GDP
Macrotrends. (n.d.). South Africa GDP (gross domestic product). https://www.macrotrends.net/global-metrics/countries/zaf/south-africa/gdp-gross-domestic-product
GDP
Macrotrends. (n.d.). South Africa GDP (gross domestic product). https://www.macrotrends.net/global-metrics/countries/zaf/south-africa/gdp-gross-domestic-product
demolished
拆除
ballroom
宴会厅
communities
Benneyworth, G. C. (2019). Land, labour, war and displacement: A history of four black concentration camps in the South African War (1899–1902). Historia, 64(2), 1–20. https://doi.org/10.17159/2309-8392/2019/v64n2a1 https://www.scielo.org.za/scielo.php?pid=S0018-229X2019000200001&script=sci_arttext
“Green Apartheid”
Venter, Z. S., Shackleton, C. M., Van Staden, F., Selomane, O., & Masterson, V. A. (2020). Green apartheid: Urban green infrastructure remains unequally distributed across income and race geographies in South Africa. Stockholm Resilience Centre. https://www.stockholmresilience.org/publications/publications/2021-01-07-green-apartheid-urban-green-infrastructure-remains-unequally-distributed-across-income-and-race-geographies-in-south-africa.html
overcrowded and under resourced.
Wood, L. (2015). The environmental impacts of colonialism [Honors thesis, Bridgewater State University]. Virtual Commons – Bridgewater State University. https://vc.bridgew.edu/cgi/viewcontent.cgi?params=%2Fcontext%2Fhonors_proj%2Farticle%2F1131%2F&path_info=Wood.pdf
longterm ecological effects
Wood, L. (2015). The environmental impacts of colonialism [Honors thesis, Bridgewater State University]. Virtual Commons – Bridgewater State University. https://vc.bridgew.edu/cgi/viewcontent.cgi?params=%2Fcontext%2Fhonors_proj%2Farticle%2F1131%2F&path_info=Wood.pdf
British colonists
Wood, L. (2015). The environmental impacts of colonialism [Honors thesis, Bridgewater State University]. Virtual Commons – Bridgewater State University. https://vc.bridgew.edu/cgi/viewcontent.cgi?params=%2Fcontext%2Fhonors_proj%2Farticle%2F1131%2F&path_info=Wood.pdf
“Scorched Earth Tactics”
BBC. The Boer Wars. BBC History. https://www.bbc.co.uk/history/british/victorians/boer_wars_01.shtml
“Total war shattered the rural economy, resulting in starvation and a humanitarian crisis
Benneyworth, G. C. (2019). Land, labour, war and displacement: A history of four black concentration camps in the South African War (1899–1902). Historia, 64(2), 1–20. https://doi.org/10.17159/2309-8392/2019/v64n2a1 https://www.scielo.org.za/scielo.php?pid=S0018-229X2019000200001&script=sci_arttext
economically prosperous
BBC. The Boer Wars. BBC History. https://www.bbc.co.uk/history/british/victorians/boer_wars_01.shtml
Hereford's study of Hart Prairie
Hereford, Richard. “The Effects of Surface Runoff on Hart Prairie from Arizona Snow Bowl Facilities, Coconino National Forest.” Heal the Peaks Campaign, 2022.
Rodgers and O'Neill's concept of infrastructural violence
Rodgers, Dennis, and Bruce O’Neill. “Infrastructural Violence: Introduction to the Special Issue.” Ethnography 13, no. 4 (December 2012): 401–412.
Adam Dunstan, in "Victims of Adaptation,"
Dunstan, Adam. “Victims of ‘Adaptation’: Climate Change, Sacred Mountains, and Perverse Resilience.” Journal of Political Ecology 26, no. 1 (2019): 704–719.
In Navajo Nation v. U.S. Forest Service (2008)
Navajo Nation v. United States Forest Service, 535 F.3d 1058 (9th Cir. 2008) (en banc).
The Hopi Tribe's 2016 official statement
Navajo Nation Office of the President and Vice President, and Navajo Nation Office of the Speaker. “Joint Press Release on Hopi Settlement Agreement with the City of Flagstaff.” March 8, 2016.
The Forest Serive documents forty-one formal meetings with tribal representatives and over two hundred phone calls.
U.S. Department of Agriculture, Forest Service. “Final Environmental Impact Statement for Arizona Snowbowl Facilities Improvements, Volume 2: Response to Comments on the Draft Environmental Impact Statement.” 2005.
The Arizona Snowbowl applied to the U.S. Forest Service for permission to create snow from Class A+ reclaimed wastewater. Class A+ is waster water that has been treated so it is safe to handle but not drink.
U.S. Department of Agriculture, Forest Service. “Final Environmental Impact Statement for Arizona Snowbowl Facilities Improvements, Volume 1.” 2005.
1978 American Indian Religious Freedom Act
American Indian Religious Freedom Act of 1978. Pub. L. No. 95-341, 92 Stat. 469. Codified at 42 U.S.C. § 1996.
Wilson v. Block, 708 F.2d 735.
Wilson v. Block, 708 F.2d 735 (D.C. Cir. 1983).
Hamilton documents that this conflict did not begin with snowmaking.
Hamilton, Lawrence. “Sacred vs Profane: Conflict over the San Francisco Peaks.” Mountain Research and Development 26, no. 4 (November 2006): 366–367.
e Forest Service's own Environmental Impact Statement records extensive testimony about the religious and cultural significance of the Peaks to multiple Native nations
U.S. Department of Agriculture, Forest Service. “Final Environmental Impact Statement for Arizona Snowbowl Facilities Improvements, Volume 2: Response to Comments on the Draft Environmental Impact Statement.” 2005.
They are included in the National Register of Historic Places
Glowacka, Maria, Dorothy Washburn, and Justin Richland. “Nuvatukya’ovi, San Francisco Peaks: Balancing Western Economies with Native American Spiritualities.” Current Anthropology 50, no. 4 (August 2009): 547–561.
As Emory Sekaquaptewa put it, "the close ties between the people and their landscape, both in their ritual as well as in their practices, that give the Hopi people their identity and place in the natural world."
Glowacka, Maria, Dorothy Washburn, and Justin Richland. “Nuvatukya’ovi, San Francisco Peaks: Balancing Western Economies with Native American Spiritualities.” Current Anthropology 50, no. 4 (August 2009): 547–561.
The Peaks are the home of the kachina spirits, and ceremonies that perpetuate Hopi cultural and spiritual life are tied directly to the mountain.
Glowacka, Maria, Dorothy Washburn, and Justin Richland. “Nuvatukya’ovi, San Francisco Peaks: Balancing Western Economies with Native American Spiritualities.” Current Anthropology 50, no. 4 (August 2009): 547–561.
Glowacka, Washburn, and Richland document the Hopi relationship to the Peaks through oral tradition, ceremony, and testimony stretching back to the 13th century
Glowacka, Maria, Dorothy Washburn, and Justin Richland. “Nuvatukya’ovi, San Francisco Peaks: Balancing Western Economies with Native American Spiritualities.” Current Anthropology 50, no. 4 (August 2009): 547–561.
A.R.S. §45-141 and §45-151(A)
Arizona Department of Water Resources. “Surface Water Overview.” Accessed April 13, 2026.
Vanderwarker
Vanderwarker, Amy. “Water and Environmental Justice.” Chap. 3 in A Twenty-First Century U.S. Water Policy, by Juliet Christian-Smith, Peter H. Gleick, Heather Cooley, Lucy Allen, Amy Vanderwarker, and Kate A. Berry. New York: Oxford University Press, 2012.
Nick Estes traces the long history of federal infrastructure project
Estes, Nick. Our History Is the Future: Standing Rock Versus the Dakota Access Pipeline, and the Long Tradition of Indigenous Resistance. New York: Verso, 2019.
Börk and Ziaja, writing in the Georgetown Law Journal, document that race is the strongest indicator of water and sanitation access in the United States
Börk, Karrigan, and Sonya Ziaja. “Amoral Water Markets?” Georgetown Law Journal 111, no. 6 (June 2023): 1335–1405.
.
Chatterjee, Pratap. “The Gold Rush Legacy: Greed, Pollution and Genocide.” Earth Island Journal 13, no. 2 (1998): p. 26
proposed using up to 1.5 million gallons per day of Class A+ reclaimed water from Flagstaff to manufacture snow on 205.3 acres
U.S. Department of Agriculture, Forest Service. “Record of Decision: Arizona Snowbowl Facilities Improvements Final EIS and Forest Plan Amendment #21.” 2005.
National Ski Areas Association frames snowmaking as part of the ski industry's "20+ year commitment to protect the environment." 87% of American ski resorts now use snowmaking. As well, the industry generated $58.8 billion in gross output in 202 and supplied about 1/2 a million jobs.
National Ski Areas Association and Brendle Group. “Climate Smart Snowmaking Fact Sheet.” 2023.
Figure 10:
Note that it looks like there is an r value > 1; there isn't. It is just from the binning and dodged bars for males and females. If we needed to use this figure in a manuscript, I would clean up the axis limits.
a forum to discuss potential paths for publishing the most interesting parts.
Reminder of how to use these comments here!
The plan was to increase snowmaking coverage by 205 acres. The system was designed to use up to 1.5 million gallons of Class A+ wastewater per day to expand the season. To do so, an almost 15-mile pipeline from Flagstaff would be built, cutting down approximately 30,000 trees.
U.S. Department of Agriculture, Forest Service. “Record of Decision: Arizona Snowbowl Facilities Improvements Final EIS and Forest Plan Amendment #21.” (2005): 9-17.
Dookʼoʼoosłííd by the Diné and Nuva'tukya'ovi by the Hopi.
Hamilton, Lawrence. “Sacred vs Profane: Conflict over the San Francisco Peaks.” Mountain Research and Development 26, no. 4 (November 2006): 366–367.
western slope
Arizona Snowbowl. “About the Mountain.” Accessed April 13, 2026.
eserved.
add our instagram link https://www.instagram.com/linean_co/
03What are you trying to do?
remove section
Reply within 1 business day · No sales calls
remove
We reply within one business day with a plan, a price, or a polite referral if we're not the right fit.
remove
What do you need?AI trainingConsultingCustom softwareJust exploring
remove this section
Helping teams modernize work across business, education, agriculture, and public service.
increase the font size 4 sizes bigger
Linean
cut the gap of blank space above this by half
Quando houver deficit atuarial
Considerando que - conforme estipula o § 18 do art. 40 - a contribuição, de aposentados e pensionistas, para o RPPS somente incide sobre o valor que exceder o teto do RGPS, havendo déficit atuarial, poderá incidir contribuição sobre aposentadoria e pensões que superarem o valor do salário-mínimo.
Não confundir com a contribuição extraordinária, visto que essa, enquanto novo tributo, somente terá lugar acaso a medida de ampliação da base de cálculo, consistente em tributar o valor que supere o salário-mínimo, for insuficiente.
other activities
Helping my kids with homework.
It's time to start annotating some documents.
interesting
The Texas GLO's recovery plan was revised thirteen times between 2017 and 2023, each amendment a correction that came years after the harm had already accumulated in the lives of families waiting for help. Throughout all thirteen versions, the word "race" never appears.
Texas General Land Office. State of Texas Action Plan for Disaster Recovery: Hurricane Harvey. Austin: Texas General Land Office, 2017. Revised through Substantial Amendment 13, 2023.
And the Texas General Land Office took charge of distributing $5.676 billion in federal Community Development Block Grant funds through programs covering homeowner assistance, affordable rental housing, and local infrastructure.
Texas General Land Office. State of Texas Action Plan for Disaster Recovery: Hurricane Harvey. Austin: Texas General Land Office, 2017. Revised through Substantial Amendment 13, 2023.
Four years after Harvey, a Houston Chronicle investigation revealed that a small, predominantly white inland town with comparatively modest damage had received far more state recovery funds than majority-minority Houston neighborhoods that had lost far more.
Despart, Zach. "How a Small Texas Town Got More Hurricane Harvey Recovery Money Than All of Houston." Houston Chronicle, 2021.
Research by economists Stephen Billings, Emily Gallagher, and Lowell Ricketts confirmed the pattern at scale.
Billings, Stephen, Emily Gallagher, and Lowell Ricketts. "Let the Rich Be Flooded: The Distribution of Financial Aid and Distress after Hurricane Harvey." Journal of Financial Economics 146, no. 2 (2022): 797–819.
This is precisely how infrastructural violence operates, not through obvious failure or deliberate discrimination, but through the quiet, technical design of systems that distribute harm through their ordinary functioning.
Rodgers, Dennis, and Bruce O'Neill. "Infrastructural Violence: Introduction to the Special Issue." Ethnography 13, no. 4 (2012): 401–412.
when the infrastructural violence of the recovery system ensured that inequality was deepened rather than corrected.
Rodgers, Dennis, and Bruce O'Neill. "Infrastructural Violence: Introduction to the Special Issue." Ethnography 13, no. 4 (2012): 401–412.
Scholars Jayajit Chakraborty, Sara Grineski, and Timothy Collins found that flood extent significantly increased in neighborhoods comprising predominantly Black, Hispanic, and socioeconomically deprived residents.
Chakraborty, Jayajit, Timothy W. Collins, and Sara E. Grineski. "Exploring the Environmental Justice Implications of Hurricane Harvey Flooding in Greater Houston, Texas." American Journal of Public Health 109, no. 2 (2019): 244–250.
que la democracia era un conjunto de pro-cedimientos para procesar los problemas, pero no la panacea para resolverlos.
A manera de conclusión, considero que el texto busca explicar cómo las dictaduras militares, las transiciones democráticas y el neoliberalismo estuvieron muy relacionados en América Latina. También comprendí que los autores critican que la democracia se haya reducido muchas veces sólo al hecho de votar, sin resolver problemas como la pobreza, la desigualdad y la falta de participación social, lo cual limita a una verdadera democracia. En general, el texto me ayudó a entender que los cambios políticos y económicos de la región no ocurrieron por separado, sino como parte de un mismo proceso histórico.
compañada de medidas socialesque atenúen la pobreza y la desigualdad
Una verdadera democracia debe incluir no sólo derechos políticos, sino también justicia social y reducción de la desigualdad.
franca crisis en muchos de los Estados latinoamericanos
La idea de que la riqueza “se derramaría” hacia toda la sociedad no produjo los resultados esperados.
contradicho estas presunciones del proyecto neoliberal
El proyecto neoliberal prometía crecimiento económico y bienestar social a través de la apertura al capital privado, pero las expectativas económicas del neoliberalismo no se cumplieron completamente en América Latina.
En conjunto, estas tres partes forman la esencia del proyecto neoliberal
Este fragmento, muestra que el neoliberalismo no sólo fue un modelo económico, sino también un proyecto político e ideológico, pues lo divide al neoliberalismo en tres dimensiones: económica, política e ideológica. Pienso que el neoliberalismo intentó presentarse como la "única opción" posible para alcanzar el desarrollo económico, pero en realidad, considero que el discurso del desarrollo económico fue utilizado para justificar reformas profundas en la economía y en la política que a veces favorecían a intereses de pocos... me hace reflexionar sobre cómo las ideas y discursos ideológicos también pueden influir mucho en la organización de la sociedad y en cómo viven la democracia y su vivel de vida de los ciudadanos. El neoliberalismo prometía modernización y progreso, aunque sus resultados fueron discutidos y criticados con el paso del tiempo...
saqueo de la economía mediante la privatización y venta de paraestatales,
Cuando dice “saqueo de la economía” muestra una postura crítica del autor hacia las privatizaciones neoliberales.
os individuos no actúan directamente en defensa de sus intereses, sinoque la delegan” a través del voto
En la democracia procedimental el ciudadano tiene una participación muy limitada, reducida principalmente al voto, lo cual, deja poco espacio para la participación social directa en las decisiones importantes.
El rol del ciudadano demo-crático en su idea se concentra únicamente en el derecho periódico a escogery autorizar un gobierno para que actúe en su nombre
Muchas veces se ve a los ciudadanos más como electores que como participantes activos en las decisiones políticas, y que participan en elegir a los representantes, pero no gobierna directamente.
través de pactos políticos, se pudo desplazar a lasdictaduras militares y se posibilitó el reconocimiento de derechos políticos yciviles liberales acordes con el modelo de la democracia.
Demuestra que, muchas transiciones a la democracia en América Latina fueron negociadas entre élites políticas y militares.
fiscales
Este fragmento muestra cómo los gobiernos ofrecieron incentivos económicos para atraer inversión privada y fortalecer el modelo neoliberal.
O’Donnell considera los siguientesrasgos para distinguir al Estados burocrático de otros Estados autoritarios
Considero que el texto intenta demostrar que el neoliberalismo no surgió sólo como un modelo económico, sino también como una forma de reorganizar el poder político y social.
En este contexto, podemos hablar de una completa supresión de la sociedadcivil, mediante la abierta represión no sólo de partidos políticos de oposición eindependientes, sino de cualquier expresión política que contraviniera o cues-tionara al poder dictatorial militar y a la oligarquía dominante.
Este fragmento muestra que durante las dictaduras militares prácticamente desapareció la libertad de participación política y social; no sólo contra partidos políticos, sino también contra cualquier forma de oposición o crítica al gobierno, lo cual en sí, atenta en contra de la libertad de expresión y manifestación.
Las fuerzas armadas asumieran el poder de reestructurar la sociedad yel Estado de acuerdo a la doctrina de seguridad nacional.
Este fragmento muestra que las dictaduras militares buscaban controlar completamente la sociedad y el Estado.
Fidel Castro
Se muestra cómo el triunfo de Fidel Castro generó temor en los grupos conservadores y en Estados Unidos.
subordinación al control político centralizado.
Considero que, las organizaciones corporativas funcionaban como una forma de dar voz a los trabajadores, pero también de mantenerlos subordinados al Estado.
Brasil
Me transmite la idea de que en esa época gran parte de América Latina estaba dominada por gobiernos autoritarios, y que, muchas veces los gobiernos justifican la represión bajo la idea de mantener el orden y combatir movimientos de revelión de masas.
Estado burocrático autoritario
El concepto de “Estado burocrático autoritario” ayuda a entender que estos gobiernos no sólo eran militares, sino también sistemas organizados para controlar la sociedad y la economía.
México, Brasil y Argentina, pre-sentó limitaciones que resultaron tanto endémicas como producto del contex-to económico internacional.
Me parece importante la idea de que México, Brasil y Argentina dependían de tecnología y maquinaria extranjera para sostener su industrialización, por cierto es una crítica actual, que considero que esta dependencia económica limitó el crecimiento de los países latinoamericanos, porque no podían controlar totalmente su producción, tal como lo vemos en nuetro país.
antecedentes el triunfo de laRevolución rusa, en 1917, y la ulterior expansión del llamado campo socialista,la crisis de 1929 y la emergencia de la organización sindical obrera. Lo anteriorpermitió la existencia del llamado capitalismo embridado.1
Este fragmento explica cómo después de la Segunda Guerra Mundial existió un intento de equilibrar la economía para evitar que las clases altas concentraran todo el poder, y me parece interesante que el texto mencione la influencia de la Revolución rusa y de los movimientos obreros, porque demuestra que los cambios económicos también surgieron por presión social y política, como ha sido en cualquier tipo de cambio político.
omo diversosgrupos de poder aprovecharon las transiciones democráticas (muchos de elloscercanos a los Estados burocráticos y las dictaduras militares) para colocarseen el poder y comenzar los procesos de reformas neoliberales en la región.
Este estracto, considero que intenta demostrar que muchos cambios económicos fueron favorecidos por grupos de poder que ya tenían influencia desde los gobiernos autoritarios.
odelo hegemónico que se impuso en toda la zona
Me hizo pensar que el neoliberalismo no apareció de repente, sino que tuvo antecedentes políticos y económicos muy claros desde las dictaduras militares en América Latina.
El Estado burocrático autoritario ylas transiciones a la democracia: lasraíces de los regímenes neoliberales enAmérica Latina
Muy interesante...
This unprecedented unification, often referred to as a “modern-day Treaty Alliance,” saw tribal leaders, elders, youth, military veterans, and non-Native allies camping side by side in harsh North Dakota winters, sharing ceremonial fires, legal strategies, and direct action tactics.
Todrys, Katherine Wiltenburg. Black Snake: Standing Rock, the Dakota Access Pipeline, and Environmental Justice. Lincoln: University of Nebraska Press, 2021.
the project has locked the Navajo Nation into a narrow, colonial-defined framework of water “beneficial use” that does not align with Navajo traditional values, economic priorities, or long-term self-determination.
Bray, Laura A. 2020. “Settler Colonialism and Rural Environmental Injustice: Water Inequality on the Navajo Nation.” Rural Sociology 86, no. 3
landowners and Indigenous nations find their property and sovereignty overridden by claims of “national energy needs.”
Shoemaker, Jessica A. “Invited Essay: Pipelines, Protest, and Property.” Great Plains Research 27, no. 2 (2017): 69–81. http://www.jstor.org/stable/44685770.
Should I Run Plain Docker Compose in Production in 2026?
--remove-orphans flag during up and down commands to ensure containers removed from the YAML file are actually stopped and cleared.daemon.json (e.g., max-size: 10m) to prevent unbounded log files from filling disks.docker image prune).docker volume prune to avoid accidental data loss from detached volumes.docker-autoheal or a dedicated agent to act on "unhealthy" states.:latest. Instead, pin images using their immutable SHA256 digests (image: myapp@sha256:...) to ensure consistency across different host pulls./var/run/docker.sock provides a container with effective root privileges on the host. Minimize its use, consider rootless Docker, or use a socket proxy to limit API exposure.Additionally, the homepage explicitly acknowledges the tribe’s ongoing fight against the DAPL, featuring a call to “Stand With Us” in support of their legal efforts to protect their land and water.
“Homepage - Standing Rock Sioux Tribe.” 2026. Standing Rock Sioux Tribe. March 24, 2026. https://www.standingrock.org/homepage/.
The address centers on the concept of “survivance”—a term coined by Gerald Vizenor that combines survival and resistance—to describe how Indigenous nations do not simply endure ongoing settler-colonial assaults on their lands, waters, and communities, but actively assert their sovereignty through creative, strategic, and culturally grounded opposition.
Boxer, Elise. “American Indian Studies Association Conference Presidential Address. Advocacy and Indigenous Resistance: The Ongoing Assault against Indigenous Sovereignty, Community, and Land.” Wicazo Sa Review 32, no. 2 (2017): 91–105. https://doi.org/10.5749/wicazosareview.32.2.0091.
La contaminación es la introducción de sustancias nocivas o elementos físicos en un medio ambiente, lo que provoca que este sea inseguro o no apto para su uso
me parece util pq puedo conocer mas sobre el concepto
Judge James Boasberg, in a carefully reasoned opinion, determined that the Corps had violated the National Environmental Policy Act (NEPA) by failing to adequately address the “highly controversial” nature of the DAPL's potential for serious environmental harm, specifically regarding the risk of a major oil spill beneath the Missouri River, the sole drinking water source for the Standing Rock Sioux Reservation.
STANDING ROCK SIOUX TRIBE v. UNITED STATES ARMY CORPS OF ENGINEERS, No. 1:2016cv01534 - Document 496 (D.D.C. 2020)
The order required that the pipeline be drained and shut down pending completion of the comprehensive environmental review, effectively halting the transport of hundreds of thousands of barrels of oil per day across the Missouri River.
Kennedy, Merrit. 2017. “Judge Delivers Blow to Trump Administration in Dakota Access Fight.” NPR. June 15, 2017. https://www.npr.org/sections/thetwo-way/2017/06/15/533057862/judge-delivers-blow-to-trump-administration-in-dakota-access-fight.
.
The Department of Climate Change, Energy. “NSW Government Launches Historic Aboriginal Water Strategy.” NSW Government, October 15, 2025. https://www.nsw.gov.au/ministerial-releases/nsw-government-launches-historic-aboriginal-water-strategy.
.
“Australia’s Water Market Is Excluding Indigenous People, Study Finds.” The Guardian, July 23,2020.https://www.theguardian.com/australia-news/2020/jul/24/australias-water-market-is-excluding-in`digenous-people-study-finds
.
Davies, Thom. “Toxic Space and Time: Slow Violence, Necropolitics, and Petrochemical Pollution.” Annals of the American Association of Geographers 108, no. 6 (2018): 1537–1553. https://doi.org/10.1080/24694452.2018.1470924.
. A disaster ends only when recovery becomes possible for everyone.
Rob Nixon, Slow Violence and the Environmentalism of the Poor (Cambridge, MA: Harvard University Press, 2011), 2. Betty Hearn Morrow, “Identifying and Mapping Community Vulnerability,” Disasters 23, no. 1 (1999): 1–18.
some people are denied the resources needed to escape danger’s aftermath.
Betty Hearn Morrow, “Identifying and Mapping Community Vulnerability,” Disasters 23, no. 1 (1999): 1–18.
insurance barriers, price increases, unstable housing, and uneven public response.
Lynne McChristian, Hurricane Andrew and Insurance: The Enduring Impact of an Historic Storm (Insurance Information Institute, 2012), 2, 17–18. Gary E. Lehman, “Price Gouging: Application of Florida’s Deceptive and Unfair Trade Practices Act in the Aftermath of Hurricane Andrew,” Nova Law Review 17, no. 3 (1993): 1029–1034. Elliott Mittler, A Case Study of Florida’s Emergency Management Since Hurricane Andrew, Working Paper 98 (Natural Hazards Center, 1997), 10–11.
the image of Carmen Rivera,
John Luke, Carmen Rivera Looks over Remains of Her House in Homestead, Florida, 31 August 1992, State Archives of Florida, Florida Memory.
Andrew became slow violence not because the hurricane moved slowly, but because recovery did.
Rob Nixon, Slow Violence and the Environmentalism of the Poor (Cambridge, MA: Harvard University Press, 2011), 2.
Weak emergency coordination
Elliott Mittler, A Case Study of Florida’s Emergency Management Since Hurricane Andrew, Working Paper 98 (Natural Hazards Center, 1997), 10–11.
Higher prices for basic materials
Gary E. Lehman, “Price Gouging: Application of Florida’s Deceptive and Unfair Trade Practices Act in the Aftermath of Hurricane Andrew,” Nova Law Review 17, no. 3 (1993): 1029–1034.
waiting for repairs,
Kenneth J. Smith and Lina Liska Belgrave, “The Reconstruction of Everyday Life,” Journal of Contemporary Ethnography 24, no. 3 (1995): 244–269.
waiting for insurance payments,
Lynne McChristian, Hurricane Andrew and Insurance: The Enduring Impact of an Historic Storm (Insurance Information Institute, 2012), 2, 17–18. Catherine Wilson, “Hurricane Leaves Florida’s Insurance Safety Net Torn,” The Washington Post, December 8, 1992.
slow violence.
Rob Nixon, Slow Violence and the Environmentalism of the Poor (Cambridge, MA: Harvard University Press, 2011), 2.
lower-income communities were not only exposed to destruction. They were also exposed to the slow violence of unequal repair.
Rob Nixon, Slow Violence and the Environmentalism of the Poor (Cambridge, MA: Harvard University Press, 2011), 2. Betty Hearn Morrow, “Identifying and Mapping Community Vulnerability,” Disasters 23, no. 1 (1999): 1–18.
Government capacity also shaped recovery
Elliott Mittler, A Case Study of Florida’s Emergency Management Since Hurricane Andrew, Working Paper 98 (Natural Hazards Center, 1997), 10–11.
When prices rise during an emergency,
Gary E. Lehman, “Price Gouging: Application of Florida’s Deceptive and Unfair Trade Practices Act in the Aftermath of Hurricane Andrew,” Nova Law Review 17, no. 3 (1993): 1029–1034.
insurers failed or delayed payment,
Catherine Wilson, “Hurricane Leaves Florida’s Insurance Safety Net Torn,” The Washington Post, December 8, 1992.
Insurance was one of the clearest mechanisms through which recovery became uneven.
Lynne McChristian, Hurricane Andrew and Insurance: The Enduring Impact of an Historic Storm (Insurance Information Institute, 2012), 2, 17–18.
Recovery required access to money, insurance, transportation, contractors, temporary housing, public assistance, and the ability to navigate bureaucratic systems.
Betty Hearn Morrow, “Identifying and Mapping Community Vulnerability,” Disasters 23, no. 1 (1999): 1–18.
recovery depended not only on the severity of wind damage but also on insurance coverage, transportation, public services, contractors, housing options, and government response.
Betty Hearn Morrow, “Identifying and Mapping Community Vulnerability,” Disasters 23, no. 1 (1999): 1–18. Elliott Mittler, A Case Study of Florida’s Emergency Management Since Hurricane Andrew, Working Paper 98 (Natural Hazards Center, 1997), 10–11.
Miami-Dade County.
University of Miami Digital Collections, “Damages to Neighborhood after Hurricane Andrew,” Hurricane Andrew Collection.
Jack Beven, “TPC NHC Hurricane Andrew,” National Hurricane Center / NOAA.
Homestead
John Luke, Carmen Rivera Looks over Remains of Her House in Homestead, Florida, 31 August 1992, State Archives of Florida, Florida Memory.
recovery did not happen automatically once the storm ended.
Elliott Mittler, A Case Study of Florida’s Emergency Management Since Hurricane Andrew, Working Paper 98 (Natural Hazards Center, 1997), 10–11.
military personnel assisting with cleanup
University of Miami Digital Collections, “Military Personnel Assist in Clean-up after Hurricane Andrew,” Hurricane Andrew Collection.
damaged residential street
University of Miami Digital Collections, “Damages to Neighborhood after Hurricane Andrew,” Hurricane Andrew Collection.
Carmen Rivera standing near the remains of her Homestead home
John Luke, Carmen Rivera Looks over Remains of Her House in Homestead, Florida, 31 August 1992, State Archives of Florida, Florida Memory.
delayed rebuilding, insurance barriers, rising recovery costs, and uneven public response.
Lynne McChristian, Hurricane Andrew and Insurance: The Enduring Impact of an Historic Storm (Insurance Information Institute, 2012), 2, 17–18. Gary E. Lehman, “Price Gouging: Application of Florida’s Deceptive and Unfair Trade Practices Act in the Aftermath of Hurricane Andrew,” Nova Law Review 17, no. 3 (1993): 1029–1034. Elliott Mittler, A Case Study of Florida’s Emergency Management Since Hurricane Andrew, Working Paper 98 (Natural Hazards Center, 1997), 10–11.
slow violence.
Rob Nixon, Slow Violence and the Environmentalism of the Poor (Cambridge, MA: Harvard University Press, 2011), 2.
Hurricane Andrew
Jack Beven, “TPC NHC Hurricane Andrew,” National Hurricane Center / NOAA.
they constituted a fundamental fight for survival against a federal legal framework that systematically prioritizes corporate energy interests and state-defined “consultation” loopholes over tribal interests
Archambault, David. “THE STANDING ROCK PROTESTS AND THE STRUGGLE FOR TRIBAL SOVEREIGNTY.” Journal of International Affairs 73, no. 2 (2020): 233–38. https://www.jstor.org/stable/26939979.
All are irreparable losses that violate both the American Indian Religious Freedom Act and treaty rights guaranteed under the 1851 and 1868 Fort Laramie Treaties, which promised uninhibited tribal use of ancestral lands.
Mengden, Walter H. “INDIGENOUS PEOPLE, HUMAN RIGHTS, AND CONSULTATION: THE DAKOTA ACCESS PIPELINE.” American Indian Law Review 41, no. 2 (2017): 441–66. https://www.jstor.org/stable/26492269.
Witnesses, including tribal leaders, environmental experts, and federal officials, described how a single leak could contaminate the river for hundreds of miles, destroying drinking water, fisheries, and ceremonial sites. Beyond the pipeline, the hearing also addressed historically low water levels caused by upstream dams, drought, and mismanagement of the Missouri River system, which have repeatedly led to shortages, boil-water advisories, and health crises on the reservation.
“WATER PROBLEMS on the STANDING ROCK SIOUX RESERVATION.” https://www.govinfo.gov/content/pkg/CHRG-108shrg97093/pdf/CHRG-108shrg97093.pdf.
delayed rebuilding, insurance barriers, rising recovery costs, and uneven public response.
Lynne McChristian, Hurricane Andrew and Insurance: The Enduring Impact of an Historic Storm (Insurance Information Institute, 2012), 2, 17–18. Gary E. Lehman, “Price Gouging: Application of Florida’s Deceptive and Unfair Trade Practices Act in the Aftermath of Hurricane Andrew,” Nova Law Review 17, no. 3 (1993): 1029–1034. Elliott Mittler, A Case Study of Florida’s Emergency Management Since Hurricane Andrew, Working Paper 98 (Natural Hazards Center, 1997), 10–11.
slow violence.
Rob Nixon, Slow Violence and the Environmentalism of the Poor (Cambridge, MA: Harvard University Press, 2011), 2.
Hurricane Andrew
Jack Beven, “TPC NHC Hurricane Andrew,” National Hurricane Center / NOAA.
.
The Mabo Case | Aiatsis Corporate Website. https://aiatsis.gov.au/explore/mabo-case
.
“Australia’s Water Market Is Excluding Indigenous People, Study Finds.” The Guardian, July 23,2020.https://www.theguardian.com/australia-news/2020/jul/24/australias-water-market-is-excluding-in`digenous-people-study-finds
.
Nixon, Rob. Slow Violence and the Environmentalism of the Poor. Harvard University Press, 2011.
“death-worlds”: spaces where toxic exposure is normalized, life expectancy is reduced, and residents are systematically subjected to the power of corporations and governments to dictate who may live and who must die, slowly
Davies, Thom. 2018. “Toxic Space and Time: Slow Violence, Necropolitics, and Petrochemical Pollution.” Annals of the American Association of Geographers 108 (6): 1537–53. https://doi.org/10.1080/24694452.2018.1470924
Standing Rock is not simply a site of victimization but of world-building: a place where protectors rehearsed an alternative future rooted in decolonization, mutual aid, and treaty-based nationhood
Estes, Nick. 2019. Our History Is the Future: Standing Rock versus the Dakota Access Pipeline, and the Long Tradition of Indigenous Resistance. S.L.: Verso.
explaining how Standing Rock is a “throughline” (a living thread connecting past and present struggles) where the same logics of extraction, treaty violation, and forced removal that characterized colonial expansion from the seventeenth century to the Indian Removal Act and the taking of the Black Hills now reappear in the form of fossil fuel infrastructure
Love, Nancy S. “From Settler Colonialism to Standing Rock: Hearing Native Voices for Peace.” College Music Symposium 58, no. 3 (2018): 1–16. https://www.jstor.org/stable/26608531.
Róisín Lanigan. The Internet Has a Cancer-Faking Problem. The Atlantic, May 2019. URL: https://www.theatlantic.com/health/archive/2019/05/faking-cancer-online/588334/ (visited on 2023-12-08).
It is so crazy that people will fake having some sort of illness all because of attention. I have first hand seen the toll that cancer has on families and how much it damages them. It is completely unsettling.
Sarah McQuate. 'I don't even remember what I read': People enter a 'dissociative state' when using social media. ScienceDaily, May 2022. URL: https://www.sciencedaily.com/releases/2022/05/220523135018.htm (visited on 2023-12-08).
This article really reminded me of why I hate doomscrolling. People literally melt their brains just scrolling away - obviously I'm joking but it does feel that way. I think especially for the younger generations there will be serious consequences for this. As I said in my previous comments, social intereactions are starting to near into a dissociative state. For kids, the implications of this in the future might be insane. I hope more research is done on this dissociative state from doom-scrolling, as there could be serious mental implications (which have been sort of covered in this reading).
In what ways have you found social media bad for your mental health and good for your mental health?
I've found social media has been bad for me because I find myself doomscrolling instead of doing things that could be productive. I also find that I watch news a lot more and my anxiety tends to be higher when I do engage in more news sources. Something good that has come out of social media is that I am more informed about what is going on. Also, I have learned a lot of new information and I am able to communicate with friends and family who aren't close distance.
the opposition to the Dakota Access Pipeline was never merely political or economic; it was deeply spiritual, rooted in a worldview where water is sacred and inseparable from life itself
Zambelich, Ariel, and Cassi Alexandra. 2016. “In Their Own Words: The ‘Water Protectors’ of Standing Rock.” NPR, December 11, 2016, sec. National. https://www.npr.org/2016/12/11/505147166/in-their-own-words-the-water-protectors-of-standing-rock.
The water protectors at Standing Rock- many of them mothers, grandmothers, and young activists- embodied a form of resistance rooted not in protest but in sacred obligation, revitalizing matriarchal leadership and demonstrating that the struggle for environmental justice is inseparable from the struggle for Indigenous sovereignty and gender justice.
Dennis, Mary Kate, and Finn McLafferty Bell. “Indigenous Women, Water Protectors, and Reciprocal Responsibilities.” Social Work 65, no. 4 (October 1, 2020)
.
Perry et al. “Availability of Drinking Water in Rural and Remote Communities in New South Wales, Australia.” Australian Journal of Primary Health 28, no. 2 (February 1, 2022): 125–30. https://doi.org/10.1071/py21119.
.
“Understanding Water Trading Prices in the Murray-Darling Basin.” Aquifa. https://www.aquifa.com.au/post/understanding-water-trading-prices-in-the-murray-darling-basin
.
Hartwig, Lana D., Natalie Osborne, and Sue Jackson. “Australia Has an Ugly Legacy of Denying Water Rights to Aboriginal People. Not Much Has Changed.” The Conversation, July 23, 2020. https://theconversation.com/australia-has-an-ugly-legacy-of-denying-water-rights-to-aboriginal-people-not-much-has-changed-141743.
.
“Australia’s Water Market Is Excluding Indigenous People, Study Finds.” The Guardian, July 23,2020.https://www.theguardian.com/australia-news/2020/jul/24/australias-water-market-is-excluding-in`digenous-people-study-finds
.
Rodgers, Dennis, and Bruce O’Neill. “Infrastructural Violence: Introduction to the Special Issue.” Ethnography 13, no. 4 (October 23, 2012): 401–412. https://doi.org/10.1177/1466138111435738.
I would need eleven additional tokens for digits 0 to 9 and PERIOD.
Nope. You could just use the traditional approach where most tokenizers only have a generic tag/discriminant for all number tokens. The every-non-text-token-is-one-character constraint is arbitrary and unnecessary.
As stated in this paper, Laughlin’s 1981 data provided a poor fit to the measured response nonlinearity. But Laughlin made more accurate measurements in 1987, which provided an excellent fit to the measured response nonlinearity. See:
Laughlin, SB, Howard, J, and Blakeslee, B. Synaptic limitations to contrast coding in the retina of the blowfly Calliphora. Proc Royal Society London B, 231(1265):437–467, 1987.
This method does not require the utilization of solvents, which are easy to handle and can be produced on a large scale, yielding highly crystalline materials with few defects and high stability.
Poorly written sentence. This method does not require the utilization of solvents. Is it easy to handle, ...
Winter kept us warm
Straight up paradox where winter is being used as a weather that is sunny and hot because of the word 'warm' there.
Doomscrolling is: “Tendency to continue to surf or scroll through bad news, even though that news is saddening, disheartening, or depressing. Many people are finding themselves reading continuously bad news about COVID-19 without the ability to stop or step back.” Merriam-Webster Dictionary [m8]
I hate doomscrolling. The term itself is more of a joke, and was initally used as one. However, and I mean this seriously, its become a lifestyle for some people. I know this, as I almost fell into the trap of endlessly doomscrolling instead of working. The amount of times I've gone to hang out with close, and distant friends, and it ends with people doomscrolling instead of being in the moment is CRAZY. I think there are serious implications of it too, so I can't wait to read more to find out. Covid was a starting point for this addiction in my eyes.
But suppose a private club has a policy that excludes women from joining. How do we balance the right to freedom of association—which would permit the club to decide for itself whom to admit—against the right not to be discriminated against—which requires equal treatment of women? In cases such as this, we need to examine the freedoms or interests at stake and decide which of the two is the more crucial for securing human dignity
Kant would argue that this private club is discriminatory against women because each human being has human dignity and should be treated as such.
About Us
All interactive elements in the top navigation bar (dropdown menus, search icon, and text links) are operable without a mouse. A keyboard user can press Tab to move through each item and Enter or Space to expand dropdowns. This follows the Operable principle, ensuring people with motor impairments who cannot use a mouse can still access all site functions.
Our Community
The navigation is consistent and predictable, with clear link text like ‘Join Us’ and ‘Free White Cane Program’. The layout avoids complex fonts, moving elements, or cognitive overload, supporting the Understandable principle for users with dyslexia, ADHD, or autism.
utilitarianism asks us to look beyond self-interest to consider impartially the interests of all persons affected by our actions.
This made me think about the trolly problem through a utilitarian lens. If there are five people tied to a track and one person tied to a diverted track, and if you pull a lever you can make the trolly divert so it only kills one person, saving five you are saving five people, but you individually are making an effort to purposely kill one.
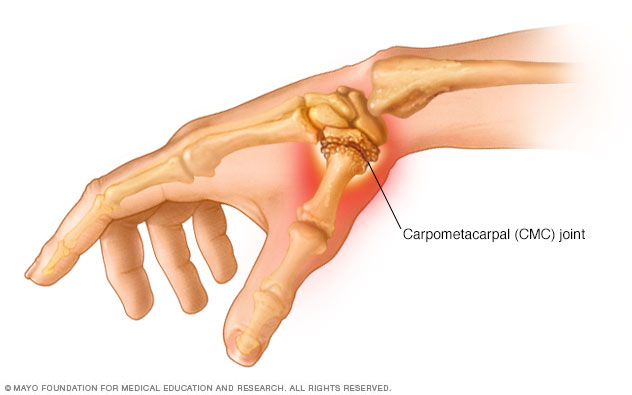
arti-culaciones MCF

primera articulació n CMC.
This protocol balances the energy consumption, improves network lifetime and achieves better throughput. The paper [12] devises a data-securing model for WSN based on a linearly complex voice encryption mechanism of a global system for mobile technology. The work presented a secure, low complexity and energy efficient routing solution for WSNs.
Le commentaire : Cette partie est importante !
FAQs
(Robust Principle) The footer is a critical part of the site's architecture, tying back to the Robust principle. The developers need to ensure this section remains adaptable and easily interpreted by a wide variety of browsers, mobile devices, and assistive technologies. The goal is to present the content in simpler ways without losing the underlying information or structure.
Crave-worthy snacks
(Foreground/Background Contrast) The banner directly below this section has text layered directly over a really busy background image of potato chips. People with limited vision might have a really hard time distinguishing the foreground text from the background there. It is a great reminder that good web design needs clear contrast, otherwise, you are completely blocking some users from reading the message.
Weekly Flyer Features
(Understandable Principle) The product grid below this header is incredibly dense with varying fonts, prices, and product images. From a structural standpoint, presenting content in a simple, predictable layout is key to the Understandable principle. Web content should be structured so that it makes sense when read in a linear fashion. If a page is too cluttered, it can cause serious cognitive overload for users with ADHD or dyslexia.
Start shopping
(Operable Principle) While digging through the site's layout near this shopping section, I realized how heavily the main site menus rely on a mouse. To actually meet the Operable standard, the main navigation interfaces need to be fully functional using just keyboard commands. Not everyone can use a mouse, especially users with motor impairments, so the navigation cannot require physical interactions they simply cannot perform.
Show some love for the moms in your life
(Perceivable Principle) I noticed this big promotional banner right away, but it made me wonder how it translates for someone using a screen reader. According to the Perceivable principle, the image next to this text needs a concise <alt> tag of 125 characters or less so visually impaired users don't miss out on the information. If it is just named something random like "IMG_098.jpg" in the code, the site is failing to make this content truly presentable to everyone.
An image of houses in the village of Savoonga during the day.Source: knom radio mission
Hovey, Davis. "Savoonga, Alaska," KNOM. 2017. https://knom.org/2023/03/01/st-lawrence-island-researchers-investigate-health-consequences-of-pollution/.
An interactive visual that explains how St. Lawrence Island became polluted and what is being done to adress the pollution.
Smith, Timothy. "Lt. Smith shows us the beauty of Gambell Alaska," Office of Coast Survey, 2015.
Department of Environmental Conservation. “Northeast Cape and Gambell Formally Used Defense Sites”. 2019. https://ejatlas.org/print/formally-used-defense-sites-in-alaska.
Apatiki, L. Pelowook, T. Sheffield, G. Ahmasuk, A. Tokeinna, R. "Photograph of garbage that washed ashore in the Bering Strait region," 2020.
Apassingok, Merle Apassingok. “Press Conference on Health & Human Rights of the Indigenous People of Sivuqaq and Future Generations.” Alaska Community Action on Toxics, March 12, 2025. https://www.youtube.com/watch?v=Isd5uuDbMbA&t=1234s.
press conference
Apassingok, Merle Apassingok. “Press Conference on Health & Human Rights of the Indigenous People of Sivuqaq and Future Generations.” Alaska Community Action on Toxics, March 12, 2025. https://www.youtube.com/watch?v=Isd5uuDbMbA&t=1234s.
slow violence
Nixon, Rob. Slow Violence and the Environmentalism of the Poor. (Harvard University Press, 2011), 152.
short video documentary
Wilson, Jim, Kirk Johnson and Channon Hodge. “Science at the End of the Earth | The New York Times”, The New York Times, August 7, 2015, https://www.youtube.com/watch?v=PA1ON0ycdRY.
From the map on the right
Kovacs, Elizabeth. “Environmental Justice Disparities in Alaska Native Communities,” D.U.Quark, 4, no. 1 (2019): 43-44. https://dsc.duq.edu/duquark/vol4/iss1/10.
St. Lawrence Island Yupik whalers flense blubber from a 65-foot bowhead whale.
McCutcheon, Steve. Sea, Lands, Rivers: Food for the village, Smithsonian Learning Lab.
Poland is now among the world’s 20 largest economies. How it happened
So, for the world’s producers of commodities and manufactures, what happens in China is a big determinant of the prices of goods, while what happens in the US is a big determinant of the costs of borrowing and servicing debt. And when these two forces are out of phase with each other, many countries around the world (and particularly commodity exporters in the global south) face booms, busts, or both in succession.
Nicely put.
pouvant exprimer une forme d’émotion via leur interface corporelle
C'est intéressant, les formes d'émotion du robot peuvent-elles être spontanées et sortir du programme initialement prévu par son créateur humain ? Le robot peut-ils devenir autonome ? C'est une question très pertinente actuellement.
volume très important d’entretiens
Nous pouvons nous demander combien d'entretiens et les conditions de réalisation de ces entretiens, les profils interrogés, le périmètre...
For more information about Kubb lawn game, see:-
For more information about float bridges, see
LLM-as-forecaster 路线基本死
所以最近关于利用大模型做时序传统任务的论文要尽快处理完。不然越来越没有机会了,
https://bafybeig26duct5lo3pyywd2pjg5nipneusuqecw7vfc3uiabi57vpft4fm.ipfs.dweb.link?filename=magyarorszag.mp4
``` There's a country Where I walked in my dream: Hungary Where I saw My own face in yours You were rich in embraces And you were loyalty in true things I'm guarding this face Hungary! I believe in my dream through a lifetime... Hungary! You're written into my heart Hungary! You're entrusted to my soul Hungary Now be the one to lead me And help me with the things I have to do Be the certainty inherent in tomorrow I only entrust you my life...
There's a country Where I walked in my dream: Hungary Where I saw My own face in yours Come on and finally once When tomorrow rises Be the one who waits me Come on and snuggle me again Hungary I've been waiting for you for a thousand years...
Come on and finally once When tomorrow rises Be the one who waits me Come on and snuggle me again Hungary I've been waiting for you for a thousand years...
Hungary! If I walk on foreign lands Hungary With me even the road turns Back to you home
With me the ancients ask you To let me continue living in you As they live inside me Hungary Get blessings of millions!
Be the certainty inherent in tomorrow! I only entrust you my life...
Be the certainty inherent in tomorrow! Get blessings of millions! ```
tomorrow
new dawn
Be the certainty inherent in tomorrowI only entrust you my life...
Be that Hungary
Le groupe gallo-roman
Exercice d'insertion d'image
langue d’oc.1
Exercice de notes de bas de page
Les
Exercice sur l'italique
citation
Exercice de citation
l’UNESCO
On peut préciser son nom complet : Organisation des Nations unies pour l'éducation, la science et la culture
Voici un résume test.
Le résumé sera à compléter après la rédaction complète de l'article.
Analyse des chartes choisies
La table des matières reste à compléter une fois les sous-titres stabilisés.
tandis que les textes non littéraires retiennent beaucoup moins l’attention
On pourrait ajouter un bref état de la recherche, éventuellement appuyé sur des données quantitatives, afin de montrer que les textes non littéraires ont effectivement reçu moins d'attention que la poésie lyrique des troubadours.
ordonnance de Villers-Cotterêts (1539)
Elle a imposé l'usage du français dans les actes officiels et judiciaires.
Brunel, Clovis. 2012. Les plus anciennes chartes en langue provençale. Recueil des pièces originales antérieures au XIIIe siècle, publiées avec une étude morphologique. Genève: Slatkine Reprints.
Il faudrait ajouter aussi les éditions originales des deux volumes du recueil : 1. Brunel (Clovis), 1926. Les plus anciennes chartes en langue provençale. Recueil des pièces originales antérieures au XIIIe siècle publiées avec une étude morphologique, Paris, Picard, LXIII-499 p. @book{brunel1926chartes, author = {Brunel, Clovis}, title = {Les plus anciennes chartes en langue provençale. Recueil des pièces originales antérieures au XIIIe siècle publiées avec une étude morphologique}, year = {1926}, address = {Paris}, publisher = {Picard}, pages = {LXIII--499} } 2. Brunel (Clovis), 1952. Les plus anciennes chartes en langue provençale, supplément. Recueil des pièces originales antérieures au XIIIe siècle, Paris, Picard, 275 p. @book{brunel1952supplement, author = {Brunel, Clovis}, title = {Les plus anciennes chartes en langue provençale, supplément. Recueil des pièces originales antérieures au XIIIe siècle}, year = {1952}, address = {Paris}, publisher = {Picard}, pages = {275} }
En 1952, le Supplément qui ajoute 191 pièces, dont 166 inédites, a vu le jour.
Ajouter aussi la référence du Supplément de 1952 au format BibTeX.
En 1926, le premier tome qui comprend 349 chartes datées du XIe au XIIe siècle paraît aux Éditions Picard.
La première version de ce recueil a paru en 1926, il faudrait ajouter la référence bibliographique en BibTeX.
Dans ce contexte, le nombre de locuteurs continue de diminuer.
Il faudrait ajouter une référence ou des données récentes pour justifier cette affirmation, p. ex. à partir des dernières enquêtes sur le nombre de locuteurs.
variétés
Il vaut la peine d'ajouter un petit paragraphe qui clarifie les différences entre les notions de langue, dialecte et variété.
Oláh Ibolya: "A Magyarország c. dalt már nem merem előadni" | PartizánPOP
nem merri
Oláh Ibolya: Magyarország (Rendszerváltás 2026 edition)
olah ibolya did not sing since 2005
magyarorszag olah ibolya 2026
History
Free Verse: The poem is written in three stanzas of uneven length
However, this notion is not confined to grid-constrained markets. France, which had a surplus grid capacity originally planned for transport and heating, is finding that AI clusters have been faster to claim, displacing the very sectors the headroom was intended to serve. 150
Indeed, it is technically possible to dynamically reallocate idle reserved capacity to other tenants, as modern AI cluster management systems can automatically assign other workloads to GPUs that are temporarily idle, and production deployments have demonstrated that dynamic sharing can increase utilisation from less than 30% to more than 70%. 137 However, the stronger the access guarantees owed to capacity holders, the less freedom the scheduler has to fill their idle periods with other workloads; we call this ‘the self-hosting trap’. Public law entities expecting guaranteed on-demand access cannot accept their reserved capacity being occupied by other jobs at the moment they submit their own workloads
I guess this is the flip side of it not being easy to overcommit capacity. I’ve seen examples of people selling pre-emptible capacity though from Nebius.
The self-hosting arrangements embedded in the AIGF governance structure also introduce a governance constraint on scheduling flexibility. Under the EuroHPC Regulation, the Union’s financial contribution, capped at 17% of the capital expenditure, entitles it to proportional access, with priority for public law entities, industrial users on EU-funded projects and SMEs
Is this the mechanism being used to provide a degree of subsidised capacity?
A multi-client AIGF producing sub-second power swings of this magnitude is therefore actively increasing the system’s balancing costs, not merely underutilising its connection
This is a key point that I haven’t seen expressed in any discussions of European data Centre build out yet. really good to actually see it named.
The hardware provided for those peaks continues drawing power through the quiet periods. Independent grid data corroborates the scale of the gap. Ember’s analysis of European TSO data revealed that data centres use, on average, only 44% of their contracted grid capacity, using 34% of it in Ireland and 30% in Norway
Does this actually include the redundancy? we saw a lower figures like this in the UK, and it wasn’t clear that it was because of their being twice the capacity made available for redundancy purposes to a data Centre
The multi-client scenario assumes a fragmented user base of enterprises, SMEs, public institutions and research organisations with heterogeneous, largely inference-driven workloads. 123 The paper concludes that given Europe’s provider landscape and user base, the anchor customer scenario is improbable and the multi-client scenario is far more realistic. This chapter takes that conclusion as its starting point. The following subsections show how a multi-client AIGF structure sharpens three bottlenecks—the utilisation gap and its energy costs, load variability and system-level impacts, and scheduling complexity—and contrast these outcomes with the anchor customer model to illustrate what the same GPU hardware would look like under a different use case.
Maybe this represents the way to standardise on making demand more amenable to to flexibility, as AIGF presumably would have some pricing power if they are offering access at subsidised rates.
Formscapes
TrailScapes
RRID:AB_477010
DOI: 10.1186/s40478-026-02240-y
Resource: (Sigma-Aldrich Cat# G3893, RRID:AB_477010)
Curator: @scibot
SciCrunch record: RRID:AB_477010
RRID:AB_2572311
DOI: 10.1186/s40478-026-02240-y
Resource: (EnCor Biotechnology Cat# MCA-5C10, RRID:AB_2572311)
Curator: @scibot
SciCrunch record: RRID:AB_2572311
RRID:AB_3678889
DOI: 10.1186/s40478-026-02240-y
Resource: RRID:AB_3678889
Curator: @scibot
SciCrunch record: RRID:AB_3678889
RRID:AB_2109953
DOI: 10.1186/s40478-026-02240-y
Resource: (EnCor Biotechnology Cat# CPCA-GFAP, RRID:AB_2109953)
Curator: @scibot
SciCrunch record: RRID:AB_2109953