Freedom internet provider on the difficulty of blocking Russian state media sites. - there is a court order to block sanctioned websites - however there is no official list of sanctioned websites - there are several lists from different MS and branche organisations with suggested sites, but this is never an official list or effort. - one of those lists, the one from Lithuania contains sites that are clearly not Russian state media (an Indian social media platform, and a generic video sharing site)
10,000 Matching Annotations
- Jan 2026
-
drive.google.com drive.google.com
-
I also picked up that Superman comic book. Each panel, complete with picture, dialogue and narrativewas a three-dimensional paragraph. In one panel, Superman breaks through a door. His suit is red, blue and yellow. The brown door shatters into manypieces. I look at the narrative above the picture. I cannot read the words, but I assume it tells me that "Superman is breaking down the door." Aloud, I pretendto read the words and say, "Superman is breaking down the door." Words, dialogue, also float out of Superman's mouth. Because he is breaking down the door,I assume he says, "I am breaking down the door." Once again, I pretend to read the words and say aloud, "I am breaking down the door" In this way, I learnedto read.
Here, Alexie explains how he started to learn to read by saying the words he believed to be there out loud. it was through this that he became verbal in his reading and learned to comprehend these words.
it was similar to how i learned to read and write, my mother bought the little Einstein DVDs and played them on the TV, it was through these DVDs that i learned more than just words, but comprehension further more.
-
I refused to fail. I was smart. I was arrogant. I was lucky. I read books late into the night, until I could barely keep my eyes open.
The author reveals why reading is such a core aspect of his existence and purpose. He came from an "Indian" family where they were set up to systemically fail, but he did not ever let that stop him, but rather give him the courage to strive for what he knew was his calling and passion.
-
The words inside a paragraph worked together for a common purpose. They had some specific reason for being inside the same fence. Thisknowledge delighted me. I began to think of everything in terms of paragraphs. Our reservation was a small paragraph within the United States. My family'shouse was a paragraph, distinct from the other paragraphs of the LeBrets to the north, the Fords to our south and the Tribal School to the west. Inside ourhouse, each family member existed as a separate paragraph but still had genetics and common experiences to link us. Now, using this logic, I can see mychanged family as an essay of seven paragraphs: mother, father, older brother, the deceased sister, my younger twin sisters and our adopted little brother
This paragraph is so beautiful and carries so much weight. The author views paragraphs as a different interpretation to each individual, being his family members, ones with different characteristics, meaning, and purpose.
-
Inside ourhouse, each family member existed as a separate paragraph but still had genetics and common experiences to link us.
Reading became a sentimental part of this family, and there is a lot of genuine love tied to literature because of the impact books had on this family.
-
-
stats.libretexts.org stats.libretexts.org
-
Descriptive statistics are just descriptive. They do not involve generalizing beyond the data at hand. Generalizing from our data to another set of cases is the business of inferential statistics,
This calls out typical mistakes that we've read out about earlier and see in today's world. I've definitely seen people treat a big mean difference or a striking chart like it automatically proves something about the world. This line is basically a warning: descriptive stats tell you what happened in your dataset, but they don't automatically justify claims about a population.
-
-
crmintler.com crmintler.com
-
By the time Prohibition went into effect, 16 January 1920,other street-gang teen-agers were coming of age to make themselves aname in organized crime: Meyer Lan
I think it's incredibly interesting that most of the organized crime that built the gangster character that we know of today started with men of a very young age. Most of the famous depictions of gangster life have the characters being a bit older-- the seasoned mob boss kind of character-- but it makes sense that in actuality, the adaptation that was needed to survive with this lifestyle came from a younger audience.
-
-
centreforpublicimpact.org centreforpublicimpact.org
-
A definition of community must be able to account for the different types of communities that exist in the world.
I am guilty of this myself, but I feel like most people only think of community in a way in which it applies to them. For example , when I think of community, I tend to think more of Lincoln than Omaha.
-
-
stats.libretexts.org stats.libretexts.org
-
Interval scales are not perfect, however. In particular, they do not have a true zero point even if one of the scaled values happens to carry the name “zero.”
This is subtle but a critical idea. It explains why certain intuitive comparisons, like saying something is "twice as hot", are actually incorrect. It shows how measurement scales affect interpretation, not just calculation.
-
-
human.libretexts.org human.libretexts.org
-
No matter the reader, no matter the writer, no matter the genre, literature is a cultural relic,
I think this is the best representation of what I believe literature is to our society. Many people say ai will change literature permanently but I believe that literature will continue to thrive
-
But... what if we were to define literature as Sidney did--a creation made from the "zodiac of [the poet's] own wit," improving upon nature itself through invention? Might some of today's internet media fall into that definition?
This changes how literature is seen because it used to mean to most people as outdated and boring, but it's how we evolved and it still exists in other types of media rather than plays.
-
As we continue to gain increasing complexity in terms of measuring brain activity and developing other tools to measure brain function, scientists may find more benefits.
I want to get back into reading, but I haven't read in my own free time for like 6-7 years. The benefits are very beneficial and I can see it helping me in my future life.
-
Indeed, if we define literature as only printed novels and poetry, perhaps it is, for all intents and purposes, dead. But... what if we were to define literature as Sidney did--a creation made from the "zodiac of [the poet's] own wit," improving upon nature itself through invention? Might some of today's internet media fall into that definition?
Literature takes forms that aren't as traditional compared to the past. I think the most prominent ways are over social media and through movies and TV because they tell their own messages too and lots of people consume them.
-
claim that literature is dead
online news and other sources of news are often seen as literature, but what the author argues here is the challenge of the idea in which literature must be a book or a poem. This is a way to connect old literature to modern day.
-
witter, Youtube, and streaming television services, literature might seem like a relic of the past. Indeed, fewer people are reading literature than ever
this reminds me of our discussion in class on how it is slowly becoming less popular today and there is a cultural shift away from the books, but not that literature is not important today or has no value.
-
writing about literature will let you exercise your critical thinking skills like no other style of writing will. Even if you don't want to pursue a career involving literature, you can use critical thinking and analysis in any field from philosophy to business to physics. More than being able to think critically, you need to be able to express those thoughts in a coherent fashion. Writing about literature will allow you to practice this invaluable communication skill.
Not only does reading help to exercise your brain but so does writing as it helps you to form your thoughts and be able to express your thoughts and feelings.
-
What if literature isn't dead after all...but thriving more than ever? What if we radically reconsider the parameters of literature? What if literature has just evolved from sonnets and novels to tweets and memes?
making the argument that literature can be vast and be placed onto more pieces than most people are consuming literature without knowing.
-
In order to write a strong paper, you will need to examine a text both subjectively and objectively.
This is very true, especially if the subject matter is something personal to you. Its important to be subjective, but also know where to be objective.
-
But... what if we were to define literature as Sidney did--a creation made from the "zodiac of [the poet's] own wit," improving upon nature itself through invention? Might some of today's internet media fall into that definition?
I feel that there should be a broader definition of literature, but not too broad. Some things on the internet shouldn't be classified as literature.
-
What if literature has just evolved from sonnets and novels to tweets and memes?
I feel like teens would enjoy literature way more but wouldn't take it seriously. It wouldn't help with reading skills as memes and brainrot contain slang most of the time.
-
What if literature has just evolved from sonnets and novels to tweets and memes?
This is a good view because evolution may look different but still possess the same intent. After all, people are still getting the word out it’s just more straightforward.
-
t if literature isn't dead after all...but thriving more than ever? What if we radically reconsider the parameters of literature? What if literature has just evolved from sonnets and novels to tweets and memes? In this textbook, we will explore how technology has blurred the lines between Literature and literature
This is like what we said in class and how literature can be defined as anything that expresses something or has meaning. In the past, we have literary works like Fahrenheit 451 and Animal Farm that is a metaphor for historical events to convey their ideas. The same can be seen for memes that embody political cartoons to satirize what is going on in the world right now.
-
writing about literature will let you exercise your critical thinking skills like no other style of writing will. Even if you don't want to pursue a career involving literature, you can use critical thinking and analysis in any field from philosophy to business to physics. More than being able to think critically, you need to be able to express those thoughts in a coherent fashion. Writing about literature will allow you to practice this invaluable communication skill.
The reason why I choose this quote is because despite being casted out of people's interest of entertainment, it's a good source of thinking. Almost everybody will not use literature in their lives but it can help increase brain activity. Leading to more critical thinking and effecting learning.
-
Indeed, if we define literature as only printed novels and poetry, perhaps it is, for all intents and purposes, dead.
Literature is a much wider range of ideas than just reading, even if we don't consciously realize it :0 People consider literature dead, but only because a lot of people don't think about how we constantly consume things that can be considered literature as well.
-
More than being able to think critically, you need to be able to express those thoughts in a coherent fashion. Writing about literature will allow you to practice this invaluable communication skill.
This shows the true importance of not only being able to read literature but also being able to understand and express what you think of it in your own creative and critical way. It tells the reader how it can prove to be an amazing skill in communication and being able to organize one's thoughts coherently.
Annotators
- ElijahChua
- derecknguy
- AriNunez19
- claryy
- MatthewCri
- AudreyOu
- chris_X
- Ethan_D
- mehtaabtoor
- MJnew
- Aysaiah
- Rapopel
- jocelynss
URL
-
-
drive.google.com drive.google.com
-
(16.5) I found this passage very interesting. Confucius is not anti-pleasure seeking, but has outlined certain sources of enjoyment that are acceptable, and some that are not. An interesting question came to my mind about pleasure seeking in the modern day. How are rituals and music different today than when Confucius was writing? Do these still maintain the same improving qualities, or have they become distractions and creators of arrogance, the negative qualities of enjoyment highlighted by Confucius? I personally believe that they are still important for improvement. I often use music to help me focus on my work and when I exercise. Even while music has changed significantly, it still holds meditative attributes that can be positive influences in our lives. However, I do think that some pleasurable practices of the modern day have drifted into the realm of arrogance. Due to social media, lots of people seek pleasure in the feeling of superiority over other people they see online. This is a modern example of what Confucius would refer to as “finding enjoyment in being arrogant.”
-
(17.16) Frequently, Confucius shows a nostalgia for the past, and a cynicism about the present, as he does in this passage. I take issue with this line of thinking. I fear that only looking backwards, and attempting to emulate an ideal past, may prevent people from solving problems that did not exist back then but are specific to the present. It is also entirely possible that, because people view the past through “rose-colored glasses,” emulating the past might not actually be as productive as it seems. Every era tends to have its problems, and if we attempt to emulate the past, we may instead revive old issues that were forgotten about.
-
(1.8) “Do not have as a friend anyone who is not as good as you are.” At first, I wanted to push back on this assertion. I interpreted it as Confucius urging us to avoid friendships with people who lack the same moral goodness. What if the person is in the earlier stages of their moral development? What if our exclusion of them infringes on this growth? While I agree that the people we keep around us are in many ways a reflection of ourselves, I do not think we must exclusively befriend people who are “as good” as we are. If that is the case, how will good morals and ethics spread through society? All the “good” people will coexist in friendship with each other and the less “good” people will be pushed away. However, upon further analysis, I also understand it to not be saying only befriend saints, but simply be careful who you choose to be intimate influences and who you mold yourself after. It is not making an elitist claim, just advising us on who we surround ourselves with. We should not be close with others who might lower our moral ceilings. I agree with Confucius here if my second interpretation of the meaning is what he intended.
-
(11.11) This feels like an example where ritual is broken for the virtues of humanity and respect. The students are almost like sons to The Master, so they show a sort of filial piety onto him. Yet, they still have moral and social allegiance to their peer. I find it interesting that Confucius ideas have rules not in writing but in feeling and respect
-
(18.2) I am not 100% certain if I am interpreting this passage correctly, but Liu Xiahui seems to be saying that if he continues to be a morally good, honest, and kind man, he will get dismissed from his position in any state that he works. It is the morally corrupt men that never need to move positions. What he seems to be suggesting is that it is “easier” to compromise one’s morals to escape consequences than it is to stay true to one’s ethical values. Liu Xiahui is an exemplary person because he values doing what is morally right over his own position. Confucius seems to be in favor of valuing integrity (even with the cost of “failure”) over convenience and safety.
-
9.23: Passage 9.23 stood out to me because it seemingly contradicts the previously established fundamentals of filial conduct. A different reading of it, however, suggests that by "esteem," Kongzi means an even greater degree of reverence than that proposed by filial conduct, akin to not only respecting someone but also recognizing their potential to become a junzi, or at least someone closer to one. Then it would make greater sense for this esteem to be withheld from those who reach "forty or fifty years and yet have done nothing of note." What is your interpretation of what Kongzi means by "esteem" in this passage?
-
4.4: While a short passage, I believe 4.4 encapsulates the fundamental theme of Kongzi's thinking that one is never born with ren or as a junzi, but must rather seek to attain these ideals through learning. By emphasizing how "one's purpose" is the driving force behind doing what is right, he is highlighting the role of our free will. Essentially, choosing to be a better person is the first step in becoming one.
-
9.19: This passage seems to relate to Confucius' view of self-cultivation as a constant task. While it may drain one of energy, the importance of continuing to cultivate oneself cannot be overlooked. What would Confucius say about someone continuing to dumping basketfuls of dirt, but these basketfuls being much lighter than they can actually carry. Would he judge their slow progress and low effort? I think that he might, given that effort is necessary to Confucius in all aspects of a life worth living. Just as with rituals, perhaps he would stress the need for full and genuine effort in every action.
-
-
social-media-ethics-automation.github.io social-media-ethics-automation.github.io
-
Anonymity can also encourage authentic behavior. If there are aspects of yourself that you don’t feel free to share in your normal life (thus making your normal life inauthentic), then anonymity might help you share them without facing negative consequences from people you know.
I think it’s worth mentioning that on anonymous sites like 4chan, anonymity both encourages and discourages 4chan. Many people strongly associate 4chan with trolling culture and therefore inauthentic, but another large part of 4chan is sharing stories(green text), which in the majority of cases have some underlying truth to them.
-
nonymity can encourage inauthentic behavior because, with no way of tracing anything back to you1, you can get away with pretending you are someone you are not, or behaving in ways that would get your true self in trouble.
People only act like themselves when they are the ones to face the repercussions of their actions. Anonymity gives people the liberty from facing consequences therefore they tend to act more irrationally. Existentialism says that people are defined by their actions, but if no one ever knew who did them, then actions don't help define anyone.
-
6.6.2. Anonymity encouraging authentic behavior# Anonymity can also encourage authentic behavior. If there are aspects of yourself that you don’t feel free to share in your normal life (thus making your normal life inauthentic), then anonymity might help you share them without facing negative consequences from people you know. 6.6.3. Is authentic self-expression good?# We can next ask if authentic self-expression is a good thing or not. But that depends, what is the authentic thing about yourself that you would be expressing? Are you authentically expressing hate or cruelty? If so, perhaps authentic self-expression is morally bad. Are you part of an oppressed or marginalized group that has been restricted from self-expression? Then perhaps expressing yourself is morally good. (See Trans Twitter and the beauty of online anonymity)
Anonymity itself is not good or bad; it is more like an amplifier that amplifies what the person is trying to express.
If a person uses anonymity to express hatred and attack others, then this expression of the "true self" is clearly morally questionable because it directly harms others. But on the other hand, for some people who are repressed and marginalized in real life, anonymity may be the only safe way for them to express their true selves. In this case, anonymity is not a shirking of responsibility, but a form of self-protection.
-
-
social-media-ethics-automation.github.io social-media-ethics-automation.github.io
-
In 2016, the Twitter account @Sciencing_Bi was created by an anonymous bisexual Native American Anthropology professor at Arizona State University (ASU). She talked about her experiences of discrimination and about being one of the women who was sexually harassed by a particular Harvard professor. She gained a large Twitter following among academics, including one of the authors of this book, Kyle. Separately, in 2018 during the MeToo movement [f7] , one of @Sciencing_Bi’s friends, Dr. BethAnn McLaughlin (a white woman), co-founded the MeTooSTEM non-profit organization, to gather stories of sexual harassment in STEM (Science, Technology, Engineering, Math). Kyle also followed her on Twitter until word later spread of Dr. McLaughlin’s toxic leadership and bullying in the MeTooSTEM organization (Kyle may have unfollowed @Sciencing_Bi at the same time for defending Dr. McLaughlin, but doesn’t remember clearly). Then, in April 2020, in the early days of the COVID pandemic [f8], @Sciencing_Bi complained of being forced to teach in person at ASU when it wasn’t safe, and then began writing about their COVID symptoms. Fig. 6.2 Tweet from @Sciencing_Bi complaining about covid, sexual harassment (Title IX) descrimination for their race (BIPOC), and mentioning that they are still posting anonymously. (screenshot source)# On July 31st, Dr. McLaughlin announced that @Sciencing_Bi had died of COVID. Fig. 6.3 Tweet from Dr. McLaughlin reporting that @Sciencing_Bi died of COVID. (screenshot source [f9])# Many academics on Twitter expressed sympathy and outrage over what had happened to @Sciencing_Bi. Dr. McLaughlin held a memorial service for @Sciencing_Bi online via Zoom. A few people showed up to Dr. McLaughlin’s online Zoom memorial service for @Sciencing_Bi, but the guests felt something was off. Afterward, Mr. Eisen began to search for any evidence that @Sciencing_Bi had been a real person. He could not find any. “The combination of the weird things that were happening on the call and looking at the tweets and seeing how much they circled BethAnn, it just became obvious to me,” he said. “‘Oh, [@Sciencing_Bi] is BethAnn.’” The Anonymous Professor Who Wasn’t - The New York Times [f10] Arizona State University confirmed that they had no professors who matched the description of @Sciencing_Bi. Dr. McLaughlin’s and @Sciencing_Bi’s accounts were suspended from Twitter for violating Twitter policies, and Dr. McLaughlin eventually confirmed that she had completely invented @Sciencing_Bi.
These things in my point of view are the things that is easily triggers people off. The account owner used someone else's identity posting things that is not true, use fake story to gather people's sympathy to make themselves popular. However people found out that it was all fake, even the account holder herself is a bully. The feeling of being used, deceived, can make people easily angered, upset, self doubt on how did us fell into someone's trap, and became a tool for her to gather fans and popularity.
-
-
drive.google.com drive.google.com
-
(Response to Rafael PL on 4.4) To Rafael's point, I think this short analect captures the flexible – and deeply personal – nature of Kongzi's teachings. The manifestation of authoritative conduct and its practical application in life can vary greatly from person to person, but it stems from cultivating a gentlement-like self – a personhood rooted in morality and uprightness. Everyone's calling is different, but so long as they maintain their values and have positive influence in their respective spheres, they can do no wrong. This also connects to my previous annotations about Kongzi's commending of living simply. Everyone's purpose is different, and material wealth and extravagance is of little significance. What matters is the development of the person.
-
-
www.biorxiv.org www.biorxiv.org
-
Note: This response was posted by the corresponding author to Review Commons. The content has not been altered except for formatting.
Learn more at Review Commons
Reply to the reviewers
We are grateful to the Review Commons reviewers for their constructive feedback, which has significantly strengthened the manuscript. In response, we have performed additional experiments, revised and expanded multiple figures, incorporated new statistical and functional analyses, and carefully edited the text to improve clarity and precision. A detailed point-by-point response to all reviewer comments, together with a summary of revised figures, is provided.
To address the reviewers' suggestions, we have conducted additional experiments that are now incorporated into new figures, or we have added new images to several existing figures where appropriate.
For this reason, please note that all figures have been renumbered to improve clarity and facilitate cross-referencing throughout the text. As recommended by Referee #3, all figure legends have been thoroughly revised to reflect these updates and are now labeled following the standard A-Z panel format, enhancing readability and ensuring easier identification. In addition, all figure legends now include the sample size for each statistical analysis.
For clarity and ease of reference, we provide below a comprehensive list of all figures included in the revised version. Figures that have undergone modifications are underlined.
Figure 1____. The first spermatogenesis wave in prepuberal mice.
This figure now includes amplified images of representative spermatocytes and a summary schematic illustrating the timeline of spermatogenesis. In addition, it now presents the statistical analysis of spermatocyte quantification to support the visual data.
__Figure 2.____ Cilia emerge across all stages of prophase I in spermatocytes during the first spermatogenesis wave. __
The images of this figure remain unchanged from the original submission, but all the graphs present now the statistical analysis of spermatocyte quantification.
Figure 3. Ultrastructure and markers of prepuberal meiotic cilia.
This figure remains unchanged from the original submission; however, we have replaced the ARL3-labelled spermatocyte image (A) with one displaying a clearer and more representative signal.
__Figure 4. Testicular tissue presents spermatocyte cysts in prepuberal mice and adult humans. __
This figure remains unchanged from the original submission.
__Figure 5. Cilia and flagella dynamics are correlated during prepuberal meiosis. __
This figure remains unchanged from the original submission.
__Figure 6. Comparative proteomics identifies potential regulators of ciliogenesis and flagellogenesis. __
This figure remains unchanged from the original submission.
Figure 7.____ Deciliation induces persistence of DNA damage in meiosis.
This figure has been substantially revised and now includes additional experiments analyzing chloral hydrate treatment, aimed at more accurately assessing DNA damage under both control and treated conditions. Images F-I and graph J are new.
Figure 8____. Aurora kinase A is a regulator of cilia disassembly in meiosis.
This figure is remodelled as the original version contained a mistake in previous panel II, for this, graph in new Fig.8 I has been corrected. In addition, it now contains additional data of αTubulin staining in arrested ciliated metaphases I after AURKA inhibition (new panel L1´).
__Figure 9. Schematic representation of the prepuberal versus adult seminiferous epithelium. __
This figure remains unchanged from the original submission.
__Supplementary Figure 1. Meiotic stages during the first meiotic wave. __
This figure remains unchanged from the original submission.
__Supplementary Figure 2 (new)____. __
This is a new figure that includes additional data requested by the reviewers. It includes additional markers of cilia in spermatocytes (glutamylated Tubulin/GT335), and the control data of cilia markers in non-ciliated spermatocytes. It also includes now the separated quantification of ciliated spermatocytes for each stage, as requested by reviewers, complementing graphs included in Figure 2.
Please note that with the inclusion of this new Supplementary Figure 2, the numbering of subsequent supplementary figures has been updated accordingly.
Supplementary Figure 3 (previously Suppl. Fig. 2)__. Ultrastructure of prophase I spermatocytes. __
This figure is equal in content to the original submission, but some annotations have been included.
Supplementary Figure 4 (previously Suppl. Fig. 3).__ Meiotic centrosome under the electron microscope. __
This figure remains unchanged from the original submission, but additional annotations have been included.
Supplementary Figure 5 (previously Suppl. Fig. 4)__. Human testis contains ciliated spermatocytes. __
This figure has been revised and now includes additional H2AX staining to better determine the stage of ciliated spermatocytes and improve their identification.
Supplementary Figure 6 (previously Suppl. Fig. 5). GLI1 and GLI3 readouts of Hedgehog signalling are not visibly affected in prepuberal mouse testes.
This figure has been remodeled and now includes the quantification of GLI1 and GLI3 and its corresponding statistical analysis. It also includes the control data for Tubulin, instead of GADPH.
Supplementary Figure 7 (previously Suppl. Fig. 6)__. CH and MLN8237 optimization protocol. __
This figure has been remodeled to incorporate control experiments using 1-hour organotypic culture treatment.
Supplementary Figure 8 (previously Suppl. Fig. 7)__. Tracking first meiosis wave with EdU pulse injection during prepubertal meiosis. __This figure remains unchanged from the original submission.
Supplementary Figure 9 (previously Suppl. Fig. 8)__. PLK1 and AURKA inhibition in cultured spermatocytes. __
This figure has been remodeled and now includes additional data on spindle detection in control and AURKA-inhibited spermatocytes (both ciliated and non ciliated).
DETAILED POINT-BY-POINT RESPONSE TO THE REVIEWERS
We will submit both the PDF version of the revised manuscript and the Word file with tracked changes relative to the original submission. Each modification made in response to reviewers' suggestions is annotated in the Word document within the corresponding section of the text. all new figures have also been uploaded to the system.
Response to the Referee #1
In this manuscript by Perez-Moreno et al., titled "The dynamics of ciliogenesis in prepubertal mouse meiosis reveal new clues about testicular maturation during puberty", the authors characterize the development of primary cilia during meiosis in juvenile male mice. The authors catalog a variety of testicular changes that occur as juvenile mice age, such as changes in testis weight and germ cell-type composition. They next show that meiotic prophase cells initially lack cilia, and ciliated meiotic prophase cells are detected after 20 days postpartum, coinciding with the time when post-meiotic spermatids within the developing testes acquire flagella. They describe that germ cells in juvenile mice harbor cilia at all substages of meiotic prophase, in contrast to adults where only zygotene stage meiotic cells harbor cilia. The authors also document that cilia in juvenile mice are longer than those in adults. They characterize cilia composition and structure by immunofluorescence and EM, highlighting that cilia polymerization may initially begin inside the cell, followed by extension beyond the cell membrane. Additionally, they demonstrate ciliated cells can be detected in adult human testes. The authors next perform proteomic analyses of whole testes from juvenile mice at multiple ages, which may not provide direct information about the extremely small numbers of ciliated meiotic cells in the testis, and is lacking follow up experiments, but does serve as a valuable resource for the community. Finally, the authors use a seminiferous tubule culturing system to show that chemical inhibition of Aurora kinase A likely inhibits cilia depolymerization upon meiotic prophase I exit and leads to an accumulation of metaphase-like cells harboring cilia. They also assess meiotic recombination progression using their culturing system, but this is less convincing.
Author response: We sincerely thank Ref #1 for the thorough and thoughtful evaluation of our manuscript. We are particularly grateful for the reviewer's careful reading and constructive feedback, which have helped us refine several sections of the text and strengthen our discussion. All comments and suggestions have been carefully considered and addressed, as detailed below.
__Major comments: __
-
There are a few issues with the experimental set up for assessing the effects of cilia depolymerization on DNA repair (Figure 7-II). First, how were mid pachytene cells identified and differentiated from early pachytene cells (which would have higher levels of gH2AX) in this experiment? I suggest either using H1t staining (to differentiate early/mid vs late pachytene) or the extent of sex chromosome synapsis. This would ensure that the authors are comparing similarly staged cells in control and treated samples. Second, what were the gH2AX levels at the starting point of this experiment? A more convincing set up would be if the authors measure gH2AX immediately after culturing in early and late cells (early would have higher gH2AX, late would have lower gH2AX), and then again after 24hrs in late cells (upon repair disruption the sampled late cells would have high gH2AX). This would allow them to compare the decline in gH2AX (i.e., repair progression) in control vs treated samples. Also, it would be informative to know the starting gH2AX levels in ciliated vs non-ciliated cells as they may vary.
Response:
We thank Ref #1 for this valuable comment, which significantly contributed to improving both the design and interpretation of the cilia depolymerization assay.
Following this suggestion, we repeated the experiment including 1-hour (immediately after culturing), and 24-hour cultures for both control and chloral hydrate (CH)-treated samples (n = 3 biological replicates). To ensure accurate staging, we now employ triple immunolabelling for γH2AX, SYCP3, and H1T, allowing clear distinction of zygotene (H1T−), early pachytene (H1T−), and late pachytene (H1T+) cells. The revised data (Figure 7) now provide a more complete and statistically robust analysis of DNA damage dynamics. These results confirm that CH-induced deciliation leads to persistence of the γH2AX signal at 24 hours, indicating impaired DNA repair progression in pachytene spermatocytes. The new images and graphs are included in the revised Figure 7.
Regarding the reviewer's final point about the comparison of γH2AX levels between ciliated and non-ciliated cells, we regret that direct comparison of γH2AX levels between ciliated and non-ciliated cells is not technically feasible. To preserve cilia integrity, all cilia-related imaging is performed using the squash technique, which maintains the three-dimensional structure of the cilia but does not allow reliable quantification of DNA damage markers due to nuclear distortion. Conversely, the nuclear spreading technique, used for DNA damage assessment, provides optimal visualization of repair foci but results in the loss of cilia due to cytoplasmic disruption during the hypotonic step. Given that spermatocytes in juvenile testes form developmentally synchronized cytoplasmic cysts, we consider that analyzing a statistically representative number of spermatocytes offers a valid and biologically meaningful measure of tissue-level effects.
In conclusion, we believe that the additional experiments and clarifications included in revised Figure 7 strengthen our conclusion that cilia depolymerization compromises DNA repair during meiosis. Further functional confirmation will be pursued in future works, since we are currently generating a conditional genetic model for a ciliopathy in our laboratory.
The authors analyze meiotic progression in cells cultured with/without AURKA inhibition in Figure 8-III and conclude that the distribution of prophase I cells does not change upon treatment. Is Figure 8-III A and B the same data? The legend text is incorrect, so it's hard to follow. Figure 8-III A shows a depletion of EdU-labelled pachytene cells upon treatment. Moreover, the conclusion that a higher proportion of ciliated zygotene cells upon treatment (Figure 8-II C) suggests that AURKA inhibition delays cilia depolymerization (page 13 line 444) does not make sense to me.
Response:
We thank Ref#1 for identifying this issue and for the careful examination of Figure 8. We discovered that the submitted version of Figure 8 contained a mismatch between the figure legend and the figure panels. The legend text was correct; however, the figure inadvertently included a non-corresponding graph (previously panel II-A), which actually belonged to Supplementary Figure 7 in the original submission. We apologize for this mistake.
This error has been corrected in the revised version. The updated Figure 8 now accurately presents the distribution of EdU-labelled spermatocytes across prophase I substages in control and AURKA-inhibited cultures (previously Figure 8-II B, now Figure 8-A). The corrected data show no significant differences in the proportions of EdU-labelled spermatocytes among prophase I substages after 24 hours of AURKA inhibition, confirming that meiotic progression is not delayed and that no accumulation of zygotene cells occurs under this treatment. Therefore, the observed increase in ciliated zygotene spermatocytes upon AURKA inhibition (new Figure 8 H-I) is best explained by a delay in cilia disassembly, rather than by an arrest or slowdown in meiotic progression. The figure legend and main text have been revised accordingly.
How do the authors know that there is a monopolar spindle in Figure 8-IV treated samples? Perhaps the authors can use a different Tubulin antibody (that does not detect only acetylated Tubulin) to show that there is a monopolar spindle.
Response:
We appreciate Ref#1 for this excellent suggestion. In the original submission (lines 446-447), we described that ciliated metaphase I spermatocytes in AURKA-inhibited samples exhibited monopolar spindle phenotypes. This description was based on previous reports showing that AURKA or PLK1 inhibition produces metaphases with monopolar spindles characterized by aberrant yet characteristic SYCP3 patterns, abnormal chromatin compaction, and circular bivalent alignment around non-migrated centrosomes (1). In our study, we observed SYCP3 staining consistent with these characteristic features of monopolar metaphases I.
However, we agree with Ref #1 that this could be better sustained with data. Following the reviewer's suggestion, we performed additional immunostaining using α-Tubulin, which labels total microtubules rather than only the acetylated fraction. For clarity purposes, the revised Figure 8 now includes α-Tubulin staining in the same ciliated metaphase I cells shown in the original submission, confirming the presence of defective microtubule polymerization and defective spindle organization. For clarity, we now refer to these ciliated metaphases I as "arrested MI". This new data further support our conclusion that AURKA inhibition disrupts spindle bipolarization and prevents cilia depolymerization, indicating that cilia maintenance and bipolar spindle organization are mechanistically incompatible events during male meiosis. The abstract, results, and discussion section has been expanded accordingly, emphasizing that the persistence of cilia may interfere with microtubule polymerization and centrosome separation under AURKA inhibition. The Discussion has been expanded to emphasize that persistence of cilia may interfere with centrosome separation and microtubule polymerization, contrasting with invertebrate systems -e.g. Drosophila (2) and P. brassicae (3)- in which meiotic cilia persist through metaphase I without impairing bipolar spindle assembly.
- Alfaro, et al. EMBO Rep 22, (2021). DOI: 15252/embr.202051030 (PMID: 33615693)
- Riparbelli et al . Dev Cell (2012) DOI: 1016/j.devcel.2012.05.024 (PMID: 22898783)
- Gottardo et al, Cytoskeleton (Hoboken) (2023) DOI: 1002/cm.21755 (PMID: 37036073)
The authors state in the abstract that they provide evidence suggesting that centrosome migration and cilia depolymerization are mutually exclusive events during meiosis. This is not convincing with the data present in the current manuscript. I suggest amending this statement in the abstract.
Response:
We thank Ref#1 for this valuable observation, with which we fully agree. To avoid overstatement, the original statement has been removed from the Abstract, Results, and Discussion, and replaced with a more accurate formulation indicating that cilia maintenance and bipolar spindle formation are mutually exclusive events during mouse meiosis.
This revised statement is now directly supported by the new data presented in Figure 8, which demonstrate that AURKA inhibition prevents both spindle bipolarization and cilia depolymerization. We are grateful to the reviewer for highlighting this important clarification.
Minor comments:
The presence of cilia in all stages of meiotic prophase I in juvenile mice is intriguing. Why is the cellular distribution and length of cilia different in prepubertal mice compared to adults (where shorter cilia are present only in zygotene cells)? What is the relevance of these developmental differences? Do cilia serve prophase I functions in juvenile mice (in leptotene, pachytene etc.) that are perhaps absent in adults?
Related to the above point, what is the relevance of the absence of cilia during the first meiotic wave? If cilia serve a critical function during prophase I (for instance, facilitating DSB repair), does the lack of cilia during the first wave imply differing cilia (and repair) requirements during the first vs latter spermatogenesis waves?
In my opinion, these would be interesting points to discuss in the discussion section.
Response:
We thank the reviewer for these thoughtful observations, which we agree are indeed intriguing.
We believe that our findings likely reflect a developmental role for primary cilia during testicular maturation. We hypothesize that primary cilia at this stage might act as signaling organelles, receiving cues from Sertoli cells or neighboring spermatocytes and transmitting them through the cytoplasmic cysts shared by spermatocytes. Such intercellular communication could be essential for coordinating tissue maturation and meiotic entry during puberty. Although speculative, this hypothesis aligns with the established role of primary cilia as sensory and signaling hubs for GPCR and RTK pathways regulating cell differentiation and developmental patterning in multiple tissues (e.g., 1, 2). The Discussion section has been expanded to include these considerations.
- Goetz et al, Nat Rev Genet (2010)- DOI: 1038/nrg2774 (PMID: 20395968)
- Naturky et al , Cell (2019) DOI: 1038/s41580-019-0116-4 (PMID: 30948801) Our study focuses on the first spermatogenic wave, which represents the transition from the juvenile to the reproductive phase. It is therefore plausible that the transient presence of longer cilia during this period reflects a developmental requirement for external signaling that becomes dispensable in the mature testis. Given that this is only the second study to date examining mammalian meiotic cilia, there remains a vast area of research to explore. We plan to address potential signaling cascades involved in these processes in future studies.
On the other hand, while we cannot confirm that the cilia observed in zygotene spermatocytes persist until pachytene within the same cell, it is reasonable to speculate that they do, serving as longer-lasting signaling structures that facilitate testicular development during the critical pubertal window. In addition, the observation of ciliated spermatocytes at all prophase I substages at 20 dpp, together with our proteomic data, supports the idea that the emergence of meiotic cilia exerts a significant developmental impact on testicular maturation.
In summary, although we cannot yet define specific prophase I functions for meiotic cilia in juvenile spermatocytes, our data demonstrate that the first meiotic wave differs from later waves in cilia dynamics, suggesting distinct regulatory requirements between puberty and adulthood. These findings underscore the importance of considering developmental context when using the first meiotic wave as a model for studying spermatogenesis.
The authors state on page 9 lines 286-288 that the presence of cytoplasmic continuity via intercellular bridges (between developmentally synchronous spermatocytes) hints towards a mechanism that links cilia and flagella formation. Please clarify this statement. While the correlation between the timing of appearance of cilia and flagella in cells that are located within the same segment of the seminiferous tubule may be hinting towards some shared regulation, how would cytoplasmic continuity participate in this regulation? Especially since the cytoplasmic continuity is not between the developmentally distinct cells acquiring the cilia and flagella?
Response:
We thank Ref#1 for this excellent question and for the opportunity to clarify our statement.
The presence of intercellular bridges between spermatocytes is well known and has long been proposed to support germ cell communication and synchronization (1,2) as well as sharing mRNA (3) and organelles (4). A classic example is the Akap gene, located on the X chromosome and essential for the formation of the sperm fibrous sheath; cytoplasmic continuity through intercellular bridges allows Akap-derived products to be shared between X- and Y-bearing spermatids, thereby maintaining phenotypic balance despite transcriptional asymmetry (5). In addition, more recent work has further demonstrated that these bridges are critical for synchronizing meiotic progression and for processes such as synapsis, double-strand break repair, and transposon repression (6).
In this context, and considering our proteomic data (Figure 6), our statement did not intend to imply direct cytoplasmic exchange between ciliated and flagellated cells. Although our current methods do not allow comprehensive tracing of cytoplasmic continuity from the basal to the luminal compartment of the seminiferous epithelium, we plan to address this limitation using high-resolution 3D and ultrastructural imaging approaches in future studies.
Based on our current data, we propose that cytoplasmic continuity within developmentally synchronized spermatocyte cysts could facilitate the coordinated regulation of ciliogenesis, and similarly enable the sharing of regulatory factors controlling flagellogenesis within spermatid cysts. This coordination may occur through the diffusion of centrosomal or ciliary proteins, mRNAs, or signaling intermediates involved in the regulation of microtubule dynamics. However, we cannot exclude the possibility that such cytoplasmic continuity extends across all spermatocytes derived from the same spermatogonial clone, potentially providing a larger regulatory network.]] This mechanism could help explain the temporal correlation we observe between the appearance of meiotic cilia and the onset of flagella formation in adjacent spermatids within the same seminiferous segment.
We have revised the Discussion to explicitly clarify this interpretation and to note that, although hypothetical, it is consistent with established literature on cytoplasmic continuity and germ cell coordination.
- Dym, et al. * Reprod.*(1971) DOI: 10.1093/biolreprod/4.2.195 (PMID: 4107186)
- Braun et al. Nature. (1989) DOI: 1038/337373a0 (PMID: 2911388)
- Greenbaum et al. * Natl. Acad. Sci. USA*(2006). DOI: 10.1073/pnas.0505123103 (PMID: 16549803)
- Ventelä et al. Mol Biol Cell. (2003) DOI: 1091/mbc.e02-10-0647 (PMID: 12857863)
- Turner et al. Journal of Biological Chemistry (1998). DOI: 1074/jbc.273.48.32135 (PMID: 9822690)
- Sorkin, et al. Nat Commun (2025). DOI: 1038/s41467-025-56742-9 (PMID: 39929837) *note: due to manuscript-length limitations, not all cited references can be included in the text; they are listed here to substantiate our response.
Individual germ cells in H&E-stained testis sections in Figure 1-II are difficult to see. I suggest adding zoomed-in images where spermatocytes/round spermatids/elongated spermatids are clearly distinguishable.
Response:
Ref#1 is very right in this suggestion. We have revised Figure 1 to improve the quality of the H&E-stained testis sections and have added zoomed-in panels where spermatocytes, round spermatids, and elongated spermatids are clearly distinguishable. These additions significantly enhance the clarity and interpretability of the figure.
In Figure 2-II B, the authors document that most ciliated spermatocytes in juvenile mice are pachytene. Is this because most meiotic cells are pachytene? Please clarify. If the data are available (perhaps could be adapted from Figure 1-III), it would be informative to see a graph representing what proportions of each meiotic prophase substages have cilia.
Response:
We thank the reviewer for this valuable observation. Indeed, the predominance of ciliated pachytene spermatocytes reflects the fact that most meiotic cells in juvenile testes are at the pachytene stage (Figure 1). We have clarified this point in the text and have added a new supplementary figure (Supplementary Figure 2, new figure) presenting a graph showing the proportion of spermatocytes at each prophase I substage that possess primary cilia. This visualization provides a clearer quantitative overview of ciliation dynamics across meiotic substages.
I suggest annotating the EM images in Sup Figure 2 and 3 to make it easier to interpret.
Response:
We thank the reviewer for this helpful suggestion. We have now added annotations to the EM images in Supplementary Figures 3 and 4 to facilitate their interpretation. These visual guides help readers more easily identify the relevant ultrastructural features described in the text.
The authors claim that the ratio between GLI3-FL and GLI3-R is stable across their analyzed developmental window in whole testis immunoblots shown in Sup Figure 5. Quantifying the bands and normalizing to the loading control would help strengthen this claim as it hard to interpret the immunoblot in its current form.
Response:
We thank the reviewer for this valuable suggestion. Following this recommendation, Supplementary Figure 5 has been revised to include quantification of GLI1 and GLI3 protein levels, normalized to the loading control.
After quantification, we observed statistically significant differences across developmental stages. Specifically, GLI1 expression is slightly higher at 21 dpp compared to 8 dpp. For GLI3, we performed two complementary analyses:
- Total GLI3 protein (sum of full-length and repressor forms normalized to loading control) shows a progressive decrease during development, with the lowest levels at 60 dpp (Supplementary Figure 5D).
- GLI3 activation status, assessed as the GLI3-FL/GLI3-R ratio, is highest during the 19-21 dpp window, compared to 8 dpp and 60 dpp. Although these results suggest a possible transient activation of GLI3 during testicular maturation, we caution that this cannot automatically be attributed to increased Hedgehog signaling, as GLI3 processing can also be affected by other processes, such as changes in ciliogenesis. Furthermore, because the analysis was performed on whole-testis protein extracts, these changes cannot be specifically assigned to ciliated spermatocytes.
We have expanded the Discussion to address these findings and to highlight the potential involvement of the Desert Hedgehog (DHH) pathway, which plays key roles in testicular development, Sertoli-germ cell communication, and spermatogenesis (1, 2, 3). We plan to investigate these pathways further in future studies.
- Bitgood et al. Curr Biol. (1996). DOI: 1016/s0960-9822(02)00480-3 (PMID: 8805249)
- Clark et al. Biol Reprod. (2000) DOI: 1095/biolreprod63.6.1825 (PMID: 11090455)
- O'Hara et al. BMC Dev Biol. (2011) DOI: 1186/1471-213X-11-72 (PMID: 22132805) *note: due to manuscript-length limitations, not all cited references can be included in the text; they are listed here to substantiate our response.
There are a few typos throughout the manuscript. Some examples: page 5 line 172, Figure 3-I legend text, Sup Figure 5-II callouts, Figure 8-III legend, page 15 line 508, page 17 line 580, page 18 line 611.
Response:
We thank the reviewer for detecting this. All typographical errors have been corrected, and figure callouts have been reviewed for consistency.
Response to the Referee #2
This study focuses on the dynamic changes of ciliogenesis during meiosis in prepubertal mice. It was found that primary cilia are not an intrinsic feature of the first wave of meiosis (initiating at 8 dpp); instead, they begin to polymerize at 20 dpp (after the completion of the first wave of meiosis) and are present in all stages of prophase I. Moreover, prepubertal cilia (with an average length of 21.96 μm) are significantly longer than adult cilia (10 μm). The emergence of cilia coincides temporally with flagellogenesis, suggesting a regulatory association in the formation of axonemes between the two. Functional experiments showed that disruption of cilia by chloral hydrate (CH) delays DNA repair, while the AURKA inhibitor (MLN8237) delays cilia disassembly, and centrosome migration and cilia depolymerization are mutually exclusive events. These findings represent the first detailed description of the spatiotemporal regulation and potential roles of cilia during early testicular maturation in mice. The discovery of this phenomenon is interesting; however, there are certain limitations in functional research.
We thank Referee #2 for their careful reading of the manuscript and for highlighting important limitations regarding functional interpretation.
Our primary objective in this study was to provide a rigorous structural, temporal, and developmental characterization of meiotic ciliogenesis in the mammalian testis, a process for which almost no prior data exist. Given this lack of foundational information, we focused on establishing when, where, and in which meiotic stages primary cilia form during prepubertal development, and on identifying candidate regulatory pathways using complementary imaging, proteomic, and pharmacological approaches.
We agree that genetic ablation models would provide the most direct means to test ciliary function during spermatogenesis. However, we believe that such functional analyses must be preceded by a detailed developmental and phenotypic framework, which was previously unavailable. The present study therefore represents a necessary first step, defining the dynamics, ultrastructure, and molecular context of meiotic cilia during the transition from juvenile to adult spermatogenesis. We are currently generating conditional genetic models to directly address functional mechanisms in future work.
Regarding the temporal coincidence between the emergence of meiotic cilia and the onset of flagellogenesis, we do not interpret this observation as evidence of stochastic or non-functional protein expression. Rather, we present it as a developmental correlation that may reflect shared regulatory constraints on axonemal assembly during testicular maturation. We have clarified in the revised manuscript that this relationship is descriptive and hypothesis-generating, and we avoid assigning direct causal roles.
With respect to the proteomic analysis, we agree that proteomics alone cannot establish function. Our intent was not to assign causality, but to provide a developmental, hypothesis-generating dataset identifying candidate regulators that are enriched at the precise developmental window when both meiotic cilia and spermatid flagella first emerge. We have revised the text to explicitly frame these data as a resource for future mechanistic studies, rather than as direct functional evidence.
Taken together, we believe that the revised manuscript now more accurately reflects the scope and limitations of the study, while providing a robust and much-needed developmental framework for future genetic and functional analyses of meiotic ciliogenesis in mammals. We would be happy to further clarify any aspect of these interpretations if the reviewer or editor considers it helpful.
Major points:
-
The prepubertal cilia in spermatocytes discovered by the authors lack specific genetic ablation to block their formation, making it impossible to evaluate whether such cilia truly have functions. Because neither in the first wave of spermatogenesis nor in adult spermatogenesis does this type of cilium seem to be essential. In addition, the authors also imply that the formation of such cilia appears to be synchronized with the formation of sperm flagella. This suggests that the production of such cilia may merely be transient protein expression noise rather than a functionally meaningful cellular structure.
Response:
We agree that a genetic ablation model would represent the ideal approach to directly test cilia function in spermatogenesis. However, given the complete absence of prior data describing the dynamics of ciliogenesis during testis development, our priority in this study was to establish a rigorous structural and temporal characterization of this process in the main mammalian model organism, the mouse. This systematic and rigorous phenotypic characterization is a necessary first step before any functional genetics could be meaningfully interpreted.
To our knowledge, this study represents the first comprehensive analysis of ciliogenesis during prepubertal mouse meiosis, extending our previous work on adult spermatogenesis (1). Beyond these two contributions, only four additional studies have addressed meiotic cilia-two in zebrafish (2, 3), with Mytlys et al. also providing preliminary observations relevant to prepubertal male meiosis that we discuss in the present work, one in Drosophila (4) and a recent one in butterfly (5). No additional information exists for mammalian gametogenesis to date.
- López-Jiménez et al. Cells (2022) DOI: 10.3390/cells12010142 (PMID: 36611937)
- Mytlis et al. Science (2022) DOI: 10.1126/science.abh3104 (PMID: 35549308)
- Xie et al. J Mol Cell Biol (2022) DOI: 10.1093/jmcb/mjac049 (PMID: 35981808)
- Riparbelli et al . Dev Cell (2012) DOI: 10.1016/j.devcel.2012.05.024 (PMID: 22898783)
- Gottardo et al, Cytoskeleton (Hoboken) (2023) DOI: 10.1002/cm.21755 (PMID: 37036073) We therefore consider this descriptive and analytical foundation to be essential before the development of functional genetic models. Indeed, we are currently generating a conditional genetic model for a ciliopathy in our laboratory. These studies are ongoing and will directly address the type of mechanistic questions raised here, but they extend well beyond the scope and feasible timeframe of the present manuscript.
We thus maintain that the present work constitutes a necessary and timely contribution, providing a robust reference dataset that will facilitate and guide future functional studies in the field of cilia and meiosis.
Taking this into account, we would be very pleased to address any additional, concrete suggestions from Ref#2 that could further strengthen the current version of the manuscript
The high expression of axoneme assembly regulators such as TRiC complex and IFT proteins identified by proteomic analysis is not particularly significant. This time point is precisely the critical period for spermatids to assemble flagella, and TRiC, as a newly discovered component of flagellar axonemes, is reasonably highly expressed at this time. No intrinsic connection with the argument of this paper is observed. In fact, this testicular proteomics has little significance.
Response:
We appreciate this comment but respectfully disagree with the reviewer's interpretation of our proteomic data. To our knowledge, this is the first proteomic study explicitly focused on identifying ciliary regulators during testicular development at the precise window (19-21 dpp) when both meiotic cilia and spermatid flagella first emerge.
While Piprek et al (1) analyzed the expression of primary cilia in developing gonads, proteomic data specifically covering the developmental transition at 19-21 dpp were not previously available. Furthermore, a recent cell-sorting study (2), detected expression of cilia proteins in pachytene spermatocytes compared to round spermatids, but did not explore their functional relevance or integrate these data with developmental timing or histological context.
In contrast, our dataset integrates histological staging, high-resolution microscopy, and quantitative proteomics, revealing a set of candidate regulators (including DCAF7, DYRK1A, TUBB3, TUBB4B, and TRiC) potentially involved in cilia-flagella coordination. We view this as a hypothesis-generating resource that outlines specific proteins and pathways for future mechanistic studies on both ciliogenesis and flagellogenesis in the testis.
Although we fully agree that proteomics alone cannot establish causal function, we believe that dismissing these data as having little significance overlooks their value as the first molecular map of the testis at the developmental window when axonemal structures arise. Our dataset provides, for the first time, an integrated view of proteins associated with ciliary and flagellar structures at the developmental stage when both axonemal organelles first appear. We thus believe that our proteomic dataset represents an important and novel contribution to the understanding of testicular development and ciliary biology.
Considering this, we would again welcome any specific suggestions from Ref#2 on additional analyses or clarifications that could make the relevance of this dataset even clearer to readers.
- Piprek et al. Int J Dev Biol. (2019) doi: 10.1387/ijdb.190049rp (PMID: 32149371).
- Fang et al. Chromosoma. (1981) doi: 10.1007/BF00285768 (PMID: 7227045). Response to the Referee #3
In "The dynamics of ciliogenesis in prepubertal mouse meiosis reveals new clues about testicular development" Pérez-Moreno, et al. explore primary cilia in prepubertal mouse spermatocytes. Using a combination of microscopy, proteomics, and pharmacological perturbations, the authors carefully characterize prepubertal spermatocyte cilia, providing foundational work regarding meiotic cilia in the developing mammalian testis.
Response: We sincerely thank Ref#3 for their positive assessment of our work and for the thoughtful suggestions that have helped us strengthen the manuscript. We are pleased that the reviewer recognizes both the novelty and the relevance of our study in providing foundational insights into meiotic ciliogenesis during prepubertal testicular development. All specific comments have been carefully considered and addressed as detailed below.
Major concerns:
-
The authors provide evidence consistent with cilia not being present in a larger percentage of spermatocytes or in other cells in the testis. The combination of electron microscopy and acetylated tubulin antibody staining establishes the presence of cilia; however, proving a negative is challenging. While acetylated tubulin is certainly a common marker of cilia, it is not in some cilia such as those in neurons. The authors should use at least one additional cilia marker to better support their claim of cilia being absent.
Response:
We thank the reviewer for this helpful suggestion. In the revised version, we have strengthened the evidence for cilia identification by including an additional ciliary marker, glutamylated tubulin (GT335), in combination with acetylated tubulin and ARL13B (which were included in the original submission). These data are now presented in the new Supplementary Figure 2, which also includes an example of a non-ciliated spermatocyte showing absence of both ARL13B and AcTub signals.
Taken together, these markers provide a more comprehensive validation of cilia detection and confirm the absence of ciliary labelling in non-ciliated spermatocytes.
The conclusion that IFT88 localizes to centrosomes is premature as key controls for the IFT88 antibody staining are lacking. Centrosomes are notoriously "sticky", often sowing non-specific antibody staining. The authors must include controls to demonstrate the specificity of the staining they observe such as staining in a genetic mutant or an antigen competition assay.
Response:
We appreciate the reviewer's concern and fully agree that antibody specificity is critical when interpreting centrosomal localization. The IFT88 antibody used in our study is commercially available and has been extensively validated in the literature as both a cilia marker (1, 2), and a centrosome marker in somatic cells (3). Labelling of IFT88 in centrosomes has also been previously described using other antibodies (4, 5). In our material, the IFT88 signal consistently appears at one of the duplicated centrosomes and at both spindle poles-patterns identical to those reported in somatic cells. We therefore consider the reported meiotic IFT88 staining as specific and biologically reliable.
That said, we agree that genetic validation would provide the most definitive confirmation. We would like to inform that we are currently since we are currently generating a conditional genetic model for a ciliopathy in our laboratory that will directly assess both antibody specificity and functional consequences of cilia loss during meiosis. These experiments are in progress and will be reported in a follow-up study.
- Wong et al. Science (2015). DOI: 1126/science.aaa5111 (PMID: 25931445)
- Ocbina et al. Nat Genet (2011). DOI: 1038/ng.832 (PMID: 21552265)
- Vitre et al. EMBO Rep (2020). DOI: 15252/embr.201949234 (PMID: 32270908)
- Robert A. et al. J Cell Sci (2007). DOI: 1242/jcs.03366 (PMID: 17264151)
- Singla et al, Developmental Cell (2010). DOI: 10.1016/j.devcel.2009.12.022 (PMID: 20230748) *note: due to manuscript-length limitations, not all cited references can be included in the text; they are listed here to substantiate our response.
There are many inconsistent statements throughout the paper regarding the timing of the first wave of spermatogenesis. For example, the authors state that round spermatids can be detected at 21dpp on line 161, but on line 180, say round spermatids can be detected a 19dpp. Not only does this lead to confusion, but such discrepancies undermine the validity of the rest of the paper. A summary graphic displaying key events and their timing in the first wave of spermatogenesis would be instrumental for reader comprehension and could be used by the authors to ensure consistent claims throughout the paper.
Response:
We thank the reviewer for identifying this inconsistency and apologize for the confusion. We confirm that early round spermatids first appear at 19 dpp, as shown in the quantitative data (Figure 1J). This can be detected in squashed spermatocyte preparations, where individual spermatocytes and spermatids can be accurately quantified. The original text contained an imprecise reference to the histological image of 21 dpp (previous line 161), since certain H&E sections did not clearly show all cell types simultaneously. However, we have now revised Figure 1, improving the image quality and adding a zoomed-in panel highlighting early round spermatids. Image for 19 dpp mice in Fig 1D shows early, yet still aflagellated spermatids. The first ciliated spermatocytes and the earliest flagellated spermatids are observed at 20 dpp. This has been clarified in the text.
In addition, we also thank the reviewer for the suggestion of adding a summary graphic, which we agree greatly facilitates reader comprehension. We have added a new schematic summary (Figure 1K) illustrating the key stages and timing of the first spermatogenic wave.
In the proteomics experiments, it is unclear why the authors assume that changes in protein expression are predominantly due to changes within the germ cells in the developing testis. The analysis is on whole testes including both the somatic and germ cells, which makes it possible that protein expression changes in somatic cells drive the results. The authors need to justify why and how the conclusions drawn from this analysis warrant such an assumption.
Response:
We agree with the reviewer that our proteomic analysis was performed on whole testis samples, which contain both germ and somatic cells. Although isolation of pure spermatocyte populations by FACS would provide higher resolution, obtaining sufficient prepubertal material for such analysis would require an extremely large number of animals. To remain compliant with the 3Rs principle for animal experimentation, we therefore used whole-testis samples from three biological replicates per age.
We acknowledge that our assumption-that the main differences arise from germ cells-is a simplification. However, germ cells constitute the vast majority of testicular cells during this developmental window and are the population undergoing major compositional changes between 15 dpp and adulthood. It is therefore reasonable to expect that a substantial fraction of the observed proteomic changes reflects alterations in germ cells. We have clarified this point in the revised text and have added a statement noting that changes in somatic cells could also contribute to the proteomic profiles.
The authors should provide details on how proteins were categorized as being involved in ciliogenesis or flagellogenesis, specifically in the distinction criteria. It is not clear how the categorizations were determined or whether they are valid. Thus, no one can repeat this analysis or perform this analysis on other datasets they might want to compare.
Response:
We thank the reviewer for this opportunity to clarify our approach. The categorization of protein as being involved in ciliogenesis or flagellogenesis was based on their Gene Ontology (GO) cellular component annotations obtained from the PANTHER database (Version 19.0), using the gene IDs of the Differentially Expressed Proteins (DEPs). Specifically, we used the GO terms cilium (GO:0005929) and motile cilium (GO:0031514). Since motile cilium is a subcategory of cilium, proteins annotated only with the general cilium term, but not included under motile cilium, were considered to be associated with primary cilia or with shared structural components common to different types of cilia. These GO terms are represented in the bottom panel of the Figure 6.
This information has been added to the Methods section and referenced in the Results for transparency and reproducibility.
In the pharmacological studies, the authors conclude that the phenotypes they observe (DNA damage and reduced pachytene spermatocytes) are due to loss of or persistence of cilia. This overinterprets the experiment. Chloral hydrate and MLN8237 certainly impact ciliation as claimed, but have additional cellular effects. Thus, it is possible that the observed phenotypes were not a direct result of cilia manipulation. Either additional controls must address this or the conclusions need to be more specific and toned down.
Response:
We thank the reviewer for this fair observation and have taken steps to strengthen and refine our interpretation. In the revised version, we now include data from 1-hour and 24-hour cultures for both control and chloral hydrate (CH)-treated samples (n = 3 biological replicates). The triple immunolabelling with γH2AX, SYCP3, and H1T allows accurate staging of zygotene (H1T⁻), early pachytene (H1T⁻), and late pachytene (H1T⁺) spermatocytes.
The revised Figure 7 now provides a more complete and statistically supported analysis of DNA damage dynamics, confirming that CH-induced deciliation leads to persistent γH2AX signal at 24 hours, indicative of delayed or defective DNA repair progression. We have also toned down our interpretation in the Discussion, acknowledging that CH could affect other cellular pathways.
As mentioned before, the conditional genetic model that we are currently generating will allow us to evaluate the role of cilia in meiotic DNA repair in a more direct and specific way.
Assuming the conclusions of the pharmacological studies hold true with the proper controls, the authors still conflate their findings with meiotic defects. Meiosis is not directly assayed, which makes this conclusion an overstatement of the data. The conclusions need to be rephrased to accurately reflect the data.
Response:
We agree that this aspect required clarification. As noted above, we have refined both the Results and Discussion sections to make clear that our assays specifically targeted meiotic spermatocytes.
We now present data for meiotic stages at zygotene, early pachytene and late pachytene. This is demonstrated with the labelling for SYCP3 and H1T, both specific marker for meiosis that are not detectable in non meiotic cells. We believe that this is indeed a way to assay the meiotic cells, however, we have specified now in the text that we are analysing potential defects in meiosis progression. We are sorry if this was not properly explained in the original manuscript: it is now rephrased in the new version both in the results and discussion section.
It is not clear why the authors chose not to use widely accepted assays of Hedgehog signaling. Traditionally, pathway activation is measured by transcriptional output, not GLI protein expression because transcription factor expression does not necessarily reflect transcription levels of target genes.
Response:
We agree with the reviewer that measuring mRNA levels of Hedgehog pathway target genes, typically GLI1 and PTCH1, is the most common method for measuring pathway activation, and is widely accepted by researchers in the field. However, the methods we use in this manuscript (GLI1 and GLI3 immunoblots) are also quite common and widely accepted:
Regarding GLI1 immunoblot, many articles have used this method to monitor Hedgehog signaling, since GLI1 protein levels have repeatedly been shown to also go up upon pathway activation, and down upon pathway inhibition, mirroring the behavior of GLI1 mRNA. Here are a few publications that exemplify this point:
- Banday et al. 2025 Nat Commun. DOI: 10.1038/s41467-025-56632-0 (PMID: 39894896)
- Shi et al 2022 JCI Insight DOI: 10.1172/jci.insight.149626 (PMID: 35041619)
- Deng et al. 2019 eLife, DOI: 10.7554/eLife.50208 (PMID: 31482846)
- Zhu et al. 2019 Nat Commun, DOI: 10.1038/s41467-019-10739-3 (PMID: 31253779)
- Caparros-Martin et al 2013 Hum Mol Genet, DOI: 10.1093/hmg/dds409 (PMID: 23026747) *note: due to manuscript-length limitations, not all cited references can be included in the text; they are listed here to substantiate our response.
As for GLI3 immunoblot, Hedgehog pathway activation is well known to inhibit GLI3 proteolytic processing from its full length form (GLI3-FL) to its transcriptional repressor (GLI3-R), and such processing is also commonly used to monitor Hedgehog signal transduction, of which the following are but a few examples:
- Pedraza et al 2025 eLife, DOI: 10.7554/eLife.100328 (PMID: 40956303)
- Somatilaka et al 2020 Dev Cell, DOI: 10.1016/j.devcel.2020.06.034 (PMID: 32702291)
- Infante et al 2018, Nat Commun, DOI: 10.1038/s41467-018-03339-0 (PMID: 29515120)
- Wang et al 2017 Dev Biol DOI: 10.1016/j.ydbio.2017.08.003 (PMID: 28800946)
- Singh et al 2015 J Biol Chem DOI: 10.1074/jbc.M115.665810 (PMID: 26451044) *note: due to manuscript-length limitations, not all cited references can be included in the text; they are listed here to substantiate our response.
In summary, we think that we have used two well established markers to look at Hedgehog signaling (three, if we include the immunofluorescence analysis of SMO, which we could not detect in meiotic cilia).
These Hh pathway analyses did not provide any convincing evidence that the prepubertal cilia we describe here are actively involved in this pathway, even though Hh signaling is cilia-dependent and is known to be active in the male germline (Sahin et al 2014 Andrology PMID: 24574096; Mäkelä et al 2011 Reproduction PMID: 21893610; Bitgood et al 1996 Curr Biol. PMID: 8805249).
That said, we fully agree that our current analyses do not allow us to draw definitive conclusions regarding Hedgehog pathway activity in meiotic cilia, and we now state this explicitly in the revised Discussion.
Also in the Hedgehog pathway experiment, it is confusing that the authors report no detection of SMO yet detect little to no expression of GLIR in their western blot. Undetectable SMO indicates Hedgehog signaling is inactive, which results in high levels of GLIR. The impact of this is that it is not clear what is going on with Hh signaling in this system.
Response:
It is true that, when Hh signaling is inactive (and hence SMO not ciliary), the GLI3FL/GLI3R ratio tends to be low.
Although our data in prepuberal mouse testes show a strong reduction in total GLI3 protein levels (GLI3FL+GLI3R) as these mice grow older, this downregulation of total GLI3 occurs without any major changes in the GLI3FL/GLI3R ratio, which is only modestly affected (suppl. Figure 6).
Hence, since it is the ratio that correlates with Hh signaling rather than total levels, we do not think that the GLI3R reduction we see is incompatible with our non-detection of SMO in cilia: it seems more likely that overall GLI3 expression is being downregulated in developing testes via a Hh-independent mechanism.
Also potentially relevant here is the fact that some cell types depend more on GLI2 than on GLI3 for Hh signaling. For instance, in mouse embryos, Hh-mediated neural tube patterning relies more heavily on GLI2 processing into a transcriptional activator than on the inhibition of GLI3 processing into a repressor. In contrast, the opposite is true during Hh-mediated limb bud patterning (Nieuwenhuis and Hui 2005 Clin Genet. PMID: 15691355). We have not looked at GLI2, but it is conceivable that it could play a bigger role than GLI3 in our model.
Moreover, several forms of GLI-independent non-canonical Hh signaling have been described, and they could potentially play a role in our model, too (Robbins et al 2012 Sci Signal. PMID: 23074268).
We have revised the discussion to clarify some of these points.
All in all, we agree that our findings regarding Hh signaling are not conclusive, but we still think they add important pieces to the puzzle that will help guide future studies.
There are multiple instances where it is not clear whether the authors performed statistical analysis on their data, specifically when comparing the percent composition of a population. The authors need to include appropriate statistical tests to make claims regarding this data. While the authors state some impressive sample sizes, once evaluated in individual categories (eg specific cell type and age) the sample sizes of evaluated cilia are as low as 15, which is likely underpowered. The authors need to state the n for each analysis in the figures or legends.
We thank the reviewer for highlighting this important issue. We have now included the sample size (n) for every analysis directly in the figure legends. Although this adds length, it improves transparency and reproducibility.
Regarding the doubts of Ref#3 about the different sample sizes, the number of spermatocytes quantified in each stage is in agreement with their distribution in meiosis (example, pachytene lasts for 10 days this stage is widely represented in the preparations, while its is much difficult to quantify metaphases I that are less present because the stage itself lasts for less than 24hours). Taking this into account, we ensured that all analyses remain statistically valid and representative, applying the appropriate statistical tests for each dataset. These details are now clearly indicated in the revised figures and legends.
Minor concerns:
-
The phrase "lactating male" is used throughout the paper and is not correct. We assume this term to mean male pups that have yet to be weaned from their lactating mother, but "lactating male" suggests a rare disorder requiring medical intervention. Perhaps "pre-weaning males" is what the authors meant.
Response:
We thank the reviewer for noticing this terminology error. The expression has been corrected to "pre-weaning males" throughout the manuscript.
The convention used to label the figures in this paper is confusing and difficult to read as there are multiple panels with the same letter in the same figure (albeit distinct sections). Labeling panels in the standard A-Z format is preferred. "Panel Z" is easier to identify than "panel III-E".
Response:
We thank the reviewer for this suggestion. All figures have been relabelled using the standard A-Z panel format, ensuring consistency and easier readability across the manuscript.
-
-
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Referee #3
Evidence, reproducibility and clarity
Summary:
In "The dynamics of ciliogenesis in prepubertal mouse meiosis reveals new clues about testicular development" Pérez-Moreno, et al. explore primary cilia in prepubertal mouse spermatocytes. Using a combination of microscopy, proteomics, and pharmacological perturbations, the authors carefully characterize prepubertal spermatocyte cilia, providing foundational work regarding meiotic cilia in the developing mammalian testis.
Major concerns:
-
The authors provide evidence consistent with cilia not being present in a larger percentage of spermatocytes or in other cells in the testis. The combination of electron microscopy and acetylated tubulin antibody staining establishes the presence of cilia; however, proving a negative is challenging. While acetylated tubulin is certainly a common marker of cilia, it is not in some cilia such as those in neurons. The authors should use at least one additional cilia marker to better support their claim of cilia being absent.
-
The conclusion that IFT88 localizes to centrosomes is premature as key controls for the IFT88 antibody staining are lacking. Centrosomes are notoriously "sticky", often sowing non-specific antibody staining. The authors must include controls to demonstrate the specificity of the staining they observe such as staining in a genetic mutant or an antigen competition assay.
-
There are many inconsistent statements throughout the paper regarding the timing of the first wave of spermatogenesis. For example, the authors state that round spermatids can be detected at 21dpp on line 161, but on line 180, say round spermatids can be detected a 19dpp. Not only does this lead to confusion, but such discrepancies undermine the validity of the rest of the paper. A summary graphic displaying key events and their timing in the first wave of spermatogenesis would be instrumental for reader comprehension and could be used by the authors to ensure consistent claims throughout the paper.
-
In the proteomics experiments, it is unclear why the authors assume that changes in protein expression are predominantly due to changes within the germ cells in the developing testis. The analysis is on whole testes including both the somatic and germ cells, which makes it possible that protein expression changes in somatic cells drive the results. The authors need to justify why and how the conclusions drawn from this analysis warrant such an assumption.
-
The authors should provide details on how proteins were categorized as being involved in ciliogenesis or flagellogenesis, specifically in the distinction criteria. It is not clear how the categorizations were determined or whether they are valid. Thus, no one can repeat this analysis or perform this analysis on other datasets they might want to compare.
-
In the pharmacological studies, the authors conclude that the phenotypes they observe (DNA damage and reduced pachytene spermatocytes) are due to loss of or persistence of cilia. This overinterprets the experiment. Chloral hydrate and MLN8237 certainly impact ciliation as claimed, but have additional cellular effects. Thus, it is possible that the observed phenotypes were not a direct result of cilia manipulation. Either additional controls must address this or the conclusions need to be more specific and toned down.
-
Assuming the conclusions of the pharmacological studies hold true with the proper controls, the authors still conflate their findings with meiotic defects. Meiosis is not directly assayed, which makes this conclusion an overstatement of the data. The conclusions need to be rephrased to accurately reflect the data.
-
It is not clear why the authors chose not to use widely accepted assays of Hedgehog signaling. Traditionally, pathway activation is measured by transcriptional output, not GLI protein expression because transcription factor expression does not necessarily reflect transcription levels of target genes.
-
Also in the Hedgehog pathway experiment, it is confusing that the authors report no detection of SMO yet detect little to no expression of GLIR in their western blot. Undetectable SMO indicates Hedgehog signaling is inactive, which results in high levels of GLIR. The impact of this is that it is not clear what is going on with Hh signaling in this system.
-
There are multiple instances where it is not clear whether the authors performed statistical analysis on their data, specifically when comparing the percent composition of a population. The authors need to include appropriate statistical tests to make claims regarding this data. While the authors state some impressive sample sizes, once evaluated in individual categories (eg specific cell type and age) the sample sizes of evaluated cilia are as low as 15, which is likely underpowered. The authors need to state the n for each analysis in the figures or legends.
Minor concerns:
-
The phrase "lactating male" is used throughout the paper and is not correct. We assume this term to mean male pups that have yet to be weaned from their lactating mother, but "lactating male" suggests a rare disorder requiring medical intervention. Perhaps "pre-weaning males" is what the authors meant.
-
The convention used to label the figures in this paper is confusing and difficult to read as there are multiple panels with the same letter in the same figure (albeit distinct sections). Labeling panels in the standard A-Z format is preferred. "Panel Z" is easier to identify than "panel III-E".
Significance
Overall, this is a well-done body of work that deserves recognition for the novel and implicative discoveries it presents. Assuming the conclusions hold true following appropriate statistical analysis and rephrasing, this paper would report the first documented evidence of meiotic cilia in the developing mammalian testis with sufficient rigor to become the foundational work on this topic.
This paper will be of interest to communities focused on germ cell development, cilia, and Hedgehog signaling. It may prompt a new perspective on Desert Hedgehog signaling as it pertains to spermatogenesis. Further, this work will be of interest to those studying male fertility, as it highlights the potential role of cilia in spermatogenesis.
Further, the proteomic analysis presented has the potential to invoke hypotheses and experimentation investigating the role of several proteins with previously uncharacterized roles in ciliogenesis, flagellogenesis, and/or spermatogenesis. The finding that the onset of ciliogenesis and flagellogenesis appear to be temporally linked has the potential to prompt research regarding shared molecular mechanisms dictating axonemal formation. We believe this paper has the potential to have an impact in its respective field, underscored by the exquisite microscopy and detailed characterization of meiotic cilia.
-
-
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Referee #1
Evidence, reproducibility and clarity
In this manuscript by Perez-Moreno et al., titled "The dynamics of ciliogenesis in prepubertal mouse meiosis reveal new clues about testicular maturation during puberty", the authors characterize the development of primary cilia during meiosis in juvenile male mice. The authors catalog a variety of testicular changes that occur as juvenile mice age, such as changes in testis weight and germ cell-type composition. They next show that meiotic prophase cells initially lack cilia, and ciliated meiotic prophase cells are detected after 20 days postpartum, coinciding with the time when post-meiotic spermatids within the developing testes acquire flagella. They describe that germ cells in juvenile mice harbor cilia at all substages of meiotic prophase, in contrast to adults where only zygotene stage meiotic cells harbor cilia. The authors also document that cilia in juvenile mice are longer than those in adults. They characterize cilia composition and structure by immunofluorescence and EM, highlighting that cilia polymerization may initially begin inside the cell, followed by extension beyond the cell membrane. Additionally, they demonstrate ciliated cells can be detected in adult human testes. The authors next perform proteomic analyses of whole testes from juvenile mice at multiple ages, which may not provide direct information about the extremely small numbers of ciliated meiotic cells in the testis, and is lacking follow up experiments, but does serve as a valuable resource for the community. Finally, the authors use a seminiferous tubule culturing system to show that chemical inhibition of Aurora kinase A likely inhibits cilia depolymerization upon meiotic prophase I exit and leads to an accumulation of metaphase-like cells harboring cilia. They also assess meiotic recombination progression using their culturing system, but this is less convincing.
Few suggestions/comments are listed below:
Major comments
-
There are a few issues with the experimental set up for assessing the effects of cilia depolymerization on DNA repair (Figure 7-II). First, how were mid pachytene cells identified and differentiated from early pachytene cells (which would have higher levels of gH2AX) in this experiment? I suggest either using H1t staining (to differentiate early/mid vs late pachytene) or the extent of sex chromosome synapsis. This would ensure that the authors are comparing similarly staged cells in control and treated samples. Second, what were the gH2AX levels at the starting point of this experiment? A more convincing set up would be if the authors measure gH2AX immediately after culturing in early and late cells (early would have higher gH2AX, late would have lower gH2AX), and then again after 24hrs in late cells (upon repair disruption the sampled late cells would have high gH2AX). This would allow them to compare the decline in gH2AX (i.e., repair progression) in control vs treated samples. Also, it would be informative to know the starting gH2AX levels in ciliated vs non-ciliated cells as they may vary.
-
The authors analyze meiotic progression in cells cultured with/without AURKA inhibition in Figure 8-III and conclude that the distribution of prophase I cells does not change upon treatment. Is Figure 8-III A and B the same data? The legend text is incorrect, so it's hard to follow. Figure 8-III A shows a depletion of EdU-labelled pachytene cells upon treatment. Moreover, the conclusion that a higher proportion of ciliated zygotene cells upon treatment (Figure 8-II C) suggests that AURKA inhibition delays cilia depolymerization (page 13 line 444) does not make sense to me.
-
How do the authors know that there is a monopolar spindle in Figure 8-IV treated samples? Perhaps the authors can use a different Tubulin antibody (that does not detect only acetylated Tubulin) to show that there is a monopolar spindle.
-
The authors state in the abstract that they provide evidence suggesting that centrosome migration and cilia depolymerization are mutually exclusive events during meiosis. This is not convincing with the data present in the current manuscript. I suggest amending this statement in the abstract.
Minor comments
- The presence of cilia in all stages of meiotic prophase I in juvenile mice is intriguing. Why is the cellular distribution and length of cilia different in prepubertal mice compared to adults (where shorter cilia are present only in zygotene cells)? What is the relevance of these developmental differences? Do cilia serve prophase I functions in juvenile mice (in leptotene, pachytene etc.) that are perhaps absent in adults?
Related to the above point, what is the relevance of the absence of cilia during the first meiotic wave? If cilia serve a critical function during prophase I (for instance, facilitating DSB repair), does the lack of cilia during the first wave imply differing cilia (and repair) requirements during the first vs latter spermatogenesis waves?
In my opinion, these would be interesting points to discuss in the discussion section.
-
The authors state on page 9 lines 286-288 that the presence of cytoplasmic continuity via intercellular bridges (between developmentally synchronous spermatocytes) hints towards a mechanism that links cilia and flagella formation. Please clarify this statement. While the correlation between the timing of appearance of cilia and flagella in cells that are located within the same segment of the seminiferous tubule may be hinting towards some shared regulation, how would cytoplasmic continuity participate in this regulation? Especially since the cytoplasmic continuity is not between the developmentally distinct cells acquiring the cilia and flagella?
-
Individual germ cells in H&E-stained testis sections in Figure 1-II are difficult to see. I suggest adding zoomed-in images where spermatocytes/round spermatids/elongated spermatids are clearly distinguishable.
-
In Figure 2-II B, the authors document that most ciliated spermatocytes in juvenile mice are pachytene. Is this because most meiotic cells are pachytene? Please clarify. If the data are available (perhaps could be adapted from Figure 1-III), it would be informative to see a graph representing what proportions of each meiotic prophase substages have cilia.
-
I suggest annotating the EM images in Sup Figure 2 and 3 to make it easier to interpret.
-
The authors claim that the ratio between GLI3-FL and GLI3-R is stable across their analyzed developmental window in whole testis immunoblots shown in Sup Figure 5. Quantifying the bands and normalizing to the loading control would help strengthen this claim as it hard to interpret the immunoblot in its current form.
-
There are a few typos throughout the manuscript. Some examples: page 5 line 172, Figure 3-I legend text, Sup Figure 5-II callouts, Figure 8-III legend, page 15 line 508, page 17 line 580, page 18 line 611.
Significance
This work provides new information about an important but poorly understood cellular structure present in meiotic cells, the primary cilium. More generally, this work expands on our understanding of testis development in juvenile mice. The microscopy images presented here are beautiful. The work is mostly descriptive but lays the groundwork for future investigations. I believe that this study would of interest to the germ cell, meiosis, and spermatogenesis communities, and with a few modifications, is suitable for publication.
-
-
-
social-media-ethics-automation.github.io social-media-ethics-automation.github.io
-
6.5. Parasocial Relationships# Another phenomenon related to authenticity which is common on social media is the parasocial relationship. Parasocial relationships are when a viewer or follower of a public figure (that is, a celebrity) feel like they know the public figure, and may even feel a sort of friendship with them, but the public figure doesn’t know the viewer at all. Parasocial relationships are not a new phenomenon, but social media has increased our ability to form both sides of these bonds. As comedian Bo Burnham put it: “This awful D-list celebrity pressure I had experienced onstage has now been democratized.” Learn more about parasocial relationships: StrucciMovies: Fake Friends YouTube Series Sarah Z: How Fans Treat Creators 33 min
Parasocial relationships are really common on social media, especially with influencers and streamers who talk directly to the camera and share personal stories. It can feel authentic when creators clearly explain the limits of the relationship, like Mr. Rogers did by calling viewers “television friends.” But it can become inauthentic or harmful when creators encourage followers to believe the relationship is more mutual than it actually is.
-
Parasocial relationships are not a new phenomenon, but social media has increased our ability to form both sides of these bonds. As comedian Bo Burnham put it: “This awful D-list celebrity pressure I had experienced onstage has now been democratized.”
Parasocial relationship as a phenomenon has become quite widespread do to social media. I sure even without social media it would still have been prevalent, imagine a Queen or a president from the past who's life was well documented and shared to the public leading to many folk feeling as if they were closer to their leader. I wonder what negative effects this type of relationship could have even to a mild extent?
-
6.5. Parasocial Relationships# Another phenomenon related to authenticity which is common on social media is the parasocial relationship. Parasocial relationships are when a viewer or follower of a public figure (that is, a celebrity) feel like they know the public figure, and may even feel a sort of friendship with them, but the public figure doesn’t know the viewer at all. Parasocial relationships are not a new phenomenon, but social media has increased our ability to form both sides of these bonds. As comedian Bo Burnham put it: “This awful D-list celebrity pressure I had experienced onstage has now been democratized.” Learn more about parasocial relationships: StrucciMovies: Fake Friends YouTube Series Sarah Z: How Fans Treat Creators 33 min
The example of Mr. Rogers shows that parasocial relationship sare not automatically unethical or inauthentic. What seems important here is that he tried to clearly define the limits of the relationship, such as calling viewers “television friends” and explaining that visits were not possible. This transparency helped make the parasocial relationship feel more authentic, even though it was not a real two-way friendship.
-
In his television program, Mr. Rogers wanted all children to feel cared for and loved. To do this, he intentionally fostered a parasocial relationship with the children in his audience (he called them his “television friends”): I give an expression of care every day to each child, to help him realize that he is unique. I end the program by saying, “You’ve made this day a special day, by just your being you. There’s no person in the whole world like you, and I like you, just the way you are.” Fred Rogers requesting funds for PBS at the US Senate in 1969 Now, as children, I (Kyle) and my sister watched this program and felt the effects of what Fred Rogers was doing and had different responses to it. I asked my mom to help me send him a letter asking if he was real, to which I got a letter back explaining that he was indeed a real person:
The section on parasocial relationships helped me understand why social media makes creators feel like “friends,” even when the relationship is one-sided. The Mr. Rogers example shows that parasocial relationships aren’t automatically bad, but they can become unethical if the creator isn’t clear about the limits of the relationship. This made me think about influencers today who intentionally blur those boundaries to seem more relatable.
-
-
openstax.org openstax.org
-
For example, all vertebrate embryos, including humans, exhibit gill slits and tails at some point in their early development. These disappear in the adults of terrestrial groups but adult forms of aquatic groups such as fish and some amphibians maintain them. Great ape embryos, including humans, have a tail structure during their development that they lose when they are born.
This excerpt explains how embryology provides evidence for evolution. Similar structures in early embryos, like tails and gill slits, show that different species share common ancestors, even if those features are not present in adults.
-
Statements such as “organisms evolve in response to a change in an environment” are quite common, but such statements can lead to two types of misunderstandings.
This passage explains that there is often a misconception within the theory of evolution that individual organisms evolve intentionally. Firstly, evolution is a process that affects populations, not individuals. Secondly, evolution occurs by natural selection, where a change in the environment causes favorable phenotypes within a population for that environment to benefit from the change. Individuals with advantageous traits are then more likely to survive, reproduce, and pass those traits on. Therefore, neither the environment itself nor the individual is causing those changes intentionally.
-
As explained in Determining Evolutionary Relationships, when similar characteristics occur because of environmental constraints and not due to a close evolutionary relationship, it is an analogy or homoplasy. For example, insects use wings to fly like bats and birds, but the wing structure and embryonic origin are completely different. These are analogous structures (Figure 20.8). On the other side, the bird and bat wings are homologous because the bones are inherited from a common ancestor, while the wings themselves are analogous as they evolved independently.
This passage explains the difference between homologous and analogous traits in evolution. Similar characteristics in different organisms do not always indicate an evolutionary relationship. When similar characteristics arise from a common ancestor, they are called homologous structures. However, when similar characteristics arise due to environmental pressures, they are called analogous structures. A structure can be homologous in its origin but analogous in its function if the adaptation occurred independently. For example, in bird and bat wings, although the bones are homologous because they are inherited from a common ancestor, the wings themselves are analogous because the ability to fly evolved independently in each lineage.
-
-
social-media-ethics-automation.github.io social-media-ethics-automation.github.io
-
Early in the days of YouTube, one YouTube channel (lonelygirl15) started to release vlogs (video web logs) consisting of a girl in her room giving updates on the mundane dramas of her life. But as the channel continued posting videos and gaining popularity, viewers started to question if the events being told in the vlogs were true stories, or if they were fictional. Eventually, users discovered that it was a fictional show, and the girl giving the updates was an actress.
The lonelygirl15 case shows why authenticity is so important on social media. People felt upset not just because the story was fake, but because they trusted it as real and emotionally invested in it. This makes me realize that when platforms or creators blur the line between fiction and reality, it can easily break users’ trust, even if the content itself is entertaining.
-
As a rule, humans do not like to be duped. We like to know which kinds of signals to trust, and which to distrust. Being lulled into trusting a signal only to then have it revealed that the signal was untrustworthy is a shock to the system, unnerving and upsetting. People get angry when they find they have been duped. These reactions are even more heightened when we find we have been duped simply for someone else’s amusement at having done so.
This part is interesting because being untrustworhy proves everyone that the material is fake, they have no way to be identified, the sources might not be credible either, which angers me when something like this happens in my feed. What’s interesting here is how deeply our sense of trust is tied to predictability and reliability, when a signal we rely on turns out to be deceptive, it not only surprises us but also triggers a strong emotional reaction.
-
Authenticity is a concept we use to talk about connections and interactions when the way the connection is presented matches the reality of how it functions.
This definition elevates authenticity from "whether the content is true" to "whether the relationship is aligned": the type of interaction presented (friendly sharing, intimate confidences, genuine vulnerability) must be consistent with how it actually operates. In other words, authenticity is a kind of "contractual consistency"—what I think I'm getting is what I actually get. If an account uses "friend-like candor" to build intimacy, but is essentially just a marketing script run by a team, the problem isn't necessarily that it's a performance, but that it fails to make it clear to the audience what kind of relationship they are in, thus creating a gap between expectations and reality, and a feeling of being exploited. This also explains why some "performative" content (comedy accounts, role-playing) doesn't receive criticism: because its presentation and actual operation are consistent, and the audience knows what they are participating in.
-
As a rule, humans do not like to be duped.
This statement shifts the issue of "authenticity" from a moral judgment (you lied to me = you are bad) to a social mechanism (being deceived = a failure of the signaling system). In social interactions, people rely on various cues to determine who and what is trustworthy. When they discover they have been "manipulated" into believing something they shouldn't, they experience intense unease and anger, because this is not just a simple information error, but a threat to their judgment and sense of security. Cases like lonelygirl15 provoke a backlash not simply because the story is fake, but because the audience believed they were establishing a "real, intimate connection," only to discover that the connection had been disguised as something else from the very beginning.
-
Many users were upset that what they had been watching wasn’t authentic. That is, users believed the channel was presenting itself as true events about a real girl, and it wasn’t that at all. Though, even after users discovered it was fictional, the channel continued to grow in popularity.
Authenticity is what helps build trust between the poster and the audience. There isn't really any other way to do this other than maybe transparency but both tie together (I would argue). When you destroy that image that people had believed or built about you, it feels like a lie or deception which are not good associations to have.
-
Authenticity is a concept we use to talk about connections and interactions when the way the connection is presented matches the reality of how it functions. An authentic connection can be trusted because we know where we stand. An inauthentic connection offers a surprise because what is offered is not what we get. An inauthentic connection could be a good surprise, but usually, when people use the term ‘inauthentic’, they are indicating that the surprise was in some way problematic: someone was duped.
I would like to add that with the rise of AI and deepfakes, the term 'authenticity' has begun to lose its meaning as it becomes increasingly difficult to distinguish between real and fake. Also, with people exaggerating their lifestyle, it will also affect the viewers into thinking that they aren't living a happy life.
-
Jennifer Lawrence became a favorite of many when she tripped at the Oscars, and turned the moment into her persona as someone with a cool-girl, unpolished, unfiltered way about her.
This whole concept of authenticity as a form of intimacy is so interesting for the concept of Ethics of Care. In that framework, we can look at what responsibilities public figures have, but also what responsibilities we have for them. Yes, it is important for the public figure to be truthful, but also to entertain. So where is the line? And if we as the masses decide they didn't toe that line well enough, is it our responsibility to then forgive them and not cyber bully them to hell? Jennifer Lawrence is such a good example of this, because she has said that, in retrospect, she was initially being authentic, but the pressure of celebrity made her fall into that quirky persona as a safe place. But as soon as she got comfortable in it, they hated her for it, and made it extremely clear to her that they hated her for it. In the end, who is TRULY authentic? Who is truly being their unfiltered self 100% of the time? Not me, and I don't think many others.
-
As a rule, humans do not like to be duped. We like to know which kinds of signals to trust, and which to distrust. Being lulled into trusting a signal only to then have it revealed that the signal was untrustworthy is a shock to the system, unnerving and upsetting. People get angry when they find they have been duped. These reactions are even more heightened when we find we have been duped simply for someone else’s amusement at having done so.
Although this incident happened years ago, this is still a repeating pattern we see in social media today. Oftentimes when I come across videos on platforms such as TikTok or Instagram, people in the comments would be debating whether or not something was fabricated as "rage-bait". Rage-bait as a term is relatively new, but the concept is as old as time- whether it be used in newspapers, story-telling, movies, music, etc, rage-bait is an effective method to garner criticism, but more importantly- engagement. The more people talk and critique a work, the more it will rise in popularity.
-
6.1. Authenticity# Early in the days of YouTube, one YouTube channel (lonelygirl15) started to release vlogs (video web logs) consisting of a girl in her room giving updates on the mundane dramas of her life. But as the channel continued posting videos and gaining popularity, viewers started to question if the events being told in the vlogs were true stories, or if they were fictional. Eventually, users discovered that it was a fictional show, and the girl giving the updates was an actress. Many users were upset that what they had been watching wasn’t authentic. That is, users believed the channel was presenting itself as true events about a real girl, and it wasn’t that at all. Though, even after users discovered it was fictional, the channel continued to grow in popularity.
The lonelygirl15 example shows why authenticity matters so much on social media. What upset people was not that the story was fictional, but that the way the connection was presented did not match reality. This makes me think that authenticity is less about whether something is “real” or “fake,” and more about whether audiences clearly understand what kind of relationship or signal they are engaging with.
-
Authenticity is a concept we use to talk about connections and interactions when the way the connection is presented matches the reality of how it functions. An authentic connection can be trusted because we know where we stand. An inauthentic connection offers a surprise because what is offered is not what we get. An inauthentic connection could be a good surprise, but usually, when people use the term ‘inauthentic’, they are indicating that the surprise was in some way problematic: someone was duped.
I found it interesting that authenticity focuses on how a connection is presented and how it actually is. Even if a connection is weakly maintained or surface level as long as it is up front it could still be seen as authentic.
-
As a rule, humans do not like to be duped. We like to know which kinds of signals to trust, and which to distrust. Being lulled into trusting a signal only to then have it revealed that the signal was untrustworthy is a shock to the system, unnerving and upsetting. People get angry when they find they have been duped. These reactions are even more heightened when we find we have been duped simply for someone else’s amusement at having done so.
One thing that stood out to me was the idea that people react so strongly to inauthenticity because we hate being duped, not just misinformed. The example of lonelygirl15 shows how upsetting it feels when the type of connection being offered (a real person’s life) doesn’t match what’s actually happening. This made me realize that authenticity isn’t just about truth, but about trust and expectations in relationships online.
-
Early in the days of YouTube, one YouTube channel (lonelygirl15) started to release vlogs (video web logs) consisting of a girl in her room giving updates on the mundane dramas of her life. But as the channel continued posting videos and gaining popularity, viewers started to question if the events being told in the vlogs were true stories, or if they were fictional. Eventually, users discovered that it was a fictional show, and the girl giving the updates was an actress.
One thing that stood out to me was the idea that people react so strongly to inauthenticity because we hate being duped, not just misinformed. The example of lonelygirl15 shows how upsetting it feels when the type of connection being offered (a real person’s life) doesn’t match what’s actually happening. This made me realize that authenticity isn’t just about truth, but about trust and expectations in relationships online.
-
To describe something as authentic, we are often talking about honesty, in that the thing is what it claims to be. But we also describe something as authentic when we want to say that it offers a certain kind of connection.
This sentence really clarified authenticity for me. It’s not just about whether something is “real” or “fake,” but whether expectations line up with reality. That helps explain why joke accounts or surprise parties don’t feel inauthentic in a bad way because the mismatch isn’t harmful or misleading.
-
Authenticity in connection requires honesty about who we are and what we’re doing; it also requires that there be some sort of reality to the connection that is supposedly being made between parties. Authentic connections frequently place high value on a sense of proximity and intimacy. Someone who pretends to be your friend, but does not spend time with you (proximity) or does not open themselves up to trusting mutual interdependence (intimacy) is offering one kind of connection (being an acquaintance) under the guise of a different kind of connection (friendship).
I really like how this connects authenticity to honesty and trust as people generally view authenticity as just being as much as yourself as possible. However for this to be possible you have to feel a certain level of trust which is often forgotten when we expect someone or something to be authentic
-
Authenticity is a concept we use to talk about connections and interactions when the way the connection is presented matches the reality of how it functions. An authentic connection can be trusted because we know where we stand. An inauthentic connection offers a surprise because what is offered is not what we get. An inauthentic connection could be a good surprise, but usually, when people use the term ‘inauthentic’, they are indicating that the surprise was in some way problematic: someone was duped.
authenticity is basically “what you see is what you get,” so you can trust the vibe and know where you stand. When something’s inauthentic, it’s not just different than expected, it’s different in a way that feels misleading, like you got played. And yeah, even if the surprise could be harmless, “inauthentic” usually implies someone was tricked or taken advantage of.
-
This is not to say that there is no room for appreciating connections that are not fully honest, transparent, and earnest all the time. Social media spaces have allowed humor and playfulness to flourish, and sometimes humor and play are not, strictly speaking, honest. Often, this does not bother us, because the kind of connection offered by joke accounts matches the jokey way they interact on social media. We get to know a lot about public figures and celebrities, but it is not usually considered problematic for celebrity social media accounts to be run by publicist teams. As long as we know where we stand, and the kind of connection being offered roughly matches the sort of connection we’re getting, things go okay.
I like this point because it feels realistic — social media isn’t always about being 100% “authentic,” and sometimes the whole vibe is obviously playful or curated. Joke accounts work because everyone’s in on the bit, and celebrity accounts run by teams don’t feel weird as long as it’s clear what kind of relationship we’re actually getting. The problem starts when the account pretends it’s a genuine personal connection but it’s really marketing or manipulation.
-
-
visualizingcultures.mit.edu visualizingcultures.mit.edu
-
Floating lanterns as a prayer for the souls of the dead and a prayer for peace. The artist was 18 years old in 1974 when she responded to the appeal for pictures recalling the bomb. TAKASHIBA Harue Born in 1956 [17_30]
I really liked this image because I’ve actually done floating lanterns before, so it feels more personal to me. To me, lanterns in water symbolize prayer, remembrance, and reflection. Even though this painting is about the bomb, it doesn’t feel chaotic or violent. It feels quiet. The lanterns floating on the river suggest that even after so much destruction, there is still space for honoring the dead and hoping for peace. Unlike the hellfire or despair sections, this image feels like it’s looking forward. The light from the lanterns stands out against the dark water and sky, which makes the hope feel fragile but still present.
-
-
visualizingcultures.mit.edu visualizingcultures.mit.edu
-
On August 12, six days after the bomb, the artist came upon a skeleton sitting in a still-intact tilted barber’s chair. YAMABE Shōji 42 years old in August 1945 [01_29] detail
This image embodies the “ghost” theme not through wandering survivors but through absence. The skeleton seated in an intact barber’s chair transforms an ordinary space into something haunted. Unlike the chaotic hellfire scenes, this painting emphasizes eerie stillness. The six-day delay underscores abandonment as death lingering in familiar places without ritual or recognition. The figure is both present and absent, a physical remnant stripped of identity. The bomb leaves behind spaces and bodies that feel suspended between life and oblivion.
-
-
stats.libretexts.org stats.libretexts.org
-
A major flaw is that ice cream consumption generally increases in the months of June, July, and August regardless of advertisements. This effect is called a history effect and leads people to interpret outcomes as the result of one variable when another variable (in this case, one having to do with the passage of time) is actually responsible.
First and foremost, I'd believe that since these months land in the summer time season, ice cream consumption will always increase during this time of year. So definitely understand how this history effect stands true. The advertisements increase probably helps increase the cravings for ice cream during the hot weather, but the history effect of increased ice cream consumption during the summer shows that the advertisements don't solely tell the full story of this 30% increase, falsely giving Ben & Jerry's credit for this increase. On a deeper level, obviously Ben & Jerry's is a company who is money driven, but I don't think increased ice cream consumption will ever be a good thing or should be celebrated. Although I must say ice cream is delicious.
-
-
visualizingcultures.mit.edu visualizingcultures.mit.edu
-
NAKANO Kenichi 47 years old in August 1945 [02_33] Hiroshima in flames on the afternoon of August 6. The writing on the painting speaks of encountering “living Hell in this world.” For many survivors, the attempt to escape the firestorms that spread from the epicenters of the explosionsor the memory of someone who failed to escape these hellfires—became the image burned on the mind.
This painting makes literal what survivors repeatedly described as “hell.” Unlike the distant parachute image, this perspective places the viewer inside the inferno. The flames erase the sky and cityscape, transforming Hiroshima into a Buddhist vision of jigoku. The bodies scattered across the ground resemble figures in traditional hell as anonymous, contorted, and stripped of individuality. The phrase “living Hell in this world” collapses metaphor into reality; this was not symbolic suffering but an earthly apocalypse. Dower’s comparison to traditional depictions of hell suggests that survivors used familiar religious imagery to articulate something otherwise unspeakable.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
This manuscript aims to explore how mutations in the PDC-3 3 β-lactamase alter its ability to bind and catalyse reactions of antibiotic compounds. The topic is interesting and the study uses MD simulations and to provide hypotheses about how the size of the binding site is altered by mutations that change the conformation and flexibility of two loops that line the binding pocket. Some greater consideration of the uncertainties and how the method choice affect the ability to compare equilibrium properties would strengthen the quantitative conclusions. While many results appear significant by eye, quantifying this and ensuring convergence would strengthen the conclusions.
Strengths:
The significance of the problem is clearly described the relationship to prior literature is discussed extensively.
Comments on revised version:
I am concerned that the authors state in the response to reviews that it is not possible to get error bars on values due to the use of the AB-MD protocol that guides the simulations to unexplored basins. Yet the authors want to compare these values between the WT and mutants. This relates to RMSD, RMSF, % H-bond and volume calculations. I don't accept that you cannot calculate an uncertainty on a time averaged property calculated across the entire simulation. In these cases you can either run repeat simulations to get multiple values on which to do statistical analysis, or you can break the simulation into blocks and check both convergence and calculate uncertainties.
I note that the authors do provide error bars on the volumes, but the statistics given for these need closer scrutiny (I cant test this without the raw data). For example the authors have p<0.0001 for the following pair of volumes 1072 {plus minus} 158 and 1115 {plus minus} 242, or for SASA p<0.0001 is given for 2 identical numbers 155+/- 3.
I also remain concerned about comparisons between simulations run with the AB-MD scheme. While each simulation is an equilibrium simulation run without biasing forces, new simulations are seeded to expand the conformational sampling of the system. This means that by definition the ensemble of simulations does not represent and equilibrium ensemble. For example, the frequency at which conformations are sampled would not be the same as in a single much longer equilibrium simulation. While you may be able to see trends in the differences between conditions run in this way, I still don't understand how you can compare quantitative information without some method of reweighing the ensemble. It is not clear that such a rewieghting exists for this methods, in which case I advise some more caution in the wording of the comparisons made from this data.
At this stage I don't feel the revision has directly addressed the main comments I raised in the earlier review, although there is a stronger response to the comments of Reviewer #2.
-
Author response:
The following is the authors’ response to the current reviews.
Reviewer #2 (Public review):
Summary:
In the manuscript entitled "Ω-Loop mutations control dynamics 2 of the active site by modulating the 3 hydrogen-bonding network in PDC-3 4 β-lactamase", Chen and coworkers provide a computational investigation of the dynamics of the enzyme Pseudomonas-derived chephalosporinase 3 (PDC3) and some mutants associated with increased antibiotic resistance. After an initial analysis of the enzyme dynamics provided by RMSD/RMSF, the author conclude that the mutations alter the local dynamics within the omega loop and the R2 loop. The authors show that the network of hydrogen bonds in disrupted in the mutants. Constant pH calculations showed that the mutations also change the pKa of the catalytic lysine 67 and pocket volume calculations showed that the mutations expand the catalytic pocket. Finally, time-independent componente analysis (tiCA) showed different profiles for the mutant enzyme as compared to the wild type.
Strengths:
The scope of the manuscript is definitely relevant. Antibiotic resistance is an important problem and, in particular, Pseudomonas aeruginosa resistance is associated with an increasing number of deaths. The choice of the computational methods is also something to highlight here. Although I am not familiar with Adaptive Bandit Molecular Dynamics (ABMD), the description provided in the manuscript that this simulation strategy is well suited for the problem under evaluation.
Weaknesses:
In the revised version, the authors addressed my concerns regarding their use of the MSM, and in my view, their conclusions are now much more robust and well-supported by the data. While it would be very interesting to see a quantitative correlation between the effects of the mutations observed in the MD data and relevant experimental findings, I understand that this may be beyond the scope of the manuscript.
Thank you for the careful evaluation and constructive comments. Regarding the suggestion of a more quantitative correlation with experimental observables, we agree that this would be valuable, and we have noted it as an important direction for future work.
Reviewer #3 (Public review):
Summary:
This manuscript aims to explore how mutations in the PDC-3 3 β-lactamase alter its ability to bind and catalyse reactions of antibiotic compounds. The topic is interesting and the study uses MD simulations and to provide hypotheses about how the size of the binding site is altered by mutations that change the conformation and flexibility of two loops that line the binding pocket. Some greater consideration of the uncertainties and how the method choice affect the ability to compare equilibrium properties would strengthen the quantitative conclusions. While many results appear significant by eye, quantifying this and ensuring convergence would strengthen the conclusions.
Strengths:
The significance of the problem is clearly described the relationship to prior literature is discussed extensively.
Comments on revised version:
I am concerned that the authors state in the response to reviews that it is not possible to get error bars on values due to the use of the AB-MD protocol that guides the simulations to unexplored basins. Yet the authors want to compare these values between the WT and mutants. This relates to RMSD, RMSF, % H-bond and volume calculations. I don't accept that you cannot calculate an uncertainty on a time averaged property calculated across the entire simulation. In these cases you can either run repeat simulations to get multiple values on which to do statistical analysis, or you can break the simulation into blocks and check both convergence and calculate uncertainties.
We thank the reviewer for raising this point. We would like to clarify that we did not intend to state that error bars are impossible to obtain under AB-MD. In fact, we reported error bars for several quantities derived from the AB-MD trajectories (we also broke the trajectories into blocks and calculated uncertainties for RMSF in our first-round response as you suggested). However, these data are closely related to your concern about comparing quantitative information without an appropriate reweighting of the ensemble. Therefore, in the revised manuscript, we removed quantitative analyses that were calculated directly from the raw AB-MD trajectories. Instead, the quantitative comparisons are now obtained from MSM analysis. We report pocket volumes and key interaction metrics for MSM metastable states, with corresponding error bars for these MSM-based quantities (Figure 6 and its supplementary figure).
I note that the authors do provide error bars on the volumes, but the statistics given for these need closer scrutiny (I cant test this without the raw data). For example the authors have p<0.0001 for the following pair of volumes 1072 {plus minus} 158 and 1115 {plus minus} 242, or for SASA p<0.0001 is given for 2 identical numbers 155+/- 3.
Thank you for this comment. As noted above, we have removed the table from the manuscript, and the pocket-volume results together with their error bars are now shown in Figure 6. To address the concern raised here and to avoid making the same mistake in future analyses, we re-examined how the statistics were computed. We believe the very small p-values were caused by treating per-frame MD values as independent observations in two-sample t-tests. Because consecutive MD frames are strongly time-correlated, they do not satisfy the independence assumption, which can greatly overestimate the effective sample size and lead to artificially small p-values. For the SASA, a p < 0.0001 is reported even though both values are shown as 155 ± 3. This is due to rounding, which can hide subtle underlying differences.
I also remain concerned about comparisons between simulations run with the AB-MD scheme. While each simulation is an equilibrium simulation run without biasing forces, new simulations are seeded to expand the conformational sampling of the system. This means that by definition the ensemble of simulations does not represent and equilibrium ensemble. For example, the frequency at which conformations are sampled would not be the same as in a single much longer equilibrium simulation. While you may be able to see trends in the differences between conditions run in this way, I still don't understand how you can compare quantitative information without some method of reweighing the ensemble. It is not clear that such a rewieghting exists for this methods, in which case I advise some more caution in the wording of the comparisons made from this data.
At this stage I don't feel the revision has directly addressed the main comments I raised in the earlier review, although there is a stronger response to the comments of Reviewer #2.
We thank the reviewer for reiterating this important point, and we agree with the underlying concern. Although AB-MD generates unbiased trajectories, the ensemble of simulations does not represent an equilibrium ensemble. As a result, statistics computed by simply concatenating all AB-MD trajectories should not be used for quantitative comparisons. In the original version, we acknowledge that we reported several quantitative descriptors directly from concatenated AB-MD frames, including (i) distributions of χ1 torsions, (ii) mean pocket volumes and SASA, and (iii) percentages of some key interactions. We agree that this was not appropriate given the adaptive sampling protocol. In the revised manuscript, we have removed these quantitative analyses.
We retained RMSD and RMSF analyses, but we have revised their wording and clarified their purpose. RMSD and RMSF are used only to summarize the structural variability and residue-level mobility observed across the collected trajectory segments and to motivate the selection of structural features for MSM construction. The manuscript now states: “Because AB-MD adaptively seeds new unbiased trajectories to expand conformational sampling, RMSD and RMSF are used here to summarize the structural variability and per-residue mobility observed across the collected trajectories.”
Regarding the reviewer’s question about reweighting, the Markov state model (MSM) provides a principled framework to obtain the stationary distribution π from the transition probability matrix T<sub>τ</sub>. The resulting π<sub>i</sup> gives the equilibrium weight of each microstate i, and the corresponding discrete free energy can be written as F<sup>i</sup>=−k<sub>B</sub>Tln(π<sub>i</sup>). PCCA then coarse-grains the microstate space into a small number of metastable states. In the revised manuscript, quantitative comparisons are therefore derived from the MSM at the level of these metastable states, rather than from unweighted counts of concatenated AB-MD frames.
Accordingly, we have revised the sections “E219K and Y221A mutations facilitate proton transfer” and “Substitutions enlarge the active-site pocket to accommodate bulkier R1 and R2 groups of β-lactams”, and we have added new figures in Figure 6 and its figure supplement. The adjustments to the quantitative analyses do not affect our original conclusions.
The following is the authors’ response to the original reviews.
Reviewer #1 (Public review):
Summary:
This manuscript uses adaptive sampling simulations to understand the impact of mutations on the specificity of the enzyme PDC-3 β-lactamase. The authors argue that mutations in the Ω-loop can expand the active site to accommodate larger substrates.
Strengths:
The authors simulate an array of variants and perform numerous analyses to support their conclusions. The use of constant pH simulations to connect structural differences with likely functional outcomes is a strength.
Weaknesses:
I would like to have seen more error bars on quantities reported (e.g., % populations reported in the text and Table 1).
We appreciate this point. Here, the population we analyze is intended to showcase conformational differences across variants rather than to estimate equilibrium occupancies. Although each system includes 100 trajectories, they were generated using an adaptive-bandit protocol. The protocol deliberately guides towards underexplored basins, therefore conformational heterogeneity betweentrajectories is expected by design. For example, in E219K the MSM decomposition shows that in states 1, 6, and 7 the K67(NZ)–S64(OG) distance is almost entirely > 6 Å, whereas in states 2 and 3 it is almost entirely < 3.5 Å (Figure 5—figure supplement 12). These distances suggest that the hydrogen bond fraction is approximately zero in states 1, 6, and 7, and close to one in states 2 and 3. In addition, the mean first passage time of the Markov state models suggests that the formation and disruption of this hydrogen bond occur on the microsecond timescale, which is far longer than the length of each individual trajectory (300 ns). Consequently, across the 100 replicas, some trajectories exhibit very low fractions, while others display the opposite trend. Under such bimodal, protocol-induced heterogeneity, computing an error bar across trajectories mainly visualizes the protocol’s dispersion and risks being misread as thermodynamic uncertainty, which is not central to our aim of comparing conformational differences between wild-type PDC-3 and variants. We therefore do not include the error bars.
Reviewer #2 (Public review):
Summary:
In the manuscript entitled "Ω-Loop mutations control dynamics of the active site by modulating the 3 hydrogen-bonding network in PDC-3 4 β-lactamase", Chen and coworkers provide a computational investigation of the dynamics of the enzyme Pseudomonas-derived cephalosporinase 3 (PDC3) and some mutants associated with increased antibiotic resistance. After an initial analysis of the enzyme dynamics provided by RMSD/RMSF, the author concludes that the mutations alter the local dynamics within the omega loop and the R2 loop. The authors show that the network of hydrogen bonds is disrupted in the mutants. Constant pH calculations showed that the mutations also change the pKa of the catalytic lysine 67, and pocket volume calculations showed that the mutations expand the catalytic pocket. Finally, time-independent component analysis (tiCA) showed different profiles for the mutant enzyme as compared to the wild type.
Strengths:
The scope of the manuscript is definitely relevant. Antibiotic resistance is an important problem, and, in particular, Pseudomonas aeruginosa resistance is associated with an increasing number of deaths. The choice of the computational methods is also something to highlight here. Although I am not familiar with Adaptive Bandit Molecular Dynamics (ABMD), the description provided in the manuscript suggests that this simulation strategy is well-suited for the problem under evaluation.
Weaknesses:
In the description of many of their results, the authors do not provide enough information for a deep understanding of the biochemistry/biophysics involved. Without these issues addressed, the strength of the evidence is of concern.
We thank the reviewer for pointing out the need for deeper discussion of the biochemical and biophysical implications of our results. In our manuscript, we begin by examining basic structural metrics (e.g., RMSD and RMSF) which clearly indicate that the major conformational changes occur in the Ω-loop and the R2 loop. We have now added a paragraph to describe the importance of the Ωloop and highlighted it in the revised manuscript on lines 142-166 of page 6. This observation guided our subsequent focus on these regions, as well as on the catalytic site. Our analysis revealed notable alterations in the hydrogen bonding network—especially in interactions involving the K67-S64, K67N152, K67-G220, Y150-A292, and N287-N314 pairs. These observations led us to conclude that:
(1) Mutations E219K and Y221A facilitate the proton transfer of catalytic residues. This is consistent with prior experimental data showing that these substitutions produce the most pronounced increase in sensitivity to cephalosporin antibiotics (lines 210-212 in page 8 of the revised manuscript).
(2) Substitutions enlarge the active-site pocket to accommodate bulkier R1 and R2 groups of β-lactams.This is in line with MIC measurements reported by Barnes et al. (2018), which showed that mutants with larger active-site pockets exhibit markedly greater sensitivity to cephalosporins with bulky side chains than others (lines 249-259 in pages 10).
Furthermore, we applied Markov state models (MSMs) to explore the timescales of the transitions between these different conformational states. We believe that these methodological steps support our conclusions.
Reviewer #3 (Public review):
Summary:
This manuscript aims to explore how mutations in the PDC-3 3 β-lactamase alter its ability to bind and catalyse reactions of antibiotic compounds. The topic is interesting, and the study uses MD simulations to provide hypotheses about how the size of the binding site is altered by mutations that change the conformation and flexibility of two loops that line the binding pocket. However, the study doesn't clearly describe the way the data is generated. While many results appear significant by eye, quantifying this and ensuring convergence would strengthen the conclusions.
Strengths:
The significance of the problem is clearly described, and the relationship to prior literature is discussed extensively.
Weaknesses:
The methods used to gain the results are not explained clearly, meaning it was hard to determine exactly how some data was obtained. The convergence and uncertainties in the data were not adequately quantified. The text is also a little long, which obscures the main findings.
We thank the reviewer for the suggestion. We respectfully ask the reviewer to specify which aspects of the data-generation methods are unclear so that we can include the necessary details in the next revision. Moreover, all statistics that are reported in the manuscript are obtained from extensive analyses of 300,000 simulation frames. The Markov state models have been validated by the ITS plots and Chapman-Kolmogorov (CK) test. The two-sample t-tests were also carried out for the volume and SASA.
Reviewer #2 (Recommendations for the authors):
(1) Figure 1D focus on the PDC3 catalytic site. However, the authors mentioned before that the enzyme has two domains, an alpha domain and an alpha/beta domain. The reader would benefit from a more detailed description of the enzyme, its active site, AND the location of the mutants under investigation in the figure.
We have updated Figure 1D and marked the positions of all mutations (V211A/G, G214A/R, E219A/G/K and Y221A/H), which have now been highlighted as spheres.
(2) Since in the journal format, the results come before the methods. It would be interesting to add a brief description of where the results came from. For example, in the first section of the results, the authors describe the flexibility of the omega loop and the R2 loop. However, the reader won't know what kind of simulation was used and for how long, for example. A sentence would add the required context for a deeper understanding here.
At the beginning of the Results and Discussion section we now state: “To investigate how the mutations in the Ω-loop affect PDC-3 dynamics, adaptive-bandit molecular dynamics (AB-MD) simulations were carried out for each system. 100 trajectories of 300 ns each (totaling 30 μs per system) were run.”
(3) Still in the same section, the authors don't define what change in RMSF is considered significant. For example, I can't see a relevant change in the RMSF for the omega loop between the et enzyme and the E219 mutants in Figure 2D. A more objective definition would be of benefit here.
Our analysis reveals that while the wild-type PDC-3 and the G214A, G214R, E214G, and Y221A variants exhibit an average per-residue RMSF of around 4 Å in the Ω-loop, the V211A and V211G variants show markedly lower values (around 1.5 Å), and the E219K and Y221H variants exhibit intermediate values between 2 and 2.5 Å. In addition, the fluctuations around the binding site should be seen collectively along with the fluctuations in the R2-loop. Importantly, we urge the reviewer to focus on the MDLovofit analysis in Figure 2C, where the dynamic differences between the core and the fluctuating loops is clearly evident.
(4) In line 138, the authors state that "Therefore, the flexibility of these proteins is mainly caused by the fluctuations in the Ω-loops and R2-loop". This is quite a bold statement to be drawn at this point. First of all, there is no mention of it in the manuscript, but is there any domain movement? Figure 2C clearly shows that there is some mobility in omega and R2 loops. But there is no evidence shown in the manuscript that shows that "the flexibility of these proteins is mainly caused by the fluctuations in the" loops. Please consider rephrasing this sentence or adding more data, if available.
We have revised the wording to take the reviewer’s concern into account. The sentence now states: “Therefore, flexibility of PDC-3 is predominantly localized to the Ω- and R2-loops, whereas the remainder of the structure is comparatively rigid.” To further explain to the reviewer, the β lactamase enzymes are fairly rigid structures, where no large-scale domain motions occur. Instead, the enzyme communicates structurally via cross correlation of loop dynamics ( https://doi.org/10.7554/eLife.66567 ).
(5) I guess, the most relevant question for the scope of the paper is not answered in this section. The authors show that the mobility of the omega- and R2-loops is altered by some mutations. Why is that? I wish I could see a figure showing where the mutations are and where the loops are. This question will come back in other sections.
We have updated Figure 1D to mark the positions of all mutations (V211A/G, G214A/R, E219A/G/K and Y221A/H) as spheres. The Ω- and R2-loops are also highlighted. All mutations map to the Ω-loop, indicating that these substitutions directly perturb this region. Notably, K67 forms a hydrogen bond with the backbone of G220 within the Ω-loop and another with the phenolic hydroxyl of Y150. Y150, in turn, hydrogen-bonds with A292 in the R2 loop. Together, the residue interaction network (G220– K67–Y150–A292) suggest a pathway by which Ω-loop mutations propagate their effects to the R2 loop.
(6) The authors then analyze the network of polar residues in the active site and the hydrogen bonds observed there. For the K67-N152 hydrogen bond, for example, there is a reduction in the occupancy from ~70% in the wild-type enzyme to ~30% and 40% in the mutants E219K and Y221, respectively. This finding is interesting. The question that remains is "why is that"? From the structural point of view, how does the replacement of E219 with a Lysine alter the hydrogen bond formation between K67 and N152? Is it due to direct competition? Solvent rearrangement? The reader is left without a clue in this section. Also, Figure 3B won't help the reader, since the mutated residues are not shown there. Please consider adding some information about why the authors believe that the mutations are disrupting the active site hydrogen bond network and showing it in Figure 3B.
We appreciate the comment and have updated Figures 1D and 3B to highlight the mutation sites. The change from ~70% in the wild type to ~30–40% in the E219K and Y221T variants reported in Table 1 refers to the S64–K67 hydrogen bond. In the wild type, K67 forms an additional hydrogen bond with G220 on the Ω-loop, which helps anchor the K67 side chain in a geometry that favors the S64–K67 interaction. In the variants, the mutations reshape the Ω-loop and frequently disrupt the K67–G220 contact. The loss of this local anchor increases the conformational dispersion of K67, which is consistent with the observed reduction of the S64–K67 occupancy. Furthermore, our observation that the mutations are disrupting the active-site hydrogen-bond network is a data-driven conclusion rather than a subjective inference. Across ten systems, our AB-MD simulations provided 30 µs of sampling per system. Saving one frame every nanosecond yielded 30,000 conformations per system and 300,000 in total. All hydrogen-bond and salt-bridge statistics were computed over this full ensemble. Thus, the conclusion that the mutations disrupt the active-site hydrogen-bond network follows directly from these ensemble statistics.
(7) The pKa calculations and the pocket volume calculations show that the mutations expand the volume of the catalytic site and alter the microenvironment. Is there any change in the solvation associated with these changes? If the volume expands and the environment becomes more acidic, are there more water molecules in the mutants as compared to the wt enzyme? If so, can changes in solvation be associated with the changes in the hydrogen bond network? Would a simulation in the presence of a substrate be meaningful here? ( I guess it would!).
Regarding solvation, we observe a modest increase in transient water occupancy associated with the increase in volume of the pocket. The conserved deacylation water molecule is the most important and is always present throughout the simulation. Additional waters enter and leave the pocket but do not form persistent interactions that measurably perturb the hydrogen-bond network of the Ω- and R2-loops. We agree that simulations with a bound substrate would be informative. However, our study focuses on how Ω-loop mutations modulate the active site of apo PDC-3 and its variants. Within this scope, we find: (i) Amino acid substitutions change the flexibility of Ω-loops and R2-loops; (ii) E219K and Y221A mutations facilitate the proton transfer; (iii) Substitutions enlarge the active-site pocket to accommodate bulkier R1 and R2 groups of β-lactams.
(8) I have some concerns regarding the Markov State Modeling as shown here. After a time-independent component analysis, the authors show the projections on the components, which is different between wild wild-type enzyme and the mutants, and draw some conclusions from these changes. For example, the authors state that "From the metastable state results, we observe that E219K adopts a highly stable conformation in which all the tridentate hydrogen-bonding interactions (K67(NZ)-S64(OG), K67(NZ)N152(OD1) and K67(NZ)-G220(O) mentioned above are broken". This is conclusion is very difficult to draw from Figure 5 alone. Unless the macrostates observed in the MSM can be shown (their structures) and could confirm the broken interactions, I really don't believe that the reader can come to the same conclusion as drawn by the authors here. I would recommend the authors to map the macrostates back to the coordinates and show them (what structure corresponds to what macrostate). After showing that, it makes sense to discuss what macrostate is being favored by what mutation. Taking conclusions from tiCA projections only is not recommended. I very strongly suggest that the authors revisit this entire section, adding more context so that the reader can draw conclusions from the data that is shown.
We appreciate the reviewer’s concern. In the Markov state modeling section, our objective is to quantify the timescales (via mean first passage times) associated with the formation and disruption of the critical hydrogen bonds (K67(NZ)-S64(OG), K67(NZ)-N152(OD1), K67(NZ)-G220(O), Y150(N)A292(O), N287(ND2)-N314(OD1)) mentioned above. Representative structures illustrating these interactions are shown in Figures 3B and 4A. We agree that the main Figure 5 alone does not convey structural information. Accordingly, we provide Figure 5—figure supplements 12–16. Together, Figure 5B and Figure 5—figure supplements 12–16 map structures to metastable states, whereas Figures 3B and 4A supply atomistic detail of the interactions. Author response image 1 presents selected subplots from Figure 5— figure supplements 12–14. Together with the free-energy landscape in Figure 5A, these data indicate that E219K adopts a highly stable conformation in which all three K67-centered hydrogen bonds (K67(NZ)–S64(OG), K67(NZ)–N152(OD1), and K67(NZ)–G220(O)) are broken.
Author response image 1.
TICA plot illustrates the distribution of E219K with the colour indicating the K67(NZ)-S64(OG), K67(NZ)-N152(OD1) and K67(NZ)-G220(O) distance.
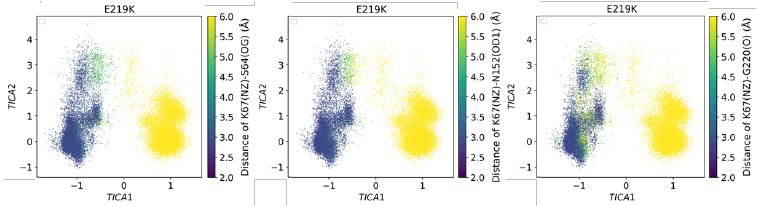
(9) As a very minor issue, there are a few typos in the manuscript text. The authors might want to take some time to revisit their entire text. Examples in lines 70, 197, etc.
Thank you for your comment. We have corrected these typos.
Reviewer #3 (Recommendations for the authors):
This manuscript aims to explore how mutations in the PDC-3 3 β-lactamase alter its ability to bind and catalyse reactions of antibiotic compounds. The topic is interesting, and the study uses MD simulations to provide hypotheses about how the size of the binding site is altered by mutations that change the conformation and flexibility of two loops that line the binding pocket.
However, the study doesn't clearly describe the way the data is generated and potentially lacks statistical rigour, which makes it uncertain if the key results are significant. As such, it is difficult to judge if the conclusions made are supported by data.
All necessary data-acquisition methods are described in the Methods section. The Markov state models have been validated by the ITS plot and the Chapman-Kolmogorov (CK) test (Figure 5—figure supplement 2–11) . The two-sample t-tests were also carried out for the volume and SASA (Table 2).
The results section jumps straight to reporting RMSD and RMSF values; however, it is not clear what simulations are used to generate this information. Indeed, the main text does not mention the simulations themselves at all. The methods section mentions that 10 independent MD simulations were set up for each system, but no information is given as to how long these were run or the equilibration protocol used. Then it says that AB-MD simulations were run, but it is not clear what starting coordinates were used for this or how the 10 replicates were fed into these simulations. Most importantly, are the RMSD and RMSF calculations and later distance distribution information derived from the equilibrium MD runs or from the AB-MD simulations?
Thank you for pointing this out. We have added “To investigate how the mutations in the Ω-loop affect PDC-3 dynamics, adaptive-bandit molecular dynamics (AB-MD) simulations were carried out for each system. 100 trajectories of 300 ns each (totaling 30 μs per system) were run.” to the Results and Discussion section. We didn’t run 10 independent MD simulations per system. We regret the typo in the Methods section that confused the reviewer. The sentence should have read – ‘All-atom MD simulations of wild-type PDC-3 and its variants were performed.’ Each system was equilibrated for 5 ns at 1 atmospheric pressure using Berendsen barostat. AB-MD simulations were initiated from these equilibrated structures. All analyses, apart from CpHMD, are based on the AB-MD trajectories.
If these are taken from the equilibrium simulations, then it is critical that the reproducibility and statistical significance of the simulations is established. This can be done by calculating the RMSD and RMSF values independently for each replicate and determining the error bars. From this, the significance of differences between WT and mutant simulations can be determined. Without this, I have no data to judge if the main conclusions are supported or not. If these are derived from the AB-MD simulations, then I want to know how the independent simulations were combined and reweighted to generate overall RMSD, RMSF, and distance distributions. Unless I misunderstand the approach, the individual simulations no longer sample all regions of conformational space the same relative amount you would see in a standard MD simulation - specific conformational regions are intentionally run more to enhance sampling, then the overall conformational distributions cannot be obtained from these simulations without some form of reweighting scheme. But no such scheme is described. In addition, convergence of the data is required to ensure that the RMSD, RMSF, and distances have reached stable values. It is possible that I am misunderstanding the approach here. But in that case, I hope the authors can clarify the method and provide a means of ensuring that the data presented is converged. Many of the differences are clear by eye, but it is important to know they are not random differences between simulations and rather reflect differences between them.
Thank you for raising this important point. In our AB-MD workflow, the adaptive bandit is used only for starting-structure selection (adaptive seeding). After each epoch, it chooses new starting snapshots from previously sampled conformations and launches the next runs. Each trajectory itself is standard, unbiased MD with no biasing potentials and no modification of the Hamiltonian. In other words, AB decides where we start, but does not alter the physics or sampling dynamics within an individual trajectory. In addition, our goal in this work is to compare variants under the same adaptive-bandit (AB) protocol, rather than to estimate equilibrium (Boltzmann) populations. Hence, we did not apply equilibrium reweighting to RMSD, RMSF, or distance distributions. However, MSM section provides reweighted reference results based on the MSM stationary distribution.
In the response to reviews, the authors state that the "RMSF is a statistical quantity derived from averaging the time series of atomic displacements, resulting in a fixed value without an inherent error bar." But normally we would run multiple replicates and get an error bar from the different values in each. To dismiss the request for uncertainties and error bars seems to miss the point. I strongly agree with the prior reviewer that comparisons between RMSF or other values should be accompanied by uncertainties and estimates of statistical significance.
Regarding the reviewers’ suggestion to present the data as a bar graph with error bars, we would like to note that RMSF is calculated as the time average of the fluctuations of each residue’s Cα atom over the entire simulation. As such, RMSF is a statistical quantity derived from averaging the time series of atomic displacements, resulting in a fixed value without an inherent error bar. We believe that our current presentation clearly and accurately reflects the local flexibility differences among the variants. Nearly all published studies report RMSF in this way, as indicated by the following examples:
Figure 3a in DOI: https://doi.org/10.1021/jacsau.2c00077
Figure 2 in DOI: https://doi.org/10.1021/acs.jcim.4c00089
Supplementary Fig. 1, 2, 5, 9, 12, 20, 22, 24, and 26 in DOI: https://doi.org/10.1038/s41467-022-293313
However, in response to the reviewers’ strong request, we present RMSF plots with error bars in our response letter.
Author response image 2.
The root-mean-square fluctuation (RMSF) profiles of wild-type PDC-3 and its variants. Blue lines show the mean RMSF across 100 independent MD trajectories for each system; red translucent bands denote the standard deviation across trajectories. The Ω-loop (residues G183 to S226) is highlighted in yellow, and the R2-loop (residues L280 to Q310) is highlighted in blue.
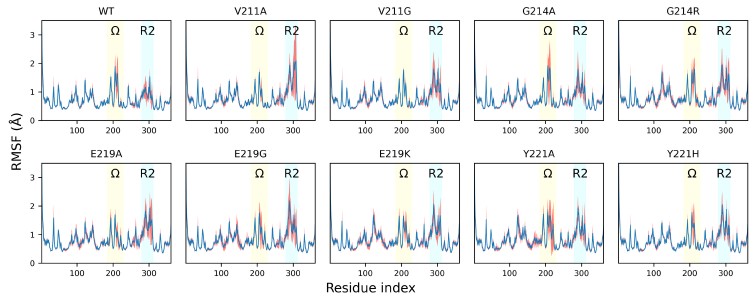
It was good to see that convergence of the constant-pH simulations was shown. While it can be challenging to get absolute pH values from the implicit solvent-based simulations, the differences between the systems are large and the trends appear significant. I was not clear how the starting coordinates were chosen for these simulations. Is the end point of the classical simulations, or is a representative snapshot chosen somehow?
To ensure comparison, all systems used the X-ray crystal structure (PDB ID: 4HEF) with T79A substitution as the initial structure. The E219K and Y221A mutants were generated in silico using the ICM mutagenesis module. We have added the clarification in Methods section: “The starting structures were identical to those used for AB-MD.”
Significant figures: Throughout the text and tables, the authors present data with more figures than are significant. 1071.81+-157.55 should be reported as 1100 +/ 160 or 1070 =- 160 . See the eLife guidelines for advice on this.
Thank you for your suggestion. We have amended these now.
The manuscript is very long for the results presented, and I feel that a clearer story would come across if the authors shortened the text so that the main conclusions and results were not lost.
We appreciate the suggestion. We examined the twenty most recent research articles published in eLife and found that they are either longer than or comparable in length to our manuscript.
-
-
sacred-texts.com sacred-texts.com
-
how difficult it is to meet with a fair creature, of whom one can say, ‘This is, indeed, the one; here is, at last, perfection.’ There are, indeed, many who fascinate; many who are ready with their pens, and who, when occasion
Tô-no-Chiûjiô is complaining that parents and friends always talk up a girl’s skills in music and poetry, but when you actually meet them, it’s a letdown.
-
but if one single tear bedews her cheek, she is no longer strong in the sanctity of her vow.
emotions such as crying are seen as a moral failure.
-
She is not only incapable of sharing these with him, but might carelessly remark, "What ails you?" How severely would this try the temper of a man
male irritation
-
But how often such girls as these are conceited about their own accomplishments, and endeavor unduly to disparage those of others!
shows that female talent is admired but also distrusted/ distasteful when the woman themself is self aware of the talent.
-
-
nmoer.pressbooks.pub nmoer.pressbooks.pub
-
but the former sounds less aggressive and accusatory,
don't be rude
-
Peer review can feel scary because you may feel uncomfortable sharing your writing at first, but remember that each writer is working toward the same goal:
It's important to remember this if you are scared to share your work
-
-
socialsci.libretexts.org socialsci.libretexts.org
-
Development is multidirectional
Development isn’t linear; people can show increases in some areas and decreases in others (e.g., better problem-solving but slower reaction time).
-
-
www.biorxiv.org www.biorxiv.org
-
Note: This response was posted by the corresponding author to Review Commons. The content has not been altered except for formatting.
Learn more at Review Commons
Reply to the reviewers
Reviewer #1 (R1)
R1 General statement: Here, Escalera-Maurer and colleagues, present an up-to-date distribution of homologues of Hok toxic proteins belonging to the well-annotated, but otherwise functionally obscure, hok/Sok type I toxin-antitoxin system, across the RefSeq database. Although such computational analyses have been done in the past, the authors here find many more hok homologs than described before, and they categorise their distribution based on whether they are encoded on chromosomes, plasmids, or (pro)phages. These computational analyses are in general tricky with T1TAs, as their toxins are quite short (~50 amino acids, as is the case for Hok), which is why the authors here used three separate approaches to expand their search (nucleotide-level BLAST, protein-homology, or both combined with Infernal). The authors cluster the Hok homologues they find based on a 60% sequence identity cut-off (expanding the known clusters in the process), and proceeded to test 31 candidates belonging to 15 sequence-clusters for their toxicity in Salmonella Typhimurium LT2, showing that 30/31 were toxic upon induction. An interesting finding from their endeavours is that hok/Sok homologues are enriched within prophages and large plasmids, but are not enriched near bacterial anti-phage defense systems (in contrast to the SymE/SymR T1TA). The findings suggest that hok/Sok are indeed sometimes linked to phage and plasmid biology, although they might not be antiphage defenses per se (they have been clearly shown in the past to be addiction modules, and this is still clearly true).
Authors' answer to R1 General statement: __We do not state here that hok/Sok are not anti-phage defense systems, but we simply observe that they do not cluster with anti-phage defense systems. We have also observed (unpublished data) that known defense systems do not systematically cluster together with other defense systems. Therefore, strong association with other defense systems would have been a strong indication of their function in phage defense but the fact that we did not observe any association with defense systems does not exclude they are involved in phage defense. __
R1_C1: My expertise lies towards the experimental side of the authors' work, I thus cannot comment on the accuracy/robustness of the computational analyses performed here. The authors do a fine job in clearly stating their findings overall; I could follow most of the conclusions, and I deemed that most of them were supported by their work. Additionally, I find that this paper is a missed opportunity to uncover even more novel biology connected to the interesting hok/Sok T1TAs. The paper does not provide a new framework to think about what is the function of the chromosomal/prophage hok/Sok T1TA systems, although I realize that this is very difficult to accomplish, especially when considering that hok/Sok systems have been around in the literature for almost 40 years.
Authors' answer to R1_C1: We agree with the reviewer, as we indeed performed this analysis having in mind to clarify the role of hok/Sok systems. However, we still believe that our strong survey of Hok loci put in light their enrichment in various mobile genetic elements, such as prophage and large conjugative plasmids, which is indubitably linked to their function. In addition, our study will guide future experimental efforts in uncovering the function of these systems, for example by helping researchers to select relevant homologs to test for a specific function.__ __
R1_C2: My major comment is in regard to the Hok toxicity assays (Fig. 2). The authors state in the discussion that "Hok peptides originating from chromosomes are as toxic as those from plasmids", but I believe that the way that they tested their constructs might not have allowed them to see toxicity differences between the two groups. Specifically, using the multi-copy plasmid pAZ3 (pBR322 origin of replication; ~15-20 plasmid copies per chromosome) to induce the different Hok toxin homologues in Salmonella Typhimurium LT2 with arabinose might have masked toxicity differences that would otherwise be apparent on the chromosomal expression-level.
Some of the authors themselves have previously used the FASTBAC-Seq method to study the Hok homologue from plasmid R1, a useful technique during which a toxin is integrated in the chromosome, in order to study their toxicity under natural levels of expression. I believe that an ideal scenario would be to apply FASTBAC-seq to some of the 31 Hok homologues described here (e.g., a subset of plasmidic vs chromosomal Hok homologues) to shed light on potential toxicity differences between the Hok clusters. This would increase the value of the presented study.
Alternatively, the authors could employ an L-arabinose concentration gradient to titrate the expression levels of the Hok toxins in order to potentially see different toxicity levels from the different homologues. However, this is not going to work in the system as they are using it now for two reasons:
- a) the S. Typhimurium LT2 (STm) used here has its arabinose utilization operon intact (araBAD), which means that Salmonella can catabolize arabinose to use it as a carbon source. This catabolization process interferes with the arabinose induction (i.e., Salmonella eats arabinose instead of using it as the Hok inducer). To ameliorate this, the authors could delete the araBAD operon in STm, rendering STm incapable of catabolizing arabinose, and repeat the experiments in that strain. Or use E. coli BW25113 as the expression host, which already has the araBAD operon deleted (it is not clear to me why the different Hok homologues would not be toxic in E. coli, as the different Hok homologues are widely diverse in sequence, as the authors found here).
- b) Even with the araBAD operon deleted, the arabinose induction would be bimodally on or off in the population, due to the bimodal expression of the arabinose transporter (AraE; see Khlebnikov et al., 2002). This would again not allow for titratable arabinose-inducible expression from different concentrations of arabinose. The solution for this would be to co-express a separate plasmid with araE, which would render every cell the same in regards to arabinose permeability, and thus the system would be titratable (as explained in Khlebnikov et al., 2002). Therefore, if the authors would be interested to go towards this route, they would have to first delete the araBAD from STm, then transform STm with an araE plasmid, and redo the experiments. In addition, I would propose to the authors to use the drop plate method (agar plate-based), which is more sensitive compared to the liquid assays employed here.
Having said all that, I understand that all this experimental work would be strenuous and time-consuming, and although I would like to see it happen, this is not my paper. I would be content therefore if the authors toned down the claim that plasmidic vs chromosomal Hok homologues have the same toxicity, and discuss that chromosomal levels of toxicity are an important caveat that has not been explored here.
__Authors' answer to R1_C2: __ We thank the reviewer for the detailed suggestion on how to better assess toxicity differences by using an araBAD deletion mutant overexpressing araE. We repeated the arabinose induction assays using drop assays and strain BW25223 with plasmid pJAT13araE and our pAZ3 based plasmid carrying Hok CDS homologs. However, we obtained similar data, not being able to distinguish between the toxicity of chromosomal versus plasmidic CDS, even using different concentration of Arabinose. This is probably because low concentration of the Hok protein are sufficient for activity, but here we are bypassing all post-transcriptional silencing by the native Hok mRNAs by expressing directly the protein, and we are using a multicopy plasmid. We now included 0.01% arabinose induction drop assays in the manuscript as the data obtained with other arabinose concentration did not provide new information. In any case, we are still not accessing the native expression levels for the following reasons 1/ chromosomal level of toxicity were not explored here and 2/ only the toxicity of the coding sequence but not the full mRNA was tested. Indeed, we do not know the exact sequence of the hok homolog mRNAs and this is beyond the scope of the study. These remarks were clearly added in the discussion.
We agree that the sentence "Hok peptides originating from chromosomes are as toxic as those from plasmids" was too strong and we have added the caveats of our experimental design in the discussion. While we indeed did not compare the toxicity of the peptides, we still showed that chromosomal Hok can be toxic upon overexpression, which would not be the case if the sequences were degenerated.
The reviewer also suggests the use of the FASTBAC-Seq method, that we previously used to study Hok from the R1 plasmid, which is a method to study toxic type I toxins at the native expression level. While FASTBAC-Seq identifies loss-of-function mutants of the systems, it does not allow to determine a difference of toxicity between systems per se. In addition, FASTBAC-Seq was always done in the context of the full mRNA, not only the coding sequence, and these sequences are presently unknown for most homologs.
Other comments:
__R1_C3: __a) There is barely any discussion of the Sok component (RNA antitoxin) of the homologues; why is that? Could you please discuss Sok differences across the homologues, or at least explain why this is not discussed at all in the paper (e.g., in the discussion)?
Authors' answer to R1_C3: __It is not trivial to identify the Sok RNA sequence, this is why it was not done in this study, a paragraph was added in the discussion explaining this. __
__R1_C4: __b) In the results section, the Hok clusters are referred to as 62 in number ("Because Hok sequences were too short and variable to construct a meaningful phylogenetic tree, we clustered the Hok sequences with a 60% identity threshold and obtained 62 clusters"), but then in the discussion section, the cluster number becomes 74 ("We highlighted the high sequence variability within Hok peptides by obtaining a total of 74 clusters with 60% identity (Fig. S7)."). Which one is the right number, and why is there a discrepancy?
Authors' answer to R1_C4: We apologize for the discrepancy between the number. The first number corresponded to the Hok hits from the refSeq and we then added the Hok hits from the plasmid and virus databases (performed later in the manuscript). We clarified this information both in the result and discussion texts (61 clusters from RefSeq and 79 in total, 74 was a typo).__ __
__R1 Significance: __The most well-clarified aspect of the paper presented here is the distribution of Hok homologues, with the novel aspect of the location in which the hok/Sok T1TAs reside (i.e., chromosome, plasmid, or phage). There is room for the molecular genetics part to be developed further, as I discussed earlier, however this study is the most up-to-date characterization of the diversity of Hok homologues, and will be of interest to the T1TA and the general toxin-antitoxin field.
__Reviewer #2 (R2) __
R2 General statement: The authors examined how the Hok toxins are spread across bacterial genomes. The manuscript including its figures is hard to read and understand. I commented figure 1 in details, but similar comments apply to the other figures. Overall, the data lack clarity and precision. Finding information about sequences, clusters in the supplementary materials was not easy. The manuscript should be thoroughly revised. In addition, I believe that other aspects should be developed to expand the interest of the study, such as the co-occurrence of multiple systems in chromosomes, on plasmids and whether they are able to crosstalk. This might provide some evolutionary insights into the biology of these toxins.
__Authors' answer to R2 General statement: __We designed all figures according to established standards for scientific data visualization, although we recognize that different presentations may work better for different audiences. In our detailed response to Figure 1A, we explain how UpSet plots are constructed and interpreted, which we hope clarifies the visualization approach for the full dataset. We are open to discussing specific improvements if the reviewer has suggestions for enhanced clarity. To address concerns about accessibility, we want to clarify that all sequences are compiled in Table S1 with their clus100 identifiers, making them easy to locate. We are open to reorganizing supplementary materials if a different structure would be more user-friendly. Finally, we agree that an extensive analysis of co-occurrences and crosstalks would be valuable. However, predicting crosstalk bioinformatically for all genomes presents challenges, as it would require predicting RNA:RNA interactions between hok mRNA and Sok sequences, which are currently unknown. Given these limitations, this analysis was beyond the scope of the current study.
R2_C1: The introduction lacks information regarding the Hok protein (size, structure prediction, localization) as well as a bit of explanation about the reason of looking at these toxins. The description of the potential roles should be a bit expanded.
Authors' answer to R2_C1: Following the comment from the reviewer, we have provided additional information about Hok in the introduction.
__R2_C2: __When the authors talk about 'loci', they mean genes encoding Hok homologs if I understand correctly. They did not look for the Sok sequences (hok-sok loci).
__Author's answer to R2_C2: __Indeed, we did not look for the Sok sequences and we are only describing Hok homologs loci, that could either encode or lack a Sok homolog.
__R2_C3: __It is not clear what the authors did with the sequences for which they could not detect a start codon and a SD (although it is unusual to refer to SD in the context of protein sequence)
Authors' answer to R2_C3: The peptides were annotated by extending the initial hit until the first start codon. Therefore, all annotated peptides have a start codon. Shine-Dalgarno sequences were annotated when confidently predicted, to provide additional information. Sequences were not excluded based on the presence or absence of the SD.
__R2_C4: __Figure 1A is not clear. The total of the bars equal 32,532 which is the number of 'loci' detected by the combination of the different methods. However, it is not clear to me how many are redundant. For instance, I suppose that all the 8483 sequences that were retrieved using blastn and Infernal were retrieved using MMseqs2, blastn and Infernal. So, what is the actual number of sequences that were found? When the authors talk about 1264 distinct peptides, what do they mean? What are the numbers on the X axis (18209, 2260, 27728)?
Author's answer to R2_C4: Figure A1 is a very typical "UpSet" plot, as indicated in the legend (A. Lex, N. Gehlenborg, H. Strobelt, R. Vuillemot and H. Pfister, "UpSet: Visualization of Intersecting Sets," in IEEE Transactions on Visualization and Computer Graphics, vol. 20, no. 12, pp. 1983-1992, 31 Dec. 2014, doi: 10.1109/TVCG.2014.2346248). Those plots are a data visualization method for showing data with more than two intersecting sets. The Hok sequence hits were obtained by 3 different methods stated on the rows (MMseqs2, blastn and Infernal, therefore the number 18209 is the number of hits by the MMseqs2, 22680 the number of hits by blastn and 27728 the number of hits by Infernal). The columns show the intersections between these three sets. For example, the mentioned 8483 sequences (second column) were only found by blastn and Infernal but not by MMseqs2. The actual total number of sequences found is indeed 32 532. The 1264 distinct peptides are peptides with different sequences. After removing false positives, degenerated sequences and small peptides, we obtained 1264 unique Hok sequences that are found in the 32532 bacterial loci.
__R2_C5: __About Infernal: first the authors are stating that only 8% of the sequences are lost when not considering the mRNA structure - which they seem to consider as negligeable. Then in the next section, they state that Infernal is the best tool at identifying clusters that are not detected otherwise. Seems a bit contradictory.
__Authors' answer to R2_C5: __We appreciate the reviewer pointing out this apparent contradiction, we have clarified this part in the revised manuscript. Infernal uses both sequence and structure information simultaneously for homology detection. While only 8% of Infernal's hits are detected uniquely when structural information was considered, these sequences account for 9 additional clusters with notably high sequence diversity, which would otherwise have been undetected. Therefore, we believe that Infernal is the best tool to capture novel cluster diversity.
__R2_C6: __Cluster determination. The threshold was put at 60% identity. What is the rationale for the 60% identity? Given that the Hok sequences (like toxins and antitoxins from TA systems in general) are highly variable, this leads to a high number of clusters. I'm not sure of the relevance of these clusters. Are there any other criteria to define clusters?
Authors' answer to R2_C6: We selected 60% identity as a balance between capturing sequence diversity and generating interpretable results. We also tested 70, 80 and 90% and obtained 128, 221, 377 clusters, respectively, which would be too many for a meaningful visualization and interpretation. The best clustering method would be constructing a phylogenetic tree. However, as explained in the discussion, because the high sequence diversity prevented the construction of a reliable phylogenetic tree, clustering was used as an alternative strategy to identify and interpret patterns of sequence variability.
__R2_C7: __The authors claim that most of the Hok diversity is found on chromosomes. However, the number of chromosomal Hok is higher than that located on plasmids, which might be related to the different sizes of the different replicons ie, chromosomes being larger than plasmids. Is there a way to normalize by determining the density per size?
Authors' answer to R2_C7: We do not claim that chromosomes contain most of Hok diversity, as this would be indeed influenced by biases in the databases. We are just describing that we found most of the diversity in chromosomes, but we cannot conclude whether this is a true representation of the frequencies in nature.__ __
R2_C8: '46 of the 62 clusters contained 10 or less distinct sequences and might be in the process of degenerating'. The authors also linked this with SD detection. Please explain. From what was indicated earlier, I understand that sequences with premature stop codons or short sequences (Authors' answer to R2_C8: We did not remove sequences for which we could not predict the SD. Indeed, lacking SD is a sign that the hok mRNA might not be able to play its biological role and would be indicative that the sequences have degenerated. To evaluate this hypothesis, we experimentally tested 5 sequences without a predicted SD and two of those were not toxic (see Table S2). In order to assess if the low abundant clusters contained degenerated sequences we experimentally tested representatives from some of the clusters with only one Hok CDS and found most of them to be toxic.
R2_C9: 'Only 7.3% of the unique sequences were found on both plasmids and chromosomes'. From this observation, the authors conclude that 'there is little stable transfer from chromosomes to plasmids or vice-versa'. I don't understand what this means. Do they mean identical sequences? The fact that sequences differ from chromosomes to plasmids does not rule out 'stable transfer'. What do they actually mean by stable transfer? Once the gene is horizontally transferred, it is fixed and vertically transmitted? Same comments apply to the inter-genera horizontal transfer by plasmids.
__Authors' answer to R2_C9: __Due to the impossibility of constructing a reliable phylogenetic tree, we used identity of sequences across different localizations or genera as our marker for recent, stable transfer events. We define stable transfer as the persistence of sequences in an unchanged form following horizontal transfer; long enough to be detected in current databases. Our approach likely underestimates total transfer events, as sequences accumulating mutations after transfer would not be captured. We would expect to observe numerous identical sequences across plasmids and chromosomes if frequent exchange were occurring, unless rapid mutation after the transfer prevented their detection as identical sequences. We have added a sentence to clarify this in the manuscript and removed the term stable transfer.
__R2_C10: __I don't understand the next section about 'family'. What do the authors mean about 'family'? Genera? The same apply to the next section about the Y to C recoding. Did the authors do point mutations in the conserved amino acids/codons to test whether they are important for toxicity? Some Hok variants lacks some of the conserved amino acids and are toxic (under overexpression conditions in Salmonella). What about T18, C31 and E42?
Authors' answer to R2_C10: Families (Enterobacteriaceae, Vibrionaceae etc... ) and genera (Escherichia, Salmonella etc...) refer to the taxonomic categories. Following the reviewer comment, we experimentally assessed the toxicity of Hok from R1 plasmid after mutating the conserved amino acids to alanine residues. All the mutants were found to be toxic under our expression conditions.
__R2_C11: __The prevalence of Hok in chromosomes or on plasmids might depend on various confounding parameters, such as the size, number of sequences available among others. The authors should find methods to correct for all that.
Authors' answer to R2_C11: Normalization would indeed be needed if we were comparing the prevalence on chromosomes vs the prevalence on plasmids. Here, we do not claim that Hok homologs are more prevalent in plasmid or chromosomes and only describe where we found them.
__R2_C12: __Link with defense systems. The threshold was set at 20 kb. Why this threshold?
Authors' answer to R2_C12: The size of defense islands in a previous report was approximately 40 kb, by setting up a 20 kb threshold we searched for defense systems in a region of 40 kb adjacent to each of the homologs (https://doi.org/10.1126/science.aar4120). If the specific homolog was part of a defense island we would expect that it is less than 20 kb apart from any defense system.
__R2 Significance: __The paper in its current state appears to serve the role of a data repository rather than a thorough and original analysis. It requires extensive revisions before it can be of interest to experts in the toxin-antitoxin field.
__ ____Reviewer #3 (R3): __
R3 General statement: In the manuscript, "The Hok bacterial toxin: diversity, toxicity, distribution and genomic localization," by Escalera-Maurer et al., investigate the distribution of Hok type I toxin proteins across bacterial species. The Hok-Sok type I toxin-antitoxin system was first described on plasmids where it serves to maintain the plasmid in a population of bacterial cells: translation of the hok mRNA is prevented via the small antitoxin RNA Sok. Upon plasmid loss, with no new transcription of sok, the highly stable hok mRNA is translated into a small protein, killing the plasmid-less cell. Homologues to the system were identified in the chromosome of E. coli in the 1990s, and subsequent analyses have identified identical systems in other bacterial chromosomes, though they are close relatives to E. coli. Given the increased number of bacterial genomes sequenced, the group examined how widespread Hok may be across bacteria. They used a combination of BLASTn, MMseqs2 (protein) and Infernal (RNA) to identify, as best possible, all possible homologs. They then used sequence identity cut-offs to form Hok "clusters," and identified key features of the cluster as well as tested toxicity of overproduction of 31 homologs in a strain of Salmonella. Overall, though a variety of bioinformatic predictions and analyses, the manuscript identifies an expanded number of Hok members not previously identified and broaden the species it is found in, supported that Hok is not associate with defense systems, and provides additional support that horizontal transfer of hok genes is likely via plasmids (where hok is presumed to have originated).
Major comments: There are some areas of the text that are a bit too definitive (these can be fixed or better explained in the text) and a few questions raised about the analyses and interpretations.
Authors' answer to R3 Major Comment: As suggested by the reviewer, we rephrased parts of the manuscript.
__These are the specific comments: __
Introduction R3_C1: First paragraph: "Toxin production leads to the death of the cell encoding it" For many chromosomally encoded systems, toxicity has only been observed via artificial overexpression. This is an important point, as for many systems, a true biological function remains unknown. Further, add caveats regarding toxin function (for systems with validated function, they are involved in...). Again, there are still many questions for many t-at systems, in particular the Type I systems.
__Authors' answer to R3_C1: __Indeed, the function of type 1 TA, in particular chromosomal ones, is still a matter of debate. While for hok/Sok R1, we previously showed death by expression at the chromosomal level, this was not shown for all TA (Le Rhun et al., NAR, 2023). We added that it could lead to the death or growth arrest of the cell instead and added the reviewer changes to for the function part.
__R3_C2: __Introduction: type I's are more narrow in distribution, but much of this is due to their size and lack of biochemical domains. Again, please clarify more here.
__Authors' answer to R3_C2: __We added the reviewer suggestion to the text.
__R3_C3: __Introduction: while Hok's have been found on chromosomes, in E. coli strains, there is clear evidence that many are inactive. This comes up in the discussion, but it is worth including briefly in the introduction.
Authors' answer to R3_C3: We have now added in the introduction that in the K12 laboratory strain, most chromosomal hok/Sok were found to be inactive.
__R3_C4: __For the predicted transmembrane domain: it would be worth to include a box/indication as to where that is within the peptide (with the understanding it may not be exact). Is there more/less variation here? I'm assuming all clusters/family have a predicted TM domain?
__Authors' answer to R3_C4: __When predicting the TM domain using DeepTMHMM - 1.0 prediction (https://services.healthtech.dtu.dk/services/DeepTMHMM-1.0/), 227 out of the 1264 unique Hok sequence are predicted to have a TM (transmembrane), 7 a SP (signal peptide) and a TM and 1025 have a SP. When predicting the TM of the consensus sequence (most abundant amino-acid) shown in Fig. 1D, region A8 to L25 is predicted to be inserted in the membrane, with the Nterm inside and Cterm outside.
__R3_C5: __What is the cutoff for being a Hok? Did they take the "last hit" and use that in additional searches to see if more appeared? If that was done, and the search was exhaustive, this really important to add for the reader.
Authors' answer to R3_C5: The MMseqs2 search was performed using 5 iterations as indicated in the M&M, meaning that the hits of the one search were used to search the database again five time in a raw. Importantly, an attempt to increase the number of iterations to 10 did not significantly increase the number of hits. Therefore, at least for the MMseqs2 search in the RefSeq database, we are close to being exhaustive.
__R3_C6: __Figure S4: the authors state that there was no difference in the degree of toxicity between the clusters. There do appear to be some peptides tested that at the arabinose concentration used did not repress growth as immediately as others. If higher arabinose concentration is used, does that eliminate these differences? OR are many of these suppressors-if diluted back again, do they grow as if they are non-toxic in arabinose?
Authors' answer to R3_C6: As suggested by Reviewer 1 (R1_C2), we performed titration of arabinose in a system overexpressing araE in a ΔaraBAD but were not able to find difference of toxicity in our conditions, see also our answer to R1_C2.
__R3_C7: __Discussion: "because non-functional homologs are expected to quickly accumulate mutations..." is a bit problematic. Hok is highly regulated-as are some of the other well-described type I toxins. In MG1655, while the coding sequence may be intact, there are other mutations and/or insertion elements that prevent expression (and be extension, function. Given the lack of consensus data for type Is, it is best to provide more context for this. If the authors wish to argue that they should quickly accumulate mutations, it would be good to provide additional rates/evidence (even for other loci) from the Enterobacteriaceae.
__Authors' answer to R3_C7: __We agree this statement might need to be supported further. We have removed this sentence to address this concern.
__Minor comments: __
__R3_C8: __For the sequences used in the search: please provide the sequence used in addition to the reference to the T1TAdb. Was the full-length hok mRNA, including mok, used? Please provide the nucleic acid sequence (and include description of whether full-length, etc.) in Materials and Methods or in Supplemental.
__Authors' answer to R3_C8: __Sequences and code were deposited on https://gitub.u-bordeaux.fr/alerhun/Escalera-Maurer_2025. This files named curated_Hok.fasta and hok.fa, corresponding to Hok protein and mRNA sequences respectively are available in the file "T1TAdb input".
__R3_C9: __60% identity was used for clustering. Did this become a problem-meaning separation of same property amino acid?
__Authors' answer to R3_C9: __We checked amino acid signatures for each cluster (Fig S2), but could not find anything relevant.
__R3_C10: __Fig. S2: for the clusters shown, please add in HokB, HokE, etc., to better correspond to Figure 1 in the main text.
__Authors' answer to R3_C10: __The clusters were annotated according to the suggestion.
__R3_C11: __Fig S1: this figure is challenging to orient-what are the numbers (8_10_85)?
Authors' answer to R3_C11: The figure was generated using the CLANS tool, with each unique sequence retrieved by our analysis shown as a dot. Hok homologous sequences are in red and cluster together, the outlier clusters are annotated with the numbers corresponding to their 60% identity cluster. We understand that separating the number using an underscore could lead to confusion, therefore we have now separated the numbers using a coma.
__R3_C12: __Please make a separate table or sheet for the experimentally tested peptides. Table S1 is quite large and a separate table/sheet would make this easier to find. If possible, please give the files names a more descriptive title (Table S1 in the name for example). This may be an issue with Review Commons but the individual file names were non-descript and the descriptions on the webpage did not indicate what the file contained.
__Authors' answer to R3_C12: __We named the files Table S1 and File_S1 to S7. We added a table S2 with the experimentally tested peptides. Note that identical peptides can be sometime found in several bacterial loci.
__R3_C13: __Figure S9: the black arrow for Hok is hard to see-it appears that the long grey bar going through multiple loci is indicative of Hok. Perhaps label this differently to make it easier on the reader (the line initially seemed to be a formatting issue and not indicative of the position of Hok.
__Authors' answer to R3_C13: __We have now added a new label to indicate where is Hok, and clarified it in the figure legend.
__R3_C14: __While the authors focused on Hok for this approach, which is fine and appropriate, can they comment at all about where mok is there in these new clusters/sub-families? Sok potential?
__Authors' answer to R3_C14: __We added a paragraph about Mok in the discussion.
__R3 Significance: __Overall the paper is a sound bioinformatic exercise and is improved with the testing of numerous "new" Hok proteins. Most of the comments can be done with some clarifications and maybe some additional analyses and/or verification which should take minimal time. The authors are over-emphatic at points as indicated and need to be more careful and precise with their language.
In terms of advancement, it advances the distribution of these systems and adds to the depth of sub-classes. The audience will be more specialized to those who study these systems.
Expertise: I have been studying type I toxin-antitoxin systems since the mid-2000s. We published a study examining (and mentioned well by this article!) the distribution in chromosomes of type I toxin-antitoxin systems, identified brand-new systems (that were chromosomally-limited at the time). My lab has continued to study regulation of type I toxins and distribution of chromosomally-only-encoded systems (so not Hok).
-
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Referee #3
Evidence, reproducibility and clarity
In the manuscript, "The Hok bacterial toxin: diversity, toxicity, distribution and genomic localization," by Escalera-Maurer et al., investigate the distribution of Hok type I toxin proteins across bacterial species. The Hok-Sok type I toxin-antitoxin system was first described on plasmids where it serves to maintain the plasmid in a population of bacterial cells: translation of the hok mRNA is prevented via the small antitoxin RNA Sok. Upon plasmid loss, with no new transcription of sok, the highly stable hok mRNA is translated into a small protein, killing the plasmid-less cell. Homologues to the system were identified in the chromosome of E. coli in the 1990s, and subsequent analyses have identified identical systems in other bacterial chromosomes, though they are close relatives to E. coli. Given the increased number of bacterial genomes sequenced, the group examined how widespread Hok may be across bacteria. They used a combination of BLASTn, MMseqs2 (protein) and Infernal (RNA) to identify, as best possible, all possible homologs. They then used sequence identity cut-offs to form Hok "clusters," and identified key features of the cluster as well as tested toxicity of overproduction of 31 homologs in a strain of Salmonella. Overall, though a variety of bioinformatic predictions and analyses, the manuscript identifies an expanded number of Hok members not previously identified and broaden the species it is found in, supported that Hok is not associate with defense systems, and provides additional support that horizontal transfer of hok genes is likely via plasmids (where hok is presumed to have originated).
Major comments: There are some areas of the text that are a bit too definitive (these can be fixed or better explained in the text) and a few questions raised about the analyses and interpretations. These are the specific comments:
Introduction
First paragraph: "Toxin production leads to the death of the cell encoding it" For many chromosomally encoded systems, toxicity has only been observed via artificial overexpression. This is an important point, as for many systems, a true biological function remains unknown. Further, add caveats regarding toxin function (for systems with validated function, they are involved in...). Again, there are still many questions for many t-at systems, in particular the Type I systems. Introduction: type I's are more narrow in distribution, but much of this is due to their size and lack of biochemical domains. Again, please clarify more here.
Introduction: while Hok's have been found on chromosomes, in E. coli strains, there is clear evidence that many are inactive. This comes up in the discussion, but it is worth including briefly in the introduction.
For the predicted transmembrane domain: it would be worth to include a box/indication as to where that is within the peptide (with the understanding it may not be exact). Is there more/less variation here? I'm assuming all clusters/family have a predicted TM domain?
What is the cutoff for being a Hok? Did they take the "last hit" and use that in additional searches to see if more appeared? If that was done, and the search was exhaustive, this really important to add for the reader.
Figure S4: the authors state that there was no difference in the degree of toxicity between the clusters. There do appear to be some peptides tested that at the arabinose concentration used did not repress growth as immediately as others. If higher arabinose concentration is used, does that eliminate these differences? OR are many of these suppressors-if diluted back again, do they grow as if they are non-toxic in arabinose?
Discussion: "because non-functional homologs are expected to quickly accumulate mutations..." is a bit problematic. Hok is highly regulated-as are some of the other well-described type I toxins. In MG1655, while the coding sequence may be intact, there are other mutations and/or insertion elements that prevent expression (and be extension, function. Given the lack of consensus data for type Is, it is best to provide more context for this. If the authors wish to argue that they should quickly accumulate mutations, it would be good to provide additional rates/evidence (even for other loci) from the Enterobacteriaceae.
Minor comments:
For the sequences used in the search: please provide the sequence used in addition to the reference to the T1TAdb. Was the full-length hok mRNA, including mok, used? Please provide the nucleic acid sequence (and include description of whether full-length, etc.) in Materials and Methods or in Supplemental.
60% identity was used for clustering. Did this become a problem-meaning separation of same property amino acid? Fig. S2: for the clusters shown, please add in HokB, HokE, etc., to better correspond to Figure 1 in the main text.
Fig S1: this figure is challenging to orient-what are the numbers (8_10_85)?
Please make a separate table or sheet for the experimentally tested peptides. Table S1 is quite large and a separate table/sheet would make this easier to find. If possible, please give the files names a more descriptive title (Table S1 in the name for example). This may be an issue with Review Commons but the individual file names were non-descript and the descriptions on the webpage did not indicate what the file contained.
Figure S9: the black arrow for Hok is hard to see-it appears that the long grey bar going through multiple loci is indicative of Hok. Perhaps label this differently to make it easier on the reader (the line initially seemed to be a formatting issue and not indicative of the position of Hok.
While the authors focused on Hok for this approach, which is fine and appropriate, can they comment at all about where mok is there in these new clusters/sub-families? Sok potential?
Significance
Overall the paper is a sound bioinformatic exercise and is improved with the testing of numerous "new" Hok proteins. Most of the comments can be done with some clarifications and maybe some additional analyses and/or verification which should take minimal time. The authors are over-emphatic at points as indicated and need to be more careful and precise with their language.
In terms of advancement, it advances the distribution of these systems and adds to the depth of sub-classes.
The audience will be more specialized to those who study these systems.
Expertise: I have been studying type I toxin-antitoxin systems since the mid-2000s. We published a study examining (and mentioned well by this article!) the distribution in chromosomes of type I toxin-antitoxin systems, identified brand-new systems (that were chromosomally-limited at the time). My lab has continued to study regulation of type I toxins and distribution of chromosomally-only-encoded systems (so not Hok).
-
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Referee #2
Evidence, reproducibility and clarity
The authors examined how the Hok toxins are spread across bacterial genomes. The manuscript including its figures is hard to read and understand. I commented figure 1 in details, but similar comments apply to the other figures. Overall, the data lack clarity and precision. Finding information about sequences, clusters in the supplementary materials was not easy. The manuscript should be thoroughly revised. In addition, I believe that other aspects should be developed to expand the interest of the study, such as the co-occurrence of multiple systems in chromosomes, on plasmids and whether they are able to crosstalk. This might provide some evolutionary insights into the biology of these toxins.
Introduction:
The introduction lacks information regarding the Hok protein (size, structure prediction, localization) as well as a bit of explanation about the reason of looking at these toxins. The description of the potential roles should be a bit expanded.
Results:
When the authors talk about 'loci', they mean genes encoding Hok homologs if I understand correctly. They did not look for the Sok sequences (hok-sok loci).
It is not clear what the authors did with the sequences for which they could not detect a start codon and a SD (although it is unusual to refer to SD in the context of protein sequence)
Figure 1A is not clear. The total of the bars equal 32,532 which is the number of 'loci' detected by the combination of the different methods. However, it is not clear to me how many are redundant. For instance, I suppose that all the 8483 sequences that were retrieved using blastn and Infernal were retrieved using MMseqs2, blastn and Infernal. So, what is the actual number of sequences that were found? When the authors talk about 1264 distinct peptides, what do they mean? What are the numbers on the X axis (18209, 2260, 27728)?
About Infernal: first the authors are stating that only 8% of the sequences are lost when not considering the mRNA structure - which they seem to consider as negligeable. Then in the next section, they state that Infernal is the best tool at identifying clusters that are not detected otherwise. Seems a bit contradictory.
Cluster determination. The threshold was put at 60% identity. What is the rationale for the 60% identity? Given that the Hok sequences (like toxins and antitoxins from TA systems in general) are highly variable, this leads to a high number of clusters. I'm not sure of the relevance of these clusters. Are there any other criteria to define clusters?
The authors claim that most of the Hok diversity is found on chromosomes. However, the number of chromosomal Hok is higher than that located on plasmids, which might be related to the different sizes of the different replicons ie, chromosomes being larger than plasmids. Is there a way to normalize by determining the density per size?
'46 of the 62 clusters contained 10 or less distinct sequences and might be in the process of degenerating'. The authors also linked this with SD detection. Please explain. From what was indicated earlier, I understand that sequences with premature stop codons or short sequences (<40aa) were removed from the analysis earlier. Lacking an SD is a sign of decay? Were these sequences lacking SD not discarded before starting the analysis? Did the authors experimentally validate some of these sequences?
'Only 7.3% of the unique sequences were found on both plasmids and chromosomes'. From this observation, the authors conclude that 'there is little stable transfer from chromosomes to plasmids or vice-versa'. I don't understand what this means. Do they mean identical sequences? The fact that sequences differ from chromosomes to plasmids does not rule out 'stable transfer'. What do they actually mean by stable transfer? Once the gene is horizontally transferred, it is fixed and vertically transmitted? Same comments apply to the inter-genera horizontal transfer by plasmids.
I don't understand the next section about 'family'. What do the authors mean about 'family'? Genera? The same apply to the next section about the Y to C recoding. Did the authors do point mutations in the conserved amino acids/codons to test whether they are important for toxicity? Some Hok variants lacks some of the conserved amino acids and are toxic (under overexpression conditions in Salmonella). What about T18, C31 and E42?
The prevalence of Hok in chromosomes or on plasmids might depend on various confounding parameters, such as the size, number of sequences available among others. The authors should find methods to correct for all that.
Link with defense systems. The threshold was set at 20 kb. Why this threshold?
Significance
The paper in its current state appears to serve the role of a data repository rather than a thorough and original analysis. It requires extensive revisions before it can be of interest to experts in the toxin-antitoxin field.
-
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Referee #1
Evidence, reproducibility and clarity
Here, Escalera-Maurer and colleagues, present an up-to-date distribution of homologues of Hok toxic proteins belonging to the well-annotated, but otherwise functionally obscure, hok/Sok type I toxin-antitoxin system, across the RefSeq database. Although such computational analyses have been done in the past, the authors here find many more hok homologs than described before, and they categorise their distribution based on whether they are encoded on chromosomes, plasmids, or (pro)phages. These computational analyses are in general tricky with T1TAs, as their toxins are quite short (~50 amino acids, as is the case for Hok), which is why the authors here used three separate approaches to expand their search (nucleotide-level BLAST, protein-homology, or both combined with Infernal). The authors cluster the Hok homologues they find based on a 60% sequence identity cut-off (expanding the known clusters in the process), and proceeded to test 31 candidates belonging to 15 sequence-clusters for their toxicity in Salmonella Typhimurium LT2, showing that 30/31 were toxic upon induction. An interesting finding from their endeavours is that hok/Sok homologues are enriched within prophages and large plasmids, but are not enriched near bacterial anti-phage defense systems (in contrast to the SymE/SymR T1TA). The findings suggest that hok/Sok are indeed sometimes linked to phage and plasmid biology, although they might not be antiphage defenses per se (they have been clearly shown in the past to be addiction modules, and this is still clearly true).
My expertise lies towards the experimental side of the authors' work, I thus cannot comment on the accuracy/robustness of the computational analyses performed here. The authors do a fine job in clearly stating their findings overall; I could follow most of the conclusions, and I deemed that most of them were supported by their work. Additionally, I find that this paper is a missed opportunity to uncover even more novel biology connected to the interesting hok/Sok T1TAs. The paper does not provide a new framework to think about what is the function of the chromosomal/prophage hok/Sok T1TA systems, although I realize that this is very difficult to accomplish, especially when considering that hok/Sok systems have been around in the literature for almost 40 years.
My major comment is in regard to the Hok toxicity assays (Fig. 2). The authors state in the discussion that "Hok peptides originating from chromosomes are as toxic as those from plasmids", but I believe that the way that they tested their constructs might not have allowed them to see toxicity differences between the two groups. Specifically, using the multi-copy plasmid pAZ3 (pBR322 origin of replication; ~15-20 plasmid copies per chromosome) to induce the different Hok toxin homologues in Salmonella Typhimurium LT2 with arabinose might have masked toxicity differences that would otherwise be apparent on the chromosomal expression-level.
Some of the authors themselves have previously used the FASTBAC-Seq method to study the Hok homologue from plasmid R1, a useful technique during which a toxin is integrated in the chromosome, in order to study their toxicity under natural levels of expression. I believe that an ideal scenario would be to apply FASTBAC-seq to some of the 31 Hok homologues described here (e.g., a subset of plasmidic vs chromosomal Hok homologues) to shed light on potential toxicity differences between the Hok clusters. This would increase the value of the presented study.
Alternatively, the authors could employ an L-arabinose concentration gradient to titrate the expression levels of the Hok toxins in order to potentially see different toxicity levels from the different homologues. However, this is not going to work in the system as they are using it now for two reasons:
a) the S. Typhimurium LT2 (STm) used here has its arabinose utilization operon intact (araBAD), which means that Salmonella can catabolize arabinose to use it as a carbon source. This catabolization process interferes with the arabinose induction (i.e., Salmonella eats arabinose instead of using it as the Hok inducer). To ameliorate this, the authors could delete the araBAD operon in STm, rendering STm incapable of catabolizing arabinose, and repeat the experiments in that strain. Or use E. coli BW25113 as the expression host, which already has the araBAD operon deleted (it is not clear to me why the different Hok homologues would not be toxic in E. coli, as the different Hok homologues are widely diverse in sequence, as the authors found here).
b) Even with the araBAD operon deleted, the arabinose induction would be bimodally on or off in the population, due to the bimodal expression of the arabinose transporter (AraE; see Khlebnikov et al., 2002). This would again not allow for titratable arabinose-inducible expression from different concentrations of arabinose. The solution for this would be to co-express a separate plasmid with araE, which would render every cell the same in regards to arabinose permeability, and thus the system would be titratable (as explained in Khlebnikov et al., 2002).
Therefore, if the authors would be interested to go towards this route, they would have to first delete the araBAD from STm, then transform STm with an araE plasmid, and redo the experiments. In addition, I would propose to the authors to use the drop plate method (agar plate-based), which is more sensitive compared to the liquid assays employed here.
Having said all that, I understand that all this experimental work would be strenuous and time-consuming, and although I would like to see it happen, this is not my paper. I would be content therefore if the authors toned down the claim that plasmidic vs chromosomal Hok homologues have the same toxicity, and discuss that chromosomal levels of toxicity are an important caveat that has not been explored here.
Other comments:
a) There is barely any discussion of the Sok component (RNA antitoxin) of the homologues; why is that? Could you please discuss Sok differences across the homologues, or at least explain why this is not discussed at all in the paper (e.g., in the discussion)?
b) In the results section, the Hok clusters are referred to as 62 in number ("Because Hok sequences were too short and variable to construct a meaningful phylogenetic tree, we clustered the Hok sequences with a 60% identity threshold and obtained 62 clusters"), but then in the discussion section, the cluster number becomes 74 ("We highlighted the high sequence variability within Hok peptides by obtaining a total of 74 clusters with 60% identity (Fig. S7)."). Which one is the right number, and why is there a discrepancy?
Significance
The most well-clarified aspect of the paper presented here is the distribution of Hok homologues, with the novel aspect of the location in which the hok/Sok T1TAs reside (i.e., chromosome, plasmid, or phage). There is room for the molecular genetics part to be developed further, as I discussed earlier, however this study is the most up-to-date characterization of the diversity of Hok homologues, and will be of interest to the T1TA and the general toxin-antitoxin field.
-
-
en.wikipedia.org en.wikipedia.org
-
In mass communication, digital media is any communication media that operates in conjunction with various encoded machine-readable data formats. Digital content can be created, viewed, distributed, modified, listened to, and preserved on a digital electronic device, including digital data storage media (in contrast to analog electronic media) and digital broadcasting.
Webpage is not complex at all, but relatively simple and easy to understand. You have a term, and the definition right below it. Whats great here is that any other terms within the definition will include a hyperlink to understand that term as well. Making the flow process easier for consumers.
-
-
human.libretexts.org human.libretexts.org
-
An example of a metaphor is when someone says, "My love, you are a rose." An example of a simile is "My darling, you are like a rose."
Metaphor: relating things to another thing Simile: Same thing but using the word like
-
Study the time period which is also part of the setting
This is important because sometimes authors might leave out context that would seem obvious at the time but needs more explaining now.
-
Are any elements repeated that may suggest a theme?
The reason why I choose this quote is because a theme could mean differently for different people. AN author could have a fix theme for a story but for others, it could mean something else. Which is why themes are different depending on the readers belief and interpretation of the story.
-
How does the time period affect the language, atmosphere, or social circumstances of the novel?
The reason why I choose this quote is because dialogue is critical to understanding the story. If a character is speaking differently than they should in a specific time period, then the story becomes unrealistic unless it's non-fiction. But regardless, dialogue sets the tone of the story.
-
“who, what, when, where, how?”
basic questions, but that help give meaning and provide understanding to the full story :)
-
How is the plot structured? Is it linear and chronological or does it move back and forth?
Something I never really thought about but now reflecting I find very important to how a plot is perceived
Annotators
URL
-
-
open.library.okstate.edu open.library.okstate.edu
-
Adapt your writing to meet the needs, interests, culture, and background of those who will be reading your documents.
This statement stuck out to me because I feel as though many writers don't keep the readers in mind when they have a story to tell.Usually they convey the message in the most easiest way they know how but this might not grab your readers attention. Thinking about what interest your audience will make your writing stand out.
-
-
Local file Local file
-
In both countries, censorship was imposed, the dissemination of newswas manipulated, and propaganda was issued; on the whole, however, thiswas a break with peacetime practice for Britons but a continuation of it forGermans.
Quote for cinema's? While () as the land of (), Britain too utilised censorship and propaganda to ensure morale. Neverthless, like Germany, () British cinema was not (), with films often utilising () to present messages. As such, we could argue that British and German civilians on the home front experienced a similar sort of experience in the cinema, where comedies and () were appreciated and increasingly made. Nevertheless, while Britain imposed newsreels within cinema, Germans saw themselves with (by blah) over 40 mins of news, something that the British did not face.
-
-
chem.libretexts.org chem.libretexts.org
-
Practice
on question 2 how do these numbers relate to the well.I understand the repulsion between nuclei and the attraction of electrons and nuclei and how this relates to equilibrium, but I do not understand how the numbers relate back to the model
-
-
www.americanyawp.com www.americanyawp.com
-
There was a South of slavery and secession — that South is dead. There is a South of union and freedom — that South, thank God, is living, breathing, growing every hour.
yes that south is had die, but that south created some of the worst conditions for blacks in america by developing the white supremacy thoughts
-
-
-
Author response:
The following is the authors’ response to the original reviews.
Reviewer #1 (Public review):
(1) The authors devote significant effort to characterizing the physical interaction between Bicc1 and Pkd2. However, the study does not examine or discuss how this interaction relates to Bicc1's well-established role in posttranscriptional regulation of Pkd2 mRNA stability and translation efficiency.
The reviewer is correct that the present study has not addressed the downstream consequences of uthis interaction considering that Bicc1 is a posttranscriptional regulator of Pkd2 (and potentially Pkd1). We think that the complex of Bicc1/Pkd1/Pkd2 retains Bicc1 in the cytoplasm and thus restrict its activity in participating in posttranscriptional regulation (see Author response image 1). We, however, do not yet have data to support this and thus have not included this model in the manuscript. Yet, we have updated the discussion of the manuscript to further elaborate on the potential mechanism of the Bicc1/Pkd1/Pkd2 complex.
We have updated the discussion to include a discussion on the potential consequences on posttranscriptional regulation by Bicc1.
Author response image 1.
Model of BICC1, PC1 and PC2 self-regulation. In this model Bicc1 acts as a positive regulator of PKD gene expression. In the presence of ‘sufficient’ amounts of PC1/PC2 complex, it is tethered to the complex and remains biologically inactive (Fig. 1A). However, once the levels of the PC1/PC2 complex are reduced, Bicc1 is now present in the cytoplasm to promote expression of the PKD proteins, thereby raising their levels (Fig. 4B), which then in turn will ‘shutdown’ Bicc1 activity by again tethering it to the plasma membrane.
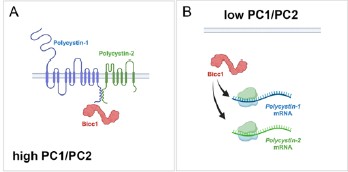
(2) Bicc1 inactivation appears to downregulate Pkd1 expression, yet it remains unclear whether Bicc1 regulates Pkd1 through direct interaction or by antagonizing miR-17, as observed in Pkd2 regulation. This should be further examined or discussed.
This is a very interesting comment. Vishal Patel published that PKD1 is regulated by a mir-17 binding site in its 3’UTR (PMID: 35965273). We, however, have not evaluated whether BICC1 participates in this regulation. A definitive answer would require utilization of the mice described in above reference, which is beyond the scope of this manuscript. We, however, have revised the discussion to elaborate on this potential mechanism.
We have updated the discussion to include a statement on the potential direct regulation of Pkd1 mRNA by Bicc1.
(3) The evidence supporting Bicc1 and ADPKD gene cooperativity, particularly with Pkd1, in mouse models is not entirely convincing, likely due to substantial variability and the aggressive nature of Bpk/Bpk mice. Increasing the number of animals or using a milder Bicc1 strain, such as jcpk heterozygotes, could help substantiate the genetic interaction.
We have initially performed the analysis using our Bicc1 complete knockout, we previously reported on (PMID 20215348) focusing on compound heterozygotes. Yet, similar to the Pkd1/Pkd2 compound heterozygotes (PMID 12140187) no cyst development was observed when we sacrificed the mice as late as P21. Our strain is similar to the above mentioned jcpk, which is characterized by a short, abnormal transcript thought to result in a null allele (PMID: 12682776). We thank the reviewer for pointing us to the reference showing the heterozygous mice exhibit glomerular cysts in the adults (PMID: 7723240). This suggestion is an interesting idea we will investigate. In general, we agree with the reviewer that a better understanding of the contribution of Bicc1 to the adult PKD phenotype will be critical. To this end, we are currently generating a floxed allele of Bicc1 that will allow us to address the cooperativity in the adult kidney, when e.g. crossed to the Pkd1<sup>RC/RC</sup> mice. Yet, these experiments are beyond the timeframe for this revision.
No changes were made in the revised manuscript.
Reviewer #2 (Public review):
(1) These results are potentially interesting, despite the limitation, also recognized by the authors, that BICC1 mutations seem exceedingly rare in PKD patients and may not "significantly contribute to the mutational load in ADPKD or ARPKD". The manuscript has several intrinsic limitations that must be addressed.
As mentioned above, the study was designed to explore whether there is an interaction between BICC1 and the PKD1/PKD2 and whether this interaction is functionally important. How this translates into the clinical relevance will require additional studies (and we have addressed this in the discussion of the manuscript).
(2) The manuscript contains factual errors, imprecisions, and language ambiguities. This has the effect of making this reviewer wonder how thorough the research reported and analyses have been.
We respectfully disagree with the reviewer on the latter interpretation. The study was performed with rigor. We have carefully assessed the critiques raised by the reviewer. As presented below, most of the criticisms raised by the reviewer have been easily addressed in the revised version of the manuscript. Yet, none of the critiques seems to directly impact the overall interpretation of the data.
Reviewer #1 (Recommendations for the authors):
(1) The manuscript requires further editing. For example, figure panels and legends are mismatched in Figure 1
We have corrected the labeling of Figure 1.
(2) Y-axis units and values are inconsistent in Figures 4b-4g, Supplementary Figures S2e and S2f are not referenced in the text, genotypes are missing in Supplementary Figure S3f, and numerous typographical errors are present.
In respect to the y-axis in Figure 4b-g, the scale is different for each of them, but that is intentional as one would lose the differences if they were all scaled identically. But we have now mentioned this in the figure legend to make the reader aware of it. In respect to the Supplemental Figure S2e,f, we included the panels in the description of the mutant BICC1 lines, but unfortunately forgot to reference them. This has now been done.
We have updated the labeling of the Y-axis for the cystic indices adding “[%]” as the unit and updated the figure legend of Figure 4. We have included the genotypes in Supplementary Figure S3f. The Supplementary Figure S2e,f is now mentioned in the supplemental material (page 9, 2<sup>nd</sup> paragraph).
Reviewer #2 (Recommendations for the authors):
(1) Previous data from mouse, Xenopus, and zebrafish suggest a crucial role for the RNAbinding protein Bicc1 in the pathogenesis of PKD, although BICC1 mutations in human PKD have not been previously reported." The cited sources (and others that were not cited) link Bicc1 mutations to renal cysts, similar to a report by Kraus (PMID: 21922595) that the authors cite later. However, a more direct link to PKD was reported by Lian and colleagues using whole Pkd1 mice (PMID: 20219263) and by Gamberi and colleagues using Pkd1 kidneys and human microarrays (PMID: 28406902). Although relevant, neither is cited here, and only the former is cited later in the manuscript.
Thanks for pointing this out. We have added these three citations.
We have added these three citations (PMID: 21922595, PMID: 20219263 and PMID: 28406902) in the indicated sentence.
(2) In Figure 1B, the lanes do not seem to correspond among panels, particularly evident in the panel with myc-mBicc1. Hence, it is difficult to agree with the presented conclusions.
We have corrected the labeling of the lanes in Figure 1b.
(3) In the Figure 1 legend: "(g) Western blot analysis following co-IP experiments, using an anti-mouse Bicc1 or anti-goat PC2 antibody as bait, identified protein interactions between endogenous PC2 and BICC1 in UCL93 cells. Non-immune goat and mouse IgG were included as a negative control." There is no mention of panel H, although this reviewer can imagine what the authors meant. The capitalization differs in the figure and legend. More troublingly, in panel G, a non-defined star indicates a strong band present in both immune and non-immune control.
We have corrected the figure legend of Figure 1 and clarified the non-specific band in the figure legend.
(4) In Figure 4, the authors do not show the matched control for the Bicc1 Pkd1 interaction in panel d, nor do they show a scale bar in either a) or d). Thus, the phenotypic severity cannot be properly assessed.
Thanks for pointing out the missing scale bars, which have now been added. In respect to the two kidneys shown in Figure 4d, the two kidneys shown are from littermates to illustrate the kidney size in agreement with the cumulative data shown in Figure 4e. Unfortunately, this litter did not have a wildtype control. As the data analysis in Figure 4e is based on littermates, mixing and matching kidneys of different litters does not seem appropriate. Thus, we have omitted showing a wildtype control in this panel. However, the size of the wildtype kidney can be seen in Figure 4a.
We have added the scale bar to both panels and have updated the figure legend to emphasize that the kidneys shown are from littermates and that no wildtype littermate was present in this litter.
(5) "Surprisingly, an 8-fold stronger interaction was observed between full-length PC1 and myc-mBicc1-ΔKH compared to mycmBicc1 or myc-mBicc1-ΔSAM." Assuming all the controls for protein folding and expression levels have been carried out and not shown/mentioned, this sentence seems to contradict the previous statement that Bicc1deltaSAM reduced the interaction with PC1 by 55%. Because the full length and SAM deletion have different interaction strengths, the latter sentence makes no sense.
The reduction in the levels of myc-mBicc1-ΔSAM compared to wildtype mycmBicc1 in respect to PC1 binding was not significant. We have clarified this in the text.
We have corrected the sentence and modified the Figure accordingly.
(6) Imprecise statements make a reader wonder how to interpret the data: "More than three independent experiments were analyzed." Stating the sample size or including it in the figure would save space and improve confidence in the data presented.
We have stated the exact number of animals per conditions above each of the bars.
(7) "Next, we performed a similar mouse study for Pkd1 by reducing the gene dose of Pkd1 postnatally in the collecting ducts using a Pkhd1-Cre as previously described40" What did the authors mean?
The reference was included to cite the mouse strain, but realized that it can be mis-interpreted that the exact experiments has been performed previously. We have clarified this in the text.
We have reworded the sentence to avoid misinterpretation.
(8) The authors examined the additive effects of knocking down Bicc1, Pkd1, and Pkd2 with morpholinos in Xenopus and, genetically, in mice. While the Bicc1[+/-] Pkd1 or 2[+/-] double heterozygote mice did not show phenotypes, the authors report that the Bicc1[-/-] Pkd1 or 2 [+/-] did instead show enlarged kidneys. What is the phenotype of a Bicc1[+/-] Pkd1 or 2 [-/-]? What we learn from the author's findings among the PKD population suggests that the latter situation would be potentially translationally relevant.
The mouse experiments were designed to address a cooperativity between Bicc1 and either Pkd1 or Pkd2 and whether removal of one copy of Pkd1 or Pkd2 would further worsen the Bicc1 cystic kidney phenotype. Thus, the parental crosses were chosen to maximize the number of animals obtained for these genotypes. Unfortunately, these crosses did not yield the genotypes requested by the reviewer. To address the contribution of Bicc1 towards the PKD population, we will need to perform a different cross, where we eliminate Pkd1 or Pkd2 in a floxed background of Bicc1 postnatally in adult mice. While we are gearing up to perform such an experiment, this is timewise beyond the scope of the manuscript. In addition, please note that we have addressed the question about the translation towards the PKD population already in the discussion of the original submission (page 13/14, last/first paragraph).
No changes have been made to the revised version of the manuscript.
(9) How do the authors interpret the milder effects of the Bicc1[-/-] Pkd1[+/-] compared to Bicc1[-/-] Pkd2[+/-] relative to the respective protein-protein interactions?
The milder effects are due to the nature of the crosses. While the Pkd2 mutant is a germline mutation, the Pkd1 mutant is a conditional allele eliminating Pkd1 only in the collecting ducts of the kidney. As such, we spare other nephron segments such as the proximal tubules, which also significantly contribute to the cyst load. As such these mouse data support the interaction between Pkd1 and Pkd2 with Bicc1, but do not allow us to directly compare the outcomes. While this was mentioned in the previous version of the manuscript, we have expanded on this in the revised version of the manuscript.
We have expanded the results section in the revised version of the manuscript highlighting that the two different approaches cannot be directly compared.
(10) How do the authors interpret that the strong Bicc1[Bpk] Pkd1 or Pkd2 double heterozygote mice did not have defects and "kidneys from Bicc1+/-:Pkd2+/- did not exhibit cysts (data not shown)", when the VEO PKD patients and - although not a genetic reduction - also the morpholino-treated Xenopus did?
VEO PKD patients are characterized by a loss of function of PKD1 or PKD2 and – as we propose in this manuscript - that BICC1 further aggravates the phenotype. Yet, we do not address either in the mouse or Xenopus experiments whether BICC1 is a genetic modifier. We are simply addressing whether the two genes show a genetic interaction. In the mouse studies, we eliminate one copy of Pkd1 or Pkd2 in the background of a hypomorphic allele of Bicc1. Similarly, in the Xenopus experiments, we employ suboptimal doses of the morpholino oligomers, i.e., concentrations that did not yield a phenotypic change and then asked whether removing both together show cooperativity. It is important to state that this is based on a biological readout and not defined based on the amount of protein. While we have described this already in the original manuscript (page 7, first paragraph), we have amended our description of the Xenopus experiment to make this even clearer.
Finally, we agree with the reviewer that if we were to address whether Bicc1 is a modifier of the PKD phenotype in mouse, we would need to reduce Bicc1 function in a Pkd1 or Pkd2 mutants. Yet, we have recognized this already in the initial version of the manuscript in the discussion (page 14, first paragraph).
We have expanded the results section when discussing the suboptimal amounts of the morpholino oligos (Page 6, 1<sup>st</sup> paragraph).
(11) Unclear: "While variants in BICC1 are very rare, we could identify two patients with BICC1 variants harboring an additional PKD2 or PKD1 variant in trans, respectively." Shortly after, the authors state in apparent contradiction that "the patients had no other variants in any of other PKD genes or genes which phenocopy PKD including PKD1, PKD2, PKHD1, HNF1s, GANAB, IFT140, DZIP1L, CYS1, DNAJB11, ALG5, ALG8, ALG9, LRP5, NEK8, OFD1, or PMM2."
The reviewer is correct. This should have been phrased differently. We have now added “Besides the variants reported below” to clarify this more adequately.
The sentence was changed to start with “Besides the variants reported below, […].”
(12) "The demonstrated interaction of BICC1, PC1, and PC2 now provides a molecular mechanism that can explain some of the phenotypic variability in these families." How do the authors reconcile this statement with their reported ultra-rare occurrence of the BICC1 mutations?
As mentioned in the manuscript and also in response to the other two reviewers, Bicc1 has been shown to regulate Pkd2 gene expression in mice and frogs via an interaction with the miR-17 family of microRNAs. Moreover, the miR-17 family has been demonstrated to be critical in PKD (PMID: 30760828, PMID: 35965273, PMID: 31515477, PMID: 30760828). In fact, both other reviewers have pointed out that we should stress this more since Bicc1 is part of this regulatory pathway. Future experiments are needed to address whether Bicc1 contributes to the variability in ADPKD onset/severity. Yet, this is beyond the scope of this study.
Based on the comments of the two other reviewers we have further addressed the Bicc1/miR-17 interaction.
(13) The manuscript should use correct genetic conventions of italicization and capitalization. This is an issue affecting the entire manuscript. Some exemplary instances are listed below.
(a) "We also demonstrate that Pkd1 and Pkd2 modifies the cystic phenotype in Bicc1 mice in a dose-dependent manner and that Bicc1 functionally interacts with Pkd1, Pkd2 and Pkhd1 in the pronephros of Xenopus embryos." Genes? Proteins?
The data presented in this section show that a hypomorphic allele of Bicc1 in mouse and a knockdown in Xenopus yields this. As both affect the proteins, the spelling should reflect the proteins.
No changes have been made in the revised manuscript.
(b) The sentence seems to use both the human and mouse genetic capitalization, although it refers to experiments in the mouse system “to define the Bicc1 interacting domains for PC2 (Fig. 2d,e). Full-length PC2 (PC2-HA) interacted with full-length myc-mBICC1.”
We agree with the review that stating the species of the molecules used is critical, we have adapted a spelling of Bicc1, where BICC1 is the human homologue, mBicc1 is the mouse homologue and xBicc1 the Xenopus one.
We have highlighted the species spelling in the methods section and labeled the species accordingly throughout the manuscript and figures.
(14) “Together these data supported our biochemical interaction data and demonstrated that BICC1 cooperated with PKD1 and PKD2.” Are the authors implying that these results in mice will translate to the human protein?
We agree that we have not formally shown that the same applies to the human proteins. Thus, we have changed the spelling accordingly.
We have revised the capitalization of the proteins.
(15) The text is often unclear, terse, or inconsistent.
(a) “These results suggested that the interaction between PC1 and Bicc1 involves the SAM but not the KH/KHL domains (or the first 132 amino acids of Bicc1). It also suggests that the N-terminus could have an inhibitory effect on PC1-BICC1 association.” How do the authors define the N-terminus? The first 132 aa? KH/KHL domains?
This was illustrated in the original Figure 2A. The DKH constructs lack the first 351 amino acids.
To make this more evident, we have specified this in the text as well.
(b) Similarly, the authors state below, "Unlike PC1, PC2 interacted with mycmBICC1ΔSAM, but not myc-mBICC1-ΔKH suggesting that PC2 binding is dependent on the N-terminal domains but not the SAM domain." It is unclear if the authors refer to the KH/KHL domains or others. Whatever the reference to the N-terminal region, it should also be consistent with the section above.
This is now specified in the text.
(c) Unclear: "We have previously demonstrated that Pkd2 levels are reduced in a complete Bicc1 null mice,22 performing qRT-PCR of P4 kidneys (i.e. before the onset of a strong cystic phenotype), revealed that Bicc1, Pkd1 and Pkd2 were statistically significantly down9 regulated (Fig. 4h-j)".
We have changed the text to clarify this.
(d) “Utilizing recombinant GST domains of PC1 and PC2, we demonstrated that BICC1 binds to both proteins in GST-pulldown assays (Fig. 1a, b)." GST-tagged domains? Fusions?
We have changed the text to clarify this.
(e) "To study the interaction between BICC1, PKD1 and PKD2 we combined biochemical approaches, knockout studies in mice and Xenopus, genetic engineered human kidney cells" > genetically engineered.
We have changed the text to clarify this.
(f) Capitalization (e.g., see Figure S3, ref. the Bpk allele) and annotation (e.g., Gly821Glu and G821E) are inconsistent.
We have homogenized the labeling of the capitalization and annotations throughout the manuscript.
(g) What do the authors mean by "homozygous evolutionarily well-conserved missense variant"?
We have changed this is the revised version of the manuscript.
Reviewer #3 (Public review/Recommendations to the authors):
(1) A further study in HUREC cells investigating the critical regulatory role of BICC1 and potential interaction with mir-17 may yet lead to a modifiable therapeutic target.
(2) This study should ideally include experiments in HUREC material obtained from patients/families with BICC1 mutations and studying its effects on the PKD1/2 complex in primary cell lines.
This is an excellent suggestion. We agree with the reviewer that it would have been interesting to analyze HUREC material from the affected patients. Unfortunately, besides DNA and the phenotypic analysis described in the manuscript neither human tissue nor primary patient-derived cells collected once the two patients with the BICC1 p.Ser240Pro variant passed away.
No changes to the revised manuscript have been made to address this point.
(3) Please remove repeated words in the following sentence in paragraph 2 of the introduction: "BICC1 encodes an evolutionarily conserved protein that is characterized by 3 K-homology (KH) and 2 KH-like (KHL) RNA-binding domains at the N-terminus and a SAM domain at the C-terminus, which are separated by a by a disordered intervening sequence (IVS).23-28".
This has been changed.
-
-
www.americanyawp.com www.americanyawp.com
-
All men are created free and equal.” Taught now to live and work for the common weal, and not for the maintenance of a royal family or an overbearing aristocracy, not for the continuance of a social system which ranks them beneath an arrogant class of drones, children of Russian and German serfs, of Irish evicted tenants, Scotch crofters, and other victims of feudal tyranny, are translated into republican Americans, and are made in one love for a country which provides equal rights and privileges for all her children
in that time of the 1800's man write that all men are created free and equal but don't do what they write when they practice seprated but not equal to the black of america
-
-
Local file Local file
-
But their economic preparations were also a function ofattitudes toward war in the interwar period itself. For this reason preparationswere extensive in Germany, much less so in the United Kingdom, and almostnonexistent in the United States.
However, such differences in initial female employment numbers and the voluntary beginings of womens work may be due to, Purseigle notes, Germany's position as aggressor, German women working in higher numbers due to mobilising earlier
-
-
social-media-ethics-automation.github.io social-media-ethics-automation.github.io
-
Separately, in 2018 during the MeToo movement, one of @Sciencing_Bi’s friends, Dr. BethAnn McLaughlin (a white woman), co-founded the MeTooSTEM non-profit organization, to gather stories of sexual harassment in STEM (Science, Technology, Engineering, Math). Kyle also followed her on Twitter until word later spread of Dr. McLaughlin’s toxic leadership and bullying in the MeTooSTEM organization (Kyle may have unfollowed @Sciencing_Bi at the same time for defending Dr. McLaughlin, but doesn’t remember clearly).
Authenticity is vital for maintaining public trust and the lack of it cause audiences to lose faith in the credibility of the information they receive from an individual or organization if they deem it inauthentic. However the norm is inauthenticity, where personas and identity are carefully crafted in order to boost engagement, this however is a very risky endeavor as it cause the groups that already agree with the perspective of the poster to stick around while those who value integrity and truthfulness to leave once they see the fakeness.
-
Sockpuppet (or a “burner” account): Creating a fake profile in order to argue a position (sometimes intentionally argued poorly to make the position look bad)
I have never heard this term before, but I knew of this before reading this chapter. It is nice to put a term with the process of creating a burner account.
-
6.3.1. Inauthentic Behaviors# Inauthentic behavior is when the reality doesn’t match what is being presented. Inauthenticity has, of course, existed throughout human history, from Ea-nasir complaining in 1750 BCE that the copper he ordered was not the high quality he had been promised, to 1917 CE in England when Arthur Conan Doyle (the author of the Sherlock Holmes stories) was fooled by photographs that appeared to be of a child next to fairies.
These historical examples show that inauthentic behavior is not new, but has long been part of human communication. What changes in digital contexts is the scale and speed at which inauthenticity can spread, especially when platforms and automated systems amplify misleading representations.
-
6.3. Inauthenticity# In 2016, the Twitter account @Sciencing_Bi was created by an anonymous bisexual Native American Anthropology professor at Arizona State University (ASU). She talked about her experiences of discrimination and about being one of the women who was sexually harassed by a particular Harvard professor. She gained a large Twitter following among academics, including one of the authors of this book, Kyle. Separately, in 2018 during the MeToo movement, one of @Sciencing_Bi’s friends, Dr. BethAnn McLaughlin (a white woman), co-founded the MeTooSTEM non-profit organization, to gather stories of sexual harassment in STEM (Science, Technology, Engineering, Math). Kyle also followed her on Twitter until word later spread of Dr. McLaughlin’s toxic leadership and bullying in the MeTooSTEM organization (Kyle may have unfollowed @Sciencing_Bi at the same time for defending Dr. McLaughlin, but doesn’t remember clearly). Then, in April 2020, in the early days of the COVID pandemic, @Sciencing_Bi complained of being forced to teach in person at ASU when it wasn’t safe, and then began writing about their COVID symptoms.
This case illustrates how perceptions of authenticity on social media are often built through personal narratives and shared experiences. When accounts gain trust through identity claims and moral authority, questions of inauthenticity can have especially high stakes, affecting not only credibility but also the communities and causes connected to that account.
-
-
human.libretexts.org human.libretexts.org
-
But undocumented immigrants, and immigrants more generally, are often baselessly blamed for increasing crime rates – including, repeatedly, by President Donald Trump. In the second and final presidential debate, Trump again claimed undocumented immigrants are rapists and murderers.
Why is this such a widespread belief? Is this another example of human's fear of the unknown being exploited for another human's personal gain?
-
Using a statistical method called regression analysis to examine the data, we found that as the size of the undocumented population increases, the property crime rate decreases, on average.
I find it surprising that undocumented immigrants make a community safer and bring down the crime rate, but it sounds probable.
-
People from all social groups and backgrounds commit crimes.
In 2010, 35% of all African American men in the U.S. had a felony conviction. All races commit crimes in similar proportions but minorities are more likely to receive harsher treatment from police and the U.S. justice system.
-
-
www.biorxiv.org www.biorxiv.org
-
Author response:
Reviewer #1 (Public review):
The authors analysed large-scale brain-state dynamics while humans watched a short video. They sought to identify the role of thalamocortical interactions.
Major concerns
(1) Rationale for using the naturalistic stimulus
In terms of brain state dynamics, previous studies have already reported large-scale neural dynamics by applying some data-driven analyses, like energy landscape analysis and Hidden Markov Model, to human fMRI/EEG data recorded during resting/task states. Considering such prior work, it'd be critical to provide sufficient biological rationales to perform a conceptually similar study in a naturalistic condition, i.e., not just "because no previous work has been done". The authors would have to clarify what type of neural mechanisms could be missed in conventional resting-state studies using, say, energy landscape analysis, but could be revealed in the naturalistic condition.
We appreciate your insightful comments regarding the need for a biological rationale in our study. As you mentioned, there are similar studies, just like Meer et al. utilized Hidden Markov Models to identify various activation modes of brain networks that included subcortical regions[1], Song et al. linked brain states to narrative understandings and attentional dynamics[2, 3]. These studies could answer why we use naturalistic stimuli datasets. Moreover, there is evidence suggesting that the thalamus plays a crucial role in processing information in a more naturalistic context while pointing out the vital role in thalamocortical communications[4, 5]. So, we tended to bridge thalamic activity and cortical state transition using the energy landscape description.
To address these gaps in conventional resting-state studies, we explored an alternative method—maximum entropy modeling based on the energy landscape. This allowed us to validate how the thalamus responds to cortical state transitions. To enhance clarity, we will update our introduction to emphasize the motivations behind our research and the significance of examining these neural mechanisms in a naturalistic setting.
(2) Effects of the uniqueness of the visual stimulus and reproducibility
One of the main drawbacks of the naturalistic condition is the unexpected effects of the stimuli. That is, this study looked into the data recorded from participants who were watching Sherlock, but what would happen to the results if we analyzed the brain activity data obtained from individuals who were watching different movies? To ensure the generalizability of the current findings, it would be necessary to demonstrate qualitative reproducibility of the current observations by analysing different datasets that employed different movie stimuli. In fact, it'd be possible to find such open datasets, like www.nature.com/articles/s41597-023-02458-8.
We appreciate your concern regarding the reproducibility of our findings. The dataset from the "Sherlock" study is of high quality and has shown good generalizability in various research contexts. We acknowledge the importance of validating our results with different datasets to enhance the robustness of our conclusions. While we are open to exploring additional datasets, we intend to pursue this validation once we identify a suitable alternative. Currently, we are considering a comparison with the dataset from "Forrest Gump" as part of our initial plan.
(3) Spatial accuracy of the "Thalamic circuit" definition
One of the main claims of this study heavily relies on the accuracy of the localization of two different thalamic architectures: matrix and core. Given the conventional or relatively low spatial resolution of the fMRI data acquisition (3x3x3 mm^3), it appears to be critically essential to demonstrate that the current analysis accurately distinguished fMRI signals between the matrix and core parts of the thalamus for each individual.
We acknowledge the importance of accurately localizing the different thalamic architectures, specifically the matrix and core regions. To address this, we downsampled the atlas of matrix and core cell populations from the previous study from a resolution of 2x2x2 mm<sup>3</sup> to 3x3x3 mm<sup>3</sup>, which aligns with our fMRI data acquisition. We would report the atlas as Supplementary Figures in our revision.
(4) More detailed analysis of the thalamic circuits
In addition, if such thalamic localisation is accurate enough, it would be greatly appreciated if the authors perform similar comparisons not only between the matrix and core architectures but also between different nuclei. For example, anterior, medial, and lateral groups (e.g., pulvinar group). Such an investigation would meet the expectations of readers who presume some microscopic circuit-level findings.
We appreciate your suggestion regarding a more detailed analysis of thalamic circuits. We have touched upon this in the discussion section as a forward-looking consideration. However, we believe that performing nuclei segmentation with 3T fMRI may not be ideal due to well-documented concerns regarding signal-to-noise ratio and spatial resolution. That said, we are interested in exploring these nuclei-pathway connections to cortical areas in future studies with a proper 7T fMRI naturalistic dataset.
(5) Rationale for different time window lengths
The authors adopted two different time window lengths to examine the neural dynamics. First, they used a 21-TR window for signal normalisation. Then, they narrowed down the window length to 13-TR periods for the following statistical evaluation. Such a seemingly arbitrary choice of the shorter time window might be misunderstood as a measure to relax the threshold for the correction of multiple comparisons. Therefore, it'd be appreciated if the authors stuck to the original 21-TR time window and performed statistical evaluations based on the setting.
Thank you for your valuable feedback regarding the choice of time window lengths. We aimed to maintain consistency in window lengths across our analyses. In light of your comments and suggestions from other reviewers, we plan to test our results using different time window lengths and report findings that generalize across these variations. Should the results differ significantly, we will discuss the implications of this variability in our revised manuscript.
(6) Temporal resolution
After identifying brain states with energy landscape analysis, this study investigated the brain state transitions by directly looking into the fMRI signal changes. This manner seems to implicitly assume that no significant state changes happen in one TR (=1.5sec), which needs sufficient validation. Otherwise, like previous studies, it'd be highly recommended to conduct different analyses (e.g., random-walk simulation) to address and circumvent this problem.
Thank you for raising this important point regarding temporal resolution. Many fMRI studies, such as those examining event boundaries during movie watching, operate under similar assumptions concerning state changes within one TR. For example, Barnett et al. processed the dynamic functional connectivity (dFC) with a window of 20 TRs (24.4s). So, we do not think it is a limitation but is a common question related to fMRI scanning parameters. To strengthen our analysis of state transitions and ensure they are not merely coincidental, we plan to conduct random-walk simulations, as suggested, to validate our findings in accordance with methodologies used in previous research.
Reviewer #2 (Public review):
Summary:
In this study, Liu et al. investigated cortical network dynamics during movie watching using an energy landscape analysis based on a maximum entropy model. They identified perception- and attention-oriented states as the dominant cortical states during movie watching and found that transitions between these states were associated with inter-subject synchronization of regional brain activity. They also showed that distinct thalamic compartments modulated distinct state transitions. They concluded that cortico-thalamo-cortical circuits are key regulators of cortical network dynamics.
Strengths:
A mechanistic understanding of cortical network dynamics is an important topic in both experimental and computational neuroscience, and this study represents a step forward in this direction by identifying key cortico-thalamo-cortical circuits. The analytical strategy employed in this study, particularly the LASSO-based analysis, is interesting and would be applicable to other data types, such as task- and resting-state fMRI.
We thanks for this comment and encouragement.
Weaknesses:
Due to issues related to data preprocessing, support for the conclusions remains incomplete. I also believe that a more careful interpretation of the "energy" derived from the maximum entropy model would greatly clarify what the analysis actually revealed.
Thank you for your valuable suggestions, and we apologize for any misunderstandings regarding the interpretation of the energy landscape in our study. To address this issue, we will include a dedicated paragraph in both the methods and results sections to clarify our use of the term "energy" derived from the maximum entropy model. This addition aims to eliminate any ambiguity and provide a clearer understanding of what our analysis reveals.
(1) I think the method used for binarization of BOLD activity is problematic in multiple ways.
a) Although the authors appear to avoid using global signal regression (page 4, lines 114-118), the proposed method effectively removes the global signal. According to the description on page 4, lines 117-122, the authors binarized network-wise ROI signals by comparing them with the cross-network BOLD signal (i.e., the global signal): at each time point, network-wise ROI signals above the cross-network signal were set to 1, and the rest were set to −1. If I understand the binarization procedure correctly, this approach forces the cross-network signal to be zero (up to some noise introduced by the binarization of network-wise signals), which is essentially equivalent to removing the global signal. Please clarify what the authors meant by stating that "this approach maintained a diverse range of binarized cortical states in data where the global signal was preserved" (page 4, lines 121-122).
Thank you for highlighting the potential issue with our binarization method. We appreciate your insights regarding the comparison of network-wise ROI signals with the cross-network BOLD signal, as this may inadvertently remove the global signal. To address this, we will conduct a comparative analysis of results obtained from both our current approach and the original pipeline. If we decide to retain our current method, we will carefully reconsider the rationale and rephrase our descriptions to ensure clarity regarding the preservation of the global signal and the diversity of binarized cortical states.
b) The authors might argue that they maintained a diverse range of cortical states by performing the binarization at each time point (rather than within each network). However, I believe this introduces another problem, because binarizing network-wise signals at each time point distorts the distribution of cortical states. For example, because the cross-network signal is effectively set to zero, the network cannot take certain states, such as all +1 or all −1. Similarly, this binarization biases the system toward states with similar numbers of +1s and −1s, rather than toward unbalanced states such as (+1, −1, −1, −1, −1, −1). These constraints and biases are not biological in origin but are simply artifacts of the binarization procedure. Importantly, the energy landscape and its derivatives (e.g., hard/easy transitions) are likely to be affected by these artifacts. I suggest that the authors try a more conventional binarization procedure (i.e., binarization within each network), which is more robust to such artifacts.
Related to this point, I have a question regarding Figure S1, in which the authors plotted predicted versus empirical state probabilities. As argued above, some empirical state probabilities should be zero because of the binarization procedure. However, in Figure S1, I do not see data points corresponding to these states (i.e., there should be points on the y-axis). Did the authors plot only a subset of states in Figure S1? I believe that all states should be included. The correlation coefficient between empirical and predicted probabilities (and the accuracy) should also be calculated using all states.
Thank you for your thoughtful examination of our data processing pipeline. We agree that a comparison between the conventional binarization method and our current approach is warranted, and we appreciate your suggestion. Upon reviewing Figure S1, we discovered that there was indeed an error related to the plotting style set to "log10." As you correctly pointed out, the data should reflect that the probabilities for states where all networks are either activated or deactivated are zero. We are very interested in exploring the state distributions obtained from both the original and current approaches, as your comments highlight important considerations. We sincerely appreciate your insightful feedback and will make sure to address these points thoroughly in our first revision.
c) The current binarization procedure likely inflates non-neuronal noise and obscures the relationship between the true BOLD signal and its binarized representation. For example, consider two ROIs (A and B): both (+2%, +1%) and (+0.01%, −0.01%) in BOLD signal changes would be mapped to (+1, −1) after binarization. This suggests that qualitatively different signal magnitudes are treated identically. I believe that this issue could be alleviated if the authors were to binarize the signal within each network, rather than at each time point.
Thank you for your important observation regarding the potential inflation of non-neuronal noise in our current binarization procedure. We recognize that this process could lead to qualitatively different signal magnitudes being treated similarly after binarization, as you illustrated with your example. While we acknowledge your point, we believe that conventional binarization pipelines may also encounter this issue, albeit by comparing signals to a network's temporal mean activity. To address this concern and maintain consistency with previous studies, we will discuss this limitation in our revised manuscript. Additionally, if deemed necessary, we will explore implementing a percentile-based threshold above the baseline to further refine our binarization approach. Your suggestion provides a valuable perspective, and we appreciate your insights.
(2) As the authors state (page 5, lines 145-148), the "energy" described in the energy landscape is not biological energy but rather a statistical transformation of probability distributions derived from the Boltzmann distribution. If this is the case, I believe that Figure 2A is potentially misleading and should be removed. This type of schematic may give the false impression that cortical state dynamics are governed by the energy landscape derived from the maximum entropy model (which is not validated).
Thank you for your valuable feedback regarding Figure 2A. We apologize for any confusion it may have created. While we recognize that similar figures are commonly used in literature involving energy landscapes (maximum entropy model), we agree that Figure 2A may mislead readers into thinking that cortical state dynamics are directly governed by the energy landscape derived from the maximum entropy model, which has not been validated. In light of your comments, we will remove Figure 2A and instead emphasize the analytical strategy presented in Figure 2B. Additionally, we will provide a simplified line graph as an illustrative example to clarify the concepts without the potential for misinterpretation.
Reviewer #3 (Public review):
Summary:
In this study, Liu et al. analyze fMRI data collected during movie watching, applied an energy landscape method with pairwise maximum entropy models. They identify a set of brain states defined at the level of canonical functional networks and quantify how the brain transitions between these states. Transitions are classified as "easy" or "hard" based on changes in the inferred energy landscape, and the authors relate transition probabilities to inter-subject correlation. A major emphasis of the work is the role of the thalamus, which shows transition-linked activity changes and dynamic connectivity patterns, including differential involvement of parvalbumin- and calbindin-associated thalamic subdivisions.
Strengths:
The study is methodologically complex and technically sophisticated. It integrates advanced analytical methods into high-dimensional fMRI data. The application of energy landscape analysis to movie-watching data appears to be novel as well. The finding on the thalamus involved energy state transition and provides a strong linkage to several theories on thalamic control functions, which is a notable strength.
Thanks for your comments on the novelty of our study.
Weaknesses:
The main weakness is the conceptual clarity and advances that this otherwise sophisticated set of analyses affords. A central conceptual ambiguity concerns the energy landscape framework itself. The authors note that the "energy" in this model is not biological energy but a statistical quantity derived from the Boltzmann distribution. After multiple reads, I still have major trouble mapping this measure onto any biological and cognitive operations. BOLD signal is a measure of oxygenation as a proxy of neural activity, and correlated BOLD (functional connectivity) is thought to measure the architecture of information communication of brain systems. The energy framework described in the current format is very difficult for most readers to map onto any neural or cognitive knowledge base on the structure and function of brain systems. Readers unfamiliar with maximum entropy models may easily misinterpret energy changes as reflecting metabolic cost, neural effort, or physiological variables, and it is just very unclear what that measure is supposed to reflect. The manuscript does not clearly articulate what conceptual and mechanistic advances the energy formalism provides beyond a mathematical and statistical report. In other words, beyond mathematical description, it is very hard for most readers to understand the process and function of what this framework is supposed to tell us in regards to functional connectivity, brain systems, and cognition. The brain is not a mathematical object; it is a biological organ with cognitive functions. The impact of this paper is severely limited until connections can be made.
Thank you for your insightful and constructive comments regarding the conceptual clarity of our energy landscape framework. We appreciate your perspective on the challenges of mapping the statistical measure of "energy" derived from the Boltzmann distribution onto biological and cognitive operations. To address these concerns, we will revise our manuscript to clarify our expressions surrounding "energy" and emphasize its probabilistic nature. Additionally, we will incorporate a series of analyses that explicitly relate the features of the energy landscape to cognitive processes and key parameters, such as brain integration and functional connectivity. We believe these changes will help bridge the gap between our mathematical framework and its relevance to understanding brain systems and cognitive functions.
Relatedly, the use of metaphors such as "valleys," "hills," and "routes" in multidimensional measures lacks grounding. Valleys and hills of what is not intuitive to understand. Based on my reading, these features correspond to local minima and barriers in a probability distribution over binarized network activation patterns, but similar to the first point, the manuscript does not clearly explain what it means conceptually, neurobiologically, or computationally for the brain to "move" through such a landscape. The brain is not computing these probabilities; they are measurement tools of "something". What is it? To advance beyond mathematical description, these measurements must be mapped onto neurobiological and cognitive information.
Thank you for your valuable feedback. In our revisions, we would aim to link the concept of rapid transition routes in the energy landscape to cognitive processes, such as narrative understanding and related features. By exploring these connections, we hope to provide a clearer context for how our framework can enhance understanding of cognitive functions and their neural correlates.
This conceptual ambiguity goes back to the Introduction. At the level of motivation, the purpose and deliverables of the study are not defined in the Introduction. The stated goal is "Transitions between distinct cortical brain states modulate the degree of shared neural processing under naturalistic conditions". I do not know if readers will have a clear answer to this question at the end. Is the claim that state transitions cause changes in inter-subject correlation, that they index moments of narrative alignment, or that they reflect changes in attentional or cognitive mode? This level of explanation is largely dissociated from the methods in their current form.
Thank you for highlighting this important point regarding the conceptual clarity in our Introduction. We appreciate your feedback about the motivation and objectives of the study. To clarify the stated goal of investigating how transitions between distinct cortical brain states modulate shared neural processing under naturalistic conditions, we will revise the manuscript to explicitly define the specific claims we aim to address. We will ensure that these explanations are closely tied to the methods employed in our study, providing a clearer framework for our readers.
Several methodological choices can use clarification. The use of a 21-TR window centered on transition offsets is unusually long relative to the temporal scale of fMRI dynamics and to the hypothesized rapidity of state transitions. On a related note, what is the temporal scale of state transition? Is it faster than 21 TRs?
Thank you for your insightful questions regarding our methodological choices. Our focus on specific state transitions necessitated the use of a 21-TR window. While it’s true that other transitions may occur within this window, averaging across the same transitions at different times allows us to identify distinctive thalamic BOLD patterns that precede cortical state transitions. This methodology enables us to capture relevant dynamics while ensuring that we focus on the transitions of interest. We appreciate your feedback, and this clarification will be included in our revised manuscript. We would also add a figure that describe the dwell time of cortical states.
The choice of movie-watching data is a strength. But, many of the analyses performed here, energy landscape estimation, clustering of states, could in principle be applied to resting-state data. The manuscript does not clearly articulate what is gained, mechanistically or cognitively, by using movie stimuli beyond the availability of inter-subject correlation.
Thank you for your question, which closely aligns with a concern raised by Reviewer #1. Our core hypothesis posits that naturalistic stimuli yield a broader set of brain states compared to those observed during resting-state conditions. To support this assertion, we will clearly articulate the findings from previous studies that relate to this hypothesis. Additionally, if appropriate, we will provide a comparative analysis between our data and resting-state data to highlight the differences and emphasize the uniqueness of the brain states elicited by naturalistic stimuli.
Because of the above issues, a broader concern throughout the results is the largely descriptive nature of the findings. For example, the LASSO analysis shows that certain state transitions predict ISC in a subset of regions, with respectable R² values. While statistically robust, the manuscript provides little beyond why these particular transitions should matter, what computations they might reflect, or how they relate to known cognitive operations during movie watching. Similar issues arise in the clustering analyses. Clustering high-dimensional fMRI-derived features will almost inevitably produce structure, whether during rest, task, or naturalistic viewing. What is missing is an explanation of why these specific clusters are meaningful in functional or mechanistic terms.
Thank you for your questions. In our revisions, we will perform additional analyses aimed at linking state transitions to cognitive processes more explicitly. Regarding clustering, we will provide a thorough discussion in the revised manuscript.
Finally, the treatment of the thalamus, while very exciting, could use a bit more anatomical and circuit-level specificity. The manuscript largely treats the thalamus as a unitary structure, despite decades of work demonstrating big functional and connectivity differences across thalamic nuclei. A whole-thalamus analysis without more detailed resolution is increasingly difficult to justify. The subsequent subdivision into PVALB- and CALB-associated regions partially addresses this, but these markers span multiple nuclei with overlapping projection patterns.
This suggestion aligns with the feedback from Reviewer #1. We believe that performing nuclei segmentation with 3T fMRI may not be ideal due to well-documented concerns regarding signal-to-noise ratio and spatial resolution. Therefore, investigating core and matrix cell projections across different thalamic nuclei using 7T fMRI presents a promising avenue for further study.
(1) Van Der Meer J N, Breakspear M, Chang L J, et al. Movie viewing elicits rich and reliable brain state dynamics [J]. Nature Communications, 2020, 11(1): 5004.
(2) Song H, Park B Y, Park H, et al. Cognitive and Neural State Dynamics of Narrative Comprehension [J]. Journal of Neuroscience, 2021, 41(43): 8972-8990.
(3) Song H, Shim W M, Rosenberg M D. Large-scale neural dynamics in a shared low-dimensional state space reflect cognitive and attentional dynamics [J]. Elife, 2023, 12.
(4) Shine J M, Lewis L D, Garrett D D, et al. The impact of the human thalamus on brain-wide information processing [J]. Nature Reviews Neuroscience, 2023, 24(7): 416-430.
(5) Yang M Y, Keller D, Dobolyi A, et al. The lateral thalamus: a bridge between multisensory processing and naturalistic behaviors [J]. Trends in Neurosciences, 2025, 48(1): 33-46.
-
Reviewer #3 (Public review):
Summary:
In this study, Liu et al. analyze fMRI data collected during movie watching, applied an energy landscape method with pairwise maximum entropy models. They identify a set of brain states defined at the level of canonical functional networks and quantify how the brain transitions between these states. Transitions are classified as "easy" or "hard" based on changes in the inferred energy landscape, and the authors relate transition probabilities to inter-subject correlation. A major emphasis of the work is the role of the thalamus, which shows transition-linked activity changes and dynamic connectivity patterns, including differential involvement of parvalbumin- and calbindin-associated thalamic subdivisions.
Strengths:
The study is methodologically complex and technically sophisticated. It integrates advanced analytical methods into high-dimensional fMRI data. The application of energy landscape analysis to movie-watching data appears to be novel as well. The finding on the thalamus involved energy state transition and provides a strong linkage to several theories on thalamic control functions, which is a notable strength.
Weaknesses:
The main weakness is the conceptual clarity and advances that this otherwise sophisticated set of analyses affords. A central conceptual ambiguity concerns the energy landscape framework itself. The authors note that the "energy" in this model is not biological energy but a statistical quantity derived from the Boltzmann distribution. After multiple reads, I still have major trouble mapping this measure onto any biological and cognitive operations. BOLD signal is a measure of oxygenation as a proxy of neural activity, and correlated BOLD (functional connectivity) is thought to measure the architecture of information communication of brain systems. The energy framework described in the current format is very difficult for most readers to map onto any neural or cognitive knowledge base on the structure and function of brain systems. Readers unfamiliar with maximum entropy models may easily misinterpret energy changes as reflecting metabolic cost, neural effort, or physiological variables, and it is just very unclear what that measure is supposed to reflect. The manuscript does not clearly articulate what conceptual and mechanistic advances the energy formalism provides beyond a mathematical and statistical report. In other words, beyond mathematical description, it is very hard for most readers to understand the process and function of what this framework is supposed to tell us in regards to functional connectivity, brain systems, and cognition. The brain is not a mathematical object; it is a biological organ with cognitive functions. The impact of this paper is severely limited until connections can be made.
Relatedly, the use of metaphors such as "valleys," "hills," and "routes" in multidimensional measures lacks grounding. Valleys and hills of what is not intuitive to understand. Based on my reading, these features correspond to local minima and barriers in a probability distribution over binarized network activation patterns, but similar to the first point, the manuscript does not clearly explain what it means conceptually, neurobiologically, or computationally for the brain to "move" through such a landscape. The brain is not computing these probabilities; they are measurement tools of "something". What is it? To advance beyond mathematical description, these measurements must be mapped onto neurobiological and cognitive information.
This conceptual ambiguity goes back to the Introduction. At the level of motivation, the purpose and deliverables of the study are not defined in the Introduction. The stated goal is "Transitions between distinct cortical brain states modulate the degree of shared neural processing under naturalistic conditions". I do not know if readers will have a clear answer to this question at the end. Is the claim that state transitions cause changes in inter-subject correlation, that they index moments of narrative alignment, or that they reflect changes in attentional or cognitive mode? This level of explanation is largely dissociated from the methods in their current form.
Several methodological choices can use clarification. The use of a 21-TR window centered on transition offsets is unusually long relative to the temporal scale of fMRI dynamics and to the hypothesized rapidity of state transitions. On a related note, what is the temporal scale of state transition? Is it faster than 21 TRs?
The choice of movie-watching data is a strength. But, many of the analyses performed here, energy landscape estimation, clustering of states, could in principle be applied to resting-state data. The manuscript does not clearly articulate what is gained, mechanistically or cognitively, by using movie stimuli beyond the availability of inter-subject correlation.
Because of the above issues, a broader concern throughout the results is the largely descriptive nature of the findings. For example, the LASSO analysis shows that certain state transitions predict ISC in a subset of regions, with respectable R² values. While statistically robust, the manuscript provides little beyond why these particular transitions should matter, what computations they might reflect, or how they relate to known cognitive operations during movie watching. Similar issues arise in the clustering analyses. Clustering high-dimensional fMRI-derived features will almost inevitably produce structure, whether during rest, task, or naturalistic viewing. What is missing is an explanation of why these specific clusters are meaningful in functional or mechanistic terms.
Finally, the treatment of the thalamus, while very exciting, could use a bit more anatomical and circuit-level specificity. The manuscript largely treats the thalamus as a unitary structure, despite decades of work demonstrating big functional and connectivity differences across thalamic nuclei. A whole-thalamus analysis without more detailed resolution is increasingly difficult to justify. The subsequent subdivision into PVALB- and CALB-associated regions partially addresses this, but these markers span multiple nuclei with overlapping projection patterns.
-
Reviewer #2 (Public review):
Summary:
In this study, Liu et al. investigated cortical network dynamics during movie watching using an energy landscape analysis based on a maximum entropy model. They identified perception- and attention-oriented states as the dominant cortical states during movie watching and found that transitions between these states were associated with inter-subject synchronization of regional brain activity. They also showed that distinct thalamic compartments modulated distinct state transitions. They concluded that cortico-thalamo-cortical circuits are key regulators of cortical network dynamics.
Strengths:
A mechanistic understanding of cortical network dynamics is an important topic in both experimental and computational neuroscience, and this study represents a step forward in this direction by identifying key cortico-thalamo-cortical circuits. The analytical strategy employed in this study, particularly the LASSO-based analysis, is interesting and would be applicable to other data types, such as task- and resting-state fMRI.
Weaknesses:
Due to issues related to data preprocessing, support for the conclusions remains incomplete. I also believe that a more careful interpretation of the "energy" derived from the maximum entropy model would greatly clarify what the analysis actually revealed.
(1) Major Comment 1:
I think the method used for binarization of BOLD activity is problematic in multiple ways.
a) Although the authors appear to avoid using global signal regression (page 4, lines 114-118), the proposed method effectively removes the global signal. According to the description on page 4, lines 117-122, the authors binarized network-wise ROI signals by comparing them with the cross-network BOLD signal (i.e., the global signal): at each time point, network-wise ROI signals above the cross-network signal were set to 1, and the rest were set to −1. If I understand the binarization procedure correctly, this approach forces the cross-network signal to be zero (up to some noise introduced by the binarization of network-wise signals), which is essentially equivalent to removing the global signal. Please clarify what the authors meant by stating that "this approach maintained a diverse range of binarized cortical states in data where the global signal was preserved" (page 4, lines 121-122).
b) The authors might argue that they maintained a diverse range of cortical states by performing the binarization at each time point (rather than within each network). However, I believe this introduces another problem, because binarizing network-wise signals at each time point distorts the distribution of cortical states. For example, because the cross-network signal is effectively set to zero, the network cannot take certain states, such as all +1 or all −1. Similarly, this binarization biases the system toward states with similar numbers of +1s and −1s, rather than toward unbalanced states such as (+1, −1, −1, −1, −1, −1). These constraints and biases are not biological in origin but are simply artifacts of the binarization procedure. Importantly, the energy landscape and its derivatives (e.g., hard/easy transitions) are likely to be affected by these artifacts. I suggest that the authors try a more conventional binarization procedure (i.e., binarization within each network), which is more robust to such artifacts.
Related to this point, I have a question regarding Figure S1, in which the authors plotted predicted versus empirical state probabilities. As argued above, some empirical state probabilities should be zero because of the binarization procedure. However, in Figure S1, I do not see data points corresponding to these states (i.e., there should be points on the y-axis). Did the authors plot only a subset of states in Figure S1? I believe that all states should be included. The correlation coefficient between empirical and predicted probabilities (and the accuracy) should also be calculated using all states.
c) The current binarization procedure likely inflates non-neuronal noise and obscures the relationship between the true BOLD signal and its binarized representation. For example, consider two ROIs (A and B): both (+2%, +1%) and (+0.01%, −0.01%) in BOLD signal changes would be mapped to (+1, −1) after binarization. This suggests that qualitatively different signal magnitudes are treated identically. I believe that this issue could be alleviated if the authors were to binarize the signal within each network, rather than at each time point.
(2) Major Comment 2:
As the authors state (page 5, lines 145-148), the "energy" described in the energy landscape is not biological energy but rather a statistical transformation of probability distributions derived from the Boltzmann distribution. If this is the case, I believe that Figure 2A is potentially misleading and should be removed. This type of schematic may give the false impression that cortical state dynamics are governed by the energy landscape derived from the maximum entropy model (which is not validated).
-
Reviewer #1 (Public review):
The authors analysed large-scale brain-state dynamics while humans watched a short video. They sought to identify the role of thalamocortical interactions.
Major concerns
(1) Rationale for using the naturalistic stimulus
In terms of brain state dynamics, previous studies have already reported large-scale neural dynamics by applying some data-driven analyses, like energy landscape analysis and Hidden Markov Model, to human fMRI/EEG data recorded during resting/task states. Considering such prior work, it'd be critical to provide sufficient biological rationales to perform a conceptually similar study in a naturalistic condition, i.e., not just "because no previous work has been done". The authors would have to clarify what type of neural mechanisms could be missed in conventional resting-state studies using, say, energy landscape analysis, but could be revealed in the naturalistic condition.
(2) Effects of the uniqueness of the visual stimulus and reproducibility
One of the main drawbacks of the naturalistic condition is the unexpected effects of the stimuli. That is, this study looked into the data recorded from participants who were watching Sherlock, but what would happen to the results if we analyzed the brain activity data obtained from individuals who were watching different movies? To ensure the generalizability of the current findings, it would be necessary to demonstrate qualitative reproducibility of the current observations by analysing different datasets that employed different movie stimuli. In fact, it'd be possible to find such open datasets, like www.nature.com/articles/s41597-023-02458-8.
(3) Spatial accuracy of the "Thalamic circuit" definition
One of the main claims of this study heavily relies on the accuracy of the localization of two different thalamic architectures: matrix and core. Given the conventional or relatively low spatial resolution of the fMRI data acquisition (3x3x3 mm^3), it appears to be critically essential to demonstrate that the current analysis accurately distinguished fMRI signals between the matrix and core parts of the thalamus for each individual.
(4) More detailed analysis of the thalamic circuits
In addition, if such thalamic localisation is accurate enough, it would be greatly appreciated if the authors perform similar comparisons not only between the matrix and core architectures but also between different nuclei. For example, anterior, medial, and lateral groups (e.g., pulvinar group). Such an investigation would meet the expectations of readers who presume some microscopic circuit-level findings.
(5) Rationale for different time window lengths
The authors adopted two different time window lengths to examine the neural dynamics. First, they used a 21-TR window for signal normalisation. Then, they narrowed down the window length to 13-TR periods for the following statistical evaluation. Such a seemingly arbitrary choice of the shorter time window might be misunderstood as a measure to relax the threshold for the correction of multiple comparisons. Therefore, it'd be appreciated if the authors stuck to the original 21-TR time window and performed statistical evaluations based on the setting.
(6) Temporal resolution
After identifying brain states with energy landscape analysis, this study investigated the brain state transitions by directly looking into the fMRI signal changes. This manner seems to implicitly assume that no significant state changes happen in one TR (=1.5sec), which needs sufficient validation. Otherwise, like previous studies, it'd be highly recommended to conduct different analyses (e.g., random-walk simulation) to address and circumvent this problem.
-
-
momentummag.com momentummag.com
-
In 5 yrs Paris has doubled cycling's modal share, and become 5th ranking city in the Copenhagenize urban cycling index. Last summer ([[Paris Versailles 2025]]) we noticed the difference (compared to [[Paris 2021]] but also thought a lot still looked improvised not embedded yet. So probably some way until it's truly ingrained
-
-
www.biorxiv.org www.biorxiv.org
-
eLife Assessment
This study investigates whether heavy metal stress can induce maize-like phenotypic and molecular responses in teosinte and whether these responses overlap with genomic regions implicated in domestication. By combining copper and cadmium treatments with quantitative phenotyping, gene-expression analyses, and expanded assessments of nucleotide diversity across a key chromosome 5 interval, the authors provide an integrated view of how abiotic stress responses intersect with domestication-related traits. The significance of the findings is valuable, as the work offers meaningful insights for the subfield of maize evolution and stress biology by extending heavy-metal response analyses to teosinte and linking them to domestication-associated loci, although the evolutionary implications remain indirect. The strength of evidence is solid, with appropriately designed and quantitatively supported experiments that broadly support the claims, but do not yet establish a causal or historical role for heavy metal stress in domestication.
-
Author response:
The following is the authors’ response to the original reviews
Public Reviews:
Reviewer #1(Public review):
In this study, Acosta-Bayona et al. aim to better understand how environmental conditions could have influenced specific gene functions that may have been selected for during the domestication of teosinte parviglumis into domesticated maize. The authors are particularly interested in identifying the initial phenotypic changes that led to the original divergence of these two subspecies. They selected heavy metal (HM) stress as the condition to investigate. While the justification for this choice remains speculative, paleoenvironmental data would add value; the authors hypothesize that volcanic activity near the region of origin could have played a role.
The justification of choice to investigate the effects of heavy metal stress is not speculative. As mentioned now in the Abstract, the elucidation of the genome from the Palomero toluqueño maize landrace revealed heavy metal effects during domestication (Vielle-Calzada et al., Science 2009). Our aim was to test the hypothesis that heavy metal (HM) stress influenced the evolutionary transition of teosinte parviglumis to maize.
(1) Although the paper presents some interesting findings, it is difficult to distinguish which observations are novel versus already known in the literature regarding maize HM stress responses. The rationale behind focusing on specific loci is often lacking. For example, a statistically significant region identified via LOD score on chromosome 5 contains over 50 genes, yet the authors focus on three known HM-related genes without discussing others in the region. It is unclear why ZmHMA1 was selected for mutagenesis over ZmHMA7 or ZmSKUs5.
We appreciated the depth and value of this comment.
Maize phenotypic responses to sublethal concentrations to heavy metals – copper (Cu) and cadmium (Cd) in particular - are well characterized and published, and in agreement with our results. In the first section of the Results (pgs 7 and 8), we added pertinent references to clearly show which observations are already known. By contrast, teosinte parviglumis responses are in all cases novel. To our knowledge this is the first study that analyzed in detail the phenotypic response of teosinte to sublethal concentrations of heavy metals, specifically Cu and Cd. We have now emphasized the novelty of these observations (pg 8).
To address the fact that we only focused on three known HM-related genes without discussing others in the statistically significant region identified via LOD score on chr.5, we have added a full section that reads as follows (pgs. 11 to 13 of the new version):
“Large-scale genomic and transcriptomic comparisons indicate that many HM response genes were positively selected across the maize genome.
To expand the results well beyond the analysis of the three genes previously described, we performed a detailed analysis of genetic diversity across the 11.47 Mb genomic region comprised between Z_mSKUs5_ and ZmHMA1. This additional analysis reveals general tendencies in the quantity and nature of loci that were affected by positive selection during the teosinte parviglumis to maize transition in a region identified via LOD score on chr.5. We compared nucleotide variability by using 100 bp bins covering loci composed of two 30 Kb segments up and downstream of coding sequences, respectively, and the coding sequence itself, for 173 genes present within the genomic region comprised between ZmSKUs5 and ZmHMA (Figure S1 and Supplementary File 6). Two types of statistical tests (ANOVA and Wilcoxon) were applied to nucleotide variability comparisons using the entirety of each locus. The Benjamini-Hochber procedure allowed an estimation of the false discovery rate (FDR<0.05) to avoid type I errors (false positives). Although some individual loci appear as differently classified depending on the statistical test applied (22 out of 173 loci), the general differences in nucleotide variability are consistently maintained within the subregions described below. We found that 166 out of 173 loci show signatures of positive selection and are roughly organized in five independent subregions of variable length. The first six loci are consecutively ordered in a 402 Kb subregion that includes ZmSKUs5. A second group of 13 consecutive loci expands over a 1.44 Mb subregion that contains NRAMP ALUMINUM TRANSPORTER1, also involved in HM response through uptake of divalent ions. A third group of 17 consecutive loci expands over 1.28 Mb; eleven contain genes encoding for uncharacterized proteins. The fourth group is composed of 57 consecutive loci expanding over 3.22 Mb and contains genes encoding for DEFECTIVE KERNEL55, AUXIN RESPONSE FACTOR16, and peroxydases involved in responses to oxydative stress. The fifth group contains 12 consecutive loci expanding over 713 Kb and contains ZmHMA1. An additional segment of approximately 1.17 Mb and containing 25 consecutive loci that were positively selected expands away from the ZmSKUs5-ZmHMA1 segment; it also contains several genes encoding for peroxydases. Although multiple loci include genes that could be involved in abiotic stress and oxidative responses, these results suggest that multiple factors other than HM stress could have played a role in the evolutionary mechanisms that affected the genetic diversity of chr.5 during the teosinte parviglumis to maize transition.
To further analyze the possibility that HM response could have played a role in maize emergence and subsequent domestication, we analyzed large scale transcriptomic data corresponding to independent experiments aiming at understanding the response of maize roots to HM stress. Six available transcriptomes were selected for in-depth analysis because they presented a fold change strictly higher than 1, and their results were supported by false discovery rates (FDR<0.05). These six transcriptomes (Table S5) included HM response datasets corresponding to growth conditions that not only incorporated Cu, but also lead (Pb) and chromium (Cr) that were not included in the substrate of our experiments. Transcriptional profiles were obtained from roots of plants at different stages: maize seedlings (Shen et al., 2012; Gao et al., 2015; Zhang et al., 2024a), three week old plantlets (Yang et al., 2023), and plants at V2 stage (Zhang et al., 2024b; Fengxia et al., 2025). A total of 120 genes shared by all six transcriptomes were found to be differentially expressed under HM stress conditions (66 upegulated and 54 downregulated; Figure S3), including ZmSKUs5, ZmHMA1 and ZmHMA7; 52 of them (43.3%) are located in maize loci showing less than 70% of the nucleotide variability found in teosinte parviglumis, suggesting that they were affected by positive selection (Yamasaki et al., 2005; Supplementary File 7). Of 18 mapping in chr.5, twelve are within the 82 cM that fractionates into multiple QTLs under selection during the parviglumis to maize transition. Interestingly, five additional loci containing HM response genes completely lack SNPs within their total length in both parviglumis and maize, and 19 additional loci lack SNPs in at least one 30 Kb segment or their coding region (Supplementary File 7), suggesting the frequent presence of ultraconserved genomic regions in many loci containing HM response genes. When this same analysis was conducted in a set of loci comprising 63 genes previously identified as differentially expressed in response to abiotic stress not directly related to HM responses (hypoxia; nutritional deficiency; soil alkalinity; drought; soil salinity), 18 loci (28.6%) showed less than 70% of the nucleotide variability found in teosinte parviglumis. Only one of them maps in chr.5 and none contained segments or coding regions lacking SNPs in parviglumis or maize. These results suggest that in contrast to other types of abiotic stress response genes, loci comprising a large set of genes that unambiguously respond to HM stress caused by chemical elements of diverse nature were affected by positive selection during the parviglumis to maize transition, irrespectively of their position in the genome.”
The detailed analysis of genetic diversity across 11.47 Mb of chr.5 in the genomic region comprised between ZmSKUs5 and ZmHMA1 in presented as Supplementary File 6.
The analysis of genetic diversity in loci encompassing heavy metal response genes shared by six transcriptomes and abiotic stress controls are described in Supplementary File 7.
In the Discussion (pgs. 21 and 22), we added a paragraph section that reads as follows:
“Although loss of genetic diversity is usually the result of human selection during domestication, it can also represent a consequence of natural selective pressures favoring fitness of specific teosinte parviglumis allelic variants better adapted to environmental changes and subsequently affected by human selection during the domestication process. This possibility is reflected by widely spread selective sweeps affecting a large portion of chr.5 that contains hundreds of genes showing signatures of positive selection. The analysis of 11.47 Mb covering the ZmHMA1ZmSKUs5 segment confirms the presence of large but discrete genomic subregions that were positively selected during the teosinte parviglumis to maize transition. Although several contain genes involved in HM response and oxidative stress, the diversity of gene functions does not necessarily favor abiotic stress over other factors that could be at the origin of selective forces affecting these regions. By contrast, a large scale transcriptomic survey indicates that genes consistently responding to HMs (Cu, Cd, Pb and Cr ) show signatures of positive selection at unusual high frequencies (43.3%) as compared to loci containing genes responding to other types of abiotic stress (28.6%). Our identification of HM response genes affected by positive selection is far from being exhaustive. Nevertheless, it agrees with the expected effects of a widespread selective sweep caused by environmental changes that influenced the parviglumis to maize transition at the genetic level. Of intriguing interest are 24 loci that partially or completely lack SNPs in both teosinte parviglumis and maize, suggesting possible genetic bottlenecks occurred before the teosinte to maize transition. Examples of other edaphological factors driving genetic divergence either in the teosintes or maize include local adaptation to phosphorus concentration in mexicana and parviglumis (Aguirre-Liguori et al. 2019), and fast maize adaptation to changing iron availability through the action of genes involved in its mobilization, uptake, and transport (Benke and Stich 2011). Our results reveal a teosinte parviglumis environmental plasticity that could be related to the function of HM response genes positively selected during the teosinte parviglumis to maize transition. Previous studies have demonstrated that transposable elements (TEs) contribute to activation of maize genes in response to abiotic stress, affecting up to 20% of the genes upregulated in response to abiotic stress, and as many as 33% of genes that are only expressed in response to stress (Makarevitch et al., 2015). It is therefore possible that the HM response of some specific genes that influenced maize emergence or domestication could be mediated by TEs influencing or driving their transcriptional regulation.”
The mutagenic analysis of ZmHMA7 and ZmSKUs5 will be included in a different publication.
(2) The idea that HM stress impacted gene function and influenced human selection during domestication is of interest. However, the data presented do not convincingly link environmental factors with human-driven selection or the paleoenvironmental context of the transition. While lower nucleotide diversity values in maize could suggest selective pressure, it is not sufficient to infer human selection and could be due to other evolutionary processes. It is also unclear whether the statistical analysis was robust enough to rule out bias from a narrow locus selection. Furthermore, the addition of paleoclimate records (Paleoenvironmental Data Sources as a starting point) or conducting ecological niche modeling or crop growth models incorporating climate and soil scenarios would strengthen the arguments.
We think that the detailed analysis of genetic diversity across 11.46 Mb covering the ZmSKUs5 to ZmHMA1 genomic segment – and its statistical validation - provides a precise understanding of the selective sweep dimensions in chr.5.
We do agree that lower nucleotide diversity values in maize are not sufficient to infer human selection. Because many HM response loci show unusually low nucleotide variability in teosinte parviglumis (see the results of the transcriptomic analysis presented above), we cannot discard the possibility that natural selection forces related to environmental changes could have affected native populations of teosinte parviglumis.
To further explore the link between environmental factors, natural or human-driven selection, and the paleoenvironmental context of the parviglumis to maize transition, we revised paleoenvironmental and geological records and added results in two sections that read as follows (pgs. 17 to 20):
“Paleoenvironmental studies reveal periods of climatic instability in the presumed region of maize emergence during the early Holocene.
It is well accepted that temperature fluctuations, volcanism and anthropogenic impact shaped the distribution and abundance of plant species in the Transmexican Volcanic Belt (TMVB) during the last 14,000 years (Torrescano-Valle et al. 2019). The TMVB has produced close to 8000 volcanic structures (Ferrari et al., 2011), transforming the relief multiple times, and causing hydrographic and soil changes that actively modified the distribution and composition of plant communities in Central Mexico. Detailed paleoenvironmental data for the Pleistocene and Holocene is available for several lacustrine zones located within the 50 to 100 km range of the region currently considered the cradle of maize domestication (Matzuoka et al. 2002; Figure 5a). In Lake Zirahuén (102°44′ W; 19°26′ N and approximately 2075 meters above sea level; index [i] in Figure 5a), pollen, microcharcoal and magnetic susceptibility analyses of two sedimentary sequences reveals three periods of major ecological change during the early and middle Holocene.
Between 9500 and 9000 calibrated years before present (cal yr BP), pine forests seem to have been associated with summer insolation increases. A second peak of forest change occurred at around 8200 cal yr BP, coinciding with cold oscillations documented in the North Atlantic. Finally, events occurred between 7500 and 7100 cal yr BP shows an abrupt change in the plant community related to humid Holocene climates and a presumed volcanic event (Lozano-García et al., 2013). The environmental history of the central Balsas watershed has also been documented by pollen, charcoal, and sedimentary analysis conducted in three lakes and a swamp of the Iguala valley (Piperno et al. 2007). Paleoecological records of lake Ixtacyola (8°20N, 99°35W and approximately 720 meters above sea level; index [ii] in Figure 5a) and lake Ixtapa (8°21N, 99°26W) indicate that an important increase in temperature and precipitation occurred between 13000 and 10000 cal yr BP. The pollen record of Ixtacyola showed that members of the genus Zea were already part of the vegetation coverage by 12900 to 13000 cal yr BP, suggesting that some teosintes – likely including parviglumis - were commonly found at elevation areas where they do not presently occur. Lake Almoloya (also named Chignahuapan; 19°05N, 99°20E and approximately 2575 meters above sea level; index [iii] in Figure 5a) in the upper Lerma basin is only 20 Km from the crater of the Nevado de Toluca that is responsible for creating the late Pleistocene Upper Toluca Pumice layer over which the Lerma basin is deposited. Pollen records indicate the presence of Zea species by 11080 to 10780 cal yr BP. As for other locations, an important period of climatic instability prevailed between 11500 and 8500 cal yr BP (Ludlow-Wiechers et al., 2005). Humidity fluctuations occurred until 8000 cal yr BP, with a stable temperate climate between 8500 and 5000 cal yr BP. Although pollen and diatom studies are often difficult to interpret at a regional scale, the overall results presented above suggest consistent periods of Zea plants present in periods of environmental and climatic instability that correlate with the history of volcanic activity during the early Holocene, as described in the next section.
Temporal and geographical convergence between volcanic eruptions and maize emergence during the Holocene.
Current evidence indicates that the emergence and domestication of maize initiated in Mesoamerica some time around 9,000 yr BP (Matsuoka et al. 2002). The current location of teosinte parviglumis populations that are phylogenetically most closely allied with maize are currently distributed in a region located between the Michoacan-Guanajuato Volcanic Field (MGVF) at their northwest, and the Nevado de Toluca and Popocatéptl volcanoes at their east and northeast (Figure 5a; Matsuoka et al. 2002). Precise records of field data indicate that ten accessions were collected in the Balsas river drainage near Teloloapan and Sierra de Huautla (Guerrero), at approximately 100 km south of the Nevado de Toluca crater. Three other accessions were collected near Tejupilco de Hidalgo and Zacazonapan (Estado de México), at approximately 50 to 60 km from the Nevado de Toluca crater (8762, JSG y LOS-161, and JSG-391). And four other accessions were located in Michoacan, at a location within the MGVF (accession 8763), or at mid-distance between the MGVF and the Nevado de Toluca crater (accessions JSG y LOS-130, 8761, and 8766).
The most important source of HMs in ancient soils of Mesoamerica is TMBV-dependent volcanic activity through short- and long-term effects related to lava deposits, ores, hydrothermal flow, and ash (Torrescano-Valle et al. 2019). The Nevado de Toluca volcano produced one of the most powerful eruptions from central Mesoamerica in the Holocene, giving rise to the Upper Toluca Pumice deposit at 12621 to 12025 cal yr BP (Arce et al., 2003; Figure 5b). The pumice fallout blanketed the Lerma and Mexico basins with 40 cm of coarse ash (Bloomfield and Valastro 1977; Arce et al. 2003). A second eruption dated by 36Cl exposure occurred at 9700 cal yr BP (Arce et al. 2003; Figure 5b), and the most recent eruption occurred at 3580 to 3831 cal yr BP (Macías et al. 1997). During the early and middle Holocene, the Popocatéptl volcano produced at least four eruptions dated 13037-12060, 10775–9564, 8328-7591, and 6262-5318 cal yr BP (Siebe et al. 1997); three other important eruptions occurred during the late Holocene, between 2713 and 733 cal yr BP (Siebe and Macías, 2006). In addition, the MGFV is a monogenetic volcanic field for which 23 independent eruptions have been documented during the Holocene, 21 of them located towards the southern part of the field, in close proximity to the region harboring some of the teosinte parviglumis populations most closely related to maize. Three of these eruptions occurred in the early Holocene (El Huanillo 1130 to 9688 cal yr BP; La Taza 10649 to 10300 cal yr BP; Cerro Grande 10173 to 9502 cal yr BP; Figure 5b), and three others during the initial period of the middle Holocene, between 8400 and 7696 cal yr BP (La Mina, Los Caballos, and Cerro Amarillo; Figure 5b). On average, a new volcano forms every ~435 years in the MGFV (Macías and Arce, 2019). No less than 16 other eruptions occurred between 7159 cal yr BP and the present time (Figure 5b). Soils of volcanic origin (andosols) are currently distributed in regions north-west from the Nevado de Toluca and Popocatéptl craters, in close proximity with teosinte parviglumis populations most closely related to maize (Figure S5). Although modern distribution of teosinte populations may differ from their distribution around 9000 yr BP, and unknown populations more closely related to maize may yet to be discovered, this data indicates that the date and region where maize emerged is convergent with the dates and locations of several volcanic eruptions occurred during the Holocene in that same region.”
(3) Despite the interest in examining HM stress in maize and the presence of a pleiotropic phenotype, the assessment of the impact of gene expression is limited. The authors rely on qPCR for two ZmHMA genes and the locus tb1, known to be associated with maize architecture. A transcriptomic analysis would be necessary to 1- strengthen the proposed connection and 2- identify other genes with linked QTLs, such as those in the short arm of chromosome 5.
Real-time qPCR is an accurate and reliable approach to assess the expression of specific genes such as ZMHMA1 and Tb1, but we agree that our results do not allow to establish a direct regulatory link between the function of Tb1, the pleiotropic parviglumis phenotype under HM stress, and the function of ZmHMA1. We also concede that the large transcriptional analysis of HM response in maize (presented above) does not allow to elucidate a possible connection between these two genes. We have substantially downplayed our conclusion in this section by modifying the end of the section in pg. 17, that now reads:
“These results do not allow to directly link the regulation of ZmHMA1 expression to the function of Tb1; however, they open an opportunity to further investigate the possibility that under HM stress, the formation of secondary ramifications in teosinte parviglumis could be repressed by transcription factors of the TCP family, including Tb1.”
This is also emphasized in the Discussion (pg 21) as follows:
“Under HM stress, we also show that Tb1 is overexpressed in the apical meristem of teosinte parviglumis, suggesting that formation of secondary ramifications is repressed by Tb1 function under HM stress, as in extant maize. At this stage we cannot discard the possibility that Tb1 upregulation in parviglumis reflects a more generalized response to abiotic stress; however, the expression ZmHMA1 is downregulated in W22 wild-type maize meristems in the presence of HMs but upregulated in teosinte parviglumis meristems, suggesting that a specific regulatory shift relating HM responses and ZmHMA1 function occurred during the teosinte parviglumis to maize transition.”
On the other hand, the transcriptional analysis the identification of 52 additional HM response genes showing signatures of positive selection occurred during the parviglumis to maize transition; 12 of them map to chr.5 within the region having linked QTLs within the short arm of chr.5. So far, genes involved in HM response and oxidative stress represent the most prevalent class of genes identified within the genomic region showing pleiotropic effects on domestication and multiple linked QTLs in chr.5.
Reviewer #2 (Public review):
Summary:
This work explores the phenotypic developmental traits associated with Cu and Cd responses in teosinte parviglumis, a species evolutionary related to extant maize crops. Cu and Cd could serve as a proxy for heavy metals present in the soils. The manuscript explores potential genetic loci associated with heavy metal responses and domestication identified in previous studies. This includes heavy metal transporters, which are unregulated during stress. To study that, the authors compare the plant architecture of maize defective in ZmHMA1 and speculate on its association with domestication.
Strengths:
Very few studies covered the responses of teosintes to heavy metal stress. The physiological function of ZmHMA1 in maize also gives some novelty in this study. The idea and speculation section is interesting and well-implemented.
Weaknesses:
The authors explored Cu/Cd stress but not a more comprehensive panel of heavy metals, making the implications of this study quite narrow. Some techniques used, such as end-point RT-PCR and qPCR, are substandard for the field. The phenotypic changes explored are not clearly connected with the potential genetic mechanisms associated with them, with the exception of nodal roots. If teosintes in response to heavy metal have phenotypic similarity with modern landraces of maize, then heavy metal stress might have been a confounding factor in the selection of maize and not a potential driving factor. Similar to the positive selection of ZmHMA1 and its phenotypic traits. In that sense, there is no clear hypothesis of what the authors are looking for in this study, and it is hard to make conclusions based on the provided results to understand its importance. The authors do not provide any clear data on the potential influence of heavy metals in the field during the domestication of maize. The potential role of Tb-1 is not very clear either.
Thank you for these comments. We have now emphasized our hypothesis in the abstract and the last paragraph of the Introduction (pg. 6):
“To test the hypothesis that heavy metal (HM) stress influenced the evolutionary transition of teosinte to maize, we exposed both subspecies to sublethal concentrations of copper and cadmium etc…”
A comprehensive panel of heavy metals would not be more accurate in terms of simulating the composition of soils evolving across 9,000 years in the region where maize presumably emerged. Copper (Cu) and cadmium (Cu) correspond each to a different affinity group for proteins of the ZmHMA family. ZmHMA1 has preferential affinity for Cu and Ag (silver), whereas ZmHMA7 has preferential affinity to Cd, Zn (zinc), Co (cobalt), and Pb (lead). Since these P1b-ATPase transporters mediate the movement of divalent cations, their function remains consistent regardless of the specific metal tested, provided it belongs to the respective affinity group. By applying sublethal concentrations of Cd (16 mg/kg) and Cu (400 mg/kg), we caused a measurable physiological response while allowing plants to complete their life cycle, including the reproductive phase, facilitating a comprehensive analysis of metal stress adaptation. Whereas higher doses impair flowering or are lethal, lower Cu/Cd concentrations do not consistently show conventional phenotypic responses such as reduced plant growth (AbdElgawad et al. 2020; Atta et al., 2023)
Based on comments by both reviewers, we present now a large transcriptional analysis that incorporates HM responses to lead (Pb) and chromium (Cr), in addition to Cu. Results show that many genes responding to Pb and Cr were also positively selected across the maize genome, suggesting that HM stress led to a ubiquitous rather than a specific evolutionary response to heavy metals (please see our response to Reviewer#1 and sections in pgs. 11 to 13) .
Real-time qPCR is an accurate and reliable approach to assess the expression of specific genes such as ZMHMA1 and Tb1, but we agree that our results do not allow to establish a direct regulatory link between the function of Tb1, the pleiotropic parviglumis phenotype under HM stress, and the function of ZmHMA1. We also concede that the large transcriptional analysis of HM response in maize (presented above) does not allow to elucidate a possible connection between these two genes. Therefore, we have substantially downplayed our conclusion in this section by modifying the end of the section in pg. 17, that now reads:
“These results do not allow to directly link the regulation of ZmHMA1 expression to the function of Tb1; however, they open an opportunity to further investigate the possibility that under HM stress, the formation of secondary ramifications in teosinte parviglumis could be repressed by transcription factors of the TCP family, including Tb1.”
There are two phenotypic changes clearly connected with the genetic mechanisms involved in the parviglumis to maize transition: plant height and the number of seminal roots (not nodal roots). These changes have been now emphasized in the Abstract and the description of the results.
Regarding the possibility for HM stress to represent a confounding factor in the selection of maize and not a driving factor, we expanded the genomic analysis of genetic diversity well beyond the analysis of the three genes under initial study, to cover a segment of 11.47 Mb comprised between ZmSKUs5 and ZmHMA1. We compared nucleotide variability by using 100 bp bins covering loci composed of two 30 Kb segments up and downstream of coding sequences, respectively, and the coding sequence itself, for 173 genes present within the genomic region comprised between ZmSKUs5 and ZmHMA (Figure S1 and Supplementary File 6). The full analysis is presented in a new section pgs. 11 and 12. We found that 166 out of 173 loci show signatures of positive selection and are roughly organized in five independent subregions of variable length. Four out of five subregions contain more than one HM or oxidative stress response gene within loci showing signatures of positive selection. Although multiple factors other than HM stress could have played a role in the evolutionary mechanisms that affected the genetic diversity of chr.5, large scale transcriptomic data corresponding to independent experiments aiming at understanding the response of maize roots to HM stress allowed the identification of 49 additional HM response genes within loci showing positive selection across the genome, a proportion (43.3%) far greater than the proportion of loci containing response genes to other types of abiotic stress not related to HMs (28.6%). These results are described in detail in pgs. 12 and 13 (Figure S3 and Supplementary File 7). These results provide strong evidence in favor of HM stress and not another factor driving positive selection.
We now provide precise and pertinent paleoenvironmental data on the potential influence of heavy metals in the field. In sections pgs. 17 to 20 we review paleoenvironmental studies revealing periods of climatic instability in the presumed region of maize emergence during the early Holocene, and data indicating that the date and region where maize emerged is convergent with the dates and locations of several volcanic eruptions occurred during the early and middle Holocene in that same region. Please see responses to Reviewer#1 for details.
We agree that our results do not allow to establish a direct regulatory link between the function of Tb1, the pleiotropic parviglumis phenotype under HM stress, and the function of ZmHMA1. We also concede that the large transcriptional analysis of HM response in maize (presented above) does not allow to elucidate a possible connection between these two genes. Therefore, we have substantially downplayed our conclusion in this section by modifying the end of the section in pg. 17, that now reads:
“These results do not allow to directly link the regulation of ZmHMA1 expression to the function of Tb1; however, they open an opportunity to further investigate the possibility that under HM stress, the formation of secondary ramifications in teosinte parviglumis could be repressed by transcription factors of the TCP family, including Tb1.”
This is also emphasized in the Discussion (pg 21) as follows:
“Under HM stress, we also show that Tb1 is overexpressed in the apical meristem of teosinte parviglumis, suggesting that formation of secondary ramifications is repressed by Tb1 function under HM stress, as in extant maize. At this stage we cannot discard the possibility that Tb1 upregulation in parviglumis reflects a more generalized response to abiotic stress; however, the expression ZmHMA1 is downregulated in W22 wild-type maize meristems in the presence of HMs but upregulated in teosinte parviglumis meristems, suggesting that a specific regulatory shift relating HM responses and ZmHMA1 function occurred during the teosinte parviglumis to maize transition.”
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
While the dataset generated provides an interesting foundation for hypothesis testing on HM stress and domestication, the current data do not sufficiently support the conclusions of the manuscript.
(1) The description of maize and teosinte architecture under HM stress is well presented.
However, traits like shoot height, leaf size reduction, and biomass loss also occur under other environmental stresses such as drought and salinity. Additional evidence beyond shoot and root architecture would help validate the link between tb1 expression and specific ZmHMA genes under HM stress, or whether it reflects a more generalized stress response.
We have already addressed in detail this point in the public response to Reviewer#1.
(2) The nucleotide variability analysis is interesting, but I would have liked to see additional information to clarify the choice of the data selection and the strength of the conclusions with human selection.
We have already addressed in detail this point in the public response to Reviewer#1.
a) The choice of Tripsacum dactyloides as the outgroup to determine nucleotide variability seems to be distant, and I wonder whether other combinations with a closer outgroup or multiple outgroups were tried to provide a more accurate context.
Nucleotide variability in Tripsacum dactyloides is used to graphically illustrate an external reference and not as an outgroup in the extended analysis of genetic diversity at the locus and genomic level. We did not used Tripsacum dactyloides as an outgroup in our statisticalm analysis. We could have indeed a closer teosinte subspecies as an outgroup, but at this stage no data warrants that environmentally-related selective pressures could have affected genetic diversite in other teosintes. This possibility in currently being investigated.
b) Evolutionary differences not related to human influence could affect the results. The phrase "order of magnitude difference in π values" needs statistical validation (e.g., confidence intervals, p-values).
We agree and have eliminated the sentence, as it is no longer relevant at the light of the detailed genomic analysis of genetic diversity prsented in Supplementary File 6.
c) The comparison with ZmGLB1, a neutral control locus, suggests that domestication-related changes in nucleotide variability are specific to the three candidate genes. However, the concept of neutrality is complex, and while ZmGLB1 may be considered neutral in this case, the argument does not address the possibility of other factors, such as linked selection, that could influence variability in these genes. Referencing Hufford et al. is insufficient and would require a deeper argument.
We also agree with this comment. We think that the influence and consequences of linked selection are now well documented for 11.46 Mb analyzed in chr.5 (pgs 11 and 12) in the main text and Supplementary File 6).
(3) The statement: "Our evidence indicates that HM stress revealed a teosinte parviglumis environmental plasticity that is directly related to the function of specific HM response genes that were affected by domestication through human selection" is not supported by the presented data. The rationale for the specific Cd/Cu dosage used is unclear. A dose-response gradient would better demonstrate the nature and strength of the plastic response.
Previous reports support the rationale for the specific HM dosage in this study; Cu/Cd dosage response gradients have been conducted in maize (AbdElgawad et al. 2020; Atta et al., 202), but since no studies have been conducted in teosinte, we reasoned that it was important to apply the same treatment to both subspecies. We have now emphasized this rationale by adding the following in pg XX: “Whereas higher doses impair flowering or are lethal, lower Cu/Cd concentrations do not consistently show conventional phenotypic responses such as reduced plant growth (AbdElgawad et al. 2020; Atta et al., 2023)”.
We agree that the statement raised by the reviewer needed revision at the light of our results. We did revise the statement to accurately reflect our current evidence as follows: “Our results reveal a teosinte parviglumis environmental plasticity that is likely related to the function of HM response genes positively selected during the teosinte parviglumis to maize transition.”
(4) In maize, TEs are known to influence gene expression under abiotic stress, including for tb1 (PMID: 25569788). Since the author appears to make a causative conclusion between ZmHMA1, TB1, and HM stress, I would have liked to see a whole-transcriptome analysis and not a curation of two genes to determine whether other factors, such as TEs, can have that would lead to similar outcomes.
We agree that is definetely a possibility that we have not investigated at this stage. However, we added a pargraph to reflect this pertinent suggestion:
“Previous studies have demonstrated that transposable elements (TEs) contribute to activation of maize genes in response to abiotic stress, affecting up to 20% of the genes upregulated in response to abiotic stress, and as many as 33% of genes that are only expressed in response to stress (Makarevitch et al., 2015). It is therefore possible that the HM response of some specific genes that influenced maize emergence or domestication could be mediated by TEs influencing or driving their transcriptional regulation.”
(5) I would suggest that the authors carefully review the tables, figures, and the corresponding legends. For example :
a) Table 2 is called before Table 1, I would therefore suggest changing the numbering to reflect the paragraph order.
Thank you for your help, we did change the order of the Tables in the new version.
b) In Table 2, it is not clear whether the P value applies to the mean difference between WT and the mutant zmhma1, either in the presence or the absence of heavy metals. In addition, the authors need to use the P-value to estimate the differences between WT in the absence vs presence of HM, and WT in the absence of HM versus the mutant in the absence of HM (idem for presence).
We did address this issue in detail and added P-values and specific pairwise comparisons to that Table (now Table 1). Data are presented as mean ± standard deviation and were tested by a paired Student’s T-Test. When the effects were significant according to T-Test, the treatments were compared with the Welch two sample T-Test at P < 0.05.
c) Table 1 and Table 2: Indicate what type of statistical test was used and the number of plants used for each experiment (n). Also, I recommend the use of scientific notation for the P-values.
The statistical tests have now been indicated, scientific notation has been added to the P-values; the number of plants and biological replicates are indicated in the Methods section.
d) Lines 202 and 204: I assume Table 1 should be called instead of Table 2.
This error has been corrected.
e) General: In the text, when significance is highlighted along with measurements, the p-value needs to be added.
We have added the P-value along the measurement for all significant differences.
f) In the text, it is also mentioned that "the expression of ZMHMA1 was significantly increased in the presence of HMs (Figure 3c)". We are looking here at an RT-PCR, which is qualitative and without a robust quantitative comparison and statistics, I cannot conclude this assessment based on the presented evidence. No statistical measure is indicated here.
Panel 3c is not RT-PCR but a real-time qPCR, showing relative fold-change, normalized to actin, with a 3-technical triplicate per 3 biological replicates). We have added error bars (SD) and P-values represented by asterisks (calculated with Student's t statistic) to support significant differences (P<0.05 and P<0.01). ZmHMA1 expression was significantly increased in the presence of HMs only in teosinte; there was no significant difference in maize.
g) Figure 3 should at least have the gene name in the figure to quickly understand the figure panel. The key conserved domains should also be identified.
We agree and apologize for the omission. The gene names have been added adjacent to the structures.
h) Sentence at lines 459-460 lacks words and punctuation.
This unfortunate rror has also been corrected.
i) Figure S1, the reference Lemmon and Doebley, 2024 should be Lemmon and Doebley, 2014 to harmonize with the text.
The correct year is 2014. We have corrected this error.
Reviewer #2 (Recommendations for the authors):
(1) The narrative should be clearer, starting with a clearer hypothesis that is later sustained or not in the results, and then discussed in the idea and speculation section.
Thank you for the comment. We have clarified the hypothesis, it is included in the abstract and the last paragraph of the Introduction. We hope it is now clear that the evidence presented supports our hypothesis
(2) Focus more on traits that are relevant, for example, nodal and seminal roots.
We modified the text to emphasize three relevant traits. In the case of teosinte under HM stress, absence of tillering and increase in the number of female inflorescences. In the case of the zmha1 mutant under HM stress, differences in the number of nodal roots, and differences in height.
(3) RNA-seq in Cu/Cd stress could make the work much more useful and complete.
As previously mentioned, we have incorporated a large scale transcriptional analysis on the basis of six transcriptomes statistically validated (Table S5). Please see sections pgs. 11 to 13 for details.
-
-
social-media-ethics-automation.github.io social-media-ethics-automation.github.io
-
And they also talked about the advertising motives behind supporting social causes (even if some employees do indeed support them), and the advertising motivation behind tweeting openly about how they are basing their decisions based on advertising.
I never see them doing this as an issue, as behind the screen of who is running the brand account is also a human, and that being able to break free from professionalism and connecting with people online, and express their opinions can be seen as a good thing, but also it can be a brilliant marketing strategy. With many brands being able to relate to and speak about the problems today help show that the corporation also cares about the people, which can help them gain more supporters and customers. But sometimes, if the topic is really controversial, or something that shouldn't be said online, some companies will still speak on it, which may hurt their companies image.
-
-
-
blogger Fabrizio Ferri Benedetti on their 4 modes of using AI in technical writing. - watercooler conversations, to get code explained - text suggestions while writing/coding (esp for repeating patterns in your work - providing context / constraints / intent to generate first drafts, restructure content, or boilerplate commentary etc. - a robotic assembly line, to do checks, tests and rewrites. MCP/skills involved.
Not either/or but switching between modes
-
-
social-media-ethics-automation.github.io social-media-ethics-automation.github.io
-
How do you think about the authenticity of the Tweets that come from others in Trump’s campaign?
Unfortunately, I believe in the authenticity of others in Trump's campaign. I don't believe in the truthfulness/factuality, but I believe a lot of MAGA genuinely believes the literal lies they are saying. There are definitely many who are lying through their teeth for their own benefit and know it, the leaked group chats basically prove that. However, every movement needs true believers, and never underestimate the power of brainwashing.
-
In 2016, when Donald Trump was running a campaign to be the US President, one twitter user pointed out that you could see which of the Tweets on Donald Trump’s Twitter account were posted from an Android phone and which from an iPhone, and that the tone was very different. A data scientist decided to look into it more and found: “My analysis … concludes that the Android and iPhone tweets are clearly from different people, “posting during different times of day and using hashtags, links, and retweets in distinct ways, “What’s more, we can see that the Android tweets are angrier and more negative, while the iPhone tweets tend to be benign announcements and pictures. …. this lets us tell the difference between the campaign’s tweets (iPhone) and Trump’s own (Android).” (Read more in this article from The Guardian) Note: we can no longer run code to check this ourselves because first, Donald Trump’s account was suspended in January 2021 for inciting violence, then when Elon Musk decided to reinstate Donald Trump’s account (using a Twitter poll as an excuse, but how many of the votes were bots?), Elon Musk also decided to remove the ability to look up a tweet’s source.
This analysis intrigued me, and it was the first time I realized that a data scientist could reasonably infer which tweets were likely to have come from Trump himself and which from his campaign. This shows that even seemingly simple metadata can contain very strong behavioral signals. This made me realize that platforms are not neutral technological Spaces, but systems that are influenced by power, economic interests, and individual decisions.
-
-
human.libretexts.org human.libretexts.org
-
1.3.186) means Act 1, Scene 3, Line 186.
I didn't know this but it seems really helpful and makes a lot of sense!
-
but the bulk of the production's effect is generated through the dialogue and its visual devices, and since the play's script dictates what the characters will say and often, through stage direction, its production strategies as well, the script is crucial to a successful performance.
A lot of the time stage direction is very important to the overall story of a play because it also conveys indication to the audience about what is going on.
-
Although a novel’s narrator can describe in detail the thoughts and impressions of its characters, a play's effects depend much more heavily on what the characters say and do.
The reason why I choose this quote is because is an authors writing is part of telling a good story. But when the story becomes a play, it helps make the emotions feel more real. Emotions are needed to help make a play feel "real" to the audience. Making the story more impactful.
-
-
edgeeffects.net edgeeffects.net
-
Misogyny dictated these bounds, but one beneficial side effect was that common plants were preserved by women even as they were dismissed by male scientists.
Everything is worth archiving! I'm glad these women were able to conserve "common' plants disregarded by their male counterparts. We never know how impermanent seemingly permanent things are. It reminds me of chestnut trees! Once everywhere in Appalachia, they nearly all died due to chestnut blight.
-
-
human.libretexts.org human.libretexts.org
-
When teachers use any of these terms, it usually means engaging with a text on a deeper level than when reading for entertainment or information: that is, looking not just at what is written (the message, also known as content), but how it is written (the methods used to shape the message, also known as form).
Literary analyses are meant to get the readers thinking and look into the author's purpose into why they made the text the way it is. Not getting readers to think it is simply just words written but meaning.
-
-
bookshelf.vitalsource.com bookshelf.vitalsource.com
-
Akhenaten’s theology was closer to monotheism because, unlike traditional Egyptian deities, the Aten could not be imagined as taking on human or animal form. If this were not controversial enough, Akhenaten also celebrated his new religion by representing himself in a very unconventional way. In a complete departure from the divine virility of his ancestors—which even his ancestor Hatshepsut had emulated—Akhenaten had himself pictured as a normal human being with distinctive features: a family man enjoying the company of his wife, Nefertiti, and their children. This emphasis on his own humanity was an extension of his theology, which honored the life force within every being. But it was very dangerous to the ideology of royal power. The pharaoh was not supposed to be an approachable man with a quirky personality. He was supposed to be a god
It demonstrates one of the greatest religious transformations of Egypt. It is notable because it demonstrates that a pharaoh could threaten established religious traditions, which had both political and spiritual repercussions.
-
-
blog.hubspot.com blog.hubspot.com
-
If you work for a small business, it’s likely difficult for you to compete with the major brands in your industry, many of which have millions of dollars to invest in campaigns. Fortunately, there are plenty of opportunities to outrank the big players through strategic digital marketing initiatives. For instance, you might use long-tail keywords to create high-quality content that ranks on search engines. Search engines don’t care which brand is the biggest, but they do care about prioritizing content that resonates best with target audiences.
I find this point interesting because it shows that online marketing gives smaller businesses a fair chance to compete with bigger brands. You don’t need a huge budget to be successful if you use smart strategies like targeting the right audience and creating content that actually connects with people. I think this is a good point because search engines focus more on relevance than brand size, which allows smaller or newer brands to still get noticed and grow.
-
-
nmoer.pressbooks.pub nmoer.pressbooks.pub
-
while, however, nevertheless, but, and similarly, as well as phrases like on the other hand and for example, can serve as transitions between sentences and ideas.
strong transition words
-
-
bookshelf.vitalsource.com bookshelf.vitalsource.com
-
Consumers often seek out retailers that stand for the social values that they embrace. In a recent survey, 66 percent of consumers indicated they would consider switching to a new product if it meant they could purchase from a company dedicated to a brighter future.36 Such considerations are important to stakeholders other than consumers too. In line with its long-standing reputation for taking a stand on various societal issues, Ben & Jerry’s launched an ongoing campaign, “Save Our Swirled,” to bring awareness to the need to heed the UN’s Paris climate accords.37
This excerpt highlights the growing importance of social values in consumer decision-making. Many consumers prefer to support retailers whose ethical, environmental, or social commitments align with their own beliefs, and are even willing to switch products to do so. The example of Ben & Jerry’s illustrates how companies can actively promote social and environmental causes as part of their brand identity, appealing not only to consumers but also to other stakeholders who value corporate responsibility.
-
-
www.kjzz.org www.kjzz.org
-
DINGMAN: Duane says that from the time he opened his typewriter shop, until about 2008, he probably repaired about 50 manual typewriters. But in 2008, something changed. His phone started ringing off the hook. Parents were calling to say that their daughters wanted a typewriter for Christmas — could they bring in an old one for him to fix up? Or did he have any for sale?JENSEN: So I asked ‘em, I said why are you interested? They said, “She watched this movie called “Kitt Kittredge.”
-
-
site.xavier.edu site.xavier.edu
-
Duane Jensen, J.C. Business Machines, 26th St. & Union Hills, Phoenix, AZ. 602-992-7611. M-F 9-4, but call first. "We fix business machines, including all typewriters. Manual portable, manual uprights, antique or vintage typewriters. We carry every typewriter ribbon manufactured. Basic cleaning / tune up is between $38.00 - 48.00 (walk in). On-site service. Basic repair / clean $68.00 - 88.00." Read a story about an experience with Mr. Jensen here. Website: http://www.phoenixtypewriter.com/
Duane Jensen's death was announced by his family on 2026-01-25.
-
-
nmoer.pressbooks.pub nmoer.pressbooks.pub
-
Research shows that not only do students encode the material at a deeper level than students who do not do this step, but while you organize your annotations, your brain is reviewing the information, so you will spend less time studying later.
Studies show compression stays long enough to implement later by organizing and note taking
-
The third planning strategy is setting a purpose for your reading. Knowing what you want to achieve from a reading assignment not only helps you determine how to approach that task, but it also helps you stay focused during those moments when you are up late, already tired, or unmotivated because relaxing in front of the television sounds far more appealing than curling up with a stack of journal articles. Sometimes your purpose is simple. You might just need to understand the reading material well enough to discuss it intelligently in class the next day. However, your purpose will often go beyond that. For instance, you might also need to read in order to compare two texts, to formulate a personal response to a text, or to gather ideas for future research.
being punctual with assignments and having in mind the instructor's rules to the assignments will guide you to be successful.
-
Managing your time – Now that you know how many sections make up the entire reading assignment, focus on setting aside enough time for reading and breaking the assignment into manageable chunks. For example, if you are assigned a seventy-page chapter to read for next week’s class, it is best not to wait until the night before to get started. How you choose to break up the reading assignment will depend on the type of reading it is. If the text is dense and packed with unfamiliar terms and concepts, you may need to read no more than five or ten pages in one sitting so that you can truly understand and process the information. With more user-friendly texts, you will be able to handle more pages in one sitting. And if you have a highly engaging reading assignment, such as a novel you cannot put down, you may be able to read lengthy passages in one sitting.
It is always good to read in parts, annotate what you have read, but if you have a good memory and remember everything at ones than that's also okay. I like to split my readings in different parts than reading all at once and remembering everything.
-
Finding the main point gives you a framework to organize the details presented in the reading and relate the reading to concepts you learned in class or through other reading assignments.
Understanding and identifying the authors main point will not only help you summarize the text, but it will help you become a better writer. You can use their style of writing as a reference when brain storming on how effectively get your point across when writing a paper.
-
research shows that cramming and procrastinating have to do with emotional dysregulation that can be helped with good time management skills.
I tend to want to do everything at once and have it all completed as quickly as possible, but that only overwhelms (emotional regulation) me and allows room for mistakes. Giving myself realistic time lines allows me to better manage my time and being successful with my tasks.
-
-
socialsci.libretexts.org socialsci.libretexts.org
-
provide a more accurate understanding of the political, cultural, historical, and social contexts that shape our current-day experiences of race, identity, community, and the disparities that plague our society.
i do feel like that this does create a sense of community or belonging especially among POC, because we are able to see our people and cultures represented and learn things about our history etc that we may not have even known as well as advocate and become aware of the different issues not only in our communities but in others communities. because its not taught in basic school curriculum.
-
-
drive.google.com drive.google.com
-
but it is the struggle to have blacksperceived as agents, as people with cognitive capacities and even with anintellectual history-attributes denied by modem racism-that is for methe primary reason for writing this book
I wonder how this assertion of agency compares to that of other assertions like subaltern studies.
-
Delany's appeal todayis that of a supreme patriarch
His liberatory ethic is characterized by masculine notions of citizenship and ownership, the ability not only to lead but to subject others to your will
-
-
www.kennedy-center.org www.kennedy-center.org
-
mental representations of what is observed or imagined; symbolic, allegorical, and metaphorical representations; careful observation of the world;
I love art for many reasons, but one that I have been thinking about the most as of late is how it prepares people to be good humans in their adult life. I am a firm believer that if we all made sure to include some kind of creative or artistic endeavor we would all be more connected to our humanity. Studying art teaches us how to observe the people and their intentions. We think more about the "why" and put ourselves in other peoples shoes. When we critique art, we often go beyond what we see in a painting, and also take into consideration the artist and their life, where they were from, and what techniques they used. Teaching kids how to do this can help familiarize them with this high level of "seeking to understand", that can only help them later on.
-
Arts integration provides opportunities for students to learn to be open and responsive to diverse perspectives
I think that art is one of the most common ways students are taught about diversity. They are able to explore different cultures, eras, and perspectives in a way that appeals to their senses. When children see portraits, touch sculptures, or listen to music, they can be exposed to a whole new world that they otherwise wouldn't have access to. Beyond human diversity, it's also interesting to see how art can put on display the diverseness in our personalities. I love art projects that have the same prompt, but give students the freedom to make it their own. Doing a gallery walk and getting to see all of the differences is so fun.
-
-
www.whatsbetter.today www.whatsbetter.today
-
The Wind (Ruach) provides the energy
Pneuma Mechanics. The Hebrew Ruach and Greek Pneuma both translate to "Wind," "Breath," or "Spirit." In John 3:8, Jesus describes the Kingdom-led life as being like the wind—unpredictable in origin but undeniable in effect. Trying to "be the engine" is an attempt to operate by the Sarx (flesh/effort). The Sailboat mode is the biblical technology of being "led by the Spirit," where the power source is external and infinite.
-
-
bookshelf.vitalsource.com bookshelf.vitalsource.com
-
Consideration of both genetic and social influences provides the most complete understanding, and efforts to integrate the two should continue.
I feel like this isn’t usually stressed enough. People always say nature vs. nurture, but the combination of the two is so important.
-
This shift emphasized the importance of social characteristics
I feel like this was definitely an important shift that was made since it makes sure to look at not only the biological causes, but also the social patterns. The biology of it all doesn’t always give the full picture and sometimes the social characteristics can really help.
-
-
saalck.pressbooks.pub saalck.pressbooks.pub
-
What does your receiver need to know?
The receiver needs to know that I am not going to make it into work today to help set up for the project but that I will do all I can to try to arrange for me to help remotely or make a list of tasks to do when I come back
-
-
saalck.pressbooks.pub saalck.pressbooks.pub
-
The outcome that I want to achieve is to directly portray to my boss that I have food poisoning and that it randomly came about. The relationship I want to have with my Boss is a professional relationship but also I would want him to get to know me on a personal level and how I am a hard worker and this project meant a lot to me. 3-5 adjectives I would want my receiver to describe me as is Formal, responsible, sympathetic, hard working, and effective communication
-
-
go-gale-com.offcampus.lib.washington.edu go-gale-com.offcampus.lib.washington.edu
-
the most often challenged were those in the Harry Potter series, for its focus on wizardry and magic and "Satanic influence."
It is incredibly interesting that witchcraft and secular humanism and the like are demonized but the bible isn't.
-
-
social-media-ethics-automation.github.io social-media-ethics-automation.github.io
-
She highlights the emotional labor of keeping emotional expressions consistent with their public persona, even when they are having different or conflicted feelings.
Influencers, and even ordinary users, often create and maintain a consistent public or social media persona. For example, a beauty influencer might present themselves as a dedicated skincare expert who puts an extreme amount of effort into their routines, hoping others will follow their example. This kind of carefully curated identity can feel authentic to viewers but may blur the line between genuine self-expression and performance.
-
-
nmoer.pressbooks.pub nmoer.pressbooks.pub
-
In academic settings, the reasons for writing often fulfill four main purposes: to summarize, to analyze, to synthesize, and to evaluate. You will encounter these four purposes not only as you read for your classes but also as you read for work or pleasure. Because reading and writing work together, your writing skills will improve as you read.
as we read, we can write and annotate what you need to know to have the article. analyze the article think about questions about the article. compare to the rough draft. rewite and put it together.
-
After thoroughly reading the assignment sheet, you might not have questions right away. However, after reading it again, either before or after you try to start the assignment, you might find that you have questions. Don’t play a guessing game when it comes to tackling assignment criteria–ask the right person for help: the instructor. Discuss any and all questions with the person who assigned the work, either in person or via email. Visit him or her during office hours or stay after class. Do not wait until the last minute, as doing so puts your grade at risk. Don’t be shy about asking your professors questions. Not only will you better your understanding and the outcome of your paper, but professors tend to enjoy and benefit from student inquiry, as questions help them rethink their assignments and improve the clarity of their expectations. You are probably not the only student with a question, so be the one who is assertive and responsible enough to find answers. In the worst case scenario, when you have completed all of these steps and a professor still fails to provide you with the clarity you are looking for, discuss your questions with fellow classmates.
I do feel like sharing ideas is not bad, when we need help, we don't always walk to the instructor maybe we just ask a classmate and have ideas and become to one another.
-
An assignment sheet may be lengthy, but resist the temptation to skim it. Observe and interpret every detail of the text. Moreover, it is essential to focus on the keywords of the subject matter being discussed. It would be unfortunate to hand in an incomplete or misguided assignment because you did not properly read and understand the guidelines. Since you can easily overlook details on the first reading, read the assignment sheet a second time. As you are reading, highlight areas where you have questions, and also mark words you feel are particularly important. Ask yourself why your professor assigned this task. How does it relate to what you are studying in class? Pay attention to key words, such as compare, contrast, analyze, etc. Who is your audience? Should the paper be written in a formal or informal tone? Is there documentation required?
perfect annotations go based a good idea or a note that you might have taken during the article reading.
-
-
socialsci.libretexts.org socialsci.libretexts.org
-
The two years of life are ones of dramatic growth and change. A newborn, with a keen sense of hearing but very poor vision,
This Is very true a lot of newborns are very good with listening and I've read before that them listening is a way their vocabulary increases because a lot of the words at birth until them are very familiar to them and easier to use and understand.
-
-
geo.libretexts.org geo.libretexts.org
-
Refer to Table 3.3. Use Your Critical Thinking Skills: Why does Table 3.3 have a specific location for 60°N but no location for 60°S? Hint: look at a globe, an atlas, or Google Earth! Use Your Critical Thinking Skills: If a specific location had been provided for 60°S, would daylength at that location be similar to the daylength for 60°N? Explain why or why not in one to two sentences. Other than the poles, which of the eight locations has the greatest variation in daylength throughout the year? Explain why in one to two sentences. Tip: for each location, subtract the largest daylength number from the smallest to find the variation in daylength at that latitude. Other than the equator, which of the eight locations has the least variation in daylength throughout the year? Explain why in one to two sentences. Canberra and Tokyo are both located at a general latitude of 35°, however, their daylengths are different from each other on each of the four dates. Explain why in one to two sentences. On which date(s) does Palmas have a longer daylength than the equator? What are the seasons of these dates for Palmas? Does the equator have seasons? Why or why not?
a: the globe shows the South Pole plus ocean. No specific land exist in that area. b: no they have different daylength due to planet earth tilt. c: based on the numbers in the table of eight locations, greatest variation in daylength is white horse, Canada 60 degrees north due to the earth position to the sun. d: according to the table, Quito, Equador 10 degrees south has the east daylength variation. e: as I mentioned in the previous answers, the position of the earth to the sun makes the daylength to be different. f: February 16 and November 16 Palmas has longer daylength. Fall and Winter. g:No, beacause the equator always remain in the center.
-
-
nmoer.pressbooks.pub nmoer.pressbooks.pub
-
es, academic writing has rules about format, style, and objectivity that you must follow, but this does not mean you can write boring, impersonal prose. You can — and should — develop an authorial voice no matter what subject you choose to write about.
writers write articles come up with ideas to attract the audience and have new ideas for the people writing and annotating.
-
For example, you update your status on a social networking site with the awareness of who will digitally follow the post. If you want to brag about a good grade, you may write the post to please family members. If you want to describe a funny moment, you may write with your friends’ senses of humor in mind. Even at work, you send e-mails with an awareness of an unintended receiver who could intercept the message.
I have always cared to have really good grades, and be on time with all my homework's, sometimes I could fall behind but I will attempt to be on the right track.
-
Although the audience for writing assignments—your readers—may not appear in person, they play a vital role in the development of your writing. Even in everyday writing activities, you identify your readers’ characteristics, interests, and expectations before making decisions about what you write. In fact, thinking about audience has become so common that you may not even detect the audience-driven decisions. We will spend this chapter focusing on the role the audience plays in your writing by reviewing information presented in the textbook Successful Writing.
the writers are not physically here, but we still attempt to complete the assignment the best way I can.
-
-
nmoer.pressbooks.pub nmoer.pressbooks.pub
-
Because literacy narratives and memoirs often deal with events that happened early on in your life, you may be wondering, “But what if I don’t remember all the details?” That’s okay! Chances are that you won’t remember every word you spoke or what the weather was like, but it is important that you tell the emotional truth.
This passage says it’s normal not to remember every detail from early life events. What matters most in a literacy narrative is being honest about how the moment felt and why it mattered, not recalling exact words or perfect descriptions.
-
-
drive.google.com drive.google.com
-
The object is not really lifelike but byconvention and repetition has been made to appear so.
This quote emphasizes that what we accept as "realistic" is often the result of repeated social conventions rather than an accurate reflection of reality. This suggests that reality is learned and reinforced through cultural practices, media, and representation. It challenges the assumption that realism simply mirrors the world as it is.
-
Our common distinction betweenappearance and reality goes back, fundamentally, to the early use - ‘thereality underlying appearances
Williams highlights a long-standing tension between what things seem to be and what they actually are. This distinction is central, which often looks beyond surface-level behaviors to uncover deeper social meanings, beliefs, and power relations. The quote reinforces the idea that reality is not immediately visible, but must be interpreted through context, culture, and history.
-
-
modulovalue.com modulovalue.com
-
ZIP handles random access but originated from MS-DOS and has inconsistent Unix permission support. What we lack is something like "ZIP for Unix": random access with proper ownership, permissions, extended attributes, and ACLs.
Why/how do these make for a line-item criterion in a list of concerns for things necessary or useful to a compiler?
-
-
bookshelf.vitalsource.com bookshelf.vitalsource.com
-
Teacher knowledge and skills as well as teacher attitudes impact the school context. Emergent bilinguals have a greater chance for success in a school with adequate numbers of highly qualified teachers with background in second language acquisition, second language teaching, linguistics, and cross-cultural communication. The presence of counselors with training in working with second language students is also essential. School resources, including bilingual libraries and adequate technology access, are important for success as well. As is true in all schools with strong parent involvement, programs that include the parents of culturally and linguistically diverse students result in better outcomes for emergent bilingual students.
As a teacher you can't do everything that everybody wants you too and you can't get every certification and take every class and spend 28 hours in a 24 hour period dedicated to school things. It's just not possible, but it seems like this is almost expected of teachers. I keep hearing from my education that we need to keep doing more and more and more and we need to do this and this and this and then I go talk to teachers who are in the classroom teaching and one main message I keep getting from them is make sure you are taking personal time for you and meeting your own self care needs. The students will not benefit from you burning out. So it is also reasonable to not be certified in things like this, but rely partially on those who are and ask for help from them when you don't know where to go. All you can do is your best and sometime you don't always have 100% to give to your best, so just give whatever percentage of whatever you have and show you students you care for them and honestly at the end of the day if you student know you love and care for them then in my book you have won big time.
-
Other Trump immigration policies included the deportation of parents and sometimes families who have lived in this country for years without legal status. Many of these deportees have contributed in important ways to their communities.
If we are held to the law then others should be too. I don't understand what is wrong with this? I think its sad that people have to feel this fear, but in this example it literally says they have lived in the county for years without legal status, meaning they had years to become legal and go through the processes to do so. They are bringing this stress on themselves by not going through the right process. Now I'm not negating the fact that they have contributed to America in many ways, wonderful, I'm glad they have, but that has literally no connection to the fact that they didn't go through the correct processes to become legal, so why is the author of this book using it as some excuse. It sounds like to me this author is saying because they have contributed to the community they have the right to stay in it. And if that is what he is saying then I firmly disagree. We are talking about critical literacy in one of my other classes and I think this is an issue that demand a ton of it, because so many people are misinformed and aren't critically literate hence why they are misinformed.
-
-
www.gutenberg.org www.gutenberg.org
-
He can have no true regard for me, or he would not have listened to her; and she, with her little rebellious heart and indelicate feelings, to throw herself into the protection of a young man with whom she has scarcely ever exchanged two words before! I am equally confounded at her impudence and his credulity. How dared he believe what she told him in my disfavour! Ought he not to have felt assured that I must have unanswerable motives for all that I had done? Where was his reliance on my sense and goodness then? Where the resentment which true love would have dictated against the person defaming me—that person, too, a chit, a child, without talent or education, whom he had been always taught to despise?
Lady Susan is an absolutely intriguing character, albeit heavily dislikeable to me personally. That being said, she's spectacularly written; the characterization within this passage alone is outstanding! It goes without saying that Susan does not like being played with, and that is exactly what is being described here. She already has a large amount of unwarranted distaste for her own daughter, of which she convinced Reginald to believe, and is now infuriated by the mere thought of her daughter going behind her back and, in her head, twisting what she has convinced Reginald of. In short, she does not like being made out to be the bad guy. Though a very heated letter, I did sense some interesting contesting feelings towards her daughter, Frederica. Though Susan's dislike of her child is palpable, there seems to still be a sense of caring. After all, she is knowledgeable enough to realize Frederica is romantically pursuing Reginald, but Susan is angered by Reginald toying with this likely one-sided romantic interest from her daughter. It is an incredibly interesting family dynamic between mother and daughter, especially with Reginald involved.
-
-
new.express.adobe.com new.express.adobe.com
-
The first Queen seems still to have had prospects; not yet fallen into sexuality, she looks outward, if only upon the snow. The second Queen is doomed to inward search that psychoanalysts like Bruno Bettelheim censoriously define as "narcissism," but which is necessitated by a state from which all outward prospects have been removed.
This quote is incredibly interesting to me. I had never thought of the first Queen and the second Queen as such opposites, but I love how it used the window as an outward reflection, and the mirror as inward. I will say, I don't love how women in all of these stories, specifically older women, are seen as witch-like and evil, but I guess it was a different time...
-
-
-
Humans have such a long history of using storytelling to connect to one another that it seems like an instinctual motivation and desire. “Insofar as we account for our own actions and for the human events that occur around us principally in terms of narrative, story, drama, it is conceivable that our sensitivity to narrative provides the major link between our own sense of self and our sense of others in the social world around us” (Bruner, 1986, p. 69). Storytelling is part and parcel of human socialization—a tool for making us known, both to ourselves and to others. In fact, anything we experience that does not get structured narratively does not get remembered
This stood out to me because it shows that storytelling isn’t just a teaching strategy, but also how people naturally make sense of the world. If we remember stories better than facts, it makes sense to use storytelling more often in class.
-
-
gemini.google.com gemini.google.com
-
It merges real-time data from a custom WAL with historical data from S3 in a single pass
This is like super similar to what Estuary is doing, but i guess custom built for super huge AI traces
-
-
scholarlykitchen.sspnet.org scholarlykitchen.sspnet.org
-
Deeper disclosure is possible: version-controlled authorship history (git-style) showing what human wrote vs. what AI generated.
The commit log becomes the disclosure - forensic, auditable, transparent. Not a vague "AI-assisted" disclaimer, but a traceable record of human-machine co-authorship.
Example: every commit with "Co-Authored-By: Claude Opus 4.5" plus commit messages explaining what was asked, proposed, reviewed, and approved.
This reframes the "crisis" as an opportunity for unprecedented transparency in collaborative authorship.
-
-
-
But in others, includingIdaho and Colorado, judges use the scores to guide their sentencing
Wow, kinda surprised to know this
-
When we ask Google Mapsfor directions, it models the world as a series of roads, tunnels, andbridges. It ignores the buildings, because they aren’t relevant to the task.When avionics software guides an airplane, it models the wind, the speedof the plane, and the landing strip below, but not the streets, tunnels,buildings, and people.
Great analogy, wondering where it's going
-
-
www.earlymoderntexts.com www.earlymoderntexts.com
-
because those arguments imply that there really isa world, and that men have bodies and so on (no sane personhas ever seriously doubted these things), but •because inconsidering these arguments we come to realise that theyare not as solid or as transparent as the ones that lead usto knowledge of our minds and of God
There is more transparent reasons to believe in God whom we cannot see than bodies that are right in front of us
-
Forwe cannot conceive of half of a mind, while we can alwaysconceive of half of a body, however small; and this leads usto recognise that the natures of mind and body are not onlydifferent but in a way opposite
This stands out to me because I have never thought of it like this before. The inability to have half a mind intrigues me.
-
But since some people may expect arguments for theimmortality of the soul in this place, I think they should bewarned here that I have tried not to put down anything thatI could not precisely demonstrate
Only writes philosophical concepts and arguments that he can prove without question (when asked to clarify at times if needed)
-
So, providedwe remember that our minds must be regarded as finite,while God is infinite and beyond our comprehension, suchobjections will not cause us any difficulty
But how do we know for sure that God is infinite? Or is this something we must assume for the arguments to work?
-
your authority will induce the atheists—whousually have smatterings of knowledge rather than intelli-gence or learning
claims atheists are not 'well-educated' but and don't have the insight a philosopher does. Can nonbelievers be philosophers then?
-
Hence, whatever the quality of my arguments may be,because they have to do with philosophy I don’t expect theywill have much effect on people’s minds unless you grant meyour patronage
Reader must be open to ideas and not seek to argue / disprove but consider the statements and their legitimacy
-
Now, I completely disagree with this; I think that almost allthe arguments that great men have put forward on these twoquestions (when properly understood) are demonstrations,and that it would be almost impossible to discover anynew ones
Philosophers have proven these points to nonbelievers, but they have failed to 'properly understand' according to Descartes
-
And in the same place, in the words ’that whichmay be known of God is manifest in them’, we seem to betold that everything that can be known of God can be shownby reasons that have no other source but our own mind
Our own mind. Reminds me of the philosophical concept where I know I am a concise being but I can't know another person is because I don't know their mind and state of consciousness personally.
-
But this argumentcannot be put to unbelievers, because they would think it tobe (as the logicians say) circular
This argument has the assumption that the one hearing it already believes in God and the soul as a theologist would, however this may not be the case which is why philosophy is a more successful approach at explaining these concepts.
-
but in the case of unbelievers,it seems that there is no religion—and hardly any moralvirtue—that they can be persuaded to adopt until these twotruths are proved to them by natural reason
From what I understand of this: Philosophers do a better job proving to nonbelievers the existence of God and the soul than a theologist does. Is that because nonbelievers are more receptive to philosophers? Or is it their approach?
-
-
www.americanyawp.com www.americanyawp.com
-
The article Rose Davis is sentenced to a life of slavery" is a record about how Rose was sentenced to become a slave for the rest of her life. In the article, it seems like Rose white was seeking her freedom. So she went to court. It might have been easy, since she was born to a white woman and her father was a black man, but unfortunately, Rose Davis was sentenced to life of slavery instead of being set free.
-
-
sites.google.com sites.google.com
-
il miglior fabbro
"The better craftsman". Eliot and Pound were not only collaborators, but also friends. This passage refers to Dante Alighieri's tribute to the Provencal poet Arnauld Daniel in Canto 26 of the Purgatorio.
-
-
Local file Local file
-
non-odoriferous precursors and specific enzymes of Corynebacteria that transformthem into volatile substances:a specific Zn-dependent Na-acyl-glutamine aminoacylase(N-AGA) triggers the release of the acids 3M2 H and HMHA from Na-acyl-glutamineconjugates secreted in the axilla [13], and a b-lyase releases sulfanyl alcohols fromsynthetic cysteine conjugates and from axilla secretions
Human armpit sweat is initially odourless because it contains non-odoriferous precursors — molecules that do not smell on their own. Body odour only develops when skin bacteria, particularly Corynebacteria, break these precursors down.
These bacteria produce specific enzymes that act like biological scissors. The enzymes cut or transform the odorless precursor molecules into small, volatile substances that can evaporate into the air and be detected by the nose.
Overall, human body odour is not produced directly by the body, but results from an interaction between odorless sweat molecules and bacterial enzymes on the skin. People tend to have the same types of odour compounds, but in different proportions, which explains why individuals smell different.
-
-
visualizingcultures.mit.edu visualizingcultures.mit.edu
-
It involves an issue that has been with us since World War II and was abruptly thrown to the forefront of popular consciousness by 9-11: when, if ever, is it appropriate to identify civilians as a legitimate target of war?
This question pushes the discussion beyond history into ethics. It frames Hiroshima and Nagasaki not just as wartime events but as ongoing moral problems that remain relevant in modern conflicts involving weapons of mass destruction.
-
-
elite-ai-assisted-coding.dev elite-ai-assisted-coding.dev
-
OpenHands demonstrated strong capabilities, particularly for complex refactoring tasks. With better configuration and more explicit instructions about development workflows, it could likely match Copilot's reliability. The open-source nature also makes it attractive, since the entire system can be self-hosted and confugred for every team's or project's needs.
openhands useful but likely needs more explicit instructions than others
-
OpenHands: Capable but Requiring InterventionI connected my repository to OpenHands through the All Hands cloud platform. I pointed the agent at a specific issue, instructing it to follow the detailed requirements and create a pull request when complete. The conversational interface displayed the agent's reasoning as it worked through the problem, and the approach appeared logical.
Also used openhands for a test. says it needs intervention (not fully delegated iow)
-
When an agent doesn't deliver what you expected, the temptation is to engage in corrective dialogue — to guide the agent toward the right solution through feedback. While some agents support this interaction model, it's often more valuable to treat failures as specification bugs. Ask yourself: what information was missing that caused the agent to make incorrect decisions? What assumptions did I fail to make explicit?This approach builds your specification-writing skills rapidly. After a few iterations, you develop an intuition for what needs to be explicit, what edge cases to cover, and how to structure instructions for maximum clarity. The goal isn't perfection on the first try, but rather continuous improvement in your ability to delegate effectively.
don't iterate for corrections. Redo and iterate the instructions. This is a bit like prompt engineering the oracle, no? AI isn't the issue, it's your instructions. Up to a point, but in flux too.
-
A complete task specification goes beyond describing what needs to be done. It should encompass the entire development lifecycle for that specific task. Think of it as creating a mini project plan that an intelligent but literal agent can follow from start to finish.
A discrete task description to be treated like a project in the GTD sense (anything above 2 steps is a project). At what point is this overkill, as in templating this project description may well lead to having the solutions once you've done this.
-
The fundamental rule for working with asynchronous agents contradicts much of modern agile thinking: create complete and precise task definitions upfront. This isn't about returning to waterfall methodologies, but rather recognizing that when you delegate to an AI agent, you need to provide all the context and guidance that you would naturally provide through conversation and iteration with a human developer.
What I mentioned above: to delegate you need to be able to fully describe and provide context for a discrete task.
-
isn't just about saving time — it's about restructuring how software gets built.
not just time saving, but a restructuring. So, any description of how the structure changes (before / after style) further down?
-
While interactive AI keeps you tethered to the development process, requiring constant attention and decision-making, asynchronous agents transform you from a driver into a delegator.
async means no handholding, but delegation instead. That is enticing obviously, but assumes unattended execution can be trusted. Seems a big if.
-
-
www.eleanorkonik.com www.eleanorkonik.com
-
Further ReadingI’m not gonna pretend to be an expert here (any more than I’m an expert Obsidian plugin developer :p) but here are some resources that helped me figure out Claude CodeKent writes a lot about how he uses Obsidian with Claude Code.This is an incredible hub of resources for using Claude Code for project management, by someone who also uses Obsidian.This take on Claude Code for non-developers helped solidify my understanding of how it all works; it hallucinates less, for one thing.Eleanor Berger has fantastic tips for working with asynchronous coding agents and is incredibly level-headed about the LLM landscape.This article does a great job of breaking down all the nitty-gritty of how Claude Code works.Damian Player has a step-by-step guide on using Claude Code as a non-technical person that goes into more depth.Here’s a tutorial from a pro that breaks down best practices for using Claude Code, like the importance of planning and thinking things through, and exactly why a good CLAUDE.md file matters.
Links w further reading wrt Claude Code and Obsidian. Most of these are links to X. Ugh.
-
My favorite kind of problem is a solvable problem. I know a lot of people who just brute force or deal with their issues, but I try to notice pain points and deal with them. This isn’t just an AI thing, this is a life thing.
Interesting point, and fair enough. Start from the friction points. Like w open data [[Open data begint buiten 20200808162905]]
-
But these days I’m not generally trying to do things faster, I’m trying to do them with less attention. All these searches and tasks run in the background, which means they actually get done. When I had to actively sit there and click through things, half of it never happened because something else more important would come up, or I just didn’t feel like doing grunt work just then.
Speaks of how the purpose is not being faster but gtd with less attention on things you don't want to free up attention for. As long as you keep it away from your own key things I suppose. The periphery of what you pay attention to. The many little side projects on the someday/maybe list, the ones just out of reach. Enticing promise! This is the lure ofc.
-
Setting Claude Code Up in ObsidianI was genuinely surprised at how easy the terminal plugin was to install for Obsidian. In Obsidian, I went to community plugins, searched for “terminal,” and installed the Terminal plugin by polyipseity. Then I clicked the “open terminal” button on the left-hand side. That’s it.There’s a dedicated Claudian plugin (subtly different from the Claudsidian solution people), but the Terminal felt a little higher fidelity to how I’m used to doing things, and a little simpler to understand. Plus, Claudian looks great but honestly I don’t think I can live without plan mode, which the readme says it doesn’t currently support. Plan mode is nice because it asks questions, really thinks things through, and can be trusted not to do dumb destructive things.
There is a terminal plugin for Obsidian that you can connect to Claude Code (apparently). She advices against the Claudian plugin bc it lacks plan mode (i.e. not immediately act)
-
If you have been following along with me for years you know I don’t hype things just because people are hyping things. But Claude Code finally has made AI a core part of my processes instead of just a thing I use sometimes as an extra source or bonus spell checker or quicker way to reformat files.
She feels Claude Code is now a core tool in her workflows
-
-
bookshelf.vitalsource.com bookshelf.vitalsource.com
-
Both societies practiced what scholars call “gender parallelism,” in which “women and men operate in two separate but equivalent spheres, each gender enjoying autonomy in its own sphere.”
Is this taking into account the aforementioned “chosen women?” If not, is the idea of gender parallelism in Incan society based specifically on how both men and women could do the same types of labor and be in high ranking positions? Was there an equivalent for the “chosen women” for boys in Incan society? Furthermore, did the Aztec society have their own version of chosen women?
-
-
social-media-ethics-automation.github.io social-media-ethics-automation.github.io
-
There are several options for how to save dates and times. Some options include a series of numbers (year, month, day, hour, minute, and second), or a string that with all of those pieces of information written out. Sometimes only the date is saved, with no time information, and sometimes the time information will include the timezone. Dates turn out to be one of the trickier data types to work with in practice. One of the main reasons for this is that what time or day it depends on what time zone you are in. So, for example, when Twitter tells me that the tweet was posted on Feb 10, 2020, does it mean Feb 10 for me? Or for the person who posted it? Those might not be the same. Or if I want to see for a given account, how much they tweeted “yesterday,” what do I mean by “yesterday?” We might be in different time zones and have different start and end times for what we each call “yesterday.”
This discussion shows that dates and times are not neutral data, but depend on context such as time zones and perspective. When platforms define concepts like “today” or “yesterday,” they are making design choices that can shape how activity is measured and interpreted, which becomes especially important when analyzing user behavior or automated posting.
-
-
social-media-ethics-automation.github.io social-media-ethics-automation.github.io
-
In order to understand how a bot is built and can work, we will now look at the different ways computer programs can be organized. We will cover a bunch of examples quickly here, to hopefully give you an idea of many options for how to write a program. Don’t worry if you don’t follow all of it, as we will go back over these one at a time in more detail throughout the book. In this section, we will not show actual Python computer programs (that will be in the next section). Instead, here we will focus on what programmers call “psuedocode,” which is a human language outline of a program. Psuedocode is intended to be easier to read and write. Pseudocode is often used by programmers to plan how they want their programs to work, and once the programmer is somewhat confident in their pseudocode, they will then try to write it in actual programming language code.
This explanation of pseudocode is helpful because it lowers the barrier to understanding how bots are structured without requiring prior programming knowledge. Framing pseudocode as a planning and thinking tool emphasizes that building bots is not just a technical process, but also a conceptual one where ethical choices can be made early, before code is even written.
-
-
data-feminism.mitpress.mit.edu data-feminism.mitpress.mit.edu
-
Consider Christine Darden, who, after speaking up to her division chief, heard nothing from him but radio silence. But then, two weeks later, she was indeed promoted.d-undefined, .lh-undefined { background-color: rgba(0, 0, 0, 0.2) !important; }1Amanda Christopher and transferred to a group focused on sonic boom research
A thing to note is that women of color have been socialized to not rock the boat. From personal experience, in PWI I walk on egg shells, because I know that if I make too much ruckus with certain individuals, I might be blackballed from the entire organization.
-
-
wrd.as.uky.edu wrd.as.uky.edu
-
But because by then I had been writing for so long, I would eventually let myselftrust the process -- sort of, more or less. I
This was interesting to me. I feel like life has a pattern to it, like writing does. And if you can kinda figure out that pattern than it helps you roll with the punches easier.
-
-
www.trans-lex.org www.trans-lex.org
-
But they were not intended to legalise a pretended association for the purpose of 33 enabling an individual to carry on his own business with limited liability in the name of a joint stock company."
whys this case going against them ijbol
-
-
publish.obsidian.md publish.obsidian.md
-
Confucian morality is secular rather than religious, which left room for the Emperor to be a representative of Divinity and claim “the Mandate of Heaven” without the Chinese Empire becoming a theocracy.
I believe this text explains that Confucianism focused more on ethics and social order than on worship or religion. Because it wasn’t religious in nature, it allowed the Emperor to claim divine approval through the “Mandate of Heaven” without turning the government into a religious state. This helped keep political power and religion connected, but not fully merged.
-
The imperial courts sent thousands of highly-educated administrators throughout the empire and China was ruled not by hereditary nobles or even elected representatives, but by a class of men who had received rigorous training and had passed very stringent examinations to prove they were qualified to lead.
It's interesting to see how China established a system where training and talent mattered more than your birthright and lineage in order to rule. Seeing that in places like Europe, for example, your lineage is often what determined who would be next to lead the country. Despite both continents being so close, it is an interesting thing to see how much they differ in this aspect. There also comes the question that if this was also a tactic used to ensure conformity, especially politically and ideologically, due to everyone being trained in the same manner. This would ensure that across their empire, the same ideologies and beliefs could be enforced.
-
One who is outstanding in ruling the people should do his best to treat the people as his children.
From this text i believe a ruler should care for the people the same way a parent cares for their kids. It suggests that leadership isn't just about control or power but responsibility, protection, and genuinely wanting what's best for those being led.
-
“He who knows all the answers has not been asked all the questions.”
I think this quote discusses how there is no possible way of knowing everything and anything because just as quickly as we learn something, something is created. So maybe to truly be knowledgeable, you don't have to know everything, but act as if you want to by asking those questions.
-
“If you think in terms of a year, plant a seed; if in terms of ten years, plant trees; if in terms of 100 years, teach the people.”
I think there could be many ways of interpreting this quote. But for me at least, it's about how the fruits of our labor are directly related to the effort and energy put into the labor. It is much more difficult to teach people than it is to plant a seed, which is why they have different outcomes.
-
-
visualizingcultures.mit.edu visualizingcultures.mit.edu
-
Usually, however, the answer is clear. Mothers attempt to nurse dead babies, or carry a dead child on their back. Infants try to nurse at the breast of a dead or injured mother.
The wording used was heartbreaking, but it reflects the sad reality that many mothers and their children faced after the bomb. On one hand, mothers tried to nurse their children, unable to fully process the catastrophe that had occurred or the possibility that their baby might be gone. On the other hand, infants were too young to comprehend what was happening, relying only on instinct and the comfort of the person they recognized.
-