMm4G6/GZ3TGT5JXHflS4NQyj2dGlB/BvmKGCi0CVPjFaxbkpYmWAInfYJBKF1cr+MM6hiqpfI7IEnB7QMt3cmT1iaqu3WzGbmW8eD81xkDq+XtNDGc8CgDZrCg8E4nGvGcTKWmTgmnFibTZa+7Wi5VzCZ9xmBBs1vIQolORVNwHg12ZtUpzkA7KGMO9IzXl97723c5Y4lHPJSp3RTQtoXBIeeUcBAu8LP/XCr5cSXmx1YTfiWCAOTteNTtWJx6S87+6Ro+MmzCFSTTwwPln3vRzoDlvE3A2hSQqsPfp1Thn8GiRdXdi2k4yy/ZaO95WqIVQhepbAansTzpdEUR6uGXeR2j3t7eg80nx5qcqezylNgW7MLOtNpDOhX9Nkol1Kdn2c3Y9Mgb6FzlaHSBt2HVHH/W3EOFE4uNfq/KGtzit2R0owugZKD6+fbWHKcnFA0tzCxAS0cq+0/ztH+VpKmket1FdlIFZOqlL8gkUPDzAvNwD1HurZP4JzcIQ9ltt80w6u5anr028Iz68QTq5+DCTt6YkSgh52SFSy3BknMmZk0ug6qkUAqmr4DVXio6ZtOa11DFiUprV3cwCd/sMdRNCZQd8notf3v7Ee6o1D0wzX1oLhOYp+Yzn9jMuzMQeJJzBhcZfl3XizGC92SKaTo65snmToXYtIbBShdCeQvlcnKL1F55z9q72AcVaBBWeE1No2S2Kw2kVnI7biDjQcPjdnfpWVSsF88sUK3aqsZo6c8KchyOr72Qu5D82sUGUHV3v8mTz39b5T0pdfHTy4ktOfvOHR5I6tA1weSS0I0Ph7sm5Jdqx7ug+DLnT+1V4zmcgI3VzF32hH9XyycVk5vhRc0unlH+8/P9DSg1Wy0/0fkKnfy0X3u7w0PDXUmiUzMNLRP82A16FSuuhO2ORCkzr6DZLbE8rR4mGeuckl95eS6hZvgcR7958aUHhAJKRdOLugdOPLXsCM0TibkcGJh+R4S61NIPwdFeAX5r8G6i0BMsA0+Qc7RpWaPpzl7+mshAadTDX9h5Rv2NCvXWGp+ruoQRFI3ECVkF0iR63gg1T9obzGZ+Kb+LaHkEEfy2H1YnNFA1qDyJft2diswtq1Py66YHF/WeDG8M+usVazbp8/5nji591FicZImAYLXSm6fCMd3squ5CFqpetCyFvpzqDvRpydH6LnKZZr5bOwPlQs3b4KzVOqggYVFIfQaXPgq2icz4jhKCRC0vgSlIUUpXUaPEPxXb8siFM1bIpmUoUOvqnlBcZQhe48HFjS7alT8e/vje4vT3r1/f3/Tnx9yFK3JClphSnD/Nrd+lghpVyEPprA5JRQCpPLPEjNprbnmhT2uYrbjUlw0Uu3I9vpjnONxBeU+pn2/2894mGZ+SC8dw6VJkwtthShOL0xwPw0/9IZ/5m78ppHaBUtaA5ClKVfrjgk3C8bWfg52Oyy2UwuoR3Gse4qR0SkjEFJsAKfblKiuYJDRIesQru4wX1PIdmt/RpJediTosvG5wkRce+pYn2co+umiFcvuEPteMjYFB6nv3jJCMyKhRIk+Y+6ZCuQF0PP7sMNXfcUheAh3P8F2gCk8x/Fsh4TQzSJ+uvOzcbvrxKf+mMbtQ6JQ8wmkXqj/aH6jYnOS84dRSA2TUNBOT5sOr399hv8u8k5LfuXU0CIdbKQAnGvDClnfrcdI7KgPV9/e3wed9h0oxiaFClGCMKcwnk63M+J3ohN93VSoq0qT//laEgnAlY/kteWTK/XFBeF+g7dMm3U47qFt340P3K//r44pWAARh4EsgC4SogUGB/f9Xlm26pVsi+H7M2207Pce6xVkBZp31hKbUDI8VFGHDygD1navHG0S1YbDmshVGWj7FT0Q51kw1mkr6g7pePbVkV0BKYkI8jOrF46fwDxOTFVx9UUNJDwkr7Od32Bqai2DF/J3tmmWripy5qTG56VWJ3rXr3QX0M4aYMXbdMBhgGl9yPBtgslC6ATSwbs5Tt8ovAAAAAElFTkSuQmCC"></image><image xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAATMAAAD4CAMAAAC33vGvAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAADVUExURUdwTHOC/2OJ/128+q9r/1KR/4Z7/12d/ppz/5pz/7ln/1es/VGe/2PM+GTR92PQ97hm/7hn/2TQ+GTQ92TQ+GLQ9woLMzDMpP///0lKaLhm/2A5mqNv/4SFmefn62XT9zFMmU9AmUFbsywrUBQcQHeB/0BGmTdulY14/2VLs1KS/2OJ/8LCzJycrV/C9z13ryg9gDNilg8TK1Oj/Vmz+y5XmCqwkzc1gEQwgChMfRpZX2VmgHV2jB9ybBc5USaZhidveNHR2bOzv4tPyiKFeU2extDR2frxEswAAAAWdFJOUwDf39/g398fH9+O39/fiWBWxcXvoD03A0W7AAAKZElEQVR42u2dC3ubOBaGPbvZTSa7ubcGOQE6gzJtOuBMk5bt2MZ+7Dj5/z9pdeEisCQgduPU+V5wik3kjF4fSQcMml4vZ//98fnFMBiu8pUvq9zLZZXPfGniT7mYueYrW/hq41O+8NXGVb7w1cpALnwV7J0dnvY0HF8Ew8fbjxlf+GLkG1++iUf+s+ADW76Jh9xmS2v+yBeO/Cn+yTZr/MbX3yrP5IZY2nPJV7aYuXlg3g4PasZOLoLHaR8YCW8Ge9VYO4axZmsPg0NF2bvgI5w0c6lIO4ayrtJOoKy9NNmn7R8NIaMtD4MD2TIpXLQeCGTrPLqFio6BdhIgy+gUaKxHO0Zv1gneOM8f4aFT4zzr9S7QnXVMN3o9JGdwBmdwBmdV6GiZOIxkNqJw1obRzFFIRnDWxDRxauyutc04o6mjIaVwZlaWOFoSCmcdle2qtA04MyvbUWkbcJY6FlI40+UYjpUpnK2S2J0llqLE84jc8vItyrZC+drYK8he6efPSWjoJryFH8XE9OfiuWXvCzprCDPHsaRpsetGsjausuVneyO3IK9n+Yqne7/Qlztj7c78/WK6bWezJmczS5zlgiZuvsU0LkpnfobijD/lNZ/olfmRb97JdvMf8y07o04j5o+VsiqERUiRbGtcOhvXCrDfEvWfl9FYDds5Ff/od/oe7YeeKUhfzpm+aSZpu8YZZSFRNjc3s2h1Jtry6rvNZVmqvIca0tm7cXl0q860TXN2d6dIW5pLT2TfwyqUdW1Kd2ZzFmqdRZHsE32Ns7LN8+Aeb9WZbtRM7xitRs5MEasQ8cWnP1E6cIuzST5kaDt7ndB5+WaxYZB4MWeVFlkqqxwbWIqzz5yZemLmYtGhLcoxkjtbZKmF4ow/56NDaHxPT+dEaa/jdUeBDTpL7qai+2LKqmeGLMWFo5C3GyJqKh3Wcw2vX8815tQaZqHNGdGOEdtxxl1lyqpttd/QoRHebihvbqHa5spcY6LUnHVaT74lznhOoWl61Th7ei3ORJucsh/1sdRSXEiKRX2YIkrU+lr6M8/YvLiyub4TCMvPKXo1Y4Do+ytDZlNSy+vis8dceiCx0p3ZnLEe0CUmZdoQjMpEd7HtMWC5MmCmThdnzNK4TDgitaexOVtoc32h7Ck0DAx+2d+RrTob1bOMZZcDTjmIZXXw+ZHNopUznmNp6s0SFjeqjA40F0jzw1AarTsEbPrYKU26HDtldcnGSjFOjivOPCKhpTP+dDzXjo38qDWSBcLiE4nLQHPjrOh4u86aj9ETe/mn4pQGr7JqwnpeQ3fMuLo3csvcJbafE/lZzgXlVZkUHY3fb+dsruuRVp1N1Hf05C5/8lOfcxSmyqZXtqliT61t5s/1yVn5+8UvkLHSM1DieROy/jcU6zubrhVmb/PcNr5Dec7Qie/qNikN3wl3loZrD2zScI3Lc9K01WupdvZGjR91zd4M1+y1a6Gjmbw2dIlrQwGcwRmc/SzOcMtrd2e4F7Eb/F5E3PPaDX7P63vcW90FcW/1/gUaZwdu9uRcEQi09kc7cq4IBFqXEWDvAHPfdE00TjHHUldlh5jL6/nKeKQNMWlQU5ZRnTOO9WlHwS2GT7uxQX0ezP3jI2UOTPsUmHIezPrklzU+5NNfdpgD88Mf6jyY+pkvzdNhZnNfdpn+Up0C0zYRJhM2WJkDU8Qan2vVwFcD91paTLXKZ1ttRdNUq9ftJlotJlttTTHTKuPs8FRnDAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIC3y/7Ju/OjnH8a+Y+W/67yr2b+YeOXJv7dil9bsPfrnoUz0/8e/eQ8CIaPt535na+/F/+yha8/jBu+5kv12Q1ffwQPD/x/K39aN/b+PHj8Qvv9/l0faKDfrwZ71Vg7CYZ/Q4wdZu1QUXYcPFJIaYy1B0XacfARRtpwWUh7H9xCRztuBtlIcDREw2zbPK/kQHASoPtvTShb59EjVLTnYSB6M4RZp0A75YMmRHTp0XjjPEfT7NY4z3q9CyQa3dKNvV4P+WzXvBbO4AzOdt0ZnY6WsySZzdIpnLVimjoKSwpnjTE2c2rsrrUNORs5GlI4swTZ0nHekLRNOKOJYyChcKZn6RhZwpmW1LEwgjNdy3SsTOFslcTubGaxTQipb4WEhNluUpC/EuYvmN/Riz3z3jHfG27f2dRxnhto1HVdWcG42IpcN84EuQVRVsDLX/Bj7eBCfLlXr2Ui97re1p3NmpxZAo0JmogNXptiizQ7K19SKUr4OqNl4XjLzqjTiDnf8LL//rCwwLdoYcAPM5TfZ89IGZb1j2BORLBpQmnM/wSRZSfbdTbS52Vpq6GTMC15fYqtqIwaf9WxV2vBtaYeyvdYrO708yKxIQ5fztlSn8reLdvkaLwivJYLZkpGTlzGiM2Zp3XmeZPsk4h0YZZ1c/yPTrbqTNedJXeMcjhNrB3aWHRi87HUMS8bXUOcjY0fhHbnorTs6eLwJZ3pcn+uTHVpLj0RNWF2YiqCgyqeuLNaasHDiz9n0Tg3tPY49rUDxFP5YZDVD2NrzhIqerGUGasegZpLMy9PwhxhscZ6GaKEgDJurg59nqFHGpvGVL8SwO4rccY6sbtUp8zijFeF8pZGeU9GuJRxK2e+IXElvq/PNV6nM2fEbKX8Rz3NtRQXpkRkEN5MIzkm2NomTz3GvjbXyMrNdZHml5/G1ttmUpPGqKcfljGAN8sJEeOY6NB8pTa2MWCib4DlaEwtY4C18IuPmyOdMuv5ICZmEcvg4tmGmkLYnIW6UGH9vwyluSYKJ2UJf90jgc3mtCPRpXU5HcQiK5K18YokrdkZ0XVJRfz4SgtXE94iRt1wq85qfdco7Xg2KC4OAIno3GkbZ0SbT5DsqMgrC4aR7yl/yAtZ4rv+Aefax+hJ0+FmYi0ukoNxcaAeaY64tcfoulDhWvy575bdfVz+Ip2Xg264ZWdpkzP7mVpa1n9RPU9jdabNNWjs1s72eIrcYm+07hm09c85rhVmrHvJjhJZ2/I8L6wcPuYUWUL2fGJKNEgcRYtYeRMvJtW9UUxewXna0Tph9kbPbdvPOs76cKbrRizDwE5+wbmR74Sfc472jTszRlqym9dUbeYaF7202Y5eGbSpa6nSt/EV+kad9emyGmvp7l60t8lrQ6e5tiSd7vJ9epu+Bpky+jsOrtuGMzh7vc5wj1h3Z7gXsRv8XsR3Q3jowtUhv4V/ChHtEfdW76ND68LNgE8WcYzpNTpk7XKuiP0LjAIdw4z3aF8go22ikU8YdI5hoO0AcFZMSniESGvDd3XauP3z4BYDQVP3fzk4q8y0dxwMP8Ka3Vhlmj0Rau+C4PH2y9/d+atc+LoR/icfYs0Wvnbmu3x8lwtfn8flw6AWZPlUq+cXwTAY6viq5f7rvYHP959b8GcD12y9vuaPJj5df2rDlXhcNTO4GlQ5OzwwTlH7Ojjgq/ixusvEvixz8EOoWvo/aS69LkeOBHwAAAAASUVORK5CYII="></image><image xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image><image xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAg0AAAB5BAMAAABIGg9ZAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAPUExURW6O4VuS5ESX6EdwTMLW9Q2YIQAAAAAFdFJOUzozKwBmW62ctQAADJNJREFUeNrtXVmS5LoNFJ4vAHIuIFIXKOtdwB++/5ksMhMgtXR3TYRjSpStXmp6/ioDSyKx1PR3/6w5TQ9+ZPtSmcKkIS+7N/6a+r9m0cdioCIbCFp+pqBBVJYvcFgfDcGk1R4KGEHKoxovcVif6w9af7Q+fNmwCCFe4PDPxxpD+bU5hFRzAATFHMrLXyccnmkNUk1BaBNa4XAQQmgW4TjMDw6QE9/8hoUoYdAQymtY9ji8HhsZangsCEwlRBYzgHNs3wWKsMNhfaxTKGODOQSj5GYRROKvHoeneoVqDQ6VP2lzCakwbCCUCLE0HNZHWgNjJPiTTuoBssSFagwFi/Cr4TA/NTZINYkJrCHAK4ozKJCoKaOGSuDw2HKiAEE+LYBCQKIC0uYeh9cjIVDQBvOIAAKlCAv4rn4RxHB4JpGmX8AWgkyk00yZQhBoEAWHxyZNZMvCG7ZKm4mi8qfQnKI8/wAOrwdCUNiT4F/BY2RBAnGSAcLgAA6PzBZIFYIgicoKWAQ9WEN1jILD42JDlRuQMKVUFB4awKQ1HJH4VXB4XHjQyUtLsEhEBJQWTBXaw5BiweH1OBDIpaFEqoA2VCB0D0Ch1SFtQBQc5sdFBnLpqXpEsMqqhgakiyMUcdlw0CcWFRUDYcZU+oQCiN4mYnGLLVBuODwKBZOdWnAMproEg6GPDPhJv17T+iAY8GVcUhwLGANZ9M4YSngoL6/pUfostFiB7jJZbIDaQPXl9GxQpPyaHpMuKMGJGUPxCAgOxOD0pA2BlCoST8FB8aVTq7HVckWNkufgEBEeQtzyhb6m+VFeIabBAYzqCRK8yJbeGBAeUgEizc/AAbUlEkaLDtIe6nDNJBIdY/uVYozzpA+BYXItksTB2hSBWeIYIFJEhNis4Sk4QHiCBmXJkvmSRnFIFDGSOlQUtu/5GXEBwlMDgW6hnUWcs0WMBkPK8yNgIJGezCmsgSlQXvQqVRS3qGAUONL8CBjUs0R5zxORgDB/toeSKPFKi0iPwIH0oQpQlUqiPwF7+IpEuldsPlEAGT5KqiUJsUZmFWDUWtqeLnYVZkJdgdeH2AOUF2ndbKEwLXosrCA2AILyLwbKOHy+0MacJpUWFC1TUpjdcYbQbCHaM7Y9uPym8AtPmtUiaA96ri+RMJgp8AxuD+L8wR9THPQkx6KkQD0BDAhEHjpOSiNOLWmKNXE1BLlKFMnVFwAxvF9oI1GC+pJsuuZJMR4pR/JUrcDMwdwi67gwgC94v8aauBBgAlo2e4tIHR7VCmgPeeg4qd67a5ILIgP+OleXyA+MCx4kt+gwbn2hzBUqVKhNddloNb3h1KaIFOktT2RAEMeNkxUE9CrUdRaX4Cbj0np2iZooYA2eKzYghrYHaYKDjTcwJFyUFK3IhjvUsFBjRN4sYsT4wDEXYZwsUIRA6hSCtzBlL0yzW+EGUeND3l5yAWRUe4A8ry5EThx2odRwpdG3NFGLzBoVNhQAw3D2QH3eQmQb8nCDCMcWZqIGBwDwy5IFAuWo9gAE6BU2GhlYX6lepArIT4yNNAjPFeWPeTh3EE6Oe7PGM0Wnvumhzg6sshN5U3QUtjBZnjHtAbMudebHJn5EaAeip24NqEN0GtXMIRcYRqsvpB8PNe4ktmKDAKlXfdxo3MkcooJQjKH+NZ49cHbeJoXZ0W/zHXoBQbR2xfYrh05yqG5RnWI0Xi2cGJ8cBWHGtMHIkxIJb6hQJMuYSBEkUMRiIByURJoUSru5cXZz5aQ1MD7QAlxtSDmU2FBjZKzf80i2UEcjbaFEfO2OksuhV4H37/LTTm5IiA25GkMaLj5UuUF2Ez+uOlw08JIFydSV2bADugaRGAcH9bkn27bqdJeqQ6LS1v0EWIIw3ZRIeIcRByMPw9SbahC0vRLq0YDgspebCIULssyYzR5ye4bxCzUKFSZ1ddoXjA5gUHBJhAKyg9GGWljV6BiTUalRcBDbRUV4MOZkgy56ZQwpspUbUv84fxrNHtQqCtJJsf5lVech0R9Hn7yiIJWOLsgWIBAWkDCHwUGdQVkfl4PjtqNtc9Mn/QnSk8uxhTXUZJlqSdFbwyB+4RNg1SdEu1GfptJ3hVXskkT0rFl4dU5AIR+fm+OApqVPgNn7t7VcudClY19wNwZp9TZjJLNmHMovlAqctS+tjRvCUZY22pDgEF18yHCOqxh5fxzERuhdfDLi4JtWehBkI4dkqUZiCqyEx8qgGSIz2fQYOFCW5nhsXbyzyy5oYX4z8mMFdhGl4REJjQp4Rsp5bxJ39wvxn70IF1yGlL3egCGw5GIs/CGSNKSupBjEHsig7UdVdmcL5GKGniIc2xQ1SlpHt0XGCO4wkl9M3sGz4a9WXl5xSHBpCpHBgOi1uGoKF+ZwXxx0ajtWXEfl+QrIDSLXOyVgT8E0hhS9mQtjuPSKe/uF2mkTFpjWyFVpw/OHfg3YUxEijVSTONX60hAYKj6gfyvt2NFhEG5vDpEdzGRkcqsrM3IFayvQp5Svn/v6han00rZrvHl3PTzuXVyvsVlbVUuwsiqdycNdcRCxS2ji87KWL+2Uh5wRQG2F19SGXaroYCwyjmMPYoqLn/jpJEi4xQmJ6FMedTy2qNHBa+0c41dv/9b2QGmai/seHdRRuBBkI8QnUMhqFxk9K0pOMboaOU58wCYB5lx6CJTdS9klCtu7c102NsZA7SWTRKWBcFDxxhXsgK0K263RwxAYFs5Ma8hQ5LbMCd6UKwYpj+cXcryFxjSxHw/dD0unLmHWfg0CY0aV5X2rcXDA/DwvWTA2TNpk2eOKduKWNjFwZ4A5eIsitcbVADiISw6mTPuqmXCs4cQdIse/QtPeyBzcEmL82S1uZg+IDSI+FqnBt0quVghsZz/6OGTfwKRC/5NL3DM+qLZJuLZ6xqRxNgYbHqfiEPrZBpRUJjl8YxTLrXDgPgm3Cezekc/8hItRYW/uQ3hKTqWpu8AnfnCL9Yb2IC08KrpWtp98sZGabDzWFglipzfsCsxx/EK8miAS1q3ZK9JXkx6+XdNNw5lXvBUjb2YPYiMvuLGs0p2MPC8SRLtZYGuY1SjaRGDjDOlbx1jwa75NxsSosPgKARt3gkRx3DiL3s6N0dZsMhp3FCNT6iYcvo0Nt/ILm4/l1p2YW+Du0eXKXWSJbaHBpsCSxYY3faKgcQsclPOxk57vCoNAXi2qR68rmtYAAKJN/+R3yMOt8kV3uKD86q/p6um0S/IGXvDeHRsVyWaFU/6eM3hsuEt8EBtw6I5Mh3Z6vY2N66mLWUl0yMHHnqBHo8p671nuFx+MO7SxcfEFm4MiW1NFaGcsjDjAGdC1+rHKXMwtlhvgIG09e2rz0nZ6PZxXzpLPOBwa+41A5Tci5LLPF+t8gyBJSdYKbLUiUwNnf05DP7FdL4BLcDKSSlxi4+o3nGP+rDEYcaj/Cp4mfO7nijWk/mYB6opsfhHTO8WlmcF6n3zRPrRGqkAtfhVPrhfvfLrBfKJLmkgS0GffjZDL5+OkmkRvkYHfXlPJ1Y46s2M7a9J1bbKP+nwDxdL/c62/l8/7hUrrWOGQrPDW9sXp0NhfyPMbN8iXTqTSO3Xm4m6xfNgeVLpuLjcpJjuw/MWkcLC33teXbODl3dz4bz0fjpP4hBI2KuwDnbCkbd3tQ5/CdhD76QYrs5M1LuMbhrD2LrJ+MG+yYTV1zUsJ/cdTXNz5MQz8XgGHx12OpBiZ3iyuKiSfjpNqgz/On/yebpDTHJx1rPxugW/lgjdwzuNHt1i+AmT+WGhQHLFoFzMDJxyuj8JRd7Gjsuhn+maNUaj8LnlY7fvDfNK2SsTX74KNeui1JIuVu8ahM1uXkWMeuRHq9JMxLB0oH6svdOKQLCY96kcydNevMPWyl+iTqQ2gTyHtbnmwafMej7aEuezK7g/5hdqHe3FR3VfvwuXofKfBQXaJrZ9NtoC0md6ssnfm8KF8YZ/s1ZOo4FT6esAB6yTcRo1On/w7xjdow3qyiqX992f8Qu0eHE7C+dUj4f35wyZB8hWbhOjYL+UaAvEtBrXUr0OwWP64PTh9nLoeJsQW8S3My4npyPWaMuJQdSjUE5Y6f5Qi1/MfK+uLj9iDtDPTqm3bzK/Jng7JRj8d2rZy6Qyp61vG/H69vZcm10Km5j+cLNUnhYFHu311/Ykt/V1di5PdRm6tqyjB/aYkue5gmf94YMBHWpExlGtPtmOjeqYNybaKWvOy7WhjDjD9XkW17PW4hcDcru//X7A5L9dTmkOYQ+cyIaedF3m/a57+/f9ne/61+9zq/93n9R8IWADrpY5DmAAAAABJRU5ErkJggg=="></image><image xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAxkAAAJ1BAMAAABHjCBZAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAqUExURUdwTAQEHwECIAABIAECIAECIAECIAsVPAECIBghSw8cQxkPPgkKLCcUSUzXulAAAAAHdFJOUwAXSd/CoYCo7AcqAAAgAElEQVR42uxbTW7jOhL2Ed7MEeYKnUUvskv6Bsk7wCzeFQiIOoAXdQ8D9gEICN5lIQLmoaZ+viIpu9PtTqdtY2BGlvhTrFJ99UNKdlare7mXe7mXe7mXe7mXe/nt8q97uZHy1193a9xSWa3+cy83Uv59t8ZNldXqn3u5lcLWeL1CeXl9/YbKizXkr1b18vLami/Wsi7tA4tvtfelNkDQs3xpfY2fVz5d/sty8Fuv8I/K3/9drZ6eH+TgTzs9obPVdOS5HR3tguho9vMxiR7Pz0/PNlsq+uHOh1Z9wgnNZxt50pOOPYDGJ9mhDeUvBI3lAxg9YMrTcyP/A/Kt2gYfoPAJJsvG8+s/q9WXe7mR8vz3alW+yt+bHHx6fKNHbWknSSfJ3+ObnL96v5DZFCPXQaNrc6wTY6AqPI3Lo1zftFL0+qa1gsG3smiiai2Z9/gGKp9vx6OPKzsTYP2PLqLyfqyjny5fJbdByH10/Qn1rzgqeA9sjfV1C9VaXl5pOWqt/CscqWOYT0joiPmnyT+P6Dvl4dvVrZEVlrxADRhk8t4Fcg5TRif5IV0nqOeGfmXWS8+0/jPyad1IzvPLL9e3xrGO0Cd3HmqoyAB14Kjmjmbnk11/NbcN+rgwb8x6C3yGfDNt1312+fJ6PWuoIjnD7zpoMlUMaOGneeGSJ9khV8RzlbDwdQjJnetW+j8hv82nm44NOk6v5A6Z++TrursrW6u5uuImNq3jGb6ZKz4V2myAConO80Ug02/Kh32yo2/BZyJqD51hkSvFRm6uyXfpkFJbRUgr2dsOBxQnqk7uGtePAZQBrINyFCOVPMMUef1h+Z2tOku14X7lzzcYG3RsFoKLVlcGRAR7KRmR546MzJ/NkNaVARH1q3VvDDLLCxuLEnd86qT9unwzO5GbyCX47VVWt7lueE7IUD/3mqiz4qJIkMHla6qibioCC3CCWxtXA8PDbmEO94Cc4fPkJvqg/H69aB7hwzVaz7HI1WLDF1IglYGE95BpZsoVw61wK7sH5wzI4XvwSALObZ2oIuDaFkTm9wat8rHxD8hfr8EEJl0X5EkimDVTtxfodmg3ERuevyt+mQwZgvqqJXkSJ8sVpqPGEIiRO4Q0ZwQCGGZBzqqFWhGAcwcTVU9HBHxEvhsUQVncMB73XQD6wnQrsdFC1lya1tVNgVZvF4VyrQgWVbjoEFSneib3YzrBBxVsEhxKL7CZkZWPySfK1Ey/ppotxYLmalxbt+fE9Q82vFfaU1FdFavq2ZQvVV8gpoeMiZLN1QGnoUQVQbL5hRrszh0xswgW5UvNOL8sv5Mq/Aq6WrAgg/nyRjezbrSHK2wtc6ebqFT1qzoJVqxhgb5AZt3QLjqvVECEQtuW5m1AGBiwENBDpsa03l+S30p3M9DIDVy6YOue1G9j3WhPwnXrU+9a/LywYoDWvVLfnkpbhtR3uak9hp1SwPkNM2OmU6kg5pSomN1goVLQb13y9yH5LtoErEu1QvUV8qTmi89t7al8iSC4UjZIMs6kOJh67qprbSk6BpN5vvRmQ8N7C3l0wJVtTJnJ4aTq1JUKsfMB+WbY7J5gliGXR75pqG8C2uPiDawb5A9LtvUxrCw3wFmpwmVpuwAf6EmeOZB9rEYtMBpsilJBJmr+6hyLN1SUR8m58lv2qysVcqIZw24NhvUUaBsreu+x/NKxYa/U6v48AytVVAGEhQg5yMAonjEA2iI2iicpndJB7bZAUJBxMpOZFxh7I1Eu58tvDLr7p8YdPJEdqdTH8/yjr0EuGBu29fZdpqPmSrv6paoO9KwDSb71gUaxxHyA1/PAglF8FuU6sSNeyD1TPoKpNwdCtmD5MXMgyiy48Fz4Xqa6eGwQ3okQeURXpXpAjgFatGgx5d1C77bIbVSh9vHz5R+NOYucux5qFUK287c4N/K8UZ/AOkB+hOLnFfpkuvfnd3YiQjZDqe8i6aqxQXUFx8bQ1ts/iP6tFOrymiSto+8LrxQbvoJblso/zTT/R/bArsqCA7mKrr1ukH/52h6sFmXW8/hz9ebfQ2d+h8mflm9b8pqsrrtu4LW15amujHMZRzmVWU58jNann7nosDesOs9SlcO6jdo7R5DNldEMUkgCeRmdpZH+gnzvgmQ7dLrWTi3WHjLr+6qTndXFYsNfYdaH384YepgiqqbCYXqhC3SzO+7sRtT6DIceR+2c0aXUI/A1nrPxM8Y+Ubu8fa78FkZzgVSbaazE/EXttNh3+Wvh937pcKHYyOvuS5pSHwDUxeC7Za6Oq8q4m7qnK0hCOcND1ZfddfUzu9fXcPFYAVrw39EiwSaNZRxbxJwpH11WNynVlKOZysjmeV6kPqrb3Gs/i/ui0eUpuPRY04WrWaBrB6e1kBSKj3oWquiPbpy5dMmsca15JvQpzXicIb+KMXuZfDCcESC4HbNO8Qt2VYiO7z4AXvR5o74khCk6pZvLj4uGl3kGCEHG7HxMgeLEcrYSlmKagLmbdMzoHfkLQWO3lJjhx24dMWuNXYYr2N0TXf0dbnZj5FLzcKc+IxysFkLwrhE+x33ckn45dEYwk8Aygo0wCKi2mQJs6OhkprHEHG+18R/JN956Fzo0Nr41SuejHqw5vt3FK9N8+qbqQrHRvvj3pz3Lwyd4jqJxQZ0NZZew9McAmByMRmR9QQ0QgLd3KZm0YJxiti0+DIKfyQ/Ke6wCQ50nhpphK9yoR1gzB77J+u46ftF1A78HaBtbdUS97xD57zCP6TDuwz6EOCeu81ic93GOPDDvgzVS2HOL+w5BRuK4l+FDEAqhFSquKCs+C92e60l70mGOSiysRj0HoeMpJqbJn0XMsXzuVw6zyLEogd1Gv869d2jAFV+amj3W339ddZnYwC+dSg2NLpQZk8AqHhTdEA7s+Pu9qhkOUdKR4BQEjiTqB6YLgiOTsq5yTUom+AmMMk/oNLHwOYrLJg2SoMwsESFOmEMQ6mDmaPK5lx0iLeXHWZt8z6MYh09sGZUThIOOixw7dYbqosO/Q8cvsa4TG3m9XoaGLsmjQqqpWu4/ChiJQZ3TxA4czFRMsTcCcWdGRo2h0An4jPQ+pJTELsxtLxTi+GhI3PFY0lCIMoEdPMqEKCHC2Eos7VVMJz8eJPss5Scxntqf/0ZxDo0PiTWNqSgxFZH5QsAKVdNcNQe+mD39guNi6wYeNTIWDUQye3FkXdJeUGZPlIQVBCnRWEDTpBHF/zEu2vMog7Ib9SqUQZAReyRJM8oj4CpTuD9pS848Syxk6zKzjiZzryHWyRcjLuX7AAsT84stR7EfYiNYrJmcGLDy+wo04pnEvu/CL+qu+A7Xf2DhmapoRkl6+w6VXkVjhVigimGjYGA8pP1kNohhsu6kM+SzEZ+VKdEKAiClarZ9UlzVAtYP2akZxkxhIkIvXwf2Js9uT284atwdNChTstA5iAdE2Mn2E8FfHpTiP1Q8eXV4kdhoP2Ar/pTKzsIudjioA8aQgisZTe/k7hwMvBABcxiikQyaMByzIRj8cGHz1YSPIq/sB3PcISTGeAjVaIFNqwi7fOMSYyffYmOAXc1kg91+jcYQsJIEDcl9QIiMeN8g5sh1m3u9PdXyMdyX72DulTYTozwMYZhY3V0IouVm4oYAM6TNkJR4txnEDmyQsBMYJ568G4QuRh5igoEDhMnjbscDfBHC3Ubm8VAcmGmSg/+mIU0Tk4QNnxPLT3FX5TP3nYgSbpCfNNBMPt8Xy9aW+YmmMgsT7gzR86ScRpjJ3uBYaBRs+K+2buALP93aWmTo+speJhAzeptdHCYBb9hx2U5RQB22A4O52224vduFKU0bQXPDUA8MsuAsQDOGPMTo8mcj+G8UwY0QcIXPO2HGLBlK5jBMPIHNsxNLsVyx22azrfIHpom7LfdW+SoPI9vElCxIGE1BbnSoiTYioDz7Jn+WnO3dg9mDcm7/CXXh2Mj4iXf3An3UHY3c7UZwDKxu2k6qtiC0HTZbBnIrASLIs89vuSndbI/tJMYQmwQ13LCdJqYWGwjUjBt3pq0EyFZtlJgbY6146iCjyBfhoZJElMSKyucJTCHyxAaQH3XmVkZ2OsBhwxEtoTRM7EaJ5wkd21hiJ4gvBF8BdUNoG13/ppy+997wYu9w12v/0d//qLu65MR1JrpsqgwLmAc2IKl4piTXPKfke9+h6mMDntV8fU53SwaSTDIh5g5JANuSbHfr9L8cun1byFTwgqQnzSa5T8LiqJtyg5zp4NHyJQzyPYW9toXNfId+YrtM5IB80f3Sk8QEzsid9tLzjzx/OdolvOBMk51fzyHbWwLiHwwuHJM9A2ADvoFlAjmIPpF2wOTGbQ/z3ncWYz/31SurY8OT4RYvRCjtX7H7REjh3kAcuT/cM24QPyBlIZNGbkyTHxn1Dc2FEHK4kNTD2BqhSynaCezEcXJVjw6TDlkUB3omHQQ9il0H0DPaaDq0dimjnlYvRUCxBbBGIpjQgSrqrKFYNOt3ozEXmlVaBfY8m+qHJmCNGTRzMGcMFDaRnSH27htjaVPd2UHqOzb0txhF0VFpNlpfUFnxwl02ZCMvYWIs4I6pfTdoOJNsdijy9A1nGRQkA9mhygRfZL+IM7G7VZ97yF3Z4UWH+6fYVDeuH9xvMXdGXLdSpRS7bd6jErcTVHdishajpJJT93k/6zp5/9Hb6L6x/Tka7Ptit0JLT9A+DJnOygUzeMmcFS/FhR8+oGaOI+y4QnPBdEdXHYaNZ8RwdQGYhqh2qjVgL6rCUFEyliofqcRaUhxTls0xKT3yWKL8pFIpxFLJVXZN0r6SGnWMclyIEKV1QodJdlX5kN2gVpW2ueJwko885TGmKG95krHGKD940/PLmSO645h0o8aosgtniGM2Di/47J8+j47lpajCGmhz0K6ggbtTRX7yPMf+LjiyHjZYn9ADhuJSABtKzjoJDeKUeMsgay5TrFlIUzPICIIJoyI2s1BZWsirCv2i0BWsq5N8ET4KebOQvqCvDJVAd7YtGELGLeQrCS8NyKMoffDB88tPBaOwI+h5ZAhhROQVTRwu4UQyZsVUksvJk4GmtC/AG7T+oObX0XSHVUNYSvZOja+BDV1hqTV4zL4oNAzoP4VEKQndOF1BTSElSACqYl5Wo09SgGT8ADegPviYiQ22KeBJFBInjKGMEajJuwwqh0fwTtCHjwwSy84qrJR+2CHjCVuln0wGuSLpKuevo14Tz5EVLXkEYHGxeZoy+puiMgE3Gl7ESqYxvh3UWdd8iuZkFwtsVvc3dOmJFybs6DZTXUCiJNwXJY7QiczIioxaFQ/4wwc+gSMhXSZFAZhJQQMYcAgABkOAmIIKORZTyGSBHtXR7B1UjqGCjxgNfBEuYQf6hCxXgENCczkRJkbOHCVjRog0y8TbhCbGE+OEaZkjTerjwuXwtNOP8/kpMVwWUpmvAYsKoTwYtSNEQZ1C5nx3ClXM3ugvCiAhETlEfoBhEDEpkAEKFqVtBGMC6K9cjEAcjlJTgJWBo3D8ZJ058VXOsX9W8cbTjNyhE4LvlIwyvm6S8RXMoXTEsFNT+80MEGklvy/KEC3rsTWLN8GRNbBhKy7d9YOzITbGQJMRM5sCJfPOlYh6n5z7lC/KFuymnoBAJ+3RRmawUgUtRn7HBvfKhA0ksfIJfQPAE1uXBUiyIoOsCTnauUKJPjr722ilADzQTcDaSGxUbBRCjygx7Q5fH/YubF3mwDwFeD4/CRswqZQZiIrQOaXDB1MG1IghgAKgof8FoVHIy1ckhGSntAxsG0jjoH+yn1/tmHUoGC5qQ92tW4Eb/qbb8o5BOEqCcIvXF2CDBrBLWQsIivRSBkVKL5oJQEmzjokOUR5dVO1cc/hjL1bVG/seMRRnFElX0eJARoYl9CtQGSSlrwgiUD0pvVKogfSXraoEF4JIDzZAF7QG6+S3YLd8Vm0KfsYZWzgoVAw/M47UCEpLM2WAYjHp+cE+NJEuOBW+2vlxKcrTDI6hqTTJAJHKvmjwpuYHgLOhQz1BBNHc57CVPE/DhkbTzb5lXoDIKCaL5Z7lhmv4JTSrIR6EciHJD6enkAdMkUYJNAkKDjRkm4SPgB06ChrKT8KeX0GPVLSTUSLbsiP2Ru+Ihjw/21YexDb4p+fnuImXYPyIXX6aOZAoVmlqASCFok/VB5kxtMqgS8PG+jHcvWXEPWIo0NgcqTU4q8gBzkElHGQBBY4TjfTyjwAtDT4REMFakVXaNXEqg+iETgyxjQ0GmDBShgqLZhkmca/RW8fMztvsJ846CTgfKBdNsrYXlE0xqxpgh8sCfpgGGQYLWJ2s1m3vi2tWt6k8s6GOH/Jto3puIJbRQjkSjAezzWqqFJU8Wefo8pWjkcxeKdy/DvYZtZm3nY1Bdoxnsq+H2Piu00MZVilLKSCJGONHMwGifTFDmZ4kEKK21VH1uNUsXi6vBdVXiuHue7XIbotaDA3TQeTqfFxQBnPxkEnZfE/btGjV8ML+0bdj6AxWJmRrS8xgenPnQeUPj1QOnRuLlTf57gqUIToENJKZHWBFMWzQQo/U6oRGbop864H1Xh994wCuFcM971vEUJAxvMDBjV1EhJhCJ0hesKdR3BpSVsRXeZR96jsNc3qNoVfguiP23SEbtUaVUtYo+pFMePMth+z2leqU4lqEPsdRzSotILHq6OtnK6zkizs2UFMJL0gs8DqNEOxdK8R3KPbaseg96+vo+R0j+qBvnllVkUpLHzzHdk2m3oiPQnvauLDwk/LCzF0ojmZVrW5T6TMEvCx9hxoYuKo0Dt8jxQNf6cHjmVnnQKH6ys0/MWBE9dAVHMjewufoDsfFnrXyHGxYpmmzk2sSpZFKTeG3kuTxr/oFFtwLMSqg7K5SV+iRAQGqc1ccEFZLo4rLY2/Sf2vojV7UdtECaGRksjpQf92LHk4wGzkGNZeDsaO7/MqWom/KjLIZzKhybFxuno60AjbOqsU11YRM0wmFHshqMCJxaCbof/Y1v6N7VJsbL1STNIMXb12PQ1h5XZW5f3fLxlfRG3xcnWHj5SQm1cTIdHoQSVYbYWEAxLAwqoJFDbILrKbKo7l/4EXRWJUu6ljkONbWG+cfFhfRLCwKpyiqQvh55aB9y+s18D3ihBbrNNe8KxB30anAy8KyMht3o6GRtph//Xoqe6iWrttjBpZiKs+fmtaHmx3zOxywg/PbHHmNKX/AJcoqjeaEkJuF20CyNHFbHPfU9cb6MVxdSmOSCpkmkaZj8kjImww4fFUize/Q+vA7Rn5MXsWmO3qghCwpS68jetJJ/T/mxs+Xp/jiZ5YZukV12mxZ0Ncs9vA5OTI/Wo79ESii+0lZoeHq2xIniopsISsL4wIbR3XGW258bX9jv3DFETbc/m9zHBihuobG/HX6z2trd3X91Jq68gEZIIlNeRhHLMVhFq4tcboCxzo2VeMFwulblirn/JqHPIe/5hWXURVPWfa3FrBqYcMjHQ5i49RjuEsbdx2bar/MNW2HYShFExt/6yu6lLIUrllVzfsrzcJlZCSb/zd0bGhN1e0TWdfQG8TGWR8kAr0haiOleB+jOHxV+j9cnbwbJskWuY8NHARGKD0xWDRG0iMjR8tw2FOrbkqqvh0b+6tl4iKpsJAO1ZnM3M1fFvTzN7Pl7YhVbqEqK68I7o0v47jMb+SWjnVseL7p5img34+Nc2eG2lRYtBJj/FjE8L+sSOi/5iawTG/3IFUPqmfygg4Hl/x7nfr+Kev+ut74d/OCZQIFNW2fC+B++7T/uJth6Q1LLDavb6m7WT/KvwYNx4bXKVzObd34qnpjUWgocwOrgT4dpvoG/hy+oMFj0+JWrkBDN8Tc1bgW8VBtuBLnco7Txtbxq7+xui9+toU0fD4Q9cYY69vMmL+Pqo+Rez1nHi1eaFaV1kmrSVW8erFHRrreOPlTqPdPqRnxoCEePMC1RsyDH75VMn2LaOtehpclKjaWLrhVi1pdVcy3JVWNHfubB/Gskt9oJdH0/qA3UIz/TmjkI5P4aeo9LwpTWgg3X2f+mIIt7oV7guPYkn+n9uiw/br5DXuaoZV9noiNLYvzFkSdv2kmL0KA80NYG1tNUDdmu2Wrhb/UFZoXv0rE4uEElhg3vWGu33nlOlzPxO52eBgH1z/GP0pRz19uND9ChuUGjHCVe+1pPxVXS62BuOHmeFU1Yv+8Zk29gXyjPuwdVXYbroOtOX6qQGH+YrP5MarGlbYXjXjd+pId6mV4xYh9db1xdAOXSzguN/8AYrV1f63wU/TGtEVicv1qkUe6fbZqwa1cd7+V+EQGmRGX2PCakaY39vsbtbFODLc/RAE2FVYla935/Ww8rOz7HT6pv9X7bjGqfLvGw/0M8/tMVFm14TBc6Y1Le1TxitjYL01cPkJBXPFYeVvzw2h4WMtzt2x4bKGQuAiKlO7zuRJvuVis4dj4UuVWibturaE9fKctFd9ytQBAnD6qCeZvQ8X8SWS4QUuMxEWAqtUYthL1hTFlcqopDsOG/3eU/co21QIbWPInzEhJY7jP9+Y+62pEJsJbwNYllYcHTW14TVteGFV8kJCV8JyehA0rUKdNBWsCjxQcpsJ7SeuK/S+Jp9wKCy39Ghaiqpe0WehW2XBl3noFz2nT4lR3kZG1sHHpRdF8tsiUPsCLrzsXDwZGNgNXl3pqYKRbU6WbVJbX6OZtOTZs+FozPqTqZv3+Gr64/WMBlFNhqbiwo8Qc76vTD0+0lz4QDHGOaOVt7ottFRgtp2GMcUh09TFs+jrM3eWy+KfY6+bFPdtEWTVwNWyK1ySbr6l4+D9117KsNrZDP5uUYd5n4B/wpjx2aVM9pqAyx5XwA/TXXOstA+bwMJtzSXcqmSUo2pKW1lp6QzK0z29drZWSzjZSp0BxQvDEoDCMyjjC6VjETU7DhaNwbvBxg6Oghmg5vO02JD55sWycnonECzsN6qlYWu7YlJRwFjUpAxriExVxkcqsX2TX9AFNrCsGKDe21X6fd4+3VM8EqW1m3GnI4pXEGknV/jL0kWaGA3LW2XpHFRTj1lSdiZQL1I1aXyoMxpZaqh2JTVL7poQ4zVu9nRkSaDrujKG1W5WXWcAR3oen8FSJYty1TWcXxwtx1B3DXS0X6GmBBjvr5lOf9vGIyLQHCpBAA97ghmBAHueGt1TmpiAdLvdUfWGcqjctDdpaLBZLnMXXumo6zfg6tW+IBahOGRpjhgCk5gy31YhwLNJF0WAWbqX8nUO0fSm4F+91L87QCN5rGHIDnc/S/wNJR9+q1AQcJDDRCZxiri3P39rcQj7fbrBggIGqKQPvontxFAwgTpX3pIeFj74+94ZCduCy8gPZhUMawbay18gRuT3LDPHTk624zBvle6pec4Pcq5fLZd5nZn6273iIZg9F0mAoMyQF0xdzthJc6gyciriIdLiLIMM8tzYs0FN9+Q2U4aXaoqsFzqs//oEaYbcgLxYFJDfjVR8P4iAP1tVIWIO7ipLYM7voIvuNXtQ0yIreEoNnk0Tz8Opb0xYIhvwAkJ0fyHDhjELZ9SkSooZIebz464T4uZDjD7z7K81Rjx0umejlTMBIeqWHen9m8ePUGJ1QqZ1OFkky9yUw6hRMzH650yIeWiqW/ZXNDeZT1YKoD8HAkwJE/JxaJc38TbcvBkUeKnCIMKcgQTY+Qp7+dJ3tmlj3d5B7Wme3zYro/uQ0rGxiOyK33dlTfWrFpEkB7riolELTZ/gblQSekpCcjYAdK2kkFgs5HUuM6LosR73+CoZIQ47i9ZM9mmN+w+A5vSkjTo9W8CQkdM0NFckgkS3SQ2Tsg2vPVMwNVu8fLTfqwro/yw0+t7hFMc26+Z6l0N5RPt6XGSyQsf/MdzT6iPi6z/0rrjxZFf5ggroSeHj6k01sX7Ru1KpQPuJhvgXeTGBNbIHP6fleqvEdnwUi2uNmods6EjJVOvheioSDxr9/RGn2VXwWt8PJfIoVcwN9zX/uhJGMVchZ0Zx568h6SdkIAqPDRAHnstHJ5s+tFIhp2PeF60bgU9GJvyW5RKdrMswfU9HB4oEIVWaPHQuH2OUJeUqKhozh194pPjillBFV72Of+Ql/KrO1OPJ53o5M+6+9KLN83+3Ui9U+8EY1hhSOraYxFOwCZqRn+/7TRAknLpXkBh6SHZ6I+njNhKeQH64c+luh7m9Jx6soOeQbat+VHqdX67ij6JoYTu2UaS/JdmnimQpKGq4bB86NlY/iH8CpVE3z5zdeH9NYvB6DduY3TSzdlakTrBGC3tXohMlHi3S1emc5qRV6KmmpZPYrfZum7+XK+1A20CaaXLvJ73c9c+/UzpIiydRLNmeYS17jOGECiHOGBCVdxkMAdRXE0tGHcGe8fN0wlgKqadAQaZfzz1QM8B9KGlptpyhHciB4jmfwqca2M/qt0AxNnxxII3VxTazmBtHbFtsK7Vf/fdNe/PRiMARaTkZkk6TITRDlR70MgEUkXdZv7KWobGS93eRXBkTYVFQTK8eTmWuI59sRUd+vb8BU7et8tJd3TO6JJwxbCQeSJIULMrEAH9fviuPB3lRmh3sgs7DjhZimUG74KhbJbfu8NhVmEaDjkfHbjgk06uHSaE4YVqjUqalWynYaWd4pjIhuxRWn6tWeqi48i39Jg4uTD1m35by5rVC+IzgzBy810crT8BDdugp6C04Nsa1rurZc0lgILoJVw/UCBIywXry0V/RoFkdJLIr318291sRFIXVhFOoT1fhaKQU9AKMiUwtwyYoqQOk+/AWbkbr+gI967zgV7cXZmxg26b+5CnQ7RxRAg8FTn6oCRmwdsHBkZyRcwW07A6g6Sg4G0xlPx+nv6IZI411sIY76l+3F0Uev2uPpFnjB0vwN71Vq1N9IBQHcQwXzLx3CM3e7BhROLl8ZL6TxzxgjolD+p+77y8tNhbiGglQt/g4tLp6Khut6ca4AACAASURBVN08wZi9jEtaiPB4+E1u3BPdLFU9Clew26oLiEjuJCBK32HZX+0HOEr7Gjqf6oh0KuRT5VQkGKcHyzgfsAO7P0eZ4ed/INsrJTqNdA0Noayo6GO/qCwcfp1Upr+COFX9FflU2FMh13C326TnhvH2LSkhE7fNf+aKzg+V23+p2zDkm+ulwPZU0zZx7V6oi7pW8cIYbu3n/rCnQk3shl5eCsfPOTCQ7MZTstHPpowgzzhjn1+swqtO1chUw7tO0sNIuIvDPyNkpDBOVRspGluKBar31+qlcOe//NMbkwjMWlhNn3X7HalTPH+PdK/Ts1813jNRSyW7JroUq672H7i/QfOm+hqu6AomXlWHz5aJCzBEVX0sG2uEyznyizTilE97cImIVLLp4x1TVynNsAub2JXcNCvvFf2lpvbY2/1ZDX+y/Z7kc+vpb/FUtJGy6du9VHnICH6ROYsvXiBNTeQExYPzglU0WDeyrsVVu0/hKOyHK5QR4YwcDr+3SwZG1vDf869PO1d+MBICyRynPDeSqisFRA+G6EbXSZf7DPzuGQyR3LCyYS4jKz3A8QGvaBIaElA1/Dm2f5CHu5drxDM+Pu0LadEorxDUu1METITd6jJDeCE3oMIqUKDpyZJybupkHsVjbvSRTlVoL24tLtYNBEYQU381EKf5PGFEpqH3AnTwzkHTp84IKfAR0jUYveLKrT9IC5ud2saL2KGOy3qjLn6bxo9orXDbtMXc8MOL72KJ3E2bIjKCVAzzNwpnye00Mq+UYIrKJjhhRVNG1WnJkF5XHypdNjHTsDCfKpjaD40E5QbRFHabC8VA+4nWKsXcgOQIlV7tA/vfzabSxHpJ1300i1NmZH2oKiPwHPSI1pXkKMIZEXdiNJlckI36/XXjfYEBdacHMTmysw1+qLqRkgEZfKMBU4lhEDpDI5gnC3F7GWJR2fBHsr/6Q7q/gKhzbgzT334N/9rp6k/O36DsTmNNOTXEZj0NRoIpqwQmhjh22+VK934Ul04uaEXmZ338gLapNrIhzeJ0W3x3RWhWHK5lQ/TkjPRYKeiXRmODMVZ7RfdaKRm9Y8jQsRHODYXTaTP+iyjqfXmciidxFTcdV8shZcVL4QdwdfiRavQWchMOvgbsFtz4eULvatsMmfh47OOfKq7jC2VF0zGUg1wKqsvfbYqk6NUSp789einsSjZQt5bgttLQIBj/PKkHW3I1PtxGbBlFlzjYx2Y/pOEeKTfkonVpDPcr3t9Ae+K8T2QyCbO3SKcHQ+Kn2l0oI/XbIHSIYZhqqbqQGxYP+nCPW3Xno/hH9htaxWUTu9j+XW4quviQimfElWkDRu5fKSxdDSxk9lQS+to1sLCzcFAgtHQwRqIPVafrDZo3fpFt97lgoIiXAhkUUx3/i7lR0cWHOUwmn2/JoPETWOBHXl0x42ChWCVEDjRMcdElJajL1TiEXZOY4TqFhxk8fdHdH4053FOttthSDT3VMP09kQXtXGnROE4I0crFrh/zekmPIMMNuq0gUpQNAt+GBawM4zhxmF5AGlzSbpfVxLI3mdeN5YFyY494OowAp8uvvX3rG5aS7cHx1QJVlNlPZlGop8nybWohv1Q6ZVh7K+9UYOFaPC689Eppm1S9P8wb3TBvINfwPXSc9r6HCnTma4yTkKKCSSH0lFxMlq6Rnw0U4ZpRVcJo6+xngQ31nVpIf6vLpsIc9dp5uOgWVqGV3hr/3rv31YXviTqyeA0eqsH5C8y+EzQpUhj94IKIINitCPxsx2TTn38YwQ0ePCVzQ9T7nhvbJfGpZPgrTlJQexNeZiQ1dIHQUwVrnZyipeo3ImQKSqduVPJSdSwyqww1tFso9aXpSzkPHqkbKIrlGwO7GYfx02P5BGZzpN5Tbr6tzGf5lcZAzs1MqMm0cks6xHBo5TAI9yh7cWJFn9EUCuvF8YQygoa7d5AU7ohIGtUNqRkBD6FSDn66wWgKN2QaTg1x9g63VFLLHVAf2eEeP5Mb5KMuri+LJTJ4AAvHrGzD9oH2Vodw6afSaPSzK9SR43nHhwPhzgm5isNGZ0xDH8ZlE1u6bigNF8vGb8yNDf2bK73r89wQmCrFmhHrOCOFjNQmxarSRYtL8KCSCVUXzqIyB6mkjHfnucGAel1c2+QWrEj93G6rak+3MNNz78/LXZZpAdQOzEhsht3aNcsxEJKuigM601pyf9sZbstVwzm46qKu18UvBsACuj+ybuO6MZSNw2JLp2k+cusd4majGXvqZPdySW4cYo7oaYrN1jlzR2ht47LBbp9y7YFSg4CR/viRukFl/EtvmmGHu99s1ngME52727Pn5/TmWOi8ISqNpJbP8cSMe+PdrhuqDKg62WsEQpsOfvJi6Uu18uvievChLIbLNwYQqDqQH+4WD1ojZLieo0i0j42KXL1TE1QaTUBuVfdqvCkIxlMwBRVmUTHxesOLhfZULhbntbjcQuEOtyjXsBatmXINV3RhLm2SXdFqX+tZHyzgxrn1zAgcT1NZgiiYsm/E8wSFqjLXIwmJoyG6avKBIyw4ZPorvBeXnoq5hsuhiu/pNs1ujvHhYTa6HaIWM1W796NxMQI0wDc4oZA8sz1I/BsDb02bzA+VOIWRQPnXUXV/VMP7sro/G8XRSwG5hhuY5dz7wxmkDa6fzhhVjCEIjeldsxpQ5QmhRnyvqizAbZg1srLTzbYt5kZvqGFftKfqzWdkCMbf5bKqNvee+5t72ycGntLWXl7cBZGQWW7cSo9KlxucF1Q18mjFFD4LNYp2Hi7nRundn/mMrFhOQ17RO6a33TA0uqPHepBUmHQbbvZ4zcg1Mpq6gCw20qQbt9I8zblQIiEYlQmcwkOl7sQj2V/RuiHmxMJu+7tYLpDAgxju7sV26jn6s+gsG7u7NAoHeBeVvlf1dQaDMBNakHQJieVGrBvMbpN5gxvcD3DU9dzfipRmuw1MlvHTW8cNUGmfeYLB1ePgopxJ0xRoQW6F65mNkRAeK0uOapwbcpmm/4iPuhOq0J14SyLMTdO8RjY8PdHfJneoDwccXKmfg9xSKkbKt+WvJtFnOptnho/hneCGgYZ7cLOwr7J68Tqw234jiNtlVG9AYbJhasJSoxlDIu6MrqVixLlNE5it8Ak71eibt6evNSgmXZRvrM7Pixf14KnHvobo+TnkxjBx8DB++h9x17KbyM6EH7sj0/thwQvgiLVlo38dgc4epOEF8jY/5frq4qbpkAk4OaOZnC1FuW7f5bvf95/UEl4cR48sjN7YpJh4pIRhHl6YPI/JoakoHVwoOCk4MVo8ldDFe2NG3A73VMWiVyJr2HNvmB3yVnTR0d96kfqsuug6fufFNZUxwpPLCj+Eyy+DV1IYTWby/Bu6hhg3yPFhn2pTFR99/HfPyBXVSfCeAU5vWNRURSLvvuWPE7mVQ3hIxeiWwR2agE83HK5Z01xuLQa6cGLXvsMln9hqhXn4X2c8dNTEiKI6bIIV8pt5Btzxw5qoieAEXvze1hPMpM0VYQsoIqmo/URMrwt/Y8MHDvparFb1qSoHWxs+oaf9fLitwixuNWOLpWExwqtoed4tGhYQa6mSX1Bh4OBHinhNhuEZgcO9cULpNYufVRGJ9lR038iH+COYwvfb22h6Lv4YLjB056GoM8b96Q9A9CQQT5Ghcs2tDeF8mzVpYuH9VT7NpjfWsOZG5f1RS3X6G8jSrFzjMT9v7F7V36JsQCBvmxuHOIze0tFGWxFOhOr5wgdp1YBdlUeJ+DeK77CDAT9PfG4CZuTSv25sGiU94jaRCI86DHx2bKswgzvh52LMcOHQ5Oyb28XlSJBsMPhOsF1IFc+r4B03bvDtr1LNLtin9tYZWYsbyngiblOhu/iycPfnS5KDlS1zRuEofgyHv8ytS/vcoMFLECh60r9G1LfaoZsqzo3gTJv42HR9Lza/xIndCJ7q+nMEDTPuuqUFeH1b19W6WUOM4gBm0/1UnkF3ehFJBIWNLpunapBYtGfxem4C7a+yac69/Tc20lIR5PH0t97FI2mw/sJaRJa3W4RiC5Jl49uQFzvbpOTXawC8aJ48VNJKoXYMof5H8ajqXGYxwHyBc9fbHx/G12ipPhimUG9/sV8o4CqqChaxgYkwFt2LHsUFFHRKygmv4mxYopcEQo0M4AMvRbyivZjEqhVmb6whP1W4xI7VfjER8pOqeHz50GewW778CYZqG7N7plT/uT1r3IziIlIv5CWj9w2+hNvUh41IUnDbyZaGtb9d98ca0pqSUQpUNzg33vNxO+Pb+/kvIfnqFJ5ROJgVvm1XhTUPFCaSrZGK8/ai5lEms3gS2Xo9vxorWf5/sOlP542aGue+9w0GN8pZ/NpT0bFpBoe7e7ih2n07OaKRwzMJPtszxfoVFgvQyuYWVEkP4SE0W9vpD2DQdZM+8B+EY+S7OMxQakvVuqF04sQ6jPpAuXGgDveQdz0qhj5UAhHZyswtKHQ+veqDVZahIQXUPiPpN2OG9LU8aSAcOm2Y58P58ita0Wu1iaWy8d8HXZuowz088m3f3X2Udg/0tVvVdYHmsHnubrMb9zyB6a7/Ln/uupoy4QrdEuoaPbirRlLhNgvFnz8bQHiqkVJfreiNUM04N1bU4r6aTeP72ggsNPYhzllU9fEAKjSZvHxHJi8Vvfa1mATZEPph3AMUGouBusO9hRr20FE/qzgxgdT/rgqp2r9v+7S4uL5mpYQ3YDazs7RPP9/dE+L03XD7eOozeW7NC0waAzaHulBnwsDb20WMgjrXDapUqvm5Gsh/40ji/f8+/+0eTY2oqFuI7cgDtYUjGZZUIox+DyIi9QJ/eOIG3Lat37a9raV7mht13Bj+oMP9jbpxXl94M8J8cWLv530XVfus/VR7Amdh1WzytnHZviFgQ8VNbDGN1fqxF/9K1Um8ZoRubpPW8RFn8fG0YeLfurtWdL3ESm5cO9zwUa6z+HXeePGtLyvkVo0UBZxQFCoijVT0416ct2DCwlDdf5qDhtgIVLWdapgsb1QNho5/XDfexvpQrScKrF0wIxWksMHtbyB54kxWQYfZreHnE9NCWX4QPdKnKoqXZRRvsoXdedLtLYDothwsHpFgsIRBHirUDh8M2qe/mUts/1l8o3uqU3XRCqxq36GnUoqM6hUWlUsAaYZXhlAEu9PfSnMLcGfR5ZQ7hgftZjkMSYPh/AWYg1n3VIrgOXfU4KmJWPmfQjSr6sRlCcCze8aTJSI7AsCNNmsoMsEoAXFBPNLhnuV9CqC+GtjToREkHGlASCw5HNHMXHs7z+K8GxHi3/BBZeOQX2oVlOW1UhXPbW7AU9ncAr7CP/OuXLvbkIqH6SR3yRgU0MafP4ekYf0xfwPs/fVESa/P7Q9yuNxSEQ53X15kTOpfKScmEtsf8zbJyu+79exzVnGKoeLOqvi1lFE0uF7U7bnWD4aMDA1B+e1NGcrd6wYs5ngW/2/k29/7DwRYd4+2tuZwouIuoqnK0MJ4szqff6+UNZaEqBHcEyULqQDsc0LtGAb1XJzUjfE8khnKBYJI577zBjpcOrOs6l2c/P5mCscXi9zPb44ZTrOwwaMXyLF5gGe57xIXgk5+9Im3MB1/0ahLdKsdkhTJIoIGV9g0/Xuquhhz0M8TAaoORwK3PcKK3X07IGhrBRuyjcYKF40EMZpxY8a8VJ4A2aCJYH4a7fyN9hbrQX6bGNPmNLtNKhqw6HN/nRE9/l2UL54oNw75VfVC9oRiUYYq7kFsghHRETzep5IxeA26qp4kg05Kxw3kQI1H0nuf/xsmsaTur5r2zYGjC39D8VRjbXGvT9U+5sPLloYaEDd/T9TZvHPf/YYqWNVIBrUFhEqhCAnFuxaKAJgnhj8uHklluxsthQs0WPvnhgjikhfmnnVG3l+Bm9L79xaAHchPZfOj1sWtUwO7o5SAvZRtQmRvC16fQtAVOsX9bLB8aLfplBteY7LVteiFw5XcqD6xA3Gb6NiUX9fb8jU8T2WHydwSSmAqy3bHwqHI2hZjRTFGgE3fspOSxaBykVnlBTmRgsE+IfoynnX669tT0evIA8d4GUhm8ljpG3lyh909Z1mYLRxQV41CRxZqRoRuhZL05y99SQ58mACDyeuYWKEC0bmO49NHaLiY++wYxZVUr03d91Sg71NukCLStYrv3/fXh+oFhSMLsFDQIUDsFI/xzKyQ50tGvi+Q0EgkmH7IIBAENE5pkNxISV+o5Et4LeLoqUaDqPeexc8bAVSxssXx+hJfg/GtWDxO0MiWG2owKqqFvI+KU2BnXriCi9jtzJDheymrG/L5y/yX5FfJjVN1UHaan1Y7uszidW1Yg3Gt4h+0Ub/2VFQ4XgBQFwlPaW69qiqKuZk4LHkwISIuCI7UpwdwjHy6G8SmsBnBncSIYA3VKQjh6JsbazOnodw4lj2OQM8u4AiGOe8iGAIpVDShI8zEmUEjKRgd0s/ldgYPWAhy4UZqyLRxuxYZGWtIz9Sb6XZPXHv7+ItLcgx/T7Q1PDymbPH57Y0IgpEhyj1JCxQMzH3LwGdFTQkJvAUi6GlJqwd3tw5VGNyrpQPHSTAjKjLZ19MM5ouC/FwNRG7ax5eU8KhU5FawIoulRvZo57wgr8MzuPW1DqWTPBdc0sByoy5xrb8a3D6dcbjXbyUBqja3wt297uJC3/g4UYcLZ9L8TC1DtXEQH6aWnm9e1KKuGpdEbaEf0jr+FMMSyo2vfvDKBw8+LZLPk0lu2CV23R8zgvtGDQfpU5FW9AsWVXFrNicxRr8Q0X1t/EIP3bDoIXgo4SQtLCAyd6N3CjaD27NldZzrxnhxCqxd91QblacipyDy3wBK4S6eaveP6ZJNol5ghe7YF0ULPWajIN9hu7KqjndAdtTKQXZTsqEFOd+xkB2EKjj4Dk5/p8ptUuXuc+95A5qfJE58Wq2uz8CxHMrWr0aehhTJ4Pc5S0t1ivOs1/mjknk4lGSPVAPttMWt3vokNxqAyA3kE2fxwQ8cv7GnUn/xa45+VN+mPe8rntzdqhR3doEQGji7aSiJaUZCMpnsVMXqIDUsOUB0DZBFBxjBnEeX3igHboN2260zTR8cri5G6Nmk419Vtd89mRseXS3Xm4YBC/N0KxVn9ucBrGNd4FYCWW1zgwPYOqlC+RsBSHNR8Lmhi6ra4f6CN402VYQkWg3koXx8zz/Ttb+NRXSE16imM+7cV0zx6G7RUKXChKwYkj/zGTiEC0JAmiRHJnPrKQlNcKe/k1KUMYqfe3NizwaK5tzYl2drhUUcNbZuDm+0wPI8JmSqjCcPUz0uCSk8uMM3Pv6UXHlABNIgt7872cGIEZY1pPnrpsPtlRu8qeK6Qf7i8XhtqQ5P3E1pFLL3/XHakaL8fE92CjrDov41Ed5ObnErN28Z+Dx7aW4l4jYj4i8Oh7mKNOurwVMX6gjG9ctBwoalxIeRnw96zUUHR8+NRgJ8AuLEuGFGcUqMf/QE7q7gClxTPEgTCUF7upTwKxFH3Wfe39tFyU0OF90LT4XcqF6Y6bh/L087ixssgQEiRtJQJpnKJBSZ/2bRIepCVoCx9RpsRchjADonoHX86P3ADyMNx9HmjXPfnqq+VGra+/GXPM2oy3mazkhUOZfpdkqVEpbW56pML7xXD18zCdXgw6HkjORIfW1IwlwkBicVPWPa28FjoH4DqGqdmL1Pfn/X7LiW8fzzFldYr4xNwHbK3/nU9ieXe4U8iPdxgmWfrc6L3xQCWStGTM0eSiXBwmJqgIUpAqzTNVUvHO5ZQNGr4SMc6dmYv/19fjs/smqIbBXEFtW6T3xN8nJDBSOTIKuRYJspq9uDyqFzsUjyYA2N9ahO5rfJAf7GWMGGm1shvT4YdZTxU9U1JPr+8diqvux+VjVgMorrEtrboiqFMRvr9daNGso6QdWeSxmgGWJCkaArga2UvDae/+CXMmMUMg0Vjo0yBjrfxTceT3WNBV2bcgW3PW3k4FiYtUn0mJ0Ian6Za21tAC/CW+LcaO/fg9DAMWQAE9Icwue3IdPSYbN4pZpt1ut1d774Bnq4dIolx4eWvf/TA1MWn7KaFnpciuoss2RsEjRF5tDOug4JshYUBRG7hAep51/8iCASZvELdEY6z+IbLRxj5TaRVnSsRfxZ2hVchqLWjfo4Ud3mpciCAbIpbosBVvFSU04YQY4aN6nw5RPVUPcbP+vf8jTbyAr3WsdJR31PzjSxqdS7f0sNLIMFPhV9BY83ILY4mxrBrhoTYSN36JMNCMMIhfQ63DBmFusGpj++i19+wWOgrsYUM0Ls/Q8axR9S7t59HY5tbPTxGuwzTEbzHUcT9rIEyjlNTRN1K+K2t9JUCZVsKQZptqcavfkix+Lc+S4uKpNcxgmHe7xBfv5gCs+Ae2Yn/6XTt4lWxJkKrr67sBa1fUjxqKnQrGibEIRHx3DTw2Vdw5obvXuqMwytWSt6YKlo6qkW6sbDkJ0oS5E8Q+wrSI6S7/ECAkSG68AxOM9Ej80ZXGbIg6VqhUAVhociUvWARWRyM2Oj1Y33x8nx9zJWGua+KunlJ3VVCrf1imyATZWFYFiPK3TXdm8r68IWvant1e1DlRaeKTi9O/rGRTDq/yfuSnIcx5n1sQVQPoAXvoCZ6DVBGm9N2NDexm9foG7zFIyRsmRLSoCZhezOrh5Q1ZHBmL6hNWZEXLT6DmXUf37bU7FtAwLZqs2tkPNxzPBLFTxQ4XBOWExzOmydQp9dddsLztw3VrVVpMHDzqQk3tYaT1Xo+0Q16zvAqOPl77/fZwbtCoVimQz6mUc+H+f06YViKYZL1YoqyGEvMMjZjt/VqW9t4ZCt4f3FOiPtZ3FcG8IvoYM91a0khzfXpn/7ijjbvQqBKSYWr2AMldKPY3p3og4sqOpq9RZFd9L2nKfwYGgyzibGioDc2UFZRO1fU5BCG81PRvDcXwO8noUv/vM7ikCkYMjch4gEgerQRsqCO/2UYcnXPgSjB3GjDur0gy2TSCNU7DHHDVZY1+DiKbYTqCGt1Jtym07q9wf0jeHSXwJIW8Tf79HFCpnBIWLxmqjPJceAKdDTJZk0lJxRK+JJR4X72VDho/jZ+pgQMwcOyY27dbRufRc/KYTnjkax/nK++l/EgtnHAu9UGyZLW4pLJu0ybjjHT5VOf7IQcYo+0Ood3mDoa4uGAKpeNG78AZ5K2Bt433iCyCSq2v/8IjlQrOIsUUjlsZLONgq9cl47xDENQGwb5rch2OAGA5Xi2Gyb+qzdAy1xed54NPZQZpnJO83ioIe7GxXtVTqSbanPQgg4p6iChUtc18oxgCTA0LkkCO+YpBE6nfDCzNzttgaDVyN3hrf9AV/cGD7kMRg30Iq+vvtZb+isvFciUxT3pTOTj/GpivP9VBAtndJGuWQ1vzpDzRAMtBn3JpSAzR8mN17vhn8tZvETCxsCm2YoXpjx5v1uvrjnZaHBeOrEZwSOFkmWLqklmYGGEDjHsZ5wsP1T4NQIW3Oi4icfZG140nfq1Dg32JvmXnJjDMelTBv7iAEE1jEEDe6nyLgvJrGJm9H+Yu8rmfVSVTcYg8AYKmHmu7n3KWx+q2hn+ED3xdY9FUrwilb0PaOu4c4i7s0iXXHpqWpwmUiWvvi0h1RzlwyfTwa+0NktlXvjgH/5CBNwG0nwmLXhsbVv04mxhgewpumhqSrGTUsHjX9fEiOeRR9voiGiBEuSjJzLD6diFbVJgAwY/Eg5Z8PxLQJu5VPFPdWLR792d3Hc4D9ER/0w/hZvl2hYmP823v2icTcRJpMXU7IUP8jaslkGi1cIy9KGRApHMN2s237LqFDT9tp0QB31mfGvVU/1EMgImAyAzOQOurivwmF6KT686n4wLqxtNTdCqlzZjepXMI/UzoK9iOCRW9NrBt3Wom6Y2x8M430ACM8NFlX/9jVUrB1J0zgWbp01CJjg46K7qzMc/TeNYWLF8FKEodBWEG+2eq+6N1l9qscfeSi/WNfwecg96IxcdukoeAWzlc1tMk6vJDI8pTD5OWpfgTinMBGqEAtqLheu2908zcbpoOi2slFXQFXjHa4uqjpU7r5dfnYGhBiWtMWNqnGLuZEERrWgkhACY59V1DbUdJnAba3uR1bBnr9t1CtZwxfdN47HtjtcJWGOHcWQx2/Lm98jpeBNGWeUiI+KeeZQpHlSuBOWRiCpBBFJUBcT9uab9lAIEtmbH/34g63emTEAfSZiBhrjcM1qBHIDNiPXa9qjpM7pQLkRCVrIRu3C8JsBsrGwMKMSKkd2q/JsAJ5C6AurcJ0fP/IBBarelbsbzhulpRaI+hMO4yjB81/cA58ifh/hO8+IRydrMlHX8R9luJMcl6ZiLsRaCkYALChLaU8YHGYGxaMWUzihM+nf3DdYvO1+y6ClUNj7e0q4vWxQbqh5ojc2ln4OyWaMM8Ta1arpdJaGX3GO99TuMGlw2XxR7MUff6Cl8Dgyef/eldwAVPT1Z2d7Sxr10c58lg+Oi0M/M2c4PfoVocIkXnCdqIbQ8EfgA7e7o5J/IZfk6FmAVYJRUNETwngb7baXke4G4OeNfSn3SYkw0tPbokF2Gj5+uGa4ZA3ZUyewKfnSIp5DhUVw2wfBYOMx/vEwMGXgbo5NrfFU4oVZhPQgN4rd+/9tqhnkgMw3JgO9TURh4n2hX9CsCMkywqWp4mqBdUKVK+xNw83A0r/EAfMrS8Uo6VHnxpt2WxuMOs3i8E3RPUHZAsykrzs26eyjwc59QidjhYQ4p4/n2KYv8fU7kVgFEwICv1Hm4Gpgt+7L//T5nw8lCj09VGM7NbZVQyfnJljdNff7I/oGXpvuhb1/S6Bqv/2l8nJ3PbMam7gosqjL7OCnvpVJVyGVcCe9UYyWCkFv4L+ZwmXU6MfIYAnnRRWi2yr8TivtNrYXP4y/onxDqpnfNvhF0n+2Qkckrb2viAAAIABJREFUUhgZkfAJHCKG4HT/1sWtIyX60Fk0Aul3yj1j/SNFzxwcnfGNkh73Wc8bf8LfKCKTiG4rdQNEX3ZcmyIT/Fh7SinhrJcg/a2fyKGXNopBhWKAxXw+8QF3xr6y23VwDZoTXC8wKuDlBn3VswgbHqiKH+sy3sR/A7CfVDe6Q98D1tBLh/vflt6WzU2i9rZkp6EAkThL0xAaWajYroaD7Fw9WzjhMm0OS6CcAEnNLGGBh2roCvnxJSj1Y2vMyKPQDQlP9b8CbrtsAvD4Wt3Fik+J8RKJo89dNEiwM71ZmRgtF1EFCYY2tnHwc/WbRvVijAf2t+NnT4pIdN44Ttk0TeYNmf4O5anK6VKoZn5j+Y6METmr7y4Z7lJuLND6uIyzts6kaNjXSoWFw44lCG7iS+HO9MmB6IZcjHmK0AiLKbyONZumleanaCkg8PPnssHSOmrVMMKqRA9IVC28PlRv/nBBnBt4LZIq6JTQKrF6WI7Gb0pHyTGoGVkqCLiIic4IDxwNeX+Po/pvwJM5oP9GBE7sKsYAX189UQNYYicJwY8sLOPMwtCp8VJJDQaylQ1VKeGJD3s1W2mtQIKJgfnH8jj1hdLVZnmkYNwYq0bPOiOvGceHNpxYXhoWsTDAU8XrdQNIHWUjRX0KwVRno6XqP6F1hA5e7I5VyEWEI2kEDzX+P2xHhrgyfOc+0IABJbyQuXIJR3cfh78XPlMHvv0191B+nI58Fh9ASc+Bx8D664a3zHCCJSTVVRVXcB9nDkxyATdCR443U0zpc67mxjiLMvzKzXD1irDehfRdKMEpxaQv6DaKRxGZnIziLTixZRQn5GdfQNEpXX7OcXU86OZ6rrW4vXpR+yXXPnNfMk5kpe1JneDRS264Cr+2bS31Nn1nWYgE+DLnHs/iw/hQdbyneqeaNbv9URUfJw6QfQHe37Y9FWaF+CKbouH9vMOMM/QlNXd17N3XCclSisZk4AgrMSGM9nHl/z8Xnh6rRXmiymfuS4M7sHbbjGtvG8wI8cVLLHqYxeP6eYOWUyRYb7US5NqXvJ/ztAx0Y0q2k2KFI4wFPVOWoF9fxMO6FirIlS/kHgaMskUPVMAhSiU56s3I48U66o9muXGit4qghsDfv8TbWMR//Mr2VtzamcJUCdV/8G0giEjg/ZR16MO1uRPN1PD9ZQqf/84YAkgI+o+M5bHLDrpbjMVhfKXGL6By0iWW7Rf/IjeojI/pCrkBNTeu3U2pmcM5plq+gtXx/BvYUwiv5sbkallVMpYJndNn6rsqQpi+Uq4uGvAkuV4WVFAnO8yN/IRT7GHghfp7OBrxN0o8DpgbXQbIyJoVrmfELc8a4th+JuMlw0BetG6Q3S38YJEKS+2rFrcKSAjbJnEaublM5IB/kR0UcRoAx/iMdeNQkWKbY9Qf6hM75kYPd3HvV5Zxsihjt3bhvsrGUHrbqogTjE1eKF5MJaW8smmGW+poV+nrYAnnas3jHjxT2EmVSRxzY+jwnZJwTDHRTXa4ovpCNlo5XWFtuFaQ20JERBpdFTz9gpJnMLhbJiCrfIhSj4NcMnhruH7qnoSxp1EjX3jsCOWFwtwYuiH39/7+fPG5CeXC2mJGBL4DeCrQbisuWuO8seYW643LiZ/q7OCucF4qQbtbXk5xksieUPwBuhkpvDB/R5qlAmAOYAfVc1wgM7ICRjKkDXsvmqfq1N5jwNAwu3vOCeRw1/S4uA7BYKjQLUtHRpaciktGWNRT1Y+VeC5JUOQaPlcxvoLRIQiO3iNIsnwpCQGlZEyNUjUgNOPge89Dfh4G81A92s8bD7X7Az1cIJrFtHIQJ8Fhc4BlVIKXcMQFUbbEFz+rbqvkY2fO3mFCdA1LFSRUUwavfXtFI5TYSELwZ8mVJ5kMPF9VONrexXUUL4uq8dvlUhZVKx4qMmw3Zw1dUFk9kZmQ0GKKQyHu7Iy25a5Wa4b7er9wBidin61MX/YlKn0p6ZdO8yLTXD6M40Zm1ZfHY4ZP0yA3ToiowocKfiMoox5XLEPOdHw9+wqYwAuqBchtEJG8MAmFGlHL6FfZl6zdTgWt2+N0gb0UDOI9Hl67G75aPW0L4aWCPw2kwCrChsca+tmgpzqJHi4cvYacb/Dx82UU96ogeRbtKfXT8BwJH99iQjY/KB7pgj5VaJjh1LDBWUjh1+NS7TSDsJC+0/MeDBfQ3Pa4DenLJ1Zz+KmxYnTD0D/vr+FgdO3/IjdeD739BRw3PuGpvKjc+rNuRCqZvAUdz6RuP9Xh1Vlen8mNqTnctvNeqTxjLC4ATAhYIHrdrGdOjI5zo0wcmBsnnMTt5rCRh7JgDYcD5Ea6Xa/Rf7/4eTKckVmDaoZmRFz0akcvLBfE18QogfHhO5h7X1ixJjSwc0cjHs19PX0Js0a5Lt1KvaBlOq4RBziOw2rk8Je58WA4Fd/FARX9MTeYLMN1gxODp3C69vkZKjJPGjxuTIj5nfGXqcxkvi5DnD2VTwa+cufjJkom8r4Mf5IbzzEW3X2APZXoGj6a76lwNUK5UdaGt4LgSd+XIvRWeYxJEpl620XN9LfykaR6MCwksW6kVmy3lcjnOALU1OrYB0vc0kZ1lBfZPFTjT0JLBU/VIJqfzfdUxdMM14ZAbuq6NPa3H2s4GTCdWen2bH2YlKZvmDMLqVGZyxSvOHaW4eFvKk+4FJMpDzNjxeave17TMrowQJCwirtSxcdvwydgcYf78/XiJW773HigAiu+lAOc/vINLObSJ08z5ryiUk8ykx8P4LgSWZBLsBpHTjbnshTpjEPcBuFON0Gw9ZYTUE5KYzHPUi4oI+AtoL8eIDfy+B3Jrr1TdeIWmBELp4K6kRCkEFcJu4hWwv8Td23JbetIdNmoArQAf2gDoheAArkAlEn+y1Xkoob9blCUQ+fWMJ6ZOM79matOo1/n4Y4aXaMDVl5HDT+J58YPPAbn6HNgBRdP3TWqByLUOlByVQsQvlDwOHEgoNtd8Og0wzOlsziKRf0bTizowD5nWKn3w89Iw2KGWFa/G+aMejCVV/gUG5rsSa8iVCjNbY52zsjnWlpDIybbeNAjBbgd/HGsEAMKBn2juGxBAX424NsW3uGu98v5Gx8fL7mB4LYfeX/F3LE6k9kRNbAfdoW8taWho+EE8HGp5RvHvxkykmwKA5/2mBYQBUqlcahQO+rEP0NRX8wMRe4bl++pjL+Pe6q+jF/jG+02/WMzCWdxtuKGjf7YMo4BhQz3DK1SgrDJzN1E/rqfuPTFIypTgvUH3V7hSbJbRqLKLnUDHqqtij/nGaZxaHEF+Akk5X+Aw13l1gTcpgScpB9OsYU5mqaOLrc+Fco7uDCJYkXf81GjlRERu0rvtZTDeSa4JlQlQCGVCR78Ms3gKQWXGwN8o5yA0v6EfQmoSTxXjcdO1f4abpO4oawLuL2nfsQd7ufPgredu7wSYYNxbEYNKK+I9CguMzvlyOzk0EUbPZ87fjfr3RrlpUJYCC5weYNeAUwFLSPvQWhBpSUkhel5q1tucHIcWQX9/7GG37pSJ/zOVjfK1/tjUycytw+TupVNuh5eyxtJl958yQhUKKvboN9DbCxc4/mikZsfa9DDBa8HORUSrnWpk6L1iHx71gq5MSvRjEEK3xfnxioyk2GLB3Biy3g8bQgincs3rdJLp1W8U2TCG4qfmgV42I40VA65ppem+JsCDomQmLLE5DGeN/gPglRsjomLR9qehWWBHS4N4+Iy8I9uf2QwV8ELEyHqB0CFzjhMhaa+IpCd0jvLuGMvjayOWMazlIufUMb8jJF/QQrIjrHEG9pMjVRKmZYh9NkTT6COrmJQIVm2Sh5wL6I4XIzF9/e1dePDVO0TiPBAbnxBLMobuYRXuXouHKZU/wLYkQ1VL3xLoiIHO4N7Dl/rFZfPPldRSWMy3uFuMGgwEvVXfNmoQ7JKDi/Xss1bT67ibPd3bW7g5Hf3PrHAF+/fT38iyV34sCHA52Loqe6YpxH9wS+yC3U2E6agnj85hLNHDS00Qv2mQx8vb0c6LGmtzhwS2RrS18RP1RLm7bFaZxGL3h9iL9rh0rWJ7hthqxvd17Hsi6QFY9Ef3i+O1NG7N6YBsVfMrdcqNFVC2UXl1lbmdGpkx3HFLoAO4DJvBG2iqm6pqKWivcj2SG1hgdPf05S7149dV3WRzoiJhW25gQeOg/NGEY5f6cxuphDJzIxNugMqE00Y1FVF8QzovWZhUHOToBYnp/upbBM4P0rw+WcOALW0Phjw3yG5rzBBfmxN1dbjQm7MAlG/Ojdoh7symwa4ZuO4fZ6fW2Z8vgNRCdRThI6I2sc97jE2gUgzjLeNrTFZ1JzIYkZ9HmbLlaUyfq0K2hkfLup020jg0lYHDv7CdHlu43hd4DAuuYEAjmt5f3jfoAYXG44Ibu9vlA15h955ZOGDJ3HKivJGiVtcFHMfd2B06qle0FO/q+AOVRiDNLOIJqzVvVIcgNFqBhWOKUE4MBYSDOI2XT5v3EUqGqRHn9uDO/Sl/HCJLexQ3TBnDCUiZaNrRnAi6Mvutlf4sxCW2I86GzAt/2L4q0RWgq+BAJ05ETgkMDgkEIsmJZkykt+K4O8AhltvMHBgQMhh7h/MG3dVC4O/SmMcyEareyEx8cD3ePTOTFHAbOKpUQ4R0Dp0uAreM/Y5xvAfRJ+jMiqrKFbUIRi8NlMjG7mEUH4kPmtwTCbaHLa5gSujS/dUTG2i3CCMehj67vNdbnQPjxPpWwGq7p2ZRjY1VS925CSH1Twx21H8bBFncAjJh8D9KGNvS30un7xhBEwhWEKkJOkxhYmjspUNbHHpME6n2PvVufFtJgPw/3CgA0f/eBn/OoN5an8rWyrtpcrenyzLPcM26NpO5WzmcH4zlePJmhGbDXpMdneFxCBGgFaNyVbpu3Fj+9q+1eUGwVisbtxJFfVizMguN9BjoBk4Oscoe4g+W2/X1529SdcuC9vjkglXCJGJSfrxb7SeK+M6U5L3ilkyhEpwDZVM4H72m4IEBADqOG8sN4qHGnBc2lO53AD/jXHceqotHG+kh9VoFEl+gp16aGPbHcJuiR3QtFOGm7IJ4xdItva1kimcZ/BtmuDjhrVSEgD+RmGgtJi0gsy3OaxcOHBreK3OiC5x2WHgBv8qPFUfVgxRK4RC3tsZ/C2HKaqEpFicuFsfLQpVUvUkOGS3LITUyIDuHIkakHH0k5KB+xDop7S/5U8+BVyITAkTA7/meZ7Syit1goxcjDW8u2MTiL5sNbAHclNP6l9eZ0dWuEWsyh7iONOJ8vCuaIi8Dj1YZKyh3HxQr5BykY32+rvEqIpk48UIPlaDfva6CYmK3OGvacJaoTWjguBNXee64qqKMSP367GGd5nF14B1vB9Znqp72RbqM8UtrT8yHZmUZQa06aThdFVFEj3nHZPsN2psVTQqmIbBf6D/yMY+G76ts8Vw0PclQYObnsti0m1YNu4X91SaGwEX6miG8upMykAR58D7EPddxiW8oZT1skjHmhH8OxWyI+f/pDmVD7ZTTdFnhAgAdbL7uBWLsP+qmhd14pjgrWnLjXVepKW6X89tkvvGDXe4MUEV/+pJ9nm3L/TQKdoUsizYm1mDKfo090WLg8gl5CjIznxGTDUeBCtSP5Ury34hUgS3VpnlI2MS9LMNfFNKUi0mfquW7bFCzAjXDZa2uHzeMBMtPIsN8Bf9q3Ru4GBtVRH0pPsr3sQV4akXpm6fFmTgx4eNXkVugyJ2JCfyH5fo0W+v6IQkjS2CRWh9K0SZJGmR6+tFQ7op+GHScXwLxm1xamHXewyobS80VQvC24BQ8xC2WVGWPitBi3tAz+y+rjcDoANBTxk1DHPLYmCC2Il67DsDSGfnJkOzeVczbafclKG1fNLEqD4uWjxg4oAvmf8oHFfWDRJhYm7Tgr167MvXAQuze5i5pe0LRbWwHItIRnXSyH1jxCSiIb6R+tWuKhEyJwTh7UmVsI/fariliRYNDQc2uXVatmAwQ3lRSiy1VN+X9lSMUgDM47L9S46opdc/vtiWGuPQdSp6K/AQ1BJhedVDcdWow187g6sad7uR+rM4XnzRDaHjtyKpKol6NrEYD6s4tbWTvFXbb5eEayqF8Ly2VNdoKaAnKt/+nojg+ez1+NftrBwY8GlGo6W8ccViWMLuqKGuP9FkROIJRKEq4Wbv+sCdrBD62twYdp9/kFxw8YCfudGF3Jhn2FPNNv1dP298cBUHaQu8xA6FqZV+2ugcrcw0ucUo3P3iVfKop4oq8WL9reodHdHF3r9a+qwJoy9X3jpCA1JljWvI56Z2b/M3xAuQ6pAWUjXwrcK2almetzY3Lp/FV+twt2ZkgHAIY1wckenFMsplUfUKLhzlUHeYfDT8TUN1Xfw+Kv/W80f5MjWIZEiVo2ttLn0Sj4nO35ALGImBHyqqILC/pbJRn+4wvhM2vNZ/AxgDC9SNoYDMpGfwC14Hu1pja3SSGwccgcjyz2rh4IGFYkn9V+rbmdezkiIsx0YdrtUKINLYzlafqjSkLDcNeq64gMwTjH/pSS/Ven+hxF7C+/N88efWMMb+C6oBNFWdTuFUNUTNk38lMejyVu6WViEGEOECHtvnnwfA+Oe2NobWbgnrRdSd1K588xmDKwmdv7VaQFZQO0UdFj1bWzjWuTF8uHqHezdvmicoIuURq7iyAh5FCbDeWLFXzaMj6kwUSe7oLQNEdIqkRPYinqezozrd7SxgKaILpMxcgAy1QbopFhSZgs1+/Kt7rqCEy5eJt12bG9/mL46CSICK3obxT/7whbDx6B7uFK7XV1mKvOFcOuSzAUTMuC8cqHj+satKHm0rj1WGh2h4AU351nZKNuphVvicwAo+T1A5lvXmPJQv76mIhnmnKg51I47k3FRs2pCYOB6yFI3+iCHAox5d/UQLTA9MivLMJx3BG3EL5fXVoHfvRLspg7HVZisliw8+NNE/mxJXjO3JgmcrTSAzsjwX3qjf+fTnisc1eCqRUb/hzm0ErOGnLqYerCXStbYaakxdXtwUs0E9+72KJ8l/MWUj7EeHfEZSFWV1cCGCqDaSEpFNlO7P6cWqboVu89724Y/S1XJTBS9VnfC1WkUPd/0He6pvcdFCzkDA3ACB4lKUiMziR6WI+nOngmCHG5HMoeCliLpiOSEwq935HLSQHrQUvdgR3V5rzbXaElHWgw6NkGqzmBrlnRomODrBL9BRTVtizPNtgeQ4Xhte4qEsqUFa0RXFouWpYrG8zlcNdNYovQtGOfJ7FQBV77kaZoyVvVbOacIMNrSsNFyDFO+QkupTDELp44N3sN2gjnz4u4HGcJxAIEOgyZ01N+4CxL22p/pWWUPscOGTK2NfBG/IIXFlozT6qoe4BDOc8TwNFqy342v+Wf3cUM/ZuDKRNUMia9Lzkdtm72QXvhbgiVEYYe6bKCx1GuDRgroBVWPLja3DXaRurEiJvRpPpVQzeKjSsyJmBKh/TPF7FMWJiEEWYdmMPfPq3sfMvqwPlWzTndzOGWmdvTo9i3aOoklfGd+myka6Emm7KVvXyv9GfKL0ugFLwwl3uHNraH0tJ9ZSA1a4T5LS2+o4s49Jrv7B3li9vVWdc7Y8WozkRg0s6BMlxOOd6vCZs5+QNIIKpwaZtaPidBLQyIKUinZrO3DZhpeKs4PLBv5ua6nmaXWP1Z5Pc8W88XG/62okkJIe1Q1xG2VamfKYOhv9unLg+qrlOzd6CbylCqSYx5/vmexwBtYMPSdgIfFlskDaICg1knBk9ceMCq+T/QfHjUE73ImjsmBuUAnXJe7V3CbajMksPlNujAOSAgSaQHnSO22X9wrpjBLJ9F69SIkIhew0xVJXuhwGle/EQTzKRAGrKepu/0fc1eQ4jvTKYwvIrAPUwhdo9wESkg6QD5JnXcZn36Av85RkBEnJcpV6ozZmqn+AWYwTmSSD8dM5KWTOYauEHrcdz6QnoochF0WesaVy3Gr31LtxYRzKudomhi9SMD4Jva0Vjl6t8jCI979WYaO6bNpNCseKKXizmUgj2dKvwFXkqAAZxzdh3lDDBF0vVXI6XfvN88hauQM1RAY9/HactYTr4TxyAw6f4Z06/27cPbVXEg+6Dlejp8UqDoOq8N6Eltc9t3qM3qYNH4wckuj/VX4KXtqPNDFbvIIQrM5gKGjDeRAJfy83Y5KTwMbVgBA9BRIU2p8fradqRcN7qi2Ke5Z+g+7EInJoIO5vRQcZ3W67Pjf2vLqr53VbwDFnaEz7oAcx0JiN41465iGCnrhiCldXndJNchPI0vHtEu+DjhsjJu6MIWNUkFAaq6mihM98uJa6UR+PWMLvn2fn/cnuD0RcncUbiGsCS27+aEHs4/cbvUbhQ2UbjcHFlsacKgerBjZ9xfTH2fynSrfSV0Yfl0zizoTZDxcAyEiepxmfatX81upGAHEvz/MzBpQagd1fG8aXsjHCLhoLP4WohoCJXPsAp2+ICSzibpjHKYNOCek1EPGISKNhaBR/b4ApQaUgyM9GXGtXQTFbg6QUQK8yh+PPs96Pm/S41fQbAqirP+3ZPFybxSHgGKZ+4CJDdxvtTH4p53a9XtqcCXjoIT2gcwi9ELVNe6GWB+zxjGPLnBl7n6ItQp1qOBH50kd5qia/GxjA8SPr/IfCYaP4fSNSPsXz00DDr8f/lv+nIanWDDQFZFuGJF4uNt4o/FIJsMhgkx+Is6Z/PSb202T2UpOp+pQNktoCaeoCqVB3f6jj0keNfKhG/brtUcocwFnM24u1jBvttWqskXYcFxH+Xc7tqZS+9YTS7KP7WgpluxuDqcmoLQvCsr7fd8pLpIqkENrO7aumYBlz8+cVU9nILC2YLBm0njtYsel5jAGVmlgvRn2mlnlPz2HCEKi3wtht8gFIZXfjfjLXUI1GzNmwOVuMy1fKwIDg0LZaaziprd+sNdQsL2YpuuMRVBrlGzP6nSk8dxy4CRfi64adjl6NdoEMQp8UOJ8MvMVXji5Kh47l2x/9akgFn+Mi9rJOUD5FE2tcQ4Fwvzr17uYwHpw9+z5CIv2umsncwNY0Ty7DHQnhS3XQ2sUohQXvVbG7oeEZ42rdN1IoUzFrs59iEWdTqwVEq4hU8QbkPt1M4d9w1J2k/uim9tTgZvS/+l/XF6X+m9C4ZBLYtM59NaJIOSoNWLvrsGCMavZs2j0jPTe3ihrF3y1eVbZKIzd+7TCWm5AVFKm6ZuIKtr1Rc7scs7xU8lTdjVB17iweOOqC4QoRVw8kVI7e7Suu/b7vkTm74K0agsuO8XbMku2o/ph2IRXQVKVsiUENVsN5TLNrAbLeh2kmnxB4LZ8n//WmdSP2VACpztyLf7qa5usp++ZlFhfOiPHZVD3ex3zqYSc9YFgHW7px/Z4z22HqTjV8KtOcXvaujoYEbohXcJYKVGmp3hn7jNjnZmtz2+zHnoq29uvCcRafijxcuRxJxr/fV6r8BFAfDDCEjcWr1Q6yd+VtGlY+O4lNVOqOE0WcwaDTHoPApw75ruAjjOqpU6NJhXzz+BnohJgyvK0lWMXPTWHD2y3ejdO5hmZs2MlevOn3rVJcwX4OMWWvgkthsplSP1aMrrg+4KVqp2PhS27ArXVDkjPU7HlnqTQ5iF4xZtRJJ2/dbhBADMfQ/uqBnsq4hltHpFPmjfsnUns/VIeZss7ifdAi96SJUBx+3TP0HIYUy/fKgCol398dBUVIfq50qyipCfPbFShKK8xdULtmE7g2CoJ+9Vj34YuXKSPcjSnM4143bjf3wz29pzIUtyEjZXkJ2rwxxLsRcuM0RODVwCJ4tNnFsJ2fJbYjB5nK+/RjzaiM6EsdpTMl540DmGvGdNizPfhYbfy2yzLHTirU8kdrqOaw+2vLv3Xg+zl+uDyM51I2lvFvbJnWQoymraePfterByO/mKTTL89ddoBNpUQs/S8U4SXolvEuYQ+uTkcc9ALZdhaDvIIzmMn3nFalIvOfad5+4vgnXf89Cs3O0f1dIjIiHSWsO2PAJXd+/atuxpPKEnsqqspIhO7SKo06/cWWqdJSp0NQIjvaLnhG+meUI2mXInH0Ft8EckUIGPrl4Nhxk5nj8XDvNnHRPplraLDhh4qblsMQIi7MRtaDXz/sXAuzq8cFCatwgrdBL/PXJUMPYoRKoFb41W+klR20AJOTnbV8t1dpnPfbqFUh53P11CZXy/jZdUOhqjuyMBv3Xg5jWAedDIZSvaY5FG9wm0g/YFQIVAwutzSTPHAqGVlkiDZJasBW7G6AHZKDDmCEUGk2zi1ICJvLgW1TPIqHzuI+cIgFz/3UWVwv4yoPZZn+WlH+PfRrMXIg3m5vB4y5U6B6ejiWxgaknyHC7VnEP6Tqo18FR6Ts8NFHl8u0ERyknfr6NO1/0FI9MG48z/fgEbcwz6bJDTUciVSt0sKBUe1I/EpkUq0obe4d6TPgcUu21r4mrd/MJctyNA0mFHCqrKidrWRLzZ4IhyhaXrV4OBaycyjL5Zj1ocK88dzEb5w2iyO36UO8FKZlGBfzbj8OhiMzo7rfM62XuY/EW88adQCXKdXlyCUpOSkcknksCJfB2J3XdnhyE9xTRztd3etldFgzOqkq/+5eDdYNzOJ4rM6dxRH4AEC9lgZUDf36pVII3aJf+83Up72tByqm+FC9+IClH8dwRsTJrRD+4Kg8kc6G8FU/NaOhqjpLTMSmamWtxsXYPwbBDB+zMqMfrlC+nO8zcl/LMFtTNQq79ooIID0I6GZ2KYYJHlRDwQq2GCiiOo0ATh1sb3OgFbK/7YLs2PqpLs5/YzUF2VyJnvuI8V3NmIka3h4PzBun6zfuLuCQs2jyiFHKtAln3PPo+jYWWYEqr97ud9S5Qj8dTlEs7kPf0RsSf8HK7dXbzmLMlIODbquzRrVBT89CNDQvPe/DjmM5C7OZPDlfnJpYvxvzs8XVAAAgAElEQVR1TGnqm9wMTe5g12N4dT0CKQEa/TUrHWghZEzpr4KQjR2S+R9lzWswaKrGPBNjrumoZx4JitiSD+2j3/y+cqxwqnN7qgv04k/YvnTdl0rN9G4ohUdhdJVrbPkJybZ+yVkibj/cdSVK9A+KmDJF+pN6qgpJChNIZR8Vxj5vbp1rO+pP8Nfe1Ys1LCJF/AacSjLmPi+nZ5rpwCFZmB8yZ7VpfBj4VA1vuGzBMqGQQeXnUEw2Q+5t+gtMpNrVyDwkVIoUvSOzEc/1ELRijPzd7BD6rJvY8fsDaQNHux56GM+z74Y0cFg2CU71UTvdVSiijv6WRJGXpXiKWSdBrcHj8LTXQ4lYUUxgZSOBfU7JUrdjcRRIz+NM8nk4jkOfm/RU7SxuVsbPx3A/A6IuL/Wo5E/2UqYPH/aJt4nrPriChWhLivSP24HB4TbTharCigrbjOqIrc8bs9JElLkTtkyV2z6s/r4bxYXchvFP78b9eXbGQDv8C8zbvkTctPTyrVUFXYeBJ0Nsqfp1XjhsC1flG9s+Z+m8b6bKhiOSXYCs6JTZCnta4kSAhHTarFQc8dZxKiFB3HAO39YPWXA8b7bf0DzrU3m4F8/tXV6qm/Cp2jRu0e0DPYiHN8Sd4vFYMpN7/g88JI8sXKPkDFaFzSGhtqi+DNSWSKGWbs18DXxblIsq0/YuOeTHRyv0VPdXAcdJ+eJSOb7oai9N1W9O4wP1fa/drW01urVrQgnSVxbv8ra/TVs9eLYON3tGWbbk6XYkM2oHd3q6+gYI0kQbk8DonM2lxfrxJNqGQ0HDh3IN/0Gm2f2T0aRaOLIYhAwmzb8apW1PcTkUJLivBX4pxdSGsve1v8UKKw+jwt2lwYRVOlvE9HWzxjesXDsbUmjbJT5TsxT145+HQ+oXDUO5nFo3GkylRFydN6qsmyZs/65cMlEa3u8R2rjRYPir+9V3a1lZ+s5B0k2rNH9Jx/ClRpSQKTqR1dbVYPdcQTlH+a7zppUaj7VUD1lxmAGrVo5TvdvE9sfuxodMfyAdXPu4gH2FqCw+YBhMPDPoxGdr8BKjHN4RptZktkzr4U6pOtlGb/FkC+wc66dmc6mf5YqMWK/aED7PRwZAagaej11p0ym7P4/RkrJR5ama6JFnSv1r/4azg/oNgQCgqq6EtPB0lI9gHrf0O8Iva5dCKC9ddswRfKSSaY4t7U+f/0KL24a/h9kabgD1k+9Gy3wQ4d8I/0ixT3gTHFfij0Szna64rMyDTr4RwaZuS19A9Jgql5KmzlQTCti2D2bobdIYKUEGKiVXRehSE/ur/PYo/qxb3DkIOE7PprnL3WiDZ7saqhjQYfzKROQr9367kksECCRPRqZcw2fA7x1Vg7dqNQVT1j63aDmvXY6ZSw7cVvq3UPo61sq74SfwZrGxXIo/L2CVo4bi3fb5L3Aquxvdo41/ZdSrwYCswe3rd9T6QT2zH0yWur+CDYVcm2DTGY23mWtSpcf1Eq4QLhHbyb/++kOx+PMCHM6B3oYcrXNxqotnmkG70s4C8x8lNHsXgzDIELiFzmmznKz3bmApFvjC4SLB6RZZr2PMNMlbxyl9nVQFPmG8qHWuB4a9//BS2WP1sOnv/+Sdupyd92fz39PN25avcURPNZgP1X6QH8H0l9XGPtU2vftTCuI+scBDTBnbqYRY8LqxnIo+YNQt5S25sx4r5DfyqbhtUpvJs/UbcFKH5yfqxjTYtehJFtnINUylMXhUOBhUGgKUTB1+1MVCfbhxR8YOQqawDF97f2mbK63UBPx2dk0AhsD83UP1Z/tchW3TRdPez+yp7qCMQKPcNYJb4sRB6u0bhGpF2nFUBGkargjfjUUOhPVozdbpHdBW17fe0cVFHURYMuZgoypLjLqmnf9QNOSZ+hP3G7ML+O/Pf8E1/OTd+PqQWXwiMsIKfu23CRsgtOEwMPwZOpUYHpDeq109lUZvQ6nJyQnaPZWOZlS1W3W2LrGsZosuIj+uNSqeqOm7t+k/jhqbu/FsdQPmbfdTMdwLJMroqcRoBIWjD6Hhr4z0gkI+pFXCJXrbWDfSEQuqAl6IGHHnxFisqvD5/xN3NTly8zj02AKkPkAWPkEuIHzlAwiwW+s2pvsGQt1l2iIfSalctldKkOTLYmbThEzy8f0E4+VJ0YnGbJgZnfMqBJFbcEiRqsjr+FyhGZCFw1LUx72NjXfx/+2uaL79HL3SPb3JyIqWXsjyVzatP3XZwRrSmKNDOuOST4voyUJnxxZWPcGyNSF82NAtZhbLnK7fmVeObEhVghqacJqhnJH9SQLD3ft43CGPhZfxv6+hiuIlqX4JmorcwOddB/dvNWaUuBsgEpD6JCKK0KdqDsrdAfvZqCmTEhLS9SSlq182mweTDS2IOxTD3YhPNTGGuxdjAaR+KPHrUEP1H5Z0DYjDL2kJvsep6gcpOjV5bszQrSn9zoqS7AyxSAiilrkuhH6jcktUZ1Y07ImHvo2piXv/oJ1sjiIm+4uoalMPz0XAV4pMwZwJdPDt7u1PayEXDWY/B30xi4rClT1lakJ3JSqN3jSuRlt+F1lKQk/jkxX84qVQs0n/BYb7I7G9CX3j73//HckDRBke2RFM87E8cjR4pvLvTdF92+ATCDuMoYfZWuQtrYfnsoonW6qHvtmiIOlFI/OmZwhqWHJ7cDIg7ugsTJqqVKLszDL+UPrt642pc5JsX4aXKdbfIe2whQhbeUpumbHIs4aqsvwlOKiyMmAxMHq4RKZ+e3juagTvNk2n2Ybq/tjZAo/DVSs93zmB9Y5gZqry/SmcX8edDCDfHWBTPfiZ9ErX5JmgCAshhDsqNeMY3mplrp5GkVZRYarfv8pqoCrjhzvcS4EkyvhSfdVd3EUpAp7Gf82uwbkB/YDLvqpo3/HUoc03ScikKkv8/wq2ayhgOAcYFRIuwm9jxU5+i49AXaNkwIbZPppPkD/lvrGNnKm4iXMXdzTh7pkPrOE4jAGKD8kQIHoCBWvI0U8Xuni+88G3gl09I37sznrszA23EJboc9DbhphWGFn+DQQ959xs5J/cyCu/bdvrMf6+IZyRD6K37Z+q2RoXHuJU8WGCNchqNcIZPUZ3L1RD/LgT5ltvA6r3KWvpcNtVeZ70PGYcl+ZbKDq/hEx/+ImUgmV8/RY9zb/ww92MeZur258Hh+dw7Yt6XYqWmyCRvFEpt9dXcYENU9Awy2C9dVwjDKCJCslwki2jJ9izFbxoqwBQVTq06pOVf59yGB//NriL175R7TwoUfRxbAwWIbxUdQAXgRFcDQE6nag6/GrHCFMSfzBpGKmJdt1dwAI88YT8bLaMsyU8rw1OWAvCTVxOThBibnRwGpwxME2Thpp90TJugaq/mhxuSmG4hbaJk5Qpvmd7vkhqGPog/FzehlBuUwimKjOVZJaIV/Hk7m6u6d2eUbD31YtfzhivBBhR5d/GX6rBHPVq+mpmqt8f2UKKmscxpU2auL1rqMxPQap4y+nWCP2CQFVIC4gES63OyJBXjelruFLLGVB4QBPh4ap2jaLgSJUMfP68IiNjMga2iZVmNFPtRiP+KIgXZDY5MD2Mj2S0vaJ5GfENVgguQ4Cf5+/w5J0FqHbAigwk+UM1L6tomBY2rsDN6Rygwj2Df+qFP09rwf0PbbyxRBr9NqoKkzPmMFNFzrF8HJITov521MZh78J4+vtmEY988oJTsQaC+6gYi5H1gVy4cM+YtXP3RUjnBJHCI1X7NtamHN8cozW1Av5B+RvTJuKmr/qJsQ5Uf1+NPT0ENM3eZy+sDSv6TWyAprSLI1h0Bp8K3WnDGKu2/rYrvlzvKpHxOFZMtvR52n9nLB7axb9VMD64b0x8iCXJAGgKMR5aSbJyBhQF91BvbnoV/oWWcGaXkPhxxGCc0hO39mgcXWo5ZkyzHLeUAKGHqwZe+pNGUQx3/2TRtlGUNLLXYxvPNSSq4Z8fKsb+ofqgmSq+E4b3AbBCLVSrHe8uu7gJnKlFmGuWBoid+jiE7slCssQeYAEMkbCsx+ZGh8Atr321HDnTeCvD1cvbIGBkG6ptqnyqCRguQFw8jP+ONLDGJv3hQS2M0ZvogOivuIUeQTM82waDWQVLSmhUGvAthMXtKjzPN3ufLHfF7OGMotc1gxq6tnG5N03T8H1j4uUPbgr1RzjHh39D1/GGJtKomaj7N2GJd0ARsZ+q8nCD3HYRyKpBXit+u6qvqsLo81nnxicqG5CqEIJby0NfqkrF/fmGDPMf+OFyF//a+VTCi8ZQ9WKVHtVJ0hjeyvLnhC8ST17H/j+SYNEYRPxKF8AE+FYNC/e+sVRUKgGgsrNUeEsRwc3V7OG/Hylu21SJvNp9Y98ALYY7NGNgU9cXXJtol3jnB63laN0L29PSaZBfbPDCPWG+7nqVix6TMQFzVr/EHiIc8AoFUzoDbYtpGVwT+zakGDlj8ajKP+SadQHjA/YNwUbqiOuqYjweNvEoBCpR+THlUyMVjXDj7AOlcXH8VxK1ZYI8nwUbrrVTZWVAMioACGfSJXRbtA6Zm0fBPxu6IeWhTMP1G4wa7ruOJD7EN+WQbUOjLp3AIjwZH1JB3vqjcwJThUN8isnoXvlxIBdcw7CCYCIhnCsCMoideZVvFcZZ85Wi+1/OLWqIWLNpGt03QFMAa4SU392Hin09IwfPREks43OflCJebBliCMaSAKoKh8xQ+17EzsVQp+juPTNjR/MBLkQa2U64DBnWX1KLUixP4RsOVVN/3hh1Fyf3NjK2oMTK44cRTZafdHAEOjgcwzuXtuPzq5yVgg+q9KuhTFwSvmMYOD0ArKV/zOmOQkPqQO+EB928ohAF/82aw/FjcpQHvo2NkJE/tP2JeZtt4z2/sGV7IuHEt87DFxZUtGt4AgbhaeS66ZbbBltXrGLmSWIyaRap9eHuoFo+8mGyKitufRkEhVoLnbgQicIz1Vg+1WaRkSoY5y+Vb2eqaIhtDZRuUXR/p4mLd0VfNnob3l5e7Ti1JvBDON6n/USFt0NVMeznlcuBHr5XJQuiWxpCFeox+PYHDBeCcSLxxKPdj0TJdg+HNtybYMWrkco7tUuoWah8TXJKu7WiPv7vzDZseAzh0jsEJaCnQWBtfQ9PgkZyxiupf7/KMLc/g/eNKlEmI/WPD+wbsR+qoiUnRHv1MwlANzOq2WIHYIh32AJ7UCQYTxfEYM3Jvo07B6YsaGF9G0xLkCdRsHdIl0EX/+lZCkMw3PZtfFDfeIFG1GNHMh2cKMvY8pblZTe3jZ12iyjLYMKQjYrJxvcRsW2FMBx2Iu+ETNwWZJgqmHUJMCSkMKOR17eR1Wrk55D6OUAvrgPul1xiY4wHXyqCqTBRQQLbDFGXz0Ku4gm2LrRyOJVqKH3KiVADyeyJPNC5Cssd6XEReV8ptFmgIOZXxpJO3hbUOHrm57CcWLANndAUGuDQR143qHMQf4cvsBI6GtWH+IK1o3RbT3te5BSB3hgdIxUsVrkU9DBOC5ItR6SgJIUeCT8L+zh0Ia9eep+ybwx9G9bVvsKGPFT1Gwe5tHmNVXTil+e8vfLFvhb+nesR/ez9HjZKcksaqexMtSp3io99mikaTk0SDEdEC1J/CybyzOaXAu42u2l0/sbGKO5vF6+J1mbfiH0TZ68dOnE8UA+4Qd+kF3pMt8mEMvE6/pIZ4KSDM8FwNi4i4YZqxghf9XMFdEo+VtLIkRWr+8Z4DHczQbGViNsXgxGRJurEWcvb28OUM3x0iijb3dGBSR1lMS2SMarOnn1KXDjoG1nYzyCw2WmW592mdVA9iKX+PdFQNf6+UYcq2v4EN+x3cZgQi7WnGLvoMn5CS+heh0y3nEPNn6n0apWAq98KHH1dkRAQjrOXXrnP+Gjp8ldMC5cvlYBV3/DSq6afYzMGfsuxqUJZgapu/aPtm1wsVMrkkeeAH7Q/N7OQRR2OqqyhCfJauoMfh1hi52BnkRP0XKk5db6lV8B37/ZtSCFQj9wwcRGjPPRtTKwYF3HTByvGTTEYJ+E2DoICPNIjGxAbrDBefqViCngmKXlnDuDGgyqtgt8uCFG0nmzroSV3fvE6ylQdOSx1m8Yz2y8VUapwFx+r3yCkCqb2eBsuRk2cacRl5uJHAU3RkERsFeI5NwHRZHUFZF148JogAG1AsBb1RN7RlnFjCc/CtS04Lq1P+TRRLcxcxbpY9ragWgy+bxCGOxkibrQXDpuvYY6wvIOb9NcbOUAxiq1qYHN0h7zXJhNL/dGlEL/vY05601hOIPSiqld8tGgJlyu4lqA8URKVN8H1k90thnJGTIrWD7HbnGW3AUV/NLnhrQTW30apZPt2dAeXegAOmYOMUFSKGZ7DJn33ligg2x4CHVOtg7buWobfv8w6XrmfKEb7sxrhh0tX8Ym3P5aMk/A4NtaF3MANNUHVlo0ntD8l3ybrOrwbGIpZQjA5fiY4QOJFVS0TLmtSmB0CSIT+8GbBv57N12rNIm+i6Kath9RHzFTVSW/6aQyqogFGuAKiDxcFDUecNMb11xNuEj6bWypZJHb8KWctC0XcJ1y2cK6aySp7leatRyf6k83bQBOX6x8NVYIaDsWpNnodex//krt4BdVfr7B1mHKPhu4JaOrW4hdpDZ8rehuJD7LD6TrX6gWWrxpMTeAqaBnCe2pI1gMTa2UwROWsV436JgqVQ6Yqc+OQoWro29gmycKkHv6Bt+EVtWUTQ6+aJkkd9ZbveetlJGvsYhwT6nTFLXw1fkcyVvUBTIclUUYI3kaW1t3OUXbAfTLxkJER3MVJUDGWh0vJdibUTN6GV1zEWudFXcP9q9fOudlqsISESmwzIDrdAtekjkdwA1NV3yU8VfhJZO4bqxLYCCahqtTeXd/F71+1QLKjUEH65W9Q3h/IbQSMEE6lfEPPTJEo7Fs13MEDuQlRgUPFOUBkPWyOGrzzObXJS8TphD/6xR0cqLls4qTTKADPbQfPsvZp4yD05BukkX/Bw62qv6lt4yoaiOpdaAph81/9bbSQ8sHpqkGcQi8MqsZNRDlUZOuSmiCTU++KrLoMQxLhglgshJ/H87k/jvLklcO+DZEMjOaMqAfPF4i43lzHPWxedPfjduFfGDv+cttAgIBn9VLUYjhrPMzFAEDFZJH5rUde0fN3EVOX3JTCoCH0OvZ/1m+V8j+/JQ7ln3DU/ygRlzyRquLbY8bls1+MdrzlzBnPcbw34fRIzsMxUD419Jc+tfF9TgJfZbZN2jFuHDVo12OPhCKfJ61F0bXvyfV4CjHa1OPnz2icCrv4VMVNcvvj7YIBXPlQWTXy/4m7mtzGeSV4bAFkDuCF70HAPgAfrOE6BuIbELrLi9hV3U1FlpRvoQSTzGAQzCKcJrur60fdRMKBZipx38fFX3BpTLnLxKKpCxARMAvX89oXyoBqIr5CdT7lGYuLasKwMU3tOIz/+TXS2mKRTXoGhtuUhi+GmrW7an4XmMYrAK4Tztz8sJF+tWyKMCBO2Vyocpc32qzZ4Ds8ClGkxb1mCi3fm7IVo0E7qJCbvnFyuG2rCbRUUicCjiiI+2pI1Z9w1J8X9rc6/A2EQG50rfepZcbd0Slj50ACNWVSF3kYPHgel57cHC8iqVNRfdkOpmGpPpxkBOumRjRR7emWd1z+gNsNs/j/KN+3p+Oc3CZ9NxBNynVTcJmjgpU4sqfoZw5dUr0LFQyQgqezBTuI7ADDjI6q8UTylm6meIG+9LR4v2v/ZHD0m/BwTBNKw9eG8hRk/Dt9L/66umUTQFypDdjWt+UGNhoMVlwlGm75HjXxa3vEszqI6BjYgVSjmqv6V+O+3dq6ZJNqSqaqn+6WQi/VTmL+UzsUTOP07m5T8enzBqbx3tgQi1dhpN9uAHUdE/o9Sye9N1GILjmcXsR5gRQKHIJoLKoE0E/txwVUjXCo3WGwQiZOGu1U5BgqHo+CrIF/9oxfulSzM3Z/7UONRlqO1iAXle41wk07KqRc4sSUnLD7fkvYjPyempwJr8Z9WJizqYQpjyZ7dUfxeGuv41if1fMReFdxzzexp2rHMS27qi8Gvr8uz1N1f3Qnvopzt41/st1oZSG14dVMQSGq8MOpYjvHr4k1UgNF4jAs+Do+E9mZszHh5C2ZzSHmICaMyn8mPjUKECWobftN6mMSlArVUnp62zLv/aS9OCgjn436OajxiwJVCbURkoVk2dY17CMiykGXxFGQb/Fa3Af3jDvPo/uYuQpnCO/jSC9l2JScRVmpjVYU7c2Y8KViVHTGLwu7sNN4uO1AmrXF54c7jMSwEyZkKeEzUDsT1qNGf1Jvc9dXRXiDiUa5W4c7MD2OOR/6+Y/m3Fm49sPYV3kSOBdpa9veTx7w0s5ickCVsRRO98N9esGADuOObhjA9DQwXYVl7wSXae1shLojczgPJN/dczEYEjLKOlxrY8wbHVVVUVkdna6y/aqe+Fz4irfbCkWB6hDQ6sdi/PoXvobgjJBPFUgaCUaEhk5Afes98TbtuYlkc60X/lTOnub5w2A1It7S5S/Fd/u+ni1S2eya5FVJOnIIpcg5FPRS0lRNtuFoVur/FMM9WdskiQ9XpqHYZjxJwgaOY9DBDwhV2t32Bf1y1zyHGbtNHec2+Pgf5CCbc8LyIPJ6cEPhIrw6IlvpRo2qTzjAW3RUrThsM/5FF55m+3KxAfAMnApJQeRTNTeF9iA45o4CuBT6cd74mXEZ3r3hAttmhJUl102NvpmKEeHUwGw5g2/Ct07vXTv3EEdIIEyIXd8kHa67q9zEofr9y/PP9Bum/BsGdbxNJBXaViMFo9SmrXsq2LiR9bseRoheSJmiJra3W+qhXoXR2VisXFN1dOtWRQrnR8PQKbmveBClPdvu4ZgmPPQ0t4Bk4HX5C21Tg0Y+vpsqtLgtfZfAYZ8k4Bx3jjFFeBIRHp7Qhw9u6+dn8TbrPbILCx93wzRcoEbxY7j7mGwVXgquKU5/RQ8Ethd/VhuMQ3ldXVMlKFSgVH+RBtQlyK1URm/ymcIQPUtE5nC3epU4ZFFZWppD80kfO6O8+8rSr3RWCUUXTYvzmEjzLDZ9T8XKg9/QTvULRFy4jFxP1cRe1S5Mxg0RjA+3m3p/MgTWKTbUNy+97Wj77FddfktrpRGXme4VcMqLmgA0CkUdB5E3hj2j7VAn47paJevoekmGDndL4aOwNnyQ1tVxeM7hGhIZ6WpjSCCmp+BAkRQcVBX229tFYczrJRq8iG7/+0gGH8BLpFAKRNP74haOPoKIQHN05zvFwwB0a9BUmRYH4u4pQCMkG56u+3sqpI4FR5J0LA1zZ2A1eNCD2FkcAUUGxrbP73Xb+oXhDlcRzBp5kVLWTIdHmoHdj3o/g29rAyBun8mOhBB6nXxDBSCX07jFYUqH8/ybngokBdBG4MPq3T1NzrSfIBC6oGQnuZTiiNRdoqNqLtB3TW2/ozbitomIY93WWszCovRvhhI8MXdP0/TjqgLjsFBrBubn5exZ3GubYN4msBSI6rymLOtygKnIrm0CkzTkOlJsHSbpGiHgIvyw7gNLxHm8rHe3LtISklcSC/WisjU4GitFp/r6mCY3/r0s8f1c3V9jcEEvLlxDaM2ExTP0/S1ciPco6Y4RCmRQKAmSFocAATf/IQo5IwrZJGRxY8VUug1Thc+tU/ZVGzam+vOn//MN1zCtr+s/BUbOm8WVFa0JHBw4GFpGVpvIZ0xXlsLRFayomHAi0es1eltuNUyXnir3LtBxffnKVwPrPuca6W4qLF3LhJV4fXMgZVEbrz+ojS7t/YNjAvg6VMPy2Rhug+1hd9dMmacg9ITctOHCL4xqg0RTF7Wgyj0/RLxF3quQRZQEBbLuX4u7p3g/VXc9Leuk3WM0fnEX1cnapgs56p+aozUoLTqtWHSnI8HIfRBmFtd0sfiEdyEe8XHID28jKdcVv+T39KlqdFssAKtTaHSTRuFhaItb10tDmly4dz+XOsxTuIYmNWNP1UZo1c6oRiBxv5SUX7iHUQXrpIaOPGXBZaPcUw/tb1sEUNwnTRVv3tlqg/tYTyqccE/VOu29HdzEerLh9Vyc6smb6lOCtPTZ4I7JuOlWG2FHVha8PR5oIkniLbkO57sxCwQy438eMCy8R0sReGzZJVQyoVkbHYo+UfKqF1VdvaUIjlQ5ENIU5n2Thw1P8cN9ojo+xSzsw2jqCz/JBLv6lIZD/oVZEnnniI3gpZYPVxtIAoI2gBZtB3iE6okOdnktdL/1G6bvg8EIzr62vq8NRUZGACOvjsBzOaGnuqgI02sGhqRukjeleg78PPRwWAJsjLaLzYMjQg90dnkoaefezPKcJvm+7qxTFqsNOEW6fUZVvJZMdBzGu/Noz7jSFNp5XDte9BmckUtjqgAaGZzTiA8RCN2RhLCrD0/kpWcXHG7BcZpRPT/ieL9hlDCfyLjw2MkrRvUMCKdHfRndJTURDFEmepmmzcLAugmooXp3nz2LPxn3/tFwKrMoXqAiwg8JwfVUab+rymAloMVVyQbnjKH1U4+Ro0bONFeNeXv2w8WkGFV1qtcmK8N+SXZ8qJHNw+BV5QQ13TN+gq/h5aqGuEYaQUEkjH6yfeJNxTEj7HhJRrVNEDBEfHaWwTOWyDTq9jXqCjy/tQErY2f5bK1VdcCtQoXFNq5lqzaKQ9RF3HS2tul5fToM91PMFPqrShzaABiGIRxZ+iEKOYmkTKaOqComb7pDH+is6z4Gn7yTWpYfoHqtvRicOLreTqVfhK89G/Ppff/r/3TBsQiZO03b9PxBqBpUH+7lGmSJ7Govk3l7ZllztPtp0LEjqGveA0a3NE0wAnTeWsKKIZuI+0oZu3WfW4QX6GW2j0Jfce2pvq6v8zHchlNdmDA397iY/wLNRrp1OIxvD2UAdWHItFudabj5bmQRUwi0qUP6W+z81vMUrZmF9VSFy47jeeKSws+Ob5IAACAASURBVCZcG9ydI6mYN1QUe/p+g+bEvXpfPJGG0Hvu+B83xsC0pRHI0WwkwWnTZTijR+X1vgMEadIZPBaPPQ0TfW5h5llXhK9VyQi7lYGBgzeVjOJ/sN9AGsr8bMyIumw4WgYjPaGp1beoxYMhWbLiyMk5e7Y6CY5SGNuIYYkO5su9oUNWlkiFtYs6cRu1UIZwoaLvoiIKVDki7kviF8/DqVCL9orb7i/oQ76iYgp780ammaRgIYlrJU+fGnzWCSi36KviY1PHVDVQA/Fw6o2uiEihemlnAF9bN6nty/X0DGU6VNFMgT3VwNAT77dK2/StR1yzOCJCkeXRyOoimXTtp7LwR0OqzK5wwzahA9ML3EQ65HZ01BBloR84kgZq+bQgxNNcT+XhNmPDqwiUP/RhSJY7GhYmkseEl5lMaKEoxGh4IR1FsgS40zeSkX75raoMli66bTW7IxA9VR1Ajb4S2Y7VhhSHOOKezsOlauClmgF1DEMlcLXBxKwVVnp4i1K1MVyTw+mVboM4txpyEI/RTD03p3AX/IoYE+OGkOY5UfKKlcaBy2ouqW759/2/9C80sTZufOj/f2Pg9prLFHZXfsl4VI2QHnKKtPZUmltT6mfB0LNVxs5BEJ+q5BZaIzV2VHSeyHT41TAQlxfVVch/J/KpbC/+qaA6x7+brTV06XToI5KvE4UfwtpwJm3OZrXxEsBQ0Hkvr2/Di/N3KYSnKPjjRcWNBixdDh/GJE2VeVs8T66NzvPzUy2RsPxOttrwbs9hDyxM0DHJskmWrjEuZZcCF1LYx8TwSPvCDR4bJvFCokKHoTvpzOGqsHkD74bYTEpP9Tx1Fm/6jSuDm4ZPLjhonpDs0UjDMZKhzuIpamUMD+/x4hNHR9oQ9+lYed26UF6KSk76EqGiZ0UVCiEx3GPnUZSn8GJu76l68Uuj/qp326cCI8DPO+vCQxAVn3gz3Pk+jHsj3H7/3d0Wf2PMYvDSOql7NrHGnlee4SPqZVFA/Jyc7fBva2NuAJw/saj3z503vk//pUGx5t7WyiOpWRuhKp/q99ZM4Xv0u1NFM7NDmjzcW0IzFhm4iIr1s7IK79tckTqaOh8HUqyvZVdbp9+fR4ep8yE/jYcrfmGKGlIyILOfRQGFhZAv7ZsnRJjmQbKR1XnYwl9HpoY/VFO2wdqB+JUBDsrA7cMC2nGUokPG8VF8cRav5/k8XOmoyIsmMiJB4fqUY+lnoQLb8BS4U83zdu6pAg26Y2+coJmKLeJyNJXfVqwG4JDR+ez0tB3WBqCRX5UHzuNLjmOBjZyUoWx+uNrgcuAbboFq5PALR2iUhiRtxAajZ0+9jc7IgjxDvabidlpZKa421EgEaw1uNf5DR6XzhtTGv4V84zw/XIlfbCguGVWUCEDCBKXfEScL3W64vQZIbUb3HKPydoCmP+wNz1twYbF3w7jo5kNFxHD6D6/GSm34ieOEvfhTvaJn9f7w0c3cvq/FZ9rDqaSTClHtiKPZWKhCYIaovudwccmbwxxsIR7f9LfVKWCZ7FPVTERWfkpMWOjBf91TMSvocu68AVD9BT/cD1xVdKHS+jgi2HB5fvDzlFVfsNqQ6hiZm9xuqcc+PWS053u0ZPZ+3ydRDq42yvQLwLC7q5T7efWyv/8TdyVJjttK9NiMIHgALXSFXnOhA+CHCK7BCOoCHTDvYiBnSKLmD3d0l6uqvXGlEzm9oQ2eamGqWRSLcT7v6W/NiBdloXvXseEJpATewZH12gkMmrWPaD1lJsDdrop0PVUf3chHyjXcKLq8FYxtNkBc9hc/tsVT8bEpspgCHSkMpm00lws2otlFGDp14fXIFmfgrdKS2T2APuJJ/LGVw/Wbdd3cVkC292uG0mJFL6wWGmngTbOIWlglbNhZUZGxti57ooAEP3/PGkgkQeVBzkJUjybDnZlE3VMJTQ84ArPgC2vmq7JfiTTzQTyEFruyXNixKbdJCByCUuBwqEobg2/7l5rcUUR3UJdbjn7q6ucYXzjJ0MGeiu6uJZOKThnPcFnhVrCERKoV80fJAZAqNrUmw7+meyorMqnh6JnfRLBbfpyebqkoI9DyhPjJ3tVXJjOMs/zU6Wpt628Gv2RrB8uyXd00CCfyUUOlDa4oVGE42s0biKZi3h8I4lquWc8KbboNee7v4Bg7BX68uKI6K0HcxGRCnMjpzFKrzj9xVkwKSjeTn3gGbJIbHyWGzH96F2+9wxX/DVELi/QqqcVi11kB+xdW6SpErLlxhfVkNWKR2jHNlN8hbRCkzVaNpO4m2OEmFTtKHyQHodsubNyra9wmOiOLQj+HKBxlCYYscStVyf7RSzVKQLzTK9O5kuYWZ0UZyp+t0lXKU+4c+lSJ3hGTkT+rG/MmuxG9NrXl/akZioIUGJjApOSRVan6l4aN0Z9cL8ho4+RutI9UitgwNfbnDMU/M5vs2k9jph3hPH/6TlFuBCI3XdprRaNP8IWYfxAMQX+S/FFfnZEe1nEnWtC90eIW+zgpG2ci+uUOSu1gvZjA3t/eGnXueaqcf1T/mafw+cNw0PwXDCm2YW4shrx/AUw0c80Y5qlH8hdRVE7qOB04UNzTOGUJEhoh6aen1z4FJwgKfbIbEfUNoHh8WsdN3WDQSOPcEH9x1ERST1fsanvpqOjV6p+tcJ3qezoSsqhSg2QLgSrgyVLDofmr2xNnmwUmMlVucYaILCfwL1JD6sYKBhz5d+OeSla4lwstRthlTtUk+1dP4kTVZ8NqAql7w9eQOZw2hsJoeq5DzDC2OyqSwH5NjID+OBY0cIjln+XStNBRZzdMFFMoO1zpqfgGSwfAV9aFWjYwG0aZ/MyoYQhNKEQl7JmH44Y2utzdbgYEndi4b0tfhUM9GJm/0Vi7TSHq6jCOV9drtn7/StnocB8y0gKXcOk683WsYVHrc1NUznv38IREjbsOo7ozFKBI+kluVEaxDXG44r8h6E9dGYqD2TMvdwOkwuOGbkVGofsJw49XhpAOL1TyyWhX6M2PpRJSssKqXyTHLPY0R8DhttxTEX/jyg5FhnFepr962+Bt4Ymn8M64IyMHUwSp3JkAuLjEPd8fxJNBtdF5QzWPrNIOTxqfB0IeqnW63BFEarKnUlh0JBxuFO1UXKbr8PcS/BZvsCfX2dzA35OxOznjU3VW9qvbO/npETbJ3laeq00VC78tGuapYguOxrc/o8Bq1lT2oRrlUP6ClIVHIiyayXVn31cDOK9uPZMC6LXyTxV3yE3RODnwKXwSZv78VW/LyaFiClDEW97+pKNCCdYLD3+GN1Bd/x5L543gO0NAhRN5njj1PXEkSaXOTOZderQUEW9Fc2Ca6t3t/G0crgn8ijVsWDcQFX2TG4hle0qasZA2o4FUQLeeqfr5yRKLZHKr9gYIbdYi++1tUonu+Wr6E0Ew8aj+LjdS3VM1xuGSGyYR/zq5/pEPhzigPO2nHJfx3gs9WZTSXSVBrKhbCy70uwho1UqvvMLZ65V9kHUVu32+ptKeit0wG84bBOBZiL8hiKr+2l3jBe0EIogTpE1yYzRiq5VT1hlncOenO7tCY9xec2fs0Y9d4zaypf76qWIa5kWsghrqqC9HXo0cmdtkAW6dYtSfX/xMWPKH85X9TGcgCmKUxZKSDyfwJEKS1lJReAFqMPPF1FehRiqH8WNbbtMiDOWI4x/fYsfROvI+N6EZXd+J9Q/1umeeNU5KMCPAJwsZXpm57wJ32CvuBrgzy/i3zV82uH/12kQiPI354stBRD+pjA8VcUZz46FZFgGphMAMF3FhXI5eXYAc663yunBn1JhlxGCjkxtemWyo0meo273xD6Cf5Nx0ODS9bywVnqoTViyemkbr1L77WI1C8iPCjOtGEc27ssgCbdVJZnA6wbp7Ii/JIkYMr0yN/GhLtaX0bf2Wp4r8ULhuLC3rBjVVcOBYAKbQMWOc9oXaUT0b/zzlC0yALAs2GoVuxFGdWevWi1iC21tUJYlIIm74JAZ+hGJjKc85/SI5YN4IbKN8bMyJPfIvxIyY/hbCoWT98SkQmtfobK54XcLZpowaKrCa2fXWqDpc0oGm3NA11cYiknP6RQXX3GAhPUyNlhh1rBuwNgTMSDQkZc6Ney3VeKtgbx1hvfe3hGS+gJ+Uv4ELw/ODAp6YWWZIlwYnktjJYd62X+XG9N/sqRZSwzUszKH2huuf79DJnMlj2RhRMK+rPP1ExOIMNfwGeXtTyWeGP4uIYRImk1DDlamffpIXcollN5T2nNhFBXGRMhCVbNbR2e9VM4EO1D3d2HvNjevMwIWIqB/5yvFyD56Aeykayc1JPKlYwvyTlgpm8SKJdIH7Rus91UEsxo1tr9na9tUr1D8C33poapEP23cEojrVLosncZHz0kWd9yvGzOJsNIsXbRfepIvy8DbP2/az3DBMs3oz8n/PDVjhCoGD+PucHGTp93zwQ4LGyRw4nCqtQlflqzUVJsbJoyLY+c68MV97wAr9tT6/isjtPM8/6W8NSP1yA1Jvwm06io2WWj4IdeZlQpMTNapeJw9hCBBKhBODRgzZkdwn+qn/K6Nw6zmcPZnmeftRKP6IrP3lfwg1XJp6DECZovlviPFS7Q3r5ml8tjV0IGDPXrCqfiRLEUezHwVGq4XbyQ6DLJxFPEEXVawpsn1InrkTjsR38fWGMdCipyJMNK3UO9GZFAr/vo24BshIivQinlA3uJNwNlTmxU+P/QRmkb0V6xOLZzOG4duveiqLGcHcaMttwngg2tDoTHbPzcPv3MRxAOyuGDQFwYNEJqdjhuTGLooqWWNevcKisgibAG0qWzj/ICYWo4650ZbbRG4oR5Vv05mjf2ohXuPa2NrB6rR50b9l0VWv6gnWH3l3m65ThzFwNxNgSuk3ZYOCoaL2EIpmeKojzn8HFkUaUIQndm/9cqbRRf2KevSzswY/Veo4yrcmd2f8s1jcpG68eBPnhSHv0X+QGn8U3mbuTW1zA/3+juAUay4cbwTDG+G8ruNkYAFDWzkwJMyjmZ5iRUiHiiZxbanw5ZqTgaf/Ij2IvI+5UT1TbfBUtDg82J36IOS/t/LDiccG5YZDW4HpelOld3GGHPoH+9vqBCgyYXbkYKfXH+BGZvS0vlA8Wt/+jpwciyBx33yobNnwRBBnYRGwq+7sRVyaqyslqse2J4TErSxgLXVj29IvZo4/s4h+0q3p2BiHu6Cs4UGdm4au2o+8GRYWJaaaIWptaumngGi6xOZ/BNShfXCJTXTlmMwUKJa8iXnJX9YOQYwQmqp6qhp5mh144hiiium9XcqdwnDVSQ7M3NfO30BG1in4/NN3+U+gWDxkX8pVtrLkndR8dN5+sLGqCMq8NGx43zhanAJ5cHS7z1Up8BFI5WWFMlAceJ0CPdVK9mXe5yCsgx9CCc+K3VTwJQZuDUTcyJ9AGOSdWh8dOogtbuj7m7HlTelL1AhjFFaxClqWY3MtBetpfVHqX7wNBTW/A3/As20seTH4/BlEwg0+dMHFroQDJr9AeeHDRB7VOT1W/KoEBUNCkVkfg0dkR5JUHox3VSl9eXP6A0XcaEw29m1aiDOec2NhEA+yOOo+VwsJdsB1XRlwTZj/wsUcjIFMBSAc0Q/5Q04KCEj+TihpEui24VYo5BiFsJcggsSdOTfmmrvPjBryIf3uoVJn0kNzrWi8NjEtFl2tI/+QORKxoycqmrQYOgkMN8W5o41lFo/B5Ucqh2NdS6rkojEw3S8HIZSft4MHC94FqeKQIOHh+EGwZa7kiSELJE+1fbM/RBlwxiiAMG17PdyD0GkGbKo4PzAQsX6sUN0NMwc/hhKjnAz5xYo5N8rX2OmuoMZT2quQE2JFWao1v2KYFKvP/+WenHIDBoN+6uFefhgDs2RXuXR6YmOB9Cl45A/dmkJQd/HGubGQ7gsX8oLiof/5pXJEJKBpswXpg1+Wj65EaY0+xvJY5XeqW4cQoIDnT/ya3ynID4iIL91UgDKeEweECwJ1VQEBAvn707SbIokVFVAc+kadihS7P/k1IbRtqrxQWu5wBVDF1ybSKMZnKPLaKkpmRPyKXjMJS4S/zsEo/++Xap6zojxVg4NkgLqx5oDlgp8/CyUrpphreHAT/ehXKuT66zZBkuoZkl/TZC0wafrAufz9cp62SXIjiHTbsXXdsHQzCEeH8eAfMmQDfyn4NxwRMRwuf3ft1gi/SzhizLlR/oTcbeXSsUL1KI9TTo+An8NyLidJ/liCU+IBibHW6XJ/GiSqk8oiTYJbMJv2D/pbuG6EC1lhLoe2dUM8H5jCj9lBjxIkyTDIzx09ryFJMFfyh1Ai5DiH8leQGzkPcj8FWRHK7JcTo3wSS3sLP/MyfMB/vKfE4L4q2Gcq7HI6VEt9ri9QySorvNVQAWCEHiri0ixte6oj1w5c41IhHxAgTf0SN02RHq9BWiqs6x7+xse1RKkUi6FkRsmP/EituZrkH35psHKSTLFU7jKDlDIeAvpHlw+QG6uEY4U/OKnfyRAWvJAt+8xyxXL7eCscfytRQ8mNQ/PcAKdg9Rmg5SF2VQgioRoOMwhBriI/ZzCUY8bktMiDd3nAQqkaISdCfqHyQ5WzIJd1D7/LKM6P0YoBgA8YA/xm0GCsDztettQi86YJFe21oM+vlw9KDe5v8Z2qmqpGs7gYm/FyhMDRVDtwqJCNCf0F98CwZsytE2RMTpL886eqUT4bSm7klFnzq5ULS06PCA+XD7ggCRgAyg1elMCfVT6H2O2KLExWv3uarhwHckD+vtLcbjOrLSnp71hNf432VEf4w+fYQST1Ij1M3QVD0VGGYEzwMwpb+fccfOnhu+WZWgco5iUzMAgBPjrIjVxEXHnT4AtMB5g5wioPF0Yq/MOdVdhZmsyTQn1oHrwygPiXvWtJchtHoqoblH2D0qIu0H2E7rnB9AFq0VfwRIx2WgwjjCv0PRwh1soLZQQwB3C4fZchMt/LBClKVeUZU+oJwrLE4gei8uHlDyDwXBH544WcCMDQro0ea73LSFTLrDHQecjhiUNOxnoMTCzVTpj4bAGOVHocKz+UFQMXBgQG4/GhvlWroeGHvldQdirxfw7i3Vm40feuuTzgyBlKKmO3bvZnF+Z4jmWEJtOBmhWppuHLtzlQvrTLCxzaCT+X96nE1lFOXPK9+ENnNnvYB3OxFJ2IAm2IqBsU5rZy/SiVE/qy2E91V65Ka7D3lRtHi7Ay1JMJ26yF4XJo4Mk9jfrU3ZrmTNpBcM++kK9NlfRso0xU9n9QOVFLfbHucJt9B0vEaj/sFbhhOXWb+pOjcZUgH5Al+QAfq+zIDMOnqCUBDmSO2nItlSn6yDPQzKW6WsqLAZ0hTNwNDnDOxhQvdVaJQzE69PCnMuxHD3rk84ner81qgM+gSjNi1Kz6AMgXo4S92TLvXy0Or82kXUNLFvapFJHEHic5YrQ6usk5NY9hsdsxHjGIPK1FVIotUp6xvx9Icewzy/DXQJysFmOQ90739Xa8kCi5HMCW9r1HgPgSJPBTOT/M1+fJU5s23uSb0+TLNywaeGiXFkc2XRbmBjwqe2rc1xlHT8fORW3iJxC7ducxjlVBHzF5TK9WQt8GW6K4DPwYFMHRRF9f1WRWLMCKAnqYUBiX9zQl2cxJ745XnvG0vsYcxpwP4+toGdlv0Wloy8HbnwckkAejoROFnWqqBbmBrCENh4JxRHeHtXhN8JoW0w2dB9GoEhzR9wpC1lG9veHUQ/Am/YENJdiCXdigt+96y9VXhoFlD3rOF3oLJ6uRc0KlA0aR+vStXNMUzBgqd27IZH25xWJxsQVYJFbvVXpUmVtPOcdLH036/scRkJQjkKn7MqLHrBfX/FtlRw1vd72+VQey2spsB40mhlcGTIZBFUxWB9ggKYzXs9NHAxQ95eKQrEaJqeryNZg14EMQrlX1hcEGRqfLFbgh/sAZnscMfpiQjQ6GgcJE7WTYlMFQqMAVo1wFP+gmk3Ktp6/6qRQIuKjcqatCcfnu3rmjVIFxByYHuF7GjB5eMJyx+TCxWabAF6ajAotRW2wBDDZknBlZyqcS9o4X8elfjuUfZj3QB2W6SiExVlgWxU6pcJhqq4joHBkZixYMf/Y6hrFiUq+qWCglzHlRB6bBwyijb6QKVZzD0eg44MC0fI+pnvuXnrONoQ/PDkXMvXPa07QwN5QeWPfdyr9c0keIHZrJJ8DXd4NmR6j0vfIh26m7KuAq+iMpUdsehW971FIAnr53Cw8qldyWohqr94wWcl2+a/j/nDHwg9F7fnERwUPovow5LZI6/deIxQWjqmxCXjhWhc1/Z1w5YmdFwDCBuEkQ2P/Mq6GU6gGNqTKYUdjVWbwp2mRpARC5UQrW3uuJ2wEs+VQZA9/Lw0iVP7MsVcZ/GiL5BbqwJmdG6caLWS+Zw0XEEbO4eUxOCcOQHEP63LULuhAyaLAd5L1zZK3jwH6z6SYqnGJgKDB49fljyWAIfTKtog+1VaiuCt1g+mCMXYwkfx7qyzOTMTzFsmFwnM1q6M+XydPJi3LDhyooPTQGLGWMSLi+cwfa3ZaUb4pxAg1P1VSQogROoAUOEzfVbBlo5RwMUj0GUIwxFrt7yNgzZX9g14nprH9bNwo9aFvDLPtMLz7BSDcJ/xbr+8PY+Go3akeHtGDU6UfIi4nsj2WmzO4kENOtvpwigz0ZGACn3nBx4/4R1j4bXWBgJtwxXEqwhbsP2RPDflImPcUWl0tX40YHbqRugEOn5S2pRWQEz/+o5BmYJgj1ferH6qt+fOpxTk+zn0fhZTjIRdMtkZyk8gIsPB1w4Eaa0dDpOj4Vk2MY52aW/Acg8Ep4VP4ffdNabI83o0t25nx0D43sgOfGEL8N9Gn02zQYgeMIQ2uVaZKlWq7vz7khmK1KJ3Uo0pqQ9MPRCPRz/bJP4eFYq+3D6GRyg4G9abZwv6h6zIHzkN8DPGBTOPeROxGqE2xFUrmKT5XCr4oZkvSe1KAVzkJpGsyf2EwTZfZjkerjfZZXYzAQ0wAiz7q02g1QmKnpHQzv2ZCpplrSboiFOzYHqKpO5YcpLDFYAg7VZH4siJPQwFM5JdQcuRJgfYuF6o03n7xB9+6wwReD2S8Bh7NjmoOB41zCnUJ/+IkNX85upOgHtOwh2GGkLRgzbRyxuawSSAPqFKa54I8U+suGbOJ/AK0AWFrMYk0pICQcCDHmyRxG6RzAg1Q/UqE5N+Anw+7nMP5AJiP2aZqTjBYkvQ43+M+QoAlJlipJXBI+uZzQHZKMKgVnqbkpsDcJ/1WWRjMeSACBSFJJEANuEherRV7pA/Tj3Qz0ixsgj3lyyc3ZNY1uyiHNNNnFfCq6VBGSw3rwmcCEYcKFDdvAsGgRzR+NVTcZR0KlGZaJ8rUDIIN4vOWkcAzcPEljtAwwOesDnI90+uzBKM3MSS18okmuyQ2OOrREsjQFwuwKnvKg9A0Pkyyok/jRCSmAY1RS7rClyIp1BAdQmH0SJ6RxTRq/AXDEUyf/hSvd6D2oBdiMqeVYkBs0Hql5wMaLJjMHODpLGJhgYV+gnhKRUyKZ6mI8aSCQaHQBaDgSwQE6QRIYJEO9My4WPmsiUIIgkinKwscX03egYrbMhtCMx+4szg1ha7A1J/DiqqUApdCQtEiBSvaMc23mFrpA+9bFdqSpQ42RzaTh5ChQV4WkLA6SkEsgIrWXGSHzwwtY6oeoNt8Q4IBn6FaQ+da6EDfS5PEahwTyhsBxs55hBAqYikGH1xffxPFiXnNgJ3AisXqzDQVAxycx65wjUlqVqSgBSlihwvmcUmwW0IRxEwOmcwES8kBJYjriWUAWtRsJqlLnrEJ3uSAASWlqUpJLFM2Jrb44SEo4oChEsEHTEVOYpeVcx/q5qDDqpUGvZgzWCLrM/ou7yh4J8WkIeIBgVHHjR2eNX9xNO2CvYzcmQ3MdDqGJwJqyMCuWTEmpa3SVngNZ2mELKhuyVetTXNqeu0ZNwUhJobeCfuGDJ0weWzrzG0rrJpAbmLEReQXTYHAs6HbTfei4MJCER3MlbqT4SJ4qgUlItCnw/BipsumG0unwlBTXkaBbItRbCV2MHRYLswH5xAVmyjY7YkEmQWVyi95yneasM+ww0D7cu8LIKdnwKPeuG8eBYFRUMRWxSJeuzg2Jvo5WVVHdCBGScP8MK9y/hPRgCTFozniUoPX4FV2oss6PAw51GDCLnZoeSEqCp8448/GSYOpSYNs588Czzi1Qo8hoazC1o9/4vBVfPt5oup88PkdPlLUcjrwCd5BMid/gEu98XZfE0YyJlwpr66je3EQYfaIRROsgeYyJ4fIVshaDu6XRWO55u6XDjmSuAyhuqyziR804Nle1G6MeD8PClFXCLBikCQDDAyBUXh10F+cF5zJtTBULoxoHLjXfFyRwVQcbFUa9c8UHO0VWAaYUqlG6xp1rTJW5Dm6T7FeNHki+NjcalZXiWYJIYY0H/HiUIo1Maf0ow+YBEZIB5kIAizrUUC7sDyZitEMYEQtjA6a4zUJ9tFiOPt0GbtEuJbKaZpEVizuVs7rqej5V+y7BElh06fwndBE1hh/iKVAJ/dP5rxTnXuqaBERQAwLqwn+wjD/UIq90PB2RwMVYQtPCm2jRTC2RoG/T2SBsSW6k0+E8Yy+PLJ6ePZPOSVGDnMb6vCKN6k9uoxqKQXQSlaaWuiQCQe+YXGpqE0m+A5eAEKnhadNY0kUpLcgNuRSfOzwRIY78QFj3jj1maQZbmfhunjaWpjfYkXBP25WapMaRa9QpjJTQxrEPVXx+ITcvI1rS+TAllcbMvR27cQ6fkfTnenFHx5LLOTmWMq8CRot4R91ykcRprFM5A6Hnwt33aHUilV9gJ93JuKlb4Mb8+MNGBPIqxF4aCnHaxTWP9un50o202qyaPAE4PIKm/0hSGB2n9Smlb4obci5BMC/Wt0DDxOh/zAAAAmdJREFURnwR2AYPmVw5GQPYMHD00EWaWoPU1EbenjfatxRvzMYejTq/wJn0FmBGQklngR+P4BgDMH+3cor/mPSjYPc1kFzVbrwyA3/uiMy3epkYne+kbZriJadWpUlNX2L3a1vOrXDjpduW220e303Yvw43bgOCF+4pXUTuO+7+xrjx1ytyebf8/3DjNkS9IENnuCGv1JvplTqlDVBH40Q8T+35aB9H0o0y1F17loyvZ29r28PazZ5zev3knNNzkUwcdQ9346rYacmUbtPZmrxHZtQt3A6Umcrx579tNp9HZf95tpzZ/aqD+9jY7/f+aUe4A8dn/+kFeLGSvV+598onNe79JC2f8c6rYmNv97if+V77dVEFNllve+f87r3f8OfpjYx3TYX308+/bTY/reVGyi8DGr+s5UbKr79vNn9fy3XKr9Mdvz1tNr+v5UbK04DG9unx8XH79PSk77bZbD+OjsbBp9hqLmyv2zbnNJXg1XzgTNvTfsQ5vteq2T61e/2c+HP8BdNqRt/kd3CyN77y8eS6yVnz185VNb7VRjiP2+12QOP99v1DfT1sh836pn9he/vwUE/gH9sH7I9L3rcXNtfhJK/Fz9TXQ3z4mXjZx7C3Ocf+fO9fGF/ir6b26cWTaprK4uqH5ou3cTP8YZM94x2TI+3rpKpWWiYWvReV7bvN5t1abqa8AY27BW7n7t39pe+7P3NT9+3Z9/ff9yvuePRu9h7u7evv3iSKu7vxrd+9u3Rzd5u1rGUta1nLWtaylrWsZS1rWcta1rKWtaxlLWtZy1rWspb/bBSMglEwCkbBKBjOAADUd4nSeo14OgAAAABJRU5ErkJggg=="></image><image xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAc0AAAEzCAMAAABkA6hGAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAC0UExURUdwTJ6cu52bvJ+ev52bu52bu////////9jS4f///5mcv7m+1ZGawf///zSoU////////5WdwP///////////////////1OR1FuR4v///12R4f7//////3GJ2jqV6P///1Ws7jaK7////1Oh5SaL4+xdLepCNXKJ2upCNTSoU/u8A/e8BjSnUqq0Jf///1Ws7nKJ2iem7h+P4SKa5xyD2hp61etENBdx0EKF9Pu7BcPf9oPC8OxpV/QAAAAudFJOUwBgTCGAc00ZCBAzNkA7/R/jETHCQYAoIH+ZX2CnxptIzr5w+N4wcKXLmHTQZOpr4Yf4AAAVd0lEQVR42uydC3fauBLHsQ2O8BGw8nqx1/dgoHnQtN3QhKS9Tb7/97qS/ECyHpZk6Dl7j/60CWBjW/ObGY2E3E4mXl5eXl5eXl5eXl6/SWA+nxvttwKuZ2A+CVZzb/Kr6WYRY2Wr4T2zeOF4jgXzyZXzUbwGtY7jbZpmcZxqQmu9qpmsnU9CCK7WwNO8qlZxvGJ/SzVvAADn04A6tj3N6yprIaYtsTnTxTXPb3oA2H2G3mUUn2kC33deo9OMs3MpRKKQdKILQOnOV1vcnwJCgWiN31pPAN66oK9aOtghyCfXeOc4BW3Ep/QQW0ACO8OZNp3Q7hk/xzTnmVlH7WXboaU83HhB+tA5jVXan2KciyzeLharhmaW4fcpTpbmGve+a7p3nZozuiG+qcmS06wxzmyR4tdZdxKviyrlC5uMvlwTKCk1N9iSTNx0dTVNyn8Vbzmaq5pjVyeRLhIjJS9TjLR2mjbT0p1SXdnldQGabd7dYj6NtddkB54miSlAyDA007r7nbeJe41fr+KbbEsOBno0Mz7He10n07avFvGq5Uzf4ml2FQ1Ds+4UsbZtFUw6SrCO57Qe5mkuOPBeF9OcFiptFeROM4vTNVVT24DtFtdL+PCrFYlaT/P3aNum2pQaf8tk2kGa2zbpdpkWANDN/hCMAFdPBKGn+buGKC01avy2CppwNG/qCO7TXFBGa1K51jEOutErKZMI60W8zboUXtP3NK/ac5Lhx7aGOseDiHQbb+c8TUAGnjcCzRu6d9aMUOIUf7KbZQAxpbWKG+IpjVe8v6d53a4zJdXLoh790ZkB+nxdD+9X2Zq+nWGa5C2Q1RAyMiLBA0y88zqjIYefb9fn2aC0/mDWbKQv0gzTvMlo5zzP/ATfdQQA+xw4f9Jb0svLy8vLy8vLy8vLy8vLy8vLy8vLy8vLy0slgCIidPkjI3pkBP49B76aLcDVLpk5RxIGs2mrWRBGFzobiCB/4ORSBoqS8HzgaRBc7MCIs0UQwOhSRo76RkbXQAmD2tAJ9UdspoA0Y7x5QIKvnj8wbk0AxzciIiRnIWwPDAPShHC83RGc1bZgD4xtcYF4EYw8u4iRe4YJZAeNCOFw1KkQtvhMCHKUBGOtAyA5cNI/MCE8G2cdhS1oQ0YeeDYVgzwitgiji7KcQaCk4d4GFCpjBSXE7GNYBgoLJKQ1zrbAlxUkQJELxvg2NTJS5oLgQjxRoDUrSFx5Aqh0ks5wyNXkuksiTpS4mlxnVmJ2N0cB4aAtwkvk22Q21HJCxcE4Eb5AMEgFOrnfkCfjXRyMQ0weDWcb5GILLUszDmaWGa5cEQZje+TQJHkAB+NERs3GxonsbREa2QLae4lBEkKha6o6X5qhQ0BLqwMjy9Rs7JoADdtsbfXIlH9oEgC8l5gZOZnNxuC0MKWd1ZF52sBWT6xCPjTf1QZnYh4ZydTSFpH5ru44E5vQtjkTsklzILDAaUUIWvQP0DCXuNjiOkbuf9QuY5ifSbUnypECUXKVcLPAmdgVBtFoWyhd2xEnmtmlf3JhRh8AkgbAYn/3TLWvSmFrMI0M6VgWIKYfiGyrPFPjWQcbcItOh6DGTTALoB4bVDzz2pdOHpnYj2fMwt7asY2DOXAw8sxhejx08AGjJvRtnlfPou5Kez8xdCYHc7rEAzTxEziNHMIscADjMlYNh68O9YgXz3JVOZ/q4FVsbmQbF5sb+UnkNHNkjwbNQofzkKwOrGye759V4sMznKKr2BxbNBn0P6fZOgM/mc2cKprANtcq8yy6fyC6d/U23rGSu2eNCs5PAruYv5xtHG2OmxoN+Z9rfWrXVIU7oofPP1p9flC4gd42HJTkWa/Cwjah60hsyDZiYkO5XJZ+4ux/tmkomElZ/uAk5TmQlhLWHXMamSdDnEEwNh2ipCwT6xDph2a++a7QprRKVOHUdSYAWBVCSNaX3H/+0dfne9vgZOOAwDyd6N/28Xw6fnwcj8f6TfynZG0TjQpNVNziIx939SFhwtgmtMmXSphY0CAoxoemZXCGkqt4+CHTvV3PyZmmOp0aZi3Q54+nRu8t4lyRpIcqZfHUu2OjHULFbmfqgH0ipQbm90PfFpFhmrIOztAiNENDmLLg1GVE1jRli7H+jX8dn54YmlTPe7OM2DMNPPZx5h3M45HEaDmUilTbtDQ3uirBYptBvBmXtRKvMYepK1c409ydWBGoTxzNmvTpHOlAE/U9F6qOfZwMTKJiAnMjBxQ6NyuacArUI6Mxa0Mi8zGn2Lj7M0E8Orm/r2tbKUycBaCJm5Snvt6fxNg8nfYmRu9HEGFXIS5YOe2q24q7LmAcQVY0NVEfziZjZB7ZYqLtCqCH1kToswKmzujslrs+TDY0GZpMcKqN3s8mFUWW997geCIjo4tbrGhqjB4Eo2gaO4OYAx5kmVU5g6A2OtN3i6F5roCwPk6S4FQbvW+aBl6RKxItD5MvtQc6HYbmpi5vNxqaSqOj6bhVPsaJWqTxVVnBWp0IMRuq00vv0Sba4+n0gnXegoaN3t9QttBanj2aO1Ojixs6mkVZHA5lWRRFCTcqmsrCNZqOW+MDTL1ByAFffv36RdPsyBOxLbhreJ2hNTQ/yAZuSzmYnYQzotszODph0KNZmaYT8YwNzU0O2y4ARHBSKGgqQxBOJ+NkOkYRWvD1F8X5MPZE8OzoCQVV61T/aWgeG5jNFqxqMITEbFCw7G53uwGaykgRK7qGZm/ap5lT2Ig1CLxKt2l+AKEFv6gM86zmRMz75UubTTt1NF/OlOnf/WAICfkMiWWPlqYqhCRppqHZn5QtFDRVnj2apmEZJLTgvqb59Xzle17Q8ERMC4oXTicZzVZ3g72QkLZ2u2Kno1kYdg5ILAFK2aQP3lNBU0VNkyg//cVqNRmVqtEgzeq/vEpTmucWVAIzNc2XwfoKznSJViJomBAjFc1S2HVjR1M9E/KfJ05/jJsYFGh+qWl+UdIsDN1mZk3zlf5EQzQF94EDNNFw/6jqT8su0eaHA/lVbMIu1W6ME+L/Gc3X8+ODjDDfmdHm+/uRbnmpf+bWNCfaPCsMUOxpNvwwvRC/IjuFl6I5+cTC/HP1r6P5JOi23lLvMLGnCa2KIE2mldPcNDRx9wmbV7mCpnWmnUzSPzo7/D32Kxj7frNw6DfPsF5fJDTPKF9fX4f7TdF9tD1nYk5T3m/WRVBJV/6WZd6VQRZVkO6L7FWL85PNHI+Z26CGJhJq2jt5FWQwQikwJEqK/JDQbLcSDde0spaVO4tEq65p5SOUgo6CziI8L1bTEv1Nu0zdf/ZgOv0gnIjOHvyzfBT23Nc0c8MWMDFbvjISab7XG2qg1WALZJSTvLpV0CyNJyNV400Sm0nReD0qyfxhftnx5l8Ypva/igwNB6z9E4EvFOZy2TdZXsO8M20BE0JIT/OD3VwMmkAyLJzYhKZmLihUjDeJBx++bw6Hw+Y72QfKRyjKWc7BwX/6aX6RuSDhRLjjXBL1g7PpPyuXedo9S1OoaY8szdx+nlZbB5UWaUs9T3uAsFs/kpQbxTyt8hsI025v9DxtP2+ByT/LpQRn0RRB0NTRWaMXr2/0QX68UWgNzlvyvN1CHnfM56GFoxbGBa3G0TXfoRh9v6mENm7pgTwdGXb9ybLFiUSYd+ZJhPEn9MrApI+O5tvbW7cRv6gMLCA9pRznLrdxdPGUdjSVbgJm4Sia5mvExBM9tjiX32pjoG/LpWJeT5PRWaPvCUeq5ndL860hXG95fcsN3ETOOZH1nblVoIjLYizXHoRj+73RHxcXhKHlWY9Y9bNXWa+p+1adtRrscDVAO5q8mOOrl2WoPL1ZQ6vvNLUlSTBqXZAmn47rOIH5YlzJNbA4z3rDeVbwdc0Sbs7o+x42Bc2cuSz11+3ydcioOprA1FUUwgpEOU1FFaRxEzCFvyXRSoMgl+JcijC1q8nYqM+NaBZmRb0sCHIhMHeJGTFd1CMpzVL6bbV2OXswZtGezYdl65Dl0ZlbmYZfCF++/fz5dn50NJk3f+4NTcP5EIJlWVRip7lDDqYRup1Svsj9IKGpjaAxVa3VGjF5N/RNYPkouVj9/XHcbVMV4fWzZdrSbN6kb7Ohr7/5h7szI5etPNhBpVkTreGgEPOiZhOKc2MzJhyx2D20uoNTZjkwyR95lrk0ayX6Lpm1TdWwpOpodu9wMAduzOvdNpX3Q3MHXbOW8b2vRZ/mwM1niXNwWt4frLqlDH1rgT5+k1/p0K2rvG2qn2cxNFuYiVlxJbcNLHa3zSIvyT9zYux/FvdyFTzNwZtmXW/ztf63L7R+k9sFtc42lYYmV2ENu6M0hvI8R0OOO2RT8/ssy4PV5yLHFdL2//BB4PLvmJj4ce/Ww1JJk7uPxMCPI7f7If/X3rkuJ64rYdTAxLEpUhK+5BxvuyZV1Pz2+z/ebkk2MXfLmAxor1WZAoKRTX/qi2Civq2Vx9++bg9svpnbx3qfX/zAS0ZuTXT0Fv73+6yafzbe/rGckofG7DE0aZ5sR8TRxH8foolzYMJWSuNi0sk8Wf8+UfPP/6dczIS3OW7SLieExFEXM2WebCbu/bP2Ps9m3DxZntT8f4Zq/t6cBPDFSHdIvB1o+xh3GBkovPeSm+Rl9i382j5CzPMHbjd1VVVNVf5zWmOtxqrku4Pa6D0IE/9dcjdjVd94ijlxlSrvdeX1Bpazv1cvB3LvNXmAmN67Dnq8Qe9dcqd+WO+1M6zPZq9eB3vtY+yz0a/XLrlecnop5HPwHWJaOUeeKfHeGXbsRsZrv93Mx1vdc4/x8TuHbxfeu+QmY4Vf3vX/T0ZaffXmWzKN3CV/9P7v3lb33YneTNjlOFv47v8/8lJ8Z8l5q69HvM8pa4MRxpnSNmLU1awWE6b5+tcoWyym2GIz1/y/caabTUDWb9MCwM02KqaXzHbSDLwxUUY0Nrlk9esXlGzeJvVBSm7225Hot5ilCZ9ttnTxOtbTuyLZnkHbK1pO7eey3VzrAbVd3u4lc80WqytaTm5Zsr3aaGu1mK31lH0P5zuemV5o95zGKHa2kdh2s/g12eRuopzv95Ws7xvY2mL9CFusL9litXmbo4HgodnNuQbvItmuXX++ewO56de4Hnb5swPfPRdNWznTMm8wcGKbEi42d0Ys24Dw2BaLu1v89X0CD6aKM/JiFc2MvWDTOHSxFOz9t3n6qa5sK8+3fmDb23I9x8DGwb8Hdpc/S9vTA1sselvMMHBie4Z+28IOvH5Qi9zVamMu/s2cbT1n09bENA7tBl6uZx/YjmwGTma1xeLbFjMOvB0YebPeRgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACGTyE+SYIfnFumj4/P6cR+xitK4wGDPrWbckV0/7ksOQM3nV3OcQolKUPPp8VEINV/IN9NMpR/FR5ZESm5cZ1K5V2Sm9lFZipovpWb88fVVxPHHZ/z1FceftvaJC7mbRlFGFfQSan65mtakxVjcMHFCqjgWH40L8cvP+AM1XyVvdlg1TUzNnGaFyPf+mbqyN0HNF4y05ubTLVaMfFLJZuK2qBmEmhJ1C9QMRc0PezdFzRdcb76fqGlyp7mHmq9T035a3s/4phRE6j2Lv1DzZdTcf057Jm/K6jP+UgVqvkyoTZLE/LPfeFl9hzepUkmU9N+G8Y0YAAAAAAAAAAAAAAAAQMiYLw/hx3jg3ySKkO/w44im82uJkn9T0VkFxSv/Oglaoicx9mmZwTEx4hPlT8RETqJsgNEWMQMqhhAzIDkRM6Bgi5gByUk1G1Jli9ECkpM4G1AlRJwNyTkxWEByEmdDqmuxVkDOSdYMyTmxVUBlLa4ZUqilBgop1GKpkEItlgoo1JI2Qwq1pE3UhOdMnNgJNeE5yyDsFJCalLT4JqAmEGkB30RNYL0JT6Qmn+yFpCZl0GvAN2L/uSKIxBlSoCVxoiY8Z9ok1AblmjhnWGpS1QZT0eKcQWVNnDMs18Q5Q3JNytqgXJNYG5RrEmuDEpNYG8JSEzmDS5qkzuDERM6gxETOoMQkd4ZRALFQCVJMom0oURb3DMwx0TM4LdHz78fYubs20hvur0mZRo/ANsbFuj8q5OO64wIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADARNQ9Lx5uC5Eqld5zEXe8PGiKRrdtXo3SqWn1dCPGut19n7Vty4Nnq7yjuTWObnUUtW2FdKf+UrUdu6vuUGTmRg6LJ5+qlEljdCzUOTXz/jr0jWEyOUah5gVva3W1K40tiyuH5VaIqMzzO6J0ruUUqm3rC2rWlpvTpdIlvnmWWsRUA8e5oeYcgf2ymj7DoOZZlVrVZyN7r5D85XJoWWZp2dgHdSm5qiyLKC5Fgl0ZR7V7Qu7XVqKyNI/SXZM3dR8RZQA7TCmpNpVXK/knI7ZtI78yaprh6/RUTWWer+xI3Y0Zu+zHtudEzTNZ89gjy+8cKvbSNo+pPqeVJi4b3SuXbAtzP+9eJg8ye3ybu/mh7DCZS7WxHGAEVF1yVPKgccOnJ2qa59xllN155XdubBNI7DlR82xFUR2JqavGCWVsV5Wm3pV6U3wzz+teTeNeua1XhmoqbVJws58gupXydNfaU4j8Nriq3Lwuz41vdsOXezWrqpEfZQ5s89IKqiszpQ7HRs3LSWyYvVJtJ3/WOnu1mftVtFet6tTcOfNnB2o2LlZXfT1VmVdKwnU3TXeyep83zanUXvt9TVvY51I3tZR168q8KrYHadQc65tFt04RXdKos7PTSO8fdOs963XFgZpialOT7vrFjoyWSSiXG5UaCY/VrKJen07N0qL6y5CLa1w2sEdmRVHYNI+a4/JmZ2kjzn5FVw3VrAZq1sdqtnvK3tN3sRybtzuj6Ima5bGaR2Wv6g6xF7LTfcZFzRE1rdSyqlezGah50TdP1JT7bsVYZ/1aVsqlSpw4r8xh96gpr8p3dY1v3lpv5mm/8OyzWDr87OyWb9qaVHeR1tzP6n7VsWu1lnSXSdljxrpHzaZP4qh5wzn1Lq7NkqN2hWUR59aMB75pnhCXO/FNeV1TFE1fvOSFqvX+s7nURMbUFcFFL2BmTlifVbNwXFCzdJ9Coua1zJkPk53qslMTHfnm7mC9+a1m94LGytUtVrUaTJXcDaG/3dGe8Kya/ee0Z9Qs3HNE2pvB1thRN25VocySXtuaVGunprbq1Hkuv5YHYndto26szceumXXuzN63Q+n8+9uYndY2qrqh5CAzsJLVq+4fRI3uyrBGd+TmudhOrcGF1Lo7j5LzN/urgzOfhw+/5vLdnP7wePW4KMIXmgAAAAAAAAAAAAAAAAAAAAAAAAAAQfEvTUPTBH0zBT4AAAAASUVORK5CYII="></image><image xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAlUAAAJOBAMAAABsOe/ZAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAkUExURWyO4ECR6iQkJUCX6Hpv51eT5K5T6AgICAMDA71Z7MVp8c268unKkLYAAAAMdFJOUzkjASolMicKGDI9VhWjmXEAACAASURBVHja7F1Nc9s4EuVhcd+KD5tjwvEWrqNlodbH+QlIUKiZn7FGpQr6B9LPXqBfNwCSUJL58K6IiLYlWaYk8PF1o/t1g57mx/ad24fpgcEDqwdWD6weWD2wemD12B5YPbB6YPXA6oHVA6v/wRa+vYv+P3/+n8RKp6+QN53u6Rf8GuhRfsxP53vaRZ6m19FTGiPFX+t3/lvAu+NB2g9P8K75jXX+md/s8/Vfh1V+u3LKcRj8BA9b/pBvZxrfzE9rIQqNNODZvAezR7enWt421BNE+/Lrw1t//p/ESmNkM5/GgDHNcZazilMolNCaX8BnbWbagBflzzjTQlTNG70q1id4N3n9jE97i88HhH8SK5wxzRYx61norOVEgcVp8PgTTm451hCE5Ixz2Ysf4ECZByGEYuyhGGCY5R30m3w+vVvZ4w9iRQfCJ2TW6Z2EW7xFPZcRER+CeJGylbPI+6X7sHJRnU3rFbWiZsLM4U0+P2oB61vTylewCvNcXahOSMU5hpjeOob8MI0u5PGmP0W6wwjyH3EUefjpccj2EulNouyb34ee1/R++SvwTz2EyK+V397o8+cQm/Oh9R/kVWFvpGFiRPXkZgTzh9LB5KOINNJAA5SjoPFgj4BR6cjox9iQlUau8fKIT8lj59fSoYS3+nzaHxNC+Aq1bmLFsy5xnc45DTKfDgws4l7jnNPRyfhoQHnf2DwZtUBNw6UvOueRXq3x1pktzBR8C6UIiTf6fNgqYTnXGfJ3YFXsWfMp4SFpGS9hGHAIMmyML9LhR5iOLscc5JAjkwAHHUI5LAYh8lvigLUcOQ74L//8/EZ0O2umV/idWPEEQ+OLMoR8j1Mqw4my0RDkLxofL8/jkCIPXMMz8TN4WeA3KmiUjXfEEb3F5wM6AV5rieR+D6++PNlp9C0doVL+9A8iBzxNpuwtsG5g9fzz2CipBNJkVb6ZlP3bu3zMmCE5MNXfjdXT8JRS9J3Rou3feTaLCC0QzH8vVr8MD1SyPmsTVOmBJXqdGCzMl6E7GfawGhyqhBBurC28UupXAos8PdKB78Lqp+EdOplfhktlvHj7nF1WjBy19pLpPVbP409/Fp4dxJqsV16lh++IWFGyhu/h1d9/AAsEUjQNKrAq4fUJxIqwwk5IusPqn8M79QxRAYu2BFUCS/0LxOJcYB9k7bCaxjdBYMZuSmWUfEbMKzHCSMn6N3n1YewAFKGCraTiqdCnTfnPYoRx7iWGW6xGj6xoBlQTnDr5KfJVGbV0V7x7Dku/5dsH9lbEJTJB8Mr6qbh1RbTy/l1jhLpWZPpYjR6xc7CgpoTN5MmrK4IrmWG6TVPh+Vxd1ld5pQePFqydSlylrFJCqswpm31WmKuWE77urz4M7agQh4JYmPg4XqCQIUOVvLu+XC4AaxdjrbEa1gQRVikFbWHyElllkCw4RXB9motKOH+DV2OLMDQFloSZUMp3+Z4MkIxQn5lYBJe+idVP49LKEq9KdCVoASzP0yDNhBd27lzhvoXVf0aO1UlfAGCcLHsEoTBD3n6dz5GJRWKyvoXVL4NaHyl7BJPoC9ll0QTIMXvZPqWgIRHrwoLDbRscWoVpY/Xsoege5meLCXo/Z3/FvIq3bfB5UFYpChmg7BVxz5NDF1pVcr07Z2JdOMRahe4tVuMKokoCK7gpCda9WuMErK6JV+dS0b3BqxEjUSAk85+tmpUiqIhc1bN74z+fL9dELIC1nglbrAZ07VZyZgVqkdlZ9lCqRUm20/l6viIppGLhj4KVQqTQlCAUp8teTLBFyyVamU/n6zX5q54RtljZEf2UWkl7JQ1UGzdVbHDJrJKZMIXuDbVarAaUQUWCsUVZl0gBvspukEpb4tWVZ8JNgNVgpacBwULrQssrxOuqQylDZpihSlNhBkuvZfcGq+fBgLICUzv7WfHoSm2jhQRWJtaVeHUWh/VD8YozZSrJe3+LV8bxnXtPvJIEuhWSR+WVLUkgpzWUykxtNqPaGZDMz+SZELyqE2GPV/rjYEEoSqZQ1zN2MDvFtLIdXuUf85LBunBSqFsNa0xeWVVj9qpU0SzoO4kNzX/OOfohqK5SoQhd3x4+juWrrKQ30uZBSEG7WlHKZZxghk541Tj36rBG5JWFqKC4FC+yHifNfp8vE2COwnbjrlc2wjPrfT0b/DgOp9BdJVqV4MRhaAcnQ7xKk2AC7KWBKq7qqY0NDsIr7q7isKrqVaoA1eY3Ds6KwUq8MuDVlaWGVmgYjle2QNakNZQre7pRu6zGUQxKIUO2w/fg1fVc2tY6McPzIOYHgCa13nxth9nYHzt3IlX6eQ9ecYUi9HPnj4NAxRUb0WBYhKnS+sqhYx5MIGVO5RsHrCRq0GFQf6VqZatRFjxHWKUUuIrYs2N3CBoMbQWqi/TLjMgrxQ1DdipuvWpVe2GBzc8wYrK9wLVfruTc89KvDq++HB+q0oiWAfNtJQLZjd3pMGSAsEKDyF3mQW4ummswOg6vpLmf/ftUcIK6rvZ6VeEVrJDJ9SKxKElYbY2wYKXnL9MIzt2CVdarpsXD9vRiwsk5gISbBNVyLQEWJc+hJs+ClT46r+DOuWJalRiUIJTfIWYYKopAiVFLxmsp/koi96gHmwelEVSVZlC0d9gK1tatl1iBHVWmWMZLaMWizFwb3QtW4cj6la2SsWqCUPHqVlocd9kyR1aZULBGx7y6li6sGrgPMw9ChEmw+QmMEkqVnj27JpVhDcbAAsmvL4lYVwmwaCLsasjhwP6K/BPlgF7BXSmIe1zZ2isLRa0yjpFyC3n2yisE7nke1EPximzQi6jnMRf2jc8hs6EE0HHCTLZnANVS/FVEQtjNnT8e1vg2a5EmXg2hWF5oxXXDQijLC8aIdJzdessr7mmIzaVAqg0ecx5EL7aq4l5RixEn9Hy6lwSQs0A2vgRW9VcSNOSFl4PwqqlBSJN/7e0o/Xpqa4HCK0RVhlwVOfb0y28FrDN3jep93P58UAu0vM4GGgxr68r2mzsw/UFdh7fynmmVbpa0bXm1z3HSM6+HtEAW12tEVaoQTZ9/27NgDMehCN1zZEXBFczQFH91uc2rI8btSsTiVllnYa8XKaBaClduZPJbyK0vApbwqsQM9dJFR47boSzIKjcR12Wd6U6yMnBXRtR1iUIXskExwWYe3AajB46vLJuhWpkgutbh0FWnDp+DBobJEZ2yBWbbSzhlbi3rfDD25sEjxu0wv6l0eMAGJzSvr5Hi2hZqgEwvDkKZV9kC8/bbdaUi40qTx+YVahAiFitoC2rfBLqilZHWofxgEZeeaZV+wVxY9StaPtHNcQ7prxioSZa5iW9HamN7UqiIe+TVOQckn+7IXS3NPHi5rNenHrSfgUyvsCqvx51kyZbalbYgEhuW1ksO6E0TKrARuuqvpPujozPoQ/HKTiVQ4ARQilvK7rR1VyMGV5QFBKBGoJKt5oNcIMyX9NvnOK8HQoovSAGfXrusbO3ZU+u+PWkcYqTECplSlDW7ddwOYhFU4bixqC3N2LaqnyiberUvbZkiggpUjn26g7QAjHiT3o9r6XHf8SqFXK/HIZWdpqpV1fU1vRTQsbInyroEofnBkqMqR34dc2DeXkp8dV5DNR+x98NKBMrL4WvXkLd2v8qGFWPD2bIkN0uxvMZbVV5dLhWssNMZDsEr214/zjYX7lDK9tvQBCwn6p64dUf+KoNmCqkqr0qB8Mi8slizpeqFhqQnG2Go3U6ARnxVptVSGhcWygEzRkY41forjhlC1B1/dYCYAVfOseWKFPmKMKzqWV5m6rfaHlFJqoAJs0UcO8+DWzNcdppMOGotlefApglG/LrtyHrOV6+O79IQQ3G6WU2BK16VfLCnX90/r8oaeDa+SbwVp4J2n9c4J90dEBc4VKDJz1Rtod3WtQnd7f24e39li7hnIVl5Ufb68YLzrkoLtbjFSkzDK/JaBTGBioupYV93Th7s9f55xZUaXGRoapqxZdmW2i4aIaurkpWUIBaW+BqM9rzi1QB6Pl5tQiZABomr8Nb6m0IMyeuekmSyQuYT7hfh1cplncym96Ob49y3v5KO9VIypXCUq1uS/Nm1+iK1iGqBUuEiPwWQ3A1elYaGng3evb9SbclULtzBPXtqFVgVrZhLymjaE28FsYr8+84Al6el5oPlGjzHmgdFreKZsBHXle0tG+EqIFBDALrwNGhEXK+U4genhNSKV+za9aHmQStf66Z1K8tstlVAWWfKSyGEVdUEJVIwG1I9nRpeIRjtrF26Xw2Z+4XQ4tgUtlQllb0hFnvnmz6YYoBEK8QJpkWKbl5WsWg43hqT1UKkqn9yI8w2qDK8fFJiUPZVrQG6rVs/EVJPy7LRr3r54N2utQRK1QSbrvUirqttf7EkM7W1qjQtkKzQ2U7ZANPNSw2vOMfp9Is+36cByjWZ+ALi1QJtx/xcmQO5copUEP4J1mfE+NwerVMzD2ItvQ7dftHXO/VWaPEQZV16/HeXGFo5K4nS69oRbkVz1GS1bGMrwHRq4yteZNJd53WXvLK1Z4FJ5cuSJI9riW9pxZekcLLIhlWYWnww+wD0ib4SYCeZBy+NJhP2NfrXe/RVzCsO1K3oVLarrZs9r3J5ufgqTm6Q2Oyc1lP2V6fl/Sq+ov/HdJT4ypaqFrfqlQvo9C42JE2gXsT1fOUhhonxcWYTVmWnfqIp8Kn1V7hgWIjd6zO83p+r4n/2U/7vT6nB1wKE2qyvwU1JAhdXK6a7RLmJq04UMOTvbY2+E7ffny4qlxmS9bhY3s3lQPtf6q6gNZKchzYMhlyLuuS6YSDXLYyhf8J3NBhD/41pPsgcA6GZ/OytsvQkuUqV3ct01/RsOsPsTcjy89PTU28L2s1tNbl6BWKXapVYPbutVUuZatUqSr0y7cFNXh2Pb8/wOZHPSVCor4RJTBdXoarMY1muvh4sMFifM2o5gPPHnME/h28PaEOA4YMhdq7eOJJI13lusuWUqDuY3+OkSn21okrVsOgcrPNKW/t2eE1RRl1fSKoT+8Hw7AhcYcLajsIQLyIdghItCmDoA7V80xFsoZqmTV4dX1MEMXaWU5jNeFvwZycJU9Wk0nXOJWbZV6W9Yc8lUGPEEdS8IjeLq8e3H6pe8dQkc6Bqt56NdU7uFXtFGCuRV1FGFRw95wpsELTdfq3At4+5Blt1fzk+bs/GFEbkMGCLc++fuiJiECQIF0pkELrqRRBap3IlH30P/oSzmjOPcxh8lbP6wVgpNj8Dw5atSoUH3IgvNv3SQhKPFL2HDYVrakh04oCNz11e2d0vx6tX4YRdbp15qtCg63KVxOur8hySzJlSdiXzBOxK+zjJNTjyJWhqO96Dh84rSqrKWaUeOlyjerRQ5P1X8bxZvrW7RVKYtC5Yy7Om1XV6MI/DnFGjzaufbIr8ej2ut2HOjNh5Lp4t/Jle3wOhaiHHjYhSMLhlGNANYGjcQmxlqsVpHAflRVvBenV40Zfr21/HqOhkTM9A3XQiqufzRahT8mp5KBu8QNGhaK2ey4SnGCpobZ+jJaEi69rrq+f78f0o1YqLu8VVIkvb6XFV1mMbMLrAdZYtbI8gvW0awDJ3YDuN509Lyiy13cFXPw4RqcDzEK1SiY2c8FWuN2/ivDIOOpRc+FkzxvykmQiIEl6ntBrGVWvi2O/BrH4LGMmtAkjrWoydpGXD7MICIFiLLZgKzcCyxaKTho3yqtfJWFOng/Wd+QgiwYQxlhazJ/Io0mNep1WEjDb15HpEiJRiaHGaM2t47mfDr1u96OsheNHM7HrYInbH7aSAVi8sGmLlnrQhKo+N7PB7I0DWyIzMHKlOJ9Ow6NtR8yory56DbcrzAjzvvQxvCgD4KI3miNn46LF8Iz+ZJ1AM7ek89HNeb44f8lF0MmQhQPshTp01YRa3obWVI5z2mOZbwtR+s9q4pC0ZymwMN7tGZNacWOPTWoDlzdH/OERW8fZJshBXUYzYLnj9U5kjWYIVycdKOBmmYnj2xiIGfuRERVjzPwwaqrddD7qj8AzZAAe7VZgFDKEnZAoeOoUpURXQFp071SGS0rNXE792WqknrNW/nd05+usx9AzZsg1IKx2Rxxo81yfNDOeyUZrCqz0hw8S4oYVpKVeT1X78+gPmcQg0nETxUTVUlS2aepteImUYZcHXMWKgskaIj9eVi0K1lCvQfbY/+Gu3l3owvr151LPkg1yaRM9e1/5DCX3URtBE7jpvDmJMW93xJEwy3YvjZo7+9cB5Fbor8XSqKPSGb8h755BxPIGtSN0J4bEwBEco/iOely965XzM3x+N9DN+MtB+XP8QffvSm8hB7ZoyXMD4QF7aYXyv75eQ3uV9CDv2cvl2qe/nbyndzt/ez/W2/Fducf7iJ/W5nOeYxfgxcUfnU3UybFz7Z+jb2djxZLyaGuXwFE63/P6Un063OZK3p9zw1/z9lG8sF0WRb5+negOB2mRZtxlMfMTLklb0ab/P8UN0fb+ae7Sv2X55eTusDhkGOydM4bzLnq75hNb3OffmxFoy7L1Vs1SMvnZJs/clRu/pVi5lzrFLSXVOqzl8l7Kk2Pyn5dfHkmZDtHKGXzva2sPOTWR+8ciuKfTEGEks8crzz3JWl5p/oz9/3+BZOB/Ip+Vf5vRaDm39++NSyyWWy3wy68f85/ZxucRYPy7xfIlPna6vccgbru/tx7GqFHC6vJyV5As7dZ2Ke/u/QKP8xmnsaUxLkWrU6fyrRAw7M4fazy7JDK93Bo+TV0FAKDoTPHea3QhBDkNQKtFf+PZLoieC8OPfPqnv47h+yEfRM1BNUkc5sFYwqMgbZYzM6gKtL3miwCrKmOBXgZrkkXju+zhuXh0EXwXZbSNyK+UYfGUMk+wM2tvbj0ckREyrjZxSdqLFUHTVx3F1MseYo18oUbv+LrNRRd205sV9tohjqBqLC3nFE81xl/SDQIYfO3Yeh/s4LwfF7YGfzCfxHTKq0Lxxs0pqVp+MPi3q7DfVbpy+4mhAJvsuNLo+iGtf3xyvsMfjdtoVGFoLp6o9hbusBc+ZtLEfKq1LyGYVJFMzbLunFx1bT5X5Bq1XsI4+ov4qsJo2iPUXS/m2MzfC8NHbrxrZPxWoxFGK1oHIrVGT9qCbskF1Mp/w3nk92Lwz9eN5npIV2XBK27ZuUlEd7fwXoRSi+BqzNBth++Lum6BtoOegaNXesGn9cPxVMM2bystt1P5yG64EPysYn2BGokZxNsZQ1869N/H3iCiNVlsLfwZPz/BQ/ipzUzCrhEGyahOpAvtLCpWI+AikA0mts6lER/6oFYuTa47becWLvh7rHsxYWGbcZ3OLUnA2CwtjwBegrIuQm5BdPThgxWH4lDzmyt5Aw2DmJmSb0NV7O/947BlkxSMwaMg+ta5u4jQgYdaQRKAFmrgpJX2B0qVaqQqLAnfubIqub4fC7dbOkZG6ManI2fGflbzi8STSGHOj2brp+C9AAlIjNyVGyCCbBES9WD/38upxc6kBpo70/KOtSZCFZs9+XRclVZw+sWmCp05kwjj6r8AxyrzEnFKtj8pimWjzivxFHS+L7w8M1eLPdEIvvvIRzDn7Y2/qvlfR4KrozbP1F3SPyb0AWc9Owj5OKLoGlUP+lP7ggfwZMovT6gk7O42VhzdzKn6OHC1WGuPxZ6QLZRcwcMhYyzBNLP6Yf690Mp5m+zFvZ97WSSWr9mttwPFtZ7kK9EOGfMGEbkKVcjkYdLbiqClF+QQJpPGO/snzOAfaE5f1CpRHYKjOEsoiz+VS01bEx4WqFJ3m8jmrkW5Akn1g0hIxi88rQcOLM8P7iHq1glVowsN4aLvbNMEhVJbgQawQYWv1BUXFPebITFW79uzIRNMVPfd9Z38/zl+POYMnWcKFUS6dulnty02p4hAmFdBiQAIuMcnT0mLwFHr2yJOVg8ZpbH9HXum+Cc8j5fv9s6pJFgIsjbVRWncE2lV32mh+GZ9smeRKrnBBWvISHshEx0EK1nO/x8TdC3Dv2p6xLfBk9GhN7p+zU6qKuoXajZ187el+m7gP17kgTRAfIzoERtvoxKBzE7/EdOAAfn0hgI1hvliMF5zFGrRypBr3dbtbCr0aJvWSFyadAUc1HyIuwGGYWmINpl79gu/Hw/WibD+rXh4gYILTg+B84pb72tAjRfVfTzsJFbmeszpU3jS4/BZle8uqYTOP83AfyEBdwFPfhjCWAtWwxklcjSubM+nkt3RtInj1knZExxOrQ4WrGmSyZMABnH+ePzdg9NH3YD6xPtuI9jAhHzaKUExxJeP/zADU+O+VPaQ+AqTLFE53/bXAtYANg50fpEP48uh7MDMLE4xBDOuqstd7Z59etamI1ictpf3OA1x18JyJo8UJlF+tXI30vboHHz03EYJsIDnZmRvPeYF2b/EoJQp7tJZycGoqe+FiQo9ffNG+a4YJSbWU9+Uz9bo+11ft9d55RU8bo6pCqIK3hxIGYJxXUfeVJWuVnTxGgY0F+scMH75hiRefP7oIt7j9kT5FWFfdfSoTe44BtNmam6SvpUtIot9Snszjz+TVIvjvpuBGquvIq3HjU/RYvp1Z0JP24NvaFheDYmV15YPHLXjDwyQ8mcvagW+S+W+4xjRlNo4dX4Atq5b8os/zygfy+kA9A7XgA0R6Os3ceszB3eym919VpYIw68lx/5omqVTUopHnzBKsaRTQQMFqlWrocLvKZB7JM7BRhXqwN8oYUcqrTRFVVmmgDV+Fqtoxn+3zisHnaI8dg/Xlh7Nq5LRS3P6JevWwXUKZjJqsHk19FzayBdjVF1kdn3hPCzeV+U3jdf/a90CgCpCK34RLZNo5HFs6Deaj9arx7a7P9p3mUhl8YrWN2F9mV4eWquqEMDsSi9l/EN23shn9Fsig50+A+kBpNQyjDdazarZ3vcLudAZpllKmbKpaL2yHvlkzJNsPmriRaXUZO/VbWuL9BQhqRprpL0uE+kCN/Rnk1WfePq+X/9/pAJ6Cgezq1+s14jmvwCvI/FayQ7nJ75Pi+osKqgb4MAzt+Mk1OLh5JXMT1wf4twdorMwMhHB6a0O5Ik2bJAPyUZ16izG2cjhQNWqa9ABGKlEcpJ3Puo/jzePcBbcHO5uLfUmh7iwYLiwITZauYoUC6BeHhqE4CVIwjPo44Y08DqBgNlF77vdNeLzob8cM1F1mxH5CH6IGXH5hdQBTqsXgdGX2ECpVwqSe1ItsPESX35JW/LqZ+KW8Ludf59X1Ib4fNAtolttkmKs6Nk2i3+Pxh4itbgmh8sWfMKhQHwFh1IlQwFvw61jxHL2XV79d18dDNdSSCOBCiQZdb6yGYg9wAQ14uMQUr1EjL8DJCIaUeRnoCjRwauyB1Rd5dXfMoKp1qu7o2ezaX8ogM3laiT1F0luwOKTeFIUgVmqBatQk9UrCNH2RVz9/dvDqbjwDoYUMH9pgHWjzOquKeKXxrkAGoDGBWEieUo9cPGJnETMKWufbz3JVX9creN2/3tsPOVCXOZ9EtY6VSe4QEoH2VLqNLayr0s0apWs/TKBBuV7Rc0beNO1pPI6AoP8WK6lX1zt7DoTTSRQe6qeT7ZxNt9m0wkQH5HpSxXraMfanxp/ynlHqOli9gWmqafgPsZJ65XnQ/da80nEkrPU2bqHBG91KtdN1iKn4ngKbG8pRZWfSdBhB5319+jhW/+vq1au7q/E33oMBSkdFVhqqvHIc4jegLCGpRTV7Ef/1zMLEbBUllVR0fS0LTTwOw3+MlcmrO89NBJbviU+T0Vh5eD1pXkVdP5KSy+vppEgEpxcNqT7Qaxmn71/CpfVKlgm93m8Pb8AmhJDtklNnWaCEqVZZAiRPQZDFnqs4hP3jELv2wzBMAjunfUzl5pXOWt5PhxxOcKZY5t1kC2yu//B2LbuN5DrUm/qAgpTFbJPu9looFzDLEeIAubsCBP+I72A8f5D72bck8lBUSU5cZafTjyTdPYsRKIo8PDynNg0MIY9tRLNQhJlCw9Ytj0pFxIo/9RgEyojm63Na3sGPK5qZ34i3Tzv4IGAraZdn8dW6ctaoJ6sILcAuG8o1BuqUVhofksvslxuSep3bGb9qxNU34aITE0I7iBU2QVDlrzhm/WdeZ8sC7A2hXuBTzFUH+pKiClXVmo8cV9Ul/B1aYZMoHOeBfLURGKDMpPe8U9kuWyKh5VaGddKC04gqgebuXyBW12uG9A7+Ply0k2qdPRCmQhlN2kEsAqK/iZE1cLWee8CxpZ4DecKK/MlH1W+KK97H2e9/V25XXSDMNToRseraQr0Z1gtqr7SZ1Q33M2ZAxwwQlF8+10u9blbfQeEUPZ9/j55MB99qxRw61QvMso6rc3uQrS2hC401sM4bSGpUQ4BCSlamvz2rU02h7uDlGvfjvP/78ecExV7lhidSAg0QZszCaAzsjRgDDg2hL55tqWPiKnSQNLXqoKhUfV8IYLXewW+JqwnyLxgv50XTrkGxGsVkQzWB5eUbdQNI22wMHTuBrBhL79HWmA356l/hqp1/w97ExPtbnVJWnXgjoiJ4kO7lKcsY5/OClGNYDCAcWI1JvzENtHKrnAbL/YaPZT94bu7jfFN9lZa+uVjAxKZ0rRYLEm6Y0S3LOVWLuE4p1pvMPuu5quL5X39jqf4ZztDCr/YPjyvYUO6mzFugsU3X4lhhYHoa1SZ8G1YYhsypGhb0s8GAK9RvCStT5KvLNe3oh8fVJH2z2EsxXKwN3kQ8R+C9UNorMmI8tuYQRplqqAUIqhZufQHdZ/WVGk2od/DBccX0da6oCmGmmgoz5sIKS0ls73bFrgwgjIEcjGMqf6ISM1Tc35mvoBX2/XEFy6RdPq9T1zX0eYPSX8dqaVjM35dR5RzWamQBwnAPuP6c5L66+h1saxs+tG7vOoDG2TfwJIJfkwasQl4cyXNmiOeAalUxZgkF5XENrvbDcwAAIABJREFUVVjMZeyZgL0xsp4kri7wias1Uh7aD07gORKysAPTuGtz0WCwCDV/pezVRqwcZHlTwuplU4v4Z27DGaFsrXL7paX78eOxN1BUBHaQWzhdN80dRe0rjHlgE8aKhs2zZFhGqBG8KYek/Z35SmZeDV+Ah/bOEwmwo7E5Mb4+LdZHRpAWTnnDBr+uDmwGlahkQTKxFQy/f67/KmU5dUNN+RK+vhcWoJFSdD5/IyYz7ZTLaVIzzjrGmb0XsDoS1E78KMUU/6bvYLaH4KFNnikLAOrWNcvX85Vo73wrX3QSeE92kqbWIDCTYbhf1rLrYZD97oqLbbJlZ2bsyfDdrc1T5Yv4XtDb9/smzvDjsXG1E/reKQ+Yl1Z4Mto68WQ55M3l1takgWxA9rTpMwvUbIopdV29Ve8gA6PNHd6/H5arNNOR46q5QDKO4hqovNCxuVxFFcBPbnAYghmwqSXJx204pSpf/StSrPvv9HyZEFQ7TAJrHCZgKVDJV2XQKjQc8IjWkTtmx0Id2HegeeDNF69O/3ZRXwn/qubWPr88bG6zE8U9wYuryipkUhpVo8BAuWJfsrENzCH08h9dvEFmNWZ7To8n5b21UjNkvug39YMTO3ey+KViOTYkPMasXsXtDe/XsG18ybFKBRUEhooJBNOFVp1W+e+MRVjpun1p6PXw2QQUT9AAQkauBVnBhzmzFmAL0WiWXc7nnKzYKYkWIMzGdB7Daf5h63yV9iau8Pr+fkyq0uIwWM3tlpw02Ybgn9nhO2StvVCWCjCzgXU8hlrgFt/c28gzUPwH1sZn0D8tatG2RsqPhxWhu4lFh0j8srW/dRpHbcwCTRjpbFrlgpi5cbPMtD36P3a3BtTyH7o5rAwH1vz5SXE/rmlHn/cvjzgq7gRFdIglv5ZmsCELw4xBLwVeUwJFdaBI2IawBL6GW4t1L6k9RlXvX9+1dDTVot/Ab592rDqklgMlS9VVKMMwmug/gLs31lxQXoZfkLDNNT7xGizUecvnZqu4amrvXO6PK6yQdLsO43gmO06VEzoM0Qe1OzIArWrcvwF0haEfyrGpW1sqOFWm2/mgfMpWB8rxUjPAjv78DXV7B4PFJCcHV5au5sSEPI3IPI8gMjoVZmwWyxAklDM4QHu3HdWn/8h5jirbv73/r3QBbWDI5wfk9ozDRAMWvoMlf48pxtBm0nTssaVxyctHMa56JW0yIFH1t3WBrjowy2cTXz9KVvEPnMUdTHrIH0386gH9YCfMbNh98w2canfFkTUqxPaOTqsp3WjU4qReBXT3IArx41A0zXx2C0xmH5ctz4+NK+gOEXDcafF1dQfp/eMd02yIzgR/YaMVgJUQ9oxSzOHqE3Ob9a+fdRxcPj5/Pn4ZCyyf4wo8mZYvwN09jujI7UShcKpIHpD7kvqKhIzHq8qpbpD3r1cb8XRoN6IKi39ivLx66bwcFaLpe//2vkhXl8fzrwhcyM42onepiqsg/L1UhQ6jBmKGurtRLsw0iSesCmom2y+gVz/nc5rfwHhy6bzeyllqW6/v/HJvxa70CTGQr2TXR4iJU8bKsjCMw2iKh2F+Y0ap5BYy/cz0a7saPqkDfedTYrc5rqx9Kjnbkc/w2LiCiRSV6wD3FjSrgH5Z5AkHNkG/9ghSY6Nk43pVV90wgmhFFpedMTXNlYKzHFHz6ZVxxbpqLa7afn93j4N1N3CtplOFGgdg7Mo0nnPV2BYPMKzGa/iwiK+3hoPdqK6c7bmgSkfl534wpnhbxNVnump39DiUo5Sk/4mFh6ZWCSqshXEUsa+GehzrMQ1GlgEJOO6hbmLW5yoqqjzDCilheXr+4oOYvtRx9XGtbj//9wHcNOVr2tZn4tIhAzHMQ2tW64YRPq6rsNy27qSaGKi3AGHmk+RSlP7gafEOXmp++/N5/+uO5mbKH6d2YAXxnFQijgwaj611XFEBze3fQLu4dBPNLR2NK4uqXCk4y2c2f7gUV0c6svwOXghDbu7EvdzTCRZc42mp0IsZYAksZDs3Mr8bm2s2hW5Hud29CYKxjtoaeyBwz6ffY9WQbiOf1cdn9dVmXJRpHkoDGqm9tsrN2oTledUjU9YEzcDegt3v1l87CiwqERitSkiDNfGUXjlnvb0XXtj7tr/zy/bnb9KK4rAe6RqM7DzbgpV3U5HQKK1ZB+UcOaqvSgVzJbN7/I0Ae2gD05eUv95UP3h1N/z5n+0rp3xYPLg5dV1jPRAKTehuILku45va6SclKChWGWFi37JdWh6a8/N/TmGUDslSUJn0BB4oa/HHUxlX7d3wTfkKR0VOEXBqablK5QuYEL7c1zAhNJTFAjYhKL8zZda18PLrVYKr7iBDxrFENyk7cUDN6T2F1au1y7hqeChs5hRRTUUmXJKqyiFzlp0Ngq+fdCM41kYHQzaAEL1wXhpxZkuumj9RdRAPyHhHOT11OP5A9y8+iP5JfOKYA3luadC9bLyDHe9HdCIlUO8xj3CVQr5iqfVQmyiavBaoJEGzuMKmGaDFT44rSxBMzOtcjBI8o+IK/WBTt/bXJnSvg6o/aGlTXa8HyHmM4juSpWGGpYy/U24QWgrG9NtY695ROUWlJ3Uz85eG0rucU/o4Fu/gFT2Z/bZ8NU2i1qvMkha0bPalFq4/g6GyFR9qmQUemrJ1mzEQ7bgd2nOap0BVOnc2PVVUnivRVDLQR4Rn/viz4DM0e5wNvD7a4tqBD9NBqHeqDypg5zuoCbNSpw/avY2dowozlv7rB9Cpr/RJyS9LT2CqPGM8HWx8CJ3ElD3OvxSf4XJtf3BTfSWbSSym2jXsAuFtypoLJ7XI3AAXUISKIlOmNzrnbjkos8jr+JFgPTM/db3NCUtXC2W+Yn0GFVb6Hfy1IaymTAkFvle4CwvDES4RQyjXkprEIWP0kinkLdezhoBVMZxnLSD2uTqYC/Xi48jH9bbkybR4fevzVVfsuyG51zr1Y2aEBlERD3ktIiwoC/DlTBsRorGwYg6vxa3S2/dKt9Ak4MUQHuMlwNRh2eNbyZNp7piszlcdTANVuXCqLAODiqsgvlJkjHRVHNQJdY9V6M2tJYLqqyOhA5NlrgfoKbSoD/ibFE/xpNJvf5S8vrav5Za4ksiCWPZC/nLEGiVOaghB+06WsoSwYwEO2mMZohea8SqaHldTXFV5xx1OvIIHOSNJWVQy+Lc/y12A1g7veWU/2InwyURy4h3a5lr2JBT78Wz4E0JzHO+cCFcNShNmPQ7DgAL9xo2xXwSWHFI6p6O+g9jHaXEgn/c/11eiHVyT1D5zhYOOIKaNzB0S6sIysxuRpHBZEsbcugloCpjKILhoUOO4qiLsKiFWto/HQzn9mI7vaBUmQ4X7/tLqB9fNnTOFtivsIk5LPatEdEwj5gxZtYBQI42gy9tbPFleMV02eUzq6dHrGW+xVKkf0lGlSOKsHk/IHw/HlLVUXOXRxP5uPsM0lU4IpD6UGVeYcTHPEfKgtBMRwlJKDmR1yljcMDNk9fXLZxoIqOe3TwoFLxmLLh9du/nb13hMh/Qnfy12ly7nO/e8KFHtTp0aMrdMc1nshN4/wDGQcQwVtpCVdEhtSJpA92l1YMpvbWbrHSwjCzadmaWiyiGakNXjTfTHYzq6Iq4+LsqiSr+DG/LVpMT9F/N4VlrIsLEYSY3NZW8h0BrV3phblBurfBU7mAiqp5Cav2Ys3dFdfLVSrEuuQnU1H9hfSy2Lph7yrxW3jweCmfB/mloCmCOKUJiPMAY6tvxeSRbGYBtCW0GsOSmvemTUUZ4y01yGUjBJWHkcV7yPRW5nK6FzS2f7+fnn2kpUzwMrvr88fKwqjrWkAS9gqJVhnCb59wiqdZWVl3rKp5/xh2M22nxESOcx0pDZj/Y1/R57Z4mrj+s622v2cWCJnj0jhBVT2JVxLQr/kczMBrYQ1O0Tdj/Pa8TrznwdTzmjeSH3exwb4krGy1IexOM5WNRW/vgaP7+9L3Rrm3rIP1cUDGrCBYOb6YpZ0iiSQyDEZPtJBRmjwtKU7A01KAeV873AeHTvDqr3O3ppalI4xYx1PMTsbnO+Igpkoouet9bt3WJ62sELtsjsBFnB2y1bJS2xPUXyMMJaELGFL2/fEih2CVVPV+7VgqwX8zlmzJKj5Aqmg5s/zc9g/PyfEhdt+U2cV7yDfEqilEYKfN1UkUJHUeodYJUUlkLGA6ho2ZhFBHndV3G1/EtGXeY/fv0/Z1eTWzeSg73RAYSqC4ynE2OWgqR9CtII3V4JELTvhZBFLz0zsW/gPk6OOCr+V6n0nl5ed/ycLAmSRX78+JH4jS4Ce2AvSlVsI/Sp/auOFcOAFrN+RfnKNM8Pzwfptlslwid03uZQri98VsNciSjdPefNLTnNScJoj9LWvcH2KEHVJkehsaRaiH+jSt2WV6P/d8pvf9FT2L/yDgpmtQoxtMCJWdRkVk3nQAsV3Sq7QEm3AK+AxgbV2puYWFc57G46tlGMxiDJalRwL+aoWK9LBMYvk6/oLsAv94OVrtzI0QhJU5WpF1aZyK+0G0GqqomlGqEPWQ0B5wo3fu6u1Xj+om7GQ6ngvRnTxGJhpEIdf8YyAYvScYTf/rD7OCS+8/Kr7yAaCcxl9k4zxnEvIuzRoVYkr9MGSX/A1+nUq5zkZJ7/I8aKJGxoaFyQrnkvGoZggHUtFfbPgCEY7YV+hT+/GZ7MZ7IS92jdXjHZw97WWA8XvntBq8DHSHy9PZzKdazi3+ppTuKjPehVDUJVtRZUbCKXDCCwUKAUhb4Exgr4a1KLIrX2vbQ/+OUqujCn09Nj18zCC+ZeLsVhUSFbr93B3IYQmOtDG6+DZZ4/KExMyMvIyKcBQbF6YL+K5tr/8ZvRdPo4uSV0aY5j5EI5Wc3zXDgFC5VVu5p5BPmVXThtREOHGexyEvBBPpqXnw2hL4i0d8wD9d7aaRximsL/o5U6DED4m+YrOQF67J0v+BVdZWa1Xl3OTXTSEGGQofwtv3KChuqxg/p+WZUM6wPzO5CsHsnrHt5ETupQL3D8dZ7Q4vg1jPvf4zsoEWhxBtnH+VW/SvA9nHBVx2PMCbqer04uyQoX7o8wtt4aov9laCHAHjwujxDlTCfwtc7+MOI6eALpBRzpEzgMwx8GQwbPOvrVy/13sKLjgSTYayaCa5bYVzkvpTqhhp/dZhKh2gS2NWlduge2AIlV5Qj1jDgxoMVIquKuD0wVq/TUVB6yfBjlo/UVcbYLfnWpbpfDzHy+usqUColrLGMbTe3FnaQm0aeo3cVZYJ6pAnY3NP8LvBQYktqTuj9vDIcVFnw6egrFVh+MMzyX9Bku5Ss5iLeqoEdG9xfFeq3YVUcgExNvUoHepr77BqbZKoKe0C3TvCY44sBojkqAYn7+dttwZgeQj35J6/aPM93au/UVSvAxJro+af1ZFYBQYITCleGeTuXmigstUx3ZsVom+l9cMj10zXt7g+CLADEMDjNEjO1xR8U6tjVp4krr9jN90Wvv4GyY/6RpVR2XIzhf0eU7HUgcTOWsQgz3y+4iDOP5BUQ2aDRQQ+SqoJk9UNwBljcEMU6g168b7Sevr0q7S/ffQRpFrJXebKkyThoKZa+LuQfLZxAOSqqOxKy0E+Sb5/X13C5iAcCXZbgqSL4adayMs4hxr60wuZs6QeMxjcH3k1uN9/xqrswqZaW6Q1UmvLCsvWHOLsJdSM9FCH2vMZek3OWhaWPyusOIE4A9MFkBBqgjg5+Q5dmZyEhsIG/86s9cr6+IIX+9PMBZZz0Lm+l5LLKaxPIUzMw+HECXjQiFrC71NWRMHgPSko0HWAG8yztC9fZo80JWGMmvKCaHsRtGaQrHQ75SBOv58bqdeQtPqxyOKIFWKMOu2uLM+C8s2pBSodAcm7p+WHM21DWzP/EhlNE72gvG72CdDsZatpTKwy+rGT7eMzELg/X9uMKhNcDxOs8ZZoVI6KL8ITM57QtX3WDNuxUdx0doxkBWRxpxiHB6/IPvH7DUxZ0CzR2ioYb9PyjRw4ADwdyn4uc1xfpKd5fu1u0VifDpQbyiAJhObpQY2pcmzHz/oGnk7uSVqY2zHoUL8S4ot7jznKxq7ZQ9v31Ba6nTj5k7Eyjz8ljdPmODUxncKlVVJRiG5P0XGpv2KhLaZ+xZJ0dy0VCCGl8nggYkLBh2VaOVAsGfPngIQUH1ond5Mtsh/qxffXzyHn0Bv7rpVwQukArfSlPBDAulQrQ3F5nbA7jgZHYqgo4MsN8rrVye5AMl95AWCjB6EFCd/Ap7vzCWLXT0q09dyCn41dc7u5QKMBwHpz3zQnnMvFL5WdiPx72I9gDvNVfHgV4YaKwZAN9uFBI2TWmk8YsFJyLqaDr4DAWbTZqvdrf6KONXd/LV/KT7gdI352fRucEB4hCOJDiv9xnPsWnlSgTndXe5XI/oFPzBWXxHVD0Oxv3fRqFgj9whe6QMCQRz4lvTt1Tr/uW9xIF8+3oTtDJK7PNhPZdP5/YKs1P12aeGagjcU911vGbjHngCg1ZXIB8H1ShzG7H+rIMwhTo2lVd0/fSz2XxFlKJiDP7vZg2qQbgWnYr6m0UFoJfsCIm1Fg5t7HzrgYKhYdjF8fMnfD2d/VH0YQlKU4gA4MuZT23RVMk7+HF2Y/3Mr2aBGcxV5lRdfJG7ZS0Nmlex05K7lajU161ZjLgFhTbH7Tb8V17bqiOs5yylPwbgQMxi8qpwJ6mjvabX35N28GTm9eUcjKlYYFzQ0JyRtup8S64Lo5rOctT+QnyhbUzBfuUFtPtIsvRQC//T1OyjgRfCIGHnbxhr2g21aT/4yezaUgye9oOzRCEjoXNhlXIRvVDU9l/6oqhqw8PAxhRXF+c2jVDRBhjEe2icBQetefUBvYnHgGgknEjEuj0ULTaxW43qV7f2cb6cChAJg91IG2vd0NOUeREpaMLWW6aIHneTWqGE1rdVZ115vMVIKLjSkCxrIazQUbGO/pXAn1Au5JaayF5TxpN5e3t/YO5cyeqb0a0vENNWnUaIvGp6WEpOBbYCMdROxsyX0XWsoGriNcIEQoDjYPZrorEGO6YhYL30+u1GmnZj7VGofkUSKc/XeX25SkXhxGK/WihG7ny3B2J2y9Zilfr6suaXk7zOk3gtEhhTqBku7mjwELAJ9AKA+hvZKsOvuHc2pG3Dk/l6uvrNl26MXsycHONaLbggZztTYjbtxss7CPmdL9o8II/dkNYz//SyrAWTZKWheZOZuKwqZaoNfWpPVtGxXrN7qW9X+aKs1VTJ8c7j9Xg+iUfkNAWOj4tcclu4STDje4ZqnBkEesZCvWzgUuQhf4GB0CGujQyQ1324WX2SU21TismIsY6ziXO+KN7ZWE3NXiXEUNzSNVcQkJaG1Jg+4aQ1IBOq9w8e2EyqhVtMi5JBdrUQEB1rL/j6yD0yQXvDSbmwGeeKKWv//na4oXCNL1rxQKJSQfZ0cLPI+NTIOqrkwlIQdOTdCBkGXrMU7tZI1MGeFrTMHWb0aCCMww5SOlIdBYQ5KUQh9MCxYgRG471md8PfirpqP87kVZ9kGFG83bKIogBd+Qbw+ChRQbQYI6Z6CbFyB8iYuFbU2Ixar9Nvo+ep+63oI6faosEmaHCitSRfffIifVFP5kv5IokMbp5IU6dKLmwsQokBx2IJ2nyVmacRZt2GbpW5q1PTQONAL0IdNAis1UzgWViDeuFzFOMPCirIURP1gli5v+Zz54JfFesr2Wme5TDzAV5YFz6Z1MKSYKtFaELgI0pMa1LVldGpWTIlDhqrEgoGOo41Dv+ILqToS9ByKpRrBHgGwVRbNNw0HWcTJZ3tL2XdbGwFZ6NFm6l69HJneEXJBeoC0wGqo3qdOxvezX1sZoOLNgHFQAPrLHhMT4HGpByAnmFiXyxBo1dt5FOIMWR+hbOJ0o31lx8nyLGInxypjjRl5uqq5ZFgf0jrJHvi7LXOyzciqOdr6AUE0SpdmcSIxERFHQ19dSe98jZJiQC/kl9Fz9r+zPJV6YZCgd+uplJlnaoqCoyzppVKhZbOcNVmk4Spxhc00oLOmVFbVoIwuhUGHA1OsQC1qLq/UanjE7hRGGICe/1d17xw3/m5oN/+41CGyhkztNNanp6aA1N9K0K0R1V/uRWhx7cekKYIJDOkoqBSsKt+QFwWgd22gTnGhVphonoKraPGin412R7n/WzmVarbhWw1p3LQ2YbgKqeAetl4W1KltPRovDnYcnXEFUi7H/YgZGpDCKhAfB2nduiSw+1qAWpPyFEbNjmQ518zDbqS9s5LFoOzHI+gE7qFI/JEpRVWTMtOtWRKAo3Ir8vk9N5NjSYHYSCpN0HFOpjjiIBVJ9xrPyiw4HMEBgtP7P0QXSDf2qboVtP4l2AytDVRuoX98tvPkgyRwKHgUvYR7OVqGSQrWpMvlewo6Gid6o6cqkNrOd3y5jeQg6+x8DoyFcyuiMGL/Vhsl8G5sEoYIfawFtXe+fNThdVua/xWIm6cZPWMwScrlXvWaqlc71PguGG+o9N7gQ+uUFKiQp5CzSMuU61TP2OLz1EpVnn0bfL8SakAyWrClPWqt89YD7l0q/FneU9wtuyhZJdEiAuAGrflvQiayFsKX+1c3Ty6xlyzmWLyGi0OGqMO0jpRGzNEr8utpX5FpQJaLhprSvLVqV7f839+5uACQccsxDfn94AWQq8AXe8XnAkmz2AjIai4MbbN1zihgHs2sI8UsP4MUlzRBL4eaAWeYAXfnUXfNmm+Akth5EFNCrXVZvMVx2BRy+IfP1OyB8mxq2ZhNpVn7UuuQ1l+L01XRHU0PTOgC+68EXQncAzN5X0ineqxVc5izueIetLbbGApePzAQGQsyO3Ta86TKc5xNAYrhI5N0lq5uakMcpwSQ3uR31vy+6ZUgLYi6XgD4EtkVAOrx5FfocqsrNiQcFxAQlqgPrA4CER/mhjT23QgAZE3UYFlcVEUKnor3RLKagZe/9ZO8Khc3/fUNdMtLszqKq5KSySMH5P8Xv3AAXQC10lIPHjHAo5BpluwrYzWgWTlT2t1LhDIOpSi4He002b86vPv/KyljcF/2XFEpfcDqRPM124WBmJwo0uOdmasGGewBdeQ8uydZ7BJCY4DEUKZMdTYNVxgYRPKLlYKR6faTBXK2B4EIdsrfnZzHfPVhb0JQ9GeRVQg0eBTVR2i7/V9LuzoHEsQxV2bxikr+9Lbx0B7FCqWRS5J7Ezvl8Vu2qD0xbIKy8+NiimsrihHjdO25TGo5VWhZjAxSP0y1urCDj0q1y865TLjiD4l8EkjKMpDd1T9nSEuUBdYa7sM4wddMKVQ7CQEz/K5djfRYIitUyk6Td+jS+0/pu/fX3Pt6NI+zvN/fyoc+iQgH9YKuVzasopfsRJtm9btRlqcFJoIXHiorPIOjrHQoilVoDVxhnQmr7V6KFtrS3E9eP0obUVn0ij8/ppr3Rdxhn8mqgKQrGh8Cl5VHaUw/0/ZFeTGjSRBXfoBDfID1syszgSbDyCaIAZ7aoAgsA8gCFg+uQ1Y+oH8AsPP0EX/W1ZlRFZWsdhsecZjjY+JrGRmZERktLsRIGZIL5GwXGEaPO6axFSGva4XybwfYercQbHWmUxHtlWn9aDcY1CWYAmuwELVuZRa/m92fzxbnOFtq2f46924y8kNMz36fUjuLTbhfIsehk2VJJWyQymNALpe3fsJxEKQTSj2XC13yu4beLJwwu206iRGoMSwrfKR8qPzPLu/TfLq+vKS4Wxff75b/fdFNRLJbNPgfBn7hSb4Dq19QgO4x8Pet60EeNO6LezEXJY8iyQfv+B3SaWb9KLbDDQPvAhpgb87dp/uhxkv0NSr19C2r7UA7NvB4HuQHRemwfWlmyEAxyMvCDZ1PDZXwamQWsobkarSbQQFNgFY15J+NnLJoKrZSKwJGChaUSlQ+ulzcZqWnJr9T8/x7GyPc5h65efBA1eCYhlDssdlTDyIGtlz1U14fUP+dq49r1h8YsUcKpaSsf3vI7LpRHY/Vbq5BQTRdDy9+OvHDnQJU+fgK/fDnPQM2d3E4yNnHKwCZRcoW/nDim48qmFvTZV81INWquSiE3QVjttUu4nVqupU3E4EZ2/FNqdVrCqTR22cUL0ZnJlX/RQgGLTry+NbfrvESnvRDP9qqVeCM1xoBgbCFSN1WAtPjUIQF25WCwmyjYXiUezfAapgdyIpRdehMviHR02of3QnMZHzWVVmyXo98XVUJ1+xzC9ft5acmm1e8Q0+ZuZBhzMc+AHEbUple2S8rcwXcOChqfjIYg1xRNgzf8agAsItN+KwrJeGtN524jJkwlNufwMBenJklnRiuXI5Nc2zRO3538T+6mXbkxwuxxc0V2Ny6QYOFWrBYHWnw5oZWpiVfLXN9IjO2YgPqPv0ebE3h5vYbgjMYnpXlfklICCYvovzCh/A2X8BXZQkTi6vNFavKytWi7f7/gp5pWquy/qAmX+H9ai3y7A9jTc3lZFTolbdDYS2dsZRj3UVL6O5OhnxQ9ltUYZIVOgkjQIAgxbUzzb4CPr//jflt+fvLr2D8Phw4LGbnKnHQDAm2Hpk+nUp7lV9NOYn90yBABe8myp9FrjmQkEvSTAuIRaRxU2O1MhgoVlH/9kxRr6mS5laUivOK9XR53gyxEUDN1QvU17i27A89ZbMgc16DjQOYDti5ioOG+1zRE15iiQRTjvpfGB0wbzJ6+dqecJeS9rObuYqYmnYXUr5oPXos54THb2bndf7wR90S2PfPirGfslcHI69YpqVZ3YVpFzF/e57rWrjo85KTAkNdwiapDbVw1sStjQJoAyhgeq1AfVvb4mPjxYK1xzNg+IDmeOLXv/WvHrQW9aHNK1GHhEczQXBFWxVxV7sn/EqdJd/9MjIsQ1/UufNeq68xlO7RVfAahmQldJnBVNY2qy+R21Hg+XS6znpGbI85KUKfp7xAAAgAElEQVS2cyE/CsiXYjHmki4vuARf8eRsEpVcx3DgrbgPMvZZdSqIXh0JsB+xrIEj4a1peQKejnZdq5W06x7+7H2NcvHxqSXP0PZXr2qZmfPe+ecQbQX1QGVCjGnkNqWIKaE8XTHTCq4EZcO1sz0trJnq0RD2zvEHUP1zWNTLXGfVK27ca71SaB2D4NwJvDfLf3wjGueVw2RetzhFT//RvDK+oePKqWJUaugYYeuNdamQm7BgDxX1J3YR3p4Jqxvl+juxTdQwlHBFK8NkU6azzWQ2EbrdQifVzyFS+AFN6XO8m/B2fU+rvPr+bj+DuEtySQ+e6slTMmgzlk3iFwqJ/BKpojAEhd3PIM4bBLuTNvIlVBQ0T59VbQ0o/j07BRln0IrOk/RTvgudGSdXtZ5THvLa29DdPnuH9NTYFpqWvTHWOvZ4YFSuBFmo1YOvwOu7JSUprJ14OJVUSZhO5uPXinWO42l3xsqxTKsV2bJggaKWT2H+02adeeU/iD7Znq2v2oank8fbg9PxBbjxIV1IDGZ/Wg/Nmp0tVgIq5IKXwF2ToGQfWB7eyP9oxEjsFc63pPCTToFSz5FXAVDve2SWAAszuwb//zavwEPO3MdxN9bf9e73ZW3AIEt5crPHgLOvP4LksVuN4B1uvR60qlqeywUtrQ1Ts7jFwejEVahzt8HF1hVEh2Z9MqDCLB0C4yZBk2hNIVb02c7rcXzffpFDZopcEYwZgqiSvKu6pl9FKj2tgmfv/RcDK24kXJt+NudGTF7p18/DDDm8yjSiunnwgbPFSj98rFO9H3T8f1P86uklp8f5x+fVwwMElYcVGuo3qNTK6zX5OlrKEwrVjl1N63f85Fp1HQITrTweV46zgGACF7TMtlYT6I3Ybum7E8B40k4dSdZLRslg+PxvYsWavZcq+JWAMWqtk7SiQzg3XOehYx+qsOU67rMdTR/aKmgFwY3klukWTniAZa5mTRoqYjHc/XU2TPLmXFahyKN598F6/l884+T7K1+vzJn0gz2li30gL96gURiaZv36iqIyRzawla/2YRggViBZBT7MUc+NnGkCQ2y9zEghlNQB2pDuHzoXk14bBgmdBEvhBlOvcI8+27e7vLpQdpO5ZR3whWCFWdfrTXOtLpicbIr9K4H2Um57tC71LFSnM/XL5AyVK6yKvTqVk6LKnXSvxXEGk3P4Dsp4k34H38jZvqb91eNP9lfhhosV6jbYoTahUg2JTwzZxnoWvapu1ipTtZzZJfwpnC9aJ/eEjfEsNzY31IBhsMEfs64fXEg6F55uRqQmdqHyq0/z6vX17XXz/uCSV8avIsGOB+J8Bowh2SpuGEwTWqmMq9pqQCsz2YhCYuk31U21BNMDLSjpxt2GvCYoAgOtil+/GbVqlo9gb16d/H2f5tUveO/kuGqub1e5/OUSX3xrBvXg0za0bjIvkI71FmK/SyMoxhT0WtC2KvEQN2DV2b7CXt9g2JgqXRadFJuqGVtT+bGPUmuev/2JfWvzepzv7/oA4XGVwOyDMbbSS7qxRr6qg6/H7gusct9BQlaFJWa36l6slLQyh1jJDpC4umnWfZVCfk0c/XxeTSZO83oezGEypm9PUirMzaOaHNdqHJqBQ8lfqOgSU+xiobwoXELHtXRYHa9OdtC6eXHuhivMZEQ2Peu83ZfOaKikVOkPCFKI1BKrP2aP85bfD7rZ+YHGHgebVkMwmIOmWZVcyU0u2ZyCbCWTYHHP+WpJKX/HRijsZ7WG4RlTmQLp37/lctJ3gdeonI5u8sD6jAVEL/9MITw2VBor4qJ5j5Sf70L7Z4wusXf2ONppGbPgyg4ahyPqT+xOW5h/QcNFOIZHkcSS4gTz4ly3LprJXtdaXGmFXkHensLs+Cd6jmms0DM8XbPe0V8+cCP2cEhwK0qUxLyJi5smdooRuv8xTIL0irnTpaJVo3pYfoWGoeNKPm8cF76CEJdOFoBxvQLg9NmGKpNTUaxYr15y9+h/fIRjngaNgThiGMfA4hsyN79rQ7iiA+a+jtKl0rmsqA0MDmkdRptOtLlSz8vsLp4IDEhCIPGZEXCataozaKGsb8TqlQvCa+4e/dJf0Qos/QbS4VHJ2QPf3xBxs+vYU+AuEp/fR1QIVUGEwfg+A9+jMjDdMfcWYEBeTfYbKCQ09AfEilHW+81YAWfIcYr8HidclY8A0XAab0Q+DbzONUQCCTUXF7e0HbZVEfbxMi7j+PIpuIlj/WD6hKzIlDULb85iehheZAbUTgFh6jN5Ffor7ryumTf45QMg+zhaU+hmDIzHMA+uTk1Vyo7BeY2iuEW1snx/XD9vwzUS9xfKbDySj52JFomz3EIEHjvaK/RVs21BCVzNt94gRbxPWS9W1KsYOUZRb4KkWT6Aw4pI68fAKizl78Fh3OKU/HXxqODIbA4gwESg3SIN9WEQ5PK9nya7CMSAQ/pCb+ab7XrF/WB27/zl3W8DV/4LeiTP6nSjvBKIj5FCSn1G/e2xdbORj7Y24igHdWCb/fyxZycFZuq1TRcOjP5LTtq0FS9Tr1628PbHq88rKeyX6CvIS4Lir0pR87CyDdXDSexB9+u6R1/OYjYU0xdcahVornQbv9EnkFncdWYG9P3npIqICfTZ+cb7s/WK+8Fsf7XUq0BhSPxoiS/UlqO9tqowGvm7rieRPoRlBHVcbaCEtmgVyrx3QLDMAdcxahUCTyHklSno/e169Xqrb//xcUg97AcefWuCVLAOpzYGy2SX+QbH0YviE/fjnQG7ptQZxkPcReDdndo2Y0s4kV4cyP2BCyqdAVp2rLYmUGi3U8vOOBtYn9M7f7BdP2Tv6TZoE+SEy5CYVQjGoA/Rp1S111hVvqVCbhWtuH6p93rHJRdHm5TpAUyvVxXzZH4JqYoQaD/PfQRa7cUK+8GnLCZz/etjVH5aZOqhjtChZV+dbwkqXWAxN6RvzLiKLk3m+Umz0JLtfyYVpt30j4Mifgq0BfTqM5ZbuqyZ+2nz6WX6q18vW5q4p8fvbh5cW/1D+mZuTsX0tALdlWnYbww3lcar1cuKvqOq1MixjJJKvoDlyhN0CujeBBSmp4hZACtul3VZ0+8mVdRfvby9bfJkXF4JcHwwgWooAEdeYbyJLAulsVK7mON9RpiVGvBVeo4SvBjwh4xoK6sNpDEo6NeBMItQzfwKskD1O6H6GvXtr1u96NOT69svyZFYkohID60VvUpdQwHu+a3EXiNaxTgffNePxBXoUCH37+Cok3mHE132oAcEphBqu660+plkmFuZtURq/h3q1a8XPw/mvHcef3xcxozX+ICjbyLVVZC9iYnsLloFiaH7EAO814PcFHhMh7xqcZ/MgMXxLlDmv45DYAfh1txZNdJE/st0x+Nzv37b76C+wcxu4u8PaUNjmWCgZ6vRavINLPSU5ycVSr6qV8GlEAzazv9xVp9ewMhZTQQFN2QNCR0bTSh4C0IcQpZNNwO1/P72JxoHDf0qrleHMePH15gzSjXuySfBopoLV+SrmybjlelEsYEAduzgPm+914b7pe2OgnlSclVH3QgAz1kKFur6Xq36/TV5g6/bt8+url7FQl2/jTCcR61XCTetNucWg6Z57w2Wx6ItKdJ1D/AUbDy8TTa0lMHQOK5VE5BitO5EQvnxI2mPbVXf3/kGv/1JONuZm8XXpV6lzuyNbgQH8wlceeuIySMd/u9Rf5e6RBWyR6W8UI7M60tuZQZamFQJaIG9jooRREo1XDer+lcXqd8pzvByzdX2pV7Ftml094DPsT//3SQK8IImx4WxLNxC2W2y6QWgolVLudhYldKk/KJ50pc4qWzy/4xdQY6k2A5cjVTbFFxgavTVa5SZB0AgLoDqBggJzS5bX9X/BFUnQEXXBWYOkPf7yXOE7QcPyGqpq3tr+Rk7HI5Q8llPpj8pjk/8SLj6UfPqM2ik3NJY38/7HytJdtXVCTgDbREW1cpLEGWn7Bi4Kt05Zc7j01h6T8WM10ULfFAjrNedgaG9Yxo3jFSzDy3MpWoIJWtY4Vfv768JXbXbn/eobSfhUbRQMNssTZSMScu1oFjdPGkJW5oH5QxgnXARUUJavK7qdAvqVvLk7Al1oW841YSGobNmodsrUoMGLcqrjy39q9vPeyzLzkv5Vgn/V+Gmtdc4r07FEw9wrVmfzVeUq5xCedfWKl9U9iYeBhvlZLsI9UpxPO6sRiZWSK1xsLx6n/fOyTuv2+ujv0pIQnMUVDfdpSmJjs1iZP3E4ZvdfQu6LgADbdAp/zyv5JPOk7Y27Uzmq0fM+hpoFQB2XTYfvMHwe36Ev18+o2PL20Ze+QfYOv3scwQwtGev2MS1jVdBKZ5aNsNfsRIDvFPtgWMBQoUhWqWEeaniyLVp7ehBGAEbx0Dbm2rC78dXcBznv/9e4VdpXPSPWI/P9LNVlD2+vMkowrDWwtw1DgwL6VKuwE3Nw75/UFpIbE5VwKNDc+V3gMobkukPV8zdfrDGUNRHhMzeoNOtXeMM7/+5xw27mlm/qUH6SrsQ/fr5pF7Wex66mcHsEFsw33hjWQXdIcmrak/ysqv9zrRGSgFj6D0RdCer+Pc459WjXPXfX7EP7y2hxTrn1QK3CsjxW6vyVgshlKKwuRlc9me0e0tzIrlQWfxkrsLibxrmv3xDw1i38hTQIctReerMq/6gCx3t+c1DzujyCjcmt5S2oXwH4/WpVzq+nmN9OcrqnPUz+ITjtwyBWODoWsIlVvA2re2aeYXB4H7SuGiaXOSAkt1/lFfy/EaN2jiEoH1/HXphB7w9tkqnzuOVi4goVE6rQphpqqxzfB8hVkAhp4oqElYNHz84TAkmc0mI7tWm98xS1Rj/DKstLuGP2vZRHt8QQjW4nmHXY/2+9KlUxxvyjqP1KS5JzJ1SIlXsN1alsWlLL5QN7CoyrE7db1HKKtpwkS6LC+bG9hG7tWqOjaaV/G/4++UzPsh5TfOvvIi9M17ECfjaeNgOKuG3+AQ7rRA2qEJ8PCShQBM0jcs6JRYDLpqSZ93Q3KvYwvZOef0GLa3C13AYxt9f1l8JzpDQZ5jxdmMRtU6YnaSY6AnGjm9P4gvKizmdqjLaM8tukAvmlP9Ip54/TKyusftJrpZ7u4vYR4uHHs2CTIPyBB/BYl79glHcLTUPuu+gFCpcNbeq77FwnHLacs+y03J14hIK0YVUPojIVWCozcBCldYSUO31uLfSW62GGOj+++MXkF/BULHmX7+/FhbrybxivYKn7ts1FuNr/YSTyfvzQuNPiAqUmXfZqLhoZsm6UKSpXB9xNY7g4RTR/KgsMeu4XT54hoIqDAO/gBKpx3fwZclVS+p+3KMTcLc+RRuaWt9kqlVxbAyUQS1NddL0OqLkriavPS878Q2MtDxY12sH7UWHSFtN1Ui0SluFYcCX8PsrujFJ6TO83vQ7CA2GuFpFU3NBiXG7UnpmIzjXqqKyM/kyd7pWuVOKKVNk/4aMGEPZiYFiqaUHpzvFfXAo6DCwr0KcxmmIvoO/tnTV/HcQoSLjSn0XV6rQ7p4kTXlc392A6RjqVhVcq0uTE6jrDZWYrtG8EmChVuIshZmazvKq2U0r7GzQJoSghff3CNVAnMHp1v5I4aIqcIUFDoxiAYi6FSp1wKhdeNpnxhQODS3Vk6R0pFC1wsPhaRUbMTdefr2pl9KEXpdiv10fMfqFgo5/DdI2THOk4nolBjk/Un4Td4/HQGguFKqFsEfhodBMQasnkJjidKIbc2a9lYyCl1IPKLc4aZ2pZMvSxg6P9BhX75d3MHX5DKJRH5Bdj6hN83fwJd55zdXqtpFXdEd441ElebTX+FLw7GyUtjlEi208jt+QV5cIDA3+I3mYceoto4guUqfoGm2nPMC3Q0SzFziO+BSO+jPn1eDz6n3zfvAW6lXLlsH5uCxtF5WeffZKfMVzKPsMrJ9OplZ4Upvvst72q4YuWl17IX8ECttT3gd2R4yF8AIlrQb06wjV/NcSF/2R2OPcrL8yulXbWl5ptLLCLpWsaU/YTWVxtcrm+UY2grMMQxn5fEMqVBfOeVqfsLHTeLRXvV7kUuyk21ktYw4UuJg91SQNwzC/wcn6dpAgkzv6kFdXGwbNGOF6va6Qq4J1nTPgYVrNq2a4TJUn1yxAqSlXFKZMMYdU+iTihCKTIFBx1FtJ+8niLnnF/JKQTdozeI2UZL0CxEf/xXY12hiRL/OfwOOZubT770p47M4bvQbIl5t/YILD3vGQkmpfKrvnWKDNPmg14g+7hPAAp0E+g+EZ2hvcvpsIHEgJ1pu4ImjTbjifLW8yd6q7a7ioQbxgH0+mVbgRpBFlWW4aunWghULPv6m5NwVlqLPXd4BTYfyTvEJW9VqqkFeLniGpFYZ6RWJMEMVsqdx7XSsdq5FSGG7SKEO2UmCgvjhyqjwxTOLtludLHkMT4cfenKUhYAVggW9x5/vX94hRrxjM/DOhZE1RvZpX9B9bHMhHvRKEXfKKntaybG4XHQOVC61nf2bPPEvKFRD3wP2IbCSqhUZavnTWUCFxdWcRzlCncHHTHU2ClleD+wnD4Cgv8BEunXE+Vb99CxclbAX53hXvn2CM+XOB9p8dk/5hSOLu5EuSHXlAUq3VBLpa+3Vho9W9LFDZTXXdIbo3AKvSis5OYQrfvlFCFf7hZhzY8Kb6duIMIbfOEDuWT2DkZlaoLHt2iLFnlliXuaBXYctVlR5ggKjcJWVFqU4RjXVWbrbB8ga3lJvhGnvXKbBZly5hkJou0XoU+GjGEf321O3Sz3uo57a9OStD9LwxM/NCIgVdFWswlPvAU2SIpzPzGl1QHQFQrmwViFsI1qv+MLGAKWi82Kz3HAblFU7fLw7q+0jzr26vgiGbpSA+f9cIjsl0ECwccvWUM2VpyvVe2or9VVlvLLr4BJtGhaysbafk8x5pYYxDZRHiN3CSnv0Rr3Fdr5KaAz/vLZdd8gAj6bTlRYn7Cm4GK8MywlQLARyfdGYW+tAlLOW3GUS1KV82zvdAZkDLq1RiGWVBgjW6qWaSSOExhmDFebWxH/wR8HZoMLxdIyG+mEOEN1hkamV29BmUa0ppFy4CXJXSXV14TLllQAKNbMyDTh3GyX3hMnB3uQz4hW0CvoASpTlMw/zrEarJ+nYC7um9Mw7BJaveVIzBgXyQFvCSMYcsWrzBXIWzMxp3lpRp4pFutQKtuI7gbKM8D3A8nHROtw1YsbKPxGFsApwYrfAER80rSvymcIbbj7/uyjmGHB/ubpbFXU/ASfw/OpAoKd57cfqqVrJITkuf6NZmvaWGCB07UOqf9LsTc/jwDVyXaqXiE5S+IWSV5tVHOAbY5NY++iuWq/aswk0Lww1y2VHYz4fShbOgMQgMpdgHgm+FXqGiHWWiTuEgQs1HnH4O1GBUlLDZ69Z1xPE96OO/6KpC4zAhXN8vdjeBe+dbau9szrrq1bzQ14FNrJnpbrdX/iJArFpweyrqckIJrYK5Rk4Z6DKpVEFOaO1PTl3N2p2VmUf4R5iVR0miceQLxCN0efW5m1ev/7176Ep4/6uvoFzLEzsunrBJz8F4nPXS/EFzjRlHXHPLDQcgGCaBll03Th8GzCHpGA4ZfIMubNg4hIKOFtRS6/sl8iwOs/MtcefF02bdSbRrNrsz3KD7cHE4CYauHfD6KTc7ktIJ6pQx2bGJSGmcCDuVqNeziD24SvekA8h71ngKsoCqFf6WH63tn+/btwAzfnV1jCtczLeLxuq85P0/0S1k6iD4yMF5yRUdVNYpLQHnLBUZxodk6vR0uWkikG+HaOWKVc/xRrrQkFUDo2X91bZncfgOtm+KsrdrQVrReFQeUXZIO0ZHGlJpPr+BbyCP3iiZlhLMZpWK9PeC8EJtQo77YRJ2xzCAPMT98gRYT8s6iheCtdqlvid29Lf/3d+0YTCn2IjHl1mkQDneCFXmDcwEZghjziWclKh94KU0N5IFetyY9AL3gTjOBQTa7ZPRBs8aYquO/pPNlQDHk5SrxRvUe5y1r2W4m7hCm5YU0fOCdZydzybadHAikTl5HRGLEdVeIRCBbyV2CPnaiquxwZk2sDDeAnKMstVFGtkJln/IK+lAw9Z0CoDxiA5B4jS6UE1+54W+PeUbfm8dxEDacRtbKSmH4Ukie5lhMxi28henrsotM5Cry7Jl59pGznMV32uUj90dHUVYXrEDndsDfv9YsSROwzJW6guQ6tsD3u6sXGQvr9NgYQofwg9Fy5Adw6FQ9ahw2QUhsPACy21JHQbMriQo0UT7kZQ0oYuT1it27AKCjppZiFCUVm7G4T3Oa8qz+I4BB+zQaIEKW+tog7rnzSU1HYRj4dKenFwhBHVIuMrTwcIyIhJf7014YYcROmi8InLVhJ0N2gbJro16pcBomiej7H+qHbfXSOTKjJQCsz+1wAmr+iz+DM7XESXAYy+tk9M6N6/jNU6HlSnaUNoyN+Sw80b3kLc+9KvJJuwi2Lm7n3Fdrz5E92ODfzWLzLXqerMS2CEiegrw1RP70yonox1WG3WsXF+mbr6VPOskv2oFRDtPHmp2WDGaWg5amELvOaJTGEa2Vok3+OvX+6YXtuTVo28Pl5XnhbFuYYJ8dF/cX+AUyrYy7qzioDnWzCowV27Kn1hdBwWtUZXCfSA0cDvsFY5Sq4aJZctl05iKlQltp/PqUavOb+irrn4xKNK9MAkixPecBhjNuWJxK1kzs7HKUyIVGifeyPd6a4rmfYNsNZLj0WtfNXp2hwQHaMzq59vvcWbd2qRP3F93komuCaVVU25Cz66Mj/2WnZyYIAZWORfPCwfActladcDXzdsNsDF0F5p+/yJitJI1xFiM7eOHKRGnafo3qu0fm/6DP+8BjdFidV3xaA1m3yTSZgt8IYALubEXTk4LWiadanM3r6dJ9hHEDTOTa5OPJqsarVQjIFAZAUOvPkzL6r7uGTZ01R6xC/MgsavloUSmnZXNzE9Ihwa9Cirx2SmJLibq2ijaTly8sybUw1a9HgkuhMQT3dWoFAb0oMPktstIqriw/zv/WedVQrf20V8FoN0rQ1+jUZAbr2wnr9yZoG9Ec3Tudk2ZI7UuG+pDhI4bK1dm7KbScttT4IAuVPjYLFhoQYGzJ/PqH+Wq7fjR/3n/P2NXdOO4kgNjMKQEbh8O8y3YE4BgQQkMLgPBgH4HeNi9CPYiECBdAivPp3H5ndWsItmtbnk83p35JthsdrFYBQW188eOcIVwnU9nNVPKTCWauGUPz5uggtJEq6eXmnJ8XcFCiaxjaUQjwxbMl4dj6h7vQT6S+VaeZRaYC9ISflbGSvPqM6/7sdEYqPAYvwXJXzgb5fEbyoWCHIep4NV2urRdr4M1syeI9mpZ7Xe5jGzldC+LHQPgKhT126RkBbkEJ/esGQ/zSnUgP3P6V6ac9uEFfLVjII2hOZjfOEn208ZfgB6YlPaTwjFtfmxjVugdbfDAYO9uthxYInuM+lx2iJUMmWc8lQ1sTyK1JVWoV39sN7zk+bLNcVQ2JplINFQjgrRA9XKdBFzHU612wzrlEh77BfBCnZ3g+LxyVKvBI8b9we4kONiKLYxWrPbdpwvXM1hffxLux+dbqb9yR/AjMX+jh4QqVjQvmHxNeDg3VzShF2+ykX8wD3wK9iqrOjippluv+PFQGMiPWuHJhREmmpGG5lxVX5ft6/PKzehzfl7/k9egXITepKtqzgnv+MW4uRXkGLLQXlVAXAQ7cj6yJ1EFBVzLTsWTV6CxtAv+DXgjFOPP3pS2oJpX/43zqrCP85f0DGfI2H/srCqlsp9PzXeotK3MmQN43Ng2iVhstE6oycQqeoMYFJGRVbcOdR1yMeVFkpEb35Ntb2lRl6dztlvHNfj8ar3Stv3Hzt/5+R580H/4fa/BcKbW8enQeKrydoKCWtVQFzAOkXRZOTrtoIQPUx7i3hb1mg6GzCN3kiJsYeTDxoY1cZVawxkMxX2xvIJeX1bb8O0fDwxPbWmXWAzo7EAZDpdvKrenG3LKlPhgzCUcPlL4LvvOii431rDT4vR2mFRCXxgVNebzRoBiDAbnKY6XHr9wBNdsXu3vwb+1Z9j53oih2flUfVNYgBzHE4l8EbzQmqxAxhEPor207ZQRoPx2TKs+TwlFu2D89ck4VrPBxkljta7h33YE7R789VsNCDO6Hw8BRM/vu1CRcEz93kLTsDuX7eUEcZ3WdIhCZhXtSHrqZhuDAa7Cqu0/HOy7+VDJJQia/8yalT2DM6Kl9+Bv0BkKefV8D74rh+E9tkdQpJ1v5yMS0fa8MdfFwKUVclqnyjqtcT0S2V7TzQbGwKFNPxwtkUzKyb55NrYUKynvuT59axQkXJJby5Lcg1m8/fPtrwccGBPVCgh7UJj9tTlCRboV7Ck9heHCQfNlNxOk9B4VMFnXO1XpDRPUwuNmND0KRaxupDfKwRuzbegzPiGvli1crhdVTCbLZ3jQKi/i0lJI2/blX865aPh2InZ8CvpydQ0B346OSnu5JvPvjPQEhmjbpi+RsoU5xAOoQKj0V/t3jbSfsxSqLVyL5dUh1vfMqzMMzfaa45ZXJQZDtTeXr/UIdjS0lo1KRULr/fq3an/59Qh4T/bFTWZum0qxSoY2o6JV2dZqDb3Cdv62P31tlz2vH9m8+pf6xb7HjoJbw3By/giHW821wlbhYSMp1ol8L/R625Sc3fMOpHS9CsVALm0gieFgM+Lm84qAFR6B4z5S6ywdVTiFAY/ZovX1J+Jf5WcTW15BOSZel4f/4lndSV6FS+YRMF88uVegaFsVNAWUQDQ4Fu2gKxEvdClInk2g9Rl5NeIndwRXVK0QLJ7B39AKc9dgdA8+PngDphYlaNrDaPBUHQ8GtyyqauYVVOyJWymFqC1YeA5UrcdDEEx2qveWFgQ5uBnj0Snuv9K7Br3VuoaqvobP1w6Tye5NPI7avUQAACAASURBVN7f49WbM20qray/0gOjZJOKQisQs41uUNnbtFrh7A3keahmPTR0+uHV/kgA2HUij35K8gp0xym6BFfA63zfyDl0sVIBrAzO8OPtnw+6EL8nq4J6AIO+R/VqpUsEKyqx1W3NSLfmqmCXBxh6E4U2N0oN2XAwPBVBCtuIeHZUN+Ouz7teYQWogF49BGmRj+uvwJPJzSae/ZWsdO3GgtqEfstMSZaZtwNYKZPdBPkubd4aj4P4Tu0QelUQICiT7UINs3LkBayZSpOQmQVKUcfvlZm1/fn1xziQvwqamds9mGoLVBatBp5mR1TaSAWybclpB3+Wsuz1/sk8aHMFHZRh6NWKclAB6OxBnIhZTYqEupkNHoJJv7BqkGZpFFZ87vN9l1c/cm+c7R6MKFeA+UhgINJ3RBHdoiPHThF2cYHr1J0LgcpsdA2d+sf3FBqX4RYIakN54VuUPMgemnXTdJ53QChyarU3YECuQrAWV69UzGLvG77Vq51oBXSuSNE+0A51Q/ln09Cowk4rb78EPK7jMxjeyRhJDJ3bZ3b7XLfhaCK/KZyQY0U22lgexi+8A7d6JZ/kHvz1GzhDtmeQvIqWBSsTb2Kw9pW9SpUF6toJqMlhDEPBTSutzSiH9irwaMuBMrqB8uxQ1J+dnOyzTbjA858K6B5uv0UAK/RV4bPel68Yv3rL4QyfP/5+fESKx1VEJxKOdr63qvbcfwxwKsP4Yt/v3T7XQNjKyRVi5UaNEIqbXDdegW4eqJzsecpAMIs7gZZUvmdQz+K3jJZF6K9iwXGlPIIh+gpov0i/EN6BFWVj1BzIatZ1jxuDP9Rb085xoIrw9UWVtBE6OqOBVko0HncvQQnXjLyyMD2zavF5VfTUC/Uq4yahfrGgXDWvxxLQIQoHD9NT3oaF6engPJTUYqMfzA+voKgzcUF+UqKxQDEzK9U47ifx4RRu5xD5hHDdtzPoe9EDr0bBGZzAh16CegNWxxCDhOranlSYTyWhzaOyBhZz2U0GHbpAgePDYuXuQdeGBjYo1gMzSYVeYZ5x9JhX9yWk1eLeOL9+FvurrV456Ko5qz0JNI9fEdpr0Rs/ccSsdtYhTn7n1GeYkNghx9erejb0xYcDezcoNE26onSbQO3gfGvcw6C8AVeZn+onhOy+3n1elbXu5T2Y6NLqAgBA9teAzDM8lVCv4GbGmQR0Y3IQH+02WK+kQnFXl4unQ6lhGN0OF/VOwEfLXIPyCkQTam2oJFbUM5Twq89n3x75pjesVU3kUPItJe3aRFaNe9w6X8rLTjMNDRbFHcnju0XTm+zW90Q+mtveQlGfdv36yh50naOyHpqHOK+4P1jMq8Qutjor+R9MoqxoReW25aFyTOLxKRCurrATrOuc1D8HgpzJM0CRMfqtNOW6UXLWhOQmbpvm2B0SqSU6gVu5kpKV1qvPbF79+5H27BREoZXECxaDSMyZzGP4dspnR8fetnvKMdDQ2AxB3Sj7bGmfbkrfm0wijdS9ac8I1XnpqrBV1DCEUN3dGfyJt3M+r6JRl78Dz69pRC32muvaKNq1A2Q6Om50eRkUdRve2iyAC70J0ebDNU1EY/AKHClmNe+nNkuY1rBjAAozy+Fb7vg/wa/y2obP/ioaDFZen13kBUqlvYkdm9u2aZ0QA4bN17bbQ8eDs5S3gnW7RWq0Q4m9YPMIlX52qF5crBb+XoDq8epb8N8zUM9ydfe4aMF/8C307YnCTnVWTT4CMkfNFZj/qkq7KQu0EPjQI7hpaV8TK12sB+pTsAvSvb5aDWVlY9XboxzFNLO72mNWeOGsC1+CbNcXiZmr7cRkfmZ1Px67wSAE/78jhMIF1Paq6qGyf7ONba6Q7q1zcgK6rcs7kKrZYHr0BwLsk71xxlHIjnIPTpl2fUEjusZ1Xe6/RQpWhPXFlO2If/WIQhWq1FntXKrva9mT7mHziLpV26ms1r/5J4W0uvWxlEAhWpPjGussUGSH7OEcw1Yr6XvsEkJNX9C13+93n1eSWHv+1fMM/vORYqJOj2iLVPWNGG2H7nK6qPsiLsKtrarrnE5F70EGRdlVVeCWHXiNqj4rHHYMbW5SrKYsH9uCtRgOM6+o6c+QSXKl9Spf20O9cstKobtS3YoXy0oNmcd4Cko3eqIOJs9fHR1EtFb0sDYSA1fky+6d481p9Op4C1P5lOe4arXiHGJefWZtlT1kmM8rrqVmPapCfxUN5xt7NBfZtI05TwUxou0SBI2BC/MXBKvNQQy4AoHx9SqCgjXB7AGc3CwC5QqtwsRJRMKfXVmtFpndEFcI15/8hLy6+/6quGMCnCEadnFd99y8UvdoHe8DVqimhKL1SjzAE3IaN9+orQM9AVBjhiMfIHEesY5dXsyZU7gKpu5xBRR3yam7VKz7PTqDvwo8mYC3n5PlZmp8nL6zJwG0OFF5rI1mVZddb4y/oEoVN0yZi4KhHOC4mTwkT3Yk49Vya4nhPbn+QlEP3VV8BpNNryivPhzQfvbCtEf1qjLKFfX4YpnHGmbNBROXvlPBmIGivTfbEsyk1mTuipMNbiAkNzkNj5RlpX0V34B3ya27hC3qRf9jvgA/cnh7YhNkyjEvoJiqpt4xmVfeJI9oVR0jo4P2C4P3JvGO331Bo2LCWJ68GGpkizhTyuBbBC2W0r5GQ5vt+N0X5lUoWl/JLsDPrG5t1F+pMB9mEoc4Xxsyq63ZsjekszuluYtG6RqLhzqZVWN7aAvaH+kbC3FvhP8BVYeyA64VVCuMTu+hUrGrCn+ExPJ5FQrWZ14zU+sVvbqoNfctT6UAszeyhyO0Kxh6hql8BwPiOnEF0lbBrDZMgq8vaetMshyhroHzDe8agWGmdL4lN1803VoIGzNG/Fhtl/Yqo5HyGfIqNRc02f/m2AG1BsYg7DRn4CJ28hKqvIa2qs0N9jnSS5t0ldmk5AiFjtltG+VZxaix3IChauEWTPKqqEn+pvWqIiRqs/nTy7HgqYHYY6sjCR5ELqFec6KFnVkIctRFJGawKU5WAAXYAo8hCKEJe2F1NH+7A2VkI68b/SxJvQo+vG85rfvQX/nX4JnWnrIyf8hOCwBDILMHt9iTL+wt93U7j8kMGiqs6mIDFURaU8IsuUtx1kVwHWhodtdtwRdhmlGomFfr3R3DNK/ympmuXul0Hl6oLzVEa9Vb3eDj1q8q1cyr3SlEammzYGxH4xMN+Smzsz6ArNVs6hRxVq0gWwFgn/GeQa1a0VithXr1+ZnxAN3wK7dZ6TbgqhcWJRe/WBJvwHXxHnjKiultBZz8UFslKXDTRhDZpRF1E4lxprpqusEF1h4gUMGrcPWtPHppXv12uwAZDNneg04Ws/IbAC/MmlXy/9raXjMslNqcxmOnAuPmJ98Pfr7VH7D+uSEPtmPh/NleBPoqRApdwupa9jiv2IvmfC2ZVw1eN9KKVpJTTWldHlW+VVptXfu9yvragpqW57P3nKE6/iyGETnh7JEOw6Pp742khe6Y2QEElQjNMbTA9ur/jF3NiuvYEZ4wZJNZCXmXzWACJmQl2i8gbLQeuJvA3QqBdhNv2iEP0P0EItIbyN40NBcm9svFOvXVz5HqyH1/587yUOeo6qvv5xId2AxngE/R69aZcaRvB0NbzNmL1Y6BVs3Ym2Ylm6fJWr7EKcUOc9xWWep/xd46a0v5ln0qGjszd7KPjwuM9DbA1sFeoEc9tFXR7Zu/V+jbt67H7yd3DKBeUcR87kq4CiuZn65fXoZRR6OtYba6UTC0nLFoIay0/nJMUEuSj9mnQqyaRBnfLnGYkdkLI3ULF3RVqKRh0YjOcFEy+fX4V//WukJIArnyFV/JbaZmVNeC+h0UVz4n1rpiQoMYFzYm0TOVb4NY9C6isS89BHqWjxgUFKDViCEw/B7ds0IugLvz2nHfDo8PwthfiBoz766KWQQq4QwGkJHSElPaGA/FLTxKC3pkPmiFuvKuYWsbhi6yFad2YbmTHwzFmFErHNL8pZrVFTS8Oy83nOZBtsUUa8wvMa72GeLl2buQndOorgIqundQUUaPDfexgcvciml90xr/k469BGK642CwUEtdCKstqatL+r06r/gUcV0VsLnS2fmZWpBE4DkcwXJNkgjYwiGRUlnjBrKyS8zA1CymfmKd3UH2hp1N59BheF7uDXWPbx2BDGOyrjgXwOF+MM6AkwLvCkyi/JmmZOE0x+bQJEDdL6i0PDKrTBD7ZpaAV6lvYCehLWq70C0advlLpTbcLqBxnzVWybrydRPormxIQpqhlkdm9mXmUBjCqovs0w6LHAk4MRyNFRgv6BPUmM6YxsCulxaCDm41CrtKcQXhwuCc3Ft4N7jo+TXqRmM+A6ubjY9v/oX8KUIXZD/odVfO6uZYWRsUprBLXfmvlSp0G+6leM3cOSIuViZZfB0ozOgUldNfuX37Sd8rA16lrl9hKdqgfOy1t4LNle2u5knNtreqWR+vZnxVUtPVCSumbzA4z5jsuHoLlhVg9gug9bW3/W2tv9qG/irn5Fg9qSdEbZD3MkmMNW27bAVnobqaP6y8Y1p0MaN2cQc547s1IzO1DL2+VzMGH+HsGJ0BrtPiVOCFIV1XkC55uonTLqqrnF3Hi2e5gmaVShtUcRxPWWKKvQd1C+KgXSkxZi2ZMhbp9jCpaL2VIIPHuHz4+4KdxGX1vTqnNbxhxkFIAqd15c8AmemBKgInJtRVUYL6X4poXlH2jfVt0gAJerOaiv0qmsqX7Kpnr6zlIY/o5trTEfSFcbArrqG/2I7Bv4NXM+OcV7Thpr/Klfj4lemG1TclmLTlsZg2Enu1QdnM8RiRLElGpXGv906K+R6dWBYKeWFpu0BrLlVwDYLDhKcq9VbFPcP7ayJD4fF/pu9gDojhJZt+Me2qeKZD3dh9M402JSeU0DkdIspjzaiVROo2FWxQ6hUTFKgk+ul337IwopsxrgZBROWlMmR/LLkSVbXgFJ1efd/aTw7AeZGAXR89ZpChRAhjYDAoS3s+DO5Ld31zNP4e4P2vpOMhFRYB34/aQmvlO8npUZmlzQAYZkj1Vva9MpYDO2+PQ0Vl2GlPdqh4sYLn+ARbZZvY61Ebhv3iK6gOj4zxGb5VOmoDvVXfcq/QMSLj8K0GOTAADOG9Sh7Ubfpl6yrh3056nJxtDNVj50tp4GWWlZrThWl5Xy7kb8bEnhepFEwCZIHN0xwb2laYadZfvPVMYkZ4pNE8OPDOhutqXKkq+x08Y+/s5gJ8IlaJDR9XpOASapYHLCZYMExT8z7TLPAjbyQ28UbCEI6PVc1BZpWmLj6pK0qZ6hiKWTKtVCZ/6YU9RCX15K2ybzs97Un9IIvBNbRy1ZOIeFaP88l4MfH4DGbiIFqKZ9M8ML2qNGR+qqIjH1K1boLSSSazBH23CyUX2ipdy9NMM/KCOXlat3Ba+l6Bz7A9JfhXOUFXBdn/p+oqX9oS7Rnjo379AC7txsu4rhXiq9g0hs2aKs8PsxNnK6PT5YJqW4fuSOtA2Z4GtApktIs3N1+d3YTwGVxeX1HwxutFoOPnSYzCZ8cqteSeYV8uLa5kKaicdiAL61p52kU0pmPvJMG6nW+4aBTUXlRkESOGwSHViOp7dX6nZeou4fvBFNGXLFvTVqK2AMYw6epRWgdOSGBq2qKqgLCDoG0wBo698Vp2SbDWSbDTcpqNgpAIWpYVPoGXZL9+xQ2c/pK6en17W8nH+Xxhz9WXF7aO+WISajCHjmKIKVXiKIJw87xXHJdnDcGE8linJ0GDs3N8fOu5qrKZDr1WQqAdxrUP4C2UldbV2xtdQs/LYuLJ8Jv+uIEvaBhWLAZIWUmKcG4YqLc6lNiebjxdMzwruA+tVaiUcg5lHnsDyjHberSepRXxQmVx00f43ugU1k2q6mrfq3OaA/no23Ox50NQyYqZ9kbtGDKN3WA3e0TeLMw9FF4wNnPiXui7gbWxvY6x9ej71u6aB6axSxd6GQwUOszXW8uPoPkOnsnTaZfGGfLcpm98QU6SHTJyZ4+mm9L6sm88Rq25fkd+r9zxptWotzZ+2rkRXbIdRR0B5Bi0odV24YramnukTEe1c992jc3LmCSzyucLoDFzaTO2hi45eaqM1LoBh5FAQYw4Es1VR6nWS+dQTUbve0047ZZmoQOje6A6GrFNsqT0zbrPNSYupyjMgzDyhUHts0e9CNV1YM0ujzelXEHPPs1EpVfww6xNKELl9+sml6TDYl6UzeorDr4VljZAQ+lJTzztN/kM3kLbHvVXb68momo2D1qcPXumBp/cwHjbHNjHJjP2QN3CIaGYl4ygplJLWh+NQTJQC9Jj03F4i2u+AOMFqG64DR3Sq9OoaZjuocVFkxqT3d8/CyDIkOA8j2EkY77QMex5cM6OwEMpqysC2olyJdvmx+EcWSa4FmTW8kkZ52wJu1nKvnurIVGK4+h0V1dTX9eov4Jl5snz2Z7mQaJ88Lo5cQ0LNWjfGBYDmwuUxwM97vysl5F3YQUnBqAyys9uRAWXjibp0KebQK4++gqS8cLQq6scc9llf+PU1HT5bvhP+16dzwk/5MAXtdHNxTOaKEx8C/FuYgaDcQ/dLDC+uK4Ij6nSddXxBdQUQbCNxWIuNmIPT9ZFOvbwc7ykd6d6B+O6el/LH3z0DIb3mD1L2JVMs9KyruD8X8LmY+7dVEtdocXivA1olVzusSSeSsfueOqYXdf0DbxY24UxQYnRm4ev4e0WvVcEIZ88X7Xwtktc1/MEqmIyQ8mJ7xGCN3TCcUjHdSwFxySo0cNJ614+qUYgPtRVFw3Nozh/DaJRgu3CJXX9btwohM/gjapLc3hfz4m8id32b58aF5SvZOzmLJTIdCdhOB8UsVsCkdEzq9AvVGZ2rmp62glrT7iItjDkY25M5NXULuxVxxETzsVAouMl+VrZ8ppwhn/GOi8zPEcZVS8S1/WF+I0DHZghagcf+xIsIte1QgKbK7HErOuUqFlElcyJsYFTYvBhIfYRJhXjOOqqeUx16zcFQwMccwvT8/2/0WripFHYtq52H+LeFCA+VwpXzDUAKpkvOfxtuowHBKdvFvE3YFwxMQ0OA4xd1U5EM7vY251g2/qWvbI9HSOKx9oseMWzDvzqPX6vvP3g4+cHfQefcERz9nukgyrYjWEj5k1eXUmwvBEpiY5rxcEXn8GuNQhfv9AoDb1Q96JVM4F8K1+/q+BWt/Cv6/U+s7+alhO7RV2dvk/+TYV53PP1DWqeMZv2YJjHARHdkPX/UgFQ4djqKFiwdqfBls2zpWGAH2bvOqzqpMMyyoHxda+oGOC7TWUVYIbwxt/eZ/nOHmd7u/3rz4+6YpLoupV2lmlEiS67SAO3ZMZIAmpdc64nbeUxAKZF4KCGCszOKF8/0zWPYj87s0mTqXlIDszyMQxX8Mcs8sX1Ntyefv2UusqT+2aprALGjxpGXDLEUGIvuAyY1zY0DIQNp3T51xCWAqJtZmbMIkNw1P0priHW8Ws8D54Ap7Ka/vM6XcHrH7PIl9126+ydd1MzymKJfG3ZFULNNoEVg+xmYbRT+oZno82p6bVgMRXnI1RNMoO4U/SqNWnyyw4U58QHxYYeuH9Dch/ITRbAvnscG346uZl604dwonxgxlnpGTaxozbnCh6xmSc7lMMsNZbbKo4pmZr2Iye5VJVvxtcqzGcsoQGzd0vkSrnsMjL7AN+NywrTjdSVfAbfFvBVVFfb79/CV3DFYiA3lhVlmJ+NqT1z+Urj9xiFbnAIHCCGxgB8dZKhzRlKmowwY4cOJl6jj3xVR25D/W3glX/dpK6uH/HLPulS3bo6/eOT01Dzr8TsgkkbVPO0b95zZGVZeio4NmnnW1hX3FNVLjOm5e+gGFy1PpfdQAz4Bo5jvDtNoqFX3k3cwjT4x3sEiwZIxtnjPJquD33Xi1X4Knz8kK4kVCL2QuH+ahNLldRHtIbPVaOEK29q7oyFr/p79IvD4qBFRdl70LNHf3HDLSjNNrepqKiurujagyr1HLszzOrq9P0bX8BilceQFeEziCCqEEJFpMd9ifypjRePoPE3opNoUgLwlj0YDBjDS9SYmzYaUfMoRk1D6lG3LzqO6iLvlXRXQtn2MosfR7Xb/fqD1l2r3v/gEGFu5nDPjT5W5SJ5Q3aDZt/cwEYU2+bKjX4znCvJ050pb0aJmSKMfRBaTBKKQUHxJ3D6V2hEr3IF+bmyZRX1oo8n/5dv+Arm+VpAglE4l+pOS4np4fPnUj7gxmAS5hteS9S+TMLGyXc26WZJIxrhwAB7f2IbD5el6C0Cr0JrhYO7ma9guITUNLheYdvt6+73D8DHz9PTJ7HEwQpwrOHczJsWQqWa2R6IR+B1RJ105GtVAtezy9xythGxriF7jKCwD+uP+xXDYLRHldAlm+48q6vHm//LZ/bEyHBi+2+YIYPZeaOsK5zO3kBXlSUyMHrMDJmGdQCu/hR8D/KNaU3wqdxC4XoojY8NBcYxiYVSMQHeI6z9cVQ3LStGkHdb19vw8ev0+q/7z89jduHlazMr5StI8pKEG0olbfuxalhSspr/LYZ8egGduEUYERk4Jr09jSfBK51W6BekrOhlP0dCgGVdnU6/3//0tLXaS3s1ndSBPdqxb97M3ituQyXeuoaoC6J5VzHfmLqSVVfbLcPRg+ZGbOut+wIxrcY1dCFcvSu1pnd52AnpI4WJgRmis5q+j6+v/7n/eLZEFc9HY3bF1tCb4yKvkiWocg3pDsISM+HE0NHQHAlQJXlqKdRll7lBZuZxZXNzm5E+Hj/u9/+9v7/P9vPqNzDHGR5P/mNUfFzD+8dP048/T3/89vjzt5/8H3/5f3tXkOW2DUP3cwxucILeqWtuwBOE6g2ynPX0BpnL1cT/AEFZsmeapOmLzWRsWSLxwU8AAilZxt+LbbyOt9fx9vr68jJebfNS3l7fXka5vNn/N9t8G//Tg29uBuLrKa9Zxr58e//7pHx7v1fCAdMKsl11Ply/KmNWPSr88f5w5XVlaj6q79CuxrznQpaXS/3tdy1ftj+/bF//2r7i7fJxPe6ZqKYF5Cu7QsQaRI17Sx+02Ncs40Gs53k77Go85b0/Ll/hgyr5NHhlV2BrnDLb1h+TqvQEyHy36I4rOxWi3qVBf1QXNKeyR7dnD9zlVyNzr2BqVP+dw/ttquLJomd5ex0PJq/KkDW+5tQeMGK1RrNan49y7YNFxZ1wrM1vD0dW97UYv+ujnseri2khHe30wvG934dywHErUdNpVnIar8ZxJqPN/LBvj2NanUXn6vENH6zmgpJyLJhWx6Pfjfax7f9bx/uW91rltGebH+DVDbXwDw1M/pTjgLGn7xCmvM7UpuOvxVsz5P4Z97OvonYP7TYVPPdBpA2aE9Lm9HT7Hn5fuuN9WehJuxOdkDErNqiWSJxyXAZFTLQVfWgEhMbSfbu7KLvUjiWW7nHlsECAWsbuN8jISs2OK/GVGXPCxqw0VNkUupkq6mw0njO9lhdquxTuCRP1tj11mFJ1m73ecnOKvlDvde2D7821bXOLhlnMte7GZovArjsXvLIrmV7oedZMtrDRZthPPXVVF3Z09mUOILTGoUkre9IzhS6FwHjMEtXIIH6i3xI6JiqrQvOjw6Zugajmawd49tx5vBJcqvcca9hiN74Q59XxJvAwWbNadKPxNLK36/mGsQuP8MpDxGV/x1uqlH1EXV4+0uIAhrXxFJ65zGo0nrQgRdnarpz2PVW13vZBGTlWGFYiS6fGiGW2JN3d0th7stbYTkmdzoCRenuxGOR9vbelg+pB1onraQMj52ql4bI91DFUBDQR2YtOEb0lAbEQw5ngLhE9iu12/blqBHij2sG1sweOhgO0Bu2xo68KoufeZW7FQMK8cDWAAGy8qXtGuEiPkWMLdfKhGLyBI6J52HpU67As9UenoZHqDFaKhavb8coufGmdlkUFQBiJ6ZCfuIlNcMDVQufaXjuHOTt1uBvPJN4T/PSf+gB0R+s0Zpo7DVDdg9iWwMSmrxN/jger+apCD/eLFQZZMtEju/KlGdFcYE7T1gK5+b4YFQSdTlXpBjzaVbNOLb0Grdjyn1zp64BNOfNDyzJCBK0zAwWp7EyLA23pLLJQKbd9sCJkXSLWniw9EqppSLp+pLQsZH5wMvsRQM/v+84157+vvPedxu1cwR1kHee2fb5wmDO4H44AL/qzSjtmfc9Ou9mrdtLqrrhb0Da3EdwLcze2w7bsnoeqj1bs3v/dstXNeGW3F4mdDeUn2tb/sIilCmZS8hGuKq9/jbsbZHwnrKrWnz+ev5gk6yit6mAqeGpXcEP7sRNb+xOcF8UuS9ufvUGoZW3KY4Iczl4qNMCFI6Pd9aAU1qUAICFaYIRcpgv4CH6A+YEFv57hj7ESUlbkKK6f+yCy+3E28NOCiUKXhpYWApGFCXDsGCtK5S28qM2Bm+dWUe86NB7NBVmNH3SZ4kZXj/Blh68B5lmSJvyqZ/jijNoPfI7Zzcftipd0Clbn7W4Rnh8qL08L8YrMoar22wxqu2TQLLKYkj2jbBygbUwrKRWq4s8NB50BrsgRviZ8qFO5C5srvt7DR/NaTyg548rYpWowTQV4MXsdAuGlVfCJtJqm5rOhrw0cV/LHT10oBmEM4sWcLIwOVceN5CZquH7BT7oqPtZT/LrglwVfr/DNlE7xC+RYMv4JrqJBJBs2tGacZqQYsYKhBis4zOtEIt62YsJur6VQI0rTwiY0Yi3ktzBAFbeqY3yQk/BrwpdyjV/P8Qtfz83qzK7E7waBWtHvSg5AHxADpkYdjPPCQ3Uf8j4Jl/dnW5dFnKhbovEOHwT9IHwmAKn/H45XThcTeYyLUAVqZJ7KbKRK8bTEwREmhQcYM8O5OfhsV7CyBi6Kc+TyyiG+t3V84WSXGdIn8dFLnAJr+RxX0QY5rMmssYm+DiAM7wAAAQBJREFUec4Wc3Lx2qglnAX4uRX95fRBvB1sQ+hjMWuVIMDFrfhpj+OLTG3lM/hB1ylRd7gS16JOtokO98QoCkKBYNaNCZJTWOoMvG7Z6Hzl8pBMJcsc1cgG3a6O8COuVie5BH5U/iC+h5xb5bZdrQumMUg1nylLWa02BlqcYz+IoDw7zzjq419XW02YJcZlwc/rS6SuBn75JP5a9bu4WiZA+9njPHjDhndhM7/VfUCVo1hwF39f+bP498oHuapzsM7mRMti4Wmv76gmB5DRn0P8eoZf/w3+D/HB7ypSfm35Qfj/CVe/SXly9eTqydWTqydXT66eXD3Lk6snV7+Mq38AvvCKQsMZ9jkAAAAASUVORK5CYII="></image><image xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAsMAAAMGBAMAAADl8JTCAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAbUExURQABIAABIAABIAABIA0WPhYgSQECISARQwoMMPPhF08AAAAEdFJOU4636dRwBhFvAAAgAElEQVR42uxbbZLrqhG92UFS2UFS9RaQbAEKLUAq/38/VCxAqvICVFyWHfqc7gbNeO6X7XnJi/FYQtB004dDCyHPly+v9Pz0t1d6YvqrIPzvV3pm+vLlL3//xys9Mf2zQfyv337La1n5zeuKb5G/wuLsdVaPc6GkCfIjB2mCb8YJud+ZQX4tmi8mypxJuJCL6tmUjK1Xa1n6BSs/xT6BeO+/A7auAvH6Sk9Mv78gfnp6QfyC+AXxK70gfkH8gviVXhC/IEbK46n4xan4Zit9xHQxv/ASbdwryg1999kf0tn+fwPE9iS5+qO59TufOo1nTn+EZzs6k9XZPPgmD7C5I1xyRy0rcCVbowfYL286kMf2fyjEvRudbSwrWlpGltB3Q2LthOu0zCdOl3U9u/2W2a7hPvtOeA5DUYmSf5jRD4fY+aA4FO18sbKxf+x1yadJXpxJug/VN6J066n0WavmbL/KCVcGLO6yn3vA6GNd3gahT2ZxZ8mabR+quA9ZO8gNMaUStrIGnHLfzSsWHIhAzqyl/6WcMHbCFZ3N5SH2i1HFAx/H/geJ/GiIswVP9mF1L4rjAWiKwaRjMmym5ryOHkLYI68Bgc1LC8DZ0cvZxrXbvtO+T4o1+z5tMcafbxOfATGczta/wj1Vbr8qgWzekor0NhtKRW9lhdNcibauuv0r8nkgXNYLbTZM7GK7vev99tnafbF7IMz/IbG4ZGexTemiPbeN7eJb5UVHo1ih7qWzrTbK+bT5zgmqm+h9t7xrFvnsAbyUe+275GqjVYpNIl+2fBrEjL86F40k2R1lx+0tQ/fIXzMUE6cblc7k1cjmGu2dir+06KQzLN3k0IOft2+rQsaZooHajfzI3e6hEBed0v1ljb3icT4Mpe08wN/hcjLmPI6OXlQjbB+8EdHiWPhLJYPxl+znXPNIcNWUO4u1yadAnP1eV05vxzpcdZilt9Nt9vXKHgwISH0Hr0YKS1VDxy/ZX/t8Weup3vw5LSU/h8VDZJRsPbnbKmTqZXF8Lae5af2vo6/u2A0URjGf5nXEx3AoA+t/zn6p7+xWcLjYjbS/J/7WCvlhEHPlY+EuWw/b3KqVUNdc9V4E7NwHA8fL7NgObAspufJsOUGg+osVt1ajcii51359Mwo+grrE+RQW6+1mYM/Q36ocI1Cal0v+ASHIZR4lA3eqtq2Zhe50oZ7Ks6AOzZnxmnahBTH0F+yTtHrJGekSZ36vvC8/F+LSn7ewxoLTzh3tvQBEVMBsAlJHDyxblKhVASB0LITuajV02iDSoaA116wg/qx970f1/pTO+B6Tvrc+fiCLh+USYwScIsKStc7LRekEYgHoSZZmSku+OHM53S2vzaEYNWxUtEi5zmymol+xrw7oWFdK9UHhdPE13zMhzr7H6DeF0UtjEIroj2a9QBGorMoOG1PudCSZizXMymc0UrDA1ZyJhiKTf8U+DYHspLfFGgxAHQKFPXuXJ7LYnjn63aPqbacOcOm8V6+LkXvAsjghnf2lnvBwTBQSbVPMglJ2sKw3rZ+0r+NYbPoMCi146YAjNGbb13teoBiWSbm7WeqPpneS+VRYamfot1qdNJQfFPxYYOyAmh606v0324bnR+uKR0BcjMOcxMWIUv9kaYhQJL+uQortV+Wnrij6Y1z5Gfb+z4OuC5o8vO16PMSFu7h6p6t/Rvp+GIt0eVNthzXfisaPCRTrsCVQ/58A1vWgbys/L1BwG4D3gjPG0yF/ddKMnHHEaZISVOB7mJBVHQeFeX24oNVMdlTVdRr0QIXkDjf1sf3BJsUOb6nZj0MFH0Ry/uiO9wCI9Sf2RdfxDu5EL0cUDLLD+j1N9BHjMGLMNlWbTK2y9lppgKYigBpHHZj4wBDHenjjj+zTpo+CjRe44XAj1Xdw+9ot55vB+BGxWLd+Rv5aB5Vt4JvnLR2dm29rFYtgDDxMhyF1hFpDDVP7Hm8TJJWicgxK+O/aP1V6uWqqRx9QqBqWh0U3idZbwfgBLC72DsyfoNS5AG+bhy2rHQ8jENOIi0JFDEKXnULHhSCEk/ygcRzLQTIczNRv258G+ye4PaKoQwcCXx1BVh7ffry7G2K+etEtqE5h96i75/55Xs9EtcKBMMJg40LfwuHo9xR8LE1nsMPRTdbp+/YP0xdkVLr90N2op9k4gGxbnTf3Ne+FmHOj5L6UOBEJnZtCCAc+Ez5HQLel/JisimSDe4GoQkYKJ63yg/yhCScIjnaN4+F61d4Rvmt/olbkre+cAIpwH0PNgMy2PaWL4xsP0XezmL/00A0fSTav4ZegO7Hj3mdAYN7IcRoQJmqALQCFg0AozJM7ffh4UAT4sc1Emx1hNAjftk85s6/DpQPtCIfjFK2nHpL1CWR9Sizm7o8tJixAtB5+RfevIbVc+BpSCFuYNinZwoZjCl+Pjd/2CU1SBKQUsnItQ9RUtOJrU9AqNqn62hTIUWSDHltdCiiVS2mkdfji4lv2RXsIV7Tp0+dE9kkHKCjkurrz57xsv/l6KMT2z4/ZEK7VAqL0ZUOv0fkEnmyh+dNKmqutVDoMEBMOAuhGN67i2CZyqTWQmiugSq16E7kpHtQ5CfwHxrCN6dZUibSMFOC4trqrwE+yfmz/QBG7Qbg5AYmxhRObPgq+Lr5t2zTnW7+PfcCirRR/dG7RiWNNjmna9CTOJS/ZAonN2oQPyiHSjteg0pamYM02lW7fyMJtFEx2aiapztV/ZJ/Hq4lsQacQ0zGeLarZekMjctYfj7wLFffHYqyK+62uSpi4kgbAgHgQN+3+pqXB0NySgyO+pmRjg/ZAR/4SQEnhjD21qcqNWrctOepA2mxpj0b71qgdYxjHqs3BjaPjpX6rDcHWMdM03PJu/Sj27kDhvzNQjBm3ZDInI8TcPiFFnGMrakflYXAgkoLZ5GcyKcWRbZJmtglO9i2mqP5vVNoOqcltAhZwnxvN20eUuexoP6ETM7uhaHpXwsD6dOL0ZIsTPiL5o3R5wtNdHnbYKp+YMMXmjdRNMSZAEWbxpMHXrsXveZZvK4nbJUYgNBOL5l+raCVJr5toEpGEVi01BdKaVORlq2yqW34T/VGGZwawok5QTk0lNL6xH6LEGlGbIrUljAKQ32zmjfFnwBoo29KNv894/xbvLoiz/eJU34ZJmJARRl/EBTgpeDXXG4lSAjjwMYFVDTcQT9xtgyJ+Ayfh2YXt5WomdMKmGSFgFq43xTuBxvC1URCFMo4CuOAedowIygNHVPph9pvSCAMXCRMYTGiV9uh6hCdxmJED0LoM50O9Pn6U/P5XK3ezeF1rX07I0vSKvjSA2Oek0AnZIh1uTkRxTjgj9JTCXYQuMrHh5Z6ijlDDar6IWBNuuVmkifzOwDAbJgHwIDagnmWCKZm/N3QuyKn9hnDr207duJRhSGTHhsnBCimFRgaj2W+/etM7qr8Y5+/KH7xoK/4iFCtizqjWrQtcBI0aJgIEUJ134AqkF7rbeEX0Ne2AQVgfBVZIhIa/uAw8Z0E9hiXOS4pEcBa9F72wNFylhvCiZkFwqlHtKF9aRvDGBIi4dSQgfhF4ASzC2TlWyANLPca18dvHj/sfoIu9dscj3Vfc52bGzZ6WqGgt7rQBzIolmgSl5kXrUKzKljjfULrEpZdFFb3gYl7e2ufFAvuBDbQskesyeDAJWugEkBGODCpz4Iy0ZeTk0RjvLf0fXR4Fsf5QUJ+dsZpApFT6BDrX4TGAiQAuJYvi7XKCThLbXgiKtrn0QaCmdmoz5qKtlj2qwp2GLyO+S6QCu9o5RMRvmd+kaOOFOCXxPoLJM+OTgWw72rUMGxUPDBTFEea+YQhcMsC7heHzsuyXJeF6j/vcUJBJjYtW1bIzIGIFSnaWJDngUoSpKkaISL20B4jEUJRxgHaVxnmB/A6D7JOol6Ik/Vt2WIlAFD0bUlii+jJHiyZkcQidycNOhSKcH7tos002bLBt8rCPOYfe7nNDh/wlZZaZYC4+7SWLmSvYNDhITMiSuotUbAIUxBaAxNmxgOkiuCx7JC1xPTcp0bwQY0gp+FG7EkUbsOdcihhNHTCO3MUjk8aihTfGoMu9fs8jyLzf5ceui3ULSG538oZGpo1SeAH1cLoIBHCpFezN7xlsWgQHAXhfREYckgYCDWUvGg6EgOBno/UCZJswMBQlFznJ0nrhJy07pf/D3dUmu43jwMsRZR+ALN3ApQOYVTmASuNjrwk0Pig5U7sr/pDkTCZ+fnl2DMEg0Gg0qly69gkq/ArtXm7/l2vS3Ftf62tc/hA1U/Id+eQ0Uxa5nqn9S5PFDuQlsLFWeF87/OiTHjLxGlv7gq/JYccB8/teaYKx2gfza4hS+E18v/M1QXsT309eM0Xl/94T7rBDiU34MhA7fjNQe6LmevIRb5Zg/y2FTdculHg+m7BO4vfsyuL1/DPtGeW1+PGpeXJ5s3HbD9R2R5y3+T1ieTNvKXbGIMFABSJGlsGfHZ3i8HGHWMwY5p+WEOMQqu3XJA4it8pfcDxlw+ERMYfdOFaXwvEAngXv+9pHzMwXCj8tHlk4xuvzfB9pr1LgwhWvNeHn8E2+0qVWCUBVr6B4Mv7M8vEjTUP4/BanlkLGorEzV17bWHH4uNOsmOFsRlLeHMNKVVPwaYdQCjvyZ5zkXbMZJxgSf6FOaitx4wnXRO6zuapfIvxtjgXts1PEjatdNapVf6hO0eQVz1rlpSd5SeKLTHI2ip2LHQBcAsHG6seGBjEddtv6OBwoABUvH8GIubSgbM6Jt29+zMaa3ALx24Vd30zjHmim8o+DPPtkz1DteeMPRScum1dSPyb9CLWgI35PfL1aOCowMs5WJOzs1mSJG/dLAjI/OFB4KF646HhLeVbl328GnOIftXuU7d6Zegqml1jjD/gl+226SX07enpvWXyjf8XJ45VEHBy8JC4sB2+ResnSocTZm5x3iwBBP1qkBzMKzthWsXALxUksrG+js0L9ixW37//nrfbf6K5V3Rn65627zJtLEm6kVpYAwUkj/pPfVslQM7LWeNr9mNfteXfExMwWnEPh8WjQzIRYtn9z/36r/72VRn2r7r6wq1xhbQqWJ8lAJGgwKGixGJGCvXho0iaD3gqy8U2K2DLmNv2Pf///fd36159nNybkE7hbFFCRP/zA0+rj1YeKg8fdqjyr9hLv9C1G35zPT+W6t7qLG+EuECTUfWR426K9fjjxPMTEOiqP2k56P0S5XNm+vXVJa6WMipPg10gw5BZyY+VdDfNisTAHigZjPhjBNAtPF7dvF4X5DiHJENxEWlNdnAAQNBIvjvWzgPFUvXi70y0jl5OaD1hqQW7MdIuGy89KmB+YUaza7/gW6o/E2EShe5nWq1KOwfoAAdeUCs/YsKpFMixQKNLGMFtrHAqyWO9mZAHm8BU6LVJPOzQvvY95NYGb4yaevXpuCUXr7D9aB+1+QaJolU0Ku1gel7J6MWLxD57xgONuFjhe8ol7WjgmyaUAz9cOGNtYD7wfxLYDJhbVKGvbNRzzkW961hVka1ZgW2rc45lMptjw2o5SVVZlqfxpVMqc7xWHf4AXZMee91Alb1uGH3fQsZq14/FI5DDm7WyrAULKDyk+Qn0nQzwWKLrz7lgBLUo7YuPG4qNKudz5llGIaINXSB4bLx6XF68CAwEsfjYeWr3raUcd7iYdEOmBoLskE3miDzIwL4YcEUOZXNxx73G6oXlJaxD08QJOYQSsxb14VF686hiNxgkmnt2rttMWiBoZb45bS0UrjxYpHKTYxYnjjaUPbPwnMY1junEYZtBNaF3FWGHBixc07zYgxaHqTplADMi/pfV8wzgBxhFsXOSkQ4OJKYXBi2fRAo42PlZ6vBAouK2UOFDcE6HQkAzDFmdBuhcLBXZet6yrY8edyKNJQvF4JGSONzMwOT2G20tFSViBF6RMivWjp92AvNi0DK3D39iCtUx3jcPCCvIgoY1+ZG0NagsE2GG9O5VIENorEx9vl01oe8kQNyOGeluJU7anBYphbKA1pMXMUxnafT5Pq4NgZ+s6l2IMcm30L+rF87qjZh7qepjQBw/RtGmW6WZxOJR2yIwV/mEw0xiwWt2pFw8roFUjQeBiIeXeLBgrz8rKZmMGsZ2T9fkjmtnT2g5lFIrIo63EM0g3sjA5LJHllCOzsMZiRAofOJ+39fORAno2LFMwCrpTceetOukiFaMFxbmplDderKrjg6gqPujxvYI8V0+3CsZIJfLOi8GCFSeW5l3nxetrWGPJ0mLz4setUHiuMQrqOSrOzfR5vpgWh+JuHROL23NgNvepsbjcJi/OSu0W7FIb+8rK9AlNwMWpzyjWcXgxIHnxYk7aplvYt7ezcKyyMOaDeQ2Q33vxmNJjfqlA+7ey4VH4e9TP4PvogYcJGp0fdNarwGzCVXFxvHEZxay9O/bipF6cS7mLjYvOZAIBChQgigPbOuIoApMWi9chsXhV6TChvr4laas3CBJkWVvRqWz+bS27kBWT8uQ/T5VWeY0DM1/qxTxyx7Pv0x18mCwfztkxecwRq7iABoqexS1dj1GDCKshbTLn0TKKm5x0OThxMdmATmbAJv1hYJvRBSI/j4OBpLxrM3dvtLXugEtoVowGhzFTHMM0L3ayyjKYmTkHL+bp5welOvWjjBdFiIsOTGO0o6DdnHfSIp0XP322cRgbKIiRcpc/Xz2jIOOsSVYB62afzu3EKkI+0c67yC9exx13MobQYjGrTt0BaCMY2xJhfNUp60ggpkSKyLsX23rkIaWHIm2ceLNe1aWZKuiDEppJXm/woCgbeuvE1n4W3W7s+egXng/x4s8igjV08QKaPCSb/JBLvrTp56SdfQnHXkGbF687MbxjeLEedy0vbnJsFw4UJrUi0jZk87hWd2zQCfSUsk/oPrFCa+DcnY/d8WkHzO/yMHxRfqCxqnIMwipe00sEGUQB3f5BHeiX0QYb0tb0n6bLEzNFqUniMPw3h8a+O3HHZ1NlRw0VY6q7Wbv8wIub+ud1OfLWkjOxBFI/dsumUHUAyMykiPzHQvFrk1McbixhyvybtzQBxXzlUw6xwoQRQ5OudBJ9IrmavP/87HmD42iDnhfzqEd6l4uOmGd35IBQWE+/kAlKinW1a+fl8xNMlaCbOY7CjequiVs1mdbrgZk+tIhkLQfeZag4ikdgUvljl7iyvBj953UcmLla7y615ihd1onR9BRwjbTdrP1QyYrNiWFj7yy5Num6vsapqsw2TdMyCtbYpumC5x1k8iylsKGvVPbnnII/5MqkUTdTFle+BoGZ62sOPAqoDF7OjZ3x4+KwyNeggefUFLGwuK7EYnLBwdAe3Yi/HsMonEfxzSgejapyxfMObSSbmiHKW0qKjTsrUiy2ds3zBavE1nUdqjY4O/n1Hxkyv5iFgfh4sxnKPz7hrAZOm0DBMdjOO+tAYwZ6DF6s26u+z7pwdfduFfT7mlkxRpHAKCYq5BpWTMAsgvkgk2jJGgVBR0fkP7u20tEC+hUp3JUrj+lSxxxqZjygUrqdprqmaZE8IV7skdjOu/mzqvrrqHEaF8wUHOhaHWh0OAzuEWjNTW3CKWBhioajJhQkiucmr+vyS+u4pM0Frv5ZRLb4et0NVUcukHCkkAr3sE+STFjKOqRsAZK3AV0E40GjjU1+aVZ56AfL5l8oSuQAYNp6AKf9dJjEpqyDB+cNziZb78aKJXxgYZ4evZymSs5uXah4uMJd2bmxysij29Edd7YVgbkqo5r8LMCk590f1lS5CphJRqsK+RqU1zbs1siayBaFhWdleo4uxM2SH6M60LwezNuj/Lm5UHFHFoKVkGL9fNp7sFQaIU5IVuGamR+HMuehez2i1mCjcLMu//nDcJz3sonb4uVc2TSSkjVDAV8CiXcvXpIYGUyVUUnbatM0Kg2U8mWOOnLRUWvmF4vEZefB2g8VY0tpR/1OhE8nXDMP82IOFW3j9fP7guVCeTHmQSHrmnMYHo8bmYKl1Y2zBOW02TqhNIqBsqSrVR5fG7etqW9ZkHGBky53liaoTZBtgkrYgUWKrRk2ARv3K8MWmVj6QPLjtQ4U1/XRRmXJX4I2SEWn53zwiygMLjpMHNBLBdckTqi5HaNY4q7iUdVdXK/Eu5WuQqMg11Qix4j7+YKYqwnuI1vvcgowMcX1NMZ9HUVVWU26xqgqbTXgdP5sgmyXRHZkLXf7quIqNvZaykZ1FXpKzh1FPi72eK2jqCoBongKNVO8uJ7bfR2dICWsYVdZaNFJNW2BAst/taSzwo5i/RymzMfN3bHwwmorrOgSkpmkQkpiaIA+2edCe2RNAYkcgAlNJnxFprAonoa0DYvFcR+CLKapZ5b88A6osn0gklusSUfZvdhyCe3tI2kjVB4Utqwvvp5GDrt5CAy0Sl4M+WLNKOjckVgk4LM5cw5ROEf+ZcgmJKXoLZx0+beRgbzJv23eHRKusTFz6UBXXj93heoumxTNRl5ik00oTmxpsc7Z5TAR9gx7PX6MehwFM3VAt23ObRvDT5wX5w5/xyRS3k/dNmMrrcrVfwDFh+GDFCKxD+j+WI15mKqisbhlFBFgOWWPA0vAbJhOcrjd9mNtOhPirkVi9+IuZwsc+bEwkFO4EYsfpx27y56vkSUWSoEn2ixHBroWSzzIp4RY7BjFEr14HqlfjEGE1WMxr2KtJw8URcmsON9KmHEm4wbikMMBJ6QJciw+lnbdjiUuPQZup2EykNq4DYU92gexnjRKaJ8DMHzRFYxRJMVdOAcesS1oDIlbDsZeFtuMyfNKGxWxw3mxjXqklE8LZmr1hhMvBxvHndIR/1FvVhzeUmP4bzzxnqapMlauf4Y6kHaWHg86Z9KWNw0lSdoK7dO1spsO1fwY7Q7AxV2Q0PpZGktNPmKgcI10PVadHm1S8vms6YQNxxTv5GsiEQBMhS4xfRsCgrTuYFtKES8GChSHPYZNLL3ChuKF0ZGT5sU5pJNZf2UHgpBHBBHBHDkTwsVEzzkbC4giLzNoiA1sj/KzfYxH8Uiy+/msgUJheMWGkVdo4ubJmoLv4tFwV8GFQoSgPUTR5hA+A2EgATtkOc3yT5vQnc6aF+cgCgYL284vhYgJ+YStxxX7ouWh6CUpxtbXd8ZUQc9j1FAYd/hXW+7aFiLI/vXzObFNImkObLlw4AVuqJeUtLFPHootSQ5QWys9tAH92StEH14Yb9Ojb9CBTnnUFVM90LFF2+ysQ3XZ4WFL1kw4BaFY53GprzwWoBS/+VYHe3cm/PpMp2bJkwXjUM79vlHIiA2lMO/dhQiPEx+jFw+bHp1fgfz6vb2Jt06c9LijIApmeKbXdx37RxlVNjiT0PgguG/uVYGeMhP2DJy218C8GBTuJDptp5umoeKjHErBdL62e27uCZg6IUoKTtgR9zcvdq3Bkaoqry4vBkv+pO1R6vUZ9Y4hbaUr6ALvRxWKs3qx1iJd/zlsM/8Pc1eb67YRAy+3jH0ALt4FCkMHsIEeQFB07Fq7HHJW8suz642lFAHS/nrYMhQ5nI81gftNNtA0ueVH+r1EmR+MNChB1UYVI5VDif2DUyhTJ9gOQWxWo8VZm7vSmQYKcyUdulfx7xFR5odd7kqPsMliA/yE4XMB273dmsjOqrhe+KW9Pi+HpfPZCMZVFNYNzJwmdhssbKBD9WKlnQN0eEfhZc2/tOqtlo1JnbCdwP4x0BjlG7TMdHYC9+zuS917cYEz9fp1OGsgjdeG1XMOXGJFs7JVLnBihyfUN47EoDxX8dIrCnresRfDz7H04nsrrjDQcR45cHg1VYcAalMN9GfrbodPXsATgIgVZ4/m+rGq4n7eQAPbDRYO93JZOtY4USe0HLHvdgwFI54oFL4x+wNX15SKxFfRc1O+0lRxJMYPXR2u/IXvc3EN0L0e7eqh2fnv93+SS2ZoZFv728UtCUMwZjTgx7ruFD5SFDLwpV+auWUsDb57nI4EtAmdmzOcBN1NkNM5xG0QEi14Dhl7fAfhE4lFCMtc7HqaeauneT8pLJSNUi5Lt4OVsUQYlTOqzFI74xxKbrnxO4hr1ox1M6/55Y71ucOln3HNNBC/uAzGR3pfbVgUOILC/wcKUUPiQyxDPEwb3Ez4nFyQi7OHbPDi4ZGj41sC3UsY1yyBCIfTNmJCI+MqKuO41olmBidIJmrbXfInFeG9jo/836DFbzu/urlujQq7HacTy+qxI8CHnxrkH782w5XYZKI2TbhmSaOItW0V5CW/Unu856pST/yVqXKSg1353SxQ3fwH3cFNn5mfEto6kAXNNYUcapJskbaxHO/Ipq2bWUItYvTic13kDwS0SYtSwIwmu61H1tZciXgTflCKY79KoPJA6Zk1WCncc28wE624pLte9SYHuUCDJRhcCTQPcsLMYE00ZOIQjNrc1nReDQGC8uqxFDJlFHdzuLIIXe/FpW8daWjTVRVL86FLmST7a852is1DK9FViJC5WT0SLkvFhXu49OK0FW+gocbTVOfXQxUx+ztjycvRjrOk5pbUkgTNYAkKJVvoHLcQvn6M8KMwHsVl6hxSPHFS2NdhhjZt/wXAj5vSaGvAX0cHmOZKGCwlGIZRb2hx+U0Vd/3cBb0YeXeHAoE0LvnGh3chOR/26/6cDbE09DK5kgZVrGBwb+biYMmbnKalA70fjOlVvPx0p68jFLAGLZ7+S8QkGT0lxM7iVYzVzltBxB4Ep00eoECWFz+4KqwPjwLmQNVt8GhZbJLDTEkaWzDJmdHhutup8ycEzEt7Xbc0l5TWzNdIYiM7iqHr0IYqTuP13ikWW9LbQbpEJlmHgZg+Cbc8FUYj2Hu0rnVYPCgAwTYQ5Rv/WKt47snMHMi2BokIR8EohGA2AMZqz2xX/RQwccqETSg0M05NoRuHrh7XxuI0tqYqHRfoy2WI7a5Gbd8OMrJl4Gqaw2mC1zl2bGsx53AAACAASURBVEzhYAX/FJjdcW5gK1Ra6+7mM109+vlRgG81L7e7enA8DD6BGzQ704QDUH1VRM2EiZWbIiiEB64XdeBYN+fR5YHHfyjctdd2V794rqe5ptsBljvx5VntuK8xsLWS51aL63elhjCIbx+Kl49K2swUo3NVulVxHKBnM9c9jMoc2ZbGbQWGKesAn4xhjede7wzqwLsyjJlSk4VQ+gR5cM+XrhPF0tlJ6lF78W3/9/X0UHCBHJmALFfpMqceKeErdIIFtMbHDfW7Po6OsAa69+JfxpPvZJYADfQEmXmSAzlcaWR9IdM58k9SWGMSSJz4i2f0KhF6Vdl+6VywVIC26ZHjx5tVbJaZVaC7QPJ7jxSmwqWAgwzsJ4UIIUvbgcP22XzuDGXDwcNfWRJJzVdXj3l2wPjS0Y9imdhgdF4Q+SNMxYCKPYDKjD6YnUJEwQCJXdqBKtaQ2aEDb3c7O0CfiakyTd0SERDbCFGY7pwDLcwUtDwJ8OEXP/hM/vBKx1CGMt3uTh0AElo4dDNOUND2XObYVZ942+h8npzCrfVydztIL8bWrBkpHVsNrstmUpvagSrG6CCBAj0AKMwy041ruh35zb7YXnhJMz/CeVRQxJ5IDhJbyut4VgLlJUwEo2dQ820rV9fETMaLqzy3C9I2EUoBh6t9z6MSWLz6AAES25q0rXSrSxGG4kIZM6iRxyudrNpEpEDXqbgrXgwrvDK07Tq1CWSMoticgcLXSUIeONRQ+ped9QUyxmi9gQY15mGNnKYObYDku2EUUcWpDm26e5cQogl6FqqPE40LP6lE/eIRC7Otd+oLtKb0uE+M8caFJN9TZh60wRrsocuZf/ezh4f65A0xxSXOWYgnKPTC2I3dQ7dZoNPj3SMIVyZtbFlXb3kDuYPxvRX/ruZAu56T3MNcjKodhrn1mYnIFiZhirszSb/4jX/4da4zm4EUfaNT4gJdbEnLWLyvnEa8iq05OF6sDLf5tuHiL1c6IzrJ9TIVptBvK5iGNkPaLj3ZQBGMCXPd/T53Gk3CcExoDrSxTGlwS40uXO901INbNI13Zt0Cxg1GMVx6mYiVsRgs+Sof1T3NdUPB6NJ9swkL1kSjc27bcRXqh8V2DMM8sD0YJ6pVAtEGu1NVApH/2ptupbR1gOaK8Doszcp7XQxroExoAw8/ok3IVkxTPne/zkR97Scz9ze2qLAdKdwNJ17dEFOCtO0TRWIQ0wyWvEPY/hGvqPLT9240D7G43bWfvLfD2Fx3V8DMHTuxhIUV/MsDBsocma1xrEvwuaO8jljs+NaxYai0XCAb2oa+VTzEelfSzEsa29feRUzCmQpM0HVOc2JHBPJ+jvVC03esqse9+IycMCfJXy69/Ivr3wdKp7nKnpEIEoR4cbWBE15188Gr4tuImanzsJMFnf+j+v24xmqaJsy83wV6Ah2o6u7uy91pRxzIDs6hJbeDqJvlsg2mcdUgcaZHJp9tlT/Nw9ElnEZh+3M/HsUFLtzlBrucR3eKHhWC1yQ3KDHJGPPWAQgWCBp0NaFh7dlf9YXP5HPeSxRG/OLlOnrdkzeozppArHPyDyCDEi50dlmHU39MRZNCm9T4u/5QxnMbTjN03O6Cb7VgmV87lLESe8KS6xqOVQ7H3CzBvVTMDKwf5/tcu8V989IjqCo1xWqyRMGp9+pRnV+rRHcXYMLDOnJsHcJBB+Buu0m8hj1NSs5p0yQPHZb0j01iDEZbtXPs1osnR9rORetR4u6+9vnORYaEUBCu3TnyillF6RysNFAya1RiFP+pVYxOuBrhJT/V3Nt+51HqxcvPumOauUQWueJjJ2vdTANPOPpD6T6sv5X087yGXlw+d3b16Lp6kNvgeDrt4XOuXsRgD1sV2zRBWufsFqMIOzAXNoBrDVtNn8GKK0RRiZl0f+6ZZn5xt8E65uzh1i9xUvKXFtUIWqutgg3iw8xc4K+UoluEmlGefeE1VaXPRDGQ/VL92ukOVnjiNzuxAFElbwSK88ns6uEpHdUHYcNu/eMZ6TFEMbqpSs8qrv/D7I2R6/G1SxcOG0wA8va6eWVJE4tz7cbmpuRM7S0yrD8U8blhqqCKuy3QfrorSv70+QcW95uQ7BMFIfC50ZE3ZphJPKpDUhye9cFa98f9meLiwZLvF4wZ6TQA5WU3OpCPatmdzHOdfpt5Ih4VjHjMbbF3gHr57P48RoAurOTb2Ma30swXwtUU4TSfvo5qc+lw20ZjBya68Deez5Xl2rAnonSfBybYjGJlJd+xikl3Vywd7914j3HCEbaKEWsTXCf8zOohuI6/s9mdwJtGnvzWnYNH0QbodunFmCjM+fWartdSRrcPf+Xs6Oy0V2FmsdCO4YwqHDgUXqMW+yWPAbYfoWKrYh+Me55H3VTFKNyVJH/7cKtwBLOJQ1GipUSvELcCwweveUH9jlX17NRW8OK5H9I2edS2eckvvip7XOyyA0CseVaNtAM+iYata/L0dxjnvtqL632/YJm/iocYkLae2134Fxc084O3O9ZziItn3PxHaTBObv1DgbhQz0RDfuA4+uRLu+rugVv/26IwJ8nvwmlTjBOC7MXMiVSJ88gVRhNuiUm9QX48g34zr5kLHn/uLt3iXSvegQTdgrTdTp/LWJL13VkgPsD3jmVffHAGUmmTMvuu6SstGJkeyT3EyOCql9F5JGMWUVhlS3+UihnJwnnllKLr9Dp4NTKIGTkHrx3r1qK7eON5M7N1sIiuzMx/6/Kxy1hs9s8NGbMRJq0SGEGagGGKMCLxQicugR4lm8aHNnBV+snMgyU/XotiST/TKBSqAxAn3P15MfnJ34iSSFEXcmYBP0X/Tw0bu9gOS9PsQv6h21wcUdvFDE9vn6xg9RzR0BrYkTR4KbD+CZdGQyjc6G7jXPXs4y7vi7nNHD+sUfRS8l8coyhfu+tn/SgkZ487sFZs0GXeslyt92JgY0g4LCckvbx3FA2/ubTVZjxMG5782wFAQzjXLC5iX18f/N6tWrHyGCFsXxWG8MK3I7qLsl+KvlLH1SthDk3YOnr0/Qt00UCfq+HHxyhtHoYrCgYxbc+cShViGbIVxIOHNdir09rZPIGWB05x5Mdc3CedZoAZnieF3X/sk360iCXMJ0Q2nj9oENl6BUPCdHdeVe3zMHGT6WFXj6EvJG8vPCAHeuG0fUKIICAQI5ucIAl63yzkpq3GBTKlqNv+pLR95PQkhok2EfrcyWOg+0Dy02WgdJp/K1H0Y5i8Qkmes9s1Ik00nD2qw52EN6PH3yq58P8/NL4MUVTF8zexHh2YmYvb4BKIsIwUtw8QrqT5Ded9c/FYwe/kSCxuk6vapGY/VHD8VMNjKGnS2ZmZuED3i04ZjCV/H1fS4l98+tB9lLgp9kfS3kYVp8hZwznJaT4akqTXXne016VenGJmmy3urpuSn3Mxi3/xB478nvoFuqCfOHASJY1BnDpwZDbHO58kSP6lL7diH9mWPlEUS+7B3W0uHpj9WthAn6KqSMbarMhN2rDX3KSxtAXPVlM3wpSXx4jmtm+92NQ0fOXvxcysU5st0GXvOP39z52E1qDuzprJv1wz5QO6V2PCZBGhapzho6/OEiO6hEFAyYG2oVAzL5dO3kATZW2PM1KK9UMlrI4AxSTcoPHkuu+gcESSy0MH15+7w0hdInJHk+sQ7n+l7TjaLRgzJEundCpfu9tfb8XhOgHllyHxLVgcFFeBTBT0wFAbvIRKjKs/jAXNPFdV2Azn165e8paHUK8e1Uv+bwd7iD+ymrgjNuYclIlEwg5TImmjIPcxQl/vwI7/eOOoA4XtHvOlH794Ik1YHYz1+ndDGyUTDzO3KYx2FQ2TRstmRTQgsPgAelTa0748vzif453PZ/OjiMPStKJxv31YsuyUwhv82zZtQlg8cHjsHmkFsDWnjv94uxYct3EYejkJ4QFo5AaDHCAB5gCCm2Pv2OLnUfa0TkJvt+h00e52V6OhyMf3ccxSXXPfAybiActd7iR5JVJsLvEnz51HCqrZ4HKVrqeXYjVWmmwbKk5WNb50jMskJQziPX6hTqy75n64FOqyCBt1r7S6iCXGWM02e9Cfdfbg/y1Ct1Z1nBB13XYzWrRnMyIK6r0GQcc7a1EynMLIQHuCpU95FLrkl+CAMwFj1zfr9Kyq5xp9BAsmWipnoqotTSkVV3V84JTJAzDJ5g095qbf3PLjltVRdPsQ2Hrc/we4uELbBvwfCHQGakqpYIhgF7jC9PxKnRgmZ8gdLR0uJjVVmb/SAuNNFPZT7e+XVXfH/0NjXNWdWJZ1YeUcK0W/rywosXKA+BXq5d5q37BM09IQWcTSKPX4ZOtxE/aruSVczxWPsm7rmOEWT8MiqaigWW3LufhgBw9dPTwyE04czXRK+pFCMGaqXf/y2VLdXedbLRpoPrOZmGRgnsDdrkCk2qZZU2+wPnBwDQ0xv9SsUdkbnkWHYGCm5EDneWbOrm38XoGgk6e7aq2a1oyKRMzItCqw3C9VtxtD3uJL6GUTvLg5WNHWEzZto+oQwiF/VChm192J1uPEtph99pgCtRV9nxlWdcYFZL2wYLt2qJWIsNpm/4yaMNLpLvcWz8rM1I6iLq4qp+I/LINzNVIxONJMFfPIi2nsCiOPog7E+EPIj5fkBmOHhWIugiVVLG38+j+z/PBbfCnLYulxLpFC3jghFqNnyoRyGenWNNpgu9Tg11Ycbfhp6ywgUuprI9tAp/oX9+ZPdXc/f85d7CgeJx2udA96i1kxY2zYLPErxOCqZXm1bMBjzVpDPMIVzxvWYPHJwxIFoaf4iA00q9ajSEdxkolY1SpR1aDGGWy8883MJhhC64ZjPYSuwSpUz9bQCvve6cWt/ZJi9fkG+qlaj3v3XzpbniQSxkm1txNqlArDVhQpgt5N7Pto/3OwGxajtlby0UPg4o198UeEq6XquB/FyqM4yxuIjU1svmsi22d0tuPqghmz3a9gvFbfGOfsbMmKB4VhWuN/YIBOs8JTaeNSi++LTdtZEmh2IqYFoYw7UeCngGeKq8cDCf5ou0YFRzqYQlw22oUIUChuiRRu7ygsL57PKsWhWxNTYjnkAvwq0yU5qUrIPzzOzPXwwKF/Q9Al4882r12e7m4hcWsO9He5XNa8u+tZl5gx2WcKC/0JLrHc4El6i2FM/nseVdkiaXCffeNBBEF3KOTvrh/D7PFBLZ417c6zR88wrqk84cp58szsgV4FAIWnQ5jabpPgc+gO0/aVI1KgTU49rD1uyrjKWvJrPs0ibVznp1PxCSkW7ASrQBJkf+csBsWVX+z2+4fcaHag4kaKZMIvt1ZAsPRM5RcruXjW3d0igT7Hf0mjs9W1cZuV5L40tmxW8/KKb907LOLNMI1rENveOff19vWVeYtn3Xp0pO2M+bmih5Xp6xjZmCD6CkTBYNAIio5Dg134AIRXL9BysYVIMZDa5qRbLK8dCdJ2zTdLYFvbMejrjKAyVaO/QwSKWmh7AuDRTg02+c3Jrc6sIqMBKTIkk4e2bBu6VUbG0kowXoDTey2nIPL21JmHoN9is/Uo2BB7HhVvkwDr8RLR9mpGK/Fqd9aVOVzNwhzMGT1QFHaXzInrCcdr+iTWVGf3Ga2xX/M+GDI6ik7SrxfgLZMt4ENNOPILVQUa4zMcrtaQYr7fp/tp/YSSrCw+dJP5pZXYPJ+lo1CWK1hPHGP8NKCpSMVo5Dt+Mhx5fe38wUuTNkrG0my+NWfcYnOwEv1itPQwB3NHiovd5RpBn5f64eaLud3q0d3ZtGj7LYYA3TlpgHZOW1fyn7YdhbcOBc6mqlPER7jDZoUJypmjHphjO4EH3fCQO0hhwx3ZDP2VhLSJ7E5irH7qxJqwlEq4YstqrTH5vbq1do0xKIqzmTq0gkT0hUJMA22CAqMtOHCLn6O2FM+8XI+bclVEPLr8eWva3SO7CGskVTT/meJOP3AnJJNKa/GrHGIDgGhA2pphmfrgkcrMlW/1/BJ/3SRbUgMzuxHeI1uIwD5zOIMNje6QDg/cCTaPxhIKxTHL5/CBRvUBje+h7fjJDYzz5DTIabuX1aftmnvAbPa51aIBdcFvXXABFmYFP239+xKzsv+lHdea0GLB2Gz7ZfAocIuVDJRl1w+5HiuDe9lAn/Dc9UfNXMzN7K5giqhQrdgYxLW+Q2ttQ70g2zcbEKSfCPssNAQzb8/cpDBc8guHO3GxVPUeC6G4OlU7WJiXIE+qKsMtbr12iL8G4gLc6v9Sp/GAw3tnnLYUNtAc6ED3wpdcmM38Uvpun923cb3IO2RXsH2uMdTgyMwxklAIHjntghttmgqz6zdX0ixmpiFBalyzUtpOWPKzwZcoSupVuaKQnK0nRuCH36oWtJ2ay7ZdExEp4EDWGCf2xb2jWL5YFluVa2q7ZvsOSbGrtbp1ynqs+t5Zv6YMeAbZwStambYpGNBSNBqsEpoYMFnGUt/wZybodiXCc/nX/5xw7cU4tViIipxDyEFPuNTNqPlNCFui6oqjvuD00+KXf4N6S8PFpgGdb2gRrTbcKOX/hJnpySntzzI/X/OCtnmCjk08PeweR5Wz+U2wYBXFfVxBUsfHnjzaAdtgYi4BkO/DiTTGhEbnWYol2y31/eglNSlMlTnKTXGrpRp6NTdoVKtR8a16TRpK4+qowfA8HjD8M90x0xbQYpeQltoIjo6ldFOVrJ6tukbJbi9bprMZI8DoLC53VfMtq8PEx/BL320Y+BDrL8HEB5qlTrp6trBWSk1tXOuEJCLwghcnYxT9YD1OQoY6cxJULS64IPBre6RYGdpOR0zbDUiD7/0Wd5a8vng55Ne1GvfxrrRLW7iv95xbbORLjkCmeCJMIaUDWGxVDT5GwUE9QlfzH8mliyr8ot8tM8kcrha3QW0obnkYhTnXrLeY0zphH58nsM5FIjzHAMaROFwPv3JbqjZF0CL+JkKoyCyu3Fz3lmX5IUjbLGYJ62OXyaKQDlgaN2smbLPvcR3FkHe7xep8d+yQQ70FGjEBR4VG8rb99k0iwpyqWFKftpVHsbLkH6nvHY7QmDM8WYfWjcNMfBuqAh/CL1sZr2yLS9D2t1a67d/iND8KMX7tWWyXNOprxV2HD3WCxBe1bdRKrDlJ6ospZgj1pQcvIj+NIkHeHkEaBhUTIkCCbjcJTLQlnVd97tq2XdIwClce9DG6jkkzIbROI30qEKvY6dr1mJ5jIzlAjsoeWNTUhvtZQGU+90DzNHNdVdM0jSh+pLQT1f3Y2Inx6BY/Ca/KhORczAr6df57KyO9tQDOtjM2DwLeZbRTIsWzB8ZnWeGpD/eqJrmX+zUNo2Cnw8uaXyRJfo0LSGfA+EfxNa6HM5K2UAUFYgrteeA1Pd/+3BUZ7vruLusW30BOs+yV7kllguGxcw2u7+yKKJ9NSF6MrM2vEH+iD1hBn5TIIP7lk2K0lebk114m5ltmpKBtlpa698iw/KiYNlw1u26K8V+wrXNfJfZx+TBA4eIC8jGaYKYmVNzR9lZ36uvSUNBz3x3os+gUd19aLvE9oU5waNkmY04E3/ISFGBK/eF4vEc6CtoOebTzCtJf+Zs9io1i/s+cd4t16/G9sm/yYqxUxqiTBvOQEMgWP6632P0RoBOuR1riAaIMPJS/mFT4jt+ECGpMmkS46vpcDVlaveSXM358DPx0L9fu/qNObFNIEnWUuAoNs4ZLy0eHZ4K1hv7YdONsbOI9Flax39IczbwJXpyVsRRjrNYz/tQKD9AJO2AV6zMqQzmAQIV1rc/FFR7Hxua2KxONuGX8QINobHThzgQz5zWM7faUXMyf/7drGjohye8xzCcK7OQ8pUzIjMHvsSdosxiloXOD6ZoGoQduPbpNW5pn5vLieZr5ZemLE9piBiaQE6zGUB/Wi1zUdCK6NR6bnmm4lIR6g8EaaHu5tSKb16By5OekWtwxCknQ/V7WoxnpNCJ11mA1TVZj1M6Ypat1bkUNlTgEOdd/FGGwUaKxtWh+jfeqBdz05sY18tx9ZRGuunHhE3GgZc1/TQAnukF8VfjdSzHcYpmclenK76ap+fqi0WCksicI2/wqWuHp5JG3Hl1KsaSZL0Ztn6JA7KerpsQ1jB0yOyO9yuQc2gwfSpCg+DVP49KItEA0tF5C9jzJdKeQ/POJdv2p/sV4i0sGWAz3uHaeVR1Uom7Db5s7zTvAIz/UUozMS9oBfNrvoIb7LxXo2uak1EZBgaQWL5D8essej09LsbXE6jgxuYmVU9mKemGWV8UckRbhUwRtlOXt95mbEEs27qst7/JyoL8gzbxjFPzhzCHIj6E/+uIJAYjZ2FUVXDGFD6/xgLppOr6vayhipOg76sWE9oxVxObc7CjmRKRND/gmcXer2OPx+ZKf9TJX90QoUos33VoZk08O3eVxgdQ2szTtIBi00TWSkF9tgF5l5qv1VxqFG8JHWy+J18/wHy0RKuzQ724+Ua0Mq5bcn8CXHMtp5LI5M6WFfph2DRRUCkIx18OyR3NEYfDaCUu+fgxPiLCj6sFKxyYnbPRAsMSMfh71iJajjZObFN62Qdnob6qxAuvRQLhK6ihuEhWGeDFPjw+eO8+umyo4E9sbt7FZslvsJoJ8vEw4f3UcpVsby8mW1oaBCBq3vRzwM5fCPWtevOjuLvX+GSRfdeExKf4jzAlP7ShbFbkX45dT1Xy/Yer8DW1tDy52UFlF0CEwPstcV8gqT0udKPxJpKCBbEDVjghQx+PVt8r0X2yuHvzSQPerSL/hzcX0QN39U9yrgpJ/Vg10DkYhXKDZXEkvHxNVBCeGnAMXKIXAYZPMeHlwiOLvh90cbG/2le7HSlur4rY/OF8WilkT8ivhLf5KxCjchbv8KWve3duIfIWpQ1HMCZYck2fOqD98EakzBGcf7NtoaMXol93RPtwWN31E+t6Z0/ntluVw9fOXu3CTJKc8Pm6IJVsN8R/E1yypo6/2q6NqB1aijQaDibjLj0dJOxi9vXU/d5juPxf+W4V3uBzN3HqYc833+qV6md524db9kcQEMpAF1RLBth5q4QommC9hbDH0i4ZL3AjZlzSO2lIj7Kz7IdsCen5m8ii+bt5RrI3xh0ibivVl8DDx12DqobFf6u/Kcsb1OBIfzHK9UDTa5xGH6tyGm15GV5XbV94G+vali6WVJH9ZF0vX92eOqUcd6Gs3md5giqsOp8TzS10Eoa9S2aBqu1K79ju48W1ElW9Vj84CU2QZ19wwxar8+fmPWVbQbyPyrGgxYBNCzISz/Y+1a0GS44Shl4OCA9DlG2z1AdiqPoAyzLGDEBKC7tmhZ3AljnftzUdWhPT03pMV15SKypumuLOzUe6Xoc3LCnr918lWt2mhXS0Y/uAJWuS5fEF3XwNmNoyCXrsvnju52cq+HvWR24zg8Vb5EAdrzUcuuV1uKiHj6WXrKPMaJHLGUTlx+NkEqWIUbbE0XCn+BpJ/MI+CTgrm8W77rGmzbbSzMkMHEeUqn7Axi2v2hllPGr3mGO5++cu6AOcvH1x2uyx+niP8/QGgnTuK7xZLtvEngnhOyPd8gyoo/3IjPncNmbfv0GEVX1AGCOcVU8/TPHV4BnmSjvwoqBgLXvxc1lFU45onnVjCf2QePf59iFEI4Sfo5Z0Cf5rHkvKItwPMZmeLhD+jlDAQW6HjzZ+ATx/l04nSuJkl0GnhRR0FaxsJLnbl0vaX67ugYIltUxCF3FdTLvHiijDBD/RXELv/W7d/rhsJuAjj9OxS/gROd9DIr4/nePFuSUeBMoREnpn/Phk6wqZprpbV5WwRtkkjXFUzzYn/LrjmBZ2Alpxwgb1pVlb//EWgMtEWS4dyfn0yCLTIp43xYk8XgL6kqgjLqg0fnRObZhVrWZ2dP+TzOphgTvukP2GLnFJQikWSrUc17P95rMrinVUIlMX43oX4aZ0QoWjY+Ixo6E472zY7M8OqHSi/+833JGK2afQvd9UVlEs1eesvzPMsbt6JDMQtG2XxQn6xyuJYjpnf92ljaILqbmeK2c4d8LK5nuiwPexjJ09btirRWxJfs1iGnwMnvTC2Eqn8WlciX04iKOfXfR1LXrmSAv4rfGaVwA7F1Z9G7TosG+8bBVIokzDT0dfsxLhxRWmFriHz55VzP+NhJnsTmxYklSA/vToEvc6/eNc0eYP2SzF8truzW5Xr13OB+lyr2brDw2xLIyYIE5UCuqoLPa2yiz/8RdDy0hLHklKupDXhxVIqqtX5MryYjV8h4Vz5qeGHVShb09mNBlbMElTT88TZOj+MbXDKzKtWDbx+IS/fvZLHBcn0DiSLaxqvc+EWDXTlcP/mEfrfhwGuItFNTR0iGNVm8RXFDMHcPSNxodCHF54TpzpeW+HDHI4aifKrESWHo65Hm/PrYHX+nWKpGtfw6u6+DKEGVq6UNwi+u+vcbjo3bbMK8p95DAP5GoZpBF50ZzAS5RMIIp+wu4gY8lSfOznr8Xjsy8x1NdLGriqfgxTMY9uEodKOHRhFYqMId9eG32+fe2dG0Gfr/NW0dxXvo1/914ePaLKHT9qm7fFY1lGQyrxw2vAJIPOlf582xZYJxWG4iCKU7doHWxaQB2PCu0XHy6C96IRFxq/wS1/R91x/cbmj89rHPO1B84iu091jWS3++WGZOdAFoHivoQhyDleOwYet2ed2tqNNmmT4juhdkJhbiL5N43jCKylI7D53FOQnOnNE7CnIRezg7Wht2zpC0Fd98bMWCvIQsx9cFLT1vKXYPtdrYHqwM0ILbF7mhmWi4R2A6SWA1TuiNcPjWsmfGhH9qfzi5Rz23EyQd1DOYgeHQuTLzmNf2xc/hDdYmZm/n+ITlTmhXzqqwU12oJwwbw3O8IJUfNYq+rPbuTfSU/iSxiXMrnwqlo5NqUfpVNjPKqTtpykbyQnvrjdQy14RO2+b7tb04rm5LXXeHpNMid3jpgAAIABJREFUK/+HLtefdqIjEkToxFHrMBYJwBHaYU+M2PHBxjUU4Me+aPSoD16pFInw4ntgpu2rMjExjRwBqw1F10yUUqwMHO37+utFfOtbFwZwjri/TH3Xp3ms1cLVxQcuhjvjV/aI3tfixR4ILw53XbgbRbsSgNjB0aqNqLo4bPgert523IXamucBnDitvaC/jiOpflWsMS7F2OUiEQtK5yBBehLhqggbHwtZ8gJnejqIELffW2esbP/u4Ttngr6bHZTIjidnpc8Nk+H0qqHwIwO7C3M/LjsFzLukZmcHruz60QFQU1UIaVtmrrv/qNeuKBvdfUS+I7luygd6684mCaG4S9l3e2c45Sd00tpX1Rra705FiB1lcoJSJ/Lbji9dohjnOg2JCVcVMF7GL3485FKYp47i3w0DJss9sbx8QR0VVa6uVhgUzTm+dy63b1XOflQuX7MCL5/H1I0lMX8cW9KXtTZ2bUcvRNgXD9ClThRXle326KGuSLQqrHw92HGijh0hzPYQvSrUX2mVThQ12f13BLYkoXYm0mo/msNh45YS0a08YxSP50Ihwr6r9ShRM+83xVZh8nKJcTyxxgld37qqBbNvWYI9FOkVTqmtiU957dusERVrIpXvEVjDaozBd+iMjdRqpARpV5WfrlJ8b9dflvxIRKIDir8frDtC2LQZfz/Z9cLb0DyWhDkRZvwQmie877oI/6c2ybiy5cAJo0Y65y6lc3kLgdNY8632ZVSVXTZL8KRjbHdta4Iw2dQz1xR2CgXShO0wudZ/xdU2Lzx//NUk2AFCecaIjGuCIZii9G0No+AsXmWuyz7nRYhAp8JmoTYWJ1nbzNi0y5KCixsMz5ENdvJSEnRYJehlnL+y+xFXoMaUwLQlEf1R8MyYsH44h1gxrpSOmBLkvvgA5lH8DNu7bw0d+VCYXKf5vVWJrez1R+8U0w5d1r2dsXI220yJGEccqDsVcbEO7SGfhsGXmCpyd8RmmLpmzOr8M+kp7Necx/syc138Oz3r6GH+y9PdvOquyTo0pTgEzYnX69Du1syc8OAVGuxfEFQaSg9drI/WTeSApvLU5dKcf+wxwDmBvckpTJcxl5rr7loCTSx5Y90W/33Us21CKm6UeN6KypU1MRa0RsRKb+pEl8i9Y+4Y35c4hdNkzLJL4s9hgB3SVaJoPR5nm7bvRw9RLJGv8CwOZJXJ0jbI68JAmwjigtD3EO9t7nxrzlp/BmYweR3JP171wfjRcbgyz+U6XP8niBTpGCMu33EDn6uxume+r4GB2C2BMPnCKZi+F29ZxxgEArL9G6e4mHzCjv3L9RwdJgYP+APY9FdcQXOYCr97Y6Kix+N/Y27ZsD7k8Joj/07lD0t8vb5lvgqjqFcbq5oGL23/2mlb0qD8wthq+3R2Rp9OMoqZ8o6N6U9Hv9o+GbwZ79X5ni1Ut3TClaggG/UVddCLBajA3hhynThy08ZbfhrvFj13CgYiDXQw/yYX0Oxkvmn0B01zu6fOqmOXrTBPc13hhd/PiyLxoql2ObyuFIac3C6R0i6WJhU3/dgal7b4ELj4Z7wA9M1dD/HMLHcxC6ftzpJfeGzUJDcd46ZLBS/tTHMuv3VJQtfhVm8v9qHQ+/P7mJsJdyoy8YiRsAr83vlqq/KkWvygLO4B4+8MHZs3EBbj3BbPKvktU6yqW3zQNmxGlQomwdvqKNj0zuEdycqPXYUfpzq4KNgJ1MoZ6iCHO6WUKJk9acFSwi1eTmJcj+Za8Ux9Fi/yBqoDdJ08fp0NE7VYHZ0ZMMywdXtR1RCH0HtOvEPj4a+/nDw+wI+bj9L+OirJDl84JAoSYdCVFR6OHIfDLuMoWw94Cpj5JN7gviiL+eqEL/9yhXH1O4+x1XFOPFT6KtErRS2ztlv62r86CT+eY9WKRvBXZEHfoZ/0sYNYP1+8EejrKMuP/Na5siBNStqITcUqZiaPHtQXo9ugtfGfncd/2mGJju5q2mQXjFEaO5nt3kq/RkoxCHdtRnxrQK2UGF3DvD4whxN2afm9K70ypnIOO3Vtvvkv/Sw7YyUycyBXFRyg/023xVsYtKLhrDvQO7oQ7j1z5+J8/hkYpzuEImopjolZaweCEgpzywF35cXLgcY/sWlLNN7xubvHokLRzHWxeuUsLgzj3/e1WKhW1mo3thFjs/p0dpB981u1KHRge+f5BRc+bNAB88TLRpUM4TyuyFjKqJGLMmHzZfmc4+wKnIwdRc7ipCH5VYVCeBQFL8bWarYUB3VZIpxQtgb7BLUTtTMmgqoHBj8yh8W50V+gw8A2E551HZWiLT9PwBv2xggDlR45IbaZ8rcuwqssonfl04b/LNzxhwkNtO2tgKxtC6VN7e3EejTYgck2VS3gUmE7W1h4tDvKH7ldw4CW1X4q/YRz+ZHLOV14g7Hs7o7Op22h0Tnf9Xiikj9O6u7ES1BIg3pwbiimRHk8cGBvBavZLJ3xTd9j87Vmp7Koq0TXwp8Akiy50iB7bCPyj3OzkRCRByzGzeh8H82Bvr6IwGeg6ebEhMWVVcXCNiSzy+GmdTZWjCfU8S/7dnL2zYHGXLTFY70QaoqwUwifIEwCO+MY6QuwDMd4IEcF67MpvMyjnXcdFncLsvgpCl1XKA+TTvF01XnjrVJoATZB6w7k8kwja09V48HmAE660HM+H/zkcScRHTVrpQriUJc/PgjIBBw+EqITOfLxQIwi4fCxP4Sssq4W83RXTgriAP07ZRWvKobtuNqbbZqOwFLRab9R/5e8XCyhe2ZmN4fEihFHGjCAKnHZOiPCCQXLdK5gFxFbChJAp6cSIgx8q+8sP3bNuCpJN9FRsL+HXApsB2jURKdcVILeJYW5tlgJyP2fkwaMiqXopUHGly22DQnOHdR4YEanMvjhcjT/JuB6lDEKPhb2WOVf/Kh7JcC9UghTJmJijdla4m3EgIQCbzsTq/AOn4COAtg4KP60curGQGJL1K9KrMc9iIV55HJArME80OWygFmcDMYYKW5Qsjg/dql2FAuzuDTGFaNAKkeccF8KG4tEaxZv1o7bJNHrN4B47IfDi4yEniPR6Rj9CQcC80JeDgcmc2nPqrqjdsSIxBPQhphmxBQ2mMXctpVCPGobVyBtzH2dPGNl67lAchTctDFms7Fio3jZ7N8hEoNXkLAiqfmXxsQVIi4i2GQqznYI6IagZcLhI2dxrECbx7EahXf4LTHSVl24lymW9qbQLY3xOxvuMLjeWbtt48i8VdNRw4uPiv7YKb62P+89rrSLoxQBIierJyhN5M6F9poc9Wu44UCgOJVa7Mo2JJZaLM6vO+/uVmAUrLsTW1KM1b/3AIUVw1Euw93sXA0FmZCpaPFhRr94WnWetBygfwPgROMu/fD/rF0Nct02D7wcOeQBqMkNPD4A/ZkH4Dy+Y3/EAuCfpCd1rEwbO07aSREUBBa7C0dLJOxBAQclhBfBjwApCjbSNcYp5pQli5PqEL6+H/WjeDdRWLTmpnGNFZrKDgDC2YP5QDmTrEaU7cbxd9SGBb/0O3n52mPUOc0xUuGGZTRaCof1R8TSDmNIjXt99yi9OYvfisnvmSoP0Aa/VdrowlUeM2eiebsezM7dwlxfuJbF4WMaH9pnD6/aiAPNh6qanI6JmISwZeFd4p9gJiYFs/4JRKw7MEznWNtiMcKruTzIaR7T3fEGWthA8GkjgnG49dgtWsb5DJjtpvxtYfdfNDN53YX6Q6GBH8aN7HrjEMUwJadEzMtEyGYB0uaQzYTM/+ZEe+lSIhWUGub0pvlOlqOLSfQfeRTt3F3NhDrbfbxnrkxXuw1HJcK4GB2OW84nJASdCB9bttEUNw9gxK5AjF3etJ92mUtCDXfsKw9K6OIAYtb4e/QWEVlcP6BcZPPuSNv7+0H16Ncs5DcMUfxcNhQNYGvv3Sw9MOqh2xCh2+SJmfLqPy1NF5NcFtGlBlhQPI0IOxwHnYIbKXsJmkcW09rDgXE1THffTzEzVRLGuzsix9AB3QtjFduOgdlBerDy2FY17uC1ZD8DxJO/mlSJg36iEwiN64FPOmAkFOJa/Gj4qP9htTR4xzgR6jGrPAyyGGsloJndLOHBoxNDX/w27EdxB8W0SosXxM3MQHGrFJLFN3bOK3c1H6xJT6wENX0HeRIvN3jBT8c0PBpjIm2DNRHBp6iDNCGaiUYVyuGsXBVRLL0eYcl3TtubxGeYG9z2c0F2FYHoZAG0N9ruLUUXz1zzJvze8m4SyuR1wkOtRTeUo1EwnjrfglkKomdwiUlFkwhXi8jhAnEjjXqJEM2URmLm92PSxqEWZy9aj3DeUYQhi0M/KzGKD6RGqKp83CeF+UbK9XyXzXoN5dRIyTW2tuOYUwOBuYJetYL9KAESNaa/ANywDqk/JnACDQVSGMVYAIpnMIoX12LUCfTFMXJD8e+WwqPLGHXk6Nt9uzhNhOut3ehFM3Zt/oS52cmbMwTnlKnNrTIcrGq0M3URxHlF2iK4NOOhwUu1b6uVYirGX49JG1EovnWAZr/sn+uLjKG7LY1o/OiaGwY7q3Bfy9gw+DwDFXloJ3ZMCrQPRV440SShOcNiH9wUx7Q2oBL0dXrrXAFxhTR3cCVNY6F4PYS06dFGIG2ZrPA+LpasOHswdW2bXCe2hbgtS9GRDX+hwl1wTG/GVD3huhbJd2fMQGIDwwf5zGNJpMkO+q9CvAmaRhD4OnBHLJvom/cUY/Fpewxpm442UitTe7afE96gnu0IjSgYtoZjzkfKZVmncJteYbzmCXpzwAs8JQx2c1H9tHi1TaH2gSJNzRjhwmQIzRv+6NAWc5NM5ZqIFKQdTWV87h7qKEaWvB4ptnfYQKHdIJ5wTAWJ23JJ6/Fy0e7DcsiMh3nMsqnzZlB79BbPO+ZpAwBKrK7DFr9ISaZk9ejRIpz6scfDD9G6JfqTKHkY79Yd9J+u0/TdHUEhtLr7YIbXCrDQrHoKt44tCP7e60T4jxNdXtqH/FmQNCj1HYTNmC9ojtYIp8jS8sQYfQQVNvJ8ApyiNiLkIpako6gRfm7rMWs98JtkkP2CzBaaZHQ6Bd/oa4IUh84W1M7NftI4m2bG74+Mtf1+DOlsKo8GQmhWLLulyls7BXhiommgQsFaGmonqAiTBoGIKo4QoGEDzfPz67EsVg00WPIxbGe1OHQZ2OQStnVfJT7PqDtRtSC9TcNcYcyTaz5T3WbcB7xLJk2I9oACmmNGFtdigIeQzDIxdCC4FFpsoWulqF1FDTtD8rz3eIpwNWIUuD1av8Xjvth2ejwvk7bQyEAzQGElZ+3A/LkmUfidtsOvJOPhi76BaAxMxPaLhLKG/wsofNAa0BoUOcyPnNjgOaolkJOWWiTf4znz566ZCx1IGFdZGuPwGWezrWMLIweoL5yFXdVOgd07djBuQseTohMSn3fEibEgF4Yjai5nnpExyTlEvA7QEdQUH7FNQjH+xf6ugJiZkixI8Tx9P+j8OnYU8d+HvljxSz12sM1lOAyHlKwZbXPtjbnOrJhwXoCLk/WpKm7VWynLppTfuoKTEiYD73GGWZho2IBOFPq+loj6s166Yj0E/eA18667w/IOvMHDns121/ihY9uWix12FuSOp8mvZo/OP/FmUSrug5s7cZiDiZ1NxiuXCK5kbR273UUQKlJb2VHxjQRfJIwdBL0hiaNQVV5PXjP/YrKyLPkdY5nhYna2OnccmUCzrWsYCnC4IKesIcwzrjnjyHlBgvTCTNOAkQ8NkVSYJ+EwuVHCOvqUpjguFRg66HuPuY4evFqJGaRQzuBThKtmdM5tGyNjh9Y14kvcztnNSPxkytY8rNpVsIuxzrfjSTNdO5uVvj01booRA5OHbxVy2jH1kqAfaklqSA21bjR7oFOLjllBBSJH9G4eoHxbjz45QHf1KBbQke7dnR5ZCo1tFRo7ZRvTWOD3MMJqUyG29569SdjsF+Bt/ZZ4q5+g3Ug8hkT5egTzhz7wqOzQTRQe6epTV4uxrDwIafNJKW3PLfl1Ay0gRdIsPnN1lSLSsngbq0RTMOoHtlq6w8fM8/SWT3kSvUZH8faZ1QqJWgZM0kTPdgTKEwEerghegHhqkmOUTx1/y+iM3+Vd2oP39XrI4aq5cItbf7TuZD1qh53H6P4ziw9sC7Vpnh736a4TXJl3uzpzsPVQHCg2x9yM1pdsGpHFGE2IGMgVGFMe8YQYeStF0rhbRFOd+Hpu6/FiceO7OTry2bRzpE1ZmWF3VEI/Uda28idudMV5sFzzn6rDeo4G4Luc6ACiw3TBjLYYvFbafsTIEBBA+KhDB8mhuRK7/Esb0oTmmPvi94OGjryDlp7NcxKHTwQrveuzHe6dzYCwzdiavXI0GC9c9iHP73uN3g97QX8yemKv6ARQ+Axtc4LnEuVtYX4QjyQca3ykKTpScGVzl7rO/CkNNJuIiYsYgwo/9mQLrbTiDmPOh+ysULWVK6hrvGu58wpW+gW3yIfaXT2T5LRaiOcERJpZKa6O2SjUAEv60s+BGp8jD3eucGM88CieO7XNfbHacEdzcowtdOeJbZQdNDpbv+lsTfPkv1OB80TqGadmf9oz0+4oai9cE5JttE2GQj8W7sco0IQM04ThQBcsaNIKXkTHZEEenh00YVGlje3oxOu56U5Z8pRqDo3YzxkOH/RSStjscmnNiJmuXS7O3ACM84Gk2X9G4/Szwre+aI8ExVcUrUGmOptpaOb9BleGwn4feOiAa1KZIFmCikcLkpjZrw+tR1+jQ7SeFDyCM20Xgulp5757DnY041ebpeHWWrhuJjAlrGoOP5vqHrPaXNcpRXr2iLnmIIEmmI3XR+jRKIHZ30NijAmPdB9lqBNvWd29nuwoXuJfTE1mIGrmEmE57tMqSD/ascr2+ezatEiydwa7I1njITjh+5SXjDg881VybJPETaU4rO4AB9NOI0I3U1ihS9mLlEZxTtxp0OrJC4X76HbKn89YtYN3OGN13LNZvtvKzbBCbEOQTSsRsuKQrL4teM4thP6Qk9JCjgSVwBfsPDm+mNlIY0fBJXMERuTxqMmzVxzWdSmiY3NA2gBRRCoUFOGitMHHOorv1rXx5FF/t/yUnXBUGtbWbKDHmaPT4u3dJJ6rbL7xy2KLvrBanXf9F7qGCmHRQVR4AMS85qAZGmWDV6XyV+SuLQ9yGmAUD6lH+zE2gJkct+OhQyjb7bVjtqBdb9p1S8x9h3wqxc2LDbRv5KCuvZsKiMuzXQJvMoh+WTAQkyIBO30emzPPJRxSfuowQDPIltgu4a114llmJryB3upHgZ6iYRH7ti2oi26nE7eDEjI8H5mOXttsd5gynzxr89fA+EONKMyslD8c2IOxWTzhQnS9lelVEmFePWNdF+Nv4a4YEAWtPNKAFz+1u6N/z1sPxnucCovbrkw0eGIbBHbbcLZjON6qqzsJbLjXq+V1rlsKsT9a6SseJHMINhz09EWmvlNiO+xDo4YX6DGPddwQGx3xeDXaRo/XgzeWvhsbyIsstgbt31orbHdsVALQcuNnGOxG68Zw5YiZ588HoNgvFtxcNkqeIk2+uaCzcufG8jpymPAw2wYz5RfiXBQJI/0Dc60w1hW0yULhLoPlx+xc87czVo02yLXYnPm02SYCozjP3j/TyqPfIr4HEufeTuxwTb9TmIMdyPB7n6PxY2dEJGqEPlzYuzjxog70bX7l9MHjMUQ6itKatsa4emY9OjjXQEtlfuy6grZKoRiUB7ZdcQ2D6521szr0xs3h7PelIN9ypUFYi8j2xSo3IcAF8BvVDcf758jSOmQ1hmdGMh0mD44y4MwW4Sd3d19NPprfQn4lw/7F/TW0s5c9xpTLZr4APw114Y4MLC90qzwUDL/bT+euIIe2Ocrw4fgQCnZ1nLueZ+WoSBpTq1Al/C8/cYJO8N/ctKGl+B/vlRbJ0p+9gXj0ePNwtHZtVpVg0+kZqs1mQjE50o1ZdXm21R/Bxbsz2etwx2sOPYCbhP1K62hSyWC9kdCsgV7lIJzh101ymW8gJEHdOI+5c0upOTo+OHq8vluMPb89AW7y9tDg1fZazPc/9g2xHVu2YK79n/20yMhHfjUTfJm76ta084sRWw9VgEFFk9itGB2DQd9GNkuRyWyJMtlh6BC/D8KL/VgnnuRRDLMH240EleCuTktckVnKyDKlrROKwwBhWnN9VdRPF7T9qn4+Fj32ls2JRB/y5si8VxxHibAkRo7CoQa5DLGuvGwxymjH6E9N419GM3l5l7rw7vUg0vbWqxP1N6sbuTmFQzsrMarsNjP5T9gGxLdH7ybvNR80wn66+jyF3Sc+AMZe2yxmjOBY0taZ+uIM1g8z4UlBA1StsJwmaV+c4q9L0lCMdUJYg1/PZrG4JdBwb8XpPKxm5lZM4xme2HanO3TmsDNL0H6uEnnMar8nwY+xTY0CFLkmOx6oaZVEO2cYCDro9akvYx48+D9I5cjBjDpEm986GEpO0wj9Vk7bi2+nPDXdvXotBlpLSfhvPndp2wX4DcVhWz3vehZ3HvGNFG5eNP+n7Wpw4+Zx6OVkSAdwrpDMAbyRDiCM5thrvkdKlD2ZmUW9LVqkab4PraBS5PtjPaStpaeMR4eGGQFEZLIhEp55H/SQowUm51Qw0cmAvWkbkQz/AS3aMJUUe+xkfr6NRMfLBFfjFldLrllMJ2GF2EIFdfHlSAMaxPMIvbOe7Q1i/Mw6XtMUsp1O5gO1cbR59x0HZgSFCfYDZ2NhXyZQMfqzhAigoqOd1A8UiAzouKkys1FHcTPu7sIFQGJdl56NLcV/Fo9SjGdvGZFsK4Tay0iKD27Bj9MBve3XOs52euTmVR7BpxEbpLmXVc3+qWzdCuh+rcXQshSof/SN209XAQqp3DnyPueaI3uKMdxd6Fi6af6S3uLHUx1FX946x+jOWjYj7FypeIcAeZIuzQ3yWa6JOxw7CtfYtQkdqqxyQJDHVjO8ipigt1hIcUhjAVpfz1qePzyETUpxxo0uXV3c37urRg/kUdy79nULFn3QV6yNQKuvg3Ji2J1n2nk8eC/OuR6lVvUcvHQOSCC7rHGN/uZL8SiahgAvYyPLLIOzQEHM3mavhjqxRbif8W3c4kLWQ0Vt98umu5sxS7YBSAl92wS2uvfOO3LHJTYjuRmVlpeCV09yOIxiCOHrROk/n7lBLxW1bGC9JQJzgU7I40dHUo1qStLxWXUT0r7hM2jn+Oi13k88bhduCuOu7XtXZuJQrCtep65CtWxexzaW+yjtvHzG6Z9yfw7nfIY7cXJdJQgQjd6kov+vQriNVtANVaLRRlM2a9aK3tm9CGeW5f0L8eThuQsCZqKleJL58U+32HcUFa9dWK1QmMnZ7RWdlGwjHNNwNkuzciG6LwnRehSsPVNrptEYq4C4ImYiau8GQ3M0VwFSEDbEh0XGj+JcA29xhAkaTZzc54ITjhw8VJrJtvj7svilm62xslrMdN1lopasazP7TMcxwzDbUVj8Xq/td1xOWkyv4v4bzizKhcaxElAia2UPI9IlsJpqM0sdYPgs5plGUALIWoSJRgaU3BTM3PTB8wvj7/+P6c41xn53a19+sPixY+EA7QKX1j7Qre/tX3WqBc534PLj6/TVpZtEtWJIR0azAWxholvjPqWgsDufMrnKQQdl3FlSTOja9jPOiGtTdrSMVJWrnrt+ibVOoKayOIyY4r7OzlF2PQHa5mdziX6Sse0m6Oou9rMQ89pdSQ7mjM04u4hIYmSvcVdroZoqkUWCGalPdOQ9iANJOW5KfOT9fD1IcTs3xv/23O3fLQqvRH+JvXyC3L/JrFw6QliDW8eo3P77oO3U72v9w+x1OG51GLRqDg9EsAlkIaNERBqx/PmBU1T0EJE+BCPrpPB2gj8SnMj7SZudhn1xIcn/uDCW9N7D+pXk37qnThnnL5uZhyp+5Hv44Xkd8M/HwaPpEPBx3v81JpOo4VUoFM1ExgX1QwozXreGKkHyrhDY7HOGnrmydkUKcRYsU0oGS8V8i29XYRS216NSN7j/Yx+P3Trzon6xaE8UdPvs5vTn5U8V27OGYobZ/LbWw2ebHnCjST/Q0Aw1BLEKDUJgC7yplQZfBANNk4st87MMeXJ7pWQkrRN9s8ftwlt8H02b/J3Qsi0unnixwuyiJ8h66Bn3Ury+D1g6BM7UMCfFn2Cffnv5wEX/66DLyWmWEVKf5gO0cCVSibk1qwwNp5z7oWv12IBQsFB0f67JBi8hlpgmfx99MfN1v3xP0UP5bdXoDGV2o1JP0n3t3bdnbZ6g0x9JbGnUYi5n5dovIsYFSdvAylIE/S+2DtUIKgFa+wDCKSPySLeM9kLqcDaG1JP8l+HFgOyo4VZ/bseBVr+BWH1guL1jfevMd4T1fyU6DjhEmqPaxntYQ5nIffBJSRN0gRfDaYDra8OFK75llN+9OOyfyhvs5SgSOaObyAAzpVL83O7382aPfxa/fo9bvITV0n8cMG+pgta1CcsfFofFB7fe+a1YO0yupJqe7MN1B76Ne9xYLCrbYJH9RGLwoqmQ/EsW4qLLDkqPp9n6ZKcnu7cX++XNGacsH7fx2mnmx/06kv+7c/yaS4qLqmPHcNdNi6rCIDzGetyeKbguf62Cr8/V2/WVSoVdGkuxOAe4BwziNWzw2WT5iVgPNoR6SBkuys+FQSQZJ8rxA+9dJjtaTEghmPx+iR/3u7mgL6jFdy0Ue6mo1FFEuZDDiTtDxsvkn9F2TU94WZaPBo7TFvLqAKADqpkOr94IsYpYxSh4RcRRi5h4Yzo8lfBMGERvXMw2sx8rdCq4wWSjpSuWS1zYten4/Lh0D/Tg+HHCNkAP58xqmlc3coyY4i5os/3vr1riNGmsfPzaYQtYCuctgT4wjOsBhb+Hq4Pa4kiurm7asBH2KR2G5xwXlXnemtDP8tTJCT8y3B6+pbgsS557M3OWAAAgAElEQVS7eB9D08YjUymm1eSxomqZd5Wv6zCIrk7Jtn6+gKa+iJyYpkCkeGiOINdKiMtAbzHXdABzK5py16L66mibYSEWgwIrhFzbKGV4wy3uaqD286Mx3JfBQLe+2KN29atimbanvMuA1nU9c8/rVItfp9TUk3D7NOkdykmk15bXGPoUjB20gwmuVrlrTT2M1ALSPq4dMdviYFUYD5s0w5lvXYTxo7fFhZA8F0FfpS/ulULDTKxpM3vd1+Gx01VgPaU4+Fr8wcUdEQiTA6Ge4Ymzipvb6/BKVex536RtMJMXSf3KSEGVw/c+QjB4oZdqjjo9799wofejzkQqrKUwEOiabCC3QdeSzoPuQxlZS1/+seuybUfbLSMtfl0+C5vwuGXyFzs9u/dVlxoMCfeWGJNSNTNe1K8ddg+G97Rm+QgqDcKcwZ4tAmHDVX4oWtx+Hr+/hIsPxrt/3evh9MW8jxo62sG1RYM+PMA2EfzLnFLz3slYj9l3p1KSRmhK08mZ2a7IqZJuWOLL2UpEiiQCk5V0loMLei8JASAEVYLi7kBx2H/Ix7mwFjcrxk4lf7/MPWoqedhpdPYYrrCOVGi3PO1as67CBGwuS2V9ba87LIR/IgAYepU2t25V3zWo2SK310mNkGNFlCvzwbRP2x+4HFNuVjTkkNNeGKRwyGAXgRxjuqNWZb/FZJa+LwvX/b7P3F0XtdneVn3xvqZ4TM/cBTfXra8LRX2qh09H59IAgqIubwb2Dia5T3sFconG8a5pHqmBat11wNkjskJE9G2bHHADyob7bCdsaRTKK92vQ9rsjJNS0KrzGWOHCyBVO6MTp5hScPlkcvb8xrjG6d2KXECWWqZlwWLBQMYIaFE6ymjX+Oax9KJB0APOUBNvm8E/hfMzSGgpEQJR6GunMgq9xXM80L82bea7q0befa0j/KerXlmNF78+SRco9TqxvhJjHqWBU0uc/mrm9L/ismwGwBsaVBUc5r66yCwPOeqsXYQJBeV3AKvJse59m9zdBsIDDUUjaCzT88/Mjl4wQN/YUnybUkWlKgrKLyM6Hscd+ubWAcezW/twv3N68qQdEl5PO+3G0ZetOz04vm3wfGAnPOTxYOzgHe1GfRgNclRSL1OuLddWThlvXil61OKB5uRBscrt+1IY6HYbcHHoUNuRc7bgsDD2Hjj8Z+3r7N43E4fe+KiX0N8V60bljhn9fFRNFXu3usHnTI6ZGSmqJibARs8zy3G2aExpKGLRlkLOWoE2OWkNG1RT2GVGhJsj+XstDrrRbiTqdlrpsIFm6R6liexYX1FK2qvNGu7TlpSuJSZhV4qFxOvZYrVwlaxGZpACpWg0dHTOGaaOvWujODCDWUKVkHON+tDJrzWQ4rcMr8d1+cV3zSXFHiulLdQANkzl9tixnTBOyRY7L4bHr+F1KX6OWM4empOErfVCgRQPqca41wWmpKQkM4SajVmCVT0d+jO1E2gfCu7txnMV5iMrCZ0NL24av/S4bGE8vL6Pm5OqMFTJ+jUXUePXX4auBAomUFnDq80Syau0D8lrT5M90tYLckt6gTfBM8Uzg3//KM6iJaarLg7P7UApLJ5Gut9UUBUUo4gEMu3dU/lrd+g+vi9s2mwRdO26wcUimFbnAztYO8boYUawD0Kgazo47dKssKrHucMCJmwZSgrUSQh2DIIfmATRCVIetRjjzLYNdzVioss01Ww59wKRWSP6g/f7q5WC1N39otFjSLg1l9Q9bYYWT0lLbsGo7RX9ZD1u+svIUc+bAr1qGIyHnCdMBgVvncDDEMmL2DKEzZCIprSdwW5ZZRU87MafMfAp9CP1OPcTLiPOUcnR+yWFwlK4k3UUAwcytHjxu34Wu8C6zW50FO/IjjqDaH8Z7A5leOucaMRKezD6wCj0eJt5Q1tTj8HGbG0pJ2XTaLZcIf4RPpT4RNZbXEadkOFDLvKth+teJ+G+GQxUNVPCYEtNFpxF8S7OSs0dn3RrR4vHUWhVk+vdWlLgEqUYE5tOepH5xCIYjEhiQ7rSpgZc1bSFzeBh6SYK3rdMQlRKMW7xfsCp1+Js3N0NnIcJjK8KrvkeyTVsuIxaMk38V19CrCjbiMkcuR5reAVkpono8EsD/9h4MCqFFmjF3BtRIET/FITEJxq/FOehdJBejgiOozF0IleOy4DbFGTzt5iH/FukGivJf93oIQ/eozdtQc0eS9/3w07tyz7AIm0TqGj/thxWKb1riv+MME9HzTE4JJjqKrfQAO1pSAkTexgTgRAfGIrT/sjpZ1UPZ/J1zXo1u7laNuTEM0Mzi20eZV98u2r00Jw22TqhHGfXAw2x9rSW0bOj7lz/POQpZa2e0IjjZF1NNaHb3itmD7xpuupyCxWHq0462uyQ1NjMf7BxzAMgQSdubpU4hU7PDfhEQwkRDjp3sO2O6eMq7u4+Ca76LV70gO0iE6NfHDxh2+wsCPoTRrQOw3g6S4JqOGeoRN02U537rsREvBJpmCFaWOOQXioX2igpBnukgE/TDg33mDqVrMetDYUIKX6+r5UNWjhQ7QN0v7k9fRRH+zVR/Cb/+SABaLLM1IOvvL7n+yCNwBOHJYCR+1q1UdNLXBmJQNhXpVVRYeG9DEuV2HCgcqGFHAVczFNuoKBN//r4UdLjok1hBsnfHn72WEdewmK5d+s6NROL23q5vlNaTYViisx9ZlGy3YDshquZkoo+f8J/btyuhm6DzZr2wnp/A/o2bO4AuaTG3L2FoDAlK04hNxv3WFXy3RR27S3+L2tXgts4DgQ/J0F8gPKFxA8QIj6AMP3sNauqm03Zjo0dDbJzbWaBZTjN7uo6gLTdvBZrsjBt3Ve0ZIuKXK6cA5X4JXuiPFPQlSf+S2G4LszvSazHQoux7UfoSaIdphvaJa8Nk7GqIOYwEtuCdo3dG2EJvHC8xZlnrgH6N5/s0/Ydw9jsFndHCo9l/OpCO88OlPfE/IFGKWBqHgKW3tQIB9oo54C+mfmhhNnpviR3V1kG2k6fRhPOhG+XNqv73dRA7PbWkZupt65t7gwuPktO051rimAg08jYRmkU2k1R1dhv8fou8vLooBTEzS89rCpX+Ai93PT40bkRslC52pVu/LOZjFFiJTpY7fc/gOrbrjIOOvO9U8tW2b7VdL/EPx6ydOJi6WqJCGADFcXUmc6uV+I44ClZ1BjGLj6Y/+ZtH0kUPkyn8bdIfmcv0WjaMAHCXg70QC1CMzwdmegzdc81Gnjs6DEIEgMSajNezYLW0E6QQhGevK2pR3+tMb6dmRT2ffVIhKItcq/CnvZz8Fqaojz0LXliVHcEJ+ijNQK+32CkzHFDt5fuSvcZLrUbi6U+BYwLve6AZqZsHfFCfR3pE5ibuUfa/YRJ3t5D46b3LkBt55kl9FwPYwPNXiNgyNYT19zMyubnues8pnfMiYfe4nCnn7RwbCpYl7PcVArc4zd5ICQ2wtUoxGi+dqqbUSQqua7s0sgZxHHjkAFoqimmq0rV6g6V+LTp7t6g+HMnLQKRzFl76PlrzNBegz2mIxTzB4rc8oR29ejYOEjtWlNRC5OdC2XO9PWhtyu87bbM4A6ja2Mc2YxURQoFtR1CitVPsBgTcNtD10ZO2+1ylkX0t4Uslc5pC+6j6xgcGBJGDWZbP8kCc9JEGUrCU0t+QG5IWsSbtlFrB1cf3GvIzJkbgafvXiWAbNJSm44eXCZV2y63oblql79n6yd2tcfOVLnf4Yrx7gJt41ky82A36P/LRvZZu+G2LfenQHqN29A/isWhFhxTwaZgcp4CgW3zf0W3n/ZH6NJISVIRh81Tk6iRkUi03VxRJzbiDzvWdnzv+i3W98KMTQJ9kzn5mWYJ5vyatLub1y5V8lv8FV1eOXusn4W3lld92QtFnvyV8jbJM7A9ggtSDtrlpqMgFAbkDNbJz7rdbUwem4LsQMXc2BpbR+G3eGd7HJZ3P41IcTlXsdTJr3GAXm2lFBq2yQnb68iwWt+amfv7pkf1sTIfyjaD1Rbf5jUjtjKZVByUKwBAPNZA15avNpCIbcddrvgRAiV1FDsX+6zFVXBmr8UXOp1fTho9rp3UliLQZuqvMDx/9eWzbnFgvc5/rkRHq8YyKsyLjXvJOYGLtxQgDVO/2GLdE+xTJksZBq9NyhlLQ8na7sOBDZMF/xFwqdEZxyvhna+W7rW4leJHO4p/9KO4hLa4KCjUOMUhwaMf7mzxKNORofJRxSiP5fkgAINVIyMC9degseJLpfUEkYlkfB/b10m+2CaNBHmBpDL7Zh3azkma/UPviHmj/RZfM8Qel7MSdLW6u/RIhEl52V9hk/Q1QkCrxbau3d1jfcNiS9PBv+NxMR3ReMn0YaUETJggECSjMEtZlIfrHrnckhbAxth0tIeO2MNOQrFh8Nm2z7zKnO+EUTSYQh5i13NY8lc75WAmz07MWuPZB7up7/d5uO+nu3QYPdL0BNU8cKyKmonMQwbqsyTCxIW2gcDlQROUOxUXHQTXBFOgk+AHGEDsKISu8UazndAVl0Q3e9b25fv7tOiUBmZy9Ehd7OFL6OA/YVA8+2Gjbq8flYfiqyS3yJx6onYZdLuFBKBl4tTcfoa9KMINGtyDnu1eDMqmxw4tMc+1Ytmxa+vB7aiuL8iC6iq8XfYDbriGajFarNNk5he1FLcLRw8ixgpQGa2sZt+Fzn6LTWX3Sb+WnqqTnn+2bFNgS4ygDobdTjQZJfzOKFE8etX8fggPS5EEmrYu6e7cqniLuVRi11Zt6XH76TbcJ1nhPW49vGeLSObkbsVTqMOsxH8SVdJbW5WHtf4i4zUGipa8BIYxLEhNHpoVdtCfvgWlGcT4bIxALDv27HyJPRtEnPsoQj4QQIoLb/F5kLwlQZcbFbqr8STmB0c2t2OzUhG+/UnHtM1zUbhoOaCZ+ImJyb0UI01Jilx0YXJMgcfgNrmxUpY15kZmG83X0KARzvQHbT/cYtvx6+hhwjSYyZ/VF99s62FT7RzRzDX2wy57Dl5hc48OfMOfGNUyDxhbeBUXyEYTDX5oBJ0mJX/lxVo3Smw2GLBh2tsXKWYgn6F8Mdujxi1Hr8XGVGGdYM/22/zwfi4KnThvsRQb42R6Gm8lhr54kB+8TXd+kR76pPh265QFXhNlmoxWXBinhG9FIR1s2orsEDLdPjZ3ASIrXpAllDOLHjQrwMMtJlmFfdtPyKc55xbjK4U2cCC/To5JzA87palnrc3vbvDx7gIsLWkY8564AqHaSli+LGYs2CwFp8w2AuuOXcvQbpmSF5jT0PyAPYSNbrt4E8LaxI03inw3/SDQdnkINP+3rYdbv4qbOQdW2xpjcmlnta6+F33HFkzTg4PrGKL0RF8+yaERuWpMGkb/xiRAwj4FBrl0n3CqYN5T0zgvoP60waPA/UegBLsGNcPeUVS/w9x7dOkdvV/P8gYyNlCySjz7fjQkrlmZWE130AGg9e3A/Ge6j6twjZS5ad9Ba9eN1DX4gzHSDi+fAkUbvS0Jk9hpi5ltiVSNvpZtjcQG2fpi/bD7Enpp4TS/v7jFXJCeeIujxRWHiq/oa96ddHvKz/vlfjqiQSVFF4rpYKdSpKzrHioLDVPQGmcGJoEIyzLc3YgtRgKWrk2fb8WhSsxBQEJmS4ZRWC3Opja/P6Ext/FMyw/PgTYOt2ysaLI7j+sO1WFjss3vqdspFooynm55+viBHrj4Z7XTFSElefQizMFGG0Fa3e0tJZfkQFExFwN8bClqXZw6ChULrT1+ftQXn4K0XZyqEtceWt1bqfiyKLvQVByhn1fVwmnZ5ehHnJ5mBm5Q27XeGaPcBO1GhslS6zawaJInDSwR3G2UxHgulVBaF717Vgt4qatGvOzjnbGt+OABpLj2NLaTdnfuXKPF2uxKhEMr0b0bRWd5g8WXF6NxKccQpRLyDmBxt5lHMT7AL27OHoxfbB7EgDp5jSsnDfqM7p4owSdsUdmojr1nGziyLUbtoAUY/9x+rw+WmSfIzK/cepTJX7x57lKlafVVv8kP3kZUDfhECT++EHiUHj3DqQ7tBEIPmss5RLmoIRIcmCdYzwrMUs2gSOxbeNGyMQR1wjm2yPq89nUohNouN3OTP7Fpuxlc7MI71N8Q4DrkEAfpwas376BP6mv+EjkT45HjpUtUh1ZxiyvcwhjUOtVNlPiNJmFBIiq7uwUlg1s7YJlyVzJmlUFrQuL9GrN7XgAYY7wD0nYGJH8xyZKkCHYwa48ajZ5htg2dQ2jgO57VNIYzjnSVR47xxvbNUiUqJXcwxARbAnT41hjLJUw5VRvwH7dybZeY99h0ScZzJWLR5zoViT0u70QbPJElH5ceSQ2FNDO9k4hhxL7df0PdPhjbidxaHtX7/lm5XWDt+NkDw4ctUf/Frsxm5eRpSXjY4JZLNwQznfCRYrfTzfa+9bbNdqSIxrQV9OXBbvDfZebXQJM3/egIAa3mEWYg2/qRCCxNQ5jdI3ZZWII3p64Vdm2FwWD3MyxNO9O8czcaklYgbyJNdONc/FyCjfs1XuyETYBQ45GyYjhUYXW6diLFidEp3771MKqKuPBjU+EVmY3wMcf1T2b8wAJKzwNR+i8bBiHqdmvYaNnYbrEZ27VTTsw9cGcasCe4bl6ERWx2xnvtA9zu8FrehyfPaPKdSHFa9igJAy4fDSrowX20t8LGj/9w6xxBtjKaP6fB6MpcJ5IRMskpXiggXcQnbm9fdkwNhm3NjgbECSid77Wi6EjhYN51M7rF+1CL1TSLm7kl1eILE+Mvp4GZpFF4nQhE7VFBY5t+J23Pf17kJ5TiQWh++FpsqBQwLudSqXFdYQPdaCkFeyVOHimIDdwQkyAmjlLVwjR21emtnV5lYoTw4IXVkmxJo3PN/99AXwQX97ZNpWIaDEg98WD9xGb70RUzRWOg8sBk0ReD0WoL+zQQBtu0waiODVtnGmESkhfmYy6kDV5rd3mTZa4TfzolRaNzPTQUjle0/3pDM5V4N8zQ/xgYL4WuVcm18wZjwZh6lJLxWf7SipZnFIlyzJnQ/RXKtg2MFXJWTH2r+nAv1DucU4jEq1DQhh+DM9dHtea+q6veDfumX+2GgDauR3yCbnyr2/VENlCICvPZY1zdUXbAIfrvgeOliWN6ZpeSDiQgrJQamXijYwoT7TbqzbeF0eUtt8ftq/zgtROtOVvdrc5D8ZcuP97g4TKLJv+bWzwNuZknYRQWFWbTXQfl58H4brLpeR2a4vXl0s6XHZG8NvYP0UNsY4DoAu5EFjcTnXKWyagHZBMHKrBrxMdieDEZ20QujTfhD5yN077u9zosGGhC13b9NcXSGX0xfS1uo9ZjnsbUwNn3oO4/sb43bjx6eaTpABYX9xALbVymac1GA7wMy/jMEEbmAYrm2nmBk+bnXT53BNgaDbP2vYa9crVv6zqQuVuBNqs2Th6Xy4nZo1ft7kpYfBzRie6lGwaP9QOMLVzj8vRTNlMdMEei4Q+JV7oNcBOcdN2aTZIvIkHeGW9UzTQ3D1zjZILQPnX0WxxPts8haEFMs3QTo+1yzgD9HVIb9d4JGD7c5Wn2iJ8PLD7SgYdZRjfM0T5MlXgJAumtkdZgCL3JI6wq1a52NvEiELPRtJUM6BJcA96ta4jVId5nXWArx3QcvBxK8Qm3mIF3/Lu7TnHk+JodXTMHivdpgb1OlIFEVY4Lfy8RWeQUaA6g7WgXGYFK2OovToQ3k2LFk2f8qjHmG07h05s8Jky8Hwqvy/jjhnSnN9BS6+15EPQ/MTNdBF2cxG0r0i+RtGezbYyE4j9atvTS2uqJbUrpBaMdcKWtVYUHdEE9hjlYgh/N4iGtRlxb9NTJC2xxRZLtnX09l32SDvSJcKnb22gWTHA6jzTuf4XkffIwNc1Ef0yvyK6ys63SX3LRkUecHtDMB9l57rD8ovAZlIpMrWjbMsOSf6kQ7ndVx66cKjx4tMfk2+eGuXselxveyh1aN1z53IxVbr9m+XFWXwww8xummcmfuzWSiR3DDODl54zXMi4/jnLoaPysSEYzet2Q3kGiWisPu5XkSnvtDrOhiVh0U5f/WLu23MZxIHg5CuQBtFdIdABhxAMQpo+9YfWTtGzLCfOxCwwQDMBptLqr6+E82LQ+s2KY2Q8a3TdPTqStF99I7DEtpPhrqOKFGcZ6SOo8HBkPesV5TV3/HeiufS+O0qAJyIRzB2waIysRKna83GYKPSUJJZ4BeTrYMb1KPmlR5uLDDndGTfH/zfq4kj7KSNttmhUeb9CEtCkdSP09pIa9V7yjUKzvZorSeao8ap6TCXAxAKOWYyR2MSvqGjEl8upBXvzywRPiK8xTIqwQqn7hqmNKKIRpRVz7WubrXfvsIQSod0v4M0vex7tKFY8OmbRD64X/Kam4nBOyH4TO/MAxmHkudeUamBkPI4q0p8z6A1w88q5OmFFPHmQcWElvoLJxM9nuGIL9J894g0zO/Hf/3oTUNlGxtIlfv52gw6qmbNwfls7Fan1t7pEGIXkKT1bnYBNxFppKI2bDWlCix+Ga0tAfmtMYlxAVAsyJD7iYiwdCTyQWe0w+bzygFHIibZMfzcUcBT2HDWQgkGGZ7lMnfO2wiFUNHUivisCSw93SWXmzVr9xLmEzkSSIhpy1Wzump2Uf85pko8PCEdkAmpmWkVShVdBhY62dV2+HaFZcRyUSAaEy2xQYiHMnuIrJwXgweuXtQw0F3zCLB3PtdGJB2o9r5ogQmDrRUMvCx9BM5MtMwI9YugrDaj/EibiKVYLovkR/eygHyK/MYy2LDP3O6SnzolMar6jvxUGVM9qGJZZxXcM7E90H3/2HZ+6IK9J8M6UnxUAamnbCa9Tt1i6icNkwSiQGgKJOFGRFjBtSPLLhaZx1oPuHRyQexjaGgVJTmm/3baKX/EZDmyp0vYPxMBjbxe79QFF6PaPH3Mrwf428zDuZQXMZE4E7NCRNzH7ER4WelfW3WOcKufAL/ZJXNZl1jV7l789jPxZHPNgDCUt+ynbHqwdLG0sSknZPT7EqXt66T4xk4pKeSZeIYZWDoMMAJBpaUeHbiGm4RIZ+KKQqWHwdxYg2WR0NYLtoY6qudP6If9qB/RePXaJRxbR7zPrc+QXasYGGBJqVu4RLg19fXDnwhUsnLaL4y38075TmYd6GCSwdCGPMgV2rMoJ+ongs5cyOxEjvMWdXkRzp4TObicqh+v3Do/Ed7oYTNHzEssPkp7mqaBpb0evy6kRKEgYvcUrr6wI+cUtJp1pS6xVtkABjZef0OtTzTx8uaMSCDFs4IPmDwZaGUuyantxwCO0ZR98d8rAz+3em3yIl/6immWX5obe7fmzjC7TMxZ9c7ZLZbZeHnSQ681FRIrQXLbRHt3RcgtiimKhku9rR/zJLkwicUO2czg7+APr8amf/LnulQPObnPlvs2CgzaexBTswG1+bq9hHwq8vBuEnsv0SRvRYD6ctUYLOdjt5WdUWdgmTZz4ksdI5s98dZUkA0RSjRhw9DseZ4Fft9rv6dDq24AkMxl/bxCqWrG3JsWI3D89REfvGtz5Lzo8tPZd9ucNzVEfzHYf9PTCBjSjxgYBishbMh1hjRrWfqAT4COWVz6FVIcuarTMoVHScThQ//TqkKsvd9jVxLjajczakYEDCJLlsksnRB8sl8vYz3s9jHCZmO3hN/NRpQIxEW6mBrzUiW6CvG9I7MuehsCn/AT+aWI/a3TorG+bK3tFx2J6NFmLBxFHQPU/+7+TXLt+Vtg+fHbhYa7C0n+WleCa4nLVkl2fT3UU+iVL6F4jawNgKvnNUxdFZ8FuQqLbjSimXzRXhcM242thQD+Wyvf4RKj30o9+NHzWvim8aspQSO1Io+LPaEY/pKhcjXDs1Uno6eARNROFXhy6syAmvItkWq13hzYMOHm2MyBKkJEBl9iqvQz3Y9HnfvzLRge7ytZsmMyelubglFB2MdVaTSHhD2ZbwBoyX+8YDEehcCJ2Q5YOxGBRXOJDuVfY7Vexz4BdYxJGQePaA50iqbBLcwUnQLMNe4UCs0v3mA+mUz9325WiD4L4WpaQ4JEg4Kh3Ran2rVCp9jqvfTYoq9uFyR9Xcto5a+OYcOTGbmnEVroo1CpDY0IV3OTnbOdlq+XiNsI1VLJnxo0n0nEbh82kcF3Nxw5pCxacdwyNs5ZGj3Y9w1QP1FIyL4aId60i8X4lMzHLGrM65Us6I6sDMth/q+8zNWE75LOK48sq48TWGsToYe7+Ev2EU8r3zFO7gHSjEcZfGtuWKwEMPoieRrqbYT6T8IhoQ7kiY3NpgTA9OlzyHEAu8plBmE5ez5XO/Gfd3UKW9vnpjApKIDHTfZm13mzbju5kDBQlRMkOgK4RibcR89ijplCzopEusZ9xhrl121ts0lK3x2iqJGbO0CPg2kppGTIBEM3NY0rBSt/VaeqFHSEfB6gEH45mNgmIxN28mb1EHNrCJ8fabOLAz2PgMB6o8SxTY3tH5OXCFhkTGKa1LNHPBGKNmkpNlChKsdnIcFePy6Bko1oazffReNmXzXLnXjX3atrlDW3e7W+U9HVR8US1aunmtDHyVhzv/TkZhACcqdxWgbO5Rs1g1sv1PpqWZ1frwn1Azfh2LVXHQ1+m7foxQhO9vXu8muQ2O8tEi3GGLQNA57nmwxIl+pgfU0nNuN6OWQa5zbLEN48CgDDaxHMU7twYMt9HoTFNqt9zJ9Sj7SePN87ITN+tpvrY5R/4vUqyDDaTy0dUAtkWZsINsf33lodJZa4/Zz0VIKsQcTnxJIuStoIqhPQCTIhPORo9PebhwJW7jRKziO+OqOHfnuod70pt+XIU4OM3QcetO0A6S15udpfyo2eBF7X7ys9vAY6udQcJO6GXhYoVSiYBL5sIb+ydkgoISpTAeNFRUTpOwRqzqmXx5LCaefLlbiK4nBP01KYzZKmYm72kXjksAACAASURBVEwpgjllXv4pmjRzcvmwnHga1qSCaxQzFRjmhkgkqyCWxPzMkv9F2WoSOaysNSVQaePIx8Wf7AjGt/usSEGkWOHEf/MCXTUhHTH49UUkvDImyonKXLDkwvpm4weStV0it6XMEfB0dG7LBll7CEmQi5m+gbDf3+VlVad/yJSsC3O98tDVr3ebJrzOmSjMXLc4LT8tdf8twR/xeKpYro3I6SRVKSmZmIc2PjmTK01DMnckDyfYj0bRzpTdXUWBTUQRGdTq1gy/yA0LdX3XLdhlDNe7bWIVMzXz7s0SXP4Mn0YvIfFp4LCV8/m4MG5BNq6FP3bEVmmfv9aVq/jTBLZoZPASktymvPv5J0AvhisCG69VccqtbtmrFzcPTU+pGgM9SYiwWRUXRzAWbvwqbrqrj6ta37gtOf5EOcUrPEuw0ScaZAm0HbE+bVMO7K9EI7FEgmEEppBLFn/xRb/mQdxcB0LK5XfOlhr/9TWJJf+lKcUGZrKiQ+Iv35qPPhhiJvvePd7/n6OazSIh01xMuzLnklsgLkqbjkmRzKvAARIxOTu45sMZ81/+3kECnZLclbZt2nanVBXiqlgrVu2M41I8eeT0oFN8eMvO5LUGyQ6sHC0BwIf0HE1HHihEickpIp2hmwdnzaAdMzacHfvH3/U//Wl/qSXUTNzubndXxWyGZ+mt6yI36QvemHIYTcHno3RvHqVis9u6M9BNGL5CjEsafSYCpZ3zsvlqR7h8e8YER4Tu8Fn1rG8Xj3qtF7ffDEQb/Km62zZlu7sxq61/Yaf2COyT6eR2r9DM/pxU/D3alTTn+DQECOK6ABo8CjnC/adVbuXwpGZpzum4nF5H7triq6RLnfMSPB4Y8Vc/eIf6yc/0o7g9rHfsrq37s756ePm44SQeMD1q8UjLzx7bNA7DNiySg3wBxIYo+Oa2hDjtnY76mDJAsoqE8UTzQ1BHczWnqb/pExqeQjT5Sa4q7BLdLdDssuQwCmoVy8t7UglD2aYXkwQLk4qUdYXpBF3uYEFa088bBnLxYBVNS+xgwnbkFxaQmNtvNqarsSiufvGqaJYwt/VW53OqODUwU5E2qeL1o3iU5BpFKd2QPNrgZciRoBSFcXyF9ACREu3ckbkVU8ZlooyqeGAePuDBFp19oCzQJnOun5ZxZSkCzcW3WedRU4UJTT74E7SLtnujek7n8pl+WkueBeR2kob60FZHkeWFnTCFatXqmoe2wCYIO+s39kOGiT4LxV8yPmgU5Dj4MxffRu/Xv1O4CZIPZpcQ3FS89i5A61PJs7qGpQcSt15FW+KB1jEZCrKxFcdoE84WQGeTzFYmxVcKRzngA50pwk6GCTHQFbbx571YKcZ3SVlyWv4/rh58gLYLtJqEdQPbSxfSFBzxhzvD872uDWh4/khxgYiVSI07UUFyay7mMcrFjpnE2DkCSZ+NnqKSg9pXcB3/4EqvqBI/2qbYr7lJYUqkSMFdPUwzejGG5sSSWJ889aa6cAIC5AbPNVw7KBkXH7bS+gWlD+9WxTs3YSwe+6HpzlqDh1olfLY4Wx5QAylu//51zNcpVUwDhVugF7l92MC2vndnKz2ZrZyTtvGFi7RwSIArLvtspAJkTU5JClHEwM6urdYQ6gMNAtPfs+vDv9vsRBnSBmMYMN7muao8xplTGPSipgnBOIPPvaCdwd2Z2rn/EmIWTpSYjfMRWcajpNm4itY70X9h76AwO2DEuxzpJISRWfHCZHNczHp8hrW5IKttCtLm7BKSO3uslpQbuJqv2E+U1COW48lj978RuWFEaheNx1Yj+IIEY7LmmbgpeSfNAc/ByiPusZ56JpW53ifoBP39zZ4fboP+43mUIYoekVdDK8kOXC8h8aPSIzleEG6fDR6GqzYsihsY33y2sSAXDGxt86CdDr3YyphzymMr4/2g3Oyssi8+JclS/bt3plb8D5qlaRbRm/cl7TqFgsUXDCjSoAstQ+xBj8GRdSPGNfx9GZgb+TfmTASKUCVMlLB4asUUNiw6T0Ym2FZUckRHQ5qPJoqjJtrubgNZ5W9VbA7RRhscAHkd2Jbw0kelDJZ35XTGKDviO8jZjnoCjceZGPGccxnblcnIgmShgqeMDO/snYix1geV18edQklt95miMIPknRneanjm8nogLid8ifMU7bE1gzeRMo1yuIBSKjx5ke5irhRItt8itUHXbmr7nTVf1QUEDu5Kv4KLD4tuxG63zTvyKwpU3Fzss+xeETLTqcfoKGDqjWrgr63gcI00JwPChC8Ys9soz3lH6tcOn+1258BUbLilDg61Hp8xis96cajbvW7zwtg2PvLfhDeYesoPHUkVoFgu8QQJsCynUoRIDrpMWmvdInOodoMuK7NWCuHItDpnceLHyZ+DJLJEZGumaHWBHfW3RVwNML7NSzPHgVQh+WKapXVxYsblvQosnes6/B+lPP7S/6ydUY7kKBBEL0fJHKClukHLB6AHDoBMHXuaiMwEbFdVd5vR7OystNoPhk0nmREvQIivLCsvU/nvp3N1NvuowO2gafDIGq6q7e8LzdAZhd2ldJj7lL++PQxg/FgV7DPLoDs6+ftX9HB9P95PMnP/5uhYYnbNcY1xoA7RuLlKuRfOfbDA477DXniOc0xKignQ1fmapqwJ4qOPx/3TIEhQbTaU36bd4k9r2qR4NjGQZhF/PK/E/kQemPvUS9dExbq642Kp0p6ra38RCTfWzYuXNejCtjg0wAf3HfB4DGbynVfp0teO7NeJPIrPdQid8OYfdRJ90AWvPS3Fvq1C85kC6PCvJ65HFyrlv9/MyLzE562UzJjc3LCj6tiHQVRjAilGEYpEGtAI8dKMomxNJj9HNvhptdgiEZwt+Bvd8Yf0u7fr6SAvj8BrzXDG+sILFf2cuXhmACbU2klduTxP1N8lWJLl2EBceT7b8q7QPfqYJbhCY2KgNpgbXTvlFpN7e7MY7cY+Xl7LvldpsgQ3P+n3r5WslLHrL/WJgXXHgt44CAlIJIMoEFUntCj7uZgLiUwrs3y0wfHf1qM8ZAu8WyfJBttIvpNmSsfm3ECMvz1x7vvWVPjn4woZ/MCYy2jnAH8SsPyQxUNxxRg2YuMXkcUH6FOkQbavXUo7+GWJMV4qFnzdJTWFfU41hZnXo+09CDW3INcXpfjQV7wy75szd5HFncfVXdhYkMO2QEsRCbNCQ+HYpBV86NIQgqtpdelaHVaB8sNiluYkIuDtYSHF7Yz76eVvKB/5yHXVuw4N5vfzAs84TDAZAl9H8iTmcuDmCnoK14qFj4ukk4tvRsWC+vDYoaounTKA/Y/HNMfSNt7ibprpXAtKub1fduSmz95PMbv5Wvv2Ba8pgnxfiIZNUy8RbxcMiFkjPOBAKFzwD8rhdokNJpguXOKEt4c4dGfVYoNm4oT9IP3pfDOvKSqjLyyfUNlGNCamEinha5c9ZD7oJ3iFudSn4xkwjxCJ/1kU+qN+8p9J4H9VjDUKGjiKCRvord3ijvlhDlJ3e5+o7Q9lOA++MD949nVOwVBRXGY+7UIWyWsNHE1mJoewrR5qxnSixs7I01knP20dXeK1OiEKOeko1lmmMMX1r928uPH47fn88Vqo4vtCkQ9NcivQ38fnnTbGIGtXTZvkDuegUkEJPGDwDGZvKbKVwFKUiviiKMHLncS49iDkaiLhyqJT+LlrUdA3d3vrxh2XG0MFPuI9jJSZgnifQaxxhE8kIajw4tKAUE0Hi1biOiZeWtKwjSLUg1uunzMmbUV95nMSdLc+U7DTDd7azRW3x+1Hy323O3K7v8nQd1IwqiGXsKoKA0IEMcuEU/tBUAT0AjJxijoD0tFlw1iV2PTFl25xaai2aTOKjSP5be9Ycs6QSz+YsNXbPECBdm6l3qXEK5zYFmfP0pyoTqmceEl5LrIPDQtJCdRhdhDMdEzouFwmUIgeShGbxaPYmn9UsSpdT/F2VOxPx5m7TVKzJIHwGiC28tRcwjhK3SD30OZ0FgGQ5A4X8OL9SKUptlZq+qoUr5y6hoWto1LlEriG4Sm25denBx/Qtzck3ZFo9+Q71+61VIzvG49pscJrfPAKbKS8qpTiQ8cEihJft6TWRVjowcgWvfq1ix2HexZEbMPzzj53vYh73IzeXjn3vZ10HolLR6FK/ZxxeJy4XBb3PqF3ngpuiyUXtyhfzkUH8d3UJpYx+OviD6UDrbIcnafMHMO22yF/vNzu+/2Wf+8pP3F9yDiIHCAJynV4NMuLuT5IwI1HDFh0mpECzPaS2jJez1orw++VmOfSTPHorpOAjhtfMeuxULRe4mcj4uE3vj38vLONB/b3WHB4ATnWNgIhP8EJLl7ZKYtx7+psIkighCxHyz7V8iUL87dv6MejrBMl3DIKejTaYO53dzfn3sox/QBTOQlKcf0bjyskyonrNhRxSkkSlIRMTPM+k2egv6zvvZh0GiFwDmMTN/Xr9TLRxCrrRHANk8J2fbG76TfvRUfh+7G7e4Z6xaI5N9K2BB7IjA0ToIXvvMLqbH5nqDGhsIqQFQstdwR5pN5efvmca6Wg8W4bt/wXYQk94qqL9viw1sK5N1Ro3w0k8kmZ1geHBNrVFqLuoKt42yOJsdCrZC9nMZPjNtcJUI7ywFNQksIB48y+uPeFycpjmzJp23qImG2gxWju3m+Usjtu8g8fPG0oKGklkyJXjxJW+tBiOvoaYRbtYFahMBwFVzWkFjhcbEBcYke8++WPrwNYhUCKaYulVebF207DPUA+3ubC5z1Wd8c1X5xo10xi8d1KLLAvLuzWCuRWsvNIKZSlqTLhx0XswaKLuWJRxCYMjlM+d9KriJ1mzhho/Wxbj07UZhGNzyvEwL/Mzrz7/bq0/xML9NeBZYW8qowaofkoZCwV5gc64QtWU3lIxqkqnMCnGLuwwDS1TOgD2tYe28zd3TCT3782Pl6+nPPBc5fdUVCx6BA5wR26LH7xdSpPUE1tL2BpTGJjBN21OqLDIjJBE8UXDVgrv/bW/WAO9K9altaRrjtvd2dbj6aRf+M/6K/umbtDQUs2L64dW8icvPmFCqDcYIJQC1azaI54dCj1LojNrhPFx1aO/3qZ7/g5NsYPNd59zhJcMSt22+3uPpqt/L3HI4+Pun7Z4WW17+lyLoLKDBC94neFyGKm4wpPMIoYU/IDI3M7jKeUUkvpOCQO//5r97WrxGIfnYT8sHnxw6SZ7mckoP3K2Z+mwg9/BkVYSx75rVmMjXaHwd0mMcxxxBZIqcGsLUQLbi0DojgNT7t0fUgh6a4rnHcTmjaBDR47Ck2W+Lg99+Qe1kZ+RFn5NiAqNjKGxqo+8io9JXg54bpYQrQdBxVGDas1YpH0Dh6vNsQNAZ3mVWO7xY9Zw0xx0+gBW+5EPzJ+Nmfrru0e8+FHpWbQzSj9HWnhOom7O+Ia6yyNgZd101E3IZJ8ICbRsLPXtcHPlZbi3v3aOREeEqG7zouxao8P6yhk63x7OQrKXcN2jBc9fhiDWu0C6nIKFX1HJRCcueA0kgQEY0dVWeUkGjYziUoxTjt70qUKcR9vMYkU/bj4aqFoPoSnPduP5UDHSgJLudYJYlOq54u2MCFZgfwa6Lbj5eUQCBA2Wp6LTOQF69qcuDP74tjd4lFhfLFQfBKrkus4s5sDvX50dCtRr3EeTSuYeys0t/rMCsQJs6moYmKEweMvGc9zxlYnazQsMbVjiYzuUIlrstzWX2FTjs3E/TjLlCHFTnF1UcK9dcW4/9/dgMUvQlL8LlkiP8+XkDrh60iNjMGAIRumar5lZ2fYk8Ki7Hg4wcTsnJT8nGYNie+7v1OYuanxbtIYqOGXXN8YO/fx+1LxanRf4M+VPCUneRKBimIGemgSjUIGk5yr6uI7elUatBOXxj/30UD6SEXkQB0k+toYyD53vg2Mb++0bCPtNY+qFe+aS1c64oArb9r4Kp2oLtIgBGihuhrSvI4yOWkrSLOzM07iUho0Ur8HAZ0eb1TSudpHZ8ESWkMhpLbcCsVTNVDeAX/yGRZB6FXohL3AzAvTyh2A2tW3L0JXbu2MkRAl2g5G0ZgDFYP80ClqKf2VKXgfbvLXfqCJWyzA/nkSbuKX6h0e9h7vF3d+t7bz59ZcfQliPwdKcdb8y8BVaKhnjjUHg+zweGaQnZbhRgVLQ/rB5Snx/ulRtnmLpVVzPXjCI6nt3dYuu31VOJO8dttRIhwBrCl4O/sQNKFKlh106Dr4aBbKq4p86pJlHBiQzeSZf2oo7ueHXbf8RX3m08ZA9Wu3tTrR3eOn6LDshpCqfAwO9J1+rThCwgiDRmhSJh8TczZ6nV2QyCowMRfScxcJvhTvgTTEbX1XLjYTX7t6TIfuQ2jyM92j65gp6DrGx+3lIyPvNVb9RH5Uq1TQHYKeg4pdM1hhyA6EA4x+59peyAcw1zsFIdDSZmw2YUsxThtOfNltFrfHum7zbvEnxa+r6Cj8frH0phC3S9v5lg5OJQ4lEsUpYJhDsc3JD3cePlgQPEcTRcMjyiKSeLnFpRtOlPiXduLe3+N7d43RedvWY9Ytpk6eI/n+Bf0q68efVIQTX2MVZ4uXXMbGJL2ib0vfFYMhNJiyMVJCCB8SfwmweTB7fopxAMdPfzyzVDBlaZ1FuNrUd7dDfgwl4uP5xqNXsj1fUXspEyCG8XXnuNrHui5FCCiK06WH+EULL3NZksEQ2knHdEA3Xhyz2WJJlCo44ykG3TYI8jSF+e6Qn3duXcxa7jfPfj93S3p/uRTl644rOyyfMTdm/Ay8YQujcC1HqdFGU7dDKnaZ09/74ru0FPdxsZTSkVwzgTaoHUWX8Pr2Wzd2F/6VExo0K+ZVSXKHY29GKoKkt9K7D/cBuPxCpOmJgjuH0sUhxde+TYZM49/jmIx5WZnZxzaOszb3pqHYS4p3p5+QKsHEy3rJCzMD8chjr4yBvNrtuOgQhArmbAuXdkledUWi10y+feFwv570xYJ+XaexgRptMHfQzPeLu9wpL/PxYufjGg/zCMf8d8onQGQrUGRSZ8yGIgpALKJmBNVYxdM33Z/rxP10Bd13FJPco5wmEYYnhSK398bH7R1d0HWRSc+Wo1VMnALPHTEeVotTy8YVUE2icjDIjC1EoyJIgHYy1faVnZKcr9The2uLY520Jf3ezVgsCbfG8LodM8nedh/PJFZ+dI/mHmVl2RJVMZHFxa9LPJgasdbHENlbKf4uG1HomEvAFF6YYCkddqB/k1jdT+YU/bWmQbeM1JoZyA+pFMPn7q1uO9slPpTkEddf8IgGtgrRa9kvICYAh4DSKw1xhceL1kr2dfy5qMSq+Z2vSTLvz8bFkXSgf5rHNnF312qxxU48Jznmc+/zyT8327OItgsC1r5vMcowDxlQQbQVtZeIQizGMYPPpsESRz7H5XXHF2rFmdlD48yn9MWbORGofnV5lFK81lr5UXI13vIsp1v4gg5skGtvAciSI/lOtRNMKc/QFWMYBBZFqxCSptR3E3s90O9mmF8nj+hS7aN6yFOaNv5JdSvo/KMaMVr0B0bb/j9QrCg7TokB/qlGsCBSIBNQOCnIkWHwvL7qeFZQ40ULWCewuttVPn3ejSFLF+G65h5FVJj/EeejB9P4AZ1pbVxozztMJULJQjV3HAFBXdwS7ZLxwiTyQJPg4zAY7iIl0oWR/FfXst1HxtWjve7WKWOgT5TirbvF+XVr7BsX/vm74z9rV4PbNg9DL+fBPMAH6AZFDmDVOoAQ+dhfxEdSlOwkTexhG9Z1xQBVoMjH94NPQ2LHxuY8aMCbn2eNSi/mcy5qYM7imVlwttquJZHlSlKVjssfxIketRNx5GX2rXGt+rff3zHY4/R6tNLkLPDOGSb8ey99xpCROw/d3J44hdz4qElY3ITEpJlTRrmXmDXMg6foJSUdNgyKb86Cw1Gf3St15y4xS0pWaYSg0+vRzdXivlX492dLtu6Ny65jKyQwBdtYKS++bkJArOJNkqiU6jZ6lUOek8BAet4tufXj6NbdZXZXtxukm878KlHYXdWNA4X7zU6UOuTn+IuyxzI4GAUnXIsH9Pocj4vNErcR0JZPuNe8uTP2j1kTd+4eJ+aOtWvZeiaFKGp+LpI23n7cLZa1B/2F05Z1uNuJcruF/zIpfoncKsJ+H1gbe47OMkjXwgsb6JmRtiTE7WJOxattlMq5Z87ztkPPvKr+SwcNxQXMTM1YIvo7f7t3SNiPdWkSUK2eOWjxhfQWC5dNqy9+0WrPHvqJqmhMlqE0NBDp5Ow8HHJsPIqkSv777ZK++KaZEzzdTcP27r9DwzAMz13oMx25mUNAs9SNaEqqL5dslAXbZ971F4l6JlaVLyuHd5RkWzpzh++6uDNzHY/QIe62Ho/v41Zc1vbtslu8id0g9dzX/97PIJlcndh54NXuuHpus5VVXfSDl8I6UolnVH9+CfiZsUuqY9w8JxM+22bUSG3foRRhBxCH0HM03WbpGkj+5rQe4lyT33OIs2aMHnUXWR23/bg3a2mujioqrENO1aRUQez3Fx6kx6lDV0klnQEogm+IY0Mqdhzu29Z7654EM2+q9fjLCdPLD3uhLlZ2Cxdl5m4/uoraWNRdXQYBiM83iVK0QKEEX6WStPwWE3aU1UlzP73DwQ/QcVeCew73zpDiJKfNbvEAA72KQCAnK6e9zYdkXxaUkcTnqXEpUymaP2yeCKIHgyN0QQLNnFabOuRP5YJLHMYRJLgDN+sakTaef+6EP8vqEcEoXt/iHhnO1EtrBjPHLAsP88us3q7skAmzD6yTQATStJQFCGZBWqA4N9oB++P95g63qxzXJ5cYtfi+bdeZJcCW3hsY/xFso2fFYlc8wH99NBFsGs+w8TwjdlT5xIT4YT7t+tjZ2zZ3cv3zxo1DYyxtRSPAqsx8uza10QjGtNvyv5R65DEEvlUM4n5NcHg0xVD0z3BDwGJftnZTQtZo7SRmtGtVFlaS/hCtaEOGz1GBQodn7prmRy12ZvJXaKDvN4WLobrzpeLf0zkjT2+M+dmuWEpyrs4edZlUKl7BgYwz8CCcMEhAK9zveL+0rJpT3lkAfbsQjXqB41793ANteouH5JQLdnfbrU3Q70VhHTZBwwBCA0eldr/onaUw1ANmnwRQXiX/XRwoEmIlikLG8mM11ooed/q6UmDz3NCfuHaocUqmRLgGo7iLzNzp7vJ7cYfrielo81/acAfhc502BNTkFSmiW+vqQ0zwYN0Ifzaut2yguziVSzc2l+97Yrd5jo7YFtp0ZyjFFbX4ZizuXYbuHyDM/PzTGeEdOnFk+FkxNWhBAg2wtlXSnklingViA1Nw1XIsylGJBfv4BvvqsKOxhZGtIrf4Mh6Fzh33u+juaMqZ/jB7ZPMNG/3wLPRyVv1+4hevklUqfsnOHoR+rWTTKKFDrsVhgXJG0wL11pbVU+NP015jx5Bvh15UTrNdl+vR4mkOzYGmZ/pyN210/BT/reCJjsTtlSpFhQWj8Ft6/FxcemuBAprHudkyca0Wl+afewL7CfgtWqU48KRYt99fffBu19xipskr4Yqml4e857Ht0rQZsl8azoa5gzWNjGkmlwQ/TyqhYQrQAp8wXGXhqVgtLpfaANkxx579yrXY9kpXdRSGZo4Oxv+eF+LDdo08LcUePG7d6sUlDmwtvLBjPJ4JFJWeIuxXweULZElGFyzNo6aZqXxTig3yiX56Hi+y5jZ2sruzt3gzi6tMTzPed+a5XfeR90fPQgPmqOjThxQ2rhS1Fc6yttN4FFbccQQNmCpKF9R1h3cD+rSrCL0+SQtx7IB5rRPd4HG7YHf3Y3gxqUQ3v4MohvGZOvTCNcpcKoi1HjNH5GqwD4eLzrMFwnOMEoytVumNhTcBtMK51Zx85KwX7nq36JfQnb/uJX2xqxN0xAs+lvD3Htt7dEKPfoFUqYApn7lGZEzNqMdJYxlrVBXKBP9WnGixSUY/1uTGgbIdRmLQMEQ/Gp6tZiJcZ4XXjEm3bR/c+KKrIHoio8mTpwomYnkSyGwcpG3Scg2fEYNBHvMW2XWotMOcMaVKfMsq7g422D3GCYe4OvqgeTpeZPkhfCvIRwmKJZre14npqEUjR8is58pP3cLdGlVHqyVzAI1WY77MOjvzWcu4sVbLeM7xaBnP65nVqNbgGG3fETveYKe8y0k8rn4usuv/uTkpP/Ep02s6Zge/t2Q7cmo7QsQE6eKDzWrShLhR0H8mnK+VYjEizRg8VufsIQumsq7nQyWODl5RitDeu2TaxtMD9F1k5nc/P9PbEZq8W83YuGUvQGBCMSsaJ84ZZfhngl9YgldmUs/tAiqmLJNM+ex0jF8owGJviRCsJzba4LCFLqvFu/5cFCkopbgZGNOfMYphon6GYcz81BXwXgkAUJa1KGY75LiyC9DCfkvL2riXwhv0WpoTKFu0pjh2yFBwOBDoQJflQN/vrhbLkp9eE16zdxrMx+4JHD/DIppFTFUmwjaUj5d/pqR+d/Djf4x1kuIBwhVytJOH4Mv6XT8xvH1t9YFuLXT/9nHAmsV20S2+oXFT2uDra2ynnOlwKCFrh7HtIE4iBtNVLGsmYWEmtxZl9cHKOT+y3Me4rAYqBhZ/i1HEfu08sOS7MsJ+FAmktmtgoJZi9aZO7Kt0JiVUdNyUnNpXsENx5uAO9tnWTAl23ZY3DkxBZFjViwwjXekmjgxUvhbu2+wccHW7quFY8mm7DdzX07f4R9HMLWc68PnpVV/0HCzOjgQkEcT6WU79SdX7dYaMhjdLCmQSu98ZSGxOxWLwUdyqo3xVK6JbP8ceQm7DtLXFl9bimxC4Md5NQ+7EK6JgzwrKg+c5ye758b6xFxDbxid4pxRoGHU/OsMSCBAF6sFsgVXJtRTfTnaeoOJutOoaw9pZUiSNxmTLj2s20GbUppD8axpFfkEF8pv9hTX8NAt9e+JuW6sIFQAAG9VJREFUYoJ2hmSm02B4gp6GNc8yP6fUmPClOHv+r3lW0RfeaBv+vk5w12aruyt2dzA5d6PH20RyTwaiLhSzNW4lm40jUIrEKTQwDuOZg3Uek6aL1peOWdxij6CTc1LHuxPdhNvPxW6WC6v/vGl0KzOzcIjuZcY1GrZtBOPnx0xH1Mt9gmBqcwd/JAMgpHaT2LIZTaXA6HVF6sG8ynRXWtyBoRIlnfIMC+toyqY0+eC5QnXrUd+7n/tFeXdw6XUn/I4I5MS5uY+vyk4vWpviCXDxIlmiiV84WNYoMx4k43XGDIJ8XMuA79q3ckJtF3tqSgRYEZ3W3P0TRYEGu8Fz9ks8ejQaxZP1Xe6XS093/BJVPlX4PSeEERMzrhJBkpskR5vZ8BxhVZd4TLeaea1fTc0VkDBcrakPPt14hK5h65484EKN0y1klWKmmRdRuO8NMX51i3c63F1Byc73TrX7pKWXm2KC47aFajM6sc4MZGLn0XKem4rfr+3Sl4OdK8zBSrN1a8FvRw3LvOQW32FJIZslucRvttC5hYN5fGLslhf5qyR7j/oGEqzCZiEUC0ohXUXhQ17LYvd3hCPKt0hxGBYdDEm0KbrVD4+0bRfV4puX8m+Dl/zT+bm3fR2h4wpPENOruP4uTAEqfJ4cu8bauwkc+brr4McPcdqcopQWP4GY/2j5slsLsSsaHlmLfPjSxAV77TgrjBGKayB5a9x05/G0L6bOmp+HuTy4kjbxF/q1mb8IJKtHOYaBFdhrWe0F11ns+ue5Gf8c3eITSHHsgXf3YdCBL4wT9HaVC7fc423bnHw0vwLknQh6rwFRiZLWZN4icf828wa6ziOLhNmRtBHqooLdUqNMqCt00dX+R6LcjrPddW1aFIKU4QEwBr34Vyyir3ruhJmJOvxmx0/usmZvnelXHhlGupX1Wh0HSRIQ8O7BfBsAJk57El48q2i0FW6+223X8Q0+Eb3MOaxrf6LBPYNRmu9HLU637SqzhJsEeyBXyAQ1+e8ixnzstwT/CaG8VpsaDkMQD+jKh+WIRm0qmM0mvYOY1fgX7lsIPgxk4uCgzSDdWwix394V64svCinuth70coJuQwf1rBXy+39hWfEbR8z5gQeTsrXZEVrjk3Q5Ck82UYOtGi8KZW5qEoTv6YHBIInYI/MGa/a6MGO/XuLT9jMSKfLbCU+vdN65tjUjFb7DCYAQY8LTgoBtSb9EKjHeuQpj8vTBvuYW9GwFonwP/sRdnxzRPATfdIT2zXAHfFlHcRvizJ+dsCdu259oxC9FcFdq6CWkjBnLaOaz8bHWtKp50hzXeVaoOOmao0Xalf72fqAu969dPMIyhQYkt7gbPlRQ83PRLZYYq80SEY5bCuois4d+ghwQb4Y1DEtU5vbjMSNhybOCHwvRurKryWCwUlErhBlnK/SqMpCK1+9eu+FKazsR1m4dHXdl4tpbvLUzfocEUde6DcRBv/bIanKeuUwUxoUnFkArxAYCBXcPTAKaOVbbBTM2rfkZHVhHTPFNsdTi6CZrk/L3dKCTAl13xvQmjir/Rbdb7RAWMYZe4AxUJeZAiqUdxsauHuxafcO4exOOStGo55I8BfNDIqbntroZI3TGKmscR5L6v86PtnjrV3dXZI+q7m7bl4EBLFaThAa45e4bIIUiP5phYo4VRB6T5NDUYWR+THqJXRJ4vbTCDlrEB9ILq/edl+R+pfFoC1ED34NOHlydpR5Hq/ek2sarMpbkjDeHZubnt3SnwHXOpNDvWxxNvcUZt5jLQhZ4jaXPMnhMsrRbZPSAAsFCcmXJfyL5wA8fTtLo2ouR1VaaOdBPY2eeh4GEYPzSNLN/8TIdGeNp+Z0sBrMwHSjDfnRJGlfOYHHmPR0L7ZgUv6QWUZVclPaHOOZQIkbXMBvoYsTLFzrGysqbJfP8uEza2Eht3O4e4g+7iS7veFYFrUQGn7iu8ERnx33aQtyyLbJXYnwC3PiqtWMCEF48lZanU0rcDq3sJHYNbYu7yS+lMv9P27Vgt43DwMvprXgD3cDRASQbB+ATeewVMAAIyk5TO6ySOrGS9ANPQXxnnPJjzMCVio+CVcV3dL9fGA0bNX9IoWdoNJbMpWKxOPtfTj3OexBcQ6HtgURPKsXKQuHzVdSyunfadss38fGyx8iY0YvowmNja5A2XzyIrv+2NiFoUOHl6Y+jKo37Nfft/yJZRtcbmUVHaYZMrsrQzHNW2SrsipqqEpt5w5RgMV7+8viQH2EJ9ff9iu6ub6cTx3br/A+UFMWjpuRBR1ajlV8jM8WdsNxL0DSTyuKXUGwLOxiWENKme0pFpoyzruRuJkkDqrCZTAsszLn+ljC+wXhpgxM46PbGiddLhdlkZh0njNlPcEe1sFe7SvlKf/eSj2LDlAWxLAr6/bLRKISCcMbsLaxQDH2JB0QQvBpv5EC/q8XHUYrdQbybr5DTLrhjgp55HbjrYexADuIXnjh1zvi7nbtwHoqWx4yhwUlYrU4Qy07jZH077X48wqqoDrtSQHEprftc3kBu22zeH5c47dHfWMwdO+Sp1LZ3N6TSZii2mnz9bgs6Srr2OyE5lChgVdmvE+biJPXitJkOmFaMBcVseWHIfECDRmcxvQ7/WTyxRFvvIZywruiOpIODNnnWjcQSZSa4Gr096nwU1Vkz8w8bYU8bNqknQedAYtI8WkbkixCzTWg7ywTQrPrlHBtnpaHw5n7QrPqNN16+a/cbir1n6rAum09mrmNIxA5tLP3oi3OIh9Ol3pZCbz8JjGXhACx4oMeUfvPUHHExOfgHUEyq92yNOt8s/3D+5yLz470NaXQs8MVA8aXrobKNI2sUYuZbddaPZ3fcbXO8YgzDU1UZRUETXMXa/mBaNvYgWN8HAzc1OW1wC/qWaPFZYt8FK5+ec36uxQlBRHL7Hs/DuBUG2cZjYAdaUGxzFPXlKn83t50vkh6dp0Brf1P7yqi27O6TifxoAq0b5aeF2Qlzote01h6tGtSGgOg90z6eBJT2wPChiZ+iWL+CA/J8oSt5jWIQiiEF3VaWUIDIf6pY5m9zu02Za7KUJpiaho0rK43EiZ1pIODE45nBB81KLCjk5o9Aza+CgZ8vKO2R26NFEGiP9kVkrxeff/qXgniYmvltlTn55ihS+jN52LX10XdANi2ybTpjBaJi6HlsjZutWO9ZImVZBXs4M41PUHwSE/dT2UsMjPdQcFsMxRoZXwptdY1kg7/egb45T5txXOX0zQRx7j1DKwzFvQ5ufk7QLd9AyD+hSCyF9+zCa0yQWSBILLtK2BKlQE3zrpGX68TgErsdZu3d+V59JbpVhrR1N1B0QscG67pe1sLShbA8aAjGVyCkJNKqM0YV4JmbpEUyaF4LYwgny56VOn4zvvitnW+oB5G3O97P8JYQi11v7D2KLxwgRnOuO2FDykCuFSa+ODKJ5SddiZxaXTM37uLQHsVGWJJKRJqkqjZp2mHrM6Sr+49p0yEVD4hRrehYzX8zF38dpVgazUdD8bLEGSGCrsdxr+OCNpkb9MZH9rZHGGhNPfFovkqG9XpKWxaypQzBctYczbwIPUsXWtpKxRithPCOCE0P3U9SFTbqi/I/O4z9uq74pI+7dDvQSPqWfiOsdaDXcWyDqwYUxktqIcWFUjuZohJ4MlN6MeiWHNEJwRoX5WfoaQs1G+JgOe+KtPe5Kj+BIhOs0KVV2Sgs8NMH+XPMJ8I8kBodLsLccuCyEQu7auOodZrAcYW9MDOYk//YhFWEc8xB3Ktwm9mSaAJrTSISOjZuS1PCXOYElZ8Nhj5DY7ayalaVUMkMNBRvpHn7NX1ennp3vgJtKF56FEt2dxtWo1iNg6nNv3bDFCnwLV2mXoF22HprdBWzqdEIpeukalVKWAydZ3zg9SQZxszSGAWKlakYLWidoqCPImJnmVhil2nx1YNl6RPAHW6pNpHi20AUS3tUam3K1jaB+yOnVlnzBr/g3Clgs40Sav2NZzCzNJPkSyz6fH52hhSZeGoeSkoYe03Y7dhmpB7W46fX66LlkxLQi8pFjDaefHHxgvwxiG1Q44m2tIRxlRwWdXOy2mVWCzuIvQmSwsw2P9kKyIpLFkZdnt4mBMRM5yjpdNatMCbStS1n6DIa+7bSQess1gfLuQ7gttixmMtYPETe9051uxNEGFcGCpyOuqSrQVmOTGy+2yEgTk4lnzVqLpx4s7+gBN4l4lFYptYlaX8UoVhhD5y1Ii+QyZCHN3r+Yvu4XRmz/G2bPy6GaiHi2inVycw97IwuhmPxxJCLH4ZiyT1Uz3yNlAnws2pQrb45PZPNA3kQoRx6SQaMWUjpfDjfy2nXJIvmIpCSuUyhYTHNRVUPIMUWUdxCuIcRv77hKZ6H4BcPh/dXYyx7P6iiE9zDahQ3l7LS3CPZCCwbLjvfhJWRlRk2T25/b5KIqDbuFCFx5LXnE8VMzpYFxQRjJ6GLTxpTCHOmdJV0btuGt10g7NEn1Z8vj+5tcFsitX2/5twFix6IJ44ha+aR4yoQf7TkDSaFMtCkMtH6BncNJNsejrgNVmWU14g9BIdtGyfQ4hNm9wsasAmWkfRZrdg17mzsyuz8Vul4eR7+ebVoLiXMfb+2n+tFevS39WKMtR3CD3SiOGdfN08aJ0DGNYEBL1serStkOqOVmDs+sVUzJ3Si6ZF52TXzTcKoIKNbfETaKF4qn6KlTPBbRbVGbY5+qBC2PK98LLEQ5N9kw8Uqnzusdwd37FNX2YflsxJGC4w1iIMHzuqfc4LhXfCRy5eEb+ehmu2M1BKT9FvmwVcyEHfXJmbfSjH7Ukvw9Ox7j410v+yCRTt7S3S31Zrd/qMUH2gbd9wJcc3hY21eM1a8anish57Ze/JXQFCceL8jq+XZ87Kz4N+CY4nTqOfH8xeHEuybT6chxmarOo7VsLLxgVKFHX7mlj/TrloeYR5l77zy44VOGFCMmG0dieJVJ2BXrVOouGDDqxoWJkQAYaZO4bl+4wnmxENAYlsYndEhHnkqFQddphSEO+JlroJabue0mfRDYXN5vNjpeDVwvMdt6MXXHZIwya/a8ziGmVg1agzFXm9ra+cIMVC/UE8NLyJGdtizaWF2BS6XKBKPJhDunWhl/5z56AtGrfR0OeMg2qUgKf2UYOXRcW77ANBynQMg5eq/zmWO8MXHrc3KO4pzcLs2Uaguw8ybe7edzLiwL6X4zvkzW5ndwyYpB/sTyT4eyEESzWb0RCaxbU5D0UxvLdfsr3ZrlnDEdaMApfgg0NFVi0c0liSB1phCLGxITalZMsD61c3sBvYvl8t7hV/mkaZkKM7yD8MZyK6affVEkBUsAcrFVh0/2a1ZLgtMbRQzVpQ9t7vW2QY4iuiLLTbO+RsjTjk46tRd/VOyM1LMzuFGPn1NMy6rOxBHezm3O/zgrnmeXbaxDW2qAh69A+FL8WJ/mQoWbfAf94ue4AAT29bS6u0lD900Qg6glqE3vd+5bb6VDbwK3Ep+k79MHH9woHz+5PmlIj9S4EiCQ86I7WaB+KyM/WhQl0Z5/nb5+LqbFPISG5A3ivO122schOIwPVjVUeDDM6L1RnWb26PdyvKT/DKpheU+scXraflK/RnnAK4ANs5Edt1SzCBVpqHivPKlBCm8d8ZXns0b9Ynlb3C/s8sce9ytPhPUpge96CYWg8n0HfiuF2Nns6X+kJx353dVyup4z8/FrGJGteYJGn3Q+9TnI1PJ6jD4TNTKpjFelbfXSvcX7iH2OzZG8SpL/LduuniEowjUKlYOyhoi14t1q9lR7yjY1X+fD0Xu8+NpW/3FR6iE9fykiI+AoRXLrDFHzUEbmtE9lWCvi501cqamrvv7PrWFE/ATxzE2gTYbBxVSoDhXQ6Y+2Fuu6hhyzeEx6atBaukq2K1ZxbeAVQEt7kG9rxiK+Q0eQ7EOWEMWNhGGL1w10+a6yyer/svTaYhXWmrFtZt7HVJp0wRPYByCY8GczQjV3sJq5WpepBkee0/8WRFvrIDmeI1kcFfOFPn3yLax8gWfj1/mjCu40ogMzzPCaJIxT5Q4KFY+/94vf3uVhmJVTRnqKCTKhquwGc1stWMxtefV+sagrbCxRiBuejzU9sCPGIpmdwEUA9MFwAaK5c1tLDei3+BXROqiMeF2NWOU4kwr6JPWnlqYLO/oi0ADTNxIE5zDJlyOYr8S8uwK1V111xrr4aZ9mnGaiSepEPUUpyFeQA2u9id+uJMefl9FBLuuh2DmmjN/Q0hM7KPRsXyQYJuFV/T3r6XMEb4YUZtuh62deXP9LymK4Y6z21dhah8aas3PqAeu+oEUuRZMwLRm7i94Zob1yqYmc9EtAlF/yWZFrY40ZvbDkEw16B076+tUGxFFpwE9CMWHLyUwhqM/fnFlt36E9dM3dFf59lMKXqOZ/IDbEFhTCPbMyojpij1t8bMJoTepvJ8hTF7F1DGgC4bHOIrDr6oCCT9c6el57s2bmukMzeXF78M2XQ3pgub1fF7KymgmwJrKWjTcUEC7m9aOlRX7iUyiSZd9iX5wHTBwqj6RuR7PfmLAlHwoal4zkH9ylQbmtQb3ITiSl+Kgg+7iuvnG/fz8q3xJsKFBh7kOy2f0SnOV0ENRXPUQDH7kGqrp1+pxJ19UGu+LbYe0GXj1isU/vtArq0JTycb++vo6z5z7mcaSGP/E9Nfq3wnHcicPnlvOHQ7F3Mql5MC2DamWvkhoKO0B/A9xmbtnPzHAxGvoMQkRLAILtfO/NLX/3uuJotO8B9sXnJUkuPpiO8MCSAKBa9w1H10N2hSgjbBDiqXaYGH6HB2oKzKgz4UQ/f5qfmK93Y7RjuJCm3DI4seqpjUbvwjmxpv7EL/Mb3f+rMgfyLCu5y9B872uJ745+qAWrNwxsVosMyxFExq6Vkcb1sWsCS+G+SzR3pCZ1zggP+6409Ul88cYJBSnzPnIoR/Cmbjib4R3nMU1HpSHapodP+D3uFpZv2iMv90rC4cpPwQmKuOjOs3OLwcD/P/2riA5biMGKk9w5Qc+6J7Kj3QA8QKWXgDx2SGBbgDD5UpyzJXjFOm1ajkccncbPQ3MDDiEi0QP3NXldRTsHN9DFwfuYUK8duvtzmGxYtw4EY7IwizA27HYEgc/uvA5TWGXUBqOjcZCe9433xCPTrrrve/OaDSa0K+awc+ZJhbPrtc88OYHdLtd2egoV7xm15HXCP5eMYYH3X5DGUMSjoKsV/Z7D+KLqt4K8VkQM0vTuORVoEW+LhgmyiU20ReMkXxVFCw5kGQF+KKAWZu13AqhCovDBduo32ixTT3Ma7lbeCaXnePTlDx3j7hKhLIQEQkj7Airc6iJrEVXcoPfIjZhM4y7aB7i7pLDpRRA0CdjwTslJporuylkAnWLxYr3Tm00hbwOOEta+8kbpI7m9snb1M4ck+K6Nt8N69n1WBcacWX0q7vC9VrORBD+LeOQV/I2BkKW6mRG0VzxuC+7HbfaqcgBj09iccXG0iQ3Ws+C4QsWG1afDSJbzK4qZqcMsAaRS7qbdTTnWLKNYDEei7mzZZEZfsAR95Yw27QR1gHnhK5/q7n7gBkhR5fwiPgyon5DYP6Kfn2Komeo2MO0OJ6mG49SsSaoKRiyJOk0cfTb9tIicaaZ2pKQIczeRle0pCdqJf5+8hwuR9l0vB1MQes59MMX7YmGPWniqh6K3ASClPD1oCtGHIiam4zPftyVCr/gjhSfyOJIhBWtVY0jEseTKaRUOJWzYQ4OL8npBdjlUnDKp/URQhI9JEQWoUf0686bTEQLZu3t3xTE91YwKRRaQyfGwDJkfIqwe0Y0skzF7NhXq7s7bDe9fy7EhpelHGP9K3Ivpmbhd6kYw1Ewt/JpFc/opbxrXRuoKVPIezsIlda8DyWYv2ERoIdWQUE24fCiCUNYK7LxGsJCre5j/M3QUpsvP4bmTC12zoZPNQkGElrCVFJtkjsJbzvaoxOQxPYHJQ1hmrutLx9OFkhbg2KTlEUCZkd3irh97tG6x4QDuQ0pORWNR1fWMhfTjqTiJIht6OkhRIxprICBgkGoElMheWWPLmv0prDDGUq0OzMsgpFFeLv0i2mFFVrZxHrVMNeYUHISO6QdM5LWOjYZVuom9evF4jvJ/g6Px2ixSQ00WXTYXYSRMCQFrxCtRB0pHiwBOjgn/iN7TjJtzItpjnAEXE2nc7zLDQUlfJ+HHC1KoRcN0HELC5rGXP3TxZBfNpj2nhSfNLHUuRzIEgkSEcPKjD2M6XDJ78BUlPjDQkC+kB3kBEequGv50GUfWwdFJYRaFi095wBW88tuBAnHCA2OjgaDJBPR+/CcGhfnbGn09jjlBEpalZCpJsVTRiNCmUY0n8qdZM9hWW0v9nzEks7J9AolWYfg4YEDTLYZgvreCYW4Sxcb1gdnVI5jtjMhRkZiYqypG0HSHCixZKZ0LYAaGDUaNbDiHoQIfJV+Dg1iYRMYirOKlg3J0rKpWAAK45AVTg5jVroLjXalkepqd4l8MostuzeZTqBSz8MxDCzz6ace53CBDBMgxDXUWQ8IB3gE0nDbHxP1uTJfGrghDGSp9liXFWynF+gsFj3aKj6sCJ0yZQ8XCt0ttuI/Fe6JDA+ilB0o0JQX0pM3rqGeUXTiQwIa4dAskm9UKE5ENq1mygTSZozmEFJoUKxDlCijpJduMWWCpNIv0+Jb9xcw5HSAaT4rT6gsKNNa5qLVk1aZXVTT6u4g0UAqSwQ3qxmjEtA0TAjJiMfdh9mznaWr0PLT2tWluq4V2ij6A18KcTKyxAA7miNzkOxmEOAagmtM5orxq0YUPKyvLq/S8Wa/snUEaBpL7isApvZnG2iMhng3mPGuvmQZ+6YlPzSiyB9cSEjLLtDC2woplX1+R07QlCuxbPStqkk1h/zBJSQR9EEtmQnVvbKmrzB2S7X8agWE6X21uddUOX0Pkoew2Eazamd2W/2plecU6y7Xo6tJlZuwMVdD0TRaMWyfT8PoRrROplIDPmEskhSgZzR4XOEEkkKjPtoeIxSyI9ptRJPDRu2njKMd+Xt6c8ykPLNBPmi3Mp8OfG0z5cV1rFiX94Vr9lH5AVoTDlLfSNN/pE+QL9fibLjHQYe9P4Ny77i+90NklPSjz7JmKdNm6ZytSKtbaZr2mrGnWfEz28NYfCj/zd0dfUE7vIDuYyK9Mdfh9ewQ9Vp8ah/4lMjbGL/0NxUFju3x10D8Ed711ew9h/yvxvvuRZGtdWmp1QFOOx0f1Wb03L+AxXqUK/Q+iDre2HATX3xgKj3e0wNIDndsP8pymLTXQskfosNXsfhTUJ14it1Lb/z8R9nH17f/MsQnpB/9rMl+xFD2Exf5LSH+vbYL4gviC+JruyC+IL4gvrYL4gvia7sgviD+v0D8/UWej173947rP3+0PxZxr3ZbrSxtNcdLPI+Htj95RhW91DnboVb1JXd9/7mdgA3vWk3JL9aufAe9/JkrxH9+v7ZHbk9PT9/+vrYHbn+tEP/x7doeuT1d28O3fwCQdoBnJ42qsAAAAABJRU5ErkJggg=="></image><image xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAMIAAACeBAMAAACSg8iOAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAtUExURUdwTP///////////////////////////////////////////////////////81e3QIAAAAOdFJOUwAgTT8RfN/DMKRgCe+QbDiZxwAACExJREFUaN7tm/9rG/cZx5/Tnb46DifHcxwCQjp5Q12GUZQ0BAJGsb2OEs9ooyu0FOGmq9eUcaiUJSNsh61mbKxgHCjLBEEYlm1lDCMn+2GQX1yWNhi6I4PBxn4QdWTdWeb0/A17ns+dZUeWE2WDsbF7E9+Xz/O5z+vzeZ7Pc4LnCICv/08pcdLTu3yQB7jo/a3lRdOZ2jG1X0JEI+Wf2iVH5qEsnZMA5QXRpNXK/RMy09PTNXEpnT6cMEmja2N0rXYR7i08k5DqXEqpwwmBMZC0DAxotS7CZLYvgjJMHj55LEMBubhYAym/psYHRk8CDIwOuwTqFkmlQaF/3CQICvWQyqeH+yEMUCRycQ7IPU1L0YS1bG6SwzOkaRcEQcqAnNTIp1DWtKwgDOQ0TQ1w/2fFIR4fpkBK6QFaQ618di2nBtIZNZeZLicl7eRUWhAGNPJHWZXHpPTJYylBkNPTx1L9rIFmniHO3Qu8HCVdo6AG6JhbgLtjkfOgaIJAhyF1aOFuMnKaXcUEigBdDWX72UuXgFasMkFKx+NDpwJjwvlyCgbica0mCGU1l7+b5FHjcZdQXmTjULbPvTSZdqPJ2XEqcMojsKs9wtBCGuRTQ8L7HoH79k/IUVh5DZn79+/XOoRI+pLiEQJjGdrOubycqXleKn9IfaFfgpSh9Ys40LZVOwSZs8AlyJkxsqY5MSKdOAzE+4zDNExeoH0opach96GS20fI1CY9QkSjRtoTgczAkEsg41QKJlOX+nkvXaJBcuzi2j3eWR2CQrs95xIUygIo85q0VHovH0B+Zj5Io6Q/cDKrsDYKMHqeAkJvhDil9jD88bwa50tKiDi/XU+KDnFYBLpd++AGGeLD/g+UL1++fPny5cuXL1++fPny5cuXL1++fP0PS3neMq/0D3GaL/S0nnun2xI0cEN9LkIUF/iE7jjK/DLA1QKd3fuJxq5ld0IGIjafNeszT8zJUSPnOwTQ10HGEv1luwncC+AlfH/0957xUAXxiduzEGrtEeoNsm/BIOa7CWGbj1WaP08BJLfDRXfS+U6AFPUAATyCJLy72oQw7ojhuMEj8AiCoOAcL/QLOGFaN2Hwe8exDYH5n5gtFWQDr9PgOr4P38f5JDn6EYy8BYNXgvMB3ZkHfN20kvTwbAtm7TbMtuGnaBU8Ag1XkOkxFQK4zBNXI6ZlWmrU0hELMaTYlKDoXKWuVdRxhUK1AmCs83xD7SDGqAHIgNu8HgfGf2DRyJL5UG+7BEl/aNgy9VJp/a7PQlY2Zpai+J6kr8fwLcnYlohd3IrhXyVj3fVScRsMG5a26U54qZ0vtql5ENX6X1AtbkVpik5eEEKWGsEV4aUgT46d2eARosSsb8aobWIn6qhwpBmyyLPbLiHRBHTyxbldwrI4QQwLRtYsGKXY63StCgIdwCg9QaiuAyw1ozTeKhFUSOyEnUrldivR3NtLYTvQMrLG8i6hsBvKmw5Ul/ERSL9+DbOCUGxVKsa6MMfIAHDsrDHHc9xPCJEXsTWxs0cIOi/vkD+yXQTQ37Vh9U+YFanlEqr88BfCLOE6r2Dz4BpCNn/R5zVIwy5BxnObE9ct6CbU7R3aUJgPWakveYT6Nj2rumaDh8C5IjludXs/gYIGgV8doU6JhpcPplGaabUOEFZxkzLC5l0U8wgc1p8nXfMS/lJ6FVdmLFXW5/YTZPJfYof2mlKl2Iqkr2MhjI8Fwd5HmKWcGqQ8WnpMbwiX8JKtKnqJJ0mBMMV7KYC2wfmwR4CidQvnFMP6Ea7InEq0qXCB3w9EGMTP1Q4hRLtKxgYNXzE9gowPbztqED/jmZ0w0abzcXRuwhOEmI4befimSTkNRZE2MxZwlhBBqeIeYRAp4c0tiOh43SPQcPhD7iXWLp0Wb6SpAz8TijBEFvky3s+PSuTG3vXUjV6P+fLly5cvX758+fLly5cvX758/aek5A+2hd7uXI68/RxjfeUygNxdLP+6ySXXLh3l0t/I/Py1FUg0n4OwusNlticJko7Y1bRLSJDFUv9tQhTvyPqD3gT7zAtY+pcIbuHoovD/0RbARHN/AZwtLoFMxhYROnWmeOe5Qz5oeATJwHeBC9grPEyb2k5DRLTFDHyPS2luCZYJ9c1E8w28DOErxxvKG9hS6Tk76dXLDyWMOFcxG8CHXMKEI84dNn3NuUaLK9pkkXRHN3YJ1c2EheioIVtvhPEafpcL3w2vXn4oofgAjNJIKy+qk0HEa9+ieH8ExUaMq+xzUVyRdI8gmXMJfCWIhRD++c74Do0waOW/0Xbr5U8hfAIX1dVNAP0RNb5KO+YRFxbD7bAovCa8yEDCqhi4wrfmcog6vFiA2WbUyitxrpfPNA8nTOCPx6C6UanoJW598W/YjDqVyi/so1alUm2ON/bt1ssiH/QSF19h8jvFZgw3vg1uvbwXQXxLKQRNtJJcv8aS2z5uhfnOmuVjk2vBIXe3/i4F+wiDompbRHzHrZf3IIy3RZ0fpl7DB9U3uWoN8MJHAGEranEB/GiLDsOrj3fXIMbYI9Sbi3SjfNmwRb28136dtTnDsl/NwvgmB4ouxLpCdhC5AM5xiH281NyNQxfBXKY4BD6mzeHWy3sQwvh3qW5B/QGsbs3QRuFIzzhJpd5U6OmRFn9sKTbCTrazl7oJxSbNIohuvbwHQbyDPoEl65a5HMQrty2Vvebo/EHIqphvQtW+hY9ktA2jp5fsW2ZTxitG262X90q5EyZ/W6SE3MjDb5HzlzMYP6U1G2ir/FHrU4Bz2JrtRXgZLfLeb9B5xauX91JE/Kc+t9i9WwTf3+Ze/6z3wzAl3lriObe7r/8q/RN/suiwXqHjUwAAAABJRU5ErkJggg=="></image><image xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAlUAAAL4BAMAAACgqh+xAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAeUExURTqW62qP4RYWFkuV52t36KFZ5wQEBLtW68Nj8FNHj2q0bHcAAAAKdFJOUyQ4Ai0kJhUuOyNVzUmgAAAgAElEQVR42uxd26rbShIVCOZ5RMGgeTvx2TTkTUWHQB73OTAf0gyC/Rn6CUP+9qjr1i1ZcmRLsr1jK4n39lX28qrVq6qrO8XhdSw8/iheGLywemH1wuqF1QurF1av44XVC6sXVi+sXli9sHodL6xeWL2w+l2wctllOkL2I7jslsGz6NaQ3UVXnd0fJp4xvmGf82+NlYvvQk/GV5zcFuQu+cEPS/fHmxx/Bn5+vE1ei95skE8or0pXBmdw/DDHL7Hx+fXJm2Fl31Z8Z85Nfat6a8i/eJe+QHqAG37bQclgpxi+5sQXtv78bnB+frobEO5qrPibkW9LXly/NP66QuICXw32gIM8Qj9CyLiiYNGtdD3eFYLdEJQcge+Rz7r2/Ad5Kp2f/gkR3Fqs+HUcv/8DwRXSeezL4beqV53dqe9N4JAnxAc6p++PrysOdoM82x4kT1l/fr50/BLuIChn578KK2d6IkwOEvGO36yTNyIMZz44/lXuTLHGpJEnCktDcM4CO71aQvPgVNOCfcCV53d6TxBxk3cjL75Kr4hQdmYmuEamvrjepUQOKnDOYkgUVHgR5J3ycwPjdjBFCfJi8myN3WCjwZrzJwnjFwyHkPEqXIHV//+unuOAb/8TUoucXYrVG1ZPc0D1779E0NwVevX3MyEVjx/MKx5x3SVYvT8JTggV0gXiDzEc02nDPFbv1fMd8IMHSDZ4i7H686lA6omFAH0kfo845e5/AVZvz6JThFIEq/8b0TrmxAqLsHp/EjqlXyJo/eW3EBioufHwBKsvzzMAIv+N5Iq8gu8iWYelvHpCWe+R4oAMekyXHcZYfXkarVJSQcU/+uN7aC1bXMCr56IUCq8Irh671pFiBcsoz2H15UkQQmZWdKGsWDEGEf8lxAqTSfQIq/dn4RNDBRyPFIDxhm9taAkurUecwertKcRK/kZJJ4hUsfqL1ng1UYUfYnXjEOzuGIPEq8gpCj5iVv/zu2vbEKRm5s5i9f4MWqVIUWoTtYp/EmD/jbxibv2KV8tZ0K0jzTV3dptQEkWniFIVAcZIoe+J5dvIq3Za3gdY/Xn5B6uvfnC3+JNvdn7JbMCyQVCXxcNgHAl7bZ8zWQOsvmwjP/WdxOn8+bViJT40XkAlSAmr+ov/RGIpr9wZrN73+Xz1Y4g/EK0gOSyFSQ4S96ZnlTjS87ya/3T1TchRz+K62fmz+JPClQ6CTKwoWFHcw4RtyLF6u/13fUOuofhOdqLiFng0BEXLAxwJLObVmRh8pIJovfULsqlSqBCrjFNgghXFvWWLdVryy7H6cpMvuLsXwSI8FTt1SwRVqnqk+qMnlv9O4t6m2ddprN7vEBk3Ai6VQTkUMQ/AynviFjHrR+QVq3s4o1e/s2vnvE9MulZhxKx7+rUHrP8XB0JJoA/nYhAfgSA7VT6TvLNmmVtHDj/0Ytx9y2D17n1cbMixuoNI3+ybAXaemGkUg+V5BOQQjFdaBeukLW5HrOpLtKbek1XKJhCKsVJVhJCAJVB5748sWI7CcGAbMqzeln/i7kYobnR+zMpVY1elh6dRsP/JWLUxBEcG65dYXQxC92CqZwrF9VC0clUmWCTsRC4PyitpB9wNq8dzFGCSznJuI2B/qVJFFz2r4jAoMejYNsyOg2+7x9MekNa/DkCbjUi1YnIOmtowVMqv40erRaxRxe82vLpJFjNTgpFcmeNP5rZYoDhfJqGK0Rd/UV6xuo8ywkmsut2ipr45jGhRCGJI2XqiaToyryAOgn0UHo8ShUGbwS/nVb1aebpVMF1+ftB0WSmVssBYEWU119DzniPRcwxK+nzVONjdWa+vOD+k6RqQJCeJFKuUSLraBfpxbCOxQst1GXd4BL261TCIaQ7Q4g8H8ac2lIOQQzDxKqzB6nMU+1JJXYt7MCqua/yRVQCRdsDjh6DlZJpwEVb1BcDUWwt0vV7ikRNBnl3O5FysAqHjOb9RUhGvPix9Djv69osp0F2v9QtKCzybJfOA47wmVvhM1yUeyYsaWM65gWCNsarvFDI7VGHAqnsoKWCF+USElz+osu6JX+IZOAZvyKt7eVKUOgxIBGZBKPUqrcRkwSe/HD8+PlJpdMZffV3LjHrpw7vLAOsuR4pbhuSatixYwQpR9FxdqMoX69Wx/VBiHe7gGeobR6Amf1LT439gdT7vNRFMvAK+QOVVzAgP7eF39ldoAahFBalZWcE4K4KyrrPCK1zHnx9SwYpg5WPhvbCqdwSLeVQJs/KpLVDB0hGQM2jByWKQ1D1wVeZwmV59MrigUphw5BXQJ0ZxUYHxQZ8dRxsIx5PPs7zqfim2W5uB5QXV7oxZBysuVNaEhoMmD2/ldYpCMu6QYfVTB0JpargmBrtLP/d9jBrwihHByERdGxa8lEFR7JSghDmveBTkHttNtb17JGEXBwoyaaN19cpclc6ZEmgagZij1WP107KcwUx9wsrtr1c7w8pyzroOqutELEylGLC2NMsH/TgG2zRP/5vWZFB7hipuaoRM1pNfQHUIqcY3PCKtsnKfe4hxcC/AQNthqpGgR36xYSc7OhaqTNttduKwaBy8kQGoN0QJjVeQLXjLCgsebDKCLajXyszIM7C894LlLpvH6VZITnczyQNrXdf1IiclGLGgouswkqlTveLFJoc7+vZd9F3yZK4UV7aOMjehMq/MMIHK1QRcUa9kKieEgcOa0Kt68wDpLgvD7vLzAw+Aac1INbKgnM6IPMGpSp3qFRFr0Nx3j3Gw3pyHmCeCYHmgh7zBKhrQbPJmUGQY5zhH9e3TXjS8rf3M9R3GBolASC3GtsYmeQatVuGwAKPXEmCNL3tepcrocPXEb+CvIC/HoKxzw5Qt545d9B1mdF3roh/5rFdY46+6vUKxuxIpniO1EEQrHms7NkphPYXdBFI9qcqyIV61Nkd4pV51W2pMt1F8ysgHWXVPe6t8rlSgZQXQP3PyntWvnHMb98nUd49ClMU22cIRzwmh1+UQXsqgefQNuVX2xGoaz3qls6kz/ipsqlfdzWBCa9pD7UdDA0orxmZCrW9hKgYJMq23t2mvhmm9qh+HOfUSrUL17LrGVKtVaFM24jrRW1PoVBYYkeoFq2lSrY/GwUt8e72aON1uhAStfWozDNgaG0yU0tKxt9QPp0nVUCD62M9wtLpoCIvqDFvUQbt9A5NK6hmvbGI5n4FQyYKcUqd4lT6OgmVJvv2ovj0cwsP4q7XhDbpnRzXY8iSvxKRSAmgpFPyJrJcs7syr1NBwmNQr97m8qE5D6DokSP1CmQtNyTLAmRyQw4+ZleWDi+qi9SfBS+YBq8r2z6l0OYSxKitRwWTaTONfWZKwxxhUg0WbD8xo+9e7xVx9uVJq9cVqCbqHR77GVKrrcM57RlFv/JBXrbUiH8IqL1o/RgRKU5Uss8GsWV1IRasCtWtW9XzKWJVEK+ZVr1tWv9IQDHt70f2zZS5XaW+/dMGYWUeZBNTMhlX9JBcU/8nkKikhjLz6sPrVbv0M3dbBeaZejFW+OZNlzF5h4m4969qbl/XoQCkMS4Ir+SupIZ/1V7NlqW73aFwONmZbBySTkHkrkKWmqPZzSrAIHBkEJQiTvzpZTb+5v1q06Ue3goRc90xbwqQdYWhOK+/CVveJqb3q1CqQSFHeHHlVJn91Zo7+8/grgUlaG7OprUqzPgULZYIZJ2tWJf9tuHJVcjpYJn/Vjnb/eKh1XosHQF6Oq+Uq7USjTn9v6fLASk1OR1ASSFCRs4pwFSXp1ZHlan1d9H6+AWzOhg277h7AmaCqlPgrtuyW50x5haYxsAiwcsCrcJ5XG4FQ75Yta2cjgM1EcH3d6zDopRFUpwEnC3uMFCeCHHxRrYr+gnhlncgz4+CKPpnuNkzTltmMV5TRSA0mV3WJQAELTpBqRK1KDUG1Dk1jvOL91SZ7ij7DPI56KVk6YlOnWWXdumZl2sbPCJVlNV5FvezFqj9Ur9qTRXHzWN1Ik7rLxKrK63ua2VRgK5dRJ0mVVzM2tCQ5j7yiKCxjFPZAFRmvTlzD5+EVWv0FMd/pi2p4wK5Bwy/xaioLZH+u7pPtemRYQXCNeBUW69VDFWcwzZqmiUDNAr1YBV2/jLM9C6U5dgKLzJX3hQh8M6gzXNMns/O0TL1U1y3oskRZcmUrFouXSqu9J8oKDBUhRWgVvVhFUvW/xBgUXrVrazL3MlZYZStsqtF2FAyUQgUz7UJDZS/TaCiyRUGY+tuFV1flg/VdtSrtwyTrIoZrJyHnlZdUeWpqS0oKjag6/RZJFc1VvLS51DO8WuivujuEIapW6e6pKWGudGuTrL4uoYdzlFLHwAGYjpK96JhXny0GeRBElP3D0ybGEamhDYXkQU+GQDOeUlMo9SgErCLOeUk+eE6vvu7OpPr64U8mIRBOtgWlorF0n3mfuQSYra97n/Kaxg/xKo66iHdjXnX7ixomt6AbUgw1Ki+GAvfuzRTWtQhTSh2GzJVA1MRhsCiVV8mJnu8Xvf7zdhfd3l0k7CDsAvvPInQyApNZH7LpNAmk0guzikowjRj2Ilr3ougBK5hXH1oXPcuret+AuoZY3N1fpUWmNnPqKXk2sy6FdpwuGZepEFM20ppWekWNeEW+PePVXD/DmrpovdMTrA8tr8XkG8jpAhsY8wpm+mBkdqt3n3ylKGj8Y73qrzGv2rR26TONg6hNjrp8crAYXpyCLaC0Nd9zLpRL7JLZSByyXBUUitT7IXXRsLxPZp8gqy+gGM9s8foa+c+3dJsvS2/Qkho/SG9Ou2B41OtppB5UrXrEKUZgobyy6UF3WM+rm2gVaJYsBQbZisI60YY9C9IzhLP5cqP+ykbBoQ0tJCCVV6fbpOw+j7PGnVF3lXRZWVlBq6CD6Muaqqar642VjLP4U19VsLKPeOXW5IP3SQIRq7QVRZXsZ7LpFHa67cIUqRqdgxd1bxpLb1jXCS5W955XP62x70Z1hvof5q4lx3EchhrQCQxtvC40AsxOgm6URYA6Ri5RQN92OibfI2VbtpM4qdTUNHo2sxAomno/PnsBER0+c066xCE6vVe0VapFAxiqJyV9HGU1jqGp66SuKGdoYn07z2p4a2VJH2eOnGHrcMOTqxk/gXkZWhixUDxoFFcobFOd/JnGO5hcXemOjq9f8TvfNSi4ZFAXXdX704qqiAG5HFuSvSBzqIB7eCgLrDBO7WNN/RscdG7/vjyXOTC8pZt7xtSuH182TuiodWUjQlyWzBaA60Exdg4LegGDlpWc1L+z+jZf6o734K+gxFMmPkY4RzxlU6LL0Cn2JWxQNkEFjoETaOeGBenr8gzkT80PvmZuHw7sUpyuFLKK/gpaJraZITJ+l8jlQJlVqOYFGRTS2KgSenugf/Dcnts/R9eXie31GrRgfqR4EzdGFwoTl2jlibojJUJWxSFV8q+c1nhM44Dl/INNjv6puroe2taJ7ZGz8fhesTEUUXvtgb1ANytvv1LVlL4B5ZQC7iB99Lfm7kWQe/rVuzmJEdPrSZpSvF74aM4GxNisHue9KiRq0BxmBYp5vHYyKoRWvzo292M4eFjIGBTi9AJmzge02FRqvWUqUPVo4G5QVZ3SzPIMlCvY1d/BFZ3Mn189pdnYoKGEMTqDm2tWGo4d1wBj4QClrlRmHHBaMilYUYV5XW3pkF++SmPY8baRrx/dgLRP1jiMt0EsQXvjpEApmigXPAijddVVJ9VNuInT23LVrncddWSAuD5v4IeP/mFjmr1lZYdzuAmtpdKO0lU0YJf07qXZfKWHtXduH7YqY8uUeX1oVMB4NdViuwWB2ZQdcgvLIrwXABkDTyiO2Ar6CEz2Yg5VXf14j8npE3XI8pZRYF3D/HPvvFsc0sE/rLxqRK6gV89De0HvIeZ1d/3Qry4uqOgj8XYqO2zr+U1YDMOIjlNYDwF5VVxUghZpVSqxSsWhVSnhIzhewjA7LPgmzjNl31a/ur6tpHrCeUCMFzZJIUQ8VvxDXqyrQJVxVypyeeHiTd6DzLL4Wu5XL3zjXHeWlLt7UyLeIesErfKiGyI5gD0BE+3ssKRT6YmFblZY3gswxvyef8s/+EDFaaCHboTNpHE4xZcVEXtZE/cpCZbU7JzGh+HPD7+Do1j00/nB7LTrdfCeC/SAQg05vaXEvOuMYH/rTFR0A0rVZ3KbGb5tT9VH6/pi1eh7E9L2E5wdomMGCjSxUXseAssqOqaG4F6Lqhf98dTEcl1tzQxPdvnhvrJSdy4+jHmWvQfcWPZF6H+tSB9D5W3G+SixUxRYDqpnQPbOu/vV9YHCktYEvUcfZ51eUgr1bzkzArOFZQVo/4EoJ0P/BHWXK/jvYk7r6pg7OLziEW3CbIEa+lh9F/vsd0vx8VyTXrFpk1BKh8WlYhn14iRkpLCunvHwDsdettXCYpyHD7Uq2ALkUqD1xOK66iOQ+SqEs1QI2bmh/l9d/b1YXb0qt/Z65HcQaJaWVKmYL04IvHe5lE2ZthAVRYlCYXPUR2F8RV1Xh+QhD7uObnjgAvLiIVBHmrsjcBTq4+Yk0F3LrBeVaYaUSkkp9iessxEWFsU6XUr/uK7vhTWWVaOG2CGIPnywv4uqRwh7aVJfyqhqaUnEQEq00aeCB/XNC/AXMduXR97Oh12uYfP/FsE3555x2f1srtJyUrqwGT00g7OKgQ5j0xITjhomOpuvxlfO+bzX73z9PUWDWJll+HQZFSXOV5UBv4pN8XFI8Eoo8OdGUL2DqaIMJ/jV1yJ+9evvQc2oyBIn10fr7Ln2kmBcz5DIrD4Fk6O+oCmCuCGUyjox9ivmM5wez4F8fZnF3t2/nHu/ssx7lOi90S4Vm5OVDqEhmfJj/GvS7l5GzYfeQ1E2cGaYTaN3nNXwXJva+7LBjkUKH2dp2byCmtpb1qjngC9ggbm5U9PSbWQIGNyFLqQv9eezcdHeHVV0e6trKREVH9mprXQYzQt6WhcvoM8Y6VcygcozECCgENEOF52seF57D17f3Kx6i2jS9W48rEw/ib9+DHlc7VZBwWSZ3PUy6tzuhJDa42d4+2dmpOCccqaSiP3Kr6FEDnRuK/+9/wZ4Mun5cQglsEDBqJBf9XewEox+0B3MtjlXOfrcT7P9zX+qDxyCfTPkPXGgKrR20YFqNZV4CbW923dQciCPzme4HjEtEBGNvQt0dCtObWGSi8As7UE9ESPWWrqdV1cmM4K2qVtdyXfw2yJ+m7Po789XSICOzOHz4mPzkTACs+pWeclQwmQdmJRGtBjiBu3oCd9A/Q5evH/wVRjywr7q3SWXo6FXWlb8DLoMdtN9xHVcIVRhmCZpH2UNaiux41LpTBjr6q+bRU8fly9auZm5jNlLQ7EKPRd7M2eOC8txMdKd8Di+XT/zLSWbE+we3v651dWPN+R8Xm8HtICe5bzfse5YBZzEwjoEj61Td6wWHGRcifqRPT2kZC1L8xnc2qXTaW+/Gt50TpE6GXwDEQKdbWU8JKL+zbw2rQcdGoyAGGdTi2KgDQAGE9F+6GGdnu1Xw3NfvWEDs4KYKEfdsWEBmFHHA42q4DtnWXxlij69coKol1CbS1KCxv32M+Fx2t/B83/3ltJw0FSRMTEoszzPqcAmWOpp15CF5OJi1P6G95/H9UjYy2EFaCH9LHqD208f1q8ALED6z3TjqlNFr4xBMN8ys8yEjwIaMMjrjxIsaVFsVVJX43cQvPP5fGnuDf/vqeJ4/qhE0e6OqMasTFjEdVztZpWMZA60zYeulmFRCwk8Zvy1utJMp0/yD0aPx0RkF9akDeUKNiDkVdgYz5rqRVPcY1lGhdCZylYtOYbJXCYBWCtndX0f4JA1Kk1qK5K0cYu+dX+8lx/nUsokYy7wDajPZb2EhZN6FYkCrag6mMQ34R28Dazv2Fn0nrVl3EbZcwzN1U7KolbmiBYV14QwYuiyeb3Iwzh0BdJ28eHAZZlSMglWCpZTNBW4//p7MNs+5qhgqJ2T7SLhhtPsp4XpFBpM3EEySy5fCZ5eZp+yulIrfWe884zIObJfDY/1qh5cRA/mRqFQewTSLOGB0KZfHrO6xof65m7iWmALyfqV+ry0X51vw+jX6Xisb3j8AJHvj67ufRJm1aWove1tTp4H5Muv2GMGxyUy5CTlBO2o2kxcTpFkkr9uT9xQHc6wu7qyLhuuLeDFRwtwH3PJJbZis5WNF1lHYdCVF9bq0yYhl6ij7zmZF+Di9lp+hl4UjVxd884G4D5/OWebQZ0sNC+L0UDYBD6Ni+MAcQ9hxKlltZYn82NC5A/DkBktFxdC+PQC4pRWBAulwNMcbEgYVVWG6klHN68Xv4m1x+Sykdf35930jfQp8b5xZRmQGHMKIt1Kstnbvi45qBF6IRBD2V59DTt8Ac3ilfBrdXW33/n6UqQmEzuWo9JDKz6JQX0SudowFRfLynKISDNruBU8E+oFJ7wQnIO39k3ctc9r+1E4PH9S3B4fe+pCC1ZSOoRdTys3lcZUFycMDWhXZkZFvE5wn76ZgcL2xOnG4g0e5/qmLqU5fMxs0uQ0Jgkw1KogGLNF3HAKpWkeTxx7KGtvVwimw4tw6ptAyO/pFbm1w+PXr+/ZqChfdzg7SNNMPj4uwjDBIqETbaijE8eHxnTGmfpqSnWN3cb2n29ifW/Ya7kXiuGmKcWPb1XVI90Ry5idMnThO8gWpXdPjqobgeKSqrZOGoKW1DT3Lukek/MswP0OPcPwkg+h5qD0E7F/HBPBCpIw44q+sSJvQrIXjaURUQSTYJ+XD6BcwDTrVy6v7/wBuCh5m9z3xjB7ZIF2mwyNVW5uG2Z2feIfKoRxRrjEdI8UWo64Kk9GNnqdfntuz+Rtam+EH0QLeGVlbVpoTMA1dHG0gfH15kRV7DOhQWlbT3NfKvMZvk4v5wevO7+DvW27yXSSGL/MIDBw8XEpvTcghq+k6goCg7E7qJgxoomWupV8B39Mq7Z4B1/GOw8thF1HKxfEgCBaUcNgcUQrV8Dp+/VJY+kCykM4JiLgC4imHrrF5u73AjR7+59jh4Lrnr4eKcwGz9xnMzhzBuUK+cZiZtsemFBjxXiI4JA9EF2pa/24fROXr9Ys+t+basqaeyTQbrZmAjCxoMiczn8BYFAvoNVVUeYP0DGS0xDIYPEVqeV3vmxlWfx5Z2fXWV3piGiDqEsad/uY8wq8F0qa9KqEBO2gN0/dzZp6rAhDwx3OutI45AP2pQ6HfApt3VuFw4AFLCiu5YCdUIrfdGrZvQouGF4cmDLHaKJGWbm6Ol/e8R3cw0dkjdeBiQTr0Yth7EpFrFlIMB2Q5EqI0Hazp0uvsFJKK/3K9AzntSzW4Q1TKOYE6o2d3B/R4opURbcbfXJcgQ9BiobkDnYFmmL//WPg3CQ/ppnPcGr7cZbfg9ety3d9BJiI0bBQn5o25eQN4WviMKRKE+WgpXOZmKwrG0BXj2rS27+234PXF1/A3qXMEWRXZI/5xobCZHcX3aYpd2JAYer9EZ17/qXk5vSwclbgcVY0kAf1q+vOtp65RFB9usUoCa0my2NvOQIJ7KGsDKoKHSjnFKbRaWvtymcbNvelPntW13v6VUbasVsM61sWnzS5YY5Pnjo1dtkCF0IAmK7cgxZY6DZ+fiAYPZ2e25d6fbr/R0IMGUdWryXB60+N4G0yAlhx4qyOiLkOjxrRHIdQIXqpCxv9ij6vOixsbb4aXvMNzDwxDU+LJqBlGntzz3AFGVsgZlr66RgKHba+fpPezrr6+tWMFEFDoV+35UD1LBrBQ8SFpGMmZ8MUX9wMmpwfUGWzoSMFuH1elVattavxuPfgsFlYcEb0E7MNKXncxrw8hiZbJF9kKckoXAh2UkiZ089e4nGlzbpCxu+YkPJ1n65vOO4G5t55BHv5+OGFk/2KYfdOjs1tG3z+cZc1I2kLhNgQO87YiO26Ok0Ckd/um1A2UINW+yojhhifCa6WHs4C67md34XTgVs5paHQ1KB1O38wt083c7y3XykVoY7KCPsp4WIHG7f5LS8GdVSEiP1TqKR74JXd5dvTr/5y58v5dDr9FjeBDS697Tutpius28hGyS/tZg627lt/ReXfedUCQ6HtfNLOurqAmzhyFn2ok3HjtwmIbPGpq6vc2EpiwJWREcFkxsAUNOpYg3x3X0HPeV1Gs+VKnszdy8+G3UWFZw3XowPfK3AxYwxdC0hDNowrq6Ap0CRsAqg/U1mlvWdlhzUyE+eX+gc3ZtGIzAq4KYvP8XVZcnFZEINIGFP5S0UFx0J0mmSvCMz+qvLZ0afLq96D97xvwAyOR2QAA9e2xNJSGisRb9EU/xN3RTlu6zBQgHQBQz/7XTwE2D8JuoCA55PkI0CPkUsU6G1fbHFIyrG8luPkpdvutv0TKIocDmesFKGYLRNjyNBbaHqiSmtHA7+6fvysPBtJFEUP+Fj7xCVD0gLta1R/UmQnXdpSr5skSuNWilBAVj0hRXH1t9qbuHye3w6yceTdXA3wJWFZIZL8M3KsLmBgTZjE9pQhqIUIgaxs33mpWnSaO1/+r36wtM2lF9RTwSJhTwnLe76Ca4q0WJdMlXtEUBRQuEgEc+QjvfOtyZO5ruKii8fu6yhqGqElUGAYpRWa8Idcu7aWBxlIyA9MX+BeGQ6VWA7sTFdqb4Li6tq1u3QK1hf1cFAN5Qfhr6vHMG4p8yY0f0TQlqjCQpIalYbOlKU9FPbzr06sG1iDD2QrbpjVJkkleqns0isdufL2SdFe5u4sIECz+HDs/tV80Ukx8/P5Kg4i2wvtcSWlLVIeWGuOTSCUFk6V2U2SPVODsY1hxYX+s4KYBVCG66dzux9YyorNv2VF19cy2bEpj1YELhOLOc7UPcnsoC1Y3QPaw3FVS2Z+6qzQ1QyiaS9z0whBAR1Xa0bWNH8Qt7cUZC0XR2WIkx3s8TvIzI/WruX3m2urAciVLq7YutPHDaDf+EwAACAASURBVJFe1tKhJTe8gGbKV6J0grXlDsT4x7r983cwDgO2lAbWF6dOMA3s5Bm1aP3KchIWIywJqer+xmJaaq0meJgjdfvvm+IznKxt+LXnoPjqQdU/+YFYfAtZfx8bfP8kaxCpHkNwP8j7W0dOKqs9ryI58Pk5jh+qWVclVCiWGyKHklpCveI7XFftgYxPX6jXV3qcGr5q1e0nwnwR8nsEgGKRhKsqZtH+8GG6Ai+PJDWPh/is0XTsrthy5bfM6ClffVKfAdJDEbAVW1qrcUTSC80+roj0BuVOmSrpAN6ugTNJLxSqr6Cp9p239fru7yquyNCaGO0saxwZYvDazGztGQz1XkSoF735Ch5CYso5TX9W+84/Yn1fzcv21X+aUTH4tFjhIIObgeVU47qWnEXDrKBQ7SXPXroloI5VVo7+FL2+pdr9zvrq/loZCqNYUQ1li2ZC2X1iq8rUNH/DDhfAK2qXU6WzcCiwSq6q7iDt8O7HkL9evn3s3hLRPj8tNDPnPzUtmiH3FTi5K4c8w5Lixi6MF/efVMZPf/4o3dopX30yt0fmOsZqJsgrlbDTjXG1YA+sExPExaZeYa7lKI7WDHk6sCyaTsRn+FB95WshsEqINsFqo4jV+5YELR8VmAtsVIllCALZiZZtjx1T+XIVT6btAfr9Hu4VyP5KM7vm+3OlQKjx2jDCMi17PjqjhvGBgHUrt8923b/pF13Bau58uTT5DD17qfedJ+qpXKC4IiPU6AVZBzFty69F1BwLicEk1g5gbw2qqcCbPXAN83RieX4H/95Ee2c9X30Pb7qEOKTir6E1iFLyQvpfl54Nyili0pxNRaU3aRw02KUwTFdVlekNLCl+jqvfe+ur02i1ivEY4d1SuRAPokO7MZGAoxT5n5aF70IbUkby4SCuMKV0h9uYZUZ/vbX3B1/M7fcmhQ+yodGLnAc6QFZA8etyVqCFBnamhGCviA7BL/ZQX5NLaLlyXo9vWeSvSlx9DheN2LyR2moQRQG6gbIfH59TO4+4LFYok9qfVPoK1h67glJcuXrf+fZBXNQDiwGyUO5hkvUbJmf7Rg1KJA+9cDoZehoxtRau47GmpiR1N6f1x2k5jSF/DmeIPLSRDJ8Y5mMbEr+pwU48WiLxCXs2UF4nnd6DVSilKCqt8nNcfe4ORmzeALUiTY/E20nCDH1SiiENxxDUuo0Sc5TFpN0M40ZfM/1Q0lau4uq6fAh3n9XXoZLd81Te14QPbay7oekYVFxx8akFQg0tutkDJVWmStTxgc3vIMXV9Uks7Clf3U88Ma/HXVGb3ES2tQZFtMHjC7zGlSqQnSv2Y6VniSoqFaY0NUWUm6tR3qMvdeg53hxf28lKmw0rAm1tnpQ2hE9EmyLJNy14UunO9tYLWYLJ8fk5p/QZbm2ezMn5Ck8gaOzSNuvZlmfZbN+geSgtD8MEqwCesTk0B8Slm2/f47QkrrJTuh/TQ7h7f/D+4mnFSmJ8iFwpiLKjpmX751Vmq2GFJPve1vCmTX9ZVUIJgcX5qryFcz/I2b3lJfTduF33w2GF2h3ZCnaLTHOU/Vz/3AjyZJ5CyyouTECtYI8Dobn8zshXpcbKvBN3WVgJtePqdUQUHO2a7kiMUIzm01Kyt1YrtEEUKiqR+qApHn11aJZ7iP5GIQ0Kb78W8Z336n54VkDxDMeIsgcbvfnUkoIOymAjYEfJVHuBXI0eeAQJVcgUSdO36TMnsCzespfr9foZfQbC92qXN7BiPLRP4oYAn8jJ2eoRtGAs6HAKuwv1qsHRKF+e/1t0IEsp+vZ9nOjVnlIENUbpgCVC+FaN3ghXSMAUBGYPihRqlI5Hf3nF8URB9fjMGUvy1e3Ffeee2U2EdD2jMazpQWzjmBqcmJDUlLlsCobaHgmnFFSPs/ecshOIXVdX82lNeDv5LrX1+i5n7i75MuNSu98D+8MmVOm+kawCi58E2R5hKSsjNXt/q6xLUAopw4/gnMiy+HlNY5y36jOoigELXbh9qVqV9/VE4qliIIdFVnZkZrYR8kIvz8qV186p3hmJfo6rKbAq/8HXecj3XRU7Y6Kx+iQ4R8SYNkx0pbKyIlGoednWdNKHnCpES7aaTqdgVwYZyzned57L9jMw5K8t2MrDxjOKHi2w4+RFp6nBdmQPIBJin6nsasxlK85C6KmsnBpFEA6TAV3N9UNextXlvd4cHjJE1Uoza4Z6n5T4SWxgMShDZ9g4yRKzNTI5PTo4dYCs6KAyavY8x5XSZ/j1EV4fx1UaFMAgunJaLNSvHBUPJBIkVY1oD1ljOrtAGpjO+ZvQUDmz+d9QNWieTJPf/n1WZeXBDlUmSmUeHz3rFWKAs1QfwvDGgsEu2AIv2qiUbjubm8zQ8RRIQBfooFwu9RV24i5Nj6p/Tn0EgTHI2vcAzULqmOMGHBpq5YVq4ybAIakfrUK3jLevQFZZITI5134Tv/a/g/dDsGj0UIOOsh6RCtBASd3HmJ7LKxrcEOEYprDsk0spHRoeu+G9zI0xvYF090rC0i3hmB14fddbW5N8J5/hvq+7waBZjeQTy8uJUppf01a1qRLrtdXYFBK0nXk9q/GWMYwYO4NCdL6Dj7/l2od3U6/vjCUl4niwQXOlvwAP3dU+ELaTxElTbmVW/IU7IXaQG3MGtue4pzH0EJZkNbpR5av36yF7BbhTUh8qlJ2mp7KnG9fkF8BhMDKLQFAZazq5/pnrULp5fPcoV5WiYTTjlK9+a872ddMv9f4K2sdc/1otRpT4lLTVisK4KIonWvquXN2s0fYjO48rq4YGfDQqP7muKm+hexyWrq9utY3JyXEVxf9bW7gwugC5tMb0JvB5MWwVnsXq7U/qhK3YwqCr1FF0RI4xmXH+yvAFKIYTJ/nE3duPYIyDV/uCSjIN+AJpgS3wBSipBujp1FOuulMO+0EYQhgoo1NhWmKJA2ucvhBXhCG3tKO33sF7V1ghwQOHGfQaCfCFuNx/CwyyK0cuq7h7AZJDwe4Gjp3C1UseL28gTogewXFOVmPBZJQe8mlxtTo/LYclG7qym0S8Y/KRXy1FbRAyHxUMyq7MHrl4nLBKQ1MeQ66p5gs4Pv73kdTH6dzquDo6d/7an97hLF8N5mmJBLXVE+Wf3TsDLqNajmAPhP5ZBDd+YHg4JCsUC4+DGh8nhbT1Rwjub5s7024Ea/cqoXHPPgienRZ9w0M+BRAebVBbXIZFxcsUtQuwogmgcbiAnNOlEkWCL/jVDz68J/jjeHkCOaIG5u+RNRALjPsVjyn5sto1F8DxQZJVdkazFxBS82FN2Wp0+FRxtVy2PDFfefZwwWEN7Knrlx4Iq1Muy/KX0wklFOyAYba9D1qzGpA6gK3rsaBb+bDkwO2t9RVN5Lmw8sp2sVAd+S8rVOOkjFuUbTVEesUoKfT1gMKEMU6iCvDClNvnukry1d/fqh98w9yZnSkH7Aoq5LjaPI1pnR3KNrBocPS6jVUyoV1NYMWEITgmSzyVIBvny/dI76PGRa8bmuT/nFG2M9lR/BZl+5QUBjbt0WUniUWaoFTIQjF7Mzvj6zkLi6i0hHT/6HxKrprewrHCGW6/evRF733PIDTs9VMYsXVaSTvG5c4b8TsS3G2ku2H6bDi2b0ojiExDQSTx+dCW2UrNB5cSdHvi6r7zpODQ5atBMxOzqWjf1D/B8CYFmt0E8Qm0PbQ0J3yqgug5lOwyhDBGVVVUMDz+gefOpR/8MV/djyZ2iauFDkriKZc2tK5l64WVxlC7GggaU+/I7+AYCyW7oHrT6WX0zmYq0p0+qPkWjsxneFO+YluSKq4q72GanhLY55fYcWBZK2sBW/HkBom9s2JndlVWqB4Xn+MipPI4p66sNDNn+Gpzlnrf6KDvP5PTqG4nMnsUa9i2BBhZMNMauE26uaF5vO0TSHMYx2eCGSiuMG2emsBFopr7nHF0wutrcyCXcdW5Ls9bgtB3HKq9yhJN690NaTpCziqJE6wC2juIjplBGAMMJhvDbeBoClrs6rgqL6HTXLXLO3S244CszgL2jIeSv40Xr5unZS5LU1M9lAhqLwK1VeeYixI46nXB9eYYG/X1o+phfAQXz+gvJEF3Ok+Gsjp0GLSl/BA9ekDv46qNfIHZa55HUDT2zt6GJ1wCG8uopswhxtHUuYomE6W+EnJtY5b6/VpmJ2GPOCjVL881Q8txOIjFMA9Qk4qqIBKFXQRjcIQICy3NTcnqriT40T3l9ildjcrf+fbrLXv0QK3UvjwdWeK1NxGreDa6UbT/ZCo7ymNCadkwWEz3EE3zPATMVRWKcmE6qsx38Hordfu5+840j9BKmFoOmgY3seG1CLMyQx0hpLLFuHM/IOrUyHR+8gg4NjwvNa6uP8v7h2ZwjivtWXx5eY9+bc5VZI4BNAC5YkcEVS7E50GzhXBvwGXkeUToZA9lNeUq6hSZRoAMWs0ziOr5o8z++P3vwm/i8iZcFCiMxvhEuT6pprAecrGkDh+XonmY0EVcKMdDI3jJUVyBGl2FUpZydAEd8hXjDL/Wte63z+pre3rjQeWrKP8enEcflXeZf5Z3hC+CUQSGohIajKllaPcv3HBcOdAc3YRUPeoCARbmbMUP4XRaudKOvpytWxtVXImoAI3jtfhJXE1V4T/Wrl1HchwGGrB+oKFk4sVhgMssuCcXYDnYuH0/MMEC9xn7Ewvc315bJIuUbM/I3T278wwFiiKLxapxLNhDSvKADG3bObkCE3VcrAMB7ZgSigcwcbUuiZ2SO/RkNoYTT99BzxMJfwFwrMgVJqiGdbyZn44wPUW26sBlH84RaCNIaPKLDCKUjmawdcroFFdTPre1vlJ90Tyc+Hw1X9SrNVcop/K+sDoNFYVPtMWzBPs4FDYI/akFLrPAHHlt0gAMkqtqBCblHCVxxf3gv9ggfClPxsvcRgYSl2CExmWnORws6VJzI9PTsZORPJ1Y1oAeunPNDW25UVElAIMMmJPD0ye4MbU3ublZ38HkJqnbP3894sP79n3C4ruHOVe+f/C78cf2sCK/QILs5Tzi7F6EGW1FKbFQgWa2QiwQ0Pz05Vo9Unxd0zUl4/ny2rjyKlgBH5cg4v5cMqDDqSE+CPGhVx7U6bQX0ZO+jZrmUF1FVfGQ9CVVqBZVVGO5REeVUpLvOh+Upbj3tnz11ngLGV0QQdpCVaCgw/iKlMayHjKPZyzU6lT0rcnKGbGhYhbIAxwL7ukFzD8xdEUXEXNnFnT6Aut7O8MiCgXMh71mWacUi9hgtHXqoTzP5MdeDTagRNudnQjy/gOHlbKrHNecBtnjxO7kiFJune8fElefG8PUZ+sr8qYEhciKVcDP7MhFUMgw95PqqCZV3xYoq7ZXVobub4cQPOgqOkBtASljpXRl9OoaSwj5x8t6HK/D5lLjUV8/u0sSxj2QnabMHTD2+48qvHCaPiTcUAFiJLU7javiqHLGooiiIFM9mZdrkovMnDXoAkdbaP9biI+loEGKGUfBYiDELqjV0HZOUNFxElcrWiyn4/Sg6NAoqzvJ7Hwbk3kH31+ps+05nnxZiNLzZ6YSB5ppPQqG7Ivej1ZSoJNitD9RL/DFw0U043geBVI1xa0f/ZuS/pBrBsuT+XGw5/X3M5iMh97/GNQhlmeC4atdEh5zsVuZUQsFENO3bZtGITxGIRaD3WHamkjVuZMTQkxd6Q5elX/F88FX3EF/UY9Y+H+PkIyxHrE7i6dUr4sYnwLIA8byZyj/kUF12Z4EBgrsBXfPyXCQL+D9T3xM+eOnvoOfr9wNt4KYpVpT5eXJb6Gv5hFYIxmN1ZTs2rRP450ifODOSlZfywWXimrdUeNnUlQOKvlW1O0N88G3pqJU5R1pm+RSq1sd9oEwu+lJZ7UrrbngHdjIiYGAFVNiyh2bVPbLjCzkfJV4iJprrCkf1/THAO5Zt/YFOAN3NRdMmrMBcTBsq6Cw1YbvKLbowl3oFLnCCzicQPY0V/EbGIURKvxibZlx4ySuYuI2h95BA/W9js8QeDDIjfMlCHdPinaGrcL+TJBYxyox3g2Go93MIIoyq+lixOakY5QvFlSYXHrm2mqiJ/H+h3sRek32409tHP6Kuh0hpQhDIQgWjo2HIYACG11rWn1icGOEKGRkanDQtJ5Ykay4qIp1MNFxTWV9RXH1/qq48v5iXSRAiWGpULP+7WtbLtnSNao64Fmd2juNyrZyEMDk4U0yTWCOJE5ZuVSn8Jr4pOS4jA7kYVydy1diEEuWG1xdjUa/PtiSPWwE04yyADT+jSNl25QLbyD6P+xsiVRMcVS5PeaUZYKqDK46rl7ilxqYyhd4Vzf4y+ghGoOVyp3FN9XMlowusnJGMbQptCxwLONT1FXVCyjleg6f/HOMyPCas0w/+KuwBfg2X/3+gh3qrVHs+sMoCqIjtOyPBKF73eeCEl8/iKLOibUIaJsICNNJvW7oHQSAUlxxiHGJ4DbhZfmi71/iDL/P9sxm0sVSMaqCMu6BMaLFjkqUaXwy5jqxoxsL1ZNSpmm9lUmqBWUOEfLCHxNKBv2Yyn3n95foFHm2cLkY/+8s4iteNyxH63c7wt56J/WqXd8NmMo3rgZG5exZHHR9BCsuGg9LqWRnFl9VMKS06F7qce98Jl95NDaXyk4Q9bo5Hl88goO4eGatwnJRvu/Ojm0giyZ09igUq87ioHwwjgZbZQdI2Woq8tUvIX+8P6/74VmSLxhzLgOGwtQ6HDFEZfOt2FAqsvrwfUgxz9+xLkxnyGimrmI4nUCY3NFMiYkethdM0zJLvlKs70XcD/YThJuL4cVYGm3YSsWwexmGqIbzf2pFnhdNdTkJ2ILkKIuEMm+B89M1pvL9S+mWZsKv/vv1Qs62V/U0UNnHUQV2dGl3l5lGGsednBQv5nZkqduuExN1giOkmK6C1gWQESoMoaEcUVe9eMhWyfAZ3kvK9mNnJZvfQmIIqjAH1uOusseA3TdxmRpUWKBXf5u++Q3kWkFcI0S+o6tnNmAP0X/n0s4xcb5Cbj/k1p7DRSGeVu7pBk3rXpEGv6Gm9Uw8LsQXTvSClUwv15/ymVyxPUmRJQB7nabKl3DRun0jbvjYHeS87pUc4y3I51W7d2dLl+NKyipuAHsGkYfhTBfIU1Mu2HcuoGF4yCQiVpXCklMVfsrrOFoznPMFeDtYfZP3UKSOddolnvI7GENv7ZnV+xSGXE2XMBYEBkMz3qyaJueKMtSWCjZZzev/5brYfpCE1Vp4fW/f3kCqP4VPO6rnm8ZVzU8brDnCqAVoJ6P5E5UV82FM14wVkuL9U5Y/z5mn9ZSm7XnluJp/qj7Dr+Oa4e8zILsK04ZQWr+ph5I/VHbsCyFaqkJ7zLn6hn7ZqQA7S1pFpRrbvREevbsrz+JtQ7N5BtOtyFcv4Mkw51E3b4ynPKY3x/unbH7aU+GupSiXn43NoEOjrKfjtvNlunz4kjY9Dd/A2zJ/GAwZBoT7mpmtPrwmW9XiobCQ8KPKxlQbuvnujeWarpHVOdXXsEx9hO6QZQ5RC8ikWU5TYO8VHx/Twke2Vu4GQn6yFvVK+qBlLuKl2RVwyex7xt+W+w++Y69KaX0j0UrWIDpn5GGI5miTlQBW1xxfV51KVA8hHdZtLduXrNcHT70nNX5p6QYCasDYwSSizL7jO8yvnkSUJK2OweP2vbeoRNoOKh608GaY65Sv5PKZ0CpOa6akvtD3JakU64t84gSNsWslYuWyp+phQ4uN5Efs2gwIq6EJN4YwL/KW22x5Rzki+uYiYQmubm1uVFrdlsRHpbgoewk9ofsRPHSbdKFSPTc4rsK4k6xG09eMBXKlgoXfRVYE0x+78TsMK1uLJsZjqCG8HrU2OcCW+xd9ByWuPh/bx0GlwIZvxqnZHtd2eMPz0wFEIux+5/JKpvJ9+7Ar8va3jCJ4Gmi7wCT84rg3irjJaeXbtz6Dy/rHOq6emHl5ENQgtXrxUBYQKVof9te5mDkk6b2X3M7V1QkxHSeyz7iEsQJCuWY3YGhyVUhxnTDLBZzTPMMf5+l8JdiChw0xozHsaGaHp9tN3V6mN6PS2EE6bqirHOiOnbJoOwB8SceBSZ7B/BBOBvqcyi/rUV0pU61HtdYM7Lv0ZL4KSmjn5pkEBnT9lL1Pw7g3Fhx4BVWcdEX4pG9XFBB9XsdTeKivU1nFq0gyYY71rMY+fvQll6H0Eq4ZXvPVZwsH8vc3Ay+m81HRriNUDLnGsCOHKSCM1KPsqWQWlM7s6UIFDOtuzpBCdXc5j7quzDG+HhTsuJH3k5pXnAH7zoce6+9/tdZWnsfO65NI5ovaB3p/IAQ2QLIp378evSAP5BssBJ0ukcAiQne4kitWJ6PiCvWYOd+9D5u37pG1TAuVWdjHYYWUB33igrIYghf+v2zfiPdNGI8y+wCdirGQaxJlj2ZwQZckLIW9nNrU/CGToOb8uQbUbaFXcC3Y5/XMlvnnn8pU71GcAbUVe7/ReY0wKBlHLOv6mpc2SI3VF4KFw3CCbwVvDTXg4i+6QKllO4eTu7rpqGXmNvB+SIuAWBJXnwI0PNHj8FpXrqsuo/WIDWqmtEGtIIDSl57DtKPUtwxvhGilRKIYZUfezrgUXbdZfdpWWDc+KI60tWCwOENtN3H6DgYvjWB+/wxq5UPleOM3g3kI0RbCvZqm+kZNuWig41gDMhhFcMlQHxGDCugBc7Ka+dd7fBndjyfyFU8DvZecVXq5sCGC3zknrhlU/cTsVBqCWmO+0kbQKl8W6mgO4y1Lor2iqbktSSqq+Xqj/L4QiGW1LI7ewUb8ypfeCMVGiRyWrxdQodoLyodVOW4u2Y2gR4w64iq46zyNiMRCk6jaS1f5tNbXb17WM5oRV/8V/s6Px1UQhpp6y8tcHsvyfjxUrOhUjs8cVdP6t8DGUl1pgnedKIQKx9+MTZm/XuT0D/pc66lZm8J8VHMCfvWFb/h372AwZbu8gBeoFwqdPex3NxxRYIpWwHHfiljZRS4xtym2vTmjxxqt4nLho0jt+RbOOZryUc35HSyk7p/wUGDyEIELF5AdIbCzKwfWg8mARhDGXOIR205fiCoERm2NwfeoU46J/m2pjnNxAZeVwUA99HUNtGX9oFr0X94f/PHg/mAw/ghBd5V4GCHmXLV8aC+B1TNFW08L27l93xxZ2AuMpVCoZRrng+LueOIRThlTawu4vofzzNXVP7M8ivoOSlw9pCcDppWvNLS1YPC7O+AGgxl0TV4Vodv2bWisJVvy2KOsp8yR90txD+t8fpNJxGzh43xm/8yz1bJ4dCcueBVQ0415SBEpPzvs8PcEX+hVCYxuYHu5rgtK1viH5KxiMr1N7pQjszvc/iBwoex0P60PqhXWy5d/Saa+ev9u3/l7jQ+trbzZu1HZtL3N5l5J2vBQGno1EGwo2kWzPqoAbSwExUFtjHJAV7eZlDJItaC5WVM8F6OU5lWv74v9wb++R9ozwMcchnwNVT2Uh4KbabMaKI3KIBKdmHaqh/DRoL7AGIOCe0pdYAEPrkOLFxAIzG2RacRMpUKustL9EkKK9fPxO2j7QTWqFB5f0DWJrSy7XD5IMfRcsHctgtAQxxbWf7QX0Ao0OTMtdUfwOmX2NbaWmb6vmWu9f3P+vYqrHz/O6qphlzIEtbeWUpTBUCNV4Ssi0QDiv6699fr+tUEynNhlcKNcUHQ3CfQFJjlONlMtOonIR7Q+hPm8clTlCzjPy4Lc3tQP/v6S9+EvtbM1g6Fj2NzAAQXDIFYSeAIBLjRMmllLR1a48lFBq95SHTMn7f7jFHepHYuUn1ceRNCfqLmR+uqP8Q1/DBfdWwMH39GLV/OBNwmflKzoinR23zaOkIVTPIRdCVaBkZZi0rgqGAv1RHChWWDOVLOGVu4H1dfywXwVoI0ZLgXtUUhXe17NA5fsPRhq0gp2XK83DiREfJbYs3Z6+j9nV5DrNrIDDUgXMLT56+DjAdmpIb+9AKvxD+DuC2QRIBcaYG77rWYVyZbalpw3mSQzS4JNkcVi1eLWzDPFHO/EGTb1qkQpSWsVWa+SZBVnnWi+S7/lO/ghv13R0BpfYHWfAh9hJc0+Mk5TjVppE9qdFmXvKf51MZPvql/g6Qjxl9Ks93e7dXs+se+Vwf4t8CfWzEmAYwQrSr3a3nm9yKv/vEsrEaXVjh3+JHAIMs3x7SjYTfwCmjs66THn1P1n48bMdAKaob2HdalR0xaVCm1NgkBAS36hYZBIyfycSs/wx1RS2nvnNzxkw0Nr4jGootyl7hEGDsy4LMHl22TkhVNvcFbJ0It5RBRXKUfhm21ts2PtpVKw1gf3kEChRyiYFcEYpFU2ne1tYn3QtxMP3fiZeU52W7ywG/18g7yqthLvCruHrcwoQjJICxa1VKX13GLrtyWRDhqlDS3fvseK8fH9IVhZsL5/VNvww1sAOt5coXe8sX5zNwBNtmNHEpGd6X7g9jZfVNeK6/iLGJtaUi2aVjYD3hVct/WD/paAgjKv5CtYXmIkZRsaKZ/Og2ovHwbOglbdg8mMhwY1hr7yqwiDfgQdgnVOKpT+ppX85aJ3lDykpFTFrrO6l2fIdU0ZmKP0U/oFXGGZ54iTNveDn+vJBBhv6EWJiqahXu0nZnd5OnUQIXJd6Ol9hDoMXyrL6tk0h5Y2DWZLXy+dp+SZIArlf6FjR3l/RkvvvL5wu/QRLjpQctw6K0d7ZJ8QGr7Wpgg2ipcZpXXGc73V7KdmFKsLZRcW37ODcLUONrd7M1pRWtJUNZ5SrNaR+VmqYs7R7SYA9n20myA11MliBrVGEKJ2CE3omPaUk6nxjabYdDgJzuCisRMlOcafMi898woVfa6+gd+gwUTG6yGtqOy48ADX15dywsdQW4btKvXkd3DQ+5ugdFpgMV4OumoYOtzfEOZTxxtd23wgcDzDSH4dA52UsZKyCfJtTkii4VYR+wAqcAAAIABJREFU/8giAl0CUqt0C3GtVjkl8wXYCm0f1quBE3OotNP8AnUy3+awpx53RGM43AAPPVJg6M0MCKZcXk/Aa6Tx+zfXKjG2Yl4Ag5aqjneI1ydV6plVWep8zpVe36e+AMHfdYWqX9Dzm0aUcPzNHddFrdI/k9UBjWE7BS52n2tPcR0E/e4m8VchwkhxjwtSabFmIWozSpxBdCDl1vKjeXDAGfhw3es8urF5CA129gQ7XWU9Amc/w882kYqeXUN1b0NNcZXI3ml4LGjY0XouFizpPLURzaVlL9Ey36XXmMyLvAp21mUSov5KYmrCMd2kZulcdGnLflZcDqQF88W7UGPcH9xQw6pnu36vVvLRdw2lZyj0PQzM1mJxxImub//6mC8atFwFUfW4qpI9P4GNYI1mt2HyVmPlItEdtVWKWkmkZnMYVp1Cd21TZumbTTepdJ/3JPfxEcWdL46QqMQoo7mSebDiM3yYV0HPdcXSxZ91GTFmqPenenQjtyXKieGAc1q8V81zd5enaBjmesbZEv2BKtxTIndI1lxx81PeYI61TlEbv3qFiwYP8RmJQdnakyr/b8XZ7QCVtEeYeZ6o7QqBkm88m5qHIzvK5pRPrzXalHbhseD1gd8hcVmQSvwSrm9QcAbFrz6qV4E34IayT/xDGgY4m4XGRtC7I0inYBJzJ3X4KNtEmaYiwg7FvXLVBarHrd8AoakqWCkRKn7+lUCDAAvlX5T4NXTrG1Sdolc4w8/XAw6Vm9CN0v8NO9Rhp84OiG9iRk2ewnD+7k0pabMeyRvGvpCX5ikxN63r34tAVKVSJeRVRNBKXBisbN/CZ6jyuh/UPc4rTaf/vmYeg79wrTzNgkMYNlRaDRXO3jyFYQTE150K1wzdBRN2NAugWTUE+rn++vHG9NlXpQRgRgblZe3QDTJeQ7YwozLyyu1xXsw4L+pVhTAYGgNlAagXNiw36HljSvbSrI/nnh+dbi7ODs9BC06j4iW8EHlHydlvxRfQgy5W0EteSW9Vcmut7H/+6jsYjJwGKRSP8XmDvKHWz2aotKxTkBYT83imCeXS1Hr26kAQgyB1dfa1XXpPbd7XgD2koQK6IBGUXjTLM7S9M/qrk5hMUIkdxWOuwdwqVWVnL8PQOdbxxfFDzwoL9LAaBhSDSNXS/l4ftN9uA2ULGDH9pWdClTA5sCoLcMzWqkQLefWXfTs/g7q/YSM6URQz7BkMnckLaLg8EDoeD4EA1x3RyhE9vF2Z6oPq7Vb5BN6eoSloaNKNs65q0I/mNTYStfU/ZM555lUlOtD2tfzZPr8Zrpue3cyIX0qhcBYcnTsQSlV3To+dMpjE9kT6yw/NiztHcn9NiuyBiI3Ve3SAlSHHvg0tYN+K9f0Rwb6vj/LKCQsY1j6Z5LigMLUYim5OYSpoAJ8+wxPVva8nQey6lr7WaeqL6UEDXndhS0a1KjwYzn8lrSLLFH5JXjlNp19fX6d4MoIsaM0avO0GLk+bedWpowvtYalIO36mRgu9hY0AGBGZRTkxdqFbzcwJcXILCM2kvKJ6PqkkWNJfVXcT5/c4A6091SpvUoghcN9cfwQZJ/Tt2i9A5Lg7pB1T2Z+qadzd9DXTCjpNXlkcNxGuwSp9KE6ZGbACfwoGKkVdBhyEyn0HX2uFtbm1A+F2NKLT4HgxU9h/AwsCQ10Bf9I82v7mXXE3TSsgVzovLxev56GYHoabu7s5lWcn/XrZlkr3+U0muyIxiNQaJQbLetHPvoMI1GCyaWbWzLP55rYZR+DQeqQKCgHRcyIM9JJnYl1mX9b7hWPOArXCSnOhdAsP5hXmQA8tKASD/EKonl9D9YnDd/CUbq0KrXLIUYr21ar5EFpEWidD5KULT9EXlPBoxiQFwFqW7YILxerWkh1au4XCWohbHkxiq84HKE+wbLzYi5ZofdC3B7XqAnI11Gr/OBfcKvKNmlfsRc35dCQserwSFMCqpFFfi4WqpuN92dxQRpzEs2GQ5yjwVQTXStJr0TiV+Eio1jE6A79y3hwnfcMH1Tsucv9mQaXp1FCsgBjfCHcS53967vG54xtuT0ljV3hBTuR5oeQDlkzCA/NL+ZMI6JpCWaEq6ROQVbE0o/kvcVFlqA3ai+r5DemPoeWZPkIWTF1vzETijG7hTN0FOxLsKaXTq2WgWpZtR+dHNCKt0EJtGagvMvuk0ie4hnHnE3dij7OxtqYmiuDrLVmBeiUIPNT8brozOmCzz65ZWTHK5Judm8YrgTRZBK6xeWhsLK+kA3WYsQYrlpzL9JtYvRpP6qqFqxnrKpPPmMc0Aw876KozV7PJtQzQQelOZBXxKvWGdY0Cj7jmGXZlt6oBvcmOOboNqsuqQoaRaGFaloYhZTzNNbXMd+n316vbpV29GryrC4lEgZ4bIbyaBLux69wsqPq9x8eCvROtxw3AbGpNlXFuSxt0/fTdE5lDKSm8nnRns6QsLHb9+kmRXwNVssrtcdRL6NgXYFDgijK+kx1K4PCtsW/uYL9oSnzWXB2rYTpzvHn20021PKV2/X1fp+TPiFtKbUNtF5izDjWl0qPaJ0muNa9Up+j3S1y0qldYyvP4RhpRyS0cN0/Dbtc8qrjHSH1aPzKf0+Ob7U7XWeg6Dp9qG6sBwr26jldO9ip4wnvTXLWgxIwZsJJR/NmuUr8OZ5xBj7qC2r+pWSWoaUNo2ymRGSo2zZX8yeH61EgMTl68d7RQ9qHNjj26qRBsmLRfBHJ9U5r2zAdY4uff4DsPhZ9vlMc3bl3TMDQjpdyFMhVeJhU7Hk8iMeBZERL14yDsKBdK+u+O40lXkJEGd27J4mW9ekwclxPaLWZWcrWdHgpHuh9q5EJnz6kSTuOWuVJ7FLyKoRJzayXGcHVzYnujRtZ2d+p162c7Tep9vB4utx7RiLNarbJSFhRez/9Dm5Wlu5I3+K/l1Y9D3Vr1fxs2EqKTs1QaWqOgSqGMK6tPCtUIbavxeNMMZ67+4gAGxzeGTMxc39zcKrWFb8Ls0X0Fk3t50QqVTDc5sxWV/uof+gJsWoZ3fTtltNEzkOehlkpNdXbdSEwV2+NiBiWHj1DBUIZp2S5PF4yDVqweCr9QSi7aRl7ps1kfoDSlJVBRBhxprnK0vPolW/pDPsNwHUy8yf1cHTl0qMcbubuZqktBAqLdOWMEgMbVQXM5CayuI1o0j1gVLqgV4n6SUDo4Vok5pfNyzNV3sPbUO7wFCOavW53MV5pgLTYtDyRI6NMLHHahh2vBeTam9uyVQ9ULQTWzt+FKkesI5hUv3nCgZJ0nkgmfwcxWNGU3O7/xx7F6NSjp6hoqy1jV8A0tjR27wFmTaTLRJjsWPECOe2wFVeYf2xtTqjAlzB0rBpv4JEckSq1ijx45CBJbYLDcZzAyr/6YLcDxjQn5HrTqCkBCDT5umBCPFC/ECYCumsfxeCu40+LjBfhGjJ10x9lNzfGWDFkACBo51RBhx940s2FgmYqWVvYd9H37O5xhUFelq1nAoW0IvNXdiux01GcfmwpX4wmEj/7ovRP4v+hYY82VO6S816onUVSZSqV6YAjM3Adm8LKxNWVepWa9os72j0O/iUH9dZVyNfArOAS3kBh2eAzWg15eR32nTq2bwYzBVXNFd1zUPqmS633wE8jKjl2gCMRIg/6Mh8w5HApz5APM/B0he86D/A7+frXz8vpXCJaj8mlxN0/dMLX0jnnYZTqrx75TfaW0ShvdufbkIs+xb7qPLApaqaqHP0sC319rerS2yqq6zoN2mHqcV0HlHnWDM5nOVUvlqhtNval+gt0xxK7H8kaKmSnQvmhnJWX91lu7ft9OOPoFjG5pKgGKynJEiQJ0hfpevcF/zfvsBd7OWAXaCm7V01Q/LTi11aHac0Fslfe6FqUTRxIU1aHTxrz1RcCJRINw9aCoapFztNsk/JbzQvI6UWMklk6BqYUzQOL3LadoMK8ER/5XL4kwNYXsmVcO4RMmrfVXbzEryKzqVt4vBHvtqGbTaFqq6wicUH4TAuUw6DZbvl1gq9DoRem79OtNXv30uphQWrXDeXcogd38sLU0440EvWK77qTKcW+r+d7sI8o916IqFWuwbr2iVnd/RMnlVinrD3cWUfKoyir2okBBc8qbn1rTqd2LKn4VWK+uw7W+KVEf1LYRcdd5MSISrrpTFWu2Zl0FHtdOobblUgbtfT/cJFGhjdazI1zWUskfmkmKxMRmz1B2Xj/e7XHcmVK4en5ooKPS3najc/IefmYez/tT8v1dHMR+cdgxJD166FR49x/cbemUk2qCv3YO0Tjs7BhaiVXj7QffwYHUf55K+ENwSqhtbptlh9rVvZVQHlWHYTz2xrv0/2fs6lIkx5lgQfkCjS8wLEvDvJVx1buhLb4DtIzf+6FvYOYmA3vbr8vKiAzZkl2zA9O9j0JOZUbGjwd+Dx52SquYXcQw+R73BClYBypgTKpY42QHpRcp7O6UvYPf4t9+ire3zv/P0z1LfCuzO07u0K5TcuDqNEKiMUqM7bnskukKZ7Ug+tg9grCFISi6Cr/lWo2cZ4SOFmxBsa9V+b1K72Ddr8/idSlWApXPbTFJj9lS2tWDwUWV19cTdSGUp/2XlfQRg+Dw4R7jtjO9g8QeVkM5A4xH3wfqYzixooep9mfe5E0c+1+1LZKbGdble4nCcHOjM63Wqx5GjycUUTTrDXA+UtNIDfXE73EbCjsG05pG7k5dGE90L51S6hYC16fTVLxYm7yJo3qlXgxKPWakUiHYk96h4s1up/Satc7QMBuPcTeZAcrWrOmeGcrZdjlx97iOmKLPNM+f/heCfHdxCuWvcJM38U+Nf0XTmDfxepTi3jGqq90dFrWCCNV1/7RXbK6oeRPsuBmlV7D8lo8NzQpbmzA+/D7pfBP8F5tqYrlQbe7V1/m96oDzUX1Dfih0XV3JNOZ2u2XToBK0b6dWFU2WYDbkUnlj0hYibuKH0R0ppuSmecVjUKrixKOKXG6VDmz+qVfSYB3zrzqrVW8eGavayjIz5gY8xq/VDb7QL5HTBritZht5T5IfrWLdpVg92/X7U3e6enkkUO8pjQAiOplJzGTtFalDcapdrEV5MpbVeIIztG9tngZOrVKvOSUbQm1WrNxt/EW5PA6Khd31lAML+47xAduvh10ureoB8HqYJiFYTcVmvbibeH8/fgeNcSwsbaMQAYnB1RKWNpao4kprV+tyYhxjUJV9ggN9BcZmx/k3TeUusxM0f9eQJDBPRafCsUq7ruK1mte/WX91lO/ctbJBlWRPhm+0taxKeQX7jTX7ayMOuDHmFpOlR4yN+cmJSjDaptnUSebqwfnGaFZgrXNtc9Rabfqr9xTwfMBDBn3BblY6MpizsyNtcx6fybp6c4e+in3omRHDAIHgIBKJCx5CU75Z4uLW2tGSkj7VRSCZwxigoGxsctirF2vbX31VPZ2e94rBZp1qSnqjMxy4s9Nhrpdgz9s5e6ERk9V0XEqMQYB8g2a92RqromRx15xUlKMgMc4zdn5H2F+v5fl3+fkK/8Ko6Osg++y3d6FsQ1O8Wc8giVJht8D01LtfmAF+S33oS9EIDH0zMGYw54Vm4DOYw1aWgmAeVrQ0Do9UzAEtrDYxU5TG87i3ynHReqbe1y9zs+9oIfqGgbntwKetpJSkxFg4gnEreHtpDkSoPDlpYyNC3WH0vx+FlfwnmR73Ddkq/TL5RxiPitWMekW8/YyzjcPq1GtO9adtt4tNT+a9fZ7mgmHwYIUzUBzRiPRNmwUwtAtZi+vS9JGewcTiCxPsmeRaTTABk6pevVnLPG9whsywL/8Guw5O9piceywGu66WUYJ7lTQA4LEbN/RFPjvS8YZtNiVcVu9+qT6zEeeOqj7Cgm9yqlVkmU93Ktau1lzBGf455iG3QjxO3UJH4VthL3iFsjIZoche4pXaPlCtK86hI/Xy+JtjC9HlXJFSEtHmxtSrR5lsztuF2dr2tV7Bn+HgGyTh461T7IrOfPt983qXPCMBxGP58E5zpxAKhCyXHefRKAyNd1efFJ26mbic1AqHCoPWn8Licc1+q+a1Xv3971ts1ao4g5Ur+whb+EKrkqQtCSXSXuIC5n9G+TjzWcWey2KBhsZ9dTgOFmZm++6wZMZLCF0NCDFBIONYGG7m7W/0gfz+OvPrw1JC7Wk1Wandcoh6sLNvLpnHbbpdz0/L8nQBF1+aXHdjpoWNSCP0IYxqKM7XL1JxM/lAyK18bWxe1ilnk1FV44vC46PFsqtVpK884Fyx5ZKxOZkRvUZm5wc4IEhQGI+DiyQIMdD86zMm1Nj2p/DojRORBfMSKLH3MmhhWpvQxWo7+Qzf79UZ5xdy5ume9tZ7HLht5DNcxnorj5cXaszZLNiQzN4gl6TJDa7WYvUxWq3SGef51X1Ey88Noz+BaZ814gOM6YMEey/UkJhZj+6vu2xXfYp+3sGuJam2c++05PDR9X3ZjAGirl6Sp8AfOulGh43JsVWpLG9xoDTCCXx3gxcmchfEdiiKOjCx/YUIE6aDYuX4FRCsf74O+Awau/Emzpgo7W23NVu9AebTCLhzI6JGoKsBQRIN/qWrx9a0EE6h6K4sC+8xRnkCI+yZQrYvDce9wrL+tMyaEnfAZ+g8B4c4nyFWLYGrrpBwre7spim53U5TdZHekuXpkvyP5JY7Iknyl9BCy0IYN9IRvVdpeI5Tvb9atm3D5J5O3/UZ55eFezqXtre0rg7KrnanVLrCyP6SRTCe3Cvy2C80mqO9uPP+DbpqtrERgV5p6Nj9Uk0QxytgFV/ArVi7hCfzdaBL7YCMol9gHjhIDKVIF/E8vvrI/Mr6BkrdwfNuVCTxLOfNmOcIaniuCnQN34OlKvRbIfgiMJQnm3n9+tIPW/7VVyXH5OtfMGlNj9q3PjizZdi7N5l/IWTgqV24niVbDyqZB+24YXXn1DzuZIJJIHg389lImtXzYxspIGG1wioilFsq/2X9cZ6f8+B/3yfz4HqvjCOqFKKWh5RrK69w0ca/gNqZT3I+C1qGC0LMGD1MF8xmlFQuY7ADYyfdykU3k2HrxkQLpjg9mpZnL1vpvP4qZTs7rLxedW/StBvGTuZHBWq3CYehurZyfm2JQ1MBNQ71SHnktghwfMe2GTsJNTOeRPkm+5o4vVarcvzq6zA3nONNpsIBtNDuilVCjblJNWvoczJfAz47XFAs/XS8jIMBouk2ZcxjGqUFUQiKubEfkujelD60vVQLdjeLFatlyvGrA91EyxCczq3sO4X42m4TJpEQ5J8qBeLj1Tjap+R/6L+5ab4w5pT9aFECnsLLsGGedj4xUCXxOoVY+wQX3XXh9AS/qmstfxuRYZNZ2ZUg9twUDL0V3ENPSFeDZw5Dz4XEYYrgWKVk1RzQt6+PXvIPfXjfTk3zBOperI01frfmfC78+XfDk3mv9FcIIlYrBqYrtd0+o8R8fK99L8kbWKK+ajguNwsu/7a3Gejem6d3ol4Jjz24Nj7R0OIkiFUsDszasVvPsKR6BVy0Vq/e/01qCc3dSBBfWdpMk6seoNVVD+votFJGbAqVF93bqCy+wRyIdp3oYwQEk47LrhVtQuFpRaZHPICMZz6Bc5F/Vc8Sai20MvOtUBpRm3MYnCDTS+jGi56Y+aQj8lP7BIeNaeHaU5n188OMC7VSjYDaabXAuaY42yzb65Wu1a5eFf1Ff7cJuupF4Awz7V3L7qbjdP1nGPGJE8PAxxBSEq5t5F4VxpuMnh1DyKiOyVcu+AOI2SbuljWFXmG2o1vUp+j7q9a3r/VKPMGccNWV+6qbNAsXicp7dWpu4Azm/h40QREvaCtYJPFZX/XweI1UyY1qnPp1fIAhHPehy2xH5ScpeambcI7tvVpnm7ecyMew+b07O92hM0b7K1mVurrh9qahv9UAu4o7H8AUWwarwui8tCiCG4P3qu3nzMvl92v9aUaPlfshVzIU0r0y66ZWc+AcFt3ADPz86NB+LhQcFLdKhhWqO2WIblO3hF4pfJOy/jPzL6676pBVBcaa13ulPtu1/iotUFs6XfW4Wv4dtn5Q8ANLoyDNjq8kH58Q2ofLReIpG7HXweXa7gWf311a4JA/ZAUdAB/NhgDDFFani5zPMqNTcDTZd/Qbam1+r7q3TXR63xUSgnJPIncdv77AjBEjIrCIMo/HpFJKR+Rt1f2TtOyEhAYvVdhBwILIfqhvbHAoc+Fl/Pl/f3LuR30eNF67qHbbntGVG472lVw+utNe3Hb8VNRlS/kmzz2FFV8zqBv0M+vULJrcMUbI2aP6xBzNyrMONFaqZuIxa8swbfJSa7kAndrZd0r+70qtKDTzPVIFwSC6ndjspJWgMf9FojRk8HEzFmwLU23PonMntlURhLR4TItZto8fgL5lUR7yAYb8W2wrusyLYeeHklget1tmInphP3o4Mls6l6F7LlWiCQoN2XcLZwPZY3AXsGT0FaS0s6KX0b1JQNAZHLXZ79VMHvLBzivdK+rmPSho5592g77ZkeMspfK8ZRgumhPkSYIjct/GmgsKfWK8X9d+wbwE6oqbZcOK0bNbwGfwPK8azvCLG69uE0BVNHy8mYNv2ktQr3RuDW1c2gRdNVnejbsvNCvId8/oVpHUGE84xRwYqckNhxQr33HJizjbDVuW9V5JTlw1Nzy3PSbzqt0LnC1Q0NJv7IgEC72eWMyZi7Y6gQlF+97UjXU+IWTWMBLP2TDcKkwnUpIZDejMVjS9jcssuUvVHX3aO8vOuWMXus8J6nshEbmR72n8jUl1B3g8NoovAIhZYQbXM48f4WHy0wd8ekenrbuhOG11ihdr8QYdLTsrfHoO12Z+ltylp79oDRdVsRIM/9u20GFdaRsD35hMTHI7UcxbYwVLvkvO9nAHe+nUHylQPhjx0ePwQkqcAi3GaB7hhXZ99vI18zOc9R18stXquGirVAa3uCrE7PaZVMnDEU6hdvUOTcVqbLIFDpGYexZOGQODmYU/ZH07fWJO+DAoTAJgzbPdqzldLtfj5FucvLa3Njor46ovqLoszCzXdZkS9exWefiw7LooADffpnHn9e/+xupYmAneQLeCZvB4XZOm55kbigRoPYv7c+9Mo+0qX1SoaYzMM8vVvbryCltM+mhfz93A+AA2IlQS0yaYXN3HXUQshDcjoT0A6kEadilTYS+1Aby35Ie3Pn/Pk/v5I/eKkcVF31qwrp7/ARXdevnegLOnatUL+x9rieth/iJDggrsUPFewJADPObTkiPosJrkEe7vn9CFWNFEbNhW/Ah/Dso6+ecn+Pwj/VU9m+M3R8GO3VXnuFW3JbWn6n5xf/bbK42oJ3QNeWuFkMrGRkL1Vg13JC6uCsHwgdUWP0DBYGKF6DHvGI8LjmutYOv9+qP91XdVl/qL7jH0/S9lMN5o47uWKfFnf8FulQJdkohGOapxzCQl2mIF5lPGkfL4MMm2eYpH6MJCqHjWPQ4gvwTP/HyBy58s17LKv9q507IdzXoGWzUnvYRFMN4ut+sLTFq3Z8ey2QgyabIZBv36UtV6gKEWSLYyqvYUoixQqbQJBWnSXOi0ZhdLzOulsj9ZruV71bdWaHxktJdcx+0lxPTce2E/AvkahqZf1EN7dDeixorWR9azf0ZGmbnwzdem/2fsalIrx4Gwwb7A8C4QmibQOz+c7A2xmQNEQvssAn0A4dwgN2iY205bVV9VSZbsFx4MPUshl6q++n7UWtyvVUQmZIS0WAIygdoqHNcm+0FaptbzcfReCaG94qDWCyJqML6HJbuARO3U3CmTls5pKCLfEGK9iKuAMe2VLde5NDfkULF8k/sLGfymF2sr8lJb9UqhK4ZFq2ZzknANYWX/SGWfjR0Ksz50eTOoE9+yGFnlaumOpl9XN/ZV6KGrOPbWmyqlW8k/IjVVNN/gz9yr57YPpKYLcsz8vb7qApNhxxkMR3S81EnM9rCYGmOCXOaaZxof16s0oo7z8KRbyCCYw8AcKldLF4L0AG77feKjikWOSZsnY30riCN6M+jVCD676MC5C1Ug5mTIGcyJiSOmOLJrQCy5VRirY/bhW9ZF7Y394r1Zc60ekq4agSiYjjTvrtJgYz7AkOqVakza+8Gb3iriiLZ4DLnpcTdKYtcDiUqDOKjNeUDQoG7/B6UgVyqTxeXdIqTQ80VEji0IZEzNOn2IqWFPR2XwK947V3f0T7xzNhSZAx7D4Teybx4lsvISEh3KOJdMIoEoT5pv3go++wsB7QqIyppL3kErlW8RQ4PRkgiAvFf2EFCvts3kxD2f3SsK1pWRGWFwt3IzLzklvSibx9Mg1IFjwAe40hakR6Eev9jCzqFAe1Uv/djhL66ZZS2YPR7PLOL9S9DCxrVqv1apZxCb7bwVzXjIN6RKSLxgkalkPCtMqMv0kLJZtl2G92HBUPn85tpConAN1TQg/BCGUOvW8xolH+XxCUznBT/kD96mPjdnHA0FnyrLrlGsYyxF9DHDR46JVXZMZ13TxOTqEKirgYtC/jfJEau7lJzGjL0XdQuRKNqmZd8re9iEz0B4e+MdxPN3Y9eY2+0gBOdaldnMyVbwtGeX/EVD97CsYylZxj474QqvAvGpXYxHJqWXvaBvrAOV6hiDN8w0OiQfpV1IJ7XfKxkHiQT5o8W/4gHwnm0FcwvDLF4QOHv/AOdxzsIX5+y4eL45foEMXZliJWYCqlHyYhi61kp6zHns++UiWaX9+vY2i+7Vf5zDm5qrOtb38aQGhnd2Hp8O9pj85Um1mjRj/srpUdWCQ5cJ5jUhz1jYv68v4pvNBO1MdcMp1hxvs/qan0As6GnCHeJ7la5WegX/tqPbRgVrxxl+kxagMP6w9+puZ+eKw0dPGxzq1yeo3zo2u7qUCs5w1+GLtXSGGMOBSjb1zYFI69jbQ5mOkoegnl8lMzRkg3PQEZphGeqq0su336ZHiU+gAAAgAElEQVS9cfh7XJF7hs8LLcDPm4jm5T7V0HYac6iiT+p7/KDnv46Cw2zwBa5XwzGqOS1u3nnTtaAR9SJRcsJgXy/aUd3IBzq7mKabSI0o37At5zN8tPiizGXXmMGyso8SnS65Eg8EVoo2qeusM7Q2Dfw79uy8aM4kguleCUFbucbuRMwcpXCJRGI/I7/p85dq+/4h2j1Oq7/a9zi3u1wrNaa95d3VKN6YkjIPOu14rQI3dHasuhZVSKBnT0tT1sw7OI27RYLRVXTjwApdmxB7TjeO1Cqka7V5FPa9Dd0Pi3sGyXzJClbZtwv1imCGey2mhGq6RWPGRy3BGGNILCLsbpCQV0IMNAK+clazWyTJWq1PJDewwcoWwlXkpiqBVjQdpkIVqV7FdFzpsLYi+6zuQfeTGdqMGt8rzegI11XEKvXg0vYXtGNMNeL73w0qPgXVKucR0TGxscfCllbkJCehQM7VDys2y1YAx0P70FSm+CO0ePtz0+P3+SndqH9gdHVMoOrFXQD7QRsGfun2yHmVw6A3Spj/wzCUe/lXCR5+XZzF+HSD6ow+1wJXIded6sVCucqa9bCliXC/U3yvPjEPZqNzea8msUUhCvK9MguOgsd0eq8e8cVE+9nNJiNIoNAlCyd5W8UvhrJcxFzAYyOPO+VaOq5oXj5RUxJ1QYfAyCMzAX77Wf3BPMiBXh/1/moCJlpNoBKIoadwCeqq2GKghbYP6sQg/tCiJyGHqx1if5lLM0y111msFS3VLLHW8a6tIjlAfiwSzKblVKfClt7ALatXpSV5dq8m4ceY/I2j8794tPegPF5B7TwNWn52uZfXsPQX1sqzlItkuhhyFjHC9NowHOXMwTTsIRugsTbVa0XXCaU9ewcLs/vsXsFwTpoGg8lw084Ng80DfyiJWOwdGRld7Ac4LEMulHcwwnxdlJptll1g0/r1jO8fLS8NaxtmLgBYoIPy1GGlv9y39rn+Dj4JkwhGA41IiV7eQAYZ+utn0ILIeyHvkGot98ruJVJj9U7IlbEaB4HIsYrEtlauoaIM1mAuRhR2A1j5VKb2o6KeIb9XdZ7MLw1QnyadnfNFl8H4RjaIHs8+QRCJ2K+JwxFQsOAqIIQ+JVytshhUgjZZ8nGUmXfOGEEXVyqUWwnliEbMgZEa0HRO+2Mo98rWq5YfMuLT2e6jfrHEZqdHvvUljWFW9xixL+wWXc0vxa6ZUirhcIzARYoxA91jNVfrnGnFHI/IY03+AqaLtR/UljqHNDv/Bk/m4yQnDj6+7J9WaAapXR8psxk8BqIyXPmhzHJCVtsM9ZvZn75Zra5b1C9Nn0IjUAIlxjWV33KxoilYPh0SWquELkRcKu6vfj9wr7i03+9H1lVv7hWdERf1/oJ2nLkS6UsI9HgmX525zFFidihHbthtsyRMudalikfSleDFfFTyBqZeHcUqcn/1R4N4G/wrbE7vdVUJ7B4zHWr/yAcovDQdBsWV1qLsL5bDwNYeyYx2ybXMMCCqerFHK4sAwB5CQLXaFFzfr1PkI9rwn8Rn+HPuh/zxxMwrENWK/HTROPcjXGkVW7jMI4ZzzJx7iIojbb6USK6hi8SYLX45chjciQ1tKP3SvNlDCFwc6KQSJKMfodyrD8Kv6ng7moV7LY0YO3kTFsTYcX9h0c54KHP/lZ42w15uN7l6y3c3BMKkeMp82SVqZttcuTbZKnh0CZ43gtqw0xkFLuuhqFcfn2e6CR4E6SGs8q4Ya590gzM+cqPIzRdxSh3RGKCBO9KOIdJ9VfoCzOXA39NStdZb9qDnhJVEulNSqzYgyKas01nBpyjtnRv7wV8yDurOOWdq42JJoaIB57y0y4JLMrsWEc4vyzxbnM8+g3gI03hD2m+D8HlXMz+JGXRlCUWRoeMttej0BGIEzM8ryx9s7QefpsOi2QaCy5c4TUbffAnIzFmsi+J7QhNNFnO54erKHalzCEYQTwG19XDeuDA03R1V8rZfKh83XKjNR+oW0tHp35fwGU79ZNSO4VYOOb0xPu4mOabHuHzaXBE3rZOWvSYBX/XrY4cd2DyaUK4Wwh5y+VZUkj/RaDe/cXuVxuUQDp/gpl6snyd7Zyb+36rO/7y+2V3aaSc/sTt0G2gfIL7RrVc6rUXSXNjC4u/3x42oZulaYw/p3PEF/lv1AIsZHTsWfkS8DfRMTKP+Mx6OyuCi7Uy9HRf9R4LmCytfgkKJeCy+mFfUf0ZDu/xaWVVJqZR/g/3JAvGNTYn1irTTaR0KVqxYfGATTwuImAoWbyICDYMxPyuj87KqiRK/Yl3XrbT/HzUDR8hEfffgVmLIqO2arYuqPpSuTQ4mDM7mmBnljbHzWNtte7C0/4QxYLIhhG8L0n9m9crovNKM05gHtbjfD2A7iXBseux1moSdaPLNPCJK3o68/3cCYYTxofb1sGAgsxjXEspjDGTgSuC9TfalEb9avcr5ok1cdGL68dFop88cV3uNFnw4WimjHWtCEEJvXmy7gFxrEd5AAAfX3ppnaKxxaA2VKFV1vISxcqX4XhW61OYeZ7pPU7269yQFV/i4F3fa/oT7z6NgNwxdYV+46GLQPoLvnGrtlsXl2ZRqG+p81dgqFB1DYA0XlIEbd5+BihS/hdvxXkkr2tw7/5rA+shXE6MEght7AeAxl9t5EES7IbP3WGgzmDWgOzn0ZRUnopVZteLjq5V9dQdnK0MIDYVenoq7mZm5ZaCbFQ/1SprR58+WxuRJgnbL5FiS4PRgMXSjiQTvrwkfFRKDbJoJxcr3N28OLFHsT71anzAxrQVaHTxDU8e+T8sEs/P31/4GC55Mw6foJ1Lny818z7SPXsy0O2iVxvFCMT+IyxX1pLNm6y5z3YYIaYJqx5dYRQjVFXuBI9kqHOSVfF5+I4oVnxajC7Xz+v5SXPQzh2TKb1ANPu5VLp9dd135CwwQ7GYuV2obQ22oTUznnmr/CN8N5ZFxvRVUx4oXZshFbuJWSJv4tLsxqNUW+YxC/V59qn6whTOw0c7xHRzpK4R5E3vsdACQr4iPMMjU1byGI+T0NB4DyWOAoxdXpAVBr9t2+M+WEwESVJ/pIhgIjcUYiGtF9eqaf3UXh/ZbEcQ4SlpJL1baV/FvZjU/z11OD4VGghISXky0vOPdoEnUJb03RKi+8f0dsD1QPQwEw80Vt+qV728/LM3HeW7m0f94Is3uwbki2wxO0oVeeT4ifUoidqUPnXXPZWeclcbBtGSWRF0yp+V0IEolcdUEs5AtTkEbimDQ8phDN6tR1aW/+k+97lt5qZZGZOcbmB6rOR/Aq/HcYWDAlguZ6Z3JNCvWzeYdfDemhcbYykntqoChMXPj0//BW2bq0SOjoFvc6uf1bfH2Zw5rrNcrprcfGGr8AWrLDjvfB+IqOyG1U3/VIfkN7r1v2lxlOwl+B723IgnvrVz3LJFE+4VAcpsEYCW0ql7Zv/aT+vpK9UrVlmd7nCrMN05ZrEvfyWa+NeTMqkE1NLXZ3KuZqbTFN4hqtazGZ875PB2oGG9iaYRpGKL7l5dUp4TpxbBhc1q5UmV/9fmjpR/EjHM/Rs0DPNYIqq7vHwTaB7GmTRNgNyPIRT7AF+uG4kA3BhjjBZJx4m28tm2zrTSQK7tnKMZzRxUb3cL2/f29Zf3VZyPX8vnXgcSAciVm7ZKzC/zqnEk0D2z3SCyZdFAL75yPCaic4QLnNJdJlDA4u1bOTchQhmjadsi4IphW9AKG4lalb5DeQctvr3PVmNdXwnwIohqnyXyDD2q72EBtNoLKQXyI5sNKQq20BQ6VlaBYrDp3pau0bSgfl9mb1sv6F45Ms89O8lKfjCvfzabAQa7U9WYaPO8YBjgRmb2gejdxukvGaHccPuyIQSRpeV6u1KplvbrmiuoAhoWXbOTlxKpl/Ru//StUvL0kNBS8vkMqOGvgEHDG8HF34eU7Zx5qqO2L3K5UrmS6eYH8Rp/C5GGf1StXLG3WlgUfN+ogXG2sj9iY7FitVF+oV9ke5/NE71z1Xf2fsatJrZxXogZ7B98GQtNc6JkfN5kbrsW3gMh4fgeBnj9j78B3BW+9L1bVqSrJkpwQOvRUyKWqU+env4sMjj7ChpXzF1j7IEHEDa9QDUW0G6wEzimfL9ylT95KvGtQswTdOJfvrCztWIAYoXgQyM40xyXt2HfqGDY6LdEP8oiTq1fPt3u6IIS9AN0rWqHKiHMZxNjgC0QywmjZ/xrlkppcMcTADSgTr8B+LEVTGtOmGRpdLlYkIZl5FzFnwRiuVnSvZI1T0U3k0oLEIFpzSvor5mNnslCN46qEmo14C83Y7FhY4sUSUx9CNWSvMI4XC8wss+Eu0NpUiTHnY9pkGAz1Cveq4of8W9D2dIdqvLRb5Fxf+guosAs8vsj2PydtDmnpH9Zp1YuHoWR0xZ/gnCLsWthndqngpuG03IouFfULW5gHbb7zs4Qhn6yuWvVisLGVfdu2zaVSaYCVdgN/2tEkNz+s/o0jwKF7k02XU8sY6hlccbrRCWcmjbxlsXO/UEBh6BXcd75XsLLgGSfvz/CfDOeDlxJtbGLYX2bs8gfYGeP/QTzmdB/xLl72Xih9XhM9DdnDWe2pL4W3zExp52fQqrhYRpIOgTTcUL3atghvr3jv/MnwPvq7eHzc71rcq7bHnXhpSwoO5PPGwrCgFfS2t/KKxvj/TlCD59IiBNWDyYcY8EHsthRHm/AEbqFhOE7L5HlV9DgnTyLixMBHtGGLWjGobS9zlTjWpTPGmIiTGONll+fQ5uDO7sAjUuqV3qUayG60XFNg76lJxQzm0JLrrLhgHcdm71XMaKj1V62JYkw8BqpL1E6TldS2whjHkB3DaGHj44yEzK6W406M2Y3hcR6KMXCxnBNrBJnAt865hv24UOFS7Ttdr4TfXsZFs6CMjY7VlN0f+fI15pB4hdoNGqf0TtPNu0lAteUKRHbvp5q5cZrLNZuOYUKZmtf8Sdkavx8fofIZbsX+6vY7s5FQw8cmCiTur3naA4LNGhPbPDLS9+B+4WFMGDzluGj+InP/+TcjvlmmkzAiVKrjP6utVkQOXfLFim4UXS+6V/wJPsk6+pavV0m2S493EOrKxnZXFRvRAV47kqnUwLJiGLuSPbsDiW+UcZkd+RCk5HIcBjkxsWEQ3wXeR8wy1mSgULpeG5X3fYv2zrf6fjD2x+ScBFgM9D8JmjdAQ6f8NAneCKv55MQo/zsgo58gPY7TyHnW7l/n/VTwFZgj0TfulfnB3mbObG24Xd9C77BT7YpKezF3KWmtWhXDHbxH+wlWwPZOHJxofI7N7AehHadGDAHlU2d25jyy/8LFTt58j7OY9qqSaylgoeGIwr/hXoWv8PteIYe3lrH+56wEZ8bVXdwY+p/I5hlsjxObrev4aJwxP/3Di8vcCCN7eQKxyaG9oEu9PUS/HJnKhU+Qp0CZludsrxBuE9+rbaO+XaThZb++t1SrJAYW4bA0V+naJJql82xn3+iia8zY0vowBh5YDGOhPAnKvmtSMNSXpLpWTzIR1YMpxxOv5tfczLzjXh0nFf5u8eyc7xmef+5pXAk2zoE8RAmfPZJj80elIdd0WNC/jaZlCA3W4+TFwLJKtFZeDoxtCy8UJSI9XawRrXaemY+Q28+NH8OjYTh2XvApwr16lnjI/5yahp7Uu9Q0XPbrJoxR+KHcioKY9m4u1qdRdnlKf3Op9qYo1F30GYQL9AzTXnkE4S8XnxRKFd+r4/Pb8A7+Nffq9qv0DsbbZjbUtgw1DnBu6w0D46EDbPmaiHTVsS3fAyRt/v304ylrQ7j/07lUJQpdVZKQNAlEj7m4jGeAXe7VhnplfLZLWN/baTVPui62ie6lW2ivnccHhBIPlLnBfkRDZO3h4m9Qs6csvODw/RVjrcUpm4z+zRcImvaJ6bhxbT9Oij9C7tpN3x604RUOZDZql3PNwBK9GHA6IMe8yhmEf0yD89G0J8XdKzOGYKtgLTASEvOvyy+55oyuEgDfOnE5l1NaTjvmcDrrTt8eXal9t/MgAr2udl54CJGhLkvnpu64OqjtOG4VaZXg9yh91btAod/j4KfcKys+FR8isZjLMbQX5agxK+bwwWQ1Mw84c/4D5OtEtWpDbT/YV5yp9yx479z+xGuJFtJKxfigQq17ogyduBiG7gqJgmLflFBq5ZCcxJQ4s0QFmyiSgc9nn+NJzQR4rqE1Vx4JDTSP0FZtQBl2ul7HvforrWiF336CGBS60tCu+kcIQ20Zb6wjWIyxu2RHf5yQCiUmJMVOBc+K2eajUxM6qQUf3aflvOJCr8CoKJd1ulbra5NdKmEyt2zffrtnmvaeEWRSVV5rStRFVN3mIJwf1T6t0+oe3j9KQpWUEm/1b1W/40WCk9gxRr1PVl6ermcaO3+A9O3hHdzRX30Z7VLxXqWkK7EkasiMAfb//WWuWafCrm40QSU0CRovXyeP4DjKIOichdkl/tudkOPpFLtobGJI8rac30B68egZ3HGf0LvTHodKe9lf1M44reExgCxKsFV71Y3GZL5BtxK8mVcmkZcVjmfXNDKukOnGgfR/FpQYe8cZ+Z1kA8Zu7LQPPPM8BN3bNqDG4eReXLJeWq/SoLhkdj4RZELRahqxiYYfSl93uiIMS1VdvJbveN38Htd1mW+UpS3Ji865n7j3YhpUR0fRvJ3r1cYrGzx/3LdvOuP8L2PXd+oZ/on9GGgIvGvrflnYB+3eO+vOnhLUzFL+gZ5KWobg9AjG/3SCkBdLd1wkNWKCsyMtIsD2n3O3ioge0ims+4s7rD2CkH8VuR+ne8WBjL1mxvbNleuxCCs7WcybFeoYdaJqRwS7Y6lZ4oVSp1stRiMR5hstV3lpEt2jfaf16ab/cDv6OjAZ8xBmZ+fbW+q42qNpgBD1gvs/WEv7gRlqGlJycqwYERBEMonjbk2jaUX1lHwxcZFBhhkgH2sE13nOgqG2XPE5bfJj7tXfixyT3/dcwBk/hdgMXmwFVVRCjEeBQ0HqY08iy4/hvHTmsqtlE4VIeJ/hEVlwYVoW8PhCL0r1al0MzBfjxrzlAoLFo83OB/bSGefr60c4QyuCXSNEbWDi1F7RtEWspNIStYyJAhJMStDn5FQyyDSi7MWaz+YeLCc52P7sVriK8i1HOJaVIK+c+Q3cmNfH/u23orfhW+rHwPDC3cYF/cBmANpKahdGgRiGnLjLg8vO4426s0tEs6s5HRvo6vgQw70i14U526+vqFPUrx+ntm/mJ/QMvJv4ilvRYm1vrRDcrCX6CxpDJ9pKlsCZhzAZcIJzzMdHZHk8yhwIcsyZGrOItZUkuDDlMRD36DplWWmbrk8DK2ZDty43i/sru6PP+xTFil1DULObwaoevOvwDjaDjbjuRIDTdYlmlzLmOU/CcI8x37hqOmXCYVhgl7aWqKE79VVhF2EOyN4r+Qa/aI3zvJwH7xlTootxcNCts/qhdCMAmVgA92BnAQJE4TZuU80iHbjPCEkWeKsuS45slVoK7Ewfor3gDoRvAxiz0ZlpbX+WtZZvMcDA8yCUXb2aY7ZX5uPMpU3YtBBX0ml58eVzUV4stFywbvI1V8yIeKXb04ncCpfMRpBK1CarG9swfJf21/c7CB09f4JXMw7pSsjNHm1Vz3ZzbY2cpq5zHclvjD3toBgf0GP/AUNMulgT6yoRUFJgHc/GkYhX8jrgoLXKcthX3XHt6Eq3c22PeMi32r1qWQ3eIrlEnD6anyzn5V7ZdM9RQytP5YrN/7lX4MTvyDbNZffNQvyn5kqYC3NWdLoRbUHgdT6cVQsW/dUZp8pDjpc4WDVHLu19e5HXJcJBC/JRs/CwxOMHWbMLxCDT4CRZqN6olTKh8otIlBg6Zq//mV3l1ux5ySbCNKD7aw0f4PEJmhnnVvZDfjshoqmb7w8Ugx2SqJrY8HgAk2FgPNSbFsto5idu3WVe9kn48JxzZ18AxwS2MbFo52Ro3vkNFCAGh3XcrBe176Fn0D3OV6ziNfPgLTbaAbMdKrgWy6624hANGycan63PB+PHjwQ0HuFdwSAflXYrrKxlxUL3HXJhWU0CX6scLVuw0KSw03lt4TeWWj6vcNEWWbu87JKIz8uOnaObh7isawLOe+zycUB9n+Er5OP6np1HsQNzxQfQqpVkKTHBtpcL+5xfn+68Z7ZjYPj88A6icb89vyoedElkENO145DPthZzDey4MUiMgRkecVQXib8d+fgCPJ6MAm7StXM+vwVp1qKUny3KHj+CsmemydlcLJ0GX3bG4Xt1q9R2Mh2/h6/PbryaKlHbONl3cMiEAEcM7ZMHkGPAxbSCqpRHoHwpn2sRbdJkeAwseptlGzgX9EkgeaAD3UwnGt+rpFwV7pWwHhsl1PZXSQmdfIVHYQ99w2BcwQboBd8j2qMIBokcKmmxPuNEZCPygMwk6cwroPY5waw2Q/LQzmq1V4r+mHpV5rff4iUOXa+2N0a+fd36uCN2DC29krl5iIsVe825Ub2InO1BaWpW55izK2aSlg6EPTmmdGbeZHljRkGMg2nf/nWNybSt6JUajUT9QRScGK4iOFYl88xQQ836MH2o+mKyV8XxGpZIDDoQLshLXxabSjJl69XKfgLEodWnj2rVcUbfh7SH9uoV3auLd7DVjRcrnFsRwRUL1qBuvkJlAEk7CHAe1vffmR0OpZoF6GpEdLp4iFbGQEPmUymX2H8tp6X8thkau6lWZmx+HW3WS/TONQ3vn0Rc0sN6nINd2ivesWHTDnHG7mPMMh+VecxkWi+UK7Znd6WeYVEF+LRArUv4Xm53intFt+k0NaOuh3sVGUGWvMKMAOCOxKBjidPgC2zrHtGirVQfQ/jyDUOw+YATg/uQzYS3bI9JmFfC9PC5NzByFojtjeFwnKdoU/+eYlffnyDKu8H6jsDUX89bDRdttWRxulk4rh+FzXNOULBCMR7teU+w4/dDcjfGSQOtETDvaiAfTYFTHHa6zhknTF7Ib1GnYKEYakePIwv3SnZeZQ7kW/oFCoLciJ1oW80qkTjUJt52iWYpzZTwtOzi9BuxmPNwhnY+D1gtJqXZPIKEiuaUXHRcqzYMOB86La7vsh+0nO1cL/q8YWqGXEIQZNl1tVfRlYOJrBzV8lECJaL9jWeYXT0+RmAMNaWSTjgsp8TFypoK7MCuuFqtMW680wN41Kod9ypq3C/ewVaiOHjV1fY4sItsJVbgqEX0qKT298jkwwe0gVfOkihIsLGTNPDMSnBRzfykAbHGEPO8ktjRVgHAItrjKij7cVrhoEJ/pZhMWQvwR6hERGUw3uPXshLZScDjw+zlO8reiL5BclBzmANHca3wkQDcFS3m/s/Y2aRIjixBWCCdYdZN86i1oKr2gpJ4BygJ7XPR0PsR0glaOkFB33Yqw93MPaSIyEyGomcr4sfD3eyzRUUMgB3bMHBJ9fhWKvjsvayn1B7WlZxcel6x15d7D/7wQoZXtwNbS8Ipr6pKn4Mxbi52zH/KLhxRYDGG2MTHZuzKHOuL51UgbjFMBM8iInYWVvjeNrv9dv1Ch9RW940Y3oOqFy3pZHheKVzAFaJQtLdlc6W+Bh0+TZRqAVoBtfZIfsyIS1DZFbq0TMs+JND1s5H+ZwUxzG77rSn+wr7aMMKezG7WfMh/4YPxbKeu75ZcV62tK1ExoHv8KDCoM107gXONQ9o3/enRLCyG3mm0wx3Y06s0FiVXs2n5nEViPTskdiVVQPEvLQZXN+iT+ZB1Fc5483m9/Lqz7gs9GZxWdTRvLo2cO7sFYZVgwdA3mdE8HbvqBf/+iyuQoMd0YuzsCHPzFEUozQVWxX4trbRhFT7VdllXt5Je9IfF4LwakAGQzBKQoSHVPp5KdL3Brvo3Z8IZ3gbGT41MyXOYueSyWuxzLcYa8GLaJTGV31yXHZNTa8WEd6CuKylG3Xl1+ljXnkxdQ8vg5f8lpXaj00GsKwtD1cyu5rKqBq6r0UYTE0n2ugeHJG+cCUrL5KynbLPP52fNanrHy9oK38k2ocxxqG8PLeTCPYjeVf3qkSjPh4I3LgwObb4PJM1/QPIh6RsAajsoZpQWm4DY+2ehS9Jd5jkNQdn0S1HpYcPT/WDBroXodvi38y2fR/9dtyOCSr/UK11dYsMpGLsase12UQhVx5zr7gyZk+wbFXuYkMEPuwox6RojONkluGLOFVftcrJbH9R12bVwD2NBKdi3+3n1pA75f1ReqceLer5HCjXXaWiq5uKWENPg29nfLBMJPAWNM6c9hjE36loMnrbCx5WSZ292CfL+W+19s+MhGPYfd6Hcg55lccvXV3V0rosJ50GyBEhEShtoGCphjfazD1zxHpGxEi+by+G+UGtle1BHE4uSVnUDzknvzS57cNviZeUuwYOfSu5By2rMa7ZrQDJfrYFc1U/wrkCbq5oT/7/r+qhk+BwhqbWTPaZj5t42p0nzXcUnQYsatLFcu6G7yPh4WHlN2rb74iocVVoz+NlEVieDUbOxV0Np5exdGchO11Cl1kREIh0M6sf6GB1JFOo0s3RNiADI7EAE6uo5Hz7WpH2YOeUTNKP8SRCjWg/5RrKu5F/fv3/wIMQo9SXfF/VBu7VPgmufWlfeVIJ8wSagYz4cvMmtq0ij7YTaY0yriBGGFB2vKNvXU+bbDneg1xpH7dDwwaQrKq0Z3YO/DXafzTF5oRvc6/mCBad+4LA0L2oXR82HJ87bGTAAbtOAoh0h4BAxjIk0F/NzTbCAT8rPztfsev0RK8CzXT4POjH+R67a3WuZ93mpULR+RXIe7RLlZ7O7B7vuBB7vrmFBSh4nZQf8wnhd5YN1Ce8N1EI0+lLX4MYecryi/KN550vwiHt9IUMh7x+02K6KA8IyzVfVMSZ7jNNK5KRquo9T2f4uCEOXQEUPXNpVyV24gEUEV6WZSRywgvgTzm7si8n0VP7VXjAAACAASURBVGtQdPgO/I+rr3497LfDAKDzU6VEt8+E7XYuupk4e3cHSs0+fqiYdqC5y6eaDald6LgCBrEXFZFCVpORCJs9CrfL4jq0I7rF+9BkyOIxyawrUcW0BjOU9I2HlGj1gDeRuQsfq8cDJzKVKOjq012DxD0mmAIRjoE8gcmlWSekMbau+J3sGgxd411LK/+1/rHssxi9k57j3D9V5ZjaZe1x4+bOMIPjMdi4kE9zWI7s9Q3+eaPOyiTAcHaYKx+6MbPDHq2sza0rL8yWY12vPllT24YOA9bV34g5kOPJ1D4rQckxZeijTQY7csE8wElezh8RTLvXtw2fOJMLNhvyvi4YlvhmXnV8c/aTbJjdqHhoh4LBCWn1+2x2UPFsx4z++x78ectpP2o7r4jaqcomnE4DzoBTY8Ug45vE+wYBg2hfjVS0j/8fOJAY0jJ2w3xAd3zPSEjNI4AOdfQT6x4f2uQTDcNxqhl8TNxLVlNUE/nYkn4sHdG6WIN2VeOTrtlnUEHtW6ix3l2GemSs7LELNRY8DRG1TajQQqUcr9q3WhIQFNFYsWrHJcjH4GlJhd8fzJ1vBe3HD0bnadGgKrX6iVjwxlrtLgkunOzdSSL6fQ3qTeiBqxm1I+v2mdo0zrxWlWhPZ6gAVpXauWwXcgfKoY5PFi8txC7dCvyre91eUyFavVZ6YBU8zg1NOE4j6hIGr3ha3YHT/UO9DzGYCKd7zgM3UyODOlTVQ9dAa48s3K/qIb6aU7/gm/hte/CWfQ/Kp1L86hPrqgN6lRPnqndihh6nlj50PtU4H0JddAvqBpzYuxoTpEcH80WWktBWEwJt+JRYgfISpIyPY+b9+q1+Z/Cil/pKHbsRm68uPXIYWHn/VH0Foj3gAl0EYzCo/TgmoGAlBcPiBZBQiSJIcL5+J4yYrXFsPT4ZB8pnuqyvP+YfjOVX8XuwJRKFY9Tya7A55ch6QG34Xp2H/w9mAEA/Rv3NFjafEH4s3mMpZei0GDJNRURzSvRoHomdR1YosEy+UFxXWX77Czsy0cS5LkXINpYLXvnZfGcZONF0ftC/5K6CiGIce/+p5kj8v1ifz5CFF3TaDmY2pOyRn1lm86YO3ZLn1deXPQdvSV+qDiVCYWVDnCcicLR0cIl59J923Ia8B5GpBAOOw8eMBX0ons6LWCxXhEikLRI7CCi7X1Mi88DYJnVauTnOy69baV29KhisrQkyLHSuGmMMNA3Qvj1lDN91VXehfGiTb3jXJTUN5r8ZkGyWwWizLXoP9VwkFiHhlVf/qTUXbBbPA+uI2saX8+q3Q/xm5oNOy+fa7W3ZidpEx1Usj2n6U2ylxVsT8NEbkijnJrGTXWE76pefII9x3ws1KBqisrJWStMOUae5RmhqXXHunNFfvSie79WJ2+WB8whfAaT9fT/2lnUd3oIgM2ivfXjjeRUO95GAnUEUH/46XC7ueS9nJ1ZgTqE9Nsxuwro6fC3KdlX6bP9rEVWl/pWEVzJtt3omGBzsVRHz9ZWbzvstCNoqyAKOi3lRqGXafOwgRzywOVEw8HSXp43wmg5M5FWllt6CPK/CzOunfxJGdXtNtHat5MfKTq0c6AqhLubXpaa2PwFqP0cVyHyOMOxGAVRjsrewOIczYvIYz7Wk07k28DDxIoyE7JmvxPPKLaziuhI8X1t5Q3gZo1Y59HHXMVVCNO2O+ajJlVAdDyqVYRzxAOBjHsmgu3CiTelaWW2srdjk02nNZn2r3XWQs/UV3jiZ80qqK/lm5sIpQudwBQZXCchgplHzDhxNI+6ZFTQ5KcN1XVmxvnAyOFPEoN3Qk+x/X90OtL4xJLQH+lZ7fmGxbpdkjlxfFDSG1hnCHwm1eQVqBo4f4HQpw7xmcMhVCKPE+G/aBj5fstPJt1ospPKa5bmBWKhdvsNpaQ+7A/fSuroVZl7ot9dtREVpi655fQpqp11F7do9Ph1Xg2lFobzqnVNC/uYkHxQ8TjPV7Moad7WoqDwA+7eu6AGpx7Efhc9kfQZyP35mPXEwDT7xqXy6i40kLOAsGFHhAACaVoW0dAxOmAuOJT/J7FkMMpUIzb5cQtduC8pNJuQdKI/Bfc88BjHH+dJllVlXLy/6qaracVcfkqIBcIrEV8h0iYVEA7UMCHbpR9NcXV7NiwPU4t8zEhLu3P8zEnOj5S1aWZv5TnUacZR+f2yMIzDWQp8B7Jj6IbsC25CJQb3T/5+H8zofHEfNoLKskj6CYg655BvJcpk1d1FlfMu1eUxe2sWppDLHLfNo9nMc48lk6/Zao9RRtJfQq7r/dM6s9VVQsas8Da+ct5534AiNTK9+QW3ywV2ZJzFwfsqZxEUeSpvS6rShK7pWhyn/M4/my7o6IQdO/fbILhE24SODJRzObjQvykco1biiWGRR+G8dBnsLDjmnhNKOJ+bEzutJ8bHTJ7Hjg5FsxbI93QuN63bVIf96OYkgo3swvAYrQ+209ZPKR9+5sqzr3lMMTaRtLpzJt0VPusc51v0rl9Y88wlpjJH+rWW1mZ0kCDw2k4Zm63b6cfKs+x8qJ3pFEdrWxahdDQuCDTXoY3pKH7v+6rLUPvsns0rYXhgNOT6kE8F1ZcHZjOipOeW/0RwX2YbQpaHL517P6XV1ysfJ6JCVHyOyK75viiHO7kMp+7i3mzAFcJKoeexASNTKAde8ETX8ZoVdKTrXN8uz1tHpekROpWwb5voedLlLt8y6Qo46EhNK0LnGHKldHK8kY5vOF6ID+IW9gK44cGYE1QmlrcHWi0vL8xztebmgVoEOtebxHtvkxSS47Q9qhu+6/e+vR0zyF6/mq13cfJslonTYiU3FkoEZAFZfycF+D0t4H8jmG/y4eboOu67NUYuqXOc0ssLEj3YLYha4Hxya7s+tq1/5s/2H+eEcTzRbubs8xs4CUcl7dHK+oacBFSEJvVM+4lPlqETL7ChXKvqYxS14Mb8pVUctu874pgc8y/WtcA/GmzC/rlhcteUGg9okGnZjmnPA2ckI947MrmgyOIwuNnbMcXZmMPksH2E9kwV2w42vO78Sj3YVOYZ7cH90XplY7We+zyBYbRV91PUj9bHSdiKpdq/4mI/YCDeo5KM/eXBoVzoZAGZ8KD8dxAWYlKcpWw47EPER1GaHPvKjB473TfDxnHoP6sxZi6u6PMLhQcUnTuMdS5GYz53vWFS+Es1R+WJZnzTapWBYloxNcHXpEd9HPEor0hf24+FV+AfecJGq5ftXNbvH2hZtH40F5aQSsLbnjzM07z6NEJ22tPpseHPfgpzgxN32xXf7tN03oR86repsvmhp8XD2+BOpQoFOe7gFfX0V+u1JL4C8BxEtUT/qx3B+gzi43viYkUhtQMvqHQP6QVrHwPIlqePnJEY3kUABuqTATRg14w6EQX7bTWv86B78nZRAXvqibdVa4HUx5KyJKU6VVQy9s85/uGTiUYA7g+KISGdP6bRnw/1Py0yEkwLaNWD+8mxmLkLklUdN9URDBn3RL4P8prkfOkqV0UQxmrhBcl5TnWw4ijJ8uxCivTptGF1Qwr8TH4PjdRYxuy8HIt/savY5fgx6Hu3utqEo1J76PbeuguQK/ua2zBjAxBmR81XPUhRhQW8fUjB88lxXD44Jr4h8HKdrmy+OowpHu0oYlov9DdCYzYvZOWq2dbUfD1fW11e8rrL9dgtyrtQFUHI5N2iINiaq7TVtvrvoGFCu+4xP9mPGNFxgIRg6qK3WCQVWvKgwEkTq6cn8LVidY3tyXdl08FbUM7QoQ+v6kaAWdDB5EMoMtaecr090jy007z/izl43chyLwgJKz7DboeEFCtisgLVzASU9gSUwbVRgwPkQ0oSdtOQHWO/6bbek+09SVEkbjGfgmUFjEoKkLu895ztiF6TY5tTN7lTKmXfK4dyv9NntvjLYwgcrhkf31UVV7Zccf7wiQR8ng2vpMeyrF91puAKU6K3lMSoyK1rW/7eZFoPHIEYEzSVkV4LDlGaoGCqnYbvFt/c7yDkABftwcspjXLbK5M0r0tXV7iuVCS6OCckYbNPGSvp3DjbzMeJKIPaDDJyxCuWlmqZpeuQYPvwdpCr0ciq2zRKSFiRUBk20N6ngyxUv7IpOXs6p0srrJcO7HQJQKaVEeUoGDqmUXKBRsT1QwffghfWbLXG39436ipW1BfCJ8ng+biLXsquQjwlN0TccDF5BpP02u5VqDthtWM2XGjjrDsO8fr3nbmjqwqKQdFUvEAXl8YphkrwJ0Lff1vQMUjFsRcEVEpcQOQBUYgnqjxtshyL639rm07xHp8rRjseCDmOCfBRqzdCYUd/rg9zsD15Xcl9RCzndvxJ75WkTa69TgyzBAnVXisjQis25jfT/baIj42ztAD0ZZzyoPtIwYBXK8n/UsiNlLjNpju+rZbVyPOQz5VfC1X4q1iZeVcFdBla3w1IVZNmNADIMPVaRxJBE1UWh4FQqcLanwNNWIL5I2OnD/Ai5rsaHN9bvB3jIt6d/nYhrf9medJmWaFEqSm1ZKU27kI8bdbNTzCd6lbK5ShxZOf906EJ1CYo2DSTYp8SAVRqhPnhfUU8GVUW3Vf0V5UpcLisIi5IqhUK4j6Z5VderPNFW8sDh549sAhWzPTqO9HRRsvVI0N5+5I012gJra9S8tq/WcwGeVLjEJpiIJX3KLYE5OBUXoi88RsUUdSxFa0JEo6LWIh8dM0SNZJQqhn4ld0MlI9BbcG+5IN/BD5pNrN9XJzXqyrgGK3kOqqu9Zjc4eAAkAEDy8tR1RZnEiUQXH1BXHc6/esXxtTOJkdTsQz/I/AaO4DA8XohiffW+qW8vRNW+SfoAIVE8nS/DoE+0N0Ojb0mTAFdXi88cuq/aZDXqGffo5M3sw6HEqEjHg4q7YeON5i88XLffNvYVNtpPGVp7qRiZIG5nzkAl/Bjjw2kojZFScKgp03WpldLkR4qCI8+u67yZy4+M97DhQCj1QEPX8PhSmfvqltEhs2Fi2VWXB2JRAz84Z5zhIHXpXTXXlsIS3hYpEZrAmzbm03pzEj0B+WAwLwjf+HYHPS30Y5iDKdzQfd9Bs69uyTkOJ+8Wl40Y54qj1NVXsMKs+VLJrrAMXQJkSXJcWzFRDB53VqWGvRjnGN4Us8ZlXxkLKsJ7hx01g/SvAutS2L86iWf3lPkEIqrdokTpurJFwytnACCvvW1JUNspQuZqbQXUJjKgAhTMNPn6XjTalhyKvCY9ldizr/BBeF69r5ZcuO2cz0V/jAHhRsqwEKIZH/PKffYawHzNcgRllMoTnDaadoGEAYRECjgOi+Ut9R+RfL0GF9IXcNi1Uuq+sgcw6l8hQOZUnLKaWrTgVHapCL0atkOpz07r1ZINh1cqqWUQ7iqV7b1D1I5LWEpE7KGsguR+G3fdV2xeyuwr1j6iSO2yegJL0shgp6HWxDkTCidwvjqIBk9WCz5MVYKbClS0Pmb+E51d6WIowgX6C3teN4m+6IqeYfal6vH8dhQHG5dqiRkMns0UCm76C8D5QFB0KrTLqWcgFfKo+fBBCuOoQI+9pcwR7H/Hu1nr20nPkNaLctG+NUMtqRat9FOQcolDKMMymW/YggMGr7YztLk27sZwwjXRmzAO3CdGEv3Q25WiZYLfu4oG5n7k9Qwk59uQ1JYSyyjVFWWWwL56YTf4G2hEMeEFp/N4XTGntknOBp0o/DwTiZSjZAiXqlfIGJ6eDjvPoPBk3jO6PqZdXdBneVnbVVVRsF2p0JyPWfeIT8ErQXYwuGvOBW8M7qrtYpCoOnseBqiIjiHTvPkKQo9BoioVsgIGEg/orXL7al3XR2iUB7CPpeiQOYOjgjS4qBmDao9akVc7js9b1dQ6E+zSqywJl+DyBVDMUc8Ed1bu4h+cdTJr+4qb7ZctQV+JiwWZjEKSAdOErRqureJjauYjsmO6tsuEK9m0Lu8TArWBIhJ0cUU4zN2fQeUftA7esM8AqSVbKDX8CpL0uC65vKpIoHbVGRzIXV2qq/s1VbO3iyaoTcpSoi0A1OhzVvg40kIR4UNwfPgB3H+1T1/Wu7Si66Pu8SWH1WYykeC1a8W118qrN4zapckEthe6mjnRnX3icI3geNLlaYDDWeAp+fEYJ6ZTjMtwYF+9LxdWiL8K76tTdijIdRUq1LAlWkv3uEoSyBUXTOCYwFPbZIl6tKESntbFQ4lR/cI6dFKc453HUGWfwds5qeuTIPVTlveBZDD1EUSATFXV15Tq47WWcKVWKANR99gMUT3rHjs0DPahT4LB/zwVxCp0GIRzfOS++kBW2Gqv76lg3Ef+skLZY4F90VIBZGDmfCVoBQv/21qDdlqK6rLXlaEwUGTl/MJxPc8G42HziPw0dauTC3waDiyY5AJksoTOyox6yY6c9b6qbQ6HPJ2xsHqtX1UcDmeCi1S0W9GoYXVF+0o0Vy4wSvQsjhl0oNJ4cGcFfIY1nxfNUfO1FXQXKPMavV34bF74mJLc9UJ4zIbS5hcnHAehNkmFtsqaJ5k2smmNqwtiwJe2sRrg0GBeocb31u125pXkbD/rOWr6trKonYpCZImRWYZ3VcsitVpHzrdsmW8Ss3nvTGwXSWNSArWR4zdkgEq51mAD379a0kIOA57tGcQjeMmzj4FAjiEvJNWmV7MsGN9YMu7qOOCMZFeJPp8OQlWBxL03EMOBsWBjr9HQ02D21f5DKLmWGZ/XM0uvtpD2SyVa1CpgiZ274aj5FX0AeLV3bRjuEuo9nGU5kf7fed/HjA9KjQ1jp7Bk2NmNUR7edxWQk1wrHElcirUSq9IlQ1lYy1LFbL6XgA/WEmuHDyHmgsfaR6/ItPQR5GizBDqGRXxKGyP+m/FIzfBNNQN8B2/JOc4zGuHwprpkUGoVq6+Id1WbVp9KsIQoHBh5kZ4PUTtNm8lD9XhZkUrUpxKbA8IHz+WH6VhttazVh8ydTeSL2VcUpp5vilJdJUKGmqUMc2hXZQfOaIQTmigjamNKtOMXoTwGncf4DTbPRypRmgqKjGHil/PuJftkuWjujfP881RsEn2pf4WfwUKgj5VYu0Sk1uAMRxC1NUYS/4HqK+Pb1ZQB12G94CVbMMz1hHpBz5tZon10X30uJ/CLos9W5s7zg5B21SUL9YV6XQuQq4DHMLdE30jHoN43y0iCFMgr6GOeDNL4hkaoPhHbHGIxsV7YpdK2a/XNlzvOJlK9vtsTzyWyWiIc49hgiUqRdl5UfOV8Al8bQTK0II5RJud0OYqzZ0/PZlgYZxNKBhJo96PNFBzHw/tKjXGCKGyzr84Fwu3zqfNMZFjCiUv1wLkGM+cGMyXAQA9VFqnamwjH4DvNI8ITSI4u51yCXdFTVOVgpcckTjuwsZS8PTObuBcNp8vqXFAkatwWnQ9gGfgr9QwHilGmOGGDAT+DKe2jqUX5vpoFo30wGiTWo260B6kb07E761uc4WeL/bBrRZyBTAeLxzd0t2MvpoySPilrnmk74lkib1cTixfk+CGVCCeCnU8kVnJWkOrH0C118BR+fnxru/Mqrw+6fafNaBdp85V1rfgV8Cl8EQ8ODJ2bV+kwiA+8Tec3az6mU8mxBHCKzc1DPEAdx/FodfU5V6IfqRMY3FdPcPxO+dqK9hSNJlB/jEYAbrZzsotQH20oanSze8s+phPoRCmakmkHQHuWxRz7Dtrp4Gqvbz6EpGjI1VYVypDl4axCOGhXta8qW4Ir0U5lOLcpJoNn47xKAGC+gLMeVML3qinqyEzMo1W7SqiyL+dwrX7SIp3ywnYOkS14V1WVrq/emCjaKtsg4kObDs9hY+JKvI0tYZrvEl6ZkKiNWLcHEcT7VUT2ujL76rw2H7xXWBuzwZIhMhUifZUNJ4FkuK/Qa03lQtuKvL3rVqEooGfn9QIrKn4CXUJ+zLVVzwPnYRgfYxKtDAd1R2b1DD5ndlUpKQAlB+4WfFkp7qpUofeqYYGoaY4auZzT7Bj5dwGoOe9jJtFI4UryvIGv4DgdvthN8FmESLFrdT7/45TfVqWS1JZKTQStPt2NabAdE7pw5IHTrKCiiRatWL7Y5XOmzUc0MLyw4OkszP8jpejycNbhOOv11VK6P6J/LPQTp16oosoHd99NjZYU8QynQ/kVFldNvEjKWHL/jZ5dF8JjBgGJDsqyOzHn6uh19fvbJChkejLnpYeV032Qnq/kFgPhMRepKF3upH2EqeAs1G7lCLZvinUViIl01TDDVomNmSD5UqCSDoAbg0DB3T8fMqAH+tX6vjovJVa2vILJIFOcaF9VwHCq6BiCbZdyALQ+BiurMG1Xm3apy9chEAU7DJFUe9RqPsF8DA8AwdI/f/v+UF/B5Qie18/g8/lnNsKyomI0NKOm/F0tTeiZM8AxskkfnFd1g1fAuT5w4IyIxxQ+u8pzmaZDE1TaVpy5FA6do7Waq/rcaxBnqGwLpzw4Rn2Y9+AbzZuxw9fAnYX5sYmb3am4IFEw9C5K6yIdEeL5xoHVMcMwHC6v7ttK9hVl45xXz+B90z1t+JwrnNKLWpvKUMY4LW9B0F3NppJXOYIt2ZXCM+hESuRp5NWTC0B07YpiOODfRv1PUS7DoToUPoJcXd3W+QxLv+++WLmXc1WqPl89t2RqRvSRZWkuqkB3BdxxqRi6RgyDnT6HLtxgiD8Gn0TUDx1o3iXMY4IxiGN+ONCN+dCaPmsESNxX81rmWqIsQoYLngQyVGDpMLg60vPhl3AFz+ewYKdfeFt1ofof8U0hF3PgbK7h0GWlL3bUiubrqyVD7p+5qM8S4SiLEc5kqdcW5IRqjwZNONpXErbbfSwq8hyVgBJRF3oAxt7ojqd5g03sBB8ONGPmI/jxoSTI2bfzDTbWe175QTJRyHep61I7Bq+17rYLEwU7xy3p05ouVYk6JRTFF07nQ20Mud9GGjhT+NT/8b5Z6Npf0uWLwnGSZ3AWlL6vjlF5X1Uk6KtKHOKU5pEDX0CQtTe12Cv5NdimDiEPBpcZDgV2mdcgdtlJzLCs2LJeaJI41Lf6+gYZA9Eycd51y93teGOd338mRaI8oVdkBv4GRhWW6h037CppV/oL3pRZnloMvk+gdtR9JVbwSVl2D3VimEXO3eP83b7YwBZzBe7G+//9tWhNv/Dne+bmf8+//7qf/8A//r38xf8pf/Qn/tGf88/n5+evz1/T33/898evaf77/tf954f6/sEj8Ivxj0RuDz6E/2vvynIj12HgXfjBg/FDgC4kILd9sVjcZLnbeZnBAImdpGNrI1ksUYvd3WdeTaQ++2uLAw/mdvycjiNpaN6Y29WAd97B1YSphiYh09PiBIkj2u0NrcJdEw/9VADzXT9SzXX9ONG8uXHep2M/E3p4ez10y2oku1ja+h6v3T7DfDBkgiVGK0X80K7rzbOphqr7qUk/LrP2qyLQ8sOIXrKnYR9tDkJq3XGM2equvQmh2qaHesSGr1nC8+ao1tslSqncdN44bV29iVdszKq86mDV3LIY+qcSVF3nzFEGJ0YH1WU0w607/xooBp45N52Z6BuWqF5pw4oEYgpvi2vI/3Cez+LNpePFviY80ZnPbx3cY3VARceYad3QjIZ3GlaXuhrwTO2mMKDBDpjb0YG1XHMQTVs3Zrbm4KVihqj5wJwzFyQDvlIGWbiAfJViOnZ0eeXbcOOcl/VrJvjNnGEuCkXBimjUGmAHGcZwR5mDG3ypggfgNFVgDCiJLLXUkW5oBjF2wC5vqDWLizBcKT4pMV2BRiEtUGsRIdEXGvwyjMc5EFu4one8krnZoBFLfCxEdwFDhkPXUqgfeqGsQ6nuZbpFb0c9Q9W8XX3B5uTw8t3iFNrsuf4wS81LwDHYb7E37LHE01DF+KwBobd9UHm1RqwgVkAw/P8IfQLMUGTkulag4pSKJj/4xajXPSX0s4KGY0od4aHRFhnDWN08shMmDO/mDEot1tXjdnqwMmmv94hhv+aO7ZSjJQ4vNOhLkbPAvoaKIqpX1XrGbZxppe/dXfaPr/rgRFRmbD+D9bMPkfhanPfxCtNR0t2uXwaVfacznxc4F1hFxPplaB0WY4NBNhTaYCW+N8MT599EKv3mXdoOgxd9kHSZLZznWT+dU0oq7LLf5JXBhZg1g11ZSv9AoI7eg+6Hz1/lW1gxtpLtECUnTuPXZvc5yf+lsqXKUqFti6GlXdOv6jRT8VLoWWG1QnyuwLzZEH3FK7FbOjYaQvSB2sRNT7AcUFGaZP+al5Km+VhgRunldybGdUNlltx0/ZVWLjRYcFZSVfZsbkt5tQKjmEjsWgnf7oN4Bl7EJmYTLVJHmN7qEDi0Ol+ksIG9kl/tOeI2OC83xiWIchaon+S6DsGzhZI6P5AgFSlYcp9X69wBH0NTOG6pDhum+vrVA7kzAh8PfqFx7R5SOzoirWPHPovJLQbMJo1dBEer1souaORjskqIvoLVjFe2iiaS9WiuOVyTUlNGy+yuIcFA5KVOKVWudyq4ApsCF0LrQRkn2S5tbvFKeyHtteQEUCuCV/XOtcPd15btfHIln1t1wMZrr+Xb3h54xcRfwoptQqaA+5RLp2qxEhCQb+uq5HTPtPK0Zi1eTgnbUqmNQnzTVnZVUxS2uoLppyLFL6jzklcTMdLvGTeK4gNp9Nz1cpn+oik694iaiHvkpvo8LgrF1I6iXS/ByXsuv4XRxS3E0XgtG1B7qzpX2g+BN2K7eEeExhUJgZ+w8azQEEeKP/CFL3Znm+PqlV16elEcLLDd3NxJXL7eSpnxIqSI2WwP5gE2A4LJ5ZPvrMvmZsR9rCCFYDjr81nTLhWmpgCVsFYihRIC9qea4VbliU7xotlBF0psphBnNzQD74JhhCIhmxC4NoQnQi2k83Vcv9EHlVfq1jmdDcqwxUDN5uR4zOeEg2mO8Txl2yQDp7ynO1g5YDK0ULNUmKAiJ38GS1VRCeKK+xkODPn4eB0JYMvrygAAAX9JREFUOf8PKwwI7DoylVGVDZEcFSFSzA5jJLrPZIx1gQIVkd0R0MxYluHxHjULSxDG6SQMoPLC9jCC4U+cfUhFPsVamel7vEI84LDbAobfFYJ/7S2vbAQyB6+8sr8FLMRHAfnQhi7PxHFITHLHBK+Eo3nIJ+NpkL7Kj91g+h5WTAviXFG02OtKAjkQhAvgnsYOZuEPNGYKfnlqeSsfeTQkGzXKcsO7lEvggJCq/GhX6Lu8Cl3RFU8rAVk7bdbLn/mStajRgmFfrkvL4M1lpcpOk+g8QusGHVtrfO4mSf4dmO5jdWaUR31xCCTU5rRXcR5ZYw7DK0u59E/yocGbluugmoBxtbyCUCIbJ/kUrJQ/iZU7sN48S7uIUvuqLN2VkoreQPU7p0izm+2lQkJXm3LVs7wljlwK+mNY3Tlkw8D3dV7QmN+3td0g/yoY/wCrf3rI32v6x2F1J9A+WP3148HqwerB6sHqwerB6sHqOR6sHqwerB6sHqwerJ7jDVb/AYETxl4kmFmdAAAAAElFTkSuQmCC"></image><linearGradient id="__lottie_element_205" spreadMethod="pad" gradientUnits="userSpaceOnUse" x1="-141.20899963378906" y1="0" x2="141.16000366210938" y2="0"><stop offset="0%" stop-color="rgb(10,11,51)"></stop><stop offset="16%" stop-color="rgb(10,11,51)"></stop><stop offset="32%" stop-color="rgb(10,11,51)"></stop><stop offset="66%" stop-color="rgb(10,11,51)"></stop><stop offset="100%" stop-color="rgb(10,11,51)"></stop></linearGradient><clipPath id="__lottie_element_207"><path fill="#ffffff" clip-rule="nonzero" d=" M205.8350067138672,63.98500061035156 C205.8350067138672,63.98500061035156 -12.609999656677246,63.98500061035156 -12.609999656677246,63.98500061035156 C-12.609999656677246,63.98500061035156 -12.609999656677246,104.85399627685547 -12.609999656677246,104.85399627685547 C-12.609999656677246,104.85399627685547 205.8350067138672,104.85399627685547 205.8350067138672,104.85399627685547 C205.8350067138672,104.85399627685547 205.8350067138672,63.98500061035156 205.8350067138672,63.98500061035156" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_210"><path fill="#ffffff" clip-rule="nonzero" d=" M174.55599975585938,-2.0810000896453857 C174.55599975585938,-2.0810000896453857 23.426000595092773,-2.0810000896453857 23.426000595092773,-2.0810000896453857 C23.426000595092773,-2.0810000896453857 23.426000595092773,27.266000747680664 23.426000595092773,27.266000747680664 C23.426000595092773,27.266000747680664 174.55599975585938,27.266000747680664 174.55599975585938,27.266000747680664 C174.55599975585938,27.266000747680664 174.55599975585938,-2.0810000896453857 174.55599975585938,-2.0810000896453857" fill-opacity="1"></path></clipPath><mask id="__lottie_element_215" mask-type="alpha"><g transform="matrix(0.9999780654907227,0,0,0.9999780654907227,464.6819152832031,287.99652099609375)" opacity="1" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M152.25,-172.9600067138672 C152.25,-172.9600067138672 -152.25,-172.9600067138672 -152.25,-172.9600067138672 C-164.81500244140625,-172.9600067138672 -175,-162.77499389648438 -175,-150.2100067138672 C-175,-150.2100067138672 -175,-92.95999908447266 -175,-92.95999908447266 C-175,-92.95999908447266 -175,156.78399658203125 -175,156.78399658203125 C-175,165.71800231933594 -167.75799560546875,172.9600067138672 -158.8249969482422,172.9600067138672 C-158.8249969482422,172.9600067138672 158.8000030517578,172.9600067138672 158.8000030517578,172.9600067138672 C167.73300170898438,172.9600067138672 174.9759979248047,165.71800231933594 174.9759979248047,156.78399658203125 C174.9759979248047,156.78399658203125 174.9759979248047,-92.95999908447266 174.9759979248047,-92.95999908447266 C174.9759979248047,-92.95999908447266 175,-92.95999908447266 175,-92.95999908447266 C175,-92.95999908447266 175,-150.2100067138672 175,-150.2100067138672 C175,-162.77499389648438 164.81399536132812,-172.9600067138672 152.25,-172.9600067138672z"></path></g></g></mask><linearGradient id="__lottie_element_227" spreadMethod="pad" gradientUnits="userSpaceOnUse" x1="-142.30499267578125" y1="1.0509999990463257" x2="141.0229949951172" y2="1.5779999494552612"><stop offset="0%" stop-color="rgb(10,11,51)"></stop><stop offset="16%" stop-color="rgb(10,11,51)"></stop><stop offset="32%" stop-color="rgb(10,11,51)"></stop><stop offset="66%" stop-color="rgb(10,11,51)"></stop><stop offset="100%" stop-color="rgb(10,11,51)"></stop></linearGradient><mask id="__lottie_element_231" mask-type="alpha"><g transform="matrix(0.9999780654907227,0,0,0.9999780654907227,426.64227294921875,140.40353393554688)" opacity="1" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-175,-17.25 C-175,-29.81399917602539 -164.81399536132812,-40 -152.25,-40 C-152.25,-40 152.25,-40 152.25,-40 C164.81399536132812,-40 175,-29.81399917602539 175,-17.25 C175,-17.25 175,40 175,40 C175,40 -175,40 -175,40 C-175,40 -175,-17.25 -175,-17.25z"></path></g></g></mask><clipPath id="__lottie_element_245"><path fill="#ffffff" clip-rule="nonzero" d=" M32.1349983215332,-4.735000133514404 C32.1349983215332,-4.735000133514404 -3.569999933242798,-4.735000133514404 -3.569999933242798,-4.735000133514404 C-3.569999933242798,-4.735000133514404 -3.569999933242798,31.858999252319336 -3.569999933242798,31.858999252319336 C-3.569999933242798,31.858999252319336 32.1349983215332,31.858999252319336 32.1349983215332,31.858999252319336 C32.1349983215332,31.858999252319336 32.1349983215332,-4.735000133514404 32.1349983215332,-4.735000133514404" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_251"><path fill="#ffffff" clip-rule="nonzero" d=" M888.7219848632812,255.41799926757812 C888.7219848632812,255.41799926757812 888.7219848632812,269.6340026855469 888.7219848632812,269.6340026855469 C888.7219848632812,269.6340026855469 930.2631225585938,269.6624755859375 930.2631225585938,269.6624755859375 C930.2631225585938,269.6624755859375 930.2631225585938,255.44590759277344 930.2631225585938,255.44590759277344 C930.2631225585938,255.44590759277344 888.7219848632812,255.41799926757812 888.7219848632812,255.41799926757812" fill-opacity="1"></path></clipPath><linearGradient id="__lottie_element_257" spreadMethod="pad" gradientUnits="userSpaceOnUse" x1="772.10302734375" y1="191.1820068359375" x2="1121.6729736328125" y2="193.52099609375"><stop offset="0%" stop-color="rgb(102,212,247)"></stop><stop offset="16%" stop-color="rgb(90,179,251)"></stop><stop offset="32%" stop-color="rgb(77,146,255)"></stop><stop offset="66%" stop-color="rgb(132,123,255)"></stop><stop offset="100%" stop-color="rgb(187,101,255)"></stop></linearGradient><clipPath id="__lottie_element_397"><path fill="#ffffff" clip-rule="nonzero" d=" M79.7969970703125,-5.060999870300293 C79.7969970703125,-5.060999870300293 41.79499816894531,-5.060999870300293 41.79499816894531,-5.060999870300293 C41.79499816894531,-5.060999870300293 41.79499816894531,33.0890007019043 41.79499816894531,33.0890007019043 C41.79499816894531,33.0890007019043 79.7969970703125,33.0890007019043 79.7969970703125,33.0890007019043 C79.7969970703125,33.0890007019043 79.7969970703125,-5.060999870300293 79.7969970703125,-5.060999870300293" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_430"><path fill="#ffffff" clip-rule="nonzero" d=" M329.1780090332031,-22.073999404907227 C329.1780090332031,-22.073999404907227 -22.47800064086914,-22.073999404907227 -22.47800064086914,-22.073999404907227 C-22.47800064086914,-22.073999404907227 -22.47800064086914,90.74400329589844 -22.47800064086914,90.74400329589844 C-22.47800064086914,90.74400329589844 329.1780090332031,90.74400329589844 329.1780090332031,90.74400329589844 C329.1780090332031,90.74400329589844 329.1780090332031,-22.073999404907227 329.1780090332031,-22.073999404907227" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_439"><path fill="#ffffff" clip-rule="nonzero" d=" M126.69499969482422,-3.2660000324249268 C126.69499969482422,-3.2660000324249268 87.16100311279297,-3.2660000324249268 87.16100311279297,-3.2660000324249268 C87.16100311279297,-3.2660000324249268 87.16100311279297,32.05500030517578 87.16100311279297,32.05500030517578 C87.16100311279297,32.05500030517578 126.69499969482422,32.05500030517578 126.69499969482422,32.05500030517578 C126.69499969482422,32.05500030517578 126.69499969482422,-3.2660000324249268 126.69499969482422,-3.2660000324249268" fill-opacity="1"></path></clipPath><linearGradient id="__lottie_element_469" spreadMethod="pad" gradientUnits="userSpaceOnUse" x1="831.0469970703125" y1="256.96600341796875" x2="1177.9189453125" y2="265.45098876953125"><stop offset="0%" stop-color="rgb(102,212,247)"></stop><stop offset="16%" stop-color="rgb(90,179,251)"></stop><stop offset="32%" stop-color="rgb(77,146,255)"></stop><stop offset="66%" stop-color="rgb(132,123,255)"></stop><stop offset="100%" stop-color="rgb(187,101,255)"></stop></linearGradient><clipPath id="__lottie_element_483"><path fill="#ffffff" clip-rule="nonzero" d=" M166.24099731445312,-1.4709999561309814 C166.24099731445312,-1.4709999561309814 137.7480010986328,-1.4709999561309814 137.7480010986328,-1.4709999561309814 C137.7480010986328,-1.4709999561309814 137.7480010986328,28.815000534057617 137.7480010986328,28.815000534057617 C137.7480010986328,28.815000534057617 166.24099731445312,28.815000534057617 166.24099731445312,28.815000534057617 C166.24099731445312,28.815000534057617 166.24099731445312,-1.4709999561309814 166.24099731445312,-1.4709999561309814" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_545"><path fill="#ffffff" clip-rule="nonzero" d=" M215.81700134277344,-3.75600004196167 C215.81700134277344,-3.75600004196167 179.5240020751953,-3.75600004196167 179.5240020751953,-3.75600004196167 C179.5240020751953,-3.75600004196167 179.5240020751953,32.10900115966797 179.5240020751953,32.10900115966797 C179.5240020751953,32.10900115966797 215.81700134277344,32.10900115966797 215.81700134277344,32.10900115966797 C215.81700134277344,32.10900115966797 215.81700134277344,-3.75600004196167 215.81700134277344,-3.75600004196167" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_601"><path fill="#ffffff" clip-rule="nonzero" d=" M32.1349983215332,-4.735000133514404 C32.1349983215332,-4.735000133514404 -3.569999933242798,-4.735000133514404 -3.569999933242798,-4.735000133514404 C-3.569999933242798,-4.735000133514404 -3.569999933242798,31.858999252319336 -3.569999933242798,31.858999252319336 C-3.569999933242798,31.858999252319336 32.1349983215332,31.858999252319336 32.1349983215332,31.858999252319336 C32.1349983215332,31.858999252319336 32.1349983215332,-4.735000133514404 32.1349983215332,-4.735000133514404" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_665"><path fill="#ffffff" clip-rule="nonzero" d=" M79.7969970703125,-5.060999870300293 C79.7969970703125,-5.060999870300293 41.79499816894531,-5.060999870300293 41.79499816894531,-5.060999870300293 C41.79499816894531,-5.060999870300293 41.79499816894531,33.0890007019043 41.79499816894531,33.0890007019043 C41.79499816894531,33.0890007019043 79.7969970703125,33.0890007019043 79.7969970703125,33.0890007019043 C79.7969970703125,33.0890007019043 79.7969970703125,-5.060999870300293 79.7969970703125,-5.060999870300293" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_689"><path fill="#ffffff" clip-rule="nonzero" d=" M126.69499969482422,-3.2660000324249268 C126.69499969482422,-3.2660000324249268 87.16100311279297,-3.2660000324249268 87.16100311279297,-3.2660000324249268 C87.16100311279297,-3.2660000324249268 87.16100311279297,32.05500030517578 87.16100311279297,32.05500030517578 C87.16100311279297,32.05500030517578 126.69499969482422,32.05500030517578 126.69499969482422,32.05500030517578 C126.69499969482422,32.05500030517578 126.69499969482422,-3.2660000324249268 126.69499969482422,-3.2660000324249268" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_695"><path fill="#ffffff" clip-rule="nonzero" d=" M337.4010009765625,153.6009979248047 C337.4010009765625,153.6009979248047 -29.985000610351562,153.6009979248047 -29.985000610351562,153.6009979248047 C-29.985000610351562,153.6009979248047 -29.985000610351562,280.4779968261719 -29.985000610351562,280.4779968261719 C-29.985000610351562,280.4779968261719 337.4010009765625,280.4779968261719 337.4010009765625,280.4779968261719 C337.4010009765625,280.4779968261719 337.4010009765625,153.6009979248047 337.4010009765625,153.6009979248047" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_701"><path fill="#ffffff" clip-rule="nonzero" d=" M166.24099731445312,-1.4709999561309814 C166.24099731445312,-1.4709999561309814 137.7480010986328,-1.4709999561309814 137.7480010986328,-1.4709999561309814 C137.7480010986328,-1.4709999561309814 137.7480010986328,28.815000534057617 137.7480010986328,28.815000534057617 C137.7480010986328,28.815000534057617 166.24099731445312,28.815000534057617 166.24099731445312,28.815000534057617 C166.24099731445312,28.815000534057617 166.24099731445312,-1.4709999561309814 166.24099731445312,-1.4709999561309814" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_713"><path fill="#ffffff" clip-rule="nonzero" d=" M215.81700134277344,-3.75600004196167 C215.81700134277344,-3.75600004196167 179.5240020751953,-3.75600004196167 179.5240020751953,-3.75600004196167 C179.5240020751953,-3.75600004196167 179.5240020751953,32.10900115966797 179.5240020751953,32.10900115966797 C179.5240020751953,32.10900115966797 215.81700134277344,32.10900115966797 215.81700134277344,32.10900115966797 C215.81700134277344,32.10900115966797 215.81700134277344,-3.75600004196167 215.81700134277344,-3.75600004196167" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_719"><path fill="#ffffff" clip-rule="nonzero" d=" M32.1349983215332,-4.735000133514404 C32.1349983215332,-4.735000133514404 -3.569999933242798,-4.735000133514404 -3.569999933242798,-4.735000133514404 C-3.569999933242798,-4.735000133514404 -3.569999933242798,31.858999252319336 -3.569999933242798,31.858999252319336 C-3.569999933242798,31.858999252319336 32.1349983215332,31.858999252319336 32.1349983215332,31.858999252319336 C32.1349983215332,31.858999252319336 32.1349983215332,-4.735000133514404 32.1349983215332,-4.735000133514404" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_725"><path fill="#ffffff" clip-rule="nonzero" d=" M1035.7130126953125,289.3900146484375 C1035.7130126953125,289.3900146484375 1256.175048828125,289.3900146484375 1256.175048828125,289.3900146484375 C1265.27294921875,289.3900146484375 1272.64697265625,296.7640075683594 1272.64697265625,305.86199951171875 C1272.64697265625,305.86199951171875 1272.64697265625,604.7100219726562 1272.64697265625,604.7100219726562 C1272.64697265625,613.8079833984375 1265.27294921875,621.1829833984375 1256.175048828125,621.1829833984375 C1256.175048828125,621.1829833984375 1035.7130126953125,621.1829833984375 1035.7130126953125,621.1829833984375 C1026.614990234375,621.1829833984375 1019.2410278320312,613.8079833984375 1019.2410278320312,604.7100219726562 C1019.2410278320312,604.7100219726562 1019.2410278320312,305.86199951171875 1019.2410278320312,305.86199951171875 C1019.2410278320312,296.7640075683594 1026.614990234375,289.3900146484375 1035.7130126953125,289.3900146484375" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_731"><path fill="#ffffff" clip-rule="nonzero" d=" M79.7969970703125,-5.060999870300293 C79.7969970703125,-5.060999870300293 41.79499816894531,-5.060999870300293 41.79499816894531,-5.060999870300293 C41.79499816894531,-5.060999870300293 41.79499816894531,33.0890007019043 41.79499816894531,33.0890007019043 C41.79499816894531,33.0890007019043 79.7969970703125,33.0890007019043 79.7969970703125,33.0890007019043 C79.7969970703125,33.0890007019043 79.7969970703125,-5.060999870300293 79.7969970703125,-5.060999870300293" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_737"><path fill="#ffffff" clip-rule="nonzero" d=" M1035.7130126953125,289.3900146484375 C1035.7130126953125,289.3900146484375 1256.175048828125,289.3900146484375 1256.175048828125,289.3900146484375 C1265.27294921875,289.3900146484375 1272.64697265625,296.7640075683594 1272.64697265625,305.86199951171875 C1272.64697265625,305.86199951171875 1272.64697265625,604.7100219726562 1272.64697265625,604.7100219726562 C1272.64697265625,613.8079833984375 1265.27294921875,621.1829833984375 1256.175048828125,621.1829833984375 C1256.175048828125,621.1829833984375 1035.7130126953125,621.1829833984375 1035.7130126953125,621.1829833984375 C1026.614990234375,621.1829833984375 1019.2410278320312,613.8079833984375 1019.2410278320312,604.7100219726562 C1019.2410278320312,604.7100219726562 1019.2410278320312,305.86199951171875 1019.2410278320312,305.86199951171875 C1019.2410278320312,296.7640075683594 1026.614990234375,289.3900146484375 1035.7130126953125,289.3900146484375" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_743"><path fill="#ffffff" clip-rule="nonzero" d=" M1035.7130126953125,289.3900146484375 C1035.7130126953125,289.3900146484375 1256.175048828125,289.3900146484375 1256.175048828125,289.3900146484375 C1265.27294921875,289.3900146484375 1272.64697265625,296.7640075683594 1272.64697265625,305.86199951171875 C1272.64697265625,305.86199951171875 1272.64697265625,604.7100219726562 1272.64697265625,604.7100219726562 C1272.64697265625,613.8079833984375 1265.27294921875,621.1829833984375 1256.175048828125,621.1829833984375 C1256.175048828125,621.1829833984375 1035.7130126953125,621.1829833984375 1035.7130126953125,621.1829833984375 C1026.614990234375,621.1829833984375 1019.2410278320312,613.8079833984375 1019.2410278320312,604.7100219726562 C1019.2410278320312,604.7100219726562 1019.2410278320312,305.86199951171875 1019.2410278320312,305.86199951171875 C1019.2410278320312,296.7640075683594 1026.614990234375,289.3900146484375 1035.7130126953125,289.3900146484375" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_749"><path fill="#ffffff" clip-rule="nonzero" d=" M126.69499969482422,-3.2660000324249268 C126.69499969482422,-3.2660000324249268 87.16100311279297,-3.2660000324249268 87.16100311279297,-3.2660000324249268 C87.16100311279297,-3.2660000324249268 87.16100311279297,32.05500030517578 87.16100311279297,32.05500030517578 C87.16100311279297,32.05500030517578 126.69499969482422,32.05500030517578 126.69499969482422,32.05500030517578 C126.69499969482422,32.05500030517578 126.69499969482422,-3.2660000324249268 126.69499969482422,-3.2660000324249268" fill-opacity="1"></path></clipPath><clipPath id="__lottie_element_755"><path fill="#ffffff" clip-rule="nonzero" d=" M166.24099731445312,-1.4709999561309814 C166.24099731445312,-1.4709999561309814 137.7480010986328,-1.4709999561309814 137.7480010986328,-1.4709999561309814 C137.7480010986328,-1.4709999561309814 137.7480010986328,28.815000534057617 137.7480010986328,28.815000534057617 C137.7480010986328,28.815000534057617 166.24099731445312,28.815000534057617 166.24099731445312,28.815000534057617 C166.24099731445312,28.815000534057617 166.24099731445312,-1.4709999561309814 166.24099731445312,-1.4709999561309814" fill-opacity="1"></path></clipPath><linearGradient id="__lottie_element_767" spreadMethod="pad" gradientUnits="userSpaceOnUse" x1="-141.49000549316406" y1="0" x2="142.6719970703125" y2="0"><stop offset="0%" stop-color="rgb(114,51,180)"></stop><stop offset="16%" stop-color="rgb(114,51,180)"></stop><stop offset="32%" stop-color="rgb(114,51,180)"></stop><stop offset="66%" stop-color="rgb(114,51,180)"></stop><stop offset="100%" stop-color="rgb(114,51,180)"></stop></linearGradient><clipPath id="__lottie_element_769"><path fill="#ffffff" clip-rule="nonzero" d=" M181.73800659179688,129.3000030517578 C181.73800659179688,129.3000030517578 12.916000366210938,129.3000030517578 12.916000366210938,129.3000030517578 C12.916000366210938,129.3000030517578 12.916000366210938,165.2100067138672 12.916000366210938,165.2100067138672 C12.916000366210938,165.2100067138672 181.73800659179688,165.2100067138672 181.73800659179688,165.2100067138672 C181.73800659179688,165.2100067138672 181.73800659179688,129.3000030517578 181.73800659179688,129.3000030517578" fill-opacity="1"></path></clipPath><linearGradient id="__lottie_element_790" spreadMethod="pad" gradientUnits="userSpaceOnUse" x1="-172.4530029296875" y1="0" x2="174.91799926757812" y2="0"><stop offset="0%" stop-color="rgb(102,212,247)"></stop><stop offset="16%" stop-color="rgb(90,179,251)"></stop><stop offset="32%" stop-color="rgb(77,146,255)"></stop><stop offset="66%" stop-color="rgb(132,123,255)"></stop><stop offset="100%" stop-color="rgb(187,101,255)"></stop></linearGradient><radialGradient id="__lottie_element_806" spreadMethod="pad" gradientUnits="userSpaceOnUse" cx="128.86700439453125" cy="144.10499572753906" r="335.8024487125565" fx="128.86700439453125" fy="144.10499572753906"><stop offset="0%" stop-color="rgb(218,140,255)" stop-opacity="1"></stop><stop offset="20%" stop-color="rgb(200,108,244)" stop-opacity="0.699999988079071"></stop><stop offset="41%" stop-color="rgb(182,77,232)" stop-opacity="0.4000000059604645"></stop><stop offset="53%" stop-color="rgb(113,116,234)" stop-opacity="0.3799999952316284"></stop><stop offset="65%" stop-color="rgb(44,155,236)" stop-opacity="0.36000001430511475"></stop><stop offset="75%" stop-color="rgb(77,148,230)" stop-opacity="0.47999998927116394"></stop><stop offset="85%" stop-color="rgb(110,142,224)" stop-opacity="0.6000000238418579"></stop></radialGradient><linearGradient id="__lottie_element_807" spreadMethod="pad" gradientUnits="userSpaceOnUse" x1="0" y1="-88.41400146484375" x2="0" y2="97.3550033569336"><stop offset="0%" stop-color="rgb(255,255,255)" stop-opacity="1"></stop><stop offset="50%" stop-color="rgb(255,255,255)" stop-opacity="0.5"></stop><stop offset="100%" stop-color="rgb(255,255,255)" stop-opacity="0"></stop></linearGradient><clipPath id="__lottie_element_872"><path d="M0,0 L246,0 L246,255 L0,255z"></path></clipPath><clipPath id="__lottie_element_897"><path fill="#ffffff" clip-rule="nonzero" d=" M32.1349983215332,-4.735000133514404 C32.1349983215332,-4.735000133514404 -3.569999933242798,-4.735000133514404 -3.569999933242798,-4.735000133514404 C-3.569999933242798,-4.735000133514404 -3.569999933242798,31.858999252319336 -3.569999933242798,31.858999252319336 C-3.569999933242798,31.858999252319336 32.1349983215332,31.858999252319336 32.1349983215332,31.858999252319336 C32.1349983215332,31.858999252319336 32.1349983215332,-4.735000133514404 32.1349983215332,-4.735000133514404" fill-opacity="1"></path></clipPath></defs><g clip-path="url(#__lottie_element_195)"><g clip-path="url(#__lottie_element_725)" transform="matrix(1.379991054534912,0,0,1.379991054534912,-1062.4317626953125,-261.09478759765625)" opacity="1" style="display: none;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path fill="rgb(59,58,94)" fill-opacity="1" d=" M1449.2750244140625,468.6369934082031 C1449.2750244140625,468.6369934082031 1100.8489990234375,820.8629760742188 1100.8489990234375,820.8629760742188 C1100.8489990234375,820.8629760742188 1100.8489990234375,792.8629760742188 1100.8489990234375,792.8629760742188 C1100.8489990234375,792.8629760742188 1449.2750244140625,440.6369934082031 1449.2750244140625,440.6369934082031 C1449.2750244140625,440.6369934082031 1449.2750244140625,468.6369934082031 1449.2750244140625,468.6369934082031z"></path></g></g><g clip-path="url(#__lottie_element_737)" transform="matrix(1.379991054534912,0,0,1.379991054534912,-1062.4317626953125,-261.09478759765625)" opacity="1" style="display: none;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path fill="rgb(59,58,94)" fill-opacity="1" d=" M1449.2750244140625,468.6369934082031 C1449.2750244140625,468.6369934082031 1100.8489990234375,820.8629760742188 1100.8489990234375,820.8629760742188 C1100.8489990234375,820.8629760742188 1100.8489990234375,802.8629760742188 1100.8489990234375,802.8629760742188 C1100.8489990234375,802.8629760742188 1449.2750244140625,450.6369934082031 1449.2750244140625,450.6369934082031 C1449.2750244140625,450.6369934082031 1449.2750244140625,468.6369934082031 1449.2750244140625,468.6369934082031z"></path></g></g><g clip-path="url(#__lottie_element_743)" transform="matrix(1.379991054534912,0,0,1.379991054534912,-1062.4317626953125,-261.09478759765625)" opacity="1" style="display: none;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path fill="rgb(59,58,94)" fill-opacity="1" d=" M1449.2750244140625,468.6369934082031 C1449.2750244140625,468.6369934082031 1100.8489990234375,820.8629760742188 1100.8489990234375,820.8629760742188 C1100.8489990234375,820.8629760742188 1100.8489990234375,792.8629760742188 1100.8489990234375,792.8629760742188 C1100.8489990234375,792.8629760742188 1449.2750244140625,440.6369934082031 1449.2750244140625,440.6369934082031 C1449.2750244140625,440.6369934082031 1449.2750244140625,468.6369934082031 1449.2750244140625,468.6369934082031z"></path></g></g><g class="png" transform="matrix(0.667044997215271,0,0,0.667044997215271,331.0806579589844,125.07731628417969)" opacity="1" style="display: block;"><image width="597px" height="760px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAlUAAAL4BAMAAACgqh+xAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAeUExURTqW62qP4RYWFkuV52t36KFZ5wQEBLtW68Nj8FNHj2q0bHcAAAAKdFJOUyQ4Ai0kJhUuOyNVzUmgAAAgAElEQVR42uxd26rbShIVCOZ5RMGgeTvx2TTkTUWHQB73OTAf0gyC/Rn6CUP+9qjr1i1ZcmRLsr1jK4n39lX28qrVq6qrO8XhdSw8/iheGLywemH1wuqF1QurF1av44XVC6sXVi+sXli9sHodL6xeWL2w+l2wctllOkL2I7jslsGz6NaQ3UVXnd0fJp4xvmGf82+NlYvvQk/GV5zcFuQu+cEPS/fHmxx/Bn5+vE1ei95skE8or0pXBmdw/DDHL7Hx+fXJm2Fl31Z8Z85Nfat6a8i/eJe+QHqAG37bQclgpxi+5sQXtv78bnB+frobEO5qrPibkW9LXly/NP66QuICXw32gIM8Qj9CyLiiYNGtdD3eFYLdEJQcge+Rz7r2/Ad5Kp2f/gkR3Fqs+HUcv/8DwRXSeezL4beqV53dqe9N4JAnxAc6p++PrysOdoM82x4kT1l/fr50/BLuIChn578KK2d6IkwOEvGO36yTNyIMZz44/lXuTLHGpJEnCktDcM4CO71aQvPgVNOCfcCV53d6TxBxk3cjL75Kr4hQdmYmuEamvrjepUQOKnDOYkgUVHgR5J3ycwPjdjBFCfJi8myN3WCjwZrzJwnjFwyHkPEqXIHV//+unuOAb/8TUoucXYrVG1ZPc0D1779E0NwVevX3MyEVjx/MKx5x3SVYvT8JTggV0gXiDzEc02nDPFbv1fMd8IMHSDZ4i7H686lA6omFAH0kfo845e5/AVZvz6JThFIEq/8b0TrmxAqLsHp/EjqlXyJo/eW3EBioufHwBKsvzzMAIv+N5Iq8gu8iWYelvHpCWe+R4oAMekyXHcZYfXkarVJSQcU/+uN7aC1bXMCr56IUCq8Irh671pFiBcsoz2H15UkQQmZWdKGsWDEGEf8lxAqTSfQIq/dn4RNDBRyPFIDxhm9taAkurUecwertKcRK/kZJJ4hUsfqL1ng1UYUfYnXjEOzuGIPEq8gpCj5iVv/zu2vbEKRm5s5i9f4MWqVIUWoTtYp/EmD/jbxibv2KV8tZ0K0jzTV3dptQEkWniFIVAcZIoe+J5dvIq3Za3gdY/Xn5B6uvfnC3+JNvdn7JbMCyQVCXxcNgHAl7bZ8zWQOsvmwjP/WdxOn8+bViJT40XkAlSAmr+ov/RGIpr9wZrN73+Xz1Y4g/EK0gOSyFSQ4S96ZnlTjS87ya/3T1TchRz+K62fmz+JPClQ6CTKwoWFHcw4RtyLF6u/13fUOuofhOdqLiFng0BEXLAxwJLObVmRh8pIJovfULsqlSqBCrjFNgghXFvWWLdVryy7H6cpMvuLsXwSI8FTt1SwRVqnqk+qMnlv9O4t6m2ddprN7vEBk3Ai6VQTkUMQ/AynviFjHrR+QVq3s4o1e/s2vnvE9MulZhxKx7+rUHrP8XB0JJoA/nYhAfgSA7VT6TvLNmmVtHDj/0Ytx9y2D17n1cbMixuoNI3+ybAXaemGkUg+V5BOQQjFdaBeukLW5HrOpLtKbek1XKJhCKsVJVhJCAJVB5748sWI7CcGAbMqzeln/i7kYobnR+zMpVY1elh6dRsP/JWLUxBEcG65dYXQxC92CqZwrF9VC0clUmWCTsRC4PyitpB9wNq8dzFGCSznJuI2B/qVJFFz2r4jAoMejYNsyOg2+7x9MekNa/DkCbjUi1YnIOmtowVMqv40erRaxRxe82vLpJFjNTgpFcmeNP5rZYoDhfJqGK0Rd/UV6xuo8ywkmsut2ipr45jGhRCGJI2XqiaToyryAOgn0UHo8ShUGbwS/nVb1aebpVMF1+ftB0WSmVssBYEWU119DzniPRcwxK+nzVONjdWa+vOD+k6RqQJCeJFKuUSLraBfpxbCOxQst1GXd4BL261TCIaQ7Q4g8H8ac2lIOQQzDxKqzB6nMU+1JJXYt7MCqua/yRVQCRdsDjh6DlZJpwEVb1BcDUWwt0vV7ikRNBnl3O5FysAqHjOb9RUhGvPix9Djv69osp0F2v9QtKCzybJfOA47wmVvhM1yUeyYsaWM65gWCNsarvFDI7VGHAqnsoKWCF+USElz+osu6JX+IZOAZvyKt7eVKUOgxIBGZBKPUqrcRkwSe/HD8+PlJpdMZffV3LjHrpw7vLAOsuR4pbhuSatixYwQpR9FxdqMoX69Wx/VBiHe7gGeobR6Amf1LT439gdT7vNRFMvAK+QOVVzAgP7eF39ldoAahFBalZWcE4K4KyrrPCK1zHnx9SwYpg5WPhvbCqdwSLeVQJs/KpLVDB0hGQM2jByWKQ1D1wVeZwmV59MrigUphw5BXQJ0ZxUYHxQZ8dRxsIx5PPs7zqfim2W5uB5QXV7oxZBysuVNaEhoMmD2/ldYpCMu6QYfVTB0JpargmBrtLP/d9jBrwihHByERdGxa8lEFR7JSghDmveBTkHttNtb17JGEXBwoyaaN19cpclc6ZEmgagZij1WP107KcwUx9wsrtr1c7w8pyzroOqutELEylGLC2NMsH/TgG2zRP/5vWZFB7hipuaoRM1pNfQHUIqcY3PCKtsnKfe4hxcC/AQNthqpGgR36xYSc7OhaqTNttduKwaBy8kQGoN0QJjVeQLXjLCgsebDKCLajXyszIM7C894LlLpvH6VZITnczyQNrXdf1IiclGLGgouswkqlTveLFJoc7+vZd9F3yZK4UV7aOMjehMq/MMIHK1QRcUa9kKieEgcOa0Kt68wDpLgvD7vLzAw+Aac1INbKgnM6IPMGpSp3qFRFr0Nx3j3Gw3pyHmCeCYHmgh7zBKhrQbPJmUGQY5zhH9e3TXjS8rf3M9R3GBolASC3GtsYmeQatVuGwAKPXEmCNL3tepcrocPXEb+CvIC/HoKxzw5Qt545d9B1mdF3roh/5rFdY46+6vUKxuxIpniO1EEQrHms7NkphPYXdBFI9qcqyIV61Nkd4pV51W2pMt1F8ysgHWXVPe6t8rlSgZQXQP3PyntWvnHMb98nUd49ClMU22cIRzwmh1+UQXsqgefQNuVX2xGoaz3qls6kz/ipsqlfdzWBCa9pD7UdDA0orxmZCrW9hKgYJMq23t2mvhmm9qh+HOfUSrUL17LrGVKtVaFM24jrRW1PoVBYYkeoFq2lSrY/GwUt8e72aON1uhAStfWozDNgaG0yU0tKxt9QPp0nVUCD62M9wtLpoCIvqDFvUQbt9A5NK6hmvbGI5n4FQyYKcUqd4lT6OgmVJvv2ovj0cwsP4q7XhDbpnRzXY8iSvxKRSAmgpFPyJrJcs7syr1NBwmNQr97m8qE5D6DokSP1CmQtNyTLAmRyQw4+ZleWDi+qi9SfBS+YBq8r2z6l0OYSxKitRwWTaTONfWZKwxxhUg0WbD8xo+9e7xVx9uVJq9cVqCbqHR77GVKrrcM57RlFv/JBXrbUiH8IqL1o/RgRKU5Uss8GsWV1IRasCtWtW9XzKWJVEK+ZVr1tWv9IQDHt70f2zZS5XaW+/dMGYWUeZBNTMhlX9JBcU/8nkKikhjLz6sPrVbv0M3dbBeaZejFW+OZNlzF5h4m4969qbl/XoQCkMS4Ir+SupIZ/1V7NlqW73aFwONmZbBySTkHkrkKWmqPZzSrAIHBkEJQiTvzpZTb+5v1q06Ue3goRc90xbwqQdYWhOK+/CVveJqb3q1CqQSFHeHHlVJn91Zo7+8/grgUlaG7OprUqzPgULZYIZJ2tWJf9tuHJVcjpYJn/Vjnb/eKh1XosHQF6Oq+Uq7USjTn9v6fLASk1OR1ASSFCRs4pwFSXp1ZHlan1d9H6+AWzOhg277h7AmaCqlPgrtuyW50x5haYxsAiwcsCrcJ5XG4FQ75Yta2cjgM1EcH3d6zDopRFUpwEnC3uMFCeCHHxRrYr+gnhlncgz4+CKPpnuNkzTltmMV5TRSA0mV3WJQAELTpBqRK1KDUG1Dk1jvOL91SZ7ij7DPI56KVk6YlOnWWXdumZl2sbPCJVlNV5FvezFqj9Ur9qTRXHzWN1Ik7rLxKrK63ua2VRgK5dRJ0mVVzM2tCQ5j7yiKCxjFPZAFRmvTlzD5+EVWv0FMd/pi2p4wK5Bwy/xaioLZH+u7pPtemRYQXCNeBUW69VDFWcwzZqmiUDNAr1YBV2/jLM9C6U5dgKLzJX3hQh8M6gzXNMns/O0TL1U1y3oskRZcmUrFouXSqu9J8oKDBUhRWgVvVhFUvW/xBgUXrVrazL3MlZYZStsqtF2FAyUQgUz7UJDZS/TaCiyRUGY+tuFV1flg/VdtSrtwyTrIoZrJyHnlZdUeWpqS0oKjag6/RZJFc1VvLS51DO8WuivujuEIapW6e6pKWGudGuTrL4uoYdzlFLHwAGYjpK96JhXny0GeRBElP3D0ybGEamhDYXkQU+GQDOeUlMo9SgErCLOeUk+eE6vvu7OpPr64U8mIRBOtgWlorF0n3mfuQSYra97n/Kaxg/xKo66iHdjXnX7ixomt6AbUgw1Ki+GAvfuzRTWtQhTSh2GzJVA1MRhsCiVV8mJnu8Xvf7zdhfd3l0k7CDsAvvPInQyApNZH7LpNAmk0guzikowjRj2Ilr3ougBK5hXH1oXPcuret+AuoZY3N1fpUWmNnPqKXk2sy6FdpwuGZepEFM20ppWekWNeEW+PePVXD/DmrpovdMTrA8tr8XkG8jpAhsY8wpm+mBkdqt3n3ylKGj8Y73qrzGv2rR26TONg6hNjrp8crAYXpyCLaC0Nd9zLpRL7JLZSByyXBUUitT7IXXRsLxPZp8gqy+gGM9s8foa+c+3dJsvS2/Qkho/SG9Ou2B41OtppB5UrXrEKUZgobyy6UF3WM+rm2gVaJYsBQbZisI60YY9C9IzhLP5cqP+ykbBoQ0tJCCVV6fbpOw+j7PGnVF3lXRZWVlBq6CD6Muaqqar642VjLP4U19VsLKPeOXW5IP3SQIRq7QVRZXsZ7LpFHa67cIUqRqdgxd1bxpLb1jXCS5W955XP62x70Z1hvof5q4lx3EchhrQCQxtvC40AsxOgm6URYA6Ri5RQN92OibfI2VbtpM4qdTUNHo2sxAomno/PnsBER0+c066xCE6vVe0VapFAxiqJyV9HGU1jqGp66SuKGdoYn07z2p4a2VJH2eOnGHrcMOTqxk/gXkZWhixUDxoFFcobFOd/JnGO5hcXemOjq9f8TvfNSi4ZFAXXdX704qqiAG5HFuSvSBzqIB7eCgLrDBO7WNN/RscdG7/vjyXOTC8pZt7xtSuH182TuiodWUjQlyWzBaA60Exdg4LegGDlpWc1L+z+jZf6o734K+gxFMmPkY4RzxlU6LL0Cn2JWxQNkEFjoETaOeGBenr8gzkT80PvmZuHw7sUpyuFLKK/gpaJraZITJ+l8jlQJlVqOYFGRTS2KgSenugf/Dcnts/R9eXie31GrRgfqR4EzdGFwoTl2jlibojJUJWxSFV8q+c1nhM44Dl/INNjv6puroe2taJ7ZGz8fhesTEUUXvtgb1ANytvv1LVlL4B5ZQC7iB99Lfm7kWQe/rVuzmJEdPrSZpSvF74aM4GxNisHue9KiRq0BxmBYp5vHYyKoRWvzo292M4eFjIGBTi9AJmzge02FRqvWUqUPVo4G5QVZ3SzPIMlCvY1d/BFZ3Mn189pdnYoKGEMTqDm2tWGo4d1wBj4QClrlRmHHBaMilYUYV5XW3pkF++SmPY8baRrx/dgLRP1jiMt0EsQXvjpEApmigXPAijddVVJ9VNuInT23LVrncddWSAuD5v4IeP/mFjmr1lZYdzuAmtpdKO0lU0YJf07qXZfKWHtXduH7YqY8uUeX1oVMB4NdViuwWB2ZQdcgvLIrwXABkDTyiO2Ar6CEz2Yg5VXf14j8npE3XI8pZRYF3D/HPvvFsc0sE/rLxqRK6gV89De0HvIeZ1d/3Qry4uqOgj8XYqO2zr+U1YDMOIjlNYDwF5VVxUghZpVSqxSsWhVSnhIzhewjA7LPgmzjNl31a/ur6tpHrCeUCMFzZJIUQ8VvxDXqyrQJVxVypyeeHiTd6DzLL4Wu5XL3zjXHeWlLt7UyLeIesErfKiGyI5gD0BE+3ssKRT6YmFblZY3gswxvyef8s/+EDFaaCHboTNpHE4xZcVEXtZE/cpCZbU7JzGh+HPD7+Do1j00/nB7LTrdfCeC/SAQg05vaXEvOuMYH/rTFR0A0rVZ3KbGb5tT9VH6/pi1eh7E9L2E5wdomMGCjSxUXseAssqOqaG4F6Lqhf98dTEcl1tzQxPdvnhvrJSdy4+jHmWvQfcWPZF6H+tSB9D5W3G+SixUxRYDqpnQPbOu/vV9YHCktYEvUcfZ51eUgr1bzkzArOFZQVo/4EoJ0P/BHWXK/jvYk7r6pg7OLziEW3CbIEa+lh9F/vsd0vx8VyTXrFpk1BKh8WlYhn14iRkpLCunvHwDsdettXCYpyHD7Uq2ALkUqD1xOK66iOQ+SqEs1QI2bmh/l9d/b1YXb0qt/Z65HcQaJaWVKmYL04IvHe5lE2ZthAVRYlCYXPUR2F8RV1Xh+QhD7uObnjgAvLiIVBHmrsjcBTq4+Yk0F3LrBeVaYaUSkkp9iessxEWFsU6XUr/uK7vhTWWVaOG2CGIPnywv4uqRwh7aVJfyqhqaUnEQEq00aeCB/XNC/AXMduXR97Oh12uYfP/FsE3555x2f1srtJyUrqwGT00g7OKgQ5j0xITjhomOpuvxlfO+bzX73z9PUWDWJll+HQZFSXOV5UBv4pN8XFI8Eoo8OdGUL2DqaIMJ/jV1yJ+9evvQc2oyBIn10fr7Ln2kmBcz5DIrD4Fk6O+oCmCuCGUyjox9ivmM5wez4F8fZnF3t2/nHu/ssx7lOi90S4Vm5OVDqEhmfJj/GvS7l5GzYfeQ1E2cGaYTaN3nNXwXJva+7LBjkUKH2dp2byCmtpb1qjngC9ggbm5U9PSbWQIGNyFLqQv9eezcdHeHVV0e6trKREVH9mprXQYzQt6WhcvoM8Y6VcygcozECCgENEOF52seF57D17f3Kx6i2jS9W48rEw/ib9+DHlc7VZBwWSZ3PUy6tzuhJDa42d4+2dmpOCccqaSiP3Kr6FEDnRuK/+9/wZ4Mun5cQglsEDBqJBf9XewEox+0B3MtjlXOfrcT7P9zX+qDxyCfTPkPXGgKrR20YFqNZV4CbW923dQciCPzme4HjEtEBGNvQt0dCtObWGSi8As7UE9ESPWWrqdV1cmM4K2qVtdyXfw2yJ+m7Po789XSICOzOHz4mPzkTACs+pWeclQwmQdmJRGtBjiBu3oCd9A/Q5evH/wVRjywr7q3SWXo6FXWlb8DLoMdtN9xHVcIVRhmCZpH2UNaiux41LpTBjr6q+bRU8fly9auZm5jNlLQ7EKPRd7M2eOC8txMdKd8Di+XT/zLSWbE+we3v651dWPN+R8Xm8HtICe5bzfse5YBZzEwjoEj61Td6wWHGRcifqRPT2kZC1L8xnc2qXTaW+/Gt50TpE6GXwDEQKdbWU8JKL+zbw2rQcdGoyAGGdTi2KgDQAGE9F+6GGdnu1Xw3NfvWEDs4KYKEfdsWEBmFHHA42q4DtnWXxlij69coKol1CbS1KCxv32M+Fx2t/B83/3ltJw0FSRMTEoszzPqcAmWOpp15CF5OJi1P6G95/H9UjYy2EFaCH9LHqD208f1q8ALED6z3TjqlNFr4xBMN8ys8yEjwIaMMjrjxIsaVFsVVJX43cQvPP5fGnuDf/vqeJ4/qhE0e6OqMasTFjEdVztZpWMZA60zYeulmFRCwk8Zvy1utJMp0/yD0aPx0RkF9akDeUKNiDkVdgYz5rqRVPcY1lGhdCZylYtOYbJXCYBWCtndX0f4JA1Kk1qK5K0cYu+dX+8lx/nUsokYy7wDajPZb2EhZN6FYkCrag6mMQ34R28Dazv2Fn0nrVl3EbZcwzN1U7KolbmiBYV14QwYuiyeb3Iwzh0BdJ28eHAZZlSMglWCpZTNBW4//p7MNs+5qhgqJ2T7SLhhtPsp4XpFBpM3EEySy5fCZ5eZp+yulIrfWe884zIObJfDY/1qh5cRA/mRqFQewTSLOGB0KZfHrO6xof65m7iWmALyfqV+ry0X51vw+jX6Xisb3j8AJHvj67ufRJm1aWove1tTp4H5Muv2GMGxyUy5CTlBO2o2kxcTpFkkr9uT9xQHc6wu7qyLhuuLeDFRwtwH3PJJbZis5WNF1lHYdCVF9bq0yYhl6ij7zmZF+Di9lp+hl4UjVxd884G4D5/OWebQZ0sNC+L0UDYBD6Ni+MAcQ9hxKlltZYn82NC5A/DkBktFxdC+PQC4pRWBAulwNMcbEgYVVWG6klHN68Xv4m1x+Sykdf35930jfQp8b5xZRmQGHMKIt1Kstnbvi45qBF6IRBD2V59DTt8Ac3ilfBrdXW33/n6UqQmEzuWo9JDKz6JQX0SudowFRfLynKISDNruBU8E+oFJ7wQnIO39k3ctc9r+1E4PH9S3B4fe+pCC1ZSOoRdTys3lcZUFycMDWhXZkZFvE5wn76ZgcL2xOnG4g0e5/qmLqU5fMxs0uQ0Jgkw1KogGLNF3HAKpWkeTxx7KGtvVwimw4tw6ptAyO/pFbm1w+PXr+/ZqChfdzg7SNNMPj4uwjDBIqETbaijE8eHxnTGmfpqSnWN3cb2n29ifW/Ya7kXiuGmKcWPb1XVI90Ry5idMnThO8gWpXdPjqobgeKSqrZOGoKW1DT3Lukek/MswP0OPcPwkg+h5qD0E7F/HBPBCpIw44q+sSJvQrIXjaURUQSTYJ+XD6BcwDTrVy6v7/wBuCh5m9z3xjB7ZIF2mwyNVW5uG2Z2feIfKoRxRrjEdI8UWo64Kk9GNnqdfntuz+Rtam+EH0QLeGVlbVpoTMA1dHG0gfH15kRV7DOhQWlbT3NfKvMZvk4v5wevO7+DvW27yXSSGL/MIDBw8XEpvTcghq+k6goCg7E7qJgxoomWupV8B39Mq7Z4B1/GOw8thF1HKxfEgCBaUcNgcUQrV8Dp+/VJY+kCykM4JiLgC4imHrrF5u73AjR7+59jh4Lrnr4eKcwGz9xnMzhzBuUK+cZiZtsemFBjxXiI4JA9EF2pa/24fROXr9Ys+t+basqaeyTQbrZmAjCxoMiczn8BYFAvoNVVUeYP0DGS0xDIYPEVqeV3vmxlWfx5Z2fXWV3piGiDqEsad/uY8wq8F0qa9KqEBO2gN0/dzZp6rAhDwx3OutI45AP2pQ6HfApt3VuFw4AFLCiu5YCdUIrfdGrZvQouGF4cmDLHaKJGWbm6Ol/e8R3cw0dkjdeBiQTr0Yth7EpFrFlIMB2Q5EqI0Hazp0uvsFJKK/3K9AzntSzW4Q1TKOYE6o2d3B/R4opURbcbfXJcgQ9BiobkDnYFmmL//WPg3CQ/ppnPcGr7cZbfg9ety3d9BJiI0bBQn5o25eQN4WviMKRKE+WgpXOZmKwrG0BXj2rS27+234PXF1/A3qXMEWRXZI/5xobCZHcX3aYpd2JAYer9EZ17/qXk5vSwclbgcVY0kAf1q+vOtp65RFB9usUoCa0my2NvOQIJ7KGsDKoKHSjnFKbRaWvtymcbNvelPntW13v6VUbasVsM61sWnzS5YY5Pnjo1dtkCF0IAmK7cgxZY6DZ+fiAYPZ2e25d6fbr/R0IMGUdWryXB60+N4G0yAlhx4qyOiLkOjxrRHIdQIXqpCxv9ij6vOixsbb4aXvMNzDwxDU+LJqBlGntzz3AFGVsgZlr66RgKHba+fpPezrr6+tWMFEFDoV+35UD1LBrBQ8SFpGMmZ8MUX9wMmpwfUGWzoSMFuH1elVattavxuPfgsFlYcEb0E7MNKXncxrw8hiZbJF9kKckoXAh2UkiZ089e4nGlzbpCxu+YkPJ1n65vOO4G5t55BHv5+OGFk/2KYfdOjs1tG3z+cZc1I2kLhNgQO87YiO26Ok0Ckd/um1A2UINW+yojhhifCa6WHs4C67md34XTgVs5paHQ1KB1O38wt083c7y3XykVoY7KCPsp4WIHG7f5LS8GdVSEiP1TqKR74JXd5dvTr/5y58v5dDr9FjeBDS697Tutpius28hGyS/tZg627lt/ReXfedUCQ6HtfNLOurqAmzhyFn2ok3HjtwmIbPGpq6vc2EpiwJWREcFkxsAUNOpYg3x3X0HPeV1Gs+VKnszdy8+G3UWFZw3XowPfK3AxYwxdC0hDNowrq6Ap0CRsAqg/U1mlvWdlhzUyE+eX+gc3ZtGIzAq4KYvP8XVZcnFZEINIGFP5S0UFx0J0mmSvCMz+qvLZ0afLq96D97xvwAyOR2QAA9e2xNJSGisRb9EU/xN3RTlu6zBQgHQBQz/7XTwE2D8JuoCA55PkI0CPkUsU6G1fbHFIyrG8luPkpdvutv0TKIocDmesFKGYLRNjyNBbaHqiSmtHA7+6fvysPBtJFEUP+Fj7xCVD0gLta1R/UmQnXdpSr5skSuNWilBAVj0hRXH1t9qbuHye3w6yceTdXA3wJWFZIZL8M3KsLmBgTZjE9pQhqIUIgaxs33mpWnSaO1/+r36wtM2lF9RTwSJhTwnLe76Ca4q0WJdMlXtEUBRQuEgEc+QjvfOtyZO5ruKii8fu6yhqGqElUGAYpRWa8Idcu7aWBxlIyA9MX+BeGQ6VWA7sTFdqb4Li6tq1u3QK1hf1cFAN5Qfhr6vHMG4p8yY0f0TQlqjCQpIalYbOlKU9FPbzr06sG1iDD2QrbpjVJkkleqns0isdufL2SdFe5u4sIECz+HDs/tV80Ukx8/P5Kg4i2wvtcSWlLVIeWGuOTSCUFk6V2U2SPVODsY1hxYX+s4KYBVCG66dzux9YyorNv2VF19cy2bEpj1YELhOLOc7UPcnsoC1Y3QPaw3FVS2Z+6qzQ1QyiaS9z0whBAR1Xa0bWNH8Qt7cUZC0XR2WIkx3s8TvIzI/WruX3m2urAciVLq7YutPHDaDf+EwAACAASURBVJFe1tKhJTe8gGbKV6J0grXlDsT4x7r983cwDgO2lAbWF6dOMA3s5Bm1aP3KchIWIywJqer+xmJaaq0meJgjdfvvm+IznKxt+LXnoPjqQdU/+YFYfAtZfx8bfP8kaxCpHkNwP8j7W0dOKqs9ryI58Pk5jh+qWVclVCiWGyKHklpCveI7XFftgYxPX6jXV3qcGr5q1e0nwnwR8nsEgGKRhKsqZtH+8GG6Ai+PJDWPh/is0XTsrthy5bfM6ClffVKfAdJDEbAVW1qrcUTSC80+roj0BuVOmSrpAN6ugTNJLxSqr6Cp9p239fru7yquyNCaGO0saxwZYvDazGztGQz1XkSoF735Ch5CYso5TX9W+84/Yn1fzcv21X+aUTH4tFjhIIObgeVU47qWnEXDrKBQ7SXPXroloI5VVo7+FL2+pdr9zvrq/loZCqNYUQ1li2ZC2X1iq8rUNH/DDhfAK2qXU6WzcCiwSq6q7iDt8O7HkL9evn3s3hLRPj8tNDPnPzUtmiH3FTi5K4c8w5Lixi6MF/efVMZPf/4o3dopX30yt0fmOsZqJsgrlbDTjXG1YA+sExPExaZeYa7lKI7WDHk6sCyaTsRn+FB95WshsEqINsFqo4jV+5YELR8VmAtsVIllCALZiZZtjx1T+XIVT6btAfr9Hu4VyP5KM7vm+3OlQKjx2jDCMi17PjqjhvGBgHUrt8923b/pF13Bau58uTT5DD17qfedJ+qpXKC4IiPU6AVZBzFty69F1BwLicEk1g5gbw2qqcCbPXAN83RieX4H/95Ee2c9X30Pb7qEOKTir6E1iFLyQvpfl54Nyili0pxNRaU3aRw02KUwTFdVlekNLCl+jqvfe+ur02i1ivEY4d1SuRAPokO7MZGAoxT5n5aF70IbUkby4SCuMKV0h9uYZUZ/vbX3B1/M7fcmhQ+yodGLnAc6QFZA8etyVqCFBnamhGCviA7BL/ZQX5NLaLlyXo9vWeSvSlx9DheN2LyR2moQRQG6gbIfH59TO4+4LFYok9qfVPoK1h67glJcuXrf+fZBXNQDiwGyUO5hkvUbJmf7Rg1KJA+9cDoZehoxtRau47GmpiR1N6f1x2k5jSF/DmeIPLSRDJ8Y5mMbEr+pwU48WiLxCXs2UF4nnd6DVSilKCqt8nNcfe4ORmzeALUiTY/E20nCDH1SiiENxxDUuo0Sc5TFpN0M40ZfM/1Q0lau4uq6fAh3n9XXoZLd81Te14QPbay7oekYVFxx8akFQg0tutkDJVWmStTxgc3vIMXV9Uks7Clf3U88Ma/HXVGb3ES2tQZFtMHjC7zGlSqQnSv2Y6VniSoqFaY0NUWUm6tR3qMvdeg53hxf28lKmw0rAm1tnpQ2hE9EmyLJNy14UunO9tYLWYLJ8fk5p/QZbm2ezMn5Ck8gaOzSNuvZlmfZbN+geSgtD8MEqwCesTk0B8Slm2/f47QkrrJTuh/TQ7h7f/D+4mnFSmJ8iFwpiLKjpmX751Vmq2GFJPve1vCmTX9ZVUIJgcX5qryFcz/I2b3lJfTduF33w2GF2h3ZCnaLTHOU/Vz/3AjyZJ5CyyouTECtYI8Dobn8zshXpcbKvBN3WVgJtePqdUQUHO2a7kiMUIzm01Kyt1YrtEEUKiqR+qApHn11aJZ7iP5GIQ0Kb78W8Z336n54VkDxDMeIsgcbvfnUkoIOymAjYEfJVHuBXI0eeAQJVcgUSdO36TMnsCzespfr9foZfQbC92qXN7BiPLRP4oYAn8jJ2eoRtGAs6HAKuwv1qsHRKF+e/1t0IEsp+vZ9nOjVnlIENUbpgCVC+FaN3ghXSMAUBGYPihRqlI5Hf3nF8URB9fjMGUvy1e3Ffeee2U2EdD2jMazpQWzjmBqcmJDUlLlsCobaHgmnFFSPs/ecshOIXVdX82lNeDv5LrX1+i5n7i75MuNSu98D+8MmVOm+kawCi58E2R5hKSsjNXt/q6xLUAopw4/gnMiy+HlNY5y36jOoigELXbh9qVqV9/VE4qliIIdFVnZkZrYR8kIvz8qV186p3hmJfo6rKbAq/8HXecj3XRU7Y6Kx+iQ4R8SYNkx0pbKyIlGoednWdNKHnCpES7aaTqdgVwYZyzned57L9jMw5K8t2MrDxjOKHi2w4+RFp6nBdmQPIBJin6nsasxlK85C6KmsnBpFEA6TAV3N9UNextXlvd4cHjJE1Uoza4Z6n5T4SWxgMShDZ9g4yRKzNTI5PTo4dYCs6KAyavY8x5XSZ/j1EV4fx1UaFMAgunJaLNSvHBUPJBIkVY1oD1ljOrtAGpjO+ZvQUDmz+d9QNWieTJPf/n1WZeXBDlUmSmUeHz3rFWKAs1QfwvDGgsEu2AIv2qiUbjubm8zQ8RRIQBfooFwu9RV24i5Nj6p/Tn0EgTHI2vcAzULqmOMGHBpq5YVq4ybAIakfrUK3jLevQFZZITI5134Tv/a/g/dDsGj0UIOOsh6RCtBASd3HmJ7LKxrcEOEYprDsk0spHRoeu+G9zI0xvYF090rC0i3hmB14fddbW5N8J5/hvq+7waBZjeQTy8uJUppf01a1qRLrtdXYFBK0nXk9q/GWMYwYO4NCdL6Dj7/l2od3U6/vjCUl4niwQXOlvwAP3dU+ELaTxElTbmVW/IU7IXaQG3MGtue4pzH0EJZkNbpR5av36yF7BbhTUh8qlJ2mp7KnG9fkF8BhMDKLQFAZazq5/pnrULp5fPcoV5WiYTTjlK9+a872ddMv9f4K2sdc/1otRpT4lLTVisK4KIonWvquXN2s0fYjO48rq4YGfDQqP7muKm+hexyWrq9utY3JyXEVxf9bW7gwugC5tMb0JvB5MWwVnsXq7U/qhK3YwqCr1FF0RI4xmXH+yvAFKIYTJ/nE3duPYIyDV/uCSjIN+AJpgS3wBSipBujp1FOuulMO+0EYQhgoo1NhWmKJA2ucvhBXhCG3tKO33sF7V1ghwQOHGfQaCfCFuNx/CwyyK0cuq7h7AZJDwe4Gjp3C1UseL28gTogewXFOVmPBZJQe8mlxtTo/LYclG7qym0S8Y/KRXy1FbRAyHxUMyq7MHrl4nLBKQ1MeQ66p5gs4Pv73kdTH6dzquDo6d/7an97hLF8N5mmJBLXVE+Wf3TsDLqNajmAPhP5ZBDd+YHg4JCsUC4+DGh8nhbT1Rwjub5s7024Ea/cqoXHPPgienRZ9w0M+BRAebVBbXIZFxcsUtQuwogmgcbiAnNOlEkWCL/jVDz68J/jjeHkCOaIG5u+RNRALjPsVjyn5sto1F8DxQZJVdkazFxBS82FN2Wp0+FRxtVy2PDFfefZwwWEN7Knrlx4Iq1Muy/KX0wklFOyAYba9D1qzGpA6gK3rsaBb+bDkwO2t9RVN5Lmw8sp2sVAd+S8rVOOkjFuUbTVEesUoKfT1gMKEMU6iCvDClNvnukry1d/fqh98w9yZnSkH7Aoq5LjaPI1pnR3KNrBocPS6jVUyoV1NYMWEITgmSzyVIBvny/dI76PGRa8bmuT/nFG2M9lR/BZl+5QUBjbt0WUniUWaoFTIQjF7Mzvj6zkLi6i0hHT/6HxKrprewrHCGW6/evRF733PIDTs9VMYsXVaSTvG5c4b8TsS3G2ku2H6bDi2b0ojiExDQSTx+dCW2UrNB5cSdHvi6r7zpODQ5atBMxOzqWjf1D/B8CYFmt0E8Qm0PbQ0J3yqgug5lOwyhDBGVVVUMDz+gefOpR/8MV/djyZ2iauFDkriKZc2tK5l64WVxlC7GggaU+/I7+AYCyW7oHrT6WX0zmYq0p0+qPkWjsxneFO+YluSKq4q72GanhLY55fYcWBZK2sBW/HkBom9s2JndlVWqB4Xn+MipPI4p66sNDNn+Gpzlnrf6KDvP5PTqG4nMnsUa9i2BBhZMNMauE26uaF5vO0TSHMYx2eCGSiuMG2emsBFopr7nHF0wutrcyCXcdW5Ls9bgtB3HKq9yhJN690NaTpCziqJE6wC2juIjplBGAMMJhvDbeBoClrs6rgqL6HTXLXLO3S244CszgL2jIeSv40Xr5unZS5LU1M9lAhqLwK1VeeYixI46nXB9eYYG/X1o+phfAQXz+gvJEF3Ok+Gsjp0GLSl/BA9ekDv46qNfIHZa55HUDT2zt6GJ1wCG8uopswhxtHUuYomE6W+EnJtY5b6/VpmJ2GPOCjVL881Q8txOIjFMA9Qk4qqIBKFXQRjcIQICy3NTcnqriT40T3l9ildjcrf+fbrLXv0QK3UvjwdWeK1NxGreDa6UbT/ZCo7ymNCadkwWEz3EE3zPATMVRWKcmE6qsx38Hordfu5+840j9BKmFoOmgY3seG1CLMyQx0hpLLFuHM/IOrUyHR+8gg4NjwvNa6uP8v7h2ZwjivtWXx5eY9+bc5VZI4BNAC5YkcEVS7E50GzhXBvwGXkeUToZA9lNeUq6hSZRoAMWs0ziOr5o8z++P3vwm/i8iZcFCiMxvhEuT6pprAecrGkDh+XonmY0EVcKMdDI3jJUVyBGl2FUpZydAEd8hXjDL/Wte63z+pre3rjQeWrKP8enEcflXeZf5Z3hC+CUQSGohIajKllaPcv3HBcOdAc3YRUPeoCARbmbMUP4XRaudKOvpytWxtVXImoAI3jtfhJXE1V4T/Wrl1HchwGGrB+oKFk4sVhgMssuCcXYDnYuH0/MMEC9xn7Ewvc315bJIuUbM/I3T278wwFiiKLxapxLNhDSvKADG3bObkCE3VcrAMB7ZgSigcwcbUuiZ2SO/RkNoYTT99BzxMJfwFwrMgVJqiGdbyZn44wPUW26sBlH84RaCNIaPKLDCKUjmawdcroFFdTPre1vlJ90Tyc+Hw1X9SrNVcop/K+sDoNFYVPtMWzBPs4FDYI/akFLrPAHHlt0gAMkqtqBCblHCVxxf3gv9ggfClPxsvcRgYSl2CExmWnORws6VJzI9PTsZORPJ1Y1oAeunPNDW25UVElAIMMmJPD0ye4MbU3ublZ38HkJqnbP3894sP79n3C4ruHOVe+f/C78cf2sCK/QILs5Tzi7F6EGW1FKbFQgWa2QiwQ0Pz05Vo9Unxd0zUl4/ny2rjyKlgBH5cg4v5cMqDDqSE+CPGhVx7U6bQX0ZO+jZrmUF1FVfGQ9CVVqBZVVGO5REeVUpLvOh+Upbj3tnz11ngLGV0QQdpCVaCgw/iKlMayHjKPZyzU6lT0rcnKGbGhYhbIAxwL7ukFzD8xdEUXEXNnFnT6Aut7O8MiCgXMh71mWacUi9hgtHXqoTzP5MdeDTagRNudnQjy/gOHlbKrHNecBtnjxO7kiFJune8fElefG8PUZ+sr8qYEhciKVcDP7MhFUMgw95PqqCZV3xYoq7ZXVobub4cQPOgqOkBtASljpXRl9OoaSwj5x8t6HK/D5lLjUV8/u0sSxj2QnabMHTD2+48qvHCaPiTcUAFiJLU7javiqHLGooiiIFM9mZdrkovMnDXoAkdbaP9biI+loEGKGUfBYiDELqjV0HZOUNFxElcrWiyn4/Sg6NAoqzvJ7Hwbk3kH31+ps+05nnxZiNLzZ6YSB5ppPQqG7Ivej1ZSoJNitD9RL/DFw0U043geBVI1xa0f/ZuS/pBrBsuT+XGw5/X3M5iMh97/GNQhlmeC4atdEh5zsVuZUQsFENO3bZtGITxGIRaD3WHamkjVuZMTQkxd6Q5elX/F88FX3EF/UY9Y+H+PkIyxHrE7i6dUr4sYnwLIA8byZyj/kUF12Z4EBgrsBXfPyXCQL+D9T3xM+eOnvoOfr9wNt4KYpVpT5eXJb6Gv5hFYIxmN1ZTs2rRP450ifODOSlZfywWXimrdUeNnUlQOKvlW1O0N88G3pqJU5R1pm+RSq1sd9oEwu+lJZ7UrrbngHdjIiYGAFVNiyh2bVPbLjCzkfJV4iJprrCkf1/THAO5Zt/YFOAN3NRdMmrMBcTBsq6Cw1YbvKLbowl3oFLnCCzicQPY0V/EbGIURKvxibZlx4ySuYuI2h95BA/W9js8QeDDIjfMlCHdPinaGrcL+TJBYxyox3g2Go93MIIoyq+lixOakY5QvFlSYXHrm2mqiJ/H+h3sRek32409tHP6Kuh0hpQhDIQgWjo2HIYACG11rWn1icGOEKGRkanDQtJ5Ykay4qIp1MNFxTWV9RXH1/qq48v5iXSRAiWGpULP+7WtbLtnSNao64Fmd2juNyrZyEMDk4U0yTWCOJE5ZuVSn8Jr4pOS4jA7kYVydy1diEEuWG1xdjUa/PtiSPWwE04yyADT+jSNl25QLbyD6P+xsiVRMcVS5PeaUZYKqDK46rl7ilxqYyhd4Vzf4y+ghGoOVyp3FN9XMlowusnJGMbQptCxwLONT1FXVCyjleg6f/HOMyPCas0w/+KuwBfg2X/3+gh3qrVHs+sMoCqIjtOyPBKF73eeCEl8/iKLOibUIaJsICNNJvW7oHQSAUlxxiHGJ4DbhZfmi71/iDL/P9sxm0sVSMaqCMu6BMaLFjkqUaXwy5jqxoxsL1ZNSpmm9lUmqBWUOEfLCHxNKBv2Yyn3n95foFHm2cLkY/+8s4iteNyxH63c7wt56J/WqXd8NmMo3rgZG5exZHHR9BCsuGg9LqWRnFl9VMKS06F7qce98Jl95NDaXyk4Q9bo5Hl88goO4eGatwnJRvu/Ojm0giyZ09igUq87ioHwwjgZbZQdI2Woq8tUvIX+8P6/74VmSLxhzLgOGwtQ6HDFEZfOt2FAqsvrwfUgxz9+xLkxnyGimrmI4nUCY3NFMiYkethdM0zJLvlKs70XcD/YThJuL4cVYGm3YSsWwexmGqIbzf2pFnhdNdTkJ2ILkKIuEMm+B89M1pvL9S+mWZsKv/vv1Qs62V/U0UNnHUQV2dGl3l5lGGsednBQv5nZkqduuExN1giOkmK6C1gWQESoMoaEcUVe9eMhWyfAZ3kvK9mNnJZvfQmIIqjAH1uOusseA3TdxmRpUWKBXf5u++Q3kWkFcI0S+o6tnNmAP0X/n0s4xcb5Cbj/k1p7DRSGeVu7pBk3rXpEGv6Gm9Uw8LsQXTvSClUwv15/ymVyxPUmRJQB7nabKl3DRun0jbvjYHeS87pUc4y3I51W7d2dLl+NKyipuAHsGkYfhTBfIU1Mu2HcuoGF4yCQiVpXCklMVfsrrOFoznPMFeDtYfZP3UKSOddolnvI7GENv7ZnV+xSGXE2XMBYEBkMz3qyaJueKMtSWCjZZzev/5brYfpCE1Vp4fW/f3kCqP4VPO6rnm8ZVzU8brDnCqAVoJ6P5E5UV82FM14wVkuL9U5Y/z5mn9ZSm7XnluJp/qj7Dr+Oa4e8zILsK04ZQWr+ph5I/VHbsCyFaqkJ7zLn6hn7ZqQA7S1pFpRrbvREevbsrz+JtQ7N5BtOtyFcv4Mkw51E3b4ynPKY3x/unbH7aU+GupSiXn43NoEOjrKfjtvNlunz4kjY9Dd/A2zJ/GAwZBoT7mpmtPrwmW9XiobCQ8KPKxlQbuvnujeWarpHVOdXXsEx9hO6QZQ5RC8ikWU5TYO8VHx/Twke2Vu4GQn6yFvVK+qBlLuKl2RVwyex7xt+W+w++Y69KaX0j0UrWIDpn5GGI5miTlQBW1xxfV51KVA8hHdZtLduXrNcHT70nNX5p6QYCasDYwSSizL7jO8yvnkSUJK2OweP2vbeoRNoOKh608GaY65Sv5PKZ0CpOa6akvtD3JakU64t84gSNsWslYuWyp+phQ4uN5Efs2gwIq6EJN4YwL/KW22x5Rzki+uYiYQmubm1uVFrdlsRHpbgoewk9ofsRPHSbdKFSPTc4rsK4k6xG09eMBXKlgoXfRVYE0x+78TsMK1uLJsZjqCG8HrU2OcCW+xd9ByWuPh/bx0GlwIZvxqnZHtd2eMPz0wFEIux+5/JKpvJ9+7Ar8va3jCJ4Gmi7wCT84rg3irjJaeXbtz6Dy/rHOq6emHl5ENQgtXrxUBYQKVof9te5mDkk6b2X3M7V1QkxHSeyz7iEsQJCuWY3YGhyVUhxnTDLBZzTPMMf5+l8JdiChw0xozHsaGaHp9tN3V6mN6PS2EE6bqirHOiOnbJoOwB8SceBSZ7B/BBOBvqcyi/rUV0pU61HtdYM7Lv0ZL4KSmjn5pkEBnT9lL1Pw7g3Fhx4BVWcdEX4pG9XFBB9XsdTeKivU1nFq0gyYY71rMY+fvQll6H0Eq4ZXvPVZwsH8vc3Ay+m81HRriNUDLnGsCOHKSCM1KPsqWQWlM7s6UIFDOtuzpBCdXc5j7quzDG+HhTsuJH3k5pXnAH7zoce6+9/tdZWnsfO65NI5ovaB3p/IAQ2QLIp378evSAP5BssBJ0ukcAiQne4kitWJ6PiCvWYOd+9D5u37pG1TAuVWdjHYYWUB33igrIYghf+v2zfiPdNGI8y+wCdirGQaxJlj2ZwQZckLIW9nNrU/CGToOb8uQbUbaFXcC3Y5/XMlvnnn8pU71GcAbUVe7/ReY0wKBlHLOv6mpc2SI3VF4KFw3CCbwVvDTXg4i+6QKllO4eTu7rpqGXmNvB+SIuAWBJXnwI0PNHj8FpXrqsuo/WIDWqmtEGtIIDSl57DtKPUtwxvhGilRKIYZUfezrgUXbdZfdpWWDc+KI60tWCwOENtN3H6DgYvjWB+/wxq5UPleOM3g3kI0RbCvZqm+kZNuWig41gDMhhFcMlQHxGDCugBc7Ka+dd7fBndjyfyFU8DvZecVXq5sCGC3zknrhlU/cTsVBqCWmO+0kbQKl8W6mgO4y1Lor2iqbktSSqq+Xqj/L4QiGW1LI7ewUb8ypfeCMVGiRyWrxdQodoLyodVOW4u2Y2gR4w64iq46zyNiMRCk6jaS1f5tNbXb17WM5oRV/8V/s6Px1UQhpp6y8tcHsvyfjxUrOhUjs8cVdP6t8DGUl1pgnedKIQKx9+MTZm/XuT0D/pc66lZm8J8VHMCfvWFb/h372AwZbu8gBeoFwqdPex3NxxRYIpWwHHfiljZRS4xtym2vTmjxxqt4nLho0jt+RbOOZryUc35HSyk7p/wUGDyEIELF5AdIbCzKwfWg8mARhDGXOIR205fiCoERm2NwfeoU46J/m2pjnNxAZeVwUA99HUNtGX9oFr0X94f/PHg/mAw/ghBd5V4GCHmXLV8aC+B1TNFW08L27l93xxZ2AuMpVCoZRrng+LueOIRThlTawu4vofzzNXVP7M8ivoOSlw9pCcDppWvNLS1YPC7O+AGgxl0TV4Vodv2bWisJVvy2KOsp8yR90txD+t8fpNJxGzh43xm/8yz1bJ4dCcueBVQ0415SBEpPzvs8PcEX+hVCYxuYHu5rgtK1viH5KxiMr1N7pQjszvc/iBwoex0P60PqhXWy5d/Saa+ev9u3/l7jQ+trbzZu1HZtL3N5l5J2vBQGno1EGwo2kWzPqoAbSwExUFtjHJAV7eZlDJItaC5WVM8F6OU5lWv74v9wb++R9ozwMcchnwNVT2Uh4KbabMaKI3KIBKdmHaqh/DRoL7AGIOCe0pdYAEPrkOLFxAIzG2RacRMpUKustL9EkKK9fPxO2j7QTWqFB5f0DWJrSy7XD5IMfRcsHctgtAQxxbWf7QX0Ao0OTMtdUfwOmX2NbaWmb6vmWu9f3P+vYqrHz/O6qphlzIEtbeWUpTBUCNV4Ssi0QDiv6699fr+tUEynNhlcKNcUHQ3CfQFJjlONlMtOonIR7Q+hPm8clTlCzjPy4Lc3tQP/v6S9+EvtbM1g6Fj2NzAAQXDIFYSeAIBLjRMmllLR1a48lFBq95SHTMn7f7jFHepHYuUn1ceRNCfqLmR+uqP8Q1/DBfdWwMH39GLV/OBNwmflKzoinR23zaOkIVTPIRdCVaBkZZi0rgqGAv1RHChWWDOVLOGVu4H1dfywXwVoI0ZLgXtUUhXe17NA5fsPRhq0gp2XK83DiREfJbYs3Z6+j9nV5DrNrIDDUgXMLT56+DjAdmpIb+9AKvxD+DuC2QRIBcaYG77rWYVyZbalpw3mSQzS4JNkcVi1eLWzDPFHO/EGTb1qkQpSWsVWa+SZBVnnWi+S7/lO/ghv13R0BpfYHWfAh9hJc0+Mk5TjVppE9qdFmXvKf51MZPvql/g6Qjxl9Ks93e7dXs+se+Vwf4t8CfWzEmAYwQrSr3a3nm9yKv/vEsrEaXVjh3+JHAIMs3x7SjYTfwCmjs66THn1P1n48bMdAKaob2HdalR0xaVCm1NgkBAS36hYZBIyfycSs/wx1RS2nvnNzxkw0Nr4jGootyl7hEGDsy4LMHl22TkhVNvcFbJ0It5RBRXKUfhm21ts2PtpVKw1gf3kEChRyiYFcEYpFU2ne1tYn3QtxMP3fiZeU52W7ywG/18g7yqthLvCruHrcwoQjJICxa1VKX13GLrtyWRDhqlDS3fvseK8fH9IVhZsL5/VNvww1sAOt5coXe8sX5zNwBNtmNHEpGd6X7g9jZfVNeK6/iLGJtaUi2aVjYD3hVct/WD/paAgjKv5CtYXmIkZRsaKZ/Og2ovHwbOglbdg8mMhwY1hr7yqwiDfgQdgnVOKpT+ppX85aJ3lDykpFTFrrO6l2fIdU0ZmKP0U/oFXGGZ54iTNveDn+vJBBhv6EWJiqahXu0nZnd5OnUQIXJd6Ol9hDoMXyrL6tk0h5Y2DWZLXy+dp+SZIArlf6FjR3l/RkvvvL5wu/QRLjpQctw6K0d7ZJ8QGr7Wpgg2ipcZpXXGc73V7KdmFKsLZRcW37ODcLUONrd7M1pRWtJUNZ5SrNaR+VmqYs7R7SYA9n20myA11MliBrVGEKJ2CE3omPaUk6nxjabYdDgJzuCisRMlOcafMi898woVfa6+gd+gwUTG6yGtqOy48ADX15dywsdQW4btKvXkd3DQ+5ugdFpgMV4OumoYOtzfEOZTxxtd23wgcDzDSH4dA52UsZKyCfJtTkii4VYR+wAqcAAAIABJREFU/8giAl0CUqt0C3GtVjkl8wXYCm0f1quBE3OotNP8AnUy3+awpx53RGM43AAPPVJg6M0MCKZcXk/Aa6Tx+zfXKjG2Yl4Ag5aqjneI1ydV6plVWep8zpVe36e+AMHfdYWqX9Dzm0aUcPzNHddFrdI/k9UBjWE7BS52n2tPcR0E/e4m8VchwkhxjwtSabFmIWozSpxBdCDl1vKjeXDAGfhw3es8urF5CA129gQ7XWU9Amc/w882kYqeXUN1b0NNcZXI3ml4LGjY0XouFizpPLURzaVlL9Ey36XXmMyLvAp21mUSov5KYmrCMd2kZulcdGnLflZcDqQF88W7UGPcH9xQw6pnu36vVvLRdw2lZyj0PQzM1mJxxImub//6mC8atFwFUfW4qpI9P4GNYI1mt2HyVmPlItEdtVWKWkmkZnMYVp1Cd21TZumbTTepdJ/3JPfxEcWdL46QqMQoo7mSebDiM3yYV0HPdcXSxZ91GTFmqPenenQjtyXKieGAc1q8V81zd5enaBjmesbZEv2BKtxTIndI1lxx81PeYI61TlEbv3qFiwYP8RmJQdnakyr/b8XZ7QCVtEeYeZ6o7QqBkm88m5qHIzvK5pRPrzXalHbhseD1gd8hcVmQSvwSrm9QcAbFrz6qV4E34IayT/xDGgY4m4XGRtC7I0inYBJzJ3X4KNtEmaYiwg7FvXLVBarHrd8AoakqWCkRKn7+lUCDAAvlX5T4NXTrG1Sdolc4w8/XAw6Vm9CN0v8NO9Rhp84OiG9iRk2ewnD+7k0pabMeyRvGvpCX5ikxN63r34tAVKVSJeRVRNBKXBisbN/CZ6jyuh/UPc4rTaf/vmYeg79wrTzNgkMYNlRaDRXO3jyFYQTE150K1wzdBRN2NAugWTUE+rn++vHG9NlXpQRgRgblZe3QDTJeQ7YwozLyyu1xXsw4L+pVhTAYGgNlAagXNiw36HljSvbSrI/nnh+dbi7ODs9BC06j4iW8EHlHydlvxRfQgy5W0EteSW9Vcmut7H/+6jsYjJwGKRSP8XmDvKHWz2aotKxTkBYT83imCeXS1Hr26kAQgyB1dfa1XXpPbd7XgD2koQK6IBGUXjTLM7S9M/qrk5hMUIkdxWOuwdwqVWVnL8PQOdbxxfFDzwoL9LAaBhSDSNXS/l4ftN9uA2ULGDH9pWdClTA5sCoLcMzWqkQLefWXfTs/g7q/YSM6URQz7BkMnckLaLg8EDoeD4EA1x3RyhE9vF2Z6oPq7Vb5BN6eoSloaNKNs65q0I/mNTYStfU/ZM555lUlOtD2tfzZPr8Zrpue3cyIX0qhcBYcnTsQSlV3To+dMpjE9kT6yw/NiztHcn9NiuyBiI3Ve3SAlSHHvg0tYN+K9f0Rwb6vj/LKCQsY1j6Z5LigMLUYim5OYSpoAJ8+wxPVva8nQey6lr7WaeqL6UEDXndhS0a1KjwYzn8lrSLLFH5JXjlNp19fX6d4MoIsaM0avO0GLk+bedWpowvtYalIO36mRgu9hY0AGBGZRTkxdqFbzcwJcXILCM2kvKJ6PqkkWNJfVXcT5/c4A6091SpvUoghcN9cfwQZJ/Tt2i9A5Lg7pB1T2Z+qadzd9DXTCjpNXlkcNxGuwSp9KE6ZGbACfwoGKkVdBhyEyn0HX2uFtbm1A+F2NKLT4HgxU9h/AwsCQ10Bf9I82v7mXXE3TSsgVzovLxev56GYHoabu7s5lWcn/XrZlkr3+U0muyIxiNQaJQbLetHPvoMI1GCyaWbWzLP55rYZR+DQeqQKCgHRcyIM9JJnYl1mX9b7hWPOArXCSnOhdAsP5hXmQA8tKASD/EKonl9D9YnDd/CUbq0KrXLIUYr21ar5EFpEWidD5KULT9EXlPBoxiQFwFqW7YILxerWkh1au4XCWohbHkxiq84HKE+wbLzYi5ZofdC3B7XqAnI11Gr/OBfcKvKNmlfsRc35dCQserwSFMCqpFFfi4WqpuN92dxQRpzEs2GQ5yjwVQTXStJr0TiV+Eio1jE6A79y3hwnfcMH1Tsucv9mQaXp1FCsgBjfCHcS53967vG54xtuT0ljV3hBTuR5oeQDlkzCA/NL+ZMI6JpCWaEq6ROQVbE0o/kvcVFlqA3ai+r5DemPoeWZPkIWTF1vzETijG7hTN0FOxLsKaXTq2WgWpZtR+dHNCKt0EJtGagvMvuk0ie4hnHnE3dij7OxtqYmiuDrLVmBeiUIPNT8brozOmCzz65ZWTHK5Judm8YrgTRZBK6xeWhsLK+kA3WYsQYrlpzL9JtYvRpP6qqFqxnrKpPPmMc0Aw876KozV7PJtQzQQelOZBXxKvWGdY0Cj7jmGXZlt6oBvcmOOboNqsuqQoaRaGFaloYhZTzNNbXMd+n316vbpV29GryrC4lEgZ4bIbyaBLux69wsqPq9x8eCvROtxw3AbGpNlXFuSxt0/fTdE5lDKSm8nnRns6QsLHb9+kmRXwNVssrtcdRL6NgXYFDgijK+kx1K4PCtsW/uYL9oSnzWXB2rYTpzvHn20021PKV2/X1fp+TPiFtKbUNtF5izDjWl0qPaJ0muNa9Up+j3S1y0qldYyvP4RhpRyS0cN0/Dbtc8qrjHSH1aPzKf0+Ob7U7XWeg6Dp9qG6sBwr26jldO9ip4wnvTXLWgxIwZsJJR/NmuUr8OZ5xBj7qC2r+pWSWoaUNo2ymRGSo2zZX8yeH61EgMTl68d7RQ9qHNjj26qRBsmLRfBHJ9U5r2zAdY4uff4DsPhZ9vlMc3bl3TMDQjpdyFMhVeJhU7Hk8iMeBZERL14yDsKBdK+u+O40lXkJEGd27J4mW9ekwclxPaLWZWcrWdHgpHuh9q5EJnz6kSTuOWuVJ7FLyKoRJzayXGcHVzYnujRtZ2d+p162c7Tep9vB4utx7RiLNarbJSFhRez/9Dm5Wlu5I3+K/l1Y9D3Vr1fxs2EqKTs1QaWqOgSqGMK6tPCtUIbavxeNMMZ67+4gAGxzeGTMxc39zcKrWFb8Ls0X0Fk3t50QqVTDc5sxWV/uof+gJsWoZ3fTtltNEzkOehlkpNdXbdSEwV2+NiBiWHj1DBUIZp2S5PF4yDVqweCr9QSi7aRl7ps1kfoDSlJVBRBhxprnK0vPolW/pDPsNwHUy8yf1cHTl0qMcbubuZqktBAqLdOWMEgMbVQXM5CayuI1o0j1gVLqgV4n6SUDo4Vok5pfNyzNV3sPbUO7wFCOavW53MV5pgLTYtDyRI6NMLHHahh2vBeTam9uyVQ9ULQTWzt+FKkesI5hUv3nCgZJ0nkgmfwcxWNGU3O7/xx7F6NSjp6hoqy1jV8A0tjR27wFmTaTLRJjsWPECOe2wFVeYf2xtTqjAlzB0rBpv4JEckSq1ijx45CBJbYLDcZzAyr/6YLcDxjQn5HrTqCkBCDT5umBCPFC/ECYCumsfxeCu40+LjBfhGjJ10x9lNzfGWDFkACBo51RBhx940s2FgmYqWVvYd9H37O5xhUFelq1nAoW0IvNXdiux01GcfmwpX4wmEj/7ovRP4v+hYY82VO6S816onUVSZSqV6YAjM3Adm8LKxNWVepWa9os72j0O/iUH9dZVyNfArOAS3kBh2eAzWg15eR32nTq2bwYzBVXNFd1zUPqmS633wE8jKjl2gCMRIg/6Mh8w5HApz5APM/B0he86D/A7+frXz8vpXCJaj8mlxN0/dMLX0jnnYZTqrx75TfaW0ShvdufbkIs+xb7qPLApaqaqHP0sC319rerS2yqq6zoN2mHqcV0HlHnWDM5nOVUvlqhtNval+gt0xxK7H8kaKmSnQvmhnJWX91lu7ft9OOPoFjG5pKgGKynJEiQJ0hfpevcF/zfvsBd7OWAXaCm7V01Q/LTi11aHac0Fslfe6FqUTRxIU1aHTxrz1RcCJRINw9aCoapFztNsk/JbzQvI6UWMklk6BqYUzQOL3LadoMK8ER/5XL4kwNYXsmVcO4RMmrfVXbzEryKzqVt4vBHvtqGbTaFqq6wicUH4TAuUw6DZbvl1gq9DoRem79OtNXv30uphQWrXDeXcogd38sLU0440EvWK77qTKcW+r+d7sI8o916IqFWuwbr2iVnd/RMnlVinrD3cWUfKoyir2okBBc8qbn1rTqd2LKn4VWK+uw7W+KVEf1LYRcdd5MSISrrpTFWu2Zl0FHtdOobblUgbtfT/cJFGhjdazI1zWUskfmkmKxMRmz1B2Xj/e7XHcmVK4en5ooKPS3najc/IefmYez/tT8v1dHMR+cdgxJD166FR49x/cbemUk2qCv3YO0Tjs7BhaiVXj7QffwYHUf55K+ENwSqhtbptlh9rVvZVQHlWHYTz2xrv0/2fs6lIkx5lgQfkCjS8wLEvDvJVx1buhLb4DtIzf+6FvYOYmA3vbr8vKiAzZkl2zA9O9j0JOZUbGjwd+Dx52SquYXcQw+R73BClYBypgTKpY42QHpRcp7O6UvYPf4t9+ire3zv/P0z1LfCuzO07u0K5TcuDqNEKiMUqM7bnskukKZ7Ug+tg9grCFISi6Cr/lWo2cZ4SOFmxBsa9V+b1K72Ddr8/idSlWApXPbTFJj9lS2tWDwUWV19cTdSGUp/2XlfQRg+Dw4R7jtjO9g8QeVkM5A4xH3wfqYzixooep9mfe5E0c+1+1LZKbGdble4nCcHOjM63Wqx5GjycUUTTrDXA+UtNIDfXE73EbCjsG05pG7k5dGE90L51S6hYC16fTVLxYm7yJo3qlXgxKPWakUiHYk96h4s1up/Satc7QMBuPcTeZAcrWrOmeGcrZdjlx97iOmKLPNM+f/heCfHdxCuWvcJM38U+Nf0XTmDfxepTi3jGqq90dFrWCCNV1/7RXbK6oeRPsuBmlV7D8lo8NzQpbmzA+/D7pfBP8F5tqYrlQbe7V1/m96oDzUX1Dfih0XV3JNOZ2u2XToBK0b6dWFU2WYDbkUnlj0hYibuKH0R0ppuSmecVjUKrixKOKXG6VDmz+qVfSYB3zrzqrVW8eGavayjIz5gY8xq/VDb7QL5HTBritZht5T5IfrWLdpVg92/X7U3e6enkkUO8pjQAiOplJzGTtFalDcapdrEV5MpbVeIIztG9tngZOrVKvOSUbQm1WrNxt/EW5PA6Khd31lAML+47xAduvh10ureoB8HqYJiFYTcVmvbibeH8/fgeNcSwsbaMQAYnB1RKWNpao4kprV+tyYhxjUJV9ggN9BcZmx/k3TeUusxM0f9eQJDBPRafCsUq7ruK1mte/WX91lO/ctbJBlWRPhm+0taxKeQX7jTX7ayMOuDHmFpOlR4yN+cmJSjDaptnUSebqwfnGaFZgrXNtc9Rabfqr9xTwfMBDBn3BblY6MpizsyNtcx6fybp6c4e+in3omRHDAIHgIBKJCx5CU75Z4uLW2tGSkj7VRSCZwxigoGxsctirF2vbX31VPZ2e94rBZp1qSnqjMxy4s9Nhrpdgz9s5e6ERk9V0XEqMQYB8g2a92RqromRx15xUlKMgMc4zdn5H2F+v5fl3+fkK/8Ko6Osg++y3d6FsQ1O8Wc8giVJht8D01LtfmAF+S33oS9EIDH0zMGYw54Vm4DOYw1aWgmAeVrQ0Do9UzAEtrDYxU5TG87i3ynHReqbe1y9zs+9oIfqGgbntwKetpJSkxFg4gnEreHtpDkSoPDlpYyNC3WH0vx+FlfwnmR73Ddkq/TL5RxiPitWMekW8/YyzjcPq1GtO9adtt4tNT+a9fZ7mgmHwYIUzUBzRiPRNmwUwtAtZi+vS9JGewcTiCxPsmeRaTTABk6pevVnLPG9whsywL/8Guw5O9piceywGu66WUYJ7lTQA4LEbN/RFPjvS8YZtNiVcVu9+qT6zEeeOqj7Cgm9yqlVkmU93Ktau1lzBGf455iG3QjxO3UJH4VthL3iFsjIZoche4pXaPlCtK86hI/Xy+JtjC9HlXJFSEtHmxtSrR5lsztuF2dr2tV7Bn+HgGyTh461T7IrOfPt983qXPCMBxGP58E5zpxAKhCyXHefRKAyNd1efFJ26mbic1AqHCoPWn8Licc1+q+a1Xv3971ts1ao4g5Ur+whb+EKrkqQtCSXSXuIC5n9G+TjzWcWey2KBhsZ9dTgOFmZm++6wZMZLCF0NCDFBIONYGG7m7W/0gfz+OvPrw1JC7Wk1Wandcoh6sLNvLpnHbbpdz0/L8nQBF1+aXHdjpoWNSCP0IYxqKM7XL1JxM/lAyK18bWxe1ilnk1FV44vC46PFsqtVpK884Fyx5ZKxOZkRvUZm5wc4IEhQGI+DiyQIMdD86zMm1Nj2p/DojRORBfMSKLH3MmhhWpvQxWo7+Qzf79UZ5xdy5ume9tZ7HLht5DNcxnorj5cXaszZLNiQzN4gl6TJDa7WYvUxWq3SGef51X1Ey88Noz+BaZ814gOM6YMEey/UkJhZj+6vu2xXfYp+3sGuJam2c++05PDR9X3ZjAGirl6Sp8AfOulGh43JsVWpLG9xoDTCCXx3gxcmchfEdiiKOjCx/YUIE6aDYuX4FRCsf74O+Awau/Emzpgo7W23NVu9AebTCLhzI6JGoKsBQRIN/qWrx9a0EE6h6K4sC+8xRnkCI+yZQrYvDce9wrL+tMyaEnfAZ+g8B4c4nyFWLYGrrpBwre7spim53U5TdZHekuXpkvyP5JY7Iknyl9BCy0IYN9IRvVdpeI5Tvb9atm3D5J5O3/UZ55eFezqXtre0rg7KrnanVLrCyP6SRTCe3Cvy2C80mqO9uPP+DbpqtrERgV5p6Nj9Uk0QxytgFV/ArVi7hCfzdaBL7YCMol9gHjhIDKVIF/E8vvrI/Mr6BkrdwfNuVCTxLOfNmOcIaniuCnQN34OlKvRbIfgiMJQnm3n9+tIPW/7VVyXH5OtfMGlNj9q3PjizZdi7N5l/IWTgqV24niVbDyqZB+24YXXn1DzuZIJJIHg389lImtXzYxspIGG1wioilFsq/2X9cZ6f8+B/3yfz4HqvjCOqFKKWh5RrK69w0ca/gNqZT3I+C1qGC0LMGD1MF8xmlFQuY7ADYyfdykU3k2HrxkQLpjg9mpZnL1vpvP4qZTs7rLxedW/StBvGTuZHBWq3CYehurZyfm2JQ1MBNQ71SHnktghwfMe2GTsJNTOeRPkm+5o4vVarcvzq6zA3nONNpsIBtNDuilVCjblJNWvoczJfAz47XFAs/XS8jIMBouk2ZcxjGqUFUQiKubEfkujelD60vVQLdjeLFatlyvGrA91EyxCczq3sO4X42m4TJpEQ5J8qBeLj1Tjap+R/6L+5ab4w5pT9aFECnsLLsGGedj4xUCXxOoVY+wQX3XXh9AS/qmstfxuRYZNZ2ZUg9twUDL0V3ENPSFeDZw5Dz4XEYYrgWKVk1RzQt6+PXvIPfXjfTk3zBOperI01frfmfC78+XfDk3mv9FcIIlYrBqYrtd0+o8R8fK99L8kbWKK+ajguNwsu/7a3Gejem6d3ol4Jjz24Nj7R0OIkiFUsDszasVvPsKR6BVy0Vq/e/01qCc3dSBBfWdpMk6seoNVVD+votFJGbAqVF93bqCy+wRyIdp3oYwQEk47LrhVtQuFpRaZHPICMZz6Bc5F/Vc8Sai20MvOtUBpRm3MYnCDTS+jGi56Y+aQj8lP7BIeNaeHaU5n188OMC7VSjYDaabXAuaY42yzb65Wu1a5eFf1Ff7cJuupF4Awz7V3L7qbjdP1nGPGJE8PAxxBSEq5t5F4VxpuMnh1DyKiOyVcu+AOI2SbuljWFXmG2o1vUp+j7q9a3r/VKPMGccNWV+6qbNAsXicp7dWpu4Azm/h40QREvaCtYJPFZX/XweI1UyY1qnPp1fIAhHPehy2xH5ScpeambcI7tvVpnm7ecyMew+b07O92hM0b7K1mVurrh9qahv9UAu4o7H8AUWwarwui8tCiCG4P3qu3nzMvl92v9aUaPlfshVzIU0r0y66ZWc+AcFt3ADPz86NB+LhQcFLdKhhWqO2WIblO3hF4pfJOy/jPzL6676pBVBcaa13ulPtu1/iotUFs6XfW4Wv4dtn5Q8ANLoyDNjq8kH58Q2ofLReIpG7HXweXa7gWf311a4JA/ZAUdAB/NhgDDFFani5zPMqNTcDTZd/Qbam1+r7q3TXR63xUSgnJPIncdv77AjBEjIrCIMo/HpFJKR+Rt1f2TtOyEhAYvVdhBwILIfqhvbHAoc+Fl/Pl/f3LuR30eNF67qHbbntGVG472lVw+utNe3Hb8VNRlS/kmzz2FFV8zqBv0M+vULJrcMUbI2aP6xBzNyrMONFaqZuIxa8swbfJSa7kAndrZd0r+70qtKDTzPVIFwSC6ndjspJWgMf9FojRk8HEzFmwLU23PonMntlURhLR4TItZto8fgL5lUR7yAYb8W2wrusyLYeeHklget1tmInphP3o4Mls6l6F7LlWiCQoN2XcLZwPZY3AXsGT0FaS0s6KX0b1JQNAZHLXZ79VMHvLBzivdK+rmPSho5592g77ZkeMspfK8ZRgumhPkSYIjct/GmgsKfWK8X9d+wbwE6oqbZcOK0bNbwGfwPK8azvCLG69uE0BVNHy8mYNv2ktQr3RuDW1c2gRdNVnejbsvNCvId8/oVpHUGE84xRwYqckNhxQr33HJizjbDVuW9V5JTlw1Nzy3PSbzqt0LnC1Q0NJv7IgEC72eWMyZi7Y6gQlF+97UjXU+IWTWMBLP2TDcKkwnUpIZDejMVjS9jcssuUvVHX3aO8vOuWMXus8J6nshEbmR72n8jUl1B3g8NoovAIhZYQbXM48f4WHy0wd8ekenrbuhOG11ihdr8QYdLTsrfHoO12Z+ltylp79oDRdVsRIM/9u20GFdaRsD35hMTHI7UcxbYwVLvkvO9nAHe+nUHylQPhjx0ePwQkqcAi3GaB7hhXZ99vI18zOc9R18stXquGirVAa3uCrE7PaZVMnDEU6hdvUOTcVqbLIFDpGYexZOGQODmYU/ZH07fWJO+DAoTAJgzbPdqzldLtfj5FucvLa3Njor46ovqLoszCzXdZkS9exWefiw7LooADffpnHn9e/+xupYmAneQLeCZvB4XZOm55kbigRoPYv7c+9Mo+0qX1SoaYzMM8vVvbryCltM+mhfz93A+AA2IlQS0yaYXN3HXUQshDcjoT0A6kEadilTYS+1Aby35Ie3Pn/Pk/v5I/eKkcVF31qwrp7/ARXdevnegLOnatUL+x9rieth/iJDggrsUPFewJADPObTkiPosJrkEe7vn9CFWNFEbNhW/Ah/Dso6+ecn+Pwj/VU9m+M3R8GO3VXnuFW3JbWn6n5xf/bbK42oJ3QNeWuFkMrGRkL1Vg13JC6uCsHwgdUWP0DBYGKF6DHvGI8LjmutYOv9+qP91XdVl/qL7jH0/S9lMN5o47uWKfFnf8FulQJdkohGOapxzCQl2mIF5lPGkfL4MMm2eYpH6MJCqHjWPQ4gvwTP/HyBy58s17LKv9q507IdzXoGWzUnvYRFMN4ut+sLTFq3Z8ey2QgyabIZBv36UtV6gKEWSLYyqvYUoixQqbQJBWnSXOi0ZhdLzOulsj9ZruV71bdWaHxktJdcx+0lxPTce2E/AvkahqZf1EN7dDeixorWR9azf0ZGmbnwzdem/2fsalIrx4Gwwb7A8C4QmibQOz+c7A2xmQNEQvssAn0A4dwgN2iY205bVV9VSZbsFx4MPUshl6q++n7UWtyvVUQmZIS0WAIygdoqHNcm+0FaptbzcfReCaG94qDWCyJqML6HJbuARO3U3CmTls5pKCLfEGK9iKuAMe2VLde5NDfkULF8k/sLGfymF2sr8lJb9UqhK4ZFq2ZzknANYWX/SGWfjR0Ksz50eTOoE9+yGFnlaumOpl9XN/ZV6KGrOPbWmyqlW8k/IjVVNN/gz9yr57YPpKYLcsz8vb7qApNhxxkMR3S81EnM9rCYGmOCXOaaZxof16s0oo7z8KRbyCCYw8AcKldLF4L0AG77feKjikWOSZsnY30riCN6M+jVCD676MC5C1Ug5mTIGcyJiSOmOLJrQCy5VRirY/bhW9ZF7Y394r1Zc60ekq4agSiYjjTvrtJgYz7AkOqVakza+8Gb3iriiLZ4DLnpcTdKYtcDiUqDOKjNeUDQoG7/B6UgVyqTxeXdIqTQ80VEji0IZEzNOn2IqWFPR2XwK947V3f0T7xzNhSZAx7D4Teybx4lsvISEh3KOJdMIoEoT5pv3go++wsB7QqIyppL3kErlW8RQ4PRkgiAvFf2EFCvts3kxD2f3SsK1pWRGWFwt3IzLzklvSibx9Mg1IFjwAe40hakR6Eev9jCzqFAe1Uv/djhL66ZZS2YPR7PLOL9S9DCxrVqv1apZxCb7bwVzXjIN6RKSLxgkalkPCtMqMv0kLJZtl2G92HBUPn85tpConAN1TQg/BCGUOvW8xolH+XxCUznBT/kD96mPjdnHA0FnyrLrlGsYyxF9DHDR46JVXZMZ13TxOTqEKirgYtC/jfJEau7lJzGjL0XdQuRKNqmZd8re9iEz0B4e+MdxPN3Y9eY2+0gBOdaldnMyVbwtGeX/EVD97CsYylZxj474QqvAvGpXYxHJqWXvaBvrAOV6hiDN8w0OiQfpV1IJ7XfKxkHiQT5o8W/4gHwnm0FcwvDLF4QOHv/AOdxzsIX5+y4eL45foEMXZliJWYCqlHyYhi61kp6zHns++UiWaX9+vY2i+7Vf5zDm5qrOtb38aQGhnd2Hp8O9pj85Um1mjRj/srpUdWCQ5cJ5jUhz1jYv68v4pvNBO1MdcMp1hxvs/qan0As6GnCHeJ7la5WegX/tqPbRgVrxxl+kxagMP6w9+puZ+eKw0dPGxzq1yeo3zo2u7qUCs5w1+GLtXSGGMOBSjb1zYFI69jbQ5mOkoegnl8lMzRkg3PQEZphGeqq0su336ZHiU+gAAAgAElEQVS9cfh7XJF7hs8LLcDPm4jm5T7V0HYac6iiT+p7/KDnv46Cw2zwBa5XwzGqOS1u3nnTtaAR9SJRcsJgXy/aUd3IBzq7mKabSI0o37At5zN8tPiizGXXmMGyso8SnS65Eg8EVoo2qeusM7Q2Dfw79uy8aM4kguleCUFbucbuRMwcpXCJRGI/I7/p85dq+/4h2j1Oq7/a9zi3u1wrNaa95d3VKN6YkjIPOu14rQI3dHasuhZVSKBnT0tT1sw7OI27RYLRVXTjwApdmxB7TjeO1Cqka7V5FPa9Dd0Pi3sGyXzJClbZtwv1imCGey2mhGq6RWPGRy3BGGNILCLsbpCQV0IMNAK+clazWyTJWq1PJDewwcoWwlXkpiqBVjQdpkIVqV7FdFzpsLYi+6zuQfeTGdqMGt8rzegI11XEKvXg0vYXtGNMNeL73w0qPgXVKucR0TGxscfCllbkJCehQM7VDys2y1YAx0P70FSm+CO0ePtz0+P3+SndqH9gdHVMoOrFXQD7QRsGfun2yHmVw6A3Spj/wzCUe/lXCR5+XZzF+HSD6ow+1wJXIded6sVCucqa9bCliXC/U3yvPjEPZqNzea8msUUhCvK9MguOgsd0eq8e8cVE+9nNJiNIoNAlCyd5W8UvhrJcxFzAYyOPO+VaOq5oXj5RUxJ1QYfAyCMzAX77Wf3BPMiBXh/1/moCJlpNoBKIoadwCeqq2GKghbYP6sQg/tCiJyGHqx1if5lLM0y111msFS3VLLHW8a6tIjlAfiwSzKblVKfClt7ALatXpSV5dq8m4ceY/I2j8794tPegPF5B7TwNWn52uZfXsPQX1sqzlItkuhhyFjHC9NowHOXMwTTsIRugsTbVa0XXCaU9ewcLs/vsXsFwTpoGg8lw084Ng80DfyiJWOwdGRld7Ac4LEMulHcwwnxdlJptll1g0/r1jO8fLS8NaxtmLgBYoIPy1GGlv9y39rn+Dj4JkwhGA41IiV7eQAYZ+utn0ILIeyHvkGot98ruJVJj9U7IlbEaB4HIsYrEtlauoaIM1mAuRhR2A1j5VKb2o6KeIb9XdZ7MLw1QnyadnfNFl8H4RjaIHs8+QRCJ2K+JwxFQsOAqIIQ+JVytshhUgjZZ8nGUmXfOGEEXVyqUWwnliEbMgZEa0HRO+2Mo98rWq5YfMuLT2e6jfrHEZqdHvvUljWFW9xixL+wWXc0vxa6ZUirhcIzARYoxA91jNVfrnGnFHI/IY03+AqaLtR/UljqHNDv/Bk/m4yQnDj6+7J9WaAapXR8psxk8BqIyXPmhzHJCVtsM9ZvZn75Zra5b1C9Nn0IjUAIlxjWV33KxoilYPh0SWquELkRcKu6vfj9wr7i03+9H1lVv7hWdERf1/oJ2nLkS6UsI9HgmX525zFFidihHbthtsyRMudalikfSleDFfFTyBqZeHcUqcn/1R4N4G/wrbE7vdVUJ7B4zHWr/yAcovDQdBsWV1qLsL5bDwNYeyYx2ybXMMCCqerFHK4sAwB5CQLXaFFzfr1PkI9rwn8Rn+HPuh/zxxMwrENWK/HTROPcjXGkVW7jMI4ZzzJx7iIojbb6USK6hi8SYLX45chjciQ1tKP3SvNlDCFwc6KQSJKMfodyrD8Kv6ng7moV7LY0YO3kTFsTYcX9h0c54KHP/lZ42w15uN7l6y3c3BMKkeMp82SVqZttcuTbZKnh0CZ43gtqw0xkFLuuhqFcfn2e6CR4E6SGs8q4Ya590gzM+cqPIzRdxSh3RGKCBO9KOIdJ9VfoCzOXA39NStdZb9qDnhJVEulNSqzYgyKas01nBpyjtnRv7wV8yDurOOWdq42JJoaIB57y0y4JLMrsWEc4vyzxbnM8+g3gI03hD2m+D8HlXMz+JGXRlCUWRoeMttej0BGIEzM8ryx9s7QefpsOi2QaCy5c4TUbffAnIzFmsi+J7QhNNFnO54erKHalzCEYQTwG19XDeuDA03R1V8rZfKh83XKjNR+oW0tHp35fwGU79ZNSO4VYOOb0xPu4mOabHuHzaXBE3rZOWvSYBX/XrY4cd2DyaUK4Wwh5y+VZUkj/RaDe/cXuVxuUQDp/gpl6snyd7Zyb+36rO/7y+2V3aaSc/sTt0G2gfIL7RrVc6rUXSXNjC4u/3x42oZulaYw/p3PEF/lv1AIsZHTsWfkS8DfRMTKP+Mx6OyuCi7Uy9HRf9R4LmCytfgkKJeCy+mFfUf0ZDu/xaWVVJqZR/g/3JAvGNTYn1irTTaR0KVqxYfGATTwuImAoWbyICDYMxPyuj87KqiRK/Yl3XrbT/HzUDR8hEfffgVmLIqO2arYuqPpSuTQ4mDM7mmBnljbHzWNtte7C0/4QxYLIhhG8L0n9m9crovNKM05gHtbjfD2A7iXBseux1moSdaPLNPCJK3o68/3cCYYTxofb1sGAgsxjXEspjDGTgSuC9TfalEb9avcr5ok1cdGL68dFop88cV3uNFnw4WimjHWtCEEJvXmy7gFxrEd5AAAfX3ppnaKxxaA2VKFV1vISxcqX4XhW61OYeZ7pPU7269yQFV/i4F3fa/oT7z6NgNwxdYV+46GLQPoLvnGrtlsXl2ZRqG+p81dgqFB1DYA0XlIEbd5+BihS/hdvxXkkr2tw7/5rA+shXE6MEght7AeAxl9t5EES7IbP3WGgzmDWgOzn0ZRUnopVZteLjq5V9dQdnK0MIDYVenoq7mZm5ZaCbFQ/1SprR58+WxuRJgnbL5FiS4PRgMXSjiQTvrwkfFRKDbJoJxcr3N28OLFHsT71anzAxrQVaHTxDU8e+T8sEs/P31/4GC55Mw6foJ1Lny818z7SPXsy0O2iVxvFCMT+IyxX1pLNm6y5z3YYIaYJqx5dYRQjVFXuBI9kqHOSVfF5+I4oVnxajC7Xz+v5SXPQzh2TKb1ANPu5VLp9dd135CwwQ7GYuV2obQ22oTUznnmr/CN8N5ZFxvRVUx4oXZshFbuJWSJv4tLsxqNUW+YxC/V59qn6whTOw0c7xHRzpK4R5E3vsdACQr4iPMMjU1byGI+T0NB4DyWOAoxdXpAVBr9t2+M+WEwESVJ/pIhgIjcUYiGtF9eqaf3UXh/ZbEcQ4SlpJL1baV/FvZjU/z11OD4VGghISXky0vOPdoEnUJb03RKi+8f0dsD1QPQwEw80Vt+qV728/LM3HeW7m0f94Is3uwbki2wxO0oVeeT4ifUoidqUPnXXPZWeclcbBtGSWRF0yp+V0IEolcdUEs5AtTkEbimDQ8phDN6tR1aW/+k+97lt5qZZGZOcbmB6rOR/Aq/HcYWDAlguZ6Z3JNCvWzeYdfDemhcbYykntqoChMXPj0//BW2bq0SOjoFvc6uf1bfH2Zw5rrNcrprcfGGr8AWrLDjvfB+IqOyG1U3/VIfkN7r1v2lxlOwl+B723IgnvrVz3LJFE+4VAcpsEYCW0ql7Zv/aT+vpK9UrVlmd7nCrMN05ZrEvfyWa+NeTMqkE1NLXZ3KuZqbTFN4hqtazGZ875PB2oGG9iaYRpGKL7l5dUp4TpxbBhc1q5UmV/9fmjpR/EjHM/Rs0DPNYIqq7vHwTaB7GmTRNgNyPIRT7AF+uG4kA3BhjjBZJx4m28tm2zrTSQK7tnKMZzRxUb3cL2/f29Zf3VZyPX8vnXgcSAciVm7ZKzC/zqnEk0D2z3SCyZdFAL75yPCaic4QLnNJdJlDA4u1bOTchQhmjadsi4IphW9AKG4lalb5DeQctvr3PVmNdXwnwIohqnyXyDD2q72EBtNoLKQXyI5sNKQq20BQ6VlaBYrDp3pau0bSgfl9mb1sv6F45Ms89O8lKfjCvfzabAQa7U9WYaPO8YBjgRmb2gejdxukvGaHccPuyIQSRpeV6u1KplvbrmiuoAhoWXbOTlxKpl/Ru//StUvL0kNBS8vkMqOGvgEHDG8HF34eU7Zx5qqO2L3K5UrmS6eYH8Rp/C5GGf1StXLG3WlgUfN+ogXG2sj9iY7FitVF+oV9ke5/NE71z1Xf2fsatJrZxXogZ7B98GQtNc6JkfN5kbrsW3gMh4fgeBnj9j78B3BW+9L1bVqSrJkpwQOvRUyKWqU+env4sMjj7ChpXzF1j7IEHEDa9QDUW0G6wEzimfL9ylT95KvGtQswTdOJfvrCztWIAYoXgQyM40xyXt2HfqGDY6LdEP8oiTq1fPt3u6IIS9AN0rWqHKiHMZxNjgC0QywmjZ/xrlkppcMcTADSgTr8B+LEVTGtOmGRpdLlYkIZl5FzFnwRiuVnSvZI1T0U3k0oLEIFpzSvor5mNnslCN46qEmo14C83Y7FhY4sUSUx9CNWSvMI4XC8wss+Eu0NpUiTHnY9pkGAz1Cveq4of8W9D2dIdqvLRb5Fxf+guosAs8vsj2PydtDmnpH9Zp1YuHoWR0xZ/gnCLsWthndqngpuG03IouFfULW5gHbb7zs4Qhn6yuWvVisLGVfdu2zaVSaYCVdgN/2tEkNz+s/o0jwKF7k02XU8sY6hlccbrRCWcmjbxlsXO/UEBh6BXcd75XsLLgGSfvz/CfDOeDlxJtbGLYX2bs8gfYGeP/QTzmdB/xLl72Xih9XhM9DdnDWe2pL4W3zExp52fQqrhYRpIOgTTcUL3atghvr3jv/MnwPvq7eHzc71rcq7bHnXhpSwoO5PPGwrCgFfS2t/KKxvj/TlCD59IiBNWDyYcY8EHsthRHm/AEbqFhOE7L5HlV9DgnTyLixMBHtGGLWjGobS9zlTjWpTPGmIiTGONll+fQ5uDO7sAjUuqV3qUayG60XFNg76lJxQzm0JLrrLhgHcdm71XMaKj1V62JYkw8BqpL1E6TldS2whjHkB3DaGHj44yEzK6W406M2Y3hcR6KMXCxnBNrBJnAt865hv24UOFS7Ttdr4TfXsZFs6CMjY7VlN0f+fI15pB4hdoNGqf0TtPNu0lAteUKRHbvp5q5cZrLNZuOYUKZmtf8Sdkavx8fofIZbsX+6vY7s5FQw8cmCiTur3naA4LNGhPbPDLS9+B+4WFMGDzluGj+InP/+TcjvlmmkzAiVKrjP6utVkQOXfLFim4UXS+6V/wJPsk6+pavV0m2S493EOrKxnZXFRvRAV47kqnUwLJiGLuSPbsDiW+UcZkd+RCk5HIcBjkxsWEQ3wXeR8wy1mSgULpeG5X3fYv2zrf6fjD2x+ScBFgM9D8JmjdAQ6f8NAneCKv55MQo/zsgo58gPY7TyHnW7l/n/VTwFZgj0TfulfnB3mbObG24Xd9C77BT7YpKezF3KWmtWhXDHbxH+wlWwPZOHJxofI7N7AehHadGDAHlU2d25jyy/8LFTt58j7OY9qqSaylgoeGIwr/hXoWv8PteIYe3lrH+56wEZ8bVXdwY+p/I5hlsjxObrev4aJwxP/3Di8vcCCN7eQKxyaG9oEu9PUS/HJnKhU+Qp0CZludsrxBuE9+rbaO+XaThZb++t1SrJAYW4bA0V+naJJql82xn3+iia8zY0vowBh5YDGOhPAnKvmtSMNSXpLpWTzIR1YMpxxOv5tfczLzjXh0nFf5u8eyc7xmef+5pXAk2zoE8RAmfPZJj80elIdd0WNC/jaZlCA3W4+TFwLJKtFZeDoxtCy8UJSI9XawRrXaemY+Q28+NH8OjYTh2XvApwr16lnjI/5yahp7Uu9Q0XPbrJoxR+KHcioKY9m4u1qdRdnlKf3Op9qYo1F30GYQL9AzTXnkE4S8XnxRKFd+r4/Pb8A7+Nffq9qv0DsbbZjbUtgw1DnBu6w0D46EDbPmaiHTVsS3fAyRt/v304ylrQ7j/07lUJQpdVZKQNAlEj7m4jGeAXe7VhnplfLZLWN/baTVPui62ie6lW2ivnccHhBIPlLnBfkRDZO3h4m9Qs6csvODw/RVjrcUpm4z+zRcImvaJ6bhxbT9Oij9C7tpN3x604RUOZDZql3PNwBK9GHA6IMe8yhmEf0yD89G0J8XdKzOGYKtgLTASEvOvyy+55oyuEgDfOnE5l1NaTjvmcDrrTt8eXal9t/MgAr2udl54CJGhLkvnpu64OqjtOG4VaZXg9yh91btAod/j4KfcKys+FR8isZjLMbQX5agxK+bwwWQ1Mw84c/4D5OtEtWpDbT/YV5yp9yx479z+xGuJFtJKxfigQq17ogyduBiG7gqJgmLflFBq5ZCcxJQ4s0QFmyiSgc9nn+NJzQR4rqE1Vx4JDTSP0FZtQBl2ul7HvforrWiF336CGBS60tCu+kcIQ20Zb6wjWIyxu2RHf5yQCiUmJMVOBc+K2eajUxM6qQUf3aflvOJCr8CoKJd1ulbra5NdKmEyt2zffrtnmvaeEWRSVV5rStRFVN3mIJwf1T6t0+oe3j9KQpWUEm/1b1W/40WCk9gxRr1PVl6ermcaO3+A9O3hHdzRX30Z7VLxXqWkK7EkasiMAfb//WWuWafCrm40QSU0CRovXyeP4DjKIOichdkl/tudkOPpFLtobGJI8rac30B68egZ3HGf0LvTHodKe9lf1M44reExgCxKsFV71Y3GZL5BtxK8mVcmkZcVjmfXNDKukOnGgfR/FpQYe8cZ+Z1kA8Zu7LQPPPM8BN3bNqDG4eReXLJeWq/SoLhkdj4RZELRahqxiYYfSl93uiIMS1VdvJbveN38Htd1mW+UpS3Ji865n7j3YhpUR0fRvJ3r1cYrGzx/3LdvOuP8L2PXd+oZ/on9GGgIvGvrflnYB+3eO+vOnhLUzFL+gZ5KWobg9AjG/3SCkBdLd1wkNWKCsyMtIsD2n3O3ioge0ims+4s7rD2CkH8VuR+ne8WBjL1mxvbNleuxCCs7WcybFeoYdaJqRwS7Y6lZ4oVSp1stRiMR5hstV3lpEt2jfaf16ab/cDv6OjAZ8xBmZ+fbW+q42qNpgBD1gvs/WEv7gRlqGlJycqwYERBEMonjbk2jaUX1lHwxcZFBhhkgH2sE13nOgqG2XPE5bfJj7tXfixyT3/dcwBk/hdgMXmwFVVRCjEeBQ0HqY08iy4/hvHTmsqtlE4VIeJ/hEVlwYVoW8PhCL0r1al0MzBfjxrzlAoLFo83OB/bSGefr60c4QyuCXSNEbWDi1F7RtEWspNIStYyJAhJMStDn5FQyyDSi7MWaz+YeLCc52P7sVriK8i1HOJaVIK+c+Q3cmNfH/u23orfhW+rHwPDC3cYF/cBmANpKahdGgRiGnLjLg8vO4426s0tEs6s5HRvo6vgQw70i14U526+vqFPUrx+ntm/mJ/QMvJv4ilvRYm1vrRDcrCX6CxpDJ9pKlsCZhzAZcIJzzMdHZHk8yhwIcsyZGrOItZUkuDDlMRD36DplWWmbrk8DK2ZDty43i/sru6PP+xTFil1DULObwaoevOvwDjaDjbjuRIDTdYlmlzLmOU/CcI8x37hqOmXCYVhgl7aWqKE79VVhF2EOyN4r+Qa/aI3zvJwH7xlTootxcNCts/qhdCMAmVgA92BnAQJE4TZuU80iHbjPCEkWeKsuS45slVoK7Ewfor3gDoRvAxiz0ZlpbX+WtZZvMcDA8yCUXb2aY7ZX5uPMpU3YtBBX0ml58eVzUV4stFywbvI1V8yIeKXb04ncCpfMRpBK1CarG9swfJf21/c7CB09f4JXMw7pSsjNHm1Vz3ZzbY2cpq5zHclvjD3toBgf0GP/AUNMulgT6yoRUFJgHc/GkYhX8jrgoLXKcthX3XHt6Eq3c22PeMi32r1qWQ3eIrlEnD6anyzn5V7ZdM9RQytP5YrN/7lX4MTvyDbNZffNQvyn5kqYC3NWdLoRbUHgdT6cVQsW/dUZp8pDjpc4WDVHLu19e5HXJcJBC/JRs/CwxOMHWbMLxCDT4CRZqN6olTKh8otIlBg6Zq//mV3l1ux5ySbCNKD7aw0f4PEJmhnnVvZDfjshoqmb7w8Ugx2SqJrY8HgAk2FgPNSbFsto5idu3WVe9kn48JxzZ18AxwS2MbFo52Ro3vkNFCAGh3XcrBe176Fn0D3OV6ziNfPgLTbaAbMdKrgWy6624hANGycan63PB+PHjwQ0HuFdwSAflXYrrKxlxUL3HXJhWU0CX6scLVuw0KSw03lt4TeWWj6vcNEWWbu87JKIz8uOnaObh7isawLOe+zycUB9n+Er5OP6np1HsQNzxQfQqpVkKTHBtpcL+5xfn+68Z7ZjYPj88A6icb89vyoedElkENO145DPthZzDey4MUiMgRkecVQXib8d+fgCPJ6MAm7StXM+vwVp1qKUny3KHj+CsmemydlcLJ0GX3bG4Xt1q9R2Mh2/h6/PbryaKlHbONl3cMiEAEcM7ZMHkGPAxbSCqpRHoHwpn2sRbdJkeAwseptlGzgX9EkgeaAD3UwnGt+rpFwV7pWwHhsl1PZXSQmdfIVHYQ99w2BcwQboBd8j2qMIBokcKmmxPuNEZCPygMwk6cwroPY5waw2Q/LQzmq1V4r+mHpV5rff4iUOXa+2N0a+fd36uCN2DC29krl5iIsVe825Ub2InO1BaWpW55izK2aSlg6EPTmmdGbeZHljRkGMg2nf/nWNybSt6JUajUT9QRScGK4iOFYl88xQQ836MH2o+mKyV8XxGpZIDDoQLshLXxabSjJl69XKfgLEodWnj2rVcUbfh7SH9uoV3auLd7DVjRcrnFsRwRUL1qBuvkJlAEk7CHAe1vffmR0OpZoF6GpEdLp4iFbGQEPmUymX2H8tp6X8thkau6lWZmx+HW3WS/TONQ3vn0Rc0sN6nINd2ivesWHTDnHG7mPMMh+VecxkWi+UK7Znd6WeYVEF+LRArUv4Xm53intFt+k0NaOuh3sVGUGWvMKMAOCOxKBjidPgC2zrHtGirVQfQ/jyDUOw+YATg/uQzYS3bI9JmFfC9PC5NzByFojtjeFwnKdoU/+eYlffnyDKu8H6jsDUX89bDRdttWRxulk4rh+FzXNOULBCMR7teU+w4/dDcjfGSQOtETDvaiAfTYFTHHa6zhknTF7Ib1GnYKEYakePIwv3SnZeZQ7kW/oFCoLciJ1oW80qkTjUJt52iWYpzZTwtOzi9BuxmPNwhnY+D1gtJqXZPIKEiuaUXHRcqzYMOB86La7vsh+0nO1cL/q8YWqGXEIQZNl1tVfRlYOJrBzV8lECJaL9jWeYXT0+RmAMNaWSTjgsp8TFypoK7MCuuFqtMW680wN41Kod9ypq3C/ewVaiOHjV1fY4sItsJVbgqEX0qKT298jkwwe0gVfOkihIsLGTNPDMSnBRzfykAbHGEPO8ktjRVgHAItrjKij7cVrhoEJ/pZhMWQvwR6hERGUw3uPXshLZScDjw+zlO8reiL5BclBzmANHca3wkQDcFS3m/s/Y2aRIjixBWCCdYdZN86i1oKr2gpJ4BygJ7XPR0PsR0glaOkFB33Yqw93MPaSIyEyGomcr4sfD3eyzRUUMgB3bMHBJ9fhWKvjsvayn1B7WlZxcel6x15d7D/7wQoZXtwNbS8Ipr6pKn4Mxbi52zH/KLhxRYDGG2MTHZuzKHOuL51UgbjFMBM8iInYWVvjeNrv9dv1Ch9RW940Y3oOqFy3pZHheKVzAFaJQtLdlc6W+Bh0+TZRqAVoBtfZIfsyIS1DZFbq0TMs+JND1s5H+ZwUxzG77rSn+wr7aMMKezG7WfMh/4YPxbKeu75ZcV62tK1ExoHv8KDCoM107gXONQ9o3/enRLCyG3mm0wx3Y06s0FiVXs2n5nEViPTskdiVVQPEvLQZXN+iT+ZB1Fc5483m9/Lqz7gs9GZxWdTRvLo2cO7sFYZVgwdA3mdE8HbvqBf/+iyuQoMd0YuzsCHPzFEUozQVWxX4trbRhFT7VdllXt5Je9IfF4LwakAGQzBKQoSHVPp5KdL3Brvo3Z8IZ3gbGT41MyXOYueSyWuxzLcYa8GLaJTGV31yXHZNTa8WEd6CuKylG3Xl1+ljXnkxdQ8vg5f8lpXaj00GsKwtD1cyu5rKqBq6r0UYTE0n2ugeHJG+cCUrL5KynbLPP52fNanrHy9oK38k2ocxxqG8PLeTCPYjeVf3qkSjPh4I3LgwObb4PJM1/QPIh6RsAajsoZpQWm4DY+2ehS9Jd5jkNQdn0S1HpYcPT/WDBroXodvi38y2fR/9dtyOCSr/UK11dYsMpGLsase12UQhVx5zr7gyZk+wbFXuYkMEPuwox6RojONkluGLOFVftcrJbH9R12bVwD2NBKdi3+3n1pA75f1ReqceLer5HCjXXaWiq5uKWENPg29nfLBMJPAWNM6c9hjE36loMnrbCx5WSZ292CfL+W+19s+MhGPYfd6Hcg55lccvXV3V0rosJ50GyBEhEShtoGCphjfazD1zxHpGxEi+by+G+UGtle1BHE4uSVnUDzknvzS57cNviZeUuwYOfSu5By2rMa7ZrQDJfrYFc1U/wrkCbq5oT/7/r+qhk+BwhqbWTPaZj5t42p0nzXcUnQYsatLFcu6G7yPh4WHlN2rb74iocVVoz+NlEVieDUbOxV0Np5exdGchO11Cl1kREIh0M6sf6GB1JFOo0s3RNiADI7EAE6uo5Hz7WpH2YOeUTNKP8SRCjWg/5RrKu5F/fv3/wIMQo9SXfF/VBu7VPgmufWlfeVIJ8wSagYz4cvMmtq0ij7YTaY0yriBGGFB2vKNvXU+bbDneg1xpH7dDwwaQrKq0Z3YO/DXafzTF5oRvc6/mCBad+4LA0L2oXR82HJ87bGTAAbtOAoh0h4BAxjIk0F/NzTbCAT8rPztfsev0RK8CzXT4POjH+R67a3WuZ93mpULR+RXIe7RLlZ7O7B7vuBB7vrmFBSh4nZQf8wnhd5YN1Ce8N1EI0+lLX4MYecryi/KN550vwiHt9IUMh7x+02K6KA8IyzVfVMSZ7jNNK5KRquo9T2f4uCEOXQEUPXNpVyV24gEUEV6WZSRywgvgTzm7si8n0VP7VXjAAACAASURBVGtQdPgO/I+rr3497LfDAKDzU6VEt8+E7XYuupk4e3cHSs0+fqiYdqC5y6eaDald6LgCBrEXFZFCVpORCJs9CrfL4jq0I7rF+9BkyOIxyawrUcW0BjOU9I2HlGj1gDeRuQsfq8cDJzKVKOjq012DxD0mmAIRjoE8gcmlWSekMbau+J3sGgxd411LK/+1/rHssxi9k57j3D9V5ZjaZe1x4+bOMIPjMdi4kE9zWI7s9Q3+eaPOyiTAcHaYKx+6MbPDHq2sza0rL8yWY12vPllT24YOA9bV34g5kOPJ1D4rQckxZeijTQY7csE8wElezh8RTLvXtw2fOJMLNhvyvi4YlvhmXnV8c/aTbJjdqHhoh4LBCWn1+2x2UPFsx4z++x78ectpP2o7r4jaqcomnE4DzoBTY8Ug45vE+wYBg2hfjVS0j/8fOJAY0jJ2w3xAd3zPSEjNI4AOdfQT6x4f2uQTDcNxqhl8TNxLVlNUE/nYkn4sHdG6WIN2VeOTrtlnUEHtW6ix3l2GemSs7LELNRY8DRG1TajQQqUcr9q3WhIQFNFYsWrHJcjH4GlJhd8fzJ1vBe3HD0bnadGgKrX6iVjwxlrtLgkunOzdSSL6fQ3qTeiBqxm1I+v2mdo0zrxWlWhPZ6gAVpXauWwXcgfKoY5PFi8txC7dCvyre91eUyFavVZ6YBU8zg1NOE4j6hIGr3ha3YHT/UO9DzGYCKd7zgM3UyODOlTVQ9dAa48s3K/qIb6aU7/gm/hte/CWfQ/Kp1L86hPrqgN6lRPnqndihh6nlj50PtU4H0JddAvqBpzYuxoTpEcH80WWktBWEwJt+JRYgfISpIyPY+b9+q1+Z/Cil/pKHbsRm68uPXIYWHn/VH0Foj3gAl0EYzCo/TgmoGAlBcPiBZBQiSJIcL5+J4yYrXFsPT4ZB8pnuqyvP+YfjOVX8XuwJRKFY9Tya7A55ch6QG34Xp2H/w9mAEA/Rv3NFjafEH4s3mMpZei0GDJNRURzSvRoHomdR1YosEy+UFxXWX77Czsy0cS5LkXINpYLXvnZfGcZONF0ftC/5K6CiGIce/+p5kj8v1ifz5CFF3TaDmY2pOyRn1lm86YO3ZLn1deXPQdvSV+qDiVCYWVDnCcicLR0cIl59J923Ia8B5GpBAOOw8eMBX0ons6LWCxXhEikLRI7CCi7X1Mi88DYJnVauTnOy69baV29KhisrQkyLHSuGmMMNA3Qvj1lDN91VXehfGiTb3jXJTUN5r8ZkGyWwWizLXoP9VwkFiHhlVf/qTUXbBbPA+uI2saX8+q3Q/xm5oNOy+fa7W3ZidpEx1Usj2n6U2ylxVsT8NEbkijnJrGTXWE76pefII9x3ws1KBqisrJWStMOUae5RmhqXXHunNFfvSie79WJ2+WB8whfAaT9fT/2lnUd3oIgM2ivfXjjeRUO95GAnUEUH/46XC7ueS9nJ1ZgTqE9Nsxuwro6fC3KdlX6bP9rEVWl/pWEVzJtt3omGBzsVRHz9ZWbzvstCNoqyAKOi3lRqGXafOwgRzywOVEw8HSXp43wmg5M5FWllt6CPK/CzOunfxJGdXtNtHat5MfKTq0c6AqhLubXpaa2PwFqP0cVyHyOMOxGAVRjsrewOIczYvIYz7Wk07k28DDxIoyE7JmvxPPKLaziuhI8X1t5Q3gZo1Y59HHXMVVCNO2O+ajJlVAdDyqVYRzxAOBjHsmgu3CiTelaWW2srdjk02nNZn2r3XWQs/UV3jiZ80qqK/lm5sIpQudwBQZXCchgplHzDhxNI+6ZFTQ5KcN1XVmxvnAyOFPEoN3Qk+x/X90OtL4xJLQH+lZ7fmGxbpdkjlxfFDSG1hnCHwm1eQVqBo4f4HQpw7xmcMhVCKPE+G/aBj5fstPJt1ospPKa5bmBWKhdvsNpaQ+7A/fSuroVZl7ot9dtREVpi655fQpqp11F7do9Ph1Xg2lFobzqnVNC/uYkHxQ8TjPV7Moad7WoqDwA+7eu6AGpx7Efhc9kfQZyP35mPXEwDT7xqXy6i40kLOAsGFHhAACaVoW0dAxOmAuOJT/J7FkMMpUIzb5cQtduC8pNJuQdKI/Bfc88BjHH+dJllVlXLy/6qaracVcfkqIBcIrEV8h0iYVEA7UMCHbpR9NcXV7NiwPU4t8zEhLu3P8zEnOj5S1aWZv5TnUacZR+f2yMIzDWQp8B7Jj6IbsC25CJQb3T/5+H8zofHEfNoLKskj6CYg655BvJcpk1d1FlfMu1eUxe2sWppDLHLfNo9nMc48lk6/Zao9RRtJfQq7r/dM6s9VVQsas8Da+ct5534AiNTK9+QW3ywV2ZJzFwfsqZxEUeSpvS6rShK7pWhyn/M4/my7o6IQdO/fbILhE24SODJRzObjQvykco1biiWGRR+G8dBnsLDjmnhNKOJ+bEzutJ8bHTJ7Hjg5FsxbI93QuN63bVIf96OYkgo3swvAYrQ+209ZPKR9+5sqzr3lMMTaRtLpzJt0VPusc51v0rl9Y88wlpjJH+rWW1mZ0kCDw2k4Zm63b6cfKs+x8qJ3pFEdrWxahdDQuCDTXoY3pKH7v+6rLUPvsns0rYXhgNOT6kE8F1ZcHZjOipOeW/0RwX2YbQpaHL517P6XV1ysfJ6JCVHyOyK75viiHO7kMp+7i3mzAFcJKoeexASNTKAde8ETX8ZoVdKTrXN8uz1tHpekROpWwb5voedLlLt8y6Qo46EhNK0LnGHKldHK8kY5vOF6ID+IW9gK44cGYE1QmlrcHWi0vL8xztebmgVoEOtebxHtvkxSS47Q9qhu+6/e+vR0zyF6/mq13cfJslonTYiU3FkoEZAFZfycF+D0t4H8jmG/y4eboOu67NUYuqXOc0ssLEj3YLYha4Hxya7s+tq1/5s/2H+eEcTzRbubs8xs4CUcl7dHK+oacBFSEJvVM+4lPlqETL7ChXKvqYxS14Mb8pVUctu874pgc8y/WtcA/GmzC/rlhcteUGg9okGnZjmnPA2ckI947MrmgyOIwuNnbMcXZmMPksH2E9kwV2w42vO78Sj3YVOYZ7cH90XplY7We+zyBYbRV91PUj9bHSdiKpdq/4mI/YCDeo5KM/eXBoVzoZAGZ8KD8dxAWYlKcpWw47EPER1GaHPvKjB473TfDxnHoP6sxZi6u6PMLhQcUnTuMdS5GYz53vWFS+Es1R+WJZnzTapWBYloxNcHXpEd9HPEor0hf24+FV+AfecJGq5ftXNbvH2hZtH40F5aQSsLbnjzM07z6NEJ22tPpseHPfgpzgxN32xXf7tN03oR86repsvmhp8XD2+BOpQoFOe7gFfX0V+u1JL4C8BxEtUT/qx3B+gzi43viYkUhtQMvqHQP6QVrHwPIlqePnJEY3kUABuqTATRg14w6EQX7bTWv86B78nZRAXvqibdVa4HUx5KyJKU6VVQy9s85/uGTiUYA7g+KISGdP6bRnw/1Py0yEkwLaNWD+8mxmLkLklUdN9URDBn3RL4P8prkfOkqV0UQxmrhBcl5TnWw4ijJ8uxCivTptGF1Qwr8TH4PjdRYxuy8HIt/savY5fgx6Hu3utqEo1J76PbeuguQK/ua2zBjAxBmR81XPUhRhQW8fUjB88lxXD44Jr4h8HKdrmy+OowpHu0oYlov9DdCYzYvZOWq2dbUfD1fW11e8rrL9dgtyrtQFUHI5N2iINiaq7TVtvrvoGFCu+4xP9mPGNFxgIRg6qK3WCQVWvKgwEkTq6cn8LVidY3tyXdl08FbUM7QoQ+v6kaAWdDB5EMoMtaecr090jy007z/izl43chyLwgJKz7DboeEFCtisgLVzASU9gSUwbVRgwPkQ0oSdtOQHWO/6bbek+09SVEkbjGfgmUFjEoKkLu895ztiF6TY5tTN7lTKmXfK4dyv9NntvjLYwgcrhkf31UVV7Zccf7wiQR8ng2vpMeyrF91puAKU6K3lMSoyK1rW/7eZFoPHIEYEzSVkV4LDlGaoGCqnYbvFt/c7yDkABftwcspjXLbK5M0r0tXV7iuVCS6OCckYbNPGSvp3DjbzMeJKIPaDDJyxCuWlmqZpeuQYPvwdpCr0ciq2zRKSFiRUBk20N6ngyxUv7IpOXs6p0srrJcO7HQJQKaVEeUoGDqmUXKBRsT1QwffghfWbLXG39436ipW1BfCJ8ng+biLXsquQjwlN0TccDF5BpP02u5VqDthtWM2XGjjrDsO8fr3nbmjqwqKQdFUvEAXl8YphkrwJ0Lff1vQMUjFsRcEVEpcQOQBUYgnqjxtshyL639rm07xHp8rRjseCDmOCfBRqzdCYUd/rg9zsD15Xcl9RCzndvxJ75WkTa69TgyzBAnVXisjQis25jfT/baIj42ztAD0ZZzyoPtIwYBXK8n/UsiNlLjNpju+rZbVyPOQz5VfC1X4q1iZeVcFdBla3w1IVZNmNADIMPVaRxJBE1UWh4FQqcLanwNNWIL5I2OnD/Ai5rsaHN9bvB3jIt6d/nYhrf9medJmWaFEqSm1ZKU27kI8bdbNTzCd6lbK5ShxZOf906EJ1CYo2DSTYp8SAVRqhPnhfUU8GVUW3Vf0V5UpcLisIi5IqhUK4j6Z5VderPNFW8sDh549sAhWzPTqO9HRRsvVI0N5+5I012gJra9S8tq/WcwGeVLjEJpiIJX3KLYE5OBUXoi88RsUUdSxFa0JEo6LWIh8dM0SNZJQqhn4ld0MlI9BbcG+5IN/BD5pNrN9XJzXqyrgGK3kOqqu9Zjc4eAAkAEDy8tR1RZnEiUQXH1BXHc6/esXxtTOJkdTsQz/I/AaO4DA8XohiffW+qW8vRNW+SfoAIVE8nS/DoE+0N0Ojb0mTAFdXi88cuq/aZDXqGffo5M3sw6HEqEjHg4q7YeON5i88XLffNvYVNtpPGVp7qRiZIG5nzkAl/Bjjw2kojZFScKgp03WpldLkR4qCI8+u67yZy4+M97DhQCj1QEPX8PhSmfvqltEhs2Fi2VWXB2JRAz84Z5zhIHXpXTXXlsIS3hYpEZrAmzbm03pzEj0B+WAwLwjf+HYHPS30Y5iDKdzQfd9Bs69uyTkOJ+8Wl40Y54qj1NVXsMKs+VLJrrAMXQJkSXJcWzFRDB53VqWGvRjnGN4Us8ZlXxkLKsJ7hx01g/SvAutS2L86iWf3lPkEIqrdokTpurJFwytnACCvvW1JUNspQuZqbQXUJjKgAhTMNPn6XjTalhyKvCY9ldizr/BBeF69r5ZcuO2cz0V/jAHhRsqwEKIZH/PKffYawHzNcgRllMoTnDaadoGEAYRECjgOi+Ut9R+RfL0GF9IXcNi1Uuq+sgcw6l8hQOZUnLKaWrTgVHapCL0atkOpz07r1ZINh1cqqWUQ7iqV7b1D1I5LWEpE7KGsguR+G3fdV2xeyuwr1j6iSO2yegJL0shgp6HWxDkTCidwvjqIBk9WCz5MVYKbClS0Pmb+E51d6WIowgX6C3teN4m+6IqeYfal6vH8dhQHG5dqiRkMns0UCm76C8D5QFB0KrTLqWcgFfKo+fBBCuOoQI+9pcwR7H/Hu1nr20nPkNaLctG+NUMtqRat9FOQcolDKMMymW/YggMGr7YztLk27sZwwjXRmzAO3CdGEv3Q25WiZYLfu4oG5n7k9Qwk59uQ1JYSyyjVFWWWwL56YTf4G2hEMeEFp/N4XTGntknOBp0o/DwTiZSjZAiXqlfIGJ6eDjvPoPBk3jO6PqZdXdBneVnbVVVRsF2p0JyPWfeIT8ErQXYwuGvOBW8M7qrtYpCoOnseBqiIjiHTvPkKQo9BoioVsgIGEg/orXL7al3XR2iUB7CPpeiQOYOjgjS4qBmDao9akVc7js9b1dQ6E+zSqywJl+DyBVDMUc8Ed1bu4h+cdTJr+4qb7ZctQV+JiwWZjEKSAdOErRqureJjauYjsmO6tsuEK9m0Lu8TArWBIhJ0cUU4zN2fQeUftA7esM8AqSVbKDX8CpL0uC65vKpIoHbVGRzIXV2qq/s1VbO3iyaoTcpSoi0A1OhzVvg40kIR4UNwfPgB3H+1T1/Wu7Si66Pu8SWH1WYykeC1a8W118qrN4zapckEthe6mjnRnX3icI3geNLlaYDDWeAp+fEYJ6ZTjMtwYF+9LxdWiL8K76tTdijIdRUq1LAlWkv3uEoSyBUXTOCYwFPbZIl6tKESntbFQ4lR/cI6dFKc453HUGWfwds5qeuTIPVTlveBZDD1EUSATFXV15Tq47WWcKVWKANR99gMUT3rHjs0DPahT4LB/zwVxCp0GIRzfOS++kBW2Gqv76lg3Ef+skLZY4F90VIBZGDmfCVoBQv/21qDdlqK6rLXlaEwUGTl/MJxPc8G42HziPw0dauTC3waDiyY5AJksoTOyox6yY6c9b6qbQ6HPJ2xsHqtX1UcDmeCi1S0W9GoYXVF+0o0Vy4wSvQsjhl0oNJ4cGcFfIY1nxfNUfO1FXQXKPMavV34bF74mJLc9UJ4zIbS5hcnHAehNkmFtsqaJ5k2smmNqwtiwJe2sRrg0GBeocb31u125pXkbD/rOWr6trKonYpCZImRWYZ3VcsitVpHzrdsmW8Ss3nvTGwXSWNSArWR4zdkgEq51mAD379a0kIOA57tGcQjeMmzj4FAjiEvJNWmV7MsGN9YMu7qOOCMZFeJPp8OQlWBxL03EMOBsWBjr9HQ02D21f5DKLmWGZ/XM0uvtpD2SyVa1CpgiZ274aj5FX0AeLV3bRjuEuo9nGU5kf7fed/HjA9KjQ1jp7Bk2NmNUR7edxWQk1wrHElcirUSq9IlQ1lYy1LFbL6XgA/WEmuHDyHmgsfaR6/ItPQR5GizBDqGRXxKGyP+m/FIzfBNNQN8B2/JOc4zGuHwprpkUGoVq6+Id1WbVp9KsIQoHBh5kZ4PUTtNm8lD9XhZkUrUpxKbA8IHz+WH6VhttazVh8ydTeSL2VcUpp5vilJdJUKGmqUMc2hXZQfOaIQTmigjamNKtOMXoTwGncf4DTbPRypRmgqKjGHil/PuJftkuWjujfP881RsEn2pf4WfwUKgj5VYu0Sk1uAMRxC1NUYS/4HqK+Pb1ZQB12G94CVbMMz1hHpBz5tZon10X30uJ/CLos9W5s7zg5B21SUL9YV6XQuQq4DHMLdE30jHoN43y0iCFMgr6GOeDNL4hkaoPhHbHGIxsV7YpdK2a/XNlzvOJlK9vtsTzyWyWiIc49hgiUqRdl5UfOV8Al8bQTK0II5RJud0OYqzZ0/PZlgYZxNKBhJo96PNFBzHw/tKjXGCKGyzr84Fwu3zqfNMZFjCiUv1wLkGM+cGMyXAQA9VFqnamwjH4DvNI8ITSI4u51yCXdFTVOVgpcckTjuwsZS8PTObuBcNp8vqXFAkatwWnQ9gGfgr9QwHilGmOGGDAT+DKe2jqUX5vpoFo30wGiTWo260B6kb07E761uc4WeL/bBrRZyBTAeLxzd0t2MvpoySPilrnmk74lkib1cTixfk+CGVCCeCnU8kVnJWkOrH0C118BR+fnxru/Mqrw+6fafNaBdp85V1rfgV8Cl8EQ8ODJ2bV+kwiA+8Tec3az6mU8mxBHCKzc1DPEAdx/FodfU5V6IfqRMY3FdPcPxO+dqK9hSNJlB/jEYAbrZzsotQH20oanSze8s+phPoRCmakmkHQHuWxRz7Dtrp4Gqvbz6EpGjI1VYVypDl4axCOGhXta8qW4Ir0U5lOLcpJoNn47xKAGC+gLMeVML3qinqyEzMo1W7SqiyL+dwrX7SIp3ywnYOkS14V1WVrq/emCjaKtsg4kObDs9hY+JKvI0tYZrvEl6ZkKiNWLcHEcT7VUT2ujL76rw2H7xXWBuzwZIhMhUifZUNJ4FkuK/Qa03lQtuKvL3rVqEooGfn9QIrKn4CXUJ+zLVVzwPnYRgfYxKtDAd1R2b1DD5ndlUpKQAlB+4WfFkp7qpUofeqYYGoaY4auZzT7Bj5dwGoOe9jJtFI4UryvIGv4DgdvthN8FmESLFrdT7/45TfVqWS1JZKTQStPt2NabAdE7pw5IHTrKCiiRatWL7Y5XOmzUc0MLyw4OkszP8jpejycNbhOOv11VK6P6J/LPQTp16oosoHd99NjZYU8QynQ/kVFldNvEjKWHL/jZ5dF8JjBgGJDsqyOzHn6uh19fvbJChkejLnpYeV032Qnq/kFgPhMRepKF3upH2EqeAs1G7lCLZvinUViIl01TDDVomNmSD5UqCSDoAbg0DB3T8fMqAH+tX6vjovJVa2vILJIFOcaF9VwHCq6BiCbZdyALQ+BiurMG1Xm3apy9chEAU7DJFUe9RqPsF8DA8AwdI/f/v+UF/B5Qie18/g8/lnNsKyomI0NKOm/F0tTeiZM8AxskkfnFd1g1fAuT5w4IyIxxQ+u8pzmaZDE1TaVpy5FA6do7Waq/rcaxBnqGwLpzw4Rn2Y9+AbzZuxw9fAnYX5sYmb3am4IFEw9C5K6yIdEeL5xoHVMcMwHC6v7ttK9hVl45xXz+B90z1t+JwrnNKLWpvKUMY4LW9B0F3NppJXOYIt2ZXCM+hESuRp5NWTC0B07YpiOODfRv1PUS7DoToUPoJcXd3W+QxLv+++WLmXc1WqPl89t2RqRvSRZWkuqkB3BdxxqRi6RgyDnT6HLtxgiD8Gn0TUDx1o3iXMY4IxiGN+ONCN+dCaPmsESNxX81rmWqIsQoYLngQyVGDpMLg60vPhl3AFz+ewYKdfeFt1ofof8U0hF3PgbK7h0GWlL3bUiubrqyVD7p+5qM8S4SiLEc5kqdcW5IRqjwZNONpXErbbfSwq8hyVgBJRF3oAxt7ojqd5g03sBB8ONGPmI/jxoSTI2bfzDTbWe175QTJRyHep61I7Bq+17rYLEwU7xy3p05ouVYk6JRTFF07nQ20Mud9GGjhT+NT/8b5Z6Npf0uWLwnGSZ3AWlL6vjlF5X1Uk6KtKHOKU5pEDX0CQtTe12Cv5NdimDiEPBpcZDgV2mdcgdtlJzLCs2LJeaJI41Lf6+gYZA9Eycd51y93teGOd338mRaI8oVdkBv4GRhWW6h037CppV/oL3pRZnloMvk+gdtR9JVbwSVl2D3VimEXO3eP83b7YwBZzBe7G+//9tWhNv/Dne+bmf8+//7qf/8A//r38xf8pf/Qn/tGf88/n5+evz1/T33/898evaf77/tf954f6/sEj8Ivxj0RuDz6E/2vvynIj12HgXfjBg/FDgC4kILd9sVjcZLnbeZnBAImdpGNrI1ksUYvd3WdeTaQ++2uLAw/mdvycjiNpaN6Y29WAd97B1YSphiYh09PiBIkj2u0NrcJdEw/9VADzXT9SzXX9ONG8uXHep2M/E3p4ez10y2oku1ja+h6v3T7DfDBkgiVGK0X80K7rzbOphqr7qUk/LrP2qyLQ8sOIXrKnYR9tDkJq3XGM2equvQmh2qaHesSGr1nC8+ao1tslSqncdN44bV29iVdszKq86mDV3LIY+qcSVF3nzFEGJ0YH1WU0w607/xooBp45N52Z6BuWqF5pw4oEYgpvi2vI/3Cez+LNpePFviY80ZnPbx3cY3VARceYad3QjIZ3GlaXuhrwTO2mMKDBDpjb0YG1XHMQTVs3Zrbm4KVihqj5wJwzFyQDvlIGWbiAfJViOnZ0eeXbcOOcl/VrJvjNnGEuCkXBimjUGmAHGcZwR5mDG3ypggfgNFVgDCiJLLXUkW5oBjF2wC5vqDWLizBcKT4pMV2BRiEtUGsRIdEXGvwyjMc5EFu4one8krnZoBFLfCxEdwFDhkPXUqgfeqGsQ6nuZbpFb0c9Q9W8XX3B5uTw8t3iFNrsuf4wS81LwDHYb7E37LHE01DF+KwBobd9UHm1RqwgVkAw/P8IfQLMUGTkulag4pSKJj/4xajXPSX0s4KGY0od4aHRFhnDWN08shMmDO/mDEot1tXjdnqwMmmv94hhv+aO7ZSjJQ4vNOhLkbPAvoaKIqpX1XrGbZxppe/dXfaPr/rgRFRmbD+D9bMPkfhanPfxCtNR0t2uXwaVfacznxc4F1hFxPplaB0WY4NBNhTaYCW+N8MT599EKv3mXdoOgxd9kHSZLZznWT+dU0oq7LLf5JXBhZg1g11ZSv9AoI7eg+6Hz1/lW1gxtpLtECUnTuPXZvc5yf+lsqXKUqFti6GlXdOv6jRT8VLoWWG1QnyuwLzZEH3FK7FbOjYaQvSB2sRNT7AcUFGaZP+al5Km+VhgRunldybGdUNlltx0/ZVWLjRYcFZSVfZsbkt5tQKjmEjsWgnf7oN4Bl7EJmYTLVJHmN7qEDi0Ol+ksIG9kl/tOeI2OC83xiWIchaon+S6DsGzhZI6P5AgFSlYcp9X69wBH0NTOG6pDhum+vrVA7kzAh8PfqFx7R5SOzoirWPHPovJLQbMJo1dBEer1souaORjskqIvoLVjFe2iiaS9WiuOVyTUlNGy+yuIcFA5KVOKVWudyq4ApsCF0LrQRkn2S5tbvFKeyHtteQEUCuCV/XOtcPd15btfHIln1t1wMZrr+Xb3h54xcRfwoptQqaA+5RLp2qxEhCQb+uq5HTPtPK0Zi1eTgnbUqmNQnzTVnZVUxS2uoLppyLFL6jzklcTMdLvGTeK4gNp9Nz1cpn+oik694iaiHvkpvo8LgrF1I6iXS/ByXsuv4XRxS3E0XgtG1B7qzpX2g+BN2K7eEeExhUJgZ+w8azQEEeKP/CFL3Znm+PqlV16elEcLLDd3NxJXL7eSpnxIqSI2WwP5gE2A4LJ5ZPvrMvmZsR9rCCFYDjr81nTLhWmpgCVsFYihRIC9qea4VbliU7xotlBF0psphBnNzQD74JhhCIhmxC4NoQnQi2k83Vcv9EHlVfq1jmdDcqwxUDN5uR4zOeEg2mO8Txl2yQDp7ynO1g5YDK0ULNUmKAiJ38GS1VRCeKK+xkODPn4eB0JYMvrygAAAX9JREFUOf8PKwwI7DoylVGVDZEcFSFSzA5jJLrPZIx1gQIVkd0R0MxYluHxHjULSxDG6SQMoPLC9jCC4U+cfUhFPsVamel7vEI84LDbAobfFYJ/7S2vbAQyB6+8sr8FLMRHAfnQhi7PxHFITHLHBK+Eo3nIJ+NpkL7Kj91g+h5WTAviXFG02OtKAjkQhAvgnsYOZuEPNGYKfnlqeSsfeTQkGzXKcsO7lEvggJCq/GhX6Lu8Cl3RFU8rAVk7bdbLn/mStajRgmFfrkvL4M1lpcpOk+g8QusGHVtrfO4mSf4dmO5jdWaUR31xCCTU5rRXcR5ZYw7DK0u59E/yocGbluugmoBxtbyCUCIbJ/kUrJQ/iZU7sN48S7uIUvuqLN2VkoreQPU7p0izm+2lQkJXm3LVs7wljlwK+mNY3Tlkw8D3dV7QmN+3td0g/yoY/wCrf3rI32v6x2F1J9A+WP3148HqwerB6sHqwerB6sHqOR6sHqwerB6sHqwerJ7jDVb/AYETxl4kmFmdAAAAAElFTkSuQmCC"></image></g><g transform="matrix(0.9999871850013733,0,0,0.9999871850013733,120.25196838378906,11.165985107421875)" opacity="1" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,399.87701416015625,530.9299926757812)"><path fill="url(#__lottie_element_205)" fill-opacity="1" d=" M-119.43599700927734,-22.22100067138672 C-119.43599700927734,-22.22100067138672 119.43599700927734,-22.22100067138672 119.43599700927734,-22.22100067138672 C131.70799255371094,-22.22100067138672 141.65699768066406,-12.272000312805176 141.65699768066406,0 C141.65699768066406,0 141.65699768066406,0 141.65699768066406,0 C141.65699768066406,12.272000312805176 131.70799255371094,22.22100067138672 119.43599700927734,22.22100067138672 C119.43599700927734,22.22100067138672 -119.43599700927734,22.22100067138672 -119.43599700927734,22.22100067138672 C-131.70799255371094,22.22100067138672 -141.65699768066406,12.272000312805176 -141.65699768066406,0 C-141.65699768066406,0 -141.65699768066406,0 -141.65699768066406,0 C-141.65699768066406,-12.272000312805176 -131.70799255371094,-22.22100067138672 -119.43599700927734,-22.22100067138672z"></path></g></g><g class="png" clip-path="url(#__lottie_element_207)" transform="matrix(0.665991485118866,0,0,0.665991485118866,455.6453857421875,485.0458984375)" opacity="1" style="display: block;"><image width="194px" height="158px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAMIAAACeBAMAAACSg8iOAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAtUExURUdwTP///////////////////////////////////////////////////////81e3QIAAAAOdFJOUwAgTT8RfN/DMKRgCe+QbDiZxwAACExJREFUaN7tm/9rG/cZx5/Tnb46DifHcxwCQjp5Q12GUZQ0BAJGsb2OEs9ooyu0FOGmq9eUcaiUJSNsh61mbKxgHCjLBEEYlm1lDCMn+2GQX1yWNhi6I4PBxn4QdWTdWeb0/A17ns+dZUeWE2WDsbF7E9+Xz/O5z+vzeZ7Pc4LnCICv/08pcdLTu3yQB7jo/a3lRdOZ2jG1X0JEI+Wf2iVH5qEsnZMA5QXRpNXK/RMy09PTNXEpnT6cMEmja2N0rXYR7i08k5DqXEqpwwmBMZC0DAxotS7CZLYvgjJMHj55LEMBubhYAym/psYHRk8CDIwOuwTqFkmlQaF/3CQICvWQyqeH+yEMUCRycQ7IPU1L0YS1bG6SwzOkaRcEQcqAnNTIp1DWtKwgDOQ0TQ1w/2fFIR4fpkBK6QFaQ618di2nBtIZNZeZLicl7eRUWhAGNPJHWZXHpPTJYylBkNPTx1L9rIFmniHO3Qu8HCVdo6AG6JhbgLtjkfOgaIJAhyF1aOFuMnKaXcUEigBdDWX72UuXgFasMkFKx+NDpwJjwvlyCgbica0mCGU1l7+b5FHjcZdQXmTjULbPvTSZdqPJ2XEqcMojsKs9wtBCGuRTQ8L7HoH79k/IUVh5DZn79+/XOoRI+pLiEQJjGdrOubycqXleKn9IfaFfgpSh9Ys40LZVOwSZs8AlyJkxsqY5MSKdOAzE+4zDNExeoH0opach96GS20fI1CY9QkSjRtoTgczAkEsg41QKJlOX+nkvXaJBcuzi2j3eWR2CQrs95xIUygIo85q0VHovH0B+Zj5Io6Q/cDKrsDYKMHqeAkJvhDil9jD88bwa50tKiDi/XU+KDnFYBLpd++AGGeLD/g+UL1++fPny5cuXL1++fPny5cuXL1++fP0PS3neMq/0D3GaL/S0nnun2xI0cEN9LkIUF/iE7jjK/DLA1QKd3fuJxq5ld0IGIjafNeszT8zJUSPnOwTQ10HGEv1luwncC+AlfH/0957xUAXxiduzEGrtEeoNsm/BIOa7CWGbj1WaP08BJLfDRXfS+U6AFPUAATyCJLy72oQw7ojhuMEj8AiCoOAcL/QLOGFaN2Hwe8exDYH5n5gtFWQDr9PgOr4P38f5JDn6EYy8BYNXgvMB3ZkHfN20kvTwbAtm7TbMtuGnaBU8Ag1XkOkxFQK4zBNXI6ZlWmrU0hELMaTYlKDoXKWuVdRxhUK1AmCs83xD7SDGqAHIgNu8HgfGf2DRyJL5UG+7BEl/aNgy9VJp/a7PQlY2Zpai+J6kr8fwLcnYlohd3IrhXyVj3fVScRsMG5a26U54qZ0vtql5ENX6X1AtbkVpik5eEEKWGsEV4aUgT46d2eARosSsb8aobWIn6qhwpBmyyLPbLiHRBHTyxbldwrI4QQwLRtYsGKXY63StCgIdwCg9QaiuAyw1ozTeKhFUSOyEnUrldivR3NtLYTvQMrLG8i6hsBvKmw5Ul/ERSL9+DbOCUGxVKsa6MMfIAHDsrDHHc9xPCJEXsTWxs0cIOi/vkD+yXQTQ37Vh9U+YFanlEqr88BfCLOE6r2Dz4BpCNn/R5zVIwy5BxnObE9ct6CbU7R3aUJgPWakveYT6Nj2rumaDh8C5IjludXs/gYIGgV8doU6JhpcPplGaabUOEFZxkzLC5l0U8wgc1p8nXfMS/lJ6FVdmLFXW5/YTZPJfYof2mlKl2Iqkr2MhjI8Fwd5HmKWcGqQ8WnpMbwiX8JKtKnqJJ0mBMMV7KYC2wfmwR4CidQvnFMP6Ea7InEq0qXCB3w9EGMTP1Q4hRLtKxgYNXzE9gowPbztqED/jmZ0w0abzcXRuwhOEmI4befimSTkNRZE2MxZwlhBBqeIeYRAp4c0tiOh43SPQcPhD7iXWLp0Wb6SpAz8TijBEFvky3s+PSuTG3vXUjV6P+fLly5cvX758+fLly5cvX758/aek5A+2hd7uXI68/RxjfeUygNxdLP+6ySXXLh3l0t/I/Py1FUg0n4OwusNlticJko7Y1bRLSJDFUv9tQhTvyPqD3gT7zAtY+pcIbuHoovD/0RbARHN/AZwtLoFMxhYROnWmeOe5Qz5oeATJwHeBC9grPEyb2k5DRLTFDHyPS2luCZYJ9c1E8w28DOErxxvKG9hS6Tk76dXLDyWMOFcxG8CHXMKEI84dNn3NuUaLK9pkkXRHN3YJ1c2EheioIVtvhPEafpcL3w2vXn4oofgAjNJIKy+qk0HEa9+ieH8ExUaMq+xzUVyRdI8gmXMJfCWIhRD++c74Do0waOW/0Xbr5U8hfAIX1dVNAP0RNb5KO+YRFxbD7bAovCa8yEDCqhi4wrfmcog6vFiA2WbUyitxrpfPNA8nTOCPx6C6UanoJW598W/YjDqVyi/so1alUm2ON/bt1ssiH/QSF19h8jvFZgw3vg1uvbwXQXxLKQRNtJJcv8aS2z5uhfnOmuVjk2vBIXe3/i4F+wiDompbRHzHrZf3IIy3RZ0fpl7DB9U3uWoN8MJHAGEranEB/GiLDsOrj3fXIMbYI9Sbi3SjfNmwRb28136dtTnDsl/NwvgmB4ouxLpCdhC5AM5xiH281NyNQxfBXKY4BD6mzeHWy3sQwvh3qW5B/QGsbs3QRuFIzzhJpd5U6OmRFn9sKTbCTrazl7oJxSbNIohuvbwHQbyDPoEl65a5HMQrty2Vvebo/EHIqphvQtW+hY9ktA2jp5fsW2ZTxitG262X90q5EyZ/W6SE3MjDb5HzlzMYP6U1G2ir/FHrU4Bz2JrtRXgZLfLeb9B5xauX91JE/Kc+t9i9WwTf3+Ze/6z3wzAl3lriObe7r/8q/RN/suiwXqHjUwAAAABJRU5ErkJggg=="></image></g><g class="png" clip-path="url(#__lottie_element_210)" transform="matrix(0.6659910678863525,0,0,0.6659910678863525,455.646240234375,496.54595947265625)" opacity="0.9999970527921651" style="display: block;"><image width="194px" height="158px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAMIAAACeBAMAAACSg8iOAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAtUExURUdwTP///////////////////////////////////////////////////////81e3QIAAAAOdFJOUwAgTT8RfN/DMKRgCe+QbDiZxwAACExJREFUaN7tm/9rG/cZx5/Tnb46DifHcxwCQjp5Q12GUZQ0BAJGsb2OEs9ooyu0FOGmq9eUcaiUJSNsh61mbKxgHCjLBEEYlm1lDCMn+2GQX1yWNhi6I4PBxn4QdWTdWeb0/A17ns+dZUeWE2WDsbF7E9+Xz/O5z+vzeZ7Pc4LnCICv/08pcdLTu3yQB7jo/a3lRdOZ2jG1X0JEI+Wf2iVH5qEsnZMA5QXRpNXK/RMy09PTNXEpnT6cMEmja2N0rXYR7i08k5DqXEqpwwmBMZC0DAxotS7CZLYvgjJMHj55LEMBubhYAym/psYHRk8CDIwOuwTqFkmlQaF/3CQICvWQyqeH+yEMUCRycQ7IPU1L0YS1bG6SwzOkaRcEQcqAnNTIp1DWtKwgDOQ0TQ1w/2fFIR4fpkBK6QFaQ618di2nBtIZNZeZLicl7eRUWhAGNPJHWZXHpPTJYylBkNPTx1L9rIFmniHO3Qu8HCVdo6AG6JhbgLtjkfOgaIJAhyF1aOFuMnKaXcUEigBdDWX72UuXgFasMkFKx+NDpwJjwvlyCgbica0mCGU1l7+b5FHjcZdQXmTjULbPvTSZdqPJ2XEqcMojsKs9wtBCGuRTQ8L7HoH79k/IUVh5DZn79+/XOoRI+pLiEQJjGdrOubycqXleKn9IfaFfgpSh9Ys40LZVOwSZs8AlyJkxsqY5MSKdOAzE+4zDNExeoH0opach96GS20fI1CY9QkSjRtoTgczAkEsg41QKJlOX+nkvXaJBcuzi2j3eWR2CQrs95xIUygIo85q0VHovH0B+Zj5Io6Q/cDKrsDYKMHqeAkJvhDil9jD88bwa50tKiDi/XU+KDnFYBLpd++AGGeLD/g+UL1++fPny5cuXL1++fPny5cuXL1++fP0PS3neMq/0D3GaL/S0nnun2xI0cEN9LkIUF/iE7jjK/DLA1QKd3fuJxq5ld0IGIjafNeszT8zJUSPnOwTQ10HGEv1luwncC+AlfH/0957xUAXxiduzEGrtEeoNsm/BIOa7CWGbj1WaP08BJLfDRXfS+U6AFPUAATyCJLy72oQw7ojhuMEj8AiCoOAcL/QLOGFaN2Hwe8exDYH5n5gtFWQDr9PgOr4P38f5JDn6EYy8BYNXgvMB3ZkHfN20kvTwbAtm7TbMtuGnaBU8Ag1XkOkxFQK4zBNXI6ZlWmrU0hELMaTYlKDoXKWuVdRxhUK1AmCs83xD7SDGqAHIgNu8HgfGf2DRyJL5UG+7BEl/aNgy9VJp/a7PQlY2Zpai+J6kr8fwLcnYlohd3IrhXyVj3fVScRsMG5a26U54qZ0vtql5ENX6X1AtbkVpik5eEEKWGsEV4aUgT46d2eARosSsb8aobWIn6qhwpBmyyLPbLiHRBHTyxbldwrI4QQwLRtYsGKXY63StCgIdwCg9QaiuAyw1ozTeKhFUSOyEnUrldivR3NtLYTvQMrLG8i6hsBvKmw5Ul/ERSL9+DbOCUGxVKsa6MMfIAHDsrDHHc9xPCJEXsTWxs0cIOi/vkD+yXQTQ37Vh9U+YFanlEqr88BfCLOE6r2Dz4BpCNn/R5zVIwy5BxnObE9ct6CbU7R3aUJgPWakveYT6Nj2rumaDh8C5IjludXs/gYIGgV8doU6JhpcPplGaabUOEFZxkzLC5l0U8wgc1p8nXfMS/lJ6FVdmLFXW5/YTZPJfYof2mlKl2Iqkr2MhjI8Fwd5HmKWcGqQ8WnpMbwiX8JKtKnqJJ0mBMMV7KYC2wfmwR4CidQvnFMP6Ea7InEq0qXCB3w9EGMTP1Q4hRLtKxgYNXzE9gowPbztqED/jmZ0w0abzcXRuwhOEmI4befimSTkNRZE2MxZwlhBBqeIeYRAp4c0tiOh43SPQcPhD7iXWLp0Wb6SpAz8TijBEFvky3s+PSuTG3vXUjV6P+fLly5cvX758+fLly5cvX758/aek5A+2hd7uXI68/RxjfeUygNxdLP+6ySXXLh3l0t/I/Py1FUg0n4OwusNlticJko7Y1bRLSJDFUv9tQhTvyPqD3gT7zAtY+pcIbuHoovD/0RbARHN/AZwtLoFMxhYROnWmeOe5Qz5oeATJwHeBC9grPEyb2k5DRLTFDHyPS2luCZYJ9c1E8w28DOErxxvKG9hS6Tk76dXLDyWMOFcxG8CHXMKEI84dNn3NuUaLK9pkkXRHN3YJ1c2EheioIVtvhPEafpcL3w2vXn4oofgAjNJIKy+qk0HEa9+ieH8ExUaMq+xzUVyRdI8gmXMJfCWIhRD++c74Do0waOW/0Xbr5U8hfAIX1dVNAP0RNb5KO+YRFxbD7bAovCa8yEDCqhi4wrfmcog6vFiA2WbUyitxrpfPNA8nTOCPx6C6UanoJW598W/YjDqVyi/so1alUm2ON/bt1ssiH/QSF19h8jvFZgw3vg1uvbwXQXxLKQRNtJJcv8aS2z5uhfnOmuVjk2vBIXe3/i4F+wiDompbRHzHrZf3IIy3RZ0fpl7DB9U3uWoN8MJHAGEranEB/GiLDsOrj3fXIMbYI9Sbi3SjfNmwRb28136dtTnDsl/NwvgmB4ouxLpCdhC5AM5xiH281NyNQxfBXKY4BD6mzeHWy3sQwvh3qW5B/QGsbs3QRuFIzzhJpd5U6OmRFn9sKTbCTrazl7oJxSbNIohuvbwHQbyDPoEl65a5HMQrty2Vvebo/EHIqphvQtW+hY9ktA2jp5fsW2ZTxitG262X90q5EyZ/W6SE3MjDb5HzlzMYP6U1G2ir/FHrU4Bz2JrtRXgZLfLeb9B5xauX91JE/Kc+t9i9WwTf3+Ze/6z3wzAl3lriObe7r/8q/RN/suiwXqHjUwAAAABJRU5ErkJggg=="></image></g><g transform="matrix(0.9999955892562866,0,0,0.9999955892562866,-347.3302001953125,-41.7615966796875)" opacity="0.00004887166292562029" style="display: none;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><g opacity="0.3" transform="matrix(1,0,0,1,727.1950073242188,527.135009765625)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.746000051498413,-3.003999948501587 C1.2699999809265137,-3.2950000762939453 0.7089999914169312,-3.440999984741211 0.06499999761581421,-3.440999984741211 C-0.5960000157356262,-3.440999984741211 -1.1790000200271606,-3.2950000762939453 -1.6820000410079956,-3.003999948501587 C-2.1760001182556152,-2.7130000591278076 -2.562999963760376,-2.306999921798706 -2.8450000286102295,-1.7860000133514404 C-3.11899995803833,-1.2649999856948853 -3.255000114440918,-0.6620000004768372 -3.255000114440918,0.026000000536441803 C-3.255000114440918,0.7049999833106995 -3.115000009536743,1.3009999990463257 -2.8329999446868896,1.812999963760376 C-2.5510001182556152,2.325000047683716 -2.1610000133514404,2.7260000705718994 -1.6670000553131104,3.0169999599456787 C-1.1640000343322754,3.2990000247955322 -0.5920000076293945,3.440999984741211 0.052000001072883606,3.440999984741211 C0.828000009059906,3.440999984741211 1.4780000448226929,3.2339999675750732 1.9980000257492065,2.819000005722046 C2.5280001163482666,2.4040000438690186 2.880000114440918,1.8880000114440918 3.055999994277954,1.2699999809265137 C3.055999994277954,1.2699999809265137 1.6540000438690186,1.2699999809265137 1.6540000438690186,1.2699999809265137 C1.5219999551773071,1.5789999961853027 1.3179999589920044,1.8259999752044678 1.0449999570846558,2.010999917984009 C0.7799999713897705,2.187999963760376 0.4490000009536743,2.2760000228881836 0.052000001072883606,2.2760000228881836 C-0.44200000166893005,2.2760000228881836 -0.8790000081062317,2.111999988555908 -1.2580000162124634,1.7860000133514404 C-1.628999948501587,1.4509999752044678 -1.8359999656677246,0.9660000205039978 -1.8799999952316284,0.3310000002384186 C-1.8799999952316284,0.3310000002384186 3.2290000915527344,0.3310000002384186 3.2290000915527344,0.3310000002384186 C3.23799991607666,0.2070000022649765 3.240999937057495,0.09700000286102295 3.240999937057495,0 C3.25,-0.10599999874830246 3.255000114440918,-0.2029999941587448 3.255000114440918,-0.29100000858306885 C3.255000114440918,-0.8650000095367432 3.121999979019165,-1.3899999856948853 2.8580000400543213,-1.8660000562667847 C2.5929999351501465,-2.3420000076293945 2.2219998836517334,-2.7219998836517334 1.746000051498413,-3.003999948501587z M-1.8539999723434448,-0.621999979019165 C-1.7740000486373901,-1.1779999732971191 -1.5540000200271606,-1.593000054359436 -1.1920000314712524,-1.8660000562667847 C-0.8299999833106995,-2.1480000019073486 -0.41600000858306885,-2.2890000343322754 0.052000001072883606,-2.2890000343322754 C0.5550000071525574,-2.2890000343322754 0.9750000238418579,-2.1389999389648438 1.309999942779541,-1.8389999866485596 C1.6449999809265137,-1.5390000343322754 1.8259999752044678,-1.1339999437332153 1.8530000448226929,-0.621999979019165 C1.8530000448226929,-0.621999979019165 -1.8539999723434448,-0.621999979019165 -1.8539999723434448,-0.621999979019165z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,739.4810180664062,527.0549926757812)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0.5220000147819519,-3.3610000610351562 C0.010999999940395355,-3.3610000610351562 -0.4440000057220459,-3.242000102996826 -0.8410000205039978,-3.003999948501587 C-1.2289999723434448,-2.7750000953674316 -1.5240000486373901,-2.4570000171661377 -1.7269999980926514,-2.0510001182556152 C-1.7269999980926514,-2.0510001182556152 -1.8329999446868896,-3.203000068664551 -1.8329999446868896,-3.203000068664551 C-1.8329999446868896,-3.203000068664551 -3.0769999027252197,-3.203000068664551 -3.0769999027252197,-3.203000068664551 C-3.0769999027252197,-3.203000068664551 -3.0769999027252197,3.3610000610351562 -3.0769999027252197,3.3610000610351562 C-3.0769999027252197,3.3610000610351562 -1.6740000247955322,3.3610000610351562 -1.6740000247955322,3.3610000610351562 C-1.6740000247955322,3.3610000610351562 -1.6740000247955322,-0.027000000700354576 -1.6740000247955322,-0.027000000700354576 C-1.6740000247955322,-0.7149999737739563 -1.5069999694824219,-1.24399995803833 -1.1710000038146973,-1.6150000095367432 C-0.8270000219345093,-1.9850000143051147 -0.39500001072883606,-2.1700000762939453 0.125,-2.1700000762939453 C1.1660000085830688,-2.1700000762939453 1.687000036239624,-1.534999966621399 1.687000036239624,-0.26499998569488525 C1.687000036239624,-0.26499998569488525 1.687000036239624,3.3610000610351562 1.687000036239624,3.3610000610351562 C1.687000036239624,3.3610000610351562 3.0769999027252197,3.3610000610351562 3.0769999027252197,3.3610000610351562 C3.0769999027252197,3.3610000610351562 3.0769999027252197,-0.4099999964237213 3.0769999027252197,-0.4099999964237213 C3.0769999027252197,-1.3890000581741333 2.8469998836517334,-2.125999927520752 2.388000011444092,-2.619999885559082 C1.937999963760376,-3.114000082015991 1.315999984741211,-3.3610000610351562 0.5220000147819519,-3.3610000610351562z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,733.552978515625,527.0549926757812)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.5360000133514404,-1.9589999914169312 C-0.5360000133514404,-1.9589999914169312 -0.6549999713897705,-3.203000068664551 -0.6549999713897705,-3.203000068664551 C-0.6549999713897705,-3.203000068664551 -1.9119999408721924,-3.203000068664551 -1.9119999408721924,-3.203000068664551 C-1.9119999408721924,-3.203000068664551 -1.9119999408721924,3.3610000610351562 -1.9119999408721924,3.3610000610351562 C-1.9119999408721924,3.3610000610351562 -0.5099999904632568,3.3610000610351562 -0.5099999904632568,3.3610000610351562 C-0.5099999904632568,3.3610000610351562 -0.5099999904632568,0.21199999749660492 -0.5099999904632568,0.21199999749660492 C-0.5099999904632568,-0.32600000500679016 -0.421999990940094,-0.7450000047683716 -0.24500000476837158,-1.0449999570846558 C-0.05999999865889549,-1.3539999723434448 0.18400000035762787,-1.5700000524520874 0.48399999737739563,-1.694000005722046 C0.7919999957084656,-1.8259999752044678 1.1399999856948853,-1.8919999599456787 1.527999997138977,-1.8919999599456787 C1.527999997138977,-1.8919999599456787 1.9119999408721924,-1.8919999599456787 1.9119999408721924,-1.8919999599456787 C1.9119999408721924,-1.8919999599456787 1.9119999408721924,-3.3610000610351562 1.9119999408721924,-3.3610000610351562 C1.3300000429153442,-3.3610000610351562 0.8309999704360962,-3.2339999675750732 0.41600000858306885,-2.9779999256134033 C0.010999999940395355,-2.7309999465942383 -0.3070000112056732,-2.3910000324249268 -0.5360000133514404,-1.9589999914169312z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,708.208984375,525.7849731445312)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-2.944999933242798,4.631999969482422 C-2.944999933242798,4.631999969482422 2.944999933242798,4.631999969482422 2.944999933242798,4.631999969482422 C2.944999933242798,4.631999969482422 2.944999933242798,3.49399995803833 2.944999933242798,3.49399995803833 C2.944999933242798,3.49399995803833 -1.5420000553131104,3.49399995803833 -1.5420000553131104,3.49399995803833 C-1.5420000553131104,3.49399995803833 -1.5420000553131104,0.503000020980835 -1.5420000553131104,0.503000020980835 C-1.5420000553131104,0.503000020980835 2.5480000972747803,0.503000020980835 2.5480000972747803,0.503000020980835 C2.5480000972747803,0.503000020980835 2.5480000972747803,-0.609000027179718 2.5480000972747803,-0.609000027179718 C2.5480000972747803,-0.609000027179718 -1.5420000553131104,-0.609000027179718 -1.5420000553131104,-0.609000027179718 C-1.5420000553131104,-0.609000027179718 -1.5420000553131104,-3.49399995803833 -1.5420000553131104,-3.49399995803833 C-1.5420000553131104,-3.49399995803833 2.944999933242798,-3.49399995803833 2.944999933242798,-3.49399995803833 C2.944999933242798,-3.49399995803833 2.944999933242798,-4.631999969482422 2.944999933242798,-4.631999969482422 C2.944999933242798,-4.631999969482422 -2.944999933242798,-4.631999969482422 -2.944999933242798,-4.631999969482422 C-2.944999933242798,-4.631999969482422 -2.944999933242798,4.631999969482422 -2.944999933242798,4.631999969482422z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,715.1270141601562,527.135009765625)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.746999979019165,-3.2820000648498535 C1.746999979019165,-3.2820000648498535 0,-0.7680000066757202 0,-0.7680000066757202 C0,-0.7680000066757202 -1.7599999904632568,-3.2820000648498535 -1.7599999904632568,-3.2820000648498535 C-1.7599999904632568,-3.2820000648498535 -3.255000114440918,-3.2820000648498535 -3.255000114440918,-3.2820000648498535 C-3.255000114440918,-3.2820000648498535 -0.9269999861717224,0 -0.9269999861717224,0 C-0.9269999861717224,0 -3.255000114440918,3.2820000648498535 -3.255000114440918,3.2820000648498535 C-3.255000114440918,3.2820000648498535 -1.7599999904632568,3.2820000648498535 -1.7599999904632568,3.2820000648498535 C-1.7599999904632568,3.2820000648498535 0,0.7680000066757202 0,0.7680000066757202 C0,0.7680000066757202 1.746999979019165,3.2820000648498535 1.746999979019165,3.2820000648498535 C1.746999979019165,3.2820000648498535 3.255000114440918,3.2820000648498535 3.255000114440918,3.2820000648498535 C3.255000114440918,3.2820000648498535 0.9259999990463257,0 0.9259999990463257,0 C0.9259999990463257,0 3.255000114440918,-3.2820000648498535 3.255000114440918,-3.2820000648498535 C3.255000114440918,-3.2820000648498535 1.746999979019165,-3.2820000648498535 1.746999979019165,-3.2820000648498535z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,720.9739990234375,526.301025390625)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0.3370000123977661,-4.116000175476074 C0.3370000123977661,-4.116000175476074 -0.8930000066757202,-4.116000175476074 -0.8930000066757202,-4.116000175476074 C-0.8930000066757202,-4.116000175476074 -1.065000057220459,-2.447999954223633 -1.065000057220459,-2.447999954223633 C-1.065000057220459,-2.447999954223633 -2.190000057220459,-2.447999954223633 -2.190000057220459,-2.447999954223633 C-2.190000057220459,-2.447999954223633 -2.190000057220459,-1.2699999809265137 -2.190000057220459,-1.2699999809265137 C-2.190000057220459,-1.2699999809265137 -1.065000057220459,-1.2699999809265137 -1.065000057220459,-1.2699999809265137 C-1.065000057220459,-1.2699999809265137 -1.065000057220459,1.972000002861023 -1.065000057220459,1.972000002861023 C-1.065000057220459,2.7750000953674316 -0.8759999871253967,3.3340001106262207 -0.4959999918937683,3.6519999504089355 C-0.11699999868869781,3.9609999656677246 0.39500001072883606,4.116000175476074 1.0390000343322754,4.116000175476074 C1.0390000343322754,4.116000175476074 2.124000072479248,4.116000175476074 2.124000072479248,4.116000175476074 C2.124000072479248,4.116000175476074 2.124000072479248,2.924999952316284 2.124000072479248,2.924999952316284 C2.124000072479248,2.924999952316284 1.3700000047683716,2.924999952316284 1.3700000047683716,2.924999952316284 C0.9900000095367432,2.924999952316284 0.7210000157356262,2.8580000400543213 0.5619999766349792,2.7260000705718994 C0.41200000047683716,2.5850000381469727 0.3370000123977661,2.3340001106262207 0.3370000123977661,1.972000002861023 C0.3370000123977661,1.972000002861023 0.3370000123977661,-1.2699999809265137 0.3370000123977661,-1.2699999809265137 C0.3370000123977661,-1.2699999809265137 2.190000057220459,-1.2699999809265137 2.190000057220459,-1.2699999809265137 C2.190000057220459,-1.2699999809265137 2.190000057220459,-2.447999954223633 2.190000057220459,-2.447999954223633 C2.190000057220459,-2.447999954223633 0.3370000123977661,-2.447999954223633 0.3370000123977661,-2.447999954223633 C0.3370000123977661,-2.447999954223633 0.3370000123977661,-4.116000175476074 0.3370000123977661,-4.116000175476074z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,777.0139770507812,525.6530151367188)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.7020000219345093,4.763999938964844 C-0.7020000219345093,4.763999938964844 0.7020000219345093,4.763999938964844 0.7020000219345093,4.763999938964844 C0.7020000219345093,4.763999938964844 0.7020000219345093,-4.763999938964844 0.7020000219345093,-4.763999938964844 C0.7020000219345093,-4.763999938964844 -0.7020000219345093,-4.763999938964844 -0.7020000219345093,-4.763999938964844 C-0.7020000219345093,-4.763999938964844 -0.7020000219345093,4.763999938964844 -0.7020000219345093,4.763999938964844z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,785.1589965820312,527.135009765625)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.746000051498413,-3.003999948501587 C1.2699999809265137,-3.2950000762939453 0.7099999785423279,-3.440999984741211 0.06599999964237213,-3.440999984741211 C-0.5950000286102295,-3.440999984741211 -1.1790000200271606,-3.2950000762939453 -1.680999994277954,-3.003999948501587 C-2.1760001182556152,-2.7130000591278076 -2.563999891281128,-2.306999921798706 -2.8459999561309814,-1.7860000133514404 C-3.119999885559082,-1.2649999856948853 -3.255000114440918,-0.6620000004768372 -3.255000114440918,0.026000000536441803 C-3.255000114440918,0.7049999833106995 -3.115000009536743,1.3009999990463257 -2.8329999446868896,1.812999963760376 C-2.549999952316284,2.325000047683716 -2.1610000133514404,2.7260000705718994 -1.6670000553131104,3.0169999599456787 C-1.1640000343322754,3.2990000247955322 -0.5920000076293945,3.440999984741211 0.052000001072883606,3.440999984741211 C0.828000009059906,3.440999984741211 1.4769999980926514,3.2339999675750732 1.996999979019165,2.819000005722046 C2.5269999504089355,2.4040000438690186 2.88100004196167,1.8880000114440918 3.056999921798706,1.2699999809265137 C3.056999921798706,1.2699999809265137 1.652999997138977,1.2699999809265137 1.652999997138977,1.2699999809265137 C1.5210000276565552,1.5789999961853027 1.319000005722046,1.8259999752044678 1.0460000038146973,2.010999917984009 C0.781000018119812,2.187999963760376 0.4490000009536743,2.2760000228881836 0.052000001072883606,2.2760000228881836 C-0.44200000166893005,2.2760000228881836 -0.878000020980835,2.111999988555908 -1.2569999694824219,1.7860000133514404 C-1.628000020980835,1.4509999752044678 -1.8359999656677246,0.9660000205039978 -1.8799999952316284,0.3310000002384186 C-1.8799999952316284,0.3310000002384186 3.2290000915527344,0.3310000002384186 3.2290000915527344,0.3310000002384186 C3.23799991607666,0.2070000022649765 3.242000102996826,0.09700000286102295 3.242000102996826,0 C3.250999927520752,-0.10599999874830246 3.255000114440918,-0.2029999941587448 3.255000114440918,-0.29100000858306885 C3.255000114440918,-0.8650000095367432 3.122999906539917,-1.3899999856948853 2.8589999675750732,-1.8660000562667847 C2.5940001010894775,-2.3420000076293945 2.2219998836517334,-2.7219998836517334 1.746000051498413,-3.003999948501587z M-1.8539999723434448,-0.621999979019165 C-1.7740000486373901,-1.1779999732971191 -1.5540000200271606,-1.593000054359436 -1.1920000314712524,-1.8660000562667847 C-0.8299999833106995,-2.1480000019073486 -0.41499999165534973,-2.2890000343322754 0.052000001072883606,-2.2890000343322754 C0.5550000071525574,-2.2890000343322754 0.9739999771118164,-2.1389999389648438 1.309000015258789,-1.8389999866485596 C1.6449999809265137,-1.5390000343322754 1.8250000476837158,-1.1339999437332153 1.8519999980926514,-0.621999979019165 C1.8519999980926514,-0.621999979019165 -1.8539999723434448,-0.621999979019165 -1.8539999723434448,-0.621999979019165z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,779.927978515625,525.6530151367188)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.7020000219345093,4.763999938964844 C-0.7020000219345093,4.763999938964844 0.7020000219345093,4.763999938964844 0.7020000219345093,4.763999938964844 C0.7020000219345093,4.763999938964844 0.7020000219345093,-4.763999938964844 0.7020000219345093,-4.763999938964844 C0.7020000219345093,-4.763999938964844 -0.7020000219345093,-4.763999938964844 -0.7020000219345093,-4.763999938964844 C-0.7020000219345093,-4.763999938964844 -0.7020000219345093,4.763999938964844 -0.7020000219345093,4.763999938964844z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,791.1510009765625,526.301025390625)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M2.190000057220459,-1.2699999809265137 C2.190000057220459,-1.2699999809265137 2.190000057220459,-2.447999954223633 2.190000057220459,-2.447999954223633 C2.190000057220459,-2.447999954223633 0.33799999952316284,-2.447999954223633 0.33799999952316284,-2.447999954223633 C0.33799999952316284,-2.447999954223633 0.33799999952316284,-4.116000175476074 0.33799999952316284,-4.116000175476074 C0.33799999952316284,-4.116000175476074 -0.8930000066757202,-4.116000175476074 -0.8930000066757202,-4.116000175476074 C-0.8930000066757202,-4.116000175476074 -1.065000057220459,-2.447999954223633 -1.065000057220459,-2.447999954223633 C-1.065000057220459,-2.447999954223633 -2.190000057220459,-2.447999954223633 -2.190000057220459,-2.447999954223633 C-2.190000057220459,-2.447999954223633 -2.190000057220459,-1.2699999809265137 -2.190000057220459,-1.2699999809265137 C-2.190000057220459,-1.2699999809265137 -1.065000057220459,-1.2699999809265137 -1.065000057220459,-1.2699999809265137 C-1.065000057220459,-1.2699999809265137 -1.065000057220459,1.972000002861023 -1.065000057220459,1.972000002861023 C-1.065000057220459,2.7750000953674316 -0.875,3.3340001106262207 -0.4959999918937683,3.6519999504089355 C-0.11699999868869781,3.9609999656677246 0.39500001072883606,4.116000175476074 1.0390000343322754,4.116000175476074 C1.0390000343322754,4.116000175476074 2.124000072479248,4.116000175476074 2.124000072479248,4.116000175476074 C2.124000072479248,4.116000175476074 2.124000072479248,2.924999952316284 2.124000072479248,2.924999952316284 C2.124000072479248,2.924999952316284 1.3700000047683716,2.924999952316284 1.3700000047683716,2.924999952316284 C0.9909999966621399,2.924999952316284 0.7210000157356262,2.8580000400543213 0.5630000233650208,2.7260000705718994 C0.4129999876022339,2.5850000381469727 0.33799999952316284,2.3340001106262207 0.33799999952316284,1.972000002861023 C0.33799999952316284,1.972000002861023 0.33799999952316284,-1.2699999809265137 0.33799999952316284,-1.2699999809265137 C0.33799999952316284,-1.2699999809265137 2.190000057220459,-1.2699999809265137 2.190000057220459,-1.2699999809265137z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,751.7730102539062,525.6530151367188)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.7020000219345093,4.763999938964844 C-0.7020000219345093,4.763999938964844 0.7020000219345093,4.763999938964844 0.7020000219345093,4.763999938964844 C0.7020000219345093,4.763999938964844 0.7020000219345093,-4.763999938964844 0.7020000219345093,-4.763999938964844 C0.7020000219345093,-4.763999938964844 -0.7020000219345093,-4.763999938964844 -0.7020000219345093,-4.763999938964844 C-0.7020000219345093,-4.763999938964844 -0.7020000219345093,4.763999938964844 -0.7020000219345093,4.763999938964844z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,746.7109985351562,527.135009765625)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0.10599999874830246,-3.440999984741211 C-0.4059999883174896,-3.440999984741211 -0.878000020980835,-3.3480000495910645 -1.309999942779541,-3.1630001068115234 C-1.7339999675750732,-2.986999988555908 -2.0869998931884766,-2.7260000705718994 -2.36899995803833,-2.381999969482422 C-2.6429998874664307,-2.046999931335449 -2.8010001182556152,-1.6410000324249268 -2.8450000286102295,-1.1640000343322754 C-2.8450000286102295,-1.1640000343322754 -1.4429999589920044,-1.1640000343322754 -1.4429999589920044,-1.1640000343322754 C-1.38100004196167,-1.534999966621399 -1.2089999914169312,-1.812999963760376 -0.9269999861717224,-1.9980000257492065 C-0.6349999904632568,-2.191999912261963 -0.29600000381469727,-2.2890000343322754 0.09200000017881393,-2.2890000343322754 C0.5339999794960022,-2.2890000343322754 0.890999972820282,-2.1619999408721924 1.1640000343322754,-1.906000018119812 C1.4470000267028809,-1.649999976158142 1.5880000591278076,-1.2660000324249268 1.5880000591278076,-0.7540000081062317 C1.5880000591278076,-0.7540000081062317 1.5880000591278076,-0.6349999904632568 1.5880000591278076,-0.6349999904632568 C1.5880000591278076,-0.6349999904632568 -0.13300000131130219,-0.6349999904632568 -0.13300000131130219,-0.6349999904632568 C-1.0499999523162842,-0.6349999904632568 -1.75600004196167,-0.4490000009536743 -2.25,-0.07900000363588333 C-2.74399995803833,0.29100000858306885 -2.990999937057495,0.7940000295639038 -2.990999937057495,1.4290000200271606 C-2.990999937057495,1.7910000085830688 -2.9030001163482666,2.125999927520752 -2.7260000705718994,2.434999942779541 C-2.549999952316284,2.734999895095825 -2.2809998989105225,2.9779999256134033 -1.9190000295639038,3.1630001068115234 C-1.5570000410079956,3.3480000495910645 -1.0989999771118164,3.440999984741211 -0.5429999828338623,3.440999984741211 C0.03099999949336052,3.440999984741211 0.49399998784065247,3.316999912261963 0.847000002861023,3.069999933242798 C1.2000000476837158,2.822999954223633 1.468999981880188,2.5190000534057617 1.6540000438690186,2.1570000648498535 C1.6540000438690186,2.1570000648498535 1.7730000019073486,3.2820000648498535 1.7730000019073486,3.2820000648498535 C1.7730000019073486,3.2820000648498535 2.990999937057495,3.2820000648498535 2.990999937057495,3.2820000648498535 C2.990999937057495,3.2820000648498535 2.990999937057495,-0.7540000081062317 2.990999937057495,-0.7540000081062317 C2.990999937057495,-1.5829999446868896 2.739000082015991,-2.236999988555908 2.2360000610351562,-2.7130000591278076 C1.7330000400543213,-3.197999954223633 1.0230000019073486,-3.440999984741211 0.10599999874830246,-3.440999984741211z M1.2970000505447388,1.309999942779541 C1.156000018119812,1.6100000143051147 0.953000009059906,1.8530000448226929 0.6880000233650208,2.0380001068115234 C0.4320000112056732,2.2139999866485596 0.11400000005960464,2.302999973297119 -0.26499998569488525,2.302999973297119 C-0.652999997138977,2.302999973297119 -0.9620000123977661,2.2190001010894775 -1.190999984741211,2.0510001182556152 C-1.4119999408721924,1.875 -1.5219999551773071,1.6369999647140503 -1.5219999551773071,1.3370000123977661 C-1.5219999551773071,1.0460000038146973 -1.406999945640564,0.8029999732971191 -1.1779999732971191,0.609000027179718 C-0.9399999976158142,0.41499999165534973 -0.5429999828338623,0.3179999887943268 0.013000000268220901,0.3179999887943268 C0.013000000268220901,0.3179999887943268 1.5750000476837158,0.3179999887943268 1.5750000476837158,0.3179999887943268 C1.5390000343322754,0.6800000071525574 1.4470000267028809,1.0099999904632568 1.2970000505447388,1.309999942779541z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,762.7069702148438,525.7919921875)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M2.8589999675750732,3.0369999408721924 C2.8589999675750732,3.0369999408721924 0.8069999814033508,-4.638000011444092 0.8069999814033508,-4.638000011444092 C0.8069999814033508,-4.638000011444092 -0.7409999966621399,-4.638000011444092 -0.7409999966621399,-4.638000011444092 C-0.7409999966621399,-4.638000011444092 -2.8580000400543213,3.0369999408721924 -2.8580000400543213,3.0369999408721924 C-2.8580000400543213,3.0369999408721924 -4.697999954223633,-4.638000011444092 -4.697999954223633,-4.638000011444092 C-4.697999954223633,-4.638000011444092 -6.206999778747559,-4.638000011444092 -6.206999778747559,-4.638000011444092 C-6.206999778747559,-4.638000011444092 -3.7060000896453857,4.638000011444092 -3.7060000896453857,4.638000011444092 C-3.7060000896453857,4.638000011444092 -2.0920000076293945,4.625 -2.0920000076293945,4.625 C-2.0920000076293945,4.625 0,-2.640000104904175 0,-2.640000104904175 C0,-2.640000104904175 2.0250000953674316,4.625 2.0250000953674316,4.625 C2.0250000953674316,4.625 3.6659998893737793,4.625 3.6659998893737793,4.625 C3.6659998893737793,4.625 6.206999778747559,-4.638000011444092 6.206999778747559,-4.638000011444092 C6.206999778747559,-4.638000011444092 4.697999954223633,-4.638000011444092 4.697999954223633,-4.638000011444092 C4.697999954223633,-4.638000011444092 2.8589999675750732,3.0369999408721924 2.8589999675750732,3.0369999408721924z"></path></g><g opacity="0.3" transform="matrix(1,0,0,1,771.9509887695312,527.135009765625)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0.10599999874830246,-3.440999984741211 C-0.4059999883174896,-3.440999984741211 -0.878000020980835,-3.3480000495910645 -1.309999942779541,-3.1630001068115234 C-1.7330000400543213,-2.986999988555908 -2.0859999656677246,-2.7260000705718994 -2.36899995803833,-2.381999969482422 C-2.6419999599456787,-2.046999931335449 -2.8010001182556152,-1.6410000324249268 -2.8450000286102295,-1.1640000343322754 C-2.8450000286102295,-1.1640000343322754 -1.4420000314712524,-1.1640000343322754 -1.4420000314712524,-1.1640000343322754 C-1.3799999952316284,-1.534999966621399 -1.2079999446868896,-1.812999963760376 -0.9259999990463257,-1.9980000257492065 C-0.6349999904632568,-2.191999912261963 -0.29499998688697815,-2.2890000343322754 0.09300000220537186,-2.2890000343322754 C0.5339999794960022,-2.2890000343322754 0.890999972820282,-2.1619999408721924 1.1649999618530273,-1.906000018119812 C1.4470000267028809,-1.649999976158142 1.5880000591278076,-1.2660000324249268 1.5880000591278076,-0.7540000081062317 C1.5880000591278076,-0.7540000081062317 1.5880000591278076,-0.6349999904632568 1.5880000591278076,-0.6349999904632568 C1.5880000591278076,-0.6349999904632568 -0.13199999928474426,-0.6349999904632568 -0.13199999928474426,-0.6349999904632568 C-1.0499999523162842,-0.6349999904632568 -1.7549999952316284,-0.4490000009536743 -2.249000072479248,-0.07900000363588333 C-2.74399995803833,0.29100000858306885 -2.990999937057495,0.7940000295639038 -2.990999937057495,1.4290000200271606 C-2.990999937057495,1.7910000085830688 -2.9019999504089355,2.125999927520752 -2.7260000705718994,2.434999942779541 C-2.5490000247955322,2.734999895095825 -2.2799999713897705,2.9779999256134033 -1.9190000295639038,3.1630001068115234 C-1.5570000410079956,3.3480000495910645 -1.0980000495910645,3.440999984741211 -0.5419999957084656,3.440999984741211 C0.03099999949336052,3.440999984741211 0.49399998784065247,3.316999912261963 0.847000002861023,3.069999933242798 C1.2000000476837158,2.822999954223633 1.468999981880188,2.5190000534057617 1.6540000438690186,2.1570000648498535 C1.6540000438690186,2.1570000648498535 1.7740000486373901,3.2820000648498535 1.7740000486373901,3.2820000648498535 C1.7740000486373901,3.2820000648498535 2.990999937057495,3.2820000648498535 2.990999937057495,3.2820000648498535 C2.990999937057495,3.2820000648498535 2.990999937057495,-0.7540000081062317 2.990999937057495,-0.7540000081062317 C2.990999937057495,-1.5829999446868896 2.740000009536743,-2.236999988555908 2.236999988555908,-2.7130000591278076 C1.7339999675750732,-3.197999954223633 1.0240000486373901,-3.440999984741211 0.10599999874830246,-3.440999984741211z M1.2970000505447388,1.309999942779541 C1.156000018119812,1.6100000143051147 0.953000009059906,1.8530000448226929 0.6880000233650208,2.0380001068115234 C0.43299999833106995,2.2139999866485596 0.11500000208616257,2.302999973297119 -0.2639999985694885,2.302999973297119 C-0.652999997138977,2.302999973297119 -0.9610000252723694,2.2190001010894775 -1.190999984741211,2.0510001182556152 C-1.4110000133514404,1.875 -1.5219999551773071,1.6369999647140503 -1.5219999551773071,1.3370000123977661 C-1.5219999551773071,1.0460000038146973 -1.406999945640564,0.8029999732971191 -1.1779999732971191,0.609000027179718 C-0.9390000104904175,0.41499999165534973 -0.5419999957084656,0.3179999887943268 0.013000000268220901,0.3179999887943268 C0.013000000268220901,0.3179999887943268 1.5750000476837158,0.3179999887943268 1.5750000476837158,0.3179999887943268 C1.5399999618530273,0.6800000071525574 1.4470000267028809,1.0099999904632568 1.2970000505447388,1.309999942779541z"></path></g></g><g opacity="1" transform="matrix(1,0,0,1,0,0)"><g opacity="1" transform="matrix(1,0,0,1,749.2109985351562,563.1920166015625)"><path fill="url(#__lottie_element_767)" fill-opacity="1" d=" M119.43599700927734,-22.22100067138672 C119.43599700927734,-22.22100067138672 -119.43599700927734,-22.22100067138672 -119.43599700927734,-22.22100067138672 C-131.70799255371094,-22.22100067138672 -141.65699768066406,-12.272000312805176 -141.65699768066406,0 C-141.65699768066406,12.272000312805176 -131.70799255371094,22.22100067138672 -119.43599700927734,22.22100067138672 C-119.43599700927734,22.22100067138672 119.43599700927734,22.22100067138672 119.43599700927734,22.22100067138672 C131.70799255371094,22.22100067138672 141.65699768066406,12.272000312805176 141.65699768066406,0 C141.65699768066406,-12.272000312805176 131.70799255371094,-22.22100067138672 119.43599700927734,-22.22100067138672z"></path></g></g></g><g class="png" clip-path="url(#__lottie_element_769)" transform="matrix(0.6659970283508301,0,0,0.6659970283508301,337.3980407714844,424.38446044921875)" opacity="0.00004887166292562029" style="display: none;"><image width="194px" height="158px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAMIAAACeBAMAAACSg8iOAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAtUExURUdwTP///////////////////////////////////////////////////////81e3QIAAAAOdFJOUwAgTT8RfN/DMKRgCe+QbDiZxwAACExJREFUaN7tm/9rG/cZx5/Tnb46DifHcxwCQjp5Q12GUZQ0BAJGsb2OEs9ooyu0FOGmq9eUcaiUJSNsh61mbKxgHCjLBEEYlm1lDCMn+2GQX1yWNhi6I4PBxn4QdWTdWeb0/A17ns+dZUeWE2WDsbF7E9+Xz/O5z+vzeZ7Pc4LnCICv/08pcdLTu3yQB7jo/a3lRdOZ2jG1X0JEI+Wf2iVH5qEsnZMA5QXRpNXK/RMy09PTNXEpnT6cMEmja2N0rXYR7i08k5DqXEqpwwmBMZC0DAxotS7CZLYvgjJMHj55LEMBubhYAym/psYHRk8CDIwOuwTqFkmlQaF/3CQICvWQyqeH+yEMUCRycQ7IPU1L0YS1bG6SwzOkaRcEQcqAnNTIp1DWtKwgDOQ0TQ1w/2fFIR4fpkBK6QFaQ618di2nBtIZNZeZLicl7eRUWhAGNPJHWZXHpPTJYylBkNPTx1L9rIFmniHO3Qu8HCVdo6AG6JhbgLtjkfOgaIJAhyF1aOFuMnKaXcUEigBdDWX72UuXgFasMkFKx+NDpwJjwvlyCgbica0mCGU1l7+b5FHjcZdQXmTjULbPvTSZdqPJ2XEqcMojsKs9wtBCGuRTQ8L7HoH79k/IUVh5DZn79+/XOoRI+pLiEQJjGdrOubycqXleKn9IfaFfgpSh9Ys40LZVOwSZs8AlyJkxsqY5MSKdOAzE+4zDNExeoH0opach96GS20fI1CY9QkSjRtoTgczAkEsg41QKJlOX+nkvXaJBcuzi2j3eWR2CQrs95xIUygIo85q0VHovH0B+Zj5Io6Q/cDKrsDYKMHqeAkJvhDil9jD88bwa50tKiDi/XU+KDnFYBLpd++AGGeLD/g+UL1++fPny5cuXL1++fPny5cuXL1++fP0PS3neMq/0D3GaL/S0nnun2xI0cEN9LkIUF/iE7jjK/DLA1QKd3fuJxq5ld0IGIjafNeszT8zJUSPnOwTQ10HGEv1luwncC+AlfH/0957xUAXxiduzEGrtEeoNsm/BIOa7CWGbj1WaP08BJLfDRXfS+U6AFPUAATyCJLy72oQw7ojhuMEj8AiCoOAcL/QLOGFaN2Hwe8exDYH5n5gtFWQDr9PgOr4P38f5JDn6EYy8BYNXgvMB3ZkHfN20kvTwbAtm7TbMtuGnaBU8Ag1XkOkxFQK4zBNXI6ZlWmrU0hELMaTYlKDoXKWuVdRxhUK1AmCs83xD7SDGqAHIgNu8HgfGf2DRyJL5UG+7BEl/aNgy9VJp/a7PQlY2Zpai+J6kr8fwLcnYlohd3IrhXyVj3fVScRsMG5a26U54qZ0vtql5ENX6X1AtbkVpik5eEEKWGsEV4aUgT46d2eARosSsb8aobWIn6qhwpBmyyLPbLiHRBHTyxbldwrI4QQwLRtYsGKXY63StCgIdwCg9QaiuAyw1ozTeKhFUSOyEnUrldivR3NtLYTvQMrLG8i6hsBvKmw5Ul/ERSL9+DbOCUGxVKsa6MMfIAHDsrDHHc9xPCJEXsTWxs0cIOi/vkD+yXQTQ37Vh9U+YFanlEqr88BfCLOE6r2Dz4BpCNn/R5zVIwy5BxnObE9ct6CbU7R3aUJgPWakveYT6Nj2rumaDh8C5IjludXs/gYIGgV8doU6JhpcPplGaabUOEFZxkzLC5l0U8wgc1p8nXfMS/lJ6FVdmLFXW5/YTZPJfYof2mlKl2Iqkr2MhjI8Fwd5HmKWcGqQ8WnpMbwiX8JKtKnqJJ0mBMMV7KYC2wfmwR4CidQvnFMP6Ea7InEq0qXCB3w9EGMTP1Q4hRLtKxgYNXzE9gowPbztqED/jmZ0w0abzcXRuwhOEmI4befimSTkNRZE2MxZwlhBBqeIeYRAp4c0tiOh43SPQcPhD7iXWLp0Wb6SpAz8TijBEFvky3s+PSuTG3vXUjV6P+fLly5cvX758+fLly5cvX758/aek5A+2hd7uXI68/RxjfeUygNxdLP+6ySXXLh3l0t/I/Py1FUg0n4OwusNlticJko7Y1bRLSJDFUv9tQhTvyPqD3gT7zAtY+pcIbuHoovD/0RbARHN/AZwtLoFMxhYROnWmeOe5Qz5oeATJwHeBC9grPEyb2k5DRLTFDHyPS2luCZYJ9c1E8w28DOErxxvKG9hS6Tk76dXLDyWMOFcxG8CHXMKEI84dNn3NuUaLK9pkkXRHN3YJ1c2EheioIVtvhPEafpcL3w2vXn4oofgAjNJIKy+qk0HEa9+ieH8ExUaMq+xzUVyRdI8gmXMJfCWIhRD++c74Do0waOW/0Xbr5U8hfAIX1dVNAP0RNb5KO+YRFxbD7bAovCa8yEDCqhi4wrfmcog6vFiA2WbUyitxrpfPNA8nTOCPx6C6UanoJW598W/YjDqVyi/so1alUm2ON/bt1ssiH/QSF19h8jvFZgw3vg1uvbwXQXxLKQRNtJJcv8aS2z5uhfnOmuVjk2vBIXe3/i4F+wiDompbRHzHrZf3IIy3RZ0fpl7DB9U3uWoN8MJHAGEranEB/GiLDsOrj3fXIMbYI9Sbi3SjfNmwRb28136dtTnDsl/NwvgmB4ouxLpCdhC5AM5xiH281NyNQxfBXKY4BD6mzeHWy3sQwvh3qW5B/QGsbs3QRuFIzzhJpd5U6OmRFn9sKTbCTrazl7oJxSbNIohuvbwHQbyDPoEl65a5HMQrty2Vvebo/EHIqphvQtW+hY9ktA2jp5fsW2ZTxitG262X90q5EyZ/W6SE3MjDb5HzlzMYP6U1G2ir/FHrU4Bz2JrtRXgZLfLeb9B5xauX91JE/Kc+t9i9WwTf3+Ze/6z3wzAl3lriObe7r/8q/RN/suiwXqHjUwAAAABJRU5ErkJggg=="></image></g><g transform="matrix(1.0017281770706177,0,0,1.0017281770706177,-485.9598388671875,107.65113830566406)" opacity="0.0000017916765349923482" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0.843994140625,0.050994873046875)"><path stroke="url(#__lottie_element_469)" stroke-linecap="butt" stroke-linejoin="miter" fill-opacity="0" stroke-miterlimit="4" stroke-opacity="1" stroke-width="3" d=" M850.1690063476562,29.8799991607666 C850.1690063476562,29.8799991607666 1154.64501953125,29.8799991607666 1154.64501953125,29.8799991607666 C1167.2099609375,29.8799991607666 1177.39501953125,40.066001892089844 1177.39501953125,52.630001068115234 C1177.39501953125,52.630001068115234 1177.39501953125,465.364990234375 1177.39501953125,465.364990234375 C1177.39501953125,477.9289855957031 1167.2099609375,488.114990234375 1154.64501953125,488.114990234375 C1154.64501953125,488.114990234375 850.1690063476562,488.114990234375 850.1690063476562,488.114990234375 C837.60498046875,488.114990234375 827.4190063476562,477.9289855957031 827.4190063476562,465.364990234375 C827.4190063476562,465.364990234375 827.4190063476562,52.630001068115234 827.4190063476562,52.630001068115234 C827.4190063476562,40.066001892089844 837.60498046875,29.8799991607666 850.1690063476562,29.8799991607666z"></path></g></g><g mask="url(#__lottie_element_215)" style="display: block;"><g class="png" transform="matrix(0.667044997215271,0,0,0.667044997215271,249.3896484375,108.404541015625)" opacity="0.9999973289049107"><image width="707px" height="774px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAsMAAAMGBAMAAADl8JTCAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAbUExURQABIAABIAABIAABIA0WPhYgSQECISARQwoMMPPhF08AAAAEdFJOU4636dRwBhFvAAAgAElEQVR42uxbbZLrqhG92UFS2UFS9RaQbAEKLUAq/38/VCxAqvICVFyWHfqc7gbNeO6X7XnJi/FYQtB004dDCyHPly+v9Pz0t1d6YvqrIPzvV3pm+vLlL3//xys9Mf2zQfyv337La1n5zeuKb5G/wuLsdVaPc6GkCfIjB2mCb8YJud+ZQX4tmi8mypxJuJCL6tmUjK1Xa1n6BSs/xT6BeO+/A7auAvH6Sk9Mv78gfnp6QfyC+AXxK70gfkH8gviVXhC/IEbK46n4xan4Zit9xHQxv/ASbdwryg1999kf0tn+fwPE9iS5+qO59TufOo1nTn+EZzs6k9XZPPgmD7C5I1xyRy0rcCVbowfYL286kMf2fyjEvRudbSwrWlpGltB3Q2LthOu0zCdOl3U9u/2W2a7hPvtOeA5DUYmSf5jRD4fY+aA4FO18sbKxf+x1yadJXpxJug/VN6J066n0WavmbL/KCVcGLO6yn3vA6GNd3gahT2ZxZ8mabR+quA9ZO8gNMaUStrIGnHLfzSsWHIhAzqyl/6WcMHbCFZ3N5SH2i1HFAx/H/geJ/GiIswVP9mF1L4rjAWiKwaRjMmym5ryOHkLYI68Bgc1LC8DZ0cvZxrXbvtO+T4o1+z5tMcafbxOfATGczta/wj1Vbr8qgWzekor0NhtKRW9lhdNcibauuv0r8nkgXNYLbTZM7GK7vev99tnafbF7IMz/IbG4ZGexTemiPbeN7eJb5UVHo1ih7qWzrTbK+bT5zgmqm+h9t7xrFvnsAbyUe+275GqjVYpNIl+2fBrEjL86F40k2R1lx+0tQ/fIXzMUE6cblc7k1cjmGu2dir+06KQzLN3k0IOft2+rQsaZooHajfzI3e6hEBed0v1ljb3icT4Mpe08wN/hcjLmPI6OXlQjbB+8EdHiWPhLJYPxl+znXPNIcNWUO4u1yadAnP1eV05vxzpcdZilt9Nt9vXKHgwISH0Hr0YKS1VDxy/ZX/t8Weup3vw5LSU/h8VDZJRsPbnbKmTqZXF8Lae5af2vo6/u2A0URjGf5nXEx3AoA+t/zn6p7+xWcLjYjbS/J/7WCvlhEHPlY+EuWw/b3KqVUNdc9V4E7NwHA8fL7NgObAspufJsOUGg+osVt1ajcii51359Mwo+grrE+RQW6+1mYM/Q36ocI1Cal0v+ASHIZR4lA3eqtq2Zhe50oZ7Ks6AOzZnxmnahBTH0F+yTtHrJGekSZ36vvC8/F+LSn7ewxoLTzh3tvQBEVMBsAlJHDyxblKhVASB0LITuajV02iDSoaA116wg/qx970f1/pTO+B6Tvrc+fiCLh+USYwScIsKStc7LRekEYgHoSZZmSku+OHM53S2vzaEYNWxUtEi5zmymol+xrw7oWFdK9UHhdPE13zMhzr7H6DeF0UtjEIroj2a9QBGorMoOG1PudCSZizXMymc0UrDA1ZyJhiKTf8U+DYHspLfFGgxAHQKFPXuXJ7LYnjn63aPqbacOcOm8V6+LkXvAsjghnf2lnvBwTBQSbVPMglJ2sKw3rZ+0r+NYbPoMCi146YAjNGbb13teoBiWSbm7WeqPpneS+VRYamfot1qdNJQfFPxYYOyAmh606v0324bnR+uKR0BcjMOcxMWIUv9kaYhQJL+uQortV+Wnrij6Y1z5Gfb+z4OuC5o8vO16PMSFu7h6p6t/Rvp+GIt0eVNthzXfisaPCRTrsCVQ/58A1vWgbys/L1BwG4D3gjPG0yF/ddKMnHHEaZISVOB7mJBVHQeFeX24oNVMdlTVdRr0QIXkDjf1sf3BJsUOb6nZj0MFH0Ry/uiO9wCI9Sf2RdfxDu5EL0cUDLLD+j1N9BHjMGLMNlWbTK2y9lppgKYigBpHHZj4wBDHenjjj+zTpo+CjRe44XAj1Xdw+9ot55vB+BGxWLd+Rv5aB5Vt4JvnLR2dm29rFYtgDDxMhyF1hFpDDVP7Hm8TJJWicgxK+O/aP1V6uWqqRx9QqBqWh0U3idZbwfgBLC72DsyfoNS5AG+bhy2rHQ8jENOIi0JFDEKXnULHhSCEk/ygcRzLQTIczNRv258G+ye4PaKoQwcCXx1BVh7ffry7G2K+etEtqE5h96i75/55Xs9EtcKBMMJg40LfwuHo9xR8LE1nsMPRTdbp+/YP0xdkVLr90N2op9k4gGxbnTf3Ne+FmHOj5L6UOBEJnZtCCAc+Ez5HQLel/JisimSDe4GoQkYKJ63yg/yhCScIjnaN4+F61d4Rvmt/olbkre+cAIpwH0PNgMy2PaWL4xsP0XezmL/00A0fSTav4ZegO7Hj3mdAYN7IcRoQJmqALQCFg0AozJM7ffh4UAT4sc1Emx1hNAjftk85s6/DpQPtCIfjFK2nHpL1CWR9Sizm7o8tJixAtB5+RfevIbVc+BpSCFuYNinZwoZjCl+Pjd/2CU1SBKQUsnItQ9RUtOJrU9AqNqn62hTIUWSDHltdCiiVS2mkdfji4lv2RXsIV7Tp0+dE9kkHKCjkurrz57xsv/l6KMT2z4/ZEK7VAqL0ZUOv0fkEnmyh+dNKmqutVDoMEBMOAuhGN67i2CZyqTWQmiugSq16E7kpHtQ5CfwHxrCN6dZUibSMFOC4trqrwE+yfmz/QBG7Qbg5AYmxhRObPgq+Lr5t2zTnW7+PfcCirRR/dG7RiWNNjmna9CTOJS/ZAonN2oQPyiHSjteg0pamYM02lW7fyMJtFEx2aiapztV/ZJ/Hq4lsQacQ0zGeLarZekMjctYfj7wLFffHYqyK+62uSpi4kgbAgHgQN+3+pqXB0NySgyO+pmRjg/ZAR/4SQEnhjD21qcqNWrctOepA2mxpj0b71qgdYxjHqs3BjaPjpX6rDcHWMdM03PJu/Sj27kDhvzNQjBm3ZDInI8TcPiFFnGMrakflYXAgkoLZ5GcyKcWRbZJmtglO9i2mqP5vVNoOqcltAhZwnxvN20eUuexoP6ETM7uhaHpXwsD6dOL0ZIsTPiL5o3R5wtNdHnbYKp+YMMXmjdRNMSZAEWbxpMHXrsXveZZvK4nbJUYgNBOL5l+raCVJr5toEpGEVi01BdKaVORlq2yqW34T/VGGZwawok5QTk0lNL6xH6LEGlGbIrUljAKQ32zmjfFnwBoo29KNv894/xbvLoiz/eJU34ZJmJARRl/EBTgpeDXXG4lSAjjwMYFVDTcQT9xtgyJ+Ayfh2YXt5WomdMKmGSFgFq43xTuBxvC1URCFMo4CuOAedowIygNHVPph9pvSCAMXCRMYTGiV9uh6hCdxmJED0LoM50O9Pn6U/P5XK3ezeF1rX07I0vSKvjSA2Oek0AnZIh1uTkRxTjgj9JTCXYQuMrHh5Z6ijlDDar6IWBNuuVmkifzOwDAbJgHwIDagnmWCKZm/N3QuyKn9hnDr207duJRhSGTHhsnBCimFRgaj2W+/etM7qr8Y5+/KH7xoK/4iFCtizqjWrQtcBI0aJgIEUJ134AqkF7rbeEX0Ne2AQVgfBVZIhIa/uAw8Z0E9hiXOS4pEcBa9F72wNFylhvCiZkFwqlHtKF9aRvDGBIi4dSQgfhF4ASzC2TlWyANLPca18dvHj/sfoIu9dscj3Vfc52bGzZ6WqGgt7rQBzIolmgSl5kXrUKzKljjfULrEpZdFFb3gYl7e2ufFAvuBDbQskesyeDAJWugEkBGODCpz4Iy0ZeTk0RjvLf0fXR4Fsf5QUJ+dsZpApFT6BDrX4TGAiQAuJYvi7XKCThLbXgiKtrn0QaCmdmoz5qKtlj2qwp2GLyO+S6QCu9o5RMRvmd+kaOOFOCXxPoLJM+OTgWw72rUMGxUPDBTFEea+YQhcMsC7heHzsuyXJeF6j/vcUJBJjYtW1bIzIGIFSnaWJDngUoSpKkaISL20B4jEUJRxgHaVxnmB/A6D7JOol6Ik/Vt2WIlAFD0bUlii+jJHiyZkcQidycNOhSKcH7tos002bLBt8rCPOYfe7nNDh/wlZZaZYC4+7SWLmSvYNDhITMiSuotUbAIUxBaAxNmxgOkiuCx7JC1xPTcp0bwQY0gp+FG7EkUbsOdcihhNHTCO3MUjk8aihTfGoMu9fs8jyLzf5ceui3ULSG538oZGpo1SeAH1cLoIBHCpFezN7xlsWgQHAXhfREYckgYCDWUvGg6EgOBno/UCZJswMBQlFznJ0nrhJy07pf/D3dUmu43jwMsRZR+ALN3ApQOYVTmASuNjrwk0Pig5U7sr/pDkTCZ+fnl2DMEg0Gg0qly69gkq/ArtXm7/l2vS3Ftf62tc/hA1U/Id+eQ0Uxa5nqn9S5PFDuQlsLFWeF87/OiTHjLxGlv7gq/JYccB8/teaYKx2gfza4hS+E18v/M1QXsT309eM0Xl/94T7rBDiU34MhA7fjNQe6LmevIRb5Zg/y2FTdculHg+m7BO4vfsyuL1/DPtGeW1+PGpeXJ5s3HbD9R2R5y3+T1ieTNvKXbGIMFABSJGlsGfHZ3i8HGHWMwY5p+WEOMQqu3XJA4it8pfcDxlw+ERMYfdOFaXwvEAngXv+9pHzMwXCj8tHlk4xuvzfB9pr1LgwhWvNeHn8E2+0qVWCUBVr6B4Mv7M8vEjTUP4/BanlkLGorEzV17bWHH4uNOsmOFsRlLeHMNKVVPwaYdQCjvyZ5zkXbMZJxgSf6FOaitx4wnXRO6zuapfIvxtjgXts1PEjatdNapVf6hO0eQVz1rlpSd5SeKLTHI2ip2LHQBcAsHG6seGBjEddtv6OBwoABUvH8GIubSgbM6Jt29+zMaa3ALx24Vd30zjHmim8o+DPPtkz1DteeMPRScum1dSPyb9CLWgI35PfL1aOCowMs5WJOzs1mSJG/dLAjI/OFB4KF646HhLeVbl328GnOIftXuU7d6Zegqml1jjD/gl+226SX07enpvWXyjf8XJ45VEHBy8JC4sB2+ResnSocTZm5x3iwBBP1qkBzMKzthWsXALxUksrG+js0L9ixW37//nrfbf6K5V3Rn65627zJtLEm6kVpYAwUkj/pPfVslQM7LWeNr9mNfteXfExMwWnEPh8WjQzIRYtn9z/36r/72VRn2r7r6wq1xhbQqWJ8lAJGgwKGixGJGCvXho0iaD3gqy8U2K2DLmNv2Pf///fd36159nNybkE7hbFFCRP/zA0+rj1YeKg8fdqjyr9hLv9C1G35zPT+W6t7qLG+EuECTUfWR426K9fjjxPMTEOiqP2k56P0S5XNm+vXVJa6WMipPg10gw5BZyY+VdDfNisTAHigZjPhjBNAtPF7dvF4X5DiHJENxEWlNdnAAQNBIvjvWzgPFUvXi70y0jl5OaD1hqQW7MdIuGy89KmB+YUaza7/gW6o/E2EShe5nWq1KOwfoAAdeUCs/YsKpFMixQKNLGMFtrHAqyWO9mZAHm8BU6LVJPOzQvvY95NYGb4yaevXpuCUXr7D9aB+1+QaJolU0Ku1gel7J6MWLxD57xgONuFjhe8ol7WjgmyaUAz9cOGNtYD7wfxLYDJhbVKGvbNRzzkW961hVka1ZgW2rc45lMptjw2o5SVVZlqfxpVMqc7xWHf4AXZMee91Alb1uGH3fQsZq14/FI5DDm7WyrAULKDyk+Qn0nQzwWKLrz7lgBLUo7YuPG4qNKudz5llGIaINXSB4bLx6XF68CAwEsfjYeWr3raUcd7iYdEOmBoLskE3miDzIwL4YcEUOZXNxx73G6oXlJaxD08QJOYQSsxb14VF686hiNxgkmnt2rttMWiBoZb45bS0UrjxYpHKTYxYnjjaUPbPwnMY1junEYZtBNaF3FWGHBixc07zYgxaHqTplADMi/pfV8wzgBxhFsXOSkQ4OJKYXBi2fRAo42PlZ6vBAouK2UOFDcE6HQkAzDFmdBuhcLBXZet6yrY8edyKNJQvF4JGSONzMwOT2G20tFSViBF6RMivWjp92AvNi0DK3D39iCtUx3jcPCCvIgoY1+ZG0NagsE2GG9O5VIENorEx9vl01oe8kQNyOGeluJU7anBYphbKA1pMXMUxnafT5Pq4NgZ+s6l2IMcm30L+rF87qjZh7qepjQBw/RtGmW6WZxOJR2yIwV/mEw0xiwWt2pFw8roFUjQeBiIeXeLBgrz8rKZmMGsZ2T9fkjmtnT2g5lFIrIo63EM0g3sjA5LJHllCOzsMZiRAofOJ+39fORAno2LFMwCrpTceetOukiFaMFxbmplDderKrjg6gqPujxvYI8V0+3CsZIJfLOi8GCFSeW5l3nxetrWGPJ0mLz4setUHiuMQrqOSrOzfR5vpgWh+JuHROL23NgNvepsbjcJi/OSu0W7FIb+8rK9AlNwMWpzyjWcXgxIHnxYk7aplvYt7ezcKyyMOaDeQ2Q33vxmNJjfqlA+7ey4VH4e9TP4PvogYcJGp0fdNarwGzCVXFxvHEZxay9O/bipF6cS7mLjYvOZAIBChQgigPbOuIoApMWi9chsXhV6TChvr4laas3CBJkWVvRqWz+bS27kBWT8uQ/T5VWeY0DM1/qxTxyx7Pv0x18mCwfztkxecwRq7iABoqexS1dj1GDCKshbTLn0TKKm5x0OThxMdmATmbAJv1hYJvRBSI/j4OBpLxrM3dvtLXugEtoVowGhzFTHMM0L3ayyjKYmTkHL+bp5welOvWjjBdFiIsOTGO0o6DdnHfSIp0XP322cRgbKIiRcpc/Xz2jIOOsSVYB62afzu3EKkI+0c67yC9exx13MobQYjGrTt0BaCMY2xJhfNUp60ggpkSKyLsX23rkIaWHIm2ceLNe1aWZKuiDEppJXm/woCgbeuvE1n4W3W7s+egXng/x4s8igjV08QKaPCSb/JBLvrTp56SdfQnHXkGbF687MbxjeLEedy0vbnJsFw4UJrUi0jZk87hWd2zQCfSUsk/oPrFCa+DcnY/d8WkHzO/yMHxRfqCxqnIMwipe00sEGUQB3f5BHeiX0QYb0tb0n6bLEzNFqUniMPw3h8a+O3HHZ1NlRw0VY6q7Wbv8wIub+ud1OfLWkjOxBFI/dsumUHUAyMykiPzHQvFrk1McbixhyvybtzQBxXzlUw6xwoQRQ5OudBJ9IrmavP/87HmD42iDnhfzqEd6l4uOmGd35IBQWE+/kAlKinW1a+fl8xNMlaCbOY7CjequiVs1mdbrgZk+tIhkLQfeZag4ikdgUvljl7iyvBj953UcmLla7y615ihd1onR9BRwjbTdrP1QyYrNiWFj7yy5Num6vsapqsw2TdMyCtbYpumC5x1k8iylsKGvVPbnnII/5MqkUTdTFle+BoGZ62sOPAqoDF7OjZ3x4+KwyNeggefUFLGwuK7EYnLBwdAe3Yi/HsMonEfxzSgejapyxfMObSSbmiHKW0qKjTsrUiy2ds3zBavE1nUdqjY4O/n1Hxkyv5iFgfh4sxnKPz7hrAZOm0DBMdjOO+tAYwZ6DF6s26u+z7pwdfduFfT7mlkxRpHAKCYq5BpWTMAsgvkgk2jJGgVBR0fkP7u20tEC+hUp3JUrj+lSxxxqZjygUrqdprqmaZE8IV7skdjOu/mzqvrrqHEaF8wUHOhaHWh0OAzuEWjNTW3CKWBhioajJhQkiucmr+vyS+u4pM0Frv5ZRLb4et0NVUcukHCkkAr3sE+STFjKOqRsAZK3AV0E40GjjU1+aVZ56AfL5l8oSuQAYNp6AKf9dJjEpqyDB+cNziZb78aKJXxgYZ4evZymSs5uXah4uMJd2bmxysij29Edd7YVgbkqo5r8LMCk590f1lS5CphJRqsK+RqU1zbs1siayBaFhWdleo4uxM2SH6M60LwezNuj/Lm5UHFHFoKVkGL9fNp7sFQaIU5IVuGamR+HMuehez2i1mCjcLMu//nDcJz3sonb4uVc2TSSkjVDAV8CiXcvXpIYGUyVUUnbatM0Kg2U8mWOOnLRUWvmF4vEZefB2g8VY0tpR/1OhE8nXDMP82IOFW3j9fP7guVCeTHmQSHrmnMYHo8bmYKl1Y2zBOW02TqhNIqBsqSrVR5fG7etqW9ZkHGBky53liaoTZBtgkrYgUWKrRk2ARv3K8MWmVj6QPLjtQ4U1/XRRmXJX4I2SEWn53zwiygMLjpMHNBLBdckTqi5HaNY4q7iUdVdXK/Eu5WuQqMg11Qix4j7+YKYqwnuI1vvcgowMcX1NMZ9HUVVWU26xqgqbTXgdP5sgmyXRHZkLXf7quIqNvZaykZ1FXpKzh1FPi72eK2jqCoBongKNVO8uJ7bfR2dICWsYVdZaNFJNW2BAst/taSzwo5i/RymzMfN3bHwwmorrOgSkpmkQkpiaIA+2edCe2RNAYkcgAlNJnxFprAonoa0DYvFcR+CLKapZ5b88A6osn0gklusSUfZvdhyCe3tI2kjVB4Utqwvvp5GDrt5CAy0Sl4M+WLNKOjckVgk4LM5cw5ROEf+ZcgmJKXoLZx0+beRgbzJv23eHRKusTFz6UBXXj93heoumxTNRl5ik00oTmxpsc7Z5TAR9gx7PX6MehwFM3VAt23ObRvDT5wX5w5/xyRS3k/dNmMrrcrVfwDFh+GDFCKxD+j+WI15mKqisbhlFBFgOWWPA0vAbJhOcrjd9mNtOhPirkVi9+IuZwsc+bEwkFO4EYsfpx27y56vkSUWSoEn2ixHBroWSzzIp4RY7BjFEr14HqlfjEGE1WMxr2KtJw8URcmsON9KmHEm4wbikMMBJ6QJciw+lnbdjiUuPQZup2EykNq4DYU92gexnjRKaJ8DMHzRFYxRJMVdOAcesS1oDIlbDsZeFtuMyfNKGxWxw3mxjXqklE8LZmr1hhMvBxvHndIR/1FvVhzeUmP4bzzxnqapMlauf4Y6kHaWHg86Z9KWNw0lSdoK7dO1spsO1fwY7Q7AxV2Q0PpZGktNPmKgcI10PVadHm1S8vms6YQNxxTv5GsiEQBMhS4xfRsCgrTuYFtKES8GChSHPYZNLL3ChuKF0ZGT5sU5pJNZf2UHgpBHBBHBHDkTwsVEzzkbC4giLzNoiA1sj/KzfYxH8Uiy+/msgUJheMWGkVdo4ubJmoLv4tFwV8GFQoSgPUTR5hA+A2EgATtkOc3yT5vQnc6aF+cgCgYL284vhYgJ+YStxxX7ouWh6CUpxtbXd8ZUQc9j1FAYd/hXW+7aFiLI/vXzObFNImkObLlw4AVuqJeUtLFPHootSQ5QWys9tAH92StEH14Yb9Ojb9CBTnnUFVM90LFF2+ysQ3XZ4WFL1kw4BaFY53GprzwWoBS/+VYHe3cm/PpMp2bJkwXjUM79vlHIiA2lMO/dhQiPEx+jFw+bHp1fgfz6vb2Jt06c9LijIApmeKbXdx37RxlVNjiT0PgguG/uVYGeMhP2DJy218C8GBTuJDptp5umoeKjHErBdL62e27uCZg6IUoKTtgR9zcvdq3Bkaoqry4vBkv+pO1R6vUZ9Y4hbaUr6ALvRxWKs3qx1iJd/zlsM/8Pc1eb67YRAy+3jH0ALt4FCkMHsIEeQFB07Fq7HHJW8suz642lFAHS/nrYMhQ5nI81gftNNtA0ueVH+r1EmR+MNChB1UYVI5VDif2DUyhTJ9gOQWxWo8VZm7vSmQYKcyUdulfx7xFR5odd7kqPsMliA/yE4XMB273dmsjOqrhe+KW9Pi+HpfPZCMZVFNYNzJwmdhssbKBD9WKlnQN0eEfhZc2/tOqtlo1JnbCdwP4x0BjlG7TMdHYC9+zuS917cYEz9fp1OGsgjdeG1XMOXGJFs7JVLnBihyfUN47EoDxX8dIrCnresRfDz7H04nsrrjDQcR45cHg1VYcAalMN9GfrbodPXsATgIgVZ4/m+rGq4n7eQAPbDRYO93JZOtY4USe0HLHvdgwFI54oFL4x+wNX15SKxFfRc1O+0lRxJMYPXR2u/IXvc3EN0L0e7eqh2fnv93+SS2ZoZFv728UtCUMwZjTgx7ruFD5SFDLwpV+auWUsDb57nI4EtAmdmzOcBN1NkNM5xG0QEi14Dhl7fAfhE4lFCMtc7HqaeauneT8pLJSNUi5Lt4OVsUQYlTOqzFI74xxKbrnxO4hr1ox1M6/55Y71ucOln3HNNBC/uAzGR3pfbVgUOILC/wcKUUPiQyxDPEwb3Ez4nFyQi7OHbPDi4ZGj41sC3UsY1yyBCIfTNmJCI+MqKuO41olmBidIJmrbXfInFeG9jo/836DFbzu/urlujQq7HacTy+qxI8CHnxrkH782w5XYZKI2TbhmSaOItW0V5CW/Unu856pST/yVqXKSg1353SxQ3fwH3cFNn5mfEto6kAXNNYUcapJskbaxHO/Ipq2bWUItYvTic13kDwS0SYtSwIwmu61H1tZciXgTflCKY79KoPJA6Zk1WCncc28wE624pLte9SYHuUCDJRhcCTQPcsLMYE00ZOIQjNrc1nReDQGC8uqxFDJlFHdzuLIIXe/FpW8daWjTVRVL86FLmST7a852is1DK9FViJC5WT0SLkvFhXu49OK0FW+gocbTVOfXQxUx+ztjycvRjrOk5pbUkgTNYAkKJVvoHLcQvn6M8KMwHsVl6hxSPHFS2NdhhjZt/wXAj5vSaGvAX0cHmOZKGCwlGIZRb2hx+U0Vd/3cBb0YeXeHAoE0LvnGh3chOR/26/6cDbE09DK5kgZVrGBwb+biYMmbnKalA70fjOlVvPx0p68jFLAGLZ7+S8QkGT0lxM7iVYzVzltBxB4Ep00eoECWFz+4KqwPjwLmQNVt8GhZbJLDTEkaWzDJmdHhutup8ycEzEt7Xbc0l5TWzNdIYiM7iqHr0IYqTuP13ikWW9LbQbpEJlmHgZg+Cbc8FUYj2Hu0rnVYPCgAwTYQ5Rv/WKt47snMHMi2BokIR8EohGA2AMZqz2xX/RQwccqETSg0M05NoRuHrh7XxuI0tqYqHRfoy2WI7a5Gbd8OMrJl4Gqaw2mC1zl2bGsx53AAACAASURBVEzhYAX/FJjdcW5gK1Ra6+7mM109+vlRgG81L7e7enA8DD6BGzQ704QDUH1VRM2EiZWbIiiEB64XdeBYN+fR5YHHfyjctdd2V794rqe5ptsBljvx5VntuK8xsLWS51aL63elhjCIbx+Kl49K2swUo3NVulVxHKBnM9c9jMoc2ZbGbQWGKesAn4xhjede7wzqwLsyjJlSk4VQ+gR5cM+XrhPF0tlJ6lF78W3/9/X0UHCBHJmALFfpMqceKeErdIIFtMbHDfW7Po6OsAa69+JfxpPvZJYADfQEmXmSAzlcaWR9IdM58k9SWGMSSJz4i2f0KhF6Vdl+6VywVIC26ZHjx5tVbJaZVaC7QPJ7jxSmwqWAgwzsJ4UIIUvbgcP22XzuDGXDwcNfWRJJzVdXj3l2wPjS0Y9imdhgdF4Q+SNMxYCKPYDKjD6YnUJEwQCJXdqBKtaQ2aEDb3c7O0CfiakyTd0SERDbCFGY7pwDLcwUtDwJ8OEXP/hM/vBKx1CGMt3uTh0AElo4dDNOUND2XObYVZ942+h8npzCrfVydztIL8bWrBkpHVsNrstmUpvagSrG6CCBAj0AKMwy041ruh35zb7YXnhJMz/CeVRQxJ5IDhJbyut4VgLlJUwEo2dQ820rV9fETMaLqzy3C9I2EUoBh6t9z6MSWLz6AAES25q0rXSrSxGG4kIZM6iRxyudrNpEpEDXqbgrXgwrvDK07Tq1CWSMoticgcLXSUIeONRQ+ped9QUyxmi9gQY15mGNnKYObYDku2EUUcWpDm26e5cQogl6FqqPE40LP6lE/eIRC7Otd+oLtKb0uE+M8caFJN9TZh60wRrsocuZf/ezh4f65A0xxSXOWYgnKPTC2I3dQ7dZoNPj3SMIVyZtbFlXb3kDuYPxvRX/ruZAu56T3MNcjKodhrn1mYnIFiZhirszSb/4jX/4da4zm4EUfaNT4gJdbEnLWLyvnEa8iq05OF6sDLf5tuHiL1c6IzrJ9TIVptBvK5iGNkPaLj3ZQBGMCXPd/T53Gk3CcExoDrSxTGlwS40uXO901INbNI13Zt0Cxg1GMVx6mYiVsRgs+Sof1T3NdUPB6NJ9swkL1kSjc27bcRXqh8V2DMM8sD0YJ6pVAtEGu1NVApH/2ptupbR1gOaK8Doszcp7XQxroExoAw8/ok3IVkxTPne/zkR97Scz9ze2qLAdKdwNJ17dEFOCtO0TRWIQ0wyWvEPY/hGvqPLT9240D7G43bWfvLfD2Fx3V8DMHTuxhIUV/MsDBsocma1xrEvwuaO8jljs+NaxYai0XCAb2oa+VTzEelfSzEsa29feRUzCmQpM0HVOc2JHBPJ+jvVC03esqse9+IycMCfJXy69/Ivr3wdKp7nKnpEIEoR4cbWBE15188Gr4tuImanzsJMFnf+j+v24xmqaJsy83wV6Ah2o6u7uy91pRxzIDs6hJbeDqJvlsg2mcdUgcaZHJp9tlT/Nw9ElnEZh+3M/HsUFLtzlBrucR3eKHhWC1yQ3KDHJGPPWAQgWCBp0NaFh7dlf9YXP5HPeSxRG/OLlOnrdkzeozppArHPyDyCDEi50dlmHU39MRZNCm9T4u/5QxnMbTjN03O6Cb7VgmV87lLESe8KS6xqOVQ7H3CzBvVTMDKwf5/tcu8V989IjqCo1xWqyRMGp9+pRnV+rRHcXYMLDOnJsHcJBB+Buu0m8hj1NSs5p0yQPHZb0j01iDEZbtXPs1osnR9rORetR4u6+9vnORYaEUBCu3TnyillF6RysNFAya1RiFP+pVYxOuBrhJT/V3Nt+51HqxcvPumOauUQWueJjJ2vdTANPOPpD6T6sv5X087yGXlw+d3b16Lp6kNvgeDrt4XOuXsRgD1sV2zRBWufsFqMIOzAXNoBrDVtNn8GKK0RRiZl0f+6ZZn5xt8E65uzh1i9xUvKXFtUIWqutgg3iw8xc4K+UoluEmlGefeE1VaXPRDGQ/VL92ukOVnjiNzuxAFElbwSK88ns6uEpHdUHYcNu/eMZ6TFEMbqpSs8qrv/D7I2R6/G1SxcOG0wA8va6eWVJE4tz7cbmpuRM7S0yrD8U8blhqqCKuy3QfrorSv70+QcW95uQ7BMFIfC50ZE3ZphJPKpDUhye9cFa98f9meLiwZLvF4wZ6TQA5WU3OpCPatmdzHOdfpt5Ih4VjHjMbbF3gHr57P48RoAurOTb2Ma30swXwtUU4TSfvo5qc+lw20ZjBya68Deez5Xl2rAnonSfBybYjGJlJd+xikl3Vywd7914j3HCEbaKEWsTXCf8zOohuI6/s9mdwJtGnvzWnYNH0QbodunFmCjM+fWartdSRrcPf+Xs6Oy0V2FmsdCO4YwqHDgUXqMW+yWPAbYfoWKrYh+Me55H3VTFKNyVJH/7cKtwBLOJQ1GipUSvELcCwweveUH9jlX17NRW8OK5H9I2edS2eckvvip7XOyyA0CseVaNtAM+iYata/L0dxjnvtqL632/YJm/iocYkLae2134Fxc084O3O9ZziItn3PxHaTBObv1DgbhQz0RDfuA4+uRLu+rugVv/26IwJ8nvwmlTjBOC7MXMiVSJ88gVRhNuiUm9QX48g34zr5kLHn/uLt3iXSvegQTdgrTdTp/LWJL13VkgPsD3jmVffHAGUmmTMvuu6SstGJkeyT3EyOCql9F5JGMWUVhlS3+UihnJwnnllKLr9Dp4NTKIGTkHrx3r1qK7eON5M7N1sIiuzMx/6/Kxy1hs9s8NGbMRJq0SGEGagGGKMCLxQicugR4lm8aHNnBV+snMgyU/XotiST/TKBSqAxAn3P15MfnJ34iSSFEXcmYBP0X/Tw0bu9gOS9PsQv6h21wcUdvFDE9vn6xg9RzR0BrYkTR4KbD+CZdGQyjc6G7jXPXs4y7vi7nNHD+sUfRS8l8coyhfu+tn/SgkZ487sFZs0GXeslyt92JgY0g4LCckvbx3FA2/ubTVZjxMG5782wFAQzjXLC5iX18f/N6tWrHyGCFsXxWG8MK3I7qLsl+KvlLH1SthDk3YOnr0/Qt00UCfq+HHxyhtHoYrCgYxbc+cShViGbIVxIOHNdir09rZPIGWB05x5Mdc3CedZoAZnieF3X/sk360iCXMJ0Q2nj9oENl6BUPCdHdeVe3zMHGT6WFXj6EvJG8vPCAHeuG0fUKIICAQI5ucIAl63yzkpq3GBTKlqNv+pLR95PQkhok2EfrcyWOg+0Dy02WgdJp/K1H0Y5i8Qkmes9s1Ik00nD2qw52EN6PH3yq58P8/NL4MUVTF8zexHh2YmYvb4BKIsIwUtw8QrqT5Ded9c/FYwe/kSCxuk6vapGY/VHD8VMNjKGnS2ZmZuED3i04ZjCV/H1fS4l98+tB9lLgp9kfS3kYVp8hZwznJaT4akqTXXne016VenGJmmy3urpuSn3Mxi3/xB478nvoFuqCfOHASJY1BnDpwZDbHO58kSP6lL7diH9mWPlEUS+7B3W0uHpj9WthAn6KqSMbarMhN2rDX3KSxtAXPVlM3wpSXx4jmtm+92NQ0fOXvxcysU5st0GXvOP39z52E1qDuzprJv1wz5QO6V2PCZBGhapzho6/OEiO6hEFAyYG2oVAzL5dO3kATZW2PM1KK9UMlrI4AxSTcoPHkuu+gcESSy0MH15+7w0hdInJHk+sQ7n+l7TjaLRgzJEundCpfu9tfb8XhOgHllyHxLVgcFFeBTBT0wFAbvIRKjKs/jAXNPFdV2Azn165e8paHUK8e1Uv+bwd7iD+ymrgjNuYclIlEwg5TImmjIPcxQl/vwI7/eOOoA4XtHvOlH794Ik1YHYz1+ndDGyUTDzO3KYx2FQ2TRstmRTQgsPgAelTa0748vzif453PZ/OjiMPStKJxv31YsuyUwhv82zZtQlg8cHjsHmkFsDWnjv94uxYct3EYejkJ4QFo5AaDHCAB5gCCm2Pv2OLnUfa0TkJvt+h00e52V6OhyMf3ccxSXXPfAybiActd7iR5JVJsLvEnz51HCqrZ4HKVrqeXYjVWmmwbKk5WNb50jMskJQziPX6hTqy75n64FOqyCBt1r7S6iCXGWM02e9Cfdfbg/y1Ct1Z1nBB13XYzWrRnMyIK6r0GQcc7a1EynMLIQHuCpU95FLrkl+CAMwFj1zfr9Kyq5xp9BAsmWipnoqotTSkVV3V84JTJAzDJ5g095qbf3PLjltVRdPsQ2Hrc/we4uELbBvwfCHQGakqpYIhgF7jC9PxKnRgmZ8gdLR0uJjVVmb/SAuNNFPZT7e+XVXfH/0NjXNWdWJZ1YeUcK0W/rywosXKA+BXq5d5q37BM09IQWcTSKPX4ZOtxE/aruSVczxWPsm7rmOEWT8MiqaigWW3LufhgBw9dPTwyE04czXRK+pFCMGaqXf/y2VLdXedbLRpoPrOZmGRgnsDdrkCk2qZZU2+wPnBwDQ0xv9SsUdkbnkWHYGCm5EDneWbOrm38XoGgk6e7aq2a1oyKRMzItCqw3C9VtxtD3uJL6GUTvLg5WNHWEzZto+oQwiF/VChm192J1uPEtph99pgCtRV9nxlWdcYFZL2wYLt2qJWIsNpm/4yaMNLpLvcWz8rM1I6iLq4qp+I/LINzNVIxONJMFfPIi2nsCiOPog7E+EPIj5fkBmOHhWIugiVVLG38+j+z/PBbfCnLYulxLpFC3jghFqNnyoRyGenWNNpgu9Tg11Ycbfhp6ywgUuprI9tAp/oX9+ZPdXc/f85d7CgeJx2udA96i1kxY2zYLPErxOCqZXm1bMBjzVpDPMIVzxvWYPHJwxIFoaf4iA00q9ajSEdxkolY1SpR1aDGGWy8883MJhhC64ZjPYSuwSpUz9bQCvve6cWt/ZJi9fkG+qlaj3v3XzpbniQSxkm1txNqlArDVhQpgt5N7Pto/3OwGxajtlby0UPg4o198UeEq6XquB/FyqM4yxuIjU1svmsi22d0tuPqghmz3a9gvFbfGOfsbMmKB4VhWuN/YIBOs8JTaeNSi++LTdtZEmh2IqYFoYw7UeCngGeKq8cDCf5ou0YFRzqYQlw22oUIUChuiRRu7ygsL57PKsWhWxNTYjnkAvwq0yU5qUrIPzzOzPXwwKF/Q9Al4882r12e7m4hcWsO9He5XNa8u+tZl5gx2WcKC/0JLrHc4El6i2FM/nseVdkiaXCffeNBBEF3KOTvrh/D7PFBLZ417c6zR88wrqk84cp58szsgV4FAIWnQ5jabpPgc+gO0/aVI1KgTU49rD1uyrjKWvJrPs0ibVznp1PxCSkW7ASrQBJkf+csBsWVX+z2+4fcaHag4kaKZMIvt1ZAsPRM5RcruXjW3d0igT7Hf0mjs9W1cZuV5L40tmxW8/KKb907LOLNMI1rENveOff19vWVeYtn3Xp0pO2M+bmih5Xp6xjZmCD6CkTBYNAIio5Dg134AIRXL9BysYVIMZDa5qRbLK8dCdJ2zTdLYFvbMejrjKAyVaO/QwSKWmh7AuDRTg02+c3Jrc6sIqMBKTIkk4e2bBu6VUbG0kowXoDTey2nIPL21JmHoN9is/Uo2BB7HhVvkwDr8RLR9mpGK/Fqd9aVOVzNwhzMGT1QFHaXzInrCcdr+iTWVGf3Ga2xX/M+GDI6ik7SrxfgLZMt4ENNOPILVQUa4zMcrtaQYr7fp/tp/YSSrCw+dJP5pZXYPJ+lo1CWK1hPHGP8NKCpSMVo5Dt+Mhx5fe38wUuTNkrG0my+NWfcYnOwEv1itPQwB3NHiovd5RpBn5f64eaLud3q0d3ZtGj7LYYA3TlpgHZOW1fyn7YdhbcOBc6mqlPER7jDZoUJypmjHphjO4EH3fCQO0hhwx3ZDP2VhLSJ7E5irH7qxJqwlEq4YstqrTH5vbq1do0xKIqzmTq0gkT0hUJMA22CAqMtOHCLn6O2FM+8XI+bclVEPLr8eWva3SO7CGskVTT/meJOP3AnJJNKa/GrHGIDgGhA2pphmfrgkcrMlW/1/BJ/3SRbUgMzuxHeI1uIwD5zOIMNje6QDg/cCTaPxhIKxTHL5/CBRvUBje+h7fjJDYzz5DTIabuX1aftmnvAbPa51aIBdcFvXXABFmYFP239+xKzsv+lHdea0GLB2Gz7ZfAocIuVDJRl1w+5HiuDe9lAn/Dc9UfNXMzN7K5giqhQrdgYxLW+Q2ttQ70g2zcbEKSfCPssNAQzb8/cpDBc8guHO3GxVPUeC6G4OlU7WJiXIE+qKsMtbr12iL8G4gLc6v9Sp/GAw3tnnLYUNtAc6ED3wpdcmM38Uvpun923cb3IO2RXsH2uMdTgyMwxklAIHjntghttmgqz6zdX0ixmpiFBalyzUtpOWPKzwZcoSupVuaKQnK0nRuCH36oWtJ2ay7ZdExEp4EDWGCf2xb2jWL5YFluVa2q7ZvsOSbGrtbp1ynqs+t5Zv6YMeAbZwStambYpGNBSNBqsEpoYMFnGUt/wZybodiXCc/nX/5xw7cU4tViIipxDyEFPuNTNqPlNCFui6oqjvuD00+KXf4N6S8PFpgGdb2gRrTbcKOX/hJnpySntzzI/X/OCtnmCjk08PeweR5Wz+U2wYBXFfVxBUsfHnjzaAdtgYi4BkO/DiTTGhEbnWYol2y31/eglNSlMlTnKTXGrpRp6NTdoVKtR8a16TRpK4+qowfA8HjD8M90x0xbQYpeQltoIjo6ldFOVrJ6tukbJbi9bprMZI8DoLC53VfMtq8PEx/BL320Y+BDrL8HEB5qlTrp6trBWSk1tXOuEJCLwghcnYxT9YD1OQoY6cxJULS64IPBre6RYGdpOR0zbDUiD7/0Wd5a8vng55Ne1GvfxrrRLW7iv95xbbORLjkCmeCJMIaUDWGxVDT5GwUE9QlfzH8mliyr8ot8tM8kcrha3QW0obnkYhTnXrLeY0zphH58nsM5FIjzHAMaROFwPv3JbqjZF0CL+JkKoyCyu3Fz3lmX5IUjbLGYJ62OXyaKQDlgaN2smbLPvcR3FkHe7xep8d+yQQ70FGjEBR4VG8rb99k0iwpyqWFKftpVHsbLkH6nvHY7QmDM8WYfWjcNMfBuqAh/CL1sZr2yLS9D2t1a67d/iND8KMX7tWWyXNOprxV2HD3WCxBe1bdRKrDlJ6ospZgj1pQcvIj+NIkHeHkEaBhUTIkCCbjcJTLQlnVd97tq2XdIwClce9DG6jkkzIbROI30qEKvY6dr1mJ5jIzlAjsoeWNTUhvtZQGU+90DzNHNdVdM0jSh+pLQT1f3Y2Inx6BY/Ca/KhORczAr6df57KyO9tQDOtjM2DwLeZbRTIsWzB8ZnWeGpD/eqJrmX+zUNo2Cnw8uaXyRJfo0LSGfA+EfxNa6HM5K2UAUFYgrteeA1Pd/+3BUZ7vruLusW30BOs+yV7kllguGxcw2u7+yKKJ9NSF6MrM2vEH+iD1hBn5TIIP7lk2K0lebk114m5ltmpKBtlpa698iw/KiYNlw1u26K8V+wrXNfJfZx+TBA4eIC8jGaYKYmVNzR9lZ36uvSUNBz3x3os+gUd19aLvE9oU5waNkmY04E3/ISFGBK/eF4vEc6CtoOebTzCtJf+Zs9io1i/s+cd4t16/G9sm/yYqxUxqiTBvOQEMgWP6632P0RoBOuR1riAaIMPJS/mFT4jt+ECGpMmkS46vpcDVlaveSXM358DPx0L9fu/qNObFNIEnWUuAoNs4ZLy0eHZ4K1hv7YdONsbOI9Flax39IczbwJXpyVsRRjrNYz/tQKD9AJO2AV6zMqQzmAQIV1rc/FFR7Hxua2KxONuGX8QINobHThzgQz5zWM7faUXMyf/7drGjohye8xzCcK7OQ8pUzIjMHvsSdosxiloXOD6ZoGoQduPbpNW5pn5vLieZr5ZemLE9piBiaQE6zGUB/Wi1zUdCK6NR6bnmm4lIR6g8EaaHu5tSKb16By5OekWtwxCknQ/V7WoxnpNCJ11mA1TVZj1M6Ypat1bkUNlTgEOdd/FGGwUaKxtWh+jfeqBdz05sY18tx9ZRGuunHhE3GgZc1/TQAnukF8VfjdSzHcYpmclenK76ap+fqi0WCksicI2/wqWuHp5JG3Hl1KsaSZL0Ztn6JA7KerpsQ1jB0yOyO9yuQc2gwfSpCg+DVP49KItEA0tF5C9jzJdKeQ/POJdv2p/sV4i0sGWAz3uHaeVR1Uom7Db5s7zTvAIz/UUozMS9oBfNrvoIb7LxXo2uak1EZBgaQWL5D8essej09LsbXE6jgxuYmVU9mKemGWV8UckRbhUwRtlOXt95mbEEs27qst7/JyoL8gzbxjFPzhzCHIj6E/+uIJAYjZ2FUVXDGFD6/xgLppOr6vayhipOg76sWE9oxVxObc7CjmRKRND/gmcXer2OPx+ZKf9TJX90QoUos33VoZk08O3eVxgdQ2szTtIBi00TWSkF9tgF5l5qv1VxqFG8JHWy+J18/wHy0RKuzQ724+Ua0Mq5bcn8CXHMtp5LI5M6WFfph2DRRUCkIx18OyR3NEYfDaCUu+fgxPiLCj6sFKxyYnbPRAsMSMfh71iJajjZObFN62Qdnob6qxAuvRQLhK6ihuEhWGeDFPjw+eO8+umyo4E9sbt7FZslvsJoJ8vEw4f3UcpVsby8mW1oaBCBq3vRzwM5fCPWtevOjuLvX+GSRfdeExKf4jzAlP7ShbFbkX45dT1Xy/Yer8DW1tDy52UFlF0CEwPstcV8gqT0udKPxJpKCBbEDVjghQx+PVt8r0X2yuHvzSQPerSL/hzcX0QN39U9yrgpJ/Vg10DkYhXKDZXEkvHxNVBCeGnAMXKIXAYZPMeHlwiOLvh90cbG/2le7HSlur4rY/OF8WilkT8ivhLf5KxCjchbv8KWve3duIfIWpQ1HMCZYck2fOqD98EakzBGcf7NtoaMXol93RPtwWN31E+t6Z0/ntluVw9fOXu3CTJKc8Pm6IJVsN8R/E1yypo6/2q6NqB1aijQaDibjLj0dJOxi9vXU/d5juPxf+W4V3uBzN3HqYc833+qV6md524db9kcQEMpAF1RLBth5q4QommC9hbDH0i4ZL3AjZlzSO2lIj7Kz7IdsCen5m8ii+bt5RrI3xh0ibivVl8DDx12DqobFf6u/Kcsb1OBIfzHK9UDTa5xGH6tyGm15GV5XbV94G+vali6WVJH9ZF0vX92eOqUcd6Gs3md5giqsOp8TzS10Eoa9S2aBqu1K79ju48W1ElW9Vj84CU2QZ19wwxar8+fmPWVbQbyPyrGgxYBNCzISz/Y+1a0GS44Shl4OCA9DlG2z1AdiqPoAyzLGDEBKC7tmhZ3AljnftzUdWhPT03pMV15SKypumuLOzUe6Xoc3LCnr918lWt2mhXS0Y/uAJWuS5fEF3XwNmNoyCXrsvnju52cq+HvWR24zg8Vb5EAdrzUcuuV1uKiHj6WXrKPMaJHLGUTlx+NkEqWIUbbE0XCn+BpJ/MI+CTgrm8W77rGmzbbSzMkMHEeUqn7Axi2v2hllPGr3mGO5++cu6AOcvH1x2uyx+niP8/QGgnTuK7xZLtvEngnhOyPd8gyoo/3IjPncNmbfv0GEVX1AGCOcVU8/TPHV4BnmSjvwoqBgLXvxc1lFU45onnVjCf2QePf59iFEI4Sfo5Z0Cf5rHkvKItwPMZmeLhD+jlDAQW6HjzZ+ATx/l04nSuJkl0GnhRR0FaxsJLnbl0vaX67ugYIltUxCF3FdTLvHiijDBD/RXELv/W7d/rhsJuAjj9OxS/gROd9DIr4/nePFuSUeBMoREnpn/Phk6wqZprpbV5WwRtkkjXFUzzYn/LrjmBZ2Alpxwgb1pVlb//EWgMtEWS4dyfn0yCLTIp43xYk8XgL6kqgjLqg0fnRObZhVrWZ2dP+TzOphgTvukP2GLnFJQikWSrUc17P95rMrinVUIlMX43oX4aZ0QoWjY+Ixo6E472zY7M8OqHSi/+833JGK2afQvd9UVlEs1eesvzPMsbt6JDMQtG2XxQn6xyuJYjpnf92ljaILqbmeK2c4d8LK5nuiwPexjJ09btirRWxJfs1iGnwMnvTC2Eqn8WlciX04iKOfXfR1LXrmSAv4rfGaVwA7F1Z9G7TosG+8bBVIokzDT0dfsxLhxRWmFriHz55VzP+NhJnsTmxYklSA/vToEvc6/eNc0eYP2SzF8truzW5Xr13OB+lyr2brDw2xLIyYIE5UCuqoLPa2yiz/8RdDy0hLHklKupDXhxVIqqtX5MryYjV8h4Vz5qeGHVShb09mNBlbMElTT88TZOj+MbXDKzKtWDbx+IS/fvZLHBcn0DiSLaxqvc+EWDXTlcP/mEfrfhwGuItFNTR0iGNVm8RXFDMHcPSNxodCHF54TpzpeW+HDHI4aifKrESWHo65Hm/PrYHX+nWKpGtfw6u6+DKEGVq6UNwi+u+vcbjo3bbMK8p95DAP5GoZpBF50ZzAS5RMIIp+wu4gY8lSfOznr8Xjsy8x1NdLGriqfgxTMY9uEodKOHRhFYqMId9eG32+fe2dG0Gfr/NW0dxXvo1/914ePaLKHT9qm7fFY1lGQyrxw2vAJIPOlf582xZYJxWG4iCKU7doHWxaQB2PCu0XHy6C96IRFxq/wS1/R91x/cbmj89rHPO1B84iu091jWS3++WGZOdAFoHivoQhyDleOwYet2ed2tqNNmmT4juhdkJhbiL5N43jCKylI7D53FOQnOnNE7CnIRezg7Wht2zpC0Fd98bMWCvIQsx9cFLT1vKXYPtdrYHqwM0ILbF7mhmWi4R2A6SWA1TuiNcPjWsmfGhH9qfzi5Rz23EyQd1DOYgeHQuTLzmNf2xc/hDdYmZm/n+ITlTmhXzqqwU12oJwwbw3O8IJUfNYq+rPbuTfSU/iSxiXMrnwqlo5NqUfpVNjPKqTtpykbyQnvrjdQy14RO2+b7tb04rm5LXXeHpNMid3jpgAAIABJREFUK/+HLtefdqIjEkToxFHrMBYJwBHaYU+M2PHBxjUU4Me+aPSoD16pFInw4ntgpu2rMjExjRwBqw1F10yUUqwMHO37+utFfOtbFwZwjri/TH3Xp3ms1cLVxQcuhjvjV/aI3tfixR4ILw53XbgbRbsSgNjB0aqNqLo4bPgert523IXamucBnDitvaC/jiOpflWsMS7F2OUiEQtK5yBBehLhqggbHwtZ8gJnejqIELffW2esbP/u4Ttngr6bHZTIjidnpc8Nk+H0qqHwIwO7C3M/LjsFzLukZmcHruz60QFQU1UIaVtmrrv/qNeuKBvdfUS+I7luygd6684mCaG4S9l3e2c45Sd00tpX1Rra705FiB1lcoJSJ/Lbji9dohjnOg2JCVcVMF7GL3485FKYp47i3w0DJss9sbx8QR0VVa6uVhgUzTm+dy63b1XOflQuX7MCL5/H1I0lMX8cW9KXtTZ2bUcvRNgXD9ClThRXle326KGuSLQqrHw92HGijh0hzPYQvSrUX2mVThQ12f13BLYkoXYm0mo/msNh45YS0a08YxSP50Ihwr6r9ShRM+83xVZh8nKJcTyxxgld37qqBbNvWYI9FOkVTqmtiU957dusERVrIpXvEVjDaozBd+iMjdRqpARpV5WfrlJ8b9dflvxIRKIDir8frDtC2LQZfz/Z9cLb0DyWhDkRZvwQmie877oI/6c2ybiy5cAJo0Y65y6lc3kLgdNY8632ZVSVXTZL8KRjbHdta4Iw2dQz1xR2CgXShO0wudZ/xdU2Lzx//NUk2AFCecaIjGuCIZii9G0No+AsXmWuyz7nRYhAp8JmoTYWJ1nbzNi0y5KCixsMz5ENdvJSEnRYJehlnL+y+xFXoMaUwLQlEf1R8MyYsH44h1gxrpSOmBLkvvgA5lH8DNu7bw0d+VCYXKf5vVWJrez1R+8U0w5d1r2dsXI220yJGEccqDsVcbEO7SGfhsGXmCpyd8RmmLpmzOr8M+kp7Necx/syc138Oz3r6GH+y9PdvOquyTo0pTgEzYnX69Du1syc8OAVGuxfEFQaSg9drI/WTeSApvLU5dKcf+wxwDmBvckpTJcxl5rr7loCTSx5Y90W/33Us21CKm6UeN6KypU1MRa0RsRKb+pEl8i9Y+4Y35c4hdNkzLJL4s9hgB3SVaJoPR5nm7bvRw9RLJGv8CwOZJXJ0jbI68JAmwjigtD3EO9t7nxrzlp/BmYweR3JP171wfjRcbgyz+U6XP8niBTpGCMu33EDn6uxume+r4GB2C2BMPnCKZi+F29ZxxgEArL9G6e4mHzCjv3L9RwdJgYP+APY9FdcQXOYCr97Y6Kix+N/Y27ZsD7k8Joj/07lD0t8vb5lvgqjqFcbq5oGL23/2mlb0qD8wthq+3R2Rp9OMoqZ8o6N6U9Hv9o+GbwZ79X5ni1Ut3TClaggG/UVddCLBajA3hhynThy08ZbfhrvFj13CgYiDXQw/yYX0Oxkvmn0B01zu6fOqmOXrTBPc13hhd/PiyLxoql2ObyuFIac3C6R0i6WJhU3/dgal7b4ELj4Z7wA9M1dD/HMLHcxC6ftzpJfeGzUJDcd46ZLBS/tTHMuv3VJQtfhVm8v9qHQ+/P7mJsJdyoy8YiRsAr83vlqq/KkWvygLO4B4+8MHZs3EBbj3BbPKvktU6yqW3zQNmxGlQomwdvqKNj0zuEdycqPXYUfpzq4KNgJ1MoZ6iCHO6WUKJk9acFSwi1eTmJcj+Za8Ux9Fi/yBqoDdJ08fp0NE7VYHZ0ZMMywdXtR1RCH0HtOvEPj4a+/nDw+wI+bj9L+OirJDl84JAoSYdCVFR6OHIfDLuMoWw94Cpj5JN7gviiL+eqEL/9yhXH1O4+x1XFOPFT6KtErRS2ztlv62r86CT+eY9WKRvBXZEHfoZ/0sYNYP1+8EejrKMuP/Na5siBNStqITcUqZiaPHtQXo9ugtfGfncd/2mGJju5q2mQXjFEaO5nt3kq/RkoxCHdtRnxrQK2UGF3DvD4whxN2afm9K70ypnIOO3Vtvvkv/Sw7YyUycyBXFRyg/023xVsYtKLhrDvQO7oQ7j1z5+J8/hkYpzuEImopjolZaweCEgpzywF35cXLgcY/sWlLNN7xubvHokLRzHWxeuUsLgzj3/e1WKhW1mo3thFjs/p0dpB981u1KHRge+f5BRc+bNAB88TLRpUM4TyuyFjKqJGLMmHzZfmc4+wKnIwdRc7ipCH5VYVCeBQFL8bWarYUB3VZIpxQtgb7BLUTtTMmgqoHBj8yh8W50V+gw8A2E551HZWiLT9PwBv2xggDlR45IbaZ8rcuwqssonfl04b/LNzxhwkNtO2tgKxtC6VN7e3EejTYgck2VS3gUmE7W1h4tDvKH7ldw4CW1X4q/YRz+ZHLOV14g7Hs7o7Op22h0Tnf9Xiikj9O6u7ES1BIg3pwbiimRHk8cGBvBavZLJ3xTd9j87Vmp7Koq0TXwp8Akiy50iB7bCPyj3OzkRCRByzGzeh8H82Bvr6IwGeg6ebEhMWVVcXCNiSzy+GmdTZWjCfU8S/7dnL2zYHGXLTFY70QaoqwUwifIEwCO+MY6QuwDMd4IEcF67MpvMyjnXcdFncLsvgpCl1XKA+TTvF01XnjrVJoATZB6w7k8kwja09V48HmAE660HM+H/zkcScRHTVrpQriUJc/PgjIBBw+EqITOfLxQIwi4fCxP4Sssq4W83RXTgriAP07ZRWvKobtuNqbbZqOwFLRab9R/5e8XCyhe2ZmN4fEihFHGjCAKnHZOiPCCQXLdK5gFxFbChJAp6cSIgx8q+8sP3bNuCpJN9FRsL+HXApsB2jURKdcVILeJYW5tlgJyP2fkwaMiqXopUHGly22DQnOHdR4YEanMvjhcjT/JuB6lDEKPhb2WOVf/Kh7JcC9UghTJmJijdla4m3EgIQCbzsTq/AOn4COAtg4KP60curGQGJL1K9KrMc9iIV55HJArME80OWygFmcDMYYKW5Qsjg/dql2FAuzuDTGFaNAKkeccF8KG4tEaxZv1o7bJNHrN4B47IfDi4yEniPR6Rj9CQcC80JeDgcmc2nPqrqjdsSIxBPQhphmxBQ2mMXctpVCPGobVyBtzH2dPGNl67lAchTctDFms7Fio3jZ7N8hEoNXkLAiqfmXxsQVIi4i2GQqznYI6IagZcLhI2dxrECbx7EahXf4LTHSVl24lymW9qbQLY3xOxvuMLjeWbtt48i8VdNRw4uPiv7YKb62P+89rrSLoxQBIierJyhN5M6F9poc9Wu44UCgOJVa7Mo2JJZaLM6vO+/uVmAUrLsTW1KM1b/3AIUVw1Euw93sXA0FmZCpaPFhRr94WnWetBygfwPgROMu/fD/rF0Nct02D7wcOeQBqMkNPD4A/ZkH4Dy+Y3/EAuCfpCd1rEwbO07aSREUBBa7C0dLJOxBAQclhBfBjwApCjbSNcYp5pQli5PqEL6+H/WjeDdRWLTmpnGNFZrKDgDC2YP5QDmTrEaU7cbxd9SGBb/0O3n52mPUOc0xUuGGZTRaCof1R8TSDmNIjXt99yi9OYvfisnvmSoP0Aa/VdrowlUeM2eiebsezM7dwlxfuJbF4WMaH9pnD6/aiAPNh6qanI6JmISwZeFd4p9gJiYFs/4JRKw7MEznWNtiMcKruTzIaR7T3fEGWthA8GkjgnG49dgtWsb5DJjtpvxtYfdfNDN53YX6Q6GBH8aN7HrjEMUwJadEzMtEyGYB0uaQzYTM/+ZEe+lSIhWUGub0pvlOlqOLSfQfeRTt3F3NhDrbfbxnrkxXuw1HJcK4GB2OW84nJASdCB9bttEUNw9gxK5AjF3etJ92mUtCDXfsKw9K6OIAYtb4e/QWEVlcP6BcZPPuSNv7+0H16Ncs5DcMUfxcNhQNYGvv3Sw9MOqh2xCh2+SJmfLqPy1NF5NcFtGlBlhQPI0IOxwHnYIbKXsJmkcW09rDgXE1THffTzEzVRLGuzsix9AB3QtjFduOgdlBerDy2FY17uC1ZD8DxJO/mlSJg36iEwiN64FPOmAkFOJa/Gj4qP9htTR4xzgR6jGrPAyyGGsloJndLOHBoxNDX/w27EdxB8W0SosXxM3MQHGrFJLFN3bOK3c1H6xJT6wENX0HeRIvN3jBT8c0PBpjIm2DNRHBp6iDNCGaiUYVyuGsXBVRLL0eYcl3TtubxGeYG9z2c0F2FYHoZAG0N9ruLUUXz1zzJvze8m4SyuR1wkOtRTeUo1EwnjrfglkKomdwiUlFkwhXi8jhAnEjjXqJEM2URmLm92PSxqEWZy9aj3DeUYQhi0M/KzGKD6RGqKp83CeF+UbK9XyXzXoN5dRIyTW2tuOYUwOBuYJetYL9KAESNaa/ANywDqk/JnACDQVSGMVYAIpnMIoX12LUCfTFMXJD8e+WwqPLGHXk6Nt9uzhNhOut3ehFM3Zt/oS52cmbMwTnlKnNrTIcrGq0M3URxHlF2iK4NOOhwUu1b6uVYirGX49JG1EovnWAZr/sn+uLjKG7LY1o/OiaGwY7q3Bfy9gw+DwDFXloJ3ZMCrQPRV440SShOcNiH9wUx7Q2oBL0dXrrXAFxhTR3cCVNY6F4PYS06dFGIG2ZrPA+LpasOHswdW2bXCe2hbgtS9GRDX+hwl1wTG/GVD3huhbJd2fMQGIDwwf5zGNJpMkO+q9CvAmaRhD4OnBHLJvom/cUY/Fpewxpm442UitTe7afE96gnu0IjSgYtoZjzkfKZVmncJteYbzmCXpzwAs8JQx2c1H9tHi1TaH2gSJNzRjhwmQIzRv+6NAWc5NM5ZqIFKQdTWV87h7qKEaWvB4ptnfYQKHdIJ5wTAWJ23JJ6/Fy0e7DcsiMh3nMsqnzZlB79BbPO+ZpAwBKrK7DFr9ISaZk9ejRIpz6scfDD9G6JfqTKHkY79Yd9J+u0/TdHUEhtLr7YIbXCrDQrHoKt44tCP7e60T4jxNdXtqH/FmQNCj1HYTNmC9ojtYIp8jS8sQYfQQVNvJ8ApyiNiLkIpako6gRfm7rMWs98JtkkP2CzBaaZHQ6Bd/oa4IUh84W1M7NftI4m2bG74+Mtf1+DOlsKo8GQmhWLLulyls7BXhiommgQsFaGmonqAiTBoGIKo4QoGEDzfPz67EsVg00WPIxbGe1OHQZ2OQStnVfJT7PqDtRtSC9TcNcYcyTaz5T3WbcB7xLJk2I9oACmmNGFtdigIeQzDIxdCC4FFpsoWulqF1FDTtD8rz3eIpwNWIUuD1av8Xjvth2ejwvk7bQyEAzQGElZ+3A/LkmUfidtsOvJOPhi76BaAxMxPaLhLKG/wsofNAa0BoUOcyPnNjgOaolkJOWWiTf4znz566ZCx1IGFdZGuPwGWezrWMLIweoL5yFXdVOgd07djBuQseTohMSn3fEibEgF4Yjai5nnpExyTlEvA7QEdQUH7FNQjH+xf6ugJiZkixI8Tx9P+j8OnYU8d+HvljxSz12sM1lOAyHlKwZbXPtjbnOrJhwXoCLk/WpKm7VWynLppTfuoKTEiYD73GGWZho2IBOFPq+loj6s166Yj0E/eA18667w/IOvMHDns121/ihY9uWix12FuSOp8mvZo/OP/FmUSrug5s7cZiDiZ1NxiuXCK5kbR273UUQKlJb2VHxjQRfJIwdBL0hiaNQVV5PXjP/YrKyLPkdY5nhYna2OnccmUCzrWsYCnC4IKesIcwzrjnjyHlBgvTCTNOAkQ8NkVSYJ+EwuVHCOvqUpjguFRg66HuPuY4evFqJGaRQzuBThKtmdM5tGyNjh9Y14kvcztnNSPxkytY8rNpVsIuxzrfjSTNdO5uVvj01booRA5OHbxVy2jH1kqAfaklqSA21bjR7oFOLjllBBSJH9G4eoHxbjz45QHf1KBbQke7dnR5ZCo1tFRo7ZRvTWOD3MMJqUyG29569SdjsF+Bt/ZZ4q5+g3Ug8hkT5egTzhz7wqOzQTRQe6epTV4uxrDwIafNJKW3PLfl1Ay0gRdIsPnN1lSLSsngbq0RTMOoHtlq6w8fM8/SWT3kSvUZH8faZ1QqJWgZM0kTPdgTKEwEerghegHhqkmOUTx1/y+iM3+Vd2oP39XrI4aq5cItbf7TuZD1qh53H6P4ziw9sC7Vpnh736a4TXJl3uzpzsPVQHCg2x9yM1pdsGpHFGE2IGMgVGFMe8YQYeStF0rhbRFOd+Hpu6/FiceO7OTry2bRzpE1ZmWF3VEI/Uda28idudMV5sFzzn6rDeo4G4Luc6ACiw3TBjLYYvFbafsTIEBBA+KhDB8mhuRK7/Esb0oTmmPvi94OGjryDlp7NcxKHTwQrveuzHe6dzYCwzdiavXI0GC9c9iHP73uN3g97QX8yemKv6ARQ+Axtc4LnEuVtYX4QjyQca3ykKTpScGVzl7rO/CkNNJuIiYsYgwo/9mQLrbTiDmPOh+ysULWVK6hrvGu58wpW+gW3yIfaXT2T5LRaiOcERJpZKa6O2SjUAEv60s+BGp8jD3eucGM88CieO7XNfbHacEdzcowtdOeJbZQdNDpbv+lsTfPkv1OB80TqGadmf9oz0+4oai9cE5JttE2GQj8W7sco0IQM04ThQBcsaNIKXkTHZEEenh00YVGlje3oxOu56U5Z8pRqDo3YzxkOH/RSStjscmnNiJmuXS7O3ACM84Gk2X9G4/Szwre+aI8ExVcUrUGmOptpaOb9BleGwn4feOiAa1KZIFmCikcLkpjZrw+tR1+jQ7SeFDyCM20Xgulp5757DnY041ebpeHWWrhuJjAlrGoOP5vqHrPaXNcpRXr2iLnmIIEmmI3XR+jRKIHZ30NijAmPdB9lqBNvWd29nuwoXuJfTE1mIGrmEmE57tMqSD/ascr2+ezatEiydwa7I1njITjh+5SXjDg881VybJPETaU4rO4AB9NOI0I3U1ihS9mLlEZxTtxp0OrJC4X76HbKn89YtYN3OGN13LNZvtvKzbBCbEOQTSsRsuKQrL4teM4thP6Qk9JCjgSVwBfsPDm+mNlIY0fBJXMERuTxqMmzVxzWdSmiY3NA2gBRRCoUFOGitMHHOorv1rXx5FF/t/yUnXBUGtbWbKDHmaPT4u3dJJ6rbL7xy2KLvrBanXf9F7qGCmHRQVR4AMS85qAZGmWDV6XyV+SuLQ9yGmAUD6lH+zE2gJkct+OhQyjb7bVjtqBdb9p1S8x9h3wqxc2LDbRv5KCuvZsKiMuzXQJvMoh+WTAQkyIBO30emzPPJRxSfuowQDPIltgu4a114llmJryB3upHgZ6iYRH7ti2oi26nE7eDEjI8H5mOXttsd5gynzxr89fA+EONKMyslD8c2IOxWTzhQnS9lelVEmFePWNdF+Nv4a4YEAWtPNKAFz+1u6N/z1sPxnucCovbrkw0eGIbBHbbcLZjON6qqzsJbLjXq+V1rlsKsT9a6SseJHMINhz09EWmvlNiO+xDo4YX6DGPddwQGx3xeDXaRo/XgzeWvhsbyIsstgbt31orbHdsVALQcuNnGOxG68Zw5YiZ588HoNgvFtxcNkqeIk2+uaCzcufG8jpymPAw2wYz5RfiXBQJI/0Dc60w1hW0yULhLoPlx+xc87czVo02yLXYnPm02SYCozjP3j/TyqPfIr4HEufeTuxwTb9TmIMdyPB7n6PxY2dEJGqEPlzYuzjxog70bX7l9MHjMUQ6itKatsa4emY9OjjXQEtlfuy6grZKoRiUB7ZdcQ2D6521szr0xs3h7PelIN9ypUFYi8j2xSo3IcAF8BvVDcf758jSOmQ1hmdGMh0mD44y4MwW4Sd3d19NPprfQn4lw/7F/TW0s5c9xpTLZr4APw114Y4MLC90qzwUDL/bT+euIIe2Ocrw4fgQCnZ1nLueZ+WoSBpTq1Al/C8/cYJO8N/ctKGl+B/vlRbJ0p+9gXj0ePNwtHZtVpVg0+kZqs1mQjE50o1ZdXm21R/Bxbsz2etwx2sOPYCbhP1K62hSyWC9kdCsgV7lIJzh101ymW8gJEHdOI+5c0upOTo+OHq8vluMPb89AW7y9tDg1fZazPc/9g2xHVu2YK79n/20yMhHfjUTfJm76ta084sRWw9VgEFFk9itGB2DQd9GNkuRyWyJMtlh6BC/D8KL/VgnnuRRDLMH240EleCuTktckVnKyDKlrROKwwBhWnN9VdRPF7T9qn4+Fj32ls2JRB/y5si8VxxHibAkRo7CoQa5DLGuvGwxymjH6E9N419GM3l5l7rw7vUg0vbWqxP1N6sbuTmFQzsrMarsNjP5T9gGxLdH7ybvNR80wn66+jyF3Sc+AMZe2yxmjOBY0taZ+uIM1g8z4UlBA1StsJwmaV+c4q9L0lCMdUJYg1/PZrG4JdBwb8XpPKxm5lZM4xme2HanO3TmsDNL0H6uEnnMar8nwY+xTY0CFLkmOx6oaZVEO2cYCDro9akvYx48+D9I5cjBjDpEm986GEpO0wj9Vk7bi2+nPDXdvXotBlpLSfhvPndp2wX4DcVhWz3vehZ3HvGNFG5eNP+n7Wpw4+Zx6OVkSAdwrpDMAbyRDiCM5thrvkdKlD2ZmUW9LVqkab4PraBS5PtjPaStpaeMR4eGGQFEZLIhEp55H/SQowUm51Qw0cmAvWkbkQz/AS3aMJUUe+xkfr6NRMfLBFfjFldLrllMJ2GF2EIFdfHlSAMaxPMIvbOe7Q1i/Mw6XtMUsp1O5gO1cbR59x0HZgSFCfYDZ2NhXyZQMfqzhAigoqOd1A8UiAzouKkys1FHcTPu7sIFQGJdl56NLcV/Fo9SjGdvGZFsK4Tay0iKD27Bj9MBve3XOs52euTmVR7BpxEbpLmXVc3+qWzdCuh+rcXQshSof/SN209XAQqp3DnyPueaI3uKMdxd6Fi6af6S3uLHUx1FX946x+jOWjYj7FypeIcAeZIuzQ3yWa6JOxw7CtfYtQkdqqxyQJDHVjO8ipigt1hIcUhjAVpfz1qePzyETUpxxo0uXV3c37urRg/kUdy79nULFn3QV6yNQKuvg3Ji2J1n2nk8eC/OuR6lVvUcvHQOSCC7rHGN/uZL8SiahgAvYyPLLIOzQEHM3mavhjqxRbif8W3c4kLWQ0Vt98umu5sxS7YBSAl92wS2uvfOO3LHJTYjuRmVlpeCV09yOIxiCOHrROk/n7lBLxW1bGC9JQJzgU7I40dHUo1qStLxWXUT0r7hM2jn+Oi13k88bhduCuOu7XtXZuJQrCtep65CtWxexzaW+yjtvHzG6Z9yfw7nfIY7cXJdJQgQjd6kov+vQriNVtANVaLRRlM2a9aK3tm9CGeW5f0L8eThuQsCZqKleJL58U+32HcUFa9dWK1QmMnZ7RWdlGwjHNNwNkuzciG6LwnRehSsPVNrptEYq4C4ImYiau8GQ3M0VwFSEDbEh0XGj+JcA29xhAkaTZzc54ITjhw8VJrJtvj7svilm62xslrMdN1lopasazP7TMcxwzDbUVj8Xq/td1xOWkyv4v4bzizKhcaxElAia2UPI9IlsJpqM0sdYPgs5plGUALIWoSJRgaU3BTM3PTB8wvj7/+P6c41xn53a19+sPixY+EA7QKX1j7Qre/tX3WqBc534PLj6/TVpZtEtWJIR0azAWxholvjPqWgsDufMrnKQQdl3FlSTOja9jPOiGtTdrSMVJWrnrt+ibVOoKayOIyY4r7OzlF2PQHa5mdziX6Sse0m6Oou9rMQ89pdSQ7mjM04u4hIYmSvcVdroZoqkUWCGalPdOQ9iANJOW5KfOT9fD1IcTs3xv/23O3fLQqvRH+JvXyC3L/JrFw6QliDW8eo3P77oO3U72v9w+x1OG51GLRqDg9EsAlkIaNERBqx/PmBU1T0EJE+BCPrpPB2gj8SnMj7SZudhn1xIcn/uDCW9N7D+pXk37qnThnnL5uZhyp+5Hv44Xkd8M/HwaPpEPBx3v81JpOo4VUoFM1ExgX1QwozXreGKkHyrhDY7HOGnrmydkUKcRYsU0oGS8V8i29XYRS216NSN7j/Yx+P3Trzon6xaE8UdPvs5vTn5U8V27OGYobZ/LbWw2ebHnCjST/Q0Aw1BLEKDUJgC7yplQZfBANNk4st87MMeXJ7pWQkrRN9s8ftwlt8H02b/J3Qsi0unnixwuyiJ8h66Bn3Ury+D1g6BM7UMCfFn2Cffnv5wEX/66DLyWmWEVKf5gO0cCVSibk1qwwNp5z7oWv12IBQsFB0f67JBi8hlpgmfx99MfN1v3xP0UP5bdXoDGV2o1JP0n3t3bdnbZ6g0x9JbGnUYi5n5dovIsYFSdvAylIE/S+2DtUIKgFa+wDCKSPySLeM9kLqcDaG1JP8l+HFgOyo4VZ/bseBVr+BWH1guL1jfevMd4T1fyU6DjhEmqPaxntYQ5nIffBJSRN0gRfDaYDra8OFK75llN+9OOyfyhvs5SgSOaObyAAzpVL83O7382aPfxa/fo9bvITV0n8cMG+pgta1CcsfFofFB7fe+a1YO0yupJqe7MN1B76Ne9xYLCrbYJH9RGLwoqmQ/EsW4qLLDkqPp9n6ZKcnu7cX++XNGacsH7fx2mnmx/06kv+7c/yaS4qLqmPHcNdNi6rCIDzGetyeKbguf62Cr8/V2/WVSoVdGkuxOAe4BwziNWzw2WT5iVgPNoR6SBkuys+FQSQZJ8rxA+9dJjtaTEghmPx+iR/3u7mgL6jFdy0Ue6mo1FFEuZDDiTtDxsvkn9F2TU94WZaPBo7TFvLqAKADqpkOr94IsYpYxSh4RcRRi5h4Yzo8lfBMGERvXMw2sx8rdCq4wWSjpSuWS1zYten4/Lh0D/Tg+HHCNkAP58xqmlc3coyY4i5os/3vr1riNGmsfPzaYQtYCuctgT4wjOsBhb+Hq4Pa4kiurm7asBH2KR2G5xwXlXnemtDP8tTJCT8y3B6+pbgsS557M3OWAAAgAElEQVS7eB9D08YjUymm1eSxomqZd5Wv6zCIrk7Jtn6+gKa+iJyYpkCkeGiOINdKiMtAbzHXdABzK5py16L66mibYSEWgwIrhFzbKGV4wy3uaqD286Mx3JfBQLe+2KN29atimbanvMuA1nU9c8/rVItfp9TUk3D7NOkdykmk15bXGPoUjB20gwmuVrlrTT2M1ALSPq4dMdviYFUYD5s0w5lvXYTxo7fFhZA8F0FfpS/ulULDTKxpM3vd1+Gx01VgPaU4+Fr8wcUdEQiTA6Ge4Ymzipvb6/BKVex536RtMJMXSf3KSEGVw/c+QjB4oZdqjjo9799wofejzkQqrKUwEOiabCC3QdeSzoPuQxlZS1/+seuybUfbLSMtfl0+C5vwuGXyFzs9u/dVlxoMCfeWGJNSNTNe1K8ddg+G97Rm+QgqDcKcwZ4tAmHDVX4oWtx+Hr+/hIsPxrt/3evh9MW8jxo62sG1RYM+PMA2EfzLnFLz3slYj9l3p1KSRmhK08mZ2a7IqZJuWOLL2UpEiiQCk5V0loMLei8JASAEVYLi7kBx2H/Ix7mwFjcrxk4lf7/MPWoqedhpdPYYrrCOVGi3PO1as67CBGwuS2V9ba87LIR/IgAYepU2t25V3zWo2SK310mNkGNFlCvzwbRP2x+4HFNuVjTkkNNeGKRwyGAXgRxjuqNWZb/FZJa+LwvX/b7P3F0XtdneVn3xvqZ4TM/cBTfXra8LRX2qh09H59IAgqIubwb2Dia5T3sFconG8a5pHqmBat11wNkjskJE9G2bHHADyob7bCdsaRTKK92vQ9rsjJNS0KrzGWOHCyBVO6MTp5hScPlkcvb8xrjG6d2KXECWWqZlwWLBQMYIaFE6ymjX+Oax9KJB0APOUBNvm8E/hfMzSGgpEQJR6GunMgq9xXM80L82bea7q0befa0j/KerXlmNF78+SRco9TqxvhJjHqWBU0uc/mrm9L/ismwGwBsaVBUc5r66yCwPOeqsXYQJBeV3AKvJse59m9zdBsIDDUUjaCzT88/Mjl4wQN/YUnybUkWlKgrKLyM6Hscd+ubWAcezW/twv3N68qQdEl5PO+3G0ZetOz04vm3wfGAnPOTxYOzgHe1GfRgNclRSL1OuLddWThlvXil61OKB5uRBscrt+1IY6HYbcHHoUNuRc7bgsDD2Hjj8Z+3r7N43E4fe+KiX0N8V60bljhn9fFRNFXu3usHnTI6ZGSmqJibARs8zy3G2aExpKGLRlkLOWoE2OWkNG1RT2GVGhJsj+XstDrrRbiTqdlrpsIFm6R6liexYX1FK2qvNGu7TlpSuJSZhV4qFxOvZYrVwlaxGZpACpWg0dHTOGaaOvWujODCDWUKVkHON+tDJrzWQ4rcMr8d1+cV3zSXFHiulLdQANkzl9tixnTBOyRY7L4bHr+F1KX6OWM4empOErfVCgRQPqca41wWmpKQkM4SajVmCVT0d+jO1E2gfCu7txnMV5iMrCZ0NL24av/S4bGE8vL6Pm5OqMFTJ+jUXUePXX4auBAomUFnDq80Syau0D8lrT5M90tYLckt6gTfBM8Uzg3//KM6iJaarLg7P7UApLJ5Gut9UUBUUo4gEMu3dU/lrd+g+vi9s2mwRdO26wcUimFbnAztYO8boYUawD0Kgazo47dKssKrHucMCJmwZSgrUSQh2DIIfmATRCVIetRjjzLYNdzVioss01Ww59wKRWSP6g/f7q5WC1N39otFjSLg1l9Q9bYYWT0lLbsGo7RX9ZD1u+svIUc+bAr1qGIyHnCdMBgVvncDDEMmL2DKEzZCIprSdwW5ZZRU87MafMfAp9CP1OPcTLiPOUcnR+yWFwlK4k3UUAwcytHjxu34Wu8C6zW50FO/IjjqDaH8Z7A5leOucaMRKezD6wCj0eJt5Q1tTj8HGbG0pJ2XTaLZcIf4RPpT4RNZbXEadkOFDLvKth+teJ+G+GQxUNVPCYEtNFpxF8S7OSs0dn3RrR4vHUWhVk+vdWlLgEqUYE5tOepH5xCIYjEhiQ7rSpgZc1bSFzeBh6SYK3rdMQlRKMW7xfsCp1+Js3N0NnIcJjK8KrvkeyTVsuIxaMk38V19CrCjbiMkcuR5reAVkpono8EsD/9h4MCqFFmjF3BtRIET/FITEJxq/FOehdJBejgiOozF0IleOy4DbFGTzt5iH/FukGivJf93oIQ/eozdtQc0eS9/3w07tyz7AIm0TqGj/thxWKb1riv+MME9HzTE4JJjqKrfQAO1pSAkTexgTgRAfGIrT/sjpZ1UPZ/J1zXo1u7laNuTEM0Mzi20eZV98u2r00Jw22TqhHGfXAw2x9rSW0bOj7lz/POQpZa2e0IjjZF1NNaHb3itmD7xpuupyCxWHq0462uyQ1NjMf7BxzAMgQSdubpU4hU7PDfhEQwkRDjp3sO2O6eMq7u4+Ca76LV70gO0iE6NfHDxh2+wsCPoTRrQOw3g6S4JqOGeoRN02U537rsREvBJpmCFaWOOQXioX2igpBnukgE/TDg33mDqVrMetDYUIKX6+r5UNWjhQ7QN0v7k9fRRH+zVR/Cb/+SABaLLM1IOvvL7n+yCNwBOHJYCR+1q1UdNLXBmJQNhXpVVRYeG9DEuV2HCgcqGFHAVczFNuoKBN//r4UdLjok1hBsnfHn72WEdewmK5d+s6NROL23q5vlNaTYViisx9ZlGy3YDshquZkoo+f8J/btyuhm6DzZr2wnp/A/o2bO4AuaTG3L2FoDAlK04hNxv3WFXy3RR27S3+L2tXgts4DgQ/J0F8gPKFxA8QIj6AMP3sNauqm03Zjo0dDbJzbWaBZTjN7uo6gLTdvBZrsjBt3Ve0ZIuKXK6cA5X4JXuiPFPQlSf+S2G4LszvSazHQoux7UfoSaIdphvaJa8Nk7GqIOYwEtuCdo3dG2EJvHC8xZlnrgH6N5/s0/Ydw9jsFndHCo9l/OpCO88OlPfE/IFGKWBqHgKW3tQIB9oo54C+mfmhhNnpviR3V1kG2k6fRhPOhG+XNqv73dRA7PbWkZupt65t7gwuPktO051rimAg08jYRmkU2k1R1dhv8fou8vLooBTEzS89rCpX+Ai93PT40bkRslC52pVu/LOZjFFiJTpY7fc/gOrbrjIOOvO9U8tW2b7VdL/EPx6ydOJi6WqJCGADFcXUmc6uV+I44ClZ1BjGLj6Y/+ZtH0kUPkyn8bdIfmcv0WjaMAHCXg70QC1CMzwdmegzdc81Gnjs6DEIEgMSajNezYLW0E6QQhGevK2pR3+tMb6dmRT2ffVIhKItcq/CnvZz8Fqaojz0LXliVHcEJ+ijNQK+32CkzHFDt5fuSvcZLrUbi6U+BYwLve6AZqZsHfFCfR3pE5ibuUfa/YRJ3t5D46b3LkBt55kl9FwPYwPNXiNgyNYT19zMyubnues8pnfMiYfe4nCnn7RwbCpYl7PcVArc4zd5ICQ2wtUoxGi+dqqbUSQqua7s0sgZxHHjkAFoqimmq0rV6g6V+LTp7t6g+HMnLQKRzFl76PlrzNBegz2mIxTzB4rc8oR29ejYOEjtWlNRC5OdC2XO9PWhtyu87bbM4A6ja2Mc2YxURQoFtR1CitVPsBgTcNtD10ZO2+1ylkX0t4Uslc5pC+6j6xgcGBJGDWZbP8kCc9JEGUrCU0t+QG5IWsSbtlFrB1cf3GvIzJkbgafvXiWAbNJSm44eXCZV2y63oblql79n6yd2tcfOVLnf4Yrx7gJt41ky82A36P/LRvZZu+G2LfenQHqN29A/isWhFhxTwaZgcp4CgW3zf0W3n/ZH6NJISVIRh81Tk6iRkUi03VxRJzbiDzvWdnzv+i3W98KMTQJ9kzn5mWYJ5vyatLub1y5V8lv8FV1eOXusn4W3lld92QtFnvyV8jbJM7A9ggtSDtrlpqMgFAbkDNbJz7rdbUwem4LsQMXc2BpbR+G3eGd7HJZ3P41IcTlXsdTJr3GAXm2lFBq2yQnb68iwWt+amfv7pkf1sTIfyjaD1Rbf5jUjtjKZVByUKwBAPNZA15avNpCIbcddrvgRAiV1FDsX+6zFVXBmr8UXOp1fTho9rp3UliLQZuqvMDx/9eWzbnFgvc5/rkRHq8YyKsyLjXvJOYGLtxQgDVO/2GLdE+xTJksZBq9NyhlLQ8na7sOBDZMF/xFwqdEZxyvhna+W7rW4leJHO4p/9KO4hLa4KCjUOMUhwaMf7mzxKNORofJRxSiP5fkgAINVIyMC9degseJLpfUEkYlkfB/b10m+2CaNBHmBpDL7Zh3azkma/UPviHmj/RZfM8Qel7MSdLW6u/RIhEl52V9hk/Q1QkCrxbau3d1jfcNiS9PBv+NxMR3ReMn0YaUETJggECSjMEtZlIfrHrnckhbAxth0tIeO2MNOQrFh8Nm2z7zKnO+EUTSYQh5i13NY8lc75WAmz07MWuPZB7up7/d5uO+nu3QYPdL0BNU8cKyKmonMQwbqsyTCxIW2gcDlQROUOxUXHQTXBFOgk+AHGEDsKISu8UazndAVl0Q3e9b25fv7tOiUBmZy9Ehd7OFL6OA/YVA8+2Gjbq8flYfiqyS3yJx6onYZdLuFBKBl4tTcfoa9KMINGtyDnu1eDMqmxw4tMc+1Ytmxa+vB7aiuL8iC6iq8XfYDbriGajFarNNk5he1FLcLRw8ixgpQGa2sZt+Fzn6LTWX3Sb+WnqqTnn+2bFNgS4ygDobdTjQZJfzOKFE8etX8fggPS5EEmrYu6e7cqniLuVRi11Zt6XH76TbcJ1nhPW49vGeLSObkbsVTqMOsxH8SVdJbW5WHtf4i4zUGipa8BIYxLEhNHpoVdtCfvgWlGcT4bIxALDv27HyJPRtEnPsoQj4QQIoLb/F5kLwlQZcbFbqr8STmB0c2t2OzUhG+/UnHtM1zUbhoOaCZ+ImJyb0UI01Jilx0YXJMgcfgNrmxUpY15kZmG83X0KARzvQHbT/cYtvx6+hhwjSYyZ/VF99s62FT7RzRzDX2wy57Dl5hc48OfMOfGNUyDxhbeBUXyEYTDX5oBJ0mJX/lxVo3Smw2GLBh2tsXKWYgn6F8Mdujxi1Hr8XGVGGdYM/22/zwfi4KnThvsRQb42R6Gm8lhr54kB+8TXd+kR76pPh265QFXhNlmoxWXBinhG9FIR1s2orsEDLdPjZ3ASIrXpAllDOLHjQrwMMtJlmFfdtPyKc55xbjK4U2cCC/To5JzA87palnrc3vbvDx7gIsLWkY8564AqHaSli+LGYs2CwFp8w2AuuOXcvQbpmSF5jT0PyAPYSNbrt4E8LaxI03inw3/SDQdnkINP+3rYdbv4qbOQdW2xpjcmlnta6+F33HFkzTg4PrGKL0RF8+yaERuWpMGkb/xiRAwj4FBrl0n3CqYN5T0zgvoP60waPA/UegBLsGNcPeUVS/w9x7dOkdvV/P8gYyNlCySjz7fjQkrlmZWE130AGg9e3A/Ge6j6twjZS5ad9Ba9eN1DX4gzHSDi+fAkUbvS0Jk9hpi5ltiVSNvpZtjcQG2fpi/bD7Enpp4TS/v7jFXJCeeIujxRWHiq/oa96ddHvKz/vlfjqiQSVFF4rpYKdSpKzrHioLDVPQGmcGJoEIyzLc3YgtRgKWrk2fb8WhSsxBQEJmS4ZRWC3Opja/P6Ext/FMyw/PgTYOt2ysaLI7j+sO1WFjss3vqdspFooynm55+viBHrj4Z7XTFSElefQizMFGG0Fa3e0tJZfkQFExFwN8bClqXZw6ChULrT1+ftQXn4K0XZyqEtceWt1bqfiyKLvQVByhn1fVwmnZ5ehHnJ5mBm5Q27XeGaPcBO1GhslS6zawaJInDSwR3G2UxHgulVBaF717Vgt4qatGvOzjnbGt+OABpLj2NLaTdnfuXKPF2uxKhEMr0b0bRWd5g8WXF6NxKccQpRLyDmBxt5lHMT7AL27OHoxfbB7EgDp5jSsnDfqM7p4owSdsUdmojr1nGziyLUbtoAUY/9x+rw+WmSfIzK/cepTJX7x57lKlafVVv8kP3kZUDfhECT++EHiUHj3DqQ7tBEIPmss5RLmoIRIcmCdYzwrMUs2gSOxbeNGyMQR1wjm2yPq89nUohNouN3OTP7Fpuxlc7MI71N8Q4DrkEAfpwas376BP6mv+EjkT45HjpUtUh1ZxiyvcwhjUOtVNlPiNJmFBIiq7uwUlg1s7YJlyVzJmlUFrQuL9GrN7XgAYY7wD0nYGJH8xyZKkCHYwa48ajZ5htg2dQ2jgO57VNIYzjnSVR47xxvbNUiUqJXcwxARbAnT41hjLJUw5VRvwH7dybZeY99h0ScZzJWLR5zoViT0u70QbPJElH5ceSQ2FNDO9k4hhxL7df0PdPhjbidxaHtX7/lm5XWDt+NkDw4ctUf/Frsxm5eRpSXjY4JZLNwQznfCRYrfTzfa+9bbNdqSIxrQV9OXBbvDfZebXQJM3/egIAa3mEWYg2/qRCCxNQ5jdI3ZZWII3p64Vdm2FwWD3MyxNO9O8czcaklYgbyJNdONc/FyCjfs1XuyETYBQ45GyYjhUYXW6diLFidEp3771MKqKuPBjU+EVmY3wMcf1T2b8wAJKzwNR+i8bBiHqdmvYaNnYbrEZ27VTTsw9cGcasCe4bl6ERWx2xnvtA9zu8FrehyfPaPKdSHFa9igJAy4fDSrowX20t8LGj/9w6xxBtjKaP6fB6MpcJ5IRMskpXiggXcQnbm9fdkwNhm3NjgbECSid77Wi6EjhYN51M7rF+1CL1TSLm7kl1eILE+Mvp4GZpFF4nQhE7VFBY5t+J23Pf17kJ5TiQWh++FpsqBQwLudSqXFdYQPdaCkFeyVOHimIDdwQkyAmjlLVwjR21emtnV5lYoTw4IXVkmxJo3PN/99AXwQX97ZNpWIaDEg98WD9xGb70RUzRWOg8sBk0ReD0WoL+zQQBtu0waiODVtnGmESkhfmYy6kDV5rd3mTZa4TfzolRaNzPTQUjle0/3pDM5V4N8zQ/xgYL4WuVcm18wZjwZh6lJLxWf7SipZnFIlyzJnQ/RXKtg2MFXJWTH2r+nAv1DucU4jEq1DQhh+DM9dHtea+q6veDfumX+2GgDauR3yCbnyr2/VENlCICvPZY1zdUXbAIfrvgeOliWN6ZpeSDiQgrJQamXijYwoT7TbqzbeF0eUtt8ftq/zgtROtOVvdrc5D8ZcuP97g4TKLJv+bWzwNuZknYRQWFWbTXQfl58H4brLpeR2a4vXl0s6XHZG8NvYP0UNsY4DoAu5EFjcTnXKWyagHZBMHKrBrxMdieDEZ20QujTfhD5yN077u9zosGGhC13b9NcXSGX0xfS1uo9ZjnsbUwNn3oO4/sb43bjx6eaTpABYX9xALbVymac1GA7wMy/jMEEbmAYrm2nmBk+bnXT53BNgaDbP2vYa9crVv6zqQuVuBNqs2Th6Xy4nZo1ft7kpYfBzRie6lGwaP9QOMLVzj8vRTNlMdMEei4Q+JV7oNcBOcdN2aTZIvIkHeGW9UzTQ3D1zjZILQPnX0WxxPts8haEFMs3QTo+1yzgD9HVIb9d4JGD7c5Wn2iJ8PLD7SgYdZRjfM0T5MlXgJAumtkdZgCL3JI6wq1a52NvEiELPRtJUM6BJcA96ta4jVId5nXWArx3QcvBxK8Qm3mIF3/Lu7TnHk+JodXTMHivdpgb1OlIFEVY4Lfy8RWeQUaA6g7WgXGYFK2OovToQ3k2LFk2f8qjHmG07h05s8Jky8Hwqvy/jjhnSnN9BS6+15EPQ/MTNdBF2cxG0r0i+RtGezbYyE4j9atvTS2uqJbUrpBaMdcKWtVYUHdEE9hjlYgh/N4iGtRlxb9NTJC2xxRZLtnX09l32SDvSJcKnb22gWTHA6jzTuf4XkffIwNc1Ef0yvyK6ys63SX3LRkUecHtDMB9l57rD8ovAZlIpMrWjbMsOSf6kQ7ndVx66cKjx4tMfk2+eGuXselxveyh1aN1z53IxVbr9m+XFWXwww8xummcmfuzWSiR3DDODl54zXMi4/jnLoaPysSEYzet2Q3kGiWisPu5XkSnvtDrOhiVh0U5f/WLu23MZxIHg5CuQBtFdIdABhxAMQpo+9YfWTtGzLCfOxCwwQDMBptLqr6+E82LQ+s2KY2Q8a3TdPTqStF99I7DEtpPhrqOKFGcZ6SOo8HBkPesV5TV3/HeiufS+O0qAJyIRzB2waIysRKna83GYKPSUJJZ4BeTrYMb1KPmlR5uLDDndGTfH/zfq4kj7KSNttmhUeb9CEtCkdSP09pIa9V7yjUKzvZorSeao8ap6TCXAxAKOWYyR2MSvqGjEl8upBXvzywRPiK8xTIqwQqn7hqmNKKIRpRVz7WubrXfvsIQSod0v4M0vex7tKFY8OmbRD64X/Kam4nBOyH4TO/MAxmHkudeUamBkPI4q0p8z6A1w88q5OmFFPHmQcWElvoLJxM9nuGIL9J894g0zO/Hf/3oTUNlGxtIlfv52gw6qmbNwfls7Fan1t7pEGIXkKT1bnYBNxFppKI2bDWlCix+Ga0tAfmtMYlxAVAsyJD7iYiwdCTyQWe0w+bzygFHIibZMfzcUcBT2HDWQgkGGZ7lMnfO2wiFUNHUivisCSw93SWXmzVr9xLmEzkSSIhpy1Wzump2Uf85pko8PCEdkAmpmWkVShVdBhY62dV2+HaFZcRyUSAaEy2xQYiHMnuIrJwXgweuXtQw0F3zCLB3PtdGJB2o9r5ogQmDrRUMvCx9BM5MtMwI9YugrDaj/EibiKVYLovkR/eygHyK/MYy2LDP3O6SnzolMar6jvxUGVM9qGJZZxXcM7E90H3/2HZ+6IK9J8M6UnxUAamnbCa9Tt1i6icNkwSiQGgKJOFGRFjBtSPLLhaZx1oPuHRyQexjaGgVJTmm/3baKX/EZDmyp0vYPxMBjbxe79QFF6PaPH3Mrwf428zDuZQXMZE4E7NCRNzH7ER4WelfW3WOcKufAL/ZJXNZl1jV7l789jPxZHPNgDCUt+ynbHqwdLG0sSknZPT7EqXt66T4xk4pKeSZeIYZWDoMMAJBpaUeHbiGm4RIZ+KKQqWHwdxYg2WR0NYLtoY6qudP6If9qB/RePXaJRxbR7zPrc+QXasYGGBJqVu4RLg19fXDnwhUsnLaL4y38075TmYd6GCSwdCGPMgV2rMoJ+ongs5cyOxEjvMWdXkRzp4TObicqh+v3Do/Ed7oYTNHzEssPkp7mqaBpb0evy6kRKEgYvcUrr6wI+cUtJp1pS6xVtkABjZef0OtTzTx8uaMSCDFs4IPmDwZaGUuyantxwCO0ZR98d8rAz+3em3yIl/6immWX5obe7fmzjC7TMxZ9c7ZLZbZeHnSQ681FRIrQXLbRHt3RcgtiimKhku9rR/zJLkwicUO2czg7+APr8amf/LnulQPObnPlvs2CgzaexBTswG1+bq9hHwq8vBuEnsv0SRvRYD6ctUYLOdjt5WdUWdgmTZz4ksdI5s98dZUkA0RSjRhw9DseZ4Fft9rv6dDq24AkMxl/bxCqWrG3JsWI3D89REfvGtz5Lzo8tPZd9ucNzVEfzHYf9PTCBjSjxgYBishbMh1hjRrWfqAT4COWVz6FVIcuarTMoVHScThQ//TqkKsvd9jVxLjajczakYEDCJLlsksnRB8sl8vYz3s9jHCZmO3hN/NRpQIxEW6mBrzUiW6CvG9I7MuehsCn/AT+aWI/a3TorG+bK3tFx2J6NFmLBxFHQPU/+7+TXLt+Vtg+fHbhYa7C0n+WleCa4nLVkl2fT3UU+iVL6F4jawNgKvnNUxdFZ8FuQqLbjSimXzRXhcM242thQD+Wyvf4RKj30o9+NHzWvim8aspQSO1Io+LPaEY/pKhcjXDs1Uno6eARNROFXhy6syAmvItkWq13hzYMOHm2MyBKkJEBl9iqvQz3Y9HnfvzLRge7ytZsmMyelubglFB2MdVaTSHhD2ZbwBoyX+8YDEehcCJ2Q5YOxGBRXOJDuVfY7Vexz4BdYxJGQePaA50iqbBLcwUnQLMNe4UCs0v3mA+mUz9325WiD4L4WpaQ4JEg4Kh3Ran2rVCp9jqvfTYoq9uFyR9Xcto5a+OYcOTGbmnEVroo1CpDY0IV3OTnbOdlq+XiNsI1VLJnxo0n0nEbh82kcF3Nxw5pCxacdwyNs5ZGj3Y9w1QP1FIyL4aId60i8X4lMzHLGrM65Us6I6sDMth/q+8zNWE75LOK48sq48TWGsToYe7+Ev2EU8r3zFO7gHSjEcZfGtuWKwEMPoieRrqbYT6T8IhoQ7kiY3NpgTA9OlzyHEAu8plBmE5ez5XO/Gfd3UKW9vnpjApKIDHTfZm13mzbju5kDBQlRMkOgK4RibcR89ijplCzopEusZ9xhrl121ts0lK3x2iqJGbO0CPg2kppGTIBEM3NY0rBSt/VaeqFHSEfB6gEH45mNgmIxN28mb1EHNrCJ8fabOLAz2PgMB6o8SxTY3tH5OXCFhkTGKa1LNHPBGKNmkpNlChKsdnIcFePy6Bko1oazffReNmXzXLnXjX3atrlDW3e7W+U9HVR8US1aunmtDHyVhzv/TkZhACcqdxWgbO5Rs1g1sv1PpqWZ1frwn1Azfh2LVXHQ1+m7foxQhO9vXu8muQ2O8tEi3GGLQNA57nmwxIl+pgfU0nNuN6OWQa5zbLEN48CgDDaxHMU7twYMt9HoTFNqt9zJ9Sj7SePN87ITN+tpvrY5R/4vUqyDDaTy0dUAtkWZsINsf33lodJZa4/Zz0VIKsQcTnxJIuStoIqhPQCTIhPORo9PebhwJW7jRKziO+OqOHfnuod70pt+XIU4OM3QcetO0A6S15udpfyo2eBF7X7ys9vAY6udQcJO6GXhYoVSiYBL5sIb+ydkgoISpTAeNFRUTpOwRqzqmXx5LCaefLlbiK4nBP01KYzZKmYm72kXjksAACAASURBVEwpgjllXv4pmjRzcvmwnHga1qSCaxQzFRjmhkgkqyCWxPzMkv9F2WoSOaysNSVQaePIx8Wf7AjGt/usSEGkWOHEf/MCXTUhHTH49UUkvDImyonKXLDkwvpm4weStV0it6XMEfB0dG7LBll7CEmQi5m+gbDf3+VlVad/yJSsC3O98tDVr3ebJrzOmSjMXLc4LT8tdf8twR/xeKpYro3I6SRVKSmZmIc2PjmTK01DMnckDyfYj0bRzpTdXUWBTUQRGdTq1gy/yA0LdX3XLdhlDNe7bWIVMzXz7s0SXP4Mn0YvIfFp4LCV8/m4MG5BNq6FP3bEVmmfv9aVq/jTBLZoZPASktymvPv5J0AvhisCG69VccqtbtmrFzcPTU+pGgM9SYiwWRUXRzAWbvwqbrqrj6ta37gtOf5EOcUrPEuw0ScaZAm0HbE+bVMO7K9EI7FEgmEEppBLFn/xRb/mQdxcB0LK5XfOlhr/9TWJJf+lKcUGZrKiQ+Iv35qPPhhiJvvePd7/n6OazSIh01xMuzLnklsgLkqbjkmRzKvAARIxOTu45sMZ81/+3kECnZLclbZt2nanVBXiqlgrVu2M41I8eeT0oFN8eMvO5LUGyQ6sHC0BwIf0HE1HHihEickpIp2hmwdnzaAdMzacHfvH3/U//Wl/qSXUTNzubndXxWyGZ+mt6yI36QvemHIYTcHno3RvHqVis9u6M9BNGL5CjEsafSYCpZ3zsvlqR7h8e8YER4Tu8Fn1rG8Xj3qtF7ffDEQb/Km62zZlu7sxq61/Yaf2COyT6eR2r9DM/pxU/D3alTTn+DQECOK6ABo8CjnC/adVbuXwpGZpzum4nF5H7triq6RLnfMSPB4Y8Vc/eIf6yc/0o7g9rHfsrq37s756ePm44SQeMD1q8UjLzx7bNA7DNiySg3wBxIYo+Oa2hDjtnY76mDJAsoqE8UTzQ1BHczWnqb/pExqeQjT5Sa4q7BLdLdDssuQwCmoVy8t7UglD2aYXkwQLk4qUdYXpBF3uYEFa088bBnLxYBVNS+xgwnbkFxaQmNtvNqarsSiufvGqaJYwt/VW53OqODUwU5E2qeL1o3iU5BpFKd2QPNrgZciRoBSFcXyF9ACREu3ckbkVU8ZlooyqeGAePuDBFp19oCzQJnOun5ZxZSkCzcW3WedRU4UJTT74E7SLtnujek7n8pl+WkueBeR2kob60FZHkeWFnTCFatXqmoe2wCYIO+s39kOGiT4LxV8yPmgU5Dj4MxffRu/Xv1O4CZIPZpcQ3FS89i5A61PJs7qGpQcSt15FW+KB1jEZCrKxFcdoE84WQGeTzFYmxVcKRzngA50pwk6GCTHQFbbx571YKcZ3SVlyWv4/rh58gLYLtJqEdQPbSxfSFBzxhzvD872uDWh4/khxgYiVSI07UUFyay7mMcrFjpnE2DkCSZ+NnqKSg9pXcB3/4EqvqBI/2qbYr7lJYUqkSMFdPUwzejGG5sSSWJ889aa6cAIC5AbPNVw7KBkXH7bS+gWlD+9WxTs3YSwe+6HpzlqDh1olfLY4Wx5QAylu//51zNcpVUwDhVugF7l92MC2vndnKz2ZrZyTtvGFi7RwSIArLvtspAJkTU5JClHEwM6urdYQ6gMNAtPfs+vDv9vsRBnSBmMYMN7muao8xplTGPSipgnBOIPPvaCdwd2Z2rn/EmIWTpSYjfMRWcajpNm4itY70X9h76AwO2DEuxzpJISRWfHCZHNczHp8hrW5IKttCtLm7BKSO3uslpQbuJqv2E+U1COW48lj978RuWFEaheNx1Yj+IIEY7LmmbgpeSfNAc/ByiPusZ56JpW53ifoBP39zZ4fboP+43mUIYoekVdDK8kOXC8h8aPSIzleEG6fDR6GqzYsihsY33y2sSAXDGxt86CdDr3YyphzymMr4/2g3Oyssi8+JclS/bt3plb8D5qlaRbRm/cl7TqFgsUXDCjSoAstQ+xBj8GRdSPGNfx9GZgb+TfmTASKUCVMlLB4asUUNiw6T0Ym2FZUckRHQ5qPJoqjJtrubgNZ5W9VbA7RRhscAHkd2Jbw0kelDJZ35XTGKDviO8jZjnoCjceZGPGccxnblcnIgmShgqeMDO/snYix1geV18edQklt95miMIPknRneanjm8nogLid8ifMU7bE1gzeRMo1yuIBSKjx5ke5irhRItt8itUHXbmr7nTVf1QUEDu5Kv4KLD4tuxG63zTvyKwpU3Fzss+xeETLTqcfoKGDqjWrgr63gcI00JwPChC8Ys9soz3lH6tcOn+1258BUbLilDg61Hp8xis96cajbvW7zwtg2PvLfhDeYesoPHUkVoFgu8QQJsCynUoRIDrpMWmvdInOodoMuK7NWCuHItDpnceLHyZ+DJLJEZGumaHWBHfW3RVwNML7NSzPHgVQh+WKapXVxYsblvQosnes6/B+lPP7S/6ydUY7kKBBEL0fJHKClukHLB6AHDoBMHXuaiMwEbFdVd5vR7OystNoPhk0nmREvQIivLCsvU/nvp3N1NvuowO2gafDIGq6q7e8LzdAZhd2ldJj7lL++PQxg/FgV7DPLoDs6+ftX9HB9P95PMnP/5uhYYnbNcY1xoA7RuLlKuRfOfbDA477DXniOc0xKignQ1fmapqwJ4qOPx/3TIEhQbTaU36bd4k9r2qR4NjGQZhF/PK/E/kQemPvUS9dExbq642Kp0p6ra38RCTfWzYuXNejCtjg0wAf3HfB4DGbynVfp0teO7NeJPIrPdQid8OYfdRJ90AWvPS3Fvq1C85kC6PCvJ65HFyrlv9/MyLzE562UzJjc3LCj6tiHQVRjAilGEYpEGtAI8dKMomxNJj9HNvhptdgiEZwt+Bvd8Yf0u7fr6SAvj8BrzXDG+sILFf2cuXhmACbU2klduTxP1N8lWJLl2EBceT7b8q7QPfqYJbhCY2KgNpgbXTvlFpN7e7MY7cY+Xl7LvldpsgQ3P+n3r5WslLHrL/WJgXXHgt44CAlIJIMoEFUntCj7uZgLiUwrs3y0wfHf1qM8ZAu8WyfJBttIvpNmSsfm3ECMvz1x7vvWVPjn4woZ/MCYy2jnAH8SsPyQxUNxxRg2YuMXkcUH6FOkQbavXUo7+GWJMV4qFnzdJTWFfU41hZnXo+09CDW3INcXpfjQV7wy75szd5HFncfVXdhYkMO2QEsRCbNCQ+HYpBV86NIQgqtpdelaHVaB8sNiluYkIuDtYSHF7Yz76eVvKB/5yHXVuw4N5vfzAs84TDAZAl9H8iTmcuDmCnoK14qFj4ukk4tvRsWC+vDYoaounTKA/Y/HNMfSNt7ibprpXAtKub1fduSmz95PMbv5Wvv2Ba8pgnxfiIZNUy8RbxcMiFkjPOBAKFzwD8rhdokNJpguXOKEt4c4dGfVYoNm4oT9IP3pfDOvKSqjLyyfUNlGNCamEinha5c9ZD7oJ3iFudSn4xkwjxCJ/1kU+qN+8p9J4H9VjDUKGjiKCRvord3ijvlhDlJ3e5+o7Q9lOA++MD949nVOwVBRXGY+7UIWyWsNHE1mJoewrR5qxnSixs7I01knP20dXeK1OiEKOeko1lmmMMX1r928uPH47fn88Vqo4vtCkQ9NcivQ38fnnTbGIGtXTZvkDuegUkEJPGDwDGZvKbKVwFKUiviiKMHLncS49iDkaiLhyqJT+LlrUdA3d3vrxh2XG0MFPuI9jJSZgnifQaxxhE8kIajw4tKAUE0Hi1biOiZeWtKwjSLUg1uunzMmbUV95nMSdLc+U7DTDd7azRW3x+1Hy323O3K7v8nQd1IwqiGXsKoKA0IEMcuEU/tBUAT0AjJxijoD0tFlw1iV2PTFl25xaai2aTOKjSP5be9Ycs6QSz+YsNXbPECBdm6l3qXEK5zYFmfP0pyoTqmceEl5LrIPDQtJCdRhdhDMdEzouFwmUIgeShGbxaPYmn9UsSpdT/F2VOxPx5m7TVKzJIHwGiC28tRcwjhK3SD30OZ0FgGQ5A4X8OL9SKUptlZq+qoUr5y6hoWto1LlEriG4Sm25denBx/Qtzck3ZFo9+Q71+61VIzvG49pscJrfPAKbKS8qpTiQ8cEihJft6TWRVjowcgWvfq1ix2HexZEbMPzzj53vYh73IzeXjn3vZ10HolLR6FK/ZxxeJy4XBb3PqF3ngpuiyUXtyhfzkUH8d3UJpYx+OviD6UDrbIcnafMHMO22yF/vNzu+/2Wf+8pP3F9yDiIHCAJynV4NMuLuT5IwI1HDFh0mpECzPaS2jJez1orw++VmOfSTPHorpOAjhtfMeuxULRe4mcj4uE3vj38vLONB/b3WHB4ATnWNgIhP8EJLl7ZKYtx7+psIkighCxHyz7V8iUL87dv6MejrBMl3DIKejTaYO53dzfn3sox/QBTOQlKcf0bjyskyonrNhRxSkkSlIRMTPM+k2egv6zvvZh0GiFwDmMTN/Xr9TLRxCrrRHANk8J2fbG76TfvRUfh+7G7e4Z6xaI5N9K2BB7IjA0ToIXvvMLqbH5nqDGhsIqQFQstdwR5pN5efvmca6Wg8W4bt/wXYQk94qqL9viw1sK5N1Ro3w0k8kmZ1geHBNrVFqLuoKt42yOJsdCrZC9nMZPjNtcJUI7ywFNQksIB48y+uPeFycpjmzJp23qImG2gxWju3m+Usjtu8g8fPG0oKGklkyJXjxJW+tBiOvoaYRbtYFahMBwFVzWkFjhcbEBcYke8++WPrwNYhUCKaYulVebF207DPUA+3ubC5z1Wd8c1X5xo10xi8d1KLLAvLuzWCuRWsvNIKZSlqTLhx0XswaKLuWJRxCYMjlM+d9KriJ1mzhho/Wxbj07UZhGNzyvEwL/Mzrz7/bq0/xML9NeBZYW8qowaofkoZCwV5gc64QtWU3lIxqkqnMCnGLuwwDS1TOgD2tYe28zd3TCT3782Pl6+nPPBc5fdUVCx6BA5wR26LH7xdSpPUE1tL2BpTGJjBN21OqLDIjJBE8UXDVgrv/bW/WAO9K9altaRrjtvd2dbj6aRf+M/6K/umbtDQUs2L64dW8icvPmFCqDcYIJQC1azaI54dCj1LojNrhPFx1aO/3qZ7/g5NsYPNd59zhJcMSt22+3uPpqt/L3HI4+Pun7Z4WW17+lyLoLKDBC94neFyGKm4wpPMIoYU/IDI3M7jKeUUkvpOCQO//5r97WrxGIfnYT8sHnxw6SZ7mckoP3K2Z+mwg9/BkVYSx75rVmMjXaHwd0mMcxxxBZIqcGsLUQLbi0DojgNT7t0fUgh6a4rnHcTmjaBDR47Ck2W+Lg99+Qe1kZ+RFn5NiAqNjKGxqo+8io9JXg54bpYQrQdBxVGDas1YpH0Dh6vNsQNAZ3mVWO7xY9Zw0xx0+gBW+5EPzJ+Nmfrru0e8+FHpWbQzSj9HWnhOom7O+Ia6yyNgZd101E3IZJ8ICbRsLPXtcHPlZbi3v3aOREeEqG7zouxao8P6yhk63x7OQrKXcN2jBc9fhiDWu0C6nIKFX1HJRCcueA0kgQEY0dVWeUkGjYziUoxTjt70qUKcR9vMYkU/bj4aqFoPoSnPduP5UDHSgJLudYJYlOq54u2MCFZgfwa6Lbj5eUQCBA2Wp6LTOQF69qcuDP74tjd4lFhfLFQfBKrkus4s5sDvX50dCtRr3EeTSuYeys0t/rMCsQJs6moYmKEweMvGc9zxlYnazQsMbVjiYzuUIlrstzWX2FTjs3E/TjLlCHFTnF1UcK9dcW4/9/dgMUvQlL8LlkiP8+XkDrh60iNjMGAIRumar5lZ2fYk8Ki7Hg4wcTsnJT8nGYNie+7v1OYuanxbtIYqOGXXN8YO/fx+1LxanRf4M+VPCUneRKBimIGemgSjUIGk5yr6uI7elUatBOXxj/30UD6SEXkQB0k+toYyD53vg2Mb++0bCPtNY+qFe+aS1c64oArb9r4Kp2oLtIgBGihuhrSvI4yOWkrSLOzM07iUho0Ur8HAZ0eb1TSudpHZ8ESWkMhpLbcCsVTNVDeAX/yGRZB6FXohL3AzAvTyh2A2tW3L0JXbu2MkRAl2g5G0ZgDFYP80ClqKf2VKXgfbvLXfqCJWyzA/nkSbuKX6h0e9h7vF3d+t7bz59ZcfQliPwdKcdb8y8BVaKhnjjUHg+zweGaQnZbhRgVLQ/rB5Snx/ulRtnmLpVVzPXjCI6nt3dYuu31VOJO8dttRIhwBrCl4O/sQNKFKlh106Dr4aBbKq4p86pJlHBiQzeSZf2oo7ueHXbf8RX3m08ZA9Wu3tTrR3eOn6LDshpCqfAwO9J1+rThCwgiDRmhSJh8TczZ6nV2QyCowMRfScxcJvhTvgTTEbX1XLjYTX7t6TIfuQ2jyM92j65gp6DrGx+3lIyPvNVb9RH5Uq1TQHYKeg4pdM1hhyA6EA4x+59peyAcw1zsFIdDSZmw2YUsxThtOfNltFrfHum7zbvEnxa+r6Cj8frH0phC3S9v5lg5OJQ4lEsUpYJhDsc3JD3cePlgQPEcTRcMjyiKSeLnFpRtOlPiXduLe3+N7d43RedvWY9Ytpk6eI/n+Bf0q68efVIQTX2MVZ4uXXMbGJL2ib0vfFYMhNJiyMVJCCB8SfwmweTB7fopxAMdPfzyzVDBlaZ1FuNrUd7dDfgwl4uP5xqNXsj1fUXspEyCG8XXnuNrHui5FCCiK06WH+EULL3NZksEQ2knHdEA3Xhyz2WJJlCo44ykG3TYI8jSF+e6Qn3duXcxa7jfPfj93S3p/uRTl644rOyyfMTdm/Ay8YQujcC1HqdFGU7dDKnaZ09/74ru0FPdxsZTSkVwzgTaoHUWX8Pr2Wzd2F/6VExo0K+ZVSXKHY29GKoKkt9K7D/cBuPxCpOmJgjuH0sUhxde+TYZM49/jmIx5WZnZxzaOszb3pqHYS4p3p5+QKsHEy3rJCzMD8chjr4yBvNrtuOgQhArmbAuXdkledUWi10y+feFwv570xYJ+XaexgRptMHfQzPeLu9wpL/PxYufjGg/zCMf8d8onQGQrUGRSZ8yGIgpALKJmBNVYxdM33Z/rxP10Bd13FJPco5wmEYYnhSK398bH7R1d0HWRSc+Wo1VMnALPHTEeVotTy8YVUE2icjDIjC1EoyJIgHYy1faVnZKcr9The2uLY520Jf3ezVgsCbfG8LodM8nedh/PJFZ+dI/mHmVl2RJVMZHFxa9LPJgasdbHENlbKf4uG1HomEvAFF6YYCkddqB/k1jdT+YU/bWmQbeM1JoZyA+pFMPn7q1uO9slPpTkEddf8IgGtgrRa9kvICYAh4DSKw1xhceL1kr2dfy5qMSq+Z2vSTLvz8bFkXSgf5rHNnF312qxxU48Jznmc+/zyT8327OItgsC1r5vMcowDxlQQbQVtZeIQizGMYPPpsESRz7H5XXHF2rFmdlD48yn9MWbORGofnV5lFK81lr5UXI13vIsp1v4gg5skGtvAciSI/lOtRNMKc/QFWMYBBZFqxCSptR3E3s90O9mmF8nj+hS7aN6yFOaNv5JdSvo/KMaMVr0B0bb/j9QrCg7TokB/qlGsCBSIBNQOCnIkWHwvL7qeFZQ40ULWCewuttVPn3ejSFLF+G65h5FVJj/EeejB9P4AZ1pbVxozztMJULJQjV3HAFBXdwS7ZLxwiTyQJPg4zAY7iIl0oWR/FfXst1HxtWjve7WKWOgT5TirbvF+XVr7BsX/vm74z9rV4PbNg9DL+fBPMAH6AZFDmDVOoAQ+dhfxEdSlOwkTexhG9Z1xQBVoMjH94NPQ2LHxuY8aMCbn2eNSi/mcy5qYM7imVlwttquJZHlSlKVjssfxIketRNx5GX2rXGt+rff3zHY4/R6tNLkLPDOGSb8ey99xpCROw/d3J44hdz4qElY3ITEpJlTRrmXmDXMg6foJSUdNgyKb86Cw1Gf3St15y4xS0pWaYSg0+vRzdXivlX492dLtu6Ny65jKyQwBdtYKS++bkJArOJNkqiU6jZ6lUOek8BAet4tufXj6NbdZXZXtxukm878KlHYXdWNA4X7zU6UOuTn+IuyxzI4GAUnXIsH9Pocj4vNErcR0JZPuNe8uTP2j1kTd+4eJ+aOtWvZeiaFKGp+LpI23n7cLZa1B/2F05Z1uNuJcruF/zIpfoncKsJ+H1gbe47OMkjXwgsb6JmRtiTE7WJOxattlMq5Z87ztkPPvKr+SwcNxQXMTM1YIvo7f7t3SNiPdWkSUK2eOWjxhfQWC5dNqy9+0WrPHvqJqmhMlqE0NBDp5Ow8HHJsPIqkSv777ZK++KaZEzzdTcP27r9DwzAMz13oMx25mUNAs9SNaEqqL5dslAXbZ971F4l6JlaVLyuHd5RkWzpzh++6uDNzHY/QIe62Ho/v41Zc1vbtslu8id0g9dzX/97PIJlcndh54NXuuHpus5VVXfSDl8I6UolnVH9+CfiZsUuqY9w8JxM+22bUSG3foRRhBxCH0HM03WbpGkj+5rQe4lyT33OIs2aMHnUXWR23/bg3a2mujioqrENO1aRUQez3Fx6kx6lDV0klnQEogm+IY0Mqdhzu29Z7654EM2+q9fjLCdPLD3uhLlZ2Cxdl5m4/uoraWNRdXQYBiM83iVK0QKEEX6WStPwWE3aU1UlzP73DwQ/QcVeCew73zpDiJKfNbvEAA72KQCAnK6e9zYdkXxaUkcTnqXEpUymaP2yeCKIHgyN0QQLNnFabOuRP5YJLHMYRJLgDN+sakTaef+6EP8vqEcEoXt/iHhnO1EtrBjPHLAsP88us3q7skAmzD6yTQATStJQFCGZBWqA4N9oB++P95g63qxzXJ5cYtfi+bdeZJcCW3hsY/xFso2fFYlc8wH99NBFsGs+w8TwjdlT5xIT4YT7t+tjZ2zZ3cv3zxo1DYyxtRSPAqsx8uza10QjGtNvyv5R65DEEvlUM4n5NcHg0xVD0z3BDwGJftnZTQtZo7SRmtGtVFlaS/hCtaEOGz1GBQodn7prmRy12ZvJXaKDvN4WLobrzpeLf0zkjT2+M+dmuWEpyrs4edZlUKl7BgYwz8CCcMEhAK9zveL+0rJpT3lkAfbsQjXqB41793ANteouH5JQLdnfbrU3Q70VhHTZBwwBCA0eldr/onaUw1ANmnwRQXiX/XRwoEmIlikLG8mM11ooed/q6UmDz3NCfuHaocUqmRLgGo7iLzNzp7vJ7cYfrielo81/acAfhc502BNTkFSmiW+vqQ0zwYN0Ifzaut2yguziVSzc2l+97Yrd5jo7YFtp0ZyjFFbX4ZizuXYbuHyDM/PzTGeEdOnFk+FkxNWhBAg2wtlXSnklingViA1Nw1XIsylGJBfv4BvvqsKOxhZGtIrf4Mh6Fzh33u+juaMqZ/jB7ZPMNG/3wLPRyVv1+4hevklUqfsnOHoR+rWTTKKFDrsVhgXJG0wL11pbVU+NP015jx5Bvh15UTrNdl+vR4mkOzYGmZ/pyN210/BT/reCJjsTtlSpFhQWj8Ft6/FxcemuBAprHudkyca0Wl+afewL7CfgtWqU48KRYt99fffBu19xipskr4Yqml4e857Ht0rQZsl8azoa5gzWNjGkmlwQ/TyqhYQrQAp8wXGXhqVgtLpfaANkxx579yrXY9kpXdRSGZo4Oxv+eF+LDdo08LcUePG7d6sUlDmwtvLBjPJ4JFJWeIuxXweULZElGFyzNo6aZqXxTig3yiX56Hi+y5jZ2sruzt3gzi6tMTzPed+a5XfeR90fPQgPmqOjThxQ2rhS1Fc6yttN4FFbccQQNmCpKF9R1h3cD+rSrCL0+SQtx7IB5rRPd4HG7YHf3Y3gxqUQ3v4MohvGZOvTCNcpcKoi1HjNH5GqwD4eLzrMFwnOMEoytVumNhTcBtMK51Zx85KwX7nq36JfQnb/uJX2xqxN0xAs+lvD3Htt7dEKPfoFUqYApn7lGZEzNqMdJYxlrVBXKBP9WnGixSUY/1uTGgbIdRmLQMEQ/Gp6tZiJcZ4XXjEm3bR/c+KKrIHoio8mTpwomYnkSyGwcpG3Scg2fEYNBHvMW2XWotMOcMaVKfMsq7g422D3GCYe4OvqgeTpeZPkhfCvIRwmKJZre14npqEUjR8is58pP3cLdGlVHqyVzAI1WY77MOjvzWcu4sVbLeM7xaBnP65nVqNbgGG3fETveYKe8y0k8rn4usuv/uTkpP/Ep02s6Zge/t2Q7cmo7QsQE6eKDzWrShLhR0H8mnK+VYjEizRg8VufsIQumsq7nQyWODl5RitDeu2TaxtMD9F1k5nc/P9PbEZq8W83YuGUvQGBCMSsaJ84ZZfhngl9YgldmUs/tAiqmLJNM+ex0jF8owGJviRCsJzba4LCFLqvFu/5cFCkopbgZGNOfMYphon6GYcz81BXwXgkAUJa1KGY75LiyC9DCfkvL2riXwhv0WpoTKFu0pjh2yFBwOBDoQJflQN/vrhbLkp9eE16zdxrMx+4JHD/DIppFTFUmwjaUj5d/pqR+d/Djf4x1kuIBwhVytJOH4Mv6XT8xvH1t9YFuLXT/9nHAmsV20S2+oXFT2uDra2ynnOlwKCFrh7HtIE4iBtNVLGsmYWEmtxZl9cHKOT+y3Me4rAYqBhZ/i1HEfu08sOS7MsJ+FAmktmtgoJZi9aZO7Kt0JiVUdNyUnNpXsENx5uAO9tnWTAl23ZY3DkxBZFjViwwjXekmjgxUvhbu2+wccHW7quFY8mm7DdzX07f4R9HMLWc68PnpVV/0HCzOjgQkEcT6WU79SdX7dYaMhjdLCmQSu98ZSGxOxWLwUdyqo3xVK6JbP8ceQm7DtLXFl9bimxC4Md5NQ+7EK6JgzwrKg+c5ye758b6xFxDbxid4pxRoGHU/OsMSCBAF6sFsgVXJtRTfTnaeoOJutOoaw9pZUiSNxmTLj2s20GbUppD8axpFfkEF8pv9hTX8NAt9e+JuW6sIFQAAG9VJREFUYoJ2hmSm02B4gp6GNc8yP6fUmPClOHv+r3lW0RfeaBv+vk5w12aruyt2dzA5d6PH20RyTwaiLhSzNW4lm40jUIrEKTQwDuOZg3Uek6aL1peOWdxij6CTc1LHuxPdhNvPxW6WC6v/vGl0KzOzcIjuZcY1GrZtBOPnx0xH1Mt9gmBqcwd/JAMgpHaT2LIZTaXA6HVF6sG8ynRXWtyBoRIlnfIMC+toyqY0+eC5QnXrUd+7n/tFeXdw6XUn/I4I5MS5uY+vyk4vWpviCXDxIlmiiV84WNYoMx4k43XGDIJ8XMuA79q3ckJtF3tqSgRYEZ3W3P0TRYEGu8Fz9ks8ejQaxZP1Xe6XS093/BJVPlX4PSeEERMzrhJBkpskR5vZ8BxhVZd4TLeaea1fTc0VkDBcrakPPt14hK5h65484EKN0y1klWKmmRdRuO8NMX51i3c63F1Byc73TrX7pKWXm2KC47aFajM6sc4MZGLn0XKem4rfr+3Sl4OdK8zBSrN1a8FvRw3LvOQW32FJIZslucRvttC5hYN5fGLslhf5qyR7j/oGEqzCZiEUC0ohXUXhQ17LYvd3hCPKt0hxGBYdDEm0KbrVD4+0bRfV4puX8m+Dl/zT+bm3fR2h4wpPENOruP4uTAEqfJ4cu8bauwkc+brr4McPcdqcopQWP4GY/2j5slsLsSsaHlmLfPjSxAV77TgrjBGKayB5a9x05/G0L6bOmp+HuTy4kjbxF/q1mb8IJKtHOYaBFdhrWe0F11ns+ue5Gf8c3eITSHHsgXf3YdCBL4wT9HaVC7fc423bnHw0vwLknQh6rwFRiZLWZN4icf828wa6ziOLhNmRtBHqooLdUqNMqCt00dX+R6LcjrPddW1aFIKU4QEwBr34Vyyir3ruhJmJOvxmx0/usmZvnelXHhlGupX1Wh0HSRIQ8O7BfBsAJk57El48q2i0FW6+223X8Q0+Eb3MOaxrf6LBPYNRmu9HLU637SqzhJsEeyBXyAQ1+e8ixnzstwT/CaG8VpsaDkMQD+jKh+WIRm0qmM0mvYOY1fgX7lsIPgxk4uCgzSDdWwix394V64svCinuth70coJuQwf1rBXy+39hWfEbR8z5gQeTsrXZEVrjk3Q5Ck82UYOtGi8KZW5qEoTv6YHBIInYI/MGa/a6MGO/XuLT9jMSKfLbCU+vdN65tjUjFb7DCYAQY8LTgoBtSb9EKjHeuQpj8vTBvuYW9GwFonwP/sRdnxzRPATfdIT2zXAHfFlHcRvizJ+dsCdu259oxC9FcFdq6CWkjBnLaOaz8bHWtKp50hzXeVaoOOmao0Xalf72fqAu969dPMIyhQYkt7gbPlRQ83PRLZYYq80SEY5bCuois4d+ghwQb4Y1DEtU5vbjMSNhybOCHwvRurKryWCwUlErhBlnK/SqMpCK1+9eu+FKazsR1m4dHXdl4tpbvLUzfocEUde6DcRBv/bIanKeuUwUxoUnFkArxAYCBXcPTAKaOVbbBTM2rfkZHVhHTPFNsdTi6CZrk/L3dKCTAl13xvQmjir/Rbdb7RAWMYZe4AxUJeZAiqUdxsauHuxafcO4exOOStGo55I8BfNDIqbntroZI3TGKmscR5L6v86PtnjrV3dXZI+q7m7bl4EBLFaThAa45e4bIIUiP5phYo4VRB6T5NDUYWR+THqJXRJ4vbTCDlrEB9ILq/edl+R+pfFoC1ED34NOHlydpR5Hq/ek2sarMpbkjDeHZubnt3SnwHXOpNDvWxxNvcUZt5jLQhZ4jaXPMnhMsrRbZPSAAsFCcmXJfyL5wA8fTtLo2ouR1VaaOdBPY2eeh4GEYPzSNLN/8TIdGeNp+Z0sBrMwHSjDfnRJGlfOYHHmPR0L7ZgUv6QWUZVclPaHOOZQIkbXMBvoYsTLFzrGysqbJfP8uEza2Eht3O4e4g+7iS7veFYFrUQGn7iu8ERnx33aQtyyLbJXYnwC3PiqtWMCEF48lZanU0rcDq3sJHYNbYu7yS+lMv9P27Vgt43DwMvprXgD3cDRASQbB+ATeewVMAAIyk5TO6ySOrGS9ANPQXxnnPJjzMCVio+CVcV3dL9fGA0bNX9IoWdoNJbMpWKxOPtfTj3OexBcQ6HtgURPKsXKQuHzVdSyunfadss38fGyx8iY0YvowmNja5A2XzyIrv+2NiFoUOHl6Y+jKo37Nfft/yJZRtcbmUVHaYZMrsrQzHNW2SrsipqqEpt5w5RgMV7+8viQH2EJ9ff9iu6ub6cTx3br/A+UFMWjpuRBR1ajlV8jM8WdsNxL0DSTyuKXUGwLOxiWENKme0pFpoyzruRuJkkDqrCZTAsszLn+ljC+wXhpgxM46PbGiddLhdlkZh0njNlPcEe1sFe7SvlKf/eSj2LDlAWxLAr6/bLRKISCcMbsLaxQDH2JB0QQvBpv5EC/q8XHUYrdQbybr5DTLrhjgp55HbjrYexADuIXnjh1zvi7nbtwHoqWx4yhwUlYrU4Qy07jZH077X48wqqoDrtSQHEprftc3kBu22zeH5c47dHfWMwdO+Sp1LZ3N6TSZii2mnz9bgs6Srr2OyE5lChgVdmvE+biJPXitJkOmFaMBcVseWHIfECDRmcxvQ7/WTyxRFvvIZywruiOpIODNnnWjcQSZSa4Gr096nwU1Vkz8w8bYU8bNqknQedAYtI8WkbkixCzTWg7ywTQrPrlHBtnpaHw5n7QrPqNN16+a/cbir1n6rAum09mrmNIxA5tLP3oi3OIh9Ol3pZCbz8JjGXhACx4oMeUfvPUHHExOfgHUEyq92yNOt8s/3D+5yLz470NaXQs8MVA8aXrobKNI2sUYuZbddaPZ3fcbXO8YgzDU1UZRUETXMXa/mBaNvYgWN8HAzc1OW1wC/qWaPFZYt8FK5+ec36uxQlBRHL7Hs/DuBUG2cZjYAdaUGxzFPXlKn83t50vkh6dp0Brf1P7yqi27O6TifxoAq0b5aeF2Qlzote01h6tGtSGgOg90z6eBJT2wPChiZ+iWL+CA/J8oSt5jWIQiiEF3VaWUIDIf6pY5m9zu02Za7KUJpiaho0rK43EiZ1pIODE45nBB81KLCjk5o9Aza+CgZ8vKO2R26NFEGiP9kVkrxeff/qXgniYmvltlTn55ihS+jN52LX10XdANi2ybTpjBaJi6HlsjZutWO9ZImVZBXs4M41PUHwSE/dT2UsMjPdQcFsMxRoZXwptdY1kg7/egb45T5txXOX0zQRx7j1DKwzFvQ5ufk7QLd9AyD+hSCyF9+zCa0yQWSBILLtK2BKlQE3zrpGX68TgErsdZu3d+V59JbpVhrR1N1B0QscG67pe1sLShbA8aAjGVyCkJNKqM0YV4JmbpEUyaF4LYwgny56VOn4zvvitnW+oB5G3O97P8JYQi11v7D2KLxwgRnOuO2FDykCuFSa+ODKJ5SddiZxaXTM37uLQHsVGWJJKRJqkqjZp2mHrM6Sr+49p0yEVD4hRrehYzX8zF38dpVgazUdD8bLEGSGCrsdxr+OCNpkb9MZH9rZHGGhNPfFovkqG9XpKWxaypQzBctYczbwIPUsXWtpKxRithPCOCE0P3U9SFTbqi/I/O4z9uq74pI+7dDvQSPqWfiOsdaDXcWyDqwYUxktqIcWFUjuZohJ4MlN6MeiWHNEJwRoX5WfoaQs1G+JgOe+KtPe5Kj+BIhOs0KVV2Sgs8NMH+XPMJ8I8kBodLsLccuCyEQu7auOodZrAcYW9MDOYk//YhFWEc8xB3Ktwm9mSaAJrTSISOjZuS1PCXOYElZ8Nhj5DY7ayalaVUMkMNBRvpHn7NX1ennp3vgJtKF56FEt2dxtWo1iNg6nNv3bDFCnwLV2mXoF22HprdBWzqdEIpeukalVKWAydZ3zg9SQZxszSGAWKlakYLWidoqCPImJnmVhil2nx1YNl6RPAHW6pNpHi20AUS3tUam3K1jaB+yOnVlnzBr/g3Clgs40Sav2NZzCzNJPkSyz6fH52hhSZeGoeSkoYe03Y7dhmpB7W46fX66LlkxLQi8pFjDaefHHxgvwxiG1Q44m2tIRxlRwWdXOy2mVWCzuIvQmSwsw2P9kKyIpLFkZdnt4mBMRM5yjpdNatMCbStS1n6DIa+7bSQess1gfLuQ7gttixmMtYPETe9051uxNEGFcGCpyOuqSrQVmOTGy+2yEgTk4lnzVqLpx4s7+gBN4l4lFYptYlaX8UoVhhD5y1Ii+QyZCHN3r+Yvu4XRmz/G2bPy6GaiHi2inVycw97IwuhmPxxJCLH4ZiyT1Uz3yNlAnws2pQrb45PZPNA3kQoRx6SQaMWUjpfDjfy2nXJIvmIpCSuUyhYTHNRVUPIMUWUdxCuIcRv77hKZ6H4BcPh/dXYyx7P6iiE9zDahQ3l7LS3CPZCCwbLjvfhJWRlRk2T25/b5KIqDbuFCFx5LXnE8VMzpYFxQRjJ6GLTxpTCHOmdJV0btuGt10g7NEn1Z8vj+5tcFsitX2/5twFix6IJ44ha+aR4yoQf7TkDSaFMtCkMtH6BncNJNsejrgNVmWU14g9BIdtGyfQ4hNm9wsasAmWkfRZrdg17mzsyuz8Vul4eR7+ebVoLiXMfb+2n+tFevS39WKMtR3CD3SiOGdfN08aJ0DGNYEBL1serStkOqOVmDs+sVUzJ3Si6ZF52TXzTcKoIKNbfETaKF4qn6KlTPBbRbVGbY5+qBC2PK98LLEQ5N9kw8Uqnzusdwd37FNX2YflsxJGC4w1iIMHzuqfc4LhXfCRy5eEb+ehmu2M1BKT9FvmwVcyEHfXJmbfSjH7Ukvw9Ox7j410v+yCRTt7S3S31Zrd/qMUH2gbd9wJcc3hY21eM1a8anish57Ze/JXQFCceL8jq+XZ87Kz4N+CY4nTqOfH8xeHEuybT6chxmarOo7VsLLxgVKFHX7mlj/TrloeYR5l77zy44VOGFCMmG0dieJVJ2BXrVOouGDDqxoWJkQAYaZO4bl+4wnmxENAYlsYndEhHnkqFQddphSEO+JlroJabue0mfRDYXN5vNjpeDVwvMdt6MXXHZIwya/a8ziGmVg1agzFXm9ra+cIMVC/UE8NLyJGdtizaWF2BS6XKBKPJhDunWhl/5z56AtGrfR0OeMg2qUgKf2UYOXRcW77ANBynQMg5eq/zmWO8MXHrc3KO4pzcLs2Uaguw8ybe7edzLiwL6X4zvkzW5ndwyYpB/sTyT4eyEESzWb0RCaxbU5D0UxvLdfsr3ZrlnDEdaMApfgg0NFVi0c0liSB1phCLGxITalZMsD61c3sBvYvl8t7hV/mkaZkKM7yD8MZyK6affVEkBUsAcrFVh0/2a1ZLgtMbRQzVpQ9t7vW2QY4iuiLLTbO+RsjTjk46tRd/VOyM1LMzuFGPn1NMy6rOxBHezm3O/zgrnmeXbaxDW2qAh69A+FL8WJ/mQoWbfAf94ue4AAT29bS6u0lD900Qg6glqE3vd+5bb6VDbwK3Ep+k79MHH9woHz+5PmlIj9S4EiCQ86I7WaB+KyM/WhQl0Z5/nb5+LqbFPISG5A3ivO122schOIwPVjVUeDDM6L1RnWb26PdyvKT/DKpheU+scXraflK/RnnAK4ANs5Edt1SzCBVpqHivPKlBCm8d8ZXns0b9Ynlb3C/s8sce9ytPhPUpge96CYWg8n0HfiuF2Nns6X+kJx353dVyup4z8/FrGJGteYJGn3Q+9TnI1PJ6jD4TNTKpjFelbfXSvcX7iH2OzZG8SpL/LduuniEowjUKlYOyhoi14t1q9lR7yjY1X+fD0Xu8+NpW/3FR6iE9fykiI+AoRXLrDFHzUEbmtE9lWCvi501cqamrvv7PrWFE/ATxzE2gTYbBxVSoDhXQ6Y+2Fuu6hhyzeEx6atBaukq2K1ZxbeAVQEt7kG9rxiK+Q0eQ7EOWEMWNhGGL1w10+a6yyer/svTaYhXWmrFtZt7HVJp0wRPYByCY8GczQjV3sJq5WpepBkee0/8WRFvrIDmeI1kcFfOFPn3yLax8gWfj1/mjCu40ogMzzPCaJIxT5Q4KFY+/94vf3uVhmJVTRnqKCTKhquwGc1stWMxtefV+sagrbCxRiBuejzU9sCPGIpmdwEUA9MFwAaK5c1tLDei3+BXROqiMeF2NWOU4kwr6JPWnlqYLO/oi0ADTNxIE5zDJlyOYr8S8uwK1V111xrr4aZ9mnGaiSepEPUUpyFeQA2u9id+uJMefl9FBLuuh2DmmjN/Q0hM7KPRsXyQYJuFV/T3r6XMEb4YUZtuh62deXP9LymK4Y6z21dhah8aas3PqAeu+oEUuRZMwLRm7i94Zob1yqYmc9EtAlF/yWZFrY40ZvbDkEw16B076+tUGxFFpwE9CMWHLyUwhqM/fnFlt36E9dM3dFf59lMKXqOZ/IDbEFhTCPbMyojpij1t8bMJoTepvJ8hTF7F1DGgC4bHOIrDr6oCCT9c6el57s2bmukMzeXF78M2XQ3pgub1fF7KymgmwJrKWjTcUEC7m9aOlRX7iUyiSZd9iX5wHTBwqj6RuR7PfmLAlHwoal4zkH9ylQbmtQb3ITiSl+Kgg+7iuvnG/fz8q3xJsKFBh7kOy2f0SnOV0ENRXPUQDH7kGqrp1+pxJ19UGu+LbYe0GXj1isU/vtArq0JTycb++vo6z5z7mcaSGP/E9Nfq3wnHcicPnlvOHQ7F3Mql5MC2DamWvkhoKO0B/A9xmbtnPzHAxGvoMQkRLAILtfO/NLX/3uuJotO8B9sXnJUkuPpiO8MCSAKBa9w1H10N2hSgjbBDiqXaYGH6HB2oKzKgz4UQ/f5qfmK93Y7RjuJCm3DI4seqpjUbvwjmxpv7EL/Mb3f+rMgfyLCu5y9B872uJ745+qAWrNwxsVosMyxFExq6Vkcb1sWsCS+G+SzR3pCZ1zggP+6409Ul88cYJBSnzPnIoR/Cmbjib4R3nMU1HpSHapodP+D3uFpZv2iMv90rC4cpPwQmKuOjOs3OLwcD/P/2riA5biMGKk9w5Qc+6J7Kj3QA8QKWXgDx2SGBbgDD5UpyzJXjFOm1ajkccncbPQ3MDDiEi0QP3NXldRTsHN9DFwfuYUK8duvtzmGxYtw4EY7IwizA27HYEgc/uvA5TWGXUBqOjcZCe9433xCPTrrrve/OaDSa0K+awc+ZJhbPrtc88OYHdLtd2egoV7xm15HXCP5eMYYH3X5DGUMSjoKsV/Z7D+KLqt4K8VkQM0vTuORVoEW+LhgmyiU20ReMkXxVFCw5kGQF+KKAWZu13AqhCovDBduo32ixTT3Ma7lbeCaXnePTlDx3j7hKhLIQEQkj7Airc6iJrEVXcoPfIjZhM4y7aB7i7pLDpRRA0CdjwTslJporuylkAnWLxYr3Tm00hbwOOEta+8kbpI7m9snb1M4ck+K6Nt8N69n1WBcacWX0q7vC9VrORBD+LeOQV/I2BkKW6mRG0VzxuC+7HbfaqcgBj09iccXG0iQ3Ws+C4QsWG1afDSJbzK4qZqcMsAaRS7qbdTTnWLKNYDEei7mzZZEZfsAR95Yw27QR1gHnhK5/q7n7gBkhR5fwiPgyon5DYP6Kfn2Komeo2MO0OJ6mG49SsSaoKRiyJOk0cfTb9tIicaaZ2pKQIczeRle0pCdqJf5+8hwuR9l0vB1MQes59MMX7YmGPWniqh6K3ASClPD1oCtGHIiam4zPftyVCr/gjhSfyOJIhBWtVY0jEseTKaRUOJWzYQ4OL8npBdjlUnDKp/URQhI9JEQWoUf0686bTEQLZu3t3xTE91YwKRRaQyfGwDJkfIqwe0Y0skzF7NhXq7s7bDe9fy7EhpelHGP9K3Ivpmbhd6kYw1Ewt/JpFc/opbxrXRuoKVPIezsIlda8DyWYv2ERoIdWQUE24fCiCUNYK7LxGsJCre5j/M3QUpsvP4bmTC12zoZPNQkGElrCVFJtkjsJbzvaoxOQxPYHJQ1hmrutLx9OFkhbg2KTlEUCZkd3irh97tG6x4QDuQ0pORWNR1fWMhfTjqTiJIht6OkhRIxprICBgkGoElMheWWPLmv0prDDGUq0OzMsgpFFeLv0i2mFFVrZxHrVMNeYUHISO6QdM5LWOjYZVuom9evF4jvJ/g6Px2ixSQ00WXTYXYSRMCQFrxCtRB0pHiwBOjgn/iN7TjJtzItpjnAEXE2nc7zLDQUlfJ+HHC1KoRcN0HELC5rGXP3TxZBfNpj2nhSfNLHUuRzIEgkSEcPKjD2M6XDJ78BUlPjDQkC+kB3kBEequGv50GUfWwdFJYRaFi095wBW88tuBAnHCA2OjgaDJBPR+/CcGhfnbGn09jjlBEpalZCpJsVTRiNCmUY0n8qdZM9hWW0v9nzEks7J9AolWYfg4YEDTLYZgvreCYW4Sxcb1gdnVI5jtjMhRkZiYqypG0HSHCixZKZ0LYAaGDUaNbDiHoQIfJV+Dg1iYRMYirOKlg3J0rKpWAAK45AVTg5jVroLjXalkepqd4l8MostuzeZTqBSz8MxDCzz6ace53CBDBMgxDXUWQ8IB3gE0nDbHxP1uTJfGrghDGSp9liXFWynF+gsFj3aKj6sCJ0yZQ8XCt0ttuI/Fe6JDA+ilB0o0JQX0pM3rqGeUXTiQwIa4dAskm9UKE5ENq1mygTSZozmEFJoUKxDlCijpJduMWWCpNIv0+Jb9xcw5HSAaT4rT6gsKNNa5qLVk1aZXVTT6u4g0UAqSwQ3qxmjEtA0TAjJiMfdh9mznaWr0PLT2tWluq4V2ij6A18KcTKyxAA7miNzkOxmEOAagmtM5orxq0YUPKyvLq/S8Wa/snUEaBpL7isApvZnG2iMhng3mPGuvmQZ+6YlPzSiyB9cSEjLLtDC2woplX1+R07QlCuxbPStqkk1h/zBJSQR9EEtmQnVvbKmrzB2S7X8agWE6X21uddUOX0Pkoew2Eazamd2W/2plecU6y7Xo6tJlZuwMVdD0TRaMWyfT8PoRrROplIDPmEskhSgZzR4XOEEkkKjPtoeIxSyI9ptRJPDRu2njKMd+Xt6c8ykPLNBPmi3Mp8OfG0z5cV1rFiX94Vr9lH5AVoTDlLfSNN/pE+QL9fibLjHQYe9P4Ny77i+90NklPSjz7JmKdNm6ZytSKtbaZr2mrGnWfEz28NYfCj/zd0dfUE7vIDuYyK9Mdfh9ewQ9Vp8ah/4lMjbGL/0NxUFju3x10D8Ed711ew9h/yvxvvuRZGtdWmp1QFOOx0f1Wb03L+AxXqUK/Q+iDre2HATX3xgKj3e0wNIDndsP8pymLTXQskfosNXsfhTUJ14it1Lb/z8R9nH17f/MsQnpB/9rMl+xFD2Exf5LSH+vbYL4gviC+JruyC+IL4gvrYL4gvia7sgviD+v0D8/UWej173947rP3+0PxZxr3ZbrSxtNcdLPI+Htj95RhW91DnboVb1JXd9/7mdgA3vWk3JL9aufAe9/JkrxH9+v7ZHbk9PT9/+vrYHbn+tEP/x7doeuT1d28O3fwCQdoBnJ42qsAAAAABJRU5ErkJggg=="></image></g></g><g class="png" transform="matrix(0.6679254770278931,0,0,0.6679254770278931,275.3056640625,98.95823669433594)" opacity="0.9999973289049107" style="display: block;"><image width="597px" height="590px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAlUAAAJOBAMAAABsOe/ZAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAkUExURWyO4ECR6iQkJUCX6Hpv51eT5K5T6AgICAMDA71Z7MVp8c268unKkLYAAAAMdFJOUzkjASolMicKGDI9VhWjmXEAACAASURBVHja7F1Nc9s4EuVhcd+KD5tjwvEWrqNlodbH+QlIUKiZn7FGpQr6B9LPXqBfNwCSUJL58K6IiLYlWaYk8PF1o/t1g57mx/ad24fpgcEDqwdWD6weWD2wemD12B5YPbB6YPXA6oHVA6v/wRa+vYv+P3/+n8RKp6+QN53u6Rf8GuhRfsxP53vaRZ6m19FTGiPFX+t3/lvAu+NB2g9P8K75jXX+md/s8/Vfh1V+u3LKcRj8BA9b/pBvZxrfzE9rIQqNNODZvAezR7enWt421BNE+/Lrw1t//p/ESmNkM5/GgDHNcZazilMolNCaX8BnbWbagBflzzjTQlTNG70q1id4N3n9jE97i88HhH8SK5wxzRYx61norOVEgcVp8PgTTm451hCE5Ixz2Ysf4ECZByGEYuyhGGCY5R30m3w+vVvZ4w9iRQfCJ2TW6Z2EW7xFPZcRER+CeJGylbPI+6X7sHJRnU3rFbWiZsLM4U0+P2oB61vTylewCvNcXahOSMU5hpjeOob8MI0u5PGmP0W6wwjyH3EUefjpccj2EulNouyb34ee1/R++SvwTz2EyK+V397o8+cQm/Oh9R/kVWFvpGFiRPXkZgTzh9LB5KOINNJAA5SjoPFgj4BR6cjox9iQlUau8fKIT8lj59fSoYS3+nzaHxNC+Aq1bmLFsy5xnc45DTKfDgws4l7jnNPRyfhoQHnf2DwZtUBNw6UvOueRXq3x1pktzBR8C6UIiTf6fNgqYTnXGfJ3YFXsWfMp4SFpGS9hGHAIMmyML9LhR5iOLscc5JAjkwAHHUI5LAYh8lvigLUcOQ74L//8/EZ0O2umV/idWPEEQ+OLMoR8j1Mqw4my0RDkLxofL8/jkCIPXMMz8TN4WeA3KmiUjXfEEb3F5wM6AV5rieR+D6++PNlp9C0doVL+9A8iBzxNpuwtsG5g9fzz2CipBNJkVb6ZlP3bu3zMmCE5MNXfjdXT8JRS9J3Rou3feTaLCC0QzH8vVr8MD1SyPmsTVOmBJXqdGCzMl6E7GfawGhyqhBBurC28UupXAos8PdKB78Lqp+EdOplfhktlvHj7nF1WjBy19pLpPVbP409/Fp4dxJqsV16lh++IWFGyhu/h1d9/AAsEUjQNKrAq4fUJxIqwwk5IusPqn8M79QxRAYu2BFUCS/0LxOJcYB9k7bCaxjdBYMZuSmWUfEbMKzHCSMn6N3n1YewAFKGCraTiqdCnTfnPYoRx7iWGW6xGj6xoBlQTnDr5KfJVGbV0V7x7Dku/5dsH9lbEJTJB8Mr6qbh1RbTy/l1jhLpWZPpYjR6xc7CgpoTN5MmrK4IrmWG6TVPh+Vxd1ld5pQePFqydSlylrFJCqswpm31WmKuWE77urz4M7agQh4JYmPg4XqCQIUOVvLu+XC4AaxdjrbEa1gQRVikFbWHyElllkCw4RXB9motKOH+DV2OLMDQFloSZUMp3+Z4MkIxQn5lYBJe+idVP49LKEq9KdCVoASzP0yDNhBd27lzhvoXVf0aO1UlfAGCcLHsEoTBD3n6dz5GJRWKyvoXVL4NaHyl7BJPoC9ll0QTIMXvZPqWgIRHrwoLDbRscWoVpY/Xsoege5meLCXo/Z3/FvIq3bfB5UFYpChmg7BVxz5NDF1pVcr07Z2JdOMRahe4tVuMKokoCK7gpCda9WuMErK6JV+dS0b3BqxEjUSAk85+tmpUiqIhc1bN74z+fL9dELIC1nglbrAZ07VZyZgVqkdlZ9lCqRUm20/l6viIppGLhj4KVQqTQlCAUp8teTLBFyyVamU/n6zX5q54RtljZEf2UWkl7JQ1UGzdVbHDJrJKZMIXuDbVarAaUQUWCsUVZl0gBvspukEpb4tWVZ8JNgNVgpacBwULrQssrxOuqQylDZpihSlNhBkuvZfcGq+fBgLICUzv7WfHoSm2jhQRWJtaVeHUWh/VD8YozZSrJe3+LV8bxnXtPvJIEuhWSR+WVLUkgpzWUykxtNqPaGZDMz+SZELyqE2GPV/rjYEEoSqZQ1zN2MDvFtLIdXuUf85LBunBSqFsNa0xeWVVj9qpU0SzoO4kNzX/OOfohqK5SoQhd3x4+juWrrKQ30uZBSEG7WlHKZZxghk541Tj36rBG5JWFqKC4FC+yHifNfp8vE2COwnbjrlc2wjPrfT0b/DgOp9BdJVqV4MRhaAcnQ7xKk2AC7KWBKq7qqY0NDsIr7q7isKrqVaoA1eY3Ds6KwUq8MuDVlaWGVmgYjle2QNakNZQre7pRu6zGUQxKIUO2w/fg1fVc2tY6McPzIOYHgCa13nxth9nYHzt3IlX6eQ9ecYUi9HPnj4NAxRUb0WBYhKnS+sqhYx5MIGVO5RsHrCRq0GFQf6VqZatRFjxHWKUUuIrYs2N3CBoMbQWqi/TLjMgrxQ1DdipuvWpVe2GBzc8wYrK9wLVfruTc89KvDq++HB+q0oiWAfNtJQLZjd3pMGSAsEKDyF3mQW4ummswOg6vpLmf/ftUcIK6rvZ6VeEVrJDJ9SKxKElYbY2wYKXnL9MIzt2CVdarpsXD9vRiwsk5gISbBNVyLQEWJc+hJs+ClT46r+DOuWJalRiUIJTfIWYYKopAiVFLxmsp/koi96gHmwelEVSVZlC0d9gK1tatl1iBHVWmWMZLaMWizFwb3QtW4cj6la2SsWqCUPHqVlocd9kyR1aZULBGx7y6li6sGrgPMw9ChEmw+QmMEkqVnj27JpVhDcbAAsmvL4lYVwmwaCLsasjhwP6K/BPlgF7BXSmIe1zZ2isLRa0yjpFyC3n2yisE7nke1EPximzQi6jnMRf2jc8hs6EE0HHCTLZnANVS/FVEQtjNnT8e1vg2a5EmXg2hWF5oxXXDQijLC8aIdJzdessr7mmIzaVAqg0ecx5EL7aq4l5RixEn9Hy6lwSQs0A2vgRW9VcSNOSFl4PwqqlBSJN/7e0o/Xpqa4HCK0RVhlwVOfb0y28FrDN3jep93P58UAu0vM4GGgxr68r2mzsw/UFdh7fynmmVbpa0bXm1z3HSM6+HtEAW12tEVaoQTZ9/27NgDMehCN1zZEXBFczQFH91uc2rI8btSsTiVllnYa8XKaBaClduZPJbyK0vApbwqsQM9dJFR47boSzIKjcR12Wd6U6yMnBXRtR1iUIXskExwWYe3AajB46vLJuhWpkgutbh0FWnDp+DBobJEZ2yBWbbSzhlbi3rfDD25sEjxu0wv6l0eMAGJzSvr5Hi2hZqgEwvDkKZV9kC8/bbdaUi40qTx+YVahAiFitoC2rfBLqilZHWofxgEZeeaZV+wVxY9StaPtHNcQ7prxioSZa5iW9HamN7UqiIe+TVOQckn+7IXS3NPHi5rNenHrSfgUyvsCqvx51kyZbalbYgEhuW1ksO6E0TKrARuuqvpPujozPoQ/HKTiVQ4ARQilvK7rR1VyMGV5QFBKBGoJKt5oNcIMyX9NvnOK8HQoovSAGfXrusbO3ZU+u+PWkcYqTECplSlDW7ddwOYhFU4bixqC3N2LaqnyiberUvbZkiggpUjn26g7QAjHiT3o9r6XHf8SqFXK/HIZWdpqpV1fU1vRTQsbInyroEofnBkqMqR34dc2DeXkp8dV5DNR+x98NKBMrL4WvXkLd2v8qGFWPD2bIkN0uxvMZbVV5dLhWssNMZDsEr214/zjYX7lDK9tvQBCwn6p64dUf+KoNmCqkqr0qB8Mi8slizpeqFhqQnG2Go3U6ARnxVptVSGhcWygEzRkY41forjhlC1B1/dYCYAVfOseWKFPmKMKzqWV5m6rfaHlFJqoAJs0UcO8+DWzNcdppMOGotlefApglG/LrtyHrOV6+O79IQQ3G6WU2BK16VfLCnX90/r8oaeDa+SbwVp4J2n9c4J90dEBc4VKDJz1Rtod3WtQnd7f24e39li7hnIVl5Ufb68YLzrkoLtbjFSkzDK/JaBTGBioupYV93Th7s9f55xZUaXGRoapqxZdmW2i4aIaurkpWUIBaW+BqM9rzi1QB6Pl5tQiZABomr8Nb6m0IMyeuekmSyQuYT7hfh1cplncym96Ob49y3v5KO9VIypXCUq1uS/Nm1+iK1iGqBUuEiPwWQ3A1elYaGng3evb9SbclULtzBPXtqFVgVrZhLymjaE28FsYr8+84Al6el5oPlGjzHmgdFreKZsBHXle0tG+EqIFBDALrwNGhEXK+U4genhNSKV+za9aHmQStf66Z1K8tstlVAWWfKSyGEVdUEJVIwG1I9nRpeIRjtrF26Xw2Z+4XQ4tgUtlQllb0hFnvnmz6YYoBEK8QJpkWKbl5WsWg43hqT1UKkqn9yI8w2qDK8fFJiUPZVrQG6rVs/EVJPy7LRr3r54N2utQRK1QSbrvUirqttf7EkM7W1qjQtkKzQ2U7ZANPNSw2vOMfp9Is+36cByjWZ+ALi1QJtx/xcmQO5copUEP4J1mfE+NwerVMzD2ItvQ7dftHXO/VWaPEQZV16/HeXGFo5K4nS69oRbkVz1GS1bGMrwHRq4yteZNJd53WXvLK1Z4FJ5cuSJI9riW9pxZekcLLIhlWYWnww+wD0ib4SYCeZBy+NJhP2NfrXe/RVzCsO1K3oVLarrZs9r3J5ufgqTm6Q2Oyc1lP2V6fl/Sq+ov/HdJT4ypaqFrfqlQvo9C42JE2gXsT1fOUhhonxcWYTVmWnfqIp8Kn1V7hgWIjd6zO83p+r4n/2U/7vT6nB1wKE2qyvwU1JAhdXK6a7RLmJq04UMOTvbY2+E7ffny4qlxmS9bhY3s3lQPtf6q6gNZKchzYMhlyLuuS6YSDXLYyhf8J3NBhD/41pPsgcA6GZ/OytsvQkuUqV3ct01/RsOsPsTcjy89PTU28L2s1tNbl6BWKXapVYPbutVUuZatUqSr0y7cFNXh2Pb8/wOZHPSVCor4RJTBdXoarMY1muvh4sMFifM2o5gPPHnME/h28PaEOA4YMhdq7eOJJI13lusuWUqDuY3+OkSn21okrVsOgcrPNKW/t2eE1RRl1fSKoT+8Hw7AhcYcLajsIQLyIdghItCmDoA7V80xFsoZqmTV4dX1MEMXaWU5jNeFvwZycJU9Wk0nXOJWbZV6W9Yc8lUGPEEdS8IjeLq8e3H6pe8dQkc6Bqt56NdU7uFXtFGCuRV1FGFRw95wpsELTdfq3At4+5Blt1fzk+bs/GFEbkMGCLc++fuiJiECQIF0pkELrqRRBap3IlH30P/oSzmjOPcxh8lbP6wVgpNj8Dw5atSoUH3IgvNv3SQhKPFL2HDYVrakh04oCNz11e2d0vx6tX4YRdbp15qtCg63KVxOur8hySzJlSdiXzBOxK+zjJNTjyJWhqO96Dh84rSqrKWaUeOlyjerRQ5P1X8bxZvrW7RVKYtC5Yy7Om1XV6MI/DnFGjzaufbIr8ej2ut2HOjNh5Lp4t/Jle3wOhaiHHjYhSMLhlGNANYGjcQmxlqsVpHAflRVvBenV40Zfr21/HqOhkTM9A3XQiqufzRahT8mp5KBu8QNGhaK2ey4SnGCpobZ+jJaEi69rrq+f78f0o1YqLu8VVIkvb6XFV1mMbMLrAdZYtbI8gvW0awDJ3YDuN509Lyiy13cFXPw4RqcDzEK1SiY2c8FWuN2/ivDIOOpRc+FkzxvykmQiIEl6ntBrGVWvi2O/BrH4LGMmtAkjrWoydpGXD7MICIFiLLZgKzcCyxaKTho3yqtfJWFOng/Wd+QgiwYQxlhazJ/Io0mNep1WEjDb15HpEiJRiaHGaM2t47mfDr1u96OsheNHM7HrYInbH7aSAVi8sGmLlnrQhKo+N7PB7I0DWyIzMHKlOJ9Ow6NtR8yory56DbcrzAjzvvQxvCgD4KI3miNn46LF8Iz+ZJ1AM7ek89HNeb44f8lF0MmQhQPshTp01YRa3obWVI5z2mOZbwtR+s9q4pC0ZymwMN7tGZNacWOPTWoDlzdH/OERW8fZJshBXUYzYLnj9U5kjWYIVycdKOBmmYnj2xiIGfuRERVjzPwwaqrddD7qj8AzZAAe7VZgFDKEnZAoeOoUpURXQFp071SGS0rNXE792WqknrNW/nd05+usx9AzZsg1IKx2Rxxo81yfNDOeyUZrCqz0hw8S4oYVpKVeT1X78+gPmcQg0nETxUTVUlS2aepteImUYZcHXMWKgskaIj9eVi0K1lCvQfbY/+Gu3l3owvr151LPkg1yaRM9e1/5DCX3URtBE7jpvDmJMW93xJEwy3YvjZo7+9cB5Fbor8XSqKPSGb8h755BxPIGtSN0J4bEwBEco/iOely965XzM3x+N9DN+MtB+XP8QffvSm8hB7ZoyXMD4QF7aYXyv75eQ3uV9CDv2cvl2qe/nbyndzt/ez/W2/Fducf7iJ/W5nOeYxfgxcUfnU3UybFz7Z+jb2djxZLyaGuXwFE63/P6Un063OZK3p9zw1/z9lG8sF0WRb5+negOB2mRZtxlMfMTLklb0ab/P8UN0fb+ae7Sv2X55eTusDhkGOydM4bzLnq75hNb3OffmxFoy7L1Vs1SMvnZJs/clRu/pVi5lzrFLSXVOqzl8l7Kk2Pyn5dfHkmZDtHKGXzva2sPOTWR+8ciuKfTEGEks8crzz3JWl5p/oz9/3+BZOB/Ip+Vf5vRaDm39++NSyyWWy3wy68f85/ZxucRYPy7xfIlPna6vccgbru/tx7GqFHC6vJyV5As7dZ2Ke/u/QKP8xmnsaUxLkWrU6fyrRAw7M4fazy7JDK93Bo+TV0FAKDoTPHea3QhBDkNQKtFf+PZLoieC8OPfPqnv47h+yEfRM1BNUkc5sFYwqMgbZYzM6gKtL3miwCrKmOBXgZrkkXju+zhuXh0EXwXZbSNyK+UYfGUMk+wM2tvbj0ckREyrjZxSdqLFUHTVx3F1MseYo18oUbv+LrNRRd205sV9tohjqBqLC3nFE81xl/SDQIYfO3Yeh/s4LwfF7YGfzCfxHTKq0Lxxs0pqVp+MPi3q7DfVbpy+4mhAJvsuNLo+iGtf3xyvsMfjdtoVGFoLp6o9hbusBc+ZtLEfKq1LyGYVJFMzbLunFx1bT5X5Bq1XsI4+ov4qsJo2iPUXS/m2MzfC8NHbrxrZPxWoxFGK1oHIrVGT9qCbskF1Mp/w3nk92Lwz9eN5npIV2XBK27ZuUlEd7fwXoRSi+BqzNBth++Lum6BtoOegaNXesGn9cPxVMM2bystt1P5yG64EPysYn2BGokZxNsZQ1869N/H3iCiNVlsLfwZPz/BQ/ipzUzCrhEGyahOpAvtLCpWI+AikA0mts6lER/6oFYuTa47becWLvh7rHsxYWGbcZ3OLUnA2CwtjwBegrIuQm5BdPThgxWH4lDzmyt5Aw2DmJmSb0NV7O/947BlkxSMwaMg+ta5u4jQgYdaQRKAFmrgpJX2B0qVaqQqLAnfubIqub4fC7dbOkZG6ManI2fGflbzi8STSGHOj2brp+C9AAlIjNyVGyCCbBES9WD/38upxc6kBpo70/KOtSZCFZs9+XRclVZw+sWmCp05kwjj6r8AxyrzEnFKtj8pimWjzivxFHS+L7w8M1eLPdEIvvvIRzDn7Y2/qvlfR4KrozbP1F3SPyb0AWc9Owj5OKLoGlUP+lP7ggfwZMovT6gk7O42VhzdzKn6OHC1WGuPxZ6QLZRcwcMhYyzBNLP6Yf690Mp5m+zFvZ97WSSWr9mttwPFtZ7kK9EOGfMGEbkKVcjkYdLbiqClF+QQJpPGO/snzOAfaE5f1CpRHYKjOEsoiz+VS01bEx4WqFJ3m8jmrkW5Akn1g0hIxi88rQcOLM8P7iHq1glVowsN4aLvbNMEhVJbgQawQYWv1BUXFPebITFW79uzIRNMVPfd9Z38/zl+POYMnWcKFUS6dulnty02p4hAmFdBiQAIuMcnT0mLwFHr2yJOVg8ZpbH9HXum+Cc8j5fv9s6pJFgIsjbVRWncE2lV32mh+GZ9smeRKrnBBWvISHshEx0EK1nO/x8TdC3Dv2p6xLfBk9GhN7p+zU6qKuoXajZ187el+m7gP17kgTRAfIzoERtvoxKBzE7/EdOAAfn0hgI1hvliMF5zFGrRypBr3dbtbCr0aJvWSFyadAUc1HyIuwGGYWmINpl79gu/Hw/WibD+rXh4gYILTg+B84pb72tAjRfVfTzsJFbmeszpU3jS4/BZle8uqYTOP83AfyEBdwFPfhjCWAtWwxklcjSubM+nkt3RtInj1knZExxOrQ4WrGmSyZMABnH+ePzdg9NH3YD6xPtuI9jAhHzaKUExxJeP/zADU+O+VPaQ+AqTLFE53/bXAtYANg50fpEP48uh7MDMLE4xBDOuqstd7Z59etamI1ictpf3OA1x18JyJo8UJlF+tXI30vboHHz03EYJsIDnZmRvPeYF2b/EoJQp7tJZycGoqe+FiQo9ffNG+a4YJSbWU9+Uz9bo+11ft9d55RU8bo6pCqIK3hxIGYJxXUfeVJWuVnTxGgY0F+scMH75hiRefP7oIt7j9kT5FWFfdfSoTe44BtNmam6SvpUtIot9Snszjz+TVIvjvpuBGquvIq3HjU/RYvp1Z0JP24NvaFheDYmV15YPHLXjDwyQ8mcvagW+S+W+4xjRlNo4dX4Atq5b8os/zygfy+kA9A7XgA0R6Os3ceszB3eym919VpYIw68lx/5omqVTUopHnzBKsaRTQQMFqlWrocLvKZB7JM7BRhXqwN8oYUcqrTRFVVmmgDV+Fqtoxn+3zisHnaI8dg/Xlh7Nq5LRS3P6JevWwXUKZjJqsHk19FzayBdjVF1kdn3hPCzeV+U3jdf/a90CgCpCK34RLZNo5HFs6Deaj9arx7a7P9p3mUhl8YrWN2F9mV4eWquqEMDsSi9l/EN23shn9Fsig50+A+kBpNQyjDdazarZ3vcLudAZpllKmbKpaL2yHvlkzJNsPmriRaXUZO/VbWuL9BQhqRprpL0uE+kCN/Rnk1WfePq+X/9/pAJ6Cgezq1+s14jmvwCvI/FayQ7nJ75Pi+osKqgb4MAzt+Mk1OLh5JXMT1wf4twdorMwMhHB6a0O5Ik2bJAPyUZ16izG2cjhQNWqa9ABGKlEcpJ3Puo/jzePcBbcHO5uLfUmh7iwYLiwITZauYoUC6BeHhqE4CVIwjPo44Y08DqBgNlF77vdNeLzob8cM1F1mxH5CH6IGXH5hdQBTqsXgdGX2ECpVwqSe1ItsPESX35JW/LqZ+KW8Ludf59X1Ib4fNAtolttkmKs6Nk2i3+Pxh4itbgmh8sWfMKhQHwFh1IlQwFvw61jxHL2XV79d18dDNdSSCOBCiQZdb6yGYg9wAQ14uMQUr1EjL8DJCIaUeRnoCjRwauyB1Rd5dXfMoKp1qu7o2ezaX8ogM3laiT1F0luwOKTeFIUgVmqBatQk9UrCNH2RVz9/dvDqbjwDoYUMH9pgHWjzOquKeKXxrkAGoDGBWEieUo9cPGJnETMKWufbz3JVX9creN2/3tsPOVCXOZ9EtY6VSe4QEoH2VLqNLayr0s0apWs/TKBBuV7Rc0beNO1pPI6AoP8WK6lX1zt7DoTTSRQe6qeT7ZxNt9m0wkQH5HpSxXraMfanxp/ynlHqOli9gWmqafgPsZJ65XnQ/da80nEkrPU2bqHBG91KtdN1iKn4ngKbG8pRZWfSdBhB5319+jhW/+vq1au7q/E33oMBSkdFVhqqvHIc4jegLCGpRTV7Ef/1zMLEbBUllVR0fS0LTTwOw3+MlcmrO89NBJbviU+T0Vh5eD1pXkVdP5KSy+vppEgEpxcNqT7Qaxmn71/CpfVKlgm93m8Pb8AmhJDtklNnWaCEqVZZAiRPQZDFnqs4hP3jELv2wzBMAjunfUzl5pXOWt5PhxxOcKZY5t1kC2yu//B2LbuN5DrUm/qAgpTFbJPu9looFzDLEeIAubsCBP+I72A8f5D72bck8lBUSU5cZafTjyTdPYsRKIo8PDynNg0MIY9tRLNQhJlCw9Ytj0pFxIo/9RgEyojm63Na3sGPK5qZ34i3Tzv4IGAraZdn8dW6ctaoJ6sILcAuG8o1BuqUVhofksvslxuSep3bGb9qxNU34aITE0I7iBU2QVDlrzhm/WdeZ8sC7A2hXuBTzFUH+pKiClXVmo8cV9Ul/B1aYZMoHOeBfLURGKDMpPe8U9kuWyKh5VaGddKC04gqgebuXyBW12uG9A7+Ply0k2qdPRCmQhlN2kEsAqK/iZE1cLWee8CxpZ4DecKK/MlH1W+KK97H2e9/V25XXSDMNToRseraQr0Z1gtqr7SZ1Q33M2ZAxwwQlF8+10u9blbfQeEUPZ9/j55MB99qxRw61QvMso6rc3uQrS2hC401sM4bSGpUQ4BCSlamvz2rU02h7uDlGvfjvP/78ecExV7lhidSAg0QZszCaAzsjRgDDg2hL55tqWPiKnSQNLXqoKhUfV8IYLXewW+JqwnyLxgv50XTrkGxGsVkQzWB5eUbdQNI22wMHTuBrBhL79HWmA356l/hqp1/w97ExPtbnVJWnXgjoiJ4kO7lKcsY5/OClGNYDCAcWI1JvzENtHKrnAbL/YaPZT94bu7jfFN9lZa+uVjAxKZ0rRYLEm6Y0S3LOVWLuE4p1pvMPuu5quL5X39jqf4ZztDCr/YPjyvYUO6mzFugsU3X4lhhYHoa1SZ8G1YYhsypGhb0s8GAK9RvCStT5KvLNe3oh8fVJH2z2EsxXKwN3kQ8R+C9UNorMmI8tuYQRplqqAUIqhZufQHdZ/WVGk2od/DBccX0da6oCmGmmgoz5sIKS0ls73bFrgwgjIEcjGMqf6ISM1Tc35mvoBX2/XEFy6RdPq9T1zX0eYPSX8dqaVjM35dR5RzWamQBwnAPuP6c5L66+h1saxs+tG7vOoDG2TfwJIJfkwasQl4cyXNmiOeAalUxZgkF5XENrvbDcwAAIABJREFUVVjMZeyZgL0xsp4kri7wias1Uh7aD07gORKysAPTuGtz0WCwCDV/pezVRqwcZHlTwuplU4v4Z27DGaFsrXL7paX78eOxN1BUBHaQWzhdN80dRe0rjHlgE8aKhs2zZFhGqBG8KYek/Z35SmZeDV+Ah/bOEwmwo7E5Mb4+LdZHRpAWTnnDBr+uDmwGlahkQTKxFQy/f67/KmU5dUNN+RK+vhcWoJFSdD5/IyYz7ZTLaVIzzjrGmb0XsDoS1E78KMUU/6bvYLaH4KFNnikLAOrWNcvX85Vo73wrX3QSeE92kqbWIDCTYbhf1rLrYZD97oqLbbJlZ2bsyfDdrc1T5Yv4XtDb9/smzvDjsXG1E/reKQ+Yl1Z4Mto68WQ55M3l1takgWxA9rTpMwvUbIopdV29Ve8gA6PNHd6/H5arNNOR46q5QDKO4hqovNCxuVxFFcBPbnAYghmwqSXJx204pSpf/StSrPvv9HyZEFQ7TAJrHCZgKVDJV2XQKjQc8IjWkTtmx0Id2HegeeDNF69O/3ZRXwn/qubWPr88bG6zE8U9wYuryipkUhpVo8BAuWJfsrENzCH08h9dvEFmNWZ7To8n5b21UjNkvug39YMTO3ey+KViOTYkPMasXsXtDe/XsG18ybFKBRUEhooJBNOFVp1W+e+MRVjpun1p6PXw2QQUT9AAQkauBVnBhzmzFmAL0WiWXc7nnKzYKYkWIMzGdB7Daf5h63yV9iau8Pr+fkyq0uIwWM3tlpw02Ybgn9nhO2StvVCWCjCzgXU8hlrgFt/c28gzUPwH1sZn0D8tatG2RsqPhxWhu4lFh0j8srW/dRpHbcwCTRjpbFrlgpi5cbPMtD36P3a3BtTyH7o5rAwH1vz5SXE/rmlHn/cvjzgq7gRFdIglv5ZmsCELw4xBLwVeUwJFdaBI2IawBL6GW4t1L6k9RlXvX9+1dDTVot/Ab592rDqklgMlS9VVKMMwmug/gLs31lxQXoZfkLDNNT7xGizUecvnZqu4amrvXO6PK6yQdLsO43gmO06VEzoM0Qe1OzIArWrcvwF0haEfyrGpW1sqOFWm2/mgfMpWB8rxUjPAjv78DXV7B4PFJCcHV5au5sSEPI3IPI8gMjoVZmwWyxAklDM4QHu3HdWn/8h5jirbv73/r3QBbWDI5wfk9ozDRAMWvoMlf48pxtBm0nTssaVxyctHMa56JW0yIFH1t3WBrjowy2cTXz9KVvEPnMUdTHrIH0386gH9YCfMbNh98w2canfFkTUqxPaOTqsp3WjU4qReBXT3IArx41A0zXx2C0xmH5ctz4+NK+gOEXDcafF1dQfp/eMd02yIzgR/YaMVgJUQ9oxSzOHqE3Ob9a+fdRxcPj5/Pn4ZCyyf4wo8mZYvwN09jujI7UShcKpIHpD7kvqKhIzHq8qpbpD3r1cb8XRoN6IKi39ivLx66bwcFaLpe//2vkhXl8fzrwhcyM42onepiqsg/L1UhQ6jBmKGurtRLsw0iSesCmom2y+gVz/nc5rfwHhy6bzeyllqW6/v/HJvxa70CTGQr2TXR4iJU8bKsjCMw2iKh2F+Y0ap5BYy/cz0a7saPqkDfedTYrc5rqx9Kjnbkc/w2LiCiRSV6wD3FjSrgH5Z5AkHNkG/9ghSY6Nk43pVV90wgmhFFpedMTXNlYKzHFHz6ZVxxbpqLa7afn93j4N1N3CtplOFGgdg7Mo0nnPV2BYPMKzGa/iwiK+3hoPdqK6c7bmgSkfl534wpnhbxNVnump39DiUo5Sk/4mFh6ZWCSqshXEUsa+GehzrMQ1GlgEJOO6hbmLW5yoqqjzDCilheXr+4oOYvtRx9XGtbj//9wHcNOVr2tZn4tIhAzHMQ2tW64YRPq6rsNy27qSaGKi3AGHmk+RSlP7gafEOXmp++/N5/+uO5mbKH6d2YAXxnFQijgwaj611XFEBze3fQLu4dBPNLR2NK4uqXCk4y2c2f7gUV0c6svwOXghDbu7EvdzTCRZc42mp0IsZYAksZDs3Mr8bm2s2hW5Hud29CYKxjtoaeyBwz6ffY9WQbiOf1cdn9dVmXJRpHkoDGqm9tsrN2oTledUjU9YEzcDegt3v1l87CiwqERitSkiDNfGUXjlnvb0XXtj7tr/zy/bnb9KK4rAe6RqM7DzbgpV3U5HQKK1ZB+UcOaqvSgVzJbN7/I0Ae2gD05eUv95UP3h1N/z5n+0rp3xYPLg5dV1jPRAKTehuILku45va6SclKChWGWFi37JdWh6a8/N/TmGUDslSUJn0BB4oa/HHUxlX7d3wTfkKR0VOEXBqablK5QuYEL7c1zAhNJTFAjYhKL8zZda18PLrVYKr7iBDxrFENyk7cUDN6T2F1au1y7hqeChs5hRRTUUmXJKqyiFzlp0Ngq+fdCM41kYHQzaAEL1wXhpxZkuumj9RdRAPyHhHOT11OP5A9y8+iP5JfOKYA3luadC9bLyDHe9HdCIlUO8xj3CVQr5iqfVQmyiavBaoJEGzuMKmGaDFT44rSxBMzOtcjBI8o+IK/WBTt/bXJnSvg6o/aGlTXa8HyHmM4juSpWGGpYy/U24QWgrG9NtY695ROUWlJ3Uz85eG0rucU/o4Fu/gFT2Z/bZ8NU2i1qvMkha0bPalFq4/g6GyFR9qmQUemrJ1mzEQ7bgd2nOap0BVOnc2PVVUnivRVDLQR4Rn/viz4DM0e5wNvD7a4tqBD9NBqHeqDypg5zuoCbNSpw/avY2dowozlv7rB9Cpr/RJyS9LT2CqPGM8HWx8CJ3ElD3OvxSf4XJtf3BTfSWbSSym2jXsAuFtypoLJ7XI3AAXUISKIlOmNzrnbjkos8jr+JFgPTM/db3NCUtXC2W+Yn0GFVb6Hfy1IaymTAkFvle4CwvDES4RQyjXkprEIWP0kinkLdezhoBVMZxnLSD2uTqYC/Xi48jH9bbkybR4fevzVVfsuyG51zr1Y2aEBlERD3ktIiwoC/DlTBsRorGwYg6vxa3S2/dKt9Ak4MUQHuMlwNRh2eNbyZNp7piszlcdTANVuXCqLAODiqsgvlJkjHRVHNQJdY9V6M2tJYLqqyOhA5NlrgfoKbSoD/ibFE/xpNJvf5S8vrav5Za4ksiCWPZC/nLEGiVOaghB+06WsoSwYwEO2mMZohea8SqaHldTXFV5xx1OvIIHOSNJWVQy+Lc/y12A1g7veWU/2InwyURy4h3a5lr2JBT78Wz4E0JzHO+cCFcNShNmPQ7DgAL9xo2xXwSWHFI6p6O+g9jHaXEgn/c/11eiHVyT1D5zhYOOIKaNzB0S6sIysxuRpHBZEsbcugloCpjKILhoUOO4qiLsKiFWto/HQzn9mI7vaBUmQ4X7/tLqB9fNnTOFtivsIk5LPatEdEwj5gxZtYBQI42gy9tbPFleMV02eUzq6dHrGW+xVKkf0lGlSOKsHk/IHw/HlLVUXOXRxP5uPsM0lU4IpD6UGVeYcTHPEfKgtBMRwlJKDmR1yljcMDNk9fXLZxoIqOe3TwoFLxmLLh9du/nb13hMh/Qnfy12ly7nO/e8KFHtTp0aMrdMc1nshN4/wDGQcQwVtpCVdEhtSJpA92l1YMpvbWbrHSwjCzadmaWiyiGakNXjTfTHYzq6Iq4+LsqiSr+DG/LVpMT9F/N4VlrIsLEYSY3NZW8h0BrV3phblBurfBU7mAiqp5Cav2Ys3dFdfLVSrEuuQnU1H9hfSy2Lph7yrxW3jweCmfB/mloCmCOKUJiPMAY6tvxeSRbGYBtCW0GsOSmvemTUUZ4y01yGUjBJWHkcV7yPRW5nK6FzS2f7+fnn2kpUzwMrvr88fKwqjrWkAS9gqJVhnCb59wiqdZWVl3rKp5/xh2M22nxESOcx0pDZj/Y1/R57Z4mrj+s622v2cWCJnj0jhBVT2JVxLQr/kczMBrYQ1O0Tdj/Pa8TrznwdTzmjeSH3exwb4krGy1IexOM5WNRW/vgaP7+9L3Rrm3rIP1cUDGrCBYOb6YpZ0iiSQyDEZPtJBRmjwtKU7A01KAeV873AeHTvDqr3O3ppalI4xYx1PMTsbnO+Igpkoouet9bt3WJ62sELtsjsBFnB2y1bJS2xPUXyMMJaELGFL2/fEih2CVVPV+7VgqwX8zlmzJKj5Aqmg5s/zc9g/PyfEhdt+U2cV7yDfEqilEYKfN1UkUJHUeodYJUUlkLGA6ho2ZhFBHndV3G1/EtGXeY/fv0/Z1eTWzeSg73RAYSqC4ynE2OWgqR9CtII3V4JELTvhZBFLz0zsW/gPk6OOCr+V6n0nl5ed/ycLAmSRX78+JH4jS4Ce2AvSlVsI/Sp/auOFcOAFrN+RfnKNM8Pzwfptlslwid03uZQri98VsNciSjdPefNLTnNScJoj9LWvcH2KEHVJkehsaRaiH+jSt2WV6P/d8pvf9FT2L/yDgpmtQoxtMCJWdRkVk3nQAsV3Sq7QEm3AK+AxgbV2puYWFc57G46tlGMxiDJalRwL+aoWK9LBMYvk6/oLsAv94OVrtzI0QhJU5WpF1aZyK+0G0GqqomlGqEPWQ0B5wo3fu6u1Xj+om7GQ6ngvRnTxGJhpEIdf8YyAYvScYTf/rD7OCS+8/Kr7yAaCcxl9k4zxnEvIuzRoVYkr9MGSX/A1+nUq5zkZJ7/I8aKJGxoaFyQrnkvGoZggHUtFfbPgCEY7YV+hT+/GZ7MZ7IS92jdXjHZw97WWA8XvntBq8DHSHy9PZzKdazi3+ppTuKjPehVDUJVtRZUbCKXDCCwUKAUhb4Exgr4a1KLIrX2vbQ/+OUqujCn09Nj18zCC+ZeLsVhUSFbr93B3IYQmOtDG6+DZZ4/KExMyMvIyKcBQbF6YL+K5tr/8ZvRdPo4uSV0aY5j5EI5Wc3zXDgFC5VVu5p5BPmVXThtREOHGexyEvBBPpqXnw2hL4i0d8wD9d7aaRximsL/o5U6DED4m+YrOQF67J0v+BVdZWa1Xl3OTXTSEGGQofwtv3KChuqxg/p+WZUM6wPzO5CsHsnrHt5ETupQL3D8dZ7Q4vg1jPvf4zsoEWhxBtnH+VW/SvA9nHBVx2PMCbqer04uyQoX7o8wtt4aov9laCHAHjwujxDlTCfwtc7+MOI6eALpBRzpEzgMwx8GQwbPOvrVy/13sKLjgSTYayaCa5bYVzkvpTqhhp/dZhKh2gS2NWlduge2AIlV5Qj1jDgxoMVIquKuD0wVq/TUVB6yfBjlo/UVcbYLfnWpbpfDzHy+usqUColrLGMbTe3FnaQm0aeo3cVZYJ6pAnY3NP8LvBQYktqTuj9vDIcVFnw6egrFVh+MMzyX9Bku5Ss5iLeqoEdG9xfFeq3YVUcgExNvUoHepr77BqbZKoKe0C3TvCY44sBojkqAYn7+dttwZgeQj35J6/aPM93au/UVSvAxJro+af1ZFYBQYITCleGeTuXmigstUx3ZsVom+l9cMj10zXt7g+CLADEMDjNEjO1xR8U6tjVp4krr9jN90Wvv4GyY/6RpVR2XIzhf0eU7HUgcTOWsQgz3y+4iDOP5BUQ2aDRQQ+SqoJk9UNwBljcEMU6g168b7Sevr0q7S/ffQRpFrJXebKkyThoKZa+LuQfLZxAOSqqOxKy0E+Sb5/X13C5iAcCXZbgqSL4adayMs4hxr60wuZs6QeMxjcH3k1uN9/xqrswqZaW6Q1UmvLCsvWHOLsJdSM9FCH2vMZek3OWhaWPyusOIE4A9MFkBBqgjg5+Q5dmZyEhsIG/86s9cr6+IIX+9PMBZZz0Lm+l5LLKaxPIUzMw+HECXjQiFrC71NWRMHgPSko0HWAG8yztC9fZo80JWGMmvKCaHsRtGaQrHQ75SBOv58bqdeQtPqxyOKIFWKMOu2uLM+C8s2pBSodAcm7p+WHM21DWzP/EhlNE72gvG72CdDsZatpTKwy+rGT7eMzELg/X9uMKhNcDxOs8ZZoVI6KL8ITM57QtX3WDNuxUdx0doxkBWRxpxiHB6/IPvH7DUxZ0CzR2ioYb9PyjRw4ADwdyn4uc1xfpKd5fu1u0VifDpQbyiAJhObpQY2pcmzHz/oGnk7uSVqY2zHoUL8S4ot7jznKxq7ZQ9v31Ba6nTj5k7Eyjz8ljdPmODUxncKlVVJRiG5P0XGpv2KhLaZ+xZJ0dy0VCCGl8nggYkLBh2VaOVAsGfPngIQUH1ond5Mtsh/qxffXzyHn0Bv7rpVwQukArfSlPBDAulQrQ3F5nbA7jgZHYqgo4MsN8rrVye5AMl95AWCjB6EFCd/Ap7vzCWLXT0q09dyCn41dc7u5QKMBwHpz3zQnnMvFL5WdiPx72I9gDvNVfHgV4YaKwZAN9uFBI2TWmk8YsFJyLqaDr4DAWbTZqvdrf6KONXd/LV/KT7gdI352fRucEB4hCOJDiv9xnPsWnlSgTndXe5XI/oFPzBWXxHVD0Oxv3fRqFgj9whe6QMCQRz4lvTt1Tr/uW9xIF8+3oTtDJK7PNhPZdP5/YKs1P12aeGagjcU911vGbjHngCg1ZXIB8H1ShzG7H+rIMwhTo2lVd0/fSz2XxFlKJiDP7vZg2qQbgWnYr6m0UFoJfsCIm1Fg5t7HzrgYKhYdjF8fMnfD2d/VH0YQlKU4gA4MuZT23RVMk7+HF2Y/3Mr2aBGcxV5lRdfJG7ZS0Nmlex05K7lajU161ZjLgFhTbH7Tb8V17bqiOs5yylPwbgQMxi8qpwJ6mjvabX35N28GTm9eUcjKlYYFzQ0JyRtup8S64Lo5rOctT+QnyhbUzBfuUFtPtIsvRQC//T1OyjgRfCIGHnbxhr2g21aT/4yezaUgye9oOzRCEjoXNhlXIRvVDU9l/6oqhqw8PAxhRXF+c2jVDRBhjEe2icBQetefUBvYnHgGgknEjEuj0ULTaxW43qV7f2cb6cChAJg91IG2vd0NOUeREpaMLWW6aIHneTWqGE1rdVZ115vMVIKLjSkCxrIazQUbGO/pXAn1Au5JaayF5TxpN5e3t/YO5cyeqb0a0vENNWnUaIvGp6WEpOBbYCMdROxsyX0XWsoGriNcIEQoDjYPZrorEGO6YhYL30+u1GmnZj7VGofkUSKc/XeX25SkXhxGK/WihG7ny3B2J2y9Zilfr6suaXk7zOk3gtEhhTqBku7mjwELAJ9AKA+hvZKsOvuHc2pG3Dk/l6uvrNl26MXsycHONaLbggZztTYjbtxss7CPmdL9o8II/dkNYz//SyrAWTZKWheZOZuKwqZaoNfWpPVtGxXrN7qW9X+aKs1VTJ8c7j9Xg+iUfkNAWOj4tcclu4STDje4ZqnBkEesZCvWzgUuQhf4GB0CGujQyQ1324WX2SU21TismIsY6ziXO+KN7ZWE3NXiXEUNzSNVcQkJaG1Jg+4aQ1IBOq9w8e2EyqhVtMi5JBdrUQEB1rL/j6yD0yQXvDSbmwGeeKKWv//na4oXCNL1rxQKJSQfZ0cLPI+NTIOqrkwlIQdOTdCBkGXrMU7tZI1MGeFrTMHWb0aCCMww5SOlIdBYQ5KUQh9MCxYgRG471md8PfirpqP87kVZ9kGFG83bKIogBd+Qbw+ChRQbQYI6Z6CbFyB8iYuFbU2Ixar9Nvo+ep+63oI6faosEmaHCitSRfffIifVFP5kv5IokMbp5IU6dKLmwsQokBx2IJ2nyVmacRZt2GbpW5q1PTQONAL0IdNAis1UzgWViDeuFzFOMPCirIURP1gli5v+Zz54JfFesr2Wme5TDzAV5YFz6Z1MKSYKtFaELgI0pMa1LVldGpWTIlDhqrEgoGOo41Dv+ILqToS9ByKpRrBHgGwVRbNNw0HWcTJZ3tL2XdbGwFZ6NFm6l69HJneEXJBeoC0wGqo3qdOxvezX1sZoOLNgHFQAPrLHhMT4HGpByAnmFiXyxBo1dt5FOIMWR+hbOJ0o31lx8nyLGInxypjjRl5uqq5ZFgf0jrJHvi7LXOyzciqOdr6AUE0SpdmcSIxERFHQ19dSe98jZJiQC/kl9Fz9r+zPJV6YZCgd+uplJlnaoqCoyzppVKhZbOcNVmk4Spxhc00oLOmVFbVoIwuhUGHA1OsQC1qLq/UanjE7hRGGICe/1d17xw3/m5oN/+41CGyhkztNNanp6aA1N9K0K0R1V/uRWhx7cekKYIJDOkoqBSsKt+QFwWgd22gTnGhVphonoKraPGin412R7n/WzmVarbhWw1p3LQ2YbgKqeAetl4W1KltPRovDnYcnXEFUi7H/YgZGpDCKhAfB2nduiSw+1qAWpPyFEbNjmQ518zDbqS9s5LFoOzHI+gE7qFI/JEpRVWTMtOtWRKAo3Ir8vk9N5NjSYHYSCpN0HFOpjjiIBVJ9xrPyiw4HMEBgtP7P0QXSDf2qboVtP4l2AytDVRuoX98tvPkgyRwKHgUvYR7OVqGSQrWpMvlewo6Gid6o6cqkNrOd3y5jeQg6+x8DoyFcyuiMGL/Vhsl8G5sEoYIfawFtXe+fNThdVua/xWIm6cZPWMwScrlXvWaqlc71PguGG+o9N7gQ+uUFKiQp5CzSMuU61TP2OLz1EpVnn0bfL8SakAyWrClPWqt89YD7l0q/FneU9wtuyhZJdEiAuAGrflvQiayFsKX+1c3Ty6xlyzmWLyGi0OGqMO0jpRGzNEr8utpX5FpQJaLhprSvLVqV7f839+5uACQccsxDfn94AWQq8AXe8XnAkmz2AjIai4MbbN1zihgHs2sI8UsP4MUlzRBL4eaAWeYAXfnUXfNmm+Akth5EFNCrXVZvMVx2BRy+IfP1OyB8mxq2ZhNpVn7UuuQ1l+L01XRHU0PTOgC+68EXQncAzN5X0ineqxVc5izueIetLbbGApePzAQGQsyO3Ta86TKc5xNAYrhI5N0lq5uakMcpwSQ3uR31vy+6ZUgLYi6XgD4EtkVAOrx5FfocqsrNiQcFxAQlqgPrA4CER/mhjT23QgAZE3UYFlcVEUKnor3RLKagZe/9ZO8Khc3/fUNdMtLszqKq5KSySMH5P8Xv3AAXQC10lIPHjHAo5BpluwrYzWgWTlT2t1LhDIOpSi4He002b86vPv/KyljcF/2XFEpfcDqRPM124WBmJwo0uOdmasGGewBdeQ8uydZ7BJCY4DEUKZMdTYNVxgYRPKLlYKR6faTBXK2B4EIdsrfnZzHfPVhb0JQ9GeRVQg0eBTVR2i7/V9LuzoHEsQxV2bxikr+9Lbx0B7FCqWRS5J7Ezvl8Vu2qD0xbIKy8+NiimsrihHjdO25TGo5VWhZjAxSP0y1urCDj0q1y865TLjiD4l8EkjKMpDd1T9nSEuUBdYa7sM4wddMKVQ7CQEz/K5djfRYIitUyk6Td+jS+0/pu/fX3Pt6NI+zvN/fyoc+iQgH9YKuVzasopfsRJtm9btRlqcFJoIXHiorPIOjrHQoilVoDVxhnQmr7V6KFtrS3E9eP0obUVn0ij8/ppr3Rdxhn8mqgKQrGh8Cl5VHaUw/0/ZFeTGjSRBXfoBDfID1syszgSbDyCaIAZ7aoAgsA8gCFg+uQ1Y+oH8AsPP0EX/W1ZlRFZWsdhsecZjjY+JrGRmZERktLsRIGZIL5GwXGEaPO6axFSGva4XybwfYercQbHWmUxHtlWn9aDcY1CWYAmuwELVuZRa/m92fzxbnOFtq2f46924y8kNMz36fUjuLTbhfIsehk2VJJWyQymNALpe3fsJxEKQTSj2XC13yu4beLJwwu206iRGoMSwrfKR8qPzPLu/TfLq+vKS4Wxff75b/fdFNRLJbNPgfBn7hSb4Dq19QgO4x8Pet60EeNO6LezEXJY8iyQfv+B3SaWb9KLbDDQPvAhpgb87dp/uhxkv0NSr19C2r7UA7NvB4HuQHRemwfWlmyEAxyMvCDZ1PDZXwamQWsobkarSbQQFNgFY15J+NnLJoKrZSKwJGChaUSlQ+ulzcZqWnJr9T8/x7GyPc5h65efBA1eCYhlDssdlTDyIGtlz1U14fUP+dq49r1h8YsUcKpaSsf3vI7LpRHY/Vbq5BQTRdDy9+OvHDnQJU+fgK/fDnPQM2d3E4yNnHKwCZRcoW/nDim48qmFvTZV81INWquSiE3QVjttUu4nVqupU3E4EZ2/FNqdVrCqTR22cUL0ZnJlX/RQgGLTry+NbfrvESnvRDP9qqVeCM1xoBgbCFSN1WAtPjUIQF25WCwmyjYXiUezfAapgdyIpRdehMviHR02of3QnMZHzWVVmyXo98XVUJ1+xzC9ft5acmm1e8Q0+ZuZBhzMc+AHEbUple2S8rcwXcOChqfjIYg1xRNgzf8agAsItN+KwrJeGtN524jJkwlNufwMBenJklnRiuXI5Nc2zRO3538T+6mXbkxwuxxc0V2Ny6QYOFWrBYHWnw5oZWpiVfLXN9IjO2YgPqPv0ebE3h5vYbgjMYnpXlfklICCYvovzCh/A2X8BXZQkTi6vNFavKytWi7f7/gp5pWquy/qAmX+H9ai3y7A9jTc3lZFTolbdDYS2dsZRj3UVL6O5OhnxQ9ltUYZIVOgkjQIAgxbUzzb4CPr//jflt+fvLr2D8Phw4LGbnKnHQDAm2Hpk+nUp7lV9NOYn90yBABe8myp9FrjmQkEvSTAuIRaRxU2O1MhgoVlH/9kxRr6mS5laUivOK9XR53gyxEUDN1QvU17i27A89ZbMgc16DjQOYDti5ioOG+1zRE15iiQRTjvpfGB0wbzJ6+dqecJeS9rObuYqYmnYXUr5oPXos54THb2bndf7wR90S2PfPirGfslcHI69YpqVZ3YVpFzF/e57rWrjo85KTAkNdwiapDbVw1sStjQJoAyhgeq1AfVvb4mPjxYK1xzNg+IDmeOLXv/WvHrQW9aHNK1GHhEczQXBFWxVxV7sn/EqdJd/9MjIsQ1/UufNeq68xlO7RVfAahmQldJnBVNY2qy+R21Hg+XS6znpGbI85KUKfp7xAAAgAElEQVS2cyE/CsiXYjHmki4vuARf8eRsEpVcx3DgrbgPMvZZdSqIXh0JsB+xrIEj4a1peQKejnZdq5W06x7+7H2NcvHxqSXP0PZXr2qZmfPe+ecQbQX1QGVCjGnkNqWIKaE8XTHTCq4EZcO1sz0trJnq0RD2zvEHUP1zWNTLXGfVK27ca71SaB2D4NwJvDfLf3wjGueVw2RetzhFT//RvDK+oePKqWJUaugYYeuNdamQm7BgDxX1J3YR3p4Jqxvl+juxTdQwlHBFK8NkU6azzWQ2EbrdQifVzyFS+AFN6XO8m/B2fU+rvPr+bj+DuEtySQ+e6slTMmgzlk3iFwqJ/BKpojAEhd3PIM4bBLuTNvIlVBQ0T59VbQ0o/j07BRln0IrOk/RTvgudGSdXtZ5THvLa29DdPnuH9NTYFpqWvTHWOvZ4YFSuBFmo1YOvwOu7JSUprJ14OJVUSZhO5uPXinWO42l3xsqxTKsV2bJggaKWT2H+02adeeU/iD7Znq2v2oank8fbg9PxBbjxIV1IDGZ/Wg/Nmp0tVgIq5IKXwF2ToGQfWB7eyP9oxEjsFc63pPCTToFSz5FXAVDve2SWAAszuwb//zavwEPO3MdxN9bf9e73ZW3AIEt5crPHgLOvP4LksVuN4B1uvR60qlqeywUtrQ1Ts7jFwejEVahzt8HF1hVEh2Z9MqDCLB0C4yZBk2hNIVb02c7rcXzffpFDZopcEYwZgqiSvKu6pl9FKj2tgmfv/RcDK24kXJt+NudGTF7p18/DDDm8yjSiunnwgbPFSj98rFO9H3T8f1P86uklp8f5x+fVwwMElYcVGuo3qNTK6zX5OlrKEwrVjl1N63f85Fp1HQITrTweV46zgGACF7TMtlYT6I3Ybum7E8B40k4dSdZLRslg+PxvYsWavZcq+JWAMWqtk7SiQzg3XOehYx+qsOU67rMdTR/aKmgFwY3klukWTniAZa5mTRoqYjHc/XU2TPLmXFahyKN598F6/l884+T7K1+vzJn0gz2li30gL96gURiaZv36iqIyRzawla/2YRggViBZBT7MUc+NnGkCQ2y9zEghlNQB2pDuHzoXk14bBgmdBEvhBlOvcI8+27e7vLpQdpO5ZR3whWCFWdfrTXOtLpicbIr9K4H2Um57tC71LFSnM/XL5AyVK6yKvTqVk6LKnXSvxXEGk3P4Dsp4k34H38jZvqb91eNP9lfhhosV6jbYoTahUg2JTwzZxnoWvapu1ipTtZzZJfwpnC9aJ/eEjfEsNzY31IBhsMEfs64fXEg6F55uRqQmdqHyq0/z6vX17XXz/uCSV8avIsGOB+J8Bowh2SpuGEwTWqmMq9pqQCsz2YhCYuk31U21BNMDLSjpxt2GvCYoAgOtil+/GbVqlo9gb16d/H2f5tUveO/kuGqub1e5/OUSX3xrBvXg0za0bjIvkI71FmK/SyMoxhT0WtC2KvEQN2DV2b7CXt9g2JgqXRadFJuqGVtT+bGPUmuev/2JfWvzepzv7/oA4XGVwOyDMbbSS7qxRr6qg6/H7gusct9BQlaFJWa36l6slLQyh1jJDpC4umnWfZVCfk0c/XxeTSZO83oezGEypm9PUirMzaOaHNdqHJqBQ8lfqOgSU+xiobwoXELHtXRYHa9OdtC6eXHuhivMZEQ2Peu83ZfOaKikVOkPCFKI1BKrP2aP85bfD7rZ+YHGHgebVkMwmIOmWZVcyU0u2ZyCbCWTYHHP+WpJKX/HRijsZ7WG4RlTmQLp37/lctJ3gdeonI5u8sD6jAVEL/9MITw2VBor4qJ5j5Sf70L7Z4wusXf2ONppGbPgyg4ahyPqT+xOW5h/QcNFOIZHkcSS4gTz4ly3LprJXtdaXGmFXkHensLs+Cd6jmms0DM8XbPe0V8+cCP2cEhwK0qUxLyJi5smdooRuv8xTIL0irnTpaJVo3pYfoWGoeNKPm8cF76CEJdOFoBxvQLg9NmGKpNTUaxYr15y9+h/fIRjngaNgThiGMfA4hsyN79rQ7iiA+a+jtKl0rmsqA0MDmkdRptOtLlSz8vsLp4IDEhCIPGZEXCataozaKGsb8TqlQvCa+4e/dJf0Qos/QbS4VHJ2QPf3xBxs+vYU+AuEp/fR1QIVUGEwfg+A9+jMjDdMfcWYEBeTfYbKCQ09AfEilHW+81YAWfIcYr8HidclY8A0XAab0Q+DbzONUQCCTUXF7e0HbZVEfbxMi7j+PIpuIlj/WD6hKzIlDULb85iehheZAbUTgFh6jN5Ffor7ryumTf45QMg+zhaU+hmDIzHMA+uTk1Vyo7BeY2iuEW1snx/XD9vwzUS9xfKbDySj52JFomz3EIEHjvaK/RVs21BCVzNt94gRbxPWS9W1KsYOUZRb4KkWT6Aw4pI68fAKizl78Fh3OKU/HXxqODIbA4gwESg3SIN9WEQ5PK9nya7CMSAQ/pCb+ab7XrF/WB27/zl3W8DV/4LeiTP6nSjvBKIj5FCSn1G/e2xdbORj7Y24igHdWCb/fyxZycFZuq1TRcOjP5LTtq0FS9Tr1628PbHq88rKeyX6CvIS4Lir0pR87CyDdXDSexB9+u6R1/OYjYU0xdcahVornQbv9EnkFncdWYG9P3npIqICfTZ+cb7s/WK+8Fsf7XUq0BhSPxoiS/UlqO9tqowGvm7rieRPoRlBHVcbaCEtmgVyrx3QLDMAdcxahUCTyHklSno/e169Xqrb//xcUg97AcefWuCVLAOpzYGy2SX+QbH0YviE/fjnQG7ptQZxkPcReDdndo2Y0s4kV4cyP2BCyqdAVp2rLYmUGi3U8vOOBtYn9M7f7BdP2Tv6TZoE+SEy5CYVQjGoA/Rp1S111hVvqVCbhWtuH6p93rHJRdHm5TpAUyvVxXzZH4JqYoQaD/PfQRa7cUK+8GnLCZz/etjVH5aZOqhjtChZV+dbwkqXWAxN6RvzLiKLk3m+Umz0JLtfyYVpt30j4Mifgq0BfTqM5ZbuqyZ+2nz6WX6q18vW5q4p8fvbh5cW/1D+mZuTsX0tALdlWnYbww3lcar1cuKvqOq1MixjJJKvoDlyhN0CujeBBSmp4hZACtul3VZ0+8mVdRfvby9bfJkXF4JcHwwgWooAEdeYbyJLAulsVK7mON9RpiVGvBVeo4SvBjwh4xoK6sNpDEo6NeBMItQzfwKskD1O6H6GvXtr1u96NOT69svyZFYkohID60VvUpdQwHu+a3EXiNaxTgffNePxBXoUCH37+Cok3mHE132oAcEphBqu660+plkmFuZtURq/h3q1a8XPw/mvHcef3xcxozX+ICjbyLVVZC9iYnsLloFiaH7EAO814PcFHhMh7xqcZ/MgMXxLlDmv45DYAfh1txZNdJE/st0x+Nzv37b76C+wcxu4u8PaUNjmWCgZ6vRavINLPSU5ycVSr6qV8GlEAzazv9xVp9ewMhZTQQFN2QNCR0bTSh4C0IcQpZNNwO1/P72JxoHDf0qrleHMePH15gzSjXuySfBopoLV+SrmybjlelEsYEAduzgPm+914b7pe2OgnlSclVH3QgAz1kKFur6Xq36/TV5g6/bt8+url7FQl2/jTCcR61XCTetNucWg6Z57w2Wx6ItKdJ1D/AUbDy8TTa0lMHQOK5VE5BitO5EQvnxI2mPbVXf3/kGv/1JONuZm8XXpV6lzuyNbgQH8wlceeuIySMd/u9Rf5e6RBWyR6W8UI7M60tuZQZamFQJaIG9jooRREo1XDer+lcXqd8pzvByzdX2pV7Ftml094DPsT//3SQK8IImx4WxLNxC2W2y6QWgolVLudhYldKk/KJ50pc4qWzy/4xdQY6k2A5cjVTbFFxgavTVa5SZB0AgLoDqBggJzS5bX9X/BFUnQEXXBWYOkPf7yXOE7QcPyGqpq3tr+Rk7HI5Q8llPpj8pjk/8SLj6UfPqM2ik3NJY38/7HytJdtXVCTgDbREW1cpLEGWn7Bi4Kt05Zc7j01h6T8WM10ULfFAjrNedgaG9Yxo3jFSzDy3MpWoIJWtY4Vfv768JXbXbn/eobSfhUbRQMNssTZSMScu1oFjdPGkJW5oH5QxgnXARUUJavK7qdAvqVvLk7Al1oW841YSGobNmodsrUoMGLcqrjy39q9vPeyzLzkv5Vgn/V+Gmtdc4r07FEw9wrVmfzVeUq5xCedfWKl9U9iYeBhvlZLsI9UpxPO6sRiZWSK1xsLx6n/fOyTuv2+ujv0pIQnMUVDfdpSmJjs1iZP3E4ZvdfQu6LgADbdAp/zyv5JPOk7Y27Uzmq0fM+hpoFQB2XTYfvMHwe36Ev18+o2PL20Ze+QfYOv3scwQwtGev2MS1jVdBKZ5aNsNfsRIDvFPtgWMBQoUhWqWEeaniyLVp7ehBGAEbx0Dbm2rC78dXcBznv/9e4VdpXPSPWI/P9LNVlD2+vMkowrDWwtw1DgwL6VKuwE3Nw75/UFpIbE5VwKNDc+V3gMobkukPV8zdfrDGUNRHhMzeoNOtXeMM7/+5xw27mlm/qUH6SrsQ/fr5pF7Wex66mcHsEFsw33hjWQXdIcmrak/ysqv9zrRGSgFj6D0RdCer+Pc459WjXPXfX7EP7y2hxTrn1QK3CsjxW6vyVgshlKKwuRlc9me0e0tzIrlQWfxkrsLibxrmv3xDw1i38hTQIctReerMq/6gCx3t+c1DzujyCjcmt5S2oXwH4/WpVzq+nmN9OcrqnPUz+ITjtwyBWODoWsIlVvA2re2aeYXB4H7SuGiaXOSAkt1/lFfy/EaN2jiEoH1/HXphB7w9tkqnzuOVi4goVE6rQphpqqxzfB8hVkAhp4oqElYNHz84TAkmc0mI7tWm98xS1Rj/DKstLuGP2vZRHt8QQjW4nmHXY/2+9KlUxxvyjqP1KS5JzJ1SIlXsN1alsWlLL5QN7CoyrE7db1HKKtpwkS6LC+bG9hG7tWqOjaaV/G/4++UzPsh5TfOvvIi9M17ECfjaeNgOKuG3+AQ7rRA2qEJ8PCShQBM0jcs6JRYDLpqSZ93Q3KvYwvZOef0GLa3C13AYxt9f1l8JzpDQZ5jxdmMRtU6YnaSY6AnGjm9P4gvKizmdqjLaM8tukAvmlP9Ip54/TKyusftJrpZ7u4vYR4uHHs2CTIPyBB/BYl79glHcLTUPuu+gFCpcNbeq77FwnHLacs+y03J14hIK0YVUPojIVWCozcBCldYSUO31uLfSW62GGOj+++MXkF/BULHmX7+/FhbrybxivYKn7ts1FuNr/YSTyfvzQuNPiAqUmXfZqLhoZsm6UKSpXB9xNY7g4RTR/KgsMeu4XT54hoIqDAO/gBKpx3fwZclVS+p+3KMTcLc+RRuaWt9kqlVxbAyUQS1NddL0OqLkriavPS878Q2MtDxY12sH7UWHSFtN1Ui0SluFYcCX8PsrujFJ6TO83vQ7CA2GuFpFU3NBiXG7UnpmIzjXqqKyM/kyd7pWuVOKKVNk/4aMGEPZiYFiqaUHpzvFfXAo6DCwr0KcxmmIvoO/tnTV/HcQoSLjSn0XV6rQ7p4kTXlc392A6RjqVhVcq0uTE6jrDZWYrtG8EmChVuIshZmazvKq2U0r7GzQJoSghff3CNVAnMHp1v5I4aIqcIUFDoxiAYi6FSp1wKhdeNpnxhQODS3Vk6R0pFC1wsPhaRUbMTdefr2pl9KEXpdiv10fMfqFgo5/DdI2THOk4nolBjk/Un4Td4/HQGguFKqFsEfhodBMQasnkJjidKIbc2a9lYyCl1IPKLc4aZ2pZMvSxg6P9BhX75d3MHX5DKJRH5Bdj6hN83fwJd55zdXqtpFXdEd441ElebTX+FLw7GyUtjlEi208jt+QV5cIDA3+I3mYceoto4guUqfoGm2nPMC3Q0SzFziO+BSO+jPn1eDz6n3zfvAW6lXLlsH5uCxtF5WeffZKfMVzKPsMrJ9OplZ4Upvvst72q4YuWl17IX8ECttT3gd2R4yF8AIlrQb06wjV/NcSF/2R2OPcrL8yulXbWl5ptLLCLpWsaU/YTWVxtcrm+UY2grMMQxn5fEMqVBfOeVqfsLHTeLRXvV7kUuyk21ktYw4UuJg91SQNwzC/wcn6dpAgkzv6kFdXGwbNGOF6va6Qq4J1nTPgYVrNq2a4TJUn1yxAqSlXFKZMMYdU+iTihCKTIFBx1FtJ+8niLnnF/JKQTdozeI2UZL0CxEf/xXY12hiRL/OfwOOZubT770p47M4bvQbIl5t/YILD3vGQkmpfKrvnWKDNPmg14g+7hPAAp0E+g+EZ2hvcvpsIHEgJ1pu4ImjTbjifLW8yd6q7a7ioQbxgH0+mVbgRpBFlWW4aunWghULPv6m5NwVlqLPXd4BTYfyTvEJW9VqqkFeLniGpFYZ6RWJMEMVsqdx7XSsdq5FSGG7SKEO2UmCgvjhyqjwxTOLtludLHkMT4cfenKUhYAVggW9x5/vX94hRrxjM/DOhZE1RvZpX9B9bHMhHvRKEXfKKntaybG4XHQOVC61nf2bPPEvKFRD3wP2IbCSqhUZavnTWUCFxdWcRzlCncHHTHU2ClleD+wnD4Cgv8BEunXE+Vb99CxclbAX53hXvn2CM+XOB9p8dk/5hSOLu5EuSHXlAUq3VBLpa+3Vho9W9LFDZTXXdIbo3AKvSis5OYQrfvlFCFf7hZhzY8Kb6duIMIbfOEDuWT2DkZlaoLHt2iLFnlliXuaBXYctVlR5ggKjcJWVFqU4RjXVWbrbB8ga3lJvhGnvXKbBZly5hkJou0XoU+GjGEf321O3Sz3uo57a9OStD9LwxM/NCIgVdFWswlPvAU2SIpzPzGl1QHQFQrmwViFsI1qv+MLGAKWi82Kz3HAblFU7fLw7q+0jzr26vgiGbpSA+f9cIjsl0ECwccvWUM2VpyvVe2or9VVlvLLr4BJtGhaysbafk8x5pYYxDZRHiN3CSnv0Rr3Fdr5KaAz/vLZdd8gAj6bTlRYn7Cm4GK8MywlQLARyfdGYW+tAlLOW3GUS1KV82zvdAZkDLq1RiGWVBgjW6qWaSSOExhmDFebWxH/wR8HZoMLxdIyG+mEOEN1hkamV29BmUa0ppFy4CXJXSXV14TLllQAKNbMyDTh3GyX3hMnB3uQz4hW0CvoASpTlMw/zrEarJ+nYC7um9Mw7BJaveVIzBgXyQFvCSMYcsWrzBXIWzMxp3lpRp4pFutQKtuI7gbKM8D3A8nHROtw1YsbKPxGFsApwYrfAER80rSvymcIbbj7/uyjmGHB/ubpbFXU/ASfw/OpAoKd57cfqqVrJITkuf6NZmvaWGCB07UOqf9LsTc/jwDVyXaqXiE5S+IWSV5tVHOAbY5NY++iuWq/aswk0Lww1y2VHYz4fShbOgMQgMpdgHgm+FXqGiHWWiTuEgQs1HnH4O1GBUlLDZ69Z1xPE96OO/6KpC4zAhXN8vdjeBe+dbau9szrrq1bzQ14FNrJnpbrdX/iJArFpweyrqckIJrYK5Rk4Z6DKpVEFOaO1PTl3N2p2VmUf4R5iVR0miceQLxCN0efW5m1ev/7176Ep4/6uvoFzLEzsunrBJz8F4nPXS/EFzjRlHXHPLDQcgGCaBll03Th8GzCHpGA4ZfIMubNg4hIKOFtRS6/sl8iwOs/MtcefF02bdSbRrNrsz3KD7cHE4CYauHfD6KTc7ktIJ6pQx2bGJSGmcCDuVqNeziD24SvekA8h71ngKsoCqFf6WH63tn+/btwAzfnV1jCtczLeLxuq85P0/0S1k6iD4yMF5yRUdVNYpLQHnLBUZxodk6vR0uWkikG+HaOWKVc/xRrrQkFUDo2X91bZncfgOtm+KsrdrQVrReFQeUXZIO0ZHGlJpPr+BbyCP3iiZlhLMZpWK9PeC8EJtQo77YRJ2xzCAPMT98gRYT8s6iheCtdqlvid29Lf/3d+0YTCn2IjHl1mkQDneCFXmDcwEZghjziWclKh94KU0N5IFetyY9AL3gTjOBQTa7ZPRBs8aYquO/pPNlQDHk5SrxRvUe5y1r2W4m7hCm5YU0fOCdZydzybadHAikTl5HRGLEdVeIRCBbyV2CPnaiquxwZk2sDDeAnKMstVFGtkJln/IK+lAw9Z0CoDxiA5B4jS6UE1+54W+PeUbfm8dxEDacRtbKSmH4Ukie5lhMxi28henrsotM5Cry7Jl59pGznMV32uUj90dHUVYXrEDndsDfv9YsSROwzJW6guQ6tsD3u6sXGQvr9NgYQofwg9Fy5Adw6FQ9ahw2QUhsPACy21JHQbMriQo0UT7kZQ0oYuT1it27AKCjppZiFCUVm7G4T3Oa8qz+I4BB+zQaIEKW+tog7rnzSU1HYRj4dKenFwhBHVIuMrTwcIyIhJf7014YYcROmi8InLVhJ0N2gbJro16pcBomiej7H+qHbfXSOTKjJQCsz+1wAmr+iz+DM7XESXAYy+tk9M6N6/jNU6HlSnaUNoyN+Sw80b3kLc+9KvJJuwi2Lm7n3Fdrz5E92ODfzWLzLXqerMS2CEiegrw1RP70yonox1WG3WsXF+mbr6VPOskv2oFRDtPHmp2WDGaWg5amELvOaJTGEa2Vok3+OvX+6YXtuTVo28Pl5XnhbFuYYJ8dF/cX+AUyrYy7qzioDnWzCowV27Kn1hdBwWtUZXCfSA0cDvsFY5Sq4aJZctl05iKlQltp/PqUavOb+irrn4xKNK9MAkixPecBhjNuWJxK1kzs7HKUyIVGifeyPd6a4rmfYNsNZLj0WtfNXp2hwQHaMzq59vvcWbd2qRP3F93komuCaVVU25Cz66Mj/2WnZyYIAZWORfPCwfActladcDXzdsNsDF0F5p+/yJitJI1xFiM7eOHKRGnafo3qu0fm/6DP+8BjdFidV3xaA1m3yTSZgt8IYALubEXTk4LWiadanM3r6dJ9hHEDTOTa5OPJqsarVQjIFAZAUOvPkzL6r7uGTZ01R6xC/MgsavloUSmnZXNzE9Ihwa9Cirx2SmJLibq2ijaTly8sybUw1a9HgkuhMQT3dWoFAb0oMPktstIqriw/zv/WedVQrf20V8FoN0rQ1+jUZAbr2wnr9yZoG9Ec3Tudk2ZI7UuG+pDhI4bK1dm7KbScttT4IAuVPjYLFhoQYGzJ/PqH+Wq7fjR/3n/P2NXdOO4kgNjMKQEbh8O8y3YE4BgQQkMLgPBgH4HeNi9CPYiECBdAivPp3H5ndWsItmtbnk83p35JthsdrFYBQW188eOcIVwnU9nNVPKTCWauGUPz5uggtJEq6eXmnJ8XcFCiaxjaUQjwxbMl4dj6h7vQT6S+VaeZRaYC9ISflbGSvPqM6/7sdEYqPAYvwXJXzgb5fEbyoWCHIep4NV2urRdr4M1syeI9mpZ7Xe5jGzldC+LHQPgKhT126RkBbkEJ/esGQ/zSnUgP3P6V6ac9uEFfLVjII2hOZjfOEn208ZfgB6YlPaTwjFtfmxjVugdbfDAYO9uthxYInuM+lx2iJUMmWc8lQ1sTyK1JVWoV39sN7zk+bLNcVQ2JplINFQjgrRA9XKdBFzHU612wzrlEh77BfBCnZ3g+LxyVKvBI8b9we4kONiKLYxWrPbdpwvXM1hffxLux+dbqb9yR/AjMX+jh4QqVjQvmHxNeDg3VzShF2+ykX8wD3wK9iqrOjippluv+PFQGMiPWuHJhREmmpGG5lxVX5ft6/PKzehzfl7/k9egXITepKtqzgnv+MW4uRXkGLLQXlVAXAQ7cj6yJ1EFBVzLTsWTV6CxtAv+DXgjFOPP3pS2oJpX/43zqrCP85f0DGfI2H/srCqlsp9PzXeotK3MmQN43Ng2iVhstE6oycQqeoMYFJGRVbcOdR1yMeVFkpEb35Ntb2lRl6dztlvHNfj8ar3Stv3Hzt/5+R580H/4fa/BcKbW8enQeKrydoKCWtVQFzAOkXRZOTrtoIQPUx7i3hb1mg6GzCN3kiJsYeTDxoY1cZVawxkMxX2xvIJeX1bb8O0fDwxPbWmXWAzo7EAZDpdvKrenG3LKlPhgzCUcPlL4LvvOii431rDT4vR2mFRCXxgVNebzRoBiDAbnKY6XHr9wBNdsXu3vwb+1Z9j53oih2flUfVNYgBzHE4l8EbzQmqxAxhEPor207ZQRoPx2TKs+TwlFu2D89ck4VrPBxkljta7h33YE7R789VsNCDO6Hw8BRM/vu1CRcEz93kLTsDuX7eUEcZ3WdIhCZhXtSHrqZhuDAa7Cqu0/HOy7+VDJJQia/8yalT2DM6Kl9+Bv0BkKefV8D74rh+E9tkdQpJ1v5yMS0fa8MdfFwKUVclqnyjqtcT0S2V7TzQbGwKFNPxwtkUzKyb55NrYUKynvuT59axQkXJJby5Lcg1m8/fPtrwccGBPVCgh7UJj9tTlCRboV7Ck9heHCQfNlNxOk9B4VMFnXO1XpDRPUwuNmND0KRaxupDfKwRuzbegzPiGvli1crhdVTCbLZ3jQKi/i0lJI2/blX865aPh2InZ8CvpydQ0B346OSnu5JvPvjPQEhmjbpi+RsoU5xAOoQKj0V/t3jbSfsxSqLVyL5dUh1vfMqzMMzfaa45ZXJQZDtTeXr/UIdjS0lo1KRULr/fq3an/59Qh4T/bFTWZum0qxSoY2o6JV2dZqDb3Cdv62P31tlz2vH9m8+pf6xb7HjoJbw3By/giHW821wlbhYSMp1ol8L/R625Sc3fMOpHS9CsVALm0gieFgM+Lm84qAFR6B4z5S6ywdVTiFAY/ZovX1J+Jf5WcTW15BOSZel4f/4lndSV6FS+YRMF88uVegaFsVNAWUQDQ4Fu2gKxEvdClInk2g9Rl5NeIndwRXVK0QLJ7B39AKc9dgdA8+PngDphYlaNrDaPBUHQ8GtyyqauYVVOyJWymFqC1YeA5UrcdDEEx2qveWFgQ5uBnj0Snuv9K7Br3VuoaqvobP1w6Tye5NPI7avUQAACAASURBVN7f49WbM20qray/0gOjZJOKQisQs41uUNnbtFrh7A3keahmPTR0+uHV/kgA2HUij35K8gp0xym6BFfA63zfyDl0sVIBrAzO8OPtnw+6EL8nq4J6AIO+R/VqpUsEKyqx1W3NSLfmqmCXBxh6E4U2N0oN2XAwPBVBCtuIeHZUN+Ouz7teYQWogF49BGmRj+uvwJPJzSae/ZWsdO3GgtqEfstMSZaZtwNYKZPdBPkubd4aj4P4Tu0QelUQICiT7UINs3LkBayZSpOQmQVKUcfvlZm1/fn1xziQvwqamds9mGoLVBatBp5mR1TaSAWybclpB3+Wsuz1/sk8aHMFHZRh6NWKclAB6OxBnIhZTYqEupkNHoJJv7BqkGZpFFZ87vN9l1c/cm+c7R6MKFeA+UhgINJ3RBHdoiPHThF2cYHr1J0LgcpsdA2d+sf3FBqX4RYIakN54VuUPMgemnXTdJ53QChyarU3YECuQrAWV69UzGLvG77Vq51oBXSuSNE+0A51Q/ln09Cowk4rb78EPK7jMxjeyRhJDJ3bZ3b7XLfhaCK/KZyQY0U22lgexi+8A7d6JZ/kHvz1GzhDtmeQvIqWBSsTb2Kw9pW9SpUF6toJqMlhDEPBTSutzSiH9irwaMuBMrqB8uxQ1J+dnOyzTbjA858K6B5uv0UAK/RV4bPel68Yv3rL4QyfP/5+fESKx1VEJxKOdr63qvbcfwxwKsP4Yt/v3T7XQNjKyRVi5UaNEIqbXDdegW4eqJzsecpAMIs7gZZUvmdQz+K3jJZF6K9iwXGlPIIh+gpov0i/EN6BFWVj1BzIatZ1jxuDP9Rb085xoIrw9UWVtBE6OqOBVko0HncvQQnXjLyyMD2zavF5VfTUC/Uq4yahfrGgXDWvxxLQIQoHD9NT3oaF6engPJTUYqMfzA+voKgzcUF+UqKxQDEzK9U47ifx4RRu5xD5hHDdtzPoe9EDr0bBGZzAh16CegNWxxCDhOranlSYTyWhzaOyBhZz2U0GHbpAgePDYuXuQdeGBjYo1gMzSYVeYZ5x9JhX9yWk1eLeOL9+FvurrV456Ko5qz0JNI9fEdpr0Rs/ccSsdtYhTn7n1GeYkNghx9erejb0xYcDezcoNE26onSbQO3gfGvcw6C8AVeZn+onhOy+3n1elbXu5T2Y6NLqAgBA9teAzDM8lVCv4GbGmQR0Y3IQH+02WK+kQnFXl4unQ6lhGN0OF/VOwEfLXIPyCkQTam2oJFbUM5Twq89n3x75pjesVU3kUPItJe3aRFaNe9w6X8rLTjMNDRbFHcnju0XTm+zW90Q+mtveQlGfdv36yh50naOyHpqHOK+4P1jMq8Qutjor+R9MoqxoReW25aFyTOLxKRCurrATrOuc1D8HgpzJM0CRMfqtNOW6UXLWhOQmbpvm2B0SqSU6gVu5kpKV1qvPbF79+5H27BREoZXECxaDSMyZzGP4dspnR8fetnvKMdDQ2AxB3Sj7bGmfbkrfm0wijdS9ac8I1XnpqrBV1DCEUN3dGfyJt3M+r6JRl78Dz69pRC32muvaKNq1A2Q6Om50eRkUdRve2iyAC70J0ebDNU1EY/AKHClmNe+nNkuY1rBjAAozy+Fb7vg/wa/y2obP/ioaDFZen13kBUqlvYkdm9u2aZ0QA4bN17bbQ8eDs5S3gnW7RWq0Q4m9YPMIlX52qF5crBb+XoDq8epb8N8zUM9ydfe4aMF/8C307YnCTnVWTT4CMkfNFZj/qkq7KQu0EPjQI7hpaV8TK12sB+pTsAvSvb5aDWVlY9XboxzFNLO72mNWeOGsC1+CbNcXiZmr7cRkfmZ1Px67wSAE/78jhMIF1Paq6qGyf7ONba6Q7q1zcgK6rcs7kKrZYHr0BwLsk71xxlHIjnIPTpl2fUEjusZ1Xe6/RQpWhPXFlO2If/WIQhWq1FntXKrva9mT7mHziLpV26ms1r/5J4W0uvWxlEAhWpPjGussUGSH7OEcw1Yr6XvsEkJNX9C13+93n1eSWHv+1fMM/vORYqJOj2iLVPWNGG2H7nK6qPsiLsKtrarrnE5F70EGRdlVVeCWHXiNqj4rHHYMbW5SrKYsH9uCtRgOM6+o6c+QSXKl9Spf20O9cstKobtS3YoXy0oNmcd4Cko3eqIOJs9fHR1EtFb0sDYSA1fky+6d481p9Op4C1P5lOe4arXiHGJefWZtlT1kmM8rrqVmPapCfxUN5xt7NBfZtI05TwUxou0SBI2BC/MXBKvNQQy4AoHx9SqCgjXB7AGc3CwC5QqtwsRJRMKfXVmtFpndEFcI15/8hLy6+/6quGMCnCEadnFd99y8UvdoHe8DVqimhKL1SjzAE3IaN9+orQM9AVBjhiMfIHEesY5dXsyZU7gKpu5xBRR3yam7VKz7PTqDvwo8mYC3n5PlZmp8nL6zJwG0OFF5rI1mVZddb4y/oEoVN0yZi4KhHOC4mTwkT3Yk49Vya4nhPbn+QlEP3VV8BpNNryivPhzQfvbCtEf1qjLKFfX4YpnHGmbNBROXvlPBmIGivTfbEsyk1mTuipMNbiAkNzkNj5RlpX0V34B3ya27hC3qRf9jvgA/cnh7YhNkyjEvoJiqpt4xmVfeJI9oVR0jo4P2C4P3JvGO331Bo2LCWJ68GGpkizhTyuBbBC2W0r5GQ5vt+N0X5lUoWl/JLsDPrG5t1F+pMB9mEoc4Xxsyq63ZsjekszuluYtG6RqLhzqZVWN7aAvaH+kbC3FvhP8BVYeyA64VVCuMTu+hUrGrCn+ExPJ5FQrWZ14zU+sVvbqoNfctT6UAszeyhyO0Kxh6hql8BwPiOnEF0lbBrDZMgq8vaetMshyhroHzDe8agWGmdL4lN1803VoIGzNG/Fhtl/Yqo5HyGfIqNRc02f/m2AG1BsYg7DRn4CJ28hKqvIa2qs0N9jnSS5t0ldmk5AiFjtltG+VZxaix3IChauEWTPKqqEn+pvWqIiRqs/nTy7HgqYHYY6sjCR5ELqFec6KFnVkIctRFJGawKU5WAAXYAo8hCKEJe2F1NH+7A2VkI68b/SxJvQo+vG85rfvQX/nX4JnWnrIyf8hOCwBDILMHt9iTL+wt93U7j8kMGiqs6mIDFURaU8IsuUtx1kVwHWhodtdtwRdhmlGomFfr3R3DNK/ympmuXul0Hl6oLzVEa9Vb3eDj1q8q1cyr3SlEammzYGxH4xMN+Smzsz6ArNVs6hRxVq0gWwFgn/GeQa1a0VithXr1+ZnxAN3wK7dZ6TbgqhcWJRe/WBJvwHXxHnjKiultBZz8UFslKXDTRhDZpRF1E4lxprpqusEF1h4gUMGrcPWtPHppXv12uwAZDNneg04Ws/IbAC/MmlXy/9raXjMslNqcxmOnAuPmJ98Pfr7VH7D+uSEPtmPh/NleBPoqRApdwupa9jiv2IvmfC2ZVw1eN9KKVpJTTWldHlW+VVptXfu9yvragpqW57P3nKE6/iyGETnh7JEOw6Pp742khe6Y2QEElQjNMbTA9ur/jF3NiuvYEZ4wZJNZCXmXzWACJmQl2i8gbLQeuJvA3QqBdhNv2iEP0P0EItIbyN40NBcm9svFOvXVz5HqyH1/587yUOeo6qvv5xId2AxngE/R69aZcaRvB0NbzNmL1Y6BVs3Ym2Ylm6fJWr7EKcUOc9xWWep/xd46a0v5ln0qGjszd7KPjwuM9DbA1sFeoEc9tFXR7Zu/V+jbt67H7yd3DKBeUcR87kq4CiuZn65fXoZRR6OtYba6UTC0nLFoIay0/nJMUEuSj9mnQqyaRBnfLnGYkdkLI3ULF3RVqKRh0YjOcFEy+fX4V//WukJIArnyFV/JbaZmVNeC+h0UVz4n1rpiQoMYFzYm0TOVb4NY9C6isS89BHqWjxgUFKDViCEw/B7ds0IugLvz2nHfDo8PwthfiBoz766KWQQq4QwGkJHSElPaGA/FLTxKC3pkPmiFuvKuYWsbhi6yFad2YbmTHwzFmFErHNL8pZrVFTS8Oy83nOZBtsUUa8wvMa72GeLl2buQndOorgIqundQUUaPDfexgcvciml90xr/k469BGK642CwUEtdCKstqatL+r06r/gUcV0VsLnS2fmZWpBE4DkcwXJNkgjYwiGRUlnjBrKyS8zA1CymfmKd3UH2hp1N59BheF7uDXWPbx2BDGOyrjgXwOF+MM6AkwLvCkyi/JmmZOE0x+bQJEDdL6i0PDKrTBD7ZpaAV6lvYCehLWq70C0advlLpTbcLqBxnzVWybrydRPormxIQpqhlkdm9mXmUBjCqovs0w6LHAk4MRyNFRgv6BPUmM6YxsCulxaCDm41CrtKcQXhwuCc3Ft4N7jo+TXqRmM+A6ubjY9v/oX8KUIXZD/odVfO6uZYWRsUprBLXfmvlSp0G+6leM3cOSIuViZZfB0ozOgUldNfuX37Sd8rA16lrl9hKdqgfOy1t4LNle2u5knNtreqWR+vZnxVUtPVCSumbzA4z5jsuHoLlhVg9gug9bW3/W2tv9qG/irn5Fg9qSdEbZD3MkmMNW27bAVnobqaP6y8Y1p0MaN2cQc547s1IzO1DL2+VzMGH+HsGJ0BrtPiVOCFIV1XkC55uonTLqqrnF3Hi2e5gmaVShtUcRxPWWKKvQd1C+KgXSkxZi2ZMhbp9jCpaL2VIIPHuHz4+4KdxGX1vTqnNbxhxkFIAqd15c8AmemBKgInJtRVUYL6X4poXlH2jfVt0gAJerOaiv0qmsqX7Kpnr6zlIY/o5trTEfSFcbArrqG/2I7Bv4NXM+OcV7Thpr/Klfj4lemG1TclmLTlsZg2Enu1QdnM8RiRLElGpXGv906K+R6dWBYKeWFpu0BrLlVwDYLDhKcq9VbFPcP7ayJD4fF/pu9gDojhJZt+Me2qeKZD3dh9M402JSeU0DkdIspjzaiVROo2FWxQ6hUTFKgk+ul337IwopsxrgZBROWlMmR/LLkSVbXgFJ1efd/aTw7AeZGAXR89ZpChRAhjYDAoS3s+DO5Ld31zNP4e4P2vpOMhFRYB34/aQmvlO8npUZmlzQAYZkj1Vva9MpYDO2+PQ0Vl2GlPdqh4sYLn+ARbZZvY61Ebhv3iK6gOj4zxGb5VOmoDvVXfcq/QMSLj8K0GOTAADOG9Sh7Ubfpl6yrh3056nJxtDNVj50tp4GWWlZrThWl5Xy7kb8bEnhepFEwCZIHN0xwb2laYadZfvPVMYkZ4pNE8OPDOhutqXKkq+x08Y+/s5gJ8IlaJDR9XpOASapYHLCZYMExT8z7TLPAjbyQ28UbCEI6PVc1BZpWmLj6pK0qZ6hiKWTKtVCZ/6YU9RCX15K2ybzs97Un9IIvBNbRy1ZOIeFaP88l4MfH4DGbiIFqKZ9M8ML2qNGR+qqIjH1K1boLSSSazBH23CyUX2ipdy9NMM/KCOXlat3Ba+l6Bz7A9JfhXOUFXBdn/p+oqX9oS7Rnjo379AC7txsu4rhXiq9g0hs2aKs8PsxNnK6PT5YJqW4fuSOtA2Z4GtApktIs3N1+d3YTwGVxeX1HwxutFoOPnSYzCZ8cqteSeYV8uLa5kKaicdiAL61p52kU0pmPvJMG6nW+4aBTUXlRkESOGwSHViOp7dX6nZeou4fvBFNGXLFvTVqK2AMYw6epRWgdOSGBq2qKqgLCDoG0wBo698Vp2SbDWSbDTcpqNgpAIWpYVPoGXZL9+xQ2c/pK6en17W8nH+Xxhz9WXF7aO+WISajCHjmKIKVXiKIJw87xXHJdnDcGE8linJ0GDs3N8fOu5qrKZDr1WQqAdxrUP4C2UldbV2xtdQs/LYuLJ8Jv+uIEvaBhWLAZIWUmKcG4YqLc6lNiebjxdMzwruA+tVaiUcg5lHnsDyjHberSepRXxQmVx00f43ugU1k2q6mrfq3OaA/no23Ox50NQyYqZ9kbtGDKN3WA3e0TeLMw9FF4wNnPiXui7gbWxvY6x9ej71u6aB6axSxd6GQwUOszXW8uPoPkOnsnTaZfGGfLcpm98QU6SHTJyZ4+mm9L6sm88Rq25fkd+r9zxptWotzZ+2rkRXbIdRR0B5Bi0odV24YramnukTEe1c992jc3LmCSzyucLoDFzaTO2hi45eaqM1LoBh5FAQYw4Es1VR6nWS+dQTUbve0047ZZmoQOje6A6GrFNsqT0zbrPNSYupyjMgzDyhUHts0e9CNV1YM0ujzelXEHPPs1EpVfww6xNKELl9+sml6TDYl6UzeorDr4VljZAQ+lJTzztN/kM3kLbHvVXb68momo2D1qcPXumBp/cwHjbHNjHJjP2QN3CIaGYl4ygplJLWh+NQTJQC9Jj03F4i2u+AOMFqG64DR3Sq9OoaZjuocVFkxqT3d8/CyDIkOA8j2EkY77QMex5cM6OwEMpqysC2olyJdvmx+EcWSa4FmTW8kkZ52wJu1nKvnurIVGK4+h0V1dTX9eov4Jl5snz2Z7mQaJ88Lo5cQ0LNWjfGBYDmwuUxwM97vysl5F3YQUnBqAyys9uRAWXjibp0KebQK4++gqS8cLQq6scc9llf+PU1HT5bvhP+16dzwk/5MAXtdHNxTOaKEx8C/FuYgaDcQ/dLDC+uK4Ij6nSddXxBdQUQbCNxWIuNmIPT9ZFOvbwc7ykd6d6B+O6el/LH3z0DIb3mD1L2JVMs9KyruD8X8LmY+7dVEtdocXivA1olVzusSSeSsfueOqYXdf0DbxY24UxQYnRm4ev4e0WvVcEIZ88X7Xwtktc1/MEqmIyQ8mJ7xGCN3TCcUjHdSwFxySo0cNJ614+qUYgPtRVFw3Nozh/DaJRgu3CJXX9btwohM/gjapLc3hfz4m8id32b58aF5SvZOzmLJTIdCdhOB8UsVsCkdEzq9AvVGZ2rmp62glrT7iItjDkY25M5NXULuxVxxETzsVAouMl+VrZ8ppwhn/GOi8zPEcZVS8S1/WF+I0DHZghagcf+xIsIte1QgKbK7HErOuUqFlElcyJsYFTYvBhIfYRJhXjOOqqeUx16zcFQwMccwvT8/2/0WripFHYtq52H+LeFCA+VwpXzDUAKpkvOfxtuowHBKdvFvE3YFwxMQ0OA4xd1U5EM7vY251g2/qWvbI9HSOKx9oseMWzDvzqPX6vvP3g4+cHfQefcERz9nukgyrYjWEj5k1eXUmwvBEpiY5rxcEXn8GuNQhfv9AoDb1Q96JVM4F8K1+/q+BWt/Cv6/U+s7+alhO7RV2dvk/+TYV53PP1DWqeMZv2YJjHARHdkPX/UgFQ4djqKFiwdqfBls2zpWGAH2bvOqzqpMMyyoHxda+oGOC7TWUVYIbwxt/eZ/nOHmd7u/3rz4+6YpLoupV2lmlEiS67SAO3ZMZIAmpdc64nbeUxAKZF4KCGCszOKF8/0zWPYj87s0mTqXlIDszyMQxX8Mcs8sX1Ntyefv2UusqT+2aprALGjxpGXDLEUGIvuAyY1zY0DIQNp3T51xCWAqJtZmbMIkNw1P0priHW8Ws8D54Ap7Ka/vM6XcHrH7PIl9126+ydd1MzymKJfG3ZFULNNoEVg+xmYbRT+oZno82p6bVgMRXnI1RNMoO4U/SqNWnyyw4U58QHxYYeuH9Dch/ITRbAvnscG346uZl604dwonxgxlnpGTaxozbnCh6xmSc7lMMsNZbbKo4pmZr2Iye5VJVvxtcqzGcsoQGzd0vkSrnsMjL7AN+NywrTjdSVfAbfFvBVVFfb79/CV3DFYiA3lhVlmJ+NqT1z+Urj9xiFbnAIHCCGxgB8dZKhzRlKmowwY4cOJl6jj3xVR25D/W3glX/dpK6uH/HLPulS3bo6/eOT01Dzr8TsgkkbVPO0b95zZGVZeio4NmnnW1hX3FNVLjOm5e+gGFy1PpfdQAz4Bo5jvDtNoqFX3k3cwjT4x3sEiwZIxtnjPJquD33Xi1X4Knz8kK4kVCL2QuH+ahNLldRHtIbPVaOEK29q7oyFr/p79IvD4qBFRdl70LNHf3HDLSjNNrepqKiurujagyr1HLszzOrq9P0bX8BilceQFeEziCCqEEJFpMd9ifypjRePoPE3opNoUgLwlj0YDBjDS9SYmzYaUfMoRk1D6lG3LzqO6iLvlXRXQtn2MosfR7Xb/fqD1l2r3v/gEGFu5nDPjT5W5SJ5Q3aDZt/cwEYU2+bKjX4znCvJ050pb0aJmSKMfRBaTBKKQUHxJ3D6V2hEr3IF+bmyZRX1oo8n/5dv+Arm+VpAglE4l+pOS4np4fPnUj7gxmAS5hteS9S+TMLGyXc26WZJIxrhwAB7f2IbD5el6C0Cr0JrhYO7ma9guITUNLheYdvt6+73D8DHz9PTJ7HEwQpwrOHczJsWQqWa2R6IR+B1RJ105GtVAtezy9xythGxriF7jKCwD+uP+xXDYLRHldAlm+48q6vHm//LZ/bEyHBi+2+YIYPZeaOsK5zO3kBXlSUyMHrMDJmGdQCu/hR8D/KNaU3wqdxC4XoojY8NBcYxiYVSMQHeI6z9cVQ3LStGkHdb19vw8ev0+q/7z89jduHlazMr5StI8pKEG0olbfuxalhSspr/LYZ8egGduEUYERk4Jr09jSfBK51W6BekrOhlP0dCgGVdnU6/3//0tLXaS3s1ndSBPdqxb97M3ituQyXeuoaoC6J5VzHfmLqSVVfbLcPRg+ZGbOut+wIxrcY1dCFcvSu1pnd52AnpI4WJgRmis5q+j6+v/7n/eLZEFc9HY3bF1tCb4yKvkiWocg3pDsISM+HE0NHQHAlQJXlqKdRll7lBZuZxZXNzm5E+Hj/u9/+9v7/P9vPqNzDHGR5P/mNUfFzD+8dP048/T3/89vjzt5/8H3/5f3tXkOW2DUP3cwxucILeqWtuwBOE6g2ynPX0BpnL1cT/AEFZsmeapOmLzWRsWSLxwU8AAilZxt+LbbyOt9fx9vr68jJebfNS3l7fXka5vNn/N9t8G//Tg29uBuLrKa9Zxr58e//7pHx7v1fCAdMKsl11Ply/KmNWPSr88f5w5XVlaj6q79CuxrznQpaXS/3tdy1ftj+/bF//2r7i7fJxPe6ZqKYF5Cu7QsQaRI17Sx+02Ncs40Gs53k77Go85b0/Ll/hgyr5NHhlV2BrnDLb1h+TqvQEyHy36I4rOxWi3qVBf1QXNKeyR7dnD9zlVyNzr2BqVP+dw/ttquLJomd5ex0PJq/KkDW+5tQeMGK1RrNan49y7YNFxZ1wrM1vD0dW97UYv+ujnseri2khHe30wvG934dywHErUdNpVnIar8ZxJqPN/LBvj2NanUXn6vENH6zmgpJyLJhWx6Pfjfax7f9bx/uW91rltGebH+DVDbXwDw1M/pTjgLGn7xCmvM7UpuOvxVsz5P4Z97OvonYP7TYVPPdBpA2aE9Lm9HT7Hn5fuuN9WehJuxOdkDErNqiWSJxyXAZFTLQVfWgEhMbSfbu7KLvUjiWW7nHlsECAWsbuN8jISs2OK/GVGXPCxqw0VNkUupkq6mw0njO9lhdquxTuCRP1tj11mFJ1m73ecnOKvlDvde2D7821bXOLhlnMte7GZovArjsXvLIrmV7oedZMtrDRZthPPXVVF3Z09mUOILTGoUkre9IzhS6FwHjMEtXIIH6i3xI6JiqrQvOjw6Zugajmawd49tx5vBJcqvcca9hiN74Q59XxJvAwWbNadKPxNLK36/mGsQuP8MpDxGV/x1uqlH1EXV4+0uIAhrXxFJ65zGo0nrQgRdnarpz2PVW13vZBGTlWGFYiS6fGiGW2JN3d0th7stbYTkmdzoCRenuxGOR9vbelg+pB1onraQMj52ql4bI91DFUBDQR2YtOEb0lAbEQw5ngLhE9iu12/blqBHij2sG1sweOhgO0Bu2xo68KoufeZW7FQMK8cDWAAGy8qXtGuEiPkWMLdfKhGLyBI6J52HpU67As9UenoZHqDFaKhavb8coufGmdlkUFQBiJ6ZCfuIlNcMDVQufaXjuHOTt1uBvPJN4T/PSf+gB0R+s0Zpo7DVDdg9iWwMSmrxN/jger+apCD/eLFQZZMtEju/KlGdFcYE7T1gK5+b4YFQSdTlXpBjzaVbNOLb0Grdjyn1zp64BNOfNDyzJCBK0zAwWp7EyLA23pLLJQKbd9sCJkXSLWniw9EqppSLp+pLQsZH5wMvsRQM/v+84157+vvPedxu1cwR1kHee2fb5wmDO4H44AL/qzSjtmfc9Ou9mrdtLqrrhb0Da3EdwLcze2w7bsnoeqj1bs3v/dstXNeGW3F4mdDeUn2tb/sIilCmZS8hGuKq9/jbsbZHwnrKrWnz+ev5gk6yit6mAqeGpXcEP7sRNb+xOcF8UuS9ufvUGoZW3KY4Iczl4qNMCFI6Pd9aAU1qUAICFaYIRcpgv4CH6A+YEFv57hj7ESUlbkKK6f+yCy+3E28NOCiUKXhpYWApGFCXDsGCtK5S28qM2Bm+dWUe86NB7NBVmNH3SZ4kZXj/Blh68B5lmSJvyqZ/jijNoPfI7Zzcftipd0Clbn7W4Rnh8qL08L8YrMoar22wxqu2TQLLKYkj2jbBygbUwrKRWq4s8NB50BrsgRviZ8qFO5C5srvt7DR/NaTyg548rYpWowTQV4MXsdAuGlVfCJtJqm5rOhrw0cV/LHT10oBmEM4sWcLIwOVceN5CZquH7BT7oqPtZT/LrglwVfr/DNlE7xC+RYMv4JrqJBJBs2tGacZqQYsYKhBis4zOtEIt62YsJur6VQI0rTwiY0Yi3ktzBAFbeqY3yQk/BrwpdyjV/P8Qtfz83qzK7E7waBWtHvSg5AHxADpkYdjPPCQ3Uf8j4Jl/dnW5dFnKhbovEOHwT9IHwmAKn/H45XThcTeYyLUAVqZJ7KbKRK8bTEwREmhQcYM8O5OfhsV7CyBi6Kc+TyyiG+t3V84WSXGdIn8dFLnAJr+RxX0QY5rMmssYm+DiAM7wAAAQBJREFUec4Wc3Lx2qglnAX4uRX95fRBvB1sQ+hjMWuVIMDFrfhpj+OLTG3lM/hB1ylRd7gS16JOtokO98QoCkKBYNaNCZJTWOoMvG7Z6Hzl8pBMJcsc1cgG3a6O8COuVie5BH5U/iC+h5xb5bZdrQumMUg1nylLWa02BlqcYz+IoDw7zzjq419XW02YJcZlwc/rS6SuBn75JP5a9bu4WiZA+9njPHjDhndhM7/VfUCVo1hwF39f+bP498oHuapzsM7mRMti4Wmv76gmB5DRn0P8eoZf/w3+D/HB7ypSfm35Qfj/CVe/SXly9eTqydWTqydXT66eXD3Lk6snV7+Mq38AvvCKQsMZ9jkAAAAASUVORK5CYII="></image></g><g transform="matrix(0.9999817609786987,0,0,0.9999817609786987,464.681640625,406.1968994140625)" opacity="1" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path fill="url(#__lottie_element_227)" fill-opacity="1" d=" M-119.43599700927734,-22.22100067138672 C-119.43599700927734,-22.22100067138672 119.43599700927734,-22.22100067138672 119.43599700927734,-22.22100067138672 C131.70799255371094,-22.22100067138672 141.65699768066406,-12.272000312805176 141.65699768066406,0 C141.65699768066406,0 141.65699768066406,0 141.65699768066406,0 C141.65699768066406,12.272000312805176 131.70799255371094,22.22100067138672 119.43599700927734,22.22100067138672 C119.43599700927734,22.22100067138672 -119.43599700927734,22.22100067138672 -119.43599700927734,22.22100067138672 C-131.70799255371094,22.22100067138672 -141.65699768066406,12.272000312805176 -141.65699768066406,0 C-141.65699768066406,0 -141.65699768066406,0 -141.65699768066406,0 C-141.65699768066406,-12.272000312805176 -131.70799255371094,-22.22100067138672 -119.43599700927734,-22.22100067138672z"></path></g></g><g class="png" transform="matrix(0.665988564491272,0,0,0.665988564491272,312.90484619140625,222.76901245117188)" opacity="1" style="display: block;"><image width="461px" height="307px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAc0AAAEzCAMAAABkA6hGAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAC0UExURUdwTJ6cu52bvJ+ev52bu52bu////////9jS4f///5mcv7m+1ZGawf///zSoU////////5WdwP///////////////////1OR1FuR4v///12R4f7//////3GJ2jqV6P///1Ws7jaK7////1Oh5SaL4+xdLepCNXKJ2upCNTSoU/u8A/e8BjSnUqq0Jf///1Ws7nKJ2iem7h+P4SKa5xyD2hp61etENBdx0EKF9Pu7BcPf9oPC8OxpV/QAAAAudFJOUwBgTCGAc00ZCBAzNkA7/R/jETHCQYAoIH+ZX2CnxptIzr5w+N4wcKXLmHTQZOpr4Yf4AAAVd0lEQVR42uydC3fauBLHsQ2O8BGw8nqx1/dgoHnQtN3QhKS9Tb7/97qS/ECyHpZk6Dl7j/60CWBjW/ObGY2E3E4mXl5eXl5eXl5eXl6/SWA+nxvttwKuZ2A+CVZzb/Kr6WYRY2Wr4T2zeOF4jgXzyZXzUbwGtY7jbZpmcZxqQmu9qpmsnU9CCK7WwNO8qlZxvGJ/SzVvAADn04A6tj3N6yprIaYtsTnTxTXPb3oA2H2G3mUUn2kC33deo9OMs3MpRKKQdKILQOnOV1vcnwJCgWiN31pPAN66oK9aOtghyCfXeOc4BW3Ep/QQW0ACO8OZNp3Q7hk/xzTnmVlH7WXboaU83HhB+tA5jVXan2KciyzeLharhmaW4fcpTpbmGve+a7p3nZozuiG+qcmS06wxzmyR4tdZdxKviyrlC5uMvlwTKCk1N9iSTNx0dTVNyn8Vbzmaq5pjVyeRLhIjJS9TjLR2mjbT0p1SXdnldQGabd7dYj6NtddkB54miSlAyDA007r7nbeJe41fr+KbbEsOBno0Mz7He10n07avFvGq5Uzf4ml2FQ1Ds+4UsbZtFUw6SrCO57Qe5mkuOPBeF9OcFiptFeROM4vTNVVT24DtFtdL+PCrFYlaT/P3aNum2pQaf8tk2kGa2zbpdpkWANDN/hCMAFdPBKGn+buGKC01avy2CppwNG/qCO7TXFBGa1K51jEOutErKZMI60W8zboUXtP3NK/ac5Lhx7aGOseDiHQbb+c8TUAGnjcCzRu6d9aMUOIUf7KbZQAxpbWKG+IpjVe8v6d53a4zJdXLoh790ZkB+nxdD+9X2Zq+nWGa5C2Q1RAyMiLBA0y88zqjIYefb9fn2aC0/mDWbKQv0gzTvMlo5zzP/ATfdQQA+xw4f9Jb0svLy8vLy8vLy8vLy8vLy8vLy8vLy8vLy0slgCIidPkjI3pkBP49B76aLcDVLpk5RxIGs2mrWRBGFzobiCB/4ORSBoqS8HzgaRBc7MCIs0UQwOhSRo76RkbXQAmD2tAJ9UdspoA0Y7x5QIKvnj8wbk0AxzciIiRnIWwPDAPShHC83RGc1bZgD4xtcYF4EYw8u4iRe4YJZAeNCOFw1KkQtvhMCHKUBGOtAyA5cNI/MCE8G2cdhS1oQ0YeeDYVgzwitgiji7KcQaCk4d4GFCpjBSXE7GNYBgoLJKQ1zrbAlxUkQJELxvg2NTJS5oLgQjxRoDUrSFx5Aqh0ks5wyNXkuksiTpS4mlxnVmJ2N0cB4aAtwkvk22Q21HJCxcE4Eb5AMEgFOrnfkCfjXRyMQ0weDWcb5GILLUszDmaWGa5cEQZje+TQJHkAB+NERs3GxonsbREa2QLae4lBEkKha6o6X5qhQ0BLqwMjy9Rs7JoADdtsbfXIlH9oEgC8l5gZOZnNxuC0MKWd1ZF52sBWT6xCPjTf1QZnYh4ZydTSFpH5ru44E5vQtjkTsklzILDAaUUIWvQP0DCXuNjiOkbuf9QuY5ifSbUnypECUXKVcLPAmdgVBtFoWyhd2xEnmtmlf3JhRh8AkgbAYn/3TLWvSmFrMI0M6VgWIKYfiGyrPFPjWQcbcItOh6DGTTALoB4bVDzz2pdOHpnYj2fMwt7asY2DOXAw8sxhejx08AGjJvRtnlfPou5Kez8xdCYHc7rEAzTxEziNHMIscADjMlYNh68O9YgXz3JVOZ/q4FVsbmQbF5sb+UnkNHNkjwbNQofzkKwOrGye759V4sMznKKr2BxbNBn0P6fZOgM/mc2cKprANtcq8yy6fyC6d/U23rGSu2eNCs5PAruYv5xtHG2OmxoN+Z9rfWrXVIU7oofPP1p9flC4gd42HJTkWa/Cwjah60hsyDZiYkO5XJZ+4ux/tmkomElZ/uAk5TmQlhLWHXMamSdDnEEwNh2ipCwT6xDph2a++a7QprRKVOHUdSYAWBVCSNaX3H/+0dfne9vgZOOAwDyd6N/28Xw6fnwcj8f6TfynZG0TjQpNVNziIx939SFhwtgmtMmXSphY0CAoxoemZXCGkqt4+CHTvV3PyZmmOp0aZi3Q54+nRu8t4lyRpIcqZfHUu2OjHULFbmfqgH0ipQbm90PfFpFhmrIOztAiNENDmLLg1GVE1jRli7H+jX8dn54YmlTPe7OM2DMNPPZx5h3M45HEaDmUilTbtDQ3uirBYptBvBmXtRKvMYepK1c409ydWBGoTxzNmvTpHOlAE/U9F6qOfZwMTKJiAnMjBxQ6NyuacArUI6Mxa0Mi8zGn2Lj7M0E8Orm/r2tbKUycBaCJm5Snvt6fxNg8nfYmRu9HEGFXIS5YOe2q24q7LmAcQVY0NVEfziZjZB7ZYqLtCqCH1kToswKmzujslrs+TDY0GZpMcKqN3s8mFUWW997geCIjo4tbrGhqjB4Eo2gaO4OYAx5kmVU5g6A2OtN3i6F5roCwPk6S4FQbvW+aBl6RKxItD5MvtQc6HYbmpi5vNxqaSqOj6bhVPsaJWqTxVVnBWp0IMRuq00vv0Sba4+n0gnXegoaN3t9QttBanj2aO1Ojixs6mkVZHA5lWRRFCTcqmsrCNZqOW+MDTL1ByAFffv36RdPsyBOxLbhreJ2hNTQ/yAZuSzmYnYQzotszODph0KNZmaYT8YwNzU0O2y4ARHBSKGgqQxBOJ+NkOkYRWvD1F8X5MPZE8OzoCQVV61T/aWgeG5jNFqxqMITEbFCw7G53uwGaykgRK7qGZm/ap5lT2Ig1CLxKt2l+AKEFv6gM86zmRMz75UubTTt1NF/OlOnf/WAICfkMiWWPlqYqhCRppqHZn5QtFDRVnj2apmEZJLTgvqb59Xzle17Q8ERMC4oXTicZzVZ3g72QkLZ2u2Kno1kYdg5ILAFK2aQP3lNBU0VNkyg//cVqNRmVqtEgzeq/vEpTmucWVAIzNc2XwfoKznSJViJomBAjFc1S2HVjR1M9E/KfJ05/jJsYFGh+qWl+UdIsDN1mZk3zlf5EQzQF94EDNNFw/6jqT8su0eaHA/lVbMIu1W6ME+L/Gc3X8+ODjDDfmdHm+/uRbnmpf+bWNCfaPCsMUOxpNvwwvRC/IjuFl6I5+cTC/HP1r6P5JOi23lLvMLGnCa2KIE2mldPcNDRx9wmbV7mCpnWmnUzSPzo7/D32Kxj7frNw6DfPsF5fJDTPKF9fX4f7TdF9tD1nYk5T3m/WRVBJV/6WZd6VQRZVkO6L7FWL85PNHI+Z26CGJhJq2jt5FWQwQikwJEqK/JDQbLcSDde0spaVO4tEq65p5SOUgo6CziI8L1bTEv1Nu0zdf/ZgOv0gnIjOHvyzfBT23Nc0c8MWMDFbvjISab7XG2qg1WALZJSTvLpV0CyNJyNV400Sm0nReD0qyfxhftnx5l8Ypva/igwNB6z9E4EvFOZy2TdZXsO8M20BE0JIT/OD3VwMmkAyLJzYhKZmLihUjDeJBx++bw6Hw+Y72QfKRyjKWc7BwX/6aX6RuSDhRLjjXBL1g7PpPyuXedo9S1OoaY8szdx+nlZbB5UWaUs9T3uAsFs/kpQbxTyt8hsI025v9DxtP2+ByT/LpQRn0RRB0NTRWaMXr2/0QX68UWgNzlvyvN1CHnfM56GFoxbGBa3G0TXfoRh9v6mENm7pgTwdGXb9ybLFiUSYd+ZJhPEn9MrApI+O5tvbW7cRv6gMLCA9pRznLrdxdPGUdjSVbgJm4Sia5mvExBM9tjiX32pjoG/LpWJeT5PRWaPvCUeq5ndL860hXG95fcsN3ETOOZH1nblVoIjLYizXHoRj+73RHxcXhKHlWY9Y9bNXWa+p+1adtRrscDVAO5q8mOOrl2WoPL1ZQ6vvNLUlSTBqXZAmn47rOIH5YlzJNbA4z3rDeVbwdc0Sbs7o+x42Bc2cuSz11+3ydcioOprA1FUUwgpEOU1FFaRxEzCFvyXRSoMgl+JcijC1q8nYqM+NaBZmRb0sCHIhMHeJGTFd1CMpzVL6bbV2OXswZtGezYdl65Dl0ZlbmYZfCF++/fz5dn50NJk3f+4NTcP5EIJlWVRip7lDDqYRup1Svsj9IKGpjaAxVa3VGjF5N/RNYPkouVj9/XHcbVMV4fWzZdrSbN6kb7Ohr7/5h7szI5etPNhBpVkTreGgEPOiZhOKc2MzJhyx2D20uoNTZjkwyR95lrk0ayX6Lpm1TdWwpOpodu9wMAduzOvdNpX3Q3MHXbOW8b2vRZ/mwM1niXNwWt4frLqlDH1rgT5+k1/p0K2rvG2qn2cxNFuYiVlxJbcNLHa3zSIvyT9zYux/FvdyFTzNwZtmXW/ztf63L7R+k9sFtc42lYYmV2ENu6M0hvI8R0OOO2RT8/ssy4PV5yLHFdL2//BB4PLvmJj4ce/Ww1JJk7uPxMCPI7f7If/X3rkuJ64rYdTAxLEpUhK+5BxvuyZV1Pz2+z/ebkk2MXfLmAxor1WZAoKRTX/qi2Civq2Vx9++bg9svpnbx3qfX/zAS0ZuTXT0Fv73+6yafzbe/rGckofG7DE0aZ5sR8TRxH8foolzYMJWSuNi0sk8Wf8+UfPP/6dczIS3OW7SLieExFEXM2WebCbu/bP2Ps9m3DxZntT8f4Zq/t6cBPDFSHdIvB1o+xh3GBkovPeSm+Rl9i382j5CzPMHbjd1VVVNVf5zWmOtxqrku4Pa6D0IE/9dcjdjVd94ijlxlSrvdeX1Bpazv1cvB3LvNXmAmN67Dnq8Qe9dcqd+WO+1M6zPZq9eB3vtY+yz0a/XLrlecnop5HPwHWJaOUeeKfHeGXbsRsZrv93Mx1vdc4/x8TuHbxfeu+QmY4Vf3vX/T0ZaffXmWzKN3CV/9P7v3lb33YneTNjlOFv47v8/8lJ8Z8l5q69HvM8pa4MRxpnSNmLU1awWE6b5+tcoWyym2GIz1/y/caabTUDWb9MCwM02KqaXzHbSDLwxUUY0Nrlk9esXlGzeJvVBSm7225Hot5ilCZ9ttnTxOtbTuyLZnkHbK1pO7eey3VzrAbVd3u4lc80WqytaTm5Zsr3aaGu1mK31lH0P5zuemV5o95zGKHa2kdh2s/g12eRuopzv95Ws7xvY2mL9CFusL9litXmbo4HgodnNuQbvItmuXX++ewO56de4Hnb5swPfPRdNWznTMm8wcGKbEi42d0Ys24Dw2BaLu1v89X0CD6aKM/JiFc2MvWDTOHSxFOz9t3n6qa5sK8+3fmDb23I9x8DGwb8Hdpc/S9vTA1sselvMMHBie4Z+28IOvH5Qi9zVamMu/s2cbT1n09bENA7tBl6uZx/YjmwGTma1xeLbFjMOvB0YebPeRgAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAACGTyE+SYIfnFumj4/P6cR+xitK4wGDPrWbckV0/7ksOQM3nV3OcQolKUPPp8VEINV/IN9NMpR/FR5ZESm5cZ1K5V2Sm9lFZipovpWb88fVVxPHHZ/z1FceftvaJC7mbRlFGFfQSan65mtakxVjcMHFCqjgWH40L8cvP+AM1XyVvdlg1TUzNnGaFyPf+mbqyN0HNF4y05ubTLVaMfFLJZuK2qBmEmhJ1C9QMRc0PezdFzRdcb76fqGlyp7mHmq9T035a3s/4phRE6j2Lv1DzZdTcf057Jm/K6jP+UgVqvkyoTZLE/LPfeFl9hzepUkmU9N+G8Y0YAAAAAAAAAAAAAAAAQMiYLw/hx3jg3ySKkO/w44im82uJkn9T0VkFxSv/Oglaoicx9mmZwTEx4hPlT8RETqJsgNEWMQMqhhAzIDkRM6Bgi5gByUk1G1Jli9ECkpM4G1AlRJwNyTkxWEByEmdDqmuxVkDOSdYMyTmxVUBlLa4ZUqilBgop1GKpkEItlgoo1JI2Qwq1pE3UhOdMnNgJNeE5yyDsFJCalLT4JqAmEGkB30RNYL0JT6Qmn+yFpCZl0GvAN2L/uSKIxBlSoCVxoiY8Z9ok1AblmjhnWGpS1QZT0eKcQWVNnDMs18Q5Q3JNytqgXJNYG5RrEmuDEpNYG8JSEzmDS5qkzuDERM6gxETOoMQkd4ZRALFQCVJMom0oURb3DMwx0TM4LdHz78fYubs20hvur0mZRo/ANsbFuj8q5OO64wIAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAADARNQ9Lx5uC5Eqld5zEXe8PGiKRrdtXo3SqWn1dCPGut19n7Vty4Nnq7yjuTWObnUUtW2FdKf+UrUdu6vuUGTmRg6LJ5+qlEljdCzUOTXz/jr0jWEyOUah5gVva3W1K40tiyuH5VaIqMzzO6J0ruUUqm3rC2rWlpvTpdIlvnmWWsRUA8e5oeYcgf2ymj7DoOZZlVrVZyN7r5D85XJoWWZp2dgHdSm5qiyLKC5Fgl0ZR7V7Qu7XVqKyNI/SXZM3dR8RZQA7TCmpNpVXK/knI7ZtI78yaprh6/RUTWWer+xI3Y0Zu+zHtudEzTNZ89gjy+8cKvbSNo+pPqeVJi4b3SuXbAtzP+9eJg8ye3ybu/mh7DCZS7WxHGAEVF1yVPKgccOnJ2qa59xllN155XdubBNI7DlR82xFUR2JqavGCWVsV5Wm3pV6U3wzz+teTeNeua1XhmoqbVJws58gupXydNfaU4j8Nriq3Lwuz41vdsOXezWrqpEfZQ5s89IKqiszpQ7HRs3LSWyYvVJtJ3/WOnu1mftVtFet6tTcOfNnB2o2LlZXfT1VmVdKwnU3TXeyep83zanUXvt9TVvY51I3tZR168q8KrYHadQc65tFt04RXdKos7PTSO8fdOs963XFgZpialOT7vrFjoyWSSiXG5UaCY/VrKJen07N0qL6y5CLa1w2sEdmRVHYNI+a4/JmZ2kjzn5FVw3VrAZq1sdqtnvK3tN3sRybtzuj6Ima5bGaR2Wv6g6xF7LTfcZFzRE1rdSyqlezGah50TdP1JT7bsVYZ/1aVsqlSpw4r8xh96gpr8p3dY1v3lpv5mm/8OyzWDr87OyWb9qaVHeR1tzP6n7VsWu1lnSXSdljxrpHzaZP4qh5wzn1Lq7NkqN2hWUR59aMB75pnhCXO/FNeV1TFE1fvOSFqvX+s7nURMbUFcFFL2BmTlifVbNwXFCzdJ9Coua1zJkPk53qslMTHfnm7mC9+a1m94LGytUtVrUaTJXcDaG/3dGe8Kya/ee0Z9Qs3HNE2pvB1thRN25VocySXtuaVGunprbq1Hkuv5YHYndto26szceumXXuzN63Q+n8+9uYndY2qrqh5CAzsJLVq+4fRI3uyrBGd+TmudhOrcGF1Lo7j5LzN/urgzOfhw+/5vLdnP7wePW4KMIXmgAAAAAAAAAAAAAAAAAAAAAAAAAAQfEvTUPTBH0zBT4AAAAASUVORK5CYII="></image></g><g mask="url(#__lottie_element_231)" style="display: block;"><g class="png" transform="matrix(0.665988564491272,0,0,0.665988564491272,140.6163787841797,84.54312133789062)" opacity="0.7786332375769343"><image width="793px" height="629px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAxkAAAJ1BAMAAABHjCBZAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAqUExURUdwTAQEHwECIAABIAECIAECIAECIAsVPAECIBghSw8cQxkPPgkKLCcUSUzXulAAAAAHdFJOUwAXSd/CoYCo7AcqAAAgAElEQVR42uxbTW7jOhL2Ed7MEeYKnUUvskv6Bsk7wCzeFQiIOoAXdQ8D9gEICN5lIQLmoaZ+viIpu9PtTqdtY2BGlvhTrFJ99UNKdlare7mXe7mXe7mXe7mXe/nt8q97uZHy1193a9xSWa3+cy83Uv59t8ZNldXqn3u5lcLWeL1CeXl9/YbKizXkr1b18vLami/Wsi7tA4tvtfelNkDQs3xpfY2fVz5d/sty8Fuv8I/K3/9drZ6eH+TgTzs9obPVdOS5HR3tguho9vMxiR7Pz0/PNlsq+uHOh1Z9wgnNZxt50pOOPYDGJ9mhDeUvBI3lAxg9YMrTcyP/A/Kt2gYfoPAJJsvG8+s/q9WXe7mR8vz3alW+yt+bHHx6fKNHbWknSSfJ3+ObnL96v5DZFCPXQaNrc6wTY6AqPI3Lo1zftFL0+qa1gsG3smiiai2Z9/gGKp9vx6OPKzsTYP2PLqLyfqyjny5fJbdByH10/Qn1rzgqeA9sjfV1C9VaXl5pOWqt/CscqWOYT0joiPmnyT+P6Dvl4dvVrZEVlrxADRhk8t4Fcg5TRif5IV0nqOeGfmXWS8+0/jPyad1IzvPLL9e3xrGO0Cd3HmqoyAB14Kjmjmbnk11/NbcN+rgwb8x6C3yGfDNt1312+fJ6PWuoIjnD7zpoMlUMaOGneeGSJ9khV8RzlbDwdQjJnetW+j8hv82nm44NOk6v5A6Z++TrursrW6u5uuImNq3jGb6ZKz4V2myAConO80Ug02/Kh32yo2/BZyJqD51hkSvFRm6uyXfpkFJbRUgr2dsOBxQnqk7uGtePAZQBrINyFCOVPMMUef1h+Z2tOku14X7lzzcYG3RsFoKLVlcGRAR7KRmR546MzJ/NkNaVARH1q3VvDDLLCxuLEnd86qT9unwzO5GbyCX47VVWt7lueE7IUD/3mqiz4qJIkMHla6qibioCC3CCWxtXA8PDbmEO94Cc4fPkJvqg/H69aB7hwzVaz7HI1WLDF1IglYGE95BpZsoVw61wK7sH5wzI4XvwSALObZ2oIuDaFkTm9wat8rHxD8hfr8EEJl0X5EkimDVTtxfodmg3ERuevyt+mQwZgvqqJXkSJ8sVpqPGEIiRO4Q0ZwQCGGZBzqqFWhGAcwcTVU9HBHxEvhsUQVncMB73XQD6wnQrsdFC1lya1tVNgVZvF4VyrQgWVbjoEFSneib3YzrBBxVsEhxKL7CZkZWPySfK1Ey/ppotxYLmalxbt+fE9Q82vFfaU1FdFavq2ZQvVV8gpoeMiZLN1QGnoUQVQbL5hRrszh0xswgW5UvNOL8sv5Mq/Aq6WrAgg/nyRjezbrSHK2wtc6ebqFT1qzoJVqxhgb5AZt3QLjqvVECEQtuW5m1AGBiwENBDpsa03l+S30p3M9DIDVy6YOue1G9j3WhPwnXrU+9a/LywYoDWvVLfnkpbhtR3uak9hp1SwPkNM2OmU6kg5pSomN1goVLQb13y9yH5LtoErEu1QvUV8qTmi89t7al8iSC4UjZIMs6kOJh67qprbSk6BpN5vvRmQ8N7C3l0wJVtTJnJ4aTq1JUKsfMB+WbY7J5gliGXR75pqG8C2uPiDawb5A9LtvUxrCw3wFmpwmVpuwAf6EmeOZB9rEYtMBpsilJBJmr+6hyLN1SUR8m58lv2qysVcqIZw24NhvUUaBsreu+x/NKxYa/U6v48AytVVAGEhQg5yMAonjEA2iI2iicpndJB7bZAUJBxMpOZFxh7I1Eu58tvDLr7p8YdPJEdqdTH8/yjr0EuGBu29fZdpqPmSrv6paoO9KwDSb71gUaxxHyA1/PAglF8FuU6sSNeyD1TPoKpNwdCtmD5MXMgyiy48Fz4Xqa6eGwQ3okQeURXpXpAjgFatGgx5d1C77bIbVSh9vHz5R+NOYucux5qFUK287c4N/K8UZ/AOkB+hOLnFfpkuvfnd3YiQjZDqe8i6aqxQXUFx8bQ1ts/iP6tFOrymiSto+8LrxQbvoJblso/zTT/R/bArsqCA7mKrr1ukH/52h6sFmXW8/hz9ebfQ2d+h8mflm9b8pqsrrtu4LW15amujHMZRzmVWU58jNann7nosDesOs9SlcO6jdo7R5DNldEMUkgCeRmdpZH+gnzvgmQ7dLrWTi3WHjLr+6qTndXFYsNfYdaH384YepgiqqbCYXqhC3SzO+7sRtT6DIceR+2c0aXUI/A1nrPxM8Y+Ubu8fa78FkZzgVSbaazE/EXttNh3+Wvh937pcKHYyOvuS5pSHwDUxeC7Za6Oq8q4m7qnK0hCOcND1ZfddfUzu9fXcPFYAVrw39EiwSaNZRxbxJwpH11WNynVlKOZysjmeV6kPqrb3Gs/i/ui0eUpuPRY04WrWaBrB6e1kBSKj3oWquiPbpy5dMmsca15JvQpzXicIb+KMXuZfDCcESC4HbNO8Qt2VYiO7z4AXvR5o74khCk6pZvLj4uGl3kGCEHG7HxMgeLEcrYSlmKagLmbdMzoHfkLQWO3lJjhx24dMWuNXYYr2N0TXf0dbnZj5FLzcKc+IxysFkLwrhE+x33ckn45dEYwk8Aygo0wCKi2mQJs6OhkprHEHG+18R/JN956Fzo0Nr41SuejHqw5vt3FK9N8+qbqQrHRvvj3pz3Lwyd4jqJxQZ0NZZew9McAmByMRmR9QQ0QgLd3KZm0YJxiti0+DIKfyQ/Ke6wCQ50nhpphK9yoR1gzB77J+u46ftF1A78HaBtbdUS97xD57zCP6TDuwz6EOCeu81ic93GOPDDvgzVS2HOL+w5BRuK4l+FDEAqhFSquKCs+C92e60l70mGOSiysRj0HoeMpJqbJn0XMsXzuVw6zyLEogd1Gv869d2jAFV+amj3W339ddZnYwC+dSg2NLpQZk8AqHhTdEA7s+Pu9qhkOUdKR4BQEjiTqB6YLgiOTsq5yTUom+AmMMk/oNLHwOYrLJg2SoMwsESFOmEMQ6mDmaPK5lx0iLeXHWZt8z6MYh09sGZUThIOOixw7dYbqosO/Q8cvsa4TG3m9XoaGLsmjQqqpWu4/ChiJQZ3TxA4czFRMsTcCcWdGRo2h0An4jPQ+pJTELsxtLxTi+GhI3PFY0lCIMoEdPMqEKCHC2Eos7VVMJz8eJPss5Scxntqf/0ZxDo0PiTWNqSgxFZH5QsAKVdNcNQe+mD39guNi6wYeNTIWDUQye3FkXdJeUGZPlIQVBCnRWEDTpBHF/zEu2vMog7Ib9SqUQZAReyRJM8oj4CpTuD9pS848Syxk6zKzjiZzryHWyRcjLuX7AAsT84stR7EfYiNYrJmcGLDy+wo04pnEvu/CL+qu+A7Xf2DhmapoRkl6+w6VXkVjhVigimGjYGA8pP1kNohhsu6kM+SzEZ+VKdEKAiClarZ9UlzVAtYP2akZxkxhIkIvXwf2Js9uT284atwdNChTstA5iAdE2Mn2E8FfHpTiP1Q8eXV4kdhoP2Ar/pTKzsIudjioA8aQgisZTe/k7hwMvBABcxiikQyaMByzIRj8cGHz1YSPIq/sB3PcISTGeAjVaIFNqwi7fOMSYyffYmOAXc1kg91+jcYQsJIEDcl9QIiMeN8g5sh1m3u9PdXyMdyX72DulTYTozwMYZhY3V0IouVm4oYAM6TNkJR4txnEDmyQsBMYJ568G4QuRh5igoEDhMnjbscDfBHC3Ubm8VAcmGmSg/+mIU0Tk4QNnxPLT3FX5TP3nYgSbpCfNNBMPt8Xy9aW+YmmMgsT7gzR86ScRpjJ3uBYaBRs+K+2buALP93aWmTo+speJhAzeptdHCYBb9hx2U5RQB22A4O52224vduFKU0bQXPDUA8MsuAsQDOGPMTo8mcj+G8UwY0QcIXPO2HGLBlK5jBMPIHNsxNLsVyx22azrfIHpom7LfdW+SoPI9vElCxIGE1BbnSoiTYioDz7Jn+WnO3dg9mDcm7/CXXh2Mj4iXf3An3UHY3c7UZwDKxu2k6qtiC0HTZbBnIrASLIs89vuSndbI/tJMYQmwQ13LCdJqYWGwjUjBt3pq0EyFZtlJgbY6146iCjyBfhoZJElMSKyucJTCHyxAaQH3XmVkZ2OsBhwxEtoTRM7EaJ5wkd21hiJ4gvBF8BdUNoG13/ppy+997wYu9w12v/0d//qLu65MR1JrpsqgwLmAc2IKl4piTXPKfke9+h6mMDntV8fU53SwaSTDIh5g5JANuSbHfr9L8cun1byFTwgqQnzSa5T8LiqJtyg5zp4NHyJQzyPYW9toXNfId+YrtM5IB80f3Sk8QEzsid9tLzjzx/OdolvOBMk51fzyHbWwLiHwwuHJM9A2ADvoFlAjmIPpF2wOTGbQ/z3ncWYz/31SurY8OT4RYvRCjtX7H7REjh3kAcuT/cM24QPyBlIZNGbkyTHxn1Dc2FEHK4kNTD2BqhSynaCezEcXJVjw6TDlkUB3omHQQ9il0H0DPaaDq0dimjnlYvRUCxBbBGIpjQgSrqrKFYNOt3ozEXmlVaBfY8m+qHJmCNGTRzMGcMFDaRnSH27htjaVPd2UHqOzb0txhF0VFpNlpfUFnxwl02ZCMvYWIs4I6pfTdoOJNsdijy9A1nGRQkA9mhygRfZL+IM7G7VZ97yF3Z4UWH+6fYVDeuH9xvMXdGXLdSpRS7bd6jErcTVHdishajpJJT93k/6zp5/9Hb6L6x/Tka7Ptit0JLT9A+DJnOygUzeMmcFS/FhR8+oGaOI+y4QnPBdEdXHYaNZ8RwdQGYhqh2qjVgL6rCUFEyliofqcRaUhxTls0xKT3yWKL8pFIpxFLJVXZN0r6SGnWMclyIEKV1QodJdlX5kN2gVpW2ueJwko885TGmKG95krHGKD940/PLmSO645h0o8aosgtniGM2Di/47J8+j47lpajCGmhz0K6ggbtTRX7yPMf+LjiyHjZYn9ADhuJSABtKzjoJDeKUeMsgay5TrFlIUzPICIIJoyI2s1BZWsirCv2i0BWsq5N8ET4KebOQvqCvDJVAd7YtGELGLeQrCS8NyKMoffDB88tPBaOwI+h5ZAhhROQVTRwu4UQyZsVUksvJk4GmtC/AG7T+oObX0XSHVUNYSvZOja+BDV1hqTV4zL4oNAzoP4VEKQndOF1BTSElSACqYl5Wo09SgGT8ADegPviYiQ22KeBJFBInjKGMEajJuwwqh0fwTtCHjwwSy84qrJR+2CHjCVuln0wGuSLpKuevo14Tz5EVLXkEYHGxeZoy+puiMgE3Gl7ESqYxvh3UWdd8iuZkFwtsVvc3dOmJFybs6DZTXUCiJNwXJY7QiczIioxaFQ/4wwc+gSMhXSZFAZhJQQMYcAgABkOAmIIKORZTyGSBHtXR7B1UjqGCjxgNfBEuYQf6hCxXgENCczkRJkbOHCVjRog0y8TbhCbGE+OEaZkjTerjwuXwtNOP8/kpMVwWUpmvAYsKoTwYtSNEQZ1C5nx3ClXM3ugvCiAhETlEfoBhEDEpkAEKFqVtBGMC6K9cjEAcjlJTgJWBo3D8ZJ058VXOsX9W8cbTjNyhE4LvlIwyvm6S8RXMoXTEsFNT+80MEGklvy/KEC3rsTWLN8GRNbBhKy7d9YOzITbGQJMRM5sCJfPOlYh6n5z7lC/KFuymnoBAJ+3RRmawUgUtRn7HBvfKhA0ksfIJfQPAE1uXBUiyIoOsCTnauUKJPjr722ilADzQTcDaSGxUbBRCjygx7Q5fH/YubF3mwDwFeD4/CRswqZQZiIrQOaXDB1MG1IghgAKgof8FoVHIy1ckhGSntAxsG0jjoH+yn1/tmHUoGC5qQ92tW4Eb/qbb8o5BOEqCcIvXF2CDBrBLWQsIivRSBkVKL5oJQEmzjokOUR5dVO1cc/hjL1bVG/seMRRnFElX0eJARoYl9CtQGSSlrwgiUD0pvVKogfSXraoEF4JIDzZAF7QG6+S3YLd8Vm0KfsYZWzgoVAw/M47UCEpLM2WAYjHp+cE+NJEuOBW+2vlxKcrTDI6hqTTJAJHKvmjwpuYHgLOhQz1BBNHc57CVPE/DhkbTzb5lXoDIKCaL5Z7lhmv4JTSrIR6EciHJD6enkAdMkUYJNAkKDjRkm4SPgB06ChrKT8KeX0GPVLSTUSLbsiP2Ru+Ihjw/21YexDb4p+fnuImXYPyIXX6aOZAoVmlqASCFok/VB5kxtMqgS8PG+jHcvWXEPWIo0NgcqTU4q8gBzkElHGQBBY4TjfTyjwAtDT4REMFakVXaNXEqg+iETgyxjQ0GmDBShgqLZhkmca/RW8fMztvsJ846CTgfKBdNsrYXlE0xqxpgh8sCfpgGGQYLWJ2s1m3vi2tWt6k8s6GOH/Jto3puIJbRQjkSjAezzWqqFJU8Wefo8pWjkcxeKdy/DvYZtZm3nY1Bdoxnsq+H2Piu00MZVilLKSCJGONHMwGifTFDmZ4kEKK21VH1uNUsXi6vBdVXiuHue7XIbotaDA3TQeTqfFxQBnPxkEnZfE/btGjV8ML+0bdj6AxWJmRrS8xgenPnQeUPj1QOnRuLlTf57gqUIToENJKZHWBFMWzQQo/U6oRGbop864H1Xh994wCuFcM971vEUJAxvMDBjV1EhJhCJ0hesKdR3BpSVsRXeZR96jsNc3qNoVfguiP23SEbtUaVUtYo+pFMePMth+z2leqU4lqEPsdRzSotILHq6OtnK6zkizs2UFMJL0gs8DqNEOxdK8R3KPbaseg96+vo+R0j+qBvnllVkUpLHzzHdk2m3oiPQnvauLDwk/LCzF0ojmZVrW5T6TMEvCx9hxoYuKo0Dt8jxQNf6cHjmVnnQKH6ys0/MWBE9dAVHMjewufoDsfFnrXyHGxYpmmzk2sSpZFKTeG3kuTxr/oFFtwLMSqg7K5SV+iRAQGqc1ccEFZLo4rLY2/Sf2vojV7UdtECaGRksjpQf92LHk4wGzkGNZeDsaO7/MqWom/KjLIZzKhybFxuno60AjbOqsU11YRM0wmFHshqMCJxaCbof/Y1v6N7VJsbL1STNIMXb12PQ1h5XZW5f3fLxlfRG3xcnWHj5SQm1cTIdHoQSVYbYWEAxLAwqoJFDbILrKbKo7l/4EXRWJUu6ljkONbWG+cfFhfRLCwKpyiqQvh55aB9y+s18D3ihBbrNNe8KxB30anAy8KyMht3o6GRtph//Xoqe6iWrttjBpZiKs+fmtaHmx3zOxywg/PbHHmNKX/AJcoqjeaEkJuF20CyNHFbHPfU9cb6MVxdSmOSCpkmkaZj8kjImww4fFUize/Q+vA7Rn5MXsWmO3qghCwpS68jetJJ/T/mxs+Xp/jiZ5YZukV12mxZ0Ncs9vA5OTI/Wo79ESii+0lZoeHq2xIniopsISsL4wIbR3XGW258bX9jv3DFETbc/m9zHBihuobG/HX6z2trd3X91Jq68gEZIIlNeRhHLMVhFq4tcboCxzo2VeMFwulblirn/JqHPIe/5hWXURVPWfa3FrBqYcMjHQ5i49RjuEsbdx2bar/MNW2HYShFExt/6yu6lLIUrllVzfsrzcJlZCSb/zd0bGhN1e0TWdfQG8TGWR8kAr0haiOleB+jOHxV+j9cnbwbJskWuY8NHARGKD0xWDRG0iMjR8tw2FOrbkqqvh0b+6tl4iKpsJAO1ZnM3M1fFvTzN7Pl7YhVbqEqK68I7o0v47jMb+SWjnVseL7p5img34+Nc2eG2lRYtBJj/FjE8L+sSOi/5iawTG/3IFUPqmfygg4Hl/x7nfr+Kev+ut74d/OCZQIFNW2fC+B++7T/uJth6Q1LLDavb6m7WT/KvwYNx4bXKVzObd34qnpjUWgocwOrgT4dpvoG/hy+oMFj0+JWrkBDN8Tc1bgW8VBtuBLnco7Txtbxq7+xui9+toU0fD4Q9cYY69vMmL+Pqo+Rez1nHi1eaFaV1kmrSVW8erFHRrreOPlTqPdPqRnxoCEePMC1RsyDH75VMn2LaOtehpclKjaWLrhVi1pdVcy3JVWNHfubB/Gskt9oJdH0/qA3UIz/TmjkI5P4aeo9LwpTWgg3X2f+mIIt7oV7guPYkn+n9uiw/br5DXuaoZV9noiNLYvzFkSdv2kmL0KA80NYG1tNUDdmu2Wrhb/UFZoXv0rE4uEElhg3vWGu33nlOlzPxO52eBgH1z/GP0pRz19uND9ChuUGjHCVe+1pPxVXS62BuOHmeFU1Yv+8Zk29gXyjPuwdVXYbroOtOX6qQGH+YrP5MarGlbYXjXjd+pId6mV4xYh9db1xdAOXSzguN/8AYrV1f63wU/TGtEVicv1qkUe6fbZqwa1cd7+V+EQGmRGX2PCakaY39vsbtbFODLc/RAE2FVYla935/Ww8rOz7HT6pv9X7bjGqfLvGw/0M8/tMVFm14TBc6Y1Le1TxitjYL01cPkJBXPFYeVvzw2h4WMtzt2x4bKGQuAiKlO7zuRJvuVis4dj4UuVWibturaE9fKctFd9ytQBAnD6qCeZvQ8X8SWS4QUuMxEWAqtUYthL1hTFlcqopDsOG/3eU/co21QIbWPInzEhJY7jP9+Y+62pEJsJbwNYllYcHTW14TVteGFV8kJCV8JyehA0rUKdNBWsCjxQcpsJ7SeuK/S+Jp9wKCy39Ghaiqpe0WehW2XBl3noFz2nT4lR3kZG1sHHpRdF8tsiUPsCLrzsXDwZGNgNXl3pqYKRbU6WbVJbX6OZtOTZs+FozPqTqZv3+Gr64/WMBlFNhqbiwo8Qc76vTD0+0lz4QDHGOaOVt7ottFRgtp2GMcUh09TFs+jrM3eWy+KfY6+bFPdtEWTVwNWyK1ySbr6l4+D9117KsNrZDP5uUYd5n4B/wpjx2aVM9pqAyx5XwA/TXXOstA+bwMJtzSXcqmSUo2pKW1lp6QzK0z29drZWSzjZSp0BxQvDEoDCMyjjC6VjETU7DhaNwbvBxg6Oghmg5vO02JD55sWycnonECzsN6qlYWu7YlJRwFjUpAxriExVxkcqsX2TX9AFNrCsGKDe21X6fd4+3VM8EqW1m3GnI4pXEGknV/jL0kWaGA3LW2XpHFRTj1lSdiZQL1I1aXyoMxpZaqh2JTVL7poQ4zVu9nRkSaDrujKG1W5WXWcAR3oen8FSJYty1TWcXxwtx1B3DXS0X6GmBBjvr5lOf9vGIyLQHCpBAA97ghmBAHueGt1TmpiAdLvdUfWGcqjctDdpaLBZLnMXXumo6zfg6tW+IBahOGRpjhgCk5gy31YhwLNJF0WAWbqX8nUO0fSm4F+91L87QCN5rGHIDnc/S/wNJR9+q1AQcJDDRCZxiri3P39rcQj7fbrBggIGqKQPvontxFAwgTpX3pIeFj74+94ZCduCy8gPZhUMawbay18gRuT3LDPHTk624zBvle6pec4Pcq5fLZd5nZn6273iIZg9F0mAoMyQF0xdzthJc6gyciriIdLiLIMM8tzYs0FN9+Q2U4aXaoqsFzqs//oEaYbcgLxYFJDfjVR8P4iAP1tVIWIO7ipLYM7voIvuNXtQ0yIreEoNnk0Tz8Opb0xYIhvwAkJ0fyHDhjELZ9SkSooZIebz464T4uZDjD7z7K81Rjx0umejlTMBIeqWHen9m8ePUGJ1QqZ1OFkky9yUw6hRMzH650yIeWiqW/ZXNDeZT1YKoD8HAkwJE/JxaJc38TbcvBkUeKnCIMKcgQTY+Qp7+dJ3tmlj3d5B7Wme3zYro/uQ0rGxiOyK33dlTfWrFpEkB7riolELTZ/gblQSekpCcjYAdK2kkFgs5HUuM6LosR73+CoZIQ47i9ZM9mmN+w+A5vSkjTo9W8CQkdM0NFckgkS3SQ2Tsg2vPVMwNVu8fLTfqwro/yw0+t7hFMc26+Z6l0N5RPt6XGSyQsf/MdzT6iPi6z/0rrjxZFf5ggroSeHj6k01sX7Ru1KpQPuJhvgXeTGBNbIHP6fleqvEdnwUi2uNmods6EjJVOvheioSDxr9/RGn2VXwWt8PJfIoVcwN9zX/uhJGMVchZ0Zx568h6SdkIAqPDRAHnstHJ5s+tFIhp2PeF60bgU9GJvyW5RKdrMswfU9HB4oEIVWaPHQuH2OUJeUqKhozh194pPjillBFV72Of+Ql/KrO1OPJ53o5M+6+9KLN83+3Ui9U+8EY1hhSOraYxFOwCZqRn+/7TRAknLpXkBh6SHZ6I+njNhKeQH64c+luh7m9Jx6soOeQbat+VHqdX67ij6JoYTu2UaS/JdmnimQpKGq4bB86NlY/iH8CpVE3z5zdeH9NYvB6DduY3TSzdlakTrBGC3tXohMlHi3S1emc5qRV6KmmpZPYrfZum7+XK+1A20CaaXLvJ73c9c+/UzpIiydRLNmeYS17jOGECiHOGBCVdxkMAdRXE0tGHcGe8fN0wlgKqadAQaZfzz1QM8B9KGlptpyhHciB4jmfwqca2M/qt0AxNnxxII3VxTazmBtHbFtsK7Vf/fdNe/PRiMARaTkZkk6TITRDlR70MgEUkXdZv7KWobGS93eRXBkTYVFQTK8eTmWuI59sRUd+vb8BU7et8tJd3TO6JJwxbCQeSJIULMrEAH9fviuPB3lRmh3sgs7DjhZimUG74KhbJbfu8NhVmEaDjkfHbjgk06uHSaE4YVqjUqalWynYaWd4pjIhuxRWn6tWeqi48i39Jg4uTD1m35by5rVC+IzgzBy810crT8BDdugp6C04Nsa1rurZc0lgILoJVw/UCBIywXry0V/RoFkdJLIr318291sRFIXVhFOoT1fhaKQU9AKMiUwtwyYoqQOk+/AWbkbr+gI967zgV7cXZmxg26b+5CnQ7RxRAg8FTn6oCRmwdsHBkZyRcwW07A6g6Sg4G0xlPx+nv6IZI411sIY76l+3F0Uev2uPpFnjB0vwN71Vq1N9IBQHcQwXzLx3CM3e7BhROLl8ZL6TxzxgjolD+p+77y8tNhbiGglQt/g4tLp6Khut6ca4AACAASURBVN08wZi9jEtaiPB4+E1u3BPdLFU9Clew26oLiEjuJCBK32HZX+0HOEr7Gjqf6oh0KuRT5VQkGKcHyzgfsAO7P0eZ4ed/INsrJTqNdA0Noayo6GO/qCwcfp1Upr+COFX9FflU2FMh13C326TnhvH2LSkhE7fNf+aKzg+V23+p2zDkm+ulwPZU0zZx7V6oi7pW8cIYbu3n/rCnQk3shl5eCsfPOTCQ7MZTstHPpowgzzhjn1+swqtO1chUw7tO0sNIuIvDPyNkpDBOVRspGluKBar31+qlcOe//NMbkwjMWlhNn3X7HalTPH+PdK/Ts1813jNRSyW7JroUq672H7i/QfOm+hqu6AomXlWHz5aJCzBEVX0sG2uEyznyizTilE97cImIVLLp4x1TVynNsAub2JXcNCvvFf2lpvbY2/1ZDX+y/Z7kc+vpb/FUtJGy6du9VHnICH6ROYsvXiBNTeQExYPzglU0WDeyrsVVu0/hKOyHK5QR4YwcDr+3SwZG1vDf869PO1d+MBICyRynPDeSqisFRA+G6EbXSZf7DPzuGQyR3LCyYS4jKz3A8QGvaBIaElA1/Dm2f5CHu5drxDM+Pu0LadEorxDUu1METITd6jJDeCE3oMIqUKDpyZJybupkHsVjbvSRTlVoL24tLtYNBEYQU381EKf5PGFEpqH3AnTwzkHTp84IKfAR0jUYveLKrT9IC5ud2saL2KGOy3qjLn6bxo9orXDbtMXc8MOL72KJ3E2bIjKCVAzzNwpnye00Mq+UYIrKJjhhRVNG1WnJkF5XHypdNjHTsDCfKpjaD40E5QbRFHabC8VA+4nWKsXcgOQIlV7tA/vfzabSxHpJ1300i1NmZH2oKiPwHPSI1pXkKMIZEXdiNJlckI36/XXjfYEBdacHMTmysw1+qLqRkgEZfKMBU4lhEDpDI5gnC3F7GWJR2fBHsr/6Q7q/gKhzbgzT334N/9rp6k/O36DsTmNNOTXEZj0NRoIpqwQmhjh22+VK934Ul04uaEXmZ338gLapNrIhzeJ0W3x3RWhWHK5lQ/TkjPRYKeiXRmODMVZ7RfdaKRm9Y8jQsRHODYXTaTP+iyjqfXmciidxFTcdV8shZcVL4QdwdfiRavQWchMOvgbsFtz4eULvatsMmfh47OOfKq7jC2VF0zGUg1wKqsvfbYqk6NUSp789einsSjZQt5bgttLQIBj/PKkHW3I1PtxGbBlFlzjYx2Y/pOEeKTfkonVpDPcr3t9Ae+K8T2QyCbO3SKcHQ+Kn2l0oI/XbIHSIYZhqqbqQGxYP+nCPW3Xno/hH9htaxWUTu9j+XW4quviQimfElWkDRu5fKSxdDSxk9lQS+to1sLCzcFAgtHQwRqIPVafrDZo3fpFt97lgoIiXAhkUUx3/i7lR0cWHOUwmn2/JoPETWOBHXl0x42ChWCVEDjRMcdElJajL1TiEXZOY4TqFhxk8fdHdH4053FOttthSDT3VMP09kQXtXGnROE4I0crFrh/zekmPIMMNuq0gUpQNAt+GBawM4zhxmF5AGlzSbpfVxLI3mdeN5YFyY494OowAp8uvvX3rG5aS7cHx1QJVlNlPZlGop8nybWohv1Q6ZVh7K+9UYOFaPC689Eppm1S9P8wb3TBvINfwPXSc9r6HCnTma4yTkKKCSSH0lFxMlq6Rnw0U4ZpRVcJo6+xngQ31nVpIf6vLpsIc9dp5uOgWVqGV3hr/3rv31YXviTqyeA0eqsH5C8y+EzQpUhj94IKIINitCPxsx2TTn38YwQ0ePCVzQ9T7nhvbJfGpZPgrTlJQexNeZiQ1dIHQUwVrnZyipeo3ImQKSqduVPJSdSwyqww1tFso9aXpSzkPHqkbKIrlGwO7GYfx02P5BGZzpN5Tbr6tzGf5lcZAzs1MqMm0cks6xHBo5TAI9yh7cWJFn9EUCuvF8YQygoa7d5AU7ohIGtUNqRkBD6FSDn66wWgKN2QaTg1x9g63VFLLHVAf2eEeP5Mb5KMuri+LJTJ4AAvHrGzD9oH2Vodw6afSaPSzK9SR43nHhwPhzgm5isNGZ0xDH8ZlE1u6bigNF8vGb8yNDf2bK73r89wQmCrFmhHrOCOFjNQmxarSRYtL8KCSCVUXzqIyB6mkjHfnucGAel1c2+QWrEj93G6rak+3MNNz78/LXZZpAdQOzEhsht3aNcsxEJKuigM601pyf9sZbstVwzm46qKu18UvBsACuj+ybuO6MZSNw2JLp2k+cusd4majGXvqZPdySW4cYo7oaYrN1jlzR2ht47LBbp9y7YFSg4CR/viRukFl/EtvmmGHu99s1ngME52727Pn5/TmWOi8ISqNpJbP8cSMe+PdrhuqDKg62WsEQpsOfvJi6Uu18uvievChLIbLNwYQqDqQH+4WD1ojZLieo0i0j42KXL1TE1QaTUBuVfdqvCkIxlMwBRVmUTHxesOLhfZULhbntbjcQuEOtyjXsBatmXINV3RhLm2SXdFqX+tZHyzgxrn1zAgcT1NZgiiYsm/E8wSFqjLXIwmJoyG6avKBIyw4ZPorvBeXnoq5hsuhiu/pNs1ujvHhYTa6HaIWM1W796NxMQI0wDc4oZA8sz1I/BsDb02bzA+VOIWRQPnXUXV/VMP7sro/G8XRSwG5hhuY5dz7wxmkDa6fzhhVjCEIjeldsxpQ5QmhRnyvqizAbZg1srLTzbYt5kZvqGFftKfqzWdkCMbf5bKqNvee+5t72ycGntLWXl7cBZGQWW7cSo9KlxucF1Q18mjFFD4LNYp2Hi7nRundn/mMrFhOQ17RO6a33TA0uqPHepBUmHQbbvZ4zcg1Mpq6gCw20qQbt9I8zblQIiEYlQmcwkOl7sQj2V/RuiHmxMJu+7tYLpDAgxju7sV26jn6s+gsG7u7NAoHeBeVvlf1dQaDMBNakHQJieVGrBvMbpN5gxvcD3DU9dzfipRmuw1MlvHTW8cNUGmfeYLB1ePgopxJ0xRoQW6F65mNkRAeK0uOapwbcpmm/4iPuhOq0J14SyLMTdO8RjY8PdHfJneoDwccXKmfg9xSKkbKt+WvJtFnOptnho/hneCGgYZ7cLOwr7J68Tqw234jiNtlVG9AYbJhasJSoxlDIu6MrqVixLlNE5it8Ak71eibt6evNSgmXZRvrM7Pixf14KnHvobo+TnkxjBx8DB++h9x17KbyM6EH7sj0/thwQvgiLVlo38dgc4epOEF8jY/5frq4qbpkAk4OaOZnC1FuW7f5bvf95/UEl4cR48sjN7YpJh4pIRhHl6YPI/JoakoHVwoOCk4MVo8ldDFe2NG3A73VMWiVyJr2HNvmB3yVnTR0d96kfqsuug6fufFNZUxwpPLCj+Eyy+DV1IYTWby/Bu6hhg3yPFhn2pTFR99/HfPyBXVSfCeAU5vWNRURSLvvuWPE7mVQ3hIxeiWwR2agE83HK5Z01xuLQa6cGLXvsMln9hqhXn4X2c8dNTEiKI6bIIV8pt5Btzxw5qoieAEXvze1hPMpM0VYQsoIqmo/URMrwt/Y8MHDvparFb1qSoHWxs+oaf9fLitwixuNWOLpWExwqtoed4tGhYQa6mSX1Bh4OBHinhNhuEZgcO9cULpNYufVRGJ9lR038iH+COYwvfb22h6Lv4YLjB056GoM8b96Q9A9CQQT5Ghcs2tDeF8mzVpYuH9VT7NpjfWsOZG5f1RS3X6G8jSrFzjMT9v7F7V36JsQCBvmxuHOIze0tFGWxFOhOr5wgdp1YBdlUeJ+DeK77CDAT9PfG4CZuTSv25sGiU94jaRCI86DHx2bKswgzvh52LMcOHQ5Oyb28XlSJBsMPhOsF1IFc+r4B03bvDtr1LNLtin9tYZWYsbyngiblOhu/iycPfnS5KDlS1zRuEofgyHv8ytS/vcoMFLECh60r9G1LfaoZsqzo3gTJv42HR9Lza/xIndCJ7q+nMEDTPuuqUFeH1b19W6WUOM4gBm0/1UnkF3ehFJBIWNLpunapBYtGfxem4C7a+yac69/Tc20lIR5PH0t97FI2mw/sJaRJa3W4RiC5Jl49uQFzvbpOTXawC8aJ48VNJKoXYMof5H8ajqXGYxwHyBc9fbHx/G12ipPhimUG9/sV8o4CqqChaxgYkwFt2LHsUFFHRKygmv4mxYopcEQo0M4AMvRbyivZjEqhVmb6whP1W4xI7VfjER8pOqeHz50GewW778CYZqG7N7plT/uT1r3IziIlIv5CWj9w2+hNvUh41IUnDbyZaGtb9d98ca0pqSUQpUNzg33vNxO+Pb+/kvIfnqFJ5ROJgVvm1XhTUPFCaSrZGK8/ai5lEms3gS2Xo9vxorWf5/sOlP542aGue+9w0GN8pZ/NpT0bFpBoe7e7ih2n07OaKRwzMJPtszxfoVFgvQyuYWVEkP4SE0W9vpD2DQdZM+8B+EY+S7OMxQakvVuqF04sQ6jPpAuXGgDveQdz0qhj5UAhHZyswtKHQ+veqDVZahIQXUPiPpN2OG9LU8aSAcOm2Y58P58ita0Wu1iaWy8d8HXZuowz088m3f3X2Udg/0tVvVdYHmsHnubrMb9zyB6a7/Ln/uupoy4QrdEuoaPbirRlLhNgvFnz8bQHiqkVJfreiNUM04N1bU4r6aTeP72ggsNPYhzllU9fEAKjSZvHxHJi8Vvfa1mATZEPph3AMUGouBusO9hRr20FE/qzgxgdT/rgqp2r9v+7S4uL5mpYQ3YDazs7RPP9/dE+L03XD7eOozeW7NC0waAzaHulBnwsDb20WMgjrXDapUqvm5Gsh/40ji/f8+/+0eTY2oqFuI7cgDtYUjGZZUIox+DyIi9QJ/eOIG3Lat37a9raV7mht13Bj+oMP9jbpxXl94M8J8cWLv530XVfus/VR7Amdh1WzytnHZviFgQ8VNbDGN1fqxF/9K1Um8ZoRubpPW8RFn8fG0YeLfurtWdL3ESm5cO9zwUa6z+HXeePGtLyvkVo0UBZxQFCoijVT0416ct2DCwlDdf5qDhtgIVLWdapgsb1QNho5/XDfexvpQrScKrF0wIxWksMHtbyB54kxWQYfZreHnE9NCWX4QPdKnKoqXZRRvsoXdedLtLYDothwsHpFgsIRBHirUDh8M2qe/mUts/1l8o3uqU3XRCqxq36GnUoqM6hUWlUsAaYZXhlAEu9PfSnMLcGfR5ZQ7hgftZjkMSYPh/AWYg1n3VIrgOXfU4KmJWPmfQjSr6sRlCcCze8aTJSI7AsCNNmsoMsEoAXFBPNLhnuV9CqC+GtjToREkHGlASCw5HNHMXHs7z+K8GxHi3/BBZeOQX2oVlOW1UhXPbW7AU9ncAr7CP/OuXLvbkIqH6SR3yRgU0MafP4ekYf0xfwPs/fVESa/P7Q9yuNxSEQ53X15kTOpfKScmEtsf8zbJyu+79exzVnGKoeLOqvi1lFE0uF7U7bnWD4aMDA1B+e1NGcrd6wYs5ngW/2/k29/7DwRYd4+2tuZwouIuoqnK0MJ4szqff6+UNZaEqBHcEyULqQDsc0LtGAb1XJzUjfE8khnKBYJI577zBjpcOrOs6l2c/P5mCscXi9zPb44ZTrOwwaMXyLF5gGe57xIXgk5+9Im3MB1/0ahLdKsdkhTJIoIGV9g0/Xuquhhz0M8TAaoORwK3PcKK3X07IGhrBRuyjcYKF40EMZpxY8a8VJ4A2aCJYH4a7fyN9hbrQX6bGNPmNLtNKhqw6HN/nRE9/l2UL54oNw75VfVC9oRiUYYq7kFsghHRETzep5IxeA26qp4kg05Kxw3kQI1H0nuf/xsmsaTur5r2zYGjC39D8VRjbXGvT9U+5sPLloYaEDd/T9TZvHPf/YYqWNVIBrUFhEqhCAnFuxaKAJgnhj8uHklluxsthQs0WPvnhgjikhfmnnVG3l+Bm9L79xaAHchPZfOj1sWtUwO7o5SAvZRtQmRvC16fQtAVOsX9bLB8aLfplBteY7LVteiFw5XcqD6xA3Gb6NiUX9fb8jU8T2WHydwSSmAqy3bHwqHI2hZjRTFGgE3fspOSxaBykVnlBTmRgsE+IfoynnX669tT0evIA8d4GUhm8ljpG3lyh909Z1mYLRxQV41CRxZqRoRuhZL05y99SQ58mACDyeuYWKEC0bmO49NHaLiY++wYxZVUr03d91Sg71NukCLStYrv3/fXh+oFhSMLsFDQIUDsFI/xzKyQ50tGvi+Q0EgkmH7IIBAENE5pkNxISV+o5Et4LeLoqUaDqPeexc8bAVSxssXx+hJfg/GtWDxO0MiWG2owKqqFvI+KU2BnXriCi9jtzJDheymrG/L5y/yX5FfJjVN1UHaan1Y7uszidW1Yg3Gt4h+0Ub/2VFQ4XgBQFwlPaW69qiqKuZk4LHkwISIuCI7UpwdwjHy6G8SmsBnBncSIYA3VKQjh6JsbazOnodw4lj2OQM8u4AiGOe8iGAIpVDShI8zEmUEjKRgd0s/ldgYPWAhy4UZqyLRxuxYZGWtIz9Sb6XZPXHv7+ItLcgx/T7Q1PDymbPH57Y0IgpEhyj1JCxQMzH3LwGdFTQkJvAUi6GlJqwd3tw5VGNyrpQPHSTAjKjLZ19MM5ouC/FwNRG7ax5eU8KhU5FawIoulRvZo57wgr8MzuPW1DqWTPBdc0sByoy5xrb8a3D6dcbjXbyUBqja3wt297uJC3/g4UYcLZ9L8TC1DtXEQH6aWnm9e1KKuGpdEbaEf0jr+FMMSyo2vfvDKBw8+LZLPk0lu2CV23R8zgvtGDQfpU5FW9AsWVXFrNicxRr8Q0X1t/EIP3bDoIXgo4SQtLCAyd6N3CjaD27NldZzrxnhxCqxd91QblacipyDy3wBK4S6eaveP6ZJNol5ghe7YF0ULPWajIN9hu7KqjndAdtTKQXZTsqEFOd+xkB2EKjj4Dk5/p8ptUuXuc+95A5qfJE58Wq2uz8CxHMrWr0aehhTJ4Pc5S0t1ivOs1/mjknk4lGSPVAPttMWt3vokNxqAyA3kE2fxwQ8cv7GnUn/xa45+VN+mPe8rntzdqhR3doEQGji7aSiJaUZCMpnsVMXqIDUsOUB0DZBFBxjBnEeX3igHboN2260zTR8cri5G6Nmk419Vtd89mRseXS3Xm4YBC/N0KxVn9ucBrGNd4FYCWW1zgwPYOqlC+RsBSHNR8Lmhi6ra4f6CN402VYQkWg3koXx8zz/Ttb+NRXSE16imM+7cV0zx6G7RUKXChKwYkj/zGTiEC0JAmiRHJnPrKQlNcKe/k1KUMYqfe3NizwaK5tzYl2drhUUcNbZuDm+0wPI8JmSqjCcPUz0uCSk8uMM3Pv6UXHlABNIgt7872cGIEZY1pPnrpsPtlRu8qeK6Qf7i8XhtqQ5P3E1pFLL3/XHakaL8fE92CjrDov41Ed5ObnErN28Z+Dx7aW4l4jYj4i8Oh7mKNOurwVMX6gjG9ctBwoalxIeRnw96zUUHR8+NRgJ8AuLEuGFGcUqMf/QE7q7gClxTPEgTCUF7upTwKxFH3Wfe39tFyU0OF90LT4XcqF6Y6bh/L087ixssgQEiRtJQJpnKJBSZ/2bRIepCVoCx9RpsRchjADonoHX86P3ADyMNx9HmjXPfnqq+VGra+/GXPM2oy3mazkhUOZfpdkqVEpbW56pML7xXD18zCdXgw6HkjORIfW1IwlwkBicVPWPa28FjoH4DqGqdmL1Pfn/X7LiW8fzzFldYr4xNwHbK3/nU9ieXe4U8iPdxgmWfrc6L3xQCWStGTM0eSiXBwmJqgIUpAqzTNVUvHO5ZQNGr4SMc6dmYv/19fjs/smqIbBXEFtW6T3xN8nJDBSOTIKuRYJspq9uDyqFzsUjyYA2N9ahO5rfJAf7GWMGGm1shvT4YdZTxU9U1JPr+8diqvux+VjVgMorrEtrboiqFMRvr9daNGso6QdWeSxmgGWJCkaArga2UvDae/+CXMmMUMg0Vjo0yBjrfxTceT3WNBV2bcgW3PW3k4FiYtUn0mJ0Ian6Za21tAC/CW+LcaO/fg9DAMWQAE9Icwue3IdPSYbN4pZpt1ut1d774Bnq4dIolx4eWvf/TA1MWn7KaFnpciuoss2RsEjRF5tDOug4JshYUBRG7hAep51/8iCASZvELdEY6z+IbLRxj5TaRVnSsRfxZ2hVchqLWjfo4Ud3mpciCAbIpbosBVvFSU04YQY4aN6nw5RPVUPcbP+vf8jTbyAr3WsdJR31PzjSxqdS7f0sNLIMFPhV9BY83ILY4mxrBrhoTYSN36JMNCMMIhfQ63DBmFusGpj++i19+wWOgrsYUM0Ls/Q8axR9S7t59HY5tbPTxGuwzTEbzHUcT9rIEyjlNTRN1K+K2t9JUCZVsKQZptqcavfkix+Lc+S4uKpNcxgmHe7xBfv5gCs+Ae2Yn/6XTt4lWxJkKrr67sBa1fUjxqKnQrGibEIRHx3DTw2Vdw5obvXuqMwytWSt6YKlo6qkW6sbDkJ0oS5E8Q+wrSI6S7/ECAkSG68AxOM9Ej80ZXGbIg6VqhUAVhociUvWARWRyM2Oj1Y33x8nx9zJWGua+KunlJ3VVCrf1imyATZWFYFiPK3TXdm8r68IWvant1e1DlRaeKTi9O/rGRTDq/yfuSnIcx5n1sQVQPoAXvoCZ6DVBGm9N2NDexm9foG7zFIyRsmRLSoCZhezOrh5Q1ZHBmL6hNWZEXLT6DmXUf37bU7FtAwLZqs2tkPNxzPBLFTxQ4XBOWExzOmydQp9dddsLztw3VrVVpMHDzqQk3tYaT1Xo+0Q16zvAqOPl77/fZwbtCoVimQz6mUc+H+f06YViKYZL1YoqyGEvMMjZjt/VqW9t4ZCt4f3FOiPtZ3FcG8IvoYM91a0khzfXpn/7ijjbvQqBKSYWr2AMldKPY3p3og4sqOpq9RZFd9L2nKfwYGgyzibGioDc2UFZRO1fU5BCG81PRvDcXwO8noUv/vM7ikCkYMjch4gEgerQRsqCO/2UYcnXPgSjB3GjDur0gy2TSCNU7DHHDVZY1+DiKbYTqCGt1Jtym07q9wf0jeHSXwJIW8Tf79HFCpnBIWLxmqjPJceAKdDTJZk0lJxRK+JJR4X72VDho/jZ+pgQMwcOyY27dbRufRc/KYTnjkax/nK++l/EgtnHAu9UGyZLW4pLJu0ybjjHT5VOf7IQcYo+0Ood3mDoa4uGAKpeNG78AZ5K2Bt433iCyCSq2v/8IjlQrOIsUUjlsZLONgq9cl47xDENQGwb5rch2OAGA5Xi2Gyb+qzdAy1xed54NPZQZpnJO83ioIe7GxXtVTqSbanPQgg4p6iChUtc18oxgCTA0LkkCO+YpBE6nfDCzNzttgaDVyN3hrf9AV/cGD7kMRg30Iq+vvtZb+isvFciUxT3pTOTj/GpivP9VBAtndJGuWQ1vzpDzRAMtBn3JpSAzR8mN17vhn8tZvETCxsCm2YoXpjx5v1uvrjnZaHBeOrEZwSOFkmWLqklmYGGEDjHsZ5wsP1T4NQIW3Oi4icfZG140nfq1Dg32JvmXnJjDMelTBv7iAEE1jEEDe6nyLgvJrGJm9H+Yu8rmfVSVTcYg8AYKmHmu7n3KWx+q2hn+ED3xdY9FUrwilb0PaOu4c4i7s0iXXHpqWpwmUiWvvi0h1RzlwyfTwa+0NktlXvjgH/5CBNwG0nwmLXhsbVv04mxhgewpumhqSrGTUsHjX9fEiOeRR9voiGiBEuSjJzLD6diFbVJgAwY/Eg5Z8PxLQJu5VPFPdWLR792d3Hc4D9ER/0w/hZvl2hYmP823v2icTcRJpMXU7IUP8jaslkGi1cIy9KGRApHMN2s237LqFDT9tp0QB31mfGvVU/1EMgImAyAzOQOurivwmF6KT686n4wLqxtNTdCqlzZjepXMI/UzoK9iOCRW9NrBt3Wom6Y2x8M430ACM8NFlX/9jVUrB1J0zgWbp01CJjg46K7qzMc/TeNYWLF8FKEodBWEG+2eq+6N1l9qscfeSi/WNfwecg96IxcdukoeAWzlc1tMk6vJDI8pTD5OWpfgTinMBGqEAtqLheu2908zcbpoOi2slFXQFXjHa4uqjpU7r5dfnYGhBiWtMWNqnGLuZEERrWgkhACY59V1DbUdJnAba3uR1bBnr9t1CtZwxfdN47HtjtcJWGOHcWQx2/Lm98jpeBNGWeUiI+KeeZQpHlSuBOWRiCpBBFJUBcT9uab9lAIEtmbH/34g63emTEAfSZiBhrjcM1qBHIDNiPXa9qjpM7pQLkRCVrIRu3C8JsBsrGwMKMSKkd2q/JsAJ5C6AurcJ0fP/IBBarelbsbzhulpRaI+hMO4yjB81/cA58ifh/hO8+IRydrMlHX8R9luJMcl6ZiLsRaCkYALChLaU8YHGYGxaMWUzihM+nf3DdYvO1+y6ClUNj7e0q4vWxQbqh5ojc2ln4OyWaMM8Ta1arpdJaGX3GO99TuMGlw2XxR7MUff6Cl8Dgyef/eldwAVPT1Z2d7Sxr10c58lg+Oi0M/M2c4PfoVocIkXnCdqIbQ8EfgA7e7o5J/IZfk6FmAVYJRUNETwngb7baXke4G4OeNfSn3SYkw0tPbokF2Gj5+uGa4ZA3ZUyewKfnSIp5DhUVw2wfBYOMx/vEwMGXgbo5NrfFU4oVZhPQgN4rd+/9tqhnkgMw3JgO9TURh4n2hX9CsCMkywqWp4mqBdUKVK+xNw83A0r/EAfMrS8Uo6VHnxpt2WxuMOs3i8E3RPUHZAsykrzs26eyjwc59QidjhYQ4p4/n2KYv8fU7kVgFEwICv1Hm4Gpgt+7L//T5nw8lCj09VGM7NbZVQyfnJljdNff7I/oGXpvuhb1/S6Bqv/2l8nJ3PbMam7gosqjL7OCnvpVJVyGVcCe9UYyWCkFv4L+ZwmXU6MfIYAnnRRWi2yr8TivtNrYXP4y/onxDqpnfNvhF0n+2Qkckrb2viAAAIABJREFUUhgZkfAJHCKG4HT/1sWtIyX60Fk0Aul3yj1j/SNFzxwcnfGNkh73Wc8bf8LfKCKTiG4rdQNEX3ZcmyIT/Fh7SinhrJcg/a2fyKGXNopBhWKAxXw+8QF3xr6y23VwDZoTXC8wKuDlBn3VswgbHqiKH+sy3sR/A7CfVDe6Q98D1tBLh/vflt6WzU2i9rZkp6EAkThL0xAaWajYroaD7Fw9WzjhMm0OS6CcAEnNLGGBh2roCvnxJSj1Y2vMyKPQDQlP9b8CbrtsAvD4Wt3Fik+J8RKJo89dNEiwM71ZmRgtF1EFCYY2tnHwc/WbRvVijAf2t+NnT4pIdN44Ttk0TeYNmf4O5anK6VKoZn5j+Y6METmr7y4Z7lJuLND6uIyzts6kaNjXSoWFw44lCG7iS+HO9MmB6IZcjHmK0AiLKbyONZumleanaCkg8PPnssHSOmrVMMKqRA9IVC28PlRv/nBBnBt4LZIq6JTQKrF6WI7Gb0pHyTGoGVkqCLiIic4IDxwNeX+Po/pvwJM5oP9GBE7sKsYAX189UQNYYicJwY8sLOPMwtCp8VJJDQaylQ1VKeGJD3s1W2mtQIKJgfnH8jj1hdLVZnmkYNwYq0bPOiOvGceHNpxYXhoWsTDAU8XrdQNIHWUjRX0KwVRno6XqP6F1hA5e7I5VyEWEI2kEDzX+P2xHhrgyfOc+0IABJbyQuXIJR3cfh78XPlMHvv0191B+nI58Fh9ASc+Bx8D664a3zHCCJSTVVRVXcB9nDkxyATdCR443U0zpc67mxjiLMvzKzXD1irDehfRdKMEpxaQv6DaKRxGZnIziLTixZRQn5GdfQNEpXX7OcXU86OZ6rrW4vXpR+yXXPnNfMk5kpe1JneDRS264Cr+2bS31Nn1nWYgE+DLnHs/iw/hQdbyneqeaNbv9URUfJw6QfQHe37Y9FWaF+CKbouH9vMOMM/QlNXd17N3XCclSisZk4AgrMSGM9nHl/z8Xnh6rRXmiymfuS4M7sHbbjGtvG8wI8cVLLHqYxeP6eYOWUyRYb7US5NqXvJ/ztAx0Y0q2k2KFI4wFPVOWoF9fxMO6FirIlS/kHgaMskUPVMAhSiU56s3I48U66o9muXGit4qghsDfv8TbWMR//Mr2VtzamcJUCdV/8G0giEjg/ZR16MO1uRPN1PD9ZQqf/84YAkgI+o+M5bHLDrpbjMVhfKXGL6By0iWW7Rf/IjeojI/pCrkBNTeu3U2pmcM5plq+gtXx/BvYUwiv5sbkallVMpYJndNn6rsqQpi+Uq4uGvAkuV4WVFAnO8yN/IRT7GHghfp7OBrxN0o8DpgbXQbIyJoVrmfELc8a4th+JuMlw0BetG6Q3S38YJEKS+2rFrcKSAjbJnEaublM5IB/kR0UcRoAx/iMdeNQkWKbY9Qf6hM75kYPd3HvV5Zxsihjt3bhvsrGUHrbqogTjE1eKF5MJaW8smmGW+poV+nrYAnnas3jHjxT2EmVSRxzY+jwnZJwTDHRTXa4ovpCNlo5XWFtuFaQ20JERBpdFTz9gpJnMLhbJiCrfIhSj4NcMnhruH7qnoSxp1EjX3jsCOWFwtwYuiH39/7+fPG5CeXC2mJGBL4DeCrQbisuWuO8seYW643LiZ/q7OCucF4qQbtbXk5xksieUPwBuhkpvDB/R5qlAmAOYAfVc1wgM7ICRjKkDXsvmqfq1N5jwNAwu3vOCeRw1/S4uA7BYKjQLUtHRpaciktGWNRT1Y+VeC5JUOQaPlcxvoLRIQiO3iNIsnwpCQGlZEyNUjUgNOPge89Dfh4G81A92s8bD7X7Az1cIJrFtHIQJ8Fhc4BlVIKXcMQFUbbEFz+rbqvkY2fO3mFCdA1LFSRUUwavfXtFI5TYSELwZ8mVJ5kMPF9VONrexXUUL4uq8dvlUhZVKx4qMmw3Zw1dUFk9kZmQ0GKKQyHu7Iy25a5Wa4b7er9wBidin61MX/YlKn0p6ZdO8yLTXD6M40Zm1ZfHY4ZP0yA3ToiowocKfiMoox5XLEPOdHw9+wqYwAuqBchtEJG8MAmFGlHL6FfZl6zdTgWt2+N0gb0UDOI9Hl67G75aPW0L4aWCPw2kwCrChsca+tmgpzqJHi4cvYacb/Dx82UU96ogeRbtKfXT8BwJH99iQjY/KB7pgj5VaJjh1LDBWUjh1+NS7TSDsJC+0/MeDBfQ3Pa4DenLJ1Zz+KmxYnTD0D/vr+FgdO3/IjdeD739BRw3PuGpvKjc+rNuRCqZvAUdz6RuP9Xh1Vlen8mNqTnctvNeqTxjLC4ATAhYIHrdrGdOjI5zo0wcmBsnnMTt5rCRh7JgDYcD5Ea6Xa/Rf7/4eTKckVmDaoZmRFz0akcvLBfE18QogfHhO5h7X1ixJjSwc0cjHs19PX0Js0a5Lt1KvaBlOq4RBziOw2rk8Je58WA4Fd/FARX9MTeYLMN1gxODp3C69vkZKjJPGjxuTIj5nfGXqcxkvi5DnD2VTwa+cufjJkom8r4Mf5IbzzEW3X2APZXoGj6a76lwNUK5UdaGt4LgSd+XIvRWeYxJEpl620XN9LfykaR6MCwksW6kVmy3lcjnOALU1OrYB0vc0kZ1lBfZPFTjT0JLBU/VIJqfzfdUxdMM14ZAbuq6NPa3H2s4GTCdWen2bH2YlKZvmDMLqVGZyxSvOHaW4eFvKk+4FJMpDzNjxeave17TMrowQJCwirtSxcdvwydgcYf78/XiJW773HigAiu+lAOc/vINLObSJ08z5ryiUk8ykx8P4LgSWZBLsBpHTjbnshTpjEPcBuFON0Gw9ZYTUE5KYzHPUi4oI+AtoL8eIDfy+B3Jrr1TdeIWmBELp4K6kRCkEFcJu4hWwv8Td23JbetIdNmoArQAf2gDoheAArkAlEn+y1Xkoob9blCUQ+fWMJ6ZOM79matOo1/n4Y4aXaMDVl5HDT+J58YPPAbn6HNgBRdP3TWqByLUOlByVQsQvlDwOHEgoNtd8Og0wzOlsziKRf0bTizowD5nWKn3w89Iw2KGWFa/G+aMejCVV/gUG5rsSa8iVCjNbY52zsjnWlpDIybbeNAjBbgd/HGsEAMKBn2juGxBAX424NsW3uGu98v5Gx8fL7mB4LYfeX/F3LE6k9kRNbAfdoW8taWho+EE8HGp5RvHvxkykmwKA5/2mBYQBUqlcahQO+rEP0NRX8wMRe4bl++pjL+Pe6q+jF/jG+02/WMzCWdxtuKGjf7YMo4BhQz3DK1SgrDJzN1E/rqfuPTFIypTgvUH3V7hSbJbRqLKLnUDHqqtij/nGaZxaHEF+Akk5X+Aw13l1gTcpgScpB9OsYU5mqaOLrc+Fco7uDCJYkXf81GjlRERu0rvtZTDeSa4JlQlQCGVCR78Ms3gKQWXGwN8o5yA0v6EfQmoSTxXjcdO1f4abpO4oawLuL2nfsQd7ufPgredu7wSYYNxbEYNKK+I9CguMzvlyOzk0EUbPZ87fjfr3RrlpUJYCC5weYNeAUwFLSPvQWhBpSUkhel5q1tucHIcWQX9/7GG37pSJ/zOVjfK1/tjUycytw+TupVNuh5eyxtJl958yQhUKKvboN9DbCxc4/mikZsfa9DDBa8HORUSrnWpk6L1iHx71gq5MSvRjEEK3xfnxioyk2GLB3Biy3g8bQgincs3rdJLp1W8U2TCG4qfmgV42I40VA65ppem+JsCDomQmLLE5DGeN/gPglRsjomLR9qehWWBHS4N4+Iy8I9uf2QwV8ELEyHqB0CFzjhMhaa+IpCd0jvLuGMvjayOWMazlIufUMb8jJF/QQrIjrHEG9pMjVRKmZYh9NkTT6COrmJQIVm2Sh5wL6I4XIzF9/e1dePDVO0TiPBAbnxBLMobuYRXuXouHKZU/wLYkQ1VL3xLoiIHO4N7Dl/rFZfPPldRSWMy3uFuMGgwEvVXfNmoQ7JKDi/Xss1bT67ibPd3bW7g5Hf3PrHAF+/fT38iyV34sCHA52Loqe6YpxH9wS+yC3U2E6agnj85hLNHDS00Qv2mQx8vb0c6LGmtzhwS2RrS18RP1RLm7bFaZxGL3h9iL9rh0rWJ7hthqxvd17Hsi6QFY9Ef3i+O1NG7N6YBsVfMrdcqNFVC2UXl1lbmdGpkx3HFLoAO4DJvBG2iqm6pqKWivcj2SG1hgdPf05S7149dV3WRzoiJhW25gQeOg/NGEY5f6cxuphDJzIxNugMqE00Y1FVF8QzovWZhUHOToBYnp/upbBM4P0rw+WcOALW0Phjw3yG5rzBBfmxN1dbjQm7MAlG/Ojdoh7symwa4ZuO4fZ6fW2Z8vgNRCdRThI6I2sc97jE2gUgzjLeNrTFZ1JzIYkZ9HmbLlaUyfq0K2hkfLup020jg0lYHDv7CdHlu43hd4DAuuYEAjmt5f3jfoAYXG44Ibu9vlA15h955ZOGDJ3HKivJGiVtcFHMfd2B06qle0FO/q+AOVRiDNLOIJqzVvVIcgNFqBhWOKUE4MBYSDOI2XT5v3EUqGqRHn9uDO/Sl/HCJLexQ3TBnDCUiZaNrRnAi6Mvutlf4sxCW2I86GzAt/2L4q0RWgq+BAJ05ETgkMDgkEIsmJZkykt+K4O8AhltvMHBgQMhh7h/MG3dVC4O/SmMcyEareyEx8cD3ePTOTFHAbOKpUQ4R0Dp0uAreM/Y5xvAfRJ+jMiqrKFbUIRi8NlMjG7mEUH4kPmtwTCbaHLa5gSujS/dUTG2i3CCMehj67vNdbnQPjxPpWwGq7p2ZRjY1VS925CSH1Twx21H8bBFncAjJh8D9KGNvS30un7xhBEwhWEKkJOkxhYmjspUNbHHpME6n2PvVufFtJgPw/3CgA0f/eBn/OoN5an8rWyrtpcrenyzLPcM26NpO5WzmcH4zlePJmhGbDXpMdneFxCBGgFaNyVbpu3Fj+9q+1eUGwVisbtxJFfVizMguN9BjoBk4Oscoe4g+W2/X1529SdcuC9vjkglXCJGJSfrxb7SeK+M6U5L3ilkyhEpwDZVM4H72m4IEBADqOG8sN4qHGnBc2lO53AD/jXHceqotHG+kh9VoFEl+gp16aGPbHcJuiR3QtFOGm7IJ4xdItva1kimcZ/BtmuDjhrVSEgD+RmGgtJi0gsy3OaxcOHBreK3OiC5x2WHgBv8qPFUfVgxRK4RC3tsZ/C2HKaqEpFicuFsfLQpVUvUkOGS3LITUyIDuHIkakHH0k5KB+xDop7S/5U8+BVyITAkTA7/meZ7Syit1goxcjDW8u2MTiL5sNbAHclNP6l9eZ0dWuEWsyh7iONOJ8vCuaIi8Dj1YZKyh3HxQr5BykY32+rvEqIpk48UIPlaDfva6CYmK3OGvacJaoTWjguBNXee64qqKMSP367GGd5nF14B1vB9Znqp72RbqM8UtrT8yHZmUZQa06aThdFVFEj3nHZPsN2psVTQqmIbBf6D/yMY+G76ts8Vw0PclQYObnsti0m1YNu4X91SaGwEX6miG8upMykAR58D7EPddxiW8oZT1skjHmhH8OxWyI+f/pDmVD7ZTTdFnhAgAdbL7uBWLsP+qmhd14pjgrWnLjXVepKW6X89tkvvGDXe4MUEV/+pJ9nm3L/TQKdoUsizYm1mDKfo090WLg8gl5CjIznxGTDUeBCtSP5Ury34hUgS3VpnlI2MS9LMNfFNKUi0mfquW7bFCzAjXDZa2uHzeMBMtPIsN8Bf9q3Ru4GBtVRH0pPsr3sQV4akXpm6fFmTgx4eNXkVugyJ2JCfyH5fo0W+v6IQkjS2CRWh9K0SZJGmR6+tFQ7op+GHScXwLxm1xamHXewyobS80VQvC24BQ8xC2WVGWPitBi3tAz+y+rjcDoANBTxk1DHPLYmCC2Il67DsDSGfnJkOzeVczbafclKG1fNLEqD4uWjxg4oAvmf8oHFfWDRJhYm7Tgr167MvXAQuze5i5pe0LRbWwHItIRnXSyH1jxCSiIb6R+tWuKhEyJwTh7UmVsI/fariliRYNDQc2uXVatmAwQ3lRSiy1VN+X9lSMUgDM47L9S46opdc/vtiWGuPQdSp6K/AQ1BJhedVDcdWow187g6sad7uR+rM4XnzRDaHjtyKpKol6NrEYD6s4tbWTvFXbb5eEayqF8Ly2VNdoKaAnKt/+nojg+ez1+NftrBwY8GlGo6W8ccViWMLuqKGuP9FkROIJRKEq4Wbv+sCdrBD62twYdp9/kFxw8YCfudGF3Jhn2FPNNv1dP298cBUHaQu8xA6FqZV+2ugcrcw0ucUo3P3iVfKop4oq8WL9reodHdHF3r9a+qwJoy9X3jpCA1JljWvI56Z2b/M3xAuQ6pAWUjXwrcK2almetzY3Lp/FV+twt2ZkgHAIY1wckenFMsplUfUKLhzlUHeYfDT8TUN1Xfw+Kv/W80f5MjWIZEiVo2ttLn0Sj4nO35ALGImBHyqqILC/pbJRn+4wvhM2vNZ/AxgDC9SNoYDMpGfwC14Hu1pja3SSGwccgcjyz2rh4IGFYkn9V+rbmdezkiIsx0YdrtUKINLYzlafqjSkLDcNeq64gMwTjH/pSS/Ven+hxF7C+/N88efWMMb+C6oBNFWdTuFUNUTNk38lMejyVu6WViEGEOECHtvnnwfA+Oe2NobWbgnrRdSd1K588xmDKwmdv7VaQFZQO0UdFj1bWzjWuTF8uHqHezdvmicoIuURq7iyAh5FCbDeWLFXzaMj6kwUSe7oLQNEdIqkRPYinqezozrd7SxgKaILpMxcgAy1QbopFhSZgs1+/Kt7rqCEy5eJt12bG9/mL46CSICK3obxT/7whbDx6B7uFK7XV1mKvOFcOuSzAUTMuC8cqHj+satKHm0rj1WGh2h4AU351nZKNuphVvicwAo+T1A5lvXmPJQv76mIhnmnKg51I47k3FRs2pCYOB6yFI3+iCHAox5d/UQLTA9MivLMJx3BG3EL5fXVoHfvRLspg7HVZisliw8+NNE/mxJXjO3JgmcrTSAzsjwX3qjf+fTnisc1eCqRUb/hzm0ErOGnLqYerCXStbYaakxdXtwUs0E9+72KJ8l/MWUj7EeHfEZSFWV1cCGCqDaSEpFNlO7P6cWqboVu89724Y/S1XJTBS9VnfC1WkUPd/0He6pvcdFCzkDA3ACB4lKUiMziR6WI+nOngmCHG5HMoeCliLpiOSEwq935HLSQHrQUvdgR3V5rzbXaElHWgw6NkGqzmBrlnRomODrBL9BRTVtizPNtgeQ4Xhte4qEsqUFa0RXFouWpYrG8zlcNdNYovQtGOfJ7FQBV77kaZoyVvVbOacIMNrSsNFyDFO+QkupTDELp44N3sN2gjnz4u4HGcJxAIEOgyZ01N+4CxL22p/pWWUPscOGTK2NfBG/IIXFlozT6qoe4BDOc8TwNFqy342v+Wf3cUM/ZuDKRNUMia9Lzkdtm72QXvhbgiVEYYe6bKCx1GuDRgroBVWPLja3DXaRurEiJvRpPpVQzeKjSsyJmBKh/TPF7FMWJiEEWYdmMPfPq3sfMvqwPlWzTndzOGWmdvTo9i3aOoklfGd+myka6Emm7KVvXyv9GfKL0ugFLwwl3uHNraH0tJ9ZSA1a4T5LS2+o4s49Jrv7B3li9vVWdc7Y8WozkRg0s6BMlxOOd6vCZs5+QNIIKpwaZtaPidBLQyIKUinZrO3DZhpeKs4PLBv5ua6nmaXWP1Z5Pc8W88XG/62okkJIe1Q1xG2VamfKYOhv9unLg+qrlOzd6CbylCqSYx5/vmexwBtYMPSdgIfFlskDaICg1knBk9ceMCq+T/QfHjUE73ImjsmBuUAnXJe7V3CbajMksPlNujAOSAgSaQHnSO22X9wrpjBLJ9F69SIkIhew0xVJXuhwGle/EQTzKRAGrKepu/0fc1eQ4jvTKYwvIrAPUwhdo9wESkg6QD5JnXcZn36Av85RkBEnJcpV6ozZmqn+AWYwTmSSD8dM5KWTOYauEHrcdz6QnoochF0WesaVy3Gr31LtxYRzKudomhi9SMD4Jva0Vjl6t8jCI979WYaO6bNpNCseKKXizmUgj2dKvwFXkqAAZxzdh3lDDBF0vVXI6XfvN88hauQM1RAY9/HactYTr4TxyAw6f4Z06/27cPbVXEg+6Dlejp8UqDoOq8N6Eltc9t3qM3qYNH4wckuj/VX4KXtqPNDFbvIIQrM5gKGjDeRAJfy83Y5KTwMbVgBA9BRIU2p8fradqRcN7qi2Ke5Z+g+7EInJoIO5vRQcZ3W67Pjf2vLqr53VbwDFnaEz7oAcx0JiN41465iGCnrhiCldXndJNchPI0vHtEu+DjhsjJu6MIWNUkFAaq6mihM98uJa6UR+PWMLvn2fn/cnuD0RcncUbiGsCS27+aEHs4/cbvUbhQ2UbjcHFlsacKgerBjZ9xfTH2fynSrfSV0Yfl0zizoTZDxcAyEiepxmfatX81upGAHEvz/MzBpQagd1fG8aXsjHCLhoLP4WohoCJXPsAp2+ICSzibpjHKYNOCek1EPGISKNhaBR/b4ApQaUgyM9GXGtXQTFbg6QUQK8yh+PPs96Pm/S41fQbAqirP+3ZPFybxSHgGKZ+4CJDdxvtTH4p53a9XtqcCXjoIT2gcwi9ELVNe6GWB+zxjGPLnBl7n6ItQp1qOBH50kd5qia/GxjA8SPr/IfCYaP4fSNSPsXz00DDr8f/lv+nIanWDDQFZFuGJF4uNt4o/FIJsMhgkx+Is6Z/PSb202T2UpOp+pQNktoCaeoCqVB3f6jj0keNfKhG/brtUcocwFnM24u1jBvttWqskXYcFxH+Xc7tqZS+9YTS7KP7WgpluxuDqcmoLQvCsr7fd8pLpIqkENrO7aumYBlz8+cVU9nILC2YLBm0njtYsel5jAGVmlgvRn2mlnlPz2HCEKi3wtht8gFIZXfjfjLXUI1GzNmwOVuMy1fKwIDg0LZaaziprd+sNdQsL2YpuuMRVBrlGzP6nSk8dxy4CRfi64adjl6NdoEMQp8UOJ8MvMVXji5Kh47l2x/9akgFn+Mi9rJOUD5FE2tcQ4Fwvzr17uYwHpw9+z5CIv2umsncwNY0Ty7DHQnhS3XQ2sUohQXvVbG7oeEZ42rdN1IoUzFrs59iEWdTqwVEq4hU8QbkPt1M4d9w1J2k/uim9tTgZvS/+l/XF6X+m9C4ZBLYtM59NaJIOSoNWLvrsGCMavZs2j0jPTe3ihrF3y1eVbZKIzd+7TCWm5AVFKm6ZuIKtr1Rc7scs7xU8lTdjVB17iweOOqC4QoRVw8kVI7e7Suu/b7vkTm74K0agsuO8XbMku2o/ph2IRXQVKVsiUENVsN5TLNrAbLeh2kmnxB4LZ8n//WmdSP2VACpztyLf7qa5usp++ZlFhfOiPHZVD3ex3zqYSc9YFgHW7px/Z4z22HqTjV8KtOcXvaujoYEbohXcJYKVGmp3hn7jNjnZmtz2+zHnoq29uvCcRafijxcuRxJxr/fV6r8BFAfDDCEjcWr1Q6yd+VtGlY+O4lNVOqOE0WcwaDTHoPApw75ruAjjOqpU6NJhXzz+BnohJgyvK0lWMXPTWHD2y3ejdO5hmZs2MlevOn3rVJcwX4OMWWvgkthsplSP1aMrrg+4KVqp2PhS27ArXVDkjPU7HlnqTQ5iF4xZtRJJ2/dbhBADMfQ/uqBnsq4hltHpFPmjfsnUns/VIeZss7ifdAi96SJUBx+3TP0HIYUy/fKgCol398dBUVIfq50qyipCfPbFShKK8xdULtmE7g2CoJ+9Vj34YuXKSPcjSnM4143bjf3wz29pzIUtyEjZXkJ2rwxxLsRcuM0RODVwCJ4tNnFsJ2fJbYjB5nK+/RjzaiM6EsdpTMl540DmGvGdNizPfhYbfy2yzLHTirU8kdrqOaw+2vLv3Xg+zl+uDyM51I2lvFvbJnWQoymraePfterByO/mKTTL89ddoBNpUQs/S8U4SXolvEuYQ+uTkcc9ALZdhaDvIIzmMn3nFalIvOfad5+4vgnXf89Cs3O0f1dIjIiHSWsO2PAJXd+/atuxpPKEnsqqspIhO7SKo06/cWWqdJSp0NQIjvaLnhG+meUI2mXInH0Ft8EckUIGPrl4Nhxk5nj8XDvNnHRPplraLDhh4qblsMQIi7MRtaDXz/sXAuzq8cFCatwgrdBL/PXJUMPYoRKoFb41W+klR20AJOTnbV8t1dpnPfbqFUh53P11CZXy/jZdUOhqjuyMBv3Xg5jWAedDIZSvaY5FG9wm0g/YFQIVAwutzSTPHAqGVlkiDZJasBW7G6AHZKDDmCEUGk2zi1ICJvLgW1TPIqHzuI+cIgFz/3UWVwv4yoPZZn+WlH+PfRrMXIg3m5vB4y5U6B6ejiWxgaknyHC7VnEP6Tqo18FR6Ts8NFHl8u0ERyknfr6NO1/0FI9MG48z/fgEbcwz6bJDTUciVSt0sKBUe1I/EpkUq0obe4d6TPgcUu21r4mrd/MJctyNA0mFHCqrKidrWRLzZ4IhyhaXrV4OBaycyjL5Zj1ocK88dzEb5w2iyO36UO8FKZlGBfzbj8OhiMzo7rfM62XuY/EW88adQCXKdXlyCUpOSkcknksCJfB2J3XdnhyE9xTRztd3etldFgzOqkq/+5eDdYNzOJ4rM6dxRH4AEC9lgZUDf36pVII3aJf+83Up72tByqm+FC9+IClH8dwRsTJrRD+4Kg8kc6G8FU/NaOhqjpLTMSmamWtxsXYPwbBDB+zMqMfrlC+nO8zcl/LMFtTNQq79ooIID0I6GZ2KYYJHlRDwQq2GCiiOo0ATh1sb3OgFbK/7YLs2PqpLs5/YzUF2VyJnvuI8V3NmIka3h4PzBun6zfuLuCQs2jyiFHKtAln3PPo+jYWWYEqr97ud9S5Qj8dTlEs7kPf0RsSf8HK7dXbzmLMlIODbquzRrVBT89CNDQvPe/DjmM5C7OZPDlfnJpYvxvzs8XVAAAgAElEQVR1TGnqm9wMTe5g12N4dT0CKQEa/TUrHWghZEzpr4KQjR2S+R9lzWswaKrGPBNjrumoZx4JitiSD+2j3/y+cqxwqnN7qgv04k/YvnTdl0rN9G4ohUdhdJVrbPkJybZ+yVkibj/cdSVK9A+KmDJF+pN6qgpJChNIZR8Vxj5vbp1rO+pP8Nfe1Ys1LCJF/AacSjLmPi+nZ5rpwCFZmB8yZ7VpfBj4VA1vuGzBMqGQQeXnUEw2Q+5t+gtMpNrVyDwkVIoUvSOzEc/1ELRijPzd7BD6rJvY8fsDaQNHux56GM+z74Y0cFg2CU71UTvdVSiijv6WRJGXpXiKWSdBrcHj8LTXQ4lYUUxgZSOBfU7JUrdjcRRIz+NM8nk4jkOfm/RU7SxuVsbPx3A/A6IuL/Wo5E/2UqYPH/aJt4nrPriChWhLivSP24HB4TbTharCigrbjOqIrc8bs9JElLkTtkyV2z6s/r4bxYXchvFP78b9eXbGQDv8C8zbvkTctPTyrVUFXYeBJ0Nsqfp1XjhsC1flG9s+Z+m8b6bKhiOSXYCs6JTZCnta4kSAhHTarFQc8dZxKiFB3HAO39YPWXA8b7bf0DzrU3m4F8/tXV6qm/Cp2jRu0e0DPYiHN8Sd4vFYMpN7/g88JI8sXKPkDFaFzSGhtqi+DNSWSKGWbs18DXxblIsq0/YuOeTHRyv0VPdXAcdJ+eJSOb7oai9N1W9O4wP1fa/drW01urVrQgnSVxbv8ra/TVs9eLYON3tGWbbk6XYkM2oHd3q6+gYI0kQbk8DonM2lxfrxJNqGQ0HDh3IN/0Gm2f2T0aRaOLIYhAwmzb8apW1PcTkUJLivBX4pxdSGsve1v8UKKw+jwt2lwYRVOlvE9HWzxjesXDsbUmjbJT5TsxT145+HQ+oXDUO5nFo3GkylRFydN6qsmyZs/65cMlEa3u8R2rjRYPir+9V3a1lZ+s5B0k2rNH9Jx/ClRpSQKTqR1dbVYPdcQTlH+a7zppUaj7VUD1lxmAGrVo5TvdvE9sfuxodMfyAdXPu4gH2FqCw+YBhMPDPoxGdr8BKjHN4RptZktkzr4U6pOtlGb/FkC+wc66dmc6mf5YqMWK/aED7PRwZAagaej11p0ym7P4/RkrJR5ama6JFnSv1r/4azg/oNgQCgqq6EtPB0lI9gHrf0O8Iva5dCKC9ddswRfKSSaY4t7U+f/0KL24a/h9kabgD1k+9Gy3wQ4d8I/0ixT3gTHFfij0Szna64rMyDTr4RwaZuS19A9Jgql5KmzlQTCti2D2bobdIYKUEGKiVXRehSE/ur/PYo/qxb3DkIOE7PprnL3WiDZ7saqhjQYfzKROQr9367kksECCRPRqZcw2fA7x1Vg7dqNQVT1j63aDmvXY6ZSw7cVvq3UPo61sq74SfwZrGxXIo/L2CVo4bi3fb5L3Aquxvdo41/ZdSrwYCswe3rd9T6QT2zH0yWur+CDYVcm2DTGY23mWtSpcf1Eq4QLhHbyb/++kOx+PMCHM6B3oYcrXNxqotnmkG70s4C8x8lNHsXgzDIELiFzmmznKz3bmApFvjC4SLB6RZZr2PMNMlbxyl9nVQFPmG8qHWuB4a9//BS2WP1sOnv/+Sdupyd92fz39PN25avcURPNZgP1X6QH8H0l9XGPtU2vftTCuI+scBDTBnbqYRY8LqxnIo+YNQt5S25sx4r5DfyqbhtUpvJs/UbcFKH5yfqxjTYtehJFtnINUylMXhUOBhUGgKUTB1+1MVCfbhxR8YOQqawDF97f2mbK63UBPx2dk0AhsD83UP1Z/tchW3TRdPez+yp7qCMQKPcNYJb4sRB6u0bhGpF2nFUBGkargjfjUUOhPVozdbpHdBW17fe0cVFHURYMuZgoypLjLqmnf9QNOSZ+hP3G7ML+O/Pf8E1/OTd+PqQWXwiMsIKfu23CRsgtOEwMPwZOpUYHpDeq109lUZvQ6nJyQnaPZWOZlS1W3W2LrGsZosuIj+uNSqeqOm7t+k/jhqbu/FsdQPmbfdTMdwLJMroqcRoBIWjD6Hhr4z0gkI+pFXCJXrbWDfSEQuqAl6IGHHnxFisqvD5/xN3NTly8zj02AKkPkAWPkEuIHzlAwiwW+s2pvsGQt1l2iIfSalctldKkOTLYmbThEzy8f0E4+VJ0YnGbJgZnfMqBJFbcEiRqsjr+FyhGZCFw1LUx72NjXfx/+2uaL79HL3SPb3JyIqWXsjyVzatP3XZwRrSmKNDOuOST4voyUJnxxZWPcGyNSF82NAtZhbLnK7fmVeObEhVghqacJqhnJH9SQLD3ft43CGPhZfxv6+hiuIlqX4JmorcwOddB/dvNWaUuBsgEpD6JCKK0KdqDsrdAfvZqCmTEhLS9SSlq182mweTDS2IOxTD3YhPNTGGuxdjAaR+KPHrUEP1H5Z0DYjDL2kJvsep6gcpOjV5bszQrSn9zoqS7AyxSAiilrkuhH6jcktUZ1Y07ImHvo2piXv/oJ1sjiIm+4uoalMPz0XAV4pMwZwJdPDt7u1PayEXDWY/B30xi4rClT1lakJ3JSqN3jSuRlt+F1lKQk/jkxX84qVQs0n/BYb7I7G9CX3j73//HckDRBke2RFM87E8cjR4pvLvTdF92+ATCDuMoYfZWuQtrYfnsoonW6qHvtmiIOlFI/OmZwhqWHJ7cDIg7ugsTJqqVKLszDL+UPrt642pc5JsX4aXKdbfIe2whQhbeUpumbHIs4aqsvwlOKiyMmAxMHq4RKZ+e3juagTvNk2n2Ybq/tjZAo/DVSs93zmB9Y5gZqry/SmcX8edDCDfHWBTPfiZ9ErX5JmgCAshhDsqNeMY3mplrp5GkVZRYarfv8pqoCrjhzvcS4EkyvhSfdVd3EUpAp7Gf82uwbkB/YDLvqpo3/HUoc03ScikKkv8/wq2ayhgOAcYFRIuwm9jxU5+i49AXaNkwIbZPppPkD/lvrGNnKm4iXMXdzTh7pkPrOE4jAGKD8kQIHoCBWvI0U8Xuni+88G3gl09I37sznrszA23EJboc9DbhphWGFn+DQQ959xs5J/cyCu/bdvrMf6+IZyRD6K37Z+q2RoXHuJU8WGCNchqNcIZPUZ3L1RD/LgT5ltvA6r3KWvpcNtVeZ70PGYcl+ZbKDq/hEx/+ImUgmV8/RY9zb/ww92MeZur258Hh+dw7Yt6XYqWmyCRvFEpt9dXcYENU9Awy2C9dVwjDKCJCslwki2jJ9izFbxoqwBQVTq06pOVf59yGB//NriL175R7TwoUfRxbAwWIbxUdQAXgRFcDQE6nag6/GrHCFMSfzBpGKmJdt1dwAI88YT8bLaMsyU8rw1OWAvCTVxOThBibnRwGpwxME2Thpp90TJugaq/mhxuSmG4hbaJk5Qpvmd7vkhqGPog/FzehlBuUwimKjOVZJaIV/Hk7m6u6d2eUbD31YtfzhivBBhR5d/GX6rBHPVq+mpmqt8f2UKKmscxpU2auL1rqMxPQap4y+nWCP2CQFVIC4gES63OyJBXjelruFLLGVB4QBPh4ap2jaLgSJUMfP68IiNjMga2iZVmNFPtRiP+KIgXZDY5MD2Mj2S0vaJ5GfENVgguQ4Cf5+/w5J0FqHbAigwk+UM1L6tomBY2rsDN6Rygwj2Df+qFP09rwf0PbbyxRBr9NqoKkzPmMFNFzrF8HJITov521MZh78J4+vtmEY988oJTsQaC+6gYi5H1gVy4cM+YtXP3RUjnBJHCI1X7NtamHN8cozW1Av5B+RvTJuKmr/qJsQ5Uf1+NPT0ENM3eZy+sDSv6TWyAprSLI1h0Bp8K3WnDGKu2/rYrvlzvKpHxOFZMtvR52n9nLB7axb9VMD64b0x8iCXJAGgKMR5aSbJyBhQF91BvbnoV/oWWcGaXkPhxxGCc0hO39mgcXWo5ZkyzHLeUAKGHqwZe+pNGUQx3/2TRtlGUNLLXYxvPNSSq4Z8fKsb+ofqgmSq+E4b3AbBCLVSrHe8uu7gJnKlFmGuWBoid+jiE7slCssQeYAEMkbCsx+ZGh8Atr321HDnTeCvD1cvbIGBkG6ptqnyqCRguQFw8jP+ONLDGJv3hQS2M0ZvogOivuIUeQTM82waDWQVLSmhUGvAthMXtKjzPN3ufLHfF7OGMotc1gxq6tnG5N03T8H1j4uUPbgr1RzjHh39D1/GGJtKomaj7N2GJd0ARsZ+q8nCD3HYRyKpBXit+u6qvqsLo81nnxicqG5CqEIJby0NfqkrF/fmGDPMf+OFyF//a+VTCi8ZQ9WKVHtVJ0hjeyvLnhC8ST17H/j+SYNEYRPxKF8AE+FYNC/e+sVRUKgGgsrNUeEsRwc3V7OG/Hylu21SJvNp9Y98ALYY7NGNgU9cXXJtol3jnB63laN0L29PSaZBfbPDCPWG+7nqVix6TMQFzVr/EHiIc8AoFUzoDbYtpGVwT+zakGDlj8ajKP+SadQHjA/YNwUbqiOuqYjweNvEoBCpR+THlUyMVjXDj7AOlcXH8VxK1ZYI8nwUbrrVTZWVAMioACGfSJXRbtA6Zm0fBPxu6IeWhTMP1G4wa7ruOJD7EN+WQbUOjLp3AIjwZH1JB3vqjcwJThUN8isnoXvlxIBdcw7CCYCIhnCsCMoideZVvFcZZ85Wi+1/OLWqIWLNpGt03QFMAa4SU392Hin09IwfPREks43OflCJebBliCMaSAKoKh8xQ+17EzsVQp+juPTNjR/MBLkQa2U64DBnWX1KLUixP4RsOVVN/3hh1Fyf3NjK2oMTK44cRTZafdHAEOjgcwzuXtuPzq5yVgg+q9KuhTFwSvmMYOD0ArKV/zOmOQkPqQO+EB928ohAF/82aw/FjcpQHvo2NkJE/tP2JeZtt4z2/sGV7IuHEt87DFxZUtGt4AgbhaeS66ZbbBltXrGLmSWIyaRap9eHuoFo+8mGyKitufRkEhVoLnbgQicIz1Vg+1WaRkSoY5y+Vb2eqaIhtDZRuUXR/p4mLd0VfNnob3l5e7Ti1JvBDON6n/USFt0NVMeznlcuBHr5XJQuiWxpCFeox+PYHDBeCcSLxxKPdj0TJdg+HNtybYMWrkco7tUuoWah8TXJKu7WiPv7vzDZseAzh0jsEJaCnQWBtfQ9PgkZyxiupf7/KMLc/g/eNKlEmI/WPD+wbsR+qoiUnRHv1MwlANzOq2WIHYIh32AJ7UCQYTxfEYM3Jvo07B6YsaGF9G0xLkCdRsHdIl0EX/+lZCkMw3PZtfFDfeIFG1GNHMh2cKMvY8pblZTe3jZ12iyjLYMKQjYrJxvcRsW2FMBx2Iu+ETNwWZJgqmHUJMCSkMKOR17eR1Wrk55D6OUAvrgPul1xiY4wHXyqCqTBRQQLbDFGXz0Ku4gm2LrRyOJVqKH3KiVADyeyJPNC5Cssd6XEReV8ptFmgIOZXxpJO3hbUOHrm57CcWLANndAUGuDQR143qHMQf4cvsBI6GtWH+IK1o3RbT3te5BSB3hgdIxUsVrkU9DBOC5ItR6SgJIUeCT8L+zh0Ia9eep+ybwx9G9bVvsKGPFT1Gwe5tHmNVXTil+e8vfLFvhb+nesR/ez9HjZKcksaqexMtSp3io99mikaTk0SDEdEC1J/CybyzOaXAu42u2l0/sbGKO5vF6+J1mbfiH0TZ68dOnE8UA+4Qd+kF3pMt8mEMvE6/pIZ4KSDM8FwNi4i4YZqxghf9XMFdEo+VtLIkRWr+8Z4DHczQbGViNsXgxGRJurEWcvb28OUM3x0iijb3dGBSR1lMS2SMarOnn1KXDjoG1nYzyCw2WmW592mdVA9iKX+PdFQNf6+UYcq2v4EN+x3cZgQi7WnGLvoMn5CS+heh0y3nEPNn6n0apWAq98KHH1dkRAQjrOXXrnP+Gjp8ldMC5cvlYBV3/DSq6afYzMGfsuxqUJZgapu/aPtm1wsVMrkkeeAH7Q/N7OQRR2OqqyhCfJauoMfh1hi52BnkRP0XKk5db6lV8B37/ZtSCFQj9wwcRGjPPRtTKwYF3HTByvGTTEYJ+E2DoICPNIjGxAbrDBefqViCngmKXlnDuDGgyqtgt8uCFG0nmzroSV3fvE6ylQdOSx1m8Yz2y8VUapwFx+r3yCkCqb2eBsuRk2cacRl5uJHAU3RkERsFeI5NwHRZHUFZF148JogAG1AsBb1RN7RlnFjCc/CtS04Lq1P+TRRLcxcxbpY9ragWgy+bxCGOxkibrQXDpuvYY6wvIOb9NcbOUAxiq1qYHN0h7zXJhNL/dGlEL/vY05601hOIPSiqld8tGgJlyu4lqA8URKVN8H1k90thnJGTIrWD7HbnGW3AUV/NLnhrQTW30apZPt2dAeXegAOmYOMUFSKGZ7DJn33ligg2x4CHVOtg7buWobfv8w6XrmfKEb7sxrhh0tX8Ym3P5aMk/A4NtaF3MANNUHVlo0ntD8l3ybrOrwbGIpZQjA5fiY4QOJFVS0TLmtSmB0CSIT+8GbBv57N12rNIm+i6Kath9RHzFTVSW/6aQyqogFGuAKiDxcFDUecNMb11xNuEj6bWypZJHb8KWctC0XcJ1y2cK6aySp7leatRyf6k83bQBOX6x8NVYIaDsWpNnodex//krt4BdVfr7B1mHKPhu4JaOrW4hdpDZ8rehuJD7LD6TrX6gWWrxpMTeAqaBnCe2pI1gMTa2UwROWsV436JgqVQ6Yqc+OQoWro29gmycKkHv6Bt+EVtWUTQ6+aJkkd9ZbveetlJGvsYhwT6nTFLXw1fkcyVvUBTIclUUYI3kaW1t3OUXbAfTLxkJER3MVJUDGWh0vJdibUTN6GV1zEWudFXcP9q9fOudlqsISESmwzIDrdAtekjkdwA1NV3yU8VfhJZO4bqxLYCCahqtTeXd/F71+1QLKjUEH65W9Q3h/IbQSMEE6lfEPPTJEo7Fs13MEDuQlRgUPFOUBkPWyOGrzzObXJS8TphD/6xR0cqLls4qTTKADPbQfPsvZp4yD05BukkX/Bw62qv6lt4yoaiOpdaAph81/9bbSQ8sHpqkGcQi8MqsZNRDlUZOuSmiCTU++KrLoMQxLhglgshJ/H87k/jvLklcO+DZEMjOaMqAfPF4i43lzHPWxedPfjduFfGDv+cttAgIBn9VLUYjhrPMzFAEDFZJH5rUde0fN3EVOX3JTCoCH0OvZ/1m+V8j+/JQ7ln3DU/ygRlzyRquLbY8bls1+MdrzlzBnPcbw34fRIzsMxUD419Jc+tfF9TgJfZbZN2jFuHDVo12OPhCKfJ61F0bXvyfV4CjHa1OPnz2icCrv4VMVNcvvj7YIBXPlQWTXy/4m7mtzGeSV4bAFkDuCF70HAPgAfrOE6BuIbELrLi9hV3U1FlpRvoQSTzGAQzCKcJrur60fdRMKBZipx38fFX3BpTLnLxKKpCxARMAvX89oXyoBqIr5CdT7lGYuLasKwMU3tOIz/+TXS2mKRTXoGhtuUhi+GmrW7an4XmMYrAK4Tztz8sJF+tWyKMCBO2Vyocpc32qzZ4Ds8ClGkxb1mCi3fm7IVo0E7qJCbvnFyuG2rCbRUUicCjiiI+2pI1Z9w1J8X9rc6/A2EQG50rfepZcbd0Slj50ACNWVSF3kYPHgel57cHC8iqVNRfdkOpmGpPpxkBOumRjRR7emWd1z+gNsNs/j/KN+3p+Oc3CZ9NxBNynVTcJmjgpU4sqfoZw5dUr0LFQyQgqezBTuI7ADDjI6q8UTylm6meIG+9LR4v2v/ZHD0m/BwTBNKw9eG8hRk/Dt9L/66umUTQFypDdjWt+UGNhoMVlwlGm75HjXxa3vEszqI6BjYgVSjmqv6V+O+3dq6ZJNqSqaqn+6WQi/VTmL+UzsUTOP07m5T8enzBqbx3tgQi1dhpN9uAHUdE/o9Sye9N1GILjmcXsR5gRQKHIJoLKoE0E/txwVUjXCo3WGwQiZOGu1U5BgqHo+CrIF/9oxfulSzM3Z/7UONRlqO1iAXle41wk07KqRc4sSUnLD7fkvYjPyempwJr8Z9WJizqYQpjyZ7dUfxeGuv41if1fMReFdxzzexp2rHMS27qi8Gvr8uz1N1f3Qnvopzt41/st1oZSG14dVMQSGq8MOpYjvHr4k1UgNF4jAs+Do+E9mZszHh5C2ZzSHmICaMyn8mPjUKECWobftN6mMSlArVUnp62zLv/aS9OCgjn436OajxiwJVCbURkoVk2dY17CMiykGXxFGQb/Fa3Af3jDvPo/uYuQpnCO/jSC9l2JScRVmpjVYU7c2Y8KViVHTGLwu7sNN4uO1AmrXF54c7jMSwEyZkKeEzUDsT1qNGf1Jvc9dXRXiDiUa5W4c7MD2OOR/6+Y/m3Fm49sPYV3kSOBdpa9veTx7w0s5ickCVsRRO98N9esGADuOObhjA9DQwXYVl7wSXae1shLojczgPJN/dczEYEjLKOlxrY8wbHVVVUVkdna6y/aqe+Fz4irfbCkWB6hDQ6sdi/PoXvobgjJBPFUgaCUaEhk5Afes98TbtuYlkc60X/lTOnub5w2A1It7S5S/Fd/u+ni1S2eya5FVJOnIIpcg5FPRS0lRNtuFoVur/FMM9WdskiQ9XpqHYZjxJwgaOY9DBDwhV2t32Bf1y1zyHGbtNHec2+Pgf5CCbc8LyIPJ6cEPhIrw6IlvpRo2qTzjAW3RUrThsM/5FF55m+3KxAfAMnApJQeRTNTeF9iA45o4CuBT6cd74mXEZ3r3hAttmhJUl102NvpmKEeHUwGw5g2/Ct07vXTv3EEdIIEyIXd8kHa67q9zEofr9y/PP9Bum/BsGdbxNJBXaViMFo9SmrXsq2LiR9bseRoheSJmiJra3W+qhXoXR2VisXFN1dOtWRQrnR8PQKbmveBClPdvu4ZgmPPQ0t4Bk4HX5C21Tg0Y+vpsqtLgtfZfAYZ8k4Bx3jjFFeBIRHp7Qhw9u6+dn8TbrPbILCx93wzRcoEbxY7j7mGwVXgquKU5/RQ8Ethd/VhuMQ3ldXVMlKFSgVH+RBtQlyK1URm/ymcIQPUtE5nC3epU4ZFFZWppD80kfO6O8+8rSr3RWCUUXTYvzmEjzLDZ9T8XKg9/QTvULRFy4jFxP1cRe1S5Mxg0RjA+3m3p/MgTWKTbUNy+97Wj77FddfktrpRGXme4VcMqLmgA0CkUdB5E3hj2j7VAn47paJevoekmGDndL4aOwNnyQ1tVxeM7hGhIZ6WpjSCCmp+BAkRQcVBX229tFYczrJRq8iG7/+0gGH8BLpFAKRNP74haOPoKIQHN05zvFwwB0a9BUmRYH4u4pQCMkG56u+3sqpI4FR5J0LA1zZ2A1eNCD2FkcAUUGxrbP73Xb+oXhDlcRzBp5kVLWTIdHmoHdj3o/g29rAyBun8mOhBB6nXxDBSCX07jFYUqH8/ybngokBdBG4MPq3T1NzrSfIBC6oGQnuZTiiNRdoqNqLtB3TW2/ozbitomIY93WWszCovRvhhI8MXdP0/TjqgLjsFBrBubn5exZ3GubYN4msBSI6rymLOtygKnIrm0CkzTkOlJsHSbpGiHgIvyw7gNLxHm8rHe3LtISklcSC/WisjU4GitFp/r6mCY3/r0s8f1c3V9jcEEvLlxDaM2ExTP0/S1ciPco6Y4RCmRQKAmSFocAATf/IQo5IwrZJGRxY8VUug1Thc+tU/ZVGzam+vOn//MN1zCtr+s/BUbOm8WVFa0JHBw4GFpGVpvIZ0xXlsLRFayomHAi0es1eltuNUyXnir3LtBxffnKVwPrPuca6W4qLF3LhJV4fXMgZVEbrz+ojS7t/YNjAvg6VMPy2Rhug+1hd9dMmacg9ITctOHCL4xqg0RTF7Wgyj0/RLxF3quQRZQEBbLuX4u7p3g/VXc9Leuk3WM0fnEX1cnapgs56p+aozUoLTqtWHSnI8HIfRBmFtd0sfiEdyEe8XHID28jKdcVv+T39KlqdFssAKtTaHSTRuFhaItb10tDmly4dz+XOsxTuIYmNWNP1UZo1c6oRiBxv5SUX7iHUQXrpIaOPGXBZaPcUw/tb1sEUNwnTRVv3tlqg/tYTyqccE/VOu29HdzEerLh9Vyc6smb6lOCtPTZ4I7JuOlWG2FHVha8PR5oIkniLbkO57sxCwQy438eMCy8R0sReGzZJVQyoVkbHYo+UfKqF1VdvaUIjlQ5ENIU5n2Thw1P8cN9ojo+xSzsw2jqCz/JBLv6lIZD/oVZEnnniI3gpZYPVxtIAoI2gBZtB3iE6okOdnktdL/1G6bvg8EIzr62vq8NRUZGACOvjsBzOaGnuqgI02sGhqRukjeleg78PPRwWAJsjLaLzYMjQg90dnkoaefezPKcJvm+7qxTFqsNOEW6fUZVvJZMdBzGu/Noz7jSFNp5XDte9BmckUtjqgAaGZzTiA8RCN2RhLCrD0/kpWcXHG7BcZpRPT/ieL9hlDCfyLjw2MkrRvUMCKdHfRndJTURDFEmepmmzcLAugmooXp3nz2LPxn3/tFwKrMoXqAiwg8JwfVUab+rymAloMVVyQbnjKH1U4+Ro0bONFeNeXv2w8WkGFV1qtcmK8N+SXZ8qJHNw+BV5QQ13TN+gq/h5aqGuEYaQUEkjH6yfeJNxTEj7HhJRrVNEDBEfHaWwTOWyDTq9jXqCjy/tQErY2f5bK1VdcCtQoXFNq5lqzaKQ9RF3HS2tul5fToM91PMFPqrShzaABiGIRxZ+iEKOYmkTKaOqComb7pDH+is6z4Gn7yTWpYfoHqtvRicOLreTqVfhK89G/Ppff/r/3TBsQiZO03b9PxBqBpUH+7lGmSJ7Govk3l7ZllztPtp0LEjqGveA0a3NE0wAnTeWsKKIZuI+0oZu3WfW4QX6GW2j0Jfce2pvq6v8zHchlNdmDA397iY/wLNRrp1OIxvD2UAdWHItFudabj5bmQRUwi0qUP6W+z81vMUrZmF9VSFy47jeeKSws+Ob5IAACAASURBVCZcG9ydI6mYN1QUe/p+g+bEvXpfPJGG0Hvu+B83xsC0pRHI0WwkwWnTZTijR+X1vgMEadIZPBaPPQ0TfW5h5llXhK9VyQi7lYGBgzeVjOJ/sN9AGsr8bMyIumw4WgYjPaGp1beoxYMhWbLiyMk5e7Y6CY5SGNuIYYkO5su9oUNWlkiFtYs6cRu1UIZwoaLvoiIKVDki7kviF8/DqVCL9orb7i/oQ76iYgp780ammaRgIYlrJU+fGnzWCSi36KviY1PHVDVQA/Fw6o2uiEihemlnAF9bN6nty/X0DGU6VNFMgT3VwNAT77dK2/StR1yzOCJCkeXRyOoimXTtp7LwR0OqzK5wwzahA9ML3EQ65HZ01BBloR84kgZq+bQgxNNcT+XhNmPDqwiUP/RhSJY7GhYmkseEl5lMaKEoxGh4IR1FsgS40zeSkX75raoMli66bTW7IxA9VR1Ajb4S2Y7VhhSHOOKezsOlauClmgF1DEMlcLXBxKwVVnp4i1K1MVyTw+mVboM4txpyEI/RTD03p3AX/IoYE+OGkOY5UfKKlcaBy2ouqW759/2/9C80sTZufOj/f2Pg9prLFHZXfsl4VI2QHnKKtPZUmltT6mfB0LNVxs5BEJ+q5BZaIzV2VHSeyHT41TAQlxfVVch/J/KpbC/+qaA6x7+brTV06XToI5KvE4UfwtpwJm3OZrXxEsBQ0Hkvr2/Di/N3KYSnKPjjRcWNBixdDh/GJE2VeVs8T66NzvPzUy2RsPxOttrwbs9hDyxM0DHJskmWrjEuZZcCF1LYx8TwSPvCDR4bJvFCokKHoTvpzOGqsHkD74bYTEpP9Tx1Fm/6jSuDm4ZPLjhonpDs0UjDMZKhzuIpamUMD+/x4hNHR9oQ9+lYed26UF6KSk76EqGiZ0UVCiEx3GPnUZSn8GJu76l68Uuj/qp326cCI8DPO+vCQxAVn3gz3Pk+jHsj3H7/3d0Wf2PMYvDSOql7NrHGnlee4SPqZVFA/Jyc7fBva2NuAJw/saj3z503vk//pUGx5t7WyiOpWRuhKp/q99ZM4Xv0u1NFM7NDmjzcW0IzFhm4iIr1s7IK79tckTqaOh8HUqyvZVdbp9+fR4ep8yE/jYcrfmGKGlIyILOfRQGFhZAv7ZsnRJjmQbKR1XnYwl9HpoY/VFO2wdqB+JUBDsrA7cMC2nGUokPG8VF8cRav5/k8XOmoyIsmMiJB4fqUY+lnoQLb8BS4U83zdu6pAg26Y2+coJmKLeJyNJXfVqwG4JDR+ez0tB3WBqCRX5UHzuNLjmOBjZyUoWx+uNrgcuAbboFq5PALR2iUhiRtxAajZ0+9jc7IgjxDvabidlpZKa421EgEaw1uNf5DR6XzhtTGv4V84zw/XIlfbCguGVWUCEDCBKXfEScL3W64vQZIbUb3HKPydoCmP+wNz1twYbF3w7jo5kNFxHD6D6/GSm34ieOEvfhTvaJn9f7w0c3cvq/FZ9rDqaSTClHtiKPZWKhCYIaovudwccmbwxxsIR7f9LfVKWCZ7FPVTERWfkpMWOjBf91TMSvocu68AVD9BT/cD1xVdKHS+jgi2HB5fvDzlFVfsNqQ6hiZm9xuqcc+PWS053u0ZPZ+3ydRDq42yvQLwLC7q5T7efWyv/8TdyVJjttK9NiMIHgALXSFXnOhA+CHCK7BCOoCHTDvYiBnSKLmD3d0l6uqvXGlEzm9oQ2eamGqWRSLcT7v6W/NiBdloXvXseEJpATewZH12gkMmrWPaD1lJsDdrop0PVUf3chHyjXcKLq8FYxtNkBc9hc/tsVT8bEpspgCHSkMpm00lws2otlFGDp14fXIFmfgrdKS2T2APuJJ/LGVw/Wbdd3cVkC292uG0mJFL6wWGmngTbOIWlglbNhZUZGxti57ooAEP3/PGkgkQeVBzkJUjybDnZlE3VMJTQ84ArPgC2vmq7JfiTTzQTyEFruyXNixKbdJCByCUuBwqEobg2/7l5rcUUR3UJdbjn7q6ucYXzjJ0MGeiu6uJZOKThnPcFnhVrCERKoV80fJAZAqNrUmw7+meyorMqnh6JnfRLBbfpyebqkoI9DyhPjJ3tVXJjOMs/zU6Wpt628Gv2RrB8uyXd00CCfyUUOlDa4oVGE42s0biKZi3h8I4lquWc8KbboNee7v4Bg7BX68uKI6K0HcxGRCnMjpzFKrzj9xVkwKSjeTn3gGbJIbHyWGzH96F2+9wxX/DVELi/QqqcVi11kB+xdW6SpErLlxhfVkNWKR2jHNlN8hbRCkzVaNpO4m2OEmFTtKHyQHodsubNyra9wmOiOLQj+HKBxlCYYscStVyf7RSzVKQLzTK9O5kuYWZ0UZyp+t0lXKU+4c+lSJ3hGTkT+rG/MmuxG9NrXl/akZioIUGJjApOSRVan6l4aN0Z9cL8ho4+RutI9UitgwNfbnDMU/M5vs2k9jph3hPH/6TlFuBCI3XdprRaNP8IWYfxAMQX+S/FFfnZEe1nEnWtC90eIW+zgpG2ci+uUOSu1gvZjA3t/eGnXueaqcf1T/mafw+cNw0PwXDCm2YW4shrx/AUw0c80Y5qlH8hdRVE7qOB04UNzTOGUJEhoh6aen1z4FJwgKfbIbEfUNoHh8WsdN3WDQSOPcEH9x1ERST1fsanvpqOjV6p+tcJ3qezoSsqhSg2QLgSrgyVLDofmr2xNnmwUmMlVucYaILCfwL1JD6sYKBhz5d+OeSla4lwstRthlTtUk+1dP4kTVZ8NqAql7w9eQOZw2hsJoeq5DzDC2OyqSwH5NjID+OBY0cIjln+XStNBRZzdMFFMoO1zpqfgGSwfAV9aFWjYwG0aZ/MyoYQhNKEQl7JmH44Y2utzdbgYEndi4b0tfhUM9GJm/0Vi7TSHq6jCOV9drtn7/StnocB8y0gKXcOk683WsYVHrc1NUznv38IREjbsOo7ozFKBI+kluVEaxDXG44r8h6E9dGYqD2TMvdwOkwuOGbkVGofsJw49XhpAOL1TyyWhX6M2PpRJSssKqXyTHLPY0R8DhttxTEX/jyg5FhnFepr962+Bt4Ymn8M64IyMHUwSp3JkAuLjEPd8fxJNBtdF5QzWPrNIOTxqfB0IeqnW63BFEarKnUlh0JBxuFO1UXKbr8PcS/BZvsCfX2dzA35OxOznjU3VW9qvbO/npETbJ3laeq00VC78tGuapYguOxrc/o8Bq1lT2oRrlUP6ClIVHIiyayXVn31cDOK9uPZMC6LXyTxV3yE3RODnwKXwSZv78VW/LyaFiClDEW97+pKNCCdYLD3+GN1Bd/x5L543gO0NAhRN5njj1PXEkSaXOTOZderQUEW9Fc2Ca6t3t/G0crgn8ijVsWDcQFX2TG4hle0qasZA2o4FUQLeeqfr5yRKLZHKr9gYIbdYi++1tUonu+Wr6E0Ew8aj+LjdS3VM1xuGSGyYR/zq5/pEPhzigPO2nHJfx3gs9WZTSXSVBrKhbCy70uwho1UqvvMLZ65V9kHUVu32+ptKeit0wG84bBOBZiL8hiKr+2l3jBe0EIogTpE1yYzRiq5VT1hlncOenO7tCY9xec2fs0Y9d4zaypf76qWIa5kWsghrqqC9HXo0cmdtkAW6dYtSfX/xMWPKH85X9TGcgCmKUxZKSDyfwJEKS1lJReAFqMPPF1FehRiqH8WNbbtMiDOWI4x/fYsfROvI+N6EZXd+J9Q/1umeeNU5KMCPAJwsZXpm57wJ32CvuBrgzy/i3zV82uH/12kQiPI354stBRD+pjA8VcUZz46FZFgGphMAMF3FhXI5eXYAc663yunBn1JhlxGCjkxtemWyo0meo273xD6Cf5Nx0ODS9bywVnqoTViyemkbr1L77WI1C8iPCjOtGEc27ssgCbdVJZnA6wbp7Ii/JIkYMr0yN/GhLtaX0bf2Wp4r8ULhuLC3rBjVVcOBYAKbQMWOc9oXaUT0b/zzlC0yALAs2GoVuxFGdWevWi1iC21tUJYlIIm74JAZ+hGJjKc85/SI5YN4IbKN8bMyJPfIvxIyY/hbCoWT98SkQmtfobK54XcLZpowaKrCa2fXWqDpc0oGm3NA11cYiknP6RQXX3GAhPUyNlhh1rBuwNgTMSDQkZc6Ney3VeKtgbx1hvfe3hGS+gJ+Uv4ELw/ODAp6YWWZIlwYnktjJYd62X+XG9N/sqRZSwzUszKH2huuf79DJnMlj2RhRMK+rPP1ExOIMNfwGeXtTyWeGP4uIYRImk1DDlamffpIXcollN5T2nNhFBXGRMhCVbNbR2e9VM4EO1D3d2HvNjevMwIWIqB/5yvFyD56Aeykayc1JPKlYwvyTlgpm8SKJdIH7Rus91UEsxo1tr9na9tUr1D8C33poapEP23cEojrVLosncZHz0kWd9yvGzOJsNIsXbRfepIvy8DbP2/az3DBMs3oz8n/PDVjhCoGD+PucHGTp93zwQ4LGyRw4nCqtQlflqzUVJsbJoyLY+c68MV97wAr9tT6/isjtPM8/6W8NSP1yA1Jvwm06io2WWj4IdeZlQpMTNapeJw9hCBBKhBODRgzZkdwn+qn/K6Nw6zmcPZnmeftRKP6IrP3lfwg1XJp6DECZovlviPFS7Q3r5ml8tjV0IGDPXrCqfiRLEUezHwVGq4XbyQ6DLJxFPEEXVawpsn1InrkTjsR38fWGMdCipyJMNK3UO9GZFAr/vo24BshIivQinlA3uJNwNlTmxU+P/QRmkb0V6xOLZzOG4duveiqLGcHcaMttwngg2tDoTHbPzcPv3MRxAOyuGDQFwYNEJqdjhuTGLooqWWNevcKisgibAG0qWzj/ICYWo4650ZbbRG4oR5Vv05mjf2ohXuPa2NrB6rR50b9l0VWv6gnWH3l3m65ThzFwNxNgSuk3ZYOCoaL2EIpmeKojzn8HFkUaUIQndm/9cqbRRf2KevSzswY/Veo4yrcmd2f8s1jcpG68eBPnhSHv0X+QGn8U3mbuTW1zA/3+juAUay4cbwTDG+G8ruNkYAFDWzkwJMyjmZ5iRUiHiiZxbanw5ZqTgaf/Ij2IvI+5UT1TbfBUtDg82J36IOS/t/LDiccG5YZDW4HpelOld3GGHPoH+9vqBCgyYXbkYKfXH+BGZvS0vlA8Wt/+jpwciyBx33yobNnwRBBnYRGwq+7sRVyaqyslqse2J4TErSxgLXVj29IvZo4/s4h+0q3p2BiHu6Cs4UGdm4au2o+8GRYWJaaaIWptaumngGi6xOZ/BNShfXCJTXTlmMwUKJa8iXnJX9YOQYwQmqp6qhp5mh144hiiium9XcqdwnDVSQ7M3NfO30BG1in4/NN3+U+gWDxkX8pVtrLkndR8dN5+sLGqCMq8NGx43zhanAJ5cHS7z1Up8BFI5WWFMlAceJ0CPdVK9mXe5yCsgx9CCc+K3VTwJQZuDUTcyJ9AGOSdWh8dOogtbuj7m7HlTelL1AhjFFaxClqWY3MtBetpfVHqX7wNBTW/A3/As20seTH4/BlEwg0+dMHFroQDJr9AeeHDRB7VOT1W/KoEBUNCkVkfg0dkR5JUHox3VSl9eXP6A0XcaEw29m1aiDOec2NhEA+yOOo+VwsJdsB1XRlwTZj/wsUcjIFMBSAc0Q/5Q04KCEj+TihpEui24VYo5BiFsJcggsSdOTfmmrvPjBryIf3uoVJn0kNzrWi8NjEtFl2tI/+QORKxoycqmrQYOgkMN8W5o41lFo/B5Ucqh2NdS6rkojEw3S8HIZSft4MHC94FqeKQIOHh+EGwZa7kiSELJE+1fbM/RBlwxiiAMG17PdyD0GkGbKo4PzAQsX6sUN0NMwc/hhKjnAz5xYo5N8rX2OmuoMZT2quQE2JFWao1v2KYFKvP/+WenHIDBoN+6uFefhgDs2RXuXR6YmOB9Cl45A/dmkJQd/HGubGQ7gsX8oLiof/5pXJEJKBpswXpg1+Wj65EaY0+xvJY5XeqW4cQoIDnT/ya3ynID4iIL91UgDKeEweECwJ1VQEBAvn707SbIokVFVAc+kadihS7P/k1IbRtqrxQWu5wBVDF1ybSKMZnKPLaKkpmRPyKXjMJS4S/zsEo/++Xap6zojxVg4NkgLqx5oDlgp8/CyUrpphreHAT/ehXKuT66zZBkuoZkl/TZC0wafrAufz9cp62SXIjiHTbsXXdsHQzCEeH8eAfMmQDfyn4NxwRMRwuf3ft1gi/SzhizLlR/oTcbeXSsUL1KI9TTo+An8NyLidJ/liCU+IBibHW6XJ/GiSqk8oiTYJbMJv2D/pbuG6EC1lhLoe2dUM8H5jCj9lBjxIkyTDIzx09ryFJMFfyh1Ai5DiH8leQGzkPcj8FWRHK7JcTo3wSS3sLP/MyfMB/vKfE4L4q2Gcq7HI6VEt9ri9QySorvNVQAWCEHiri0ixte6oj1w5c41IhHxAgTf0SN02RHq9BWiqs6x7+xse1RKkUi6FkRsmP/EituZrkH35psHKSTLFU7jKDlDIeAvpHlw+QG6uEY4U/OKnfyRAWvJAt+8xyxXL7eCscfytRQ8mNQ/PcAKdg9Rmg5SF2VQgioRoOMwhBriI/ZzCUY8bktMiDd3nAQqkaISdCfqHyQ5WzIJd1D7/LKM6P0YoBgA8YA/xm0GCsDztettQi86YJFe21oM+vlw9KDe5v8Z2qmqpGs7gYm/FyhMDRVDtwqJCNCf0F98CwZsytE2RMTpL886eqUT4bSm7klFnzq5ULS06PCA+XD7ggCRgAyg1elMCfVT6H2O2KLExWv3uarhwHckD+vtLcbjOrLSnp71hNf432VEf4w+fYQST1Ij1M3QVD0VGGYEzwMwpb+fccfOnhu+WZWgco5iUzMAgBPjrIjVxEXHnT4AtMB5g5wioPF0Yq/MOdVdhZmsyTQn1oHrwygPiXvWtJchtHoqoblH2D0qIu0H2E7rnB9AFq0VfwRIx2WgwjjCv0PRwh1soLZQQwB3C4fZchMt/LBClKVeUZU+oJwrLE4gei8uHlDyDwXBH544WcCMDQro0ea73LSFTLrDHQecjhiUNOxnoMTCzVTpj4bAGOVHocKz+UFQMXBgQG4/GhvlWroeGHvldQdirxfw7i3Vm40feuuTzgyBlKKmO3bvZnF+Z4jmWEJtOBmhWppuHLtzlQvrTLCxzaCT+X96nE1lFOXPK9+ENnNnvYB3OxFJ2IAm2IqBsU5rZy/SiVE/qy2E91V65Ka7D3lRtHi7Ay1JMJ26yF4XJo4Mk9jfrU3ZrmTNpBcM++kK9NlfRso0xU9n9QOVFLfbHucJt9B0vEaj/sFbhhOXWb+pOjcZUgH5Al+QAfq+zIDMOnqCUBDmSO2nItlSn6yDPQzKW6WsqLAZ0hTNwNDnDOxhQvdVaJQzE69PCnMuxHD3rk84ner81qgM+gSjNi1Kz6AMgXo4S92TLvXy0Or82kXUNLFvapFJHEHic5YrQ6usk5NY9hsdsxHjGIPK1FVIotUp6xvx9Icewzy/DXQJysFmOQ90739Xa8kCi5HMCW9r1HgPgSJPBTOT/M1+fJU5s23uSb0+TLNywaeGiXFkc2XRbmBjwqe2rc1xlHT8fORW3iJxC7ducxjlVBHzF5TK9WQt8GW6K4DPwYFMHRRF9f1WRWLMCKAnqYUBiX9zQl2cxJ745XnvG0vsYcxpwP4+toGdlv0Wloy8HbnwckkAejoROFnWqqBbmBrCENh4JxRHeHtXhN8JoW0w2dB9GoEhzR9wpC1lG9veHUQ/Am/YENJdiCXdigt+96y9VXhoFlD3rOF3oLJ6uRc0KlA0aR+vStXNMUzBgqd27IZH25xWJxsQVYJFbvVXpUmVtPOcdLH036/scRkJQjkKn7MqLHrBfX/FtlRw1vd72+VQey2spsB40mhlcGTIZBFUxWB9ggKYzXs9NHAxQ95eKQrEaJqeryNZg14EMQrlX1hcEGRqfLFbgh/sAZnscMfpiQjQ6GgcJE7WTYlMFQqMAVo1wFP+gmk3Ktp6/6qRQIuKjcqatCcfnu3rmjVIFxByYHuF7GjB5eMJyx+TCxWabAF6ajAotRW2wBDDZknBlZyqcS9o4X8elfjuUfZj3QB2W6SiExVlgWxU6pcJhqq4joHBkZixYMf/Y6hrFiUq+qWCglzHlRB6bBwyijb6QKVZzD0eg44MC0fI+pnvuXnrONoQ/PDkXMvXPa07QwN5QeWPfdyr9c0keIHZrJJ8DXd4NmR6j0vfIh26m7KuAq+iMpUdsehW971FIAnr53Cw8qldyWohqr94wWcl2+a/j/nDHwg9F7fnERwUPovow5LZI6/deIxQWjqmxCXjhWhc1/Z1w5YmdFwDCBuEkQ2P/Mq6GU6gGNqTKYUdjVWbwp2mRpARC5UQrW3uuJ2wEs+VQZA9/Lw0iVP7MsVcZ/GiL5BbqwJmdG6caLWS+Zw0XEEbO4eUxOCcOQHEP63LULuhAyaLAd5L1zZK3jwH6z6SYqnGJgKDB49fljyWAIfTKtog+1VaiuCt1g+mCMXYwkfx7qyzOTMTzFsmFwnM1q6M+XydPJi3LDhyooPTQGLGWMSLi+cwfa3ZaUb4pxAg1P1VSQogROoAUOEzfVbBlo5RwMUj0GUIwxFrt7yNgzZX9g14nprH9bNwo9aFvDLPtMLz7BSDcJ/xbr+8PY+Go3akeHtGDU6UfIi4nsj2WmzO4kENOtvpwigz0ZGACn3nBx4/4R1j4bXWBgJtwxXEqwhbsP2RPDflImPcUWl0tX40YHbqRugEOn5S2pRWQEz/+o5BmYJgj1ferH6qt+fOpxTk+zn0fhZTjIRdMtkZyk8gIsPB1w4Eaa0dDpOj4Vk2MY52aW/Acg8Ep4VP4ffdNabI83o0t25nx0D43sgOfGEL8N9Gn02zQYgeMIQ2uVaZKlWq7vz7khmK1KJ3Uo0pqQ9MPRCPRz/bJP4eFYq+3D6GRyg4G9abZwv6h6zIHzkN8DPGBTOPeROxGqE2xFUrmKT5XCr4oZkvSe1KAVzkJpGsyf2EwTZfZjkerjfZZXYzAQ0wAiz7q02g1QmKnpHQzv2ZCpplrSboiFOzYHqKpO5YcpLDFYAg7VZH4siJPQwFM5JdQcuRJgfYuF6o03n7xB9+6wwReD2S8Bh7NjmoOB41zCnUJ/+IkNX85upOgHtOwh2GGkLRgzbRyxuawSSAPqFKa54I8U+suGbOJ/AK0AWFrMYk0pICQcCDHmyRxG6RzAg1Q/UqE5N+Anw+7nMP5AJiP2aZqTjBYkvQ43+M+QoAlJlipJXBI+uZzQHZKMKgVnqbkpsDcJ/1WWRjMeSACBSFJJEANuEherRV7pA/Tj3Qz0ixsgj3lyyc3ZNY1uyiHNNNnFfCq6VBGSw3rwmcCEYcKFDdvAsGgRzR+NVTcZR0KlGZaJ8rUDIIN4vOWkcAzcPEljtAwwOesDnI90+uzBKM3MSS18okmuyQ2OOrREsjQFwuwKnvKg9A0Pkyyok/jRCSmAY1RS7rClyIp1BAdQmH0SJ6RxTRq/AXDEUyf/hSvd6D2oBdiMqeVYkBs0Hql5wMaLJjMHODpLGJhgYV+gnhKRUyKZ6mI8aSCQaHQBaDgSwQE6QRIYJEO9My4WPmsiUIIgkinKwscX03egYrbMhtCMx+4szg1ha7A1J/DiqqUApdCQtEiBSvaMc23mFrpA+9bFdqSpQ42RzaTh5ChQV4WkLA6SkEsgIrWXGSHzwwtY6oeoNt8Q4IBn6FaQ+da6EDfS5PEahwTyhsBxs55hBAqYikGH1xffxPFiXnNgJ3AisXqzDQVAxycx65wjUlqVqSgBSlihwvmcUmwW0IRxEwOmcwES8kBJYjriWUAWtRsJqlLnrEJ3uSAASWlqUpJLFM2Jrb44SEo4oChEsEHTEVOYpeVcx/q5qDDqpUGvZgzWCLrM/ou7yh4J8WkIeIBgVHHjR2eNX9xNO2CvYzcmQ3MdDqGJwJqyMCuWTEmpa3SVngNZ2mELKhuyVetTXNqeu0ZNwUhJobeCfuGDJ0weWzrzG0rrJpAbmLEReQXTYHAs6HbTfei4MJCER3MlbqT4SJ4qgUlItCnw/BipsumG0unwlBTXkaBbItRbCV2MHRYLswH5xAVmyjY7YkEmQWVyi95yneasM+ww0D7cu8LIKdnwKPeuG8eBYFRUMRWxSJeuzg2Jvo5WVVHdCBGScP8MK9y/hPRgCTFozniUoPX4FV2oss6PAw51GDCLnZoeSEqCp8448/GSYOpSYNs588Czzi1Qo8hoazC1o9/4vBVfPt5oup88PkdPlLUcjrwCd5BMid/gEu98XZfE0YyJlwpr66je3EQYfaIRROsgeYyJ4fIVshaDu6XRWO55u6XDjmSuAyhuqyziR804Nle1G6MeD8PClFXCLBikCQDDAyBUXh10F+cF5zJtTBULoxoHLjXfFyRwVQcbFUa9c8UHO0VWAaYUqlG6xp1rTJW5Dm6T7FeNHki+NjcalZXiWYJIYY0H/HiUIo1Maf0ow+YBEZIB5kIAizrUUC7sDyZitEMYEQtjA6a4zUJ9tFiOPt0GbtEuJbKaZpEVizuVs7rqej5V+y7BElh06fwndBE1hh/iKVAJ/dP5rxTnXuqaBERQAwLqwn+wjD/UIq90PB2RwMVYQtPCm2jRTC2RoG/T2SBsSW6k0+E8Yy+PLJ6ePZPOSVGDnMb6vCKN6k9uoxqKQXQSlaaWuiQCQe+YXGpqE0m+A5eAEKnhadNY0kUpLcgNuRSfOzwRIY78QFj3jj1maQZbmfhunjaWpjfYkXBP25WapMaRa9QpjJTQxrEPVXx+ITcvI1rS+TAllcbMvR27cQ6fkfTnenFHx5LLOTmWMq8CRot4R91ykcRprFM5A6Hnwt33aHUilV9gJ93JuKlb4Mb8+MNGBPIqxF4aCnHaxTWP9un50o202qyaPAE4PIKm/0hSGB2n9Smlb4obci5BMC/Wt0DDxOh/zAAAAmdJREFURnwR2AYPmVw5GQPYMHD00EWaWoPU1EbenjfatxRvzMYejTq/wJn0FmBGQklngR+P4BgDMH+3cor/mPSjYPc1kFzVbrwyA3/uiMy3epkYne+kbZriJadWpUlNX2L3a1vOrXDjpduW220e303Yvw43bgOCF+4pXUTuO+7+xrjx1ytyebf8/3DjNkS9IENnuCGv1JvplTqlDVBH40Q8T+35aB9H0o0y1F17loyvZ29r28PazZ5zev3knNNzkUwcdQ9346rYacmUbtPZmrxHZtQt3A6Umcrx579tNp9HZf95tpzZ/aqD+9jY7/f+aUe4A8dn/+kFeLGSvV+598onNe79JC2f8c6rYmNv97if+V77dVEFNllve+f87r3f8OfpjYx3TYX308+/bTY/reVGyi8DGr+s5UbKr79vNn9fy3XKr9Mdvz1tNr+v5UbK04DG9unx8XH79PSk77bZbD+OjsbBp9hqLmyv2zbnNJXg1XzgTNvTfsQ5vteq2T61e/2c+HP8BdNqRt/kd3CyN77y8eS6yVnz185VNb7VRjiP2+12QOP99v1DfT1sh836pn9he/vwUE/gH9sH7I9L3rcXNtfhJK/Fz9TXQ3z4mXjZx7C3Ocf+fO9fGF/ir6b26cWTaprK4uqH5ou3cTP8YZM94x2TI+3rpKpWWiYWvReV7bvN5t1abqa8AY27BW7n7t39pe+7P3NT9+3Z9/ff9yvuePRu9h7u7evv3iSKu7vxrd+9u3Rzd5u1rGUta1nLWtaylrWsZS1rWcta1rKWtaxlLWtZy1rWspb/bBSMglEwCkbBKBjOAADUd4nSeo14OgAAAABJRU5ErkJggg=="></image></g></g><g class="png" transform="matrix(0.6679254770278931,0,0,0.6679254770278931,251.31182861328125,99.99404907226562)" opacity="0.7786332375769343" style="display: block;"><image width="525px" height="121px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAg0AAAB5BAMAAABIGg9ZAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAPUExURW6O4VuS5ESX6EdwTMLW9Q2YIQAAAAAFdFJOUzozKwBmW62ctQAADJNJREFUeNrtXVmS5LoNFJ4vAHIuIFIXKOtdwB++/5ksMhMgtXR3TYRjSpStXmp6/ioDSyKx1PR3/6w5TQ9+ZPtSmcKkIS+7N/6a+r9m0cdioCIbCFp+pqBBVJYvcFgfDcGk1R4KGEHKoxovcVif6w9af7Q+fNmwCCFe4PDPxxpD+bU5hFRzAATFHMrLXyccnmkNUk1BaBNa4XAQQmgW4TjMDw6QE9/8hoUoYdAQymtY9ji8HhsZangsCEwlRBYzgHNs3wWKsMNhfaxTKGODOQSj5GYRROKvHoeneoVqDQ6VP2lzCakwbCCUCLE0HNZHWgNjJPiTTuoBssSFagwFi/Cr4TA/NTZINYkJrCHAK4ozKJCoKaOGSuDw2HKiAEE+LYBCQKIC0uYeh9cjIVDQBvOIAAKlCAv4rn4RxHB4JpGmX8AWgkyk00yZQhBoEAWHxyZNZMvCG7ZKm4mi8qfQnKI8/wAOrwdCUNiT4F/BY2RBAnGSAcLgAA6PzBZIFYIgicoKWAQ9WEN1jILD42JDlRuQMKVUFB4awKQ1HJH4VXB4XHjQyUtLsEhEBJQWTBXaw5BiweH1OBDIpaFEqoA2VCB0D0Ch1SFtQBQc5sdFBnLpqXpEsMqqhgakiyMUcdlw0CcWFRUDYcZU+oQCiN4mYnGLLVBuODwKBZOdWnAMproEg6GPDPhJv17T+iAY8GVcUhwLGANZ9M4YSngoL6/pUfostFiB7jJZbIDaQPXl9GxQpPyaHpMuKMGJGUPxCAgOxOD0pA2BlCoST8FB8aVTq7HVckWNkufgEBEeQtzyhb6m+VFeIabBAYzqCRK8yJbeGBAeUgEizc/AAbUlEkaLDtIe6nDNJBIdY/uVYozzpA+BYXItksTB2hSBWeIYIFJEhNis4Sk4QHiCBmXJkvmSRnFIFDGSOlQUtu/5GXEBwlMDgW6hnUWcs0WMBkPK8yNgIJGezCmsgSlQXvQqVRS3qGAUONL8CBjUs0R5zxORgDB/toeSKPFKi0iPwIH0oQpQlUqiPwF7+IpEuldsPlEAGT5KqiUJsUZmFWDUWtqeLnYVZkJdgdeH2AOUF2ndbKEwLXosrCA2AILyLwbKOHy+0MacJpUWFC1TUpjdcYbQbCHaM7Y9uPym8AtPmtUiaA96ri+RMJgp8AxuD+L8wR9THPQkx6KkQD0BDAhEHjpOSiNOLWmKNXE1BLlKFMnVFwAxvF9oI1GC+pJsuuZJMR4pR/JUrcDMwdwi67gwgC94v8aauBBgAlo2e4tIHR7VCmgPeeg4qd67a5ILIgP+OleXyA+MCx4kt+gwbn2hzBUqVKhNddloNb3h1KaIFOktT2RAEMeNkxUE9CrUdRaX4Cbj0np2iZooYA2eKzYghrYHaYKDjTcwJFyUFK3IhjvUsFBjRN4sYsT4wDEXYZwsUIRA6hSCtzBlL0yzW+EGUeND3l5yAWRUe4A8ry5EThx2odRwpdG3NFGLzBoVNhQAw3D2QH3eQmQb8nCDCMcWZqIGBwDwy5IFAuWo9gAE6BU2GhlYX6lepArIT4yNNAjPFeWPeTh3EE6Oe7PGM0Wnvumhzg6sshN5U3QUtjBZnjHtAbMudebHJn5EaAeip24NqEN0GtXMIRcYRqsvpB8PNe4ktmKDAKlXfdxo3MkcooJQjKH+NZ49cHbeJoXZ0W/zHXoBQbR2xfYrh05yqG5RnWI0Xi2cGJ8cBWHGtMHIkxIJb6hQJMuYSBEkUMRiIByURJoUSru5cXZz5aQ1MD7QAlxtSDmU2FBjZKzf80i2UEcjbaFEfO2OksuhV4H37/LTTm5IiA25GkMaLj5UuUF2Ez+uOlw08JIFydSV2bADugaRGAcH9bkn27bqdJeqQ6LS1v0EWIIw3ZRIeIcRByMPw9SbahC0vRLq0YDgspebCIULssyYzR5ye4bxCzUKFSZ1ddoXjA5gUHBJhAKyg9GGWljV6BiTUalRcBDbRUV4MOZkgy56ZQwpspUbUv84fxrNHtQqCtJJsf5lVech0R9Hn7yiIJWOLsgWIBAWkDCHwUGdQVkfl4PjtqNtc9Mn/QnSk8uxhTXUZJlqSdFbwyB+4RNg1SdEu1GfptJ3hVXskkT0rFl4dU5AIR+fm+OApqVPgNn7t7VcudClY19wNwZp9TZjJLNmHMovlAqctS+tjRvCUZY22pDgEF18yHCOqxh5fxzERuhdfDLi4JtWehBkI4dkqUZiCqyEx8qgGSIz2fQYOFCW5nhsXbyzyy5oYX4z8mMFdhGl4REJjQp4Rsp5bxJ39wvxn70IF1yGlL3egCGw5GIs/CGSNKSupBjEHsig7UdVdmcL5GKGniIc2xQ1SlpHt0XGCO4wkl9M3sGz4a9WXl5xSHBpCpHBgOi1uGoKF+ZwXxx0ajtWXEfl+QrIDSLXOyVgT8E0hhS9mQtjuPSKe/uF2mkTFpjWyFVpw/OHfg3YUxEijVSTONX60hAYKj6gfyvt2NFhEG5vDpEdzGRkcqsrM3IFayvQp5Svn/v6han00rZrvHl3PTzuXVyvsVlbVUuwsiqdycNdcRCxS2ji87KWL+2Uh5wRQG2F19SGXaroYCwyjmMPYoqLn/jpJEi4xQmJ6FMedTy2qNHBa+0c41dv/9b2QGmai/seHdRRuBBkI8QnUMhqFxk9K0pOMboaOU58wCYB5lx6CJTdS9klCtu7c102NsZA7SWTRKWBcFDxxhXsgK0K263RwxAYFs5Ma8hQ5LbMCd6UKwYpj+cXcryFxjSxHw/dD0unLmHWfg0CY0aV5X2rcXDA/DwvWTA2TNpk2eOKduKWNjFwZ4A5eIsitcbVADiISw6mTPuqmXCs4cQdIse/QtPeyBzcEmL82S1uZg+IDSI+FqnBt0quVghsZz/6OGTfwKRC/5NL3DM+qLZJuLZ6xqRxNgYbHqfiEPrZBpRUJjl8YxTLrXDgPgm3Cezekc/8hItRYW/uQ3hKTqWpu8AnfnCL9Yb2IC08KrpWtp98sZGabDzWFglipzfsCsxx/EK8miAS1q3ZK9JXkx6+XdNNw5lXvBUjb2YPYiMvuLGs0p2MPC8SRLtZYGuY1SjaRGDjDOlbx1jwa75NxsSosPgKARt3gkRx3DiL3s6N0dZsMhp3FCNT6iYcvo0Nt/ILm4/l1p2YW+Du0eXKXWSJbaHBpsCSxYY3faKgcQsclPOxk57vCoNAXi2qR68rmtYAAKJN/+R3yMOt8kV3uKD86q/p6um0S/IGXvDeHRsVyWaFU/6eM3hsuEt8EBtw6I5Mh3Z6vY2N66mLWUl0yMHHnqBHo8p671nuFx+MO7SxcfEFm4MiW1NFaGcsjDjAGdC1+rHKXMwtlhvgIG09e2rz0nZ6PZxXzpLPOBwa+41A5Tci5LLPF+t8gyBJSdYKbLUiUwNnf05DP7FdL4BLcDKSSlxi4+o3nGP+rDEYcaj/Cp4mfO7nijWk/mYB6opsfhHTO8WlmcF6n3zRPrRGqkAtfhVPrhfvfLrBfKJLmkgS0GffjZDL5+OkmkRvkYHfXlPJ1Y46s2M7a9J1bbKP+nwDxdL/c62/l8/7hUrrWOGQrPDW9sXp0NhfyPMbN8iXTqTSO3Xm4m6xfNgeVLpuLjcpJjuw/MWkcLC33teXbODl3dz4bz0fjpP4hBI2KuwDnbCkbd3tQ5/CdhD76QYrs5M1LuMbhrD2LrJ+MG+yYTV1zUsJ/cdTXNz5MQz8XgGHx12OpBiZ3iyuKiSfjpNqgz/On/yebpDTHJx1rPxugW/lgjdwzuNHt1i+AmT+WGhQHLFoFzMDJxyuj8JRd7Gjsuhn+maNUaj8LnlY7fvDfNK2SsTX74KNeui1JIuVu8ahM1uXkWMeuRHq9JMxLB0oH6svdOKQLCY96kcydNevMPWyl+iTqQ2gTyHtbnmwafMej7aEuezK7g/5hdqHe3FR3VfvwuXofKfBQXaJrZ9NtoC0md6ssnfm8KF8YZ/s1ZOo4FT6esAB6yTcRo1On/w7xjdow3qyiqX992f8Qu0eHE7C+dUj4f35wyZB8hWbhOjYL+UaAvEtBrXUr0OwWP64PTh9nLoeJsQW8S3My4npyPWaMuJQdSjUE5Y6f5Qi1/MfK+uLj9iDtDPTqm3bzK/Jng7JRj8d2rZy6Qyp61vG/H69vZcm10Km5j+cLNUnhYFHu311/Ykt/V1di5PdRm6tqyjB/aYkue5gmf94YMBHWpExlGtPtmOjeqYNybaKWvOy7WhjDjD9XkW17PW4hcDcru//X7A5L9dTmkOYQ+cyIaedF3m/a57+/f9ne/61+9zq/93n9R8IWADrpY5DmAAAAABJRU5ErkJggg=="></image></g><g transform="matrix(0.9999780654907227,0,0,0.9999780654907227,-491.3211669921875,345.8320617675781)" opacity="0.055341690605766436" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path stroke-linecap="butt" stroke-linejoin="miter" fill-opacity="0" stroke-miterlimit="10" stroke="rgb(255,255,255)" stroke-opacity="1" stroke-width="0.75" d=" M742.6209716796875,-166.36599731445312 C742.6209716796875,-166.36599731445312 1092.6209716796875,-166.36599731445312 1092.6209716796875,-166.36599731445312"></path></g></g><g transform="matrix(0.9999769926071167,0,0,0.9999769926071167,26.8851318359375,-208.83685302734375)" opacity="0.9999996872205996" style="display: block;"><g opacity="0.8" transform="matrix(1,0,0,1,381.09600830078125,340.01300048828125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0,-1.559999942779541 C-0.47999998927116394,-1.559999942779541 -0.8799999952316284,-1.4079999923706055 -1.2000000476837158,-1.1039999723434448 C-1.503999948501587,-0.8159999847412109 -1.656000018119812,-0.4480000138282776 -1.656000018119812,0 C-1.656000018119812,0.4480000138282776 -1.503999948501587,0.8240000009536743 -1.2000000476837158,1.128000020980835 C-0.8799999952316284,1.4160000085830688 -0.47999998927116394,1.559999942779541 0,1.559999942779541 C0.47999998927116394,1.559999942779541 0.871999979019165,1.4160000085830688 1.1759999990463257,1.128000020980835 C1.496000051498413,0.8240000009536743 1.656000018119812,0.4480000138282776 1.656000018119812,0 C1.656000018119812,-0.4480000138282776 1.496000051498413,-0.8159999847412109 1.1759999990463257,-1.1039999723434448 C0.871999979019165,-1.4079999923706055 0.47999998927116394,-1.559999942779541 0,-1.559999942779541z"></path></g><g opacity="0.8" transform="matrix(1,0,0,1,371.39898681640625,347.3330078125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M3.8399999141693115,-0.024000000208616257 C3.184000015258789,-0.40799999237060547 2.440000057220459,-0.7360000014305115 1.6080000400543213,-1.0080000162124634 C0.7440000176429749,-1.312000036239624 -0.024000000208616257,-1.5759999752044678 -0.6959999799728394,-1.7999999523162842 C-1.3680000305175781,-2.0399999618530273 -1.8960000276565552,-2.3359999656677246 -2.2799999713897705,-2.687999963760376 C-2.6640000343322754,-3.0399999618530273 -2.8559999465942383,-3.5439999103546143 -2.8559999465942383,-4.199999809265137 C-2.8559999465942383,-4.935999870300293 -2.5999999046325684,-5.51200008392334 -2.0880000591278076,-5.927999973297119 C-1.559999942779541,-6.343999862670898 -0.8960000276565552,-6.544000148773193 -0.09600000083446503,-6.5279998779296875 C0.527999997138977,-6.5279998779296875 1.055999994277954,-6.400000095367432 1.4880000352859497,-6.144000053405762 C1.9199999570846558,-5.888000011444092 2.247999906539917,-5.559999942779541 2.4719998836517334,-5.159999847412109 C2.696000099182129,-4.776000022888184 2.808000087738037,-4.368000030517578 2.808000087738037,-3.936000108718872 C2.808000087738037,-3.936000108718872 5.495999813079834,-3.936000108718872 5.495999813079834,-3.936000108718872 C5.480000019073486,-4.880000114440918 5.239999771118164,-5.703999996185303 4.776000022888184,-6.4079999923706055 C4.311999797821045,-7.127999782562256 3.6640000343322754,-7.688000202178955 2.8320000171661377,-8.088000297546387 C2.0160000324249268,-8.48799991607666 1.0640000104904175,-8.687999725341797 -0.024000000208616257,-8.687999725341797 C-1.128000020980835,-8.687999725341797 -2.0959999561309814,-8.48799991607666 -2.927999973297119,-8.088000297546387 C-3.74399995803833,-7.703999996185303 -4.383999824523926,-7.159999847412109 -4.8480000495910645,-6.455999851226807 C-5.296000003814697,-5.751999855041504 -5.51200008392334,-4.927999973297119 -5.495999813079834,-3.9839999675750732 C-5.495999813079834,-2.864000082015991 -5.176000118255615,-1.9600000381469727 -4.535999774932861,-1.2719999551773071 C-3.880000114440918,-0.5839999914169312 -2.9040000438690186,-0.024000000208616257 -1.6080000400543213,0.40799999237060547 C-0.9520000219345093,0.6320000290870667 -0.328000009059906,0.8399999737739563 0.2639999985694885,1.031999945640564 C0.8560000061988831,1.2239999771118164 1.3760000467300415,1.440000057220459 1.8240000009536743,1.6799999475479126 C2.2880001068115234,1.9199999570846558 2.6480000019073486,2.2239999771118164 2.9040000438690186,2.5920000076293945 C3.1760001182556152,2.9600000381469727 3.312000036239624,3.440000057220459 3.312000036239624,4.0320000648498535 C3.312000036239624,4.76800012588501 3.0320000648498535,5.375999927520752 2.4719998836517334,5.855999946594238 C1.9279999732971191,6.320000171661377 1.1679999828338623,6.552000045776367 0.19200000166893005,6.552000045776367 C-0.9120000004768372,6.552000045776367 -1.7680000066757202,6.239999771118164 -2.375999927520752,5.616000175476074 C-2.9839999675750732,4.992000102996826 -3.2960000038146973,4.23199987411499 -3.312000036239624,3.3359999656677246 C-3.312000036239624,3.3359999656677246 -6,3.3359999656677246 -6,3.3359999656677246 C-5.984000205993652,4.440000057220459 -5.71999979019165,5.392000198364258 -5.208000183105469,6.191999912261963 C-4.679999828338623,6.992000102996826 -3.9519999027252197,7.607999801635742 -3.0239999294281006,8.039999961853027 C-2.0959999561309814,8.472000122070312 -1.0160000324249268,8.687999725341797 0.2160000056028366,8.687999725341797 C1.4800000190734863,8.687999725341797 2.5439999103546143,8.46399974822998 3.4079999923706055,8.015999794006348 C4.271999835968018,7.552000045776367 4.920000076293945,6.960000038146973 5.3520002365112305,6.239999771118164 C5.783999919891357,5.504000186920166 6,4.711999893188477 6,3.864000082015991 C6,2.9040000438690186 5.800000190734863,2.111999988555908 5.400000095367432,1.4880000352859497 C5.015999794006348,0.8640000224113464 4.495999813079834,0.36000001430511475 3.8399999141693115,-0.024000000208616257z"></path></g><g opacity="0.8" transform="matrix(1,0,0,1,381.09600830078125,349.781005859375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-1.2719999551773071,5.952000141143799 C-1.2719999551773071,5.952000141143799 1.2719999551773071,5.952000141143799 1.2719999551773071,5.952000141143799 C1.2719999551773071,5.952000141143799 1.2719999551773071,-5.952000141143799 1.2719999551773071,-5.952000141143799 C1.2719999551773071,-5.952000141143799 -1.2719999551773071,-5.952000141143799 -1.2719999551773071,-5.952000141143799 C-1.2719999551773071,-5.952000141143799 -1.2719999551773071,5.952000141143799 -1.2719999551773071,5.952000141143799z"></path></g><g opacity="0.8" transform="matrix(1,0,0,1,419.1080017089844,349.781005859375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-1.2719999551773071,5.952000141143799 C-1.2719999551773071,5.952000141143799 1.2719999551773071,5.952000141143799 1.2719999551773071,5.952000141143799 C1.2719999551773071,5.952000141143799 1.2719999551773071,-5.952000141143799 1.2719999551773071,-5.952000141143799 C1.2719999551773071,-5.952000141143799 -1.2719999551773071,-5.952000141143799 -1.2719999551773071,-5.952000141143799 C-1.2719999551773071,-5.952000141143799 -1.2719999551773071,5.952000141143799 -1.2719999551773071,5.952000141143799z"></path></g><g opacity="0.8" transform="matrix(1,0,0,1,428.8890075683594,349.6369934082031)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M4.331999778747559,-4.751999855041504 C3.5160000324249268,-5.6479997634887695 2.388000011444092,-6.0960001945495605 0.9480000138282776,-6.0960001945495605 C0.019999999552965164,-6.0960001945495605 -0.8040000200271606,-5.880000114440918 -1.5240000486373901,-5.447999954223633 C-2.2279999256134033,-5.0320000648498535 -2.7639999389648438,-4.455999851226807 -3.131999969482422,-3.7200000286102295 C-3.131999969482422,-3.7200000286102295 -3.3239998817443848,-5.808000087738037 -3.3239998817443848,-5.808000087738037 C-3.3239998817443848,-5.808000087738037 -5.579999923706055,-5.808000087738037 -5.579999923706055,-5.808000087738037 C-5.579999923706055,-5.808000087738037 -5.579999923706055,6.0960001945495605 -5.579999923706055,6.0960001945495605 C-5.579999923706055,6.0960001945495605 -3.0360000133514404,6.0960001945495605 -3.0360000133514404,6.0960001945495605 C-3.0360000133514404,6.0960001945495605 -3.0360000133514404,-0.04800000041723251 -3.0360000133514404,-0.04800000041723251 C-3.0360000133514404,-1.2960000038146973 -2.7320001125335693,-2.25600004196167 -2.124000072479248,-2.927999973297119 C-1.5,-3.5999999046325684 -0.7160000205039978,-3.936000108718872 0.2280000001192093,-3.936000108718872 C2.115999937057495,-3.936000108718872 3.059999942779541,-2.7839999198913574 3.059999942779541,-0.47999998927116394 C3.059999942779541,-0.47999998927116394 3.059999942779541,6.0960001945495605 3.059999942779541,6.0960001945495605 C3.059999942779541,6.0960001945495605 5.579999923706055,6.0960001945495605 5.579999923706055,6.0960001945495605 C5.579999923706055,6.0960001945495605 5.579999923706055,-0.7440000176429749 5.579999923706055,-0.7440000176429749 C5.579999923706055,-2.5199999809265137 5.164000034332275,-3.8559999465942383 4.331999778747559,-4.751999855041504z"></path></g><g opacity="0.8" transform="matrix(1,0,0,1,419.1080017089844,340.01300048828125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0,-1.559999942779541 C-0.47999998927116394,-1.559999942779541 -0.8799999952316284,-1.4079999923706055 -1.2000000476837158,-1.1039999723434448 C-1.503999948501587,-0.8159999847412109 -1.656000018119812,-0.4480000138282776 -1.656000018119812,0 C-1.656000018119812,0.4480000138282776 -1.503999948501587,0.8240000009536743 -1.2000000476837158,1.128000020980835 C-0.8799999952316284,1.4160000085830688 -0.47999998927116394,1.559999942779541 0,1.559999942779541 C0.47999998927116394,1.559999942779541 0.871999979019165,1.4160000085830688 1.1759999990463257,1.128000020980835 C1.496000051498413,0.8240000009536743 1.656000018119812,0.4480000138282776 1.656000018119812,0 C1.656000018119812,-0.4480000138282776 1.496000051498413,-0.8159999847412109 1.1759999990463257,-1.1039999723434448 C0.871999979019165,-1.4079999923706055 0.47999998927116394,-1.559999942779541 0,-1.559999942779541z"></path></g><g opacity="0.8" transform="matrix(1,0,0,1,390.5299987792969,352.4209899902344)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.36000001430511475,-8.880000114440918 C-1.3680000305175781,-8.880000114440918 -2.2320001125335693,-8.687999725341797 -2.9519999027252197,-8.303999900817871 C-3.671999931335449,-7.920000076293945 -4.223999977111816,-7.400000095367432 -4.607999801635742,-6.74399995803833 C-4.97599983215332,-6.104000091552734 -5.159999847412109,-5.392000198364258 -5.159999847412109,-4.607999801635742 C-5.159999847412109,-3.312000036239624 -4.696000099182129,-2.2639999389648438 -3.7679998874664307,-1.4639999866485596 C-3.7679998874664307,-1.4639999866485596 -5.831999778747559,0.6480000019073486 -5.831999778747559,0.6480000019073486 C-5.831999778747559,0.6480000019073486 -5.831999778747559,1.2239999771118164 -5.831999778747559,1.2239999771118164 C-5.639999866485596,1.399999976158142 -5.423999786376953,1.5759999752044678 -5.184000015258789,1.7519999742507935 C-4.927999973297119,1.9119999408721924 -4.631999969482422,2.063999891281128 -4.296000003814697,2.2079999446868896 C-4.872000217437744,2.640000104904175 -5.296000003814697,3.0959999561309814 -5.567999839782715,3.5759999752044678 C-5.823999881744385,4.072000026702881 -5.952000141143799,4.567999839782715 -5.952000141143799,5.064000129699707 C-5.952000141143799,6.343999862670898 -5.440000057220459,7.296000003814697 -4.415999889373779,7.920000076293945 C-3.375999927520752,8.5600004196167 -2.0239999294281006,8.880000114440918 -0.36000001430511475,8.880000114440918 C0.871999979019165,8.880000114440918 1.8799999952316284,8.680000305175781 2.6640000343322754,8.279999732971191 C3.4639999866485596,7.895999908447266 4.047999858856201,7.383999824523926 4.415999889373779,6.74399995803833 C4.800000190734863,6.119999885559082 4.992000102996826,5.455999851226807 4.992000102996826,4.751999855041504 C4.992000102996826,3.7279999256134033 4.639999866485596,2.9119999408721924 3.936000108718872,2.303999900817871 C3.2320001125335693,1.6799999475479126 2.0880000591278076,1.2960000038146973 0.5040000081062317,1.1519999504089355 C-0.29600000381469727,1.0880000591278076 -0.9279999732971191,1.0240000486373901 -1.3919999599456787,0.9599999785423279 C-1.8559999465942383,0.8960000276565552 -2.2160000801086426,0.8240000009536743 -2.4719998836517334,0.7440000176429749 C-2.7279999256134033,0.6639999747276306 -2.944000005722046,0.5680000185966492 -3.119999885559082,0.4560000002384186 C-3.119999885559082,0.4560000002384186 -2.0880000591278076,-0.5519999861717224 -2.0880000591278076,-0.5519999861717224 C-1.559999942779541,-0.40799999237060547 -0.984000027179718,-0.335999995470047 -0.36000001430511475,-0.335999995470047 C0.6480000019073486,-0.335999995470047 1.503999948501587,-0.527999997138977 2.2079999446868896,-0.9120000004768372 C2.927999973297119,-1.2960000038146973 3.4719998836517334,-1.8079999685287476 3.8399999141693115,-2.447999954223633 C4.223999977111816,-3.0880000591278076 4.415999889373779,-3.808000087738037 4.415999889373779,-4.607999801635742 C4.415999889373779,-5.4079999923706055 4.223999977111816,-6.136000156402588 3.8399999141693115,-6.791999816894531 C3.8399999141693115,-6.791999816894531 5.952000141143799,-6.9120001792907715 5.952000141143799,-6.9120001792907715 C5.952000141143799,-6.9120001792907715 5.952000141143799,-8.592000007629395 5.952000141143799,-8.592000007629395 C5.952000141143799,-8.592000007629395 1.5119999647140503,-8.592000007629395 1.5119999647140503,-8.592000007629395 C0.9359999895095825,-8.784000396728516 0.31200000643730164,-8.880000114440918 -0.36000001430511475,-8.880000114440918z M-2.0880000591278076,2.808000087738037 C-1.496000051498413,2.9040000438690186 -0.8159999847412109,2.9760000705718994 -0.04800000041723251,3.0239999294281006 C0.9760000109672546,3.1040000915527344 1.6720000505447388,3.303999900817871 2.0399999618530273,3.624000072479248 C2.4079999923706055,3.9600000381469727 2.5920000076293945,4.360000133514404 2.5920000076293945,4.823999881744385 C2.5920000076293945,5.447999954223633 2.319999933242798,5.943999767303467 1.7760000228881836,6.311999797821045 C1.2319999933242798,6.696000099182129 0.5199999809265137,6.888000011444092 -0.36000001430511475,6.888000011444092 C-1.2719999551773071,6.888000011444092 -2.0320000648498535,6.711999893188477 -2.640000104904175,6.360000133514404 C-3.247999906539917,6.02400016784668 -3.552000045776367,5.51200008392334 -3.552000045776367,4.823999881744385 C-3.552000045776367,4.48799991607666 -3.447999954223633,4.144000053405762 -3.240000009536743,3.7920000553131104 C-3.0320000648498535,3.440000057220459 -2.6480000019073486,3.111999988555908 -2.0880000591278076,2.808000087738037z M1.4160000085830688,-2.9040000438690186 C0.9679999947547913,-2.5199999809265137 0.37599998712539673,-2.328000068664551 -0.36000001430511475,-2.328000068664551 C-1.1119999885559082,-2.328000068664551 -1.7200000286102295,-2.5199999809265137 -2.184000015258789,-2.9040000438690186 C-2.631999969482422,-3.303999900817871 -2.8559999465942383,-3.864000082015991 -2.8559999465942383,-4.584000110626221 C-2.8559999465942383,-5.320000171661377 -2.631999969482422,-5.880000114440918 -2.184000015258789,-6.263999938964844 C-1.7200000286102295,-6.6479997634887695 -1.1119999885559082,-6.840000152587891 -0.36000001430511475,-6.840000152587891 C0.37599998712539673,-6.840000152587891 0.9679999947547913,-6.6479997634887695 1.4160000085830688,-6.263999938964844 C1.8799999952316284,-5.880000114440918 2.111999988555908,-5.320000171661377 2.111999988555908,-4.584000110626221 C2.111999988555908,-3.864000082015991 1.8799999952316284,-3.303999900817871 1.4160000085830688,-2.9040000438690186z"></path></g><g opacity="0.8" transform="matrix(1,0,0,1,403.9020080566406,349.6369934082031)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0.9480000138282776,-6.0960001945495605 C0.019999999552965164,-6.0960001945495605 -0.8040000200271606,-5.880000114440918 -1.5240000486373901,-5.447999954223633 C-2.2279999256134033,-5.0320000648498535 -2.7639999389648438,-4.455999851226807 -3.131999969482422,-3.7200000286102295 C-3.131999969482422,-3.7200000286102295 -3.3239998817443848,-5.808000087738037 -3.3239998817443848,-5.808000087738037 C-3.3239998817443848,-5.808000087738037 -5.579999923706055,-5.808000087738037 -5.579999923706055,-5.808000087738037 C-5.579999923706055,-5.808000087738037 -5.579999923706055,6.0960001945495605 -5.579999923706055,6.0960001945495605 C-5.579999923706055,6.0960001945495605 -3.0360000133514404,6.0960001945495605 -3.0360000133514404,6.0960001945495605 C-3.0360000133514404,6.0960001945495605 -3.0360000133514404,-0.04800000041723251 -3.0360000133514404,-0.04800000041723251 C-3.0360000133514404,-1.2960000038146973 -2.7320001125335693,-2.25600004196167 -2.124000072479248,-2.927999973297119 C-1.5,-3.5999999046325684 -0.7160000205039978,-3.936000108718872 0.2280000001192093,-3.936000108718872 C2.115999937057495,-3.936000108718872 3.059999942779541,-2.7839999198913574 3.059999942779541,-0.47999998927116394 C3.059999942779541,-0.47999998927116394 3.059999942779541,6.0960001945495605 3.059999942779541,6.0960001945495605 C3.059999942779541,6.0960001945495605 5.579999923706055,6.0960001945495605 5.579999923706055,6.0960001945495605 C5.579999923706055,6.0960001945495605 5.579999923706055,-0.7440000176429749 5.579999923706055,-0.7440000176429749 C5.579999923706055,-2.5199999809265137 5.164000034332275,-3.8559999465942383 4.331999778747559,-4.751999855041504 C3.5160000324249268,-5.6479997634887695 2.388000011444092,-6.0960001945495605 0.9480000138282776,-6.0960001945495605z"></path></g></g><g class="png" clip-path="url(#__lottie_element_897)" transform="matrix(0.7659965753555298,0,0,0.7659965753555298,279.86126708984375,511.41363525390625)" opacity="0.9999932705661607" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_245)" transform="matrix(0.7658882737159729,0,0,0.7658882737159729,279.8788757324219,511.3905334472656)" opacity="1" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_397)" transform="matrix(0.7299758195877075,0,0,0.7299758195877075,299.3709411621094,511.8205871582031)" opacity="1" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_439)" transform="matrix(0.7642555236816406,0,0,0.7642555236816406,328.6820983886719,531.6568603515625)" opacity="1" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_483)" transform="matrix(0.765990138053894,0,0,0.765990138053894,293.98956298828125,532.0745239257812)" opacity="1" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_545)" transform="matrix(0.765990138053894,0,0,0.765990138053894,260.4649658203125,532.0747680664062)" opacity="1" style="display: block;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_601)" transform="matrix(0.7659915685653687,0,0,0.7659915685653687,398.109375,532.0751953125)" opacity="1" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_665)" transform="matrix(0.7659915685653687,0,0,0.7659915685653687,363.8470153808594,532.074951171875)" opacity="1" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_689)" transform="matrix(0.7659915685653687,0,0,0.7659915685653687,328.51031494140625,532.0740966796875)" opacity="1" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_701)" transform="matrix(0.7659915685653687,0,0,0.7659915685653687,293.9893493652344,532.0748901367188)" opacity="1" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_713)" transform="matrix(0.7659915685653687,0,0,0.7659915685653687,260.46466064453125,532.0751342773438)" opacity="1" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_719)" transform="matrix(0.7659915685653687,0,0,0.7659915685653687,398.109375,532.0751953125)" opacity="1" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_731)" transform="matrix(0.7659915685653687,0,0,0.7659915685653687,363.8470153808594,532.074951171875)" opacity="1" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_749)" transform="matrix(0.7659915685653687,0,0,0.7659915685653687,328.51031494140625,532.0740966796875)" opacity="0.8207035344373758" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g class="png" clip-path="url(#__lottie_element_755)" transform="matrix(0.7659915685653687,0,0,0.7659915685653687,293.9893493652344,532.0748901367188)" opacity="1.6781316958258686e-9" style="display: none;"><image width="212px" height="28px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAANQAAAAcCAMAAAAuu2HzAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAFQUExURUdwTP///////////1Uj7lQm5////yI0Rv///////wBS/y1C6v///////1Qx2CM1RyI0R////////wBR/wBS/yI0RiIzRyA4SABR/////////1Mz1gBS/1M6zABS/1NFulMw2lUj7VNDvlQi7wBR/yM0Rv///wBS/wBR/////1NBv1JCvv///1Qy1VM7yy1C7////////////////wBS/yM0R/b2/u3p+zuZ/PWEH3c+GlUf81NDveTd/FNHt+nj+/Lv/FQ01FQ5zPv6/uV4I1Q/xNzq/9JvKFQt3oFn4qyc6qpgJr5lJuby/4O//lI/NjI4QpvL/mU+5j17/5Z87rvX/1Om/d1zI3ei/9PK9hhi/6vQ/2Gt/ZNVKUeg/NrX+V6S/8nA8mQ8I5C0/5eX9ZWJ2npTN8G18X52c6+dkOjn95++/09WX3FwcterjRYWFg+XEcYAAAAzdFJOUwDvPFz+/RvpnvjbIN9AXWDKMJU/7kCQIFqQ0JB1bsTvRd/fzI1Qv/WdbtBAdu+/xyzcOFGbOY4AAAUxSURBVFjDvZj5U+JIFMfDSFAOlRXGAyzx1tEZa3YDwXAZIJIAERA5RURmymOd6///bV8nnXCkE5iS2S+UaULb5FPv9bf7NUVper/qnY9E5r2r76kptLgRsDGM7XBjcZrep8GTUCoVOgueUjOXb3/LHw77t/Z9hq+WAUjT/PKkgeY8jC7P3KTeS7spXbtLs0Va3wrrcq2PfLXmjYxofs1qIEeAGZHHYdXbfZIa0a57dkjuISQFy00OE6Y6sAiTjRmTzSJYS6FEAlASqYTKlEiFtmeWef7wmI70YB1ECDJNwTmGIFOq7QRBFino+LiwQFGfFzR9sMgCX5ggTLU2T4Iyy0CHjQRlM/ltd0iheGy/vvb6j9C6RB9DphnogMnqpKh3g5H/NqVy+0lQR+rYiKlSHgYqV+CPlzyUhyEqQO69ewkq8AyTzcL79fHyMoHunJg96EdmHIr5YNbXFSbKpUwoBSNWfKqoRJWnYqxsmoBDycdesZMScAkRtLMDFS5VmSXgghFqxcz3NIrbVj3L5L7daJ83caAiX2KxolBEkpvFWOyLkoBWgWKvG+fn543SlW6BZoHqZ9ksemtUGbiX2TWDcjqdtumgtEA98LjjV3zDD2uuEp4YksiBxC5qKjcJq/AiHqBzf451rf04YRU+zWQyjyybZXVl2XxGkfUqPAWU5hI3g54alY9ajeBIqVSi0lAiFVk1DrWh/vs10Nx16p0SuuIk3DD2DsLTJ9lR9VWo4Fuh9nHu5RgmB5l304OuOAOPKe8QVJHjikpDtQ2CVRxqUI2c0ri6Oy+ZW8VZJpPn2ef8M5vMF3iW51mWZ6uIKW1iFY5F0FRQeNl9gJlwq9HVNatQ/bysMAkcJxQHkSJMKs3PO9pcYju6qxt7f0qnCzyfT+f5QjrNYz2mkT691Siwn9cZ5hvOw1arhV2dwpbXlZuCgKAEoSl3sREah0K/UmqoRLl7PJ9KagYae8PD98ehogUFKv1WKDx/ID4PBlfXoJBFiE2Oa4owsQQrqDqY3pXChF0CJlbJFOo5GgUoRBIF8fBK9nr9Pw2lpt8TQAmyzHGyDOHinqzSr6RQ5e6x95UwJDH9pEI02qu1o8laITqkvDTD9Puqp5+Gh42iiNycE5Q3qGhmFAEcmkYdmEooVvDhTknHQ2PvPUmqRaOv8BpTVTojP+pf4BPv0HUgh6VRtBiGv9Vi1lHvbWFLF9FcUng4NLdEa0u/VlaoDo4W9nSSpUuSlLwwqiBJwbfu0DVLZ1VLv60PW7q6+HaBSRREWRabIlB1Jyy+iAqNkmsM1inC4rsNUDUjU7sqSW8uP7TF90HdsQ0vvut4m1QWmoL48+XXr5efIjTLk7ZJ1woTrFMNjYm4TdqhJbp/cZGE14USMtRqV2l6Z8I2CS7Of2AKw2VhwjapNb6hcGkb2orcLVde/gW9VMpdOTJxQ5vDV31TS97Q0jRdbScBBQsaz8BET7OhhbnlhIsZ1Ka+of2WQ+vlyIaWGpTy33/8+D4oqMhjBcilh4fce4+m7dVCcqBeDZBMAzVSemAo09Jjy6L0+LNFIm0H5fsqULJdQx/t9FRFogrl+c0iMYzHXiZBHcymnF9CEPF4NV+r1fJVu6qJ5bwDVfKfKWplxaqcXycxbeqHFMaDl+WZHbzQ8ThQabLH7fTMTsnWj8yZjBnotT4iGyvpA9ZHZDvxEe3N8ohsrKR3+f6/w8whrJ0ZH2ZuDmH5Nw1f/+6x8yE6dg5Md+y8HdxDKbgX3KZmLt+xYoOuY5+eAv8BxhFF+FfytiQAAAAASUVORK5CYII="></image></g><g transform="matrix(1.3937740325927734,0,0,1.3937740325927734,-841.506103515625,-10.683563232421875)" opacity="0.3" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,916.7100219726562,263.1419982910156)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M2.6700000762939453,-3.8350000381469727 C2.109999895095825,-4.122000217437744 1.4600000381469727,-4.264999866485596 0.699999988079071,-4.264999866485596 C-0.03999999910593033,-4.264999866485596 -0.7200000286102295,-4.135000228881836 -1.340000033378601,-3.875 C-1.9600000381469727,-3.621999979019165 -2.5,-3.26200008392334 -2.9600000381469727,-2.7950000762939453 C-3.4200000762939453,-2.3289999961853027 -3.7799999713897705,-1.784999966621399 -4.03000020980835,-1.1649999618530273 C-4.289999961853027,-0.5450000166893005 -4.409999847412109,0.12800000607967377 -4.409999847412109,0.8550000190734863 C-4.409999847412109,1.5479999780654907 -4.260000228881836,2.1480000019073486 -3.9600000381469727,2.6549999713897705 C-3.6600000858306885,3.1679999828338623 -3.2300000190734863,3.565000057220459 -2.690000057220459,3.8450000286102295 C-2.1500000953674316,4.125 -1.5099999904632568,4.264999866485596 -0.7799999713897705,4.264999866485596 C-0.05000000074505806,4.264999866485596 0.6200000047683716,4.135000228881836 1.2400000095367432,3.875 C1.2400000095367432,3.875 1.0700000524520874,3.174999952316284 1.0700000524520874,3.174999952316284 C0.8100000023841858,3.2880001068115234 0.5400000214576721,3.375 0.25,3.434999942779541 C-0.03999999910593033,3.494999885559082 -0.33000001311302185,3.5250000953674316 -0.6200000047683716,3.5250000953674316 C-1.5399999618530273,3.5250000953674316 -2.259999990463257,3.2809998989105225 -2.7799999713897705,2.7950000762939453 C-3.309999942779541,2.315000057220459 -3.569999933242798,1.6380000114440918 -3.569999933242798,0.7649999856948853 C-3.569999933242798,0.16500000655651093 -3.4700000286102295,-0.39500001072883606 -3.259999990463257,-0.9150000214576721 C-3.059999942779541,-1.434999942779541 -2.7699999809265137,-1.8890000581741333 -2.390000104904175,-2.2750000953674316 C-2.0199999809265137,-2.6689999103546143 -1.5800000429153442,-2.9749999046325684 -1.0700000524520874,-3.194999933242798 C-0.5699999928474426,-3.421999931335449 -0.019999999552965164,-3.5350000858306885 0.5799999833106995,-3.5350000858306885 C1.5199999809265137,-3.5350000858306885 2.25,-3.2920000553131104 2.7799999713897705,-2.805000066757202 C3.299999952316284,-2.325000047683716 3.569999933242798,-1.6619999408721924 3.569999933242798,-0.8149999976158142 C3.569999933242798,-0.5490000247955322 3.5399999618530273,-0.2750000059604645 3.490000009536743,0.004999999888241291 C3.430000066757202,0.27799999713897705 3.3499999046325684,0.531000018119812 3.25,0.7649999856948853 C3.1500000953674316,0.9980000257492065 3.0199999809265137,1.187999963760376 2.859999895095825,1.3350000381469727 C2.700000047683716,1.4809999465942383 2.5199999809265137,1.5549999475479126 2.319999933242798,1.5549999475479126 C2.0799999237060547,1.5549999475479126 1.9299999475479126,1.4709999561309814 1.8700000047683716,1.3049999475479126 C1.7999999523162842,1.13100004196167 1.7899999618530273,0.9079999923706055 1.840000033378601,0.6349999904632568 C1.840000033378601,0.6349999904632568 2.319999933242798,-2.125 2.319999933242798,-2.125 C2.319999933242798,-2.125 1.440000057220459,-2.125 1.440000057220459,-2.125 C1.440000057220459,-2.125 1.3200000524520874,-1.465000033378601 1.3200000524520874,-1.465000033378601 C1.1200000047683716,-1.9789999723434448 0.6899999976158142,-2.234999895095825 0.05000000074505806,-2.234999895095825 C-0.3100000023841858,-2.234999895095825 -0.6299999952316284,-2.1589999198913574 -0.9200000166893005,-2.005000114440918 C-1.2100000381469727,-1.8519999980926514 -1.4600000381469727,-1.6419999599456787 -1.6699999570846558,-1.375 C-1.8899999856948853,-1.1150000095367432 -2.049999952316284,-0.8149999976158142 -2.1600000858306885,-0.4749999940395355 C-2.2799999713897705,-0.1420000046491623 -2.3299999237060547,0.20800000429153442 -2.3299999237060547,0.574999988079071 C-2.3299999237060547,1.0950000286102295 -2.190000057220459,1.5080000162124634 -1.909999966621399,1.815000057220459 C-1.6299999952316284,2.115000009536743 -1.2300000190734863,2.265000104904175 -0.7300000190734863,2.265000104904175 C-0.3799999952316284,2.265000104904175 -0.05000000074505806,2.184999942779541 0.25999999046325684,2.0250000953674316 C0.5699999928474426,1.8650000095367432 0.8299999833106995,1.6410000324249268 1.0299999713897705,1.3550000190734863 C1.0700000524520874,1.6349999904632568 1.1799999475479126,1.8580000400543213 1.3700000047683716,2.0250000953674316 C1.559999942779541,2.184999942779541 1.8300000429153442,2.265000104904175 2.1700000762939453,2.265000104904175 C2.5299999713897705,2.265000104904175 2.8399999141693115,2.177999973297119 3.119999885559082,2.005000114440918 C3.4000000953674316,1.8250000476837158 3.630000114440918,1.5880000591278076 3.819999933242798,1.2949999570846558 C4.010000228881836,0.9950000047683716 4.159999847412109,0.6579999923706055 4.260000228881836,0.2849999964237213 C4.360000133514404,-0.08900000154972076 4.409999847412109,-0.47200000286102295 4.409999847412109,-0.8650000095367432 C4.409999847412109,-1.531999945640564 4.25,-2.121999979019165 3.950000047683716,-2.634999990463257 C3.6500000953674316,-3.1489999294281006 3.2200000286102295,-3.5490000247955322 2.6700000762939453,-3.8350000381469727z M0.8600000143051147,0.675000011920929 C0.7200000286102295,0.9210000038146973 0.5400000214576721,1.1180000305175781 0.3199999928474426,1.2649999856948853 C0.10000000149011612,1.4049999713897705 -0.14000000059604645,1.475000023841858 -0.4000000059604645,1.475000023841858 C-0.7300000190734863,1.475000023841858 -0.9800000190734863,1.378000020980835 -1.1399999856948853,1.184999942779541 C-1.2999999523162842,0.9850000143051147 -1.3799999952316284,0.7250000238418579 -1.3799999952316284,0.4050000011920929 C-1.3799999952316284,0.09099999815225601 -1.3200000524520874,-0.20499999821186066 -1.2000000476837158,-0.48500001430511475 C-1.0800000429153442,-0.7649999856948853 -0.9100000262260437,-0.9950000047683716 -0.6800000071525574,-1.1749999523162842 C-0.46000000834465027,-1.3550000190734863 -0.1899999976158142,-1.4450000524520874 0.11999999731779099,-1.4450000524520874 C0.47999998927116394,-1.4450000524520874 0.7400000095367432,-1.3220000267028809 0.9100000262260437,-1.0750000476837158 C1.0700000524520874,-0.8289999961853027 1.1399999856948853,-0.5320000052452087 1.1100000143051147,-0.1850000023841858 C1.0800000429153442,0.13500000536441803 1,0.42100000381469727 0.8600000143051147,0.675000011920929z"></path></g><g opacity="1" transform="matrix(1,0,0,1,904.9400024414062,261.74700927734375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.5299999713897705,3.5999999046325684 C-0.5299999713897705,3.5999999046325684 0.5299999713897705,3.5999999046325684 0.5299999713897705,3.5999999046325684 C0.5299999713897705,3.5999999046325684 0.5299999713897705,-3.5999999046325684 0.5299999713897705,-3.5999999046325684 C0.5299999713897705,-3.5999999046325684 -0.5299999713897705,-3.5999999046325684 -0.5299999713897705,-3.5999999046325684 C-0.5299999713897705,-3.5999999046325684 -0.5299999713897705,3.5999999046325684 -0.5299999713897705,3.5999999046325684z"></path></g><g opacity="1" transform="matrix(1,0,0,1,893.4550170898438,261.74700927734375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0.41499999165534973,-1.4800000190734863 C0.054999999701976776,-1.4800000190734863 -0.2750000059604645,-1.3969999551773071 -0.574999988079071,-1.2300000190734863 C-0.875,-1.0700000524520874 -1.1050000190734863,-0.847000002861023 -1.2649999856948853,-0.5600000023841858 C-1.2649999856948853,-0.5600000023841858 -1.2649999856948853,-3.5999999046325684 -1.2649999856948853,-3.5999999046325684 C-1.2649999856948853,-3.5999999046325684 -2.325000047683716,-3.5999999046325684 -2.325000047683716,-3.5999999046325684 C-2.325000047683716,-3.5999999046325684 -2.325000047683716,3.5999999046325684 -2.325000047683716,3.5999999046325684 C-2.325000047683716,3.5999999046325684 -1.2649999856948853,3.5999999046325684 -1.2649999856948853,3.5999999046325684 C-1.2649999856948853,3.5999999046325684 -1.2649999856948853,1.0399999618530273 -1.2649999856948853,1.0399999618530273 C-1.2649999856948853,0.5199999809265137 -1.1349999904632568,0.11999999731779099 -0.875,-0.1599999964237213 C-0.6150000095367432,-0.4399999976158142 -0.2750000059604645,-0.5799999833106995 0.125,-0.5799999833106995 C0.8849999904632568,-0.5799999833106995 1.274999976158142,-0.10000000149011612 1.274999976158142,0.8600000143051147 C1.274999976158142,0.8600000143051147 1.274999976158142,3.5999999046325684 1.274999976158142,3.5999999046325684 C1.274999976158142,3.5999999046325684 2.325000047683716,3.5999999046325684 2.325000047683716,3.5999999046325684 C2.325000047683716,3.5999999046325684 2.325000047683716,0.75 2.325000047683716,0.75 C2.325000047683716,0.009999999776482582 2.1549999713897705,-0.546999990940094 1.815000057220459,-0.9200000166893005 C1.475000023841858,-1.2940000295639038 1.0049999952316284,-1.4800000190734863 0.41499999165534973,-1.4800000190734863z"></path></g><g opacity="1" transform="matrix(1,0,0,1,902.8200073242188,261.74700927734375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.5299999713897705,3.5999999046325684 C-0.5299999713897705,3.5999999046325684 0.5299999713897705,3.5999999046325684 0.5299999713897705,3.5999999046325684 C0.5299999713897705,3.5999999046325684 0.5299999713897705,-3.5999999046325684 0.5299999713897705,-3.5999999046325684 C0.5299999713897705,-3.5999999046325684 -0.5299999713897705,-3.5999999046325684 -0.5299999713897705,-3.5999999046325684 C-0.5299999713897705,-3.5999999046325684 -0.5299999713897705,3.5999999046325684 -0.5299999713897705,3.5999999046325684z"></path></g><g opacity="1" transform="matrix(1,0,0,1,899.02001953125,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.3200000524520874,-2.2699999809265137 C0.9599999785423279,-2.490000009536743 0.5400000214576721,-2.5999999046325684 0.05000000074505806,-2.5999999046325684 C-0.44999998807907104,-2.5999999046325684 -0.8899999856948853,-2.490000009536743 -1.2699999809265137,-2.2699999809265137 C-1.6399999856948853,-2.049999952316284 -1.940000057220459,-1.74399995803833 -2.1500000953674316,-1.350000023841858 C-2.359999895095825,-0.9570000171661377 -2.4600000381469727,-0.5 -2.4600000381469727,0.019999999552965164 C-2.4600000381469727,0.5329999923706055 -2.3499999046325684,0.9829999804496765 -2.140000104904175,1.3700000047683716 C-1.9299999475479126,1.75600004196167 -1.6299999952316284,2.059999942779541 -1.2599999904632568,2.2799999713897705 C-0.8799999952316284,2.493000030517578 -0.44999998807907104,2.5999999046325684 0.03999999910593033,2.5999999046325684 C0.6299999952316284,2.5999999046325684 1.1200000047683716,2.443000078201294 1.5099999904632568,2.130000114440918 C1.909999966621399,1.815999984741211 2.180000066757202,1.4259999990463257 2.309999942779541,0.9599999785423279 C2.309999942779541,0.9599999785423279 1.25,0.9599999785423279 1.25,0.9599999785423279 C1.149999976158142,1.1929999589920044 1,1.3799999952316284 0.7900000214576721,1.5199999809265137 C0.5899999737739563,1.652999997138977 0.3400000035762787,1.7200000286102295 0.03999999910593033,1.7200000286102295 C-0.33000001311302185,1.7200000286102295 -0.6600000262260437,1.5959999561309814 -0.949999988079071,1.350000023841858 C-1.2300000190734863,1.0959999561309814 -1.3899999856948853,0.7300000190734863 -1.4199999570846558,0.25 C-1.4199999570846558,0.25 2.440000057220459,0.25 2.440000057220459,0.25 C2.450000047683716,0.15600000321865082 2.450000047683716,0.0729999989271164 2.450000047683716,0 C2.4600000381469727,-0.07999999821186066 2.4600000381469727,-0.15399999916553497 2.4600000381469727,-0.2199999988079071 C2.4600000381469727,-0.6539999842643738 2.359999895095825,-1.0499999523162842 2.1600000858306885,-1.409999966621399 C1.9600000381469727,-1.7699999809265137 1.6799999475479126,-2.056999921798706 1.3200000524520874,-2.2699999809265137z M-1.399999976158142,-0.4699999988079071 C-1.340000033378601,-0.8899999856948853 -1.1699999570846558,-1.2039999961853027 -0.8999999761581421,-1.409999966621399 C-0.6299999952316284,-1.6239999532699585 -0.3100000023841858,-1.7300000190734863 0.03999999910593033,-1.7300000190734863 C0.41999998688697815,-1.7300000190734863 0.7400000095367432,-1.6169999837875366 0.9900000095367432,-1.3899999856948853 C1.2400000095367432,-1.1640000343322754 1.3799999952316284,-0.8569999933242798 1.399999976158142,-0.4699999988079071 C1.399999976158142,-0.4699999988079071 -1.399999976158142,-0.4699999988079071 -1.399999976158142,-0.4699999988079071z"></path></g><g opacity="1" transform="matrix(1,0,0,1,908.8599853515625,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.2699999809265137,-2.2699999809265137 C0.8899999856948853,-2.490000009536743 0.4699999988079071,-2.5999999046325684 0,-2.5999999046325684 C-0.4699999988079071,-2.5999999046325684 -0.8899999856948853,-2.490000009536743 -1.2699999809265137,-2.2699999809265137 C-1.649999976158142,-2.056999921798706 -1.9600000381469727,-1.753999948501587 -2.180000066757202,-1.3600000143051147 C-2.4000000953674316,-0.9739999771118164 -2.509999990463257,-0.5199999809265137 -2.509999990463257,0 C-2.509999990463257,0.5199999809265137 -2.4000000953674316,0.9760000109672546 -2.180000066757202,1.3700000047683716 C-1.9600000381469727,1.75600004196167 -1.6699999570846558,2.059999942779541 -1.2899999618530273,2.2799999713897705 C-0.9100000262260437,2.493000030517578 -0.49000000953674316,2.5999999046325684 -0.009999999776482582,2.5999999046325684 C0.46000000834465027,2.5999999046325684 0.8899999856948853,2.493000030517578 1.2699999809265137,2.2799999713897705 C1.649999976158142,2.059999942779541 1.9500000476837158,1.75600004196167 2.1700000762939453,1.3700000047683716 C2.390000104904175,0.9760000109672546 2.509999990463257,0.5199999809265137 2.509999990463257,0 C2.509999990463257,-0.5199999809265137 2.390000104904175,-0.9739999771118164 2.1700000762939453,-1.3600000143051147 C1.9500000476837158,-1.753999948501587 1.649999976158142,-2.056999921798706 1.2699999809265137,-2.2699999809265137z M1.2200000286102295,0.9399999976158142 C1.0800000429153442,1.1859999895095825 0.9100000262260437,1.3730000257492065 0.6899999976158142,1.5 C0.4699999988079071,1.6260000467300415 0.23999999463558197,1.690000057220459 -0.009999999776482582,1.690000057220459 C-0.27000001072883606,1.690000057220459 -0.5,1.6260000467300415 -0.7099999785423279,1.5 C-0.9300000071525574,1.3730000257492065 -1.100000023841858,1.1859999895095825 -1.2300000190734863,0.9399999976158142 C-1.3700000047683716,0.6859999895095825 -1.4299999475479126,0.37299999594688416 -1.4299999475479126,0 C-1.4299999475479126,-0.37400001287460327 -1.3700000047683716,-0.6840000152587891 -1.2300000190734863,-0.9300000071525574 C-1.100000023841858,-1.184000015258789 -0.9300000071525574,-1.3739999532699585 -0.7099999785423279,-1.5 C-0.49000000953674316,-1.6269999742507935 -0.25999999046325684,-1.690000057220459 0,-1.690000057220459 C0.25,-1.690000057220459 0.47999998927116394,-1.6269999742507935 0.699999988079071,-1.5 C0.9200000166893005,-1.3739999532699585 1.090000033378601,-1.184000015258789 1.2200000286102295,-0.9300000071525574 C1.350000023841858,-0.6840000152587891 1.4199999570846558,-0.37400001287460327 1.4199999570846558,0 C1.4199999570846558,0.37299999594688416 1.350000023841858,0.6859999895095825 1.2200000286102295,0.9399999976158142z"></path></g><g opacity="1" transform="matrix(1,0,0,1,949.2550048828125,263.9070129394531)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.4450000524520874,-3.299999952316284 C1.0750000476837158,-3.5269999504089355 0.6549999713897705,-3.640000104904175 0.17499999701976776,-3.640000104904175 C-0.26499998569488525,-3.640000104904175 -0.625,-3.549999952316284 -0.9049999713897705,-3.369999885559082 C-1.184999942779541,-3.190000057220459 -1.4049999713897705,-2.9839999675750732 -1.565000057220459,-2.75 C-1.565000057220459,-2.75 -1.6749999523162842,-3.5199999809265137 -1.6749999523162842,-3.5199999809265137 C-1.6749999523162842,-3.5199999809265137 -2.625,-3.5199999809265137 -2.625,-3.5199999809265137 C-2.625,-3.5199999809265137 -2.625,3.640000104904175 -2.625,3.640000104904175 C-2.625,3.640000104904175 -1.565000057220459,3.640000104904175 -1.565000057220459,3.640000104904175 C-1.565000057220459,3.640000104904175 -1.565000057220459,0.6800000071525574 -1.565000057220459,0.6800000071525574 C-1.3949999809265137,0.9599999785423279 -1.1649999618530273,1.1759999990463257 -0.8650000095367432,1.3300000429153442 C-0.5649999976158142,1.4830000400543213 -0.22499999403953552,1.559999942779541 0.16500000655651093,1.559999942779541 C0.6449999809265137,1.559999942779541 1.0750000476837158,1.4500000476837158 1.4450000524520874,1.2300000190734863 C1.8049999475479126,1.003000020980835 2.0950000286102295,0.6959999799728394 2.305000066757202,0.3100000023841858 C2.515000104904175,-0.08399999886751175 2.625,-0.5299999713897705 2.625,-1.0299999713897705 C2.625,-1.5299999713897705 2.515000104904175,-1.9769999980926514 2.305000066757202,-2.369999885559082 C2.0950000286102295,-2.7639999389648438 1.8049999475479126,-3.0739998817443848 1.4450000524520874,-3.299999952316284z M1.1050000190734863,0.18000000715255737 C0.8050000071525574,0.4860000014305115 0.4350000023841858,0.6399999856948853 -0.014999999664723873,0.6399999856948853 C-0.4749999940395355,0.6399999856948853 -0.8450000286102295,0.4860000014305115 -1.1349999904632568,0.18000000715255737 C-1.4249999523162842,-0.1340000033378601 -1.565000057220459,-0.5400000214576721 -1.565000057220459,-1.0399999618530273 C-1.565000057220459,-1.534000039100647 -1.4249999523162842,-1.937000036239624 -1.1349999904632568,-2.25 C-0.8450000286102295,-2.563999891281128 -0.4749999940395355,-2.7200000286102295 -0.014999999664723873,-2.7200000286102295 C0.2849999964237213,-2.7200000286102295 0.5550000071525574,-2.6470000743865967 0.7950000166893005,-2.5 C1.024999976158142,-2.359999895095825 1.2050000429153442,-2.1640000343322754 1.3450000286102295,-1.909999966621399 C1.475000023841858,-1.656999945640564 1.5449999570846558,-1.3669999837875366 1.5449999570846558,-1.0399999618530273 C1.5449999570846558,-0.5400000214576721 1.3949999809265137,-0.1340000033378601 1.1050000190734863,0.18000000715255737z"></path></g><g opacity="1" transform="matrix(1,0,0,1,960.8250122070312,264.74200439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0.004999999888241291,-0.6650000214576721 C-0.19499999284744263,-0.6650000214576721 -0.36500000953674316,-0.5989999771118164 -0.4950000047683716,-0.4650000035762787 C-0.625,-0.33899998664855957 -0.6850000023841858,-0.18199999630451202 -0.6850000023841858,0.004999999888241291 C-0.6850000023841858,0.1850000023841858 -0.625,0.3409999907016754 -0.4950000047683716,0.4749999940395355 C-0.36500000953674316,0.6010000109672546 -0.19499999284744263,0.6650000214576721 0.004999999888241291,0.6650000214576721 C0.20499999821186066,0.6650000214576721 0.36500000953674316,0.6010000109672546 0.4950000047683716,0.4749999940395355 C0.625,0.3409999907016754 0.6850000023841858,0.1850000023841858 0.6850000023841858,0.004999999888241291 C0.6850000023841858,-0.18199999630451202 0.625,-0.33899998664855957 0.4950000047683716,-0.4650000035762787 C0.36500000953674316,-0.5989999771118164 0.20499999821186066,-0.6650000214576721 0.004999999888241291,-0.6650000214576721z"></path></g><g opacity="1" transform="matrix(1,0,0,1,964.5499877929688,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.6299999952316284,-1.4900000095367432 C-0.4099999964237213,-1.6239999532699585 -0.17000000178813934,-1.690000057220459 0.09000000357627869,-1.690000057220459 C0.4000000059604645,-1.690000057220459 0.6600000262260437,-1.6139999628067017 0.8899999856948853,-1.4600000381469727 C1.1200000047683716,-1.3070000410079956 1.2599999904632568,-1.093999981880188 1.3300000429153442,-0.8199999928474426 C1.3300000429153442,-0.8199999928474426 2.440000057220459,-0.8199999928474426 2.440000057220459,-0.8199999928474426 C2.319999933242798,-1.3669999837875366 2.049999952316284,-1.7999999523162842 1.6299999952316284,-2.119999885559082 C1.2200000286102295,-2.440000057220459 0.699999988079071,-2.5999999046325684 0.09000000357627869,-2.5999999046325684 C-0.4000000059604645,-2.5999999046325684 -0.8299999833106995,-2.490000009536743 -1.2200000286102295,-2.2699999809265137 C-1.600000023841858,-2.049999952316284 -1.899999976158142,-1.74399995803833 -2.119999885559082,-1.350000023841858 C-2.3299999237060547,-0.9639999866485596 -2.440000057220459,-0.5139999985694885 -2.440000057220459,0 C-2.440000057220459,0.5130000114440918 -2.3299999237060547,0.9660000205039978 -2.119999885559082,1.3600000143051147 C-1.899999976158142,1.746000051498413 -1.600000023841858,2.049999952316284 -1.2200000286102295,2.2699999809265137 C-0.8299999833106995,2.490000009536743 -0.4000000059604645,2.5999999046325684 0.09000000357627869,2.5999999046325684 C0.699999988079071,2.5999999046325684 1.2200000286102295,2.436000108718872 1.6399999856948853,2.109999895095825 C2.059999942779541,1.7829999923706055 2.3299999237060547,1.3530000448226929 2.440000057220459,0.8199999928474426 C2.440000057220459,0.8199999928474426 1.3300000429153442,0.8199999928474426 1.3300000429153442,0.8199999928474426 C1.2599999904632568,1.100000023841858 1.1200000047683716,1.315999984741211 0.8899999856948853,1.4700000286102295 C0.6600000262260437,1.6230000257492065 0.4000000059604645,1.7000000476837158 0.09000000357627869,1.7000000476837158 C-0.17000000178813934,1.7000000476837158 -0.4099999964237213,1.6330000162124634 -0.6299999952316284,1.5 C-0.8500000238418579,1.3660000562667847 -1.0299999713897705,1.1759999990463257 -1.159999966621399,0.9300000071525574 C-1.2899999618530273,0.6759999990463257 -1.3600000143051147,0.3659999966621399 -1.3600000143051147,0 C-1.3600000143051147,-0.367000013589859 -1.2899999618530273,-0.6740000247955322 -1.159999966621399,-0.9200000166893005 C-1.0299999713897705,-1.1740000247955322 -0.8500000238418579,-1.3639999628067017 -0.6299999952316284,-1.4900000095367432z"></path></g><g opacity="1" transform="matrix(1,0,0,1,924.52001953125,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.3200000524520874,-2.2699999809265137 C0.9599999785423279,-2.490000009536743 0.5400000214576721,-2.5999999046325684 0.05000000074505806,-2.5999999046325684 C-0.44999998807907104,-2.5999999046325684 -0.8899999856948853,-2.490000009536743 -1.2699999809265137,-2.2699999809265137 C-1.6399999856948853,-2.049999952316284 -1.9299999475479126,-1.74399995803833 -2.1500000953674316,-1.350000023841858 C-2.3499999046325684,-0.9570000171661377 -2.4600000381469727,-0.5 -2.4600000381469727,0.019999999552965164 C-2.4600000381469727,0.5329999923706055 -2.3499999046325684,0.9829999804496765 -2.140000104904175,1.3700000047683716 C-1.9199999570846558,1.75600004196167 -1.6299999952316284,2.059999942779541 -1.2599999904632568,2.2799999713897705 C-0.8799999952316284,2.493000030517578 -0.4399999976158142,2.5999999046325684 0.03999999910593033,2.5999999046325684 C0.6299999952316284,2.5999999046325684 1.1200000047683716,2.443000078201294 1.5099999904632568,2.130000114440918 C1.909999966621399,1.815999984741211 2.180000066757202,1.4259999990463257 2.309999942779541,0.9599999785423279 C2.309999942779541,0.9599999785423279 1.25,0.9599999785423279 1.25,0.9599999785423279 C1.149999976158142,1.1929999589920044 1,1.3799999952316284 0.7900000214576721,1.5199999809265137 C0.5899999737739563,1.652999997138977 0.3400000035762787,1.7200000286102295 0.03999999910593033,1.7200000286102295 C-0.33000001311302185,1.7200000286102295 -0.6600000262260437,1.5959999561309814 -0.949999988079071,1.350000023841858 C-1.2300000190734863,1.0959999561309814 -1.3799999952316284,0.7300000190734863 -1.4199999570846558,0.25 C-1.4199999570846558,0.25 2.440000057220459,0.25 2.440000057220459,0.25 C2.450000047683716,0.15600000321865082 2.450000047683716,0.0729999989271164 2.450000047683716,0 C2.4600000381469727,-0.07999999821186066 2.4600000381469727,-0.15399999916553497 2.4600000381469727,-0.2199999988079071 C2.4600000381469727,-0.6539999842643738 2.359999895095825,-1.0499999523162842 2.1600000858306885,-1.409999966621399 C1.9600000381469727,-1.7699999809265137 1.6799999475479126,-2.056999921798706 1.3200000524520874,-2.2699999809265137z M-1.399999976158142,-0.4699999988079071 C-1.340000033378601,-0.8899999856948853 -1.1699999570846558,-1.2039999961853027 -0.8999999761581421,-1.409999966621399 C-0.6200000047683716,-1.6239999532699585 -0.3100000023841858,-1.7300000190734863 0.03999999910593033,-1.7300000190734863 C0.41999998688697815,-1.7300000190734863 0.7400000095367432,-1.6169999837875366 0.9900000095367432,-1.3899999856948853 C1.25,-1.1640000343322754 1.3799999952316284,-0.8569999933242798 1.399999976158142,-0.4699999988079071 C1.399999976158142,-0.4699999988079071 -1.399999976158142,-0.4699999988079071 -1.399999976158142,-0.4699999988079071z"></path></g><g opacity="1" transform="matrix(1,0,0,1,957.1300048828125,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.3200000524520874,-2.2699999809265137 C0.9599999785423279,-2.490000009536743 0.5400000214576721,-2.5999999046325684 0.05000000074505806,-2.5999999046325684 C-0.44999998807907104,-2.5999999046325684 -0.8899999856948853,-2.490000009536743 -1.2699999809265137,-2.2699999809265137 C-1.6399999856948853,-2.049999952316284 -1.9299999475479126,-1.74399995803833 -2.1500000953674316,-1.350000023841858 C-2.3499999046325684,-0.9570000171661377 -2.4600000381469727,-0.5 -2.4600000381469727,0.019999999552965164 C-2.4600000381469727,0.5329999923706055 -2.3499999046325684,0.9829999804496765 -2.140000104904175,1.3700000047683716 C-1.9199999570846558,1.75600004196167 -1.6299999952316284,2.059999942779541 -1.2599999904632568,2.2799999713897705 C-0.8799999952316284,2.493000030517578 -0.4399999976158142,2.5999999046325684 0.03999999910593033,2.5999999046325684 C0.6299999952316284,2.5999999046325684 1.1200000047683716,2.443000078201294 1.5099999904632568,2.130000114440918 C1.909999966621399,1.815999984741211 2.180000066757202,1.4259999990463257 2.309999942779541,0.9599999785423279 C2.309999942779541,0.9599999785423279 1.25,0.9599999785423279 1.25,0.9599999785423279 C1.149999976158142,1.1929999589920044 1,1.3799999952316284 0.7900000214576721,1.5199999809265137 C0.5899999737739563,1.652999997138977 0.3400000035762787,1.7200000286102295 0.03999999910593033,1.7200000286102295 C-0.33000001311302185,1.7200000286102295 -0.6600000262260437,1.5959999561309814 -0.949999988079071,1.350000023841858 C-1.2300000190734863,1.0959999561309814 -1.3799999952316284,0.7300000190734863 -1.4199999570846558,0.25 C-1.4199999570846558,0.25 2.440000057220459,0.25 2.440000057220459,0.25 C2.450000047683716,0.15600000321865082 2.450000047683716,0.0729999989271164 2.450000047683716,0 C2.4600000381469727,-0.07999999821186066 2.4600000381469727,-0.15399999916553497 2.4600000381469727,-0.2199999988079071 C2.4600000381469727,-0.6539999842643738 2.359999895095825,-1.0499999523162842 2.1600000858306885,-1.409999966621399 C1.9600000381469727,-1.7699999809265137 1.6799999475479126,-2.056999921798706 1.3200000524520874,-2.2699999809265137z M-1.399999976158142,-0.4699999988079071 C-1.340000033378601,-0.8899999856948853 -1.1699999570846558,-1.2039999961853027 -0.8999999761581421,-1.409999966621399 C-0.6200000047683716,-1.6239999532699585 -0.3100000023841858,-1.7300000190734863 0.03999999910593033,-1.7300000190734863 C0.41999998688697815,-1.7300000190734863 0.7400000095367432,-1.6169999837875366 0.9900000095367432,-1.3899999856948853 C1.25,-1.1640000343322754 1.3799999952316284,-0.8569999933242798 1.399999976158142,-0.4699999988079071 C1.399999976158142,-0.4699999988079071 -1.399999976158142,-0.4699999988079071 -1.399999976158142,-0.4699999988079071z"></path></g><g opacity="1" transform="matrix(1,0,0,1,970.1799926757812,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.2699999809265137,-2.2699999809265137 C0.8999999761581421,-2.490000009536743 0.47999998927116394,-2.5999999046325684 0,-2.5999999046325684 C-0.46000000834465027,-2.5999999046325684 -0.8899999856948853,-2.490000009536743 -1.2699999809265137,-2.2699999809265137 C-1.649999976158142,-2.056999921798706 -1.9500000476837158,-1.753999948501587 -2.180000066757202,-1.3600000143051147 C-2.4000000953674316,-0.9739999771118164 -2.509999990463257,-0.5199999809265137 -2.509999990463257,0 C-2.509999990463257,0.5199999809265137 -2.4000000953674316,0.9760000109672546 -2.180000066757202,1.3700000047683716 C-1.9600000381469727,1.75600004196167 -1.659999966621399,2.059999942779541 -1.2899999618530273,2.2799999713897705 C-0.9100000262260437,2.493000030517578 -0.47999998927116394,2.5999999046325684 -0.009999999776482582,2.5999999046325684 C0.4699999988079071,2.5999999046325684 0.8899999856948853,2.493000030517578 1.2699999809265137,2.2799999713897705 C1.649999976158142,2.059999942779541 1.9500000476837158,1.75600004196167 2.1700000762939453,1.3700000047683716 C2.4000000953674316,0.9760000109672546 2.509999990463257,0.5199999809265137 2.509999990463257,0 C2.509999990463257,-0.5199999809265137 2.4000000953674316,-0.9739999771118164 2.1700000762939453,-1.3600000143051147 C1.9500000476837158,-1.753999948501587 1.649999976158142,-2.056999921798706 1.2699999809265137,-2.2699999809265137z M1.2200000286102295,0.9399999976158142 C1.090000033378601,1.1859999895095825 0.9100000262260437,1.3730000257492065 0.6899999976158142,1.5 C0.47999998927116394,1.6260000467300415 0.25,1.690000057220459 -0.009999999776482582,1.690000057220459 C-0.25999999046325684,1.690000057220459 -0.49000000953674316,1.6260000467300415 -0.7099999785423279,1.5 C-0.9200000166893005,1.3730000257492065 -1.090000033378601,1.1859999895095825 -1.2300000190734863,0.9399999976158142 C-1.3600000143051147,0.6859999895095825 -1.4299999475479126,0.37299999594688416 -1.4299999475479126,0 C-1.4299999475479126,-0.37400001287460327 -1.3600000143051147,-0.6840000152587891 -1.2300000190734863,-0.9300000071525574 C-1.090000033378601,-1.184000015258789 -0.9200000166893005,-1.3739999532699585 -0.7099999785423279,-1.5 C-0.49000000953674316,-1.6269999742507935 -0.25,-1.690000057220459 0,-1.690000057220459 C0.25999999046325684,-1.690000057220459 0.49000000953674316,-1.6269999742507935 0.699999988079071,-1.5 C0.9200000166893005,-1.3739999532699585 1.100000023841858,-1.184000015258789 1.2200000286102295,-0.9300000071525574 C1.3600000143051147,-0.6840000152587891 1.4199999570846558,-0.37400001287460327 1.4199999570846558,0 C1.4199999570846558,0.37299999594688416 1.3600000143051147,0.6859999895095825 1.2200000286102295,0.9399999976158142z"></path></g><g opacity="1" transform="matrix(1,0,0,1,977.52001953125,262.8070068359375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M3.430000066757202,-1.9800000190734863 C3.069999933242798,-2.3540000915527344 2.5899999141693115,-2.5399999618530273 1.9700000286102295,-2.5399999618530273 C1.6100000143051147,-2.5399999618530273 1.2799999713897705,-2.4570000171661377 0.9800000190734863,-2.2899999618530273 C0.6800000071525574,-2.124000072479248 0.4399999976158142,-1.8969999551773071 0.25999999046325684,-1.6100000143051147 C-0.05000000074505806,-2.2300000190734863 -0.6000000238418579,-2.5399999618530273 -1.3799999952316284,-2.5399999618530273 C-1.7300000190734863,-2.5399999618530273 -2.0399999618530273,-2.4670000076293945 -2.299999952316284,-2.319999933242798 C-2.559999942779541,-2.1740000247955322 -2.7699999809265137,-1.9739999771118164 -2.930000066757202,-1.7200000286102295 C-2.930000066757202,-1.7200000286102295 -3.0199999809265137,-2.4200000762939453 -3.0199999809265137,-2.4200000762939453 C-3.0199999809265137,-2.4200000762939453 -3.9600000381469727,-2.4200000762939453 -3.9600000381469727,-2.4200000762939453 C-3.9600000381469727,-2.4200000762939453 -3.9600000381469727,2.5399999618530273 -3.9600000381469727,2.5399999618530273 C-3.9600000381469727,2.5399999618530273 -2.9000000953674316,2.5399999618530273 -2.9000000953674316,2.5399999618530273 C-2.9000000953674316,2.5399999618530273 -2.9000000953674316,-0.019999999552965164 -2.9000000953674316,-0.019999999552965164 C-2.9000000953674316,-0.5400000214576721 -2.7899999618530273,-0.9399999976158142 -2.549999952316284,-1.2200000286102295 C-2.309999942779541,-1.5 -2.009999990463257,-1.6399999856948853 -1.6399999856948853,-1.6399999856948853 C-0.8999999761581421,-1.6399999856948853 -0.5299999713897705,-1.159999966621399 -0.5299999713897705,-0.20000000298023224 C-0.5299999713897705,-0.20000000298023224 -0.5299999713897705,2.5399999618530273 -0.5299999713897705,2.5399999618530273 C-0.5299999713897705,2.5399999618530273 0.5299999713897705,2.5399999618530273 0.5299999713897705,2.5399999618530273 C0.5299999713897705,2.5399999618530273 0.5299999713897705,-0.019999999552965164 0.5299999713897705,-0.019999999552965164 C0.5299999713897705,-0.5400000214576721 0.6399999856948853,-0.9399999976158142 0.8799999952316284,-1.2200000286102295 C1.1200000047683716,-1.5 1.4199999570846558,-1.6399999856948853 1.7999999523162842,-1.6399999856948853 C2.5299999713897705,-1.6399999856948853 2.9000000953674316,-1.159999966621399 2.9000000953674316,-0.20000000298023224 C2.9000000953674316,-0.20000000298023224 2.9000000953674316,2.5399999618530273 2.9000000953674316,2.5399999618530273 C2.9000000953674316,2.5399999618530273 3.9600000381469727,2.5399999618530273 3.9600000381469727,2.5399999618530273 C3.9600000381469727,2.5399999618530273 3.9600000381469727,-0.3100000023841858 3.9600000381469727,-0.3100000023841858 C3.9600000381469727,-1.0499999523162842 3.7799999713897705,-1.6069999933242798 3.430000066757202,-1.9800000190734863z"></path></g><g opacity="1" transform="matrix(1,0,0,1,934.5399780273438,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0.07999999821186066,-2.5999999046325684 C-0.30000001192092896,-2.5999999046325684 -0.6600000262260437,-2.5299999713897705 -0.9900000095367432,-2.390000104904175 C-1.309999942779541,-2.256999969482422 -1.5700000524520874,-2.059999942779541 -1.7899999618530273,-1.7999999523162842 C-1.9900000095367432,-1.5470000505447388 -2.109999895095825,-1.2400000095367432 -2.1500000953674316,-0.8799999952316284 C-2.1500000953674316,-0.8799999952316284 -1.090000033378601,-0.8799999952316284 -1.090000033378601,-0.8799999952316284 C-1.0399999618530273,-1.159999966621399 -0.9100000262260437,-1.3700000047683716 -0.699999988079071,-1.5099999904632568 C-0.47999998927116394,-1.656999945640564 -0.2199999988079071,-1.7300000190734863 0.07000000029802322,-1.7300000190734863 C0.4099999964237213,-1.7300000190734863 0.6800000071525574,-1.6339999437332153 0.8799999952316284,-1.440000057220459 C1.100000023841858,-1.246999979019165 1.2000000476837158,-0.9570000171661377 1.2000000476837158,-0.5699999928474426 C1.2000000476837158,-0.5699999928474426 1.2000000476837158,-0.47999998927116394 1.2000000476837158,-0.47999998927116394 C1.2000000476837158,-0.47999998927116394 -0.10000000149011612,-0.47999998927116394 -0.10000000149011612,-0.47999998927116394 C-0.7900000214576721,-0.47999998927116394 -1.3200000524520874,-0.3400000035762787 -1.7000000476837158,-0.05999999865889549 C-2.069999933242798,0.2199999988079071 -2.259999990463257,0.6000000238418579 -2.259999990463257,1.0800000429153442 C-2.259999990463257,1.3530000448226929 -2.190000057220459,1.6059999465942383 -2.059999942779541,1.840000033378601 C-1.9199999570846558,2.065999984741211 -1.7200000286102295,2.25 -1.4500000476837158,2.390000104904175 C-1.1699999570846558,2.5299999713897705 -0.8299999833106995,2.5999999046325684 -0.4099999964237213,2.5999999046325684 C0.029999999329447746,2.5999999046325684 0.3799999952316284,2.50600004196167 0.6399999856948853,2.319999933242798 C0.9100000262260437,2.132999897003174 1.1100000143051147,1.902999997138977 1.25,1.6299999952316284 C1.25,1.6299999952316284 1.340000033378601,2.4800000190734863 1.340000033378601,2.4800000190734863 C1.340000033378601,2.4800000190734863 2.259999990463257,2.4800000190734863 2.259999990463257,2.4800000190734863 C2.259999990463257,2.4800000190734863 2.259999990463257,-0.5699999928474426 2.259999990463257,-0.5699999928474426 C2.259999990463257,-1.1970000267028809 2.069999933242798,-1.690000057220459 1.690000057220459,-2.049999952316284 C1.309999942779541,-2.4170000553131104 0.7799999713897705,-2.5999999046325684 0.07999999821186066,-2.5999999046325684z M0.9800000190734863,0.9900000095367432 C0.8799999952316284,1.215999960899353 0.7200000286102295,1.399999976158142 0.5199999809265137,1.5399999618530273 C0.33000001311302185,1.6729999780654907 0.09000000357627869,1.7400000095367432 -0.20000000298023224,1.7400000095367432 C-0.49000000953674316,1.7400000095367432 -0.7200000286102295,1.6759999990463257 -0.8999999761581421,1.5499999523162842 C-1.059999942779541,1.4160000085830688 -1.149999976158142,1.2359999418258667 -1.149999976158142,1.0099999904632568 C-1.149999976158142,0.7900000214576721 -1.059999942779541,0.6060000061988831 -0.8899999856948853,0.46000000834465027 C-0.7099999785423279,0.31299999356269836 -0.4099999964237213,0.23999999463558197 0.009999999776482582,0.23999999463558197 C0.009999999776482582,0.23999999463558197 1.190000057220459,0.23999999463558197 1.190000057220459,0.23999999463558197 C1.1699999570846558,0.5130000114440918 1.100000023841858,0.7630000114440918 0.9800000190734863,0.9900000095367432z"></path></g><g opacity="1" transform="matrix(1,0,0,1,929.47998046875,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.3200000524520874,-2.4800000190734863 C1.3200000524520874,-2.4800000190734863 0,-0.5799999833106995 0,-0.5799999833106995 C0,-0.5799999833106995 -1.3300000429153442,-2.4800000190734863 -1.3300000429153442,-2.4800000190734863 C-1.3300000429153442,-2.4800000190734863 -2.4600000381469727,-2.4800000190734863 -2.4600000381469727,-2.4800000190734863 C-2.4600000381469727,-2.4800000190734863 -0.699999988079071,0 -0.699999988079071,0 C-0.699999988079071,0 -2.4600000381469727,2.4800000190734863 -2.4600000381469727,2.4800000190734863 C-2.4600000381469727,2.4800000190734863 -1.3300000429153442,2.4800000190734863 -1.3300000429153442,2.4800000190734863 C-1.3300000429153442,2.4800000190734863 0,0.5799999833106995 0,0.5799999833106995 C0,0.5799999833106995 1.3200000524520874,2.4800000190734863 1.3200000524520874,2.4800000190734863 C1.3200000524520874,2.4800000190734863 2.4600000381469727,2.4800000190734863 2.4600000381469727,2.4800000190734863 C2.4600000381469727,2.4800000190734863 0.699999988079071,0 0.699999988079071,0 C0.699999988079071,0 2.4600000381469727,-2.4800000190734863 2.4600000381469727,-2.4800000190734863 C2.4600000381469727,-2.4800000190734863 1.3200000524520874,-2.4800000190734863 1.3200000524520874,-2.4800000190734863z"></path></g><g opacity="1" transform="matrix(1,0,0,1,941.7100219726562,262.8070068359375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.9700000286102295,-2.5399999618530273 C1.6200000047683716,-2.5399999618530273 1.2899999618530273,-2.4570000171661377 0.9800000190734863,-2.2899999618530273 C0.6800000071525574,-2.124000072479248 0.4399999976158142,-1.8969999551773071 0.25999999046325684,-1.6100000143051147 C-0.03999999910593033,-2.2300000190734863 -0.5899999737739563,-2.5399999618530273 -1.3799999952316284,-2.5399999618530273 C-1.7200000286102295,-2.5399999618530273 -2.0299999713897705,-2.4670000076293945 -2.299999952316284,-2.319999933242798 C-2.559999942779541,-2.1740000247955322 -2.7699999809265137,-1.9739999771118164 -2.930000066757202,-1.7200000286102295 C-2.930000066757202,-1.7200000286102295 -3.0199999809265137,-2.4200000762939453 -3.0199999809265137,-2.4200000762939453 C-3.0199999809265137,-2.4200000762939453 -3.9600000381469727,-2.4200000762939453 -3.9600000381469727,-2.4200000762939453 C-3.9600000381469727,-2.4200000762939453 -3.9600000381469727,2.5399999618530273 -3.9600000381469727,2.5399999618530273 C-3.9600000381469727,2.5399999618530273 -2.9000000953674316,2.5399999618530273 -2.9000000953674316,2.5399999618530273 C-2.9000000953674316,2.5399999618530273 -2.9000000953674316,-0.019999999552965164 -2.9000000953674316,-0.019999999552965164 C-2.9000000953674316,-0.5400000214576721 -2.7799999713897705,-0.9399999976158142 -2.549999952316284,-1.2200000286102295 C-2.309999942779541,-1.5 -2,-1.6399999856948853 -1.6399999856948853,-1.6399999856948853 C-0.8999999761581421,-1.6399999856948853 -0.5299999713897705,-1.159999966621399 -0.5299999713897705,-0.20000000298023224 C-0.5299999713897705,-0.20000000298023224 -0.5299999713897705,2.5399999618530273 -0.5299999713897705,2.5399999618530273 C-0.5299999713897705,2.5399999618530273 0.5299999713897705,2.5399999618530273 0.5299999713897705,2.5399999618530273 C0.5299999713897705,2.5399999618530273 0.5299999713897705,-0.019999999552965164 0.5299999713897705,-0.019999999552965164 C0.5299999713897705,-0.5400000214576721 0.6499999761581421,-0.9399999976158142 0.8799999952316284,-1.2200000286102295 C1.1200000047683716,-1.5 1.4299999475479126,-1.6399999856948853 1.7999999523162842,-1.6399999856948853 C2.5399999618530273,-1.6399999856948853 2.9000000953674316,-1.159999966621399 2.9000000953674316,-0.20000000298023224 C2.9000000953674316,-0.20000000298023224 2.9000000953674316,2.5399999618530273 2.9000000953674316,2.5399999618530273 C2.9000000953674316,2.5399999618530273 3.9600000381469727,2.5399999618530273 3.9600000381469727,2.5399999618530273 C3.9600000381469727,2.5399999618530273 3.9600000381469727,-0.3100000023841858 3.9600000381469727,-0.3100000023841858 C3.9600000381469727,-1.0499999523162842 3.7899999618530273,-1.6069999933242798 3.430000066757202,-1.9800000190734863 C3.0799999237060547,-2.3540000915527344 2.5899999141693115,-2.5399999618530273 1.9700000286102295,-2.5399999618530273z"></path></g><g opacity="1" transform="matrix(1,0,0,1,953.27001953125,261.74700927734375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.5299999713897705,3.5999999046325684 C-0.5299999713897705,3.5999999046325684 0.5299999713897705,3.5999999046325684 0.5299999713897705,3.5999999046325684 C0.5299999713897705,3.5999999046325684 0.5299999713897705,-3.5999999046325684 0.5299999713897705,-3.5999999046325684 C0.5299999713897705,-3.5999999046325684 -0.5299999713897705,-3.5999999046325684 -0.5299999713897705,-3.5999999046325684 C-0.5299999713897705,-3.5999999046325684 -0.5299999713897705,3.5999999046325684 -0.5299999713897705,3.5999999046325684z"></path></g></g><g clip-path="url(#__lottie_element_251)" style="display: block;" transform="matrix(1.3937740325927734,0,0,1.3937740325927734,-841.506103515625,-10.683563232421875)" opacity="0.39279356565821305"><g opacity="1" transform="matrix(1,0,0,1,916.7100219726562,263.1419982910156)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M2.6700000762939453,-3.8350000381469727 C2.109999895095825,-4.122000217437744 1.4600000381469727,-4.264999866485596 0.699999988079071,-4.264999866485596 C-0.03999999910593033,-4.264999866485596 -0.7200000286102295,-4.135000228881836 -1.340000033378601,-3.875 C-1.9600000381469727,-3.621999979019165 -2.5,-3.26200008392334 -2.9600000381469727,-2.7950000762939453 C-3.4200000762939453,-2.3289999961853027 -3.7799999713897705,-1.784999966621399 -4.03000020980835,-1.1649999618530273 C-4.289999961853027,-0.5450000166893005 -4.409999847412109,0.12800000607967377 -4.409999847412109,0.8550000190734863 C-4.409999847412109,1.5479999780654907 -4.260000228881836,2.1480000019073486 -3.9600000381469727,2.6549999713897705 C-3.6600000858306885,3.1679999828338623 -3.2300000190734863,3.565000057220459 -2.690000057220459,3.8450000286102295 C-2.1500000953674316,4.125 -1.5099999904632568,4.264999866485596 -0.7799999713897705,4.264999866485596 C-0.05000000074505806,4.264999866485596 0.6200000047683716,4.135000228881836 1.2400000095367432,3.875 C1.2400000095367432,3.875 1.0700000524520874,3.174999952316284 1.0700000524520874,3.174999952316284 C0.8100000023841858,3.2880001068115234 0.5400000214576721,3.375 0.25,3.434999942779541 C-0.03999999910593033,3.494999885559082 -0.33000001311302185,3.5250000953674316 -0.6200000047683716,3.5250000953674316 C-1.5399999618530273,3.5250000953674316 -2.259999990463257,3.2809998989105225 -2.7799999713897705,2.7950000762939453 C-3.309999942779541,2.315000057220459 -3.569999933242798,1.6380000114440918 -3.569999933242798,0.7649999856948853 C-3.569999933242798,0.16500000655651093 -3.4700000286102295,-0.39500001072883606 -3.259999990463257,-0.9150000214576721 C-3.059999942779541,-1.434999942779541 -2.7699999809265137,-1.8890000581741333 -2.390000104904175,-2.2750000953674316 C-2.0199999809265137,-2.6689999103546143 -1.5800000429153442,-2.9749999046325684 -1.0700000524520874,-3.194999933242798 C-0.5699999928474426,-3.421999931335449 -0.019999999552965164,-3.5350000858306885 0.5799999833106995,-3.5350000858306885 C1.5199999809265137,-3.5350000858306885 2.25,-3.2920000553131104 2.7799999713897705,-2.805000066757202 C3.299999952316284,-2.325000047683716 3.569999933242798,-1.6619999408721924 3.569999933242798,-0.8149999976158142 C3.569999933242798,-0.5490000247955322 3.5399999618530273,-0.2750000059604645 3.490000009536743,0.004999999888241291 C3.430000066757202,0.27799999713897705 3.3499999046325684,0.531000018119812 3.25,0.7649999856948853 C3.1500000953674316,0.9980000257492065 3.0199999809265137,1.187999963760376 2.859999895095825,1.3350000381469727 C2.700000047683716,1.4809999465942383 2.5199999809265137,1.5549999475479126 2.319999933242798,1.5549999475479126 C2.0799999237060547,1.5549999475479126 1.9299999475479126,1.4709999561309814 1.8700000047683716,1.3049999475479126 C1.7999999523162842,1.13100004196167 1.7899999618530273,0.9079999923706055 1.840000033378601,0.6349999904632568 C1.840000033378601,0.6349999904632568 2.319999933242798,-2.125 2.319999933242798,-2.125 C2.319999933242798,-2.125 1.440000057220459,-2.125 1.440000057220459,-2.125 C1.440000057220459,-2.125 1.3200000524520874,-1.465000033378601 1.3200000524520874,-1.465000033378601 C1.1200000047683716,-1.9789999723434448 0.6899999976158142,-2.234999895095825 0.05000000074505806,-2.234999895095825 C-0.3100000023841858,-2.234999895095825 -0.6299999952316284,-2.1589999198913574 -0.9200000166893005,-2.005000114440918 C-1.2100000381469727,-1.8519999980926514 -1.4600000381469727,-1.6419999599456787 -1.6699999570846558,-1.375 C-1.8899999856948853,-1.1150000095367432 -2.049999952316284,-0.8149999976158142 -2.1600000858306885,-0.4749999940395355 C-2.2799999713897705,-0.1420000046491623 -2.3299999237060547,0.20800000429153442 -2.3299999237060547,0.574999988079071 C-2.3299999237060547,1.0950000286102295 -2.190000057220459,1.5080000162124634 -1.909999966621399,1.815000057220459 C-1.6299999952316284,2.115000009536743 -1.2300000190734863,2.265000104904175 -0.7300000190734863,2.265000104904175 C-0.3799999952316284,2.265000104904175 -0.05000000074505806,2.184999942779541 0.25999999046325684,2.0250000953674316 C0.5699999928474426,1.8650000095367432 0.8299999833106995,1.6410000324249268 1.0299999713897705,1.3550000190734863 C1.0700000524520874,1.6349999904632568 1.1799999475479126,1.8580000400543213 1.3700000047683716,2.0250000953674316 C1.559999942779541,2.184999942779541 1.8300000429153442,2.265000104904175 2.1700000762939453,2.265000104904175 C2.5299999713897705,2.265000104904175 2.8399999141693115,2.177999973297119 3.119999885559082,2.005000114440918 C3.4000000953674316,1.8250000476837158 3.630000114440918,1.5880000591278076 3.819999933242798,1.2949999570846558 C4.010000228881836,0.9950000047683716 4.159999847412109,0.6579999923706055 4.260000228881836,0.2849999964237213 C4.360000133514404,-0.08900000154972076 4.409999847412109,-0.47200000286102295 4.409999847412109,-0.8650000095367432 C4.409999847412109,-1.531999945640564 4.25,-2.121999979019165 3.950000047683716,-2.634999990463257 C3.6500000953674316,-3.1489999294281006 3.2200000286102295,-3.5490000247955322 2.6700000762939453,-3.8350000381469727z M0.8600000143051147,0.675000011920929 C0.7200000286102295,0.9210000038146973 0.5400000214576721,1.1180000305175781 0.3199999928474426,1.2649999856948853 C0.10000000149011612,1.4049999713897705 -0.14000000059604645,1.475000023841858 -0.4000000059604645,1.475000023841858 C-0.7300000190734863,1.475000023841858 -0.9800000190734863,1.378000020980835 -1.1399999856948853,1.184999942779541 C-1.2999999523162842,0.9850000143051147 -1.3799999952316284,0.7250000238418579 -1.3799999952316284,0.4050000011920929 C-1.3799999952316284,0.09099999815225601 -1.3200000524520874,-0.20499999821186066 -1.2000000476837158,-0.48500001430511475 C-1.0800000429153442,-0.7649999856948853 -0.9100000262260437,-0.9950000047683716 -0.6800000071525574,-1.1749999523162842 C-0.46000000834465027,-1.3550000190734863 -0.1899999976158142,-1.4450000524520874 0.11999999731779099,-1.4450000524520874 C0.47999998927116394,-1.4450000524520874 0.7400000095367432,-1.3220000267028809 0.9100000262260437,-1.0750000476837158 C1.0700000524520874,-0.8289999961853027 1.1399999856948853,-0.5320000052452087 1.1100000143051147,-0.1850000023841858 C1.0800000429153442,0.13500000536441803 1,0.42100000381469727 0.8600000143051147,0.675000011920929z"></path></g><g opacity="1" transform="matrix(1,0,0,1,904.9400024414062,261.74700927734375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.5299999713897705,3.5999999046325684 C-0.5299999713897705,3.5999999046325684 0.5299999713897705,3.5999999046325684 0.5299999713897705,3.5999999046325684 C0.5299999713897705,3.5999999046325684 0.5299999713897705,-3.5999999046325684 0.5299999713897705,-3.5999999046325684 C0.5299999713897705,-3.5999999046325684 -0.5299999713897705,-3.5999999046325684 -0.5299999713897705,-3.5999999046325684 C-0.5299999713897705,-3.5999999046325684 -0.5299999713897705,3.5999999046325684 -0.5299999713897705,3.5999999046325684z"></path></g><g opacity="1" transform="matrix(1,0,0,1,893.4550170898438,261.74700927734375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0.41499999165534973,-1.4800000190734863 C0.054999999701976776,-1.4800000190734863 -0.2750000059604645,-1.3969999551773071 -0.574999988079071,-1.2300000190734863 C-0.875,-1.0700000524520874 -1.1050000190734863,-0.847000002861023 -1.2649999856948853,-0.5600000023841858 C-1.2649999856948853,-0.5600000023841858 -1.2649999856948853,-3.5999999046325684 -1.2649999856948853,-3.5999999046325684 C-1.2649999856948853,-3.5999999046325684 -2.325000047683716,-3.5999999046325684 -2.325000047683716,-3.5999999046325684 C-2.325000047683716,-3.5999999046325684 -2.325000047683716,3.5999999046325684 -2.325000047683716,3.5999999046325684 C-2.325000047683716,3.5999999046325684 -1.2649999856948853,3.5999999046325684 -1.2649999856948853,3.5999999046325684 C-1.2649999856948853,3.5999999046325684 -1.2649999856948853,1.0399999618530273 -1.2649999856948853,1.0399999618530273 C-1.2649999856948853,0.5199999809265137 -1.1349999904632568,0.11999999731779099 -0.875,-0.1599999964237213 C-0.6150000095367432,-0.4399999976158142 -0.2750000059604645,-0.5799999833106995 0.125,-0.5799999833106995 C0.8849999904632568,-0.5799999833106995 1.274999976158142,-0.10000000149011612 1.274999976158142,0.8600000143051147 C1.274999976158142,0.8600000143051147 1.274999976158142,3.5999999046325684 1.274999976158142,3.5999999046325684 C1.274999976158142,3.5999999046325684 2.325000047683716,3.5999999046325684 2.325000047683716,3.5999999046325684 C2.325000047683716,3.5999999046325684 2.325000047683716,0.75 2.325000047683716,0.75 C2.325000047683716,0.009999999776482582 2.1549999713897705,-0.546999990940094 1.815000057220459,-0.9200000166893005 C1.475000023841858,-1.2940000295639038 1.0049999952316284,-1.4800000190734863 0.41499999165534973,-1.4800000190734863z"></path></g><g opacity="1" transform="matrix(1,0,0,1,902.8200073242188,261.74700927734375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.5299999713897705,3.5999999046325684 C-0.5299999713897705,3.5999999046325684 0.5299999713897705,3.5999999046325684 0.5299999713897705,3.5999999046325684 C0.5299999713897705,3.5999999046325684 0.5299999713897705,-3.5999999046325684 0.5299999713897705,-3.5999999046325684 C0.5299999713897705,-3.5999999046325684 -0.5299999713897705,-3.5999999046325684 -0.5299999713897705,-3.5999999046325684 C-0.5299999713897705,-3.5999999046325684 -0.5299999713897705,3.5999999046325684 -0.5299999713897705,3.5999999046325684z"></path></g><g opacity="1" transform="matrix(1,0,0,1,899.02001953125,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.3200000524520874,-2.2699999809265137 C0.9599999785423279,-2.490000009536743 0.5400000214576721,-2.5999999046325684 0.05000000074505806,-2.5999999046325684 C-0.44999998807907104,-2.5999999046325684 -0.8899999856948853,-2.490000009536743 -1.2699999809265137,-2.2699999809265137 C-1.6399999856948853,-2.049999952316284 -1.940000057220459,-1.74399995803833 -2.1500000953674316,-1.350000023841858 C-2.359999895095825,-0.9570000171661377 -2.4600000381469727,-0.5 -2.4600000381469727,0.019999999552965164 C-2.4600000381469727,0.5329999923706055 -2.3499999046325684,0.9829999804496765 -2.140000104904175,1.3700000047683716 C-1.9299999475479126,1.75600004196167 -1.6299999952316284,2.059999942779541 -1.2599999904632568,2.2799999713897705 C-0.8799999952316284,2.493000030517578 -0.44999998807907104,2.5999999046325684 0.03999999910593033,2.5999999046325684 C0.6299999952316284,2.5999999046325684 1.1200000047683716,2.443000078201294 1.5099999904632568,2.130000114440918 C1.909999966621399,1.815999984741211 2.180000066757202,1.4259999990463257 2.309999942779541,0.9599999785423279 C2.309999942779541,0.9599999785423279 1.25,0.9599999785423279 1.25,0.9599999785423279 C1.149999976158142,1.1929999589920044 1,1.3799999952316284 0.7900000214576721,1.5199999809265137 C0.5899999737739563,1.652999997138977 0.3400000035762787,1.7200000286102295 0.03999999910593033,1.7200000286102295 C-0.33000001311302185,1.7200000286102295 -0.6600000262260437,1.5959999561309814 -0.949999988079071,1.350000023841858 C-1.2300000190734863,1.0959999561309814 -1.3899999856948853,0.7300000190734863 -1.4199999570846558,0.25 C-1.4199999570846558,0.25 2.440000057220459,0.25 2.440000057220459,0.25 C2.450000047683716,0.15600000321865082 2.450000047683716,0.0729999989271164 2.450000047683716,0 C2.4600000381469727,-0.07999999821186066 2.4600000381469727,-0.15399999916553497 2.4600000381469727,-0.2199999988079071 C2.4600000381469727,-0.6539999842643738 2.359999895095825,-1.0499999523162842 2.1600000858306885,-1.409999966621399 C1.9600000381469727,-1.7699999809265137 1.6799999475479126,-2.056999921798706 1.3200000524520874,-2.2699999809265137z M-1.399999976158142,-0.4699999988079071 C-1.340000033378601,-0.8899999856948853 -1.1699999570846558,-1.2039999961853027 -0.8999999761581421,-1.409999966621399 C-0.6299999952316284,-1.6239999532699585 -0.3100000023841858,-1.7300000190734863 0.03999999910593033,-1.7300000190734863 C0.41999998688697815,-1.7300000190734863 0.7400000095367432,-1.6169999837875366 0.9900000095367432,-1.3899999856948853 C1.2400000095367432,-1.1640000343322754 1.3799999952316284,-0.8569999933242798 1.399999976158142,-0.4699999988079071 C1.399999976158142,-0.4699999988079071 -1.399999976158142,-0.4699999988079071 -1.399999976158142,-0.4699999988079071z"></path></g><g opacity="1" transform="matrix(1,0,0,1,908.8599853515625,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.2699999809265137,-2.2699999809265137 C0.8899999856948853,-2.490000009536743 0.4699999988079071,-2.5999999046325684 0,-2.5999999046325684 C-0.4699999988079071,-2.5999999046325684 -0.8899999856948853,-2.490000009536743 -1.2699999809265137,-2.2699999809265137 C-1.649999976158142,-2.056999921798706 -1.9600000381469727,-1.753999948501587 -2.180000066757202,-1.3600000143051147 C-2.4000000953674316,-0.9739999771118164 -2.509999990463257,-0.5199999809265137 -2.509999990463257,0 C-2.509999990463257,0.5199999809265137 -2.4000000953674316,0.9760000109672546 -2.180000066757202,1.3700000047683716 C-1.9600000381469727,1.75600004196167 -1.6699999570846558,2.059999942779541 -1.2899999618530273,2.2799999713897705 C-0.9100000262260437,2.493000030517578 -0.49000000953674316,2.5999999046325684 -0.009999999776482582,2.5999999046325684 C0.46000000834465027,2.5999999046325684 0.8899999856948853,2.493000030517578 1.2699999809265137,2.2799999713897705 C1.649999976158142,2.059999942779541 1.9500000476837158,1.75600004196167 2.1700000762939453,1.3700000047683716 C2.390000104904175,0.9760000109672546 2.509999990463257,0.5199999809265137 2.509999990463257,0 C2.509999990463257,-0.5199999809265137 2.390000104904175,-0.9739999771118164 2.1700000762939453,-1.3600000143051147 C1.9500000476837158,-1.753999948501587 1.649999976158142,-2.056999921798706 1.2699999809265137,-2.2699999809265137z M1.2200000286102295,0.9399999976158142 C1.0800000429153442,1.1859999895095825 0.9100000262260437,1.3730000257492065 0.6899999976158142,1.5 C0.4699999988079071,1.6260000467300415 0.23999999463558197,1.690000057220459 -0.009999999776482582,1.690000057220459 C-0.27000001072883606,1.690000057220459 -0.5,1.6260000467300415 -0.7099999785423279,1.5 C-0.9300000071525574,1.3730000257492065 -1.100000023841858,1.1859999895095825 -1.2300000190734863,0.9399999976158142 C-1.3700000047683716,0.6859999895095825 -1.4299999475479126,0.37299999594688416 -1.4299999475479126,0 C-1.4299999475479126,-0.37400001287460327 -1.3700000047683716,-0.6840000152587891 -1.2300000190734863,-0.9300000071525574 C-1.100000023841858,-1.184000015258789 -0.9300000071525574,-1.3739999532699585 -0.7099999785423279,-1.5 C-0.49000000953674316,-1.6269999742507935 -0.25999999046325684,-1.690000057220459 0,-1.690000057220459 C0.25,-1.690000057220459 0.47999998927116394,-1.6269999742507935 0.699999988079071,-1.5 C0.9200000166893005,-1.3739999532699585 1.090000033378601,-1.184000015258789 1.2200000286102295,-0.9300000071525574 C1.350000023841858,-0.6840000152587891 1.4199999570846558,-0.37400001287460327 1.4199999570846558,0 C1.4199999570846558,0.37299999594688416 1.350000023841858,0.6859999895095825 1.2200000286102295,0.9399999976158142z"></path></g><g opacity="1" transform="matrix(1,0,0,1,949.2550048828125,263.9070129394531)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.4450000524520874,-3.299999952316284 C1.0750000476837158,-3.5269999504089355 0.6549999713897705,-3.640000104904175 0.17499999701976776,-3.640000104904175 C-0.26499998569488525,-3.640000104904175 -0.625,-3.549999952316284 -0.9049999713897705,-3.369999885559082 C-1.184999942779541,-3.190000057220459 -1.4049999713897705,-2.9839999675750732 -1.565000057220459,-2.75 C-1.565000057220459,-2.75 -1.6749999523162842,-3.5199999809265137 -1.6749999523162842,-3.5199999809265137 C-1.6749999523162842,-3.5199999809265137 -2.625,-3.5199999809265137 -2.625,-3.5199999809265137 C-2.625,-3.5199999809265137 -2.625,3.640000104904175 -2.625,3.640000104904175 C-2.625,3.640000104904175 -1.565000057220459,3.640000104904175 -1.565000057220459,3.640000104904175 C-1.565000057220459,3.640000104904175 -1.565000057220459,0.6800000071525574 -1.565000057220459,0.6800000071525574 C-1.3949999809265137,0.9599999785423279 -1.1649999618530273,1.1759999990463257 -0.8650000095367432,1.3300000429153442 C-0.5649999976158142,1.4830000400543213 -0.22499999403953552,1.559999942779541 0.16500000655651093,1.559999942779541 C0.6449999809265137,1.559999942779541 1.0750000476837158,1.4500000476837158 1.4450000524520874,1.2300000190734863 C1.8049999475479126,1.003000020980835 2.0950000286102295,0.6959999799728394 2.305000066757202,0.3100000023841858 C2.515000104904175,-0.08399999886751175 2.625,-0.5299999713897705 2.625,-1.0299999713897705 C2.625,-1.5299999713897705 2.515000104904175,-1.9769999980926514 2.305000066757202,-2.369999885559082 C2.0950000286102295,-2.7639999389648438 1.8049999475479126,-3.0739998817443848 1.4450000524520874,-3.299999952316284z M1.1050000190734863,0.18000000715255737 C0.8050000071525574,0.4860000014305115 0.4350000023841858,0.6399999856948853 -0.014999999664723873,0.6399999856948853 C-0.4749999940395355,0.6399999856948853 -0.8450000286102295,0.4860000014305115 -1.1349999904632568,0.18000000715255737 C-1.4249999523162842,-0.1340000033378601 -1.565000057220459,-0.5400000214576721 -1.565000057220459,-1.0399999618530273 C-1.565000057220459,-1.534000039100647 -1.4249999523162842,-1.937000036239624 -1.1349999904632568,-2.25 C-0.8450000286102295,-2.563999891281128 -0.4749999940395355,-2.7200000286102295 -0.014999999664723873,-2.7200000286102295 C0.2849999964237213,-2.7200000286102295 0.5550000071525574,-2.6470000743865967 0.7950000166893005,-2.5 C1.024999976158142,-2.359999895095825 1.2050000429153442,-2.1640000343322754 1.3450000286102295,-1.909999966621399 C1.475000023841858,-1.656999945640564 1.5449999570846558,-1.3669999837875366 1.5449999570846558,-1.0399999618530273 C1.5449999570846558,-0.5400000214576721 1.3949999809265137,-0.1340000033378601 1.1050000190734863,0.18000000715255737z"></path></g><g opacity="1" transform="matrix(1,0,0,1,960.8250122070312,264.74200439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0.004999999888241291,-0.6650000214576721 C-0.19499999284744263,-0.6650000214576721 -0.36500000953674316,-0.5989999771118164 -0.4950000047683716,-0.4650000035762787 C-0.625,-0.33899998664855957 -0.6850000023841858,-0.18199999630451202 -0.6850000023841858,0.004999999888241291 C-0.6850000023841858,0.1850000023841858 -0.625,0.3409999907016754 -0.4950000047683716,0.4749999940395355 C-0.36500000953674316,0.6010000109672546 -0.19499999284744263,0.6650000214576721 0.004999999888241291,0.6650000214576721 C0.20499999821186066,0.6650000214576721 0.36500000953674316,0.6010000109672546 0.4950000047683716,0.4749999940395355 C0.625,0.3409999907016754 0.6850000023841858,0.1850000023841858 0.6850000023841858,0.004999999888241291 C0.6850000023841858,-0.18199999630451202 0.625,-0.33899998664855957 0.4950000047683716,-0.4650000035762787 C0.36500000953674316,-0.5989999771118164 0.20499999821186066,-0.6650000214576721 0.004999999888241291,-0.6650000214576721z"></path></g><g opacity="1" transform="matrix(1,0,0,1,964.5499877929688,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.6299999952316284,-1.4900000095367432 C-0.4099999964237213,-1.6239999532699585 -0.17000000178813934,-1.690000057220459 0.09000000357627869,-1.690000057220459 C0.4000000059604645,-1.690000057220459 0.6600000262260437,-1.6139999628067017 0.8899999856948853,-1.4600000381469727 C1.1200000047683716,-1.3070000410079956 1.2599999904632568,-1.093999981880188 1.3300000429153442,-0.8199999928474426 C1.3300000429153442,-0.8199999928474426 2.440000057220459,-0.8199999928474426 2.440000057220459,-0.8199999928474426 C2.319999933242798,-1.3669999837875366 2.049999952316284,-1.7999999523162842 1.6299999952316284,-2.119999885559082 C1.2200000286102295,-2.440000057220459 0.699999988079071,-2.5999999046325684 0.09000000357627869,-2.5999999046325684 C-0.4000000059604645,-2.5999999046325684 -0.8299999833106995,-2.490000009536743 -1.2200000286102295,-2.2699999809265137 C-1.600000023841858,-2.049999952316284 -1.899999976158142,-1.74399995803833 -2.119999885559082,-1.350000023841858 C-2.3299999237060547,-0.9639999866485596 -2.440000057220459,-0.5139999985694885 -2.440000057220459,0 C-2.440000057220459,0.5130000114440918 -2.3299999237060547,0.9660000205039978 -2.119999885559082,1.3600000143051147 C-1.899999976158142,1.746000051498413 -1.600000023841858,2.049999952316284 -1.2200000286102295,2.2699999809265137 C-0.8299999833106995,2.490000009536743 -0.4000000059604645,2.5999999046325684 0.09000000357627869,2.5999999046325684 C0.699999988079071,2.5999999046325684 1.2200000286102295,2.436000108718872 1.6399999856948853,2.109999895095825 C2.059999942779541,1.7829999923706055 2.3299999237060547,1.3530000448226929 2.440000057220459,0.8199999928474426 C2.440000057220459,0.8199999928474426 1.3300000429153442,0.8199999928474426 1.3300000429153442,0.8199999928474426 C1.2599999904632568,1.100000023841858 1.1200000047683716,1.315999984741211 0.8899999856948853,1.4700000286102295 C0.6600000262260437,1.6230000257492065 0.4000000059604645,1.7000000476837158 0.09000000357627869,1.7000000476837158 C-0.17000000178813934,1.7000000476837158 -0.4099999964237213,1.6330000162124634 -0.6299999952316284,1.5 C-0.8500000238418579,1.3660000562667847 -1.0299999713897705,1.1759999990463257 -1.159999966621399,0.9300000071525574 C-1.2899999618530273,0.6759999990463257 -1.3600000143051147,0.3659999966621399 -1.3600000143051147,0 C-1.3600000143051147,-0.367000013589859 -1.2899999618530273,-0.6740000247955322 -1.159999966621399,-0.9200000166893005 C-1.0299999713897705,-1.1740000247955322 -0.8500000238418579,-1.3639999628067017 -0.6299999952316284,-1.4900000095367432z"></path></g><g opacity="1" transform="matrix(1,0,0,1,924.52001953125,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.3200000524520874,-2.2699999809265137 C0.9599999785423279,-2.490000009536743 0.5400000214576721,-2.5999999046325684 0.05000000074505806,-2.5999999046325684 C-0.44999998807907104,-2.5999999046325684 -0.8899999856948853,-2.490000009536743 -1.2699999809265137,-2.2699999809265137 C-1.6399999856948853,-2.049999952316284 -1.9299999475479126,-1.74399995803833 -2.1500000953674316,-1.350000023841858 C-2.3499999046325684,-0.9570000171661377 -2.4600000381469727,-0.5 -2.4600000381469727,0.019999999552965164 C-2.4600000381469727,0.5329999923706055 -2.3499999046325684,0.9829999804496765 -2.140000104904175,1.3700000047683716 C-1.9199999570846558,1.75600004196167 -1.6299999952316284,2.059999942779541 -1.2599999904632568,2.2799999713897705 C-0.8799999952316284,2.493000030517578 -0.4399999976158142,2.5999999046325684 0.03999999910593033,2.5999999046325684 C0.6299999952316284,2.5999999046325684 1.1200000047683716,2.443000078201294 1.5099999904632568,2.130000114440918 C1.909999966621399,1.815999984741211 2.180000066757202,1.4259999990463257 2.309999942779541,0.9599999785423279 C2.309999942779541,0.9599999785423279 1.25,0.9599999785423279 1.25,0.9599999785423279 C1.149999976158142,1.1929999589920044 1,1.3799999952316284 0.7900000214576721,1.5199999809265137 C0.5899999737739563,1.652999997138977 0.3400000035762787,1.7200000286102295 0.03999999910593033,1.7200000286102295 C-0.33000001311302185,1.7200000286102295 -0.6600000262260437,1.5959999561309814 -0.949999988079071,1.350000023841858 C-1.2300000190734863,1.0959999561309814 -1.3799999952316284,0.7300000190734863 -1.4199999570846558,0.25 C-1.4199999570846558,0.25 2.440000057220459,0.25 2.440000057220459,0.25 C2.450000047683716,0.15600000321865082 2.450000047683716,0.0729999989271164 2.450000047683716,0 C2.4600000381469727,-0.07999999821186066 2.4600000381469727,-0.15399999916553497 2.4600000381469727,-0.2199999988079071 C2.4600000381469727,-0.6539999842643738 2.359999895095825,-1.0499999523162842 2.1600000858306885,-1.409999966621399 C1.9600000381469727,-1.7699999809265137 1.6799999475479126,-2.056999921798706 1.3200000524520874,-2.2699999809265137z M-1.399999976158142,-0.4699999988079071 C-1.340000033378601,-0.8899999856948853 -1.1699999570846558,-1.2039999961853027 -0.8999999761581421,-1.409999966621399 C-0.6200000047683716,-1.6239999532699585 -0.3100000023841858,-1.7300000190734863 0.03999999910593033,-1.7300000190734863 C0.41999998688697815,-1.7300000190734863 0.7400000095367432,-1.6169999837875366 0.9900000095367432,-1.3899999856948853 C1.25,-1.1640000343322754 1.3799999952316284,-0.8569999933242798 1.399999976158142,-0.4699999988079071 C1.399999976158142,-0.4699999988079071 -1.399999976158142,-0.4699999988079071 -1.399999976158142,-0.4699999988079071z"></path></g><g opacity="1" transform="matrix(1,0,0,1,957.1300048828125,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.3200000524520874,-2.2699999809265137 C0.9599999785423279,-2.490000009536743 0.5400000214576721,-2.5999999046325684 0.05000000074505806,-2.5999999046325684 C-0.44999998807907104,-2.5999999046325684 -0.8899999856948853,-2.490000009536743 -1.2699999809265137,-2.2699999809265137 C-1.6399999856948853,-2.049999952316284 -1.9299999475479126,-1.74399995803833 -2.1500000953674316,-1.350000023841858 C-2.3499999046325684,-0.9570000171661377 -2.4600000381469727,-0.5 -2.4600000381469727,0.019999999552965164 C-2.4600000381469727,0.5329999923706055 -2.3499999046325684,0.9829999804496765 -2.140000104904175,1.3700000047683716 C-1.9199999570846558,1.75600004196167 -1.6299999952316284,2.059999942779541 -1.2599999904632568,2.2799999713897705 C-0.8799999952316284,2.493000030517578 -0.4399999976158142,2.5999999046325684 0.03999999910593033,2.5999999046325684 C0.6299999952316284,2.5999999046325684 1.1200000047683716,2.443000078201294 1.5099999904632568,2.130000114440918 C1.909999966621399,1.815999984741211 2.180000066757202,1.4259999990463257 2.309999942779541,0.9599999785423279 C2.309999942779541,0.9599999785423279 1.25,0.9599999785423279 1.25,0.9599999785423279 C1.149999976158142,1.1929999589920044 1,1.3799999952316284 0.7900000214576721,1.5199999809265137 C0.5899999737739563,1.652999997138977 0.3400000035762787,1.7200000286102295 0.03999999910593033,1.7200000286102295 C-0.33000001311302185,1.7200000286102295 -0.6600000262260437,1.5959999561309814 -0.949999988079071,1.350000023841858 C-1.2300000190734863,1.0959999561309814 -1.3799999952316284,0.7300000190734863 -1.4199999570846558,0.25 C-1.4199999570846558,0.25 2.440000057220459,0.25 2.440000057220459,0.25 C2.450000047683716,0.15600000321865082 2.450000047683716,0.0729999989271164 2.450000047683716,0 C2.4600000381469727,-0.07999999821186066 2.4600000381469727,-0.15399999916553497 2.4600000381469727,-0.2199999988079071 C2.4600000381469727,-0.6539999842643738 2.359999895095825,-1.0499999523162842 2.1600000858306885,-1.409999966621399 C1.9600000381469727,-1.7699999809265137 1.6799999475479126,-2.056999921798706 1.3200000524520874,-2.2699999809265137z M-1.399999976158142,-0.4699999988079071 C-1.340000033378601,-0.8899999856948853 -1.1699999570846558,-1.2039999961853027 -0.8999999761581421,-1.409999966621399 C-0.6200000047683716,-1.6239999532699585 -0.3100000023841858,-1.7300000190734863 0.03999999910593033,-1.7300000190734863 C0.41999998688697815,-1.7300000190734863 0.7400000095367432,-1.6169999837875366 0.9900000095367432,-1.3899999856948853 C1.25,-1.1640000343322754 1.3799999952316284,-0.8569999933242798 1.399999976158142,-0.4699999988079071 C1.399999976158142,-0.4699999988079071 -1.399999976158142,-0.4699999988079071 -1.399999976158142,-0.4699999988079071z"></path></g><g opacity="1" transform="matrix(1,0,0,1,970.1799926757812,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.2699999809265137,-2.2699999809265137 C0.8999999761581421,-2.490000009536743 0.47999998927116394,-2.5999999046325684 0,-2.5999999046325684 C-0.46000000834465027,-2.5999999046325684 -0.8899999856948853,-2.490000009536743 -1.2699999809265137,-2.2699999809265137 C-1.649999976158142,-2.056999921798706 -1.9500000476837158,-1.753999948501587 -2.180000066757202,-1.3600000143051147 C-2.4000000953674316,-0.9739999771118164 -2.509999990463257,-0.5199999809265137 -2.509999990463257,0 C-2.509999990463257,0.5199999809265137 -2.4000000953674316,0.9760000109672546 -2.180000066757202,1.3700000047683716 C-1.9600000381469727,1.75600004196167 -1.659999966621399,2.059999942779541 -1.2899999618530273,2.2799999713897705 C-0.9100000262260437,2.493000030517578 -0.47999998927116394,2.5999999046325684 -0.009999999776482582,2.5999999046325684 C0.4699999988079071,2.5999999046325684 0.8899999856948853,2.493000030517578 1.2699999809265137,2.2799999713897705 C1.649999976158142,2.059999942779541 1.9500000476837158,1.75600004196167 2.1700000762939453,1.3700000047683716 C2.4000000953674316,0.9760000109672546 2.509999990463257,0.5199999809265137 2.509999990463257,0 C2.509999990463257,-0.5199999809265137 2.4000000953674316,-0.9739999771118164 2.1700000762939453,-1.3600000143051147 C1.9500000476837158,-1.753999948501587 1.649999976158142,-2.056999921798706 1.2699999809265137,-2.2699999809265137z M1.2200000286102295,0.9399999976158142 C1.090000033378601,1.1859999895095825 0.9100000262260437,1.3730000257492065 0.6899999976158142,1.5 C0.47999998927116394,1.6260000467300415 0.25,1.690000057220459 -0.009999999776482582,1.690000057220459 C-0.25999999046325684,1.690000057220459 -0.49000000953674316,1.6260000467300415 -0.7099999785423279,1.5 C-0.9200000166893005,1.3730000257492065 -1.090000033378601,1.1859999895095825 -1.2300000190734863,0.9399999976158142 C-1.3600000143051147,0.6859999895095825 -1.4299999475479126,0.37299999594688416 -1.4299999475479126,0 C-1.4299999475479126,-0.37400001287460327 -1.3600000143051147,-0.6840000152587891 -1.2300000190734863,-0.9300000071525574 C-1.090000033378601,-1.184000015258789 -0.9200000166893005,-1.3739999532699585 -0.7099999785423279,-1.5 C-0.49000000953674316,-1.6269999742507935 -0.25,-1.690000057220459 0,-1.690000057220459 C0.25999999046325684,-1.690000057220459 0.49000000953674316,-1.6269999742507935 0.699999988079071,-1.5 C0.9200000166893005,-1.3739999532699585 1.100000023841858,-1.184000015258789 1.2200000286102295,-0.9300000071525574 C1.3600000143051147,-0.6840000152587891 1.4199999570846558,-0.37400001287460327 1.4199999570846558,0 C1.4199999570846558,0.37299999594688416 1.3600000143051147,0.6859999895095825 1.2200000286102295,0.9399999976158142z"></path></g><g opacity="1" transform="matrix(1,0,0,1,977.52001953125,262.8070068359375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M3.430000066757202,-1.9800000190734863 C3.069999933242798,-2.3540000915527344 2.5899999141693115,-2.5399999618530273 1.9700000286102295,-2.5399999618530273 C1.6100000143051147,-2.5399999618530273 1.2799999713897705,-2.4570000171661377 0.9800000190734863,-2.2899999618530273 C0.6800000071525574,-2.124000072479248 0.4399999976158142,-1.8969999551773071 0.25999999046325684,-1.6100000143051147 C-0.05000000074505806,-2.2300000190734863 -0.6000000238418579,-2.5399999618530273 -1.3799999952316284,-2.5399999618530273 C-1.7300000190734863,-2.5399999618530273 -2.0399999618530273,-2.4670000076293945 -2.299999952316284,-2.319999933242798 C-2.559999942779541,-2.1740000247955322 -2.7699999809265137,-1.9739999771118164 -2.930000066757202,-1.7200000286102295 C-2.930000066757202,-1.7200000286102295 -3.0199999809265137,-2.4200000762939453 -3.0199999809265137,-2.4200000762939453 C-3.0199999809265137,-2.4200000762939453 -3.9600000381469727,-2.4200000762939453 -3.9600000381469727,-2.4200000762939453 C-3.9600000381469727,-2.4200000762939453 -3.9600000381469727,2.5399999618530273 -3.9600000381469727,2.5399999618530273 C-3.9600000381469727,2.5399999618530273 -2.9000000953674316,2.5399999618530273 -2.9000000953674316,2.5399999618530273 C-2.9000000953674316,2.5399999618530273 -2.9000000953674316,-0.019999999552965164 -2.9000000953674316,-0.019999999552965164 C-2.9000000953674316,-0.5400000214576721 -2.7899999618530273,-0.9399999976158142 -2.549999952316284,-1.2200000286102295 C-2.309999942779541,-1.5 -2.009999990463257,-1.6399999856948853 -1.6399999856948853,-1.6399999856948853 C-0.8999999761581421,-1.6399999856948853 -0.5299999713897705,-1.159999966621399 -0.5299999713897705,-0.20000000298023224 C-0.5299999713897705,-0.20000000298023224 -0.5299999713897705,2.5399999618530273 -0.5299999713897705,2.5399999618530273 C-0.5299999713897705,2.5399999618530273 0.5299999713897705,2.5399999618530273 0.5299999713897705,2.5399999618530273 C0.5299999713897705,2.5399999618530273 0.5299999713897705,-0.019999999552965164 0.5299999713897705,-0.019999999552965164 C0.5299999713897705,-0.5400000214576721 0.6399999856948853,-0.9399999976158142 0.8799999952316284,-1.2200000286102295 C1.1200000047683716,-1.5 1.4199999570846558,-1.6399999856948853 1.7999999523162842,-1.6399999856948853 C2.5299999713897705,-1.6399999856948853 2.9000000953674316,-1.159999966621399 2.9000000953674316,-0.20000000298023224 C2.9000000953674316,-0.20000000298023224 2.9000000953674316,2.5399999618530273 2.9000000953674316,2.5399999618530273 C2.9000000953674316,2.5399999618530273 3.9600000381469727,2.5399999618530273 3.9600000381469727,2.5399999618530273 C3.9600000381469727,2.5399999618530273 3.9600000381469727,-0.3100000023841858 3.9600000381469727,-0.3100000023841858 C3.9600000381469727,-1.0499999523162842 3.7799999713897705,-1.6069999933242798 3.430000066757202,-1.9800000190734863z"></path></g><g opacity="1" transform="matrix(1,0,0,1,934.5399780273438,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0.07999999821186066,-2.5999999046325684 C-0.30000001192092896,-2.5999999046325684 -0.6600000262260437,-2.5299999713897705 -0.9900000095367432,-2.390000104904175 C-1.309999942779541,-2.256999969482422 -1.5700000524520874,-2.059999942779541 -1.7899999618530273,-1.7999999523162842 C-1.9900000095367432,-1.5470000505447388 -2.109999895095825,-1.2400000095367432 -2.1500000953674316,-0.8799999952316284 C-2.1500000953674316,-0.8799999952316284 -1.090000033378601,-0.8799999952316284 -1.090000033378601,-0.8799999952316284 C-1.0399999618530273,-1.159999966621399 -0.9100000262260437,-1.3700000047683716 -0.699999988079071,-1.5099999904632568 C-0.47999998927116394,-1.656999945640564 -0.2199999988079071,-1.7300000190734863 0.07000000029802322,-1.7300000190734863 C0.4099999964237213,-1.7300000190734863 0.6800000071525574,-1.6339999437332153 0.8799999952316284,-1.440000057220459 C1.100000023841858,-1.246999979019165 1.2000000476837158,-0.9570000171661377 1.2000000476837158,-0.5699999928474426 C1.2000000476837158,-0.5699999928474426 1.2000000476837158,-0.47999998927116394 1.2000000476837158,-0.47999998927116394 C1.2000000476837158,-0.47999998927116394 -0.10000000149011612,-0.47999998927116394 -0.10000000149011612,-0.47999998927116394 C-0.7900000214576721,-0.47999998927116394 -1.3200000524520874,-0.3400000035762787 -1.7000000476837158,-0.05999999865889549 C-2.069999933242798,0.2199999988079071 -2.259999990463257,0.6000000238418579 -2.259999990463257,1.0800000429153442 C-2.259999990463257,1.3530000448226929 -2.190000057220459,1.6059999465942383 -2.059999942779541,1.840000033378601 C-1.9199999570846558,2.065999984741211 -1.7200000286102295,2.25 -1.4500000476837158,2.390000104904175 C-1.1699999570846558,2.5299999713897705 -0.8299999833106995,2.5999999046325684 -0.4099999964237213,2.5999999046325684 C0.029999999329447746,2.5999999046325684 0.3799999952316284,2.50600004196167 0.6399999856948853,2.319999933242798 C0.9100000262260437,2.132999897003174 1.1100000143051147,1.902999997138977 1.25,1.6299999952316284 C1.25,1.6299999952316284 1.340000033378601,2.4800000190734863 1.340000033378601,2.4800000190734863 C1.340000033378601,2.4800000190734863 2.259999990463257,2.4800000190734863 2.259999990463257,2.4800000190734863 C2.259999990463257,2.4800000190734863 2.259999990463257,-0.5699999928474426 2.259999990463257,-0.5699999928474426 C2.259999990463257,-1.1970000267028809 2.069999933242798,-1.690000057220459 1.690000057220459,-2.049999952316284 C1.309999942779541,-2.4170000553131104 0.7799999713897705,-2.5999999046325684 0.07999999821186066,-2.5999999046325684z M0.9800000190734863,0.9900000095367432 C0.8799999952316284,1.215999960899353 0.7200000286102295,1.399999976158142 0.5199999809265137,1.5399999618530273 C0.33000001311302185,1.6729999780654907 0.09000000357627869,1.7400000095367432 -0.20000000298023224,1.7400000095367432 C-0.49000000953674316,1.7400000095367432 -0.7200000286102295,1.6759999990463257 -0.8999999761581421,1.5499999523162842 C-1.059999942779541,1.4160000085830688 -1.149999976158142,1.2359999418258667 -1.149999976158142,1.0099999904632568 C-1.149999976158142,0.7900000214576721 -1.059999942779541,0.6060000061988831 -0.8899999856948853,0.46000000834465027 C-0.7099999785423279,0.31299999356269836 -0.4099999964237213,0.23999999463558197 0.009999999776482582,0.23999999463558197 C0.009999999776482582,0.23999999463558197 1.190000057220459,0.23999999463558197 1.190000057220459,0.23999999463558197 C1.1699999570846558,0.5130000114440918 1.100000023841858,0.7630000114440918 0.9800000190734863,0.9900000095367432z"></path></g><g opacity="1" transform="matrix(1,0,0,1,929.47998046875,262.86700439453125)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.3200000524520874,-2.4800000190734863 C1.3200000524520874,-2.4800000190734863 0,-0.5799999833106995 0,-0.5799999833106995 C0,-0.5799999833106995 -1.3300000429153442,-2.4800000190734863 -1.3300000429153442,-2.4800000190734863 C-1.3300000429153442,-2.4800000190734863 -2.4600000381469727,-2.4800000190734863 -2.4600000381469727,-2.4800000190734863 C-2.4600000381469727,-2.4800000190734863 -0.699999988079071,0 -0.699999988079071,0 C-0.699999988079071,0 -2.4600000381469727,2.4800000190734863 -2.4600000381469727,2.4800000190734863 C-2.4600000381469727,2.4800000190734863 -1.3300000429153442,2.4800000190734863 -1.3300000429153442,2.4800000190734863 C-1.3300000429153442,2.4800000190734863 0,0.5799999833106995 0,0.5799999833106995 C0,0.5799999833106995 1.3200000524520874,2.4800000190734863 1.3200000524520874,2.4800000190734863 C1.3200000524520874,2.4800000190734863 2.4600000381469727,2.4800000190734863 2.4600000381469727,2.4800000190734863 C2.4600000381469727,2.4800000190734863 0.699999988079071,0 0.699999988079071,0 C0.699999988079071,0 2.4600000381469727,-2.4800000190734863 2.4600000381469727,-2.4800000190734863 C2.4600000381469727,-2.4800000190734863 1.3200000524520874,-2.4800000190734863 1.3200000524520874,-2.4800000190734863z"></path></g><g opacity="1" transform="matrix(1,0,0,1,941.7100219726562,262.8070068359375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M1.9700000286102295,-2.5399999618530273 C1.6200000047683716,-2.5399999618530273 1.2899999618530273,-2.4570000171661377 0.9800000190734863,-2.2899999618530273 C0.6800000071525574,-2.124000072479248 0.4399999976158142,-1.8969999551773071 0.25999999046325684,-1.6100000143051147 C-0.03999999910593033,-2.2300000190734863 -0.5899999737739563,-2.5399999618530273 -1.3799999952316284,-2.5399999618530273 C-1.7200000286102295,-2.5399999618530273 -2.0299999713897705,-2.4670000076293945 -2.299999952316284,-2.319999933242798 C-2.559999942779541,-2.1740000247955322 -2.7699999809265137,-1.9739999771118164 -2.930000066757202,-1.7200000286102295 C-2.930000066757202,-1.7200000286102295 -3.0199999809265137,-2.4200000762939453 -3.0199999809265137,-2.4200000762939453 C-3.0199999809265137,-2.4200000762939453 -3.9600000381469727,-2.4200000762939453 -3.9600000381469727,-2.4200000762939453 C-3.9600000381469727,-2.4200000762939453 -3.9600000381469727,2.5399999618530273 -3.9600000381469727,2.5399999618530273 C-3.9600000381469727,2.5399999618530273 -2.9000000953674316,2.5399999618530273 -2.9000000953674316,2.5399999618530273 C-2.9000000953674316,2.5399999618530273 -2.9000000953674316,-0.019999999552965164 -2.9000000953674316,-0.019999999552965164 C-2.9000000953674316,-0.5400000214576721 -2.7799999713897705,-0.9399999976158142 -2.549999952316284,-1.2200000286102295 C-2.309999942779541,-1.5 -2,-1.6399999856948853 -1.6399999856948853,-1.6399999856948853 C-0.8999999761581421,-1.6399999856948853 -0.5299999713897705,-1.159999966621399 -0.5299999713897705,-0.20000000298023224 C-0.5299999713897705,-0.20000000298023224 -0.5299999713897705,2.5399999618530273 -0.5299999713897705,2.5399999618530273 C-0.5299999713897705,2.5399999618530273 0.5299999713897705,2.5399999618530273 0.5299999713897705,2.5399999618530273 C0.5299999713897705,2.5399999618530273 0.5299999713897705,-0.019999999552965164 0.5299999713897705,-0.019999999552965164 C0.5299999713897705,-0.5400000214576721 0.6499999761581421,-0.9399999976158142 0.8799999952316284,-1.2200000286102295 C1.1200000047683716,-1.5 1.4299999475479126,-1.6399999856948853 1.7999999523162842,-1.6399999856948853 C2.5399999618530273,-1.6399999856948853 2.9000000953674316,-1.159999966621399 2.9000000953674316,-0.20000000298023224 C2.9000000953674316,-0.20000000298023224 2.9000000953674316,2.5399999618530273 2.9000000953674316,2.5399999618530273 C2.9000000953674316,2.5399999618530273 3.9600000381469727,2.5399999618530273 3.9600000381469727,2.5399999618530273 C3.9600000381469727,2.5399999618530273 3.9600000381469727,-0.3100000023841858 3.9600000381469727,-0.3100000023841858 C3.9600000381469727,-1.0499999523162842 3.7899999618530273,-1.6069999933242798 3.430000066757202,-1.9800000190734863 C3.0799999237060547,-2.3540000915527344 2.5899999141693115,-2.5399999618530273 1.9700000286102295,-2.5399999618530273z"></path></g><g opacity="1" transform="matrix(1,0,0,1,953.27001953125,261.74700927734375)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-0.5299999713897705,3.5999999046325684 C-0.5299999713897705,3.5999999046325684 0.5299999713897705,3.5999999046325684 0.5299999713897705,3.5999999046325684 C0.5299999713897705,3.5999999046325684 0.5299999713897705,-3.5999999046325684 0.5299999713897705,-3.5999999046325684 C0.5299999713897705,-3.5999999046325684 -0.5299999713897705,-3.5999999046325684 -0.5299999713897705,-3.5999999046325684 C-0.5299999713897705,-3.5999999046325684 -0.5299999713897705,3.5999999046325684 -0.5299999713897705,3.5999999046325684z"></path></g></g><g transform="matrix(1.3937740325927734,0,0,1.3937740325927734,-1642.6363525390625,-142.80917358398438)" opacity="0.4" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><g opacity="1" transform="matrix(1,0,0,1,-12.771031379699707,0)"><path stroke-linecap="butt" stroke-linejoin="miter" fill-opacity="0" stroke-miterlimit="4" stroke="rgb(255,255,255)" stroke-opacity="1" stroke-width="0.5" d=" M1517.1829833984375,346.947998046875 C1517.1829833984375,346.947998046875 1517.1829833984375,365.947998046875 1517.1829833984375,365.947998046875"></path></g></g></g><g class="0 0 png" clip-path="url(#__lottie_element_695)" style="display: none;" transform="matrix(0.6660119295120239,0,0,0.6660119295120239,103.37197875976562,434.5452575683594)" opacity="3.059729061760663e-7"><image width="307px" height="248px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAATMAAAD4CAMAAAC33vGvAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAADVUExURUdwTHOC/2OJ/128+q9r/1KR/4Z7/12d/ppz/5pz/7ln/1es/VGe/2PM+GTR92PQ97hm/7hn/2TQ+GTQ92TQ+GLQ9woLMzDMpP///0lKaLhm/2A5mqNv/4SFmefn62XT9zFMmU9AmUFbsywrUBQcQHeB/0BGmTdulY14/2VLs1KS/2OJ/8LCzJycrV/C9z13ryg9gDNilg8TK1Oj/Vmz+y5XmCqwkzc1gEQwgChMfRpZX2VmgHV2jB9ybBc5USaZhidveNHR2bOzv4tPyiKFeU2extDR2frxEswAAAAWdFJOUwDf39/g398fH9+O39/fiWBWxcXvoD03A0W7AAAKZElEQVR42u2dC3ubOBaGPbvZTSa7ubcGOQE6gzJtOuBMk5bt2MZ+7Dj5/z9pdeEisCQgduPU+V5wik3kjF4fSQcMml4vZ//98fnFMBiu8pUvq9zLZZXPfGniT7mYueYrW/hq41O+8NXGVb7w1cpALnwV7J0dnvY0HF8Ew8fbjxlf+GLkG1++iUf+s+ADW76Jh9xmS2v+yBeO/Cn+yTZr/MbX3yrP5IZY2nPJV7aYuXlg3g4PasZOLoLHaR8YCW8Ge9VYO4axZmsPg0NF2bvgI5w0c6lIO4ayrtJOoKy9NNmn7R8NIaMtD4MD2TIpXLQeCGTrPLqFio6BdhIgy+gUaKxHO0Zv1gneOM8f4aFT4zzr9S7QnXVMN3o9JGdwBmdwBmdV6GiZOIxkNqJw1obRzFFIRnDWxDRxauyutc04o6mjIaVwZlaWOFoSCmcdle2qtA04MyvbUWkbcJY6FlI40+UYjpUpnK2S2J0llqLE84jc8vItyrZC+drYK8he6efPSWjoJryFH8XE9OfiuWXvCzprCDPHsaRpsetGsjausuVneyO3IK9n+Yqne7/Qlztj7c78/WK6bWezJmczS5zlgiZuvsU0LkpnfobijD/lNZ/olfmRb97JdvMf8y07o04j5o+VsiqERUiRbGtcOhvXCrDfEvWfl9FYDds5Ff/od/oe7YeeKUhfzpm+aSZpu8YZZSFRNjc3s2h1Jtry6rvNZVmqvIca0tm7cXl0q860TXN2d6dIW5pLT2TfwyqUdW1Kd2ZzFmqdRZHsE32Ns7LN8+Aeb9WZbtRM7xitRs5MEasQ8cWnP1E6cIuzST5kaDt7ndB5+WaxYZB4MWeVFlkqqxwbWIqzz5yZemLmYtGhLcoxkjtbZKmF4ow/56NDaHxPT+dEaa/jdUeBDTpL7qai+2LKqmeGLMWFo5C3GyJqKh3Wcw2vX8815tQaZqHNGdGOEdtxxl1lyqpttd/QoRHebihvbqHa5spcY6LUnHVaT74lznhOoWl61Th7ei3ORJucsh/1sdRSXEiKRX2YIkrU+lr6M8/YvLiyub4TCMvPKXo1Y4Do+ytDZlNSy+vis8dceiCx0p3ZnLEe0CUmZdoQjMpEd7HtMWC5MmCmThdnzNK4TDgitaexOVtoc32h7Ck0DAx+2d+RrTob1bOMZZcDTjmIZXXw+ZHNopUznmNp6s0SFjeqjA40F0jzw1AarTsEbPrYKU26HDtldcnGSjFOjivOPCKhpTP+dDzXjo38qDWSBcLiE4nLQHPjrOh4u86aj9ETe/mn4pQGr7JqwnpeQ3fMuLo3csvcJbafE/lZzgXlVZkUHY3fb+dsruuRVp1N1Hf05C5/8lOfcxSmyqZXtqliT61t5s/1yVn5+8UvkLHSM1DieROy/jcU6zubrhVmb/PcNr5Dec7Qie/qNikN3wl3loZrD2zScI3Lc9K01WupdvZGjR91zd4M1+y1a6Gjmbw2dIlrQwGcwRmc/SzOcMtrd2e4F7Eb/F5E3PPaDX7P63vcW90FcW/1/gUaZwdu9uRcEQi09kc7cq4IBFqXEWDvAHPfdE00TjHHUldlh5jL6/nKeKQNMWlQU5ZRnTOO9WlHwS2GT7uxQX0ezP3jI2UOTPsUmHIezPrklzU+5NNfdpgD88Mf6jyY+pkvzdNhZnNfdpn+Up0C0zYRJhM2WJkDU8Qan2vVwFcD91paTLXKZ1ttRdNUq9ftJlotJlttTTHTKuPs8FRnDAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIC3y/7Ju/OjnH8a+Y+W/67yr2b+YeOXJv7dil9bsPfrnoUz0/8e/eQ8CIaPt535na+/F/+yha8/jBu+5kv12Q1ffwQPD/x/K39aN/b+PHj8Qvv9/l0faKDfrwZ71Vg7CYZ/Q4wdZu1QUXYcPFJIaYy1B0XacfARRtpwWUh7H9xCRztuBtlIcDREw2zbPK/kQHASoPtvTShb59EjVLTnYSB6M4RZp0A75YMmRHTp0XjjPEfT7NY4z3q9CyQa3dKNvV4P+WzXvBbO4AzOdt0ZnY6WsySZzdIpnLVimjoKSwpnjTE2c2rsrrUNORs5GlI4swTZ0nHekLRNOKOJYyChcKZn6RhZwpmW1LEwgjNdy3SsTOFslcTubGaxTQipb4WEhNluUpC/EuYvmN/Riz3z3jHfG27f2dRxnhto1HVdWcG42IpcN84EuQVRVsDLX/Bj7eBCfLlXr2Ui97re1p3NmpxZAo0JmogNXptiizQ7K19SKUr4OqNl4XjLzqjTiDnf8LL//rCwwLdoYcAPM5TfZ89IGZb1j2BORLBpQmnM/wSRZSfbdTbS52Vpq6GTMC15fYqtqIwaf9WxV2vBtaYeyvdYrO708yKxIQ5fztlSn8reLdvkaLwivJYLZkpGTlzGiM2Zp3XmeZPsk4h0YZZ1c/yPTrbqTNedJXeMcjhNrB3aWHRi87HUMS8bXUOcjY0fhHbnorTs6eLwJZ3pcn+uTHVpLj0RNWF2YiqCgyqeuLNaasHDiz9n0Tg3tPY49rUDxFP5YZDVD2NrzhIqerGUGasegZpLMy9PwhxhscZ6GaKEgDJurg59nqFHGpvGVL8SwO4rccY6sbtUp8zijFeF8pZGeU9GuJRxK2e+IXElvq/PNV6nM2fEbKX8Rz3NtRQXpkRkEN5MIzkm2NomTz3GvjbXyMrNdZHml5/G1ttmUpPGqKcfljGAN8sJEeOY6NB8pTa2MWCib4DlaEwtY4C18IuPmyOdMuv5ICZmEcvg4tmGmkLYnIW6UGH9vwyluSYKJ2UJf90jgc3mtCPRpXU5HcQiK5K18YokrdkZ0XVJRfz4SgtXE94iRt1wq85qfdco7Xg2KC4OAIno3GkbZ0SbT5DsqMgrC4aR7yl/yAtZ4rv+Aefax+hJ0+FmYi0ukoNxcaAeaY64tcfoulDhWvy575bdfVz+Ip2Xg264ZWdpkzP7mVpa1n9RPU9jdabNNWjs1s72eIrcYm+07hm09c85rhVmrHvJjhJZ2/I8L6wcPuYUWUL2fGJKNEgcRYtYeRMvJtW9UUxewXna0Tph9kbPbdvPOs76cKbrRizDwE5+wbmR74Sfc472jTszRlqym9dUbeYaF7202Y5eGbSpa6nSt/EV+kad9emyGmvp7l60t8lrQ6e5tiSd7vJ9epu+Bpky+jsOrtuGMzh7vc5wj1h3Z7gXsRv8XsR3Q3jowtUhv4V/ChHtEfdW76ND68LNgE8WcYzpNTpk7XKuiP0LjAIdw4z3aF8go22ikU8YdI5hoO0AcFZMSniESGvDd3XauP3z4BYDQVP3fzk4q8y0dxwMP8Ka3Vhlmj0Rau+C4PH2y9/d+atc+LoR/icfYs0Wvnbmu3x8lwtfn8flw6AWZPlUq+cXwTAY6viq5f7rvYHP959b8GcD12y9vuaPJj5df2rDlXhcNTO4GlQ5OzwwTlH7Ojjgq/ixusvEvixz8EOoWvo/aS69LkeOBHwAAAAASUVORK5CYII="></image></g><g transform="matrix(1.3937771320343018,0,0,1.3937771320343018,295.56915283203125,544.2210693359375)" opacity="0.78" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path stroke-linecap="round" stroke-linejoin="round" fill-opacity="0" stroke-dasharray=" 2 4" stroke-dashoffset="0" stroke="rgb(69,81,189)" stroke-opacity="1" stroke-width="1" d="M0 0"></path></g></g><g transform="matrix(0.000003478006874502171,0,0,0.000003478006874502171,344.10601806640625,542.7962646484375)" opacity="1" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path fill="rgb(3,100,255)" fill-opacity="1" d=" M0,-7 C0,-7 0,-7 0,-7 C3.865999937057495,-7 7,-3.865999937057495 7,0 C7,0 7,0 7,0 C7,3.865999937057495 3.865999937057495,7 0,7 C0,7 0,7 0,7 C-3.865999937057495,7 -7,3.865999937057495 -7,0 C-7,0 -7,0 -7,0 C-7,-3.865999937057495 -3.865999937057495,-7 0,-7z"></path></g></g><g transform="matrix(1.0018882751464844,0,0,1.0018882751464844,-485.4373779296875,99.56974792480469)" opacity="0.030624556934953646" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0.843994140625,0.050994873046875)"><path stroke="url(#__lottie_element_257)" stroke-linecap="butt" stroke-linejoin="miter" fill-opacity="0" stroke-miterlimit="4" stroke-opacity="1" stroke-width="3" d=" M793.969970703125,15.937999725341797 C793.969970703125,15.937999725341797 1098.4439697265625,15.937999725341797 1098.4439697265625,15.937999725341797 C1111.008056640625,15.937999725341797 1121.1939697265625,26.124000549316406 1121.1939697265625,38.6879997253418 C1121.1939697265625,38.6879997253418 1121.1939697265625,340.02899169921875 1121.1939697265625,340.02899169921875 C1121.1939697265625,352.5929870605469 1111.008056640625,362.77899169921875 1098.4439697265625,362.77899169921875 C1098.4439697265625,362.77899169921875 793.969970703125,362.77899169921875 793.969970703125,362.77899169921875 C781.406005859375,362.77899169921875 771.219970703125,352.5929870605469 771.219970703125,340.02899169921875 C771.219970703125,340.02899169921875 771.219970703125,38.6879997253418 771.219970703125,38.6879997253418 C771.219970703125,26.124000549316406 781.406005859375,15.937999725341797 793.969970703125,15.937999725341797z"></path></g></g><g transform="matrix(0.3451286554336548,0,0,0.3451286554336548,288.37506103515625,307.8779602050781)" opacity="1" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path fill="rgb(3,100,255)" fill-opacity="1" d=" M0,-7 C0,-7 0,-7 0,-7 C3.865999937057495,-7 7,-3.865999937057495 7,0 C7,0 7,0 7,0 C7,3.865999937057495 3.865999937057495,7 0,7 C0,7 0,7 0,7 C-3.865999937057495,7 -7,3.865999937057495 -7,0 C-7,0 -7,0 -7,0 C-7,-3.865999937057495 -3.865999937057495,-7 0,-7z"></path></g></g><g class="0 0 png" clip-path="url(#__lottie_element_430)" style="display: block;" transform="matrix(1.0459963083267212,0,0,1.0459963083267212,-10.29925537109375,384.169189453125)" opacity="2.5743892848595353e-8"><image width="307px" height="248px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAATMAAAD4CAMAAAC33vGvAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAADVUExURUdwTHOC/2OJ/128+q9r/1KR/4Z7/12d/ppz/5pz/7ln/1es/VGe/2PM+GTR92PQ97hm/7hn/2TQ+GTQ92TQ+GLQ9woLMzDMpP///0lKaLhm/2A5mqNv/4SFmefn62XT9zFMmU9AmUFbsywrUBQcQHeB/0BGmTdulY14/2VLs1KS/2OJ/8LCzJycrV/C9z13ryg9gDNilg8TK1Oj/Vmz+y5XmCqwkzc1gEQwgChMfRpZX2VmgHV2jB9ybBc5USaZhidveNHR2bOzv4tPyiKFeU2extDR2frxEswAAAAWdFJOUwDf39/g398fH9+O39/fiWBWxcXvoD03A0W7AAAKZElEQVR42u2dC3ubOBaGPbvZTSa7ubcGOQE6gzJtOuBMk5bt2MZ+7Dj5/z9pdeEisCQgduPU+V5wik3kjF4fSQcMml4vZ//98fnFMBiu8pUvq9zLZZXPfGniT7mYueYrW/hq41O+8NXGVb7w1cpALnwV7J0dnvY0HF8Ew8fbjxlf+GLkG1++iUf+s+ADW76Jh9xmS2v+yBeO/Cn+yTZr/MbX3yrP5IZY2nPJV7aYuXlg3g4PasZOLoLHaR8YCW8Ge9VYO4axZmsPg0NF2bvgI5w0c6lIO4ayrtJOoKy9NNmn7R8NIaMtD4MD2TIpXLQeCGTrPLqFio6BdhIgy+gUaKxHO0Zv1gneOM8f4aFT4zzr9S7QnXVMN3o9JGdwBmdwBmdV6GiZOIxkNqJw1obRzFFIRnDWxDRxauyutc04o6mjIaVwZlaWOFoSCmcdle2qtA04MyvbUWkbcJY6FlI40+UYjpUpnK2S2J0llqLE84jc8vItyrZC+drYK8he6efPSWjoJryFH8XE9OfiuWXvCzprCDPHsaRpsetGsjausuVneyO3IK9n+Yqne7/Qlztj7c78/WK6bWezJmczS5zlgiZuvsU0LkpnfobijD/lNZ/olfmRb97JdvMf8y07o04j5o+VsiqERUiRbGtcOhvXCrDfEvWfl9FYDds5Ff/od/oe7YeeKUhfzpm+aSZpu8YZZSFRNjc3s2h1Jtry6rvNZVmqvIca0tm7cXl0q860TXN2d6dIW5pLT2TfwyqUdW1Kd2ZzFmqdRZHsE32Ns7LN8+Aeb9WZbtRM7xitRs5MEasQ8cWnP1E6cIuzST5kaDt7ndB5+WaxYZB4MWeVFlkqqxwbWIqzz5yZemLmYtGhLcoxkjtbZKmF4ow/56NDaHxPT+dEaa/jdUeBDTpL7qai+2LKqmeGLMWFo5C3GyJqKh3Wcw2vX8815tQaZqHNGdGOEdtxxl1lyqpttd/QoRHebihvbqHa5spcY6LUnHVaT74lznhOoWl61Th7ei3ORJucsh/1sdRSXEiKRX2YIkrU+lr6M8/YvLiyub4TCMvPKXo1Y4Do+ytDZlNSy+vis8dceiCx0p3ZnLEe0CUmZdoQjMpEd7HtMWC5MmCmThdnzNK4TDgitaexOVtoc32h7Ck0DAx+2d+RrTob1bOMZZcDTjmIZXXw+ZHNopUznmNp6s0SFjeqjA40F0jzw1AarTsEbPrYKU26HDtldcnGSjFOjivOPCKhpTP+dDzXjo38qDWSBcLiE4nLQHPjrOh4u86aj9ETe/mn4pQGr7JqwnpeQ3fMuLo3csvcJbafE/lZzgXlVZkUHY3fb+dsruuRVp1N1Hf05C5/8lOfcxSmyqZXtqliT61t5s/1yVn5+8UvkLHSM1DieROy/jcU6zubrhVmb/PcNr5Dec7Qie/qNikN3wl3loZrD2zScI3Lc9K01WupdvZGjR91zd4M1+y1a6Gjmbw2dIlrQwGcwRmc/SzOcMtrd2e4F7Eb/F5E3PPaDX7P63vcW90FcW/1/gUaZwdu9uRcEQi09kc7cq4IBFqXEWDvAHPfdE00TjHHUldlh5jL6/nKeKQNMWlQU5ZRnTOO9WlHwS2GT7uxQX0ezP3jI2UOTPsUmHIezPrklzU+5NNfdpgD88Mf6jyY+pkvzdNhZnNfdpn+Up0C0zYRJhM2WJkDU8Qan2vVwFcD91paTLXKZ1ttRdNUq9ftJlotJlttTTHTKuPs8FRnDAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAIC3y/7Ju/OjnH8a+Y+W/67yr2b+YeOXJv7dil9bsPfrnoUz0/8e/eQ8CIaPt535na+/F/+yha8/jBu+5kv12Q1ffwQPD/x/K39aN/b+PHj8Qvv9/l0faKDfrwZ71Vg7CYZ/Q4wdZu1QUXYcPFJIaYy1B0XacfARRtpwWUh7H9xCRztuBtlIcDREw2zbPK/kQHASoPtvTShb59EjVLTnYSB6M4RZp0A75YMmRHTp0XjjPEfT7NY4z3q9CyQa3dKNvV4P+WzXvBbO4AzOdt0ZnY6WsySZzdIpnLVimjoKSwpnjTE2c2rsrrUNORs5GlI4swTZ0nHekLRNOKOJYyChcKZn6RhZwpmW1LEwgjNdy3SsTOFslcTubGaxTQipb4WEhNluUpC/EuYvmN/Riz3z3jHfG27f2dRxnhto1HVdWcG42IpcN84EuQVRVsDLX/Bj7eBCfLlXr2Ui97re1p3NmpxZAo0JmogNXptiizQ7K19SKUr4OqNl4XjLzqjTiDnf8LL//rCwwLdoYcAPM5TfZ89IGZb1j2BORLBpQmnM/wSRZSfbdTbS52Vpq6GTMC15fYqtqIwaf9WxV2vBtaYeyvdYrO708yKxIQ5fztlSn8reLdvkaLwivJYLZkpGTlzGiM2Zp3XmeZPsk4h0YZZ1c/yPTrbqTNedJXeMcjhNrB3aWHRi87HUMS8bXUOcjY0fhHbnorTs6eLwJZ3pcn+uTHVpLj0RNWF2YiqCgyqeuLNaasHDiz9n0Tg3tPY49rUDxFP5YZDVD2NrzhIqerGUGasegZpLMy9PwhxhscZ6GaKEgDJurg59nqFHGpvGVL8SwO4rccY6sbtUp8zijFeF8pZGeU9GuJRxK2e+IXElvq/PNV6nM2fEbKX8Rz3NtRQXpkRkEN5MIzkm2NomTz3GvjbXyMrNdZHml5/G1ttmUpPGqKcfljGAN8sJEeOY6NB8pTa2MWCib4DlaEwtY4C18IuPmyOdMuv5ICZmEcvg4tmGmkLYnIW6UGH9vwyluSYKJ2UJf90jgc3mtCPRpXU5HcQiK5K18YokrdkZ0XVJRfz4SgtXE94iRt1wq85qfdco7Xg2KC4OAIno3GkbZ0SbT5DsqMgrC4aR7yl/yAtZ4rv+Aefax+hJ0+FmYi0ukoNxcaAeaY64tcfoulDhWvy575bdfVz+Ip2Xg264ZWdpkzP7mVpa1n9RPU9jdabNNWjs1s72eIrcYm+07hm09c85rhVmrHvJjhJZ2/I8L6wcPuYUWUL2fGJKNEgcRYtYeRMvJtW9UUxewXna0Tph9kbPbdvPOs76cKbrRizDwE5+wbmR74Sfc472jTszRlqym9dUbeYaF7202Y5eGbSpa6nSt/EV+kad9emyGmvp7l60t8lrQ6e5tiSd7vJ9epu+Bpky+jsOrtuGMzh7vc5wj1h3Z7gXsRv8XsR3Q3jowtUhv4V/ChHtEfdW76ND68LNgE8WcYzpNTpk7XKuiP0LjAIdw4z3aF8go22ikU8YdI5hoO0AcFZMSniESGvDd3XauP3z4BYDQVP3fzk4q8y0dxwMP8Ka3Vhlmj0Rau+C4PH2y9/d+atc+LoR/icfYs0Wvnbmu3x8lwtfn8flw6AWZPlUq+cXwTAY6viq5f7rvYHP959b8GcD12y9vuaPJj5df2rDlXhcNTO4GlQ5OzwwTlH7Ojjgq/ixusvEvixz8EOoWvo/aS69LkeOBHwAAAAASUVORK5CYII="></image></g><g transform="matrix(1.3937883377075195,0,0,1.3937883377075195,204.76449584960938,341.3288879394531)" opacity="0.78" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path stroke-linecap="round" stroke-linejoin="round" fill-opacity="0" stroke-dasharray=" 2 4" stroke-dashoffset="180" stroke="rgb(69,81,189)" stroke-opacity="1" stroke-width="1" d="M0 0"></path></g></g><g transform="matrix(1,0,0,1,23.738006591796875,309.98101806640625)" opacity="0.6018093333338205" style="display: none;"><g opacity="1" transform="matrix(1,0,0,1,153.46299743652344,162.51199340820312)"><path fill="rgb(126,125,192)" fill-opacity="1" d=" M-4.631999969482422,4.052999973297119 C-4.631999969482422,4.052999973297119 4.631999969482422,4.052999973297119 4.631999969482422,4.052999973297119 C4.631999969482422,4.052999973297119 4.631999969482422,0 4.631999969482422,0 C4.631999969482422,0 5.789999961853027,0 5.789999961853027,0 C5.789999961853027,0 5.789999961853027,4.631999969482422 5.789999961853027,4.631999969482422 C5.789999961853027,4.785999774932861 5.729000091552734,4.933000087738037 5.620999813079834,5.041999816894531 C5.51200008392334,5.151000022888184 5.364999771118164,5.210999965667725 5.210999965667725,5.210999965667725 C5.210999965667725,5.210999965667725 -5.210999965667725,5.210999965667725 -5.210999965667725,5.210999965667725 C-5.364999771118164,5.210999965667725 -5.511000156402588,5.151000022888184 -5.619999885559082,5.041999816894531 C-5.729000091552734,4.933000087738037 -5.789999961853027,4.785999774932861 -5.789999961853027,4.631999969482422 C-5.789999961853027,4.631999969482422 -5.789999961853027,0 -5.789999961853027,0 C-5.789999961853027,0 -4.631999969482422,0 -4.631999969482422,0 C-4.631999969482422,0 -4.631999969482422,4.052999973297119 -4.631999969482422,4.052999973297119z M0.5789999961853027,-1.7369999885559082 C0.5789999961853027,-1.7369999885559082 0.5789999961853027,2.315999984741211 0.5789999961853027,2.315999984741211 C0.5789999961853027,2.315999984741211 -0.5789999961853027,2.315999984741211 -0.5789999961853027,2.315999984741211 C-0.5789999961853027,2.315999984741211 -0.5789999961853027,-1.7369999885559082 -0.5789999961853027,-1.7369999885559082 C-0.5789999961853027,-1.7369999885559082 -3.4739999771118164,-1.7369999885559082 -3.4739999771118164,-1.7369999885559082 C-3.4739999771118164,-1.7369999885559082 0,-5.210999965667725 0,-5.210999965667725 C0,-5.210999965667725 3.4739999771118164,-1.7369999885559082 3.4739999771118164,-1.7369999885559082 C3.4739999771118164,-1.7369999885559082 0.5789999961853027,-1.7369999885559082 0.5789999961853027,-1.7369999885559082z"></path></g><g opacity="1" transform="matrix(1,0,0,1,153.58999633789062,182.7760009765625)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-38.974998474121094,2.815999984741211 C-39.525001525878906,2.815999984741211 -40.02399826049805,2.7100000381469727 -40.47100067138672,2.496999979019165 C-40.9109992980957,2.2769999504089355 -41.26300048828125,1.9429999589920044 -41.527000427246094,1.496000051498413 C-41.784000396728516,1.0490000247955322 -41.9119987487793,0.47999998927116394 -41.9119987487793,-0.20900000631809235 C-41.9119987487793,-0.20900000631809235 -41.9119987487793,-5.015999794006348 -41.9119987487793,-5.015999794006348 C-41.9119987487793,-5.015999794006348 -40.74599838256836,-5.015999794006348 -40.74599838256836,-5.015999794006348 C-40.74599838256836,-5.015999794006348 -40.74599838256836,-0.1979999989271164 -40.74599838256836,-0.1979999989271164 C-40.74599838256836,0.47699999809265137 -40.584999084472656,0.972000002861023 -40.262001037597656,1.2869999408721924 C-39.93899917602539,1.6019999980926514 -39.50299835205078,1.7599999904632568 -38.952999114990234,1.7599999904632568 C-38.40999984741211,1.7599999904632568 -37.97800064086914,1.6019999980926514 -37.654998779296875,1.2869999408721924 C-37.332000732421875,0.972000002861023 -37.17100143432617,0.47699999809265137 -37.17100143432617,-0.1979999989271164 C-37.17100143432617,-0.1979999989271164 -37.17100143432617,-5.015999794006348 -37.17100143432617,-5.015999794006348 C-37.17100143432617,-5.015999794006348 -36.005001068115234,-5.015999794006348 -36.005001068115234,-5.015999794006348 C-36.005001068115234,-5.015999794006348 -36.005001068115234,-0.20900000631809235 -36.005001068115234,-0.20900000631809235 C-36.005001068115234,0.47999998927116394 -36.14099884033203,1.0490000247955322 -36.4119987487793,1.496000051498413 C-36.67599868774414,1.9429999589920044 -37.03499984741211,2.2769999504089355 -37.4900016784668,2.496999979019165 C-37.9370002746582,2.7100000381469727 -38.43199920654297,2.815999984741211 -38.974998474121094,2.815999984741211z M-34.974998474121094,5.104000091552734 C-34.974998474121094,5.104000091552734 -34.974998474121094,-2.7720000743865967 -34.974998474121094,-2.7720000743865967 C-34.974998474121094,-2.7720000743865967 -33.93000030517578,-2.7720000743865967 -33.93000030517578,-2.7720000743865967 C-33.93000030517578,-2.7720000743865967 -33.808998107910156,-1.9249999523162842 -33.808998107910156,-1.9249999523162842 C-33.632999420166016,-2.181999921798706 -33.39099884033203,-2.4089999198913574 -33.08300018310547,-2.6070001125335693 C-32.775001525878906,-2.805000066757202 -32.37900161743164,-2.9040000438690186 -31.895000457763672,-2.9040000438690186 C-31.367000579833984,-2.9040000438690186 -30.902000427246094,-2.7790000438690186 -30.499000549316406,-2.5299999713897705 C-30.09600067138672,-2.2809998989105225 -29.77899932861328,-1.940000057220459 -29.552000045776367,-1.5069999694824219 C-29.316999435424805,-1.0740000009536743 -29.201000213623047,-0.5830000042915344 -29.201000213623047,-0.032999999821186066 C-29.201000213623047,0.5170000195503235 -29.316999435424805,1.0080000162124634 -29.552000045776367,1.440999984741211 C-29.77899932861328,1.8660000562667847 -30.09600067138672,2.2039999961853027 -30.499000549316406,2.453000068664551 C-30.902000427246094,2.694999933242798 -31.371000289916992,2.815999984741211 -31.9060001373291,2.815999984741211 C-32.33100128173828,2.815999984741211 -32.709999084472656,2.7320001125335693 -33.040000915527344,2.562999963760376 C-33.362998962402344,2.3940000534057617 -33.61800003051758,2.1559998989105225 -33.808998107910156,1.8480000495910645 C-33.808998107910156,1.8480000495910645 -33.808998107910156,5.104000091552734 -33.808998107910156,5.104000091552734 C-33.808998107910156,5.104000091552734 -34.974998474121094,5.104000091552734 -34.974998474121094,5.104000091552734z M-32.104000091552734,1.8040000200271606 C-31.604999542236328,1.8040000200271606 -31.19499969482422,1.6349999904632568 -30.871999740600586,1.2979999780654907 C-30.548999786376953,0.953000009059906 -30.388999938964844,0.5059999823570251 -30.388999938964844,-0.04399999976158142 C-30.388999938964844,-0.40299999713897705 -30.461000442504883,-0.722000002861023 -30.607999801635742,-1.0010000467300415 C-30.7549991607666,-1.2799999713897705 -30.957000732421875,-1.496000051498413 -31.214000701904297,-1.649999976158142 C-31.47100067138672,-1.8109999895095825 -31.767000198364258,-1.8919999599456787 -32.104000091552734,-1.8919999599456787 C-32.60300064086914,-1.8919999599456787 -33.013999938964844,-1.7200000286102295 -33.33700180053711,-1.375 C-33.652000427246094,-1.0299999713897705 -33.808998107910156,-0.5870000123977661 -33.808998107910156,-0.04399999976158142 C-33.808998107910156,0.5059999823570251 -33.652000427246094,0.953000009059906 -33.33700180053711,1.2979999780654907 C-33.013999938964844,1.6349999904632568 -32.60300064086914,1.8040000200271606 -32.104000091552734,1.8040000200271606z M-28.361000061035156,2.684000015258789 C-28.361000061035156,2.684000015258789 -28.361000061035156,-5.236000061035156 -28.361000061035156,-5.236000061035156 C-28.361000061035156,-5.236000061035156 -27.19499969482422,-5.236000061035156 -27.19499969482422,-5.236000061035156 C-27.19499969482422,-5.236000061035156 -27.19499969482422,2.684000015258789 -27.19499969482422,2.684000015258789 C-27.19499969482422,2.684000015258789 -28.361000061035156,2.684000015258789 -28.361000061035156,2.684000015258789z M-23.589000701904297,2.815999984741211 C-24.110000610351562,2.815999984741211 -24.57900047302246,2.6989998817443848 -24.996999740600586,2.4639999866485596 C-25.408000946044922,2.2219998836517334 -25.733999252319336,1.8880000114440918 -25.97599983215332,1.4630000591278076 C-26.218000411987305,1.0299999713897705 -26.339000701904297,0.527999997138977 -26.339000701904297,-0.04399999976158142 C-26.339000701904297,-0.6159999966621399 -26.218000411987305,-1.1150000095367432 -25.97599983215332,-1.5399999618530273 C-25.726999282836914,-1.9730000495910645 -25.39299964904785,-2.305999994277954 -24.975000381469727,-2.5409998893737793 C-24.55699920654297,-2.7829999923706055 -24.090999603271484,-2.9040000438690186 -23.577999114990234,-2.9040000438690186 C-23.05699920654297,-2.9040000438690186 -22.591999053955078,-2.7829999923706055 -22.180999755859375,-2.5409998893737793 C-21.76300048828125,-2.305999994277954 -21.433000564575195,-1.9730000495910645 -21.19099998474121,-1.5399999618530273 C-20.941999435424805,-1.1150000095367432 -20.816999435424805,-0.6159999966621399 -20.816999435424805,-0.04399999976158142 C-20.816999435424805,0.527999997138977 -20.941999435424805,1.0299999713897705 -21.19099998474121,1.4630000591278076 C-21.433000564575195,1.8880000114440918 -21.76300048828125,2.2219998836517334 -22.180999755859375,2.4639999866485596 C-22.599000930786133,2.6989998817443848 -23.06800079345703,2.815999984741211 -23.589000701904297,2.815999984741211z M-23.589000701904297,1.815000057220459 C-23.309999465942383,1.815000057220459 -23.054000854492188,1.7450000047683716 -22.819000244140625,1.6059999465942383 C-22.57699966430664,1.4670000076293945 -22.382999420166016,1.2610000371932983 -22.236000061035156,0.9900000095367432 C-22.089000701904297,0.7110000252723694 -22.016000747680664,0.367000013589859 -22.016000747680664,-0.04399999976158142 C-22.016000747680664,-0.45500001311302185 -22.089000701904297,-0.7960000038146973 -22.236000061035156,-1.0670000314712524 C-22.375,-1.3459999561309814 -22.56599998474121,-1.5549999475479126 -22.808000564575195,-1.694000005722046 C-23.042999267578125,-1.8329999446868896 -23.298999786376953,-1.902999997138977 -23.577999114990234,-1.902999997138977 C-23.85700035095215,-1.902999997138977 -24.117000579833984,-1.8329999446868896 -24.358999252319336,-1.694000005722046 C-24.5939998626709,-1.5549999475479126 -24.784000396728516,-1.3459999561309814 -24.930999755859375,-1.0670000314712524 C-25.077999114990234,-0.7960000038146973 -25.150999069213867,-0.45500001311302185 -25.150999069213867,-0.04399999976158142 C-25.150999069213867,0.367000013589859 -25.077999114990234,0.7110000252723694 -24.930999755859375,0.9900000095367432 C-24.784000396728516,1.2610000371932983 -24.5939998626709,1.4670000076293945 -24.358999252319336,1.6059999465942383 C-24.124000549316406,1.7450000047683716 -23.868000030517578,1.815000057220459 -23.589000701904297,1.815000057220459z M-18.1299991607666,2.815999984741211 C-18.591999053955078,2.815999984741211 -18.972000122070312,2.739000082015991 -19.273000717163086,2.5850000381469727 C-19.573999404907227,2.430999994277954 -19.79800033569336,2.2290000915527344 -19.94499969482422,1.9800000190734863 C-20.091999053955078,1.7230000495910645 -20.165000915527344,1.4450000524520874 -20.165000915527344,1.1440000534057617 C-20.165000915527344,0.6159999966621399 -19.958999633789062,0.1979999989271164 -19.54800033569336,-0.10999999940395355 C-19.136999130249023,-0.4180000126361847 -18.551000595092773,-0.5720000267028809 -17.788000106811523,-0.5720000267028809 C-17.788000106811523,-0.5720000267028809 -16.357999801635742,-0.5720000267028809 -16.357999801635742,-0.5720000267028809 C-16.357999801635742,-0.5720000267028809 -16.357999801635742,-0.6710000038146973 -16.357999801635742,-0.6710000038146973 C-16.357999801635742,-1.0959999561309814 -16.475000381469727,-1.4149999618530273 -16.709999084472656,-1.628000020980835 C-16.937000274658203,-1.840999960899353 -17.235000610351562,-1.9470000267028809 -17.601999282836914,-1.9470000267028809 C-17.924999237060547,-1.9470000267028809 -18.207000732421875,-1.8660000562667847 -18.448999404907227,-1.7050000429153442 C-18.68400001525879,-1.5509999990463257 -18.82699966430664,-1.3200000524520874 -18.878000259399414,-1.0119999647140503 C-18.878000259399414,-1.0119999647140503 -20.04400062561035,-1.0119999647140503 -20.04400062561035,-1.0119999647140503 C-20.006999969482422,-1.4079999923706055 -19.874000549316406,-1.7450000047683716 -19.64699935913086,-2.0239999294281006 C-19.41200065612793,-2.309999942779541 -19.1200008392334,-2.5260000228881836 -18.76799964904785,-2.6730000972747803 C-18.409000396728516,-2.8269999027252197 -18.014999389648438,-2.9040000438690186 -17.59000015258789,-2.9040000438690186 C-16.82699966430664,-2.9040000438690186 -16.238000869750977,-2.7019999027252197 -15.819999694824219,-2.2990000247955322 C-15.402000427246094,-1.902999997138977 -15.192999839782715,-1.3600000143051147 -15.192999839782715,-0.6710000038146973 C-15.192999839782715,-0.6710000038146973 -15.192999839782715,2.684000015258789 -15.192999839782715,2.684000015258789 C-15.192999839782715,2.684000015258789 -16.20400047302246,2.684000015258789 -16.20400047302246,2.684000015258789 C-16.20400047302246,2.684000015258789 -16.30299949645996,1.7489999532699585 -16.30299949645996,1.7489999532699585 C-16.457000732421875,2.049999952316284 -16.68199920654297,2.302999973297119 -16.975000381469727,2.507999897003174 C-17.26799964904785,2.7130000591278076 -17.652999877929688,2.815999984741211 -18.1299991607666,2.815999984741211z M-17.898000717163086,1.8700000047683716 C-17.58300018310547,1.8700000047683716 -17.319000244140625,1.7970000505447388 -17.106000900268555,1.649999976158142 C-16.88599967956543,1.496000051498413 -16.716999053955078,1.2940000295639038 -16.600000381469727,1.0449999570846558 C-16.475000381469727,0.7960000038146973 -16.398000717163086,0.5210000276565552 -16.368999481201172,0.2199999988079071 C-16.368999481201172,0.2199999988079071 -17.66699981689453,0.2199999988079071 -17.66699981689453,0.2199999988079071 C-18.128999710083008,0.2199999988079071 -18.459999084472656,0.3009999990463257 -18.658000946044922,0.4620000123977661 C-18.849000930786133,0.6230000257492065 -18.94300079345703,0.824999988079071 -18.94300079345703,1.0670000314712524 C-18.94300079345703,1.315999984741211 -18.851999282836914,1.5140000581741333 -18.66900062561035,1.6610000133514404 C-18.47800064086914,1.7999999523162842 -18.22100067138672,1.8700000047683716 -17.898000717163086,1.8700000047683716z M-11.769000053405762,2.815999984741211 C-12.29699993133545,2.815999984741211 -12.762999534606934,2.690999984741211 -13.166000366210938,2.441999912261963 C-13.569000244140625,2.193000078201294 -13.885000228881836,1.8519999980926514 -14.112000465393066,1.4190000295639038 C-14.33899974822998,0.9860000014305115 -14.45300006866455,0.4950000047683716 -14.45300006866455,-0.054999999701976776 C-14.45300006866455,-0.6050000190734863 -14.33899974822998,-1.093000054359436 -14.112000465393066,-1.5180000066757202 C-13.885000228881836,-1.9509999752044678 -13.569000244140625,-2.2880001068115234 -13.166000366210938,-2.5299999713897705 C-12.755000114440918,-2.7790000438690186 -12.28600025177002,-2.9040000438690186 -11.758000373840332,-2.9040000438690186 C-11.324999809265137,-2.9040000438690186 -10.947999954223633,-2.819999933242798 -10.625,-2.6510000228881836 C-10.295000076293945,-2.4820001125335693 -10.038000106811523,-2.24399995803833 -9.854999542236328,-1.9359999895095825 C-9.854999542236328,-1.9359999895095825 -9.854999542236328,-5.236000061035156 -9.854999542236328,-5.236000061035156 C-9.854999542236328,-5.236000061035156 -8.689000129699707,-5.236000061035156 -8.689000129699707,-5.236000061035156 C-8.689000129699707,-5.236000061035156 -8.689000129699707,2.684000015258789 -8.689000129699707,2.684000015258789 C-8.689000129699707,2.684000015258789 -9.734000205993652,2.684000015258789 -9.734000205993652,2.684000015258789 C-9.734000205993652,2.684000015258789 -9.854999542236328,1.8370000123977661 -9.854999542236328,1.8370000123977661 C-10.031000137329102,2.0940001010894775 -10.27299976348877,2.321000099182129 -10.581000328063965,2.5190000534057617 C-10.888999938964844,2.7170000076293945 -11.28499984741211,2.815999984741211 -11.769000053405762,2.815999984741211z M-11.5600004196167,1.8040000200271606 C-11.060999870300293,1.8040000200271606 -10.654000282287598,1.6319999694824219 -10.33899974822998,1.2869999408721924 C-10.015999794006348,0.9419999718666077 -9.854999542236328,0.49900001287460327 -9.854999542236328,-0.04399999976158142 C-9.854999542236328,-0.593999981880188 -10.015999794006348,-1.0379999876022339 -10.33899974822998,-1.375 C-10.654000282287598,-1.7200000286102295 -11.060999870300293,-1.8919999599456787 -11.5600004196167,-1.8919999599456787 C-12.059000015258789,-1.8919999599456787 -12.468999862670898,-1.7200000286102295 -12.791999816894531,-1.375 C-13.114999771118164,-1.0379999876022339 -13.276000022888184,-0.593999981880188 -13.276000022888184,-0.04399999976158142 C-13.276000022888184,0.3149999976158142 -13.20300006866455,0.6340000033378601 -13.055999755859375,0.9129999876022339 C-12.909000396728516,1.1920000314712524 -12.708000183105469,1.4119999408721924 -12.451000213623047,1.5729999542236328 C-12.187000274658203,1.7269999980926514 -11.890000343322754,1.8040000200271606 -11.5600004196167,1.8040000200271606z M-5.247000217437744,2.684000015258789 C-5.247000217437744,2.684000015258789 -5.247000217437744,-5.015999794006348 -5.247000217437744,-5.015999794006348 C-5.247000217437744,-5.015999794006348 -4.080999851226807,-5.015999794006348 -4.080999851226807,-5.015999794006348 C-4.080999851226807,-5.015999794006348 -4.080999851226807,1.7599999904632568 -4.080999851226807,1.7599999904632568 C-4.080999851226807,1.7599999904632568 -0.5830000042915344,1.7599999904632568 -0.5830000042915344,1.7599999904632568 C-0.5830000042915344,1.7599999904632568 -0.5830000042915344,2.684000015258789 -0.5830000042915344,2.684000015258789 C-0.5830000042915344,2.684000015258789 -5.247000217437744,2.684000015258789 -5.247000217437744,2.684000015258789z M2.5209999084472656,2.815999984741211 C2.000999927520752,2.815999984741211 1.531999945640564,2.6989998817443848 1.1139999628067017,2.4639999866485596 C0.703000009059906,2.2219998836517334 0.37700000405311584,1.8880000114440918 0.13500000536441803,1.4630000591278076 C-0.10700000077486038,1.0299999713897705 -0.2280000001192093,0.527999997138977 -0.2280000001192093,-0.04399999976158142 C-0.2280000001192093,-0.6159999966621399 -0.10700000077486038,-1.1150000095367432 0.13500000536441803,-1.5399999618530273 C0.3840000033378601,-1.9730000495910645 0.7179999947547913,-2.305999994277954 1.1360000371932983,-2.5409998893737793 C1.5540000200271606,-2.7829999923706055 2.0190000534057617,-2.9040000438690186 2.5320000648498535,-2.9040000438690186 C3.052999973297119,-2.9040000438690186 3.5190000534057617,-2.7829999923706055 3.928999900817871,-2.5409998893737793 C4.3470001220703125,-2.305999994277954 4.677000045776367,-1.9730000495910645 4.919000148773193,-1.5399999618530273 C5.169000148773193,-1.1150000095367432 5.293000221252441,-0.6159999966621399 5.293000221252441,-0.04399999976158142 C5.293000221252441,0.527999997138977 5.169000148773193,1.0299999713897705 4.919000148773193,1.4630000591278076 C4.677000045776367,1.8880000114440918 4.3470001220703125,2.2219998836517334 3.928999900817871,2.4639999866485596 C3.510999917984009,2.6989998817443848 3.0420000553131104,2.815999984741211 2.5209999084472656,2.815999984741211z M2.5209999084472656,1.815000057220459 C2.799999952316284,1.815000057220459 3.056999921798706,1.7450000047683716 3.2909998893737793,1.6059999465942383 C3.5329999923706055,1.4670000076293945 3.7279999256134033,1.2610000371932983 3.874000072479248,0.9900000095367432 C4.020999908447266,0.7110000252723694 4.093999862670898,0.367000013589859 4.093999862670898,-0.04399999976158142 C4.093999862670898,-0.45500001311302185 4.020999908447266,-0.7960000038146973 3.874000072479248,-1.0670000314712524 C3.734999895095825,-1.3459999561309814 3.5439999103546143,-1.5549999475479126 3.302000045776367,-1.694000005722046 C3.068000078201294,-1.8329999446868896 2.811000108718872,-1.902999997138977 2.5320000648498535,-1.902999997138977 C2.253999948501587,-1.902999997138977 1.9930000305175781,-1.8329999446868896 1.7510000467300415,-1.694000005722046 C1.5169999599456787,-1.5549999475479126 1.3270000219345093,-1.3459999561309814 1.1799999475479126,-1.0670000314712524 C1.0329999923706055,-0.7960000038146973 0.9599999785423279,-0.45500001311302185 0.9599999785423279,-0.04399999976158142 C0.9599999785423279,0.367000013589859 1.0329999923706055,0.7110000252723694 1.1799999475479126,0.9900000095367432 C1.3270000219345093,1.2610000371932983 1.5169999599456787,1.4670000076293945 1.7510000467300415,1.6059999465942383 C1.9859999418258667,1.7450000047683716 2.243000030517578,1.815000057220459 2.5209999084472656,1.815000057220459z M8.074999809265137,1.0119999647140503 C7.789000034332275,1.0119999647140503 7.525000095367432,0.9789999723434448 7.2829999923706055,0.9129999876022339 C7.2829999923706055,0.9129999876022339 6.809999942779541,1.375 6.809999942779541,1.375 C6.890999794006348,1.4259999990463257 6.989999771118164,1.4700000286102295 7.10699987411499,1.5069999694824219 C7.224999904632568,1.5440000295639038 7.389999866485596,1.5770000219345093 7.6020002365112305,1.6059999465942383 C7.815000057220459,1.6349999904632568 8.104999542236328,1.6649999618530273 8.470999717712402,1.694000005722046 C9.196999549865723,1.7599999904632568 9.722000122070312,1.9359999895095825 10.043999671936035,2.2219998836517334 C10.366999626159668,2.500999927520752 10.527999877929688,2.875 10.527999877929688,3.3440001010894775 C10.527999877929688,3.6670000553131104 10.4399995803833,3.9709999561309814 10.263999938964844,4.256999969482422 C10.095999717712402,4.550000190734863 9.82800006866455,4.784999847412109 9.461000442504883,4.960999965667725 C9.10200023651123,5.144000053405762 8.640000343322754,5.236000061035156 8.074999809265137,5.236000061035156 C7.313000202178955,5.236000061035156 6.692999839782715,5.089000225067139 6.216000080108643,4.796000003814697 C5.747000217437744,4.510000228881836 5.51200008392334,4.073999881744385 5.51200008392334,3.486999988555908 C5.51200008392334,3.259999990463257 5.571000099182129,3.0320000648498535 5.688000202178955,2.805000066757202 C5.813000202178955,2.5850000381469727 6.006999969482422,2.375999927520752 6.270999908447266,2.177999973297119 C6.117000102996826,2.111999988555908 5.98199987411499,2.0420000553131104 5.863999843597412,1.968999981880188 C5.754000186920166,1.8880000114440918 5.65500020980835,1.8079999685287476 5.566999912261963,1.7269999980926514 C5.566999912261963,1.7269999980926514 5.566999912261963,1.4630000591278076 5.566999912261963,1.4630000591278076 C5.566999912261963,1.4630000591278076 6.513000011444092,0.4950000047683716 6.513000011444092,0.4950000047683716 C6.0879998207092285,0.12800000607967377 5.875,-0.35199999809265137 5.875,-0.9459999799728394 C5.875,-1.3049999475479126 5.960000038146973,-1.6319999694824219 6.127999782562256,-1.9249999523162842 C6.303999900817871,-2.2260000705718994 6.557000160217285,-2.4639999866485596 6.88700008392334,-2.640000104904175 C7.2170000076293945,-2.815999984741211 7.61299991607666,-2.9040000438690186 8.074999809265137,-2.9040000438690186 C8.383000373840332,-2.9040000438690186 8.668999671936035,-2.859999895095825 8.932999610900879,-2.7720000743865967 C8.932999610900879,-2.7720000743865967 10.968000411987305,-2.7720000743865967 10.968000411987305,-2.7720000743865967 C10.968000411987305,-2.7720000743865967 10.968000411987305,-2.002000093460083 10.968000411987305,-2.002000093460083 C10.968000411987305,-2.002000093460083 10,-1.9470000267028809 10,-1.9470000267028809 C10.175999641418457,-1.6460000276565552 10.263999938964844,-1.312999963760376 10.263999938964844,-0.9459999799728394 C10.263999938964844,-0.5789999961853027 10.175999641418457,-0.24899999797344208 10,0.04399999976158142 C9.831999778747559,0.3370000123977661 9.581999778747559,0.5720000267028809 9.251999855041504,0.7480000257492065 C8.930000305175781,0.9240000247955322 8.536999702453613,1.0119999647140503 8.074999809265137,1.0119999647140503z M8.074999809265137,0.0989999994635582 C8.413000106811523,0.0989999994635582 8.684000015258789,0.010999999940395355 8.888999938964844,-0.16500000655651093 C9.10200023651123,-0.3479999899864197 9.208000183105469,-0.6050000190734863 9.208000183105469,-0.9350000023841858 C9.208000183105469,-1.2719999551773071 9.10200023651123,-1.5290000438690186 8.888999938964844,-1.7050000429153442 C8.684000015258789,-1.88100004196167 8.413000106811523,-1.968999981880188 8.074999809265137,-1.968999981880188 C7.730999946594238,-1.968999981880188 7.452000141143799,-1.88100004196167 7.238999843597412,-1.7050000429153442 C7.033999919891357,-1.5290000438690186 6.931000232696533,-1.2719999551773071 6.931000232696533,-0.9350000023841858 C6.931000232696533,-0.6050000190734863 7.033999919891357,-0.3479999899864197 7.238999843597412,-0.16500000655651093 C7.452000141143799,0.010999999940395355 7.730999946594238,0.0989999994635582 8.074999809265137,0.0989999994635582z M6.611999988555908,3.377000093460083 C6.611999988555908,3.691999912261963 6.751999855041504,3.927000045776367 7.03000020980835,4.080999851226807 C7.309000015258789,4.242000102996826 7.6570000648498535,4.322999954223633 8.074999809265137,4.322999954223633 C8.479000091552734,4.322999954223633 8.805000305175781,4.235000133514404 9.053999900817871,4.059000015258789 C9.303999900817871,3.890000104904175 9.428000450134277,3.6630001068115234 9.428000450134277,3.377000093460083 C9.428000450134277,3.1640000343322754 9.343999862670898,2.9809999465942383 9.175000190734863,2.8269999027252197 C9.006999969482422,2.680000066757202 8.687999725341797,2.5889999866485596 8.218000411987305,2.552000045776367 C7.866000175476074,2.5299999713897705 7.554999828338623,2.496999979019165 7.2829999923706055,2.453000068664551 C7.0269999504089355,2.5920000076293945 6.85099983215332,2.743000030517578 6.755000114440918,2.9040000438690186 C6.659999847412109,3.065000057220459 6.611999988555908,3.2230000495910645 6.611999988555908,3.377000093460083z M13.923999786376953,2.815999984741211 C13.402999877929688,2.815999984741211 12.934000015258789,2.6989998817443848 12.515999794006348,2.4639999866485596 C12.104999542236328,2.2219998836517334 11.779000282287598,1.8880000114440918 11.536999702453613,1.4630000591278076 C11.295000076293945,1.0299999713897705 11.173999786376953,0.527999997138977 11.173999786376953,-0.04399999976158142 C11.173999786376953,-0.6159999966621399 11.295000076293945,-1.1150000095367432 11.536999702453613,-1.5399999618530273 C11.78600025177002,-1.9730000495910645 12.119999885559082,-2.305999994277954 12.538000106811523,-2.5409998893737793 C12.956000328063965,-2.7829999923706055 13.420999526977539,-2.9040000438690186 13.9350004196167,-2.9040000438690186 C14.454999923706055,-2.9040000438690186 14.920999526977539,-2.7829999923706055 15.331999778747559,-2.5409998893737793 C15.75,-2.305999994277954 16.079999923706055,-1.9730000495910645 16.32200050354004,-1.5399999618530273 C16.570999145507812,-1.1150000095367432 16.695999145507812,-0.6159999966621399 16.695999145507812,-0.04399999976158142 C16.695999145507812,0.527999997138977 16.570999145507812,1.0299999713897705 16.32200050354004,1.4630000591278076 C16.079999923706055,1.8880000114440918 15.75,2.2219998836517334 15.331999778747559,2.4639999866485596 C14.913999557495117,2.6989998817443848 14.444000244140625,2.815999984741211 13.923999786376953,2.815999984741211z M13.923999786376953,1.815000057220459 C14.20199966430664,1.815000057220459 14.458999633789062,1.7450000047683716 14.694000244140625,1.6059999465942383 C14.935999870300293,1.4670000076293945 15.130000114440918,1.2610000371932983 15.277000427246094,0.9900000095367432 C15.42300033569336,0.7110000252723694 15.496999740600586,0.367000013589859 15.496999740600586,-0.04399999976158142 C15.496999740600586,-0.45500001311302185 15.42300033569336,-0.7960000038146973 15.277000427246094,-1.0670000314712524 C15.13700008392334,-1.3459999561309814 14.946999549865723,-1.5549999475479126 14.704999923706055,-1.694000005722046 C14.470000267028809,-1.8329999446868896 14.213000297546387,-1.902999997138977 13.9350004196167,-1.902999997138977 C13.656000137329102,-1.902999997138977 13.395999908447266,-1.8329999446868896 13.154000282287598,-1.694000005722046 C12.918999671936035,-1.5549999475479126 12.727999687194824,-1.3459999561309814 12.581999778747559,-1.0670000314712524 C12.4350004196167,-0.7960000038146973 12.362000465393066,-0.45500001311302185 12.362000465393066,-0.04399999976158142 C12.362000465393066,0.367000013589859 12.4350004196167,0.7110000252723694 12.581999778747559,0.9900000095367432 C12.727999687194824,1.2610000371932983 12.918999671936035,1.4670000076293945 13.154000282287598,1.6059999465942383 C13.387999534606934,1.7450000047683716 13.645000457763672,1.815000057220459 13.923999786376953,1.815000057220459z M19.92799949645996,2.684000015258789 C19.92799949645996,2.684000015258789 19.92799949645996,-5.015999794006348 19.92799949645996,-5.015999794006348 C19.92799949645996,-5.015999794006348 21.0939998626709,-5.015999794006348 21.0939998626709,-5.015999794006348 C21.0939998626709,-5.015999794006348 21.0939998626709,-1.715999960899353 21.0939998626709,-1.715999960899353 C21.0939998626709,-1.715999960899353 24.822999954223633,-1.715999960899353 24.822999954223633,-1.715999960899353 C24.822999954223633,-1.715999960899353 24.822999954223633,-5.015999794006348 24.822999954223633,-5.015999794006348 C24.822999954223633,-5.015999794006348 25.98900032043457,-5.015999794006348 25.98900032043457,-5.015999794006348 C25.98900032043457,-5.015999794006348 25.98900032043457,2.684000015258789 25.98900032043457,2.684000015258789 C25.98900032043457,2.684000015258789 24.822999954223633,2.684000015258789 24.822999954223633,2.684000015258789 C24.822999954223633,2.684000015258789 24.822999954223633,-0.7590000033378601 24.822999954223633,-0.7590000033378601 C24.822999954223633,-0.7590000033378601 21.0939998626709,-0.7590000033378601 21.0939998626709,-0.7590000033378601 C21.0939998626709,-0.7590000033378601 21.0939998626709,2.684000015258789 21.0939998626709,2.684000015258789 C21.0939998626709,2.684000015258789 19.92799949645996,2.684000015258789 19.92799949645996,2.684000015258789z M29.641000747680664,2.815999984741211 C29.106000900268555,2.815999984741211 28.628999710083008,2.6989998817443848 28.211000442504883,2.4639999866485596 C27.801000595092773,2.2219998836517334 27.47800064086914,1.8880000114440918 27.243000030517578,1.4630000591278076 C27.009000778198242,1.0379999876022339 26.891000747680664,0.5429999828338623 26.891000747680664,-0.02199999988079071 C26.891000747680664,-0.593999981880188 27.0049991607666,-1.0959999561309814 27.23200035095215,-1.5290000438690186 C27.466999053955078,-1.9620000123977661 27.790000915527344,-2.2990000247955322 28.200000762939453,-2.5409998893737793 C28.618000030517578,-2.7829999923706055 29.101999282836914,-2.9040000438690186 29.652000427246094,-2.9040000438690186 C30.187999725341797,-2.9040000438690186 30.652999877929688,-2.7829999923706055 31.048999786376953,-2.5409998893737793 C31.44499969482422,-2.305999994277954 31.753000259399414,-1.9910000562667847 31.972999572753906,-1.5950000286102295 C32.19300079345703,-1.1990000009536743 32.303001403808594,-0.7630000114440918 32.303001403808594,-0.28600001335144043 C32.303001403808594,-0.21299999952316284 32.29999923706055,-0.13199999928474426 32.29199981689453,-0.04399999976158142 C32.29199981689453,0.03700000047683716 32.28900146484375,0.12800000607967377 32.28099822998047,0.23100000619888306 C32.28099822998047,0.23100000619888306 28.03499984741211,0.23100000619888306 28.03499984741211,0.23100000619888306 C28.07200050354004,0.7590000033378601 28.243999481201172,1.1619999408721924 28.552000045776367,1.440999984741211 C28.868000030517578,1.7120000123977661 29.231000900268555,1.8480000495910645 29.641000747680664,1.8480000495910645 C29.97100067138672,1.8480000495910645 30.246000289916992,1.774999976158142 30.465999603271484,1.628000020980835 C30.694000244140625,1.4739999771118164 30.86199951171875,1.2690000534057617 30.972000122070312,1.0119999647140503 C30.972000122070312,1.0119999647140503 32.13800048828125,1.0119999647140503 32.13800048828125,1.0119999647140503 C31.992000579833984,1.524999976158142 31.697999954223633,1.9539999961853027 31.257999420166016,2.2990000247955322 C30.826000213623047,2.6440000534057617 30.28700065612793,2.815999984741211 29.641000747680664,2.815999984741211z M29.641000747680664,-1.9470000267028809 C29.253000259399414,-1.9470000267028809 28.908000946044922,-1.8300000429153442 28.60700035095215,-1.5950000286102295 C28.30699920654297,-1.3680000305175781 28.12299919128418,-1.0230000019073486 28.05699920654297,-0.5609999895095825 C28.05699920654297,-0.5609999895095825 31.136999130249023,-0.5609999895095825 31.136999130249023,-0.5609999895095825 C31.114999771118164,-0.9860000014305115 30.96500015258789,-1.3240000009536743 30.68600082397461,-1.5729999542236328 C30.408000946044922,-1.8220000267028809 30.05900001525879,-1.9470000267028809 29.641000747680664,-1.9470000267028809z M33.090999603271484,2.684000015258789 C33.090999603271484,2.684000015258789 33.090999603271484,-2.7720000743865967 33.090999603271484,-2.7720000743865967 C33.090999603271484,-2.7720000743865967 34.13600158691406,-2.7720000743865967 34.13600158691406,-2.7720000743865967 C34.13600158691406,-2.7720000743865967 34.23500061035156,-1.7380000352859497 34.23500061035156,-1.7380000352859497 C34.42599868774414,-2.0969998836517334 34.689998626708984,-2.380000114440918 35.027000427246094,-2.5850000381469727 C35.37200164794922,-2.7980000972747803 35.7859992980957,-2.9040000438690186 36.27000045776367,-2.9040000438690186 C36.27000045776367,-2.9040000438690186 36.27000045776367,-1.6829999685287476 36.27000045776367,-1.6829999685287476 C36.27000045776367,-1.6829999685287476 35.95100021362305,-1.6829999685287476 35.95100021362305,-1.6829999685287476 C35.62900161743164,-1.6829999685287476 35.3390007019043,-1.628000020980835 35.082000732421875,-1.5180000066757202 C34.83300018310547,-1.4149999618530273 34.63100051879883,-1.2359999418258667 34.47700119018555,-0.9789999723434448 C34.33100128173828,-0.7300000190734863 34.25699996948242,-0.38100001215934753 34.25699996948242,0.06599999964237213 C34.25699996948242,0.06599999964237213 34.25699996948242,2.684000015258789 34.25699996948242,2.684000015258789 C34.25699996948242,2.684000015258789 33.090999603271484,2.684000015258789 33.090999603271484,2.684000015258789z M39.25,2.815999984741211 C38.7140007019043,2.815999984741211 38.237998962402344,2.6989998817443848 37.81999969482422,2.4639999866485596 C37.409000396728516,2.2219998836517334 37.08599853515625,1.8880000114440918 36.85200119018555,1.4630000591278076 C36.617000579833984,1.0379999876022339 36.5,0.5429999828338623 36.5,-0.02199999988079071 C36.5,-0.593999981880188 36.612998962402344,-1.0959999561309814 36.840999603271484,-1.5290000438690186 C37.07500076293945,-1.9620000123977661 37.39799880981445,-2.2990000247955322 37.808998107910156,-2.5409998893737793 C38.22700119018555,-2.7829999923706055 38.71099853515625,-2.9040000438690186 39.26100158691406,-2.9040000438690186 C39.79600143432617,-2.9040000438690186 40.262001037597656,-2.7829999923706055 40.65800094604492,-2.5409998893737793 C41.05400085449219,-2.305999994277954 41.36199951171875,-1.9910000562667847 41.582000732421875,-1.5950000286102295 C41.801998138427734,-1.1990000009536743 41.9119987487793,-0.7630000114440918 41.9119987487793,-0.28600001335144043 C41.9119987487793,-0.21299999952316284 41.90800094604492,-0.13199999928474426 41.9010009765625,-0.04399999976158142 C41.9010009765625,0.03700000047683716 41.89699935913086,0.12800000607967377 41.88999938964844,0.23100000619888306 C41.88999938964844,0.23100000619888306 37.64400100708008,0.23100000619888306 37.64400100708008,0.23100000619888306 C37.68000030517578,0.7590000033378601 37.85300064086914,1.1619999408721924 38.1609992980957,1.440999984741211 C38.47600173950195,1.7120000123977661 38.8390007019043,1.8480000495910645 39.25,1.8480000495910645 C39.58000183105469,1.8480000495910645 39.85499954223633,1.774999976158142 40.07500076293945,1.628000020980835 C40.301998138427734,1.4739999771118164 40.47100067138672,1.2690000534057617 40.58100128173828,1.0119999647140503 C40.58100128173828,1.0119999647140503 41.74700164794922,1.0119999647140503 41.74700164794922,1.0119999647140503 C41.599998474121094,1.524999976158142 41.30699920654297,1.9539999961853027 40.867000579833984,2.2990000247955322 C40.433998107910156,2.6440000534057617 39.89500045776367,2.815999984741211 39.25,2.815999984741211z M39.25,-1.9470000267028809 C38.861000061035156,-1.9470000267028809 38.51599884033203,-1.8300000429153442 38.215999603271484,-1.5950000286102295 C37.915000915527344,-1.3680000305175781 37.731998443603516,-1.0230000019073486 37.66600036621094,-0.5609999895095825 C37.66600036621094,-0.5609999895095825 40.74599838256836,-0.5609999895095825 40.74599838256836,-0.5609999895095825 C40.7239990234375,-0.9860000014305115 40.573001861572266,-1.3240000009536743 40.29499816894531,-1.5729999542236328 C40.01599884033203,-1.8220000267028809 39.667999267578125,-1.9470000267028809 39.25,-1.9470000267028809z"></path></g></g><g clip-path="url(#__lottie_element_872)" transform="matrix(0.29999932646751404,0,0,0.29999932646751404,111.72709655761719,307.7491455078125)" opacity="0" style="display: none;"><g transform="matrix(0.9999990463256836,0,0,0.9999990463256836,-300.2005920410156,-56.11981964111328)" opacity="1" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><g opacity="0.38" transform="matrix(1,0,0,1,422.4020080566406,183.8209991455078)"></g></g></g><g class="png" transform="matrix(0.6660000085830688,0,0,0.6660000085830688,23.63299560546875,35.459999084472656)" opacity="1" style="display: block;"><image width="296px" height="213px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAASgAAADVCAMAAADTop1YAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAADnUExURUdwTA0hVQoeThAjVEZRfRwoWgwhUUZIgEFKekRIfQ8kWBEfVhUlViEaWxQbUxgaVRgnWg8dUhUeWBscWRMmWR0ZVyUdXykgYywkZxgtYC8mbDQqcDkudiIqZT80fiAkYCo2YtHU3lBbgP+JTf9qTfvpTT5Kd/lN6//KTf+qTWz/UalQ57T3TY/7Tf9NqNnyTcJP6d1O7E348/9Nf03Z+HZd9PpQVf9Nz02/+pJU6l//db3Bz02Y/E10/1H/1lhl/WdwkYmRqlv/kk2r+02G/lj/p1T/vf9obaWrvk1tUn5DVCdjiWcvhV5pCsgAAAAKdFJOUwD///+r//9/P9Pxgf6aAAAgAElEQVR42uSby3LjOBJFuwATttlYoDroBRcKUVzwJ7SRdvr/LxrkA8hMkNSjyl0zEQPZkiXTqtCpmxcXCfKvv1bj4/3tc/7/HZ9vb+8ffz0c72/52GVapv+1MfIdfm3eRvzOo8s3vMMHesRvHhPd0Rvxl4xlWYDWA0wAqfvlcVT39lX9bY+mkeieR6SBj758Re/rUx4ebvwF3wF+F+gpj+DNwL/Ch74f+iGPfuyHcexHM6bM6u3jLqbuz49Et8yny7eYHxNA6uLO8OXOF1axoAlMJtSfQ31ixuB9H4c6+jHfpZbVnqre/iuYEBQhQkop8rMY91ixvISP0VCGUumEQM/akf+yj35gUQ0jfI1jwntBNX9uierjc57+MKFIdzHhfUdqilx4AikpIXHx+VJ5nmoRP3wQRJ4Q8U+BXxdiPWhqGJSmiNdoVbXM72tORU5x72N9fT+kep+o6FhMVHbdhpKUYUX2HF1WkSsucP0FhiS6CurIPkvKK0j5brSaGtflVzjFlz/rL2OiP09ACVAhoq7y6XZKz4stsS/DLUTwceQTpOhCYFqeRWWqcEBRZVi9wGo45TF/butJM/j6dhm1gBlSRBWliHZeDarFpPhUPRVXioEBFQEVeypf6ifB1COqXiSFmCyqyWrqc1mVWNwsuq/v9afUdVJtBky3N9VJIogehUT3IdbKKvUWpPYqtnbu6wdvPGpcaWrSPvU2f59KnnemCJSKhgqrLqr6U2Kqrl3KDfBguUXiUEEEMafCyYnEavUNQInKDyH14zCsMaFPfUjh7Qkm/gokTUO5UTSHYNUlnOrYvbv95KQnu2JKpeRYURKgNCTzg+Y00De6OeVOSlPjKnsqm/r4XP54KKAY3pXYxLXWPU5OHJ6KfYM5xeLURTXRNcUnT0pOCFh3UHgmS2E+WImqFN/7fFCppmt8PH5/UohUeCkymxTb6c4mgvUc5z27k0z5dbbTMrKDcXklq95rk6KIsNLUMrOTT/er7etJPvFutUYqOElOpKfKqNua50ROkUOTCuCR7CmKTGTSU5WHDlXnQO3oPUapYaiaAqNK43rpR5L6mC2U+Ku58WlrIxFJbNqOA3rdy17uWU+cxJFULEkycNTcEZOoigWF6RxJ+aEXTW3YOUvqbfkjE171dXxQkanbZ6SXvnpNRx+3SilIwAQWaFEyXANMqjUrKvaG0lY+wIgAEx8v8b7uL1u+XgAX9zhGaRcU467m1G02CaqRe0kJ4k1l8WvMWmNxW6Liqa/nJOX7aui1ALdq72P+LkL7MosFX+q45lKJBHeqjhOBb90pqPVdyVD44V1hAZpyRlSGUqm+vidMXlMaNjxqXHJCeF++aU57pgRTJEWVouOV8F7ZebUExpDpuU1Q2k2l4WQF42rNubb26iJH+blEBGxLbSQpNKlNi4rfnpvImXDBkmgJ/KAzV9cr0Xabioi8aqM0TqWU5ISXURTfo6T6gUnVbN6vWGWT+nyhC/X1m/XH7oQ6YhOXBlS0YYBzuKzsgkQC5eghyEpuS0fIiu8bVsql+ro2HvotRQGo2q776v6V2S/KMgaSU1cbvarZ21lKNWBG08VU4bLIIshk5yQwMS7n1qUntk66BJuSXgvkKP5eh/MHfc2v7zEo7M1lRrWfst1IkZLzaqZrGklB+k26R+C0niodp7QlpNQ7NqlzYEr9q6B+U1MSximKR942SNvLOio4XqnE2t8uvcugoqXhZWOTc6Xo+CfnGkPnBU/vudGpKQ0bFjUuDah/o09HjSesuRRNF5M0ldp1XVnVmdZcSQVqtRKEkxfRKEEhKFV9rl3z+R9oVICKd6+QUZs4D2tQltfX98gspY4XdqnsQ3XrWa+oKMp+QZTcpDNTned82IrgrCPHPuWCU+IyoZNg9dhGYFHBJl+/1RF+vxvMf78Uad8AeipkTnu98Nrj9d63eop1oy6UNsH2mk5NeCwmJlR9fbWY6cmlBornI7ak1knqsKeo70GkNzeRjXTFt/cNOGXarbogfQKvNgrqisX5lW2zfogQ+pOSlBNHD2hTyAn0VGe+tHbz+V8CpbbJY+q4O5fSnU3NumCJZjtTpXAVqpUjZVCuwRQcl5wTXbkaqCQfmH5L2V/nzNk/MPPdMc2vA00FEZ1PwE4+Te12OVn4tCxR6q6uenXdya4KfTvl4EZMVUtSeM7ZiC59Kao8L7sxKbWFd7ijqDjf1Jjm0+nyYuVFIsURk3LCcj2fTqfbdVoL6no6t72UaAKPKroCyG+s5lx1cE2rPGnsvKAye1Zjs1/8QFGXkxovgqrdlBIw2Ziu9Q0v7T6UJ1C1+srGgVrTeZXF8cGF7aoLTiCJqNqFDLzPj7LeM7tWyUx6B/jOoPZO85nmZZ4zrcucf+heAMUb5OjgHWUCtvDM6XyZlwV4zW3tZVDVn4p320xeP5/TycC1sancFK6irrC35Bv6ui4e173gwzg98qgMin69vKKoVE9QSZFTAXzlN7tNWIb53c5jtGc6AajmzCa9/G36ck735FxQoUDpyaiq8fOiJwxSvd0xbtPmYzOPFRQpalLymybdnynP6JFWvykfntT5KGeiQ/IpksrHUy4gUHnk/75OK2nqhJANT0hLZv3qTS7o6gt6+jOlh8mco7k6U2OD1eEJRS1VUfMNjJjBwc+nKzyZz7fucoaq6iZ8vGJrLsHP+cXaKJiVM03zJc9xccRjbgt7FGJBu79OQOqW3zIfMflgTySovuSUfTtVbiIlrShXzYqY/wilca765rANo/f2DtWj7oH6RyvqRjZ8ntjpz2d6cqm/uvDjFSoPjAgxjBSeQEWTnPIUANe5WHsBFZby0pw/w/l0hXecbIMAlGS6A7VBt0PJ1p9udUo80KQMJqWowzMeNeP/fDddqQTxSdbS6UbQsrTmMx+SP9uS0gVeTFC7FzX/2xie/+IyRmCzMKiM7jx3Pr/XeQoZVP4HrpcuNKXnKxiuvGaGs7Oe/E4XX7XAXqWDYVyZ1IFo3Zn1jEdB3ZzxJWRz4yf51RlAXRlqFlhKM0gqf+Ir2vgVjAnN/KZBZU5w4MWTsG4eQYUrSin6XOlXUNTpEtRmuW0QmDAQ9oVkIOrISRaFMwibVMK6s/nggLenZ70aD67AKD+Z8AxrYHQBWh3KDGsuAaP8SWc8GRlYJqsor+Y5/s1pJEXdABmMTDWEM1ajiU6rdkqVCrv2fVSmh1DiZl7jlXMU02Bz+aGQemLWW4qiBNSkoqgBdcFTVACUjquXWHHoJmaW2I3CZX7zhUChkPJw+eAug7rZMM4VV1nVCe6xnlw77XmxqL6PPWMas6j0Oa8Hqr3DrykKLOrK41JnRgAFERxAYWYqhywECnCIPxVQIHwBdSZQwStQW92UYBYp4RlIW80WJhWrRyU6hzopKydUr3iUAXUxx8wCKpGiiIrZswMrKk0C+BsqPY81WEoP3QoUxKV3K9sAWw06RSg0j3dwmS6LaR0MIKVEsNJ4FIN6RVGL8ahq5tc83RlQ0EoBRYGZUxy/1vAkMyDkggloXTA6VTMHIS3wKcDvQqsotf43afupoUu2bMMUQdWzqBM7VNIzXgG1vJDMC6iZgifM4uRjoKQJbBvcHBTVMZWLTpkZz3UZKWZmRB1kAe+XG051Eg+Ch8QwiaLWvd5gnDs8icqUnt7YiXU5nIBV0rX3Ox61CpygqESK6sijIsSpMxxyq+Xna8AEz84lBzzgEBQWJfP5VF4KYe1R4k7VpfZS00MvLzEKL4uJg4oHWHdHUdNLirKl1+GahWRFM2NKpfRIURnOlZYwo+4SzIQqL1pw9TthlAdZ+bqEudEB0CQQUE5n8Borw7NikuUfjp8ql+O1HjUdJKq+NkUdHoHaau2WsZSeJ54DzedjknoSX9KyLLnQmtYvXOXVSXuuW5apuRAqHzGtuuAWUghNDndPznvlrX7Wf432WSuoMZGPJ5LUQQXOX77sDHYx6zmrpptZJrtu+3wne1mYV1dD7bUJTBNTFna63/S46myDUxtUFD2RQQ1lztOKeqYXfrzfxUx0Cp1cfLB/XpiXvfJyUpjuYZqTeze3o6TqKqvwfPWFWnvqv4odCiANY9XU8WmP4usQj+aV46oWE+8h8Bk9ZQ/B7iCUky/86iQeVXY7ePQOgguqhamYPVl6BZNWFLi5EtSQpGNe5PTQo453RJXsZgtttNTLEu0FZOZqTUsrWH/aY+VET+GFRNA0DuD2k1HJPkbk2ktcdzTnHUvfgEjtgTq+cN0mIeITngjTxll09prWDT15s8W5O2zP8oUIJVPeT3RzuOXx99+ZU6TlMJw6fRyOJCkqPBhw/4yZcwkeWj3FIqVYzitIdeO8PGyg2iAVvN6RWnk5pyfZKZf0+IKoeBoATv7nT32NcQ3lWHygqSOZVIGlS+/4ip5SrbwkVwOnchZGjCs9/Yeza+1tG1eisAXpgy3LlK3EVnpVBUiKNE1v2jSp2xR9AE1b4Pr2//+e5TxIDh+SnR3KdpJdLLqnZ86cGVFMlpXhPuhIn5wRTGZdUTzLhocw1dO6EMv83czLOSCFlFIu9zj1GKSIUf1z0o/ZlDNEytQ9t0ssepQlmXD2dp1jVLIHnkYu6jlRI6cILQRKU2pOQAFSq7LRf27CqtcaZcveoJj3IUJd/JS+crdZcoMVPTatUmyynMqycqDguSnmAJvE9oFUf1ccAqk29/QApqywiTefA06rkhlFNqr3KUVCJYCS8JwPcKrP/YzzohRppyJnkFJw+ajvgIRPzVz8uZ1daKBqHYhTTTo+JyGn3FvR37OmU6u40eudhzq+henCwxwMoaQvQAlXaUqZbfWxNLntKcMeSu6bez5KlWEVBSo51zvLqGbFedf0ivLOS7zzw0D1gd8UzskdeyHoNJB2ccqJniXL5CbDocalCB350SBplGCBQmlOEZ+g7BFKiJNOPavlrE+YeybxJFCdQyI4GqSPt/M4pErpCAaeQkh4giLL5CPRgXua+gAV07EbdkNNX40wWahqVPJsSoSqAaVsTslXrhAjzaoeGBWkXidTbwiWjtUcUWxNrSNO5ZJNxmeOPUiWLHlyn2E25Avc/sI0XOFuA5ZvwypMuSm/QKMqzSdQKVPzVLNCRvVyHNV3naBUMvW6RFvnNXeekpuzVNRwvUvbcfcI8KgVt21euOciQSiJUW1IZQjFDkrjpAlVo4XimqdL3soqlIYIL5InzjwJVJeobvqzjT050cmdVVCOKlSZHehaBoByXd2B3EsAWFtGVbgg60wUVVFlWaVBqhCplf7DNyU5KHpB/8JIpYDy8o6waZP7DR2RciHnKT8uhgZZSqcKcURBEqqp3FBQDGwmkIapIIxqA5FhF3AKkhBRqsu6nmuoKjIHK1ioUUCpts+RUhormXtRC9MFlsBHTLlZQW4USiUmBbEzGEi60elKhE3SShWhAyekap9RgBOgVIBC1RlKeVnNV+g2S4QJlFz1us6bktdaQp0boLo8CVakT0rZTWI4LVCy4JXUNR3sXIQ0ZdmYNKUhKUb8eM2UqixI1h2g3dRI1VWGC/hk3KYiiVKIlGKkOpDz2B50Xr/SCbjaZObR9kwVKFQTaXj4aHkmjypKjQrc7aji4KC3kJueiE7mVZkvTamr64o5hThVgNJ8hZRSzChAie1B7wlUoFGdFKmWftBKNy7PdLBDlRCjJngSOD1VKUa8+NS/DVwM3R6fJqwTLWRSPWU/QNUPvgHRKphRmYYK0w77F82oNem5KXtd70GVrHqST70cZBqgrNksS+XPCsyBevlzjvnLadloS3ly3zMC/hfj2OgLYgsrGVrLVwSSzjt9odv0ofI0yiHUDZW7XFQ5pZQ4QKzMz/e7S4qLywsb1xfXFK85Xpl4R/GW478Ubzi+QPz8+fP3799//vz5ruMDxwPEZ4inp6evOn79+vVNxw+IjxCPGDcY72/eU9xRXHF8wrjFuL/f/yU+5SDmALf2B52MhOFsDWat+VopfzRuks90weDYynODEsCkgbo0MBFQr2EBTLAQJlgaJlgIEyL1Bhbi9IZwioB68HF6QpwsUA6nmxsPqLs7ROoKlsWJYdJA6dgzUmwOwB4kNKr1RLv10q4V/Z2AirnEmrS/vNwxTIxUQKnXTKlXr31GvWVKEVAGp5BRDqkHhOqzhupJMIqR+ghIPRqoHhOMurrzGPVJQLVHQqmeE7hLMco0LMaG962FSIX3EZzfhJlgQ1jtdpeSURIkTj0GyuLEjILUe+dwskDFjDKJ92GAUT9CRj16jHpvKYUwXUWMur/fbXOScmcO4tRrXQvcce61wbRABZmHjEJC5XsEykmUACtIPQmVxyhLqQOMYonyGfXN5B5TyiXfe0uplEbdfnJI7e53u03LjAJbLjPvTIq5MOGddxshNc5U0jXtQ5gMTteeSCUZxUAFGoWESmnUw4dIo76GGvUxxaikRsnMA6T2eW8YRWoe24Mul/0vSFZPnyo180WgDE55vgsIlap6hlAho5xIxalnGfVdI/VBIPX5IdaoHwapx7GqZzQKcy8CaveCcq/rcg1OHzGqF95A94RywML3D/JIn6jWwSr3RqF2IVLXHk7XCUYJLX8biXmy6nka9eRpOWUeIXXjazlrVKDlUqQAp92u6ZlQXVqjpHsysq5f1o9HeQcqbmi1w/BhuhyqegcYZXF6M6pRgNRT4KO+JX0UQ3VnNEoyylLq3lJqY0yrr+Zn/ijYc5pDroDzDgNv258TSLbqydy7vhipej5MgY/6Ml71HpJV76MlVMgoU/SuYo26J0oRo/Zc84BQjNSZ58w9U26SL8w5kXq0Kxuzb490EowKDMJF0nAmoPJSb4BREqfYRx3PKEmoW0coBApqXu8plGSU3xNLC+UH3ihsSJ7IRO0x8S59Z26MeSTmBqd3vj0INerLkEbFVe/XVz/zRhllcbqSdlOIlOqN3exjjerSg+CITbCFqAQmEZUQqHJhGEVQXVwGLmoAqBCpgxr1wfMHCWf+4xhGScN562sUAUUNjGv1zugCoGxH7Pq71k3qEtGAlJNAcdELGZXUcpN6rtkb06g3/6rqQdkzjEJKBYxy9uBTyCjCaTfrOfn83LMtTBdnX6jjdksaAcUwlWufUUGzdz3Y68W5N9LrfT9Q9X7FuTfKKF+kRNXb0+SAQdpaUvljFtEdt2pAylnFS/MyNippOEc0KtDyY3s9IeZH9XoDGvUp6mBE1ZNqvsWiR4zq7K1Orna9ygezDnnVkJzzRDP3bFSQewKn6wMadVyv53qYhI/ypwc3QVcc5d5t7Dhf8CzKM1E+o7rw1l2KTtZDlaWFqtx5Rkrag2vPHwRTljGN+vLvnHk8jxLOfKTqOUpxzSOoNpB7Z1KjROL12OAxWqEpoLZF3mppyll5PsKo4V4vXfXSvZ6oesnpge31xPTg5jHFKNnD3Ma5ZzNPsElUvb61eddao5lwB7jF2KcTN3tOpC4Gjfl1am435KOOZNTIhJOHB0HmeR1MnHmCTppPAirWqF4MNlVqAMU4md97weYAUSqbLPcZ5c8Phnu9V/486qhe72GQUd/c9EAyys+9K1/OQ0ZtvcwDKd/6GsX3hXuqecpzBqX1BqThpsvDW1LwVTab5btdYnYgGXVIy5/b68Ua9cNND2Tm+RqVwskChUpucm+DjIKiB8s5c2aUErsw/GJnnk1mUtm3rJlpxPIhRoW93iF/kO71vqcmnJ8TE06vh/EmnBapeB5lxPyFhanfOD5R+r1ke6Btu9igqRI9nmqCIMMJJ+3i71fZpxiFZe+46UEo5oc16skw6qtw5pJRj0ktvxtw5vtNv+iFLd8YG3Wmr3MWc9EHq7QrQGPQWJkilBp63g2PN6m823qXg/f1DiJ11Mw8vK+XdJwy9UJGhdOD/X963J9Bvhxh2pI5wCnLlsUcbYHdhhHt+W1s7jVWwjFmJRzXYQ8nzs/3XvyPL7H4Oib+T+tQvIQFVyL+mjUW+79/t+v+dOHuLANMG9PBkEgJH9UKmJRSQ5lXuhlLY89On5ntGLTpYpLZg9Fge0ld200Gpaaefq/whL9Kf117R9LRe02PGoj9hfhfxYNqZ/PZZD6vKx3Laqmjgt2Yk3ldV3Wt3/U3K4jlyVJf82W1qvQ3K/1v4D9Zw1qsFxBrHSf5ycn65CTv132+6E+tRhm3acT8jDWK7uNpmWpT9kk1gUBRyWvo98/M5P6LyQRe5q2YTApY0Q6Mwp6pUqRPs8fNFf7pdZnZLW6jwst8qcGreUHQux9LwFbDti5x5Xqt80UOrwVn3ikwamNr3pbnLGw4W2yCzZ6VgdGKZ6L4t89kaA4kVhOCa5JpgOADYTJQTZKP37kToGjfPey2pO3zABb8pIKfTH2MqoqRImjqqmKMaoGS+VzyO5FwtVwv1xSA1FqL+MISqrNukwmFpEKNas1mzdbcc0nknRJDcnrIpkGI8MAOQSeEamKiiBgVnC06Fdt/62JqGVO4TZewS66qp5XAiL5hQnk8SsLk+FRpkMrlWiO1WmtKIVKaTeu1KXin/Wm3QU5pgDYi9XxGJZ/cYJRKZRmFMOHBcASSTL5JBNWkSEAVpFxtoOKHoIg/TCPY3GQ2QbtEqyWjDKlqi840mXbEqPUSKXWCqQeMgrWQGsUNzHbLlNqiRpEd7xVSijbvJ90mj8ixw2MmzczJZ/jFzOSdTjzMvAJhGtgOPahNRS338OKzKzLl7CdSi2DyYlql5Mkmnk49k3WoUAuAyTFq02+BUb1tX2ixmCsc1B2h41zzgE7abM4yq08z5hPDRPKEuZcl9rAmeIXP8xRTkXpGlyyb+LOyKs5UsqmXhAjZtLRVclkCm/ANsVoQVhqsU8Mo9lCI1BZxIkbZ2y3tqDw5N85aDgJFhwpZajFWGYk4ClSUedNpKg2nhlCGVO6jKkiUMpKkgrZjOk2qqwOxJKwAqJJSb32yzunSdDKMWpCJ2nTAp44yjy3ndvuSnXnLs98YpX8Yu5YlV3ElGLKiWUg2tDDgVzvcq/v/n3hVT5WE6DPYxpyeWWVkZmWVBG45FYBKlKICuzmB5IhPak/DfzxOYlLJSi4VyQFOiE6+UkLZLJAsXKcOTKQ6ItQV/WmiEJUJhThdLgITEupzv1GMIkbdn080c2Pijz/5JKyin1qjcsePuZaKx2iBOTn/JzSGTaS6YWhwEphYbgOexiP3/oNQnEwFKKh76E/rBWJmFt6lKI9C1PsGON3JzE3gfBwF8mVPp5DVR7/JGrwgVaIm8eo4FRzjZqDBO1YG5pLn+3wEMG8IlA5r3E53iFIW3gofxGmlcHClGIWM2sjGYW+seBRb1Js8qjdXodvbdbaiXg4PhAu0eTxYW6IsTlVvEOU1AjwdQJUKTr6ilZeopFFABedLAk9/+pNojkveyuLLBgW9TA4FWXsAlTLqQ7q73SkegPhIeu/vgwBlRis0+yV/CtAKE0aaNUV0nsPT8QbyLlqJ0zjfd5j0w4QCPqkGuV85bFRaPjFCk1gUePkX2HluXsDLL+jlGzMKu5fb/X4jSj1pzoJmzrOVx39wp4Wfmrew6DQbOJMyPXctXPcOScQWLkbeHH608ZKo5LFvga8R/5UOsZHoZHMm4bSC8lB90BujQdHLeNSH+ASMYi8HRv0+LJceNmbuRgb8i1hIJByv5IwZND0RVIyRgcg1ACX6SypPtkhpsLHANr0jWJUXIxf/Tn9aEp85OllGrWjkCBM2xAATFr2PWhR1xIjTU1sYZBTGp0dV8zp0AjZhNA/YvgSJmQia0AkNyldWLhcnx6pzJ8WIQKuyQOVPaOF+TCYVVJXOH0A1FTaZQI44rezl6zXXvAvqDumU3xtCBQWPsbpzOJB48CCEBKqlH8rP8shvfmasD8ahvKETe/meUCf8ryf9QxLATLeC8IiJj2aQMtbdb5dTBRsLExv5SBOqDBO+MpngAyhdL7aBQUpxNGCoOJn/vvuZoFpyWXCmsvDDh3W0gl/OVc3wQPmgHarAP0/EppMTRjlKAXiPfa06jw7uK4DEnvwhnyZRHI0KZLbHfEI6TQjTyiZ1FfFlh9poFkVmjoy647z8CZRSoB7tjpWO7nS0shgnz9AE6fJ8k8kLXAARiA9plapU0MQBKHwME1PKjxWZ/EG1U70xgdifVHcr8ulMSIHowMxxaHAhnDKjbtuNGXW7Y83Lbo7KE4/qLwgvZU53Djg04N9YC9TfcZcXvE41PdU8Fp7O605OGKV2TpKT9+DbGQpwydPYEqFiNhFA52MTn2x0sj6+ElYw+b0im648Es5WvmE+2Fh5N9UeTVmQVVj1evtVyqowJXJcmVqQSwoRMSmoRxFMNpfrVWZU5pRD8z4BTJEDQsfB8XZ7yUoSmeTcUuls49LY2lJ9rOJRMCWHaICTA4Bqw3SQgdpgypI7PY5RaOQgvicz6vGPBMVo4c/Q4bsMDRwzymPK9IPbDzLRmgbj45idnNS6QTDyXOH4TXhp+PbtANNm75EGKSq9cYcQvr5gJeGKylsvTKiNhbeR8rDs6Y19nKL2jFpKfjq3OAX62SKZ/4bSw8ggSlOUG6qwydHAAUKOG2DAyeEZFk+8bYIxGKSxGfKqfc+FS1Nx8B6FGt2h9K6kPTSpDQzqupFHUc17g0nhHZDv0r/8KqNq3fUyFP5MptCJrUlqn1PV+U4YJ4dCX8o4Jodkci6lju5Ic2PiW6QzRFFa4LgvdZPySWYEY+tMDafWleIBsYktShh1I06RlaNHoZc/368doxbdemi7FsXpzD8CqYMoF5yQaZC5pjDKIUQn/KIIlcCdsOg5xsgxpzzRig5YnMt4ZYQYm1jZEXNq1rxk11cEpTqQr0Z84OZq5GRUG7zIy5FO5Z5aMPJfkh7Fg0eBqbfLAFHyhUy8ppCBKlMoTpmmxXMmZQ7S/SVMDzzpdS2lZBkTVjhj+kfT+x8UZ8udoESRPAsOAhSghEsvpCqpL8QAABaZSURBVLuNSh5ZObJJLOrF0lse1TpCz8zhgfs0OwjVIErBogTltNQRpRx/J7hOxKfkCkiu+BPBhNQ6hGg6AzqzgjTamdw0NbitUukm0R2uC2ewLpf1g5T6IKOQUAAT3qZ953DwJocCnH41cPJDz/sWhdicQygzzSD1zvEPPJS+Rb5OJRc4yk+QyAWnqFbeZnIdocQitL36dNQ0WpmNNY/GtVYeHpfVWBTmAspQ2Z+g6FHJu5FFPTEc/L6QUa1HdQcsgdUnysNIQNJzgtMgK55MIvrgGcFCbxqc64zFhVDApngousm2KaNgtM+WghGCtVYocdWjCcv2xRa1sUVtcts/JU3MUBmk4lEcpJbv72XvUCGYaidRHHw8SHfHc3LbCsPVibo7R1UO1OYAJXbw2LcoASgeDiyl0O2sqaZTSZiIkoD1lRnF9e5yISNnqBAmDpvKJ1be8/U/jQeLpKgOUougVOCSxkWN3OjOlbUqItXA9Y7+7MSc8ieSJ0UjPcQoHnr4bBo6i1EF26hymxpCwfGVsfoimC4XzZqbJRQiJRkKkBLpwcNfv7sebshU2hav0jMr5/WKApkTUcpRLBBCeYxRJLNYoIp8igBTOTV0OtNi7zge1ri15Esx8WktUAGj8PhCKtGbGSXLCsinN7MJ1UdAPQyfvjvK00Y4lFl50MUEWaGSFKAvWlIncyJ3csaiIsYA0VyKghLAA2DFys4BGvxu1+kO4GK5dWACpFR62BEDTsAngonY9C7KA5xe4lFLJ2wyn86hIlTITAJoqOyVqOm13En7AgZOdo5IoVEBWCk6Dkt45k+iE+4LM8GSTjNlzNEw6RAprXSrWLlGAyFUPiGdPkon0Z2GTWLUrzDKVr1uyVPdmYkBN8O6y8AiZFjFtDq5gZsWUB59iEWqu8jsiggSWRRfKFZAopkDwTTDdcfEd00wolP5E+FkvFyOj4RyptT9gFFLd2vdUmycHFw3Q9FY05n9PV6ZVPpidKahJAPBSbRGYEVGKCVBiQxqjLNS6zzJ0NLiMx4iZIpd5eVZdexSm+JEfOKidxeLEpB+X3AUM5edBq30llZ2NNx0duNKpbim7JkD+JRl54hEPtKWS1FeMr4UMVoSm2a1ceRTptJZdlv0RMeXe81ZjypAfQir22ejTH67d7IBS+/9bU1q2WFVdmPg4cDIfYVAIRDTiC+c5NBEmFEmcAgMVbkiuSRiYzqJLRX5tRbex2otHFKkphamL8lQm6FUptOtwYlAegqjFh2vfPfSgcQnVJ6QKLgeVIMu6hnnQrXRiY4oICWlUkrRDAsi8YgzATiS7CE4E1LnhkBV71v6umlnUOulxAMV3rZxj/e5y3iFkPp9MVQKlM4MGqxCrT7kFMNUhygzq+NU6XayS0lqHpe86hip2pW0OZd5ylhVOdleqOe1zuE2YU592VUWRS2ezOt2unsZj1r2K1RNKufxEyaCsENhcDaaV/bEAlQ+eSdCK7pLxKdS8VB5I1EJPxNvVN0L7lqBxN9NtZu6NW8rOAlWCBTT6bdSngBV3Rb0XcHkTdkD4QXmUxAgmgkLXxQm8ZSAgCrRqRCKuDUSpWLxpFkYZXZfmr0WvJOgmcsJVHs+7RglwYCNHJu8G4VN41BPw6iCzZFHlcGB0VwBy65PFaC8UR1dxFz0XIEolTPBhHGAfDy/IF3OOH2Cy2nuEKkQSiGzJr52DWptcOLVvIxSbeUvozwbD3qzuiVoMid/go9HiGzlK6udUuYKQmLjiFJkoLjeiQS5aRlZcnSa4oyePukyCxv5yku+63Xaz5smNacGJ+nyNEOp7G63qhluepfGo8SmvtsUhYw6l3zgnCWLRigniZz+oPaUPHp4RscxLFjzLJkiu5PqjkQ3oTnNKrd5Kv9Ym7U6O8U8PqzyNnYoHBrwIOq+J5QcP8KonuaK7phPkDZRewF119yfwIyyBS/yFwDiRHMVRCnGMiyYy1IUc2nsdClX0tsVlsZNYqosfOrjJHwyCYqFV3CiMflv41CvHzFzw6dvEw1C1RA7Z4peafP2mnPO/D8qOVPuTDoY1cYlFXD/i740T5zGx8ma1Dq1Nv5HKthFqM0gdWOc3rcyhto51I+tegxV6HZ5BJVjWu1i1KAarHCKyiq6CyzFBqym2mUzJ4sCbOa2j5t1RwoSCx3KmlH56sJEQwMteBeh01atUEnUrIz8pzCq6+OLndnJHunAO3xqnzK5s+JT4oAQ7SQlVtmA/GkWHzf9CtoUVLxpqksecOm6c6VpPRKdKG+9VMK7Gd3dCqHu+2zww0DhCszS2XlvhcfbWGCTj+PRgatDpWvFhzApHqkxKIYoAkKRvsia5nFuJuKzTQVAJXgrd6bpL2syEHUMiqV3L9FAUSq6+xEzZ4weYuJLs5ywcDOM/bDnfs9XbJI81R4RZedjbHsWHT7NUvUIIATL6K6UPcpMa71Cvv6LUZd2ANzi1GaD+3NX8yyj6mAQStZcmE6wqzVUzXCVyF3ds1RlL9aTubq9A9HNpeQhp+ju1rkQKV9cr7MkctgpDuuYK7Hqn8elOuqKJzDtpwYv6l1+KkYJnapsYBddMGkKUOH/nF3tchs5DrQqqY3DOVo6cj5kSZek7nn2/Z/nNMQ3CI68pzheZ392NRqNBmZ88kIeUAkVHL5sn9OfAmwqyCP8W4quuYJkqmANGjz7Wcr5CzDlPjLQllygir3mA/n0eDBQo9klwSEiAZROnlHS9jp/ME3Y7igd94QqZSqMUUPrXD5Au516l0pxQbsSX76GEXnMxSgUMsrySUOlHfnjAV8I1Ge4VkBGnb4xrWwSJe7pL/VvhRLCNPigkO9gYcG1kjtD3Z0L/1GZCgzDy9c+eenIZDIDazWjhvdAc/BQGtXqL5FAUcebKTWA/VQ3wAQ8EqReobQTiiD6ILTOKORawqs8QAacgmOLr5SdoZOZ8Tal5NzyrjaFAoFSjJrTJ+d0c+LWp/OVE5VfCkAhVv1b69OQT4W+f0ykS5iOC5eo/mrlsQWDFbwFw0OCIzYtxhes2pFv3X6qk3IFlGjUBXmU3BWi4JROX/8AQuPKQ4e5Q4USToTaUWsoVRnsQMhrVRlUxWOL415nYpWu44nPZK/Jw/BNKg8JpTUqDlnIQXHlpVclB6bAg0MmsyhePeXpA957gXXHEgWvcbDpU6XlAQBUa/VeyXQ6V3Qm+rWm/Hp1U57lkzDqolOVPjgY8+mvVzidRkX3hAR+EoUCVuHoghKF73HAh8faP85d0WWFVZYf1Gyn7lbU8AKW3DS8q5lcHqDjDNR/PvtdnpEoMgb/qPrGDa/Zy2/c7YRRykI1VlW2TlR5bVoRJtXOLsnI0oG0cgYlk0u0Gb4biUKYrI9KvE2whEK7+XWopsOeVxqjqOTK9EEAFTQDhccWflCayHUg3FnNdVh2izHksj0/kPL7zc14nlGfJOS4diF3cAKA0j/h03Rioxl3vP2Xa5EbJ06d8a/8o9IHQpXavjsFhwrTwsTuySxckFG642Hh+RjqRmsXkae+9FIYQ6lQ86uMOnSaJfg0HScqNR3neqNn7/EzSAeYVnZg8TEdZr+bz8klh7obDyWVR0Cl6NxAuyizfXnJp7F72quuYUXOQJHprOoPuEQgnduTiPh8hu112ZhwPwLjWd2qIk1lDbpbA6m8h8HJMCrxnznQKOU4T33ii0HeNDLlReRpIvP0wegQPPjT/ioeYhQSamGglIRnXW7ZxL1Qk9LrtDxpX7CxIb+yNbh5r7nD1ICy9iBFxfdSySEd//btoOZAxP3nXNCQF4qeiryQ56wq7kDKs+p1ru7EZkazi/qdXy6FMnT6pUrP5it20AuM1LdRtztqdhN7cRzxuN2pmsPsoCplOqN56r04uSUkUYDSqqtOyTjBtPl9gs7JXelhZJ7EH3TWwCcs/8cQjDNwwCiuvWLDFbKY+FR51b6papEiSrlWx25cDJRLV7pYkzqeF6gnoTyj0pxSJ+YnPjcgbqUYp2OwCjNK113Z39BHdIKGV9GDk46TREWRXOZOl0MdX7VC7fjo9HfTMN1tDqVhsqWX5qFGMTRBBaYxn0r/T6g6hdL+GkMU8tq+zvw2HoBoUbZA4KJZJasgxXnMVRGKj1bUNfl27RyUiuvEGEQaRU/lzYGWK6zSgE4vFApGPEozNaWQUKJLZ+OdegclfsmMK4EVl286MpCed7vKE1T3PjTAlteA+ry4LXpgo/ogWBvQL1Rd6euuMKN2O9Bep9YSg85jVmIWq1TO2h9kn6VYNmklN2mdcZpeyh/688uXHiV3vZSTP7AOHc/vEKWw/oogVUI1R2I1uAIeCVALOynyTDnrld3Sabi/FVtNpinHBhSUGynXdAIxl643qDvxUckSCv/HIFJBF441N4AICUVZig4LuNvBV5CpcJtbegW38iT2yQ/DN7N5iZ0Bld6nv9ec++RAY+QnmlHZFVKmAUjQ7lrZFbDh0ucO9YlFKh80O193u4Hq9523q0Kp9+TEJ9GoZMaYYNSzK5ikURsKVEnIpWkEU8NoLzlsf1UJ1KJMOfiD6iI6c2roYRqrk3KaCqr77R6EBsQnwyimVBggBFp+kBUU7Q5MTPDRFV4pmkfnXp6IUtmkKdllvaHDlH6nTDlZAyvkwUaBGIWGM1zmWWcwMgfTs/BiPsGkclB2jU1gniRO6frdUu2qJZtcJR/OdZm8E+oTy9NmnoXtYNIW6oF8EjEnEe9mmBOlB6k35QnZlACcJDD9qzCTxlC1qkN7wO7p3LMKtTy7CY9UfPHSZCOozYdQMriI11RxnVLyX7+6rpfUpc88HvWS91HhUqqQPM1jlECdoOoGrmDR1iBXiVEG1gnm37yaqcXNwlfd8ChfMTF51PGQUVx0cwrdwWm81ms0Sqd+z1KO2x1pVFX5XBmY8oxYZTvaRXFK9hht/lostJo3s3mxjnyH6aGAsq8VcYTySo5WfXeYaRQTTNMRm8iOF7ZRxc3B2hk8seK0dxA4Sal5sIhOsnVxi2Ev5ErJH+IOoPRiV6C8Zjo5Kcdi3Pk0wbdgcxdL1FlaHrHoxWd/rLyKFegeTxRp6j2mTaDsic81ukHkjeeDyaRKL5mrqM4XmG6Hws4TSxo4gzGdyGtWY82jT64YGeSsI19tCla/tVuj5NfdrZhjKI3T3aYrKOem9PwD+8H8IoWYTlB3KQFUCfAqOLWUQ32qCqIvfBomNdsAKpAnu20J5ekq5+RqxrM7dCFUx6jLTDLe1V802uE1JzS7NJxeXsj4V1CCl/UBo1Tu63dR1oavZgze1FaYFWq76bhO4pV+HH4IVMQoBdJsrxElamkKnnAT3OiE6pReLxH8iEcGKgZsMfnKIsrk1lG4iuorb+vPo8mQb/G+s/PkquNZwxlPefZuM8EFRmMUK1Tyae8rOhU1ubzi1ILF59cILk5xVadgUpGmkihZeIZTHjY8VXmNUUlGvNnct5Irb2ilXZhS4xQcRZ8EoaSAGkV0jNFZe6gBPnUBA4XOQBvMJfdr4LwGHtPIOEcrG6F0IFBGxX3ppaP9C8pVAyjR8IJsSn5/UA7TJ5ZzrLoS1l2myIDYFOxYKMvsO94mFysep51N2y18GvZ+t6u8h8GqAZXYmncLGDlxZTopXTpBy0uyCg5DlXrB7InAOtbyrNWcwzkTGcjjroGSb8KoVSeanTO4q80wFd5dCZRnVBrxicbhhKya2JE3kHasUoHvFNQ9h7znf+eGyewiuq+3vAzyRB5K4aMm4G6/GZ3TGT5JXHflS4NQyj2dGlB/BvmKGCi0CVPjFaxbkpYmWAInfYJBKF1cr+MM6hiqpfI7IEnB7QMt3cmT1iaqu3WzGbmW8eD81xkDq+XtNDGc8CgDZrCg8E4nGvGcTKWmTgmnFibTZa+7Wi5VzCZ9xmBBs1vIQolORVNwHg12ZtUpzkA7KGMO9IzXl97723c5Y4lHPJSp3RTQtoXBIeeUcBAu8LP/XCr5cSXmx1YTfiWCAOTteNTtWJx6S87+6Ro+MmzCFSTTwwPln3vRzoDlvE3A2hSQqsPfp1Thn8GiRdXdi2k4yy/ZaO95WqIVQhepbAansTzpdEUR6uGXeR2j3t7eg80nx5qcqezylNgW7MLOtNpDOhX9Nkol1Kdn2c3Y9Mgb6FzlaHSBt2HVHH/W3EOFE4uNfq/KGtzit2R0owugZKD6+fbWHKcnFA0tzCxAS0cq+0/ztH+VpKmket1FdlIFZOqlL8gkUPDzAvNwD1HurZP4JzcIQ9ltt80w6u5anr028Iz68QTq5+DCTt6YkSgh52SFSy3BknMmZk0ug6qkUAqmr4DVXio6ZtOa11DFiUprV3cwCd/sMdRNCZQd8notf3v7Ee6o1D0wzX1oLhOYp+Yzn9jMuzMQeJJzBhcZfl3XizGC92SKaTo65snmToXYtIbBShdCeQvlcnKL1F55z9q72AcVaBBWeE1No2S2Kw2kVnI7biDjQcPjdnfpWVSsF88sUK3aqsZo6c8KchyOr72Qu5D82sUGUHV3v8mTz39b5T0pdfHTy4ktOfvOHR5I6tA1weSS0I0Ph7sm5Jdqx7ug+DLnT+1V4zmcgI3VzF32hH9XyycVk5vhRc0unlH+8/P9DSg1Wy0/0fkKnfy0X3u7w0PDXUmiUzMNLRP82A16FSuuhO2ORCkzr6DZLbE8rR4mGeuckl95eS6hZvgcR7958aUHhAJKRdOLugdOPLXsCM0TibkcGJh+R4S61NIPwdFeAX5r8G6i0BMsA0+Qc7RpWaPpzl7+mshAadTDX9h5Rv2NCvXWGp+ruoQRFI3ECVkF0iR63gg1T9obzGZ+Kb+LaHkEEfy2H1YnNFA1qDyJft2diswtq1Py66YHF/WeDG8M+usVazbp8/5nji591FicZImAYLXSm6fCMd3squ5CFqpetCyFvpzqDvRpydH6LnKZZr5bOwPlQs3b4KzVOqggYVFIfQaXPgq2icz4jhKCRC0vgSlIUUpXUaPEPxXb8siFM1bIpmUoUOvqnlBcZQhe48HFjS7alT8e/vje4vT3r1/f3/Tnx9yFK3JClphSnD/Nrd+lghpVyEPprA5JRQCpPLPEjNprbnmhT2uYrbjUlw0Uu3I9vpjnONxBeU+pn2/2894mGZ+SC8dw6VJkwtthShOL0xwPw0/9IZ/5m78ppHaBUtaA5ClKVfrjgk3C8bWfg52Oyy2UwuoR3Gse4qR0SkjEFJsAKfblKiuYJDRIesQru4wX1PIdmt/RpJediTosvG5wkRce+pYn2co+umiFcvuEPteMjYFB6nv3jJCMyKhRIk+Y+6ZCuQF0PP7sMNXfcUheAh3P8F2gCk8x/Fsh4TQzSJ+uvOzcbvrxKf+mMbtQ6JQ8wmkXqj/aH6jYnOS84dRSA2TUNBOT5sOr399hv8u8k5LfuXU0CIdbKQAnGvDClnfrcdI7KgPV9/e3wed9h0oxiaFClGCMKcwnk63M+J3ohN93VSoq0qT//laEgnAlY/kteWTK/XFBeF+g7dMm3U47qFt340P3K//r44pWAARh4EsgC4SogUGB/f9Xlm26pVsi+H7M2207Pce6xVkBZp31hKbUDI8VFGHDygD1navHG0S1YbDmshVGWj7FT0Q51kw1mkr6g7pePbVkV0BKYkI8jOrF46fwDxOTFVx9UUNJDwkr7Od32Bqai2DF/J3tmmWripy5qTG56VWJ3rXr3QX0M4aYMXbdMBhgGl9yPBtgslC6ATSwbs5Tt8ovAAAAAElFTkSuQmCC"></image></g><g transform="matrix(0.9999990463256836,0,0,0.9999990463256836,-5.151672840118408,-36.89065933227539)" opacity="1" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,55.481998443603516,126.74700164794922)"><path fill="rgb(114,51,180)" fill-opacity="1" d=" M-7.070000171661377,-9.762999534606934 C-7.070000171661377,-9.762999534606934 7.070000171661377,-9.762999534606934 7.070000171661377,-9.762999534606934 C8.557000160217285,-9.762999534606934 9.762999534606934,-8.557000160217285 9.762999534606934,-7.070000171661377 C9.762999534606934,-7.070000171661377 9.762999534606934,7.070000171661377 9.762999534606934,7.070000171661377 C9.762999534606934,8.557000160217285 8.557000160217285,9.762999534606934 7.070000171661377,9.762999534606934 C7.070000171661377,9.762999534606934 -7.070000171661377,9.762999534606934 -7.070000171661377,9.762999534606934 C-8.557000160217285,9.762999534606934 -9.762999534606934,8.557000160217285 -9.762999534606934,7.070000171661377 C-9.762999534606934,7.070000171661377 -9.762999534606934,-7.070000171661377 -9.762999534606934,-7.070000171661377 C-9.762999534606934,-8.557000160217285 -8.557000160217285,-9.762999534606934 -7.070000171661377,-9.762999534606934z"></path></g><g opacity="1" transform="matrix(1,0,0,1,55.481998443603516,126.74600219726562)"><path fill="rgb(250,250,250)" fill-opacity="1" d=" M2.931999921798706,-2.1440000534057617 C2.931999921798706,-2.1440000534057617 2.1440000534057617,-2.931999921798706 2.1440000534057617,-2.931999921798706 C2.013000011444092,-3.062999963760376 1.8009999990463257,-3.062999963760376 1.6699999570846558,-2.931999921798706 C1.6699999570846558,-2.931999921798706 0.6190000176429749,-1.88100004196167 0.6190000176429749,-1.88100004196167 C0.6190000176429749,-1.88100004196167 -0.03099999949336052,-2.5239999294281006 -0.03099999949336052,-2.5239999294281006 C-0.03099999949336052,-2.5239999294281006 -0.5049999952316284,-2.049999952316284 -0.5049999952316284,-2.049999952316284 C-0.5049999952316284,-2.049999952316284 -0.027000000700354576,-1.5720000267028809 -0.027000000700354576,-1.5720000267028809 C-0.027000000700354576,-1.5720000267028809 -3.0299999713897705,1.430999994277954 -3.0299999713897705,1.430999994277954 C-3.0299999713897705,1.430999994277954 -3.0299999713897705,3.0299999713897705 -3.0299999713897705,3.0299999713897705 C-3.0299999713897705,3.0299999713897705 -1.430999994277954,3.0299999713897705 -1.430999994277954,3.0299999713897705 C-1.430999994277954,3.0299999713897705 1.5720000267028809,0.027000000700354576 1.5720000267028809,0.027000000700354576 C1.5720000267028809,0.027000000700354576 2.049999952316284,0.5049999952316284 2.049999952316284,0.5049999952316284 C2.049999952316284,0.5049999952316284 2.5239999294281006,0.03099999949336052 2.5239999294281006,0.03099999949336052 C2.5239999294281006,0.03099999949336052 1.878000020980835,-0.6159999966621399 1.878000020980835,-0.6159999966621399 C1.878000020980835,-0.6159999966621399 2.927999973297119,-1.6660000085830688 2.927999973297119,-1.6660000085830688 C3.062999963760376,-1.8009999990463257 3.062999963760376,-2.013000011444092 2.931999921798706,-2.1440000534057617z M-1.7100000381469727,2.3570001125335693 C-1.7100000381469727,2.3570001125335693 -2.3570001125335693,1.7109999656677246 -2.3570001125335693,1.7109999656677246 C-2.3570001125335693,1.7109999656677246 0.3569999933242798,-1.003000020980835 0.3569999933242798,-1.003000020980835 C0.3569999933242798,-1.003000020980835 1.003000020980835,-0.35600000619888306 1.003000020980835,-0.35600000619888306 C1.003000020980835,-0.35600000619888306 -1.7100000381469727,2.3570001125335693 -1.7100000381469727,2.3570001125335693z"></path></g></g><g transform="matrix(0.9999990463256836,0,0,0.9999990463256836,-5.000431537628174,-42.00043869018555)" opacity="0.8" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,127.7770004272461,177.23399353027344)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-14.189000129699707,2.75 C-14.189000129699707,2.75 -11.350000381469727,-4.949999809265137 -11.350000381469727,-4.949999809265137 C-11.350000381469727,-4.949999809265137 -10.074000358581543,-4.949999809265137 -10.074000358581543,-4.949999809265137 C-10.074000358581543,-4.949999809265137 -7.257999897003174,2.75 -7.257999897003174,2.75 C-7.257999897003174,2.75 -8.489999771118164,2.75 -8.489999771118164,2.75 C-8.489999771118164,2.75 -9.149999618530273,0.8690000176429749 -9.149999618530273,0.8690000176429749 C-9.149999618530273,0.8690000176429749 -12.295999526977539,0.8690000176429749 -12.295999526977539,0.8690000176429749 C-12.295999526977539,0.8690000176429749 -12.956000328063965,2.75 -12.956000328063965,2.75 C-12.956000328063965,2.75 -14.189000129699707,2.75 -14.189000129699707,2.75z M-11.967000007629395,-0.054999999701976776 C-11.967000007629395,-0.054999999701976776 -9.479999542236328,-0.054999999701976776 -9.479999542236328,-0.054999999701976776 C-9.479999542236328,-0.054999999701976776 -10.722999572753906,-3.563999891281128 -10.722999572753906,-3.563999891281128 C-10.722999572753906,-3.563999891281128 -11.967000007629395,-0.054999999701976776 -11.967000007629395,-0.054999999701976776z M-6.651000022888184,5.170000076293945 C-6.651000022888184,5.170000076293945 -6.651000022888184,-2.7060000896453857 -6.651000022888184,-2.7060000896453857 C-6.651000022888184,-2.7060000896453857 -5.605999946594238,-2.7060000896453857 -5.605999946594238,-2.7060000896453857 C-5.605999946594238,-2.7060000896453857 -5.485000133514404,-1.8589999675750732 -5.485000133514404,-1.8589999675750732 C-5.309000015258789,-2.115999937057495 -5.066999912261963,-2.3429999351501465 -4.758999824523926,-2.5409998893737793 C-4.451000213623047,-2.739000082015991 -4.054999828338623,-2.8380000591278076 -3.571000099182129,-2.8380000591278076 C-3.0429999828338623,-2.8380000591278076 -2.5769999027252197,-2.7130000591278076 -2.1740000247955322,-2.4639999866485596 C-1.7710000276565552,-2.2149999141693115 -1.4550000429153442,-1.8739999532699585 -1.2280000448226929,-1.440999984741211 C-0.9929999709129333,-1.0080000162124634 -0.8759999871253967,-0.5170000195503235 -0.8759999871253967,0.032999999821186066 C-0.8759999871253967,0.5830000042915344 -0.9929999709129333,1.0740000009536743 -1.2280000448226929,1.5069999694824219 C-1.4550000429153442,1.9320000410079956 -1.7710000276565552,2.2699999809265137 -2.1740000247955322,2.5190000534057617 C-2.5769999027252197,2.760999917984009 -3.046999931335449,2.881999969482422 -3.5820000171661377,2.881999969482422 C-4.006999969482422,2.881999969482422 -4.385000228881836,2.7980000972747803 -4.715000152587891,2.628999948501587 C-5.038000106811523,2.4600000381469727 -5.294000148773193,2.2219998836517334 -5.485000133514404,1.9140000343322754 C-5.485000133514404,1.9140000343322754 -5.485000133514404,5.170000076293945 -5.485000133514404,5.170000076293945 C-5.485000133514404,5.170000076293945 -6.651000022888184,5.170000076293945 -6.651000022888184,5.170000076293945z M-3.7799999713897705,1.8700000047683716 C-3.2809998989105225,1.8700000047683716 -2.871000051498413,1.7009999752044678 -2.5480000972747803,1.3639999628067017 C-2.2249999046325684,1.0190000534057617 -2.063999891281128,0.5720000267028809 -2.063999891281128,0.02199999988079071 C-2.063999891281128,-0.3370000123977661 -2.13700008392334,-0.656000018119812 -2.2839999198913574,-0.9350000023841858 C-2.430999994277954,-1.2139999866485596 -2.631999969482422,-1.4299999475479126 -2.8889999389648438,-1.5839999914169312 C-3.1459999084472656,-1.7450000047683716 -3.443000078201294,-1.8259999752044678 -3.7799999713897705,-1.8259999752044678 C-4.2789998054504395,-1.8259999752044678 -4.689000129699707,-1.6540000438690186 -5.01200008392334,-1.309000015258789 C-5.327000141143799,-0.9639999866485596 -5.485000133514404,-0.5210000276565552 -5.485000133514404,0.02199999988079071 C-5.485000133514404,0.5720000267028809 -5.327000141143799,1.0190000534057617 -5.01200008392334,1.3639999628067017 C-4.689000129699707,1.7009999752044678 -4.2789998054504395,1.8700000047683716 -3.7799999713897705,1.8700000047683716z M-0.035999998450279236,5.170000076293945 C-0.035999998450279236,5.170000076293945 -0.035999998450279236,-2.7060000896453857 -0.035999998450279236,-2.7060000896453857 C-0.035999998450279236,-2.7060000896453857 1.0089999437332153,-2.7060000896453857 1.0089999437332153,-2.7060000896453857 C1.0089999437332153,-2.7060000896453857 1.1299999952316284,-1.8589999675750732 1.1299999952316284,-1.8589999675750732 C1.305999994277954,-2.115999937057495 1.5479999780654907,-2.3429999351501465 1.8559999465942383,-2.5409998893737793 C2.1640000343322754,-2.739000082015991 2.559999942779541,-2.8380000591278076 3.0439999103546143,-2.8380000591278076 C3.572000026702881,-2.8380000591278076 4.038000106811523,-2.7130000591278076 4.440999984741211,-2.4639999866485596 C4.843999862670898,-2.2149999141693115 5.159999847412109,-1.8739999532699585 5.38700008392334,-1.440999984741211 C5.622000217437744,-1.0080000162124634 5.738999843597412,-0.5170000195503235 5.738999843597412,0.032999999821186066 C5.738999843597412,0.5830000042915344 5.622000217437744,1.0740000009536743 5.38700008392334,1.5069999694824219 C5.159999847412109,1.9320000410079956 4.843999862670898,2.2699999809265137 4.440999984741211,2.5190000534057617 C4.038000106811523,2.760999917984009 3.568000078201294,2.881999969482422 3.0329999923706055,2.881999969482422 C2.6080000400543213,2.881999969482422 2.2300000190734863,2.7980000972747803 1.899999976158142,2.628999948501587 C1.5770000219345093,2.4600000381469727 1.3209999799728394,2.2219998836517334 1.1299999952316284,1.9140000343322754 C1.1299999952316284,1.9140000343322754 1.1299999952316284,5.170000076293945 1.1299999952316284,5.170000076293945 C1.1299999952316284,5.170000076293945 -0.035999998450279236,5.170000076293945 -0.035999998450279236,5.170000076293945z M2.8350000381469727,1.8700000047683716 C3.3340001106262207,1.8700000047683716 3.74399995803833,1.7009999752044678 4.066999912261963,1.3639999628067017 C4.389999866485596,1.0190000534057617 4.551000118255615,0.5720000267028809 4.551000118255615,0.02199999988079071 C4.551000118255615,-0.3370000123977661 4.478000164031982,-0.656000018119812 4.330999851226807,-0.9350000023841858 C4.184000015258789,-1.2139999866485596 3.9830000400543213,-1.4299999475479126 3.7260000705718994,-1.5839999914169312 C3.4690001010894775,-1.7450000047683716 3.171999931335449,-1.8259999752044678 2.8350000381469727,-1.8259999752044678 C2.3359999656677246,-1.8259999752044678 1.9259999990463257,-1.6540000438690186 1.6030000448226929,-1.309000015258789 C1.2879999876022339,-0.9639999866485596 1.1299999952316284,-0.5210000276565552 1.1299999952316284,0.02199999988079071 C1.1299999952316284,0.5720000267028809 1.2879999876022339,1.0190000534057617 1.6030000448226929,1.3639999628067017 C1.9259999990463257,1.7009999752044678 2.3359999656677246,1.8700000047683716 2.8350000381469727,1.8700000047683716z M6.578000068664551,2.75 C6.578000068664551,2.75 6.578000068664551,-5.170000076293945 6.578000068664551,-5.170000076293945 C6.578000068664551,-5.170000076293945 7.74399995803833,-5.170000076293945 7.74399995803833,-5.170000076293945 C7.74399995803833,-5.170000076293945 7.74399995803833,2.75 7.74399995803833,2.75 C7.74399995803833,2.75 6.578000068664551,2.75 6.578000068664551,2.75z M9.435999870300293,5.170000076293945 C9.435999870300293,5.170000076293945 10.744999885559082,2.2990000247955322 10.744999885559082,2.2990000247955322 C10.744999885559082,2.2990000247955322 10.425999641418457,2.2990000247955322 10.425999641418457,2.2990000247955322 C10.425999641418457,2.2990000247955322 8.270000457763672,-2.7060000896453857 8.270000457763672,-2.7060000896453857 C8.270000457763672,-2.7060000896453857 9.53600025177002,-2.7060000896453857 9.53600025177002,-2.7060000896453857 C9.53600025177002,-2.7060000896453857 11.208000183105469,1.3309999704360962 11.208000183105469,1.3309999704360962 C11.208000183105469,1.3309999704360962 12.956000328063965,-2.7060000896453857 12.956000328063965,-2.7060000896453857 C12.956000328063965,-2.7060000896453857 14.189000129699707,-2.7060000896453857 14.189000129699707,-2.7060000896453857 C14.189000129699707,-2.7060000896453857 10.668999671936035,5.170000076293945 10.668999671936035,5.170000076293945 C10.668999671936035,5.170000076293945 9.435999870300293,5.170000076293945 9.435999870300293,5.170000076293945z"></path></g><g opacity="1" transform="matrix(1,0,0,1,127.74500274658203,175.98399353027344)"><path stroke-linecap="butt" stroke-linejoin="miter" fill-opacity="0" stroke-miterlimit="4" stroke="rgb(255,255,255)" stroke-opacity="1" stroke-width="1" d=" M-77.5,-16.5 C-77.5,-16.5 77.5,-16.5 77.5,-16.5 C79.98500061035156,-16.5 82,-14.484999656677246 82,-12 C82,-12 82,12 82,12 C82,14.484999656677246 79.98500061035156,16.5 77.5,16.5 C77.5,16.5 -77.5,16.5 -77.5,16.5 C-79.98500061035156,16.5 -82,14.484999656677246 -82,12 C-82,12 -82,-12 -82,-12 C-82,-14.484999656677246 -79.98500061035156,-16.5 -77.5,-16.5z"></path></g></g><g class="png" transform="matrix(0.4999995231628418,0,0,0.4999995231628418,150.16448974609375,75.31493377685547)" opacity="1" style="display: block;"><image width="66px" height="66px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAEIAAABCCAMAAADUivDaAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAAzUExURUdwTAICAgEBAaenpwQEBAMDAwICAhkZGQoKCgcHB/Ly8k9PT8nJyeTk5LCwsJKSkvr6+uwWQioAAAAQdFJOUwAzQAETHCYDBgzsXK3Rk3xWRxyEAAACnElEQVRYw8WY63KEIAyFlewuiqi8/9NWbrkIiG5n2rQz7Y/m25PDLekw/EkADPBlIo9nkJylMfyvGXWLMLD06fhioFsQnx4B0zkipkPJ8qeYP/NAir5kRIJIfx3BIZP/i6a3XmQkUDYFYi4cAbQg579ZZEr2pI2IAMr/fMJ3pkSIbgiRhJAdYkw/CZKLaRiJAE8YKQiCxZwZpCEQPhmgfIwqUS4ZHkEaIkCJyJDECLtMrmcqQwCWza7GGbPafckQzoCyjEwIgGVbHQsbIZlRVOLNzD4kwm7cKTxEMviqMBGecEhYXRlmKxjCCiwjEIyrhj0xSAWKiEY0CcSoI6iMNiEzDhl+VZihQCJ8GdZdxH4wsgxmBggR2xXBmQVl8EqkCHOJCKWgDIm4J0LKwEp4HUqtHYTbEuIVKhGIWMfSI7gVK2EIYFZsXYSv5IQIByTtzBt1OMfNwELQzVuIrYMwfYTlS1JBuN8jnqqArxClF/DUzr1mJ9ta9qtFHfgp2/tbq7Y7+RlZzC038aiWJ1X1K9nFhXFC3KpkrV5bwF6A7posWAe/tdCMd+/2xUsrbSxSQcvqZWzdO6t6/XIZF46ahTmBJ4QqmUNvc8lYFN56fkXZiwjMjQuGEQTZHpx7g9rDLp72uXiWsckhhrInyLrLBsM3NY0+CRmK9SjGZkCrRSkYCaL23Vq7hTZJNFvVdk1rxoitzljr1hJhKpvG9LwzRqKMqWVEwKvZ/0JiTKL5HT/UuTLC1OjCAf2QHXTZP2vdROisI84R73dzFGiMaQAF5MXS2RgQrITWXKcJMpcTTVTQmkZoukyz2VwEzWa9CZONh5jMhju5rRuzNY6p5wlT3x52i2EZB+YB4OnMHSfbNN/eHrcr/zKAR5/+X/ED5vZsdcVjAFYAAAAASUVORK5CYII="></image></g><g transform="matrix(0.9999990463256836,0,0,0.9999990463256836,129.81350708007812,98.8109130859375)" opacity="1" style="display: block;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path fill="rgb(114,51,180)" fill-opacity="1" d=" M36.85100173950195,-14.38599967956543 C40.93199920654297,-14.38599967956543 44.24100112915039,-11.07699966430664 44.24100112915039,-6.995999813079834 C44.24100112915039,-2.9149999618530273 40.93199920654297,0.39399999380111694 36.85100173950195,0.39399999380111694 C32.77000045776367,0.39399999380111694 29.461000442504883,-2.9149999618530273 29.461000442504883,-6.995999813079834 C29.461000442504883,-11.07699966430664 32.77000045776367,-14.38599967956543 36.85100173950195,-14.38599967956543z"></path></g></g></g><g transform="matrix(0.30000001192092896,0,0,0.30000001192092896,177.19320678710938,490.6830749511719)" opacity="0.03" style="display: none;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path fill="rgb(84,82,186)" fill-opacity="1" d=" M-77.5,-33.5 C-77.5,-33.5 77.5,-33.5 77.5,-33.5 C79.98500061035156,-33.5 82,-31.485000610351562 82,-29 C82,-29 82,29 82,29 C82,31.485000610351562 79.98500061035156,33.5 77.5,33.5 C77.5,33.5 -77.5,33.5 -77.5,33.5 C-79.98500061035156,33.5 -82,31.485000610351562 -82,29 C-82,29 -82,-29 -82,-29 C-82,-31.485000610351562 -79.98500061035156,-33.5 -77.5,-33.5z"></path></g></g><g transform="matrix(0.30238619446754456,0,0,0.30238619446754456,130.79591369628906,438.6265869140625)" opacity="0.003408834304328252" style="display: none;"><g opacity="0.38" transform="matrix(1,0,0,1,153.46299743652344,184.4080047607422)"><path fill="url(#__lottie_element_806)" fill-opacity="1" d=" M-82.3239974975586,-92 C-82.3239974975586,-92 82.3239974975586,-92 82.3239974975586,-92 C91.25700378417969,-92 98.5,-84.75700378417969 98.5,-75.8239974975586 C98.5,-75.8239974975586 98.5,75.8239974975586 98.5,75.8239974975586 C98.5,84.75700378417969 91.25700378417969,92 82.3239974975586,92 C82.3239974975586,92 -82.3239974975586,92 -82.3239974975586,92 C-91.25700378417969,92 -98.5,84.75700378417969 -98.5,75.8239974975586 C-98.5,75.8239974975586 -98.5,-75.8239974975586 -98.5,-75.8239974975586 C-98.5,-84.75700378417969 -91.25700378417969,-92 -82.3239974975586,-92z"></path></g><g opacity="0.8" transform="matrix(1,0,0,1,109.43699645996094,118.74400329589844)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-32.345001220703125,5.855999946594238 C-33.45399856567383,5.855999946594238 -34.41400146484375,5.616000175476074 -35.224998474121094,5.136000156402588 C-36.025001525878906,4.644999980926514 -36.64400100708008,3.9679999351501465 -37.08100128173828,3.1040000915527344 C-37.518001556396484,2.2290000915527344 -37.73699951171875,1.2209999561309814 -37.73699951171875,0.07999999821186066 C-37.73699951171875,-1.0720000267028809 -37.518001556396484,-2.0799999237060547 -37.08100128173828,-2.944000005722046 C-36.64400100708008,-3.819000005722046 -36.025001525878906,-4.500999927520752 -35.224998474121094,-4.992000102996826 C-34.41400146484375,-5.482999801635742 -33.45399856567383,-5.728000164031982 -32.345001220703125,-5.728000164031982 C-31.033000946044922,-5.728000164031982 -29.961000442504883,-5.4079999923706055 -29.128999710083008,-4.76800012588501 C-28.297000885009766,-4.127999782562256 -27.768999099731445,-3.2320001125335693 -27.545000076293945,-2.0799999237060547 C-27.545000076293945,-2.0799999237060547 -29.433000564575195,-2.0799999237060547 -29.433000564575195,-2.0799999237060547 C-29.582000732421875,-2.7309999465942383 -29.902000427246094,-3.247999906539917 -30.39299964904785,-3.631999969482422 C-30.884000778198242,-4.015999794006348 -31.540000915527344,-4.208000183105469 -32.361000061035156,-4.208000183105469 C-33.48099899291992,-4.208000183105469 -34.36600112915039,-3.8239998817443848 -35.016998291015625,-3.055999994277954 C-35.667999267578125,-2.2990000247955322 -35.99300003051758,-1.253000020980835 -35.99300003051758,0.07999999821186066 C-35.99300003051758,1.402999997138977 -35.667999267578125,2.443000078201294 -35.016998291015625,3.200000047683716 C-34.36600112915039,3.9570000171661377 -33.48099899291992,4.335999965667725 -32.361000061035156,4.335999965667725 C-31.540000915527344,4.335999965667725 -30.884000778198242,4.15500020980835 -30.39299964904785,3.7920000553131104 C-29.902000427246094,3.428999900817871 -29.582000732421875,2.933000087738037 -29.433000564575195,2.303999900817871 C-29.433000564575195,2.303999900817871 -27.545000076293945,2.303999900817871 -27.545000076293945,2.303999900817871 C-27.768999099731445,3.4130001068115234 -28.297000885009766,4.2829999923706055 -29.128999710083008,4.9120001792907715 C-29.961000442504883,5.540999889373779 -31.033000946044922,5.855999946594238 -32.345001220703125,5.855999946594238z M-23.246000289916992,5.855999946594238 C-24.20599937438965,5.855999946594238 -24.96299934387207,5.557000160217285 -25.51799964904785,4.960000038146973 C-26.062000274658203,4.36299991607666 -26.333999633789062,3.4719998836517334 -26.333999633789062,2.2880001068115234 C-26.333999633789062,2.2880001068115234 -26.333999633789062,-2.2720000743865967 -26.333999633789062,-2.2720000743865967 C-26.333999633789062,-2.2720000743865967 -24.63800048828125,-2.2720000743865967 -24.63800048828125,-2.2720000743865967 C-24.63800048828125,-2.2720000743865967 -24.63800048828125,2.111999988555908 -24.63800048828125,2.111999988555908 C-24.63800048828125,3.6480000019073486 -24.009000778198242,4.415999889373779 -22.75,4.415999889373779 C-22.121000289916992,4.415999889373779 -21.60300064086914,4.191999912261963 -21.197999954223633,3.74399995803833 C-20.792999267578125,3.2960000038146973 -20.59000015258789,2.6559998989105225 -20.59000015258789,1.8240000009536743 C-20.59000015258789,1.8240000009536743 -20.59000015258789,-2.2720000743865967 -20.59000015258789,-2.2720000743865967 C-20.59000015258789,-2.2720000743865967 -18.893999099731445,-2.2720000743865967 -18.893999099731445,-2.2720000743865967 C-18.893999099731445,-2.2720000743865967 -18.893999099731445,5.664000034332275 -18.893999099731445,5.664000034332275 C-18.893999099731445,5.664000034332275 -20.398000717163086,5.664000034332275 -20.398000717163086,5.664000034332275 C-20.398000717163086,5.664000034332275 -20.525999069213867,4.271999835968018 -20.525999069213867,4.271999835968018 C-20.770999908447266,4.763000011444092 -21.134000778198242,5.1519999504089355 -21.61400032043457,5.440000057220459 C-22.08300018310547,5.7170000076293945 -22.62700080871582,5.855999946594238 -23.246000289916992,5.855999946594238z M-13.996999740600586,5.855999946594238 C-15,5.855999946594238 -15.826000213623047,5.611000061035156 -16.476999282836914,5.119999885559082 C-17.128000259399414,4.629000186920166 -17.500999450683594,3.9790000915527344 -17.597000122070312,3.1679999828338623 C-17.597000122070312,3.1679999828338623 -15.885000228881836,3.1679999828338623 -15.885000228881836,3.1679999828338623 C-15.800000190734863,3.5309998989105225 -15.597000122070312,3.8450000286102295 -15.277000427246094,4.111999988555908 C-14.956999778747559,4.368000030517578 -14.53600025177002,4.495999813079834 -14.012999534606934,4.495999813079834 C-13.50100040435791,4.495999813079834 -13.128000259399414,4.388999938964844 -12.892999649047852,4.176000118255615 C-12.657999992370605,3.9630000591278076 -12.541000366210938,3.7170000076293945 -12.541000366210938,3.440000057220459 C-12.541000366210938,3.0350000858306885 -12.706000328063965,2.763000011444092 -13.036999702453613,2.624000072479248 C-13.357000350952148,2.4749999046325684 -13.805000305175781,2.3410000801086426 -14.380999565124512,2.2239999771118164 C-14.829000473022461,2.128000020980835 -15.277000427246094,2 -15.725000381469727,1.840000033378601 C-16.16200065612793,1.6799999475479126 -16.530000686645508,1.4559999704360962 -16.82900047302246,1.1679999828338623 C-17.117000579833984,0.8690000176429749 -17.26099967956543,0.4690000116825104 -17.26099967956543,-0.03200000151991844 C-17.26099967956543,-0.7250000238418579 -16.993999481201172,-1.3009999990463257 -16.461000442504883,-1.7599999904632568 C-15.928000450134277,-2.2290000915527344 -15.180999755859375,-2.4639999866485596 -14.220999717712402,-2.4639999866485596 C-13.336000442504883,-2.4639999866485596 -12.621000289916992,-2.250999927520752 -12.07699966430664,-1.8240000009536743 C-11.522000312805176,-1.3969999551773071 -11.196999549865723,-0.7950000166893005 -11.10099983215332,-0.01600000075995922 C-11.10099983215332,-0.01600000075995922 -12.732999801635742,-0.01600000075995922 -12.732999801635742,-0.01600000075995922 C-12.78600025177002,-0.3569999933242798 -12.946000099182129,-0.6240000128746033 -13.213000297546387,-0.8159999847412109 C-13.468999862670898,-1.0080000162124634 -13.815999984741211,-1.1039999723434448 -14.253000259399414,-1.1039999723434448 C-14.680000305175781,-1.1039999723434448 -15.010000228881836,-1.0130000114440918 -15.244999885559082,-0.8320000171661377 C-15.479999542236328,-0.6610000133514404 -15.597000122070312,-0.43700000643730164 -15.597000122070312,-0.1599999964237213 C-15.597000122070312,0.11699999868869781 -15.437000274658203,0.335999995470047 -15.116999626159668,0.4959999918937683 C-14.78600025177002,0.656000018119812 -14.354000091552734,0.800000011920929 -13.821000099182129,0.9279999732971191 C-13.288000106811523,1.0449999570846558 -12.79699993133545,1.184000015258789 -12.348999977111816,1.343999981880188 C-11.890000343322754,1.4930000305175781 -11.522000312805176,1.7170000076293945 -11.244999885559082,2.0160000324249268 C-10.968000411987305,2.315000057220459 -10.829000473022461,2.752000093460083 -10.829000473022461,3.328000068664551 C-10.817999839782715,4.052999973297119 -11.10099983215332,4.656000137329102 -11.677000045776367,5.136000156402588 C-12.241999626159668,5.616000175476074 -13.015999794006348,5.855999946594238 -13.996999740600586,5.855999946594238z M-6.375,5.664000034332275 C-7.1539998054504395,5.664000034332275 -7.771999835968018,5.4770002365112305 -8.230999946594238,5.104000091552734 C-8.6899995803833,4.71999979019165 -8.918999671936035,4.043000221252441 -8.918999671936035,3.072000026702881 C-8.918999671936035,3.072000026702881 -8.918999671936035,-0.8479999899864197 -8.918999671936035,-0.8479999899864197 C-8.918999671936035,-0.8479999899864197 -10.279000282287598,-0.8479999899864197 -10.279000282287598,-0.8479999899864197 C-10.279000282287598,-0.8479999899864197 -10.279000282287598,-2.2720000743865967 -10.279000282287598,-2.2720000743865967 C-10.279000282287598,-2.2720000743865967 -8.918999671936035,-2.2720000743865967 -8.918999671936035,-2.2720000743865967 C-8.918999671936035,-2.2720000743865967 -8.711000442504883,-4.288000106811523 -8.711000442504883,-4.288000106811523 C-8.711000442504883,-4.288000106811523 -7.2230000495910645,-4.288000106811523 -7.2230000495910645,-4.288000106811523 C-7.2230000495910645,-4.288000106811523 -7.2230000495910645,-2.2720000743865967 -7.2230000495910645,-2.2720000743865967 C-7.2230000495910645,-2.2720000743865967 -4.982999801635742,-2.2720000743865967 -4.982999801635742,-2.2720000743865967 C-4.982999801635742,-2.2720000743865967 -4.982999801635742,-0.8479999899864197 -4.982999801635742,-0.8479999899864197 C-4.982999801635742,-0.8479999899864197 -7.2230000495910645,-0.8479999899864197 -7.2230000495910645,-0.8479999899864197 C-7.2230000495910645,-0.8479999899864197 -7.2230000495910645,3.072000026702881 -7.2230000495910645,3.072000026702881 C-7.2230000495910645,3.509000062942505 -7.131999969482422,3.812999963760376 -6.951000213623047,3.9839999675750732 C-6.758999824523926,4.144000053405762 -6.434000015258789,4.223999977111816 -5.974999904632568,4.223999977111816 C-5.974999904632568,4.223999977111816 -5.063000202178955,4.223999977111816 -5.063000202178955,4.223999977111816 C-5.063000202178955,4.223999977111816 -5.063000202178955,5.664000034332275 -5.063000202178955,5.664000034332275 C-5.063000202178955,5.664000034332275 -6.375,5.664000034332275 -6.375,5.664000034332275z M-0.15399999916553497,5.855999946594238 C-0.9110000133514404,5.855999946594238 -1.593999981880188,5.684999942779541 -2.2019999027252197,5.343999862670898 C-2.7990000247955322,4.992000102996826 -3.2739999294281006,4.506999969482422 -3.625999927520752,3.888000011444092 C-3.9779999256134033,3.259000062942505 -4.1539998054504395,2.5280001163482666 -4.1539998054504395,1.6959999799728394 C-4.1539998054504395,0.8640000224113464 -3.9779999256134033,0.13899999856948853 -3.625999927520752,-0.47999998927116394 C-3.263000011444092,-1.1089999675750732 -2.7780001163482666,-1.5950000286102295 -2.1700000762939453,-1.9359999895095825 C-1.562000036239624,-2.2880001068115234 -0.8849999904632568,-2.4639999866485596 -0.1379999965429306,-2.4639999866485596 C0.6190000176429749,-2.4639999866485596 1.2970000505447388,-2.2880001068115234 1.8940000534057617,-1.9359999895095825 C2.502000093460083,-1.5950000286102295 2.9820001125335693,-1.1089999675750732 3.3340001106262207,-0.47999998927116394 C3.697000026702881,0.13899999856948853 3.878000020980835,0.8640000224113464 3.878000020980835,1.6959999799728394 C3.878000020980835,2.5280001163482666 3.697000026702881,3.259000062942505 3.3340001106262207,3.888000011444092 C2.9820001125335693,4.506999969482422 2.502000093460083,4.992000102996826 1.8940000534057617,5.343999862670898 C1.2860000133514404,5.684999942779541 0.6029999852180481,5.855999946594238 -0.15399999916553497,5.855999946594238z M-0.15399999916553497,4.400000095367432 C0.25099998712539673,4.400000095367432 0.625,4.298999786376953 0.9660000205039978,4.0960001945495605 C1.3179999589920044,3.8929998874664307 1.6009999513626099,3.5950000286102295 1.8140000104904175,3.200000047683716 C2.0269999504089355,2.7950000762939453 2.134000062942505,2.2929999828338623 2.134000062942505,1.6959999799728394 C2.134000062942505,1.0989999771118164 2.0269999504089355,0.6029999852180481 1.8140000104904175,0.20800000429153442 C1.6109999418258667,-0.19699999690055847 1.3339999914169312,-0.5009999871253967 0.9819999933242798,-0.7039999961853027 C0.640999972820282,-0.9070000052452087 0.2669999897480011,-1.0080000162124634 -0.1379999965429306,-1.0080000162124634 C-0.5429999828338623,-1.0080000162124634 -0.921999990940094,-0.9070000052452087 -1.2740000486373901,-0.7039999961853027 C-1.6150000095367432,-0.5009999871253967 -1.8930000066757202,-0.19699999690055847 -2.1059999465942383,0.20800000429153442 C-2.319000005722046,0.6029999852180481 -2.4260001182556152,1.0989999771118164 -2.4260001182556152,1.6959999799728394 C-2.4260001182556152,2.2929999828338623 -2.319000005722046,2.7950000762939453 -2.1059999465942383,3.200000047683716 C-1.8930000066757202,3.5950000286102295 -1.6150000095367432,3.8929998874664307 -1.2740000486373901,4.0960001945495605 C-0.9330000281333923,4.298999786376953 -0.5590000152587891,4.400000095367432 -0.15399999916553497,4.400000095367432z M5.2829999923706055,5.664000034332275 C5.2829999923706055,5.664000034332275 5.2829999923706055,-2.2720000743865967 5.2829999923706055,-2.2720000743865967 C5.2829999923706055,-2.2720000743865967 6.7870001792907715,-2.2720000743865967 6.7870001792907715,-2.2720000743865967 C6.7870001792907715,-2.2720000743865967 6.931000232696533,-1.1519999504089355 6.931000232696533,-1.1519999504089355 C7.186999797821045,-1.5570000410079956 7.5229997634887695,-1.8769999742507935 7.939000129699707,-2.111999988555908 C8.366000175476074,-2.3469998836517334 8.855999946594238,-2.4639999866485596 9.41100025177002,-2.4639999866485596 C10.670000076293945,-2.4639999866485596 11.543999671936035,-1.968000054359436 12.03499984741211,-0.9760000109672546 C12.322999954223633,-1.434999942779541 12.706999778747559,-1.7970000505447388 13.187000274658203,-2.063999891281128 C13.678000450134277,-2.3310000896453857 14.206000328063965,-2.4639999866485596 14.770999908447266,-2.4639999866485596 C15.762999534606934,-2.4639999866485596 16.54199981689453,-2.1649999618530273 17.10700035095215,-1.5679999589920044 C17.672000885009766,-0.9710000157356262 17.954999923706055,-0.07999999821186066 17.954999923706055,1.1039999723434448 C17.954999923706055,1.1039999723434448 17.954999923706055,5.664000034332275 17.954999923706055,5.664000034332275 C17.954999923706055,5.664000034332275 16.259000778198242,5.664000034332275 16.259000778198242,5.664000034332275 C16.259000778198242,5.664000034332275 16.259000778198242,1.2799999713897705 16.259000778198242,1.2799999713897705 C16.259000778198242,-0.25600001215934753 15.67199993133545,-1.0240000486373901 14.49899959564209,-1.0240000486373901 C13.902000427246094,-1.0240000486373901 13.41100025177002,-0.800000011920929 13.027000427246094,-0.35199999809265137 C12.654000282287598,0.09600000083446503 12.467000007629395,0.7360000014305115 12.467000007629395,1.5679999589920044 C12.467000007629395,1.5679999589920044 12.467000007629395,5.664000034332275 12.467000007629395,5.664000034332275 C12.467000007629395,5.664000034332275 10.770999908447266,5.664000034332275 10.770999908447266,5.664000034332275 C10.770999908447266,5.664000034332275 10.770999908447266,1.2799999713897705 10.770999908447266,1.2799999713897705 C10.770999908447266,-0.25600001215934753 10.178999900817871,-1.0240000486373901 8.994999885559082,-1.0240000486373901 C8.407999992370605,-1.0240000486373901 7.922999858856201,-0.800000011920929 7.539000034332275,-0.35199999809265137 C7.165999889373779,0.09600000083446503 6.979000091552734,0.7360000014305115 6.979000091552734,1.5679999589920044 C6.979000091552734,1.5679999589920044 6.979000091552734,5.664000034332275 6.979000091552734,5.664000034332275 C6.979000091552734,5.664000034332275 5.2829999923706055,5.664000034332275 5.2829999923706055,5.664000034332275z M20.47100067138672,-3.7760000228881836 C20.150999069213867,-3.7760000228881836 19.884000778198242,-3.871999979019165 19.67099952697754,-4.064000129699707 C19.468000411987305,-4.267000198364258 19.367000579833984,-4.517000198364258 19.367000579833984,-4.815999984741211 C19.367000579833984,-5.114999771118164 19.468000411987305,-5.360000133514404 19.67099952697754,-5.552000045776367 C19.884000778198242,-5.755000114440918 20.150999069213867,-5.855999946594238 20.47100067138672,-5.855999946594238 C20.791000366210938,-5.855999946594238 21.052000045776367,-5.755000114440918 21.2549991607666,-5.552000045776367 C21.468000411987305,-5.360000133514404 21.575000762939453,-5.114999771118164 21.575000762939453,-4.815999984741211 C21.575000762939453,-4.517000198364258 21.468000411987305,-4.267000198364258 21.2549991607666,-4.064000129699707 C21.052000045776367,-3.871999979019165 20.791000366210938,-3.7760000228881836 20.47100067138672,-3.7760000228881836z M19.62299919128418,5.664000034332275 C19.62299919128418,5.664000034332275 19.62299919128418,-2.2720000743865967 19.62299919128418,-2.2720000743865967 C19.62299919128418,-2.2720000743865967 21.319000244140625,-2.2720000743865967 21.319000244140625,-2.2720000743865967 C21.319000244140625,-2.2720000743865967 21.319000244140625,5.664000034332275 21.319000244140625,5.664000034332275 C21.319000244140625,5.664000034332275 19.62299919128418,5.664000034332275 19.62299919128418,5.664000034332275z M22.60099983215332,5.664000034332275 C22.60099983215332,5.664000034332275 22.60099983215332,4.288000106811523 22.60099983215332,4.288000106811523 C22.60099983215332,4.288000106811523 26.840999603271484,-0.8640000224113464 26.840999603271484,-0.8640000224113464 C26.840999603271484,-0.8640000224113464 22.665000915527344,-0.8640000224113464 22.665000915527344,-0.8640000224113464 C22.665000915527344,-0.8640000224113464 22.665000915527344,-2.2720000743865967 22.665000915527344,-2.2720000743865967 C22.665000915527344,-2.2720000743865967 28.76099967956543,-2.2720000743865967 28.76099967956543,-2.2720000743865967 C28.76099967956543,-2.2720000743865967 28.76099967956543,-0.8960000276565552 28.76099967956543,-0.8960000276565552 C28.76099967956543,-0.8960000276565552 24.48900032043457,4.25600004196167 24.48900032043457,4.25600004196167 C24.48900032043457,4.25600004196167 28.840999603271484,4.25600004196167 28.840999603271484,4.25600004196167 C28.840999603271484,4.25600004196167 28.840999603271484,5.664000034332275 28.840999603271484,5.664000034332275 C28.840999603271484,5.664000034332275 22.60099983215332,5.664000034332275 22.60099983215332,5.664000034332275z M33.8650016784668,5.855999946594238 C33.08599853515625,5.855999946594238 32.393001556396484,5.684999942779541 31.78499984741211,5.343999862670898 C31.187999725341797,4.992000102996826 30.718000411987305,4.506999969482422 30.37700080871582,3.888000011444092 C30.035999298095703,3.2690000534057617 29.864999771118164,2.5490000247955322 29.864999771118164,1.7280000448226929 C29.864999771118164,0.8960000276565552 30.030000686645508,0.16500000655651093 30.361000061035156,-0.46399998664855957 C30.70199966430664,-1.093000054359436 31.172000885009766,-1.5839999914169312 31.768999099731445,-1.9359999895095825 C32.37699890136719,-2.2880001068115234 33.08100128173828,-2.4639999866485596 33.88100051879883,-2.4639999866485596 C34.65999984741211,-2.4639999866485596 35.33700180053711,-2.2880001068115234 35.91299819946289,-1.9359999895095825 C36.48899841308594,-1.5950000286102295 36.9370002746582,-1.1360000371932983 37.25699996948242,-0.5600000023841858 C37.57699966430664,0.01600000075995922 37.73699951171875,0.6510000228881836 37.73699951171875,1.343999981880188 C37.73699951171875,1.4509999752044678 37.731998443603516,1.5679999589920044 37.72100067138672,1.6959999799728394 C37.72100067138672,1.812999963760376 37.715999603271484,1.9470000267028809 37.70500183105469,2.0959999561309814 C37.70500183105469,2.0959999561309814 31.52899932861328,2.0959999561309814 31.52899932861328,2.0959999561309814 C31.582000732421875,2.864000082015991 31.83300018310547,3.4509999752044678 32.28099822998047,3.8559999465942383 C32.7400016784668,4.250999927520752 33.268001556396484,4.447999954223633 33.8650016784668,4.447999954223633 C34.345001220703125,4.447999954223633 34.744998931884766,4.341000080108643 35.064998626708984,4.127999782562256 C35.395999908447266,3.9040000438690186 35.64099884033203,3.6050000190734863 35.80099868774414,3.2320001125335693 C35.80099868774414,3.2320001125335693 37.49700164794922,3.2320001125335693 37.49700164794922,3.2320001125335693 C37.284000396728516,3.9790000915527344 36.856998443603516,4.603000164031982 36.21699905395508,5.104000091552734 C35.5880012512207,5.605000019073486 34.80400085449219,5.855999946594238 33.8650016784668,5.855999946594238z M33.8650016784668,-1.0720000267028809 C33.29999923706055,-1.0720000267028809 32.79800033569336,-0.9010000228881836 32.361000061035156,-0.5600000023841858 C31.923999786376953,-0.2290000021457672 31.656999588012695,0.2720000147819519 31.56100082397461,0.9440000057220459 C31.56100082397461,0.9440000057220459 36.04100036621094,0.9440000057220459 36.04100036621094,0.9440000057220459 C36.00899887084961,0.32499998807907104 35.790000915527344,-0.16500000655651093 35.3849983215332,-0.527999997138977 C34.97999954223633,-0.890999972820282 34.472999572753906,-1.0720000267028809 33.8650016784668,-1.0720000267028809z"></path></g><g opacity="1" transform="matrix(1,0,0,1,153.46299743652344,172.01199340820312)"><path stroke-linecap="butt" stroke-linejoin="miter" fill-opacity="0" stroke-miterlimit="4" stroke-dasharray=" 2 2" stroke-dashoffset="0" stroke="rgb(84,82,186)" stroke-opacity="1" stroke-width="1" d=" M-77.5,-33.5 C-77.5,-33.5 77.5,-33.5 77.5,-33.5 C79.98500061035156,-33.5 82,-31.485000610351562 82,-29 C82,-29 82,29 82,29 C82,31.485000610351562 79.98500061035156,33.5 77.5,33.5 C77.5,33.5 -77.5,33.5 -77.5,33.5 C-79.98500061035156,33.5 -82,31.485000610351562 -82,29 C-82,29 -82,-29 -82,-29 C-82,-31.485000610351562 -79.98500061035156,-33.5 -77.5,-33.5z"></path></g><g opacity="0.25" transform="matrix(1,0,0,1,153.46400451660156,184.40899658203125)"><path stroke="url(#__lottie_element_807)" stroke-linecap="butt" stroke-linejoin="miter" fill-opacity="0" stroke-miterlimit="4" stroke-opacity="1" stroke-width="0.86" d=" M-82.32499694824219,-91.56999969482422 C-82.32499694824219,-91.56999969482422 82.32499694824219,-91.56999969482422 82.32499694824219,-91.56999969482422 C91.02100372314453,-91.56999969482422 98.06999969482422,-84.52100372314453 98.06999969482422,-75.82499694824219 C98.06999969482422,-75.82499694824219 98.06999969482422,75.82499694824219 98.06999969482422,75.82499694824219 C98.06999969482422,84.52100372314453 91.02100372314453,91.56999969482422 82.32499694824219,91.56999969482422 C82.32499694824219,91.56999969482422 -82.32499694824219,91.56999969482422 -82.32499694824219,91.56999969482422 C-91.02100372314453,91.56999969482422 -98.06999969482422,84.52100372314453 -98.06999969482422,75.82499694824219 C-98.06999969482422,75.82499694824219 -98.06999969482422,-75.82499694824219 -98.06999969482422,-75.82499694824219 C-98.06999969482422,-84.52100372314453 -91.02100372314453,-91.56999969482422 -82.32499694824219,-91.56999969482422z"></path></g></g><g transform="matrix(0.30238544940948486,0,0,0.30238544940948486,130.79603576660156,438.62677001953125)" opacity="0.003408834304328252" style="display: none;"><g opacity="1" transform="matrix(1,0,0,1,153.49600219726562,235.0540008544922)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M-14.189000129699707,2.75 C-14.189000129699707,2.75 -11.35099983215332,-4.949999809265137 -11.35099983215332,-4.949999809265137 C-11.35099983215332,-4.949999809265137 -10.074999809265137,-4.949999809265137 -10.074999809265137,-4.949999809265137 C-10.074999809265137,-4.949999809265137 -7.258999824523926,2.75 -7.258999824523926,2.75 C-7.258999824523926,2.75 -8.491000175476074,2.75 -8.491000175476074,2.75 C-8.491000175476074,2.75 -9.151000022888184,0.8690000176429749 -9.151000022888184,0.8690000176429749 C-9.151000022888184,0.8690000176429749 -12.29699993133545,0.8690000176429749 -12.29699993133545,0.8690000176429749 C-12.29699993133545,0.8690000176429749 -12.956999778747559,2.75 -12.956999778747559,2.75 C-12.956999778747559,2.75 -14.189000129699707,2.75 -14.189000129699707,2.75z M-11.967000007629395,-0.054999999701976776 C-11.967000007629395,-0.054999999701976776 -9.480999946594238,-0.054999999701976776 -9.480999946594238,-0.054999999701976776 C-9.480999946594238,-0.054999999701976776 -10.723999977111816,-3.563999891281128 -10.723999977111816,-3.563999891281128 C-10.723999977111816,-3.563999891281128 -11.967000007629395,-0.054999999701976776 -11.967000007629395,-0.054999999701976776z M-6.651000022888184,5.170000076293945 C-6.651000022888184,5.170000076293945 -6.651000022888184,-2.7060000896453857 -6.651000022888184,-2.7060000896453857 C-6.651000022888184,-2.7060000896453857 -5.605999946594238,-2.7060000896453857 -5.605999946594238,-2.7060000896453857 C-5.605999946594238,-2.7060000896453857 -5.485000133514404,-1.8589999675750732 -5.485000133514404,-1.8589999675750732 C-5.309000015258789,-2.115999937057495 -5.066999912261963,-2.3429999351501465 -4.758999824523926,-2.5409998893737793 C-4.451000213623047,-2.739000082015991 -4.054999828338623,-2.8380000591278076 -3.571000099182129,-2.8380000591278076 C-3.0429999828338623,-2.8380000591278076 -2.5769999027252197,-2.7130000591278076 -2.1740000247955322,-2.4639999866485596 C-1.7710000276565552,-2.2149999141693115 -1.4550000429153442,-1.8739999532699585 -1.2280000448226929,-1.440999984741211 C-0.9929999709129333,-1.0080000162124634 -0.8759999871253967,-0.5170000195503235 -0.8759999871253967,0.032999999821186066 C-0.8759999871253967,0.5830000042915344 -0.9929999709129333,1.0740000009536743 -1.2280000448226929,1.5069999694824219 C-1.4550000429153442,1.9320000410079956 -1.7710000276565552,2.2699999809265137 -2.1740000247955322,2.5190000534057617 C-2.5769999027252197,2.760999917984009 -3.046999931335449,2.881999969482422 -3.5820000171661377,2.881999969482422 C-4.006999969482422,2.881999969482422 -4.385000228881836,2.7980000972747803 -4.715000152587891,2.628999948501587 C-5.038000106811523,2.4600000381469727 -5.294000148773193,2.2219998836517334 -5.485000133514404,1.9140000343322754 C-5.485000133514404,1.9140000343322754 -5.485000133514404,5.170000076293945 -5.485000133514404,5.170000076293945 C-5.485000133514404,5.170000076293945 -6.651000022888184,5.170000076293945 -6.651000022888184,5.170000076293945z M-3.7799999713897705,1.8700000047683716 C-3.2809998989105225,1.8700000047683716 -2.871000051498413,1.7009999752044678 -2.5480000972747803,1.3639999628067017 C-2.2249999046325684,1.0190000534057617 -2.063999891281128,0.5720000267028809 -2.063999891281128,0.02199999988079071 C-2.063999891281128,-0.3370000123977661 -2.13700008392334,-0.656000018119812 -2.2839999198913574,-0.9350000023841858 C-2.430999994277954,-1.2139999866485596 -2.631999969482422,-1.4299999475479126 -2.8889999389648438,-1.5839999914169312 C-3.1459999084472656,-1.7450000047683716 -3.443000078201294,-1.8259999752044678 -3.7799999713897705,-1.8259999752044678 C-4.2789998054504395,-1.8259999752044678 -4.689000129699707,-1.6540000438690186 -5.01200008392334,-1.309000015258789 C-5.327000141143799,-0.9639999866485596 -5.485000133514404,-0.5210000276565552 -5.485000133514404,0.02199999988079071 C-5.485000133514404,0.5720000267028809 -5.327000141143799,1.0190000534057617 -5.01200008392334,1.3639999628067017 C-4.689000129699707,1.7009999752044678 -4.2789998054504395,1.8700000047683716 -3.7799999713897705,1.8700000047683716z M-0.03700000047683716,5.170000076293945 C-0.03700000047683716,5.170000076293945 -0.03700000047683716,-2.7060000896453857 -0.03700000047683716,-2.7060000896453857 C-0.03700000047683716,-2.7060000896453857 1.0080000162124634,-2.7060000896453857 1.0080000162124634,-2.7060000896453857 C1.0080000162124634,-2.7060000896453857 1.128999948501587,-1.8589999675750732 1.128999948501587,-1.8589999675750732 C1.3049999475479126,-2.115999937057495 1.5479999780654907,-2.3429999351501465 1.8559999465942383,-2.5409998893737793 C2.1640000343322754,-2.739000082015991 2.559999942779541,-2.8380000591278076 3.0439999103546143,-2.8380000591278076 C3.572000026702881,-2.8380000591278076 4.0370001792907715,-2.7130000591278076 4.440999984741211,-2.4639999866485596 C4.843999862670898,-2.2149999141693115 5.158999919891357,-1.8739999532699585 5.38700008392334,-1.440999984741211 C5.620999813079834,-1.0080000162124634 5.738999843597412,-0.5170000195503235 5.738999843597412,0.032999999821186066 C5.738999843597412,0.5830000042915344 5.620999813079834,1.0740000009536743 5.38700008392334,1.5069999694824219 C5.158999919891357,1.9320000410079956 4.843999862670898,2.2699999809265137 4.440999984741211,2.5190000534057617 C4.0370001792907715,2.760999917984009 3.568000078201294,2.881999969482422 3.0329999923706055,2.881999969482422 C2.6070001125335693,2.881999969482422 2.2300000190734863,2.7980000972747803 1.899999976158142,2.628999948501587 C1.5770000219345093,2.4600000381469727 1.3200000524520874,2.2219998836517334 1.128999948501587,1.9140000343322754 C1.128999948501587,1.9140000343322754 1.128999948501587,5.170000076293945 1.128999948501587,5.170000076293945 C1.128999948501587,5.170000076293945 -0.03700000047683716,5.170000076293945 -0.03700000047683716,5.170000076293945z M2.8350000381469727,1.8700000047683716 C3.3329999446868896,1.8700000047683716 3.74399995803833,1.7009999752044678 4.066999912261963,1.3639999628067017 C4.388999938964844,1.0190000534057617 4.551000118255615,0.5720000267028809 4.551000118255615,0.02199999988079071 C4.551000118255615,-0.3370000123977661 4.4770002365112305,-0.656000018119812 4.330999851226807,-0.9350000023841858 C4.184000015258789,-1.2139999866485596 3.9820001125335693,-1.4299999475479126 3.7260000705718994,-1.5839999914169312 C3.4690001010894775,-1.7450000047683716 3.171999931335449,-1.8259999752044678 2.8350000381469727,-1.8259999752044678 C2.3359999656677246,-1.8259999752044678 1.9249999523162842,-1.6540000438690186 1.6030000448226929,-1.309000015258789 C1.2879999876022339,-0.9639999866485596 1.128999948501587,-0.5210000276565552 1.128999948501587,0.02199999988079071 C1.128999948501587,0.5720000267028809 1.2879999876022339,1.0190000534057617 1.6030000448226929,1.3639999628067017 C1.9249999523162842,1.7009999752044678 2.3359999656677246,1.8700000047683716 2.8350000381469727,1.8700000047683716z M6.578000068664551,2.75 C6.578000068664551,2.75 6.578000068664551,-5.170000076293945 6.578000068664551,-5.170000076293945 C6.578000068664551,-5.170000076293945 7.74399995803833,-5.170000076293945 7.74399995803833,-5.170000076293945 C7.74399995803833,-5.170000076293945 7.74399995803833,2.75 7.74399995803833,2.75 C7.74399995803833,2.75 6.578000068664551,2.75 6.578000068664551,2.75z M9.437000274658203,5.170000076293945 C9.437000274658203,5.170000076293945 10.746000289916992,2.2990000247955322 10.746000289916992,2.2990000247955322 C10.746000289916992,2.2990000247955322 10.427000045776367,2.2990000247955322 10.427000045776367,2.2990000247955322 C10.427000045776367,2.2990000247955322 8.270999908447266,-2.7060000896453857 8.270999908447266,-2.7060000896453857 C8.270999908447266,-2.7060000896453857 9.53600025177002,-2.7060000896453857 9.53600025177002,-2.7060000896453857 C9.53600025177002,-2.7060000896453857 11.208000183105469,1.3309999704360962 11.208000183105469,1.3309999704360962 C11.208000183105469,1.3309999704360962 12.956999778747559,-2.7060000896453857 12.956999778747559,-2.7060000896453857 C12.956999778747559,-2.7060000896453857 14.189000129699707,-2.7060000896453857 14.189000129699707,-2.7060000896453857 C14.189000129699707,-2.7060000896453857 10.668999671936035,5.170000076293945 10.668999671936035,5.170000076293945 C10.668999671936035,5.170000076293945 9.437000274658203,5.170000076293945 9.437000274658203,5.170000076293945z"></path></g><g opacity="1" transform="matrix(1,0,0,1,153.46299743652344,233.8040008544922)"><path stroke-linecap="butt" stroke-linejoin="miter" fill-opacity="0" stroke-miterlimit="4" stroke="rgb(255,255,255)" stroke-opacity="1" stroke-width="1" d=" M-77.5,-16.5 C-77.5,-16.5 77.5,-16.5 77.5,-16.5 C79.98500061035156,-16.5 82,-14.484999656677246 82,-12 C82,-12 82,12 82,12 C82,14.484999656677246 79.98500061035156,16.5 77.5,16.5 C77.5,16.5 -77.5,16.5 -77.5,16.5 C-79.98500061035156,16.5 -82,14.484999656677246 -82,12 C-82,12 -82,-12 -82,-12 C-82,-14.484999656677246 -79.98500061035156,-16.5 -77.5,-16.5z"></path></g></g><g class="png" transform="matrix(0.5960000157356262,0,0,0.5960000157356262,126.5889892578125,466.1409912109375)" opacity="1" style="display: none;"><image width="172px" height="60px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAKwAAAA8CAMAAADMiaGpAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAB1UExURUdwTJNDz4g9zf///5lF0rmI5P///////////6dj2P///////6BI1aRK1pRD0Yk+zoo+zaBI1Z9I1P///////////4Q7zKBI1P///////4Q7zP///5FBz////5RC0P///6FJ1ZJC0JhF0qtO2H04yYo+zoU8zFmAC+wAAAAfdFJOUwBAnt8gECC/oBtbQL5kgNxj3JZwkO/v74DPu7AwMFBA+hIEAAAF60lEQVRo3tWai7aiOBBFQQlh8X6qKBpA8f8/carygKCo0NOOTFavexHxsq2cOlUJbRiTwwqj3W13jI31DxLecNxv9+iydtZ4x1GBFcbGWjOqeeRB3R2PEcLed+F6FbDhCtiFBF5UiNt1UbxmBdyOau7jqMOxRi1wBdxukTmcqnactuORXqMCdHwR2a5ZmRZCoYCNFkLr2CnWpmmi1WjBjIRYTd1t75w0raq04WMd0rWEWHfxKNc4qsg1a4Ow7Qq0QCYUYB55WIcKFkdI2yY/Dq41oYANN9jdyAPCfYsj/D3rgwI46/EhitaG0wY/hOWGpYfQlHXLnEjDBGD3v1OChWGt9FzDhqB71RGkQJv+NLCxnmucVc81UpnDKwKx3T9XM5vS01kcZpRmSwDgozSfeS0oNhor4N7puVb5aFp+H/ugresJgVDGSnFUMkaXwLqMsWLmtegDfZRFT6jlGpHloOlNoKrreiLFMrilxwMFB+4S2Bw+wOzZsJsRq66AYCdIm8EErGlYUjCWK2p7CSxl87/eABsjqq4A88hRozTkJkDeweI9C6mC06KkQVbmLIVFvYZ6YRACSIk0geAtrCd0sFgFHofdLoStUAJ6YZBq5a5KoHr5b2ENoYNs9o01scMgy2AvAGtqfWyjwxrJR9gD10EpppRkFOALh+fcmZ4OIpeoA/5GwOaIt2Uln4ETyKboZyOn1PMoK6T7gQ26Xl4UNCdj2FiDDTnqLg4huQSs/xEWBeDhD/Rbh8mRy5m2pa4dceG2zyo4cqlMTqF8NqjY6V+WnBYWsj1sJ2HjBrttEGsINqBg6w+weKM8E3brsn54k7DqLeOM3y7vrZkOH3T178zYQUT2rmA7BRsBq4/Hi2BRfXJC0Tsd14MpZtk0LD14tioJ+KMYInvKXEeGFiNLDy7+OfoQWQVrggREoQjbHrb+CEvEjNninlt5xpmEPWslgWpnqAxhKf6AI/WhfPHeR7ZTsKDUpnoDew3e1CJHCvE0lN4J2N7dREIWfAb6K8T8i19UrzS3+xMsGqyxPLLCMj1p9Ad5z/ItLBFCUYz9wWEM68q/cH8HCx7QGo+w11ewGNDCeIDdvoWVpSRX7qxgJZ0Oe/4QWQtYE2O2DBB2uwwWI+g4Dh1f8QQrv9O7yAb7oX/RI/sXYR+s6o9h+RJGBVaDvf5N2GKAzRfCNgOs5fPFYb/y/g7seWAVvfufwYpld9r3F8lXYF116Mhe5hNs18lGBipsLGFb3m77Q2+LhpXOh90+wZ4nYR31jlpozIUlau/N4mJt9zoSqOB6WQ574idsZb9PsKWwU0GTffbZTsHy2Y/CON03YwXgohECK1NtLqwj6ahGd3qAJUqq/OgkmyFVDAXsSa9gA6yl2u2xAmDRCGFVgZ0HK0u7LRCI7EPII6zH+jWQrCeqN5DNAMA6RJP0AGtUkURNdJpgX+NQCx5zDiwG5nBWXReeZ9BKFewBNmOqJ+A4tvDdrXNWbRaeLZzzQS18oq6LtM0sgN2P9pJQAfU16SMdAKz5CdYbHMlWHU7fUw+wJyVmwe2+62e5OjZ9LeBTbl4qY6wAGDp+ArDk9apaLsBOo3vYWx2W9LDlsPjyxHvDSmE7XilsufeZYK+vNuClAnzt7RRYX+51ZbRUmzEZ56My6+0Mm0UvExtFsNBy1fVl31/yz6Ib2Gh4ByJhKb4scrkVcQR/ndzRNhOOmug7CT6wXmfuItrjvY5ZOx/CuvqZE9ZFhpkUJvAUXMuXCtBFgajXb+4m0/F+h/O0byZMYPx0g4RCAbrbBvuvs36GFU83Wv3phlSA/6SAa2Iav4XlTze0HkspIHhSwP7bDxrnwIpFNxZZs7qkzwoIhQLSr2/Qz4NVTzfatm5r+OdXui38FwpQjqfDuuWrjelLwklbtKtAtwWOev3lQ5rJIoC4YwUQpYCVPRXHKKY1j2z4oADfNNY4LFm2Ak0B+8BY6wgkbppKsYbEWO9ALWC3XQsFrPp/Hykt/Gd29e+1wCXgB8T4fwzLWh3pPwaHCY6L2yqyAAAAAElFTkSuQmCC"></image></g><g transform="matrix(0.9999947547912598,0,0,0.9999947547912598,-233.36257934570312,180.001953125)" opacity="0.0029360000007613964" style="display: none;"><g opacity="0.3" transform="matrix(1,0,0,1,462.20098876953125,375.3890075683594)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0,-13.5 C7.455999851226807,-13.5 13.5,-7.455999851226807 13.5,0 C13.5,7.455999851226807 7.455999851226807,13.5 0,13.5 C-7.455999851226807,13.5 -13.5,7.455999851226807 -13.5,0 C-13.5,-7.455999851226807 -7.455999851226807,-13.5 0,-13.5z"></path></g><g opacity="0.7000000000000001" transform="matrix(1,0,0,1,462.20098876953125,375.3890075683594)"><path fill="rgb(255,255,255)" fill-opacity="1" d=" M0,-8.071999549865723 C4.458000183105469,-8.071999549865723 8.071999549865723,-4.458000183105469 8.071999549865723,0 C8.071999549865723,4.458000183105469 4.458000183105469,8.071999549865723 0,8.071999549865723 C-4.458000183105469,8.071999549865723 -8.071999549865723,4.458000183105469 -8.071999549865723,0 C-8.071999549865723,-4.458000183105469 -4.458000183105469,-8.071999549865723 0,-8.071999549865723z"></path></g></g><g class="png" transform="matrix(0.1998000144958496,0,0,0.1998000144958496,160.21141052246094,485.2845764160156)" opacity="0.0032492724142423415" style="display: none;"><image width="172px" height="60px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAKwAAAA8CAMAAADMiaGpAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAB1UExURUdwTJNDz4g9zf///5lF0rmI5P///////////6dj2P///////6BI1aRK1pRD0Yk+zoo+zaBI1Z9I1P///////////4Q7zKBI1P///////4Q7zP///5FBz////5RC0P///6FJ1ZJC0JhF0qtO2H04yYo+zoU8zFmAC+wAAAAfdFJOUwBAnt8gECC/oBtbQL5kgNxj3JZwkO/v74DPu7AwMFBA+hIEAAAF60lEQVRo3tWai7aiOBBFQQlh8X6qKBpA8f8/carygKCo0NOOTFavexHxsq2cOlUJbRiTwwqj3W13jI31DxLecNxv9+iydtZ4x1GBFcbGWjOqeeRB3R2PEcLed+F6FbDhCtiFBF5UiNt1UbxmBdyOau7jqMOxRi1wBdxukTmcqnactuORXqMCdHwR2a5ZmRZCoYCNFkLr2CnWpmmi1WjBjIRYTd1t75w0raq04WMd0rWEWHfxKNc4qsg1a4Ow7Qq0QCYUYB55WIcKFkdI2yY/Dq41oYANN9jdyAPCfYsj/D3rgwI46/EhitaG0wY/hOWGpYfQlHXLnEjDBGD3v1OChWGt9FzDhqB71RGkQJv+NLCxnmucVc81UpnDKwKx3T9XM5vS01kcZpRmSwDgozSfeS0oNhor4N7puVb5aFp+H/ugresJgVDGSnFUMkaXwLqMsWLmtegDfZRFT6jlGpHloOlNoKrreiLFMrilxwMFB+4S2Bw+wOzZsJsRq66AYCdIm8EErGlYUjCWK2p7CSxl87/eABsjqq4A88hRozTkJkDeweI9C6mC06KkQVbmLIVFvYZ6YRACSIk0geAtrCd0sFgFHofdLoStUAJ6YZBq5a5KoHr5b2ENoYNs9o01scMgy2AvAGtqfWyjwxrJR9gD10EpppRkFOALh+fcmZ4OIpeoA/5GwOaIt2Uln4ETyKboZyOn1PMoK6T7gQ26Xl4UNCdj2FiDDTnqLg4huQSs/xEWBeDhD/Rbh8mRy5m2pa4dceG2zyo4cqlMTqF8NqjY6V+WnBYWsj1sJ2HjBrttEGsINqBg6w+weKM8E3brsn54k7DqLeOM3y7vrZkOH3T178zYQUT2rmA7BRsBq4/Hi2BRfXJC0Tsd14MpZtk0LD14tioJ+KMYInvKXEeGFiNLDy7+OfoQWQVrggREoQjbHrb+CEvEjNninlt5xpmEPWslgWpnqAxhKf6AI/WhfPHeR7ZTsKDUpnoDew3e1CJHCvE0lN4J2N7dREIWfAb6K8T8i19UrzS3+xMsGqyxPLLCMj1p9Ad5z/ItLBFCUYz9wWEM68q/cH8HCx7QGo+w11ewGNDCeIDdvoWVpSRX7qxgJZ0Oe/4QWQtYE2O2DBB2uwwWI+g4Dh1f8QQrv9O7yAb7oX/RI/sXYR+s6o9h+RJGBVaDvf5N2GKAzRfCNgOs5fPFYb/y/g7seWAVvfufwYpld9r3F8lXYF116Mhe5hNs18lGBipsLGFb3m77Q2+LhpXOh90+wZ4nYR31jlpozIUlau/N4mJt9zoSqOB6WQ574idsZb9PsKWwU0GTffbZTsHy2Y/CON03YwXgohECK1NtLqwj6ahGd3qAJUqq/OgkmyFVDAXsSa9gA6yl2u2xAmDRCGFVgZ0HK0u7LRCI7EPII6zH+jWQrCeqN5DNAMA6RJP0AGtUkURNdJpgX+NQCx5zDiwG5nBWXReeZ9BKFewBNmOqJ+A4tvDdrXNWbRaeLZzzQS18oq6LtM0sgN2P9pJQAfU16SMdAKz5CdYbHMlWHU7fUw+wJyVmwe2+62e5OjZ9LeBTbl4qY6wAGDp+ArDk9apaLsBOo3vYWx2W9LDlsPjyxHvDSmE7XilsufeZYK+vNuClAnzt7RRYX+51ZbRUmzEZ56My6+0Mm0UvExtFsNBy1fVl31/yz6Ib2Gh4ByJhKb4scrkVcQR/ndzRNhOOmug7CT6wXmfuItrjvY5ZOx/CuvqZE9ZFhpkUJvAUXMuXCtBFgajXb+4m0/F+h/O0byZMYPx0g4RCAbrbBvuvs36GFU83Wv3phlSA/6SAa2Iav4XlTze0HkspIHhSwP7bDxrnwIpFNxZZs7qkzwoIhQLSr2/Qz4NVTzfatm5r+OdXui38FwpQjqfDuuWrjelLwklbtKtAtwWOev3lQ5rJIoC4YwUQpYCVPRXHKKY1j2z4oADfNNY4LFm2Ak0B+8BY6wgkbppKsYbEWO9ALWC3XQsFrPp/Hykt/Gd29e+1wCXgB8T4fwzLWh3pPwaHCY6L2yqyAAAAAElFTkSuQmCC"></image></g><g class="png" transform="matrix(0.5859960317611694,0,0,0.5859960317611694,258.20574951171875,143.2486572265625)" opacity="0.000002947207834864685" style="display: none;"><image width="172px" height="60px" preserveAspectRatio="xMidYMid slice" xlink:href="data:image/png;base64,iVBORw0KGgoAAAANSUhEUgAAAKwAAAA8CAMAAADMiaGpAAAABGdBTUEAALGPC/xhBQAAAAFzUkdCAK7OHOkAAAB1UExURUdwTJNDz4g9zf///5lF0rmI5P///////////6dj2P///////6BI1aRK1pRD0Yk+zoo+zaBI1Z9I1P///////////4Q7zKBI1P///////4Q7zP///5FBz////5RC0P///6FJ1ZJC0JhF0qtO2H04yYo+zoU8zFmAC+wAAAAfdFJOUwBAnt8gECC/oBtbQL5kgNxj3JZwkO/v74DPu7AwMFBA+hIEAAAF60lEQVRo3tWai7aiOBBFQQlh8X6qKBpA8f8/carygKCo0NOOTFavexHxsq2cOlUJbRiTwwqj3W13jI31DxLecNxv9+iydtZ4x1GBFcbGWjOqeeRB3R2PEcLed+F6FbDhCtiFBF5UiNt1UbxmBdyOau7jqMOxRi1wBdxukTmcqnactuORXqMCdHwR2a5ZmRZCoYCNFkLr2CnWpmmi1WjBjIRYTd1t75w0raq04WMd0rWEWHfxKNc4qsg1a4Ow7Qq0QCYUYB55WIcKFkdI2yY/Dq41oYANN9jdyAPCfYsj/D3rgwI46/EhitaG0wY/hOWGpYfQlHXLnEjDBGD3v1OChWGt9FzDhqB71RGkQJv+NLCxnmucVc81UpnDKwKx3T9XM5vS01kcZpRmSwDgozSfeS0oNhor4N7puVb5aFp+H/ugresJgVDGSnFUMkaXwLqMsWLmtegDfZRFT6jlGpHloOlNoKrreiLFMrilxwMFB+4S2Bw+wOzZsJsRq66AYCdIm8EErGlYUjCWK2p7CSxl87/eABsjqq4A88hRozTkJkDeweI9C6mC06KkQVbmLIVFvYZ6YRACSIk0geAtrCd0sFgFHofdLoStUAJ6YZBq5a5KoHr5b2ENoYNs9o01scMgy2AvAGtqfWyjwxrJR9gD10EpppRkFOALh+fcmZ4OIpeoA/5GwOaIt2Uln4ETyKboZyOn1PMoK6T7gQ26Xl4UNCdj2FiDDTnqLg4huQSs/xEWBeDhD/Rbh8mRy5m2pa4dceG2zyo4cqlMTqF8NqjY6V+WnBYWsj1sJ2HjBrttEGsINqBg6w+weKM8E3brsn54k7DqLeOM3y7vrZkOH3T178zYQUT2rmA7BRsBq4/Hi2BRfXJC0Tsd14MpZtk0LD14tioJ+KMYInvKXEeGFiNLDy7+OfoQWQVrggREoQjbHrb+CEvEjNninlt5xpmEPWslgWpnqAxhKf6AI/WhfPHeR7ZTsKDUpnoDew3e1CJHCvE0lN4J2N7dREIWfAb6K8T8i19UrzS3+xMsGqyxPLLCMj1p9Ad5z/ItLBFCUYz9wWEM68q/cH8HCx7QGo+w11ewGNDCeIDdvoWVpSRX7qxgJZ0Oe/4QWQtYE2O2DBB2uwwWI+g4Dh1f8QQrv9O7yAb7oX/RI/sXYR+s6o9h+RJGBVaDvf5N2GKAzRfCNgOs5fPFYb/y/g7seWAVvfufwYpld9r3F8lXYF116Mhe5hNs18lGBipsLGFb3m77Q2+LhpXOh90+wZ4nYR31jlpozIUlau/N4mJt9zoSqOB6WQ574idsZb9PsKWwU0GTffbZTsHy2Y/CON03YwXgohECK1NtLqwj6ahGd3qAJUqq/OgkmyFVDAXsSa9gA6yl2u2xAmDRCGFVgZ0HK0u7LRCI7EPII6zH+jWQrCeqN5DNAMA6RJP0AGtUkURNdJpgX+NQCx5zDiwG5nBWXReeZ9BKFewBNmOqJ+A4tvDdrXNWbRaeLZzzQS18oq6LtM0sgN2P9pJQAfU16SMdAKz5CdYbHMlWHU7fUw+wJyVmwe2+62e5OjZ9LeBTbl4qY6wAGDp+ArDk9apaLsBOo3vYWx2W9LDlsPjyxHvDSmE7XilsufeZYK+vNuClAnzt7RRYX+51ZbRUmzEZ56My6+0Mm0UvExtFsNBy1fVl31/yz6Ib2Gh4ByJhKb4scrkVcQR/ndzRNhOOmug7CT6wXmfuItrjvY5ZOx/CuvqZE9ZFhpkUJvAUXMuXCtBFgajXb+4m0/F+h/O0byZMYPx0g4RCAbrbBvuvs36GFU83Wv3phlSA/6SAa2Iav4XlTze0HkspIHhSwP7bDxrnwIpFNxZZs7qkzwoIhQLSr2/Qz4NVTzfatm5r+OdXui38FwpQjqfDuuWrjelLwklbtKtAtwWOev3lQ5rJIoC4YwUQpYCVPRXHKKY1j2z4oADfNNY4LFm2Ak0B+8BY6wgkbppKsYbEWO9ALWC3XQsFrPp/Hykt/Gd29e+1wCXgB8T4fwzLWh3pPwaHCY6L2yqyAAAAAElFTkSuQmCC"></image></g><g transform="matrix(0.9411653280258179,0,0,0.9411653280258179,400.5613708496094,171.8002166748047)" opacity="0.000012173113368589839" style="display: none;"><path stroke="url(#__lottie_element_790)" stroke-linecap="butt" stroke-linejoin="miter" fill-opacity="0" stroke-miterlimit="4" stroke-opacity="1" stroke-width="3" d=" M-175,-17.25 C-175,-29.81399917602539 -164.81399536132812,-40 -152.25,-40 C-152.25,-40 152.25,-40 152.25,-40 C164.81399536132812,-40 175,-29.81399917602539 175,-17.25 C175,-17.25 175,40 175,40 C175,40 -175,40 -175,40 C-175,40 -175,-17.25 -175,-17.25z"></path><g opacity="1" transform="matrix(1,0,0,1,0,0)"></g></g><g transform="matrix(0,0,0,0,225.62977600097656,163.5154266357422)" opacity="1" style="display: none;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path fill="rgb(3,100,255)" fill-opacity="1" d=" M0,-7 C0,-7 0,-7 0,-7 C3.865999937057495,-7 7,-3.865999937057495 7,0 C7,0 7,0 7,0 C7,3.865999937057495 3.865999937057495,7 0,7 C0,7 0,7 0,7 C-3.865999937057495,7 -7,3.865999937057495 -7,0 C-7,0 -7,0 -7,0 C-7,-3.865999937057495 -3.865999937057495,-7 0,-7z"></path></g></g><g transform="matrix(1.3937851190567017,0,0,1.3937851190567017,142.01939392089844,196.96629333496094)" opacity="0.78" style="display: none;"><g opacity="1" transform="matrix(1,0,0,1,0,0)"><path stroke-linecap="round" stroke-linejoin="round" fill-opacity="0" stroke-dasharray=" 2 4" stroke-dashoffset="180" stroke="rgb(69,81,189)" stroke-opacity="1" stroke-width="1" d="M0 0"></path></g></g></g></svg>