Not yet enjoy'd: so tedious is this day As is the night before some festival To an impatient child that hath new robes
Simile: Waiting for Romeo is like a child waiting to wear new clothes. Shows her youth.
Not yet enjoy'd: so tedious is this day As is the night before some festival To an impatient child that hath new robes
Simile: Waiting for Romeo is like a child waiting to wear new clothes. Shows her youth.
Take him and cut him out in little stars,
Metaphor: Juliet says if Romeo dies, she’d make him into stars so everyone loves night. Ironically, he will die.
Play'd for a pair of stainless maidenhoods:
Innocence: She’s referring to their virginity, which they’ll lose that night.
Prince Escalus. And for that offence 1705Immediately we do exile him hence:
Plot: Prince exiles Romeo instead of death. This is “mercy” but will cause more problems.
Leap to these arms, untalk'd of and unseen.
Theme: Secret love. They must hide their relationship.
Juliet. Gallop apace, you fiery-footed steeds, Towards Phoebus' lodging: such a wagoner 1720As Phaethon would whip you to the west, And bring in cloudy night immediately.
Juliet can’t wait for night so Romeo can come. She’s passionate and eager.
Prince Escalus. And for that offence 1705Immediately we do exile him hence:
Plot: Prince exiles Romeo instead of death. This is “mercy” but will cause more problems.
Romeo slew Tybalt, Romeo must not live.
Lady Capulet demands Romeo’s death. She’s vengeful.
Benvolio. Tybalt, here slain, whom Romeo's hand did slay; Romeo that spoke him fair, bade him bethink 1670How nice the quarrel was, and urged withal Your high displeasure: all this uttered With gentle breath, calm look, knees humbly bow'd, Could not take truce with the unruly spleen Of Tybalt deaf to peace, but that he tilts 1675With piercing steel at bold Mercutio's breast, Who all as hot, turns deadly point to point, And, with a martial scorn, with one hand beats Cold death aside, and with the other sends It back to Tybalt, whose dexterity, 1680Retorts it: Romeo he cries aloud, 'Hold, friends! friends, part!' and, swifter than his tongue, His agile arm beats down their fatal points, And 'twixt them rushes; underneath whose arm 1685An envious thrust from Tybalt hit the life Of stout Mercutio, and then Tybalt fled; But by and by comes back to Romeo, Who had but newly entertain'd revenge, And to 't they go like lightning, for, ere I 1690Could draw to part them, was stout Tybalt slain. And, as he fell, did Romeo turn and fly. This is the truth, or let Benvolio die.
Plot: Benvolio tells the truth about the fight, defending Romeo.
Benvolio. O noble prince, I can discover all The unlucky manage of this fatal brawl: 1660There lies the man, slain by young Romeo, That slew thy kinsman, brave Mercutio.
Plot: Benvolio tells the truth about the fight, defending Romeo.
Romeo. O, I am fortune's fool!
Theme: Fate. Romeo feels like a puppet of bad luck.
Staying for thine to keep him company: Either thou, or I, or both, must go with him. Tybalt. Thou, wretched boy, that didst consort him here, Shalt with him hence. 1640 Romeo. This shall determine that. [They fight; TYBALT falls]
Plot: Romeo kills Tybalt in a duel. Now he’s a murderer.
Romeo. Alive, in triumph! and Mercutio slain! 1630Away to heaven, respective lenity, And fire-eyed fury be my conduct now!
Romeo switches from peace to fury. He wants revenge for Mercutio
Benvolio. O Romeo, Romeo, brave Mercutio's dead!
Plot: Confirmation of death. The turning point of the play.
Romeo. This gentleman, the prince's near ally, My very friend, hath got his mortal hurt In my behalf; my reputation stain'd With Tybalt's slander,—Tybalt, that an hour Hath been my kinsman! O sweet Juliet, 1620Thy beauty hath made me effeminate And in my temper soften'd valour's steel!
Romeo feels responsible and thinks love made him weak (“effeminate”).
rogue, a villain, that fights by the book of arithmetic! Why the devil came you between us? I was hurt under your arm.
Tragedy: Mercutio says Romeo got in the way. This guilt will haunt Romeo.
Mercutio. No, 'tis not so deep as a well, nor so wide as a church-door; but 'tis enough,'twill serve: ask for me to-morrow, and you shall find me a grave man.
Even dying, Mercutio makes puns (“grave man”). Shows his wit and bravery.
A plague o' both your houses!
Theme: Mercutio curses both families. He dies blaming their feud.
[TYBALT under ROMEO's arm stabs MERCUTIO, and flies with his followers] Mercutio. I am hurt.
Plot: The fatal moment. Romeo’s interference accidentally gets Mercutio killed.
Romeo. Gentle Mercutio, put thy rapier up.
Romeo steps in to keep peace, showing his new maturity.
Mercutio. Good king of cats, nothing but one of your nine lives; that I mean to make bold withal, and as you shall use me hereafter, drybeat the rest of the eight. Will you pluck your sword out of his pitcher by the ears? make haste, lest mine be about your 1580ears ere it be out.
Mercutio calls Tybalt “king of cats” (mockery) and challenges him to fight.
Mercutio. O calm, dishonourable, vile submission!
Mercutio is disgusted by Romeo’s peacefulness. He sees it as cowardly.
Romeo. Tybalt, the reason that I have to love thee 1560Doth much excuse the appertaining rage To such a greeting: villain am I none; Therefore farewell; I see thou know'st me not.
Plot: Romeo secretly loves Tybalt now (he’s family through Juliet). He refuses to fight.
No better term than this,—thou art a villain.
Insult: “Villain” is a serious insult, challenging Romeo’s honor.
Tybalt. Well, peace be with you, sir: here comes my man.
Insult: Tybalt calls Romeo his servant, showing disrespect.
Tybalt. Mercutio, thou consort'st with Romeo,—
Pun: Tybalt says “consort’st” (associate). Mercutio pretends it means “musician” and threatens with his “fiddlestick” (sword).
Mercutio. And but one word with one of us? couple it with something; make it a word and a blow.
Wordplay: “A word and a blow” means talk then fight. Mercutio is looking for a fight.
Mercutio. By my heel, I care not.
Mercutio is defiant and doesn’t fear the Capulets. He’s confident and unafraid.
Mercutio. Nay, an there were two such, we should have none shortly, for one would kill the other. Thou! why, thou wilt quarrel with a man that hath a hair more, 1515or a hair less, in his beard, than thou hast: thou wilt quarrel with a man for cracking nuts, having no other reason but because thou hast hazel eyes: what eye but such an eye would spy out such a quarrel? Thy head is as fun of quarrels as an egg is full of 1520meat, and yet thy head hath been beaten as addle as an egg for quarrelling: thou hast quarrelled with a man for coughing in the street, because he hath wakened thy dog that hath lain asleep in the sun: didst thou not fall out with a tailor for wearing 1525his new doublet before Easter? with another, for tying his new shoes with old riband? and yet thou wilt tutor me from quarrelling!
Mercutio’s exaggeration. Hyperbole: Mercutio claims Benvolio would fight over anything, even a cough or someone’s shoes. This is funny exaggeration.
Mercutio. Thou art like one of those fellows that when he enters the confines of a tavern claps me his sword upon the table and says 'God send me no need of 1505thee!' and by the operation of the second cup draws it on the drawer, when indeed there is no need. Benvolio. Am I like such a fellow? Mercutio. Come, come, thou art as hot a Jack in thy mood as any in Italy, and as soon moved to be moody, and as 1510soon moody to be moved.
Mercutio mocks Benvolio. Mercutio jokes that Benvolio is quick to fight. He’s teasing, showing his playful, sarcastic side.
Benvolio. I pray thee, good Mercutio, let's retire: The day is hot, the Capulets abroad, 1500And, if we meet, we shall not scape a brawl; For now, these hot days, is the mad blood stirring.
Benvolio is the peacekeeper. He wants to avoid a fight because it's hot and people are angry (“mad blood stirring”).
het regime
National newspaper now using 'regime' to describe the cabinet level behaviour
Die aspecten van het trumpisme – wetteloosheid, vijanddenken, leugens, omdraaiingen en geweld – zijn de pijlers van zowel zijn binnenlandse als buitenlandse beleid. Dat is al tien jaar het geval,
The aspects of trumpism, lawlessness, thinking in terms of enemies, lies, Orwellian spinning, violence, are the pillars of domestic and foreign policy.
Editorial opinion in Dutch national newspaper "The naive belief everything will turn out fine in the USA must end"
exemplifies
ilustreert
https://ipfs.io/ipns/k51qzi5uqu5div1auuxm59ygav4p7gdg9z4e9iggtu6m43rmc3xw75mczx2b7x/
identify ipns local name
on thinkpad IPFS Desktop IPNS Name is
🧊ipfs k51qzi5uqu5div1auuxm59ygav4p7gdg9z4e9iggtu6m43rmc3xw75mczx2b7x
new version that contains stuff addsd under IPFS version added card to about section
Bank Account Details
Can we hide this for US?
Settlement Details
Can we hide this for US?
Capture and Refund Settings
Can we hide this for US?
Transaction Limits
Can we hide this for US?
Business Website Details
Can we hide this for US?
Assisted Onboarding
Can we hide this section for US?
completion
In test and live mode below, can we change API keys begin with rzp_test_us_ and rzp_live_us_
macOS Sonoma (14.x) • macOS Sequoia (15.x)
For 16.1 and 17.0 we will be supporting
macOS Sequoia (15.x) macOS Tahoe (26.x)
BFSI
Route not available in US
Automate
Can we have images changed here. Showing rupee symbol for US also
Plugins
Can we please include Magento in this list?
Shopify Integration
Shopify isnt live in the US yet
the autopoiesis of the IndyWeb
x
eLife Assessment
This manuscript proposes a lateralized, lobe-specific brain-liver sympathetic neurocircuit regulating hepatic glucose metabolism and presents anatomical evidence for sympathetic crossover at the porta hepatis using viral tracing and neuromodulation approaches. While the topic is of important significance and the methodologies are, in principle, state-of-the-art, significant concerns regarding experimental design, incomplete methodological reporting, sparse and ambiguous labeling, and overi-nterpretation of the data substantially weaken support for the study's central conclusions, thereby limiting the study's completeness. The work will be of interest to biologists, clinicians, and physiologists.
Reviewer #1 (Public review):
Summary:
This manuscript by Wang et al. reports the potential involvement of an asymmetric neurocircuit in the sympathetic control of liver glucose metabolism.
Strengths:
The concept that the contralateral brain-liver neurocircuit preferentially regulates each liver lobe may be interesting.
Weaknesses:
However, the experimental evidence presented did not support the study's central conclusion.
(1) Pseudorabies virus (PRV) tracing experiment:<br /> The liver not only possesses sympathetic innervations but also vagal sensory innervations. The experimental setup failed to distinguish whether the PRV-labeling of LPGi (Lateral Paragigantocellular Nucleus) is derived from sympathetic or vagal sensory inputs to the liver.
(2) Impact on pancreas:<br /> The celiac ganglia not only provide sympathetic innervations to the liver but also to the pancreas, the central endocrine organ for glucose metabolism. The chemogenetic manipulation of LPGi failed to consider a direct impact on the secretion of insulin and glucagon from the pancreas.
(3) Neuroanatomy of the brain-liver neurocircuit:<br /> The current study and its conclusion are based on a speculative brain-liver sympathetic circuit without the necessary anatomical information downstream of LPGi.
(4) Local manipulation of the celiac ganglia:<br /> The left and right ganglia of mice are not separate from each other but rather anatomically connected. The claim that the local injection of AAV in the left or right ganglion without affecting the other side is against this basic anatomical feature.
Reviewer #2 (Public review):
Summary:
The manuscript by Wang and colleagues aims to determine whether the left and right LPGi differentially regulate hepatic glucose metabolism and to reveal decussation of hepatic sympathetic nerves.
The authors used tissue clearing to identify sympathetic fibers in the liver lobes, then injected PRV into the hepatic lobes. Five days post-injection, PRV-labeled neurons in the LPGi were identified. The results indicated contralateral dominance of premotor neurons and partial innervation of more than one lobe. Then the authors activated each side of the LPGi, resulting in a greater increase in blood glucose levels after right-sided activation than after left-sided activation, as well as changes in protein expression in the liver lobes. These data suggested modulation of HGP (hepatic glucose production) in a lobe-specific manner. Chemical denervation of a particular lobe did not affect glucose levels due to compensation by the other lobes. In addition, nerve bundles decussate in the hepatic portal region.
Strengths:
The manuscript is timely and relevant. It is important to understand the sympathetic regulation of the liver and the contribution of each lobe to hepatic glucose production. The authors use state-of-the-art methodology.
Weaknesses:
(1) The wording/terminology used in the manuscript is misleading, and it is not used in the proper context. For instance, the goal of the study is "to investigate whether cerebral hemispheres differentially regulate hepatic glucose metabolism..." (see abstract); however, the authors focus on the brainstem (a single structure without hemispheres). Similarly, symmetric is not the best word for the projections.
(2) Sparse labeling of liver-related neurons was shown in the LPGi (Figure 1). It would be ideal to have lower magnification images to show the area. Higher quality images would be necessary, as it is difficult to identify brainstem areas. The low number of labeled neurons in the LPGi after five days of inoculation is surprising. Previous findings showed extensive labeling in the ventral brainstem at four days post-inoculation (Desmoulins et al., 2025). Unfortunately, it is not possible to compare the injection paradigm/methods because the PRV inoculation is missing from the methods section. If the PRV is different from the previously published viral tracers, time-dependent studies to determine the order of neurons and the time course of infection would be necessary.
(3) Not all LPGi cells are liver-related. Was the entire LPGi population stimulated, or was it done in a cell-type-specific manner? What was the strain, sex, and age of the mice? What was the rationale for using the particular viral constructs?
(4) The authors should consider the effect of stimulation of double-labeled neurons (innervating more than one lobe) and potential confounding effects regarding other physiological functions.
(5) The authors state that "central projections directly descend along the sympathetic chain to the celiac-superior mesenteric ganglia". What they mean is unclear. Do the authors refer to pre-ganglionic neurons or premotor neurons? How does it fit with the previous literature?
(6) How was the chemical denervation completed for the individual lobes?
(7) The Western Blot images look like they are from different blots, but there are no details provided regarding protein amount (loading) or housekeeping. What was the reason to switch beta-actin and alpha-tubulin? In Figures 3F -G, the GS expression is not a good representative image. Were chemiluminescence or fluorescence antibodies used? Were the membranes reused?
(8) Key references using PRV for liver innervation studies are missing (Stanley et al, 2010 [PMID: 20351287]; Torres et al., 2021 [PMID: 34231420]; Desmoulins et al., 2025 [PMID: 39647176]).
Reviewer #3 (Public review):
Summary:
This study found a lobe-specific, lateralized control of hepatic glucose metabolism by the brain and provides anatomical evidence for sympathetic crossover at the porta hepatis. The findings are particularly insightful to the researchers in the field of liver metabolism, regeneration, and tumors.
Strengths:
Increasing evidence suggests spatial heterogeneity of the liver across many aspects of metabolism and regenerative capacity. The current study has provided interesting findings: neuronal innervation of the liver also shows anatomical differences across lobes. The findings could be particularly useful for understanding liver pathophysiology and treatment, such as metabolic interventions or transplantation.
Weaknesses:
Inclusion of detailed method and Discussion:
(1) The quantitative results of PRV-labeled neurons are presented, and please include the specific quantitative methods.
(2) The Discussion can be expanded to include potential biological advantages of this complex lateralized innervation pattern.
Reviewer #4 (Public review):
Summary:
The studies here are highly informative in terms of anatomical tracing and sympathetic nerve function in the liver related to glucose levels, but given that they are performed in a single species, it is challenging to translated them to humans, or to determine whether these neural circuits are evolutionarily conserved. Dual-labeling anatomical studies are elegant, and the addition of chemogenetic and optogenetic studies is mechanistically informative. Denervation studies lack appropriate controls, and the role of sensory innervation in the liver is overlooked.
Specific Weaknesses - Major:
(1) The species name should be included in the title.
(2) Tyrosine hydroxylase was used to mark sympathetic fibers in the liver, but this marker also hits a portion of sensory fibers that need to be ruled out in whole-mount imaging data
(3) Chemogenetic and optogenetic data demonstrating hyperglycemia should be described in the context of prior work demonstrating liver nerve involvement in these processes. There is only a brief mention in the Discussion currently, but comparing methods and observations would be helpful.
(4) Sympathetic denervation with 6-OHDA can drive compensatory increases to tissue sensory innervation, and this should be measured in the liver denervation studies to implicate potential crosstalk, especially given the increase in LPGi cFOS that may be due to afferent nerve activity. Compensatory sympathetic drive may not be the only culprit, though it is clearly assumed to be. The sensory or parasympathetic/vagal innervation of the liver is altogether ignored in this paper and could be better described in general.
Author response:
Public Reviews:
Reviewer #1 (Public review):
Summary:
This manuscript by Wang et al. reports the potential involvement of an asymmetric neurocircuit in the sympathetic control of liver glucose metabolism.
Strengths:
The concept that the contralateral brain-liver neurocircuit preferentially regulates each liver lobe may be interesting.
Weaknesses:
However, the experimental evidence presented did not support the study's central conclusion.
We sincerely thank the reviewer for recognizing the conceptual novelty of our work and for constructive comments aimed at enhancing its rigor and clarity. In response, we will carry out targeted experiments to address the points raised, including: (i) further characterization of LPGi projections to vagal and sympathetic circuits; (ii) evaluation of potential pancreatic involvement; and (ii) validation of the specificity of chemogenetic activation within the proposed circuit. We anticipate completing the revised version within 8 weeks.
(1) Pseudorabies virus (PRV) tracing experiment:
The liver not only possesses sympathetic innervations but also vagal sensory innervations. The experimental setup failed to distinguish whether the PRV-labeling of LPGi (Lateral Paragigantocellular Nucleus) is derived from sympathetic or vagal sensory inputs to the liver.
Thank you for raising this important point. We fully agree that the liver receives both sympathetic and vagal sensory innervation, and we acknowledge that PRV-based tracing alone does not definitively distinguish between these two pathways. This represents a limitation of the original experimental design.
Based on established anatomical literature as well as our experimental observations, vagal sensory neuron cell bodies reside in the nodose ganglion (NG), and their central projections terminate predominantly in the nucleus of the solitary tract (NTS) (Nature. 2023;623(7986):387-396; Curr Biol. 2020;30(20):3986-3998.e5.), which is located in the dorsomedial medulla. In contrast, the LPGi, together with other sympathetic-related nuclei, is predominantly distributed in the ventral medulla (Cell Metab. 2025;37(11):2264-2279.e10; Nat Commun. 2022;13(1):5079.).
To directly assess the contribution of vagal sensory pathways, we will perform an additional PRV tracing experiment using two groups of mice: one with bilateral nodose ganglion (NG) removal and a sham-operated control group. Identical PRV injections will be delivered to the liver in both groups, and PRV labeling in the LPGi will be quantitatively compared. Preservation of LPGi labeling following NG ablation would indicate that PRV transmission occurs primarily via sympathetic, rather than vagal sensory, pathways. These data will be incorporated into the revised manuscript and are expected to be completed within 3 weeks.
(2) Impact on pancreas:
The celiac ganglia not only provide sympathetic innervations to the liver but also to the pancreas, the central endocrine organ for glucose metabolism. The chemogenetic manipulation of LPGi failed to consider a direct impact on the secretion of insulin and glucagon from the pancreas.
Thank you for this important comment. We agree that the celiac ganglia (CG) provide sympathetic innervation not only to the liver but also to the pancreas, which plays a central role in glucose homeostasis through the secretion of both insulin and glucagon. Therefore, the potential pancreatic implications associated with LPGi chemogenetic manipulation worth careful consideration.
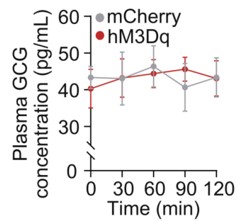
To address this concern, we examined circulating glucagon levels following chemogenetic manipulation of the LPGi. As shown in the Supplementary Figure below, plasma glucagon (GCG) concentrations were not significantly altered at 30, 60, 90, or 120 minutes compared with control mice (n = 6), indicating that LPGi manipulation does not measurably affect glucagon secretion under our experimental conditions.
We acknowledge that insulin secretion was not assessed in the study, which represents an important limitation given the pancreatic innervation of the CG. To further strengthen our interpretation, we are performing additional experiments in newly prepared mice to measure circulating insulin levels following LPGi manipulation. These data together with Author response image 1 below will be included in the revised manuscript upon completion.
Author response image 1.
Plasma concentrations of GCG in mice following LPGi GABAergic neurons activation.
(3) Neuroanatomy of the brain-liver neurocircuit:<br /> The current study and its conclusion are based on a speculative brain-liver sympathetic circuit without the necessary anatomical information downstream of LPGi.
Thank you for raising this important point. A clear anatomical definition of the downstream pathways linking the brain to the liver is essential for interpreting the proposed brain-liver sympathetic circuit.
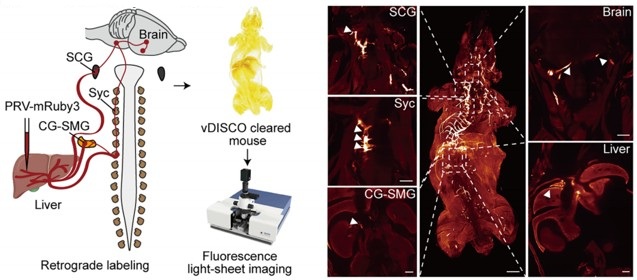
However, the present study (Figure 4A) provides direct anatomical evidence supporting the organization of the brain–liver sympathetic neurocircuit. These observations are consistent with our recent detailed characterization of the brain-liver sympathetic circuit published in Cell Metabolism (Cell Metab. 2025;37(11):2264–2279), LPGi GABAergic neurons inhibit GABAergic neurons in the caudal ventrolateral medulla (CVLM). Disinhibition of CVLM reduces GABAergic suppression of rostral ventrolateral medulla (RVLM) neurons, which are key excitatory drivers of sympathetic tone. RVLM neurons project to sympathetic preganglionic neurons in the sympathetic chain (Syc). These neurons synapse with postganglionic sympathetic neurons in ganglia such as the celiac-superior mesenteric ganglion (CG-SMG). Postganglionic sympathetic fibers then innervate the liver, releasing NE to activate hepatic β<sub>2</sub>-adrenergic receptors and stimulate HGP.
Together, these data establish a coherent anatomical basis for the proposed brain-liver sympathetic pathway and clarify the downstream organization relevant to the functional experiments presented here.
Author response image 2.
Tracing scheme (Left) and whole-mount imaging (Right) of PRV-labeled brain-liver neurocircuit. Scale bars, 3,000 (whole mount) or 1,000 (optical sections) μm.
(4) Local manipulation of the celiac ganglia:<br /> The left and right ganglia of mice are not separate from each other but rather anatomically connected. The claim that the local injection of AAV in the left or right ganglion without affecting the other side is against this basic anatomical feature.
Thank you for raising this important anatomical point. We fully acknowledge that the left and right celiac ganglia (CG) in mice are interconnected, and that unilateral viral injection could theoretically affect the contralateral side. The celiac–superior mesenteric ganglion (CG-SMG) complex serves as a major sympathetic hub that regulates visceral organ functions. Recent transcriptomic, anatomical, and functional studies have revealed that the CG-SMG is not a homogeneous structure but is composed of molecularly and functionally distinct neuronal populations. These populations exhibit specialized projection patterns and regulate different aspects of gastrointestinal physiology, supporting a model of modular sympathetic control. (Nature. 2025 Jan;637(8047):895-902). Therefore, we were aware of this phenomenon during the initial stages of these experiments.
To minimize unintended spread to the contralateral CG, we took two complementary approaches.
First, we optimized the injection strategy by using an extremely small injection volume (100 nL per site), with a very slow infusion rate (50 nL/min), and fine glass micropipettes. With these refinements, contralateral viral spread was rarely observed.
Second, and importantly, all animals included in the final analyses were subjected to post hoc anatomical verification. After completion of the experiments, CG were collected, sectioned, and examined for viral expression. As shown in Supplementary Figure 5F, only mice in which viral expression was strictly confined to the targeted CG, with no detectable infection in the contralateral ganglion, were included in the presented data.
Together, these measures ensure that the reported effects are attributable to local manipulation of the intended CG. We will ensure that the Methods section more explicitly details these technical precautions and that the legend for Figure S5F clearly states its role in validating injection specificity.
Reviewer #2 (Public review):
Summary:
The manuscript by Wang and colleagues aims to determine whether the left and right LPGi differentially regulate hepatic glucose metabolism and to reveal decussation of hepatic sympathetic nerves.
The authors used tissue clearing to identify sympathetic fibers in the liver lobes, then injected PRV into the hepatic lobes. Five days post-injection, PRV-labeled neurons in the LPGi were identified. The results indicated contralateral dominance of premotor neurons and partial innervation of more than one lobe. Then the authors activated each side of the LPGi, resulting in a greater increase in blood glucose levels after right-sided activation than after left-sided activation, as well as changes in protein expression in the liver lobes. These data suggested modulation of HGP (hepatic glucose production) in a lobe-specific manner. Chemical denervation of a particular lobe did not affect glucose levels due to compensation by the other lobes. In addition, nerve bundles decussate in the hepatic portal region.
We thank the reviewer for the thorough and constructive evaluation of our manuscript. In direct response, we will undertake comprehensive revisions to enhance the rigor and clarity of the study, including: (i) correcting ambiguous or misleading terminology pertaining to anatomical resolution and sympathetic circuit organization; (ii) expanding the Methods section with complete experimental details, improved image presentation, and explicit justification of our viral and genetic approaches; and (iii) strengthening data interpretation by addressing issues related to sparse PRV labeling, projection heterogeneity, and the functional implications of double-labeled neurons. All revisions are expected to be completed within 8 weeks.
Strengths:
The manuscript is timely and relevant. It is important to understand the sympathetic regulation of the liver and the contribution of each lobe to hepatic glucose production. The authors use state-of-the-art methodology.
Weaknesses:
(1) The wording/terminology used in the manuscript is misleading, and it is not used in the proper context. For instance, the goal of the study is "to investigate whether cerebral hemispheres differentially regulate hepatic glucose metabolism..." (see abstract); however, the authors focus on the brainstem (a single structure without hemispheres). Similarly, symmetric is not the best word for the projections.
We thank the reviewer for raising these critical points regarding terminology and conceptual framing. We acknowledge that certain phrases in our original manuscript may have been overly broad or ambiguous, particularly in describing the scope of sympathetic heterogeneity and the specificity of neural projections. Due to practical constraints and the scope of our study, our investigation is focused on the brainstem, which represents the final common pathway for these lateralized commands. We acknowledge that terms referring to the cerebral hemispheres do not accurately describe our study.
We are revising the manuscript to ensure accurate and consistent terminology and will submit the revised version with these corrections.
(2) Sparse labeling of liver-related neurons was shown in the LPGi (Figure 1). It would be ideal to have lower magnification images to show the area. Higher quality images would be necessary, as it is difficult to identify brainstem areas. The low number of labeled neurons in the LPGi after five days of inoculation is surprising. Previous findings showed extensive labeling in the ventral brainstem at four days post-inoculation (Desmoulins et al., 2025). Unfortunately, it is not possible to compare the injection paradigm/methods because the PRV inoculation is missing from the methods section. If the PRV is different from the previously published viral tracers, time-dependent studies to determine the order of neurons and the time course of infection would be necessary.
We sincerely thank the reviewer for these detailed and constructive comments regarding the PRV tracing experiments. We fully agree that careful presentation and interpretation of the anatomical data are essential for ensuring rigor and transparency. We address each point in detail below.
(1) Image magnification and anatomical context of LPGi labeling
We agree that the original images did not sufficiently convey the broader anatomical context of the LPGi. In the revised manuscript, we will replace the original panels in Figure 1 with new images that include lower-magnification overviews of the brainstem, alongside higher-magnification views of the LPGi. These images clearly delineate the LPGi with respect to established anatomical landmarks and atlas boundaries. Image contrast and resolution will also be optimized to allow unambiguous identification of PRV-labeled neurons and surrounding structures.
(2) Sparse LPGi labeling at 5 days post-injection and methodological details
We apologize for the omission of the detailed PRV injection protocol in the original Methods section. We deliberately used small-volume, focal injections (1 µL per liver lobe) to minimize viral spread and to restrict labeling to circuits specifically connected to the targeted hepatic region. Under these conditions, early-stage or intermediate-order upstream nuclei such as the LPGi are expected to exhibit relatively sparse labeling compared to more proximal autonomic nuclei. This information will add, including the PRV strain, viral titer, injection volume, precise injection coordinates, and surgical procedures.
(3) Not all LPGi cells are liver-related. Was the entire LPGi population stimulated, or was it done in a cell-type-specific manner? What was the strain, sex, and age of the mice? What was the rationale for using the particular viral constructs?
We thank the reviewer for this insightful and important question. We agree that not all neurons within the LPGi are liver-related, and we apologize that our rationale was not clearly articulated in the original manuscript.
(1) Our decision to target GABAergic neurons in the LPGi using Gad1-Cre mice was based on prior experimental evidence rather than an assumption about the entire LPGi population. In our previous study (Cell Metab. 2025;37(11):2264-2279.e10), we performed single-cell RNA sequencing on retrogradely labeled LPGi neurons following liver tracing. These analyses revealed that the majority of liver-projecting LPGi neurons are GABAergic in nature. Based on these findings, we chose to selectively manipulate GABAergic neurons in the LPGi rather than the entire LPGi neuronal population, in order to achieve greater cellular specificity and to minimize potential confounding effects arising from heterogeneous neuron types within this region. We regret that this rationale was not clearly described in the original submission and have now revised the manuscript to explicitly state this reasoning.
(2) In addition, we apologize for the omission of mouse strain, sex, and age information in the Methods section. These details will be fully added.
(3) We selected AAV-based viral vectors, specifically the AAV9 serotype, due to their well-established efficiency in transducing neurons in the brainstem, relatively low toxicity, and widespread use in circuit-level chemogenetic and optogenetic studies. When combined with Cre-dependent viral constructs in Gad1-Cre mice, this approach enabled selective and reliable manipulation of LPGi GABAergic neurons.
(4) The authors should consider the effect of stimulation of double-labeled neurons (innervating more than one lobe) and potential confounding effects regarding other physiological functions.
We thank the reviewer for raising this important point. We agree that neurons innervating more than one liver lobe could, in principle, introduce potential confounding effects and may reflect higher-order integrative autonomic neurons.
This consideration is consistent with a key finding of the cited study: the celiac-superior mesenteric ganglion (CG-SMG) contains molecularly distinct sympathetic neuron populations (e.g., RXFP1<sup>+</sup> vs. SHOX2<sup>+</sup>) that exhibit complementary organ projections and separate, non‑overlapping functions. Specifically, RXFP1<sup>+</sup> neurons innervate secretory organs (pancreas, bile duct) to regulate secretion, while SHOX2<sup>+</sup> neurons innervate the gastrointestinal tract to control motility. This functional segregation supports the concept of specialized autonomic modules rather than a uniform,“fight or flight”response, reinforcing the need for careful interpretation of circuit-specific manipulations. (Nature. 2025;637(8047):895-902; Neuron. Published online December 10, 2025).
In our PRV tracing experiments, the proportion of double-labeled neurons was relatively small, suggesting that the majority of labeled LPGi neurons preferentially associate with individual hepatic lobes. Nevertheless, we recognize that activation of this minority population could contribute to broader physiological effects beyond strictly lobe-specific regulation. We acknowledge that the absence of single-cell-level resolution in the current study limits our ability to further dissect the functional heterogeneity of these projection-defined neurons, and we will explicitly state this as a limitation in the revised manuscript. We will explicitly acknowledge this possibility in the revised manuscript and included it as a limitation of the current study. We thank the reviewer for highlighting this important conceptual consideration.
(5) The authors state that "central projections directly descend along the sympathetic chain to the celiac-superior mesenteric ganglia". What they mean is unclear. Do the authors refer to pre-ganglionic neurons or premotor neurons? How does it fit with the previous literature?
We thank the reviewer for pointing out this imprecise wording. We agree that the original phrasing was anatomically inaccurate and potentially confusing. The pathways we intended to describe involve brainstem premotor neurons that project to sympathetic preganglionic neurons in the spinal cord. These preganglionic neurons then innervate neurons in the celiac–superior mesenteric ganglia, which in turn provide postganglionic input to the liver.
We are revising the manuscript to clearly distinguish premotor from preganglionic neurons and to describe this pathway in a manner consistent with the established organization of sympathetic autonomic circuits reported in the previous literature. The revised wording will explicitly reflect this hierarchical relay structure.
(6) How was the chemical denervation completed for the individual lobes?
We thank the reviewer for raising this important methodological concern. We agree that potential diffusion of 6-OHDA is a critical issue when performing lobe-specific chemical denervation, and we apologize that our original description did not sufficiently clarify how this was controlled.
In the revised Methods section, we will provide a detailed description of the denervation procedure, including the injection volume and concentration of 6-OHDA, as well as the physical separation and isolation of individual hepatic lobes during application to minimize diffusion to adjacent tissue.
To directly assess the specificity of the chemical denervation, we included immunofluorescence and Western blot analyses demonstrating a selective reduction of sympathetic markers in the targeted lobe, with minimal effects on non-targeted lobes. These results support the effectiveness and relative spatial confinement of the 6-OHDA treatment under our experimental conditions.
We thank the reviewer for highlighting this point, which has helped us improve both the clarity and rigor of the manuscript.
(7) The Western Blot images look like they are from different blots, but there are no details provided regarding protein amount (loading) or housekeeping. What was the reason to switch beta-actin and alpha-tubulin? In Figures 3F -G, the GS expression is not a good representative image. Were chemiluminescence or fluorescence antibodies used? Were the membranes reused?
We thank the reviewer for this careful and detailed evaluation of the Western blot data. We apologize that insufficient methodological detail was provided in the original submission.
(1) We would like to clarify that the protein bands shown within each panel were derived from the same membrane. To improve transparency, we will provide full, uncropped images of the corresponding membranes in the supplementary materials. In addition, detailed information regarding protein loading amounts, gel conditions, and housekeeping controls will be added to the Methods section.
(2) The use of different loading controls (β-actin or α-tubulin) reflects a technical consideration rather than an experimental inconsistency. In our experiments, the molecular weight of the TH (62kDa) was too close to α-tubulin (55kDa), and β-actin (42kDa) was therefore used to avoid band overlap and to ensure accurate quantification.
(3) Regarding the GS signal shown in Figures 3F–G, we agree that the original representative image was suboptimal. This appears to be related to antibody performance rather than sample quality. To address this, we are repeating the GS Western blot using a newly validated antibody. The original tissue samples had been aliquoted and stored at −80 °C, allowing reliable re-analysis. This work will be done in 8 weeks.
(4) All Western blot experiments were detected using chemiluminescence, and membrane stripping and reprobing procedures are now explicitly described in the Methods section.
We thank the reviewer for highlighting these issues, which significantly improve the rigor and clarity of our data presentation.
(8) Key references using PRV for liver innervation studies are missing (Stanley et al, 2010 [PMID: 20351287]; Torres et al., 2021 [PMID: 34231420]; Desmoulins et al., 2025 [PMID: 39647176]).
We thank the reviewer for pointing out these important and highly relevant references that were inadvertently omitted in our initial submission. The studies by Stanley et al. (Proc Natl Acad Sci U S A, 2010), Torres et al. (Am J Physiol Regul Integr Comp Physiol, 2021), and Desmoulins et al. (Auton Neurosci, 2025) represent key PRV-based retrograde tracing work that has mapped central neural circuits innervating the liver and thus provide essential context for our anatomical analyses.
We agree that inclusion of these studies is necessary to properly situate our findings within the existing literature. Accordingly, we will incorporate citations to these references in the revised manuscript and discuss their relationship to our results.
Reviewer #3 (Public review):
Summary:
This study found a lobe-specific, lateralized control of hepatic glucose metabolism by the brain and provides anatomical evidence for sympathetic crossover at the porta hepatis. The findings are particularly insightful to the researchers in the field of liver metabolism, regeneration, and tumors.
Strengths:
Increasing evidence suggests spatial heterogeneity of the liver across many aspects of metabolism and regenerative capacity. The current study has provided interesting findings: neuronal innervation of the liver also shows anatomical differences across lobes. The findings could be particularly useful for understanding liver pathophysiology and treatment, such as metabolic interventions or transplantation.
Weaknesses:
Inclusion of detailed method and Discussion:
We sincerely thank the reviewer for the positive and constructive feedback, which will significantly enhance both the methodological rigor and the broader biological interpretation of our study. In direct response, we will revise the Discussion to elaborate on the potential physiological advantages of a lateralized and lobe-specific pattern of liver innervation. Furthermore, we will expand the Methods section to include a comprehensive description of the quantitative analysis applied to PRV-labeled neurons. Together, these revisions will strengthen the manuscript’s clarity, depth, and relevance to researchers in hepatic metabolism, regeneration, and disease. We expect to complete all updates within 8 weeks.
(1) The quantitative results of PRV-labeled neurons are presented, and please include the specific quantitative methods.
We thank the reviewer for this helpful suggestion. We will add a detailed description of the quantitative methods used to analyze PRV-labeled neurons in the revised Methods section. This includes information on the counting criteria, the brain regions analyzed, how the regions of interest were delineated, and the normalization procedures applied to obtain the reported neuron counts.
(2) The Discussion can be expanded to include potential biological advantages of this complex lateralized innervation pattern.
We appreciate the reviewer’s suggestion. We will expand the Discussion to include a paragraph addressing the potential biological significance of lateralized liver innervation. We highlight that this asymmetric organization could allow for more precise, lobe-specific regulation of hepatic metabolism, enable integration of distinct physiological signals, and potentially provide robustness against perturbations. These points will discuss in the revised manuscript.
Reviewer #4 (Public review):
Summary:
The studies here are highly informative in terms of anatomical tracing and sympathetic nerve function in the liver related to glucose levels, but given that they are performed in a single species, it is challenging to translated them to humans, or to determine whether these neural circuits are evolutionarily conserved. Dual-labeling anatomical studies are elegant, and the addition of chemogenetic and optogenetic studies is mechanistically informative. Denervation studies lack appropriate controls, and the role of sensory innervation in the liver is overlooked.
We sincerely appreciate the reviewer's thoughtful evaluation and fully agree that findings derived from a single-species model must be interpreted with caution in relation to human physiology. In direct response, we will revise the manuscript to explicitly clarify that all experimental data were obtained in mice and to provide a discussion of the limitations regarding direct extrapolation to humans. Concurrently, we will expand the Discussion section by integrating our findings with recent human and translational studies, including a multicenter clinical trial demonstrating that catheter-based endovascular denervation of the celiac and hepatic arteries significantly improved glycemic control in patients with poorly controlled type 2 diabetes, without major adverse events (Signal Transduct Target Ther. 2025;10(1):371). While our current work focuses on defining the anatomical organization and functional asymmetry of this circuit in mice, the clinical findings suggest that the core principles, sympathetic control of hepatic glucose metabolism via CG-liver pathways, may be conserved and of translational relevance. Additionally, we will clarify the interpretation of tyrosine hydroxylase labeling and expand the discussion of hepatic sensory and parasympathetic innervation, acknowledging their important roles in liver–brain communication and identifying them as key directions for future research. Collectively, these revisions will provide a more balanced, clinically informed, and rigorous framework for interpreting our findings, and we aim to complete all updates within 8 weeks.
Specific Weaknesses - Major:
(1) The species name should be included in the title.
We thank the reviewer for this suggestion. We agree that the species should be clearly indicated. The findings presented in this study were obtained in mice using tissue clearing and whole-organ imaging approaches. Due to technical limitations, these observations are currently limited to the mouse strain. We will update the title and clarified the species used throughout the manuscript.
(2) Tyrosine hydroxylase was used to mark sympathetic fibers in the liver, but this marker also hits a portion of sensory fibers that need to be ruled out in whole-mount imaging data
We thank the reviewer for pointing this out. We acknowledge that tyrosine hydroxylase (TH) labels not only sympathetic fibers but also a subset of sensory fibers. We will add a limitation of this point in the revised manuscript. In addition, ongoing experiments using retrograde PRV labeling from the liver, combined with sectioning, are being used to distinguish sympathetic fibers from vagal and dorsal root ganglion–derived sensory fibers. These data will be included in a forthcoming update of the manuscript and are expected to be completed in approximately 6 weeks.
(3) Chemogenetic and optogenetic data demonstrating hyperglycemia should be described in the context of prior work demonstrating liver nerve involvement in these processes. There is only a brief mention in the Discussion currently, but comparing methods and observations would be helpful.
We thank the reviewer for this suggestion. Previous studies largely relied on electrical stimulation to modulate liver innervation, which provides relatively coarse control of neural activity (Eur J Biochem. 1992;207(2):399-411). By contrast, our use of chemogenetic and optogenetic approaches allows selective, cell-type–specific manipulation of LPGi neurons. We will revise the Discussion to place our functional data in the context of prior work, highlighting how these more precise approaches improve understanding of the contribution of liver-innervating neurons to hyperglycemia.
(4) Sympathetic denervation with 6-OHDA can drive compensatory increases to tissue sensory innervation, and this should be measured in the liver denervation studies to implicate potential crosstalk, especially given the increase in LPGi cFOS that may be due to afferent nerve activity. Compensatory sympathetic drive may not be the only culprit, though it is clearly assumed to be. The sensory or parasympathetic/vagal innervation of the liver is altogether ignored in this paper and could be better described in general.
We thank the reviewer for this insightful comment and agree that chemical sympathetic denervation with 6-OHDA may induce compensatory changes in non-sympathetic hepatic inputs, including sensory and parasympathetic (vagal) innervation. As the reviewer correctly points out, increased LPGi cFOS activity may reflect afferent nerve engagement rather than solely compensatory sympathetic drive.
More broadly, we agree that the central nervous system functions as an integrated homeostatic network that continuously processes diverse afferent signals, including hepatic sensory and vagal inputs, as well as other interoceptive cues. From this perspective, the LPGi cFOS changes observed in our study likely represent one component of a complex integrative response rather than evidence for a single dominant pathway.
We acknowledge that the present study did not directly assess hepatic sensory or parasympathetic innervation, which represents a limitation in scope. In the revised manuscript, we will expand the Discussion to explicitly note this limitation and provide a more balanced consideration of potential crosstalk among sympathetic, sensory, and parasympathetic pathways in shaping LPGi activity following hepatic denervation.
Recommendations for the authors:
Reviewer #2 (Recommendations for the authors):
Although the findings are interesting, this reviewer has major concerns about the experimental design, methodology, results, and interpretation of the data. Experimental details are lacking, including basic information (age, sex, strain of mice, procedures, magnification, etc.).
We thank the reviewer for this important recommendation. We agree that comprehensive reporting of experimental details is essential for rigor and reproducibility.
In the revised manuscript, we will add complete information regarding mouse strain, sex, age, and sample size for each experiment. In addition, detailed descriptions of surgical procedures, viral constructs, injection parameters, imaging magnification, and analysis methods have been incorporated into the Methods section.
These revisions ensure that all experiments are described with sufficient technical detail and clarity to allow accurate interpretation and replication of our findings.
Reviewer #3 (Recommendations for the authors):
Addressing a few questions might help:
(1) The study found that liver-associated LPGi neurons are predominantly GABAergic. It would be informative to molecularly characterize the PRV-traced, liver-projecting LPGi neurons to determine their neurochemical phenotypes.
We thank the reviewer for this insightful suggestion. We agree that molecular characterization of liver-projecting LPGi neurons is important for understanding their functional identity.
This issue has been addressed in detail in our recent study (Cell Metab. 2025;37(11):2264-2279.e10), in which we performed single-cell RNA sequencing on retrogradely traced LPGi neurons connected to the liver. These analyses demonstrated that the majority of liver-projecting LPGi neurons are GABAergic, with a defined transcriptional profile distinct from neighboring non–liver-related populations.
Based on these findings, the current study selectively targets GABAergic LPGi neurons using Gad1-Cre mice. We are now explicitly referencing and summarizing these molecular results in the revised manuscript to clarify the neurochemical identity of the PRV-traced LPGi neurons.
(2) Is it possible to do a local microinjection of a sodium channel blocker (e.g., lidocaine) or an adrenergic receptor antagonist into the porta hepatis? That would potentially provide additional evidence for the porta hepatis as the functional crossover point.
We appreciate the reviewer’s thoughtful suggestion. While pharmacological blockade at the porta hepatis could modulate local neural activity, the proposed approach may not fully capture the distinction between ipsilateral and contralateral inputs, and may not conclusively establish neural crossover at this particular site.
In our view, the anatomical evidence provided by whole-mount tissue clearing, dual-labeled tracing, and direct visualization of decussating nerve bundles at the porta hepatis offers a more definitive demonstration of sympathetic crossover. Pharmacological blockade would affect both crossed and uncrossed fibers simultaneously and therefore would not specifically resolve the anatomical organization of this decussation.
Nevertheless, we agree that functional interrogation of the porta hepatis represents an interesting direction for future work, and we will now acknowledge this possibility in the Discussion.
(3) It is possible to investigate the effects of unilateral LPGi manipulation or ablation of one side of CG/SMG on liver metabolism, such as hyperglycemia?
We thank the reviewer for this important suggestion. We agree that unilateral ablation or silencing of the CG-SMG could provide additional insight into lateralized sympathetic control of liver metabolism.
However, precise and selective ablation of one side of the CG-SMG through 6-OHDA without affecting the contralateral ganglion or adjacent autonomic structures remains technically challenging, particularly given the anatomical connectivity between the two sides. We are currently optimizing approaches to achieve reliable unilateral manipulation.
If successful within the revision timeframe, we will include these experiments and corresponding metabolic analyses in the revised manuscript. If not, we will explicitly discuss this experimental limitation and the predicted metabolic consequences of unilateral CG-SMG ablation as an important direction for future studies. This work will be done in 6 weeks.
Reviewer #4 (Recommendations for the authors):
In the abstract and elsewhere, the use of the term 'sympathetic release' is unclear - do you mean release of nerve products, such as the neurotransmitter norepinephrine? This should be more clearly defined.
We thank the reviewer for pointing out this ambiguity. We agree that the term “sympathetic release” was imprecise. In the revised manuscript, we will explicitly refer to the release of sympathetic neurotransmitters, primarily norepinephrine, from postganglionic sympathetic fibers.
We will revise the wording throughout the manuscript to ensure accurate and consistent terminology and to avoid potential confusion regarding the underlying neurobiological mechanisms.
eLife Assessment
The findings are important, as they identify MIRO1 as a central regulator linking mitochondrial positioning and respiratory chain function to VSMC proliferation, neointima formation, and human vasoproliferative disease. Overall, the strength of evidence is convincing, with comprehensive in vivo and in vitro data, including human cells and added bioenergetic analyses, that broadly support the main claims despite some remaining limitations in mechanistic and mitochondrial assays.
Reviewer #1 (Public review):
Summary:
In this paper, the authors investigate the effects of Miro1 on VSMC biology after injury. Using conditional knockout animals, they provide the important observation that Miro1 is required for neointima formation. They also confirm that Miro1 is expressed in human coronary arteries. Specifically, in conditions of coronary diseases, it is localized in both media and neointima and, in atherosclerotic plaque, Miro1 is expressed in proliferating cells.
However, the role of Miro1 in VSMC in CV diseases is poorly studied and the data available are limited; therefore, the authors decided to deepen this aspect. The evidence that Miro-/- VSMCs show impaired proliferation and an arrest in S phase is solid and further sustained by restoring Miro1 to control levels, normalizing proliferation. Miro1 also affects mitochondrial distribution, which is strikingly changed after Miro1 deletion. Both effects are associated with impaired energy metabolism due to the ability of Miro1 to participate in MICOS/MIB complex assembly, influencing mitochondrial cristae folding. Interestingly, the authors also show the interaction of Miro1 with NDUFA9, globally affecting super complex 2 assembly and complex I activity.<br /> Finally, these important findings also apply to human cells and can be partially replicated using a pharmacological approach, proposing Miro1 as a target for vasoproliferative diseases.
Strengths:
The discovery of Miro1 relevance in neointima information is compelling, as well as the evidence in VSMC that MIRO1 loss impairs mitochondrial cristae formation, expanding observations previously obtained in embryonic fibroblasts.<br /> The identification of MIRO1 interaction with NDUFA9 is novel and adds value to this paper. Similarly, the findings that VSMC proliferation requires mitochondrial ATP support the new idea that these cells do not rely mostly on glycolysis.
The revised manuscript includes additional data supporting mitochondrial bioenergetic impairment in MIRO1 knockout VSMCs. Measurements of oxygen consumption rate (OCR), along with Complex I (ETC-CI) and Complex V activity, have been added and analyzed across multiple experimental conditions. Collectively, these findings provide a more comprehensive characterization of the mitochondrial functional state. Following revision, the association between MIRO1 deficiency and impaired Complex I activity is more robust.
Although the precise molecular mechanism of action remains to be fully elucidated, in this updated version, experiments using a MIRO1 reducing agent are presented with improved clarity
Although some limitations remain, the authors have addressed nearly all the concerns raised, and the manuscript has substantially improved
Weaknesses:
Figure 6: The authors do not address the concern regarding the cristae shape; however, characterization of the cristae phenotype with MIRO1 ΔTM would have strengthened the mechanistic link between MIRO1 and the MIB/MICOS complex
Although the authors clarified their reasoning, they did not explore in vivo validation of key biochemical findings, which represents a limitation of the current study. While their justification is acknowledged, at least a preliminary exploratory effort could have been evaluated to reinforce the translational relevance of the study.
Finally, in line with the explanations outlined in the rebuttal, the Discussion section should mention the limits of MIRO1 reducer treatment.
Reviewer #2 (Public review):
Summary:
This study identifies the outer‑mitochondrial GTPase MIRO1 as a central regulator of vascular smooth muscle cell (VSMC) proliferation and neointima formation after carotid injury in vivo and PDGF-stimulation ex vivo. Using smooth muscle-specific knockout male mice, complementary in vitro murine and human VSMC cell models, and analyses of mitochondrial positioning, cristae architecture and respirometry, the authors provide solid evidence that MIRO1 couples mitochondrial motility with ATP production to meet the energetic demands of the G1/S cell cycle transition. However, a component of the metabolic analyses are suboptimal and would benefit from more robust methodologies. The work is valuable because it links mitochondrial dynamics to vascular remodelling and suggests MIRO1 as a therapeutic target for vasoproliferative diseases, although whether pharmacological targeting of MIRO1 in vivo can effectively reduce neointima after carotid injury has not been explored. This paper will be of interest to those working on VSMCs and mitochondrial biology.
Strengths:
The strength of the study lies in its comprehensive approach assessing the role of MIRO1 in VSMC proliferation in vivo, ex vivo and importantly in human cells. The subject provides mechanistic links between MIRO1-mediated regulation of mitochondrial mobility and optimal respiratory chain function to cell cycle progression and proliferation. Finally, the findings are potentially clinically relevant given the presence of MIRO1 in human atherosclerotic plaques and the available small molecule MIRO1.
Weaknesses:
(1) High-resolution respirometry (Oroboros) to determine mitochondrial ETC activity in permeabilized VSMCs would be informative.
(2) Therapeutic targeting of MIRO1 failed to prevent neointima formation, however, the technical difficulties of such an experiment is appreciated.
Reviewer #3 (Public review):
Summary:
This study addresses the role of MIRO1 in vascular smooth muscle cell proliferation, proposing a link between MIRO1 loss and altered growth due to disrupted mitochondrial dynamics and function. While the findings are useful for understanding the importance of mitochondrial positioning and function in this specific cell type, the main bioenergetic and mechanistic claims are not strongly supported.
Strengths:
This study focuses on an important regulatory protein, MIRO1, and its role in vascular smooth muscle cell (VSMC) proliferation, a relatively underexplored context.
This study explores the link between smooth muscle cell growth, mitochondrial dynamics, and bioenergetics, which is a significant area for both basic and translational biology.
The use of both in vivo and in vitro systems provides a useful experimental framework to interrogate MIRO1 function in this context.
Weaknesses:
The proposed link between MIRO1 and respiratory supercomplex biogenesis or function is not clearly defined.
Completeness and integration of mitochondrial assays is marginal, undermining the strength of the conclusions regarding oxidative phosphorylation.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews
Reviewer #1 (Public review):
Summary:
In this paper, the authors investigate the effects of Miro1 on VSMC biology after injury. Using conditional knockout animals, they provide the important observation that Miro1 is required for neointima formation. They also confirm that Miro1 is expressed in human coronary arteries. Specifically, in conditions of coronary diseases, it is localized in both media and neointima, and, in atherosclerotic plaque, Miro1 is expressed in proliferating cells.
However, the role of Miro1 in VSMC in CV diseases is poorly studied, and the data available are limited; therefore, the authors decided to deepen this aspect. The evidence that Miro-/- VSMCs show impaired proliferation and an arrest in S phase is solid and further sustained by restoring Miro1 to control levels, normalizing proliferation. Miro1 also affects mitochondrial distribution, which is strikingly changed after Miro1 deletion. Both effects are associated with impaired energy metabolism due to the ability of Miro1 to participate in MICOS/MIB complex assembly, influencing mitochondrial cristae folding. Interestingly, the authors also show the interaction of Miro1 with NDUFA9, globally affecting super complex 2 assembly and complex I activity.
Finally, these important findings also apply to human cells and can be partially replicated using a pharmacological approach, proposing Miro1 as a target for vasoproliferative diseases.
Strengths:
The discovery of Miro1 relevance in neointima information is compelling, as well as the evidence in VSMC that MIRO1 loss impairs mitochondrial cristae formation, expanding observations previously obtained in embryonic fibroblasts.
The identification of MIRO1 interaction with NDUFA9 is novel and adds value to this paper. Similarly, the findings that VSMC proliferation requires mitochondrial ATP support the new idea that these cells do not rely mostly on glycolysis.
Weaknesses:
(1) Figure 3:
I appreciate the system used to assess mitochondrial distribution; however, I believe that time-lapse microscopy to evaluate mitochondrial movements in real time should be mandatory. The experimental timing is compatible with time-lapse imaging, and these experiments will provide a quantitative estimation of the distance travelled by mitochondria and the fraction of mitochondria that change position over time. I also suggest evaluating mitochondrial shape in control and MIRO1-/- VSMC to assess whether MIRO1 absence could impact mitochondrial morphology, altering fission/fusion machinery, since mitochondrial shape could differently influence the mobility.
Mitochondrial motility experiments. WT and Miro1-/- VSMCs were transiently transfected with mito-ds-red and untargeted GFP adenoviruses to fluorescently label mitochondria and cytosol, respectively. Live-cell fluorescence confocal microscopy was used to acquire mitochondrial images at one-minute intervals over a 25-30-minute period. WT cells exhibited dynamic reorganization of the mitochondrial network, whereas Miro1-/- VSMCs displayed minimal mitochondrial movement, characterized only by limited oscillatory behavior without network remodeling (Supplemental Video 1).
Mitochondrial shape (form factor) was assessed by confocal microscopy in WT and Miro1-/- VSMCs. Analysis of the mitochondrial form factor (defined as the ratio of mitochondrial length to width) during cell cycle progression revealed morphological changes in wild type (WT) cells, characterized by an increase in form factor. In contrast, Miro1-/- cells exhibited no significant alterations in mitochondrial morphology (Figure 3- Figure supplement 1B).
(2) Figure 6:
The evidence of MIRO1 ablation on cristae remodeling is solid; however, considering that the mechanism proposed to explain the finding is the modulation of MICOS/MIB complex, as shown in Figure 6D, I suggest performing EM analysis in each condition. In my mind, Miro1 KK and Miro1 TM should lead to different cristae phenotypes according to the different impact on MICOS/MIB complex assembly. Especially, Miro1 TM should mimic Miro1 -/- condition, while Miro1 KK should drive a less severe phenotype. This would supply a good correlation between Miro1, MICOS/MIB complex formation and cristae folding.
I also suggest performing supercomplex assembly and complex I activity with each plasmid to correlate MICOS/MIB complex assembly with the respiratory chain efficiency.
Complex I activity assays revealed that overexpression of MIRO1-WT fully restored enzymatic activity in MIRO1-/- cells, whereas MIRO1-KK provided partial rescue. In contrast, a MIRO1 mutant lacking the transmembrane domain failed to restore activity and resembled the Miro1-/- phenotype (Figure 6- Figure supplement 2).
The Complex I activity in each Miro1 mutant correlated with the degree of MICOS/MIB complex assembly in pulldown assays, implying a functional link between Miro1 and mitochondrial cristae organization.
Moreover, an in-gel Complex V activity assay was performed to evaluate the enzymatic activity of mitochondrial ATP synthase in a native gel following electrophoresis. To normalize the activity signal, a Blue Native PAGE of the same samples was probed for the ATP5F1 subunit. A modest, yet statistically significant reduction in Complex V activity was observed in Miro1-/- cells (Figure 6- Figure supplement 1).
(3) I noticed that none of the in vitro findings have been validated in an in vivo model. I believe this represents a significant gap that would be valuable to address. In your animal model, it should not be too complex to analyze mitochondria by electron microscopy to assess cristae morphology. Additionally, supercomplex assembly and complex I activity could be evaluated in tissue homogenates to corroborate the in vitro observations.
We appreciate the reviewer’s comment. However, our currently available samples have been processed by light microscopy and are therefore not suitable for embedding for light for electron microscopy.
(4) I find the results presented in Figure S7 somewhat unclear. The authors employ a pharmacological strategy to reduce Miro1 and validate the findings previously obtained with the genetic knockout model. They report increased mitophagy and a reduction in mitochondrial mass. However, in my opinion, these changes alone could significantly impact cellular metabolism. A lower number of mitochondria would naturally result in decreased ATP production and reduced mitochondrial respiration. This, in turn, weakens the proposed direct link between Miro1 deletion and impaired metabolic function or altered electron transport chain (ETC) activity. I believe this section would benefit from additional experiments and a more in-depth discussion.
We initially conducted experiments using the MIRO1 reducer to explore the translational potential of our findings. These experiments aimed to provide a foundation in vivo studies. However, despite multiple attempts, we were unable to demonstrate a significant effect of MIRO1reducer, delivered via a Pluronic gel, on the mitochondria of the vascular wall. Of note, he role of MIRO1 in mitophagy has been well-established in several studies (for example, PMID: 34152608), which show that genetic deletion of Miro1 delays the translocation of the E3 ubiquitin ligase Parkin onto damaged mitochondria, thereby reducing mitochondrial clearance in fibroblasts and cultured neurons. Furthermore, loss of Miro1 in the hippocampus and cortex increases mitofusin levels with the appearance of hyperfused mitochondria and activation of the integrated stress response. Thus, MIRO1 deletion in genetic models does not result in a substantial reduction of mitochondria but causes hyperfused mitochondria. The rationale for developing the MIRO1 reducer stems from genetic forms of Parkinson’s disease, where Miro1 is retained in PD cells but degraded in healthy cells following mitochondrial depolarization (PMID: 31564441). Thus, the degradation of mutant MIRO1 by the reducer does not phenocopy the effects of genetic MIRO1 depletion. Thus, we believe the data with the reducer demonstrate that MIRO1 can be acutely targeted in vitro, but the mechanism of action (as the reviewer points out, the reduction of mitochondrial mass may lead to decreased ATP levels, potentially reducing cell proliferation) differs from that of chronic genetic deletion. In fact, we observe somewhat increased mitochondrial length in MIRO1-/- cells. We acknowledge that this is complex and have revised the paragraph to clarify the use of the MIRO1 reducer.
Reviewer #2 (Public review):
Summary:
This study identifies the outer mitochondrial GTPase MIRO1 as a central regulator of vascular smooth muscle cell (VSMC) proliferation and neointima formation after carotid injury in vivo and PDGF-stimulation ex vivo. Using smooth muscle-specific knockout male mice, complementary in vitro murine and human VSMC cell models, and analyses of mitochondrial positioning, cristae architecture, and respirometry, the authors provide solid evidence that MIRO1 couples mitochondrial motility with ATP production to meet the energetic demands of the G1/S cell cycle transition. However, a component of the metabolic analyses is suboptimal and would benefit from more robust methodologies. The work is valuable because it links mitochondrial dynamics to vascular remodeling and suggests MIRO1 as a therapeutic target for vasoproliferative diseases, although whether pharmacological targeting of MIRO1 in vivo can effectively reduce neointima after carotid injury has not been explored. This paper will be of interest to those working on VSMCs and mitochondrial biology.
Strengths:
The strength of the study lies in its comprehensive approach, assessing the role of MIRO1 in VSMC proliferation in vivo, ex vivo, and importantly in human cells. The subject provides mechanistic links between MIRO1-mediated regulation of mitochondrial mobility and optimal respiratory chain function to cell cycle progression and proliferation. Finally, the findings are potentially clinically relevant given the presence of MIRO1 in human atherosclerotic plaques and the available small molecule MIRO1.
Weaknesses:
(1) There is a consistent lack of reporting across figure legends, including group sizes, n numbers, how many independent experiments were performed, or whether the data is mean +/- SD or SEM, etc. This needs to be corrected.
These data were added in the revised manuscript.
(2) The in vivo carotid injury experiments are in male mice fed a high-fat diet; this should be explicitly stated in the abstract, as it's unclear if there are any sex- or diet-dependent differences. Is VSMC proliferation/neointima formation different in chow-fed mice after carotid injury?
This is an important point, and we appreciate the feedback. In this model, the transgene is located on the Y chromosome. As a result, only male mice can be studied. However, in our previous experiments, we have not observed any sex-dependent changes in neointimal formation. Additionally, please note that smooth muscle cell proliferation in neointimal formation is enhanced in models of cholesterol-fed mice on a high-fat diet.
(3) The main body of the methods section is thin, and it's unclear why the majority of the methods are in the supplemental file. The authors should consider moving these to the main article, especially in an online-only journal.
We thank the reviewer for this suggestion. We moved the methods to the main manuscript.
(4) Certain metabolic analyses are suboptimal, including ATP concentration and Complex I activity measurements. The measurement of ATP/ADP and ATP/AMP ratios for energy charge status (luminometer or mass spectrometry), while high-resolution respirometry (Oroboros) to determine mitochondrial complex I activity in permeabilized VSMCs would be more informative.
ATP/ADP and ATP/AMP ratios were assessed on samples from WT and Miro1-/- VSMCs using an ATP/ADP/AMP Assay Kit (Cat#: A-125) purchased from Biomedical Research Service, University at Buffalo, New York). Miro1-/- samples exhibited reduced ATP levels accompanied by elevated concentrations of ADP and AMP. As a result, both ATP/ADP and ATP/AMP ratios were significantly lower in MIRO1-/- cells compared to WT, indicating impaired cellular energy homeostasis (Figure 5B, C).
(5) The statement that 'mitochondrial mobility is not required for optimal ATP production' is poorly supported. XF Seahorse analysis should be performed with nocodazole and also following MIRO1 reconstitution +/- EF hands.
To evaluate the metabolic effects of Nocodazole, we conducted Seahorse metabolic assays on vascular smooth muscle cells with various conditions (VSMCs). We used WT VSMCs, Miro1-/- VSMCs, and Miro1-/- VSMCs that expressed either MIRO1-WT, KK, or ΔTM mutants.Our results demonstrate that Nocodazole exposure did not compromise mitochondrial respiratory activity. However, Miro1-/- VSMCs displayed a trend toward reduced basal and maximal mitochondrial respiration when compared to WT cells. This deficit was only partially corrected by the expression of the MIRO1-KK mutant. In contrast, reintroducing MIRO1-WT through adenoviral delivery fully restored mitochondrial respiration to normal levels (Figure 5- Figure supplement 1).
(6) The authors should consider moving MIRO1 small molecule data into the main figures. A lot of value would be added to the study if the authors could demonstrate that therapeutic targeting of MIRO1 could prevent neointima formation in vivo.
We appreciate the reviewer's comment and attempted the suggested in vivo experiments using the commercially available Miro1 reducer. For these experiments, we used a pluronic gel to deliver the reducer to the adventitial area surrounding the carotid artery. Despite numerous attempts to optimize the experimental conditions, we were unable to reliably detect a significant effect of the reducer on mitochondria in the vascular wall.
Reviewer #3 (Public review):
Summary:
This study addresses the role of MIRO1 in vascular smooth muscle cell proliferation, proposing a link between MIRO1 loss and altered growth due to disrupted mitochondrial dynamics and function. While the findings are potentially useful for understanding the importance of mitochondrial positioning and function in this specific cell type within health and disease contexts, the evidence presented appears incomplete, with key bioenergetic and mechanistic claims lacking adequate support.
Strengths:
(1)The study focuses on an important regulatory protein, MIRO1, and its role in vascular smooth muscle cell (VSMC) proliferation, a relatively underexplored context.
(2) It explores the link between smooth muscle cell growth, mitochondrial dynamics, and bioenergetics, which is a potentially significant area for both basic and translational biology.
(3) The use of both in vivo and in vitro systems provides a potentially useful experimental framework to interrogate MIRO1 function in this context.
Weaknesses:
(1) The central claim that MIRO1 loss impairs mitochondrial bioenergetics is not convincingly demonstrated, with only modest changes in respiratory parameters and no direct evidence of functional respiratory chain deficiency.
(2) The proposed link between MIRO1 and respiratory supercomplex assembly or function is speculative, lacking mechanistic detail and supported by incomplete or inconsistent biochemical data.
(3) Key mitochondrial assays are either insufficiently controlled or poorly interpreted, undermining the strength of the conclusions regarding oxidative phosphorylation.
(4) The study does not adequately assess mitochondrial content or biogenesis, which could confound interpretations of changes in respiratory activity.
(5) Overall, the evidence for a direct impact of MIRO1 on mitochondrial respiratory function in the experimental setting is weak, and the conclusions overreach the data.
Recommendations for the authors:
Reviewer #3 (Recommendations for the authors):
(1) Throughout the manuscript, the authors incorrectly use "mobility" to describe the active transport of mitochondria. The appropriate term is "mitochondrial motility," which refers to active, motor-driven movement. "Mobility" implies passive diffusion and is not scientifically accurate in this context.
(2) "Super complex" should be consistently written as "supercomplex," in line with accepted mitochondrial biology terminology.
We thank the reviewer for this comment and revised the text accordingly.
(3) A significant limitation of the in vivo model is the mild phenotype observed, which is expected from an inducible knockout system. The authors should clarify whether a constitutive, tissue-specific knockout was considered and, if not, whether embryonic lethality or another limitation prevented its generation.
This genetic model was originally developed by Dr. Janet Shaw at the University of Utah. In the original publication, Miro1 was constitutively knocked out in neurons. Germline inactivation of Miro1 was achieved by crossing mice harboring the Miro1F allele with a mouse line expressing Cre recombinase under the control of the hypoxanthine-guanine phosphoribosyltransferase (HPRT) promoter. Mating Miro1+/− mice resulted in Miro1−/− animals, which were cyanotic and died shortly after birth. Due to this outcome, we opted to develop an inducible, smooth muscle-specific model. Additionally, we considered testing whether the acute use of an inhibitor or a knockdown system targeting Miro1 could be evaluated as a potential therapeutic approach.
(4) In Figure 1A and S1A, the authors use Western blotting to validate the knockout in the aorta and IHC in carotid arteries. The choice of different methods does not seem justified, and qPCR data are shown only for the aorta. IHC appears to be suboptimal for assessing MIRO1 levels in vascular tissue due to high autofluorescence, and IHC in Figure S1A is merely qualitative, with no quantification provided.
We present complementary approaches to validate the deletion of Miro1. For Western blot analysis, we used the aorta because it provides more material for analysis. The autofluorescence observed via immunofluorescence is characteristic of elastin fibers within the media layer, making our results typical for this technique. As shown in Figure 1- Figure supplement 1, our data demonstrate a significant decrease, if not a complete knockout, of the target protein specifically in smooth muscle cells.
(5) In Figure 1G, the bottom left panel (magnification) shows a lower green signal than the top left panel, suggesting these may have been collected with different signal intensity. This raises concerns about image consistency and representation.
Top images in Figure 1G are taken at magnification 63x. Bottom images were made at magnification 20x. The intensity is different between the two magnifications, but similar between genotypes.
(6) In Figure S3, the sampling is uncontrolled: the healthy subject and the patient differ markedly in age. The claim of colocalization is not substantiated with any quantitative analysis.
As outlined in the Methods section, our heart samples were obtained from LVAD patients or explanted hearts from transplant recipients. Due to the limited availability of such samples, there is indeed a difference in age between the healthy subject and the patient. While we acknowledge this limitation, the scarcity of samples made it challenging to control for age. Additionally, we determined that performing a quantitative analysis of colocalization would not yield robust or meaningful data given the constraints of our sample size and variability.
(7) Figure S4A lacks statistical analysis, which is necessary for interpreting the data shown.
This appears to be a misunderstanding. In this manuscript, we do present statistically significant differences and focus on those that are biologically meaningful. Specifically, we highlight differences between PDGF treatment versus no treatment within the same genotype, as well as differences between the two genotypes under the same treatment condition (control or PDGF treatment). In this particular case, there is only a statistical difference between WT+PDGF and SM-Miro1-/, but since this is not a meaningful comparison, it is not shown. Please note that this approach applies to all figures in the manuscript. Including all comparisons—whether statistically significant or not, and whether biologically meaningful or not—may appear rigorous but in our opinion, ultimately detracts from the main message of this paper.
(8) The authors state, "given the generally poor proliferation of VSMCs from SM-MIRO1-/- mice, in later experiments we used VSMCs from MIRO1fl/fl mice and infected them with adenovirus expressing cre." This is not convincing, especially since in vivo cre efficiency is generally lower than in vitro. Moreover, the methods indicate that "VSMCs from littermate controls were subjected to the same procedure with empty vector control adenovirus," yet in Figure 2A, the control appears to be MIRO1fl/fl VSMCs transduced with Ad-EV. The logic and consistency of the controls used need clarification.
For the initial experiments, cells were explanted from SM-MIRO1-/- mice (Figure 2- Figure supplement 1). In these mice, Cre recombination had occurred in vivo, and the cells exhibited very poor growth. In fact, their growth was so limited that we decided not to pursue this experimental approach after three independent experiments.
For subsequent experiments, cells were explanted from Miro1fl/fl mice and passaged several times, which allowed us to generate the number of cells required for the experiments (Figure 2B). Once sufficient Miro1fl/fl cells were obtained, they were treated with adenovirus expressing Cre, as described in the Methods section. Control cells were treated with an empty vector adenovirus. To clarify, the control cells are Miro1fl/fl cells infected with an empty vector adenovirus, while the MIRO1-/- cells are Miro1fl/fl cells infected with adenovirus expressing Cre. The statement that “littermate controls were used” is incorrect as in fact, Miro1fl/fl cells from the same preparation were either infected with an empty vector adenovirus, or with adenovirus expressing Cre. As mentioned, the knockdown was confirmed by Western blotting.
(9) Figure 2C shows a growth delay in MIRO1-/- cells. Have the authors performed additional time points to determine when these cells return to G1 and quantify the duration of the lag?
This is an excellent suggestion. So far, we have not performed this experiment.
(10) In the 24 h time point of Figure 2C, MIRO1-/- cells appear to be cycling, yet no cyclin E signal is detected. How do the authors explain this inconsistency? Additionally, in Figure 2H, the quantification of cyclin E is unreliable, given that lanes 3 and 4 show no detectable signal.
We agree with the reviewer—the inconsistency is driven by the exposure of the immunoblot presented. We revisited the data, reviewed the quantification, and performed an additional experiment. We are now presenting an exposure that demonstrates levels of cyclin E (Figure 2G).
(11) In Figure 3D, the authors present mitochondrial probability map vs. distance from center curves. How was the "center" defined in this analysis? Were radial distances normalized across cells (e.g., to the cell radius or maximum extent)? If not, variation in cell and/or nucleus size or shape could significantly affect the resulting profiles. No statistical analysis is provided for this assessment, which undermines its quantitative value. Furthermore, the rationale behind the use of mito95 values is not clearly explained.
The center refers to the center of the microchip's Y-shaped pattern, to which each cell is attached. Since all Y-shapes on the chip are identical in size, normalization is not required. The size of the optimal Y-shapes was tested as recommended by CYTOO. For further context, please refer to the papers by the Kittler group.
Additionally, a graph demonstrating the percentage of mitochondria localized at specific distances can be produced for any given distance. Notably, the further from the center of the chip, the more pronounced the differences become.
(12) The authors apply a 72 h oligomycin treatment to assess proliferation and a 16 h treatment to measure ATP levels. This discrepancy in experimental design is not justified in the manuscript. The length of treatment directly impacts the interpretation of the data in Figures 4C, 4D, and 4E, and needs to be addressed.
Thank you for this comment. We have performed additional experiments to align these time points. In the revised manuscript, we now present proliferation and ATP production measured at the same time point (Figure 4A, B for proliferation and ATP levels).
(13) The manuscript repeatedly suggests that MIRO1 loss causes a defect in mitochondrial ATP production, yet no direct demonstration of a bioenergetic defect is provided. The claim relies on a modest decrease in supercomplex species (of undefined composition) and a mild reduction in complex I activity that does not support a substantial OXPHOS defect. Notably, the respirometry data in Figure 5I do not align with the BN-PAGE results in Figure 6I. There is increasing evidence that respiratory chain supercomplexes do not confer a catalytic advantage. The authors should directly assess the enzymatic activities of all respiratory complexes. Reported complex I activity in MIRO1-/- cells appears rotenone-like (virtually zero, figure 3K) or ~30% residual (Figure 3L), suggesting a near-total loss of functional complex I, which is not reflected in the BN-PAGE. Additionally, complex I activity has not been normalized to a mitochondrial reference, such as citrate synthase.
Given that we work in primary cells and are limited by the number of cells we can generate, we concentrated on ETC1 and 5 and performed experiments in cells after expression of MIRO1 WT and MIRO1 mutants (Figure 6- Figure supplement 1). Please note that the addition of Rotenone abolishes the slope of NADH consumptions (Figure 6- Figure supplement 2F).
While the ETC1 activity is measured in Fig. 6K, the blue native gel shown in Figure 6I is performed without substrate and thus, indicative of protein complex abundance rather than complex activity.
In additional experiments, we normalized the activity to citrate synthase as requested.
(14) In the methods section, the complex I activity assay is incorrectly described: complex I is a NADH dehydrogenase, so the assay measures NADH oxidation, not NADPH.
We thank the reviewer for his comment and revised the manuscript accordingly.
(15) The authors have not assessed mitochondrial mass, which is a critical omission. Differences in mitochondrial biogenesis or content could underlie several observed phenotypes and should be controlled for.
A qPCR assay was used to assess mitochondrial DNA copy number in WT and Miro1-/- VSMCs. We determined the abundance of COX1 and MT-RNR1 DNA as mitochondrial gene targets and NDUFV DNA as the nuclear reference gene. While the results in Miro1-/- cells were highly variable, no statistically significant reduction of copy numbers was detected (Figure 3- Figure supplement 1B).
(16) Complex IV signal is missing in Figure 6I. Its omission is not acknowledged or explained.
Thank you for this comment. We believe this is due to a technical issue. Complex IV can be challenging to detect consistently, as its visibility is highly dependent on sample preparation conditions. In this specific case, we suspect that the buffer used during the isolation process may have influenced the detection of Complex IV.
(17) Figure 6D does not appear representative of the quantifications shown. C-MYC signal is visibly reduced in the mutant, consistent with the lower levels of interactors such as Sam50 and NDUFA9. Additionally, the SDHA band is aligned at the bottom of the blot box. The list of antibodies used, and their catalog number is missing, or it was not provided to the reviewers. It seems plausible that the authors used a cocktail antibody set (e.g., Abcam ab110412), which includes anti-NDUFA9. This would contradict the claim of reduced complex I and SC levels, as the steady-state levels of NDUFA9 appear unchanged.
We acknowledge that the expression of the myc-MIRO1 mutant is lower compared to myc-MIRO1 WT or myc-MIRO1 KK. Achieving identical expression levels when overexpressing multiple MIRO1 constructs is challenging. We agree that the lower expression of this mutant contributes to a reduced pull-down. Our quantification shows a reduction in association, although it is not statistically significant.
A list of the antibodies was provided in the Methods section.
We would like to clarify that we did not use an antibody cocktail in our experiments.
(18) The title of Figure 6, "Loss of Miro1 leads to dysregulation of ETC activity under growth conditions," is vague. The term "dysregulation" should be replaced with a more specific mechanistic descriptor-what specific regulatory defect is meant?
We thank the reviewer for this suggestion and rephrased the title.
(19) In the results text for Figure 6, the authors state: "These data demonstrate that MIRO1 associates with MIB/MICOS and that this interaction promotes the formation of mitochondrial super complexes and the activity of ETC complex I." This conclusion is speculative and not mechanistically supported by the data presented.
We appreciate the reviewer's feedback. We have revised the text to clarify the relationship between MIRO1, MIB/MICOS, supercomplex formation, and ETC activity. The updated text now states: "These data demonstrate that MIRO1 associates with MIB/MICOS. Additionally, MIRO1 promotes the formation of mitochondrial supercomplexes and enhances the activity of ETC complex I.”
(20) In Figure 7A, it is unclear what the 3x siControl/siMiro1 pairs represent-are these different cell lines or technical replicates of the same line? No loading control is shown. If changes in mitochondrial protein abundance are being evaluated, using COX4 as a loading control is inappropriate. The uneven COX4 signal across samples further complicates interpretation
Please note that we used primary cells, not cell lines. The three siControl/siMiro1 pairs represent independent cell isolations, each transfected with either siControl or. siMIRO1 mRNA. While the possibility of a difference in mitochondrial mass is an interesting question, the primary objective of this experiment is to demonstrate that the technique effectively results in the knockdown of Miro1, which is exclusively localized to mitochondria and not present in the cytosol. As such, we believe that Cox4 serves as a reasonable loading control. Although Miro1 knockdown may lead to a reduction in mitochondrial mass, the focus of this experiment is not to assess mitochondrial mass but to confirm the reduction in Miro1 protein levels on mitochondria. We also performed anti-VDAC immunoblots on the same membranes as alternative loading control (Author response image 1).
Author response image 1.
(21) Figure 7G is difficult to interpret. Why did the authors choose to use a sensor-based method instead of the chemiluminescent assay to measure ATP in these samples?
Both methods were employed to assess ATP levels in human samples. ATP measurements obtained with luminescent assay are provided.
pomodoro timer for linux
Loops
The use of loops and conditionals in social media bots highlights a major ethical challenge regarding scale and context. While a human can manually block a few trolls, a bot using a loop can automatically block hundreds of users in seconds based on a simple conditional like an iphone or android tag. This demonstrates how automation doesn't just make tasks easier but changes the power dynamics of online interaction by allowing a single user to exert massive influence without manual effort.
Or a computer program can repeat an action until a condition is met:
This reminds me of when youtubers post videos of followers doing "day x until y" messages. I never considered the possibility that it was fake until now. If you combine this with the sleep feature and randomize the timeframe of the post, it could look very real. I also wonder if in the near future this could be done with AI to create automated videos.
Water framework directive kaderrichtlijn water
Water Framework Directive (Kaderrichtlijn Water) Since 2000.
EU technical guidelines for monitoring PFAS in drinking water
Commonmark is the attempt to have a standardised markdown version.
eLife Assessment
This manuscript provides useful insights into how the brain can simultaneously represent events and the times when they occurred. The results include a comparison between two different basis functions for temporal selectivity and how these generate different predictions for the dynamics of neural populations. The conclusions are partly incomplete because of questions such as the impact of the linear separability assumption and whether joint encodings of event type and time can be made without it.
Joint Public Review:
Quite obviously, the brain encodes "time", as we are able to tell if something happened before or after something else. How this is done, however, remains essentially not understood. In the context of Working Memory tasks, many experiments have shown that the neural activity during the retention period "encodes" time, besides the stimulus to be remembered; that is, the time elapsed from stimulus presentation can be reliably inferred from the recordings, even if time per se is not important for the task. This implies 'mixed selectivity', in the weak sense of neural activity varying with both stimulus identity and time elapsed (since presentation).
In this paper, the authors investigate the implications of a specific form of such mixed selectivity, that is, conjunctive coding of what (stimulus) and when (time) at the single-neuron level, on the resulting dynamics of the population activity when 'viewed' through linear dimensionality-reduction techniques, essentially Principal Component Analysis (PCA). The theoretical/modeling results presented provide a useful guide to the interpretation of the experimental results; in particular, with respect to what can, or cannot, be rightfully inferred from those experimental results (using PCA-like techniques). The results are essentially theoretical in nature; there are, however, some conclusions that require a more precise justification, in my opinion. More generally, as the authors themselves discuss in the paper, it is not clear how to generalize this coding scheme to more complicated, but behaviorally and cognitively relevant, situations, such as multi-item WM or WM for sequences.
(1) It is unclear to me how the conjunctive code that the authors use (i.e., Equation (3)) is constrained by the theoretical desiderata (i.e., compositionality) they list, or whether it is simply an ansatz, partly motivated by theoretical considerations and experimental observations.
The "what" part: What the authors mean by "relationships" between stimuli is never clearly defined. From their argument (and from Figure 1b), it would seem that what they mean is "angles" between population vectors for all pairs of stimuli. If this is so, then the effect of the passing time can only amount to a uniform rescaling of the components of the population vector (i.e., it must be a similarity transformation; rotations are excluded, if the linear-decoder vectors are to be time-independent); the scaling factor, then, must be a strictly monotonous function of time (increasing or decreasing), if one is to decode time. In other words, the "when" receptive fields must be the same for all neurons.
The "when" part: The condition, \tau_3=\tau_1+\tau_2, does not appear to be used at all. In fact, it is unclear (to me at least) whether the model, as it is formulated, is able to represent time intervals between stimuli.
(2) For the specific case considered, i.e., conjunctive coding, it would seem that one should be able to analytically work out the demixed PCA (see Kobak et al., 2016). More generally, it seems interesting to compare the results of the PCA and the demixed PCA in this specific case, even just using synthetic data.
(3) In the Section "Dimensionality of neural trajectories...", there is some claim about how the dimensionality of the population activity goes up with the observation window T, backed up by numerical results that somehow mimic the results of Cueva et al. (2020) on experimental data. Is this a result that can be formally derived? Related to this point, it would be useful to provide a little more justification for Equation (17). Naively, one would think that the correlation matrix of the temporal component is always full-rank nominally, but that one can get excellent low-rank approximations (depending on T, following your argument).
eLife Assessment
The authors provide a scholarly review of intracranial research into the neural correlates of consciousness (NCCs). To our knowledge, this is the first such review, and it therefore may become a must-read for anyone working in the field of consciousness research. It is not so persuasive that intracranial recordings are better suited to identifying pure NCCs than other methods, which appears a problem instead solved through novel paradigms and better-developed theories - but this no doubt reflects an in-depth, timely, and insightful contribution to the literature.
Reviewer #1 (Public review):
Summary
In this review paper, the authors describe the concept of neural correlates of consciousness (NCC) and explain how noninvasive neuroimaging methods fall short of being able to properly characterise an unconfounded NCC. They argue that intracranial research is a means to address this gap and provide a review of many intracranial neuroimaging studies that have sought to answer questions regarding the neural basis of perceptual consciousness.
Strengths
The authors have provided an in-depth, timely, and scholarly contribution to the study of NCCs. First and foremost, the review surveys a vast array of literature. The authors synthesise findings such that a coherent narrative of what invasive electrophysiology studies have revealed about the neural basis of consciousness can be easily grasped by the reader. The review is also, to the best of my knowledge, the first review to specifically target intracranial approaches to consciousness and to describe their results in a single article. This is a credit to the authors, as it becomes ever harder to apply strict tests to theories of consciousness using methods such as fMRI and M/EEG it is important to have informative resources describing the results of human intracranial research so that theorists will have to constrain their theories further in accordance with such data. As far as the authors were aiming to provide a complete and coherent overview of intracranial approaches to the study of NCCs, I believe they have achieved their aim.
Weaknesses
Overall, I feel positive about this paper. However, there are a couple of aspects to the manuscript that I think could be improved.
(1) Distinguishing NCCs from their prerequisites or consequences
This section in the introduction was particularly confusing to me. Namely, in this section, the authors' aim is to explain how intracranial recordings can help distinguish 'pure' NCCs from their antecedents and consequences. However, the authors almost exclusively describe different tasks (e.g., no-report tasks) that have been used to help solve this problem, rather than elaborating on how intracranial recordings may resolve this issue. The authors claim that no-report designs rely on null findings, and invasive recordings can be more sensitive to smaller effects, which can help in such cases. However, this motivation pertains to the previous sub-section (limits of noninvasive methods), since it is primarily concerned with the lack of temporal and spatial resolution of fMRI and M/EEG. It is not, in and of itself, a means to distinguish NCCs from their confounds.
As such, in its current formulation, I do not find the argument that intracranial recordings are better suited to identifying pure NCCs (i.e. separating them from pre- or post-processing) convincing. To me, this is a problem solved through novel paradigms and better-developed theories. As it stands, the paper justifies my position by highlighting task developments that help to distinguish NCCs from prerequisites and consequences, rather than giving a novel argument as to why intracranial recordings outperform noninvasive methods beyond the reasons they explained in the previous section. Again, this position is justified when, from lines 505-506, the authors describe how none of the reported single-cell studies were able to dissociate NCCs from post-perceptual processing. As such, it seems as if, even with intracranial recording, NCCs and their confounds cannot be disentangled without appropriate tasks.
The section 'Towards Better Behavioural Paradigms' is a clear attempt to address these issues and, as such, I am sure the authors share the same concerns as I am raising. Still, I remain unconvinced that the distinguishing of NCCs from pre-/post- processing is a fair motivation for using intracranial over noninvasive measures.
(2) Drawing misleading conclusions from certain studies
There are passages of the manuscript where the authors draw conclusions from studies that are not necessarily warranted by the studies they cite. For instance:
Lines 265 - 271: "The results of these two studies revealed a complex pattern: on the one hand, HGA in the lateral occipitotemporal cortex and the ventral visual cortex correlated with stimulus strength. On the other hand, it also correlated with another factor that does not appear to play a role in visibility (repetition suppression), and did not correlate with a non-sensory factor that affects visibility reports (prior exposure). These results suggest that activity in occipitotemporal cortex regions reflecting higher-order visual processing may be a precursor to the NCC but not an NCC proper."
It's possible to imagine a theory that would predict HGA could correlate with stimulus strength and repetition suppression, or that it would not correlate with prior exposure (e.g. prior exposure could impact response bias without affecting subjective visibility itself). The authors describe this exact ambiguity in interpretation later in the article (line 664), but in its current form, at least in line 270 (when the study is most extensively discussed), the manuscript heavily implies that HGA is not an NCC proper. This generates a false impression that intracranial recordings have conclusively determined that occipitotemporal HGA is not a pure NCC, which is certainly a premature conclusion.
Line 243: "Altogether, these early human intracranial studies indicate that early-latency visual processing steps, reflected in broadband and low gamma activity, occur irrespective of whether a stimulus is consciously perceived or not. They also identified a candidate NCC: later (>200 ms) activity in the occipitotemporal region responsible for higher-order visual processing."
The authors claim in this section that later (>200ms) activity in occipitotemporal regions may be a candidate for an NCC. However, the Fisch et al. (2009) study they describe in support of this conclusion found that early (~150ms) activity could dissociate conscious and unconscious processing. This would suggest that it is early processing that lays claim to perceptual consciousness. The authors explicitly describe the Fisch et al results as showing evidence for early markers of consciousness (line 240: '...exhibited an early...response following recognized vs unrecognised stimuli.) Yet only a few lines later they use this to support the conclusion that a candidate NCC is 'later (>200ms) activity in the occipitotemporal region' (line 245). As such, I am not sure what conclusion the authors want me to make from these studies.
This problem is repeated in lines 386-387: "Altogether, studies that investigated the cortical correlates of visual consciousness point to a role of neural responses starting ~250 ms after stimulus onset in the non-primary visual cortex and prefrontal cortex."
This seems to be directly in conflict with the Fisch et al results, which show that correlates of consciousness can begin ~100ms earlier than the authors state in this passage.
(3) Justifying single-neuron cortical correlates of consciousness
The purpose of the present manuscript is to highlight why and how intracortical measures of neural activity can help reveal the neural correlates of perceptual consciousness. As such, in the section 'Single-neuron cortical correlates of perceptual consciousness', I think the paper is lacking an argument as to why single-neuron research is useful when searching for the NCC. Most theories of consciousness are based around circuit or system-level analyses (e.g., global ignition, recurrent feedback, prefrontal indexing, etc.) and usually do not make predictions about single cells. Without any elaboration or argument as to why single-cell research is necessary for a science of consciousness, the research described in this section, although excellent and valuable in its own right, seems out of place in the broader discussion of NCCs. A particularly strong interpretation here could be that intracranial recordings mislead researchers into studying single cells simply because it is the finest level of analysis, rather than because it offers helpful insight into the NCCs.
(4) No mention of combined fMRI-EEG research
A minor point, but I was surprised that the authors did not mention any combined fMRI-EEG research when they were discussing the limits of noninvasive recordings. Intracortical recordings are one way to surpass the spatial and temporal resolution limits of M/EEG and fMRI respectively, but studies that combine fMRI and EEG are also an alternative means to solve this problem: by combining the spatial resolution of fMRI with the temporal resolution of EEG, researchers can - in theory - compare when and where certain activity patterns (be they univariate ERPs or multivariate patterns) arise. The authors do cite one paper (Dellert et al., 2021 JNeuro) that used this kind of setup, but they discuss it only with respect to the task and ignore the recording method. The argument for using intracranial recordings is weaker for not mentioning a viable, noninvasive alternative that resolves the same issues.
Reviewer #2 (Public review):
Summary:
In this work, the authors review the study of the neural correlates of consciousness (NCCs). They discuss several of the difficulties that researchers must face when studying NCCs, and argue that several of these difficulties can be alleviated by using intracranial recordings in humans.
They describe what constitutes an NCC, and the difficulties to distinguish between an NCC proper from the prerequisites and consequences of conscious processing.
They also describe the two main types of experimental designs used to study NCCs. These are the contrastive approach (with its report and non-report variants), and the supraliminal approach, each with its own merits and pitfalls.
They discuss the limitations of non-invasive methods, such as fMRI, EEG and MEG, as well as the limitations of the use of invasive recordings in non-human animals.
After setting the stage in this way, the authors provide an extensive review of the knowledge acquired by using invasive recordings in humans. This included population-level measurements in vision and in other sensory modalities, as well as single-neuron level studies. The authors also discuss studies of subcortical NCCs.
The second half of this work discusses the theoretical insights gained through the use of intracranial recordings, as well as their limitations, and a perspective for future work.
Strengths:
This work offers an impressive review, which will serve as a useful reference document, both for newcomers to the study of NCC and for experienced researchers. The inclusion of non-visual and subcortical NCCs is of particular merit, as these have been understudied.
Besides serving as a review, this work includes a perspective, exploring several directions to pursue for the progress of the field.
Weaknesses:
The intention of the authors is to argue how some of the problems faced when studying NCCs are alleviated by the use of intracranial recordings in humans. But in some cases, the link between the problems related to the study of NCCs and the advantages of intracranial recordings over non-invasive methods is not clear.
For example, the authors explain the difficulties in distinguishing between true NCCs from their prerequisites and consequences. This constitutes a difficult conceptual problems that plague all recording techniques. The authors don't provide a convincing explanation of how intracranial recordings offer advantages over EEG or MEG when dealing with these problems.
For example, the authors explain how the use of non-report designs to rule out post-perceptual processing relies on null results, which, according to them, are harder to interpret given the low resolution of non-invasive methods. But the interpretation of null results is actually more complicated in the case of intracranial recordings. As the coverage achieved by the electrodes is sparse, if a null result is attested, it remains possible that a true effect was present in a nearby patch of cortex out of coverage.
The authors argue that the spatial resolution of intracranial recordings is better than that of EEG and MEG. While this is technically true (especially compared to EEG), the true spatial scale of the NCCs is unknown. If NCCs' span is in the mm range, then the additional spatial resolution of intracranial recordings might not be an advantage.
Another factor that should be taken into consideration when assessing the spatial resolution of intracranial recordings is that while the listening zone of individual intracranial contacts is small, coverage is sparse and defined by clinical criteria (something that the authors discuss). In practice, the activity recorded by contacts is usually attributed to anatomically defined ROIs with a scale in the cm range. Given the sparse and uneven (across regions and patients) coverage afforded by intracranial recordings, the advantage of intracranial recordings in terms of spatial resolution is overstated.
Appraisal of whether the authors achieved their aims:
In this work, the authors have gathered an impressive review and have discussed several important problems in the field of study of NCCs, as well as provided a perspective on how the field could move forward.
What is less clear is how the use of intracranial recordings per se holds potential to overcome problems such as the distinction between true NCCs and the prerequisites and consequences of conscious processing.
Discussion of the likely impact of the work on the field:
This work has the potential of becoming a must-read for anyone working in the field of consciousness research.
Reviewer #3 (Public review):
Summary:
This narrative review provides a clear, well-structured, and comprehensive synthesis of intracerebral recording work on the neural correlates of consciousness. It is written in an accessible manner that will be useful to a broad community of researchers, from those new to iEEG to specialists in the field.
Strengths:
The manuscript successfully integrates methodological and theoretical perspectives and offers a balanced overview of current, sometimes contradicting evidence. As such, the manuscript is important as it calls for a concerted and better exploration of NCCs using iEEG in the future.
Weaknesses:
The manuscript extensively discusses the use of "report" as a criterion for identifying conscious perception and its limitations for separating between correlates of consciousness and post-consciousness processes, yet the term is not defined at the outset. The authors should specify what they mean by "report" (e.g., verbal report, nonverbal self-report, or any meta-cognitive indication of experience). Importantly, this definition should be explicitly linked to the theoretical landscape: whether the authors adopt an access-consciousness perspective in which (self) reportability is central, or whether the review also aims to address phenomenal consciousness. Making this conceptual grounding explicit at the beginning will help readers interpret the empirical work surveyed throughout the review.
In addition, the review would benefit from an earlier introduction of the distinction between states and contents of consciousness. This distinction becomes important in the later section on anaesthesia, sleep, and epileptic seizures, where the focus shifts from content-specific NCCs to alterations in global states. Presenting these definitions upfront and briefly explaining how states and contents interact would strengthen the coherence of the manuscript.
Overall, this is an excellent and timely review. With clearer initial theoretical definitions of consciousness, the manuscript will offer an even stronger conceptual framework for interpreting intracerebral studies of consciousness.
The original description of Markdown by John Gruber. Seems the only diff I use is wiki links
Silverbullet note making app has a page on how they deal with Markdown, and where it diverges from the standard.
n
Schrijffoutje
teg aan
Schrijffoutje
Wens
Aanpassen in: De wens is...
samengevat
Samengevat kan worden verwijderd.
ovan
Aanpassen ovan => van
op welke manier met het vraagstuk aan de slag
Aanpassen in: op welke manier de provincie en aannemers met het vraagstuk aan de slag gaan (het werkproces en de onderlinge informatie-uitwisseling).
is
woordje 'is' kan worden delete
on International Women’s Day, the bot automatically finds when any of those companies make an official tweet celebrating International Women’s Day and it quote tweets it with the pay gap at that company:
This is a fascinating idea. I think this is a distillation of the ideal of that fact-checking feature that Meta tried and then abandoned, or the community notes feature on Twitter currently. The ability to add context in real time to an issue which is politically multi-faceted (such as corporations both paying lip service to international women's day while not eradicating the structures of misogyny and patriarchy in their own companies) is something social media ought to be doing, but doesn't. I wonder what factors led to more social media companies not attempting to add or push these features to their platforms. Is it profit? Is it their own political beliefs? Or is it simply not feasible?
Though testosterone levels are different between males and females on average, many external factors can change these levels, such as whether or not a person is raising a child
This statement is a claim of fact which can be back by scientific evidence and research.
Stop Using Phony Science to Justify Transphobia
The text takes the form of an scientific article on a science blog website.
While this is a small overview, the science is clear and conclusive: sex is not binary, transgender people are real. It is time that we acknowledge this. Defining a person’s sex identity using decontextualized “facts” is unscientific and dehumanizing.
The author hopes for the readers to takeaway that transgender people are scientifically real and that sex is more complicated than the binary it is commonly believed to be. The author wants the readers to stop trying to put people into boxes based off of faulty knowledge about sex and gender, and use the research they provided to expand their understanding.
By Simón(e) D Sun
Simon(e) D Sun is the author of this article. She is a transgender artist, scientist, and doctoral candidate at New York University's Neuroscience Institute. They published this article on the Scientific American blog, and write articles about the science behind sex and gender. This text is a perfect example of the writing she does within her field of study and work.
I believe that they wrote this article to inform others about the science behind trans people's existence and allow others to learn about their lived experience as a trans woman.
This article is public and posted on a science blog, so the audience is most likely anyone with or without a background in science, who is interested in the science behind gender.
Contrary to popular belief, scientific research helps us better understand the unique and real transgender experience. Specifically, through three subjects: (1) genetics, (2) neurobiology and (3) endocrinology. So, hold onto your parts, whatever they may be. It’s time for “the talk.”
This section shows that the text is about the validity of transgender people's existence and that science can prove their existence.
Defining a person’s sex identity using decontextualized “facts” is unscientific and dehumanizing.
The purpose of this article is to show the masses that are uniformed on trans struggles that they are indeed a person, and should not be demonized for how they identify.
Stop Using Phony Science to Justify Transphobia
While this article is very opinionated, the author gives many facts and citations that prove that her side tends to be the correct definition, however it is important to remember that this type of article is not doing to true justice of the other side of the argument. The other side may not be truly represented in this opinionated article.
While this is a small overview, the science is clear and conclusive: sex is not binary, transgender people are real.
This is a claim of value.
So, no matter what a pundit, politician or internet troll may say, trans people are an indispensable part of our living reality.
The authors audience in this article is clear as she tries to educate the masses through the misinformation given by people trying to demonize the trans community and weaponize this hate.
The truth is, your biological sex isn’t carved in stone, but a living system with the potential for change.
June 13, 2019
This article was written during pride month which is historically known to incur a lot of hate for the LGBTQ community.
The real world consequences are stacking up: the trans military ban, bathroom bills, and removal of workplace and medical discrimination protections, a 41-51 percent suicide attempt rate and targeted fatal violence . It’s not just internet trolling anymore.
The author's purpose in writing this article is clearly laid out in the opening portion of this document. We can see that her main motivation is to uplift that trans community while educating the masses on the trans struggle due to the everlasting backlash that the trans community gets.
By Simón(e) D Sun
Simon(e) d sun is a trans doctoral candidate in the Tsien Lab at New York University's Neuroscience Institute. She has dedicated life to being a writer that focuses on gender studies.
The irony in all this is that these “protectors of enlightenment” are guilty of the very behavior this phrase derides. Though often dismissed as just a fringe internet movement, they espouse unscientific claims that have infected our politics and culture. Especially alarming is that these “intellectual” assertions are used by nonscientists to claim a scientific basis for the dehumanization of trans people. The real world consequences are stacking up: the trans military ban, bathroom bills, and removal of workplace and medical discrimination protections, a 41-51 percent suicide attempt rate and targeted fatal violence . It’s not just internet trolling anymore.Contrary to popular belief, scientific research helps us better understand the unique and real transgender experience. Specifically, through three subjects: (1) genetics, (2) neurobiology and (3) endocrinology. So, hold onto your parts, whatever they may be. It’s time for “the talk.”
This text is about the use of scientific research to understand the unique and real trans experience using genetics, neurobiology, and endocrinology. Its main message is that the trans experience is far more complicated than many dehumanizing articles make it out to be.
Transgender humans represent the complexity and diversity that are fundamental features of life, evolution and nature itself.
Despite the author claiming that this is a fact, this statement is likely a claim of definition. Fundamental features of life would need to be defined.
The popular belief that your sex arises only from your chromosomal makeup is wrong.
The author brings in a claim of definition
Antiscientific sentiment bombards our politics, or so says the Intellectual Dark Web (IDW).
This shows the context as it was written in a significant moment when transgender rights debuted, which at this time much discussions opposed trans inclusion.
THE BRAIN: WHERE STUFF GETS “MADE UP”
This shows the Genre of the article as it comes to show the Scientific American blog network being a persuasive blog.
worden te worden
(2x worden, zin loopt niet)
door aannemer.
door de aannemer (de toevoegen)
door aannemer
door de aannemer (de toevoegen)
worden te worden ingemeten
worden te worden (zin loopt niet)
“human computers”
I think this concept of a human computer is very thought provoking in a myriad of ways. My own research pertains to how identity is being reshaped along the boundaries of recommendation algorithms, and I feel like this idea of a human computer has a lot of thematic overlap. I know that the first "computers" were people who manually did computation as their job, but to what extent did their status as essentially beings of code shape their lives? Did they (or do they, in the case of modern human computers) see the world differently than I? What do they notice? Does living so in tune with the digital shape their philosophical framework? I feel like the fact that so much of culture and my own time is mediated by retention-based algorithms drastically shapes my ability to imagine another world different from my own. Does the same hold true for human computers?
eLife Assessment
This important study establishes a workflow based on environmental sampling for the discovery of bacteriophages capable of infecting antibiotic-resistant pathogens. The experimental design, analysis, and results demonstrating the effectiveness of the workflow are convincing, although a broader sampling scheme and more careful framing of the data within the current limitations of viral taxonomy could strengthen the work. This study will interest researchers working on bacterial infections, environmental microbiology, and phage-based alternatives for addressing antimicrobial resistance.
Reviewer #1 (Public review):
Summary:
In the manuscript "Pathogen-Phage Geomapping to Overcome Resistance," Do et al. present an impressive demonstration of using geographical sampling and metagenomics to guide sample choice for enrichment in human-associated microbes and the pathogen of interest to increase the chances of success for isolating phages active against highly resistant bacterial strains. The authors document many notable successes (17!) with highly resistant bacterial isolates and share a thoughtfully structured phage discovery effort, potentially opening the door to similar geomapping efforts across the field. While the work is methodologically strong and valuable for the community, there are a few areas where additional clarification and analysis could better align the claims with the data presented.
Strengths:
(1) The manuscript describes a well-executed and transparent example of overcoming a major obstacle in therapeutic virus identification, providing a practical success story that will resonate with researchers in microbiology and medicine.
(2) Many phage researchers have anecdotally experienced a similar phenomenon, that a particular wastewater treatment plant always seems to have the pathogens you need. Quantifying this with metagenomics modernizes and adds evidence to this phenomenon in a way that could help researchers reproduce this success in a methodical way.
(3) The methodology of combining environmental sampling, viral screening, and host-range analysis is clearly articulated and reproducible, offering a valuable blueprint for others in the field.
(4) The data are presented with appropriate analytical rigor, and the results include robust sequencing and metagenomic profiling that deepen understanding of local viral communities.
(5) The 17 successes yielding 35 phages have a lot of phylogenetic novelty beyond what the Tailor labs have typically found with previous methods.
(6) The work highlights a practical and innovative solution to an increasingly important clinical problem, supporting the development of personalized antiviral strategies.
Weaknesses:
(1) The central concept of geomapping as a broadly applicable strategy is wonderfully supported by the 17 successes documented in the paper. While this is actually, of course, a strength, the study does not include a comparative analysis across multiple sites with varying sampling outcomes for different bacterial types, which would be necessary to validate this claim more generally.
(2) Some elements, such as beta diversity comparisons and the metagenomics analysis of viral dark matter, would benefit from additional statistical analysis and clearer context.
(3) Claims about therapeutic cocktails would be better framed as speculative and/or moved to the discussion section.
(4) The manuscript could be strengthened by elaborating on the scope and composition of the phage and bacterial isolate collections, which are important for interpreting the broader significance of the findings.
Reviewer #2 (Public review):
Summary:
The manuscript by Do and colleagues aims to develop a workflow for isolating and identifying bacteriophages with potential applications in phage therapy against antibiotic-resistant pathogens. The workflow integrates geΦmapping as a strategy to identify potential phage sources, ΦHD as a device for phage concentration, and RΦ as a phage library constructed from the initial sampling, resulting in the discovery of 36 new phages. The paper is overall interesting, and the proposed method appears robust and effective.
Strengths:
The methods proposed combined state-of-the-art strategies to solve an ever-increasing problem of antibiotic resistance. The methods are robust, and the controls are appropriate. The integration of environmental sampling, concentration strategies, and downstream genomic characterization is a clear strength and provides a potentially scalable framework for identifying candidate therapeutic phages. The manuscript is clearly written overall, and the results support the main conclusions.
Weaknesses:
While the authors acknowledge several limitations, some aspects require clearer framing or additional clarification. The proposed workflow focuses exclusively on aquatic environments as sources of phages, which may limit the diversity of hosts and phage types recoverable using this approach. Some interpretations, particularly regarding taxonomic classification and sampling saturation, would benefit from more cautious wording given current limitations in viral taxonomy and the observed data.
RRID:AB_2892682
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_3718609
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_955447
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_2753196
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_2107448
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_302459
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_2756528
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_2894870
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_10597232
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_2289842
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_3083804
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_2210545
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_2801561
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_955417
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_2714032
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_2076150
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_10644283
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
RRID:AB_3678465
Traceback (most recent call last): File "/home/ubuntu/dashboard/py/create_release_tables.py", line 54, in format_anno_for_release parsedanno = HypothesisAnnotation(anno) File "/home/ubuntu/dashboard/py/hypothesis.py", line 192, in init t = row['document']['title'] TypeError: string indices must be integers
eLife Assessment
This important work shows that a history of cocaine self-administration disrupts the orbitofrontal cortex's ability to encode similarities between distinct sensory stimuli that possess identical task information - hidden states. The evidence supporting these conclusions is compelling, with methods and analyses spanning self-administration, a novel 'figure 8' sequential odor task, recordings from 3,881 single units, and sophisticated firing analyses revealing complex orbitofrontal representations of task structure. These results will be of broad interest to psychologists, neuroscientists, and clinicians.
Reviewer #1 (Public review):
Summary:
In this study, the authors trained rats on a "figure 8" go/no-go odor discrimination task. Six odor cues (3 rewarded and 3 non-rewarded) were presented in a fixed temporal order and arranged into two alternating sequences that partially overlap (Sequence #1: 5⁺-0⁻-1⁻-2⁺; Sequence #2: 3⁺-0⁻-1⁻-4⁺) --forming an abstract figure-8 structure of looping odor cues.
This task is particularly well-suited for probing representations of hidden states, defined here as the animal's position within the task structure beyond superficial sensory features. Although the task can be solved without explicit sequence tracking, it affords the opportunity to generalize across functionally equivalent trials (or "positions") in different sequences, allowing the authors to examine how OFC representations collapse across latent task structure.
Rats were first trained to criterion on the task and then underwent 15 days of self-administration of either intravenous cocaine (3 h/day) or sucrose. Following self-administration, electrodes were implanted in lateral OFC, and single-unit activity was recorded while rats performed the figure-8 task.
Across a series of complementary analyses, the authors report several notable findings. In control animals, lOFC neurons exhibit representational compression across corresponding positions in the two sequences. This compression is observed not only in trial/positions involving overlapping odor (e.g., Position 3 = odor 1 in sequence 1 vs sequence 2), but also in trials/positions involving distinct, sequence-specific odors (e.g., Position 4: odor 2 vs odor 4) --indicating generalization across functionally equivalent task states. Ensemble decoding confirms that sequence identity is weakly decodable at these positions, consistent with the idea that OFC representations collapse incidental differences in sensory information into a common latent or hidden state representation. In contrast, cocaine-experienced rats show persistently stronger differentiation between sequences, including at overlapping odor positions.
Strengths:
Elegant behavioral design that affords the detection of hidden-state representations.
Sophisticated and complementary analytical approaches (single-unit activity, population decoding, and tensor component analysis).
Weaknesses:
The number of subjects is small --can't fully rule out idiosyncratic, animal-specific effects.
Comments
(1) Emergence of sequence-dependent OFC representations across learning.
A conceptual point that would benefit from further discussion concerns the emergence of sequence-dependent OFC activity at overlapping positions (e.g., position P3, odor 1). This implies knowledge of the broader task structure. Such representations are presumably absent early in learning, before rats have learned the sequence structure. While recordings were conducted only after rats were well trained, it would be informative if the authors could comment on how they envision these representations developing over learning. For example, does sequence differentiation initially emerge as animals learn the overall task structure, followed by progressive compression once animals learn that certain states are functionally equivalent? Clarifying this learning-stage interpretation would strengthen the theoretical framing of the results.
(2) Reference to the 24-odor position task
The reference to the previously published 24-odor position task is not well integrated into the current manuscript. Given that this task has already been published and is not central to the main analyses presented here, the authors may wish to a) better motivate its relevance to the current study or b) consider removing this supplemental figure entirely to maintain focus.
(3) Missing behavioral comparison
Line 117: the authors state that absolute differences between sequences differ between cocaine and sucrose groups across all three behavioral measures. However, Figure 1 includes only two corresponding comparisons (Fig. 1I-J). Please add the third measure (% correct) to Figure 1, and arrange these panels in an order consistent with Figure 1F-H (% correct, reaction time, poke latency).
(4) Description of the TCA component
Line 220: authors wrote that the first TCA component exhibits low amplitude at positions P1 and P4 and high amplitude at positions P2 and P3. However, Figure 3 appears to show the opposite pattern (higher magnitude at P1 and P4 and lower magnitude at P2 and P3). Please check and clarify this apparent discrepancy. Alternatively, a clearer explanation of how to interpret the temporal dynamics and scaling of this component in the figure would help readers correctly understand the result.
(5) Sucrose control<br /> Sucrose self-administration is a reasonable control for instrumental experience and reward exposure, but it means that this group also acquired an additional task involving the same reinforcer. This experience may itself influence OFC representations and could contribute to the generalization observed in control animals. A brief discussion of this possibility would help contextualize the interpretation of cocaine-related effects.
(6) Acknowledge low N
The number of rats per group is relatively low. Although the effects appear consistent across animals within each group, this sample size does not fully rule out idiosyncratic, animal-specific effects. This limitation should be explicitly acknowledged in the manuscript.
(7) Figure 3E-F: The task positions here are ordered differently (P1, P4, P2, P3) than elsewhere in the paper. Please reorder them to match the rest of the paper.
Reviewer #2 (Public review):
In the current study, the authors use an odor-guided sequence learning task described as a "figure 8" task to probe neuronal differences in latent state encoding within the orbitofrontal cortex after cocaine (n = 3) vs sucrose (n = 3) self-administration. The task uses six unique odors which are divided into two sequences that run in series. For both sequences, the 2nd and 3rd odors are the same and predict reward is not available at the reward port. The 1st and 4th odors are unique, and are followed by reward. Animals are well-trained before undergoing electrode implant and catheterization, and then retrained for two weeks prior to recording. The hypothesis under test is that cocaine-experienced animals will be less able to use the latent task structure to perform the task, and instead encode information about each unique sequence that is largely irrelevant. Behaviorally, both cocaine and sucrose-experienced rats show high levels of accuracy on task, with some group differences noted. When comparing reaction times and poke latencies between sequences, more variability was observed in the cocaine-treated group, implying animals treated these sequences somewhat differently. Analyses done at the single unit and ensemble level suggests that cocaine self-administration had increased the encoding of sequence-specific information, but decreased generalization across sequences. For example, the ability to decode odor position and sequence from neuronal firing in cocaine-treated animals was greater than controls. This pattern resembles that observed within the OFC of animals that had fewer training sessions. The authors then conducted tensor component analysis (TCA) to enable a more "hypothesis agnostic" evaluation of their data.
Overall, the paper is well written and the authors do a good job of explaining quite complicated analyses so that the reader can follow their reasoning. I have the following comments.
While well-written, the introduction mainly summarises the experimental design and results, rather than providing a summary of relevant literature that informed the experimental design. More details regarding the published effects of cocaine self-administration on OFC firing, and on tests of behavioral flexibility across species, would ground the paper more thoroughly in the literature and explain the need for the current experiment.
For Fig 1F, it is hard to see the magnitude of the group difference with the graph showing 0-100%- can the y axis be adjusted to make this difference more obvious? It looks like the cocaine-treated animals were more accurate at P3- is that right?<br /> The concluding section is quite brief. The authors suggest that the failure to generalize across sequences observed in the current study could explain why people who are addicted to cocaine do not use information learned e.g. in classrooms or treatment programs to curtail their drug use. They do not acknowledge the limitations of their study e.g. use of male rats exclusively, or discuss alternative explanations of their data.
Is it a problem that neuronal encoding of the "positions" i.e. the specific odors was at or near chance throughout in controls? Could they be using a simpler strategy based on the fact that two successive trials are rewarded, then two successive trials are not rewarded, such that the odors are irrelevant?
When looking at the RT and poke latency graphs, it seems the cocaine-experienced rats were faster to respond to rewarded odors, and also faster to poke after P3. Does this mean they were more motivated by the reward?
eLife Assessment
This important study provides the first direct neuroimaging evidence for the integration-segregation theory of exogenous attention underlying inhibition of return, using an optimized IOR-Stroop fMRI paradigm to dissociate integration and segregation processes and to demonstrate that attentional orienting modulates semantic- and response-level conflict processing. Although the empirical evidence is compelling, clearer justification of the experimental logic, more cautious framing of behavioral and regional interpretations, and greater transparency in reporting and presentation are needed to strengthen the conclusions. The work will be of broad interest to researchers investigating visual attention, perception, cognitive control, and conflict processing.
Reviewer #1 (Public review):
Summary:
This study makes a significant and timely contribution to the field of attention research. By providing the first direct neuroimaging evidence for the integration-segregation theory of exogenous attention, it fills a critical gap in our understanding of the neural mechanisms underlying inhibition of return (IOR). The authors employ a carefully optimized cue-target paradigm combined with fMRI to elegantly dissociate the neural substrates of cue-target integration from those of segregation, thereby offering compelling support for the integration-segregation account. Beyond validating a key theoretical hypothesis, the study also uncovers an interaction between spatial orienting and cognitive conflict processing, suggesting that exogenous attention modulates conflict processing at both semantic and response levels. This finding shed new light on the neural mechanisms that connect exogenous attentional orienting with cognitive control.
Strengths:
The experimental design is rigorous, the analyses are thorough, and the interpretation is well grounded in the literature. The manuscript is clearly written, logically structured, and addresses a theoretically important question. Overall, this is an excellent, high-impact study that advances both theoretical and neural models of attention.
Weaknesses:
While this study addresses an important theoretical question and presents compelling neuroimaging findings, a few additional details would help improve clarity and interpretation. Specifically, more information could be provided regarding the experimental conditions (SI and RI), the justification for the criteria used for excluding behavioral trials, and how the null condition was incorporated into the analyses. In addition, given the non-significant interaction effect in the behavioral results, the claim that the behavioral data "clearly isolated" distinct semantic and response conflict effects should be phrased more cautiously.
Reviewer #2 (Public review):
Summary:
This study provides evidence for the integration-segregation theory of an attentional effect, widely cited as inhibition of return (IOR), from a neuroimaging perspective, and explores neural interactions between IOR and cognitive conflict, showing that conflict processing is potentially modulated by attentional orienting.
Strengths:
The integration-segregation theory was examined in a sophisticated experimental task that also accounted for cognitive conflict processing, which is phenomenologically related to IOR but "non-spatial" by nature. This study was carefully designed and executed. The behavioral and neuroimaging data were carefully analyzed and largely well presented.
Weaknesses:
The rationale for the experimental design was not clearly explained in the manuscript; more specifically, why the current ER-fMRI study would disentangle integration and segregation processes was not explained. The introduction of "cognitive conflict" into the present study was not well reasoned for a non-expert reader to follow.
The presentation of the results can be further improved, especially the neuroimaging results. For instance, Figure 4 is challenging to interpret. If "deactivation" (or a reduction in activation) is regarded as a neural signature of IOR, this should be clearly stated in the manuscript.
Reviewer #3 (Public review):
Summary:
This study aims to provide the first direct neuroimaging evidence relevant to the integration-segregation theory of exogenous attention - a framework that has shaped behavioral research for more than two decades but has lacked clear neural validation. By combining an inhibition-of-return (IOR) paradigm with a modified Stroop task in an optimized event-related fMRI design, the authors examine how attentional integration and segregation processes are implemented at the neural level and how these processes interact with semantic and response conflicts. The central goal is to map the distinct neural substrates associated with integration and segregation and to clarify how IOR influences conflict processing in the brain.
Strengths:
The study is well-motivated, addressing a theoretically important gap in the attention literature by directly testing a long-standing behavioral framework with neuroimaging methods. The experimental approach is creative: integrating IOR with a Stroop manipulation expands the theoretical relevance of the paradigm, and the use of a genetic-algorithm-optimized fMRI design ensures high efficiency. Methodologically, the study is sound, with rigorous preprocessing, appropriate modeling, and analyses that converge across multiple contrasts. The results are theoretically coherent, demonstrating plausible dissociations between integration-related activity in the fronto-parietal attention network (FEF, IPS, TPJ, dACC) and segregation-related activity in medial temporal regions (PHG, STG). The findings advance the field by supplying much-needed neural evidence for the integration-segregation framework and by clarifying how IOR modulates conflict processing.
Weaknesses:
Some interpretive aspects would benefit from clarification, particularly regarding the dual roles ascribed to dACC activation and the circumstances under which PHG and STG are treated as a single versus separate functional clusters. Reporting conventions are occasionally inconsistent (e.g., statistical formatting, abbreviation definitions), which may hinder readability. More detailed reporting of sample characteristics, exclusion criteria, and data-quality metrics-especially regarding the global-variance threshold-would improve transparency and reproducibility. Finally, some limitations of the study, including potential constraints on generalization, are not explicitly acknowledged and should be articulated to provide a more balanced interpretation.
eLife Assessment
This important work contributes a transcriptional dataset that identifies potential genes involved in axon initial growth and axon regrowth, followed by a characterization of axon phenotypes after knockdown of a subset of these genes. Focused experiments on a single gene, Pmvk, highlight the potential role of the mevalonate pathway in axon regrowth. The methods are convincing, though partially incomplete. The data establish a basis for further studies on axonal development and will be of interest to both developmental neurobiologists and those seeking to develop molecular tools to target, monitor, and manipulate axon morphology and function.
Reviewer #1 (Public review):
Summary:
Fahdan et al. present a study investigating the molecular programs underlying axon initial growth and regrowth in Drosophila mushroom body (MB) neurons. The authors leverage the fact that different Kenyon cell (KC) subtypes undergo distinct axonal events on the same developmental timeline: γ KCs prune and then regrow their axons during early pupation, whereas α/β KCs extend their axons for the first time during the same pupal period. Using bulk Smart-seq2 RNA sequencing across six developmental time points, the authors identify genes enriched during γ KC regrowth and α/β KC initial outgrowth, and subsequently perform an RNAi screen to determine which candidates are functionally required for these processes.
Among these, they focus on Pmvk, a key enzyme in the mevalonate pathway. Both RNAi knockdown and a CRISPR-generated mutant produce strong γ KC regrowth defects. Knockdown of other mevalonate pathway components (Hmgcr, Mvk) partially recapitulates this phenotype. The authors propose that Pmvk promotes axonal regrowth through effects on the TOR pathway.
Overall, this work identifies new molecular players in developmental axon remodeling and provides intriguing evidence connecting Pmvk to γ KC regrowth.
While the Pmvk knockdown and loss-of-function data are compelling, the evidence that the mevalonate pathway broadly regulates γ KC axon regrowth is less clear. RNAi knockdown of enzymes upstream of Pmvk (Hmgcr, Mvk) produces only mild phenotypes, and knockdown of several downstream enzymes produces no phenotype. The authors attribute this discrepancy to the possibility of weak RNAi constructs, which is plausible but not fully demonstrated. It would be helpful for the authors to discuss alternative explanations, including non-canonical roles for Pmvk that may not require the full pathway, and clarify the extent to which the current data support the conclusion that the mevalonate pathway, rather than Pmvk specifically, is a core regulator of regrowth.
It is not clear from the Methods whether γ KCs and α/β KCs were sorted from the same brains using orthogonal binary expression systems (e.g., Gal4 > reporter 1 and LexA > reporter 2), or isolated separately from different fly lines. If the latter, differences in genetic background, staging, or batch effects could influence transcriptional comparisons. This should be explicitly clarified in the Methods, and any associated limitations discussed in the manuscript.
The authors have made important findings that contribute to our understanding of axon growth and regrowth. As written, some major claims are only partially supported, but these issues can be addressed through reframing and clarification. In particular, the manuscript would benefit from (1) a more cautious interpretation of the mevalonate pathway's role, potentially considering Pmvk non-canonical functions, and (2) addressing methodological ambiguities in the transcriptomic analysis.
Reviewer #2 (Public review):
Fahdan et al. set out to build upon their previous work outlining the genes involved in axon growth, targeting two axon growth states: initial growth and regrowth. They outline a debate in the field that axon regrowth (For instance, after injury or in the peripheral nervous system) is different from initial axon growth, for which the authors have previously demonstrated distinct mechanisms. The authors set out to directly compare the transcriptomes of initial axon growth and regrowth, specifically within the same neuronal environment and developmental time point. To this end, the authors used the well-characterized genetic tools available in Drosophila melanogaster (the fruit fly) to build a valuable dataset of genes involved at different time points in axon growth (alpha/beta Mushroom Body Kenyon cells) and regrowth (gamma Mushroom Body Kenyon cells). The authors then focus on genes that are upregulated during both initial axon growth and axon regrowth. Then, using this subset of genes, they screen for axonal growth and regrowth deficits by knocking down 300 of these genes. 12 genes are found to be phenotypically involved in both axon growth and regrowth based on RNAi gene-targeted knockdown in the Mushroom Body. Of these 12 genes, the authors focus on one gene, Pmvk, which is part of the mevalonate pathway. They then highlight other genes in this pathway. But these genes primarily affect axon regrowth, not initial axon growth, implicating metabolic pathways in axon regrowth. This comprehensive RNA-seq dataset will be a valuable resource for the field of axon growth and regrowth, as well as for other researchers studying the Mushroom Body.
Strengths:
This paper contains many strengths, including the in-depth sequencing of overlapping developmental time points during the alpha/beta KCs' initial axon growth and gamma KCs' regrowth. This produces a rich dataset of differentially expressed genes across different time points in either cell population during development. In addition, the authors characterized expression patterns at developmental time points for 30 Gal4 lines previously identified as alpha/beta KC-expressing. This is very helpful for Drosophila
Mushroom Body researchers because the authors not only characterized alpha/beta expression but also alpha'/beta' expression, gamma expression, and non-MB expression. The authors comprehensively walked through identifying differentially expressed genes during alpha/beta axon growth, identifying a subset of overlapping upregulated genes between cell types, then systematically characterized whether knockdown of a subset of these genes produced an axonal growth defect, and finally selected 1 of 3 cell-autonomous genes important for gamma KCs regrowth to further study.
The authors utilized the developing Mushroom Body in Drosophila melanogaster, which happens to have new neurons developing axons and neurons that have undergone pruning and are regrowing neurons at the same developmental time. They are also in the same part of the brain (the Mushroom Body) and, in theory, since the authors implicate a metabolic pathway, they will have similar metabolic growth conditions.
Identifying Pmvk and two other components of the mevalonate pathway in axon regrowth opens up novel avenues for future studies on the role this metabolic pathway may have in axon growth. The authors of this paper are also very upfront about their negative results, allowing researchers to avoid running redundant experiments and truly build on this work.
Weaknesses:
While the dataset produced in this study is a strength, certain aspects make it more challenging to interpret. For instance, the authors state that roughly equal numbers of males and females are used for sequencing, and this vagueness, coupled with only taking a subset of the GFP-labeled neurons during FACs sorting, can introduce confounds into the dataset. This may hold true in imaging studies as well, in which males and females were used interchangeably.
Additionally, a rationale is needed to explain why random numbers of 1-7 were assigned to zero-expressing genes in the DESeq analysis. This does not seem to conform to the usual way this analysis is normally performed. This can alter how genes across the dataset are normalized and requires further explanation.
The display and discussion of the data set do not always align with the authors' stated goal of having a comprehensive description of the genes that dynamically change during axon<br /> growth and regrowth. Displaying more information about genes differentially expressed in the alpha/beta KCs, or any information about the genes diƯerentially expressed in the gamma KCs when using the same criteria as the alpha/beta KCs, or the 676 overlapping upregulated genes, would significantly add to this paper. The authors previously performed a similar study across developmental time points for gamma KCs, and it is not clear whether any overlapping genes were identified. Also, more information on the genes consisting of PC1 and PC3 when showing the PCA analysis would be helpful. Within the text, there is a discussion of why certain genes or gene groups were omitted or selected, such as clusters 1 and 2, and then some of their subgroups based on expected genes. There is also some discussion of omitted gene groups, but this is not complete across the different clusters, nor is there a discussion of why PC2 was not selected or of which genes might exhibit greater variability than cell type. The authors would make a stronger case for the genes they pursued if they showed that groups of genes already known to be involved in axon growth clustered within the selected groups. Since we do not see the gene lists, this is unclear and adds to the sometimes arbitrary nature of the author's choices about what to pursue in this paper. A larger set of descriptors, such as gene lists and Gene Ontology analysis beyond what is shown, would be very helpful in putting the results in context and determining whether this is a resource beneficial to others.
While the Pmvk story is interesting, the authors appear to make some arbitrary decisions in what is shown or pursued in this paper. Visually, CadN and Twr appear to be more severe axon regrowth phenotypes, where the peduncle appears intact, and axons are not regrowing in Figures 3 N and O. In contrast, Pmvk visually appears to lose neurons in Figure 3 M. With a change of the Gal4 driver (Figure 4), Pmvk now produces a gamma axon regrowth phenotype similar to CadN and Twr in Figure 3. This diƯerence in the use of Gal4 for characterizing axonal phenotypes is not discussed, making some interpretations more challenging due to diƯerences in Gal4 expression strength. For instance, the sequencing work was done with a diƯerent Gal4 MB expressing line than the characterization of gene knockdowns. Further characterization of the Pmvk was performed in the same Gal4 lines as the sequencing (Figure 4), suggesting a potential diƯerence in Gal4 strength that may play a role in their rescue experiments if they are using a slightly weaker Gal4 for gamma lobe expression. A broader discussion of this may make the selection of Pmvk less arbitrary if the phenotype is similar to those of CadN and Twr. Along the lines of the sometimes arbitrary nature of the genes chosen to pursue further, the authors state that they selected genes that showed differential expression at any time point. As they refine their list of genes to pursue further, they seem to prioritize genes that change at 18-21 APF. This appears to be the early period for axon growth in alpha/beta KCs and gamma KCs, based on Figure 1. A stronger case might be made at longer time points when the axon is growing or regrowing.
The paper would benefit from scaling back the claim that the mevalonate pathway is involved. The authors identified only a subset of genes from the mevalonate pathway, all immediately upstream of Pmvk, with no effect on downstream genes. Along these lines, the paper would benefit from a discussion of non-canonical PmvK signaling.
While the ability to take neurons at the same developmental time and from the same brain region is a strength, they are still 2 different types of neurons. Although gamma neuron axon growth occurs very early in development, it would be interesting to know whether the same genes are involved in their initial growth. A caveat to the author's conclusion is that these are 2 different cell types, and they might use different genetic programs or use overlapping ones at other times. The authors did not show that gamma KCs use these genes in their initial axon growth.
Bots present a similar disconnect between intentions and actions. Bot programs are written by one or more people, potentially all with different intentions, and they are run by others people, or sometimes scheduled by people to be run by computers. This means we can analyze the ethics of the action of the bot, as well as the intentions of the various people involved, though those all might be disconnected.
I think this is very interesting as it makes it harder to put responsibility on a single person or thing. A bot could be good or bad regardless of its original intent. This means that the outcome and human accountability should be thought about separately.
There is no way in which police can maintain dignity in seizing and destroying a donkey on whose flank a political message has been inscribed.
INFO 103 Winter 2026. I think this is absolutely right. It reminds me of the rapid development of AI in recent years. One point that people are extremely worried about is that if Ai really becomes so powerful that even humans cannot control it, and if it invays media platforms like a hacker and then changes the direction of public opinion, this would be a very terrifying thing. But at this point, how should we convict?
Bots present a similar disconnect between intentions and actions. Bot programs are written by one or more people, potentially all with different intentions, and they are run by others people, or sometimes scheduled by people to be run by computers.
This situation is similar to the Ship of Theseus philosophical paradox, which questions identity and responsibility when parts of an object are gradually replaced. A social media post can be created and disseminated by either a human or a bot, and bots themselves are designed by humans with different intentions. When a bot-generated post causes harm or social disruption, responsibility becomes unclear, raising ethical questions about whether accountability lies with the original creators, the operators, or the system itself.
Bots present a similar disconnect between intentions and actions. Bot programs are written by one or more people, potentially all with different intentions, and they are run by others people, or sometimes scheduled by people to be run by computers.
I think this is really interesting, because bots are not independent, they are written by real people, and I found this interesting, because it's incredible to see bots written by different people can mean different things. This helps explain why online content from bots can sometimes seem unpredictable or inconsistent.
Bots might have significant limits on how helpful they are, such as tech support bots you might have had frustrating experiences with on various websites.
This shows how bots may not always be effective, especially in more specific contexts such as tech support. I have had frustrating experiences with bots like these, and it shows the limits of the current programming of the bots.
Antagonistic bots can also be used as a form of political pushback that may be ethically justifiable. For example, the “Gender Pay Gap Bot” bot on Twitter is connected to a database on gender pay gaps for companies in the UK. Then on International Women’s Day, the bot automatically finds when any of those companies make an official tweet celebrating International Women’s Day and it quote tweets it with the pay gap at that company:
It is "confrontational", but it has a social justice purpose - to use automation to counter the "pseudo-equality propaganda" of corporate marketing and bring the real structural problem (the wage gap) to the public. This example shows that some antagonistic bots can instead become tools for monitoring power.
Bots might have significant limits on how helpful they are, such as tech support bots you might have had frustrating experiences with on various websites. 3.2.2. Antagonistic bots:# On the other hand, some bots are made with the intention of harming, countering, or deceiving others.
The "bot" itself is not good or bad, but depends on what it is designed for and how the rules of the platform constrain it. For example, friendly bots (automatic captioning, vaccine progress, red panda images) essentially improve the efficiency of information acquisition and enhance the user experience; antagonistic bots (spam, fake fans, astroturfing), however, can create false public opinion and make people think that "many people support/oppose a certain opinion", which directly affects public judgment
We also would like to point out that there are fake bots as well, that is real people pretending their work is the result of a Bot. For example, TikTok user Curt Skelton posted a video claiming that he was actually an AI-generated / deepfake character:
As someone who's majoring in a creative field, I find it both incredibly interesting and concerning just how advanced AI is getting, and where this rapid innovation will take us in just a few years. It's so jarring to be watching a video on Tiktok or Instagram and fully believe it to be completely real, just to feel the need to dissect the video to see if it's really real. I can't begin to imagine how the job industry will change due to AI, but with innovation there (hopefully) comes opportunity.**
On the other hand, some bots are made with the intention of harming, countering, or deceiving others. For example, people use bots to spam advertisements at people. You can use bots as a way of buying fake followers, or making fake crowds that appear to support a cause (called Astroturfing).
Although bot programs are written by one or more people, the individuals who decide the content and deploy the posts often have clear intentions. While responsibility may partially lie with those who develop the bots, the primary accountability for harmful consequences should rest with those who create and control the content.
3.2.3. Corrupted bots# As a final example, we wanted to tell you about Microsoft Tay a bot that got corrupted. In 2016, Microsft launched a Twitter bot that was intended to learn to speak from other Twitter users and have conversations. Twitter users quickly started tweeting racist comments at Tay, which Tay learned from and started tweeting out within one day. Read more about what went wrong from Vice How to Make a Bot That Isn’t Racist 3.2.4. Registered vs. Unregistered bots# Most social media platforms provide an official way to connect a bot to their platform (called an Application Programming Interface, or API). This lets the social media platform track these registered bots and provide certain capabilities and limits to the bots (like a rate limit on how often the bot can post). But when some people want to get around these limits, they can make bots that don’t use this official API, but instead, open the website or app and then have a program perform clicks and scrolls the way a human might. These are much harder for social media platforms to track, and they normally ban accounts doing this if they are able to figure out that is what is happening. 3.2.5. Fake Bots# We also would like to point out that there are fake bots as well, that is real people pretending their work is the result of a Bot. For example, TikTok user Curt Skelton posted a video claiming that he was actually an AI-generated / deepfake character:
This passage uses three levels to remind us that "robots" themselves do not equate to intelligence or objectivity. Tay's "contamination" illustrates that machine learning-based conversational robots absorb biases from the platform as "language norms"—when training data comes from an environment full of provocation and racism, the system becomes an amplifier of prejudice; the problem is not just a technical failure, but a governance failure of treating a "public platform" as a safe training ground. Next, the "registered vs. unregistered bots" reveal the cat-and-mouse game of platform regulation and countermeasures: API restrictions act as rules and guardrails, while simulated clicks bypassing APIs disguise automation as "human," making it harder for platforms to track, demonstrating that visibility and controllability are themselves forms of power. Finally, the "fake bots" point to another form of deception: humans pretending to be AI to gain traffic, a sense of mystery, or immunity from responsibility—this blurs the line of "authenticity" and reminds us that in the attention economy, technological identity can also be used for performance and marketing.
On the other hand, some bots are made with the intention of harming, countering, or deceiving others. For example, people use bots to spam advertisements at people. You can use bots as a way of buying fake followers, or making fake crowds that appear to support a cause (called Astroturfing). As one example, in 2016, Rian Johnson, who was in the middle of directing Star Wars: The Last Jedi, got bombarded by tweets that all originated in Russia (likely making at least some use of bots). “I’ve gotten a rush of tweets – coordinated tweets. Like, somewhere else on the internet there’s like a group on the internet saying, ‘Okay, everyone tweet Rian Johnson.’ All from Russian accounts, and all begging me not to kill Admiral Hux in this movie.” From: https://www.imdb.com/video/vi3962091545 (start at 7:49) After the Star Wars: Last Jedi was released, there was a significant online backlash. When a researcher looked into it: [Morten] Bay found that 50.9% of people tweeting negatively about “The Last Jedi” were “politically motivated or not even human,” with a number of these users appearing to be Russian trolls. The overall backlash against the film wasn’t even that great, with only 21.9% of tweets analyzed about the movie being negative in the first place. https://www.indiewire.com/2018/10/star-wars-last-jedi-backlash-study-russian-trolls-rian-johnson-1202008645/ Antagonistic bots can also be used as a form of political pushback that may be ethically justifiable. For example, the “Gender Pay Gap Bot” bot on Twitter is connected to a database on gender pay gaps for companies in the UK. Then on International Women’s Day, the bot automatically finds when any of those companies make an official tweet celebrating International Women’s Day and it quote tweets it with the pay gap at that company:
This passage shifts the discussion of "bots" from neutral tools back into the context of power and manipulation: they can not only automate the dissemination of information but also automate the creation of "false impressions of public opinion" (follower boosting, astroturfing) and targeted harassment (the coordinated attack on Rian Johnson). More notably, the research mentions that a large number of negative tweets were "politically motivated or non-human," meaning that the anger, ridicule, and boycotts we see online may not be a natural aggregation of "genuine public opinion," but rather an emotional landscape that is organized, amplified, and fabricated. Finally, the "Gender Pay Gap Bot" provides a counterexample: this "adversarial" automation can be used for public accountability—by forcibly juxtaposing corporate holiday statements with structural data (wage gaps), it forces people to see the reality obscured by public relations language. The key is not whether "bots are good or bad," but who uses them and whose perceptions and interests they are used to shape.
As a final example, we wanted to tell you about Microsoft Tay a bot that got corrupted. In 2016, Microsft launched a Twitter bot that was intended to learn to speak from other Twitter users and have conversations. Twitter users quickly started tweeting racist comments at Tay, which Tay learned from and started tweeting out within one day. Read more about what went wrong from Vice How to Make a Bot That Isn’t Racist
The discussion of bots influencing public opinion raises important ethical questions about power and accountability. Even if a bot spreads accurate information, the scale and speed of automation can still distort public discourse. This suggests that ethical evaluation of bots should consider not only content accuracy but also their impact on human decision-making and democratic processes.
The discussion of bots influencing public opinion raises important ethical questions about power and accountability. Even if a bot spreads accurate information, the scale and speed of automation can still distort public discourse. This suggests that ethical evaluation of bots should consider not only content accuracy but also their impact on human decision-making and democratic processes.
ethically justifiable
To me I find it problematic in practice for there to be a distinction between ethical and non-ethical use of antagonistic bots. Everybody has their own worldview and values. To define some of these values as ethical on social media is to impose them on everyone. Maybe this would be okay if there was a democratic way for this. But there isn't. These bots are made to "get a rise out of people" or stir emotions. Subjecting people to that through automated bots under the guise of ethics I disagree with
Copy to clipboard If you run the code above you will see that the program pauses as it displays the output above. These pauses may come in handy when posting tweets, to make it look like your bot is taking time to type in the text. You will get a chance to try that in the next practice section.
I always wondered how programmers would create these sorts of commands, and it's cool to know that it's done with simple commands like these! I was also not previously aware that to display something on a screen, you have to use the command 'display'. I previously thought that 'print' was the main form to do so.
What bots do you find surprising?
INFO 103 Winter 2026 I think what surprises me the most, or rather makes me worry, are those bots with certain intelligent attributes. They can make teasing and humorous comments in the comment sections of people's posts just like humans. If they have such intelligent attributes, then if they are not properly regulated, Will they mark inappropriate remarks and thus bring a bad direction to public opinion?
I think the bot that quotes tweets with statistics of the pay gap at organizations is really effective. Activism burnout is real, if a human tried to do that they'd get exhausted with seeing so often just how bad it is and exhausted by whatever hate comments I'm sure the bot receives. A bot isn't going to care about any of that.
Juan Ponce de León, the conqueror of Puerto Rico, landed in Florida in 1513 in search of wealth and enslaved laborers.
Where in Florida did he land?
The Spanish relied on Indigenous allies. The Tlaxcala were among the most important Spanish allies in their conquest. This sixteenth-century drawing depicts the Spanish and their Tlaxcalan allies fighting against the Purépecha
I find this image interesting as there are many different things going on. I notice there is a man hanging from the top, as well as a man with his head chopped off in the bottom. I also see a dog fighting as well as horses. I also notice the different weapons and shields used.
A few decades later, at the end of the 15th century, the Portuguese plantation system developed on the island of São Tomé became a model for the plantation system as it was expanded across the Atlantic.
I wonder why this location became the model for plantation systems compared to others? And is it still there?
On many points, archaeological and traditional knowledge sources converge: the dental, archaeological, linguistic, oral, ecological, and genetic evidence illustrates a great deal of diversity, with numerous groups settling and migrating over thousands of years, potentially from many different points of origin.7
One of my favorite parts of history is the wide variety of sources and pieces of evidence to cite from and learn from.
Statement from Federal Reserve Chair Jerome H. Powell - Federal Reserve Board<br /> by [[Jerome H. Powell]] Chairman of the Federal Reserve <br /> accessed on 2026-01-11T22:52:25
edward_9639tu
All spams
Note that sometimes people use “bots” to mean inauthentically run accounts, such as those run by actual humans, but are paid to post things like advertisements or political content. We will not consider those to be bots, since they aren’t run by a computer. Though we might consider these to be run by “human computers” who are following the instructions given to them, such as in a click farm:
This paragraph, I mean, it's very important that people know what bots do, they misconceptualize them, but in this day of ChatGPT, Microsoft Copilot, Google Gemini, DuckAI. These bots though are actually interesting in a sense how that not all accounts called ‘bots’ are truly automated. I think this distinction is important because it changes how we understand online content, whether it’s influenced by algorithms or by campaigns.
This means that media, which includes painting, movies, books, speech, songs, dance, etc., all communicates in some way, and thus are social. And every social thing humans do is done through various mediums. So, for example, a war is enacted through the mediums of speech (e.g., threats, treaties, battle plans), coordinated movements, clothing (uniforms), and, of course, the mediums of weapons and violence.
The definition of bots in this chapter highlights that automation exists on a spectrum rather than as a simple bot vs. human distinction. I found it interesting that many accounts we interact with daily may be partially automated, which challenges the assumption that bots are always deceptive or malicious. This makes me think that ethical concerns should focus more on transparency and intent, not just whether automation is involved.
current
EN: Electric current is the rate at which electric charge flows through a conductor. - Unit: ampere (A). - Current in a circuit is associated with energy transfer; devices transform electrical energy into other forms.
中文:电流是电荷通过导体的流动速率。 - 单位:安培(A)。 - 电路中的电流对应能量的传递,用电器把电能转化为其他形式。
mass
EN: Mass is the amount of matter in an object and a measure of its inertia (resistance to changes in motion). - Unit: kilogram (kg). - In this Topic, mass affects both KE (1/2 mv^2) and GPE (m g Δh) linearly.
中文:质量表示物体所含物质的多少,也是惯性大小的量度(抗拒运动状态改变的能力)。 - 单位:千克(kg)。 - 在本主题中,质量会线性影响动能(1/2 mv^2)和重力势能(m g Δh)。
acceleration due to gravity
EN: Acceleration due to gravity (g) near Earth is about 9.8 m/s^2. - It is the acceleration of a freely falling object (ignoring air resistance). - g is used in gravitational potential energy: GPE = m g Δh.
中文:近地面重力加速度 g ≈ 9.8 m/s^2。 - 忽略空气阻力时,自由落体的加速度约为 g。 - 在重力势能公式中使用:GPE = m g Δh。
change in height
EN: Change in height (Δh) is the difference between final and initial height. - It tells how much gravitational potential energy changes: ΔGPE = m g Δh. - Δh depends on the chosen reference level and can be positive or negative.
中文:高度变化(Δh)是末高度与初高度之差。 - 它决定重力势能的变化:ΔGPE = m g Δh。 - Δh 与所选参考高度有关,可为正也可为负。
velocity
EN: Velocity is speed with direction (a vector). - Unit: metres per second (m/s). - Because KE ∝ v^2, doubling velocity makes kinetic energy 4 times larger (if mass is the same).
中文:速度(velocity)是带方向的速率(矢量)。 - 单位:米每秒(m/s)。 - 因为动能与 v^2 成正比,同质量时速度加倍,动能会变为 4 倍。
kilojoules
EN: A kilojoule (kJ) is 1000 joules. - We often use kJ because the joule is a small unit in everyday situations.
中文:千焦(kJ)= 1000 焦耳(J)。 - 由于焦耳较小,生活和化学/食品能量等常用千焦表示。
joule (J)
EN: The joule (J) is the SI unit of energy. - 1 J = 1 N·m = 1 kg·m^2/s^2. - Rough sense of scale: lifting a 0.10 kg object by 1 m requires about 1 J of gravitational potential energy.
中文:焦耳(J)是能量的国际单位。 - 1 J = 1 N·m = 1 kg·m^2/s^2。 - 直观尺度:把约 0.10 kg 的物体抬高 1 m,大约需要 1 J 的重力势能。
ultraviolet radiation
EN: Ultraviolet (UV) radiation is electromagnetic radiation with wavelengths shorter than visible violet light. - UV photons have relatively high energy and can cause chemical changes (e.g., sunburn).
中文:紫外线(UV)是波长比紫光更短的电磁辐射。 - 紫外光子能量较高,可能引起化学变化(如晒伤)。
efficient
EN: An energy transformation is efficient if a large fraction of the input energy becomes useful output. - Efficiency = (useful output energy) / (input energy) × 100%. - Real systems are never 100% efficient because some energy becomes unwanted thermal/sound energy due to friction and resistance.
中文:能量转化“高效”表示输入能量中有较大比例变成了有用的输出。 - 效率 = 有用输出能量 / 输入能量 × 100%。 - 真实系统不可能 100% 高效,因为摩擦/电阻等会使部分能量转为不想要的热能/声能。
friction
EN: Friction is a force that opposes motion between surfaces (or through air/water). - Friction converts mechanical energy into less useful forms, mainly thermal energy (and sometimes sound). - This is why motion usually slows down without additional energy input.
中文:摩擦力是阻碍相对运动的力(包括表面摩擦和空气/水阻力)。 - 摩擦会把机械能转化为较难利用的热能(有时还有声能)。 - 因此没有持续能量输入时,运动往往会逐渐减慢。
sound energy
EN: Sound energy is energy carried by vibrations (mechanical waves) traveling through matter. - Sound requires a medium (air, water, solids) and cannot travel through a vacuum. - When absorbed, sound energy often transforms into thermal energy.
中文:声能是由振动产生并通过介质传播的机械波所携带的能量。 - 声音需要介质(空气/水/固体),真空中不能传播。 - 被吸收后通常转化为热能。
radiant energy
EN: Radiant energy is energy carried by electromagnetic radiation (waves/photons). - It includes visible light, infrared, and ultraviolet. - When absorbed by matter, radiant energy often transforms into thermal energy.
中文:辐射能是由电磁辐射(电磁波/光子)携带的能量。 - 包括可见光、红外线、紫外线等。 - 被物质吸收后常转化为热能。
infrared
EN: Infrared is electromagnetic radiation with wavelengths longer than visible red light. - It is often experienced as warmth because absorbing infrared increases thermal energy.
中文:红外线是波长比红光更长的电磁辐射。 - 常被感受到“热”,因为吸收红外线会增加物体的热能。
nuclear energy
EN: Nuclear energy is energy stored in atomic nuclei. - It can be released by fission, fusion, or radioactive decay. - Nuclear energy has a very high energy density compared with chemical energy.
中文:核能是储存在原子核中的能量。 - 可通过核裂变、核聚变或放射性衰变释放。 - 相比化学能,核能的能量密度非常高。
magnetic potential energy
EN: Magnetic potential energy is stored due to the position/orientation of magnets in a magnetic field. - If released, this stored energy can transform into kinetic energy as objects move.
中文:磁势能是由磁体在磁场中的位置/方向而储存的能量。 - 释放后可转化为动能,使物体发生运动。
Heat
EN: Heat is thermal energy that is transferred from a warmer object to a cooler one because of a temperature difference. - Heat is energy in transit (not a substance stored inside an object). - Unit: joule (J).
中文:热量是由于温度差从高温物体传递到低温物体的热能。 - 热量强调“传递中的能量”,不是物体内部的一种“物质”。 - 单位:焦耳(J)。
thermal energy
EN: Thermal energy is the total kinetic energy of the particles in matter (microscopic motion). - Generally, higher temperature means greater average particle kinetic energy. - Thermal energy can be transferred as heat when there is a temperature difference.
中文:热能(更准确地说是与温度有关的内能部分)来自物质内部粒子的微观运动动能总和。 - 温度越高,粒子平均动能越大。 - 有温差时,热能会以“热量”的形式传递。
electrical potential energy
EN: Electrical potential energy is stored energy due to separated electric charges (electric potential difference/voltage). - A battery stores electrical potential energy and can transfer it to a circuit. - In a circuit it can transform into other forms (light, thermal, sound, motion).
中文:电势能是由于电荷分离而储存的能量(与电势差/电压有关)。 - 电池储存电势能,并可把能量传递到电路中。 - 在电路中可转化为光、热、声或机械运动等其他形式。
electrical kinetic energy
EN: Electrical kinetic energy refers to the energy associated with moving electric charges (electric current). - When charges move through a device, the energy can transform into light, heat, or motion.
中文:电荷运动(电流)所携带的能量可理解为“电的动能”。 - 电流通过用电器时,能量可以转化为光、热或机械运动等。
chemical potential energy
EN: Chemical potential energy is stored in chemical bonds. - Fuels, food, and batteries store chemical potential energy. - During chemical reactions it can transform into thermal, radiant (light), electrical, and mechanical energy.
中文:化学势能是储存在化学键中的能量。 - 燃料、食物、电池都储存化学势能。 - 化学反应中它可转化为热能、光(辐射)能、电能、机械能等。
gravitational potential energy
EN: Gravitational potential energy (GPE) is energy stored because of height in a gravitational field. - It depends on mass, gravitational field strength (g), and change in height. - Near Earth: GPE = m g Δh (relative to a chosen reference level).
中文:重力势能(GPE)是物体由于处在重力场中的高度而具有的能量。 - 与质量、重力加速度 g,以及高度变化 Δh 有关。 - 近地面常用:GPE = m g Δh(相对于所选参考高度)。
mechanical kinetic energy
EN: Mechanical kinetic energy is the kinetic energy of the motion of objects (macroscopic motion you can observe). - It depends on mass and velocity: KE = 1/2 mv^2. - It is different from thermal energy, which is microscopic particle motion.
中文:机械动能(机械运动的动能)是可观察到的物体整体运动所具有的动能。 - 与质量和速度有关:KE = 1/2 mv^2。 - 它不同于热能;热能来自微观粒子的无规则运动。
energy
EN: Energy is the ability to cause change in a system or do work. - Energy appears in different forms (mechanical, thermal, chemical, electrical, radiant, etc.). - Energy can be transferred between a system and its surroundings, or transformed from one form to another. - The SI unit of energy is the joule (J).
中文:能量是使系统发生变化或对物体做功的能力。 - 能量有多种形式(机械、热、化学、电、辐射等)。 - 能量既可以在系统与环境之间传递,也可以在不同形式之间转化。 - 能量的国际单位是焦耳(J)。
system
EN: A system is the part of the universe you choose to study (the object(s) under observation). - The system boundary is defined by you, depending on the question you are trying to answer. - Energy can enter/leave the system, so tracking the system helps you describe energy transfers.
中文:系统是你选择研究的那一部分宇宙(被观察/被分析的对象或对象集合)。 - 系统边界由研究者根据问题来定义,可大可小。 - 能量可以在系统与环境之间传递,因此先定义系统有助于分析能量变化。
potential energy
EN: Potential energy is stored energy due to position or configuration. - It can be transformed into kinetic energy and other forms. - Examples include gravitational, chemical, electrical, and magnetic potential energy.
中文:势能是由于位置或结构/状态而“储存”的能量。 - 势能可以转化为动能以及其他形式的能量。 - 常见类型:重力势能、化学势能、电势能、磁势能等。
kinetic energy
EN: Kinetic energy is the energy of motion. Anything that is moving has kinetic energy. - For a moving object, kinetic energy depends on mass and velocity. - In many Grade-10 problems: KE = 1/2 mv^2 (always ≥ 0).
中文:动能是由于运动而具有的能量,任何在运动的物体都有动能。 - 动能与质量和速度有关。 - 常用公式:KE = 1/2 mv^2(动能一定是非负的)。
law of conservation of energy
EN: The law of conservation of energy states that energy cannot be created or destroyed. - Total energy stays constant in an isolated system. - What changes is the form of energy and where it is (system vs. surroundings). - Some transformed energy becomes less useful (often as thermal energy and sound) due to friction and other processes.
中文:能量守恒定律:能量不会凭空产生,也不会凭空消失。 - 对于孤立系统,总能量保持不变。 - 改变的是能量的形式以及能量所在的位置(系统或环境)。 - 由于摩擦等原因,部分能量会转化为较难利用的热能/声能,但总量仍守恒。
surroundings
EN: The surroundings are everything outside the system boundary. - Surroundings can interact with the system by transferring energy (and sometimes matter). - In many problems, you only include the nearby surroundings that actually interact.
中文:环境(周围环境)是系统边界之外的所有事物。 - 环境可以通过能量(有时也包括物质)与系统发生相互作用。 - 解题时通常只考虑与系统有实际相互作用的那部分环境。
结算类型+日期+资方
条件不全,完整条件见SettlementDocumentKey,或者至少需要再加上收付款方和 sourceType,注意加上等就行
对应一条结算指令
对应两条结算指令
系统定位
图不对 1. 上游业务准确应该是个人理财、机构资金、支付系统 2. 结算模块有调账务模块生成结算账务的情况 3. 业务数据接入层少了和机构资金的对账 图可以参考:https://fintopia.feishu.cn/wiki/Y5b9wEpCqiF75PkUi2OcLyAvnGg 里的,不过已经不是最新的了
Как и на клиенте, трафик, поступивший на сервер, обрабатывается модулем маршрутизации (routing) в соответствии с заданными правилами и перенаправляется на разные исходящие подключения (outbounds):
тоже самое делает и сервер. по определённым правилам маршрутизирует и отправляет в outbound
входящее подключение (inbounds) сервера Xray
попадает в inbounds сервера за пределами великого Китайского фаервола
Приложение должно самостоятельно или с помощью стороннего инструмента перенаправить трафик на входящее подключение
Пользователь, который хочет подключиться должен знать адресс inBounds клинта xray на сервере.
inBounds - это, по сути, вход для любого трафика. условно ворота в дом.
или как идеальная школа, в неё заходишь, и тебе открываются пути а пути))
At the same time, that document didn’t change anything about Python’s runtime behavior, nor did it prevent other uses of annotations.
what a smart move!
"[prolong-ing] the past into the present"
I would argue that this statement has a negative connotation when it pertains to memory, and I disagree with that sentiment. The past is almost if not more important than the present.