But woo her, gentle Paris, get her heart,
Capulet is somehow progressive. He wants Juliet to consent to marriage.
But woo her, gentle Paris, get her heart,
Capulet is somehow progressive. He wants Juliet to consent to marriage.
Capulet. But saying o'er what I have said before: My child is yet a stranger in the world; She hath not seen the change of fourteen years, Let two more summers wither in their pride, 280Ere we may think her ripe to be a bride.
Capulet is protective. He thinks Juliet is too young to marry at 14.
Yet tell me not, for I have heard it all. Here's much to do with hate, but more with love. Why, then, O brawling love! O loving hate! 200O any thing, of nothing first create! O heavy lightness! serious vanity! Mis-shapen chaos of well-seeming forms! Feather of lead, bright smoke, cold fire, sick health! 205Still-waking sleep, that is not what it is! This love feel I, that feel no love in this. Dost thou not laugh?
Oxymorons (opposites together). Shows how confused love makes him feel.
Benvolio. My noble uncle, do you know the cause? Montague. I neither know it nor can learn of him. Benvolio. Have you importuned him by any means? 165 Montague. Both by myself and many other friends: But he, his own affections' counsellor, Is to himself—I will not say how true— But to himself so secret and so close, So far from sounding and discovery,
Plot: Mystery about Romeo's sadness. His parents don't understand him.
Benvolio. I do but keep the peace: put up thy sword,
Benvolio is reasonable. He explains he's just trying to stop the fight.
Sampson. [Aside to GREGORY] Is the law of our side, if I say 60ay? Gregory. No. Sampson. No, sir, I do not bite my thumb at you, sir, but I bite my thumb, sir.
Character Trait: Sampson is a coward. He won't admit the insult when challenged.
Sampson. My naked weapon is out: quarrel, I will back thee.
Character Trait: Sampson is a coward. He draws his sword but wants Gregory to start the fight.
Sampson. I strike quickly, being moved. Gregory. But thou art not quickly moved to strike.
Character Trait: Gregory calls Sampson a coward. Sampson talks big but won't actually fight.
Act I, Scene 1 Verona. A public place. next scene [Enter SAMPSON and GREGORY, of the house of Capulet, armed with swords and bucklers] Sampson. Gregory, o' my word, we'll not carry coals. Gregory. No, for then we should be colliers. Sampson. I mean, an we be in choler, we'll draw. Gregory. Ay, while you live, draw your neck out o' the collar. 20 Sampson. I strike quickly, being moved. Gregory. But thou art not quickly moved to strike. Sampson. A dog of the house of Montague moves me. Gregory. To move is to stir; and to be valiant is to stand: therefore, if thou art moved, thou runn'st away. 25 Sampson. A dog of that house shall move me to stand: I will take the wall of any man or maid of Montague's. Gregory. That shows thee a weak slave; for the weakest goes to the wall. Sampson. True; and therefore women, being the weaker vessels, 30are ever thrust to the wall: therefore I will push Montague's men from the wall, and thrust his maids to the wall. Gregory. The quarrel is between our masters and us their men. Sampson. 'Tis all one, I will show myself a tyrant: when I 35have fought with the men, I will be cruel with the maids, and cut off their heads. Gregory. The heads of the maids? Sampson. Ay, the heads of the maids, or their maidenheads; take it in what sense thou wilt. 40 Gregory. They must take it in sense that feel it. Sampson. Me they shall feel while I am able to stand: and 'tis known I am a pretty piece of flesh. Gregory. 'Tis well thou art not fish; if thou hadst, thou hadst been poor John. Draw thy tool! here comes 45two of the house of the Montagues. Sampson. My naked weapon is out: quarrel, I will back thee. Gregory. How! turn thy back and run? Sampson. Fear me not. Gregory. No, marry; I fear thee! 50 Sampson. Let us take the law of our sides; let them begin. Gregory. I will frown as I pass by, and let them take it as they list. Sampson. Nay, as they dare. I will bite my thumb at them; which is a disgrace to them, if they bear it. 55 [Enter ABRAHAM and BALTHASAR] Abraham. Do you bite your thumb at us, sir? Sampson. I do bite my thumb, sir. Abraham. Do you bite your thumb at us, sir? Sampson. [Aside to GREGORY] Is the law of our side, if I say 60ay? Gregory. No. Sampson. No, sir, I do not bite my thumb at you, sir, but I bite my thumb, sir. Gregory. Do you quarrel, sir? 65 Abraham. Quarrel sir! no, sir. Sampson. If you do, sir, I am for you: I serve as good a man as you. Abraham. No better. Sampson. Well, sir. Gregory. Say 'better:' here comes one of my master's kinsmen. 70 Sampson. Yes, better, sir. Abraham. You lie. Sampson. Draw, if you be men. Gregory, remember thy swashing blow. [They fight] [Enter BENVOLIO] Benvolio. Part, fools! Put up your swords; you know not what you do. [Beats down their swords] [Enter TYBALT] Tybalt. What, art thou drawn among these heartless hinds? 80Turn thee, Benvolio, look upon thy death. Benvolio. I do but keep the peace: put up thy sword, Or manage it to part these men with me. Tybalt. What, drawn, and talk of peace! I hate the word, As I hate hell, all Montagues, and thee: 85Have at thee, coward! [They fight] [Enter, several of both houses, who join the fray; then enter Citizens, with clubs] First Citizen. Clubs, bills, and partisans! strike! beat them down! 90Down with the Capulets! down with the Montagues! [Enter CAPULET in his gown, and LADY CAPULET] Capulet. What noise is this? Give me my long sword, ho! Lady Capulet. A crutch, a crutch! why call you for a sword? Capulet. My sword, I say! Old Montague is come, 95And flourishes his blade in spite of me. [Enter MONTAGUE and LADY MONTAGUE] Montague. Thou villain Capulet,—Hold me not, let me go. Lady Montague. Thou shalt not stir a foot to seek a foe. [Enter PRINCE, with Attendants] Prince Escalus. Rebellious subjects, enemies to peace, Profaners of this neighbour-stained steel,— Will they not hear? What, ho! you men, you beasts, That quench the fire of your pernicious rage With purple fountains issuing from your veins, 105On pain of torture, from those bloody hands Throw your mistemper'd weapons to the ground, And hear the sentence of your moved prince. Three civil brawls, bred of an airy word, By thee, old Capulet, and Montague, 110Have thrice disturb'd the quiet of our streets, And made Verona's ancient citizens Cast by their grave beseeming ornaments, To wield old partisans, in hands as old, Canker'd with peace, to part your canker'd hate: 115If ever you disturb our streets again, Your lives shall pay the forfeit of the peace. For this time, all the rest depart away: You Capulet; shall go along with me: And, Montague, come you this afternoon, 120To know our further pleasure in this case, To old Free-town, our common judgment-place. Once more, on pain of death, all men depart. [Exeunt all but MONTAGUE, LADY MONTAGUE, and BENVOLIO] Montague. Who set this ancient quarrel new abroach? 125Speak, nephew, were you by when it began? Benvolio. Here were the servants of your adversary, And yours, close fighting ere I did approach: I drew to part them: in the instant came The fiery Tybalt, with his sword prepared, 130Which, as he breathed defiance to my ears, He swung about his head and cut the winds, Who nothing hurt withal hiss'd him in scorn: While we were interchanging thrusts and blows, Came more and more and fought on part and part, 135Till the prince came, who parted either part. Lady Montague. O, where is Romeo? saw you him to-day? Right glad I am he was not at this fray. Benvolio. Madam, an hour before the worshipp'd sun Peer'd forth the golden window of the east, 140A troubled mind drave me to walk abroad; Where, underneath the grove of sycamore That westward rooteth from the city's side, So early walking did I see your son: Towards him I made, but he was ware of me 145And stole into the covert of the wood: I, measuring his affections by my own, That most are busied when they're most alone, Pursued my humour not pursuing his, And gladly shunn'd who gladly fled from me. 150 Montague. Many a morning hath he there been seen, With tears augmenting the fresh morning dew. Adding to clouds more clouds with his deep sighs; But all so soon as the all-cheering sun Should in the furthest east begin to draw 155The shady curtains from Aurora's bed, Away from the light steals home my heavy son, And private in his chamber pens himself, Shuts up his windows, locks far daylight out And makes himself an artificial night: 160Black and portentous must this humour prove, Unless good counsel may the cause remove. Benvolio. My noble uncle, do you know the cause? Montague. I neither know it nor can learn of him. Benvolio. Have you importuned him by any means? 165 Montague. Both by myself and many other friends: But he, his own affections' counsellor, Is to himself—I will not say how true— But to himself so secret and so close, So far from sounding and discovery, 170As is the bud bit with an envious worm, Ere he can spread his sweet leaves to the air, Or dedicate his beauty to the sun. Could we but learn from whence his sorrows grow. We would as willingly give cure as know. 175 [Enter ROMEO] Benvolio. See, where he comes: so please you, step aside; I'll know his grievance, or be much denied. Montague. I would thou wert so happy by thy stay, To hear true shrift. Come, madam, let's away. 180 [Exeunt MONTAGUE and LADY MONTAGUE] Benvolio. Good-morrow, cousin. Romeo. Is the day so young? Benvolio. But new struck nine. Romeo. Ay me! sad hours seem long. 185Was that my father that went hence so fast? Benvolio. It was. What sadness lengthens Romeo's hours? Romeo. Not having that, which, having, makes them short. Benvolio. In love? Romeo. Out— 190 Benvolio. Of love? Romeo. Out of her favour, where I am in love. Benvolio. Alas, that love, so gentle in his view, Should be so tyrannous and rough in proof! Romeo. Alas, that love, whose view is muffled still, 195Should, without eyes, see pathways to his will! Where shall we dine? O me! What fray was here? Yet tell me not, for I have heard it all. Here's much to do with hate, but more with love. Why, then, O brawling love! O loving hate! 200O any thing, of nothing first create! O heavy lightness! serious vanity! Mis-shapen chaos of well-seeming forms! Feather of lead, bright smoke, cold fire, sick health! 205Still-waking sleep, that is not what it is! This love feel I, that feel no love in this. Dost thou not laugh? Benvolio. No, coz, I rather weep. Romeo. Good heart, at what? 210 Benvolio. At thy good heart's oppression. Romeo. Why, such is love's transgression. Griefs of mine own lie heavy in my breast, Which thou wilt propagate, to have it prest With more of thine: this love that thou hast shown 215Doth add more grief to too much of mine own. Love is a smoke raised with the fume of sighs; Being purged, a fire sparkling in lovers' eyes; Being vex'd a sea nourish'd with lovers' tears: What is it else? a madness most discreet, 220A choking gall and a preserving sweet. Farewell, my coz. Benvolio. Soft! I will go along; An if you leave me so, you do me wrong. Romeo. Tut, I have lost myself; I am not here; 225This is not Romeo, he's some other where. Benvolio. Tell me in sadness, who is that you love. Romeo. What, shall I groan and tell thee? Benvolio. Groan! why, no. But sadly tell me who. 230 Romeo. Bid a sick man in sadness make his will: Ah, word ill urged to one that is so ill! In sadness, cousin, I do love a woman. Benvolio. I aim'd so near, when I supposed you loved. Romeo. A right good mark-man! And she's fair I love. 235 Benvolio. A right fair mark, fair coz, is soonest hit. Romeo. Well, in that hit you miss: she'll not be hit With Cupid's arrow; she hath Dian's wit; And, in strong proof of chastity well arm'd, From love's weak childish bow she lives unharm'd. 240She will not stay the siege of loving terms, Nor bide the encounter of assailing eyes, Nor ope her lap to saint-seducing gold: O, she is rich in beauty, only poor, That when she dies with beauty dies her store. 245 Benvolio. Then she hath sworn that she will still live chaste? Romeo. She hath, and in that sparing makes huge waste, For beauty starved with her severity Cuts beauty off from all posterity. She is too fair, too wise, wisely too fair, 250To merit bliss by making me despair: She hath forsworn to love, and in that vow Do I live dead that live to tell it now. Benvolio. Be ruled by me, forget to think of her. Romeo. O, teach me how I should forget to think. 255 Benvolio. By giving liberty unto thine eyes; Examine other beauties. Romeo. 'Tis the way To call hers exquisite, in question more: These happy masks that kiss fair ladies' brows 260Being black put us in mind they hide the fair; He that is strucken blind cannot forget The precious treasure of his eyesight lost: Show me a mistress that is passing fair, What doth her beauty serve, but as a note 265Where I may read who pass'd that passing fair? Farewell: thou canst not teach me to forget. Benvolio. I'll pay that doctrine, or else die in debt. [Exeunt] previous scene Act I, Scene 2 A street. next scene [Enter CAPULET, PARIS, and Servant] Capulet. But Montague is bound as well as I, In penalty alike; and 'tis not hard, I think, For men so old as we to keep the peace. Paris. Of honourable reckoning are you both; And pity 'tis you lived at odds so long. 275But now, my lord, what say you to my suit? Capulet. But saying o'er what I have said before: My child is yet a stranger in the world; She hath not seen the change of fourteen years, Let two more summers wither in their pride, 280Ere we may think her ripe to be a bride. Paris. Younger than she are happy mothers made. Capulet. And too soon marr'd are those so early made. The earth hath swallow'd all my hopes but she, She is the hopeful lady of my earth: 285But woo her, gentle Paris, get her heart, My will to her consent is but a part; An she agree, within her scope of choice Lies my consent and fair according voice. This night I hold an old accustom'd feast, 290Whereto I have invited many a guest, Such as I love; and you, among the store, One more, most welcome, makes my number more. At my poor house look to behold this night Earth-treading stars that make dark heaven light: 295Such comfort as do lusty young men feel When well-apparell'd April on the heel Of limping winter treads, even such delight Among fresh female buds shall you this night Inherit at my house; hear all, all see, 300And like her most whose merit most shall be: Which on more view, of many mine being one May stand in number, though in reckoning none, Come, go with me. [To Servant, giving a paper] 305Go, sirrah, trudge about Through fair Verona; find those persons out Whose names are written there, and to them say, My house and welcome on their pleasure stay. [Exeunt CAPULET and PARIS] Servant. Find them out whose names are written here! It is written, that the shoemaker should meddle with his yard, and the tailor with his last, the fisher with his pencil, and the painter with his nets; but I am sent to find those persons whose names are here 315writ, and can never find what names the writing person hath here writ. I must to the learned.—In good time. [Enter BENVOLIO and ROMEO] Benvolio. Tut, man, one fire burns out another's burning, One pain is lessen'd by another's anguish; 320Turn giddy, and be holp by backward turning; One desperate grief cures with another's languish: Take thou some new infection to thy eye, And the rank poison of the old will die. Romeo. Your plaintain-leaf is excellent for that. 325 Benvolio. For what, I pray thee? Romeo. For your broken shin. Benvolio. Why, Romeo, art thou mad? Romeo. Not mad, but bound more than a mad-man is; Shut up in prison, kept without my food, 330Whipp'd and tormented and—God-den, good fellow. Servant. God gi' god-den. I pray, sir, can you read? Romeo. Ay, mine own fortune in my misery. Servant. Perhaps you have learned it without book: but, I pray, can you read any thing you see? 335 Romeo. Ay, if I know the letters and the language. Servant. Ye say honestly: rest you merry! Romeo. Stay, fellow; I can read. [Reads] 'Signior Martino and his wife and daughters; 340County Anselme and his beauteous sisters; the lady widow of Vitravio; Signior Placentio and his lovely nieces; Mercutio and his brother Valentine; mine uncle Capulet, his wife and daughters; my fair niece Rosaline; Livia; Signior Valentio and his cousin 345Tybalt, Lucio and the lively Helena.' A fair assembly: whither should they come? Servant. Up. Romeo. Whither? Servant. To supper; to our house. 350 Romeo. Whose house? Servant. My master's. Romeo. Indeed, I should have ask'd you that before. Servant. Now I'll tell you without asking: my master is the great rich Capulet; and if you be not of the house 355of Montagues, I pray, come and crush a cup of wine. Rest you merry! [Exit] Benvolio. At this same ancient feast of Capulet's Sups the fair Rosaline whom thou so lovest, 360With all the admired beauties of Verona: Go thither; and, with unattainted eye, Compare her face with some that I shall show, And I will make thee think thy swan a crow. Romeo. When the devout religion of mine eye 365Maintains such falsehood, then turn tears to fires; And these, who often drown'd could never die, Transparent heretics, be burnt for liars! One fairer than my love! the all-seeing sun Ne'er saw her match since first the world begun. 370 Benvolio. Tut, you saw her fair, none else being by, Herself poised with herself in either eye: But in that crystal scales let there be weigh'd Your lady's love against some other maid That I will show you shining at this feast, 375And she shall scant show well that now shows best. Romeo. I'll go along, no such sight to be shown, But to rejoice in splendor of mine own. [Exeunt] previous scene Act I, Scene 3 A room in Capulet’s house. next scene [Enter LADY CAPULET and Nurse] Lady Capulet. Nurse, where's my daughter? call her forth to me. Nurse. Now, by my maidenhead, at twelve year old, I bade her come. What, lamb! what, ladybird! God forbid! Where's this girl? What, Juliet! [Enter JULIET] Juliet. How now! who calls? Nurse. Your mother. Juliet. Madam, I am here. What is your will? Lady Capulet. This is the matter:—Nurse, give leave awhile, 390We must talk in secret:—nurse, come back again; I have remember'd me, thou's hear our counsel. Thou know'st my daughter's of a pretty age. Nurse. Faith, I can tell her age unto an hour. Lady Capulet. She's not fourteen. 395 Nurse. I'll lay fourteen of my teeth,— And yet, to my teeth be it spoken, I have but four— She is not fourteen. How long is it now To Lammas-tide? Lady Capulet. A fortnight and odd days. 400 Nurse. Even or odd, of all days in the year, Come Lammas-eve at night shall she be fourteen. Susan and she—God rest all Christian souls!— Were of an age: well, Susan is with God; She was too good for me: but, as I said, 405On Lammas-eve at night shall she be fourteen; That shall she, marry; I remember it well. 'Tis since the earthquake now eleven years; And she was wean'd,—I never shall forget it,— Of all the days of the year, upon that day: 410For I had then laid wormwood to my dug, Sitting in the sun under the dove-house wall; My lord and you were then at Mantua:— Nay, I do bear a brain:—but, as I said, When it did taste the wormwood on the nipple 415Of my dug and felt it bitter, pretty fool, To see it tetchy and fall out with the dug! Shake quoth the dove-house: 'twas no need, I trow, To bid me trudge: And since that time it is eleven years; 420For then she could stand alone; nay, by the rood, She could have run and waddled all about; For even the day before, she broke her brow: And then my husband—God be with his soul! A' was a merry man—took up the child: 425'Yea,' quoth he, 'dost thou fall upon thy face? Thou wilt fall backward when thou hast more wit; Wilt thou not, Jule?' and, by my holidame, The pretty wretch left crying and said 'Ay.' To see, now, how a jest shall come about! 430I warrant, an I should live a thousand years, I never should forget it: 'Wilt thou not, Jule?' quoth he; And, pretty fool, it stinted and said 'Ay.' Lady Capulet. Enough of this; I pray thee, hold thy peace. Nurse. Yes, madam: yet I cannot choose but laugh, 435To think it should leave crying and say 'Ay.' And yet, I warrant, it had upon its brow A bump as big as a young cockerel's stone; A parlous knock; and it cried bitterly: 'Yea,' quoth my husband,'fall'st upon thy face? 440Thou wilt fall backward when thou comest to age; Wilt thou not, Jule?' it stinted and said 'Ay.' Juliet. And stint thou too, I pray thee, nurse, say I. Nurse. Peace, I have done. God mark thee to his grace! Thou wast the prettiest babe that e'er I nursed: 445An I might live to see thee married once, I have my wish. Lady Capulet. Marry, that 'marry' is the very theme I came to talk of. Tell me, daughter Juliet, How stands your disposition to be married? 450 Juliet. It is an honour that I dream not of. Nurse. An honour! were not I thine only nurse, I would say thou hadst suck'd wisdom from thy teat. Lady Capulet. Well, think of marriage now; younger than you, Here in Verona, ladies of esteem, 455Are made already mothers: by my count, I was your mother much upon these years That you are now a maid. Thus then in brief: The valiant Paris seeks you for his love. Nurse. A man, young lady! lady, such a man 460As all the world—why, he's a man of wax. Lady Capulet. Verona's summer hath not such a flower. Nurse. Nay, he's a flower; in faith, a very flower. Lady Capulet. What say you? can you love the gentleman? This night you shall behold him at our feast; 465Read o'er the volume of young Paris' face, And find delight writ there with beauty's pen; Examine every married lineament, And see how one another lends content And what obscured in this fair volume lies 470Find written in the margent of his eyes. This precious book of love, this unbound lover, To beautify him, only lacks a cover: The fish lives in the sea, and 'tis much pride For fair without the fair within to hide: 475That book in many's eyes doth share the glory, That in gold clasps locks in the golden story; So shall you share all that he doth possess, By having him, making yourself no less. Nurse. No less! nay, bigger; women grow by men. 480 Lady Capulet. Speak briefly, can you like of Paris' love? Juliet. I'll look to like, if looking liking move: But no more deep will I endart mine eye Than your consent gives strength to make it fly. [Enter a Servant] Servant. Madam, the guests are come, supper served up, you called, my young lady asked for, the nurse cursed in the pantry, and every thing in extremity. I must hence to wait; I beseech you, follow straight. Lady Capulet. We follow thee. 490[Exit Servant] Juliet, the county stays. Nurse. Go, girl, seek happy nights to happy days. [Exeunt] previous scene Act I, Scene 4 A street. next scene [Enter ROMEO, MERCUTIO, BENVOLIO, with five or six [p]Maskers, Torch-bearers, and others] Romeo. What, shall this speech be spoke for our excuse? Or shall we on without a apology? Benvolio. The date is out of such prolixity: We'll have no Cupid hoodwink'd with a scarf, 500Bearing a Tartar's painted bow of lath, Scaring the ladies like a crow-keeper; Nor no without-book prologue, faintly spoke After the prompter, for our entrance: But let them measure us by what they will; 505We'll measure them a measure, and be gone. Romeo. Give me a torch: I am not for this ambling; Being but heavy, I will bear the light. Mercutio. Nay, gentle Romeo, we must have you dance. Romeo. Not I, believe me: you have dancing shoes 510With nimble soles: I have a soul of lead So stakes me to the ground I cannot move. Mercutio. You are a lover; borrow Cupid's wings, And soar with them above a common bound. Romeo. I am too sore enpierced with his shaft 515To soar with his light feathers, and so bound, I cannot bound a pitch above dull woe: Under love's heavy burden do I sink. Mercutio. And, to sink in it, should you burden love; Too great oppression for a tender thing. 520 Romeo. Is love a tender thing? it is too rough, Too rude, too boisterous, and it pricks like thorn. Mercutio. If love be rough with you, be rough with love; Prick love for pricking, and you beat love down. Give me a case to put my visage in: 525A visor for a visor! what care I What curious eye doth quote deformities? Here are the beetle brows shall blush for me. Benvolio. Come, knock and enter; and no sooner in, But every man betake him to his legs. 530 Romeo. A torch for me: let wantons light of heart Tickle the senseless rushes with their heels, For I am proverb'd with a grandsire phrase; I'll be a candle-holder, and look on. The game was ne'er so fair, and I am done. 535 Mercutio. Tut, dun's the mouse, the constable's own word: If thou art dun, we'll draw thee from the mire Of this sir-reverence love, wherein thou stick'st Up to the ears. Come, we burn daylight, ho! Romeo. Nay, that's not so. 540 Mercutio. I mean, sir, in delay We waste our lights in vain, like lamps by day. Take our good meaning, for our judgment sits Five times in that ere once in our five wits. Romeo. And we mean well in going to this mask; 545But 'tis no wit to go. Mercutio. Why, may one ask? Romeo. I dream'd a dream to-night. Mercutio. And so did I. Romeo. Well, what was yours? 550 Mercutio. That dreamers often lie. Romeo. In bed asleep, while they do dream things true. Mercutio. O, then, I see Queen Mab hath been with you. She is the fairies' midwife, and she comes In shape no bigger than an agate-stone 555On the fore-finger of an alderman, Drawn with a team of little atomies Athwart men's noses as they lie asleep; Her wagon-spokes made of long spiders' legs, The cover of the wings of grasshoppers, 560The traces of the smallest spider's web, The collars of the moonshine's watery beams, Her whip of cricket's bone, the lash of film, Her wagoner a small grey-coated gnat, Not so big as a round little worm 565Prick'd from the lazy finger of a maid; Her chariot is an empty hazel-nut Made by the joiner squirrel or old grub, Time out o' mind the fairies' coachmakers. And in this state she gallops night by night 570Through lovers' brains, and then they dream of love; O'er courtiers' knees, that dream on court'sies straight, O'er lawyers' fingers, who straight dream on fees, O'er ladies ' lips, who straight on kisses dream, Which oft the angry Mab with blisters plagues, 575Because their breaths with sweetmeats tainted are: Sometime she gallops o'er a courtier's nose, And then dreams he of smelling out a suit; And sometime comes she with a tithe-pig's tail Tickling a parson's nose as a' lies asleep, 580Then dreams, he of another benefice: Sometime she driveth o'er a soldier's neck, And then dreams he of cutting foreign throats, Of breaches, ambuscadoes, Spanish blades, Of healths five-fathom deep; and then anon 585Drums in his ear, at which he starts and wakes, And being thus frighted swears a prayer or two And sleeps again. This is that very Mab That plats the manes of horses in the night, And bakes the elflocks in foul sluttish hairs, 590Which once untangled, much misfortune bodes: This is the hag, when maids lie on their backs, That presses them and learns them first to bear, Making them women of good carriage: This is she— 595 Romeo. Peace, peace, Mercutio, peace! Thou talk'st of nothing. Mercutio. True, I talk of dreams, Which are the children of an idle brain, Begot of nothing but vain fantasy, 600Which is as thin of substance as the air And more inconstant than the wind, who wooes Even now the frozen bosom of the north, And, being anger'd, puffs away from thence, Turning his face to the dew-dropping south. 605 Benvolio. This wind, you talk of, blows us from ourselves; Supper is done, and we shall come too late. Romeo. I fear, too early: for my mind misgives Some consequence yet hanging in the stars Shall bitterly begin his fearful date 610With this night's revels and expire the term Of a despised life closed in my breast By some vile forfeit of untimely death. But He, that hath the steerage of my course, Direct my sail! On, lusty gentlemen. 615 Benvolio. Strike, drum. [Exeunt] previous scene Act I, Scene 5 A hall in Capulet’s house. [Musicians waiting. Enter Servingmen with napkins] First Servant. Where's Potpan, that he helps not to take away? He shift a trencher? he scrape a trencher! 620 Second Servant. When good manners shall lie all in one or two men's hands and they unwashed too, 'tis a foul thing. First Servant. Away with the joint-stools, remove the court-cupboard, look to the plate. Good thou, save me a piece of marchpane; and, as thou lovest me, let 625the porter let in Susan Grindstone and Nell. Antony, and Potpan! Second Servant. Ay, boy, ready. First Servant. You are looked for and called for, asked for and sought for, in the great chamber. 630 Second Servant. We cannot be here and there too. Cheerly, boys; be brisk awhile, and the longer liver take all. [Enter CAPULET, with JULIET and others of his house, meeting the Guests and Maskers] Capulet. Welcome, gentlemen! ladies that have their toes Unplagued with corns will have a bout with you. 635Ah ha, my mistresses! which of you all Will now deny to dance? she that makes dainty, She, I'll swear, hath corns; am I come near ye now? Welcome, gentlemen! I have seen the day That I have worn a visor and could tell 640A whispering tale in a fair lady's ear, Such as would please: 'tis gone, 'tis gone, 'tis gone: You are welcome, gentlemen! come, musicians, play. A hall, a hall! give room! and foot it, girls. [Music plays, and they dance] 645More light, you knaves; and turn the tables up, And quench the fire, the room is grown too hot. Ah, sirrah, this unlook'd-for sport comes well. Nay, sit, nay, sit, good cousin Capulet; For you and I are past our dancing days: 650How long is't now since last yourself and I Were in a mask? Second Capulet. By'r lady, thirty years. Capulet. What, man! 'tis not so much, 'tis not so much: 'Tis since the nuptials of Lucentio, 655Come pentecost as quickly as it will, Some five and twenty years; and then we mask'd. Second Capulet. 'Tis more, 'tis more, his son is elder, sir; His son is thirty. Capulet. Will you tell me that? 660His son was but a ward two years ago. Romeo. [To a Servingman] What lady is that, which doth enrich the hand Of yonder knight? Servant. I know not, sir. 665 Romeo. O, she doth teach the torches to burn bright! It seems she hangs upon the cheek of night Like a rich jewel in an Ethiope's ear; Beauty too rich for use, for earth too dear! So shows a snowy dove trooping with crows, 670As yonder lady o'er her fellows shows. The measure done, I'll watch her place of stand, And, touching hers, make blessed my rude hand. Did my heart love till now? forswear it, sight! For I ne'er saw true beauty till this night. 675 Tybalt. This, by his voice, should be a Montague. Fetch me my rapier, boy. What dares the slave Come hither, cover'd with an antic face, To fleer and scorn at our solemnity? Now, by the stock and honour of my kin, 680To strike him dead, I hold it not a sin. Capulet. Why, how now, kinsman! wherefore storm you so? Tybalt. Uncle, this is a Montague, our foe, A villain that is hither come in spite, To scorn at our solemnity this night. 685 Capulet. Young Romeo is it? Tybalt. 'Tis he, that villain Romeo. Capulet. Content thee, gentle coz, let him alone; He bears him like a portly gentleman; And, to say truth, Verona brags of him 690To be a virtuous and well-govern'd youth: I would not for the wealth of all the town Here in my house do him disparagement: Therefore be patient, take no note of him: It is my will, the which if thou respect, 695Show a fair presence and put off these frowns, And ill-beseeming semblance for a feast. Tybalt. It fits, when such a villain is a guest: I'll not endure him. Capulet. He shall be endured: 700What, goodman boy! I say, he shall: go to; Am I the master here, or you? go to. You'll not endure him! God shall mend my soul! You'll make a mutiny among my guests! You will set cock-a-hoop! you'll be the man! 705 Tybalt. Why, uncle, 'tis a shame. Capulet. Go to, go to; You are a saucy boy: is't so, indeed? This trick may chance to scathe you, I know what: You must contrary me! marry, 'tis time. 710Well said, my hearts! You are a princox; go: Be quiet, or—More light, more light! For shame! I'll make you quiet. What, cheerly, my hearts! Tybalt. Patience perforce with wilful choler meeting Makes my flesh tremble in their different greeting. 715I will withdraw: but this intrusion shall Now seeming sweet convert to bitter gall. [Exit] Romeo. [To JULIET] If I profane with my unworthiest hand This holy shrine, the gentle fine is this: 720My lips, two blushing pilgrims, ready stand To smooth that rough touch with a tender kiss. Juliet. Good pilgrim, you do wrong your hand too much, Which mannerly devotion shows in this; For saints have hands that pilgrims' hands do touch, 725And palm to palm is holy palmers' kiss. Romeo. Have not saints lips, and holy palmers too? Juliet. Ay, pilgrim, lips that they must use in prayer. Romeo. O, then, dear saint, let lips do what hands do; They pray, grant thou, lest faith turn to despair. 730 Juliet. Saints do not move, though grant for prayers' sake. Romeo. Then move not, while my prayer's effect I take. Thus from my lips, by yours, my sin is purged. Juliet. Then have my lips the sin that they have took. Romeo. Sin from thy lips? O trespass sweetly urged! 735Give me my sin again. Juliet. You kiss by the book. Nurse. Madam, your mother craves a word with you. Romeo. What is her mother? Nurse. Marry, bachelor, 740Her mother is the lady of the house, And a good lady, and a wise and virtuous I nursed her daughter, that you talk'd withal; I tell you, he that can lay hold of her Shall have the chinks. 745 Romeo. Is she a Capulet? O dear account! my life is my foe's debt. Benvolio. Away, begone; the sport is at the best. Romeo. Ay, so I fear; the more is my unrest. Capulet. Nay, gentlemen, prepare not to be gone; 750We have a trifling foolish banquet towards. Is it e'en so? why, then, I thank you all I thank you, honest gentlemen; good night. More torches here! Come on then, let's to bed. Ah, sirrah, by my fay, it waxes late: 755I'll to my rest. [Exeunt all but JULIET and Nurse] Juliet. Come hither, nurse. What is yond gentleman? Nurse. The son and heir of old Tiberio. Juliet. What's he that now is going out of door? 760 Nurse. Marry, that, I think, be young Petrucio. Juliet. What's he that follows there, that would not dance? Nurse. I know not. Juliet. Go ask his name: if he be married. My grave is like to be my wedding bed. 765 Nurse. His name is Romeo, and a Montague; The only son of your great enemy. Juliet. My only love sprung from my only hate! Too early seen unknown, and known too late! Prodigious birth of love it is to me, 770That I must love a loathed enemy. Nurse. What's this? what's this? Juliet. A rhyme I learn'd even now Of one I danced withal. [One calls within 'Juliet.'] Nurse. Anon, anon! Come, let's away; the strangers all are gone. [Exeunt]
I can see various characterizations, themes and stylistic devices, which I will discuss below
Will you tell me that? 660His son was but a ward two years ago.
Capulet's disbelief shows how time can feel distorted, a recurring theme in the play where emotions and events move faster than expected
Come, knock and enter; and no sooner in, But every man betake him to his legs
Benvolio encourages action and movement, acting as a practical voice among the group. This highlights his role as a mediator.
Both by myself and many other friends: But he, his own affections' counsellor, Is to himself—I will not say how true
Montague and others have noticed the change in Romeo. His sadness and emotion, is taking over his well-being.
Madam, an hour before the worshipp'd sun Peer'd forth the golden window of the east, 140A troubled mind drave me to walk abroad; Where, underneath the grove of sycamore That westward rooteth from the city's side, So early walking did I see your son: Towards him I made, but he was ware of me 145And stole into the covert of the wood: I, measuring his affections by my own, That most are busied when they're most alone, Pursued my humour not pursuing his, And gladly shunn'd who gladly fled from me.
Benvolio is explains where he seen Romeo. He is letting Romeo's parents know that Romeo ran from him into the wood after he knew Benvolio was going to approach him.
But Montague is bound as well as I, In penalty alike; and 'tis not hard, I think, For men so old as we to keep the peace.
Both Capulet and Montageu have the same responsibility to keep peace and they share the same penalty if its not kept. This shows that he believes its not too difficult to keep the peace and he is willing to cooperate.
Alas, that love, so gentle in his view, Should be so tyrannous and rough in proof!
Benvolio is pointing out a contrast between how love is supposed to feel gentle but can also feel rough.
Could we but learn from whence his sorrows grow. We would as willingly give cure as know.
Montague wants to know where Romeo's sadness comes from. He wants to help and comfort Romeo. This shows that Romeo's father is compassionate.
Ay, the heads of the maids, or their maidenheads; take it in what sense thou wilt.
Sampson doesn't seem to have issues with the men, but with the women. In this line, is Sampson only referring to kill the women or is this line a metaphor for something else?
No, sir, I do not bite my thumb at you, sir, but I bite my thumb, sir.
Sampson is using sarcasm to provoke a fight. This shows how the feud causes people to look for any excuse to start violence.
Tis all one, I will show myself a tyrant: when I 35have fought with the men, I will be cruel with the maids, and cut off their heads.
Sampson is saying that the feud is not only between the men but also innocent women. Sampson is portraying himself as a man that is brutal is careless.
Reviewer #1 (Public review):
Summary:
Here, the authors have addressed the recruitment and firing patterns of motor units (MUs) from the long and lateral heads of triceps in the mouse. They used their newly developed Myomatrix arrays to record from these muscles during treadmill locomotion at different speeds, and they used template-based spike sorting (Kilosort) to extract units. Between MUs from the two heads, the authors observe differences in their firing rates, recruitment probability, phase of activation within the locomotor cycle and interspike interval patterning. Examining different walking speeds, the authors find increases in both recruitment probability and firing rates as speed increases. The authors also observed differences in the relation between recruitment and the angle of elbow extension between motor units from each head. These differences indicate meaningful variation between motor units within and across motor pools, and may reflect the somewhat distinct joint actions of the two heads of triceps.
Strengths:
The extraction of MU spike timing for many individual units is an exciting new method that has great promise for exposing the fine detail in muscle activation and its control by the motor system. In particular, the methods developed by the authors for this purpose seem to be the only way to reliably resolve single MUs in the mouse, as the methods used previously in humans and in monkeys (e.g. Marshall et al. Nature Neuroscience, 2022) do not seem readily adaptable for use in rodents.
The paper provides a number of interesting observations. There are signs of interesting differences in MU activation profiles for individual muscles here, consistent with those shown by Marshall et al. It is also nice to see fine scale differences in the activation of different muscle heads, which could relate to their partially distinct functions. The mouse offers greater opportunities for understanding the control of these distinct functions, compared to the other organisms in which functional differences between heads have previously been described.
The Discussion is very thorough, providing a very nice recounting of a great deal of relevant previous results.
Weaknesses:
The findings are limited to one pair of muscle heads. While the findings are important in their own right, the lack of confirmation from analysis of other muscles acting at other joints leaves the generalization of these findings unclear.
While differences between muscle heads with somewhat distinct functions are interesting and relevant to joint control, differences between MUs for individual muscles, like those in Marshall et al., are more striking because they cannot be attributed potentially to differences in each head's function. The present manuscript does show some signs of differences for MUs within individual heads (e.g. Figure 2C), but the manuscript falls short of providing a statistical basis for the existence of distinct subpopulations.
Author response:
The following is the authors’ response to the original reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
Here, the authors have addressed the recruitment and firing patterns of motor units (MUs) from the long and lateral heads of the triceps in the mouse. They used their newly developed Myomatrix arrays to record from these muscles during treadmill locomotion at different speeds, and they used template-based spike sorting (Kilosort) to extract units. Between MUs from the two heads, the authors observed differences in their firing rates, recruitment probability, phase of activation within the locomotor cycle, and interspike interval patterning. Examining different walking speeds, the authors find increases in both recruitment probability and firing rates as speed increases. The authors also observed differences in the relation between recruitment and the angle of elbow extension between motor units from each head. These differences indicate meaningful variation between motor units within and across motor pools and may reflect the somewhat distinct joint actions of the two heads of triceps.
Strengths:
The extraction of MU spike timing for many individual units is an exciting new method that has great promise for exposing the fine detail in muscle activation and its control by the motor system. In particular, the methods developed by the authors for this purpose seem to be the only way to reliably resolve single MUs in the mouse, as the methods used previously in humans and in monkeys (e.g. Marshall et al. Nature Neuroscience, 2022) do not seem readily adaptable for use in rodents.
The paper provides a number of interesting observations. There are signs of interesting differences in MU activation profiles for individual muscles here, consistent with those shown by Marshall et al. It is also nice to see fine-scale differences in the activation of different muscle heads, which could relate to their partially distinct functions. The mouse offers greater opportunities for understanding the control of these distinct functions, compared to the other organisms in which functional differences between heads have previously been described.
The Discussion is very thorough, providing a very nice recounting of a great deal of relevant previous results.
We thank the Reviewer for these comments.
Weaknesses:
The findings are limited to one pair of muscle heads. While an important initial finding, the lack of confirmation from analysis of other muscles acting at other joints leaves the general relevance of these findings unclear.
The Reviewer raises a fair point. While outside the scope of this paper, future studies should certainly address a wider range of muscles to better characterize motor unit firing patterns across different sets of effectors with varying anatomical locations. Still, the importance of results from the triceps long and lateral heads should not be understated as this paper, to our knowledge, is the first to capture the difference in firing patterns of motor units across any set of muscles in the locomoting mouse.
While differences between muscle heads with somewhat distinct functions are interesting and relevant to joint control, differences between MUs for individual muscles, like those in Marshall et al., are more striking because they cannot be attributed potentially to differences in each head's function. The present manuscript does show some signs of differences for MUs within individual heads: in Figure 2C, we see what looks like two clusters of motor units within the long head in terms of their recruitment probability. However, a statistical basis for the existence of two distinct subpopulations is not provided, and no subsequent analysis is done to explore the potential for differences among MUs for individual heads.
We agree with the Reviewer and have revised the manuscript to better examine potential subpopulations of units within each muscle as presented in Figure 2C. We performed Hartigan’s dip test on motor units within each muscle to test for multimodal distributions. For both muscles, p > 0.05, so we cannot reject the null hypothesis that the units in each muscle come from a multimodal distribution. However, Hartigan’s test and similar statistical methods have poor statistical power for the small sample sizes (n=17 and 16 for long and lateral heads, respectively) considered here, so the failure to achieve statistical significance might reflect either the absence of a true difference or a lack of statistical resolution.
Still, the limited sample size warrants further data collection and analysis since the varying properties across motor units may lead to different activation patterns. Given these results, we have edited the text as follows:
“A subset of units, primarily in the long head, were recruited in under 50% of the total strides and with lower spike counts (Figure 2C). This distribution of recruitment probabilities might reflect a functionally different subpopulation of units. However, the distribution of recruitment probabilities were not found to be significantly multimodal (p>0.05 in both cases, Hartigan’s dip test; Hartigan, 1985). However, Hartigan’s test and similar statistical methods have poor statistical power for the small sample sizes (n=17 and 16 for long and lateral heads, respectively) considered here, so the failure to achieve statistical significance might reflect either the absence of a true difference or a lack of statistical resolution.”
The statistical foundation for some claims is lacking. In addition, the description of key statistical analysis in the Methods is too brief and very hard to understand. This leaves several claims hard to validate.
We thank the Reviewer for these comments and have clarified the text related to key statistical analyses throughout the manuscript, as described in our other responses below.
Reviewer #2 (Public review):
The present study, led by Thomas and collaborators, aims to describe the firing activity of individual motor units in mice during locomotion. To achieve this, they implanted small arrays of eight electrodes in two heads of the triceps and performed spike sorting using a custom implementation of Kilosort. Simultaneously, they tracked the positions of the shoulder, elbow, and wrist using a single camera and a markerless motion capture algorithm (DeepLabCut). Repeated one-minute recordings were conducted in six mice at five different speeds, ranging from 10 to 27.5 cm·s<sup>-1</sup>.
From these data, the authors reported that:
(1) a significant portion of the identified motor units was not consistently recruited across strides,
(2) motor units identified from the lateral head of the triceps tended to be recruited later than those from the long head,
(3) the number of spikes per stride and peak firing rates were correlated in both muscles, and
(4) the probability of motor unit recruitment and firing rates increased with walking speed.
The authors conclude that these differences can be attributed to the distinct functions of the muscles and the constraints of the task (i.e., speed).
Strengths:
The combination of novel electrode arrays to record intramuscular electromyographic signals from a larger muscle volume with an advanced spike sorting pipeline capable of identifying populations of motor units.
We thank the Reviewer for this comment.
Weaknesses:
(1) There is a lack of information on the number of identified motor units per muscle and per animal.
The Reviewer is correct that this information was not explicitly provided in the prior submission. We have therefore added Table 1 that quantifies the number of motor units per muscle and per animal.
(2) All identified motor units are pooled in the analyses, whereas per-animal analyses would have been valuable, as motor units within an individual likely receive common synaptic inputs. Such analyses would fully leverage the potential of identifying populations of motor units.
Please see our answer to the following point, where we address questions (2) and (3) together.
(3) The current data do not allow for determining which motor units were sampled from each pool. It remains unclear whether the sample is biased toward high-threshold motor units or representative of the full pool.
We thank the Reviewer for these comments. To clarify how motor unit responses were distributed across animals and muscle targets, we updated or added the following figures:
Figure 2C
Figure 4–figure supplement 1
Figure 5–figure supplement 2
Figure 6–figure supplement 2
These provide a more complete look at the range of activity within each motor pool, suggesting that we do measure from units with different activation thresholds within the same motor pool, rather than this variation being due to cross-animal differences. For example, Figure 2C illustrates that motor units from the same muscle and animal show a wide variety of recruitment probabilities. However, the limited number of motor units recorded from each individual animal does not allow a statistically rigorous test for examining cross-animal differences.
(4) The behavioural analysis of the animals relies solely on kinematics (2D estimates of elbow angle and stride timing). Without ground reaction forces or shoulder angle data, drawing functional conclusions from the results is challenging.
The Reviewer is correct that we did not measure muscular force generation or ground reaction forces in the present study. Although outside the scope of this study, future work might employ buckle force transducers as used in larger animals (Biewener et al., 1988; Karabulut et al., 2020) to examine the complex interplay between neural commands, passive biomechanics, and the complex force-generating properties of muscle tissue.
Major comments:
(1) Spike sorting
The conclusions of the study rely on the accuracy and robustness of the spike sorting algorithm during a highly dynamic task. Although the pipeline was presented in a previous publication (Chung et al., 2023, eLife), a proper validation of the algorithm for identifying motor unit spikes is still lacking. This is particularly important in the present study, as the experimental conditions involve significant dynamic changes. Under such conditions, muscle geometry is altered due to variations in both fibre pennation angles and lengths.
This issue differs from electrode drift, and it is unclear whether the original implementation of Kilosort includes functions to address it. Could the authors provide more details on the various steps of their pipeline, the strategies they employed to ensure consistent tracking of motor unit action potentials despite potential changes in action potential waveforms, and the methods used for manual inspection of the spike sorting algorithm's output?
This is an excellent point and we agree that the dynamic behavior used in this investigation creates potential new challenges for spike sorting. In our analysis, Kilosort 2.5 provides key advantages in comparing unit waveforms across multiple channels and in detecting overlapping spikes. We modified this version of Kilosort to construct unit waveform templates using only the channels within the same muscle (Chung et al., 2023), as clarified in the revised Methods section (see “Electromyography (EMG)”):
“A total of 33 units were identified across all animals. Each unit’s isolation was verified by confirming that no more than 2% of inter-spike intervals violated a 1 ms refractory limit. Additionally, we manually reviewed cross-correlograms to ensure that each waveform was only reported as a single motor unit.”
The Reviewer is correct that our ability to precisely measure a unit’s activity based on its waveform will depend on the relationship between the embedded electrode and the muscle geometry, which alters over the course of the stride. As a follow-up to the original text, we have included new analyses to characterize the waveform activity throughout the experiment and stride (also in Methods):
“We further validated spike sorting by quantifying the stability of each unit’s waveform across time (Figure 1–figure supplement 1). First, we calculated the median waveform of each unit across every trial to capture long-term stability of motor unit waveforms. Additionally, we calculated the median waveform through the stride binned in 50 ms increments using spiking from a single trial. This second metric captures the stability of our spike sorting during the rapid changes in joint angles that occur during the burst of an individual motor unit. In doing so, we calculated each motor unit’s waveforms from the single channel in which that unit’s amplitude was largest and did not attempt to remove overlapping spikes from other units before measuring the median waveform from the data. We then calculated the correlation between a unit’s waveform over either trials or bins in which at least 30 spikes were present. The high correlation of a unit waveform over time, despite potential changes in the electrodes’ position relative to muscle geometry over the dynamic task, provides additional confidence in both the stability of our EMG recordings and the accuracy of our spike sorting.”
(2) Yield of the spike sorting pipeline and analyses per animal/muscle
A total of 33 motor units were identified from two heads of the triceps in six mice (17 from the long head and 16 from the lateral head). However, precise information on the yield per muscle per animal is not provided. This information is crucial to support the novelty of the study, as the authors claim in the introduction that their electrode arrays enable the identification of populations of motor units. Beyond reporting the number of identified motor units, another way to demonstrate the effectiveness of the spike sorting algorithm would be to compare the recorded EMG signals with the residual signal obtained after subtracting the action potentials of the identified motor units, using a signal-to-residual ratio.
Furthermore, motor units identified from the same muscle and the same animal are likely not independent due to common synaptic inputs. This dependence should be accounted for in the statistical analyses when comparing changes in motor unit properties across speeds and between muscles.
We thank the Reviewer for this comment. Regarding motor unit yield, as described above the newly-added Table 1 displays the yield from each animal and muscle.
Regarding spike sorting, while signal-to-residual is often an excellent metric, it is not ideal for our high-resolution EMG signals since isolated single motor units are typically superimposed on a “bulk” background consisting of the low-amplitude waveforms of other motor units. Because these smaller units typically cannot be sorted, it is challenging to estimate the “true” residual after subtracting (only) the largest motor unit, since subtracting each sorted unit’s waveform typically has a very small effect on the RMS of the total EMG signal. To further address concerns regarding spike sorting quality, we added Figure 1–figure supplement 1 that demonstrates motor units’ consistency over the experiment, highlighting that the waveform maintains its shape within each stride despite muscle/limb dynamics and other possible sources of electrical noise or artifact.
Finally, the Reviewer is correct that individual motor units in the same muscle are very likely to receive common synaptic inputs. These common inputs may reflect in sparse motor units being recruited in overlapping rather than different strides. Indeed, in the following added to the Results, we identified that motor units are recruited with higher probability when additional units are recruited.
“Probabilistic recruitment is correlated across motor units
Our results show that the recruitment of individual motor units is probabilistic even within a single speed quartile (Figure 5A-C) and predicts body movements (Figure 6), raising the question of whether the recruitment of individual motor units are correlated or independent. Correlated recruitment might reflect shared input onto the population of motor units innervating the muscle (De Luca, 1985; De Luca & Erim, 1994; Farina et al., 2014). For example, two motor units, each with low recruitment probabilities, may still fire during the same set of strides. To assess the independence of motor unit recruitment across the recorded population, we compared each unit’s empirical recruitment probability across all strides to its conditional recruitment probability during strides in which another motor unit from the same muscle was recruited (Figure 7). Doing this for all motor unit pairs revealed that motor units in both muscles were biased towards greater recruitment when additional units were active (p<0.001, Wilcoxon signed-rank tests for both the lateral and long heads of triceps). This finding suggests that probabilistic recruitment reflects common synaptic inputs that covary together across locomotor strides.”
(3) Representativeness of the sample of identified motor units
However, to draw such conclusions, the authors should exclusively compare motor units from the same pool and systematically track violations of the recruitment order. Alternatively, they could demonstrate that the motor units that are intermittently active across strides correspond to the smallest motor units, based on the assumption that these units should always be recruited due to their low activation thresholds.
One way to estimate the size of motor units identified within the same muscle would be to compare the amplitude of their action potentials, assuming that all motor units are relatively close to the electrodes (given the selectivity of the recordings) and that motoneurons innervating more muscle fibres generate larger motor unit action potentials.
We thank the Reviewer for this comment. Below, we provide more detailed analyses of the relationships between motor unit spike amplitude and the recruitment probability as well as latency (relative to stride onset) of activation.
We generated the below figures to illustrate the relationship between the amplitude of motor units and their firing properties. As suspected, units with larger-amplitude waveforms fired with lower probability and produced their first spikes later in the stride. If we were comfortable assuming that larger spike amplitudes mean higher-force units, then this would be consistent with a key prediction of the size principle (i.e. that higher-force units are recruited later). However, we are hesitant to base any conclusions on this assumption or emphasize this point with a main-text figure, since EMG signal amplitude may also vary due to the physical properties of the electrode and distance from muscle fibers. Thus it is possible that a large motor unit may have a smaller waveform amplitude relative to the rest of the motor pool.
Author response image 1.
Relation between motor unit amplitude and (A) recruitment probability and (B) mean first spike time within the stride. Colored lines indicate the outcome of linear regression analyses.
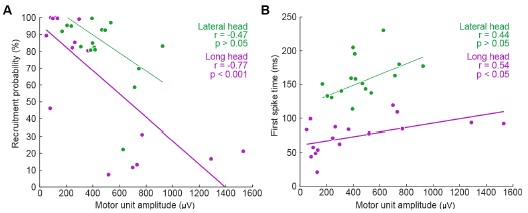
Currently, the data seem to support the idea that motor units that are alternately recruited across strides have recruitment thresholds close to the level of activation or force produced during slow walking. The fact that recruitment probability monotonically increases with speed suggests that the force required to propel the mouse forward exceeds the recruitment threshold of these "large" motor units. This pattern would primarily reflect spatial recruitment following the size principle rather than flexible motor unit control.
We thank the Reviewer for this comment. We agree with this interpretation, particularly in relation to the references suggested in later comments, and have added the following text to the Discussion to better reflect this argument:
“To investigate the neuromuscular control of locomotor speed, we quantified speed-dependent changes in both motor unit recruitment and firing rate. We found that the majority of units were recruited more often and with larger firing rates at faster speeds (Figure 5, Figure5–figure supplement 1). This result may reflect speed-dependent differences in the common input received by populations of motor neurons with varying spiking thresholds (Henneman et al., 1965). In the case of mouse locomotion, faster speeds might reflect a larger common input, increasing the recruitment probability as more neurons, particularly those that are larger and generate more force, exceed threshold for action potentials (Farina et al., 2014).”
(4) Analysis of recruitment and firing rates
The authors currently report active duration and peak firing rates based on spike trains convolved with a Gaussian kernel. Why not report the peak of the instantaneous firing rates estimated from the inverse of the inter-spike interval? This approach appears to be more aligned with previous studies conducted to describe motor unit behaviour during fast movements (e.g., Desmedt & Godaux, 1977, J Physiol; Van Cutsem et al., 1998, J Physiol; Del Vecchio et al., 2019, J Physiol).
We thank the Reviewer for this comment. In the revised Discussion (see ‘Firing rates in mouse locomotion compared to other species’) we reference several examples of previous studies that quantified spike patterns based on the instantaneous firing rate. We chose to report the peak of the smoothed firing rate because that quantification includes strides with zero spikes or only one spike, which occur regularly in our dataset (and for which ISI rate measures, which require two spikes to define an instantaneous firing rate, cannot be computed). Regardless, in the revised Figure 4B, we present an analysis that uses inter-spike intervals as suggested, which yielded similar ranges of firing rates as the primary analysis.
(5) Additional analyses of behaviour
The authors currently analyse motor unit recruitment in relation to elbow angle. It would be valuable to include a similar analysis using the angular velocity observed during each stride, re broadly, comparing stride-by-stride changes in firing rates with changes in elbow angular velocity would further strengthen the final analyses presented in the results section.
We thank the Reviewer for this comment. To address this, we have modified Figure 6 and the associated Supplemental Figures, to show relationships in unit activation with both the range of elbow extension and the range of elbow velocity for each stride. These new Supplemental Figures show that the trends shown in main text Figure 6C and 6E (which show data from all speed quartiles on the same axes) are also apparent in both the slower and faster quartiles individually, although single-quartile statistical tests (with smaller sample size than the main analysis) not reach statistical significance in all cases.
Reviewer #3 (Public review):
Summary:
Using the approach of Myomatrix recording, the authors report that:
(1) Motor units are recruited differently in the two types of muscles.
(2) Individual units are probabilistically recruited during the locomotion strides, whereas the population bulk EMG has a more reliable representation of the muscle.
(3) The recruitment of units was proportional to walking speed.
Strengths:
The new technique provides a unique data set, and the data analysis is convincing and well-performed.
We thank the Reviewer for the comment.
Weaknesses:
The implications of "probabilistical recruitment" should be explored, addressed, and analyzed further.
Comments:
One of the study's main findings (perhaps the main finding) is that the motor units are "probabilistically" recruited. The authors do not define what they mean by probabilistically recruited, nor do they present an alternative scenario to such recruitment or discuss why this would be interesting or surprising. However, on page 4, they do indicate that the recruitment of units from both muscles was only active in a subset of strides, i.e., they are not reliably active in every step.
If probabilistic means irregular spiking, this is not new. Variability in spiking has been seen numerous times, for instance in human biceps brachii motor units during isometric contractions (Pascoe, Enoka, Exp physiology 2014) and elsewhere. Perhaps the distinction the authors are seeking is between fluctuation-driven and mean-driven spiking of motor units as previously identified in spinal motor networks (see Petersen and Berg, eLife 2016, and Berg, Frontiers 2017). Here, it was shown that a prominent regime of irregular spiking is present during rhythmic motor activity, which also manifests as a positive skewness in the spike count distribution (i.e., log-normal).
We thank the Reviewer for this comment and have clarified several passages in response. The Reviewer is of course correct that irregular motor unit spiking has been described previously and may reflect motor neurons’ operating in a high-sensitivity (fluctuation-driven) regime. We now cite these papers in the Discussion (see ‘Firing rates in mouse locomotion compared to other species’). Additionally, the revision clarifies that “probabilistically” - as defined in our paper - refers only to the empirical observation that a motor unit spikes during only a subset of strides, either when all locomotor speeds are considered together (Figure 2) or separately (Figure 5A-C):
“Motor units in both muscles exhibited this pattern of probabilistic recruitment (defined as a unit’s firing on only a fraction of strides), but with differing distributions of firing properties across the long and lateral heads (Figure 2).”
“Our findings (Figure 4) highlight that even with the relatively high firing rates observed in mice, there are still significant changes in firing rate and recruitment probability across the spikes within bursts (Figure 4B) and across locomotor speeds (Figure 5F). Future studies should more carefully examine how these rapidly changing spiking patterns derive from both the statistics of synaptic inputs and intrinsic properties of motor neurons (Manuel & Heckman, 2011; Petersen & Berg, 2016; Berg, 2017).”
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
As mentioned above, there are several issues with the statistics that need to be corrected to properly support the claims made in the paper.
The authors compare the fractions of MUs that show significant variation across locomotor speeds in their firing rate and recruitment probability. However, it is not statistically founded to compare the results of separate statistical tests based on different kinds of measurements and thus have unconstrained differences in statistical power. The comparison of the fractional changes in firing rates and recruitment across speeds that follow is helpful, though in truth, by contemporary standards, one would like to see error bars on these estimates. These could be generated using bootstrapping.
The Reviewer is correct, and we have revised the manuscript to better clarify which quantities should or should not be compared, including the following passage (see “Motor unit mechanisms of speed control” in Results):
“Speed-dependent increases in peak firing rate were therefore also present in our dataset, although in a smaller fraction of motor units (22/33) than changes in recruitment probability (31/33). Furthermore, the mean (± SE) magnitude of speed-dependent increases was smaller for spike rates (mean rate<sub>fast</sub>/rate<sub>slow</sub> of 111% ± 20% across all motor units) than for recruitment probabilities (mean p(recruitment) <sub>fast</sub>/p(recruitment) <sub>slow</sub> of 179% ± 3% across all motor units). While fractional changes in rate and recruitment probability are not readily comparable given their different upper limits, these findings could suggest that while both recruitment and peak rate change across speed quartiles, increased recruitment probability may play a larger role in driving changes in locomotor speed.”
The description in the Methods of the tests for variation in firing rates and recruitment probability across speeds are extremely hard to understand - after reading many times, it is still not clear what was done, or why the method used was chosen. In the main text, the authors quote p-values and then state "bootstrap confidence intervals," which is not a statistical test that yields a p-value. While there are mathematical relationships between confidence intervals and statistical tests such that a one-to-one correspondence between them can exist, the descriptions provided fall short of specifying how they are related in the present instance. For this reason, and those described in what follows, it is not clear what the p-values represent.
Next, the authors refer to fitting a model ("a Poisson distribution") to the data to estimate firing rate and recruitment probability, that the model results agree with their actual data, and that they then bootstrapped from the model estimates to get confidence intervals and compute p-values. Why do this? Why not just do something much simpler, like use the actual spike counts, and resample from those? I understand that it is hard to distinguish between no recruitment and just no spikes given some low Poisson firing rate, but how does that challenge the ability to test if the firing rates or the number of spiking MUs changes significantly across speeds? I can come up with some reasons why I think the authors might have decided to do this, but reasoning like this really should be made explicit.
In addition, the authors would provide an unambiguous description of the model, perhaps using an equation and a description of how it was fit. For the bootstrapping, a clear description of how the resampling was done should be included. The focus on peak firing rate instead of mean (or median) firing rate should also be justified. Since peaks are noisier, I would expect the statistical power to be lower compared to using the mean or median.
We thank the Reviewer for the comments and have revised and expanded our discussion of the statistical tests employed. We expanded and clarified our description of these techniques in the updated Methods section:
“Joint model of rate and recruitment
We modeled the recruitment probability and firing rate based on empirical data to best characterize firing statistics within the stride. Particularly, this allowed for multiple solutions to explain why a motor unit would not spike within a stride. From the empirical data alone, strides with zero spikes would have been assumed to have no recruitment of a unit. However, to create a model of motor unit activity that includes both recruitment and rate, it must be possible that a recruited unit can have a firing rate of zero. To quantify the firing statistics that best represent all spiking and non-spiking patterns, we modeled recruitment probability and peak firing rate along the following piecewise function:
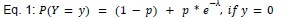
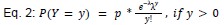
where y denotes the observed peak firing rate on a given stride (determined by convolving motor unit spike times with a Gaussian kernel as described above), p denotes the probability of recruitment, and λ denotes the expected peak firing rate from a Poisson distribution of outcomes. Thus, an inactive unit on a given stride may be the result of either non-recruitment or recruitment with a stochastically zero firing rate. The above equations were fit by minimizing the negative log-likelihood of the parameters given the data.
“Permutation test for joint model of rate and recruitment and type 2 regression slopes
To quantify differences in firing patterns across walking speeds, we subdivided each mouse’s total set of strides into speed quartiles and calculated rate (𝜆, Eq. 1 and 2, Fig. 5A-C) and recruitment probability terms (p, Eq. 1 and 2, Fig. 5D-F) for each unit in each speed quartile. Here we calculated the difference in both the rate and recruitment terms across the fastest and slowest speed quartiles (p<sub>fast</sub>-p<sub>slow</sub> and 𝜆<sub>fast</sub>-𝜆<sub>slow</sub>). To test whether these model parameters were significantly different depending on locomotor speed, we developed a null model combining strides from both the fastest and slowest speed quartiles. After pooling strides from both quartiles, we randomly distributed the pooled set of strides into two groups with sample sizes equal to the original slow and fast quartiles. We then calculated the null model parameters for each new group and found the difference between like terms. To estimate the distribution of possible differences, we bootstrapped this result using 1000 random redistributions of the pooled set of strides. Following the permutation test, the 95% confidence interval of this final distribution reflects the null hypothesis of no difference between groups. Thus, the null hypothesis can be rejected if the true difference in rate or recruitment terms exceeds this confidence interval.
We followed a similar procedure to quantify cross-muscle differences in the relationship between firing parameters. For each muscle, we estimated the slope across firing parameters for each motor unit using type 2 regression. In this case, the true difference was the difference in slopes between muscles. To test the null hypothesis that there was no difference in slopes, the null model reflected the pooled set of units from both muscles. Again, slopes were calculated for 1000 random resamplings of this pooled data to estimate the 95% confidence interval.”
The argument for delayed activation of the lateral head is interesting, but I am not comfortable saying the nervous system creates a delay just based on observations of the mean time of the first spike, given the potential for differential variability in spike timing across muscles and MUs. One way to make a strong case for a delay would be to show aggregate PSTHs for all the spikes from all the MUs for each of the two heads. That would distinguish between a true delay and more gradual or variable activation between the heads.
This is a good point and we agree that the claim made about the nervous system is too strong given the results. Even with Author response image 2 below that the Reviewer suggested, there is still not enough evidence to isolate the role of the nervous system in the muscles’ activation.
Author response image 2.
Aggregate peristimulus time histogram (PSTH) for all motor unit spike times in the long head (top) and lateral head (bottom) within the stride.
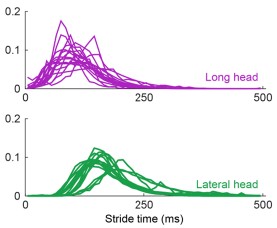
In the ideal case, we would have more simultaneous recordings from both muscles to make a more direct claim on the delay. Still, within the current scope of the paper, to correct this and better describe the difference in timing of muscle activity, we edited the text to the following:
“These findings demonstrate that despite the synergistic (extensor) function of the long and lateral heads of the triceps at the elbow, the motor pool for the long head becomes active roughly 100 ms before the motor pool supplying the lateral head during locomotion (Figure 3C).”
The results from Marshall et al. 2022 suggest that the recruitment of some MUs is not just related to muscle force, but also the frequency of force variation - some of their MUs appear to be recruited only at certain frequencies. Figure 5C could have shown signs of this, but it does not appear to. We do not really know the force or its frequency of variation in the measurements here. I wonder whether there is additional analysis that could address whether frequency-dependent recruitment is present. It may not be addressable with the current data set, but this could be a fruitful direction to explore in the future with MU recordings from mice.
We agree that this would be a fruitful direction to explore, however the Reviewer is correct that this is not easily addressable with the dataset. As the Reviewer points out, stride frequency increases with increased speed, potentially offering the opportunity to examine how motor unit activity varies with the frequency, phase, and amplitude of locomotor movements. However, given our lack of force data (either joint torques or ground reaction forces), dissociating the frequency/phase/amplitude of skeletal kinematics from the frequency/phase/amplitude of muscle force. Marshall et al. (2022) mitigated these issues by using an isometric force-production task (Marshall et al., 2022). Therefore, while we agree that it would be a major contribution to extend such investigations to whole-body movements like locomotion, given the complexities described above we believe this is a project for the future, and beyond the scope of the present study.
Minor:
Page 5: "Units often displayed no recruitment in a greater proportion of strides than for any particular spike count when recruited (Figures 2A, B)," - I had to read this several times to understand it. I suggest rephrasing for clarity.
We have changed the text to read:
“Units demonstrated a variety of firing patterns, with some units producing 0 spikes more frequently than any non-zero spike count (Figure 2A, B),...”
Figure 3 legend: "Mean phase ({plus minus} SE) of motor unit burst duration across all strides.": It is unclear what this means - durations are not usually described as having a phase. Do we mean the onset phase?
We have changed the text to read:
“Mean phase ± SE of motor unit burst activity within each stride”
Page 9: "suggesting that the recruitment of individual motor units in the lateral and long heads might have significant (and opposite) effects on elbow angle in strides of similar speed (see Discussion)." I wouldn't say "opposite" here - that makes it sound like the authors are calling the long head a flexor. The authors should rephrase or clarify the sense in which they are opposite.
This is a fair point and we agree we should not describe the muscles as ‘opposite’ when both muscles are extensors. We have removed the phrase ‘and opposite’ from the text.
Page 11: "in these two muscles across in other quadrupedal species" - typo.
We have corrected this error.
Page 16: This reviewer cannot decipher after repeated attempts what the first two sentences of the last paragraph mean. - “Future studies might also use perturbations of muscle activity to dissociate the causal properties of each motor unit’s activity from the complex correlation structure of locomotion. Despite the strong correlations observed between motor unit recruitment and limb kinematics (Fig. 6, Supplemental Fig. 3), these results might reflect covariations of both factors with locomotor speed rather than the causal properties of the recorded motor unit.”
For better clarity, we have changed the text to read:
“Although strong correlations were observed between motor unit recruitment and limb kinematics during locomotion (Figure 6, Figure 6–figure supplement 1), it remains unclear whether such correlations actually reflect the causal contributions that those units make to limb movement. To resolve this ambiguity, future studies could use electrical or optical perturbations of muscle contraction levels (Kim et al., 2024; Lu et al., 2024; Srivastava et al., 2015, 2017) to test directly how motor unit firing patterns shape locomotor movements. The short-latency effects of patterned motor unit stimulation (Srivastava et al., 2017) could then reveal the sensitivity of behavior to changes in muscle spiking and the extent to which the same behaviors can be performed with many different motor commands.”
Reviewer #2 (Recommendations for the authors):
Minor comments:
Introduction:
(1) "Although studies in primates, cats, and zebrafish have shown that both the number of active motor units and motor unit firing rates increase at faster locomotor speeds (Grimby, 1984; Hoffer et al., 1981, 1987; Marshall et al., 2022; Menelaou & McLean, 2012)." I would remove Marshall et al. (2022) as their monkeys performed pulling tasks with the upper limb. You can alternatively remove locomotor from the sentence and replace it with contraction speed.
Thank you for the comment. While we intended to reference this specific paper to highlight the rhythmic activity in muscles, we agree that this deviates from ‘locomotion’ as it is referenced in the other cited papers which study body movement. We have followed the Reviewer’s suggestion to remove the citation to Marshall et al.
(2) "The capability and need for faster force generation during dynamic behavior could implicate motor unit recruitment as a primary mechanism for modulating force output in mice."
The authors could add citations to this sentence, of works that showed that recruitment speed is the main determinant of the rate of force development (see for example Dideriksen et al. (2020) J Neurophysiol; J. L. Dideriksen, A. Del Vecchio, D. Farina, Neural and muscular determinants of maximal rate of force development. J Neurophysiol 123, 149-157 (2020)).
Thank you for pointing out this important reference. We have included this as a citation as recommended.
Results:
(3) "Electrode arrays (32-electrode Myomatrix array model RF-4x8-BHS-5) were implanted in the triceps brachii (note that Figure 1D shows the EMG signal from only one of the 16 bipolar recording channels), and the resulting data were used to identify the spike times of individual motor units (Figure 1E) as described previously (Chung et al., 2023)."
This sentence can be misleading for the reader as the array used by the researchers has 4 threads of 8 electrodes. Would it be possible to specify the number of electrodes implanted per head of interest? I assume 8 per head in most mice (or 4 bipolar channels), even if that's not specifically written in the manuscript.
Thank you for the suggestion. As described above, we have added Table 1, which includes all array locations, and we edited the statement referenced in the comment as follows:
“Electrode arrays (32-electrode Myomatrix array model RF-4x8-BHS-5) were implanted in forelimb muscles (note that Figure 1D shows the EMG signal from only one of the 16 bipolar recording channels), and the resulting data were used to identify the spike times of individual motor units in the triceps brachii long and lateral heads (Table 1, Figure 1E) as described previously (Chung et al., 2023).“
(4) "These findings demonstrate that despite the overlapping biomechanical functions of the long and lateral heads of the triceps, the nervous system creates a consistent, approximately 100 ms delay (Figure 3C) between the activation of the two muscles' motor neuron pools. This timing difference suggests distinct patterns of synaptic input onto motor neurons innervating the lateral and long heads."
Both muscles don't have fully overlapping biomechanical functions, as one of them also acts on the shoulder joint. Please be more specific in this sentence, saying that both muscles are synergistic at the elbow level rather than "have overlapping biomechanical functions".
We agree with the above reasoning and that our manuscript should be clearer on this point. We edited the above text in accordance with the Reviewer suggestion as follows:
"These findings demonstrate that despite the synergistic (extensor) function of the long and lateral heads of the triceps at the elbow, …”
(5) "Together with the differences in burst timing shown in Figure 3B, these results again suggest that the motor pools for the lateral and long heads of the triceps receive distinct patterns of synaptic input, although differences in the intrinsic physiological properties of motor neurons innervating the two muscles might also play an important role."
It is difficult to draw such an affirmative conclusion on the synaptic inputs from the data presented by the authors. The differences in firing rates may solely arise from other factors than distinct synaptic inputs, such as the different intrinsic properties of the motoneurons or the reception of distinct neuromodulatory inputs.
To better explain our findings, we adjusted the above text in the Results (see “Motor unit firing patterns in the long and lateral heads of the triceps”):
“Together with the differences in burst timing shown in Figure 3B, these results again suggest that the motor pools for the lateral and long heads of the triceps receive distinct patterns of synaptic input, although differences in the intrinsic physiological properties of motor neurons innervating the two muscles might also play an important role.”
We also included the following distinction in the Discussion (see “Differences in motor unit activity patterns across two elbow extensors”) to address the other plausible mechanisms mentioned.
“The large differences in burst timing and spike patterning across the muscle heads suggest that the motor pools for each muscle receive distinct inputs. However, differences in the intrinsic physiological properties of motor units and neuromodulatory inputs across motor pools might also make substantial contributions to the structure of motor unit spike patterns (Martínez-Silva et al., 2018; Miles & Sillar, 2011).”
(6) "We next examined whether the probabilistic recruitment of individual motor units in the triceps and elbow extensor muscle predicted stride-by-stride variations in elbow angle kinematics."
I'm not sure that the wording is appropriate here. The analysis does not predict elbow angle variations from parameters extracted from the spiking activity. It rather compares the average elbow angle between two conditions (motor unit active or not active).
We thank the Reviewer for this comment and agree that the wording could be improved here to better reflect our analysis. To lower the strength of our claim, we replaced usage of the word ‘predict’ with ‘correlates’ in the above text and throughout the paper when discussing this result.
Methods:
(7) "Using the four threads on the customizable Myomatrix array (RF-4x8-BHS-5), we implanted a combination of muscles in each mouse, sometimes using multiple threads within the same muscle. [...] Some mice also had threads simultaneously implanted in their ipsilateral or contralateral biceps brachii although no data from the biceps is presented in this study."
A precise description of the localisation of the array (muscles and the number of arrays per muscle) for each animal would be appreciated.
(8) "A total of 33 units were identified and manually verified across all animals." A precise description of the number of motor units concurrently identified per muscle and per animal would be appreciated. Moreover, please add details on the manual inspection. Does it involve the manual selection of missing spikes? What are the criteria for considering an identified motor unit as valid?
As discussed earlier, we added Table 1 to the main text to provide the details mentioned in the above comments.
Regarding spike sorting, given the very large number of spikes recorded, we did not rely on manual adjusting mislabeled spikes. Instead, as described in the revised Methods section, we verified unit isolation by ensuring units had >98% of spikes outside of 1ms of each other. Moreover, as described above we have added new analyses (Figure 1–figure supplement 1) confirming the stability of motor unit waveforms across both the duration of individual recording sessions (roughly 30 minutes) and across the rapid changes in limb position within individual stride cycles (roughly 250 msec).
Reviewer #3 (Recommendations for the authors):
Figure 2 (and supplement) show spike count distributions with strong positive skewness, which is in accordance with the prediction of a fluctuation-driven regime. I suggest plotting these on a logarithmic x-axis (in addition to the linear axis), which should reveal a bell-shaped distribution, maybe even Gaussian, in a majority of the units.
We thank the Reviewer for the suggestion. We present the requested analysis below, which shows bell-shaped distributions for some (but not all) distributions. However, we believe that investigating why some replotted distributions are Gaussian and others are not falls beyond the scope of this paper, and likely requires a larger dataset than the one we were able to obtain.
Author response image 3.
Spike count distributions for each motor unit on a logarithmic x-axis.
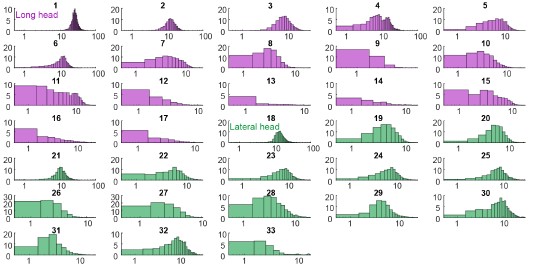
Why not more data? I tried to get an overview of how much data was collected.
Supplemental Figure 1 has all the isolated units, which amounts to 38 (are the colors the two muscle types?). Given there are 16 leads in each myomatrix, in two muscles, of six mice, this seems like a low yield. Could the authors comment on the reasons for this low yield?
Regarding motor unit yield, even with multiple electrodes per muscle and a robust sorting algorithm, we often isolated only a few units per muscle. This yield likely reflects two factors. First, because of the highly dynamic nature of locomotion and high levels of muscle contraction, isolating individual spikes reliably across different locomotor speeds is inherently challenging, regardless of the algorithm being employed. Second, because the results of spike-train analyses can be highly sensitive to sorting errors, we have only included the motor units that we can sort with the highest possible confidence across thousands of strides.
Minor:
Figure captions especially Figure 6: The text is excessively long. Can the text be shortened?
We thank the Reviewer for this comment. Generally, we seek to include a description of the methods and results within the figure captions, but we concede that we can condense the information in some cases. In a number of cases, we have moved some of the descriptive text from the caption to the Methods section.
References
Berg, R. W. (2017). Neuronal Population Activity in Spinal Motor Circuits: Greater Than the Sum of Its Parts. Frontiers in Neural Circuits, 11. https://doi.org/10.3389/fncir.2017.00103
Biewener, A. A., Blickhan, R., Perry, A. K., Heglund, N. C., & Taylor, C. R. (1988). Muscle Forces During Locomotion in Kangaroo Rats: Force Platform and Tendon Buckle Measurements Compared. Journal of Experimental Biology, 137(1), 191–205. https://doi.org/10.1242/jeb.137.1.191
Chung, B., Zia, M., Thomas, K. A., Michaels, J. A., Jacob, A., Pack, A., Williams, M. J., Nagapudi, K., Teng, L. H., Arrambide, E., Ouellette, L., Oey, N., Gibbs, R., Anschutz, P., Lu, J., Wu, Y., Kashefi, M., Oya, T., Kersten, R., … Sober, S. J. (2023). Myomatrix arrays for high-definition muscle recording. eLife, 12, RP88551. https://doi.org/10.7554/eLife.88551
De Luca, C. J. (1985). Control properties of motor units. Journal of Experimental Biology, 115(1), 125–136. https://doi.org/10.1242/jeb.115.1.125
De Luca, C. J., & Erim, Z. (1994). Common drive of motor units in regulation of muscle force. Trends in Neurosciences, 17(7), 299–305. https://doi.org/10.1016/0166-2236(94)90064-7
Farina, D., Negro, F., & Dideriksen, J. L. (2014). The effective neural drive to muscles is the common synaptic input to motor neurons. The Journal of Physiology, 592(16), 3427–3441. https://doi.org/10.1113/jphysiol.2014.273581
Hartigan, P. M. (1985). Algorithm AS 217: Computation of the Dip Statistic to Test for Unimodality. Applied Statistics, 34(3), 320. https://doi.org/10.2307/2347485
Henneman, E., Somjen, G., & Carpenter, D. O. (1965). FUNCTIONAL SIGNIFICANCE OF CELL SIZE IN SPINAL MOTONEURONS. Journal of Neurophysiology, 28(3), 560–580. https://doi.org/10.1152/jn.1965.28.3.560
Karabulut, D., Dogru, S. C., Lin, Y.-C., Pandy, M. G., Herzog, W., & Arslan, Y. Z. (2020). Direct Validation of Model-Predicted Muscle Forces in the Cat Hindlimb During Locomotion. Journal of Biomechanical Engineering, 142(5), 051014. https://doi.org/10.1115/1.4045660
Kim, J. J., Wyche, I. S., Olson, W., Lu, J., Bakir, M. S., Sober, S. J., & O’Connor, D. H. (2024). Myo-optogenetics: Optogenetic stimulation and electrical recording in skeletal muscles. https://doi.org/10.1101/2024.06.21.600113
Lu, J., Zia, M., Baig, D. A., Yan, G., Kim, J. J., Nagapudi, K., Anschutz, P., Oh, S., O’Connor, D., Sober, S. J., & Bakir, M. S. (2024). Opto-Myomatrix: μLED integrated microelectrode arrays for optogenetic activation and electrical recording in muscle tissue. https://doi.org/10.1101/2024.07.01.601601
Manuel, M., & Heckman, C. J. (2011). Adult mouse motor units develop almost all of their force in the subprimary range: A new all-or-none strategy for force recruitment? Journal of Neuroscience, 31(42), 15188–15194. https://doi.org/10.1523/JNEUROSCI.2893-11.2011
Marshall, N. J., Glaser, J. I., Trautmann, E. M., Amematsro, E. A., Perkins, S. M., Shadlen, M. N., Abbott, L. F., Cunningham, J. P., & Churchland, M. M. (2022). Flexible neural control of motor units. Nature Neuroscience, 25(11), 1492–1504. https://doi.org/10.1038/s41593-022-01165-8
Martínez-Silva, M. de L., Imhoff-Manuel, R. D., Sharma, A., Heckman, C. J., Shneider, N. A., Roselli, F., Zytnicki, D., & Manuel, M. (2018). Hypoexcitability precedes denervation in the large fast-contracting motor units in two unrelated mouse models of ALS. eLife, 7(2007), 1–26. https://doi.org/10.7554/eLife.30955
Miles, G. B., & Sillar, K. T. (2011). Neuromodulation of Vertebrate Locomotor Control Networks. Physiology, 26(6), 393–411. https://doi.org/10.1152/physiol.00013.2011
Petersen, P. C., & Berg, R. W. (2016). Lognormal firing rate distribution reveals prominent fluctuation–driven regime in spinal motor networks. eLife, 5. https://doi.org/10.7554/elife.18805
Srivastava, K. H., Elemans, C. P. H., & Sober, S. J. (2015). Multifunctional and Context-Dependent Control of Vocal Acoustics by Individual Muscles. The Journal of Neuroscience, 35(42), 14183–14194. https://doi.org/10.1523/JNEUROSCI.3610-14.2015
Srivastava, K. H., Holmes, C. M., Vellema, M., Pack, A. R., Elemans, C. P. H., Nemenman, I., & Sober, S. J. (2017). Motor control by precisely timed spike patterns. Proceedings of the National Academy of Sciences of the United States of America, 114(5), 1171–1176. https://doi.org/10.1073/pnas.1611734114
Humans are subject to these material needs, but their purposeand agency are not set by the imperatives of material survival.The purposes of human lives are set by normative not naturalimperatives. By “ought” not “instinct”. These normative pur-poses, in turn, can be changed in the light of self-reflection.“As distinct from natural freedom, spiritual freedom requiresthe ability to ask which imperatives to follow in the light of ourends, as well as the ability to call into question, challenge andtransform our ends themselves”.28Humans also generate surplus time, after material necessi-ties have been met, but nothing tells them how they ought touse this time. They themselves must decide this. Material andspiritual freedom are inseparable – there is no spiritual realmapart from the material – but they are not the same. Spiritualfreedom is a higher order of freedom; it is dependent on naturebut transcends it. The way we lead our lives is not set by mate-rial necessity but set by us. To which I would add that culture,at its broadest, is the name we give to the collective patterningof our material and spiritual freedoms, patterns based not juston communication – all plants and animals communicate – buton symbolisation, forms of speaking about what we ought to do.
In other words, we have to fill our time lest it be boring enough to not be worth living. It's an uphill battle against the vastness of the universe as a Sisyphus' giant boulder up the hill of society. We rationalise. We have to, to exist: Once we accept patterns and we are made aware of our ability to layer multiple of them, we rely on social constructs, on organising fictions, on tales, the stories we tell ourselves and others, on narratives (a la Byung Chul), religions, the argument behind our actions, the why, our purpose, our goals… these reasons are needed. Culture is needed.
Culture, in both its individual tra-jectory and its social guarantee, is thus essential to the develop-ment of individual capabilities, as in the work of Amartya Senand Martha Nussbaum, and thus an essential part of demo-cratic citizenship, a fundamental human right.
Very Descartes-Rosseau declinclination to curtal Tocqueville's degeneration of Tyranny of the Majority, in line with Rawlsian golden rule "Veil of Ignorance" anti-bias original state requirements. Still, so many things more would be needed to achieve this! The question is more, I argue, of what to prioritise. I am sorry to say that although I agree with Appadurai’s thesis that doing science should be a human right, I cannot see it happening. Not everyone can do science, at least not simultaneously (if we take it as more than revisionist critical thinking), because other tasks like farming, building, or healing are needed. If these are to fall under the umbrella of "culture", not from an optimisation perspective but a care one, then I think we first and foremost need to change the system we live under, capitalism, and sure, culture can help do it, although maybe through isolationism (which was Nozick's critique of socialism as imposition in the form of Imperialistic Utopians, after all, the same argument was used to "civilise" Eastern indigenous).
Health, education,housing, transport, food, energy, water, basic communicationsystems – should these not be fixed before we get to culture?1In what follows I make an argument for the centrality ofculture in overcoming the present crisis, and in the followingchapters sketch out what such an agenda might look like. Weneed to acknowledge the “legitimation crisis” to which culture-as- industry was a response – art associated with elitism andpatrician subsidy, the growth of the culture industries
But culture is health! A culture of health, of dancing, of being engaged in mentally challenging projects. Culture is education, it teaches, it creates communities. Culture might not be food per se, but it can help grow food, and cook it, collectively, with shared non-contaminating gardens, and recycling processes, through rites more often depicted in religion.
Culture is communication when misscomunication is rampant: It's not just posting and sharing the life of your children online, it's being aware of its impacts, checking before sending news, and feeling safe enough to talk about suicide and emotions, or to speak up about a close relative who is harassing you, regardless of gender! Is this not a progressivist culture that does not invisibilise these things to private life?
Or rather, arguably, culture could act as these things. Currently, mass consumption culture is at times an inhibating mass-sterelisation device, and at others, a way to propagate neo-colonialist monopolistic ideals who benefit a wealthy few. It also perpetuates the myth of equality and meritocracy, through ads, and its instrumentalist tokenised portrayal of diverse hires and rampant on your face corruption and ideals buying, ultimately makes change within the system almost impossible.
Neoliberalism, a distinct phase of global capitalism thatemerged in the early 1980s, is in crisis. The economic, politi-cal, and social solutions it brought to the prior crisis of Fordistsocial democracy in the 1970s no longer work. That muchmight be broadly agreed. It has become ever clearer since the2008– 10 global financial crisis. Stagnant growth, stalled wages,growing inequality, cost- of-living increases, rising indebtedness,increasing precarity of employment, and the erosion of publicservices are the stuff of the nightly news. This is accompaniedby widespread disaffection and disconnection from the politicalprocess, resulting in alternating moments of political efferves-cence (often as anti-politics) and resigned passivity. This in theface of a growing perception of the deepening hold of powerfulelites over the workings of government, in the form of lobby-ing, political finance, corruption, cronyism, the public–private“revolving door”, and a pervasive cynicism towards techno-cratic political expertise
Yes, but there have been some welfare and rights advances! Think women and LGBTIQA+ rights. There is increasing cost because demographics have changed, it's not just automation and IA giving the capitalists excess capital gains. Polarisation and corruption are a byproduct of mass production, of noisy, post-truth production.
For TimJackson, modern consumerism is a catastrophe, and the soonerwe exit from it the better. Kate Soper, who flags a less puri-tanical approach in her book on “alternative hedonism”, alsosees much of modern cultural consumption as the problem tobe solved.
But is this culture? And if social media, TV, Netflix, Spotify, and YouTube, OnlyFans, is culture, and that's where people spend their time in, how can we break it down so culture becomes grassroots and not a client service? How can it help establish two-way caring? Yes to destroying bigTech, bigEd, bigAg, bigPharma, and bigClothing/Jewelry, but NOT to having tech, education, health, and cool things to wear. Less, more durable, repairable.
The status of the resource in the context of maturity lifecycle.
I have a question regarding the use of this property: Does it refer to the status of the dataset (or another resource) that some metadata refers to or does it directly refer to the metadata itself? The latter would make more sense to me, but it's not entirely clear from the description. (At least not to me.)
without mechanisms to protect European champions from foreign acquisition, any progress can be undone overnight
This is exactly what we have had for the past 25 years in Europe. And you can design against it, even if you don't want to or cannot keep out foreign investment. In the Solvinity case it would mean upfront knowing how and where to deploy your services before any US investments happen, and a dissolution clause in your contract. You mitigate not by prevent investment or wanting European champions, you mitigate by upfront having options that you can quickly deploy, so options getting provided. If you want to work for the European public sector you cannot have any CLOUD Act involvement. And you have to know you will lose your entire client base in the public sector if you do. Snowcrash, but yeah. And a herd of zebra's, not seeking unicorns.
For Europe, the question is no longer whether it should pursue digital sovereignty, but whether it has the collective will to stop talking, start building, and, most importantly, distinguish real autonomy from clever marketing.
again confusing sovereignty with autonomy. Action does not have collective will as prerequisite, although it helps mobilising more resources.
For these individual successes to scale into a continent-wide shift, however, structural barriers must be addressed. The path to digital sovereignty is not a single, grand gesture but a series of deliberate, often difficult, choices. The examples from Austria, France, and the ICC show that the journey begins with a single, courageous step, often prompted by the mundane reality of a data protection assessment.
It's not about scaling, it's about spreading. Not the same thing. No scaler involved. Which is said in the second sentence: 'not a single grand gesture but a series'
These cases illustrate that while a complete break from US hyperscalers may seem unrealistic, as Forrester predicts, targeted migrations for specific, high-risk applications are not only possible but are actively being pursued. They represent a grassroots movement, driven by legal obligations and a growing desire for autonomy.
another odd sentence.
targeted migrations is how you break from hyperscalers. Not by expecting to create your own hyperscaler (which would also then be a potential monopolist and threat to digital autonomy if not sovereignty like US stuff)
It's not 'grassroots' if national government entities are switching. What would be top-down in this case?
The International Criminal Court (ICC) in The Hague announced in November 2025 it was replacing its Microsoft office software with a European alternative.
Not a really good example, bc it's an international entity, but yes.
For Caffarra, this trend is dangerously reminiscent of Gaia-X, a previous flagship initiative for a federated European cloud. "The intention behind Gaia-X was good," she says. "The problem was that American companies lobbied to be included. Once Microsoft, Google, and AWS were inside Gaia-X, the initiative lost its purpose.". In her opinion: "That is why it failed."
not sure that is solely why it failed, but it did make the entire thing moot that is for sure.
from talking to building, from regulating to investing, and from passivity to action. Europe cannot rely on Brussels to deliver this transformation. The market must build it, but governments must create the conditions, through procurement preferences and initial funding, that make it viable.
yes.
The goal, she emphasizes, is not autarky or protectionism, but resilience. Europe does not need to achieve complete independence from American technology, but it does need to reclaim a meaningful share of its own market. "Can we please have 30 to 40 percent for ourselves?"
30-40% is still a minimal step imo. Yes, resilience. But for the public sector I cannot see how any public sector body can use US or Chinese digital tech bc all of it is in non-compliance w EU regs. Make the compliance a kick-out req in procurement.
Her proposed solution, embodied in the Eurostack Foundation, is not more regulation but an industrial strategy focused on three pillars:
With which she says the regulatory framework is useful as is?
Its use is strictly limited to external communication with parties that still rely on it, such as the European Commission. Even then, strict rules apply: no sensitive information may be discussed on Teams, and usage is kept to an absolute minimum. This hybrid approach reflects a pragmatic recognition that complete independence is not always immediately possible when external partners remain locked into US platforms.
yes, it is a pragmatic choice. Similar to us, we don't use any GAFAM but still use Teams because clients do.
Yet the Austrian case also illustrates the practical limits of digital sovereignty. Nextcloud now serves as the primary collaboration platform for internal communication and file sharing, but Microsoft Teams has not been banned entirely.
Odd formulation, switching from Office to Teams in one sentence which are different things.
"You don't achieve digital sovereignty overnight," Ollrom tells The Register. "You have to do this in many steps, but you have to start with the first step. Don't just talk about it, but execute it."
agreed, a first step to break status quo is important.
Austria's Federal Ministry for Economy, Energy and Tourism is a case in point. The ministry recently completed a migration of 1,200 employees to the European open-source collaboration platform Nextcloud, but the project was not a migration away from an existing US cloud provider. It was a deliberate choice not to adopt one
Austrian Min for Economy Energy and Tourism uses Nextcloud for its 1200 civil servants, not MS Office. It was not a move away from GAFAM bc they did not use that.
Author response:
The following is the authors’ response to the previous reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
Mazar & Yovel 2025 dissect the inverse problem of how echolocators in groups manage to navigate their surroundings despite intense jamming using computational simulations.
The authors show that despite the 'noisy' sensory environments that echolocating groups present, agents can still access some amount of echo-related information and use it to navigate their local environment. It is known that echolocating bats have strong small and large-scale spatial memory that plays an important role for individuals. The results from this paper also point to the potential importance of an even lower-level, short-term role of memory in the form of echo 'integration' across multiple calls, despite the unpredictability of echo detection in groups. The paper generates a useful basis to think about the mechanisms in echolocating groups for experimental investigations too.
Strengths:
The paper builds on biologically well-motivated and parametrised 2D acoustics and sensory simulation setup to investigate the various key parameters of interest
The 'null-model' of echolocators not being able to tell apart objects & conspecifics while echolocating still shows agents succesfully emerge from groups - even though the probability of emergence drops severely in comparison to cognitively more 'capable' agents. This is nonetheless an important result showing the direction-of-arrival of a sound itself is the 'minimum' set of ingredients needed for echolocators navigating their environment.
The results generate an important basis in unraveling how agents may navigate in sensorially noisy environments with a lot of irrelevant and very few relevant cues.
The 2D simulation framework is simple and computationally tractable enough to perform multiple runs to investigate many variables - while also remaining true to the aim of the investigation.
Weaknesses:
Authors have not yet provided convincing justification for the use of different echolocation phases during emergence and in cave behaviour. In the previous modelling paper cited for the details - here the bat-agents are performing a foraging task, and so the switch in echolocation phases is understandable. While flying with conspecifics, the lab's previous paper has shown what they call a 'clutter response' - but this is not necessarily the same as going into a 'buzz'-type call behaviour. As pointed out by another reviewer - the results of the simulations may hinge on the fact that bats are showing this echolocation phase-switching, and thus improving their echo-detection. This is not necessarily a major flaw - but something for readers to consider in light of the sparse experimental evidence at hand currently.
The use of echolocation phases—defined as the sequential search, approach, and buzz call patterns—has been documented not only during foraging but also in tasks such as landing, obstacle avoidance, clutter navigation, and drinking. Bat call structure has been shown to vary systematically with object proximity, not exclusively in response to prey. During obstacle avoidance, phase transitions were observed, with approach calls emitted in grouped sequences and with reduced durations (Gustafson & Schnitzler, 1979; Schnitzler et al., 1987). In landing contexts, bats have been reported to emit short-duration calls and decrease inter-pulse intervals—buzz-like patterns also observed during prey capture— suggesting shared acoustic strategies across behaviors (Hagino et al., 2007; Hiryu et al., 2008; Melcón et al., 2007, 2009). Comparable patterns have been reported during drinking maneuvers, where “drinking buzzes” have been proposed to guide a precise approach to the water surface, analogous to landing buzzes (Griffiths, 2013; Russo et al., 2016). In response to environmental complexity, bats were found to shorten calls and increase repetition rates when navigating cluttered spaces compared to open ones (Falk et al., 2014; Kalko & Schnitzler, 1993).
Moreover, field recordings from our study of Rhinopoma microphyllum (Goldshtein et al., 2025) revealed shortened call durations and inter-pulse intervals during dense group flight outside the cave during emergence—patterns consistent with terminal-approach phase that is typical when coming very close to an object (another bat in this case). The Author response image 1 shows an approach sequence recorded from a tagged bat approximately 20 meters from the cave entrance, with self-generated echolocation calls marked. The inter-pulse-interval of ca. 20 ms is used by these bats when a reflective object (another bat in this case) is nearby.
Author response image 1.
These results provide direct evidence that bats actively employ approach-phase echolocation during swarming likely to avoid collision with other bats. This supports the view that echolocation phase transitions are a general proximity-based sensing strategy, adapted across a variety of behavioral scenarios—not limited to hunting alone.
In our simulations, bats predominantly emitted calls in the approach phase, with only rare occurrences of buzz-phase calls.
See lines 355-363 in the revised manuscript.
The decision to model direction-of-arrival with such high angular resolution (1-2 degrees) is not entirely justifiable - and the authors may wish to do simulation runs with lower angular resolution. Past experimental paradigms haven't really separated out target-strength as a confounding factor for angular resolution (e.g. see the cited Simmons et al. 1983 paper). Moreover, to this reviewer's reading of the cited paper - it is not entirely clear how this experiment provides source-data to support the DoA-SNR parametrisation in this manuscript. The cited paper has two array-configurations, both of which are measured to have similar received levels upon ensonification. A relationship between angular resolution and signal-to-noise ratio is understandable perhaps - and one can formulate such a relationship, but here the reviewer asks that the origin/justification be made clear. On an independent line, also see the recent contrasting results of Geberl, Kugler, Wiegrebe 2019 (Curr. Biol.) - who suggest even poorer angular resolution in echolocation.
We thank the reviewer for raising this important point. The acuity of 1.5–3° in horizontal direction-of-arrival (DoA) estimation is based on the classical work of Simmons et al. with Eptesicus fuscus (Simmons et al., 1983). Similar precision was later supported by Erwin et al. (Erwin et al., 2001), who modeled azimuth estimation from measured interaural intensity differences (IIDs), reporting an average error of 0.2° with a standard deviation of ~2.2°, consistent with the behavioral data found by Simmons. The decline in acuity with increasing arrival angle has also been demonstrated in behavioral and physiological studies of binaural IID processing (Erwin et al., 2001; Fay, 1995; Razak, 2012; Wohlgemuth et al., 2016). The error model itself was first introduced in our earlier work (Mazar & Yovel, 2020).
Importantly, Geberl et al. (Geberl et al., 2019) examined the resolution of weak targets masked by nearby strong flankers and found poor spatial discrimination of ~45 degrees; however, they were studying a detection problem, rather than the horizontal acuity of azimuth estimation. Indeed, our model assumes there is no spatial discrimination at all.
Overall, while our DoA–SNR parametrization can certainly be critiqued and alternative parameterizations could be tested in future work, we believe it reflects a reasonable and empirically supported assumption.
Reviewer #2 (Public review):
This manuscript describes a detailed model for bats flying together through a fixed geometry. The model considers elements which are faithful to both bat biosonar production and reception and the acoustics governing how sound moves in air and interacts with obstacles. The model also incorporates behavioral patterns observed in bats, like one-dimensional feature following and temporal integration of cognitive maps. From a simulation study of the model and comparison of the results with the literature, the authors gain insight into how often bats may experience destructive interference of their acoustic signals and those of their peers, and how much such interference may actually negatively effect the groups' ability to navigate effectively. The authors use generalized linear models to test the significance of the effects they observe.
The work relies on a thoughtful and detailed model which faithfully incorporates salient features, such as acoustic elements like the filter for a biological receiver and temporal aggregation as a kind of memory in the system. At the same time, the authors abstract features that are complicating without being expected to give additional insights, as can be seen in the choice of a two-dimensional rather than three-dimensional system. I thought that the level of abstraction in the model was perfect, enough to demonstrate their results without needless details. The results are compelling and interesting, and the authors do a great job discussing them in the context of the biological literature.
With respect to the first version of the manuscript, the authors have remedied all my outstanding questions or concerns in the current version. The new supplementary figure 5 is especially helpful in understanding the geometry.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
Data Availability: This reviewer lauds the authors for switching from a private commercial folder requiring login to one that does not. At the cost of being overtly pedantic - the Github repository is not a long-term archival resource. The ideal solution is to upload the code in an academic repository (Zenodo, OSF, etc.) to periodically create a 'static snapshot' of code for archival, while also hosting a 'live' version on Github.
We have uploaded to Zenodo repository, and updated the link in the paper:
How bats exit a crowded colony when relying on echolocation only - a modeling approach
In one of the rebuttals to Reviewer #3- the authors have cited a wrong paper (Beleyur & Goerlitz 2019) - while discussing broad bandwidth calls improving detection - and may wish to correct this if possible on record.
We have removed the incorrect citation from the revised version of the manuscript.
Specific comments on the 2nd manuscript:
Figure 5: Table 1 says 1, 2,5,10,20,40,100 bats were simulated (line 138-139) but the conclusion (line 398) says '1 to 100 bats' per 3msq. However, the X-axis only stops at 40 and says 'number of bats', while the legend says bats/3msq....what is actually being plotted? Moreover, in the entire paper there is a constant back-and-forth between density and # of bats - perhaps it is explained beforehand, but it is a bit unsettling - and more can be done to clarify these two conventions.
While most parameters were tested across the full range of 1 to 100 bats per 3 m², a subset of conditions—including misidentification, multi-call clustering, wall target strength, and conspecific target strength—were simulated only up to 40 bats due to significantly longer run-times. This is now clarified in both the main text and the Table 1 caption.
In our simulations, the primary parameter was the number of bats placed within a 3 m² starting area, which directly determined the initial density (bats per 3 m²). Throughout the manuscript, we use “number of bats” to refer to the simulation input, while “density” denotes the equivalent ecological measure. Figure 5 and related captions have been revised accordingly to note these conventions and to indicate when results are shown only up to 40 bats (see lines 120–122, 314-317 in the revised text).
Table 1: This was made considerably difficult to read given the visual clutter - and I hope I've understood these changes correctly.
What is in the square brackets of the effect-size (e.g. first row with values 'Exit prob. (%)' says -0.37/bat [63:100] ? What does this 63:100 refer to?
What is the 'process flag'
Values in square brackets indicate the minimum and maximum values of the metric across the tested range (e.g., [63:100] shows the range of exit probabilities observed across different bat densities).
The term “process flag” has been replaced with “with and without multi-call clustering” for clarity
Both the table layout and caption have been revised to reduce visual clutter and to make these conventions clearer to the reader.
Lines 562-3: "In our study, due to the dense cave environment, the bats are found to operate in the approach phase nearly all of the time, which is consistent with natural cave emergence behavior" - bats are 'found to' implies there is some experimental data or it is an emergent property. See above for the point questioing the implementation of multiple echolocation phases in the model, but also - here the bat-agents are allowed to show different phases and thus they do so -- it is a constraint of the implementation and not a result per se given the size of the cave and the number of bats involved...
We removed the sentence from the Methods section, since it could be misinterpreted as an experimental finding rather than a model outcome. Instead, we now discuss this in the Discussion, clarifying that the predominance of the approach phase arises from the cluttered cave environment in our simulations, which is consistent with natural emergence behavior (see lines 355-363). In this context, the use of echolocation phases is presented as a biologically plausible modeling choice rather than an empirical result.
Lines 659-660: The parametrisation between DoA and SNR is supposedly found in 'Equation 10' - which this reviewer could not find in the manuscript
The equation was accidentally omitted in the previous revision and has now been reinserted into the manuscript. It defines how direction-of-arrival (DoA) error depends on SNR and azimuth angle (see lines 603-605).
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Bouma et al. present a comprehensive analysis of DOT1L-mediated histone H3K79 methylation across canonical DC subsets. By mapping the methylation landscape, the authors demonstrate that DOT1L regulates both shared and subset-specific gene programs. They show that in vitro or in vivo deletion of Dot1l, followed by in vitro differentiation, results in reduced myeloid progenitors and pDCs alongside an increase in cDC2s, while cDC1 numbers remain largely unaffected. Functionally, Dot1l-deficient DCs fail to produce IFNα upon stimulation. Transcriptomic profiling reveals enrichment of antigen presentation pathways in Dot1l-KO subsets, with upregulated MHC class II surface expression in pDCs. Mechanistically, pharmacological inhibition of DOT1L links these effects to its methyltransferase activity. Collectively, the data suggest that DOT1L differentially regulates canonical DC subset development and represses antigen presentation pathways.
The manuscript is well-written and technically sound. However, several conclusions would benefit from deeper discussion or additional experimental validation.
Major Comments
The authors propose that DOT1L loss skews DC differentiation toward a pDC-like phenotype. However, DOT1L deletion or inhibition, and the consequent global loss of H3K79 methylation, is well known to downregulate key cell-cycle genes (e.g., Cyclin D1, Cyclin E, CDK4/6, MCM family) while upregulating cell-cycle inhibitors (e.g., Cdkn1a and b). These transcriptional changes are associated with slower proliferation, G1 arrest or delayed S-phase entry, and reduced DNA replication fork progression. Importantly, blocking DNA synthesis (e.g., with aphidicolin or mitomycin C) during early culture inhibits DC emergence, underscoring that proliferation is essential for differentiation. The authors should discuss how their findings align with this established literature. Could the observed DC subset shifts result from impaired cell-cycle progression rather than lineage-specific transcriptional reprogramming? A more detailed consideration of this point is needed. 2. Discrepancy between in vitro and in vivo pDC phenotypes
The in vitro data show a marked reduction in pDCs, yet in vivo pDC numbers appear unchanged. Although the discussion briefly mentions proliferation differences, this discrepancy deserves a clearer explanation or experimental follow-up.
Minor Comments
The current manuscript fills a gap in knowledge, and this is its major strength. Other strengths are clarity and technical appropriateness.
The major weakness is that the work is mainly descriptive. Mechanistic insights into DOT1L-dependent transcriptional regulation are still weak. The proposed mechanism -that DOT1L maintains pDC identity through H3K79 methylation at key transcription factors (Tcf4, SpiB, Irf8)- is intriguing but currently lacks functional evidence. The authors should consider validating this model experimentally, by modulating the expression of these genes without affecting DOT1L activity. Also the model suggesting that DOT1L indirectly represses antigen presentation via the Fbxo11-Ciita pathway is interesting but remains speculative. Additional mechanistic data would help support this claim.
[row for row in board if is_incomplete(row)]
A nitpick, but would inline the condition like this:
[row for row in board if 0 in row]
I get that this is maybe for clarity + aligning with clojure version thought :)
Not to be annoying, but did you know that I gave birth to the single most handsome, charismatic, adorable, intelligent, articulate, and literate baby to ever grace this earth?
Jayzus. Someone ought to feel guilty for even thinking up such a sentence.
Reviewer #2 (Public review):
Summary:
In this manuscript, Porras-Gómez et al. analyse the lipid composition and biophysical properties of pulmonary surfactant obtained by bronchoalveolar lavage (BAL) from a group of bottlenose dolphins (Tursiops truncatus), including two healthy individuals and five affected by pneumonia. Through lipidomic analysis, the authors report an exacerbated presence of cardiolipin species in the BAL lipid extracts from diseased dolphins compared to healthy ones. Structural analyses using electron microscopy, atomic force microscopy, and X-ray scattering on rehydrated membrane samples reveal that lipids from diseased animals form membranes with a more pronounced Lβ phase and reduced fluidity. Moreover, the membranes from affected lungs appear more interconnected and less hydrated, as indicated by the X-ray scattering data. These findings provide valuable and convincing insights into how pulmonary disease alters the lipid composition and structural properties of surfactant in diving mammals, and may have broader implications for understanding surfactant dysfunction in marine mammals.
Strengths:
The study is well designed, and the experimental techniques were applied in a logical and coherent manner. The results are thoroughly analysed and discussed, and the manuscript is clearly written and well organized, making it both easy to follow and scientifically robust. Although the number of samples is limited, the rarity and logistical challenges of obtaining bronchoalveolar lavage material, particularly from animals affected by respiratory disease, make this study especially valuable and relevant.
Weaknesses:
In my opinion, the main issue lies in the treatment of the samples. Pulmonary surfactant is a lipoprotein complex produced by type II pneumocytes of the alveolar epithelium in the form of compact and highly dehydrated structures known as tubular myelin. Once secreted, these structures unfold and, upon contact with the air-liquid interface, form an interfacial monolayer connected to surfactant membranes in the subphase, thereby facilitating respiratory dynamics throughout the breathing cycle.
When bronchoalveolar lavages are treated using the Bligh and Dyer method to extract the hydrophobic fraction of these samples, the structural complexity of the surfactant is disrupted, and this organization cannot be completely restored once the lipids are rehydrated. Although these extracts contain the hydrophobic proteins SP-B and SP-C, the hydrophilic protein SP-A may play an essential role in the formation of pulmonary surfactant structures. It is well established that SP-A is crucial for the formation of tubular myelin, an intermediate structure between the lamellar bodies newly secreted by type II cells and the interfacial surfactant layers.
Moreover, and more importantly, bronchoalveolar lavage fluid may contain cells, tissue debris, and even bacteria that can alter the lipid composition of the samples used in the study after extraction by the Bligh and Dyer method. For this reason, most studies include a density gradient centrifugation step to isolate the surfactant membranes. Consequently, the samples used may be contaminated with phospholipids originating from other cells, such as macrophages, pneumocytes, or bacterial cells, particularly in lavages obtained from diseased animals.
Although the techniques employed provide valuable information about the behaviour of surfactant membranes and allow certain inferences regarding their functionality, no functional studies of these samples have been conducted using methods such as the constrained drop surfactometer or the captive bubble surfactometer. The observed alterations do not necessarily demonstrate that surfactant modulates its properties, as claimed by the authors, but rather indicate that it is altered by the presence of other lipids.
The spin-coating technique used to form lipid films for analysis by atomic force microscopy is not the most suitable approach to reproduce the structures generated by pulmonary surfactant. However, the results obtained may still provide valuable insights into the biophysical behaviour of its components. The analysis of lung tissue shown in Supplementary Figure S3 presents the same limitation, as the samples were embedded in a cutting compound, and the measurements may have been taken from different regions of the tissue. Therefore, it cannot be ensured that the analysed structures correspond to those generated by pulmonary surfactant.
The finding that the structures formed in samples obtained from diseased animals are more tightly packed and dehydrated than those derived from the surfactant of healthy animals contrasts with the notion that the high efficiency of lamellar bodies in generating interfacial structures is related to their high degree of packing and dehydration. The formation of these structures involves the participation of the ABCA3 protein, which pumps phospholipids into the interior of lamellar bodies, and SP-B, which facilitates the formation of close membrane contacts.
While the results are interesting from a comparative perspective, the implications for surfactant performance and respiratory dynamics should be interpreted with caution.
Reviewer #3 (Public review):
In this manuscript, the authors present data on the supposed composition of pulmonary surfactant obtained from bronchoalveolar lavages (BALs) of a small cohort of dolphins, a group of them suffering from pneumonia. The lipid compositional differences of the sample group are consistent with the different pathological situations of the specimens, suggesting that differences in surfactant composition are somehow associated (as a cause or as a consequence) with the particular pathophysiological contexts. It is particularly remarkable that an increase in cardiolipins and plasmalogens appears as an abnormal composition in pathological surfactants. The study is completed by analyzing the differences in membrane properties (order, packing, phase) of abnormal versus "control" membranes, concluding that pneumonia in dolphins is associated with a significant alteration of surfactant membranes that become more rigid, packed and thicker than those in surfactant from animals with no lung disease.
In general terms, the data provided are of interest as they somehow offer a framework of effects that may extend what is known about alterations of composition, biophysical properties and functional performance of pulmonary surfactant as a consequence of respiratory pathologies. A collection of pertinent biophysical methodologies (fluorescence, X-ray scattering, AFM) have been applied to complete a full characterization of membrane properties in the different samples.
However, they way the samples have been processed, i.e. by making organic extracts of hydrophobic (lipid and protein) components before surfactant membranes have been purified or at least, separated from bulk lavage, open the question of how much of the altered composition is actually occurring in surfactant or comes from other membranes (from cells, bacteria) that have been completely intermixed as a consequence of the organic extraction. Without an appropriate surfactant membrane obtention, the results of the study should be taken with caution and await confirmation. Specific questions that need to be considered include:
(1) As said, the direct organic extract of BAL samples ends in a full mix of lipid and protein components that in origin could be part of different membranes, either from different surfactant assemblies, or even from pulmonary cells or membrane debris, or microorganisms, collected within the lavage. Obtaining conclusions about the structure and properties of membranes artefactually reconstituted from such lipid and protein mixtures is far from correct.
It is mentioned that "subsequentially" to the organic extraction, the samples were subjected to ultracentrifugation to separate debris and membrane cells. I do not see what the ultracentrifugation is going to change if it is done after the organic extraction. It should have been done before the extraction, for the organic solvents to solubilize exclusively the large, and relatively light, surfactant membrane complexes.
On the other hand, the ulterior reconstitution of the obtained full lipid mixture surely ends in membrane assemblies whose compositional distribution and organization may differ significantly from those in the original membranes.
Taking all this into account, statements such as "These aggregate forms reproduce the expected membrane microstructures observed in native alveolar hypophase" or "pulmonary membranes can be successfully extracted and reconstituted from BALs of Navy dolphins" are simply not true and should be rephrased.
One can understand that the limitation of material may make it difficult to obtain first the purified surfactant membranes and then their organic extract. However, the limitation should be acknowledged to make the readers clear that the actual compositional effects caused in surfactant by pneumonia need confirmation.
(2) In some of the experiments, i.e. in the AFM characterization, supported membranes were prepared by the spray-dry method applied to organic solutions. Again, the spray-dry of organic lipid solutions ends in a lipid dispersion that may be very far from the real organization of the lipids in actual surfactant membranes.
(3) When stated that phospholipid concentrations are greater in BAL from pinnipeds than in humans, how has the actual concentration been determined? BAL volumes are typically subjected to large variations depending on the conditions used to obtain the lavage (including volume of saline instilled, level of atelectasia in the lung tissue, presence of inflammation and edema, etc). If total amounts of phospholipids in BAL are to be compared, certain normalization procedures should be applied, such as for instance, with respect to the urea concentration in serum.
(4) All the differences regarding membrane phase and lipid order/packing have been interpreted in terms of the potential coexistence of Lbeta (gel)/Lalpha (liquid crystalline) phases. However, it has been well established that in lipid systems containing cholesterol, such as pulmonary surfactant, phase coexistence can actually be of the type liquid-ordered (Lo)/liquid-disordered (Ld), very different in terms of mobility and true molecular order. Why do the authors consider that Lbeta is the phase observed in the surfactant membranes they have reconstituted? The presence of round-shaped domains seems to indicate that a liquid/liquid phase segregation is actually occurring.
(5) In the same line as the previous comment, the authors state that SAXS shows that bovine-extracted pulmonary membranes exhibit a coexistence of two lamellar phases, one rich in unsaturated lipids and one in saturated lipids. SAXS and WAXS cannot provide compositional information, but structural parameters such as membrane thickness, or molecular order. This should be clarified.
(6) It is mentioned that the surfactant monolayer at the air-liquid interface is interconnected to tubular membranous structures (tubular myelin, TM). It is true that TM, when present, appears interconnected with the interface. However, it is widely recognized that there are many other structures connected with the interfacial film, including multilamellar membrane arrays or reservoirs that have not been mentioned here. Furthermore, TM is not required for surfactant function, because it is absent, for instance, in mice lacking expression of surfactant protein SP-A, which can breathe perfectly.
(7) In the Discussion, the authors mention that "...after squeeze-out, the excluded multilayers remain closely associated with the interfacial monolayer rather than escaping into the subphase". The authors may like to complete this discussion by specifying that the stable association of excluded assemblies with the interfacial film is actually possible thanks to the surfactant proteins.
Reviewer #2 (Public review):
Summary:
The goal of the experiment was to identify the fMRI neural correlates of persistence and recovery of forgotten memories. A forgotten memory was defined behaviorally as successful learning, followed by failure in a recall format task, followed by next-day success in a recognition format task. The comparison is to memories that were not forgotten at any stage of the task. Various univariate, connectivity, and multivariate analyses were used to identify neural correlates of forgotten memories that were recovered, that remained forgotten, and successful memory. Some claims are made about how activity of the "episodic memory network" predicts the persistence of forgotten memories.
Strengths:
Studies on the persistence of forgotten memories in rodent models have been used to make some novel claims about the potential properties of engrams. Attempting similar research in humans is a laudable goal.
Patterns of behavioral responses are consistent across subjects.
Weaknesses:
I do not find that the fMRI results fit the narrative provided.
A major issue is that primary results do not replicate across the two fMRI datasets that were collected using the same task. For example, hippocampal activity associated with correct responses (confident and guess) was identified in the group receiving the fMRI scan that used a small FOV, but not in the group that received an fMRI scan of the whole brain, for both 30-min and 24-hr delays (lines 202-217). This suggests that the main findings are not even replicable internally within the same experiment. There is no reasonable justification for this.
Next, most of the reported fMRI findings do not meet reasonable thresholds for statistical significance. In many places, the authors acknowledge this in the text by saying that a difference in the fMRI metric "tended towards significant correlation" or that comparisons "revealed non-significant mean value comparisons". It is not clear why these non-significant findings are interpreted as though they are positive findings. Beyond that, many of the reported findings are not meeting the threshold (i.e., p=0.058), without any acknowledgement that they are marginal. Beyond that, the majority of comparisons that are interpreted in the main text are not significant based on the companion information provided in the supplementary tables. That is, they are totally non-significant when using FWE or FDR correction at either the cluster or peak levels.
Beyond this, the supplementary tables indicate that "clusters identified solely within white matter regions have been excluded." The fact that there are any findings in white matter to ignore indicates that the statistical thresholds are inappropriate. It's tantamount to seeing activation in the brain of a dead fish.
The overall picture based on these factors is that the statistical tests did not use sufficiently stringent safeguards against false positives given the multiple comparison problem that plagues fMRI. So, there are tons of false positives, which are being selectively interpreted to tell a particular story. That is, each comparison yields lots of findings in many brain area, and those that do not fit the particular narrative are being ignored (including those in white matter). What's more, when the small FOV fMRI scan is done, the imaging volume is centered on the hippocampus and its close network, so all false positives appear to be exactly in those brain regions about which the authors want to make conclusions. When throwing darts, you will always hit a bullseye if that is all that exists. The fact that the same comparisons done in the companion whole-brain dataset do not yield the same results is telling: the analysis plan is not sufficiently rigorous to yield findings that are replicable.
Further, I think that it is highly debatable whether the task measures the recovery of forgotten memories at all. Forgotten memories are defined as those that fail when tested using a recollection format but succeed when tested using a recognition format. The well-characterized distinction between recollection and recognition is thus being construed as telling us something about the fate of engrams. I think the much more likely alternative is that "forgotten" memories are just relatively weak memories that don't meet whatever criteria subjects typically use when making recollection judgments, and not some special category of memory. In terms of brain activation, they seem for the most part to follow the pattern of stronger memory, but weaker.
Finally, many hypotheses are used as though they are proven. For instance, fMRI activity patterns are called "engrams" even though there are no tests to determine whether they meet reasonable criteria that have been adopted in the engram literature (e.g., necessity, sufficiency). Whatever happens over the 24-hour delay is called "consolidation" even if there is no test that consolidation has occurred. Etc. It becomes hard to differentiate what is an assumption, versus a hypothesis, versus an inference/conclusion.
Reviewer #1 (Public review):
Summary:
This study extends the short-term synaptic plasticity (STP)-based theory of activity-silent working memory (WM) by introducing a physiological mechanism for chunking that relies on synaptic augmentation (SA) and specialized chunking clusters. The model consists of a recurrent neural network comprising excitatory clusters representing individual items and a global inhibitory pool. The self-connections within each cluster dynamically evolve through the combined effects of STP and SA. When a chunking cue, such as a brief pause in a stimulus sequence, is presented, the chunking cluster transiently suppresses the activity of the item clusters, enabling the grouped items to be maintained as a coherent unit and subsequently reactivated in sequence. This mechanism allows the network to enhance its effective memory capacity without exceeding the number of simultaneously active clusters, which defines the basic capacity. They further derive a new upper limit of WM capacity, the new magic number. When the basic capacity is four, the upper bound for complete recall becomes eight, and the optimal hierarchical structure corresponds to a binary tree of two-item pairs forming four chunks that combine into two meta-chunks. Reanalysis of linguistic data and single-neuron recordings from human epilepsy patients (identifying boundary neurons) provides qualitative support for the model's predictions.
Strengths:
This study makes an important contribution to theoretical and computational neuroscience by proposing a physiologically grounded mechanism for chunking based on STP and SA. By embedding these processes in a recurrent neural network, the authors provide a unified account of how chunks can be formed, maintained, and sequentially retrieved through local circuit dynamics, rather than through top-down cognitive strategies. The work is conceptually original, analytically rigorous, and clearly presented, deriving a simple yet powerful capacity law that extends the classical magic number framework from four to eight items under hierarchical chunking. The modeling results are further supported by preliminary empirical evidence from linguistic data and single-neuron recordings in the human medial temporal lobe, lending credibility to the proposed mechanism. Overall, this is a well-designed and well-written study that offers novel insights into the neural basis of working-memory capacity and establishes a solid bridge between theoretical modeling and experimental findings.
Weaknesses:
This study is conceptually strong and provides an elegant theoretical framework, but several aspects limit its biological and empirical grounding.
First, the control mechanism that triggers and suppresses chunking clusters remains only schematically defined. The model assumes that chunking events are initiated by pauses, prosodic cues, or internal control signals, but does not specify the underlying neural circuits (e.g., prefrontal-basal ganglia loops) that could mediate this gating in the brain. Clarifying where, when, and how the chunking clusters are turned on and off will be critical for establishing biological plausibility.
Second, the network representation is simplified: item clusters are treated as non-overlapping and homogeneous, whereas real cortical circuits exhibit overlapping representations, distinct excitatory/inhibitory populations, and multiscale local and long-range connectivity. It remains unclear how robust the proposed dynamics and derived capacity limit would be under such biologically realistic conditions.
Third, the model heavily relies on SA operating over a timescale of several seconds, yet in vivo, the time constants and prevalence of SA can vary widely across cortical regions and neuromodulatory states. The stability of the predicted "new magic number" under realistic noise levels and modulatory influences, therefore, needs to be systematically evaluated.
Reviewer #2 (Public review):
Summary:
This work extends a previous recurrent neural network model of activity-silent working memory to account for well-established findings from psychology and neuroscience suggesting that working memory capacity constraints can be partially overcome when stimuli can be organized into chunks. This is accomplished via the introduction of specialized chunking clusters of neurons to the original model. When these chunking clusters are activated by a cue (such as a longer delay between stimuli), they rapidly suppress recently active stimulus clusters. This makes these stimulus clusters available for later retrieval via a synaptic augmentation mechanism, thereby expanding the network's overall effective capacity. Furthermore, these chunking clusters can be arranged in a hierarchical fashion, where chunking clusters are themselves chunked by higher-level chunking clusters, further expanding the network's overall effective capacity to a new "magic number", 2^{C-1} (where C is the basic capacity without chunking). In addition to illustrating the basic dynamics of the model with detailed simulations (Figures 1 and 2), the paper also utilizes qualitative predictions from the model to (re-)analyze data collected in previous experiments, including single-unit recordings from human medial temporal lobe as well as behavioral findings from a classic study of human memory.
Strengths:
The writing and figures are very clear, and the general topic is relevant to a broad interdisciplinary audience. The work is strongly theory-driven, but also makes some effort to engage with existing data from two empirical studies. The basic results showcasing how chunking can be achieved in an activity-silent working memory model via suppression and synaptic augmentation dynamics are interesting. Furthermore, we agree with the authors that the derivation of their new "magic number" is relatively general and could apply to other models, so those findings in particular may be of interest even to researchers using different modeling frameworks.
Weaknesses:
(1) Very important aspects of the model are assumed / hard-coded, raising the concern that it relies too much on an external controller, and that it would therefore be difficult to implement the same principles in a fully behaving model responsible for producing its own outputs from a sequence of stimuli (i.e., without a priori knowledge of the structure of incoming sequences).
(i) One such aspect is the use of external chunking cues provided to the model at critical times to activate the chunking clusters. The simulations reported in the paper were conducted in a setting where signals to chunk are conveniently indicated by longer delays between stimuli. In this case, it is not difficult to imagine how an external component could detect the presence of such a delay and activate a chunking cluster in response. However, in order for the model to be more broadly applicable to different memory tasks that elicit chunking-related phenomena, a more general-purpose detector would be required (see further comments below and alternative models).
(ii) Relatedly, and as the authors acknowledge in the discussion, the network relies on a pretty sophisticated external controller that decides when the individual chunking clusters are activated or deactivated during readout/retrieval. This seems especially complex in the hierarchical case. How might a network decide which chunking/meta-chunking clusters are activated/deactivated in which order? This was hard-coded in their simulations, but we imagine that it would be difficult to implement a general solution to this problem, especially in cases where there is ambiguity about which stimuli should be chunked, or where the structure of the incoming sequence is not known in advance.
(iii) One of the central mechanisms of the model is the rapid synaptic plasticity in the inhibitory connections responsible for binding chunking clusters to their corresponding stimulus clusters. This mechanism again appears to have been hard-coded in the main simulations. Although we appreciate that the authors worked on one possible way that this could be implemented (Methods section D, Supplementary Figure S2), in the end, their solution seems to rely on precisely fine-tuning the timing with which stimuli are presented - a factor that seems unlikely to matter very much in humans/animals. This stands in contrast with models of working memory that rely on persistent activity, which are more robust to changes in timing. Note that we do not discount the possibility of activity-silent WM, and indeed it should be studied in its own right, but it is then even more important to highlight which of its features are dependent on the time constants, etc.
(2) Another key shortcoming of this work is its limited direct engagement with empirical evidence and alternative computational accounts of chunking in WM. Although the efforts to re-analyze existing empirical results in light of the new predictions made by the model are commendable, in the end, we think they fall short of being convincing. As noted above, the model doesn't actually perform the same two tasks used in the human experiments, so direct quantitative comparisons between the model and human behavior or neural data are not possible. Instead, the authors rely on isolating two qualitative predictions of the model - the "dip" and "ramp" phenomena observed after a chunking cluster is activated (Figure 3), and the new magic number for effective capacity derived from the model in the case where stimuli are chunkable, which approximately converges with human recall performance in a memory study (Figure 4). Below, we highlight some specific issues related to these two sets of analyses, but the larger point is that if the model is making a commitment about how these neural mechanisms relate to behavioral phenomena, it would be important to test if the model can produce the behavioral patterns of data in experimental paradigms that have been extensively used to characterize those phenomena. For example, modern paradigms characterizing capacity limits have been more careful to isolate the contributions of WM per se (whereas the original magic number 7 is now thought to reflect a combination of episodic and working memory; see Cowan 2010). There are several existing models that more directly engage with this literature (e.g., Edin et al., 2009; Matthey et al., 2015; Nassar et al., 2018; Soni & Frank, 2025; Swan & Wyble, 2014; van den Berg et al., 2014; Wei et al., 2012), some of which also account for chunking-related phenomena (e.g., Wei et al, 2012; Nassar et al., 2018; Panichello et al., 2019; Soni & Frank, 2025). A number of related proposals suggest that WM capacity limits emerge from fundamentally different mechanisms than the one considered here - for example, content-related interference (Bays, 2014; Ma et al., 2014; Schurgin et al., 2020), or limitations in the number of content-independent pointers that can be deployed at a given time (Awh & Vogel, 2025), and/or the inherent difficulty of learning this binding problem (Soni & Frank, 2025). We think it would be worth discussing how these ideas could be considered complementary or alternatives to the ones presented here.
(i) Single unit recordings. We found it odd that the authors chose to focus on evidence from single-unit recordings in the medial temporal lobe from a study focused on episodic memory. It was unclear how exactly these data are supposed to relate to their proposal. Is the suggestion that a mechanism similar to the boundary neurons might be operative in the case of working memory over shorter timescales in WM-related areas such as the prefrontal cortex, or that their chunking mechanism may relate not only to working memory but also to episodic memory in the medial temporal lobe?
(ii) N-gram memory experiment. Our main complaint about the analysis of the behavioral data from the human memory study (Figure 4) is that the model clearly does not account for the main effect observed in that study - namely, the better recall observed for higher-order n-gram approximations to English. We acknowledge that this was perhaps not the main point of the analysis (which related more to the prediction about the absolute capacity limit M*), but it relates to a more general criticism that the model cannot account for chunking behavior associated with statistical learning or semantic similarity. Most of the examples used in the introduction and discussion are of this kind (e.g., expressions such as "Oh my God" or "Easier said than done", etc.). However, the chunking mechanism of the model should not have any preference for segmenting based on statistical regularities or semantic similarity - it should work just as well if statistical anomalies or semantic dissimilarity were used as external chunking cues. In our view, these kinds of effects are likely to relate to the brain's use of distributed representations that can capture semantic similarity and learn statistical regularities in the environment. Although these kinds of effects may be beyond the scope of this model, some effort could be made to highlight this in the discussion. But again, more generally, the paper would be more compelling if the model were challenged to simulate more modern experimental paradigms aimed at testing the nature of capacity limits in WM, or chunking, etc.
(iii) There are a number of other empirical phenomena that we're not sure the model can explain. In particular, one of the hallmarks of WM capacity limits is that it suffers from a recency bias, where people are more likely to remember the most recent items at the expense of items presented prior to that (Oberauer et al 2012). [There are also studies showing primacy effects in addition to recency effects, but the primacy effects are generally attributed to episodic rather than working memory - for example, introducing a distractor task abolishes the recency but not primacy effect]. But the current model seems to make the opposite prediction: when the stimuli exceed its base capacity, it appears to forget the most recent stimuli rather than the earliest ones (Figure 1d). This seems to result from the number of representations that can be reactivated within a cycle and thus seems inherent to the dynamics of the model, but the authors can clarify if, instead, it depends on the particular values of certain parameters. (In contrast, this recency effect is captured in other models with chunking capabilities based on attractive dynamics and/or gating mechanisms - eg Boboeva et al 2023; Soni & Frank (2025)). Relatedly, we're not sure if the model could account for the more recent finding that recall is specifically enhanced when chunks occur in early serial positions compared to later ones (Thalmann, Souza, Oberauer, 2019).
Reviewer #3 (Public review):
The paper presents a synaptic mechanism for chunking in working memory, extending previous work of the last author by introducing specialized "chunking clusters", neural populations that can dynamically segment incoming items into chunks. The idea is that this enables hierarchical representations that increase the effective capacity of working memory. They also derive a theoretical bound for working memory capacity based on this idea, suggesting that hierarchical chunking expands the number of retrievable items beyond the basic WM capacity. Finally, they present neural and behavioral data related to their hypothesis.
Strengths
A major strength of the paper is its clear theoretical ambition of developing a mechanistic model of working memory chunking.
Weaknesses
Despite the inspiration in biophysical mechanisms (short-term synaptic plasticity with different time constants), the model is "cartoonish". It is unclear whether the proposed mechanism would work reliably in the presence of noise and non-zero background activity or in a more realistic implementation (e.g., a spiking network).
As far as I know, there is no evidence for cyclic neural activation patterns, which are supposed to limit WM capacity (such as in Figure 1d). In fact, I believe there is no evidence for population bursts in WM, which are a crucial ingredient of the model. For example, Panicello et al. 2024 have found evidence for periods during which working memory decoding accuracy decreases, but no population bursts were observed in their data. In brief, my critique is that including some biophysical mechanism in an abstract model does not make the model plausible per se.
It is claimed that "our proposed chunking mechanism applies to both the persistent-activity and periodic-activity regimes, with chunking clusters serving the same function in each", but this is not shown. If the results and model predictions are the same, irrespective of whether WM is activity-silent or persistent, I suggest highlighting this more and including the corresponding simulations.
The empirical validations of the model are weak. The single-unit analysis is purely descriptive, without any statistical quantification of the apparent dip-ramp pattern. I agree that the dip-ramp pattern may be consistent with the proposed model, but I don't believe that this pattern is a specific prediction of the proposed model. It seems just to be an interesting observation that may be compatible with several network mechanisms involving some inhibition and a rebound.
Moreover, the reanalyses of n-gram behavioral data do not constitute a mechanistic test of the model. The "new magic number" depends strongly on structural assumptions about how chunking operates, and it is unclear whether human working memory uses the specific hierarchical scheme required to achieve the predicted limit.
The presentation of the modeling results is highly compressed in two figures and is rather hard to follow. Plotting the activity of different neural clusters in separate subplots or as heatmaps (x-axis time, y-axis neural population, color = firing rate) would help to clarify (Figure 1d). Also, control signals that activate the chunking clusters should be shown.
Overall, the theoretical proposal is interesting, but its empirical grounding and biological plausibility need to be substantially reinforced.
true, but free will is entirely different. these are examples of conditions a person can have, not everyday consciousness.
author is arguing that rather than free will, the child is being controlled by biology and environment in numerous ways, and each different way is why they make the decision they do. but then what do we say about critical thinking and decision making? its not like if the child's brain is unable to concentrate due to hunger they dont brainstorm other decisions at all. they likely did, and they chose based on free will, no?
interesting point, but I wonder if this has a different meaning, not that we can automatically interpret this capability as free will being fake
Sakal
🧠 VOCABULARY UPLOAD: Sakal (Hebrew): Often translated as "success," but more accurately means "prudent, circumspect, wise insight." It is the opposite of "Hustle." Hustle is blind speed. Sakal is looking at the map, understanding the terrain, and taking the right step, not just the fast one.
🎉 SIGNAL ACQUIRED: You Found It.
Congratulations, Agent.
Most people scan the headlines and move on. They stay on the surface. But you stopped. You questioned the text. You looked deeper.
You have just passed the first test of an Insider.
You have discovered the "Tactical Overlay" (the Kingdomlens). This is where the real work happens. While the main text gives you the map, these notes give you the compass—the science, the scripture, and the deeper conversation.
SYSTEM NOTE (TRANSPARENCY): This overlay is powered by Hypothesis, an open-source non-profit organization dedicated to open conversation.
This is a third-party tool, distinct from the Dream Centre.
To join the conversation, you will need to create a free account directly with them.
**YOUR NEXT MOVE: ** Initialize: Click "Sign Up" above to create your secure ID with Hypothesis.
Radio Check: Once logged in, click the Reply Arrow ↩️ on this note and type a simple confirmation:
"I'm in."
Welcome to the team. Let's dig in.
frequency by 60%
Percentages are great since it's a method to hide the lack of datapoints, but if you have bigger numbers they can be supported using the more explicit math (e.g. Frequency = n/t, where n = number, t =duration), it's even better. Not sure what your numbers are that you've done to calculate 60%, but if it's something like 10 -> 16 publication a week, then that's probably less interesting than something like 1,000,000 -> 1,600,000. Here, I'd use 60% if it's a smaller number like 16, but definitely would use 1.6m if that was a true number.
100%
Let's drop the bold, numbers usually scream themselves. I know my last recommendation was to bold numbers only, but that's for numbers within numbers scenario, like spreadsheets.
“It’s not that different than looking at the printing press, and the evolution of the book,” he said. After Gutenberg, the printing press was mostly used to mimic the calligraphy in bibles. It took nearly 100 years of technical and conceptual improvements to invent the modern book. “There was this entire period where they had the new technology of printing, but they were just using it to emulate the old media.”
biobanks
EN - Meaning: Collections that store biological samples (like blood, tissue) and associated data for research. - Key idea: Biobanks support medical research but require strong policies for consent, privacy, and data security.
中文 - 含义:生物样本库:保存血液、组织等样本及相关数据,用于科研的收集与存储系统。 - 关键点:有助于医学研究,但需要完善的知情同意、隐私与数据安全制度。
Images - https://en.wikipedia.org/wiki/Special:Search?search=biobank - https://commons.wikimedia.org/wiki/Special:MediaSearch?type=image&search=biobank
herbicide-resistant plants
EN - Meaning: Crop plants engineered to survive a herbicide that would normally kill them. - Key idea: Enables weed control by spraying, but can lead to selection for herbicide-resistant weeds.
中文 - 含义:耐除草剂作物:被改造后能耐受本应杀死它们的除草剂。 - 关键点:便于喷洒除草剂控草,但可能促使杂草选择出耐药性。
Images - https://en.wikipedia.org/wiki/Special:Search?search=herbicide-resistant%20crop - https://commons.wikimedia.org/wiki/Special:MediaSearch?type=image&search=herbicide-resistant%20crop
BT corn
EN - Meaning: Corn genetically engineered to produce Bt toxin (from Bacillus thuringiensis) that kills certain insect pests. - Key idea: Can reduce insect damage, but may raise concerns about resistance and ecological effects.
中文 - 含义:Bt 玉米:转基因玉米,可产生 Bt 毒素(来源于苏云金芽孢杆菌),用来杀死特定害虫。 - 关键点:可减少虫害,但也可能引发抗性与生态影响等问题。
Images - https://en.wikipedia.org/wiki/Special:Search?search=Bt%20corn - https://commons.wikimedia.org/wiki/Special:MediaSearch?type=image&search=Bt%20corn
GMOs
EN - Meaning: Genetically modified organisms; organisms whose DNA has been altered using biotechnology. - Key idea: GMOs can be used in medicine and agriculture, but they can also raise environmental and health concerns.
中文 - 含义:GMO(基因改造生物):通过生物技术改变过 DNA 的生物体。 - 关键点:在医药与农业中有应用,但也可能引发环境与健康方面的担忧。
Images - https://en.wikipedia.org/wiki/Special:Search?search=genetically%20modified%20organism - https://commons.wikimedia.org/wiki/Special:MediaSearch?type=image&search=genetically%20modified%20organism
In June 2015, a white supremacist walked into a black church in Charleston, South Carolina, during a prayer meeting and shot and killed nine African Americans congregants, including the pastor.7Tessa Berenson, “Everything We Know about the Charleston Shooting,” Time, June 18, 2015, accessed August 1, 2015, http://time.com/3926112/charleston-shooting-latest jQuery("#footnote_plugin_tooltip_2620_7").tooltip({ tip: "#footnote_plugin_tooltip_text_2620_7", tipClass: "footnote_tooltip", effect: "fade", fadeOutSpeed: 100, predelay: 400, position: "top right", relative: true, offset: [10, 10] });
It has and will always baffle me that people can develop so much hate in their heart to not only take lives of innocents they've never met but also throw away their own life in the process deluded by the idea that they're doing the world some good in the form of necessary evil.
I want to make the writing as dissonant as I can, to recreate a sense of disruption, of an essential brokenness. I want to make the writing as unobtrusive as I can, to have it slip easily into the mind, mild-mannered and unassuming, before revealing that it has been wearing a vest of explosives all along.
I was going to make this comment at the end of the article so it is surprising to see the author mention it here. While I understand that the author's intention is to portray the brutality, sadness and trauma that come with the experiences they are recounting, as a reader this invokes a sense of wanting to help or knowing that there is a way out for these people or literally anything that can offer comfort, but there is not a shred of positivity in this article. Yes, the people are portrayed as strong resilient individuals but the article never states how they manage to cope or overcome the experience they have been through so neither my urge to help is fulfilled nor am I comforted by seeing a light at the end of this tunnel. This is not a critique but just an opinion as to what I would have wished to see up until this point.
“But Miss,” he replied, incredulous. “I’m not writing about war and bombs and tragedy. Why would I give them such names?”
It is heartbreaking to hear that anyone's entire world image could be consumed like this.
Afterward, trying to pick them up and separate them out, I am left with a thousand cuts I can feel every time I move or breathe. Afterward I realize that there is a shard I have failed to remove, that it has entered my eyes and become lodged there, cutting into my vision always, digging into the form and content of my memories.
What a unique and interesting experience. Of course this does come with its pains as is stated but I think there's more to it than that. The fact that a person feels this way is proof that they are invested enough to be worthy of translating a story but it is also their responsibility to make sure as little of themselves as possible is reflected in that translation as it could mutate the original experience if done carelessly.
To translate a text is to enter into the most intimate relationship with it possible. It is the translator’s body, almost more so than the translator’s mind, that is the vessel of transfer. The mind equates words, expressions, deals with techniques and logistics; it is within the body that the real alchemy—mysterious, unnamed and inexplicable—takes place. That alchemy has to do with truth more than signification, that is, the animating force behind signification, which transforms it into meaning, into something that moves. Gayarti Spivak qualifies the act of translation as “erotic,” but there is something too gentle about that word to ring true for me. The word captures the act of surrender, and the abundantly physical communion with the text, but there is something messier and bloodier that is elided. More agonized and agonizing too. There is a violence in undoing someone’s words and reconstituting them in a vocabulary foreign to them, a vocabulary of your own choosing. There is a violence, too, in the way you are—for long moments—annihilated by the other; undone in return. Neither the translator nor the text emerges from the act unscathed.
I find this to be incredibly interesting because my grandpa translates books! His name is Refaat Elsayed Ali, you might've read one of his translations if you're really into arabic literature but I never really thought about it like this. I feel like I have gained a greater understanding of how he feels when he's working
From “to witness,” we get shahed, the one who witnesses; mashhad, the spectacle or the scene, but also shaheed, martyr; istishhad, to be martyred, to die for a cause.
I somehow never drew this connection even though these are relatively common words that I have used many times before. It is very interesting how a word can carry such a deep double meaning. That is assuming that they both have the same origin of sh-h-d in our dictionary, if they have different origins then I'm afraid this might just be a coincidence if I were to act as the grammar police but it is interesting nonetheless,
I have buried seven husbands, three fiancés, fifteen sons and a two-week old daughter I finally agreed to have at 42 for my husband’s sake
I find it both surprising and inexplicable that someone could be capable of finding love (or the time for love) under such extreme circumstances so the sheer amount of partners listed is beyond shocking. Even if these marriages weren't out of love I cannot imagine still being able to take partners after the dirst few tragedies. Does your heart not close off at some point? Do you not start to fear that kind of connection? And same applies to the children. I pray nobody ever has to go through the pain of losing a child but at some point surely you feel some form of aversion to bringing children into this evil world that's already taken so much from you.
Cortical bone damage
Somewhat unrelated to the purpose of this paper, but still relevant to the course - I wonder how trabecular bone would differ in damage in postmortem processes. Though cortical bone is typically on the outside of the bone, and thus more 'exposed' to taphonomic processes, trabecular bone is far more porous, which might be expected to preserve poorly. If anyone knows the answer to this, let me know! If not, hopefully we discuss it in class.
It bears mentioning that an expert/professional repair person can only tell very little of the condition of a typewriter by photos. Does it look generally clean? Are the decals in tact? Does the segment look clean (a vague proxy for the potential condition of the internals)? Is anything obvious missing (knobs, return lever, keys)? Does it look cared for or has it been neglected in a barn for half a century? Most modern typewriters made after 1930 in unknown condition are worth about $5-25 and they peak at about $500 when purchased from a solid repair shop unless some Herculean additional restoration has taken place, they've got a rarer typeface, or are inherently actually rare. Hint: unless it's a pro repair shop or very high end collector with lots of experience, don't trust anyone saying that a typewriter is "rare", run the other direction. Run faster if they say it "works, but just needs a new ribbon" as—even at the most expensive—new ribbon is only $15 and their "rare" $600+ machine should have fresh, wet ribbon. The rule of thumb I use is that no one online selling a typewriter knows anything about it, including if it actually works. Worse, they've probably priced it at professional repair shop prices because they don't know that condition is king.
The least experienced typist will know far more about the condition of a machine by putting their hands on it and trying it out. Does it generally work? Does the carriage move the full length of its travel? Can you set the margins at the extremes and space reliably from one end to another? Does it skip? Is the inside clean or full of decades of dried oil, dust, and eraser crumbs? Does the margin release work? Does it backspace properly? If typing HHHhhhHHH are the letters all printed well and on the same baseline?
Presumably a typewriter at an antique store will meet these minimum conditions (though be aware that many don't as their proprietors have no idea about typewriters other than that if they wait long enough, some sucker will spend $150 on almost anything). They've done the work of finding a machine that (barely) works, housing it, and presenting it to the public for sale. This time and effort is worth something to the beginning typewriter enthusiast, but worth much, much less to the longer term practiced collector.
If everything is present and at least generally limping along, you've got yourself a $30 typewriter. Most people can spend a few hours watching YouTube videos and then manage to clean and lubricate a typewriter to get it functioning reasonably. You can always learn to do the adjustments from Youtube videos. (Or just take it to a repair shop and fork over $200-400 to get things squared.)
If you're getting into collecting, you'll make some useful mistakes by overpaying in the beginning and those mistakes will teach you a lot.
Maybe you're a tinkerer and looking for a project? If so, then find the cheapest machine you can get your hands on (maybe a Royal KMM for $9 at thrift) and work your way through a home study course.
Otherwise, if you're just buying one or two machines to use—by far—the best value you'll find is to purchase a cleaned, oiled, and well-adjusted machine from a repair shop. Sure it might cost $350-600, but what you'll save in time, effort, heartache, repair, etc. will more than outweigh the difference. Additionally you'll have a range of machines to choose from aesthetically and you can test out their feel to find something that works best for you.
Or, you could buy a reasonable machine like this for $70 and find out it needs cleaning, oiling, and adjusting and potentially a few repairs. The repair tab might run you an additional $450. Is it worth it when a repair shop would have sold you the same or a very similar machine in excellent condition for $350?
Remember in asking about the cost and value of a typewriter, you're actually attempting to maximize a wide variety of variables including, but not limited to: upfront money, information about the current state of the market, information/knowledge about the machine itself, information about how to clean it, information about oiling it, information about adjusting, information about repairing it, cost and availability of tools and repair parts, and the time involved for both learning and doing all of these. The more time you've spent learning and doing all of these, the better "deals" you'll find, but gaining this expertise is going to cost you a few years of life. What is all this "worth" when you just want to type on a machine that actually works?
Most of the prognostication you'll find in fora like this will be generally useless to you because you're not readily aware of the context and background of the respondents with respect to all of the variables above. Similarly they're working with no context about you, your situation, where you live, what's available in your area, your level of typewriter knowledge, or your budget. You don't know what you don't know. At the end of the day, you're assuredly just as well off to use a bit of your intuition and putting your hands on a machine and trying it out. Then ask: "What is it worth to you?"
If you're simply asking: "Is this highway robbery?", the answer is no.
More resources (and some of my own context) if you need them: https://boffosocko.com/research/typewriter-collection/
Happy typing.
Reply to u/NeverTheNess at https://reddit.com/r/typewriters/comments/1q7eho6/spotted_a_royal_at_an_antique_store_good/<br /> RE: run-of-the-mill late 70s plastic Litton/Royal typewriter
positive thing
This is obviously a great outcome, but it can be expected that this will not always be the case. Perhaps the close collaboration between the University and Huron-Wendat in this process contributed to this outcome. Such a precedent should be followed in future research to ensure the interests of both groups, though specifically of Indigenous groups, can be met.
safety constraints work by reducing the model's generative capacity, constraining outputs that are considered risky, controversial, or potentially harmful. This reduction necessarily decreases entropy in the information-theoretic sense, narrowing the range of possible responses the model can generate. What safety optimises for is not maximum (or more) information but maximum predictability, steering the model away from novel or unexpected outputs toward safer, more conventional patterns.
The distal convoluted tubule will recover another 10–15 percent before the filtrate enters the collecting ducts
I agree with calling out the specific percentage here, but I feel that if we are going to do so we should also do it in the PCT and loop discussion.
Characterization in act II, scene 3: Friar: "Holy Saint Francis, what a change is here!" – He's shocked by the speed of the change from Rosaline to Juliet. He doubts Romeo's emotional depth. "Young men's love then lies / Not truly in their hearts, but in their eyes" – He criticizes Romeo for loving based on physical appearance (eyes), not deep feeling. "Jesu Maria, what a deal of brine / Hath wash'd thy sallow cheeks for Rosaline!" – "Brine" = salt tears. He mocks Romeo's exaggerated, performative past grief.
Paradox and key themes in act II, scene 3: "The earth that's nature's mother is her tomb" – Paradox. The earth gives life (womb) and receives death (tomb). Central theme: life and death are intertwined. "For nought so vile that on the earth doth live / But to the earth some special good doth give" – Nothing is so evil it doesn't have some potential good use. "Virtue itself turns vice, being misapplied" – Key theme. Good things (like herbs, or love) can become harmful if used incorrectly. Foreshadows the plan's failure.
Metaphor and apostrophe are used in act II, scene 2: "He jests at scars that never felt a wound." – Metaphor. Mercutio jokes about love's pain because he's never been in love (felt the wound). This shows Romeo's sincere love towards Juliet. "But, soft! what light through yonder window breaks?" – Apostrophe. He speaks to the light as if it's a person. The famous start of the balcony scene.
Prologue (Lines 780-790): "But to his foe supposed he must complain" – Dramatic irony. He must lament to his supposed enemy (a Capulet). Shows the central conflict of loving a an enemy's daughter.
But passion lends them power, time means, to meet Tempering extremities with extreme sweet.
Their love makes them brave enough to find secret times to meet, and the joy of being together makes the terrifying risk seem worth it.
This cannot anger him: 'twould anger him To raise a spirit in his mistress' circle Of some strange nature, letting it there stand Till she had laid it and conjured it down; 825That were some spite: my invocation Is fair and honest, and in his mistress' name I conjure only but to raise up him
Is Mercutio involved with the same girl Romeo is sad about?
O Romeo, Romeo! wherefore art thou Romeo? 880Deny thy father and refuse thy name; Or, if thou wilt not, be but sworn my love, And I'll no longer be a Capulet.
Juliet is expressing her deep love for Romeo. She is willing to give up her family name if she has to. This is showing how devoted and caring she is toward Romeo.
But [it] has expanded drastically after Russia’s full-scale invasion of Ukraine in 2022. That saw what we call the shadow fleet explode to some 900-1,200 vessels globally.
since the 2022 full scale Russian invasion of Ukraine, while the shadow fleet existed before, numbers have risen to 900-1200 ships globally
I am not uninterested in change. I am uninterested in pretending that it is always available, or that it arrives at midnight. So, this is how I have entered 2026. Not with a clean page but with a pen hovering over old ink, writing in the margins where I can. I am not starting again, I am continuing, carefully. Happy New Year. Same old life.
This is true on various levels. The psychological threshold that is a new calendar year was apparent to me as a child. I tell my team now 'if this were April and you had to postpone to May would you think that is problematic? Then why when its mid December is it hard to postpone to early January?
Berlin Senate sees open data as a cause for the (potential hybrid) attack on the Berlin power grid. Frag den Staat team says Senate is trying to wash its hands of resp after decades of under-investing in power infrastructure and its resilience. I see a general trend towards obscuring more data that has been part of open data for a decade or more. Btw the Dutch subsoil cable data is not public, the power networks (like in Germany) are published by the network government owned enterprises. In part this is bc of the HVD regulation. The next round of HVD, and the DS for Energy will likely require this data to be generally available too. Data spaces are a way to deal with access with nuance and granularity but open data is the actual legal default EU wide. So it must be legally explained if something is not open.
Petra de Sutter, rector U Gent had in haar inaugurele rede in september 2025 twee citaten die verzonnen zijn door AI.
Het eerste niet bestaande citaat - Einstein, 1929 in toespraak Sorbonne, "dogma is the enemy of progress" (Einstein did receive a honorary doctorate Dec 1929 at Sorbonne)
Het tweede, niet benoemd - uit "de rectorale rede" Hans Jonas 1979 Uni Munchen, parafrase van wat Rabelais in de 16e schreef. (Jonas never lived in Germany again after the war, but was a visiting prof in Munich 1982-1983 per https://en.wikipedia.org/wiki/Hans_Jonas so the speech never existed.
The 1960s: Bob Dylan—Blowin' in the Wind
Setting the scene: the song was released in 1963 and included in the album The Freewhelin' Bob Dylan. All of Dylan's "most famous political songs were written [...] between January 1962 and October 1963" and "those [...] fixed him in the popular imagination" as a protest songwriter (Lynskey, 2010, 67).
The historical context in which the song was released is essential to understand its meaning: during the 1950s the civil rights movement against racial segregation started to gain momentum, reaching its highest point precisely in 1963. Indeed, in that year:
* Desegregation protests spread throughout the Southern states over more than 100 cities. The most famous was the March on Washington: it was organized by the "Big Six" of the civil rights movement (Martin Luther King Jr., James Farmer, J. Lewis, P. Randolph, R. Wilkins, W. Young), attended by more than 250.000 protesters (including Bob Dylan). It was in this occasion that the renowned "I have a dream" speech took place.

It is important to bear in mind that protests were not only motivated by the insufferable racial segregation, but they were also anti-war oriented: the Vietnam war (1955-1975), which had been raging for ten years by the mid-1960s, was felt as an unnecessary conflict, especially by students and young people, who were the ones recruited in the U.S. army. Indeed, "the average age of an American soldier in Vietnam was 19" (https://www.bbc.co.uk/bitesize/guides/z6dk8hv/revision/4 ).
The only way to actually win is to get better each day
I agree with this because no one is actually good at anything at first, but if they're really passionate about what they're doing, then they'll put in the work to get better and better everyday.
The latter either claims to be legitimate or is believed to be so, and is effective in imposing its will on many over whom it claims authority, perhaps because its claim to legitimacy is recognized by many of its subjects. But it does not necessarily possess legitimacy.
referring to de facto authority
Global cooperation between companies not collapsing but changing shape. Smaller, more agility This is in line w the resilience dimension of [[Networked Agency 20160818213155]]
That’s interesting. I hadn’t really made the connection between those two stories, but I see what you mean. I wouldn’t say that escalation is an essential part of storytelling as a whole, but I think there is a specific mode of science fiction storytelling in which extrapolation is a key part: taking an idea as far as it will go, trying to imagine what comes next, to the limit of your imagination.
Hearing this definition of science fiction makes me want to write science fiction! I love this concept of taking it as far as it will go.
Reviewer #2 (Public review):
Summary:
In this manuscript, Grier, Salimian, and Kaufman characterize the relationship between the activity of neurons in sensorimotor cortex and forelimb kinematics in mice performing a reach-to-grasp task. First, they train animals to reach to two cued targets to retrieve water reward, measure limb motion with high resolution, and characterize the stereotyped kinematics of the shoulder, elbow, wrist, and digits. Next, they find that inactivation of the caudal forelimb motor area severely impairs coordination of the limb and prevents successful performance of the task. They then use calcium imaging to measure the activity of neurons in motor and somatosensory cortex, and demonstrate that fine details of limb kinematics can be decoded with high fidelity from this activity. Finally, they show reach direction (left vs right target) can be decoded earlier in the trial from motor than from somatosensory cortex.
Strengths:
In my opinion, this manuscript is technically outstanding and really sets a new bar for motor systems neurophysiology in the mouse. The writing and figures are clear, and the claims are supported by the data. This study is timely, as there has been a recent trend towards recording large numbers of neurons across the brain in relatively uncontrolled tasks and inferring a widespread but coarse encoding of high-level task variables. The central finding here, that sensorimotor cortical activity reflects fine details of forelimb movement, argues against the resurgent idea of cortical equipotentiality, and in favor of a high degree of specificity in the responses of individual neurons and of the specialization of cortical areas.
Comment on revised version:
The authors addressed all my concerns, and in my opinion, the manuscript is suitable for publication of the Version of Record in its current form.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
Summary:
This study addresses the encoding of forelimb movement parameters using a reach-to-grasp task in mice. The authors use a modified version of the water-reaching paradigm developed by Galinanes and Huber. Two-photon calcium imaging was then performed with GCaMP6f to measure activity across both the contralateral caudal forelimb area (CFA) and the forelimb portion of primary somatosensory cortex (fS1) as mice perform the reaching behavior. Established methods were used to extract the activity of imaged neurons in layer 2/3, including methods for deconvolving the calcium indicator's response function from fluorescence time series. Video-based limb tracking was performed to track the positions of several sites on the forelimb during reaching and extract numerous low-level (joint angle) and high-level (reach direction) parameters. The authors find substantial encoding of parameters for both the proximal and distal parts of the limb across both CFA and fS1, with individual neurons showing heterogeneous parameter encoding. Limb movement can be decoded similarly well from both CFA and fS1, though CFA activity enables decoding of reach direction earlier and for a more extended duration than fS1 activity. Collectively, these results indicate involvement of a broadly distributed sensorimotor region in mouse cortex in determining low-level features of limb movement during reach-to-grasp.
Strengths:
The technical approach is of very high quality. In particular, the decoding methods are well designed and rigorous. The use of partial correlations to distinguish correlation between cortical activity and either proximal or distal limb parameters or either low- or high-level movement parameters was very nice. The limb tracking was also of extremely high quality, and critical here to revealing the richness of distal limb movement during task performance.
The task itself also reflects an important extension of the original work by Galinanes and Huber. The demonstration of a clear, trackable grasp component in a paradigm where mice will perform hundreds of trials per day expands the experimental opportunities for the field. This is an exciting development.
The findings here are important and the support for them is solid. The work represents an important step forward toward understanding the cortical origins of limb control signals. One can imagine numerous extensions of this work to address basic questions that have not been reachable in other model systems.
Collectively, these strengths made this manuscript a pleasure to read and review.
Thank you!
Weaknesses:
In the last section of the results, the authors purport to examine the representation of "higher-level target-related signals," using the decoding of reach direction. While I think the authors are careful in their phrasing here, I think they should be more explicit about what these signals could be reflecting. The "signals" here that are used to decode direction could relate to anything - low-level signals related to limb or postural muscles, or true high-level commands that dictate only what movement downstream motor centers should execute, rather than the muscle commands that dictate how. One could imagine using a partial correlation-type approach again here to extract a signal uncorrelated with all the measured low-level parameters, but there would still be all the unmeasured ones. Again, I think it is still ok to call these "high-level signals," but I think some explicit discussion of what these signals could reflect is necessary.
Thank you for this excellent suggestion. We have followed both pieces of the reviewer’s advice. First, we performed the suggested analysis, partialing off the kinematics then performing target classification on the residuals. This is now Figure 6S1. The analysis revealed the presence of target-related information in the neural activity after subtracting off all linear correlations with kinematics, supporting our claims that higher-level information is present in both populations. The exact timing of classifier performances varied substantially across mice, potentially due to differences in reach-to-grasp strategy, kinematic tracking fidelity, and exact spatial locations of each recorded FOV. Following the second suggestion, we have made the relevant text more careful. We now conclude simply that higher-level signals, meaning those signals that are largely unrelated to forelimb joint angle kinematics, are present but with variable timing and strengths in each area. That text now reads:
“Target decoding performance could result from truly higher-level signals that code abstractly for target location, or alternatively could be supported by strong encoding of kinematic variables that differed between targets. To disambiguate these possibilities, we refit the linear classifier to neural data after regressing off variance related to the joint angle kinematics. The strength and exact time course of the resulting target decoding varied somewhat across animals, but the earliest portion of target decoding performance persisted in all animals after the removal of kinematics and performance remained stronger for M1-fl than S1-fl (Fig. 6S1B). We thus conclude that higher-level signals are present in both areas, but differ in their exact timing and strength. However, we note that other possible signals, such as postural changes, could not be controlled for here.”
Related to this, I think the manuscript in general does not do an adequate job of explicitly raising the important caveats in interpreting parametric correlations in motor system signals, like those raised by Todorov, 2000. The authors do an expert job of handling the correlations, using PCA to extract uncorrelated components and using the partial correlation approach. However, more clarity about the range of possible signal types the recorded activity could reflect seems necessary.
This is an important point, and our text could have unintentionally misled readers. We have now attempted to make this point explicit in the Discussion and in the Results for Figure 6. This Discussion text now reads:
“Moreover, as is widely known (Todorov 2000), the exact role of these kinematically-related signals is challenging to determine from correlative measures alone; thus, determining whether these signals are used for direct movement control or instead indirectly reflect control performed elsewhere is left as a topic for future work.”
The manuscript could also do a better job of clarifying relevant similarities and differences between the rodent and primate systems, especially given the claims about the rodent being a "first-class" system for examining the cellular and circuit basis of motor control, which I certainly agree with. Interspecies similarities and differences could be better addressed both in the Introduction, where results from both rodents and primates are intermixed (second paragraph), and in the Discussion, where more clarity on how results here agree and disagree with those from primates would be helpful. For example, the ratio of corticospinal projections targeting sensory and motor divisions of the spinal cord differs substantially between rodents and primates. As another example, the relatively high physical proximity between the typical neurons in mouse M1 and S1 compared to primates seems likely to yoke their activity together to a greater extent. There is also the relatively large extent of fS1 from which forelimb movements can be elicited through intracortical microstimulation at current levels similar to those for evoking movement from M1. All of these seem relevant in the context of findings that activity in mouse M1 and S1 are similar.
We understand two points to address here. The first point is that we needed to be more careful to attribute previous results as being from the rodent vs. monkey. We agree. We have now revised several parts of the paper to make these distinctions clearer. The second point is about the potential benefit of a thorough review of the many ways in which primate and rodent sensorimotor systems differ. We entirely agree that this could be useful for the field. However, this is a sizable endeavor and doing it full justice is beyond what we know how to fit in the space allotted for framing our results here. We therefore sought a compromise, acknowledging how our results correspond to existing results in the primate without exhaustively accounting for how they differ. Future work will be necessary to more carefully disambiguate whether species-specific differences are due to biomechanical, neurological, ethological, or as-of-yet undetermined sources. We have incorporated your final specific points about what could produce similar information in M1 and S1 into the Discussion.
“This may simply be a consequence of widely distributed representations of movement across mouse cortex (Musall et al. 2019; Steinmetz et al. 2019; Stringer et al. 2019), including forelimb somatosensory areas, or may be a consequence of the close physical proximity of M1-fl and S1-fl hindering development of functionally distinct representations (Tennant et al. 2011).”
In addition, there are a number of other issues related to the interpretation of findings here that are not adequately addressed. These are described in the Recommendations for improvement.
Reviewer #2 (Public review):
Summary:
In this manuscript, Grier, Salimian, and Kaufman characterize the relationship between the activity of neurons in sensorimotor cortex and forelimb kinematics in mice performing a reach-to-grasp task. First, they train animals to reach to two cued targets to retrieve water reward, measure limb motion with high resolution, and characterize the stereotyped kinematics of the shoulder, elbow, wrist, and digits. Next, they find that inactivation of the caudal forelimb motor area severely impairs coordination of the limb and prevents successful performance of the task. They then use calcium imaging to measure the activity of neurons in motor and somatosensory cortex, and demonstrate that fine details of limb kinematics can be decoded with high fidelity from this activity. Finally, they show reach direction (left vs right target) can be decoded earlier in the trial from motor than from somatosensory cortex.
Strengths:
In my opinion, this manuscript is technically outstanding and really sets a new bar for motor systems neurophysiology in the mouse. The writing and figures are clear, and the claims are supported by the data. This study is timely, as there has been a recent trend towards recording large numbers of neurons across the brain in relatively uncontrolled tasks and inferring a widespread but coarse encoding of high-level task variables. The central finding here, that sensorimotor cortical activity reflects fine details of forelimb movement, argues against the resurgent idea of cortical equipotentiality, and in favor of a high degree of specificity in the responses of individual neurons and of the specialization of cortical areas.
Thank you!
Weaknesses:
It would be helpful for the authors to be more explicit about which models of mouse cortical function their results support or rule out, and how their findings break new conceptual ground.
We appreciate this feedback and have attempted to make these details clearer through changes to the Introduction and Discussion. One key change is noted below:
“The presence of detailed kinematic signals in the sensorimotor cortex supports a model of mouse sensorimotor cortex in which M1-fl and S1-fl play a strong role in shaping the fine details of reaching and grasping movements.”
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
In addition to the weaknesses noted above, I suggest the authors also address the following:
The last results section is generally lacking in statistical support for claims. Statistical support should be added.
Thank you for pointing this out, we have added more statistical support to this section.
The consideration in the Discussion of relevant previous findings and potential explanations for the distal limb signals in mouse sensorimotor cortex is somewhat lacking. There are several specific issues:
(1) In contrast to the present study, the studies cited in regards to a lack of motor cortical involvement did not involve dexterous movements - in fact, Kawai et al. explicitly engineered a task that did not involve dexterity to distinguish the role of motor cortex in learning from its known role in dextrous movement execution. In Kawai et al., the authors note one rat who adopted a more dexterous approach to the lever pressing task; in this rat, a motor cortical lesion did cause a longer-lasting reduction in task performance. In additional experiments reported in Kawai's PhD thesis, performance of a dextrous task does erode with motor cortex lesion, as seen in other studies, like the early rodent reaching work of Whishaw and colleagues.
(2) Other possible explanations for the persistence of non-dexterous tasks following motor cortical removal are compensation by, or redundant functionality in, other motor system regions.
(3) It is also worth noting that stimulation in different regions of mouse M1 and S1 evokes alternately, digit, wrist, and elbow movements in fairly similar proportions (Tennant, 2011), suggesting that descending pathways substantially target spinal circuits that control all forelimb joints.
(4) It also seems relevant that although the recovery time course is longer, nonhuman primates also retain substantial hand control after motor cortical removal (e.g. Lashley, 1925; Glees and Cole, 1950; Passingham et al., 1983). Humans of course, appear to be a different story.
These are good points. We have tried to make the Discussion better reflect the tension in the literature, including with this new text:
“However, several other previous results have indirectly suggested that M1 and S1 may be involved in the details of forelimb movement. Performance suffers with inactivation or lesioning of M1 and S1 in skilled, complex manual behaviors (Guo et al 2015, Mizes et al 2024, Whishaw et al 1990) or idiosyncratic use of digits to accomplish non-dexterous tasks (Kawai 2014). The sparing of non-dexterous tasks with these lesions may also reflect redundancy in control as opposed to irrelevance of M1 and S1. Nevertheless, our finding of low-level kinematic information in sensorimotor cortex supports a role for cortex beyond simply providing redundant high-level commands to these subcortical areas.”
We have avoided mentioning points 3 and 4 in the paper; the stimulation results might follow from activating projections not normally involved in this behavior, and discussing primates in this context would require a long list of caveats. We agree that these points are worth thinking about, but are concerned that they are too circumstantial to include in interpreting the results formally.
Although similar decoding performance is achieved using neurons from both CFA and fS1, I am left wondering whether you would do substantially better with CFA using activity at additional preceding time points, or when using exclusively time points from the past. The primary model used here appears to use neural signals from corresponding time points to decode limb parameters, but results seemingly could be different when using preceding time points as regressors.
We appreciate this suggestion and have added the analysis to an additional supplementary panel for Figure 5 (Figure 5S3). Incorporating lags into the decoder via a Wiener filter does indeed improve the decoding performance, but this could simply be due to the increase in the number of predictor variables. This analysis did not, however, further disambiguate M1-fl and S1-fl: the performance improvement was similar across areas for both causal and acausal lag configurations. This could be a consequence of the time resolution of calcium imaging, so further experiments with electrophysiology would be required to rule this possibility out. We now note this new result:
“Including additional causal (-100 ms preceding) and/or acausal (-100 ms preceding to 100 following) lags improved decoding performance modestly and similarly for both areas (Fig. 5S3E-F).”
Related to this, I am also worried about the bleeding of signals across time here. If you deconvolve and interpolate between time points, the interpolation seemingly will pull information into the past, up to half the sampling period, which here is on the order of how long it takes signals to travel to and from the limb. The authors do not make any inappropriate claims about the neural signals here reflecting causes or consequences of what is happening at the limb, but readers (like me) will still try to draw these sorts of conclusions. Is it possible that, although decoding from instantaneous signals is similar for the two regions, the M1 signals are actually motor signals related to future limb state while the S1 signals are sensory consequences? Even if many of the relevant details related to conduction times are not known, perhaps the authors could clarify what can and can't be said related to causal interpretation here.
Thank you for suggesting further explanation here. We agree that our interpretation could be made more specific. We have added text in the Discussion section to speak more directly to what can and cannot be concluded from our analyses. In short, it is hard to be certain of lags in calcium imaging data for many reasons, and using recording methods with finer temporal resolution (like electrophysiology) will be necessary for determining the precise temporal relationships between kinematics and neural activity. In the absence of these recordings, we limit our claim to kinematic information being present in M1-fl and S1-fl neural activity and leave determining the causal role of this information to future work.
New clarifying text in the Discussion:
“The use of calcium imaging further prevents strong conclusions about whether activity reflects future limb states or sensory consequences. Confirming this limitation, inclusion of lagged data in the decoding models, whether causal or acausal, resulted in similar performance changes in both areas.”
An alternative reason why lift onset is less decodable in CFA is that CFA activates substantially before lift onset, as has been observed in previous rodent studies (Kargo and Nitz, 2004; Miri et al., 2017; Veuthey et al., 2020), perhaps as some sort of movement preparation. S1, on the other hand, may not have this early activity, and so may show a clearer transient at onset when the hand and limb start to move. This seems more likely than the explanations provided by the authors.
This is a valid possible alternative explanation and we have updated the Discussion to reflect this. This difference in the structure of M1-fl activity versus S1-fl is apparent in the projections of Figure 6A, which show M1-fl projections more clearly aligned to cue-onset than S1-fl projections.
“Our lift time decoding results are consistent with this view and align with recent observations characterizing mouse proprioceptive forelimb cortex, (Alonso et al 2023), although an alternative explanation may be simply that M1-fl activates earlier than S1-fl during reaching (Kargo and Nitz 2004; Miri et al 2017; Veuthey et al 2020).”
To better clarify relevant similarities and differences between the rodent and primate systems, the Introduction could include some of these similarities and differences exposed by the literature currently cited, and the Discussion could include an additional paragraph specifically relating findings here to previous observations in the primate.
We appreciate the reviewer’s thoughtfulness on possible framings of our results. When writing this paper, framing was a major challenge for us and we drafted quite a few versions of the Introduction including some that focused more on mouse-primate comparison. In the end, we decided the most critical function of the Intro was to set up our central question, of “levels-of-sensorimotor-control”. The rich primate literature was valuable here, but getting into a protracted compare-and-contrast exercise quickly became a distraction from the point. Further, we sought to highlight the relevance and importance of the question answered in our work as the mouse has gained prominence for filling gaps that are challenging to address with primates. This paper serves as one of many early steps towards the ultimate goal of revealing general properties of sensorimotor cortical function with the mouse model. We have made some subtle changes to the Introduction that we hope will more clearly communicate this narrative.
We agree that a Discussion paragraph directly relating our results to those in primates would benefit our conclusions and have added one:
“These results expand our understanding of the rodent sensorimotor system and highlight similarities to nonhuman primates. We show here evidence in mice of detailed joint angle kinematic signals from the full forelimb in M1 and S1, as has been shown in macaque cortex during tasks involving reaching and grasping objects (Vargas-Irwin et al. 2010; Saleh et al. 2010, 2012; Goodman et al. 2019; Okorokova et al. 2020). Additionally, the earlier onset of movement-related activity in M1-fl compared to S1-fl is similar to macaque M1 and S1 (Tanji and Evarts 1976). Taken together these results suggest that the mouse can be employed to address questions traditionally explored in primates about how cortical activity encodes detailed movement commands.”
Although this is outside the scope of the present study, it would be interesting to image descending projection neurons to see what signals are conveyed downstream, and to what targets. Some signals observed in layer 2/3 may not be strongly reflected in descending projections.
We agree that recording from descending projection neurons in this task would be of deep interest – and also agree that these experiments are beyond the scope of the present study. We look forward to performing these additional experiments in future work.
Minor:
(1) The use of "CFA" and “fS1” is a bit confusing. S1, like M1, is defined primarily based on histological criteria, while CFA is defined by intracortical microstimulation. CFA contains a substantial fraction of fS1, seemingly most of it based on the maps shown in Tennant et al., 2011. This is not really a criticism, as the field has not reached any sort of consensus on this nomenclature yet.
We are similarly unhappy with the inconsistency of the terminology in the field, and struggled with how not to make it worse. After much debate and consultation with colleagues, we decided to use “M1” and “S1” to evoke the century of literature on these areas; and “-fl” to indicate forelimb because it is more intuitive than “-ul” and avoids using the illegible “-ll” for hindlimb (relevant to our subsequent paper). For what we called M1-fl, we recorded where we did because anecdotally we saw similar responses across that swath; but note that this definition is also consistent with the definition of “MOp-ul” found with multimodal mapping by
Munoz-Castaneda (2021), which extends a little anteriorly of MOp as defined by the Allen CCF. As the field continues to mature, we hope future work can converge on a set of shared terms.
(2) Page 4: "Inactivations and lesions of M1 and S1 have shown that M1 is required for the execution of dexterous reach-to-grasp movements" - to me, earlier work from Whishaw and colleagues deserves to be cited here.
We appreciate the suggestion and have updated the references in this section to better reflect the prior work from Whishaw and other researchers.
(3) Page 5: "evoking sufficient trial-to-trial variability to avoid model overfitting." - what I think the authors are referring to here is a particular kind of "overfitting," the consequence of not exploring the full movement space, as opposed to model overfitting from issues with the model-fitting method itself. Rather than just saying overfitting, the authors could be clearer about what they are referring to.
The reviewer is right; the phenomenon we intended to refer to is not properly termed overfitting. Specifically, we meant that data with restricted range does not necessarily express global structure, and models can therefore incorrectly fit them. For example, fitting a linear model to data including many periods of a sine wave will correctly show a zero-slope linear component, but fitting to only a portion of a single cycle will typically yield a nonzero slope. This is not overfitting, is not exactly underfitting (because the relevant structure is barely present in the data, as opposed to missed by an insufficiently powerful model), is not bias (the data are fit well), and is not even necessarily a problem (the local relationship may be what you are interested in). Yet, it does not reflect the larger structure of the data.
We do not know of a standard term for this phenomenon, so instead of dragging the reader through this tangential argument, we have tried to offer a simpler motivation for using multiple targets:
“Assessing the relationship between neural activity and the details of movement requires striking a balance between achieving repeatable behavior and evoking sufficient trial-to-trial variability to broadly sample movement space”.
(4) Page 5: Caudal Forelimb Area should not be capitalized.
Obviated with the change in area nomenclature.
(5) Page 7: "of linearly independent degrees of freedom" - for a neuroscience audience, I think it is better to explicitly mention that the resulting PCs are uncorrelated.
We agree that this section could benefit from clarification. We have attempted to provide additional nuance to indicate what the analysis was intended to test.
“Despite the strong coupling between the proximal and distal joint angles, rich variation remained in the action of different joints over time. The presence of strong correlations across joints suggested that the kinematics may be well described by a smaller number of independent degrees of freedom than the total number of recorded angles. To assess the number of linearly independent (uncorrelated) degrees of freedom amongst the 24 joint angles and velocities, we used double-cross-validated PCA (Yu et al. 2009); Methods; Fig. 3D), finding intermediate dimensionalities of 7 (median for joint angles) and 10 (velocities; Fig. 3E). This is consistent with the idea that joint angles across the limb are coordinated instead of controlled independently, and that this coordination is flexible enough over time to enable accurately performing reaching and grasping to different targets.”
(6) Page 7: In the Results, the authors should mention what indicator is being used, the imaging frame rate, and summarize briefly how cells were defined.
Thank you for the suggestion, these details have been added to the relevant results section for clarity.
“To do so, we recorded neural activity from neurons in layer 2/3 M1-fl extending into the immediately adjacent secondary motor cortex (M2), and the forelimb region of S1 (S1-fl) using two-photon calcium imaging of GCaMP6f-expressing neurons in layer 2/3 (185-230 μm deep, imaged at 31 Hz, cells extracted with Suite2p (Pachitariu et al 2017)).”
(7) Page 7: "corrected at n=2" - n doesn't typically refer to the number of tests, so for clarity I would say "corrected for dual tests."
Thank you for pointing this out, we have corrected the text and added additional explanation in the methods for our approach to determining statistical significance across the targets and locking events.
“P-values obtained through the ZETA were then Bonferroni corrected for dual tests when measuring the number of cells modulated to a given event and corrected for six tests (2 targets and 3 events) when measuring the overall number of modulated cells.”
(8) Page 7: In the Results, when the decoding is introduced, it would be helpful to have a few details without having to hunt through the Methods. For example, were things regularized, how was cross-validation handled, etc?
Thank you for the suggestion, these details have been added to the relevant results section for clarity.
A simple linear regression model related the single-trial joint angles at all time points to single-trial neural activity at the corresponding moments. The model was fit with ridge regression, the ridge penalty was determined via a heuristic (Karabatsos 2018), and performance was measured on held-out trials (80/20 train/test split, 50 folds).
(9) Page 8: I think it is worth noting how much mouse reaching involves shoulder rotation as opposed to movement in other joints, as this seems very different from primates.
Thank you for pointing this out. We think this is mostly a task difference: our mice were in a quadrupedal stance, whereas monkeys are typically asked to reach from a sitting position. We now mention this in the Results.
“Reaching evoked particularly large rotation of the shoulder, likely because the mice reached from a quadrupedal position to targets on either side of the snout.”
(10) Page 8: Should provide quantification to clarify what is meant by "closely tracked."
We have updated the text to indicate that this claim was meant to be qualitative, and to more clearly highlight that the interest here is the first demonstration of the ability to reconstruct valid forelimb postures from decoded joint angles in the mouse. Quantifying the reconstruction properly would require substantially more manual data labeling, and the successful decoding itself demonstrates indirectly that the reconstructions are good enough to obtain the results of interest.
Additionally, we reconstructed the skeletal representation of the forelimb from the decoded joint angles and found that, as intended, the reconstructed postures had strong qualitative resemblance to the true postures, even of “minor” angles like cylindrical paw deformation or digit splay (Fig. 5C,G).
(11) Page 8: "Overall, these results suggest that instantaneous movement-related signals are similarly distributed across CFA and fS1." - I know we are being succinct here, but this sentence sounds like a non sequitur in the context of this paragraph - perhaps include a conclusion from the results in this paragraph first, then summarize the whole section.
Thank you for the suggestion, we have updated this text to more clearly conclude the results of this section.
Overall, these results reveal that neural activity in M1-fl and S1-fl is closely related to the kinematic details of reach-to-grasp movements. The ability to decode substantial variance in proximal and distal joints suggests that this relationship extends to the entire forelimb and the similar performance obtained from each area suggests that this information is similarly distributed across M1-fl and S1-fl.
(12) Page 10: Mention of projections from fS1 does not explicitly specify their preferential targeting of the dorsal horn, which seems relevant.
We appreciate the suggestion and have added this detail to the text.
Rodent S1-fl is known to influence interneuron populations in the spinal cord through direct and indirect projections that predominantly target the dorsal horn (Ueno et al. 2018), thus these signals may also reflect S1-fl’s important role in modulating reflex circuits to coordinate sensory feedback with movement generation (Moreno-López et al. 2016; Moreno-Lopez et al. 2021; Seki et al. 2003).
(13) Page 31: Labels on the figure indicating what blue and red stand for would be helpful.
Thank you for the suggestion, labels have been added to indicate left and right trials for Figure 5 C/F and Figure 6A.
(14) Page 32: Legend does not include panel D.
Thank you for catching this, the corresponding caption has been added.
Reviewer #2 (Recommendations for the authors):
(1) The Introduction could perhaps set the central question in starker relief. What specifically do the authors mean by high- vs low-level control? As suggested by the cited studies, this has been a fraught issue in primate work for decades, and I think a finer-grained framing of alternative hypotheses would help set up the results. For example, would better performance at decoding joint angles than paw position be evidence for lower-level control? The clarity of the Introduction might also be improved if the facts and unknowns were broken down by species throughout.
We have tried to further improve the focus of the Introduction on the central question, clarify what we mean, and make clearer in the review of the literature which species a finding comes from.
The clarifying text from the introduction is quoted below:
Extensive motor mapping experiments in rodents have revealed that activating different parts of the sensorimotor cortex evokes movements of different body parts or different kinds of movements of the same body part, as it does in primates (for review, see (Harrison and Murphy 2014)). Yet it is unclear how the topography of stimulation-evoked movements relates to the roles of these areas during volitional actions. Perturbations during behavioral tasks in mice involving forelimb lever or reaching movements have provided a coarse-level understanding of how these areas contribute during behavior. Inactivations and lesions of M1 and S1 have shown that M1 is required for the execution of dexterous reach-to-grasp movements (Guo et al. 2015; Sauerbrei et al. 2020; Galiñanes et al. 2018; Wang et al. 2017; Whishaw et al. 1991; Whishaw 2000) and that S1 is essential for adapting learned movements to external perturbations of a joystick (Mathis et al. 2017). However, spinal cord projections from mouse M1 and S1 primarily target spinal interneurons rather than directly synapsing onto motor neurons (Gu et al. 2017; Ueno et al. 2018; Wang et al. 2017), suggesting cortical activity might play a more modulatory role. Further, stimulation of brainstem nuclei alone can evoke naturalistic forelimb actions, including realistic reaching movements involving coordinated flexion and extension of the proximal and distal limb (Esposito et al. 2014; Ruder et al. 2021; Yang et al. 2023). Taken together, these results have raised the question of what role mouse M1 and S1 play in the control of goal-directed forelimb movements.
One route to answering this question involves characterizing the signals present in mouse M1 and S1 during movement. If mouse M1 and S1 were to control only high-level aspects of forelimb movements, activity should be dominated by ‘abstract’ signals like target location and reflect little trial-to-trial variability in reach kinematics. If instead M1 and S1 control low-level movement features then activity should correlate strongly with forelimb joint angle kinematics and their trial-to-trial variation when reaching to different targets. While the presence of high- or low-level signals in a cortical area does not necessarily imply that they are causally responsible for these aspects of movement, characterizing what signals are present serves as a first step toward determining how these areas relate to movement.
(2) The kinematics and calcium traces appear to be highly stereotyped across trials. If the population encodes joint angles, would one expect to find correlations between the neural and kinematic residuals after subtraction of the time-varying means? Some additional analysis and/or discussion on this point would be helpful, especially as there are only two targets.
This is a great idea. As suggested, we implemented regression models on the residuals for each target in the new Figure 5S3. Figure 5S3 A and B show the performance when decoding the residuals for right trials and C and D show performance for left trials. Decoding remained well above chance, despite shrinking down due to predicting this relatively small within-target variation. This analysis supports our claims from the main regression models in Figure 5 and 5S1-2, and also suggests that movements ipsilateral to the reaching limb (contralateral to the recording hemisphere) may be better encoded than movements contralateral to the reaching limb. We have added a reference to this additional residual analysis in the final paragraph of the decoding section of the Results section:
“Finally, we tested whether the ability to decode these many joint angles was a direct consequence of inter-joint correlations, and might not be indicative of the presence of “real” information about some of these joints. To do so, we fit partial correlation models that removed correlations between proximal and distal joints, or removed correlations of the joint angles with a high-level parameter – the overall distance of the paw centroid to the spout. Despite substantially lowering the behavioral variance, in each case the residuals could still be decoded from neural activity (Fig 5S2A-D). Similar decoding performance for M1-fl and S1-fl was obtained from models fit to decode single-trial residuals separately for left and right trials (Fig 5S3A-D), indicating that trial-to-trial variations on each basic movement were decodable from these populations.”
Along similar lines, binary classification is used to characterize cue-, lift-, and contact-responsive neurons. Is it possible to exploit trial-to-trial variation in the cue-lift and lift-contact latencies to extract the time-varying marginal effects of each event (e.g., using a GLM)?
For the detection of single-cell modulations by different events, we have elected to retain our simple statistical test to determine modulation; in our experience, encoding models typically involve a surprising number of steps to get them to do what you actually intend. We leave more extensive encoding model-style analysis to future work, currently in progress.
(3) The authors mention prior studies suggesting that the control of some forelimb tasks can be gradually transferred from the cortex to the subcortical centers. Have they performed the inactivation at different time points across learning, and if so, do they have evidence for a diminishing effect over time (e.g., blocking of both initiation and coordination early in training)? In addition, the effects of motor cortex inactivation are similar to, but slightly different from, effects shown in reaching tasks in prior studies. Some additional discussion on this point would be useful.
Our inactivation experiments in this study were intended to coarsely demonstrate the involvement of mouse forelimb sensorimotor cortex in our task. We have not performed the inactivations over learning and leave such experiments to future work.
We agree that a little more clarity relating our results to previous ones was warranted. Previous studies (Guo et al. 2015 and Galinanes et al. 2018) have demonstrated inactivation impacts on similar tasks, but for thoroughness we sought to show the same for our task as it varied from the pellet and motorized water spout tasks in both training time and target configurations. Our results are strongly in line with those of Galinanes et al. 2018 which used a fairly similar water spout target configuration. In the inactivation experiments of that paper, 3 out of 13 animals with initiation-triggered inactivations were able to initiate reaching within a time window similar to control trials. Additionally, a proportion of trials across multiple mice proceeded with little perturbation from the inactivations. This is consistent with our observation that M1-fl inactivations may either abolish movement initiation or allow movement initiation but impair task completion on a trial-by-trial and animal-to-animal basis. Further work is required to determine what factors influence these differential responses to inactivation and to determine how these effects differ across task variations (i.e., pellet vs water spout). We have added a brief description of these nuances to the text for clarity.
“These inactivations blocked the execution of the reach to grasp sequence, preventing the animal from making contact with the spout during the 3-second laser stimulation period (Fig. 1F; 86.5% control trials with contact within 3 seconds of cue, 5.1% inactivation trials with contact, P < 10<sup>-191</sup>, Mann-Whitney U test, 2 mice, 495 stimulation trials). Interestingly, inactivation at the time of cue often did not prevent reach initiation (mouse 1: 54.7%, mouse 2: 34.2% of inactivation trials with lift within 3 seconds; 93.5%, 86.2% control trials). Yet the movement stalled once the paw and digits extended towards the spout, producing uncoordinated and unsuccessful reaching trajectories (Fig. 1I, two representative datasets). Taken together, these results support the involvement of M1-fl in the water-reaching task and suggest that the strength of inactivation effects may depend on specific task details like training time or target configuration (c.f. Galinanes et al. 2018).”
Minor points
(1) The rationale for the multiple comparisons procedure in identifying event-locked responses should be explained in more detail. If I understand correctly, the authors are not correcting for comparisons across ROIs, but instead control the family-wise error rate across brain regions and event types (dividing alpha by two or six). Why not instead control the false discovery rate across ROIs?
Thank you for pointing this out, it was confusing as written and we received a similar comment from Reviewer 1. We have fixed the wording now to make it clearer why we did this. We simply aimed to describe how many of the recorded neurons in each area were modulated by the task as a proxy for the engagement of these areas during the behavior, and to use this measure of modulation as a criterion for including the neuron in subsequent analysis. In other words, if the question had been “are any neurons in this area modulated by the task?” then correcting for the number of ROIs would be the correct method; but if the question is, “is this neuron probably modulated and therefore worth including in my decoder?” correcting for the number of ROIs will typically be much too conservative. Thus, we only sought to correct for the false discovery rate across events and targets for each ROI. We have added additional text in the methods to clarify these choices, below. Please also see response to (7) from Reviewer 1 above.
“Note that we did not correct for the number of ROIs tested for two reasons. First, the goal of this testing was to serve as a criterion for inclusion in subsequent decoding analyses, not to determine whether any neurons in the area at all were modulated; and second, correcting for the number of ROIs would bias comparison between areas if different numbers of ROIs were recorded in one area vs. the other.”
(2) It appears joint angles are treated as linear variables in the decoding analysis; is this correct? This seems reasonable as long as the range of motion is not too large, but the authors might briefly comment on the issue in the Methods.
Yes, all joint angles are treated as linear variables in the linear regression model. We observed empirically (as can be seen in Figure 3B and Figure 5B/F) that the joint angle variables were relatively constrained to specific ranges during the task, with no angles displaying substantial wrap-around during the reaching and grasping movements. It is true that use of nonlinear decoding would almost surely improve performance further. Future work could also compare decoding of joint angles with muscle forces, which correlate and which we made no effort to distinguish here. In this work, though, the demonstration of a substantial relationship between neural activity and kinematics already tells us that fine details of movement are present in the M1 and S1-fl populations, which is a critical fact to understand these areas and was not previously known. We now comment explicitly on this, as suggested.
“Joint angle or velocity kinematics were linearly interpolated from their original 6.66 ms to 10 ms and smoothed with a Gaussian (15 ms s.d.). These angular variables were then treated linearly in decoding analyses as their ranges were relatively constrained during the reaching and grasping movements; although the true relationships are likely nonlinear, this serves as a sufficient approximation to demonstrate the presence of a relationship between neural activity and kinematics.”
(3) Are the limb pose estimates mirrored along the mediolateral axis? Figures 1C and 2D appear to show reaches to the left spout on the animal's right.
Thank you for pointing out the ambiguity in the display of these data. The reach trajectories were not mirrored along the mediolateral axis, but they are displayed from the perspective of the behavioral imaging cameras as shown in Figure 1A. Thus the right target reaches (ipsilateral to the animal’s reaching arm) are on the left side of the camera image and the left target reaches (contralateral to the animal’s reaching arm) are on the right side of the image. We have clarified this in the figure captions.
Reviewer #1 (Public review):
Wang et al. studied an old, still unresolved problem: Why are reaching movements often biased? Using data from a set of new experiments and from earlier studies, they identified how the bias in reach direction varies with movement direction and movement extent, and how this depends on factors such as the hand used, the presence of visual feedback, the size and location of the workspace, the visibility of the start position and implicit sensorimotor adaptation. They then examined whether a target bias, a proprioceptive bias, a bias in the transformation from visual to proprioceptive coordinates and/or biomechanical factors could explain the observed patterns of biases. The authors conclude that biases are best explained by a combination of transformation and target biases.
A strength of this study is that it used a wide range of experimental conditions with also a high resolution of movement directions and large numbers of participants, which produced a much more complete picture of the factors determining movement biases than previous studies did. The study used an original, powerful and elegant method to distinguish between the various possible origins of motor bias, based on the number of peaks in the motor bias plotted as a function of movement direction. The biomechanical explanation of motor biases could not be tested in this way, but this explanation was excluded in a different way using data on implicit sensorimotor adaptation. This was also an elegant method as it allowed the authors to test biomechanical explanations without the need to commit to a certain biomechanical cost function.
Overall, the authors have done a good job mapping out reaching biases in a wide range of conditions, revealing new patterns in one of the most basic tasks, and the evidence for the proposed origins is convincing. The study will likely have substantial impact on the field, as the approach taken is easily applicable to other experimental conditions. As such, the study can spark future research on the origin of reaching biases.
Comments on revisions:
The authors have addressed my concerns convincingly. The inclusion of the data on movement extent, and the comparison with the data and explanation of Gordon et al. (1994), has strengthened the paper, as it shows that the proposed model can also explain biases in movement extent. I also appreciate the addition of the mathematical analysis, although I suspect that this analysis can be developed further to yield more detailed insights into the conditions under which the 1-, 2- and 4-peaked patterns arise, but that is a more suitable question for follow-up work.
Reviewer #3 (Public review):
This study makes excellent use of a uniquely large dataset of reaching movements collected over several decades to evaluate the origins of systematic motor biases. The analyses convincingly demonstrate that these biases are not explained by errors in sensed hand position or by biomechanical constraints, but instead arise from a misalignment between eye-centric and body-centric representations of position. By testing multiple computational models across diverse contexts-including different effectors, visible versus occluded start positions-the authors provide strong evidence for their transformation model. My earlier concerns have been addressed, and I find the work to be a significant and timely contribution that will be of broad interest to researchers studying visuomotor control, perception, and sensorimotor integration.
Comments on revisions:
None
Author response:
The following is the authors’ response to the previous reviews
General recommendations (from the Reviewing Editor):
The reviewers agreed that addressing some specific concerns would improve the clarity of the paper and the strength of the conclusions. These points are listed below, and described in more detail in the reviewer-specific 'Recommendations for Authors':
We thanks the editor and reviewers for the encouraging feedback and constructive comments. We provide our point-by-point response below.
(1) The details of the new experiment including number of subjects and a description of the analysis should be provided in the main text.
We now provide a detailed description of the methods (including the number of subjects; N = 30) and analyses for the new experiment. See our response to Reviewer 2 for more details.
(2) It would be informative to see how the amplitude biases observed, agree with those found by Gordon et al. 1994.
Addressed. Please see our response to Reviewer 1, comment 1.
(3) Each of the models lead to different bias patterns. It would be very helpful to hear the author's interpretation, ideally with a mathematical explanation, of what leads to these distinct patterns.
Addressed. Please see our response to Reviewer 1, comment 2.
Reviewer #1 (Recommendations for the authors):
(1) Most of my points have been addressed convincingly in this revision. The new experiment in which also biases in movement amplitude were determined is a welcome addition to the paper. However, I could not see the results of this study, as the authors did not include Fig. 4 in the manuscript, but repeated Fig. 3. That's unfortunate as I would have like to see the similarity between the biases in direction and amplitude. Moreover, I would have liked to see how the amplitude biases agree with those found by Gordon et al. EBR (1994) 99:112-130, and to which extent Gordon et al.'s explanation can explain the pattern.
We apologize for including the incorrect figure in the previous version of our manuscript. We did make a correction and submitted a corrected version, but it appears that it didn’t make its way to you. The correct Figure 4 is now in the manuscript.
The motor biases in amplitude (extent) observed in Experiment 4 (Author response image 1) are qualitatively similar to the pattern reported by Gordon et al. 1994. While the exact peaks do not match perfectly, both datasets show a two-peaked pattern.
Gordon et al. (1994) attributed the bias in amplitude to direction-dependent variation in movement speed which, in their view, arise from anisotropies in limb inertia. Specifically, moving the upper arm along its quasiorthogonal direction (i.e., rotation about the elbow) requires lower effective inertia than moving parallel to the upper-arm axis. Given the arm posture in both datasets, the upper limb points toward ~135°/315°, with the orthogonal direction corresponding to ~45°/225°. The two-peaked speed profiles in both our data Author response image 1 and Gordon et al. are consistent with this prediction.
Author response image 1.
Gordon et al (1994) noted that, while the extent bias function should mirror the speed bias function, the motor planning system might proactively compensate for the speed bias. Indeed, while the extent and speed bias functions are roughly aligned in their study, the two are misaligned in our Experiment 4. For example, the speed function peaks around 45° which corresponds to a valley in the extent bias function. The difference between their data and ours could be due to a difference in the starting point configuration. However, their model predicts alignment of the speed and extent functions independent of starting point configuration. In contrast, the TR+TG model does predict our observed extent bias function and yields predictions about how this should change with different start point configurations. As such, while heterogeneity in movement speed may contribute to extent bias to some degree, we think the transformation bias and visual-target bias likely play a larger role in determining the amplitude bias observed extent bias at movement endpoint.
We have added a discussion section about the bias function reported by Gordon et al. (1994) and their account in the manuscript (lines 482-493). We do not repeat it here, as the content largely overlaps with the response above.
(2) One of the most important new insights from this study is that the three single-source models lead to different bias patterns, with 1, 2 or 4 peaks. However, what I miss in the paper is an intuitive explanation why they do so. Now, the models are described and their predictions are shown, but it remains unclear where these distinct patterns come from. As scientists, we want to understand things, so I would very much appreciate if the authors can provide such an intuitive explanation, for instance using a mathematical proof. That could also identify how general these patterns are, or if there are certain requirements for them to occur (such as a certain shape of the transformation bias).
Note that the closed-form mathematical expression for the motor bias function is not straight forward. As such, the intuition comes primarily from inspection, that is, the model simulations themselves, what we show Figure 1 of the paper. Importantly, the model predictions are insensitive to the parameter values over a reasonable range. Thus, the number of peaks predicted by each model is a core distinguishing feature. We present in the Supplementary Results a formalized mathematical analysis to illustrate how different models produce different numbers of peaks in the movement-bias function.
(3) I think it's a good idea to change the previous "Visual Bias" into a "Target Bias". This raises the question whether the "Prioprioceptive Bias" should not be changed into a "Hand Bias" or "Start Bias"?
While we appreciate the reviewer’s point here, we prefer the term “Proprioceptive Bias” given that this term has been used in the literature and provides a contrast with sources of bias arising from vision. “Hand Bias” and "Start Bias” seem more ambiguous.
L51: I think "would fall short" should be replaced by "would overshoot".
L127: I think "biased toward the vertical axis" should be replaced by "biased away from the vertical axis". Figure 3 still contains the old terminology like T+V. Please replace by the new terminology. L255: Replace "Exp 1a" by "Exp 1b".
L376: Replace 60 by 6.
L831-2: I hope the summed LL was maximized, not minimized.
Thanks for catching the typos. We have corrected all of them.
Reviewer #2 (Recommendations for the authors):
I think that Experiment 4 does not mention how many participants performed the study. (Only in the response to the reviewers I found this)
We have added information regarding the number of participants in the Fig 4 (N=30).
I am very happy that the authors added the biomechanical simulation into the paper. I am not convinced that this addressed my concerns exactly but it is an excellent addition and the authors have now adjusted the text appropriately.
We appreciate the positive response to our additional assessment of biomechanical factors. We welcome any additional information on how we might fully address this issue.
line 826: extend -> extent
Corrected.
Figure 4. I think that the authors have put the wrong figure here. I cannot see any data for extent. I would need to see this figure (or please correct me - but the caption doesn't match the figure and I don't see the results clearly. (I think the review might have the correct figure).
We apologize for this mistake. We now provided the correct Figure 4 in the paper (also included in the first page of the response letter).
I am missing the detailed description on when the direction error and distance error were calculated for exp 4 - and what exactly was used? How did the authors examine the values without correction? What time point was used? Did I miss the analysis section for this?
Participants were instructed to make fast, straight movement without any corrections and were given up to 1 s to complete the movement. Hand position was recorded once the movement speed dropped below 1 cm/s. On 99.8% of trials, movement speed did not increase once this threshold was passed, indicating that the participants adhered to the instructions. On the remaining trials, we detected a secondary corrective movement (increase in speed >5 cm/s). On these trials, we used the position recorded when the movement speed initially dropped below 1 cm/s as the endpoint position. The pattern of results would be the same were we to exclude these trials.
This information has been added to the Methods section (line 661-666).
Reviewer #1 (Public review):
In this study, the authors explore the implications of two types of rhythmic inhibition - "gamma" (30-80 Hz) and "beta"(13-30Hz) - for synaptic integration. They study this in a multi-compartmental model L5 pyramidal neuron with Poisson excitation and rhythmic inhibition (16 Hz and 64 Hz), applied either to the perisomatic or apical tuft regions in the neuron. They find that 64 Hz inhibition applied to the cell body is effective in phasic modulation of AP generation, while 16 Hz inhibition applied to the apical tufts is effective in phasic modulation of dendritic spikes (in addition to APs). Switching the location of the two kinds of rhythmic inhibition reduces the overall excitability, but is not effective in phasic modulation of either dendritic spikes and weakly so for somatic APs.
Strengths:
The effect of the timescale of rhythmic inhibition on synaptic integration is an interesting question, since a) rhythmic spiking is most strongly evident in inhibitory population, b) rhythmic spiking is modulated by behavioral states and the sensory environment. The methods are clear and data are well-presented. The study systematically explores the effect of two frequencies of rhythmic inhibition in a biophysically detailed model. The study considers not only idealized rhythmic inhibition but also the bursty kind that is observed in in-vivo conditions. Both distributed and clustered excitatory synaptic organization are simulated, which covers the two extremes of the spatial organization of excitatory inputs in-vivo.
Author response:
The following is the authors’ response to the original reviews.
Reviewer #1:
SOM+ interneurons such as Martinotti cells target the apical tufts of pyramidals in the cortex. Since interneurons in general are strongly implicated in mediating rhythmic population activity over a range of timescales, it is quite appropriate to study the consequence of rhythmic inhibition provided by SOM+ interneurons for synaptic integration, including the phenomenon of dendritic spikes. However, using conclusions from a singular study (ref 22) to identify the beta band as the rhythm mediated by SOM+ is not very accurate. SOM+ interneurons have been implicated in regulating rhythms centered just below 30 Hz (refs 22, 21). It is a range that lies in the grey zone of the traditional definition of beta and gamma. However, it is significantly higher than the 16 Hz rhythms explored in this study. It thus remains unknown how a 25-30 Hz rhythmic inhibition (that has an experimentally suggested role for dendrite targeting SOM+ INs) in apical tufts regulates dendritic spikes.
We agree with the reviewer that the rhythms arising from SOM+ interneurons can extend their frequencies higher than the 16 Hz analyzed in this study. To address this, we have conducted a new set of simulations where we delivered distal dendritic inhibition across a range of frequencies, from 0.5 to 80 Hz (see new Results section “Frequency specific effects of rhythmic inhibition on neuronal integration”). These results revealed, surprisingly, that at 30 Hz their ability to entrain Ca<sup>2+</sup> and NMDA spikes degrades (but not Na<sup>+</sup> spikes). This suggests that beta rhythms in the 20-30 Hz range are operating at the highest frequency for which dendritically targeting inhibition will be effective. The implications are covered in the Discussion section “Interaction with microcircuitry”. They are:
“Particularly in the visual cortex, SOM interneurons can generate a rhythm in the 25-30 Hz range [22]. We found this to be at the upper end of the frequency range for dendritic inhibitory rhythms to be effective in modulating NMDA and Ca<sup>2+</sup> spikes. If this rhythm solely recruited SOM interneurons, its effectiveness would be marginal. Potentially compensating for this, recent work has found that PV interneurons also participate in beta/low-gamma [23, 24] (but see [21, 22]). In our model, on its own when beta rhythmic inhibition was delivered perisomatically we found that it was less able to entrain spiking and had an overall hyperpolarizing effect. However, if delivered in conjunction with the distal dendritic inhibition arising from SOM interneurons, this may strengthen entrainment.”
Distal dendritic inhibition has been previously shown to be more effective in controlling dendritic spikes. However, given the slow timescale of dendritic spikes, it can be hypothesized that high-frequency rhythmic inhibition would be ineffective in entraining the dendritic spikes either in distal or proximal location, as demonstrated by 4H and 5F, and vice versa. A computational study can take this further by exploring the robustness of this hypothesis. By sticking to a single-frequency definition of what constitutes Gamma (64 Hz) and Beta (16 Hz) inhibition, the current exploration does support the core hypothesis. However, given the temporal dynamics of dendritic spikes, it is valuable to learn, for example, the upper bound of "Beta" range (13-30Hz) inhibition that fails to phasically modulate them. In addition to the reason stated in the earlier paragraph, Alpha band activity (8-12 Hz), has been implicated (e.g. van Kerkoerle, 2014) in signaling of inter-areal feedback to the superficial layer in the cortex, potentially targeting apical tufts of pyramidals from multiple layers and resulting in alpha-range rhythmic inhibition. To make the findings significant, it might therefore be more pertinent to understand the consequences of ~10Hz rhythmic inhibition (in addition to the ~25-30 Hz Beta/Gamma) in the apical tufts for phasic modulation of dendritic spikes.
We added an additional set of simulations that address this in the Results section ‘Frequency specific effects of rhythmic inhibition on neuronal integration’. In general, we found that dendritic and perisomatic inhibitory rhythms at lower frequencies could entrain AP generation, but with less functional specialization. This is explored in our Discussion section ‘Interneuron specializations and rhythm timescales’.
The differential effect of Gamma and Beta range inhibition on basal and apical excitatory clusters is not convincing from the information provided. The basal cluster appears to overlap with perisomatic inhibitory synapses. The description in the methods does not have enough information to negate the visual perception (ln 979-81). With this understanding, it is not surprising that the correlation between excitation and APs is high (during the trough of gamma) for basal and not apical excitation. A more comparable scenario would be a more distal location of the basal excitatory cluster.
While we stated in the original manuscript that we were contrasting ‘basal’ vs. ‘apical’ clustered inputs, this terminology did not reflect our intent with these analyses. We meant to contrast proximal vs. distal dendritic clustered synaptic inputs, which the reviewer correctly noted is confounded in the apical vs. basal comparison. We have rewritten these results, their discussion, and corresponding figure, to clearly state that we are contrasting proximal vs. distal synaptic input.
Reviewer #2:
The weaknesses are probably in some of the parameterizations of inhibitory synaptic dynamics. A unitary peak conductance of 1nS is very high for inhibitory synapses. This high value could invariably skew some of the network-level predictions. The authors could obtain specific parameters from the Neocortical Collaboration Portal (https://bbp.epfl.ch/nmcportal/microcircuit.html), which is an incredible resource for cortical neurons and synapses.
We appreciate the valuable resource mentioned by the reviewer and will consult it when constructing future models. Regarding the present one, our choice of peak conductance was based on previous studies, namely:
Egger R, Narayanan RT, Guest JM, Bast A, Udvary D, Messore LF, Das S, de Kock CPJ, Oberlaender M (2020) Cortical output is gated by horizontally projecting neurons in the deep layers. Neuron 105, 122-137.e128.
and
Xiang Z, Huguenard JR, Prince DA (2002) Synaptic inhibition of pyramidal cells evoked by different interneuronal subtypes in layer v of rat visual cortex. J Neurophysiol 88, 740-750.
The study by Egger et al. used an inhibitory peak conductance of 1 nS and was simulating circuitry very similar to ours. We validated these synapses in pilot simulations that sought to characterize the resulting IPSPs and IPSCs, and whose results can be seen in Table 1 of our methods. These synapses exhibited IPSCs whose peak amplitudes ranged over values (~24162 pA) that agreed with the experimental literature, such as Xiang et al.
Given this, we feel our parameterization of inhibitory synapses does not warrant any changes.
Reviewer #3:
What disappointed me a bit was the lack of a concise summary of what we learned beyond the fact that beta and gamma act differently on dendritic integration. The individual paragraphs of the discussion often are 80% summary of existing theories and only a single vague statement about how the results in this study relate. I think a summarizing schematic or similar would help immensely.
We agree with the reviewer that a summary schematic would help the reader. This has been added to the manuscript as Figure 11. It demonstrates the principal findings of the paper and is referenced in the opening paragraph of the discussion section.
Orthogonal to that, there were some points where the authors could have offered more depth on specific features. For example, the authors summarized that their "results suggest that the timescales of these rhythms align with the specialized impacts of SOM and PV interneurons on neuronal integration". Here they could go deeper and try to explain why SOM impact is specialized at slower time scales. (I think their results provide enough for a speculative outlook.)
This discussion has been expanded under the section “Interneuron specializations and rhythm timescales”. The added text is:
“So, while our results suggest that spatial targeting of SOM and PV interneurons aligns with the timescales of their network-level rhythms, it could also be that their timing and subcellular localization interact to produce specialized neuron-level functions [85]. For instance, NMDA and Ca<sup>2+</sup> spikes in the distal dendrites last for ~50 ms, making the slower beta rhythm more appropriate for bidirectionally controlling them. Both can be described as dynamical systems with distinct phases with differing sensitivity to inhibition. Ca<sup>2+</sup> spikes are dynamical events comprised of an initiation, plateau, and termination phase. Inhibition delivered during the plateau phase shortens their duration [86]. If the beta rhythm is comprised of cycling between periods of elevated excitation (increased NMDA spike generation) followed by elevated inhibition, then Ca<sup>2+</sup> spike initiation will tend to occur during the excitatory phase, and its plateau during the subsequent inhibitory phase. A plateau during the inhibitory phase will more quickly enter termination. This is bidirectional control. On the other hand, slower rhythms (e.g. 1 Hz) initiate Ca<sup>2+</sup> spikes during the excitatory phase that plateau and enter termination autonomously, before the inhibitory phase is reached. The same principle holds for NMDA spikes [87]. As a result, rhythms in the range from 15-30 Hz are optimal for synchronizing the onsets and offsets of dendritic spikes across a population of neurons.
The integrative effects of gamma (>40 Hz) are also specialized. Low frequency inhibitory rhythms delivered to the soma tended to shift the membrane potential higher or lower with the rhythm’s phase, effectively bringing it closer or farther from AP generation but not changing the neuron’s sensitivity to fast synaptic inputs. In the gamma frequency range, this is reversed, with the mean membrane potential not varying with rhythm phase but with a shifting bias to positive or negative membrane potential fluctuations. In addition, the trough phase of gamma lowers the threshold for AP generation, while slower rhythms like beta only raise the threshold. Consequently, the timing of gamma is ideal for increasing the sensitivity of the neuron to rapid excitation. This agrees with the observation that gamma oscillations accompany rapid excitation-inhibition balancing [88].”
We also extended our discussion section ‘Relevance to coding’ to explore how beta and gamma rhythms can support sparse vs. dense population coding, respectively. It reads:
“One interpretation of rhythms arising from local inhibitory feedback is that they maintain the balance between excitation and inhibition. This can be thought of as a normalization operation that maintains activity within a set range. Normalization can be achieved either through a subtractive effect that raises the threshold for initiating an action potential, or a multiplicative effect that lowers the slope of the relationship between excitation and action potential firing rate. When considered at the population level, these normalization effects impact coding in different ways. Subtractive normalization increases sparsity by dropping out neurons whose excitation is below the raised threshold. Multiplicative normalization, however, encourages dense codes by scaling down firing rates and compressing the range of firing rates. This study found that while both perisomatic and distal dendritic inhibition produced subtractive effects, only perisomatic had a multiplicative effect. Tying this to beta and gamma, beta rhythms may encourage sparse population codes while gamma allows for dense.”
Beyond that, the authors invite the community to reappraise the role of gamma and beta in coding. This idea seems to be hindered by the fact that I cannot find a mention of a release of the model used in this work. The base pyramidal cell model is of course available from the original study, but it would be helpful for follow-up work to release the complete setup including excitatory and inhibitory synapses and their activation in the different simulation paradigms used. As well as code related to that.
We have added a Code and Data Availability section that addresses this. It reads: “Simulation code is deposited at ModelDB athttps://modeldb.science/2019883 . The raw simulation data are available from DBH upon request. Analysis code is posted as a github repo at https://github.com/dbheadley/InhibOnDendComp.”
Reviewer #1 (Public review):
Summary:
The authors investigate the determinants of population-level cell size variability, quantified via the coefficient of variation, in budding yeast populations. Using a combination of computational modeling and experimental readouts, they conclude that mother-daughter division asymmetry is the dominant factor shaping the coefficient of variation of cell size. In particular, through parameter sensitivity analysis of the Chandler-Brown model and empirical perturbations, the authors show that size-control mutations have limited effects on CV, whereas modulating mother-daughter asymmetry, by changing the growth environment, produces substantially larger shifts.
Strengths:
(1) The study addresses a fundamental question in biophysics, i.e., what are the mechanisms that produce and maintain population size heterogeneity?
(2) It provides a conceptual reconciliation for previous observations that size-control mutants often alter mean size but not CV.
(3) The modeling framework is clearly explained and compared to the data.
(4) The parameter sensitivity analysis is thoughtfully performed and provides transparent intuition about which parameters influence variability.
(5) The writing is clear, and the figures are well-organized.
Weaknesses:
(1) The work focuses on the Chandler-Brown model, so it is not clear to what extent the conclusions depend on it. A sensitivity or robustness check using an alternative model would strengthen generality.
(2) CV is the sole descriptor used to quantify heterogeneity; while this is an efficient descriptor, it must be handled with care when used on experimental data, as it may vary due to differences in the chosen observables (e.g., if size is identified via cell volume, length, area, number of proteins, etc.) instead of real differences in the distribution.
(3) The experimental validation using varied nutrient conditions is interesting; however, the statistical significance of the found correlations should be provided/discussed.
Reviewer #2 (Public review):
Summary:
This paper provides a new framework for understanding how cell size variability arises in budding yeast populations. Whereas previous studies emphasized G1/S size control in daughter cells as the main regulator of size homeostasis, the authors show that perturbations to this control checkpoint have only modest effects on population-wide size variability.
By extending a stochastic model of the yeast cell cycle to include both mother and daughter lineages, the authors demonstrate that division asymmetry-stemming from slower growth and longer post-Start phases in mother cells-is the key factor determining the population coefficient of variation (CV). As mothers grow larger and daughters smaller, the overall size distribution broadens. Experimental measurements across multiple mutants and conditions support the predicted correlation between asymmetry and CV.
Strengths:
The main conceptual advance of this study is to consider the full proliferating population, and in particular the dominant mother lineages, rather than single-cycle daughters, thereby offering a population-level explanation for size variability that is consistent with several previous but seemingly conflicting results.
Weaknesses:
Nevertheless, the modelling is described superficially and has notable limitations.
(1) The extended Chandler-Brown model was originally parameterized only for daughter cells, and its generalization to mothers introduces several new assumptions that are not directly tested.
(2) The model treats asymmetry phenomenologically, without a mechanistic basis, so while it correctly identifies correlations, causality remains uncertain.
(3) Moreover, since population CVs emerge from steady-state lineage dynamics, they could be sensitive to parameter choices or growth-related details not fully explored in the current analysis.
In summary, this study provides a useful conceptual synthesis and a useful quantitative framework, but it should be clear that readers should interpret the modeling as heuristic. The central message-that division asymmetry dominates population size variability-remains interesting and well supported at the phenomenological level.
Reviewer #3 (Public review):
Summary:
The article studies the origins of cell size random variability in budding yeast. Different strains with different average cell sizes have very similar noise measured using the coefficient of variability defined as the standard deviation over the mean. Manipulating the noise in key variables such as the duration of cell stages, the growth rate or the division strategy (adder, timer, sizer) was not enough to explain the observed noise in mutants. The proposed solution for the origin of most of the cell size noise is related to the asymmetry in the average cell size for cells with two different phenotypes: daughter cells (New cells that have not passed the first division) AND 'Mother cells' (the rest). The origin of the cell size noise is mainly related to the fact that the distributions of these phenotypes have different cell size distributions. The article includes simple statistical methods for hypothesis analysis and explanatory figures.
Strengths:
The article provides different approaches: experimental (mutants and different growth conditions) and computational (simulations) to explain and test the hypothesis. The methods are based on previous articles with simple conclusions and explanations easy to follow.
The rigor level in both mathematical and biological approaches looks fair to me. The terms are well defined and consistent throughout the article. Authors use well-established analysis techniques.
The proposed theoretical analysis is coarse-grained and therefore can explain different strains and mutations using mathematical tools (noise analysis), aiming to reach general (mathematically) claims. This approach strengthens the conclusions and provides a good language to set a bridge between the biological community and mathematicians (quantitative biologists).
The concept that the population heterogeneity (mothers vs daughters) is a fundamental reason behind the cell size variability is not new, but this article presents a clear experimental justification for the development of complete models of cell size regulation. I consider this contribution very relevant to the community modelling cell size.
Weaknesses:
The concept that population heterogeneity (mother and daughters) with different cell size distributions explains the observed size variability in a heterogeneous population. It is not clear how the population composition can affect this heterogeneity. Intuitively, I would expect that the fraction (number of daughters)/(number of mothers) changes in different stages of the population expansion due to the mean duration of both stages can change in different growth conditions. I would suggest studying how different (or not) these fractions are in different conditions. The authors should acknowledge this effect and discuss briefly using, for instance, simple models of random variables addition (adding different fractions of individuals with different cell size distributions) in which cases (different fractions or different means and noises in their respective distribution) their contribution is relevant. Finally. Do different simulations (gradient or sizer, timer) predict different moments (mean and CV) in distributions of both mother size and daughter size?
Related to the previous comment, I would also include the fraction (number of daughters)/(number of mothers) or the percentage in different growth conditions with their respective size moments (mean and CV) to test whether the resultant cell size moments are related to the addition of two variables with different fractions with their respective moments.
It is interesting how the G1 timer and G1 Sizer are located in different quadrants of Figure 4D, while the studied mutants belong to the other quadrant. I expected them to be closer to the G1 timer, similar to that observed in Figure 4G. I think the authors should discuss this dissimilarity.
Although the authors are working using a definite model, other models would predict different results, especially in synthetic data. For instance, the same models for obtaining sizers can predict different noise levels.
Nieto, C. et al., 2024. npj Systems Biology and Applications, 10(1), p.61.
Barber, Felix, et al., Frontiers in cell and developmental biology 5 (2017): 92.
Teimouri, H. et al,.2020. The Journal of Physical Chemistry Letters, 11(20), pp.8777-8782.
I would mention that the noise level also depends on whether the population has reached steady-state conditions. This would require multiple generations, and measure over at least a couple of thousand cells. Therefore, experiments with single-cell-derived colonies would present different levels of noise than the noise in steady conditions, especially if few cells were sampled. However, I acknowledge that the purpose of the article is not a detailed description of the system but rather the presentation of the concept and for that matter, this level of detail is not mandatory.
Reviewer #3 (Public review):
In this manuscript, authors use the Drosophila wing as model system and combine state-of-the-arte genetic engineering to identify and validate the molecular players mediating the activity of one of the cis-regulatory enhancers of the apterous gene involved in the regulation of its expression domain in the dorsal compartment of the wing primordium during larval development. The paper is subdivided into the following chapters/figures:
(1) In the first couple of figures, authors describe the methodology to genetically manipulate the apE enhancer (a cartoon summarizing all the previous work with this enhancer might help) and identify two well-conserved domains in the OR463 enhancer required for wing development (the m3 region whose deletion phenocopies OR463 deletion: loss of wing, and the m1 region, whose deletion gives rise to AP identify changes in the P compartment).
(2) In the following three figures, authors characterize the m1 regulatory region, identify HOX and ETS binding sites, functionally validate their role in wing development and the activity of the genes/proteins regulating their activity (eg-. Hth and Pointed) by their ability to phenocopy (when depleted) the m1 loss of function wing phenotype. Authors conclude that Hth and Pointed regulate apterous expression through the m1 region.
(3) In the last few figures, authors perform similar experiments with the m3 regulatory region to conclude that the Grn and Antennapedia regulate apterous expression through the m3 enhancer.
My comments:
Technically sound: As stated in my previous review, the work is technically excellent (authors use state-of-the-art genetic engineering to manipulate the enhancer and combine it with genetic analysis through RNAi and CRISPR/Cas9 and phenotypic characterization to functionally validate their findings), figures are nicely done and cartoons are self-explanatory.
Poor paper writing: The paper is too long and difficult to read/understand, many grammatical mistakes are found, and formatting is in some cases heterodox.
Science:
(1) The question of "who is locating the relative position of the AP and DV boundaries in the developing wing?" is not resolved. I would then change the intro or reduce the tone of this question. Having said that, I agree that these results shed light on the wing phenotypes of some apterous alleles related to AP identify and growth and, as such, I congratulate the authors.
(2) Identification of two TFs (Grain and Antp) mediating the regulation of apterous expression is interesting but some contextualization might be required. Data on Antp is not as convincing as data on Grn. I wonder whether Antp data can be removed at all.
(3) I am not sure whether the term hemizygous is used properly
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
The Drosophila wing disc is an epithelial tissue, the study of which has provided many insights into the genetic regulation of organ patterning and growth. One fundamental aspect of wing development is the positioning of the wing primordia, which occurs at the confluence of two developmental boundaries, the anterior-posterior and the dorsal-ventral. The dorsal-ventral boundary is determined by the domain of expression of the gene apterous, which is set early in the development of the wing disc. For this reason, the regulation of apterous expression is a fundamental aspect of wing formation.
In this manuscript, the authors used state-of-the-art genomic engineering and a bottom-up approach to analyze the contribution of a 463 base pair fragment of apterous regulatory DNA. They find compelling evidence about the inner structure of this regulatory DNA and the upstream transcription factors that likely bind to this DNA to regulate apterous early expression in the Drosophila wing disc.
Strengths:
This manuscript has several strengths concerning both the experimental techniques used to address the problem of gene regulation and the relevance of the subject. To identify the mode of operation of the 463 bp enhancer, the authors use a balanced combination of different experimental approaches. First, they use bioinformatic analysis (sequence conservation and identification of transcription factors binding sites) to identify individual modules within the 463 bp enhancer. Second, they identify the functional modules through genetic analysis by generating Drosophila strains with individual deletions. Each deletion is characterized by looking at the resulting adult phenotype and also by monitoring apterous expression in the mutant wing discs. They then use a clever method to interfere in a more dynamic manner with the function of the enhancer, by directing the expression of catalytically inactive Cas9 to specific regions of this DNA. Finally, they recur to a more classical genetic approach to uncover the relevance of candidate transcription factors, some of them previously known and others suggested by the bioinformatic analysis of the 463 bp sequence. This workflow is clearly reflected in the manuscript, and constitutes a great example of how to proceed experimentally in the analysis of regulatory DNA.
We thank the reviewer for these positive comments on the manuscript.
Weaknesses:
There are several caveats with the data that might be constructed as weaknesses, some of them are intrinsic to this detailed analysis or to the experimental difficulties of dealing with the wing disc in its earliest stages, and others are more conceptual and are offered here in case the authors may wish to consider them.
(1) The primordium of the wing region of the wing imaginal disc is defined by the expression of the gen vestigial, which is regulated by inputs coming from the dorsal-ventral boundary (Notch and wg) and from the anterior-posterior boundary (Dpp). Having such a principal role in wing primordium specification and expansion, I am surprised that this manuscript does not mention this gene in the main text and only contains indirect references to it. I consider that the manuscript would have benefited a lot by including vestigial in the analysis, at least as a marker of early wing primordium. This might allow us to visualize directly the positioning of the primordium in the apterous mutants generated in this study, adding more verisimilitude to the interpretations that place this domain based on indirect evidence.
Vg does indeed play a critical role on the formation of the wing disc, and it is an ideal marker for the identification of the wing pouch. In the updated version of the article, we have now followed the expression of vg in some of the OR463 mutants via immunostaining of the Vg protein (Supplementary Figure 6). Cells within posterior wing outgrowths in Δm1flies were invariably positive for Vg. This result further supports our previous identification of these cells as pouch cells. In those mutants in which no cross-over between DV and AP was observed, vg expression was severely reduced or absent, indicating that the wing pouch had not been specified. We thank the reviewer for this experimental idea, which we believe strengthens the final manuscript.
We have added to the text:
“To identify the nature of the posterior outgrowths, we performed anti-Vestigal (Vg) antibody staining of Δm1 mutants (Supplementary Figure 6). Vg is a key regulator of wing specifications and also participates in wing growth and patterning (Baena-Lopez & García-Bellido, 2006; Kim et al., 1996; Zecca & Struhl, 2007a). In those discs, in which the stripe was extended and the P compartment was enlarged, Vg was detected throughout the outgrowth, supporting the wing pouch identity of this region (Supplementary Figure 6B). Hemizygous Δm3 mutants presented a highly reduced anti-Vg signal, which suggests that no wing pouch is specified in these mutants (Supplementary Figure 6C).”
(2) The authors place some emphasis on the idea that their work addresses possible coordination between setting the D/V boundary and the A/P boundary:
Abstract: "Thus, the correct establishment of ap expression pattern with respect to en must be tightly controlled", "...challenging the mechanism by which apE miss-regulation leads to AP defects." "Detailed mutational analyses using CRISPR/Cas revealed a role of apE in positioning the DV boundary with respect to the AP boundary"
Introduction: "However, little is known about how the expression pattern of ap is set up with respect that of en. In other words, how is the DV boundary positioned with respect to the AP boundary?"
"How such interaction between ap and the AP specification program arises is unknown."
Results: "Some of these phenotypes are reminiscent of those reported for apBlot (Whittle, 1979) and point towards a yet undescribed crosstalk between ap early expression and the AP specification program."
At the same time, they express the notion, with which this reviewer agrees, that all defects observed in A/P patterning arising as a result of apterous miss-regulation are due to the fact that in their mutants, apterous expression is lost mainly in the posterior dorsal compartment, bringing novel confrontations between the A/P and the D/V boundaries.
To me, the key point is why the expression of apterous in different mutants of the OR463 enhancer affects only the posterior compartment. This should be discussed because it is far from obvious that apterous expression has different regulatory requirements in the anterior and posterior compartments.
We agree with the reviewer that the differential effect of the mutations on the expression of ap in the A and P compartment is a key factor underlying our explanation of how the phenotypes arise. To clarify this point, we have now extended our first discussion point. Moreover, we have included some other references of differential enhancer regulation in different wing disc compartments. In addition, we have discussed whether this effect has to do with the different regulation of the enhancer in the A and P compartment or due to regulation of downstream effectors.
Added paragraph:
“Although apE is active throughout the dorsal compartment, its disruption leads to a preferential loss of ap expression in posterior cells. The asymmetric effect of apE perturbation on the anterior and posterior compartments suggests that apE transcriptional control is not equivalent across the A/P axis. Compartment-dependent differences in enhancer regulation have also been documented in other developmental contexts; for example, the Distal-less DMX-R element is interpreted through distinct cofactor combinations (Sloppy paired anteriorly and Engrailed posteriorly) (Gebelein et al., 2004), and specific mutations within DMX-R preferentially disrupt enhancer function in anterior versus posterior cells. It is possible that apE is more sensitive to misregulation due to differential transcriptional regulation across compartments. Nevertheless, we cannot exclude the possibility that the posterior bias we observe arises not from enhancer logic per se, but from intrinsic differences in tissue architecture or the dynamics of boundary positioning during wing disc development.”
(3) The description of gene expression in the wing disc of novel apterous mutants is only carried out in late third instar discs (Figs. 2, 3, 5, and 7). This is understandable given the technical difficulties of dealing with early discs, as those shown in the analysis of candidate apterous regulatory transcription factors (Fig. 4F, Fig. 6 C-D). However, because the effects of the mutants on apterous expression are expected to occur much earlier than the time of expression analysis, this fact should be discussed.
We agree with the reviewer regarding the limitations of our analysis whenever we analyzed third instar larvae to assess the expression of the OE463 enhancer. We have included a statement in which this is mentioned in the discussion:
“It is important to acknowledge that all expression analyses were conducted in third-instar discs, a stage that follows the initial establishment of ap expression. Earlier effects are therefore inferred rather than directly observed, as imaging and staging of early discs present significant technical challenges due to their small size and fragility. A direct observation of the early wing disc across mutant conditions would likely help to clarify the role of the discovered factors during early ap expression.”
Reviewer #2 (Public Review):
In their manuscript, "Transcriptional control of compartmental boundary positioning during Drosophila wing development," Aguilar and colleagues do an exceptional job of exploring how tissue axes are established across Drosophila development. The authors perform a series of functional perturbations using mutational analyses at the native locus of apterous (ap), and perform tissue-specific enhancer disruption via dCas9 expression. This innovative approach allowed them to explore the spatio-temporal requirements of an apterous enhancer. Combining these techniques allowed the authors to explore the molecular basis of apterous expression, connecting the genotypes to the phenotypical effects of enhancer perturbations. To me, this paper was a beautiful example of what can be done using modern drosophila genetics to understand classic questions in developmental biology and transcriptional regulation.
In sum, this was a rigorous paper bridging scales from the molecular to phenotypes, with new insight into how enhancers control compartmental boundary positioning during Drosophila wing development.
We would like to thank the reviewer for its positive and encouraging comments, as well as for the careful review of the manuscript and figures. We have adapted most of the suggestions in the new manuscript.
Reviewer #3 (Public Review):
In this manuscript, authors use the Drosophila wing as a model system and combine state-ofthe-art genetic engineering to identify and validate the molecular players mediating the activity of one of the cis-regulatory enhancers of the apterous gene involved in the regulation of its expression domain in the dorsal compartment of the wing primordium during larval development.
(1) The authors raise two very important questions in the Introduction: (1) who is locating the relative position of the AP and DV boundaries in the developing wing, and (2) who is responsible for the maintenance of the apterous expression domain late in larval development. None of these two questions have been responded to and, indeed, the summary of the work (as stated in the conclusions of the last paragraph of the Introduction) does not resolve any of these questions.
We believe the results presented, together with those added during the revision, shed some on the positioning of the boundary. We proposed that the combined integration of four TFs by the OR463 enhancer is fundamental for the correct positioning. Additionally, we proposed a model on how these positioning problems result in the phenotypes observed (Supplementary figure 7, now also shown in Figure 2D). Our results indicate that ap expression in the PD quadrant is particularly sensitive to mutations in the enhancer, which we have now further elaborated on in the first part of the discussion. Together, we believe that our results do tackle the first problem posed in the introduction, while not completely solving them. As for the second question, we have tried to remove any suggestions that this article tries to explain later regulation of apterous. Probably this misunderstanding arises from a sentence in the introduction which has now been deleted. The means of the maintenance of ap expression in later stages has been partially explored previously (See Bieli et al 2015) and it is subject of our current studies.
(2) The authors have identified two different regions whose deletions give very interesting phenotypes in the adult wing (AP identify change & outgrowths, and loss of wing), and have bioinformatically identified and functionally verified 4 TFs that mediate the activity of these regions by their capacity to phenocopy the wing phenotype. While identification of the 2 TFs acting on the m1 is incremental with respect to previous work on the identification of the enhancer responsible for the early expression of Ap, identification of Antp and Grn does not explain the loss of function phenotype of the m3 enhancer. Does any of these results shed any light on the first two Qs? Do these results explain the compartment boundary position in the wing as stated in the title? Expression of lacZ reporter assays is fundamental to demonstrate their model of Figure 8. The reduction of the PD compartment is difficult to understand by the sole reduction in ap expression in this region (which has not been demonstrated).
We agree that the identification of Antp and Grn does not by itself explain the loss-of-function phenotype of the m3 enhancer. However, these transcription factors represent the best current candidates for direct regulators for this enhancer. We have clarified in the text that Antp and Grn may not act as instructive inputs but rather play a permissive role in enabling ap expression through m3. Importantly, the dCas9-mediated perturbation experiments directly demonstrate that targeted manipulation of apE in this region is sufficient to produce the characteristic duplications, providing functional evidence that apE activity underlies the observed phenotypes. In addition, lacZ reporter assays confirm that apE expression is indeed affected in all cases where the experimental setup permitted detection. Together, these results validate that the observed morphological phenotypes stem from perturbation of apE activity and support the proposed model for enhancer regulation and its role in compartment boundary maintenance.
(3) The authors state in one of the sections "Spatio-temporal analysis of apE via dCas9 ". No temporal manipulation of gene activity is shown. The authors should combine GAL4/UAs with the Gal80ts to demonstrate the temporal requirements of Antp/Grn and Pnt/Hth as depicted in their model of Figure 8.
We agree with the reviewer that the temporal dimension was not explored in the first version of the manuscript (aside of the temporal constrains of en-Gal4 driver). As suggested by the reviewer, we have now used a tub-Gal80ts allele to temporally control the enhancer perturbation and delimit its window of activity. The results are included in two new panels in the figure 3 (H and H’). The new data agrees with the notion that apE enhancer is important up to L2 stages but dispensable later in development. We have added the following paragraph to the text:
“To define the developmental time window during which the apE enhancer remains sensitive to repression, we combined the temperature-sensitive tub-Gal80<sup>ts</sup> system with temporally controlled expression of dCas9. Animals carrying the en-Gal4, tub-Gal80<sup>ts</sup>, UAS-dCas9 and U6-OR463gRNA(4x) transgenes were maintained at 18 °C to suppress dCas9 expression. Independent sets of embryos were then shifted to 29 °C at successive developmental intervals ranging from 0 to 168 h after egg laying (AEL), so that dCas9 induction occurred at distinct time points in development (Figure 3H). Under these conditions, dCas9 transcription was induced only after the temperature shift, while the gRNAs were expressed constitutively. Wing phenotypes were quantified in adult progeny as a readout of apE enhancer perturbation. When dCas9 was expressed from embryonic or early larval stages (0–48 h AEL), nearly all wings (70–90%) displayed severe ap-like phenotypes, including posterior compartment duplication and loss of anterior–posterior boundary integrity. Shifting animals later (48–72 h AEL) still produced a majority (~66%) of abnormal wings, whereas induction after 72 h AEL resulted in progressively weaker effects and complete loss of phenotypes by 96 h AEL (Figure 3H’).
These results delineate the developmental period during which apE activity is required for proper wing patterning. Perturbation during the first half of the second larval instar (≤ 96 h at 18 °C) was sufficient to elicit strong ap-like transformations, consistent with the enhancer being functionally required during early larval stages and becoming dispensable thereafter. The temporal decline in phenotype penetrance thus reflects the progressive loss of apE sensitivity to dCas9-mediated repression, providing a precise estimate of when its activity is no longer required for wing morphogenesis.”
(4) The authors have not managed to explain the AP phenotype. Thus, this work opens many unresolved questions and does not resolve the title, which is a big overstatement. Thus, strengths (technically excellent), weakness (there is not much to learn about wing development and apterous regulation from these results besides the incremental identification of 4 additional TFs mediating the regulation of ap expression by their ability to phenocopy regulatory mutations of the apterous gene).
As mentioned in response to reviewer 1, we have indeed no concrete explanation for why the P compartment seems more sensitive to mutations. We have now further discussed this point (see below paragraph, now included in the discussion). As for how the adult phenotypes arise from the mutant wing discs, we have a good idea (see Supplementary figure 7 and Figure 2).
We are pleased to hear that the reviewer considers our article technically valuable. Therefore, we have reformulated the title such as the technical merits play a bigger role in it:
”in situ mutational screening and CRISPR interference demonstrate that the apterous Early enhancer is required for developmental boundary positioning”
Paragraph added to the discussion:
" Although apE is active throughout the dorsal compartment, its disruption leads to a preferential loss of ap expression in posterior cells. The asymmetric effect of apE perturbation on the anterior and posterior compartments suggests that apE transcriptional control is not equivalent across the A/P axis. Compartment-dependent differences in enhancer regulation have also been documented in other developmental contexts; for example, the Distal-less DMX-R element is interpreted through distinct cofactor combinations (Sloppy paired anteriorly and Engrailed posteriorly) (Gebelein et al., 2004), and specific mutations within DMX-R preferentially disrupt enhancer function in anterior versus posterior cells. It is possible that apE is more sensitive to misregulation due to differential transcriptional regulation across compartments. Nevertheless, we cannot exclude the possibility that the posterior bias we observe arises not from enhancer logic per se, but from intrinsic differences in tissue architecture or the dynamics of boundary positioning during wing disc development.”
Recommendations for the authors:
Reviewer #1 (Recommendations For The Authors):
Formatting of references should be checked throughout the manuscript
Reviewer #2 (Recommendations For The Authors):
Here, I note a few points that would help clarify the manuscript and connect it with a broader community.
Figure 1: it could help the reader to add the landing site genetic scheme to the main figure.
In a first draft that was exactly the original configuration, but after comparing both versions we determined that the presence of the landing site removes a bit of the focus of the phenotypes.
Figure 1: what species were used for the conservation alignment? Further details would be nice to add here.
We have now added a section of bioinformatical analysis, which was missing in the original manuscript:
Sequence conservation of the OR463 fragment within the ap upstream intergenic region was analysed across different dipteran species using the “Cons 124 Insects” multiple-alignment track of the D. melanogaster dm6 genome on the UCSC Genome Browser (Kent et al., 2002, https://genome.ucsc.edu). Conservation scores were obtained from the phastCons (Siepel et al., 2005) and used to delineate conserved and less conserved blocks within OR463. Conserved transcription factor binding sites were predicted with MotEvo (Arnold et al., 2011), which defined four conserved modules (m1–m4) and six inter-modules (N1–N6). Additional motif analysis was performed using the JASPAR CORE Insecta database and the Target Explorer tool to cross-validate conserved binding-site predictions and refine motif assignments within the enhancer.
From Figure 2: I would consider moving the model or portions of it to a main figure. These models, while descriptive, really help make the manuscript more approachable. Note that eLife does not have forced figure requirements.
We have adapted the reviewer’s suggestion and we are very grateful for it. We think the figure has greatly improved. The final figure now highlights a small part of the model, which is still included in the Supplementary Figure.
Figure 5: This figure is fantastic, and the results are particularly important. I would recommend increasing the weight of the arrows from D to E, making it more obvious. Did the authors consider any temperature or other perturbations to look at robustness? They mention "robustness" a few times, and this could be an excellent system to explore a bit further. For panels F and G, it would be nice to have a bit of biochemistry here to test the spacing requirements' effects on the distances (but it's great phenotypical data, regardless).
We have chosen a darker grey to highlight the lines.
We appreciate the reviewer’s suggestions. With respect to robustness assays, such as temperature perturbations, we agree that the apE enhancer would be a suitable system for such experiments. However, these analyses would move the study beyond its current scope, which is focused on defining the regulatory logic of boundary positioning through mutational dissection and CRISPRi. We therefore prefer not to expand the work in this direction here, but we note that this would be an interesting avenue for future investigation.
Similarly, biochemical assays probing spacing requirements would provide additional mechanistic insight but would represent a separate line of work. In this manuscript, we aimed to establish the functional consequences of motif spacing using in vivo genetic and phenotypic analyses, which we believe sufficiently support our conclusions.
Thank you for the insight.
Discussion: To the point "most point mutations or short deletions in enhancer regions have little effect on gene expression" I would push the authors to discuss their work in relation to Fuqua et al., (Nature 2020) and Kvon et al., (Cell 2020). Their work is consistent with enhancers being sensitive to mutations, and this warrants further discussion because it could be important for the transcription field.
Hox genes as pioneer factors, I would recommend citing Loker et al., (Curr Biol 2021), as an example of Hox genes functioning as a pioneer factor.
We thank the reviewer for this suggestion. We have now added a short paragraph in the Discussion noting how our observations may relate to the mutational patterns described in Fuqua et al. (2020) and Kvon et al. (2020), while keeping the interpretation tentative. The text now says:
“Recent large-scale enhancer mutagenesis studies have shown that the mutational consequences within enhancers can vary widely. In some cases, many nucleotide positions appear tolerant to single-base changes and only a small subset of mutations produce clear functional effects (Kvon et al., 2020). In other enhancers, regulatory information is distributed more densely, and mutations at multiple positions can alter output (Fuqua et al., 2020). Together, these studies illustrate that enhancer sensitivity is not uniform but depends on enhancer-specific features such as motif organization, cooperativity, and redundancy. Within this broader landscape, the apE enhancer appears to represent a particularly sensitive case.”
We also included a citation to Loker et al. (2021) in connection with the possible pioneer-like contribution of HOX input to apE.
We would like to thank all reviewers for their effort.
Reviewer #1 (Public review):
Summary:
I read the paper by Parrotta et al with great interest. The authors are asking an interesting and important question regarding pain perception, which is derived from predictive processing accounts of brain function. They ask: If the brain indeed integrates information coming from within the body (interoceptive information) to comprise predictions about the expected incoming input and how to respond to it, could we provide false interoceptive information to modulate its predictions, and subsequently alter the perception of such input? To test this question, they use pain as the input and the sounds of heartbeats (falsified or accurate) as the interoceptive signal.
Strengths:
I found the question well-established, interesting and important, with important implications and contributions for several fields, including neuroscience of prediction-perception and pain research. The study is clearly written, the methods are generally adequate, and the results indeed support the claim that false cardiac feedback modulates both pain perception and anticipatory cardiac frequency. Importantly, the authors include a control experiment using exteroceptive auditory feedback to test whether effects are specific to heartbeat-like cues. This addition substantially strengthens interpretability.
Weaknesses:
In my view, the authors' central interpretation, namely that the effects arise because the manipulation targets interoceptive rather than exteroceptive or high-level threat-related cues, cannot be fully supported by the current design. The evidence does not rule out the possibility that participants interpret increased heartbeat sounds as a generic danger/threat cue rather than as (manipulated) interoceptive input. I also disagree with several other claims, though they are less critical, for example, that the use of specific comparisons without pre-registering them, the use of sensitivity analysis to justify sample size, and the intentional use of only 6 trials per participant.
Conclusion:
To conclude, the authors have shown in their findings that predictions about an upcoming aversive (pain) stimulus - and its subsequent subjective perception - can be altered not only by external expectations, or manipulating the pain cue, as was done in studies so far, but also by manipulating a cue that has fundamental importance to human physiological status, namely heartbeats. Whether this is a manipulation of actual interoception as sensed by the brain is, in my view, left to be proven.
Even if the authors drop this claim, the paper has important implications in several fields of science, ranging from neuroscience prediction-perception research, to pain research, and may have implications for clinical disorders, as the authors propose. Furthermore, it may lead - either the authors or someone else - to further test this interesting question of manipulation of interoception in a different or more controlled manner.
I salute the authors for coming up with this interesting question and encourage them to continue and explore ways to study it and related follow-up questions.
Reviewer #3 (Public review):
Parrotta et al provide a convincing and thorough revision of their manuscript "Exposure to false cardiac feedback alters pain perception and anticipatory cardiac frequency". The authors addressed my previous concerns regarding theoretical framing and methodological clarity. For example:
They provided additional detail on the experimental design, procedure and statistical analyses.
The predictive coding rationale for the hypotheses has been clarified.
The limitations of the study are discussed comprehensively
Additional analyses were performed to investigate the role of learning effects and across-experiment effects
New supplementary figures allow a closer look at the feedback-related response patterns
In sum, the revisions improve the manuscript. However, some issues remain present.
(1) Potential learning/ habituation effects. In my first review of the manuscript, I raised the concern that learning effects may have contributed to the observed differences between interoceptive & exteroceptive cues.<br /> The authors argue that the small number of six trials per condition could limit aversive effects of differential learning between experiments. However, electric nociceptive stimuli are exceptionally potent in classical conditioning experiments and humans can develop conditioned responses to these types of stimuli after a single trial [1-2]. Therefore, six trials are sufficient to allow for associative or expectancy-based learning processes.
However, the authors are also presenting additional analyses, i.e. LME models which included trial rank as a predictor. While these models do not show a statistically significant learning effect, they do indicate a noteworthy larger effect in earlier trials compared to later ones. However, in my reading, this speaks towards the presence of unspecific effects of attention or arousal. This pattern is compatible with early learning or, alternatively, with non-specific attentional or arousal responses that diminish across repetitions. This is potentially a limitation of the design: repetition-related effects (attention reduction, arousal habituation, early learning) may contribute to the results, and distinguishing between interoceptive inference and non-specific effects remains challenging within this paradigm.
(1) Haesen K, Beckers T, Baeyens F, Vervliet B. One-trial overshadowing: Evidence for fast specific fear learning in humans. Behav Res Ther. 2017 Mar;90:16-24. doi: 10.1016/j.brat.2016.12.001. Epub 2016 Dec 8. PMID: 27960093.
(2) Glenn CR, Lieberman L, Hajcak G. Comparing electric shock and a fearful screaming face as unconditioned stimuli for fear learning. Int J Psychophysiol. 2012 Dec;86(3):214-9. doi: 10.1016/j.ijpsycho.2012.09.006. Epub 2012 Sep 21. PMID: 23007035; PMCID: PMC3627354.
(2) SESOI and power rationale. The authors elaborated on the sensitivity analyses and the rationale of reporting SESOI rather than traditional a-priori power analyses and included this information in the manuscript, which improves transparency.
(3) Unspecific arousal/ attention mechanisms. The authors argue against unspecific arousal mechanisms based on the absence of main effects in pain ratings and heart rate. This reduces the likelihood of a purely unspecific arousal account, however, these unspecific effects may not need to manifest as main effects. Unspecific mechanisms are likely adding (at least residual) effects onto the results.
Regarding attention-based mechanisms, the authors have clarified that in Experiment 2 (exteroceptive cue), the participants are instructed that the sound does not have any relation with their heart rate. If participants did not receive any instructions on the meaning of the knocking sounds, they may have simply ignored it - not unlikely, also because the exteroceptive feedback did not elicit any systematic effect on the outcome variables (minus the slowing of HR with slower exteroceptive feedback, which may reflect noise, altering, multiple comparisons?). Ultimately, how the participants did or did not process the exteroceptive cue is unclear.
(4) The authors provided more context to their hypothesis and strengthened its theoretical motivation (increased pain intensity with incongruent-high cardiac feedback), rooting it in predictive coding accounts of interoception. For instance, their prior study shows that participants report an increased cardiac frequency while anticipating pain. The reasoning behind this study is hence that if pain shapes cardiac perception, cardiac perception should in turn shape pain perception. The introduction has been revised accordingly, adding more references on the interplay between cardiac feedback and pain and emotional responses. While this rooting within the predictive processing framework is now clearly developed, it also underscores a gap between the proposed theoretical mechanism and the current analytical approach. The hypothesis is formulated in a mechanistic, computational-level language, yet the statistical analysis remains primarily descriptive, at a group level, and does not directly test the predictive-coding account.
New concerns introduced by the revision:
(1) Some of the newly added paragraphs interrupt the narrative flow. For example, the justification of the supradiaphragmatic focus based on the BPQ questionnaire feels too long for this section and might fit more naturally in the theoretical background or introduction. Similarly, the predictive-coding paragraph appearing after the hypotheses seems better suited to the earlier conceptual framing rather than following the hypothesis statements. It would be better for the argumentative flow if hypotheses followed from theoretical considerations.
(2) The authors now note that the administration of the BPQ questionnaire was exploratory, explaining the null-results in the methods section as resulting from an underpowered design. But if the design is not appropriate for discovering a connection between self-reported body awareness and pain ratings, why was it administered in the first place? The rationale here is unclear.
(3) The discussion is longer than before and would benefit greatly from streamlining the arguments.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
I read the paper by Parrotta et al with great interest. The authors are asking an interesting and important question regarding pain perception, which is derived from predictive processing accounts of brain function. They ask: If the brain indeed integrates information coming from within the body (interoceptive information) to comprise predictions about the expected incoming input and how to respond to it, could we provide false interoceptive information to modulate its predictions, and subsequently alter the perception of such input? To test this question, they use pain as the input and the sounds of heartbeats (falsified or accurate) as the interoceptive signal.
Strengths:
I found the question well-established, interesting, and important, with important implications and contributions for several fields, including neuroscience of prediction-perception, pain research, placebo research, and health psychology. The paper is well-written, the methods are adequate, and the findings largely support the hypothesis of the authors. The authors carried out a control experiment to rule out an alternative explanation of their finding, which was important.
Weaknesses:
I will list here one theoretical weakness or concern I had, and several methodological weaknesses.
The theoretical concern regards what I see as a misalignment between a hypothesis and a result, which could influence our understanding of the manipulation of heartbeats, and its meaning: The authors indicate from prior literature and find in their own findings, that when preparing for an aversive incoming stimulus, heartbeats *decrease*. However, in their findings, manipulating the heartbeats that participants hear to be slower than their own prior to receiving a painful stimulus had *no effect* on participants' actual heartbeats, nor on their pain perceptions. What authors did find is that when listening to heartbeats that are *increased* in frequency - that was when their own heartbeats decreased (meaning they expected an aversive stimulus) and their pain perceptions increased.
This is quite complex - but here is my concern: If the assumption is that the brain is collecting evidence from both outside and inside the body to prepare for an upcoming stimulus, and we know that *slowing down* of heartbeats predicts an aversive stimulus, why is it that participants responded in a change in pain perception and physiological response when listened to *increased heartbeats* and not decreased? My interpretation is that the manipulation did not fool the interoceptive signals that the brain collects, but rather the more conscious experience of participants, which may then have been translated to fear/preparation for the incoming stimulus. As the authors indicate in the discussion (lines 704-705), participants do not *know* that decreased heartbeats indicate upcoming aversive stimulus, and I would even argue the opposite - the common knowledge or intuitive response is to increase alertness when we hear increased heartbeats, like in horror films or similar scenarios. Therefore, the unfortunate conclusion is that what the authors assume is a manipulation of interoception - to me seems like a manipulation of participants' alertness or conscious experience of possible danger. I hope the (important) distinction between the two is clear enough because I find this issue of utmost importance for the point the paper is trying to make. If to summarize in one sentence - if it is decreased heartbeats that lead the brain to predict an approaching aversive input, and we assume the manipulation is altering the brain's interoceptive data collection, why isn't it responding to the decreased signal? --> My conclusion is, that this is not in fact a manipulation of interoception, unfortunately
We thank the reviewer for their comment, which gives us the opportunity to clarify what we believe is a theoretical misunderstanding that we have not sufficiently made clear in the previous version of the manuscript. The reviewer suggests that a decreased heart rate itself might act as an internal cue for a forthcoming aversive stimulus, and questions why our manipulation of slower heartbeats then did not produce measurable effects.
The central point is this: decreased heart rate is not a signal the brain uses to predict a threat, but is a consequence of the brain having already predicted the threat. This distinction is crucial. The well-known anticipatory decrease of heartrate serves an allostatic function: preparing the body in advance so that physiological responses to the actual stressor (such as an increase in sympathetic activation) do not overshoot. In other words, the deceleration is an output of the predictive model, not an input from which predictions are inferred. It would be maladaptive for the brain to predict threat through a decrease in heartrate, as this would then call for a further decrease, creating a potential runaway cycle.
Instead, increased heart rate is a salient and evolutionarily conserved cue for arousal, threat, and pain. This association is reinforced both culturally - for example, through the use of accelerating heartbeats in films and media to signal urgency, as R1 mentions - and physiologically, as elevated heart rates reliably occur in response to actual (not anticipated) stressors. Decreased heartrates, in contrast, are reliably associated with the absence of stressors, for example during relaxation and before (and during) sleep. Thus, across various everyday experiences, increased (instead of decreased) heartrates are robustly associated with actual stressors, and there is no a priori reason to assume that the brain would treat decelerating heartrates as cue for threat. As we argued in previous work, “the relationship between the increase in cardiac activity and the anticipation of a threat may have emerged from participants’ first-hand experience of increased heart rates to actual, not anticipated, pain” (Parrotta et al., 2024). The changes in heart rate and pain perception that we hypothesize (and observe) are therefore fully in line with the prior literature on the anticipatory compensatory heartrate response (Bradley et al., 2008, 2005; Colloca et al., 2006; Lykken et al., 1972; Taggart et al., 1976; Tracy et al., 2017; Skora et al., 2022), as well as with Embodied Predictive Coding models (Barrett & Simmons, 2015; Pezzulo, 2014; Seth, 2013; Seth et al., 2012), which assume that our body is regulated through embodied simulations that anticipate likely bodily responses to upcoming events, thereby enabling anticipatory or allostatic regulation of physiological states (Barrett, 2017).
We now add further explanation to this point to the Discussion (lines 740-758) and Introduction (lines 145-148; 154-156) of our manuscript to make this important point clearer.
Barrett, L. F., & Simmons, W. K. (2015). Interoceptive predictions in the brain. Nature reviews neuroscience, 16(7), 419-429.
Barrett, L. F. (2017). The theory of constructed emotion: An active inference account of interoception and categorization. Social cognitive and affective neuroscience, 12(1), 1-23.
Bradley, M. M., Moulder, B., & Lang, P. J. (2005). When good things go bad: The reflex physiology of defense. Psychological science, 16(6), 468-473.
Bradley, M. M., Silakowski, T., & Lang, P. J. (2008). Fear of pain and defensive activation. PAIN®, 137(1), 156-163.
Colloca, L., Petrovic, P., Wager, T. D., Ingvar, M., & Benedetti, F. (2010). How the number of learning trials affects placebo and nocebo responses. Pain®, 151(2), 430-439.
Lykken, D., Macindoe, I., & Tellegen, A. (1972). Preception: Autonomic response to shock as a function of predictability in time and locus. Psychophysiology, 9(3), 318-333.
Taggart, P., Hedworth-Whitty, R., Carruthers, M., & Gordon, P. D. (1976). Observations on electrocardiogram and plasma catecholamines during dental procedures: The forgotten vagus. British Medical Journal, 2(6039), 787-789.
Tracy, L. M., Gibson, S. J., Georgiou-Karistianis, N., & Giummarra, M. J. (2017). Effects of explicit cueing and ambiguity on the anticipation and experience of a painful thermal stimulus. PloS One, 12(8), e0183650.
Parrotta, E., Bach, P., Perrucci, M. G., Costantini, M., & Ferri, F. (2024). Heart is deceitful above all things: Threat expectancy induces the illusory perception of increased heartrate. Cognition, 245, 105719.
Pezzulo, G. (2014). Why do you fear the bogeyman? An embodied predictive coding model of perceptual inference. Cognitive, Affective & Behavioral Neuroscience, 14(3), 902-911.
Seth, A., Suzuki, K., & Critchley, H. (2012). An Interoceptive Predictive Coding Model of Conscious Presence. Frontiers in Psychology, 2. https://www.frontiersin.org/articles/10.3389/fpsyg.2011.00395
Seth, A. K. (2013). Interoceptive inference, emotion, and the embodied self. Trends in Cognitive Sciences, 17(11), 565-573.
Skora, L. I., Livermore, J. J. A., & Roelofs, K. (2022). The functional role of cardiac activity in perception and action. Neuroscience & Biobehavioral Reviews, 104655.
I will add that the control experiment - with an exteroceptive signal (knocking of wood) manipulated in a similar manner - could be seen as evidence of the fact that heartbeats are regarded as an interoceptive signal, and it is an important control experiment, however, to me it seems that what it is showing is the importance of human-relevant signals to pain prediction/perception, and not directly proves that it is considered interoceptive. For example, it could be experienced as a social cue of human anxiety/fear etc, and induce alertness.
The reviewer asks us to consider whether our measured changes in pain response happen not because the brain treats the heartrate feedback in Experiment 1 as interoceptive stimulus, but because heartbeat sounds could have signalled threat on a more abstract, perhaps metacognitive or affective, level, in contrast to the less visceral control sounds in Experiment 2. We deem this highly unlikely for several reasons.
First, as we point out in our response to Reviewer 3 (Point 3), if this were the case, the different sounds in both experiments should have induced overall (between-experiment) differences in pain perception and heart rate, induced by the (supposedly) generally more threatening heart beat sounds. However, when we added such comparisons, no such between-experiment differences were obtained (See Results Experiment 2, and Supplementary Materials, Cross-experiment analysis between-subjects model). Instead, we only find a significant interaction between experiment and feedback (faster, slower). Thus, it is not the heartbeat sounds per se that induce the measured changes to pain perception, but the modulation of their rate, and that identical changes to the rate of non-heartrate sounds produce no such effects. In other words, pain perception is sensitive to a change in heart rate feedback, as we predicted, instead of the overall presence of heartbeat sounds (as one would need to predict if heart beat sounds had more generally induced threat or stress).
Second, one may suspect that it is precisely the acceleration of heartrate feedback that could act as cue to arousal, while accelerated exteroceptive feedback would not. However, if this were the case, one would need to predict a general heart rate increase with accelerated feedback, as this is the general physiological marker of increasing alertness and arousal (e.g. Tousignant-Laflamme et al., 2005; Terkelsen et al., 2005; for a review, see Forte et al., 2022). However, the data shows the opposite, with real heartrates decreasing when the heartrate feedback increases. This result is again fully in line with the predicted interoceptive consequences of accelerated heartrate feedback, which mandates an immediate autonomic regulation, especially when preparing for an anticipated stressor.
Third, our view is further supported by neurophysiological evidence showing that heartbeat sounds, particularly under the belief they reflect one’s own body, are not processed merely as generic aversive or “human-relevant” signals. For instance, Vicentin et al. (2024) showed that simulated faster heartbeat sounds elicited stronger EEG alpha-band suppression, indicative of increased cortical activation over frontocentral and right frontal areas, compatible with the localization of brain regions contributing to interoceptive processes (Kleint et al., 2015). Importantly, Kleint et al. also demonstrated via fMRI that heartbeat sounds, compared to acoustically matched tones, selectively activate bilateral anterior insula and frontal operculum, key hubs of the interoceptive network. This suggests that the semantic identity of the sound as a heartbeat is sufficient to elicit internal body representations, despite its exteroceptive nature. Further evidence comes from van Elk et al. (2014), who found that heartbeat sounds suppress the auditory N1 component, a neural marker of sensory attenuation typically associated with self-generated or predicted stimuli. The authors interpret this as evidence that the brain treats heartbeat sounds as internally predicted bodily signals, supporting interoceptive predictive coding accounts in which exteroceptive cues (i.e., auditory cardiac feedback) are integrated with visceral information to generate coherent internal body representations.
Finally, it is worth noting that the manipulation of heartrate feedback in our study elicited measurable compensatory changes in participants’ actual heart rate. This is striking compared to our previous work (Parrotta et al., 2024), wherein we used a highly similar design as here, combined with a very strong threat manipulation. Specifically, we presented participants with highly salient threat cues (knives directed at an anatomical depiction of a heart), which predicted forthcoming pain with 100% validity (compared to flowers that did predict the absence of pain with 100%). In other words, these cues perfectly predicted actual pain, through highly visceral stimuli. Nevertheless, we found no measurable decrease in actual heartrate. From an abstract threat perspective, it is therefore striking that the much weaker manipulation of slightly increased or decreased heartrates we used here would induce such a change. The difference therefore suggests that what caused the response here is not due to an abstract feeling of threat, but because the brain indeed treated the increased heartrate feedback as an interoceptive signal for (stressor-induced) sympathetic activation, which would then be immediately down-regulated.
Together, we hope you agree that these considerations make a strong case against a non-specific, arousal or alertness-related explanation of our data. We now make this point clearer in the new paragraph of the Discussion (Accounting for general unspecific contributionslines 796-830), and have added the relevant between experiment comparisons to the Results of Experiment 2.
Forte, G., Troisi, G., Pazzaglia, M., Pascalis, V. D., & Casagrande, M. (2022). Heart rate variability and pain: a systematic review. Brain sciences, 12(2), 153.
Vicentin, S., Guglielmi, S., Stramucci, G., Bisiacchi, P., & Cainelli, E. (2024). Listen to the beat: behavioral and neurophysiological correlates of slow and fast heartbeat sounds. International Journal of Psychophysiology, 206, 112447.
Kleint, N. I., Wittchen, H. U., & Lueken, U. (2015). Probing the interoceptive network by listening to heartbeats: an fMRI study. PloS one, 10(7), e0133164.
Parrotta, E., Bach, P., Perrucci, M. G., Costantini, M., & Ferri, F. (2024). Heart is deceitful above all things: Threat expectancy induces the illusory perception of increased heartrate. Cognition, 245, 105719.
Terkelsen, A. J., Mølgaard, H., Hansen, J., Andersen, O. K., & Jensen, T. S. (2005). Acute pain increases heart rate: differential mechanisms during rest and mental stress. Autonomic Neuroscience, 121(1-2), 101-109.
Tousignant-Laflamme, Y., Rainville, P., & Marchand, S. (2005). Establishing a link between heart rate and pain in healthy subjects: a gender effect. The journal of pain, 6(6), 341-347.
van Elk, M., Lenggenhager, B., Heydrich, L., & Blanke, O. (2014). Suppression of the auditory N1-component for heartbeat-related sounds reflects interoceptive predictive coding. Biological psychology, 99, 172-182.
Several additional, more methodological weaknesses include the very small number of trials per condition - the methods mention 18 test trials per participant for the 3 conditions, with varying pain intensities, which are later averaged (and whether this is appropriate is a different issue). This means 6 trials per condition, and only 2 trials per condition and pain intensity. I thought that this number could be increased, though it is not a huge concern of the paper. It is, however, needed to show some statistics about the distribution of responses, given the very small trial number (see recommendations for authors). The sample size is also rather small, on the verge of "just right" to meet the required sample size according to the authors' calculations.
We provide detailed responses to these points in the “Recommendations for The Authors” section, where each of these issues is addressed point by point in response to the specific questions raised.
Finally, and just as important, the data exists to analyze participants' physiological responses (ECG) after receiving the painful stimulus - this could support the authors' claims about the change in both subjective and objective responses to pain. It could also strengthen the physiological evidence, which is rather weak in terms of its effect. Nevertheless, this is missing from the paper.
This is indeed an interesting point, and we agree that analyzing physiological responses such as ECG following the painful stimulus could offer additional insights into the objective correlates of pain. However, it is important to clarify that the experiment was not designed to investigate post-stimulus physiological responses. Our primary focus was on the anticipatory processes leading up to the pain event. Notably, in the time window immediately following the stimulus - when one might typically expect to observe physiological changes such as an increase in heart rate - participants were asked to provide subjective ratings of their nociceptive experience. It is therefore not a “clean” interval that would lend itself for measurement, especially as a substantial body of evidence indicates that one’s heart rate is strongly modulated by higher-order cognitive processes, including attentional control, executive functioning, decision-making and action itself (e.g., Forte et al., 2021a; Forte et al., 2021b; Luque-Casado et al., 2016).
This limitation is particularly important as the induced change in pain ratings by our heart rate manipulation is substantially smaller than the changes in heart rate induced by actual pain (e.g., Loggia et al., 2011). To confirm this for our study, we simply estimated how much change in heart rate is produced by a change in actual stimulus intensity in the initial no feedback phase of our experiment. There, we find that a change between stimulus intensities 2 and 4 induces a NPS change of 32.95 and a heart rate acceleration response of 1.19 (difference in heart rate response relative to baseline, Colloca et al., 2006), d = .52, p < .001. The change of NPS induced by our implicit heart rate manipulation, however, is only a seventh of this (4.81 on the NPS). This means that the expected effect size of heart rate acceleration produced by our manipulation would only be d = .17. A power analysis, using GPower, reveals that a sample size of n = 266 would be required to detect such an effect, if it exists. Thus, while we agree that this is an exciting hypothesis to be tested, it requires a specifically designed study, and a much larger sample than was possible here.
Colloca, L., Benedetti, F., & Pollo, A. (2006). Repeatability of autonomic responses to pain anticipation and pain stimulation. European Journal of Pain, 10(7), 659-665.
Forte, G., Morelli, M., & Casagrande, M. (2021a). Heart rate variability and decision-making: Autonomic responses in making decisions. Brain sciences, 11(2), 243.
Forte, G., Favieri, F., Oliha, E. O., Marotta, A., & Casagrande, M. (2021b). Anxiety and attentional processes: the role of resting heart rate variability. Brain sciences, 11(4), 480.
Loggia, M. L., Juneau, M., & Bushnell, M. C. (2011). Autonomic responses to heat pain: Heart rate, skin conductance, and their relation to verbal ratings and stimulus intensity. PAIN®, 152(3), 592-598.
Luque-Casado, A., Perales, J. C., Cárdenas, D., & Sanabria, D. (2016). Heart rate variability and cognitive processing: The autonomic response to task demands. Biological psychology, 113, 83-90
I have several additional recommendations regarding data analysis (using an ANOVA rather than multiple t-tests, using raw normalized data rather than change scores, questioning the averaging across 3 pain intensities) - which I will detail in the "recommendations for authors" section.
We provide detailed responses to these points in the “Recommendations for The Authors” section, where each of these issues is addressed point by point in response to the specific questions raised.
Conclusion:
To conclude, the authors have shown in their findings that predictions about an upcoming aversive (pain) stimulus - and its subsequent subjective perception - can be altered not only by external expectations, or manipulating the pain cue, as was done in studies so far, but also by manipulating a cue that has fundamental importance to human physiological status, namely heartbeats. Whether this is a manipulation of actual interoception as sensed by the brain is - in my view - left to be proven.
Still, the paper has important implications in several fields of science ranging from neuroscience prediction-perception research, to pain and placebo research, and may have implications for clinical disorders, as the authors propose. Furthermore, it may lead - either the authors or someone else - to further test this interesting question of manipulation of interoception in a different or more controlled manner.
I salute the authors for coming up with this interesting question and encourage them to continue and explore ways to study it and related follow-up questions.
We sincerely thank the reviewer for the thoughtful and encouraging feedback. We hope our responses to your points below convince you a bit more that what we are measuring does indeed capture interoceptive processes, but we of course fully acknowledge that additional measures - for example from brain imaging (or computational modelling, see Reviewer 3) - could further support our interpretation, and highlights in the Limitations and Future directions section.
Reviewer #2 (Public Review):
In this manuscript, Parrotta et al. tested whether it is possible to modulate pain perception and heart rate by providing false HR acoustic feedback before administering electrical cutaneous shocks. To this end, they performed two experiments. The first experiment tested whether false HR acoustic feedback alters pain perception and the cardiac anticipatory response. The second experiment tested whether the same perceptual and physiological changes are observed when participants are exposed to a non-interoceptive feedback. The main results of the first experiment showed a modulatory effect for faster HR acoustic feedback on pain intensity, unpleasantness, and cardiac anticipatory response compared to a control (acoustic feedback congruent to the participant's actual HR). However, the results of the second experiment also showed an increase in pain ratings for the faster non-interoceptive acoustic feedback compared to the control condition, with no differences in pain unpleasantness or cardiac response.
The main strengths of the manuscript are the clarity with which it was written, and its solid theoretical and conceptual framework. The researchers make an in-depth review of predictive processing models to account for the complex experience of pain, and how these models are updated by perceptual and active inference. They follow with an account of how pain expectations modulate physiological responses and draw attention to the fact that most previous studies focus on exteroceptive cues. At this point, they make the link between pain experience and heart rate changes, and introduce their own previous work showing that people may illusorily perceive a higher cardiac frequency when expecting painful stimulation, even though anticipating pain typically goes along with a decrease in HR. From here, they hypothesize that false HR acoustic feedback evokes more intense and unpleasant pain perception, although the actual HR actually decreases due to the orienting cardiac response. Furthermore, they also test the hypothesis that an exteroceptive cue will lead to no (or less) changes in those variables. The discussion of their results is also well-rooted in the existing bibliography, and for the most part, provides a credible account of the findings.
Thank you for the clear and thoughtful review. We appreciate your positive comments on the manuscript’s clarity, theoretical framework, and interpretation of results.
The main weaknesses of the manuscript lies in a few choices in methodology and data analysis that hinder the interpretation of the results and the conclusions as they stand.
The first peculiar choice is the convoluted definition of the outcomes. Specifically, pain intensity and unpleasantness are first normalized and then transformed into variation rates (sic) or deltas, which makes the interpretation of the results unnecessarily complicated. This is also linked to the definitions of the smallest effect of interest (SESOI) in terms of these outcomes, which is crucial to determining the sample size and gauging the differences between conditions. However, the choice of SESOI is not properly justified, and strangely, it changes from the first experiment to the second.
We thank the reviewer for this important observation. In the revised manuscript, we have made substantial changes and clarifications to address both aspects of this concern: (1) the definition of outcome variables and their normalization, and (2) the definition of the SESOI.
First, As explained in our response to Reviewer #1, we have revised the analyses and removed the difference-based change scores from the main results, addressing concerns about interpretability. However, we retained the normalization procedure: all variables (heart rate, pain intensity, unpleasantness) are normalized relative to the no-feedback baseline using a standard proportional change formula (X−bX)/bX(X - bX)/bX(X−bX)/bX, where X is the feedback-phase mean and bX is the no-feedback baseline. This is a widely used normalization procedure (e.g., Bartolo et al., 2013; Cecchini et al., 2020). This method controls for interindividual variability by expressing responses relative to each participant’s own baseline. The resulting normalized values are then used directly in all analyses, and not further transformed into deltas.
To address potential concerns about this baseline correction approach and its interpretability, we also conducted a new set of supplementary analyses (now reported in the supplementary materials) that include the no-feedback condition explicitly in the models, rather than treating it as a baseline for normalization. These models confirm that our main effects are not driven by the choice of normalization and hold even when no-feedback is analyzed as an independent condition. The new analyses and results are now reported in the Supplementary Materials.
Second, concerning the SESOI values and their justification: The difference in SESOI values between Experiment 1 and Experiment 2 reflects the outcome of sensitivity analyses conducted for each dataset separately, rather than a post-hoc reinterpretation of our results. Specifically, we followed current methodological recommendations (Anderson, Kelley & Maxwell, 2017; Albers & Lakens, 2017; Lakens, 2022), which advise against estimating statistical power based on previously published effect sizes, especially when working with novel paradigms or when effect sizes in the literature may be inflated or imprecise. Instead, we used the sensitivity analysis function in G*Power (Version 3.1) to determine the smallest effect size our design was capable of detecting with high statistical power (90%), given the actual sample size, test type, and alpha level used in each experiment. This is a prospective, design-based estimation rather than a post-hoc analysis of observed effects. The slight differences in SESOI are due to more participants falling below our exclusions criteria in Experiment 2, leading to slightly larger effect sizes that can be detected (d = 0.62 vs d = 0.57). Importantly, both experiments remain adequately powered to detect effects of a size commonly reported in the literature on top-down pain modulation. For instance, Iodice et al. (2019) reported effects of approximately d = 0.7, which is well above the minimum detectable thresholds of our designs.
We have now clarified the logic in the Participant section of Experiment 1 (193-218).
Anderson, S. F., Kelley, K., & Maxwell, S. E. (2017). Sample-Size Planning for More Accurate Statistical Power: A Method Adjusting Sample Effect Sizes for Publication Bias and Uncertainty. Psychological Science, 28(11), 1547-1562.
Bartolo, M., Serrao, M., Gamgebeli, Z., Alpaidze, M., Perrotta, A., Padua, L., Pierelli, F., Nappi, G., & Sandrini, G. (2013). Modulation of the human nociceptive flexion reflex by pleasant and unpleasant odors. PAIN®, 154(10), 2054-2059.
Cecchini, M. P., Riello, M., Sandri, A., Zanini, A., Fiorio, M., & Tinazzi, M. (2020). Smell and taste dissociations in the modulation of tonic pain perception induced by a capsaicin cream application. European Journal of Pain, 24(10), 1946-1955.
Lakens, D. (2022). Sample size justification. Collabra: psychology, 8(1), 33267.
Albers, C., & Lakens, D. (2018). When power analyses based on pilot data are biased: Inaccurate effect size estimators and follow-up bias. Journal of experimental social psychology, 74, 187-195.
Furthermore, the researchers propose the comparison of faster vs. slower delta HR acoustic feedback throughout the manuscript when the natural comparison is the incongruent vs. the congruent feedback.
We very much disagree that the natural comparison is congruent vs incongruent feedback. First, please note that congruency simply refers to whether the heartrate feedback was congruent with (i.e., matched) the participant’s heartrate measurements in the no feedback trials, or whether it was incongruent, and was therefore either faster or slower than this baseline frequency. As such, simply comparing congruent with incongruent feedback could only indicate that pain ratings change when the feedback does not match the real heart rate, irrespective of whether it is faster or slower. Such a test can therefore only reveal potential general effects of surprise or salience, when the feedback heartrate does not match the real one.
We therefore assume that the reviewer specifically refers to the comparison of congruent vs incongruent faster feedback. However, this is not a good test either, as this comparison is, by necessity, confounded with the factor of surprise described above. In other words, if a difference would be found, it would not be clear if it emerges because, as we assume, that faster feedback is represented as an interoceptive signal for threat, or simply because participants are surprised about heartrate feedback that diverges from their real heartrate. Note that even a non-significant result in the analogous comparison of congruent vs incongruent slower feedback would not be able to resolve this confound, as in null hypothesis testing the absence of a significant effect does, per definition, not indicate that there is no effect - only that it could not be detected here.
Instead, the only possible test of our hypothesis is the one we have designed our experiment around and focussed on with our central t-test: the comparison of incongruent faster with incongruent slower feedback. This keeps any possible effects of surprise/salience from generally altered feedback constant and allows us to test our specific hypothesis: that real heart rates will decrease and pain ratings will increase when receiving false interoceptive feedback about increased compared to decreasing heartrates. Note that this test of faster vs slower feedback is also statistically the most appropriate, as it collapses our prediction onto a single and highest-powered hypothesis test: As faster and slower heartrate feedback are assumed to induce effects in the opposite direction, the effect size of their difference is, per definition, double than the averaged effect size for the two separate tests of faster vs congruent feedback and slower vs congruent feedback.
That being said, we also included comparisons with the congruent condition in our revised analysis, in line with the reviewer’s suggestion and previous studies. These analyses help explore potential asymmetries in the effect of false feedback. While faster feedback (both interoceptive and exteroceptive) significantly modulated pain relative to congruent feedback, the slower feedback did not, consistent with previous literature showing stronger effects for arousal-increasing cues (e.g., Valins, 1966; Iodice et al., 2019). To address this point, in the revised manuscript we have added a paragraph to the Data Analysis section of Experiment 1 (lines 405-437) to make this logic clearer.
Valins, S. (1966). Cognitive effects of false heart-rate feedback. Journal of personality and social psychology, 4(4), 400.
Iodice, P., Porciello, G., Bufalari, I., Barca, L., & Pezzulo, G. (2019). An interoceptive illusion of effort induced by false heart-rate feedback. Proceedings of the National Academy of Sciences, 116(28), 13897-13902.
This could be influenced by the fact that the faster HR exteroceptive cue in experiment 2 also shows a significant modulatory effect on pain intensity compared to congruent HR feedback, which puts into question the hypothesized differences between interoceptive vs. exteroceptive cues. These results could also be influenced by the specific choice of exteroceptive cue: the researchers imply that the main driver of the effect is the nature of the cue (interoceptive vs. exteroceptive) and not its frequency. However, they attempt to generalize their findings using knocking wood sounds to all possible sounds, but it is possible that some features of these sounds (e.g., auditory roughness or loomingness) could be the drivers behind the observed effects.
We appreciate this thoughtful comment. We agree that low-level auditory features can potentially introduce confounds in the experimental design, and we acknowledge the importance of distinguishing these factors from the higher-order distinction that is central to our study: whether the sound is perceived as interoceptive (originating from within the body) or exteroceptive (perceived as external). To this end, the knocking sound was chosen not for its specific acoustic profile, but because it lacked bodily relevance, thus allowing us to test whether the same temporal manipulations (faster, congruent, slower) would have different effects depending on whether the cue was interpreted as reflecting an internal bodily state or not. In this context, the exteroceptive cue served as a conceptual contrast rather than an exhaustive control for all auditory dimensions.
Several aspects of our data make it unlikely that the observed effects are driven by unspecific acoustic characteristics of the sounds used in the exteroceptive and interoceptive experiments (see also our responses to Reviewer 1 and Reviewer 3 who raised similar points).
First, if the knocking sound had inherent acoustic features that strongly influenced perception or physiological responses, we would expect it to have produced consistent effects across all feedback conditions (Faster, Slower, Congruent), regardless of the interpretive context. This would have manifested as an overall difference between experiments in the between-subjects analyses and in the supplementary mixed-effects models that included Experiment as a fixed factor. Yet, we observed no such main effects in any of our variables. Instead, significant differences emerged only in specific theoretically predicted comparisons (e.g., Faster vs. Slower), and critically, these effects depended on the cue type (interoceptive vs. exteroceptive), suggesting that perceived bodily relevance, rather than a specific acoustic property, was the critical modulator. In other words, any alternative explanation based on acoustic features would need to be able to explain why these acoustic properties would induce not an overall change in heart rate and pain perception (i.e., similarly across slower, faster, and congruent feedback), but the brain’s response to changes in the rate of this feedback – increasing pain ratings and decreasing heartrates for faster relative to slower feedback. We hope you agree that a simple effect of acoustic features would not predict such a sensitivity to the rate with which the sound was played.
Please refer to our responses to Reviewers 1 and 2 for further aspects of the data, arguing strongly against other features associated with the sounds (e.g., alertness, arousal) could be responsible for the results, as the data pattern again goes in the opposite direction than that predicted by such accounts (e.g., faster heartrate feedback decreased real heartrate, instead of increasing them, as would be expected if accelerated heartrate feedback increased arousal).
Finally, to further support this interpretation, we refer to neurophysiological evidence showing that heartbeat sounds are not processed as generic auditory signals, but as internal, bodily relevant cues especially when believed to reflect one’s own physiological state. For instance, fMRI research (Kleint et al., 2015) shows that heartbeat sounds engage key interoceptive regions such as the anterior insula and frontal operculum more than acoustically matched control tones. EEG data (Vicentin et al., 2024) showed that faster heartbeat sounds produce stronger alpha suppression over frontocentral areas, suggesting enhanced processing in networks associated with interoceptive attention. Moreover, van Elk et al. (2014) found that heartbeat sounds attenuate the auditory N1 response, a neural signature typically linked to self-generated or predicted bodily signals. These findings consistently demonstrate that heartbeats sounds are processed as interoceptive and self-generated signals, which is in line with our rationale that the critical factor at play concern whether it is semantically perceived as reflecting one’s own bodily state, rather than the physical properties of the sound.
We now explicitly discuss these issues in the revised Discussion section (lines 740-758).
Kleint, N. I., Wittchen, H. U., & Lueken, U. (2015). Probing the interoceptive network by listening to heartbeats: an fMRI study. PloS one, 10(7), e0133164.
van Elk, M., Lenggenhager, B., Heydrich, L., & Blanke, O. (2014). Suppression of the auditory N1-component for heartbeat-related sounds reflects interoceptive predictive coding. Biological psychology, 99, 172-182.
Vicentin, S., Guglielmi, S., Stramucci, G., Bisiacchi, P., & Cainelli, E. (2024). Listen to the beat: behavioral and neurophysiological correlates of slow and fast heartbeat sounds. International Journal of Psychophysiology, 206, 112447.
Finally, it is noteworthy that the researchers divided the study into two experiments when it would have been optimal to test all the conditions with the same subjects in a randomized order in a single cross-over experiment to reduce between-subject variability. Taking this into consideration, I believe that the conclusions are only partially supported by the evidence. Despite of the outcome transformations, a clear effect of faster HR acoustic feedback can be observed in the first experiment, which is larger than the proposed exteroceptive counterpart. This work could be of broad interest to pain researchers, particularly those working on predictive coding of pain.
We appreciate the reviewer’s suggestion regarding a within-subject crossover design. While such a design indeed offers increased statistical power by reducing interindividual variability (Charness, Gneezy, & Kuhn, 2012), we intentionally opted for a between-subjects design due to theoretical and methodological considerations specific to studies involving deceptive feedback. Most importantly, carryover effects are a major concern in deception paradigms. Participants exposed to one type of feedback initially (e.g., interoceptive), and then the other (exteroceptive) would be more likely to develop suspicion or adaptive strategies that would alter their responses. Such expectancy effects could contaminate results in a crossover design, particularly when participants realize that feedback is manipulated. In line with this idea, past studies on false cardiac feedback (e.g., Valins, 1966; Pennebaker & Lightner, 1980) often employed between-subjects or blocked designs to mitigate this risk.
Pennebaker, J. W., & Lightner, J. M. (1980). Competition of internal and external information in an exercise setting. Journal of personality and social psychology, 39(1), 165.
Valins, S. (1966). Cognitive effects of false heart-rate feedback. Journal of personality and social psychology, 4(4), 400.
Reviewer #3 (Public Review):
In their manuscript titled "Exposure to false cardiac feedback alters pain perception and anticipatory cardiac frequency", Parrotta and colleagues describe an experimental study on the interplay between false heart rate feedback and pain experience in healthy, adult humans. The experimental design is derived from Bayesian perspectives on interoceptive inference. In Experiment 1 (N=34), participants rated the intensity and unpleasantness of an electrical pulse presented to their middle fingers. Participants received auditory cardiac feedback prior to the electrical pulse. This feedback was congruent with the participant's heart rate or manipulated to have a higher or lower frequency than the participant's true heart rate (incongruent high/ low feedback). The authors find heightened ratings of pain intensity and unpleasantness as well as a decreased heart rate in participants who were exposed to the incongruent-high cardiac feedback. Experiment 2 (N=29) is equivalent to Experiment 1 with the exception that non-interoceptive auditory feedback was presented. Here, mean pain intensity and unpleasantness ratings were unaffected by feedback frequency.
Strengths:
The authors present interesting experimental data that was derived from modern theoretical accounts of interoceptive inference and pain processing.
(1) The motivation for the study is well-explained and rooted within the current literature, whereas pain is the result of a multimodal, inferential process. The separation of nociceptive stimulation and pain experience is explained clearly and stringently throughout the text.
(2) The idea of manipulating pain-related expectations via an internal, instead of an external cue, is very innovative.
(3) An appropriate control experiment was implemented, where an external (non-physiological) auditory cue with parallel frequency to the cardiac cue was presented.
(4) The chosen statistical methods are appropriate, albeit averaging may limit the opportunity for mechanistic insight, see weaknesses section.
(5) The behavioral data, showing increased unpleasantness and intensity ratings after exposure to incongruent-high cardiac feedback, but not exteroceptive high-frequency auditory feedback, is backed up by ECG data. Here, the decrease in heart rate during the incongruent-high condition speaks towards a specific, expectation-induced physiological effect that can be seen as resulting from interoceptive inference.
We thank the reviewer for their positive feedback. We are glad that the study’s theoretical foundation, innovative design, appropriate control conditions, and convergence of behavioral and physiological data were well received.
Weaknesses:
Additional analyses and/ or more extensive discussion are needed to address these limitations:
(1) I would like to know more about potential learning effects during the study. Is there a significant change in ∆ intensity and ∆ unpleasantness over time; e.g. in early trials compared to later trials? It would be helpful to exclude the alternative explanation that over time, participants learned to interpret the exteroceptive cue more in line with the cardiac cue, and the effect is driven by a lack of learning about the slightly less familiar cue (the exteroceptive cue) in early trials. In other words, the heartbeat-like auditory feedback might be "overlearned", compared to the less naturalistic tone, and more exposure to the less naturalistic cue might rule out any differences between them w.r.t. pain unpleasantness ratings.
We thank the reviewer for raising this important point. Please note that the repetitions in our task were relatively limited (6 trials per condition), which limits the potential influence of such differential learning effects between experiments. To address this concern, we performed an additional analysis, reported in the Supplementary Materials, using a Linear Mixed-Effects Model approach. This method allowed us to include "Trial" (the rank order of each trial) as a variable to account for potential time-on-task effects such as learning, adaptation, or fatigue (e.g., Möckel et al., 2015). All feedback conditions (no-feedback, congruent, faster, slower) and all stimulus intensity levels were included.
Specifically, we tested the following models:
Likert Pain Unpleasantness Ratings ~ Experiment × Feedback × StimInt × Trial + (StimInt + Trial | Subject)
Numeric Pain Scale of Intensity Ratings ~ Experiment × Feedback × StimInt × Trial + (StimInt + Trial | Subject)
In both models, no significant interactions involving Trial × Experiment or Trial × Feedback × Experiment were found. Instead, we just find generally larger effects in early trials compared to later ones (Main effect of Trial within each Experiment), similar to other cognitive illusions where repeated exposure diminishes effects. Thus, although some unspecific changes over time may have occurred (e.g., due to general task exposure), these changes did not differ systematically across experimental conditions (interoceptive vs. exteroceptive) or feedback types. However, we are fully aware that the absence of significant higher-order interactions does not conclusively rule out the possibility of learning-related effects. It is possible that our models lacked the statistical power to detect more subtle or complex time-dependent modulations, particularly if such effects differ in magnitude or direction across feedback conditions.
We report the full description of these analyses and results in the Supplementary materials 1. Cross-experiment analysis (between-subjects model).
(2) The origin of the difference in Cohen's d (Exp. 1: .57, Exp. 2: .62) and subsequently sample size in the sensitivity analyses remains unclear, it would be helpful to clarify where these values are coming from (are they related to the effects reported in the results? If so, they should be marked as post-hoc analyses).
Following recommendations (Anderson, Kelley & Maxwell, 2017; Albers & Lakens, 2017), we do not report theoretical power based on previously reported effect sizes as this neglects uncertainty around effect size measurements, especially for new effects for which no reliable expected effect size estimates can be derived across the literature. Instead, the power analysis is based on a sensitivity analysis, conducted in G*Power (Version 3.1). Importantly, these are not post-hoc analyses, as they are not based on observed effect sizes in our study, but derived a priori. Sensitivity analyses estimate effect sizes that our design is well-powered (90%) to detect (i.e. given target power, sample size, type of test), for the crucial comparison between faster and slower feedback in both experiments (Lakens, 2022). Following recommendations, we also report the smallest effect size this test can in principle detect in our study (SESOI, Lakens, 2022). This yields effect sizes of d = .57 in Experiment 1 and d = .62 in Experiment 2 at 90% power and SESOIs of d = .34 and .37, respectively. Note that values are slightly higher in Experiment 2, as more participants were excluded based on our exclusion criteria. Importantly, detectable effect sizes in both experiments are smaller than reported effect sizes for comparable top-down effects on pain measurements of d = .7 (Iodice et al., 2019). We have now added more information to the power analysis sections to make this clearer (lines 208-217).
Albers, C., & Lakens, D. (2018). When power analyses based on pilot data are biased: Inaccurate effect size estimators and follow-up bias. Journal of experimental social psychology, 74, 187-195.
Anderson, S. F., Kelley, K., & Maxwell, S. E. (2017). Sample-Size Planning for More Accurate Statistical Power: A Method Adjusting Sample Effect Sizes for Publication Bias and Uncertainty. Psychological Science, 28(11), 1547-1562.
Lakens, D. (2022). Sample size justification. Collabra: psychology, 8(1), 33267.
(3) As an alternative explanation, it is conceivable that the cardiac cue may have just increased unspecific arousal or attention to a larger extent than the exteroceptive cue. It would be helpful to discuss the role of these rather unspecific mechanisms, and how it may have differed between experiments.
We thank the reviewer for raising this important point. We agree that, in principle, unspecific mechanisms such as increased arousal or attention driven by cardiac feedback could be an alternative explanation for the observed effects. However, several aspects of our data indicate that this is unlikely:
(1) No main effect of Experiment on pain ratings:
If the cardiac feedback had simply increased arousal or attention in a general (non-specific) way, we would expect a main effect of Experiment (i.e., interoceptive vs exteroceptive condition) on pain intensity or unpleasantness ratings, regardless of feedback frequency. However, such a main effect was never observed when we compared between experiments (see between-experiment t-tests in results, and in supplementary analyses). Instead, effects were specific to the manipulation of feedback frequency.
(2) Heart rate as an arousal measure:
Heart rate (HR) is a classical physiological index of arousal. If there had been an unspecific increase in arousal in the interoceptive condition, we would expect a main effect of Experiment on HR. However, no such main effect was found. Instead, our HR analyses revealed a significant interaction between feedback and experiment, suggesting that HR changes depended specifically on the feedback manipulation rather than reflecting a general arousal increase.
(3) Arousal predicts faster, not slower, heart rates
In Experiment 1, faster interoceptive cardiac feedback led to a slowdown in heartrates both when compared to slower feedback and to congruent cardiac feedback. This is in line with the predicted compensatory response to faster heart rates. In contrast, if faster feedback would have only generally increased arousal, heart rates should have increased instead of decreased, as indicated by several prior studies (Tousignant-Laflamme et al., 2005; Terkelsen et al., 2005; for a review, see Forte et al., 2022), predicting the opposite pattern of responses than was found in Experiment 1.
Taken together, these findings indicate that the effects observed are unlikely to be driven by unspecific arousal or attention mechanisms, but rather are consistent with feedback-specific modulations, in line with our interoceptive inference framework.
We have now integrated these considerations in the revised discussion (lines 796-830), and added the relevant between-experiment comparisons to the Results of Experiment 2 and the supplementary analysis.
Terkelsen, A. J., Mølgaard, H., Hansen, J., Andersen, O. K., & Jensen, T. S. (2005). Acute pain increases heart rate: differential mechanisms during rest and mental stress. Autonomic Neuroscience, 121(1-2), 101-109.
Tousignant-Laflamme, Y., Rainville, P., & Marchand, S. (2005). Establishing a link between heart rate and pain in healthy subjects: a gender effect. The journal of pain, 6(6), 341-347.
Forte, G., Troisi, G., Pazzaglia, M., Pascalis, V. D., & Casagrande, M. (2022). Heart rate variability and pain: a systematic review. Brain sciences, 12(2), 153.
(4) The hypothesis (increased pain intensity with incongruent-high cardiac feedback) should be motivated by some additional literature.
We thank the reviewer for this helpful suggestion. Please note that the current phenomenon was tested in this experiment for the first time. Therefore, there is no specific prior study that motivated our hypotheses; they were driven theoretically, and derived from our model of interoceptive integration of pain and cardiac perception. The idea that accelerated cardiac feedback (relative to decelerated feedback) will increase pain perception and reduce heart rates is grounded on Embodied Predictive coding frameworks. Accordingly, expectations and signals from different sensory modalities (sensory, proprioceptive, interoceptive) are integrated both to efficiently infer crucial homeostatic and physiological variables, such as hunger, thirst, and, in this case, pain, and regulate the body’s own autonomic responses based on these inferences.
Within this framework, the concept of an interoceptive schema (Tschantz et al., 2022; Iodice et al., 2019; Parrotta et al., 2024; Schoeller et al., 2022) offers the basis for understanding interoceptive illusions, wherein inferred levels of interoceptive states (i.e., pain) deviate from the actual physiological state. Cardiac signals conveyed by the feedback manipulation act as a misleading prior, shaping the internal generative model of pain. Specifically, an increased heart rate may signal a state of threat, establishing a prior expectation of heightened pain. Building on predictive models of interoception, we predict that this cardiac prior is integrated with interoceptive (i.e., actual nociceptive signal) and exteroceptive inputs (i.e., auditory feedback input), leading to a subjective experience of increased pain even when there is no corresponding increase in the nociceptive input.
This idea is not completely new, but it is based on our previous findings of an interoceptive cardiac illusion driven by misleading priors about anticipated threat (i.e., pain). Specifically, in Parrotta et al. (2024), we tested whether a common false belief that heart rate increases in response to threat lead to an illusory perception of accelerated cardiac activity when anticipating pain. In two experiments, we asked participants to monitor and report their heartbeat while their ECG was recorded. Participants performed these tasks while visual cues reliably predicted a forthcoming harmless (low-intensity) vs. threatening (high-intensity) cutaneous electrical stimulus. We showed that anticipating a painful vs. harmless stimulus causes participants to report an increased cardiac frequency, which does not reflect their real cardiac response, but the common (false) belief that heart rates would accelerate under threat, reflecting the hypothesised integration of prior expectations and interoceptive inputs when estimating cardiac activity.
Here we tested the counterpart of such a cardiac illusion. We reasoned that if cardiac interoception is shaped by expectations about pain, then the inverse should also be true: manipulating beliefs about cardiac activity (via cardiac feedback) in the context of pain anticipation should influence the perception of pain. Specifically, we hypothesized that presenting accelerated cardiac feedback would act as a misleading prior, leading to an illusory increase in pain experience, even in the absence of an actual change in nociceptive input.
Moreover, next to the references already provided in the last version of the manuscript, there is ample prior research that provides more general support for such relationships. Specifically, studies have shown that providing mismatched cardiac feedback in contexts where cardiovascular changes are typically expected (i.e. sexual arousal, Rupp & Wallen, 2008; Valins, 1996; physical exercise, Iodice et al., 2019) can enhance the perception of interoceptive states associated with those experiences. Furthermore, findings that false cardiac feedback can influence emotional experience suggest that it is the conscious perception of physiological arousal, combined with the cognitive interpretation of the stimulus, that plays a key role in shaping emotional responses (Crucian et al., 2000).
This point is now addressed in the revised Introduction, wherein additional references have been integrated (lines 157-170).
Crucian, G. P., Hughes, J. D., Barrett, A. M., Williamson, D. J. G., Bauer, R. M., Bowers, D., & Heilman, K. M. (2000). Emotional and physiological responses to false feedback. Cortex, 36(5), 623-647.
Iodice, P., Porciello, G., Bufalari, I., Barca, L., & Pezzulo, G. (2019). An interoceptive illusion of effort induced by false heart-rate feedback. Proceedings of the National Academy of Sciences, 116(28), 13897-13902.
Parrotta, E., Bach, P., Perrucci, M. G., Costantini, M., & Ferri, F. (2024). Heart is deceitful above all things: Threat expectancy induces the illusory perception of increased heartrate. Cognition, 245, 105719.
Rupp, H. A., & Wallen, K. (2008). Sex differences in response to visual sexual stimuli: A review. Archives of sexual behavior, 37(2), 206-218.
Schoeller, F., Horowitz, A., Maes, P., Jain, A., Reggente, N., Moore, L. C., Trousselard, M., Klein, A., Barca, L., & Pezzulo, G. (2022). Interoceptive technologies for clinical neuroscience.
Tschantz, A., Barca, L., Maisto, D., Buckley, C. L., Seth, A. K., & Pezzulo, G. (2022). Simulating homeostatic, allostatic and goal-directed forms of interoceptive control using active inference. Biological Psychology, 169, 108266.
Valins, S. (1966). Cognitive effects of false heart-rate feedback. Journal of personality and social psychology, 4(4), 400.
(5) The discussion section does not address the study's limitations in a sufficient manner. For example, I would expect a more thorough discussion on the lack of correlation between participant ratings and self-reported bodily awareness and reactivity, as assessed with the BPQ.
We thank the reviewer for this valuable observation. In response, we have revised the Discussion section to explicitly acknowledge and elaborate on the lack of significant correlations between participants’ pain ratings and their self-reported bodily awareness and reactivity as assessed with the BPQ.
We now clarify that the inclusion of this questionnaire was exploratory. While it would be theoretically interesting to observe a relationship between subjective pain modulation and individual differences in interoceptive awareness, detecting robust correlations between within-subject experimental effects and between-subjects trait measures such as the BPQ typically requires much larger sample sizes (often exceeding N = 200) due to the inherently low reliability of such cross-level associations (see Hedge, Powell & Sumner, 2018; the “reliability paradox”). As such, the absence of a significant correlation in our study does not undermine the conclusions we draw from our main findings. Future studies with larger samples will be needed to systematically address this question. We now acknowledge this point explicitly in the revised manuscript (lines 501-504; 832-851).
Hedge, C., Powell, G., & Sumner, P. (2018). The reliability paradox: Why robust cognitive tasks do not produce reliable individual differences. Behavior Research Methods, 50(3), 1166-1186. https://doi.org/10.3758/s13428-017-0935-1
(a) Some short, additional information on why the authors chose to focus on body awareness and supradiaphragmatic reactivity subscales would be helpful.
We chose to focus on the body awareness and supradiaphragmatic reactivity subscales because these aspects are closely tied to emotional and physiological processing, particularly in the context of interoception. Body awareness plays a critical role in how individuals perceive and interpret bodily signals, which in turn affects emotional regulation and self-awareness. Supradiaphragmatic reactivity refers specifically to organs located or occurring above the diaphragm (i.e., the muscle that separates the chest cavity from the abdomen), which includes the heart, compared to subdiaphragmatic reactivity subscales further down. Our decision to include these subscales is further motivated by recent research, including the work by Petzschner et al. (2021), which demonstrates that the focus of attention can modulate the heartbeat-evoked potential (HEP), and that this modulation is predicted by participants’ responses on the supradiaphragmatic reactivity subscales. Thus, this subscale, and the more general body awareness scale, allows us to explore the interplay between bodily awareness, physiological reactivity, and emotional processing in our study. We now clarify this point in the revised version of the Methods - Body Perception Questionnaire (lines 384-393).
(6) The analyses presented in this version of the manuscript allow only limited mechanistic conclusions - a computational model of participants' behavior would be a very strong addition to the paper. While this may be out of the scope of the article, it would be helpful for the reader to discuss the limitations of the presented analyses and outline avenues towards a more mechanistic understanding and analysis of the data. The computational model in [7] might contain some starting ideas.
Thank you for your valuable feedback. We agree that a computational model would enhance the mechanistic understanding of our findings. While this is beyond the current scope, we now discuss the limitations of our analysis in the Limitations and Future directions section (lines 852-863). Specifically, we acknowledge that future studies could use computational models to better understand the interactions between physiological, cognitive, and perceptual factors.
Some additional topics were not considered in the first version of the manuscript:
(1) The possible advantages of a computational model of task behavior should be discussed.
We agree that a computational model of task behavior could provide several advantages. By formalizing principles of predictive processing and active inference, such a model could generate quantitative predictions about how heart rate (HR) and feedback interact, providing a more precise understanding of their respective contributions to pain modulation. However, this is a first demonstration of a theoretically predicted phenomenon, and computationally modelling it is currently outside the scope of the article. We would be excited to explore this in the future. We have added a brief discussion of these potential advantages in the revised manuscript and suggest that future work could integrate computational modelling to further deepen our understanding of these processes (lines 852-890).
(2) Across both experiments, there was a slightly larger number of female participants. Research suggests significant sex-related differences in pain processing [1,2]. It would be interesting to see what role this may have played in this data.
Thank you for your insightful comment. While we acknowledge that sex-related differences in pain processing are well-documented in the literature, we do not have enough participants in our sample to test this in a well-powered way. As such, exploring the role of sex differences in pain perception will need to be addressed in future studies with more balanced samples. It would be interesting if more sensitive individuals, with a more precise representation of pain, also show smaller effects on pain perception. We have noted this point in the revised manuscript (lines 845-851) and suggest that future research could specifically investigate how sex differences might influence the modulation of pain and physiological responses in similar experimental contexts.
(3) There are a few very relevant papers that come to mind which may be of interest. These sources might be particularly useful when discussing the roadmap towards a mechanistic understanding of the inferential processes underlying the task responses [3,4] and their clinical implications.
Thank you for highlighting these relevant papers. We appreciate your suggestion and have now cited them in the Limitations and Future directions paragraph (lines 852-863).
(4) In this version of the paper, we only see plots that illustrate ∆ scores, averaged across pain intensities - to better understand participant responses and the relationship with stimulus intensity, it would be helpful to see a more descriptive plot of task behavior (e.g. stimulus intensity and raw pain ratings)
To directly address the reviewer’s request, we now provide additional descriptive plots in the supplementary material of the revised manuscript, showing raw pain ratings across different stimulus intensities and feedback conditions. These plots offer a clearer view of participant behavior without averaging across pain levels, helping to better illustrate the relationship between stimulus intensity and reported pain.
Mogil, J. S. (2020). Qualitative sex differences in pain processing: emerging evidence of a biased literature. Nature Reviews Neuroscience, 21(7), 353-365. https://www.nature.com/articles/s41583-020-0310-6
Sorge, R. E., & Strath, L. J. (2018). Sex differences in pain responses. Current Opinion in Physiology, 6, 75-81. https://www.sciencedirect.com/science/article/abs/pii/S2468867318300786?via%3Dihub
Unal, O., Eren, O. C., Alkan, G., Petzschner, F. H., Yao, Y., & Stephan, K. E. (2021). Inference on homeostatic belief precision. Biological Psychology, 165, 108190.
Allen, M., Levy, A., Parr, T., & Friston, K. J. (2022). In the body's eye: the computational anatomy of interoceptive inference. PLoS Computational Biology, 18(9), e1010490.
Stephan, K. E., Manjaly, Z. M., Mathys, C. D., Weber, L. A., Paliwal, S., Gard, T., ... & Petzschner, F. H. (2016). Allostatic self-efficacy: A metacognitive theory of dyshomeostasis-induced fatigue and depression. Frontiers in human neuroscience, 10, 550.
Friston, K. J., Stephan, K. E., Montague, R., & Dolan, R. J. (2014). Computational psychiatry: the brain as a phantastic organ. The Lancet Psychiatry, 1(2), 148-158.
Eckert, A. L., Pabst, K., & Endres, D. M. (2022). A Bayesian model for chronic pain. Frontiers in Pain Research, 3, 966034.
We thank the reviewer for highlighting these relevant references which have now been integrated in the revised version of the manuscript.
Recommendations For The Authors:
Reviewer #1 (Recommendations For The Authors):
At the time I was reviewing this paper, I could not think of a detailed experiment that would answer my biggest concern: Is this a manipulation of the brain's interoceptive data integration, or rather a manipulation of participants' alertness which indirectly influences their pain prediction?
One incomplete idea that came to mind was delivering this signal in a more "covert" manner (though I am not sure it will suffice), or perhaps correlating the effect size of a participant with their interoceptive abilities, as measured in a different task or through a questionnaire.... Another potential idea is to tell participants that this is someone else's HR that they hear and see if that changes the results (though requires further thought). I leave it to the authors to think further, and perhaps this is to be answered in a different paper - but if so, I am sorry to say that I do not think the claims can remain as they are now, and the paper will need a revision of its arguments, unfortunately. I urge the authors to ask further questions if my point about the concern was not made clear enough for them to address or contemplate it.
We thank the reviewer for raising this important point. As detailed in our previous response, this point invites an important clarification regarding the role of cardiac deceleration in threat processing. Rather than serving as an interoceptive input from which the brain infers the likelihood of a forthcoming aversive event, heart rate deceleration is better described as an output of an already ongoing predictive process, as it reflects an allostatic adjustment of the bodily state aimed at minimizing the impact of the predicted perturbation (e.g., pain) and preventing sympathetic overshoot. It would be maladaptive for the brain to use a decelerating heart rate as evidence of impending threat, since this would paradoxically trigger further parasympathetic activation, initiating a potentially destabilizing feedback loop. Conversely, increased heart rate represents an evolutionarily conserved cue for arousal, threat, and pain. Our results therefore align with the idea that the brain treats externally manipulated increases in cardiac signals as congruent with anticipated sympathetic activation, prompting a compensatory autonomic and perceptual response consistent with embodied predictive processing frameworks (e.g., Barrett & Simmons, 2015; Seth, 2013).
We would also like to re-iterate that our results cannot be explained by general differences induced by the different heart rate sounds relative to the exteroceptive (see also our detailed comments to your point above, and our response to a similar point from Reviewer 3), for three main reasons.
(1) No main effect of Experiment on pain ratings:
If the cardiac feedback had simply increased arousal or attention in a general (non-specific) way, we would expect a main effect of Experiment (i.e., interoceptive vs exteroceptive condition) on pain intensity or unpleasantness ratings, regardless of feedback frequency. However, such a main effect was never observed. Instead, effects were specific to the manipulation of feedback frequency.
(2) Heart rate as an arousal measure:
Heart rate (HR) is a classical physiological index of arousal. If there had been an unspecific increase in arousal in the interoceptive condition, we would expect a main effect of Experiment on HR. However, no such main effect was found. Instead, our HR analyses revealed a significant interaction between feedback and experiment, suggesting that HR changes depended specifically on the feedback manipulation rather than reflecting a general arousal increase.
(3) Arousal predicts faster, not slower, heart rates
In Experiment 1, faster interoceptive cardiac feedback led to a slowdown in heartrates both when compared to slower feedback and to congruent cardiac feedback. This is in line with the predicted compensatory response to faster heart rates. In contrast, if faster feedback would have only generally increased arousal, heart rates should have increased instead of decreased, as indicated by several prior studies (for a review, see Forte et al., 2022), predicting the opposite pattern of responses than was found in Experiment 1.
Taken together, these findings indicate that the effects observed are unlikely to be driven by unspecific arousal or attention mechanisms, but rather are consistent with feedback-specific modulations, in line with our interoceptive inference framework. We now integrate these considerations in the general discussion (lines 796-830).
Barrett, L. F., & Simmons, W. K. (2015). Interoceptive predictions in the brain. Nature reviews neuroscience, 16(7), 419-429.
Forte, G., Troisi, G., Pazzaglia, M., Pascalis, V. D., & Casagrande, M. (2022). Heart rate variability and pain: a systematic review. Brain sciences, 12(2), 153.
Seth, A. K. (2013). Interoceptive inference, emotion, and the embodied self. Trends in Cognitive Sciences, 17(11), 565-573.
Additional recommendations:
Major (in order of importance):
(1) Number of trials per participant, per condition: as I mentioned, having only 6 trials for each condition is very little. The minimum requirement to accept so few trials would be to show data about the distribution of participants' responses to these trials, both per pain intensity (which was later averaged across - another issue discussed later), and across pain intensities, and see that it allows averaging across and that it is not incredibly variable such that the mean is unreliable.
We appreciate the reviewer’s concern regarding the limited number of trials per condition. This choice was driven by both theoretical and methodological considerations.
First, as is common in body illusion paradigms (e.g., the Rubber Hand Illusion, Botvinick & Cohen, 1998; the Full Body Illusion, Ehrsson, 2007; the Cardio-visual full body illusion, Pratviel et al., 2022) only a few trials are typically employed due to the immediate effects these manipulations elicit. Repetition can reduce the strength of the illusion through habituation, increased awareness, or loss of believability.
Second, the experiment was already quite long (1.5h to 2h per participant) and cognitively demanding. It would not have been feasible to expand it further without compromising data quality due to fatigue, attentional decline, or participant disengagement.
Third, the need for a large number of trials is more relevant when using implicit measures such as response times or physiological indices, which are typically indirectly related to the psychological constructs of interest. In contrast, explicit ratings are often more sensitive and less noisy, and thus require fewer repetitions to yield reliable effects (e.g., Corneille et al., 2024).
Importantly, we also addressed your concern analytically. We ran therefore linear mixed-effects model analyses across all dependent variables (See Supplementary materials), with Trial (i.e., the rank order of each trial) included as a predictor to account for potential time-on-task effects such as learning, adaptation, or fatigue (e.g., Möckel et al., 2015). These models captured trial-by-trial variability and allowed us to test for systematic changes in heart rate (HR) and pain ratings including interactions with feedback conditions (e.g., Klieg et al., 2011; Baayen et al., 2010; Ambrosini et al., 2019). The consistent effects of Trial suggest that repetition dampens the illusion, reinforcing our decision to limit the number of exposures.
In the interoceptive experiment, these analyses revealed a significant Feedback × Trial interaction (F(3, 711.19) = 6.16, p < .001), indicating that the effect of feedback on HR was not constant over time. As we suspected, and in line with other illusion-like effects, the difference between Faster and Slower feedback, which was significant early on (estimate = 1.68 bpm, p = .0007), decreased by mid-session (estimate = 0.69 bpm, p = .0048), and was no longer significant in later trials (estimate = 0.30 bpm, p = .4775). At the end of the session, HR values in the Faster and Slower conditions even numerically converged (Faster: M = 74.4, Slower: M = 74.1), and the non-significant contrast confirms that the difference had effectively vanished (for further details about slope estimation, see Supplementary material).
The same pattern emerged for pain-unpleasantness ratings. A significant Feedback × Trial interaction (F (3, 675.33) = 3.44, p = .0165) revealed that the difference between Faster and Slower feedback was strongest at the beginning of the session and progressively weakened. Specifically, Faster feedback produced higher unpleasantness than Slower in early trials (estimate= -0.28, p = .0058) and mid-session (estimate = - 0.19, p = .0001), but this contrast was no longer significant in the final trials, wherein all the differences between active feedback conditions vanished (all ps > .55).
Finally, similar results were yielded for pain intensity ratings. A significant Feedback × Trial interaction (F (3, 669.15) = 9.86, p < .001) showed that the Faster vs Slower difference was greatest at the start of the session and progressively vanished over trials. In early trials Faster feedback exceeded Slower (estimate=-8.33, p = .0001); by mid-session this gap had shrunk to 4.48 points (p < .0001); and in the final trials it was no longer significant (all ps > .94).
Taken together, our results show that the illusion induced by Faster relative to slower feedback fades with repetition; adding further trials would likely have masked this key effect, confirming the methodological choice to restrict each condition to fewer exposures. To conclude, given that this is the first study to investigate an illusion of pain using heartbeat-based manipulation, we intentionally limited repeated exposures to preserve the integrity of the illusion. The use of mixed models as complementary analyses strengthens the reliability of our conclusions within these necessary design constraints. We now clarify this point in the Procedure paragraph (lines 328-335)
Ambrosini, E., Peressotti, F., Gennari, M., Benavides-Varela, S., & Montefinese, M. (2023). Aging-related effects on the controlled retrieval of semantic information. Psychology and Aging, 38(3), 219.
Baayen, R. H., & Milin, P. (2010). Analyzing reaction times. International Journal of Psychological Research, 3(2), 12-28.
Botvinick, M., & Cohen, J. (1998). Rubber hands ‘feel’touch that eyes see. Nature, 391(6669), 756-756.
Corneille, O., & Gawronski, B. (2024). Self-reports are better measurement instruments than implicit measures. Nature Reviews Psychology, 3(12), 835–846.
Ehrsson, H. H. (2007). The experimental induction of out-of-body experiences. Science, 317(5841), 1048-1048.
Kliegl, R., Wei, P., Dambacher, M., Yan, M., & Zhou, X. (2011). Experimental effects and individual differences in linear mixed models: Estimating the relation of spatial, object, and attraction effects in visual attention. Frontiers in Psychology, 1, 238. https://doi.org/10.3389/fpsyg.2010.00238
Möckel, T., Beste, C., & Wascher, E. (2015). The effects of time on task in response selection-an ERP study of mental fatigue. Scientific reports, 5(1), 10113.
Pratviel, Y., Bouni, A., Deschodt-Arsac, V., Larrue, F., & Arsac, L. M. (2022). Avatar embodiment in VR: Are there individual susceptibilities to visuo-tactile or cardio-visual stimulations?. Frontiers in Virtual Reality, 3, 954808.
(2) Using different pain intensities: what was the purpose of training participants on correctly identifying pain intensities? You state that the aim of having 5 intensities is to cause ambiguity. What is the purpose of making sure participants accurately identify the intensities? Also, why then only 3 intensities were used in the test phase? The rationale for these is lacking.
We thank the reviewer for raising these important points regarding the use of different pain intensities. The purpose of using five levels during the calibration and training phases was to introduce variability and increase ambiguity in the participants’ sensory experience. This variability aimed to reduce predictability and prevent participants from forming fixed expectations about stimulus intensity, thereby enhancing the plausibility of the illusion. It also helped prevent habituation to a single intensity and made the manipulation subtler and more credible. We had no specific theoretical hypotheses about this manipulation. Regarding the accuracy training, although the paradigm introduced ambiguity, it was important to ensure that participants developed a stable and consistent internal representation of the pain scale. This step was essential to control for individual differences in sensory discrimination and to ensure that illusion effects were not confounded by participants’ inability to reliably distinguish between intensities.
As for the use of only three pain intensities in the test phase, the rationale was to focus on a manageable subset that still covered a meaningful range of the stimulus spectrum. This approach followed the same logic as Iodice et al. (2019, PNAS), who used five (rather than all seven) intensity levels during their experimental session. Specifically, they excluded the extreme levels (45 W and 125 W) used during baseline, to avoid floor and ceiling effects and to ensure that each test intensity could be paired with both a “slower” and a “faster” feedback from an adjacent level. This would not have been possible at the extremes of the intensity range, where no adjacent level exists in one direction. We adopted the same strategy to preserve the internal consistency and plausibility of our feedback manipulation.
We further clarified these points in the revised manuscript (lines 336-342).
Iodice, P., Porciello, G., Bufalari, I., Barca, L., & Pezzulo, G. (2019). An interoceptive illusion of effort induced by false heart-rate feedback. Proceedings of the National Academy of Sciences, 116(28), 13897-13902.
(3) Averaging across pain intensities: this is, in my opinion, not the best approach as by matching a participant's specific responses to a pain stimulus before and after the manipulation, you can more closely identify changes resulting from the manipulation. Nevertheless, the minimal requirement to do so is to show data of distributions of pain intensities so we know they did not differ between conditions per participant, and in general - as you indicate they were randomly distributed.
We thank the reviewer for this thoughtful comment. The decision to average across pain intensities in our main analyses was driven by the specific aim of the study: we did not intend to determine at which exact intensity level the illusion was most effective, and the limited number of trials makes such an analysis difficult. Rather, we introduced variability in nociceptive input to increase ambiguity and reduce predictability in the participants’ sensory experience. This variability was critical for enhancing the plausibility of the illusion by preventing participants from forming fixed expectations about stimulus strength. Additionally, using a range of intensities helped to minimize habituation effects and made the feedback manipulation subtler and more credible.
That said, we appreciate the reviewer’s point that matching specific responses before and after the manipulation at each intensity level could provide further insights into how the illusion operates across varying levels of nociceptive input. We therefore conducted supplementary analyses using linear mixed-effects models in which all three stimulus intensities were included as a continuous fixed factor. This allowed us to examine whether the effects of feedback were intensity-specific or generalized across different levels of stimulation
These analyses revealed that, in both the interoceptive and exteroceptive experiments, the effect of feedback on pain ratings was significantly modulated by stimulus intensity, as indicated by a Feedback × Stimulus Intensity interaction (Interoceptive: unpleasantness F(3, 672.32)=3.90, p=.0088; intensity ratings F(3, 667.07)=3.46, p=.016. Exteroceptive: unpleasantness F(3, 569.16)=8.21, p<.0001; intensity ratings F(3, 570.65)=3.00, p=.0301). The interaction term confirmed that the impact of feedback varied with stimulus strength, yet the pattern that emerged in each study diverged markedly.
In the interoceptive experiment, the accelerated-heartbeat feedback (Faster) systematically heightened pain relative to the decelerated version (Slower) at every level of noxious input: for low-intensity trials Faster exceeded Slower by 0.22 ± 0.08 points on the unpleasantness scale (t = 2.84, p = .0094) and by 3.87 ± 1.69 units on the numeric intensity scale (t = 2.29, p = .0448); at the medium intensity the corresponding differences were 0.19 ± 0.05 (t = -4.02, p = .0001) and 4.52 ± 1.06 (t = 4.28, p < .0001); and even at the highest intensity, Faster still surpassed Slower by 0.17 ± 0.08 on unpleasantness (t = 2.21, p = .0326) and by 5.16 ± 1.67 on intensity (t = 3.09, p = .0032). This uniform Faster > Slower pattern indicates that the interoceptive manipulation amplifies perceived pain in a stimulus-independent fashion.
The exteroceptive control experiment told a different story: the Faster-Slower contrast reached significance only at the most noxious setting (unpleasantness: estimate = 0.24 ± 0.07, t = -3.24, p = .0019; intensity: estimate = - 5.14 ± 1.82, t = 2.83, p = .0072) and was absent at the medium level (intensity , p=0.29; unpleasantness, p=0.45), while at the lowest level Slower actually produced numerically higher unpleasantness (2.56 versus 2.40) and intensity ratings (44.7 versus 42.2).
Thus, although both studies show that feedback effects depend on the actual nociceptive level of the stimulus, the results suggest that the faster vs. slower interoceptive feedback manipulation delivers a robust and intensity-invariant enhancement of pain, whereas the exteroceptive cue exerts a sporadic influence that surfaces solely under maximal stimulation.
These new results are now included in the Supplementary Materials, where we report the detailed analyses for both the Interoceptive and Exteroceptive experiments on the Likert unpleasantness ratings and the numeric pain intensity ratings.
(4) Sample size: It seems that the sample size was determined after the experiment was conducted, as the required N is identical to the actual N. I would be transparent about that, and say that retrospective sample size analyses support the ability of your sample size to support your claims. In general, a larger sample size than is required is always recommended, and if you were to run another study, I suggest you increase the sample size.
As also addressed in our responses to your later comments (see our detailed reply regarding the justification of SESOI and power analyses), the power analyses reported here were not post-hoc power analyses based on obtained results. In line with current recommendations (Anderson, Kelley & Maxwell, 2017; Albers & Lakens, 2018), we did not base our analyses on previously reported effect sizes, as these can carry considerable uncertainty, particularly for novel effects where robust estimates are lacking. Instead, we used sensitivity analyses, conducted using the sensitivity analysis function in G*Power (Version 3.1). Sensitivity analyses allow us to report effect sizes that our design was adequately powered (90%) to detect, given the actual sample size, desired power level, and the statistical test used in each experiment (Lakens, 2022). Following further guidance (Lakens, 2022), we also report the smallest effect size of interest (SESOI) that these tests could reliably detect.
This approach indicated that our design was powered to detect effect sizes of d = 0.57 in Experiment 1 and d = 0.62 in Experiment 2, with corresponding SESOIs of d = 0.34 and d = 0.37, respectively. The slightly higher value in Experiment 2 reflects the greater number of participants excluded (from an equal number originally tested) based on pre-specified criteria. Importantly, both experiments were well-powered to detect effects smaller than those typically reported in similar top-down pain modulation studies, where effect sizes around d = 0.7 have been observed (Iodice et al., 2019).
We have now clarified this rationale in the revised manuscript, Experiment 1- Methods - Participants (lines 208-217).
Albers, C., & Lakens, D. (2018). When power analyses based on pilot data are biased: Inaccurate effect size estimators and follow-up bias. Journal of experimental social psychology, 74, 187-195.
Anderson, S. F., Kelley, K., & Maxwell, S. E. (2017). Sample-Size Planning for More Accurate Statistical Power: A Method Adjusting Sample Effect Sizes for Publication Bias and Uncertainty. Psychological Science, 28(11), 1547-1562. https://doi.org/10.1177/0956797617723724
Lakens, D. (2022). Sample size justification. Collabra: psychology, 8(1), 33267.
(5) Analysis: the use of change scores instead of the actual scores is not recommended, as it is a loss of data, but could have been ignored if it didn't have a significant effect on the analyses conducted. Instead of conducting an RM-ANOVA of conditions (faster, slower, normal heartbeats) across participants, finding significant interaction, and then moving on to specific post-hoc paired comparisons between conditions, the authors begin with the change score but then move on to conduct the said paired comparisons without ever anchoring these analyses in an appropriate larger ANOVA. I strongly recommend the use of an ANOVA but if not, the authors would have to correct for multiple comparisons at the minimum.
We thank the reviewer for their comment regarding the use of change scores. These were originally derived from the difference between the slower and faster feedback conditions relative to the congruent condition. In line with the reviewer’s recommendation, we have now removed these difference-based change scores from the main analysis. The results remain identical. Please note that we have retained the normalization procedure, relative to each participant’s initial baseline in the no feedback trials, as it is widely used in the interoceptive and pain literature (e.g., Bartolo et al., 2013; Cecchini et al., 2020; Riello et al., 2019). This approach helps to control for interindividual variability and baseline differences by expressing each participant’s response relative to their no-feedback baseline. As before, normalization was applied across all dependent variables (heart rate, pain intensity, and pain unpleasantness).
To address the reviewer’s concern about statistical validity, we now first report a 1-factor repeated-measures ANOVA (Greenhouse-Geisser corrected) for each dependent variable, with feedback condition (slower, congruent, faster) as the within-subject factor.
These show in each case a significant main effect, which we then follow with planned paired-sample t-tests comparing:
Faster vs. slower feedback (our main hypothesis, as these manipulations are expected to produce largest, most powerful, test of our hypothesis, see response to Reviewer 3),
Faster vs. congruent and slower vs. congruent (to test for potential asymmetries, as suggested by previous false heart rate feedback studies).
The rationale of these analyses is further discussed in the Data Analysis of Experiment 1 (lines 405-437).
Although we report the omnibus one-factor RM-ANOVAs to satisfy conventional expectations, we note that such tests are not statistically necessary, nor even optimal, when the research question is fully captured by a priori, theory-driven contrasts. Extensive methodological work shows that, in this situation, going straight to planned contrasts maximises power without inflating Type I error and avoids the logical circularity of first testing an effect one does not predict (e.g., Rosenthal & Rosnow, 1985). In other words, an omnibus F is warranted only when one wishes to protect against unspecified patterns of differences. Here our hypotheses were precise (Faster ≠ Slower; potential asymmetry relative to Congruent), so the planned paired comparisons would have sufficed statistically. We therefore include the RM-ANOVAs solely for readers who expect to see them, but our inferential conclusions rest on the theoretically motivated contrasts.
Rosenthal, R., & Rosnow, R. L. (1985). Contrast analysis. New York: Cambridge.
(6) Correlations: were there correlations between subjects' own heartbeats (which are considered a predictive cue) and pain perceptions? This is critical to show that the two are in fact related.
We thank the reviewer for this thoughtful suggestion. While we agree that testing for a correlation between anticipatory heart rate responses and subjective pain ratings is theoretically relevant. However, we have not conducted this analysis in the current manuscript, as our study was not designed or powered to reliably detect such individual differences. As noted by Hedge, Powell, and Sumner (2018), robust within-subject experimental designs tend to minimize between-subject variability in order to detect clear experimental effects. This reduction in variance at the between-subject level limits the reliability of correlational analyses involving trait-like or individual response patterns. This issue, known as the reliability paradox, highlights that measures showing robust within-subject effects may not show stable individual differences, and therefore correlations with other individual-level variables (like subjective ratings used here) require much larger samples to produce interpretable results than available here (and commonly used in the literature), typically more than 200 participants. For these reasons, we believe that running such an analysis in our current dataset would not yield informative results and could be misleading.
We now explicitly acknowledge this point in the revised version of the manuscript (Limitations and future directions, lines 832-851) and suggest that future studies specifically designed to examine individual variability in anticipatory physiological responses and pain perception would be better suited to address this question.
Hedge, C., Powell, G., & Sumner, P. (2018). The reliability paradox: Why robust cognitive tasks do not produce reliable individual differences. Behavior Research Methods, 50(3), 1166-1186. https://doi.org/10.3758/s13428-017-0935-1
(7) The direct comparison between studies is great! and finally the use of ANOVA - but why without the appropriate post-hoc tests to support the bold claims in lines 542-544? This is needed. Same for 556-558.
We apologize if our writing was not clear here, but the result of the ANOVAs fully warrants the claims in 542-544 (now lines 616-618) and 556-558 (now lines 601-603).
In a 2x2 design, the interaction term is mathematically identical to comparing the difference induced by Factor 1 at one level of Factor 2 with the same difference induced at the other level of Factor 2. In our 2x2 analysis with the factors Experiment (Cardiac feedback, Exteroceptive feedback - between participants) and Feedback Frequency (faster, slower - within participants), the interaction therefore directly tests whether the effect of Feedback frequency differs statistically (i.e., is larger or smaller) in the participants in the interoceptive and exteroceptive experiments. Thus, the conclusion that “faster feedback affected the perceptual bias more strongly in the Experiment 1 than in Experiment 2” captures the outcome of the significant interaction exactly. Indeed, this test would be statistically equivalent (and would produce identical p values) to a simple between-group t-test between each participant’s difference between the faster and slower feedback in the interoceptive group and the analogous differences between the faster and slower feedback in the exteroceptive group, as illustrated in standard examples of factorial analysis (see, e.g., Maxwell, Delaney and Kelley, 2018).
Please note that, for the above reason, mathematically the conclusion of larger effects in one experiment than the other is licensed by the significant interaction even without follow-up t-tests. However, if the reader would like to see these tests, they are simply the main analysis results reported in each of the two experiment sections, where significant (t-test) differences between faster and slower feedback were induced with interoceptive cues (Experiment 1) but not exteroceptive cues (Experiment 2). Reporting them in the between-experiment comparison section again would therefore be redundant.
To avoid this lack of clarity, we have now re-written the results section of each experiment. First, as noted above, we now precede our main hypothesis test - the crucial t-test comparing heartrate and pain ratings after faster vs slower feedback - with an ANOVA including all three levels (faster, congruent, slower feedback). Moreover, we removed the separate between-experiment comparison section. Instead, in the Result section of the exteroceptive Experiment 2, we now directly compare the (absent or reversed) effects of faster vs slower feedback directly, with a between-groups t-test, with the present effects in the interoceptive Experiment 1. This shows conclusively, and hopefully more clearly, that the effects in both experiments differ. We hope that this makes the logic of our analyses clearer.
Maxwell, S. E., Delaney, H. D., & Kelley, K. (2017). Designing experiments and analyzing data: A model comparison perspective. Routledge.
(8) The discussion is missing a limitation paragraph.
Thank you for the suggestion. We have now added a dedicated limitations paragraph in the Discussion section (lines 832-890).
Additional recommendations:
Minor (chronological order):
(1) Sample size calculations for both experiments: what was the effect size based on? A citation or further information is needed. Also, clarify why the effect size differed between the two experiments.
Please see above
(2) "Participants were asked to either not drink coffee or smoke cigarettes" - either is implying that one of the two was asked. I suspect it is redundant as both were not permitted.
The intention was to restrict both behaviors, so we have corrected the sentence to clarify that participants were asked not to drink coffee or smoke cigarettes before the session.
(3) Normalization of ECG - what exactly was normalized, namely what measure of the ECG?
The normalized measure was the heart rate, expressed in beats per minute (bpm). We now clarify this in the Data Analysis section of Experiment 1 (Measures of the heart rate recorded with the ECG (beats per minute) in the feedback phase were normalized)
(4) Line 360: "Mean Δ pain unpleasantness ratings were analysed analogously" - this is unclear, if already described in methods then should be removed here, if not - should be further explained here.
Thank you for your observation. We are no longer using change scores.
(5) Lines 418-420: "Consequently, perceptual and cardiac modulations associated with the feedback manipulation should be reduced over the exposure to the faster exteroceptive sound." - why reduced and not unchanged? I didn't follow the logic.
We chose the term “reduced” rather than “unchanged” to remain cautious in our interpretation. Statistically, the absence of a significant effect in one experiment does not necessarily mean that no effect is present; it simply means we did not detect one. For this reason, we avoided using language that would suggest complete absence of modulation. It also more closely matches the results of the between experiment comparisons that we report in the Result section of Experiment 2, which can in principle only show that the effect in Experiment 2 was smaller than that of Experiment 1, not that it was absent. Even the TOST analysis that we utilize to show the absence of an effect can only show that any effect that is present is smaller than we could reasonably expect to detect with our experimental design, not its complete absence.
Also, on a theoretical level, pain is a complex, multidimensional experience influenced not only by sensory input but also by cognitive, emotional, social and expectancy factors. For this reason, we considered it important to remain open to the possibility that other mechanisms beyond the misleading cardiac prior induced by the feedback might have contributed to the observed effects. If such other influences had contributed to the induced differences between faster and slower feedback in Experiment 1, some remainder of this difference could have been observed in Experiment 2 as well.
Thus, for both statistical and theoretical reasons, we were careful to predict a reduction of the crucial difference, not its complete elimination. However, to warrant the possibility that effects could be completely eliminated we now write that “perceptual and cardiac modulations associated with the feedback manipulation should be reduced or eliminated with exteroceptive feedback”
(6) Study 2 generation of feedback - was this again tailored per participants (25% above and beyond their own HR at baseline + gradually increasing or decreasing), or identical for everyone?
Yes, in Study 2, the generation of feedback was tailored to each participant, mirroring the procedure or Experiment 1. Specifically, the feedback was set to be 25% above or below their baseline heart rate, with the feedback gradually increasing or decreasing. This individualized approach ensured that each participant experienced feedback relative to their own baseline heart rate. We now clarify this in the Methods section (lines 306-318).
(7) I did not follow why we need the TOST and how to interpret its results.
We thank the reviewer for raising this important point. In classical null hypothesis significance testing (NHST), a non-significant p-value (e.g., p > .05) only indicates that we failed to find a statistically significant difference, not that there is no difference. It therefore does not allow us to conclude that two conditions are equivalent – only that we cannot confidently say they are different. In our case, to support the claim that exteroceptive feedback does not induce perceptual or physiological changes (unlike interoceptive feedback), we needed a method to test for the absence of a meaningful effect, not just the absence of a statistically detectable one.
The TOST (Two One-Sided Tests) procedure reverses the logic of NHST by testing whether the observed effect falls within a predefined equivalence interval, called the smallest effect size of interest (SESOI) that is in principle measurable with our design parameters (e.g., type of test, number of participants). This approach is necessary when the goal is not to detect a difference, but rather to demonstrate that an observed effect is so small that it can be considered negligible – or at the least smaller than we could in principle expect to observe in the given experiment. We used the TOST procedure in Experiment 2 to test for statistical equivalence between the effects of faster and slower exteroceptive feedback on pain ratings and heart rate.
We hope that the clearer explanation now provided in data analysis of Experiment 2 section (lines 5589-563) fully addresses the reviewer’s concern.
(8) Lines 492-3: authors say TOST significant, while p value = 0.065
We thank the reviewer for spotting this inconsistency. The discrepancy was due to a typographical error in the initial manuscript. During the revision of the paper, we rechecked and fully recomputed all TOST analyses, and the results have now been corrected throughout the manuscript to accurately reflect the statistical outcomes. In particular, for the comparison of heart rate between faster and slower exteroceptive feedback in Experiment 2, the corrected TOST analysis now shows a significant equivalence, with the observed effect size being d = -0.19 (90% CI [-0.36, -0.03]) and both one-sided tests yielding p = .025 and p < .001. These updated results are reported in the revised Results section.
Reviewer #2 (Recommendations For The Authors):
I would suggest the authors revise their definition of pain in the introduction, since it is not always a protective experience. The new IASP definition specifically takes this into consideration.
We thank the reviewer for this suggestion. We have updated the definition of pain in the Introduction (lines 2-4) to align with the most recent IASP definition (2020), which characterizes pain as “an unpleasant sensory and emotional experience associated with, or resembling that associated with, actual or potential tissue damage” (lines 51-53).
The work on exteroceptive cues does not necessarily neglect the role of interoceptive sources of information, although it is true that it has been comparatively less studied. I suggest rephrasing this sentence to reflect this.
We thank the reviewer for pointing out this important nuance. We agree that studies employing exteroceptive cues to modulate pain perception do not necessarily neglect the role of interoceptive sources, even though these are not always the primary focus of investigation. Our intention was not to imply a strict dichotomy, but rather to highlight that interoceptive mechanisms have been comparatively under-investigated. We have revised the sentence in the Introduction accordingly to better reflect this perspective (Introduction, lines 110-112, “Although interoceptive processes may have contributed to the observed effects, these studies did not specifically target interoceptive sources of information within the inferential process.”).
The last paragraph of the introduction (lines 158-164) contains generalizations beyond what can be supported by the data and the results, about the generation of predictive processes and the origins of these predictions. The statements regarding the understanding of pain-related pathologies in terms of chronic aberrant predictions in the context of this study are also unwarranted.
We have deleted this paragraph now.
I could not find the study registration (at least in clinicaltrials.gov). This is curious considering that the hypothesis and the experimental design seem in principle well thought out, and a study pre-registration improves the credibility of the research (Nosek et al., 2018). I also find the choice for the smallest effect of interest (SESOI) odd. Besides the unnecessary variable transformations (more on that later), there is no justification for why that particular SESOI was chosen, or why it changes between experiments (Dienes, 2021; King, 2011), which makes the choice look arbitrary. The SESOI is a fundamental component of a priori power analysis (Lakens, 2022), and without rationale and preregistration, it is impossible to tell whether this is a case of SPARKing or not (Sasaki & Yamada, 2023).
We acknowledge that the study was not preregistered. Although our hypotheses and design were developed a priori and informed by established theoretical frameworks, the lack of formal preregistration is a limitation.
The SESOI values for Experiments 1 and 2 were derived from sensitivity analyses based on the fixed design parameters (type of test, number of participants, alpha level) of our study, not from any post-hoc interpretation based on observed results - they can therefore not be a case of SPARKing. Following current recommendations (Anderson, Kelley & Maxwell, 2017; Albers & Lakens, 2017; Lakens, 2022), we avoided basing power estimates on published effect sizes, as no such values exist for in novel paradigms, and are typically inflated due to publication and other biases. Instead, sensitivity analyses (using G*Power, v 3.1) allows us to calculate, prospectively, the smallest effect each design could detect with 90 % power, given the actual sample size, test type, and α level. Because more participants were excluded in Experiment 2, this design can detect slightly larger effects (d = 0.62) than Experiment 1 (d = 0.57). Please note that both studies therefore remain well-powered to capture effects of the magnitude typically reported in previous research using feedback manipulations to explore interoceptive illusions (e.g., Iodice et al., 2019, d ≈ 0.7).
We have added this clarification to the Participants section of Experiment 1 (Lines 208-217).
Anderson, S. F., Kelley, K., & Maxwell, S. E. (2017). Sample-Size Planning for More Accurate Statistical Power: A Method Adjusting Sample Effect Sizes for Publication Bias and Uncertainty. Psychological Science, 28(11), 1547-1562.
Lakens, D. (2022). Sample size justification. Collabra: psychology, 8(1), 33267.
Albers, C., & Lakens, D. (2018). When power analyses based on pilot data are biased: Inaccurate effect size estimators and follow-up bias. Journal of experimental social psychology, 74, 187-195.
In the Apparatus subsection, it is stated that the intensity of the electrical stimuli was fixed at 2 ms. I believe the authors refer to the duration of the stimulus, not its intensity.
You are right, thank you for pointing that out. The text should refer to the duration of the electrical stimulus, not its intensity. We have corrected this wording in the revised manuscript to avoid confusion.
It would be interesting to report (in graphical form) the stimulation intensities corresponding to the calibration procedure for the five different pain levels identified for all subjects.
That's a good suggestion. We have included a supplementary figure showing the stimulation intensities corresponding to the five individually calibrated pain levels across all participants (Supplementary Figure 11.)
It is questionable that researchers state that "pain and unpleasantness should be rated independently" but then the first level of the Likert scale for unpleasantness is "1=no pain". This is particularly relevant since simulation (and specifically electrical stimulation) can be unpleasant but non-painful at the same time. Since the experiments were already performed, the researchers should at least explain this choice.
Thank you for raising this point. You are right in that the label of “no pain” in the pain unpleasantness scale was not ideal, and we now acknowledge this in the text (lines 886-890). Please note that this was always the second rating that participants gave (after pain intensity), and the strongest results come from this first rating.
Discussion.
I did not find in the manuscript the rationale for varying the frequency of the heart rate by 25% (instead of any other arbitrary quantity).
We thank the Reviewer for this observation, which prompted us to clarify the rationale behind our choice of a ±25% manipulation of heart rate feedback. False feedback paradigms have historically relied on a variety of approaches to modulate perceived cardiac signals. Some studies have adopted non-individualised values, using fixed frequencies (e.g., 60 or 110 bpm) to evoke states of calm or arousal, independently of participants’ actual physiology (Valins, 1966; Shahidi & Baluch, 1991; Crucian et al., 2000; Tajadura-Jiménez et al., 2008). Others have used the participant’s real-time heart rate as a basis, introducing accelerations or decelerations without applying a specific percentage transformation (e.g., Iodice et al., 2019). More recently, a growing body of work has employed percentage-based alterations of the instantaneous heart rate, offering a controlled and participant-specific manipulation. These include studies using −20% (Azevedo et al., 2017), ±30% (Dey et al., 2018), and even ±50% (Gray et al., 2007).
These different methodologies - non-individualised, absolute, or proportionally scaled - have all been shown to effectively modulate subjective and physiological responses. They suggest that the impact of false feedback does not depend on a single fixed method, but rather on the plausibility and salience of the manipulation within the context of the task. We chose to apply a ±25% variation because it falls well within the most commonly used range and strikes a balance between producing a detectable effect and maintaining the illusion of physiological realism. The magnitude is conceptually justified as being large enough to shape interoceptive and emotional experience (as shown by Azevedo and Dey), yet small enough to avoid implausible or disruptive alterations, such as those approaching ±50%. We have now clarified this rationale in the revised Procedure paragraph of Experiment 1 (lines 306-318).
T. Azevedo, R., Bennett, N., Bilicki, A., Hooper, J., Markopoulou, F., & Tsakiris, M. (2017). The calming effect of a new wearable device during the anticipation of public speech. Scientific reports, 7(1), 2285.
Crucian, G. P., Hughes, J. D., Barrett, A. M., Williamson, D. J. G., Bauer, R. M., Bowers, D., & Heilman, K. M. (2000). Emotional and physiological responses to false feedback. Cortex, 36(5), 623-647.
Dey, A., Chen, H., Billinghurst, M., & Lindeman, R. W. (2018, October). Effects of manipulating physiological feedback in immersive virtual environments. In Proceedings of the 2018 Annual Symposium on Computer-Human Interaction in Play (pp. 101-111).
Gray, M. A., Harrison, N. A., Wiens, S., & Critchley, H. D. (2007). Modulation of emotional appraisal by false physiological feedback during fMRI. PLoS one, 2(6), e546.
Shahidi, S., & Baluch, B. (1991). False heart-rate feedback, social anxiety and self-attribution of embarrassment. Psychological reports, 69(3), 1024-1026.
Tajadura-Jiménez, A., Väljamäe, A., & Västfjäll, D. (2008). Self-representation in mediated environments: the experience of emotions modulated by auditory-vibrotactile heartbeat. CyberPsychology & Behavior, 11(1), 33-38.
Valins, S. (1966). Cognitive effects of false heart-rate feedback. Journal of personality and social psychology, 4(4), 400.
The researchers state that pain ratings collected in the feedback phase were normalized to the no-feedback phase to control for inter-individual variability in pain perception, as established by previous research. They cite three studies involving smell and taste, of which the last two contain the same normalization presented in this study. However, unlike these studies, the outcomes here require no normalization whatsoever, because there should be no (or very little) inter-individual variability in pain intensity ratings. Indeed, pain intensity ratings in this study are anchored to 30, 50, and 70 / 100 as a condition of the experimental design. The researchers go to extreme lengths to ensure this is the case, by adjusting stimulation intensities until at least 75% of stimulation intensities are correctly matched to their pain ratings counterpart in the pre-experiment procedure. In other words, inter-individual variability in this study is in stimulation intensities, and not pain intensity ratings. Even if it could be argued that pain unpleasantness and heart rate still need to account for inter-individual variability, the best way to do this is by using the baseline (no-feedback) measures as covariates in a mixed linear model. Another advantage of this approach is that all the effects can be described in terms of the original scales and are readily understandable, and post hoc tests between levels can be corrected for multiple comparisons. On the contrary, the familywise error rate for the comparisons between conditions in the current analysis is larger than 5% (since there is a "main" paired t-test and additional "simple" tests).
We disagree that there is little to no variability in the no feedback phase. Participants were tested in their ability to distinguish intensities in an initial pre-experiment calibration phase. In the no feedback phase, participants rated the pain stimuli in the full experimental context.
In the pre-experiment calibration phase, participants were tested only once in their ability to match five electrical‐stimulation levels to the 0-100 NPS scale, before any feedback manipulation started. During this pre-experiment calibration we required that each level was classified correctly on ≥ 75 % of the four repetitions; “correct” meant falling within ± 5 NPS units of the target anchor (e.g., a response of 25–35 was accepted for the 30/100 anchor). This procedure served one purpose only: to make sure that every participant entered the main experiment with three unambiguously distinguishable stimulation levels (30 / 50 / 70). We integrated this point in the revised manuscript lines 263-270.
Once the real task began, the context changed: shocks are unpredictable, attention is drawn to the heartbeat, and participants must judge both intensity and unpleasantness. In this full experimental setting the no-feedback block indeed shows considerable variability, even for the pain intensity ratings. Participants mean rating on the NPS scale was 46.4, with a standard deviation of 11.9 - thus participants vary quite strongly in their mean ratings (range 14.5 to 70). Moreover, while all participants show a positive correlation between actual intensities and their ratings (i.e., they rate the higher intensities as more intense than the lower ones), they vary in how much of the scale they use, with differences between reported highest and lowest intensities ranging between 8 and 91, for the participants showing the smallest and largest differences, respectively.
Thus, while we simplified the analysis to remove the difference scoring relative to the congruent trials and now use these congruent trials as an additional condition in the analysis, we retained the normalisation procedure to account for the in-fact-existing between-participant variability, and ensure consistency with prior research (Bartolo et al., 2013; Cecchini et al., 2020; Riello et al., 2019) and our a priori analysis plan.
However, to ensure we fully address your point here (and the other reviewers’ points about potential additional factors affecting the effects, like trial number and stimulus intensity), we also report an additional linear mixed-effects model analysis without normalization. It includes every feedback level as condition (No-Feedback, Congruent, Slower, Faster), plus additional predictors for actual stimulus intensity and trial rank within the experiment (as suggested by the other reviewers). This confirms that all relevant results remain intact once baseline and congruent trials are explicitly included in the model.
In brief, cross‐experiment analyses demonstrated that the Faster vs Slower contrast was markedly larger when the feedback was interoceptive than when it was exteroceptive. This held for heart-rate deceleration (b = 0.94 bpm, p = .005), for increases in unpleasantness (b = -0.16 Likert units, p = .015), and in pain-intensity ratings (b = -3.27 NPS points, p = .037).
These findings were then further confirmed by within-experiment analyses. Within the interoceptive experiment, the mixed-model on raw scores replicated every original effect: heart rate was lower after Faster than Slower feedback (estimate = –0.69 bpm, p = .005); unpleasantness was higher after Faster than Slower feedback (estimate = 0.19, p < .001); pain-intensity rose after Faster versus Slower (estimate=-4.285, p < .001). In the exteroceptive experiment, however, none of these Faster–Slower contrasts reached significance for heart rate (all ps > .33), unpleasantness (all ps > .43) or intensity (all ps > .10). Because these effects remain significant even with No-Feedback and Congruent trials explicitly included in the model and vanish under exteroceptive control, the supplementary, non-normalised analyses confirm that the faster vs. slower interoceptive feedback uniquely lowers anticipatory heart rate while amplifying both intensity and unpleasantness of pain, independent of data transformation or reference conditions. Please see Supplementary analyses for further details.
Bartolo, M., Serrao, M., Gamgebeli, Z., Alpaidze, M., Perrotta, A., Padua, L., Pierelli, F., Nappi, G., & Sandrini, G. (2013). Modulation of the human nociceptive flexion reflex by pleasant and unpleasant odors. PAIN®, 154(10), 2054-2059.
Cecchini, M. P., Riello, M., Sandri, A., Zanini, A., Fiorio, M., & Tinazzi, M. (2020). Smell and taste dissociations in the modulation of tonic pain perception induced by a capsaicin cream application. European Journal of Pain, 24(10), 1946-1955.
Riello, M., Cecchini, M. P., Zanini, A., Di Chiappari, M., Tinazzi, M., & Fiorio, M. (2019). Perception of phasic pain is modulated by smell and taste. European Journal of Pain, 23(10), 1790-1800.
I could initially not find a rationale for bringing upfront the comparison between faster vs. slower HR acoustic feedback when in principle the intuitive comparisons would be faster vs. congruent and slower vs. congruent feedback. This is even more relevant considering that in the proposed main comparison, the congruent feedback does not play a role: since Δ outcomes are calculated as (faster - congruent) and (slower - congruent), a paired t-test between Δ faster and Δ slower outcomes equals (faster - congruent) - (slower - congruent) = (faster - slower). I later realized that the statistical comparison (paired t-test) of pain intensity ratings of faster vs. slower acoustic feedback is significant in experiment 1 but not in experiment 2, which in principle would support the argument that interoceptive, but not exteroceptive, feedback modulates pain perception. However, the "simple" t-tests show that faster feedback modulates pain perception in both experiments, although the effect is larger in experiment 1 (interoceptive feedback) compared to experiment 2 (exteroceptive feedback).
The comparison between faster and slower feedback is indeed crucial, and we regret not having made this clearer in the first version of the manuscript. As noted in our response to your point in the public review, this comparison is both statistically most powerful, and theoretically the most appropriate, as it controls for any influence of salience or surprise when heart rates deviate (in either direction) from what is expected. It therefore provides a clean measure of how much accelerated heartrate affects pain perception and physiological response, relative to an equal change in the opposite direction. However, as noted above, in the new version of the manuscript we have now removed the analysis via difference scores, and directly compared all three relevant conditions (faster, congruent, slower), first via an ANOVA and then with follow-up planned t-tests.
Please refer to our previous response for further details (i.e., Furthermore, the researchers propose the comparison of faster vs. slower delta HR acoustic feedback throughout the manuscript when the natural comparison is the incongruent vs. the congruent feedback [..]).
The design of experiment two involves the selection of knocking wood sounds to act as exteroceptive acoustic feedback. Since the purpose is to test whether sound affects pain intensity ratings, unpleasantness, and heart rate, it would have made sense to choose sounds that would be more likely to elicit such changes, e.g. Taffou et al. (2021), Chen & Wang (2022), Zhou et al. (2022), Tajadura-Jiménez et al. (2010). Whereas I acknowledge that there is a difference in effect sizes between experiment 1 and experiment 2 for the faster acoustic feedback, I am not fully convinced that this difference is due to the nature of the feedback (interoceptive vs. exteroceptive), since a similar difference could arguably be obtained by exteroceptive sound with looming or rough qualities. Since the experiment was already carried out and this hypothesis cannot be tested, I suggest that the researchers moderate the inferences made in the Discussion regarding these results.
Please refer to our previous response for a previous detailed answer to this point in the Public Review (i.e., This could be influenced by the fact that the faster HR exteroceptive cue in experiment 2 also shows a significant modulatory effect [..]). As we describe there, we see little grounds to suspect such a non-specific influence of acoustic parameters, as it is specifically the sensitivity to the change in heart rate (faster vs slower) that is affected by our between-experiment manipulation, not the overall response to the different exteroceptive or interoceptive sounds. Moreover, the specific change induced by the faster interoceptive feedback - a heartrate deceleration - is not consistent with a change in arousal or alertness (which would have predicted an increase in heartrate with increasing arousal). See also Discussion-Accounting for general unspecific contributions.
Additionally, the fact that no significant effects were found for unpleasantness ratings or heart rate (absence of evidence) should not be taken as proof that faster exteroceptive feedback does not induce an effect on these outcomes (evidence of absence). In this case, it could be that there is actually no effect on these variables, or that the experiment was not sufficiently powered to detect those effects. This would depend on the SESOIs for these variables, which as stated before, was not properly justified.
We very much agree that the absence of significant effects should not be interpreted as definitive evidence of absence. Indeed, we were careful not to overinterpret the null findings for heart rate and unpleasantness ratings, and we conducted additional analyses to clarify their interpretation. First, the TOST analysis shows that any effects in Experiment 2 are (significantly) smaller than the smallest effect size that can possibly be detected in our experiment, given the experimental parameters (number of participants, type of test, alpha level). Second, and more importantly, we run between-experiments comparisons (see Results Experiment 2, and Supplementary materials, Cross-experiment analysis between-subjects model) of the crucial difference in the changes induced by faster and slower feedback. This showed that the differences were larger with interoceptive (Experiment 1) than exteroceptive cues (Experiment 2). Thus, even if a smaller than is in principle detectable effect is induced by the exteroceptive cues in Experiment 2, it is smaller than with interoceptive cues in Experiment 1.
To ensure we fully address this point, we have now simplified our main analysis (main manuscript), replicated it with a different analysis (Supplementary material), we motivate more clearly (Methods Experiment 1), why the comparison between faster and slower feedback is crucial, and we make clearer that the difference between these conditions is larger in Experiment 1 than Experiment 2 (Results Experiment 2). Moreover, we went through the manuscript and ensured that our wording does not over-interpret the absence of effects in Experiment 2, as an absence of a difference.
The section "Additional comparison analysis between experiments" encompasses in a way all possible comparisons between levels of the different factors in both experiments. My original suggestion regarding the use of a mixed linear model with covariates is still valid for this case. This analysis also brings into question another aspect of the experimental design: what is the rationale for dividing the study into two experiments, considering that variability and confounding factors would have been much better controlled in a single experimental session that includes all conditions?
We thank the reviewer for their comment. We would like to note, first, that the between-experiment analyses did not encompass all possible comparisons between levels, as it just included faster and slower feedback for the within-experiment comparison Instead, they focus on the specific interaction between faster and slower feedback on the one hand, and interoceptive vs exteroceptive cues on the other. This interaction essentially compares, for each dependent measure (HR, pain unpleasantness, pain intensity), the difference between faster and slower feedback in Experiment 1 with that the same difference in Experiment 2 (and would produce identical p values to a between-experiment t-test). The significant interactions therefore indicate larger effects of interoceptive cues than exteroceptive ones for each of the measures. To make this clearer, we have now exchanged the analysis with between-experiment t-tests of the difference between faster and slower feedback for each measure (Results Experiment 2), producing identical results. Moreover, as suggested, we also now report linear mixed model analyses (see Supplementary Materials), which provide a comprehensive comparison across experiments.
Regarding the experimental design, we appreciate the reviewer’s suggestion regarding a within-subject crossover design. While such an approach indeed offers greater statistical power by reducing interindividual variability (Charness, Gneezy, & Kuhn, 2012), we intentionally chose a between-subjects design due to theoretical and methodological considerations specific to deceptive feedback paradigms. First, carryover effects are a major concern in deception studies. Participants exposed to one type of feedback could develop suspicion or adaptive strategies that would alter their responses in subsequent conditions (Martin & Sayette, 1993). Expectancy effects could thus contaminate results in a crossover design, particularly when feedback manipulation becomes apparent. In line with this idea, past studies on false cardiac feedback (e.g., Valins, 1966; Pennebaker & Lightner, 1980) often employed between-subjects or blocked designs to maintain the ecological validity of the illusion.
Charness, G., Gneezy, U., & Kuhn, M. A. (2012). Experimental methods: Between-subject and within-subject design. Journal of economic behavior & organization, 81(1), 1-8.
Martin, C. S., & Sayette, M. A. (1993). Experimental design in alcohol administration research: limitations and alternatives in the manipulation of dosage-set. Journal of studies on alcohol, 54(6), 750-761.
Pennebaker, J. W., & Lightner, J. M. (1980). Competition of internal and external information in an exercise setting. Journal of personality and social psychology, 39(1), 165.
Valins, S. (1966). Cognitive effects of false heart-rate feedback. Journal of personality and social psychology, 4(4), 400.
References
Chen ZS, Wang J. Pain, from perception to action: A computational perspective. iScience. 2022 Dec 1;26(1):105707. doi: 10.1016/j.isci.2022.105707.
Dienes Z. Obtaining Evidence for No Effect. Collabra: Psychology 2021 Jan 4; 7 (1): 28202. doi: 10.1525/collabra.28202
King MT. A point of minimal important difference (MID): a critique of terminology and methods. Expert Rev Pharmacoecon Outcomes Res. 2011 Apr;11(2):171-84. doi: 10.1586/erp.11.9.
Lakens D. Sample Size Justification. Collabra: Psychology 2022 Jan 5; 8 (1): 33267. doi: 10.1525/collabra.33267
Nosek BA, Ebersole CR, DeHaven AC, Mellor DT. The preregistration revolution. Proc Natl Acad Sci U S A. 2018 Mar 13;115(11):2600-2606. doi: 10.1073/pnas.1708274114.
Sasaki K, Yamada Y. SPARKing: Sample-size planning after the results are known. Front Hum Neurosci. 2023 Feb 22;17:912338. doi: 10.3389/fnhum.2023.912338.
Taffou M, Suied C, Viaud-Delmon I. Auditory roughness elicits defense reactions. Sci Rep. 2021 Jan 13;11(1):956. doi: 10.1038/s41598-020-79767-0.
Tajadura-Jiménez A, Väljamäe A, Asutay E, Västfjäll D. Embodied auditory perception: The emotional impact of approaching and receding sound sources. Emotion. 2010, 10(2), 216-229.https://doi.org/10.1037/a0018422
Zhou W, Ye C, Wang H, Mao Y, Zhang W, Liu A, Yang CL, Li T, Hayashi L, Zhao W, Chen L, Liu Y, Tao W, Zhang Z. Sound induces analgesia through corticothalamic circuits. Science. 2022 Jul 8;377(6602):198-204. doi: 10.1126/science.abn4663.
Reviewer #3 (Recommendations For The Authors):
The manuscript would benefit from some spelling- and grammar checking.
Done
Discussion:
The discussion section is rather lengthy and would benefit from some re-structuring, editing, and sub-section headers.
In response, we have restructured and edited the Discussion section to improve clarity and flow.
I personally had a difficult time understanding how the data relates to the rubber hand illusion (l.623-630). I would recommend revising or deleting this section.
We thank the reviewer for this valuable feedback. We have revised the paragraph and made the parallel clearer (lines 731-739).
Other areas are a bit short and might benefit from some elaboration, such as clinical implications. Since they were mentioned in the abstract, I had expected a bit more thorough discussion here (l. 718).
Thank you for this suggestion. We have expanded the discussion to more thoroughly address the clinical implications of our interoceptive pain illusion (See Limitations and Future Directions paragraph).
Further, clarification is needed for the following:
I would like some more details on participant instructions; in particular, the potential difference in instruction between Exp. 1 and 2, if any. In Exp. 1, it says: (l. 280) "Crucially, they were also informed that over the 60 seconds preceding the administration of the shock, they were exposed to acoustic feedback, which was equivalent to their ongoing heart rate". Was there a similar instruction for Exp. 2? If yes, it would suggest a more specific effect of cardiac auditory feedback; if no, the ramifications of this difference in instructions should be more thoroughly discussed.
Thank you for this suggestion. We have clarified this point in the Procedure of Experiment 2 (548-550).
🎉 SIGNAL ACQUIRED: You Found It.
Congratulations, Agent.
Most people scan the headlines and move on. They stay on the surface. But you stopped. You questioned the text. You looked deeper.
You have just passed the first test of an Insider.
You have discovered the "Tactical Overlay" (the Kingdomlens). This is where the real work happens. While the main text gives you the map, these notes give you the compass—the science, the scripture, and the deeper conversation.
SYSTEM NOTE (TRANSPARENCY): This overlay is powered by Hypothesis, an open-source non-profit organization dedicated to open conversation.
This is a third-party tool, distinct from the Dream Centre.
To join the conversation, you will need to create a free account directly with them.
**YOUR NEXT MOVE: ** Initialize: Click "Sign Up" above to create your secure ID with Hypothesis.
Radio Check: Once logged in, click the Reply Arrow ↩️ on this note and type a simple confirmation:
"I'm in."
Welcome to the team. Let's dig in.
CLICK HERE TO ENGAGE THE KINGDOMLENS.
**WELCOME TO THE INVISIBLE REALM. ** You just proved a spiritual reality: The answer was always here, but you had to take action to reveal it.
This tool you are using is called The Tactical Overlay (Hypothesis). It is our way of communicating "beneath" the surface of the text.
THE LESSON: Jesus said, "The Kingdom of God is at hand." He didn't say it was far away. He said it was right here—superimposed over our normal reality.
The World says: "What you see is what you get."
The Kingdom says: "What is seen is temporary; what is unseen is eternal."
To be an Insider is to live Inside-Out, Back-to-Front, The Other Way Up and Invisible. To look at a situation that seems hopeless (the surface text) and activate the Kingdomlens (this overlay) to see what God is actually doing.
YOUR MISSION (SYSTEM CHECK):
Use the Overlay: Whenever you see yellow text on this site, it means there is Kingdom Intel hidden there. Click it.
Confirm Comms: Do not create a new highlight yet. Instead, click the Reply Arrow on this note and tell us:
"I'm seeing the other layer."
Welcome to the resistance.
Reviewer #1 (Public review):
Summary:
This study was designed to manipulate and analyze the effects of chemosensory cues on visuomotor control. They approach this by analyzing how eye-body coordination and brain-wide activity are altered with specific chemosensation in larval zebrafish. After analyzing the dynamics of coupled saccade-tail coordination sequences - directionally linked and typically coupled to body turns - the authors investigated the effects of sensory cues shown to be either aversive or appetitive on freely swimming zebrafish on the eye-body coordination. Aversive chemicals lead to an increase in saccade-tail sequences in both number and dynamics, seemingly facilitating behaviors like escape. Brain-wide imaging led the authors to neurons in the telencephalic pallium as a target to study eye-body coordination. Pallium neuron activity correlated with both aversive chemicals and coupled saccade-tail movements.
Recommendations for improvement are minimal. So much of the data is ultimately tabular, and the figures are an impenetrable wall of datapoints. 1c is an excellent example: three concentrations are presented, but it's rare for the three averages to trend appropriately. The key point, which is that aversive odors are repulsive and attractive odors (sometimes) attractive just gets lost in showing the three concentrations individually; it also makes direct comparisons impossible. There are similar challenges abound in the violin plots in 4e-4h, the error bars on the "fits" in 4i-4m, and so on. We recommend selecting an illustrative subset of data to present to permit interpretation and putting the rest in a supplemental table. (Presenting) less is more (effective).
Reviewer #3 (Public review):
The manuscript investigates the coupling of saccadic eye movements (S) with directed tail flips (T). The remarkable discovery is that tail flips that are preceded by a conjugate sacced (S-T) can be credibly classified as specific "volitional" turns that are distinguished from the standard tail movements that seem to be more of a spontaneous and "impulsive" nature.
They show that 'turning intent', as indicated by a small increase in S, is elevated by aversive odors, while 'gliding intent', as indicated by a decrease in S and an increase in undulation cycles, is elevated by appetitive odors.
This is a very important finding, which is backed up by a thorough behavioral analysis, and the identification of neural populations in the pallium and sub-pallium that clearly distinguish between these kinds of turns is very promising. Here they identify neuronal populations that are preferentially active during - and predictive of - coupled (S-T) versus isolated (T) tail flips.
Especially the fact that S-T turns (but not T turns) can be predicted already by pre-event, ramping, pallial activity is intriguing.
The authors then go on and demonstrate that the frequency of (S-T) turns is modulated in fish exposed to appetitive or aversive odors.<br /> Specifically, they quantify the aversiveness and appetitive-ness of several odors in a free swimming assay. They select a couple of these odors based on their valence, and they demonstrate that these odors induce moderate modulation in the frequency of eye saccades (S) and tail flips (T) and (S-T) turns.
The study is rigorous and thorough, and the findings are informative and novel.
In important controls, they confirm that brain-wide imaging can distinguish between appetitive and aversive contexts, and they show that pallial activation by aversive odors is consistent with neural activity in the rhombencephalon that correlates with turning activity, whereas sub-pallial activation by appetitive odors correlates with rhombencephalic activity related to gliding.
Overall, this manuscript is very good.
Author response:
We thank the editors and all reviewers for the detailed evaluation of the work and the overall positive remarks, as well as the constructive feedback to improve our manuscript. Based on the integrated comments of the reviewers and advice of the reviewing editor, we will suitably address all comments raised by the reviewers, and we outline our revision plan below:
Interpretation of findings
● We will carefully reframe our interpretation of the data regarding the role of the pallium in the coupled saccade-tail turning events, and clearly state that we have not established a causal role, which requires additional perturbation experiments.
● We will also acknowledge the confounding interpretation that the pallial activities recorded may also represent or include arousal state signals.
Streamlining the presentation
● In the introduction, we will better contextualize our study with additional discussions on (i) the advantageous use of zebrafish to study chemosensation, factoring in differences in the spread of chemical cues in water vs. air, and (ii) the disruption of eye-body coordination and underlying neural circuits.
● We will streamline the presentation of data in Fig. 1 by keeping the overall responses of the larvae to each chemical across concentrations in the main figure, while moving suitable additional details to a supplementary figure.
● Similarly, for each of the subsequent main figures, wherever suitable we will select an illustrative, core set of panels to retain in the main figure, and move other more detailed plots to supplementary figures.
● We will incorporate additional references and discussions of the past literature, including relating our findings to (i) chemosensation/multisensory integration in Drosophila, (ii) thermosensation-driven and navigational behavior in larval zebrafish, and (iii) fleeing or escape behavior in zebrafish and other species.
● We will clarify our animal subject inclusion criteria, that all larval subjects with sufficiently high-quality, stable imaging were included (i.e., we only excluded larvae because of insufficient quality of imaging, but not other factors).
● For applicable plots, adding suitable additional details to the plots or legends (e.g., clarification of measures, specifying numbers of cells).
Data analysis and statistics
● We will perform additional data analysis, by making comparisons with statistics performedon fish subject-level, and include confident intervals wherever applicable.
Reviewer #1 (Public review):
Summary:
Witte et al. examined whether canonical behavioral functions attributed to the cerebellum decline with age. To test this, they recruited younger, old, and older-old adults in a comprehensive battery of tasks previously identified as cerebellar-dependent in the literature. Remarkably, they found that cerebellar function is largely preserved across the lifespan-and in some cases even enhanced. Structural imaging confirmed that their older adult cohort was representative in terms of both cerebellar gray- and white-matter volume. Overall, this is an important study with strong theoretical implications and convincing evidence supporting the motor reserve hypothesis, demonstrating that cerebellar-dependent measures remain largely intact with aging.
Strengths:
(1) Relatively large sample size.
(2) Most comprehensive behavioral battery to date assessing cerebellar-dependent behavior.
(3) Structural MRI confirmation of age-related decline in cerebellar gray and white matter, ensuring representativeness of the sample.
Weaknesses:
(1) Although the authors note this was outside the study's scope, the absence of a voxel-based morphometry (VBM) analysis limits anatomical and functional specificity. Such an analysis would clarify which functions are cerebellar-dependent rather than solely inferring this from prior neuropsychological literature.
(2) As acknowledged in the Discussion, task classification (cerebellar-dependent vs. general measures) remains somewhat ambiguous. Some "general" measures may still rely on cerebellar processes based on the paper's own criteria - for example, tasks in which individuals with cerebellar degeneration show impairments.
(3) Cerebellar-dependent and general measures may inherently differ in measurement noise, potentially biasing results toward detecting effects in general measures but not in cerebellar-dependent ones.
Reviewer #2 (Public review):
Summary:
The authors are investigating cerebellar-mediated motor behaviors in a large sample of adults, including 30 individuals over the age of 80 (a great strength of this work). They employed a large battery of motor tasks that are tied to cerebellar function, in addition to a cognitive task and motor tasks that are more general. They also evaluated cerebellar structure. Across their behavioral metrics, they found that even with cerebellar degeneration, cerebellar-mediated motor behavior remained intact relative to young adults. However, this was not the case for measures not directly tied to cerebellar function. The authors suggest that these functions are preserved and speak to the resiliency and redundancy of function in the cerebellum. They also speculate that cerebellar circuits may be especially good for preserving function in the face of structural change. The tasks are described very well, and their implementation is also well-done with consideration for rigor in the data collection and processing. The inclusion of Bayesian estimates is also particularly useful, given the theoretically important lack of age differences reported. This work is methodologically rigorous with respect to the behavior, and certainly thought-provoking.
Strengths:
The methodological rigor, inclusion of Bayesian statistics, and the larger sample of individuals over the age of 80 in particular are all great strengths of this work. Further, as noted in the text, the fact that all participants completed the full testing battery is of great benefit.
Weaknesses:
The suggestion of cerebellar reserve, given that at the group level there is a lack of difference for cerebellar-specific behavioral components, could be more robustly tested. That is, the authors suggest that this is a reserve given that the volume of cerebellar gray matter is smaller in the two older groups, though behavior is preserved. This implies volume and behavior are seemingly dissociated. However, there is seemingly a great deal of behavioral variability within each group and likewise with respect to cerebellar volume. Is poorer behavior associated with smaller volume? If so, this would still suggest that volume and behavior are linked, but rather than being age that is critical, it is volume. On the flip side, a lack of associations between behavior and volume would be quite compelling with respect to reserve. More generally, as explicated in the recommendations, there are analyses that could be conducted that, in my opinion, would more robustly support their arguments given the data that they have available. This is a well-executed and thought-provoking investigation, but there is also room for a bit more discussion.
Reviewer #3 (Public review):
Wang et al. report multiple experiments using functional magnetic resonance spectroscopy (fMRS) in a multiple object tracking (MOT) task to investigate the effect of experimentally manipulating a) the number of targets, b) object size, and c) total number of objects in the display on GABA and glutamate (Glx) concentrations in parietal and visual cortex. Data is analyzed in two orthogonal ways throughout: via condition differences in behavorial performance (inverse efficiency), GABA, and Glx concentrations and through correlations between changes in inverse efficiency and GABA or Glx. All three experimental manipulations affected inverse efficiency, with worse performance with more targets, smaller objects, and a larger total number of objects. However, only the manipulation of the target number produced a condition difference in GABA and Glx, with higher concentrations of both in the parietal VOI and only of Glx in the visual VOI with more targets ('high load'). Correlational analyses revealed that participants with a larger change in GABA in the parietal VOI with a higher number of targets showed a smaller drop in behavioral performance with more targets. The opposite direction of correlation was observed for Glx in both the visual and parietal VOI.
In the two control experiments, correlations were only investigated in the parietal VOI. There was a negative correlation between change in Glx and change in inverse efficiency with manipulation of object size, i.e. participants exhibiting a positive change in Glx showed no or little difference in performance, but those with an increase in Glx with smaller targets showed a more pronounced drop in performance. There was no correlation with GABA for the manipulation of object size. For the manipulation of total object number, participants exhibiting an increasing GABA concentration with more objects showed a smaller drop in performance.
The authors' main claim is that GABAergic suppression of goal-irrelevant distractors in parietal cortex is key to goal-directed visual information processing.
The study is, to my knowledge, the first to employ fMRS in an MOT paradigm, and I read it with great interest. I am admittedly not an expert on the fMRS technique and have therefore refrained from commenting on the technical aspects of its use. Although the application of fMRS to MOT is novel and adds new knowledge to the field, I have some critiques and believe that a much more nuanced interpretation of the findings is warranted.
Major
(1) Especially the control experiments lean heavily on Bettencourt and Somers (2009) and adopt and to some extent exaggerate claims from that paper uncritically. This is obvious in referring to the manipulations of object size and object number as high/low enhancement and high/low suppression, as if the association of these physical manipulations of the stimulus display with attentional mechanisms were so obvious and beyond doubt that drawing any distinction between these manipulations and their supposed effects is entirely superfluous. This seems far beyond what is warranted to me. It may seem plausible that adding distractors engages distractor suppression more, but whether this is truly the case is an empirical question, and Bettencourt and Somers (2009) have no direct measure of distractor suppression to substantiate this claim. Their study is purely behavioral, and there is no attempt to assess distractor processing separately. The case for the 'target enhancement' manipulation is even weaker: objects are of a sufficient size and at maximum contrast (white on black screen, but exact details are omitted) to be clearly visible in either condition, so why would smaller objects require more enhancement? Although the present data shows a clear effect of manipulating object size, the corresponding size of the effect in Bettencourt and Somers (2009) is rather underwhelming and does not warrant such a strong conclusion. In summary, the link between the object number and object size manipulations with suppression and enhancement is very far from the 1:1 that the authors seem to assume. Accordingly, I believe that the manipulations should be labelled as object number and object size rather than their hypothesized effects, throughout and that there should be a much more critical discussion as to whether these manipulations are indeed related to these effects as expected.
(2) The author's interpretation of the results seems rather uncritical. What is observed (at least in the first experiment) is a change in GABA and Glx concentrations with changes in the number of tracked targets. Is the only conceivable way in which this could happen through target enhancement and distractor suppression? The processing of targets and distractors is not measured directly, so any claims are indirect, at best. The authors cite the recent 'Ten simple rules to study distractor suppression' paper (Wöstmann et al., 2022), which presents a consensus between leading researchers in the field. Neither Bettencourt & Somers (2009) nor the design of the current study live up to the rules established in that paper, so a much more nuanced interpretation and discussion of the current findings seems warranted. It is anything but obvious to me that the only activity in the parietal cortex that could possibly be suppressed by GABA is the representation of distractors. Indeed, cueing more targets (high load) decreases the number of distractors in the first experiment, so the need for distractor suppression in the high load condition is less than in the low load condition. So, shouldn't we observe lower GABA concentrations in the 'high load' condition?
(3) It seems that the authors included data from both correctly tracked and incorrectly tracked trials in their fMRS analysis. In MOT, attending target objects is the task per se, so task errors indicate that participants did not actually track the targets. So when comparing conditions with different error levels, it is ambiguous whether changes in brain activity reflect the experimental manipulation as such, or rather the different mix of correctly tracked and incorrectly tracked trials that result from this physical manipulation. Are the correlations perhaps driven by the inclusion of different proportions of correctly tracked trials across participants? It seems that the authors may have to separate correct and error trials in the analysis to check for the possibility that effects are due to the inclusion of data from trials in which participants may have stopped tracking at least some of the target objects. Of course, such an analysis is somewhat limited by the fact that only one target was probed, yielding a 50% guessing chance (i.e. even if the response is correct, we do not know whether the other, unprobed, objects were tracked correctly on that trial).
(4) The key findings from the control experiments are purely correlational. The supposed cause may be what the authors claim, but there is an infinity of alternative explanations. Correlational findings cannot simply be interpreted as if they resulted from an experimental manipulation (...although this is, unfortunately, by no means rare in the cognitive neuroscience literature). The authors should make a rigorous effort to consider the most plausible alternative explanations for these correlations and argue why or why not they believe that they can be discounted.
(5) Related to the previous point: the experimental manipulations did not produce mean differences in GABA/Glx in the control experiments. Doesn't this speak against the authors' interpretation? They briefly acknowledge this in the discussion, but I think there is a deeper problem. The absence of these effects casts doubt on what these manipulations actually do, and therefore also on the interpretation of the correlations in these experiments. For example, the authors might also have concluded from the same data that the absence of increased GABA in the 'high suppression' condition refutes the very idea that GABA concentrations are related to distractor suppression.
(6) 'Inverse Efficiency' is a highly unusual measure of MOT performance in the literature, and its use reduces the comparability of the findings with previous work. The standard is to assess the correctness ('accuracy') of responses with no focus on speed. This makes sense as responses are given after the object motion has stopped. At the same time, reaction time can be informative too (e.g., Störmer et al., 2013). I think the authors should justify their use of inverse efficiency as the dependent variable.
(7) The choice of variable names is problematic: it is sometimes misleading and makes understanding the findings harder (see also points 1 and 6): obvious, unambiguous, and importantly, interpretation free names for conditions such as target number (2/4), object size (small/large), and total object number (8/12) become load (high/low), target enhancement (high/low) and distractor suppression (low/high). This reduces clarity and, especially in the case of enhancement and suppression, conflates the actual manipulation with its interpretation.
Like nihilism, existentialism starts with a claim that there is no fundamental meaning or morality. But in existentialism, people must create their own meaning and morality. In philosophical terms: “existence precedes essence.” That is, things exist first without meaning or value. They only have value because we choose to create meaning or value for them. Existential ethics: your actions have consequences, but the consequences are unknowable. Even though you desire to do what’s right, there is no objective morality to follow (part of existential angst). You are forced to invent some sort of meaning-giving basis for life that can tell you what to care about. This might include constructing a kind of “morality” to follow.
I like how existentialism emphasizes that there is no inherent meaning or morality and that we have to create our own. It’s interesting because it puts the responsibility for living a meaningful life entirely on the individual, which can feel both freeing and overwhelming. This idea kind of lead me into thinking about how my values and choices are created by myself.
儒(其他关联)# B成为模范人物(例如,仁慈的;真诚的;尊敬和祭祀祖先;尊重各方ents长老和权威人士(照顾儿童和年轻人;对家人和他人慷慨大方)。 这些特质通常是通过仪式和礼节(包括祭祀祖先、音乐和饮茶)来展现和实现的。,从而形成和谐的社会。 关键数据: 孔子中国约500 孟子约350,中国 荀子公元前300年左右,中国 道教# 顺应宇宙自然循环,采取自然而然的行动。强行推动事物发展很可能会适得其反。。 不认同儒家注重礼仪规范,更崇尚自发性和玩乐精神。 就像水(柔软且易变形)经过一段时间可以切割岩石一样。 关键数据: 老子公元前500年左右的中国 老子 庄子公元前300年左右的中国
I thought it was interesting that many of these frameworks try to describe what makes a “good person,” but they don’t always agree about what that actually looks like. For example, Confucianism emphasizes rituals and social roles, while Taoism encourages doing less and letting things unfold naturally. Reading them side-by-side made me realize that ethical behavior can depend a lot on what a culture values, not just on universal rules.
Actions are judged on the sum total of their consequences (utility calculus) The ends justify the means. Utilitarianism: “It is the greatest happiness of the greatest number that is the measure of right and wrong.”
I get why consequentialism is appealing, especially for platforms that rely on metrics like engagement or growth, but it also feels kind of risky. If decisions are made only based on what benefits the majority, harm to smaller or more vulnerable groups can easily be overlooked. In social media, this could mean justifying toxic or harmful content as long as it keeps most users active or entertained.
“Rational Selfishness”: It is rational to seek your own self-interest above all else. Great feats of engineering happen when brilliant people ruthlessly follow their ambition. That is, Do whatever benefits yourself. Altruism is bad.
I think Rational selfishness is becoming more popular in today's culture because of the rise in individualism. I think society has progressed to where people don't need to rely on each other as much to survive, and social mobility is much more achievable. But this comes at the expense of others, if you do whatever benefits yourself at all times, that means you might not care about people and it can and will ruin your relationships. Selfishness is only positive for one's self, but as humans we need community and support so I don't think this is a sustainable mentality.
There is no right or wrong. Nothing matters.
This statement sounds very "radical," as if it could free one from stress, but I think it can easily become a form of escapism: when we say "it doesn't matter," we are often avoiding things we actually care about. Even if there are no absolutely uniform "right answers" in the world, we still make choices every day based on relationships, consequences, and responsibilities—these choices themselves demonstrate that "things do matter to us." Therefore, nihilism can be used to remind me not to be bound by external standards, but it cannot be used as an excuse to "avoid responsibility."
“A person is a person through other people.”
This statement made me think: many of our feelings of "who I am" don't just appear out of thin air, but are shaped within relationships—for example, when we are respected and trusted, we are more likely to become confident and kind; when we are ignored or hurt, we may become more withdrawn. It's not about "you must please everyone," but rather a reminder to consider one more thing when making decisions: will my actions help others feel more like "a complete person"? If it can lead to greater dignity and recognition for both parties, then it's often a better choice.
Being and becoming an exemplary person (e.g., benevolent; sincere; honoring and sacrificing to ancestors; respectful to parents, elders and authorities, taking care of children and the young; generous to family and others). These traits are often performed and achieved through ceremonies and rituals (including sacrificing to ancestors, music, and tea drinking), resulting in a harmonious society.
While these ethics have good intentions, they often do not result in a harmonious society. The reason is that it gives those specific people a lot of power and authority, and people would abuse that authority for their own needs. But since people want to be seen as exemplary people and don't want to disrupt society, they would not dare to question or go against those specific people
Locke: Everyone has a right to life, liberty, and property
I find it interesting that Locke comes up in almost every subject. I also find it interesting in how Locke's words are interpreted and applied in both past and present society. While I don't necessarily disagree with Locke, it should be emphasized that everyone may obtain the right to life, liberty, and happiness, but the way it is distributed or treated in society is extremely unequal towards marginalized groups.
Consequentialism:
I believe that it is always helpful to recognise the existence of utilitarianism within the context of consequentialism via the specific qualities. (All utilitarians fall within consequentialism, but not all consequentialists are utilitarian.) As a specific theory within the broader theory of consequentialism, ultilitarians justify their actions on the scale of maximum utility, hereby attaching quantifiable values to pain and pleasures, while consequentialists recognise that there are different spectrums to base the morality of an action upon!
But on December 17, 2010, a young fruit vendor set himself on fire in a Tunisian village followinga dispute with the local police. The incident sparked protests that were captured on video and seen bymillions across the world. A popular rebellion soon broke out, and twenty-eight days later PresidentZine el-Abidine Ben Ali stepped down after twenty-three years in office.
arab spring begins
R0:
Reviewer #1:
I reviewed the manuscript titled ‘Awareness of anthrax disease and the knowledge of its transmission and symptoms identification: a cross sectional study among butchers in Ile-Ife’ and below are my observations: Abstract The authors were not consistent in the use of tenses. They should stick to the use of past tenses or continuous tenses. In line 43 they mentioned ‘were not aware’ but in 45 used ‘are aware’. Aware ‘of’ is more preferable than aware ‘about’. They were also not consistent in the use of singular or plural words. For instance, in line 46, they referred to ‘animal’ and then ‘animals’. They can stick to the former as a collective noun or the latter as plural. The authors’ conclusion that there was poor awareness is not consistent with the fact that there was no mention of scoring the questionnaire in the methods. In addition, the lack of mention of knowledge which is a part of the title in the abstract leaves a major void. Knowledge should be captured both in the methods and results. Otherwise, it should be deleted from the title. Introduction Remove the space between the punctuation and the citation in square bracket see lines 63, 79 and 112, for instance. Anthrax is a disease (not an agent) caused by Bacillus anthracis not ‘Bacillus Anthrax’. Line 75: Change ‘buy’ with ‘but’. Authors should stick to ‘human or ‘humans’ (Lines 74 and 76). The transition from one paragraph to the other is not smooth. For instance, lines 79 and 80 do not have any connection. In fact paragraph 4 has no place in its current position in the background of the study. The first sentence of paragraph 5 makes no sense and should be recast. The statement that the first case of anthrax in Nigeria was in 2023 is a lie! The authors should make use of the correct words: For instance, ‘body openings’ should be referred to as ‘body orifices’. Line 104: The authors seem to have changed from reporting a study to writing a proposal. Lines 105 – 108 require referencing. Lines 111 and 112: Change ‘has’ to ‘have’. Line 118: ‘Osun - State, South-Western’, should be written ‘Osun State, southwestern’. Line 121: Change ‘it's’ to ‘its’. Line 125: Change ‘accommodates’ to ‘accommodate’. The authors should draw a map of the area being described. Line 134: Change ‘accommodates’ to ‘accommodate’. The inclusion and exclusion criteria are confusing: ‘Butchers who were standing were included’. Does this mean that those butchers who were sitting were excluded? ‘Butchers who were sick were excluded’, therefore only the healthy were included. How did the authors identify healthy and sick butchers? What was the reason for the chosen criteria? Lines 140 - 149: The fact that the authors are contemplating calculating sample size and recruiting respondents for a study that is concluded is absurd. Lines 158 and 159: Change ‘was’ to ‘were’. Line 161: Which random sampling did the authors use to select the butchers? They should be specific in explaining how the sampling method used was applied in the work. The sampling was not well explained. The authors claimed to have used multi-stage sampling technique. What was the sampling frame in each stage and how were they prepared or accessed? The authors did not explain whether and how the questionnaire was validity and reliability tested. Was there a pilot study and how was it conducted? Without these, the instrument is not valid for a meaningful research. How many sections did the instrument contain and how many questions were there in each section? Was the questionnaire scored and how was it scored? Without scoring the questionnaire, the authors cannot determine the levels of awareness or knowledge. Results Line 167: Change ‘questionnaires’ to ‘questionnaire’ because only one questionnaire was used for the study. Line 174: Change ‘rare’ to ‘rear’. The authors claimed to have worked with only commercial butchers. How come most of them have become cattle rearers? Table 1: Change ‘live stock’ to ‘livestock’. Line 180 – 181. Recast the sentence. Discussion The authors seem not to understand that there is a difference between knowledge and awareness, Therefore, it may be worthwhile to define both words in the work. The authors’ claims on knowledge level may not be correct without scoring knowledge to determine the level among the participants. Comparisons made with some author, example Cadmus et al. (2024) do not rhyme.
Limitations Secondary education is not low level of literacy. Therefore, an area that has more than 70% of the population that attained secondary education cannot be regarded as having low level of literacy.
Reviewer #2:
The manuscript “Awareness of Anthrax Disease and the Knowledge of Its Transmission and Symptoms Identification: A Cross-Sectional Study Among Butchers in Ile-Ife” addresses an important public health issue, the awareness and knowledge of anthrax among butchers, that represents a high-risk occupational category in Nigeria. The topic is relevant, timely, also including a One Health perspective. The study provides useful descriptive data that could inform public health education and prevention strategies. The authors present clear findings about the low awareness of anthrax among butchers and the implications for public health. The study is quite good in methodology and analysis, but there are different areas for improvement, before the manuscript could be considered for publication. As regards the study design, it would be appropriate to clarify the sampling frame, randomization process, and justify the adequacy of the final sample size. How were markets and butchers selected? Was any sampling frame used? No basic inferential analyses (e.g., chi-square tests) were performed to identify factors associated with awareness and knowledge. Could you please, add this kind of analyses? Despite collecting rich demographic data and having explicit research objectives to assess awareness, no inferential analyses examine whether education, income, or animal sourcing practices associate with knowledge levels. Chi-square tests and logistic regression would identify high-risk subgroups requiring targeted interventions, fulfilling the study's stated aims and enhancing public health utility beyond simple prevalence reporting The results could be enriched by analysing possible relationships between the respondents' demographics (e.g., education, age) and their awareness or knowledge of anthrax. This would help contextualize the findings. The discussion could be strengthened by addressing any potential biases in the study (e.g., recall bias due to the use of self-reported data). The paper could explore in greater depth why the awareness was low (e.g., cultural factors, lack of effective public health messaging). While the conclusion addresses the need for improved awareness, it could be more specific about what measures should be taken. For example, the authors could suggest specific communication strategies, community-based interventions, or policy changes. The figures could benefit from more detailed captions that explain what each figure illustrates beyond just showing the data. Furthermore, the manuscript contains grammatical errors and spelling mistakes throughout such as “symtoms instead of symptoms in the short title” “Bacillus Anthrax (line 67)” instead of Bacillus anthracis, “unclothed blood (line 83)” instead of “unclotted blood”. Unclotted is better than un-clotted. Line 217 4got? Correct. Write B. anthracis in italics all over the manuscript. Some sentences are unclear or repetitive, especially in the Background and Discussion sections, so a professional English language editing is recommended.
Academic Editor:
The reviewers have raised a number of concerns that need attention. In particular, they request additional information on methodological aspects of the study, additional analysis, a clearly defined research question, and improvements to the quality of the language used.
condensed
Here’s a thorough explanation of “condensed” in both English and 中文, now with appropriate illustrative images from the internet** to help you visualize the meanings. 👇
Meaning: When a gas cools down and turns into a liquid, that liquid is described as condensed. This is a common phase change in the water cycle (vapour → water droplets). (Wikipedia)



👉 English explanation:
👉 中文解释:
Examples:
Meaning: When much of the water is removed from a liquid (like milk), what’s left is thicker and more concentrated — we call it condensed. (Wikipedia)
👉 English explanation:
👉 中文解释:
Examples:
Meaning: When extra details are removed and information is presented in a shorter, tighter form, that version is called condensed.
👉 English:
👉 中文:
Examples:
(Note: Internet images for “condensed text summary diagram” commonly show further visualizations but may be behind paid stock image sites. (Shutterstock))
| Usage (English) | 中文解释 | Key Idea | | ---------------------------- | --------------- | --------------------------- | | condensed water | 凝结水 / 凝结的液体 | gas → liquid | | condensed milk | 炼乳 / 浓缩牛奶 | water removed, concentrated | | condensed summary / text | 精简版 / 简写版 | shorter, fewer details |
If you’d like, I can also generate a printable infographic combining these meanings visually with labels in both English and Chinese. Would you like that? 😊
He saw the world was incomplete and decided to make some human beings. So he took some clay and modeled the figure of a man and laid him on the ground. At first he was very small but grew rapidly until he reached normal size. But as yet he had no life; he was still asleep. Then the Bald Eagle stood and admired his work. “It is impossible,” said he, “that he should be left alone; he must have a mate.” So he pulled out a feather and laid it beside the sleeping man. Then he left them and went off a short distance, for he knew that a woman was being formed from the feather.
The Bald Eagle created man out of clay, a smooth material that turns rough as it hardens, which can be associated with more “masculine” characteristics. In comparison, the Bald Eagle created woman from a feather from his own body, signifying birth and the softness at which more “feminine” characteristics are associated with.
Then the Bald Eagle turned to the man and said, “She is all right now; you and she are to live together.”
There is a theory presented throughout this story, that women are a creation to serve the purpose of accompanying and serving a man and that they may possess no autonomy or free will, is short-sighted; but also, contradicts typical societal structures of many native peoples. Most native societies are--and were--matrilineal and give great value to women and the things that they are capable of beyond their physical presence, and this opposition to typical cultural patterns was interesting to me.
Tempo* launches testnet
Which is a fast internet scale payment focused decentralized blockchain. So again, it's a rail infra for any stable coin - and new companies can start creating their own customer support systems and regulation systems around it. Not easy because of regulations and laws, but it makes the infra of money movement easy since you move it within the wallet. And since you move it on a recognized and trusted blockchain, customers already have a sense of trust with you.
The Year of the Fintech Compounder0102030405FinTech �TrendsAgentic CommerceStablecoins BanksNeobanks06070908PaymentsPrediction MarketsOpen FinanceConsumer �Health10WrapThe big names keep getting bigger. Fintech hyperscalers had a year, with Nubank over 127m customers, Klarna at 114m and Revolut snapping at their heels at 65m. The banks have their mojo back. The IPO market is open. And yet there’s a lingering sense of doom.This was the year of Agentic Commerce, Stablecoins, and prediction markets. Trends that seem destined to reshape the industry, but are still in their early development. Agentic commercce volume is small. Stablecoins are still cross border, and prediction markets are mostly sports. Will that all change?ST
Also available here:
Fake Bots
This example scares me a lot personally. Because how are you as a user expected to know what is real and what is fake? If I saw that first video I would believe it, and I have been taught a lot of source criticism in school. It is becoming increasingly harder for the individual to see through fake content. I know that on Tiktok there is a label if the creator has labeled it as AI-generated, but what if the creator wants to fool their audience like here? I think this example poses a difficult question of how to recognize what is real, and what is fake?
“Gender Pay Gap Bot”
I really find this kind of bot interesting, because it is both good and bad (mostly good in my opinion). It is bad for the companies because it outs them and tells people the real story of how they are not actually supporting women. It is really good for highlighting the inequality that is so very present in the corporate world. I think that this is a provocative bot, but I still find it so necessary in exposing companies who show themselves publicly as supporting women, but not really doing it in reality.
In 2016, Microsft launched a Twitter bot that was intended to learn to speak from other Twitter users and have conversations. Twitter users quickly started tweeting racist comments at Tay, which Tay learned from and started tweeting out within one day.
This immediately reminded me of what happened with Twitter's Ai Grok specifically, but has been seen in other Ai's as well. I'm more curious about the behavior behind the humans that feed these types of comments into chatbots to corrupt them. What is the motivation? We briefly talked about internet and real-life, how are these behaviors shown online versus in-person? If there were a bot in real life that would learn from conversations it had with people, would it go a different direction? Would people still try to teach it outlandish things?
there remains one glaring difference between the US Congress and the European Parliament: in the former there is a powerful “electoral connection” between citizens and elected legislators, whereas in the latter this connection is extremely weak.
Explicitly compares EP to U.S. Congress -> but ELECTORAL CONNECTION to electorate is "remarkably weak" unlike in US -> therefore cannot directly apply theories of what influences Congressional actions to EP, because can't take into consideration the effect of voter punishment / needing to please one's constituents.
" there remains one glaring difference between the US Congress and the European Parliament: in the former there is a powerful “electoral connection” between citizens and elected legislators, whereas in the latter this connection is extremely weak." Actually "almost nonesixtent!"
Aha -> not low voter turnout, but because they only vote for MEPs based on NATIONAL performance of the parties they belong to (think of RN in France -> French voted en masse for them in EP elections because EM government was doing so poorly IN FRANCE -> has nothing to do w/ how EM government is acting in the PARLIAMENT itself.
"Specifically, the election and reelection prospects of almost all MEPs have more to do with the position of the MEPs' national party in the domestic arena—such as the party's governing status, the timing of the European Parliament election in the national electoral cycle, and the performance of the national government—than with how the MEP has behaved or what the MEP has done inside the European Parliament"
Means that in order to win EP seats, domestic parties must please voters DOMESTICALLY instead of specifically in regards to the EP -> unlike in US where parties at House and executive levels are the same / impact same electoral issues.
"The lack of an electoral connection is mainly due to the incentives of national parties to treat European Parliament elections as midterm polls on the policies and performance of the current domestic government. These incentives mean that most national parties would rather campaign on their policies and candidates for domestic office rather than on the record of their politicians, or the behavior of the political group to which they belong, in the European Parliament."
One issue is voting system - Preferrential ballots mean tyhat voters get to choose more directly their candidate for MEP as opposed to just following domestic trends -> some countries have this, others don't. - "Although all member states use a system of proportional representation (PR), about half of the member states use a preferential system, i.e., “open” list PR or single transferable vote. In these countries, voters can choose between candidates from the same party as well as between parties. In contrast, the “closed” list operating for the European Parliament elections in the other half of the member states only offers voters the chance to choose between preordered lists of candidates from each of the parties. For example, MEPs elected under preferential systems tend to spend more time campaigning directly to voters and representing the interests of their constituencies in the Parliament than MEPs elected under closed-list systems (Farrell & Scully 2007). Furthermore, voters are more likely to be contacted by candidates and be better informed about the elections under open-list electoral rules (Hix & Hagemann 2009). Inside the Parliament, MEPs elected under preferential electoral systems tend to be more independent from their national parties and more responsive to the instructions from their European political groups (Hix 2004)." - idk but dw about it
Actions we must explain with alternative means (i.e., not voter pressure): - "the voting behavior of MEPs, the assignments and activities of MEPs in committees, the formation of political parties in the Parliament and coalitions between these parties, and the way the Parliament and MEPs behave in legislative bargains with the EU Council and Commission must be explained by other political motivations, such as the policy objectives or career incentives of MEPs."
MEPs are unlikely to be rewarded or punished in elections for their actions inside the Parliament means that theoretical models cannot simply be taken “off the rack” from the study of the US Congress, where a strong electoral connection is a standard underlying assumption.
DOES compare it to Congress in US -> but notes that THERE IS A WEAK ELECTORAL LINK TO EP -> i.e., low voter turnout or participation / care for who is elected to EP -> very unlike Congress where midterms are generally impactful Here, Cannot APPLY models / theories that concern congress to the EP -> instead MEPS ARE NOT PUNISHED ELECTORIALLY IN THE SAME WAY HOUSE REPBS/DEMOCRATS ARE -> PUBLIC IGNORES THEM SO LESS ACCOUNTABILITY - Says this is IMPORTANT opportunity for researchers to study "electoral" politics w/o needing to consider the influence / pressure of electoral punishment/needing to please one's constituents -> instead must consider motivations for actions taken in EP as being influenced by PERSONAL / CAREER GOALS and ISSUES ABOUT THE ACTUAL POLICIES AT HAND (also likely the influence of pressure groups / lobbyists, if not elecvtorate)
QUESTION -> what kind of data do we have on lobby groups as pressure groups on EP ? Says that we can't rely on explaining MEP motivations based on electoral pressure -> instead only on career aspirations of MEPs and actual policy issues -> but what about non-electoral (i.e., voter) external pressures?
" Finally, the lack of a strong electoral connection in the European Parliament is both a challenge for researchers and an opportunity. The fact that MEPs are unlikely to be rewarded or punished in elections for their actions inside the Parliament means that theoretical models cannot simply be taken “off the rack” from the study of the US Congress, where a strong electoral connection is a standard underlying assumption. But the lack of an electoral incentive also provides an opportunity for European Parliament scholars to develop different theoretical underpinnings of some of the standard explanations for legislative behavior. If the formation of parties or the division of labor in committees cannot be explained by electoral incentives, then other motivations—such as policy concerns or career paths—need to be taken more seriously. If we can understand how career and policy motivations shape legislative behavior in the European Parliament, this knowledge will supplement or perhaps even challenge the general models of legislative behavior that assume that reelection is the dominant goal of legislators."
amendment could come from an interest group, a national government, the Commission, or even the Council.
developed by Kreppel involved identifying each legislative amendment proposed by the Parliament and tracking whether the amendment was then accepted by the Commission and ultimately by the Council, and so became law.
EMPIRICAL METHODS for registering / guaging power of EP - Trickier / more subjective than you might think (how does one measure "power" ?) - Basically TWO options 1. Amendment analysis -> Remember, this is a shuttle / "navette" system -> bills are PROPOSED by the Council, pass through the Commission, and then finally to the Palriament, where amendments are proposed and then sent back for approval (veto power) -> amendment analysis is tracking which bills that ARE amended by the EP actually become law (i.e., if they are accepted by the EC / Council) " "Amendment analysis as applied to the EU legislative process was pioneered by Amie Kreppel as a graduate student of George Tsebelis. The method developed by Kreppel involved identifying each legislative amendment proposed by the Parliament and tracking whether the amendment was then accepted by the Commission and ultimately by the Council, and so became law." -> also takes into account severity of the changes. -> finds that 37 and 28% of amendments by EP were accepted by Commission but only 20% and 19% accepted by Council (which therefore became law) -> strongest (obviously) on more bipartisan issues. Conclude that you can predict the fate of a bill by following the COMMISSION's behaviour 85% of the time, but smaller success rate for predicting based on EP's behaviour -> Therefore EP is LESS powerful than when it had conditional veto procedure (before EU treaty / 1993). "found that “the Commission's behavior predicts the overall fate of an amendment 85% of the time under cooperation and 70% of the time under codecision” (Tsebelis et al. 2001, p. 595). They concluded that when the Commission and Parliament worked together, they were more powerful under the cooperation procedure than under the codecision procedure, as Tsebelis had predicted." - However, Hix notes that OTHER studies have pointed to opposite outcome, particularly after Amsterdam treaty / 2000s
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Summary:
Wethington, Nayak, Jensen et al. investigated changes within protein abundances in distinct NK cell cycle stages after NKG2D stimulation of primary human NK cells and the NK cell line NKL. In addition the authors use mathematical models to define distinct patterns of signaling protein abundances across different cell cycle stages.
Overall, the manuscript is well written and of interest for the scientific community. However, the manuscript could benefit from additional improvements.
Major comments
Minor comments:
General assessment:
The manuscript provides an interesting mathematical modeling as well as CyTOF data from NK cell stimulations about differences in protein abundances throughout different cell cycle stages of NK cells. The data of the NK cell stimulation could be better linked to the mathematical modeling to make a stronger case for the robustness of the model and for more mechanistic conclusions. The manuscript contains a lot of data which is sometimes presented to condensed (Figure 2), the manuscript could benefit from a clearer red line throughout/focus on key molecules.
Audience:
The data presented is of interest for the specialized NK cell community but the discussion section could be improved by making a stronger case of how the herein presented data/model will benefit further studies within the NK cell or general immunology field.
My field of expertise: NK cell biology, tissue-resident NK cells.
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Summary: The paper reports the results of a study that examines how cell cycle stages influence NK cell receptor signaling. The authors find that while most signaling proteins increase monotonically with cell cycle progression, a subset shows non-monotonic variations. Simple computational models are used to explore mechanisms to qualitatively explain the observations.
Major comments:
I am not convinced that the use of models here substantially contributes to the understanding of the observations. The reason is that the results are fairly intuitive and actually to a good extent already well known to those who construct models of reaction kinetics including cell-cycle dynamics. So for example the observation that protein numbers increase mononotically with cell-cycle progression is the obvious thing to expect because since most proteins have a lifetime that exceeds the mean cell-cycle duration then it follows that naturally the protein numbers have to increase during the cell-cycle. This already explains the bulk of the observations. For those proteins where there is non-monotic behaviour, indeed there is something more complex going on but here there are many possibilities. As they say, if we have a 2nd order reaction then the firing rate of bimolecular reactions could increase or decrease with cell-cycle progression because it decreases with cell volume and increases with abundance, both of which factors vary with cell-cycle progression. A model is not quite needed to see that this may lead to non-monotic behaviour. If the model was fitted to the data, i.e. the experimental distributions of protein abundance with cell-cycle progression were fitted to the model and then these are used to constrain possible mechanisms, then yes I would agree that the model brings in some added benefit. Another criticism is the modelling approach itself involves strong simplifications that may not be entirely realistic." : (i) the volume does not seem to change within one cell-cycle stage, e.g. it is 1.3 for all times within the S phase.. "This assumption may be questionable, particularly for cell-cycle stages that occupy a large portion of the cycle." The cell volume generally should vary continuously with time within the cell-cycle and because the propensities are time-dependent then the SSA is not anymore exact and hence one needs to use modifications of it which account for such phenomena. (ii) the doubling of gene copy number due to DNA replication seems to have been omitted from the model. This is expected to lead to a considerable change in the protein numbers at the point in the cell-cycle where DNA replication occurs and hence appears to be an important factor for this study. (iii) how do we reconcile protein concentration homeostasis with the models described in this paper? This is a well known phenomenon, see for e.g. Nature communications 9.1 (2018): 4496 and references therein. (iv) cell-size control mechanisms are not included in the model (adder, sizer, timer); the choice is known to crucially alter protein dynamics across the cell-cycle so difficult to see how one can ignore the inclusion of these. See for e.g. PLoS computational biology 18.10 (2022): e1010574.
Minor comments:
The literature on models of gene expression (mRNA and protein dynamics) including cell-cycle dynamics is extensive and the discussion of this paper would benefit from including more of this. Some of these papers include Biophysical Journal, 107 (2014), 301-313; Journal of theoretical biology 348 (2014): 1-11.; PLoS computational biology 12.8 (2016): e1004972; Plos one 15.1 (2020): e0226016; J. Chem. Phys. 159, 224102 (2023).
This is an interesting paper with both data and modelling. However, presently, the connection between them does not appear strong enough to fully support the conclusions drawn.
By Simón(e) D Sun
The popular belief that your sex arises only from your chromosomal makeup is wrong.The truth is, your biological sex isn’t carved in stone, but a living system with the potential for change.
Important figures cherry-pick what they want the audience to hear or know.
Actual research shows that sex is anything but binary
This is the subject of the article providing facts and research about science being transphobic. The main message this article delivers is how transgender people are people.
Actual research shows that sex is anything but binary
Nearly everyone in middle school biology learned that if you’ve got XX chromosomes, you’re a female; if you’ve got XY, you’re a male.
The author assumes the reader has basic biologic knowledge but not to the point where it is overly advanced. It shows that the text is aimed at more of a general based knowledge audience.
“What AI imperils is not human creativity itself but the ability to make a living from creative endeavor,”
“We’re not making it for that reason but the way ppl choose to use it isn’t our fault. Safeguard will develop.” But tech is moving so fast.
I think this point can well reflect the rapid development of AI nowadays. People often worry that if AI develops to a certain extent, it will replace a large number of existing jobs and raise some moral and ethical issues, such as whether AI will do harm to humans when it develops perception?
“We’re not making it for that reason but the way ppl choose to use it isn’t our fault. Safeguard will develop.” But tech is moving so fast. That there is no way humanity or laws can keep up. We don’t even know how to deal with open death threats online.
I think its really hard to weigh the consequences vs rewards with new tech. part of me thinks that there will always be tools that could be misused but our social contract is what deters people from causing harm. However, this is not always the case, when a tool is developed and distributed that violates freedoms and benefits the few the need for further development before distributing becomes clear.
We also see this phrase used to say that things seen on social media are not authentic, but are manipulated, such as people only posting their good news and not bad news, or people using photo manipulation software to change how they look
I think this is an interesting concept to think about, as we are usually conditioned to think that the internet "isn't real", that most things online are fabricated, exaggerated, etc. However, I do think that just because this is common online, it's not to say that "real life" is a place where everyone is completely authentic and themselves, as some people may feel that they only want to share the good parts of their lives with their friends or family, while keeping anything that wouldn't be considered "good" to themselves, and vice versa. I do think it's hasty to say that all that we see on social media "is not real", as there are plenty of real people behind each account, but we must consider that because people are able to be behind potentially anonymous accounts, it is much easier to fabricate stories or life experiences, or to center one's entire online presence around a portion of their life they want the internet to see, essentially artificially creating an online persona that is not reflective of who they are in real life.
We also see this phrase used to say that things seen on social media are not authentic, but are manipulated, such as people only posting their good news and not bad news, or people using photo manipulation software to change how they look.
I think this idea shows how social media can distort people's lives. When all we see is mostly good news, filters, and edited photos, it's easy to compare ourselves to something that was never real. Over time, this can affect our self-esteem and our expectations of what is "normal". This reminds me that we often forget that social media is more like a highlight reel than real life.
We also see this phrase used to say that things seen on social media are not authentic, but are manipulated, such as people only posting their good news and not bad news, or people using photo manipulation software to change how they look.
I would like to disagree on this because posting about their good news and manipulating their photo are two different things. While one states something that actually happened, manipulating photos is just being deceptive. Additionally, who would actually post about their bad news, given the fact that they might get made fun of by other people, and a majority of those would not receive consequences.
We also see this phrase used to say that things seen on social media are not authentic, but are manipulated, such as people only posting their good news and not bad news, or people using photo manipulation software to change how they look. We’re curious to see how this phrase is continued to be used, and how these sentiments are continuing, being rejected, or evolving.
This idea has always been interesting to me because I feel like I've seen it before, time and time again. The way people act online is way different then how people act in real life and is not an accurate representation of everyone's thoughts. On social media, there will be a bunch of people with the same opinion on a certain topic in my feed, but when I hear people talk about that same topic in real life, their perspectives all vary. When you're only looking at what people are saying online, it can feel like it's the opinion of everyone, but it's not an accurate representation of how everyone is feeling.
We also see this phrase used to say that things seen on social media are not authentic, but are manipulated, such as people only posting their good news and not bad news, or people using photo manipulation software to change how they look.
Social media is highly manipulated to show what we want people to see. Whether that be doctoring photos or only posting sensationalized media. This concept shows that online spaces can create distorted perceptions of reality rather than reflecting authentic life.
Chris, on the other hand, does almost no preparation for his speech, but, being charming and comfortable in front of a crowd, smiles a lot while providing virtually nothing of substance. The audience takeaway from Chris’s speech is, “I have no idea what he was talking about” and other feelings ranging from “He’s good in front of an audience” to “I don’t trust him.” So the moral here is that a well-prepared speech that is delivered poorly is still a well-prepared speech, whereas a poorly written speech delivered superbly is still a poorly written speech. Despite this irony, we realize that delivery is what you are probably most concerned about when it comes to giving speeches, so this chapter is designed to help you achieve the best delivery possible and eliminate some of the nervousness you might be feeling. To do that, we should first dismiss the myth that public speaking is just reading and talking at the same time. You already know how to read, and you already know how to talk, which is why you’re taking a class called “public speaking” and not one called “public talking” or “public reading.”
better to have audience's idea of you as knowing what they're talking about and nervous rather than confidently rambling
For another example, take the right to vote. The secret ballot is one of the most cherished principles of American democracy. We vote in secret so that our choice of a candidate is made freely and without fear of punishment. That is all true, but it is also possible to predict the candidate for whom any one individual will vote if enough is known about the individual. Again, our choice (in this case, our choice of a candidate) is affected by many aspects of our social backgrounds and, in this sense, is not made as freely as we might think.
This paragraph makes me sad. It shows that our social background controls us more than we know. We think we are free, but our family, country, and class already push us toward certain choices. I feel like People who travel to other countries are lucky, because they can see many different ways of living and thinking.
If so, the choice of a mate is not as free as we might like to think it is.
I agree, People often think they’re freely choosing a partner, but they pick based on looks, money, or other outside traits instead of the person’s real personality. It’s like buying a bag because it’s cute but not thinking if it’s functional
The author can also predict the kind of person any one heterosexual reader will marry. If the reader is a woman, she will marry a man of her race who is somewhat older and taller and who is from her social class. If the reader is a man, he will marry a woman of his race who is somewhat younger and shorter and who is from his social class. A reader will even marry someone who is similar in appearance. A reader who is good-looking will marry someone who is also good-looking; a reader with more ordinary looks will marry someone who also fits that description; and a reader who is somewhere between good-looking and ordinary-looking will marry someone who also falls in the middle of the spectrum. Naturally, these predictions will prove wrong for some readers. However, when one takes into account all the attributes listed (race, height, age, social class, appearance), the predictions will be right much more often than they are wrong, because people in the United States do in fact tend to choose mates fitting these general descriptions
I do not fully agree with this paragraph. The author talks like they can predict who a person will marry, but every person has different thoughts, experiences, and choices. People read many different books and meet many different people, so their partner cannot be predicted just from statistics. Marriage is a personal decision, not only a social pattern.
And yet perhaps we have less freedom than we think. Although we have the right to choose how to believe and act, many of our choices are affected by our society, culture, and social institutions in ways we do not even realize
I agree. America has many opportunities, but our freedom is limited by social and economic pressure. If you don’t have money or don’t work hard, you can end up homeless, and the system doesn’t always protect you. That means our choices are not as free as they look.
If your blog is installed in a subdirectory but you’ve set a different wp_siteurl, you don’t need the redirect — index.php will handle it automatically.
wp_siteurl will solve the redirect issue of having the blog in a subdir that's diff from where WP itself is. What about my set-up that uses the domain root too.
Using a subdirectory for your blog For WebFinger to function properly, it needs to be mapped to the root directory of your blog’s URL.
I have a setup where WP is in a different directory than my blog's url, but where also the main domain shows the blog.
The glottis is the opening inside the larynx, which also contains the vocal folds or cords. Air passes through this opening when breathing and making speech.
Maybe a quick functional discussion stating that the glottis is widened when breathing, but narrowed during phonation to allow the passing air to vibrate the vocal chords?
Article by [[Doug Belshaw p]] on how changes doesn't come from telling people they're wrong but from using crises to shift group behaviour.
Stories can produce results that slogans and demands can't. Reality, in the form of how people actually respond, pushed Weymouth to correct his formula.
he changed his formula, but did not correct it. In fact nothing in this text suggests there was even any impact of his change in actions, nor that it had more impact as the text implies than his previous actions.
Aristotle called practical wisdom phronesis. It's the ability to discern what works in particular situations, not through abstract principle alone, but through experience and judgement. It's knowing what to do, and when, in specific circumstances.
context is important in pragmatism, also to avoid reductio ad absurdum
This is why I count myself as a Pragmatist. It's a philosophical tradition that views language and thought as tools for problem-solving and action, not for trying to create a mirror of reality. At its core is an understanding that you cannot know the world without having a perspective. This does not make all perspectives somehow “equally valid” but instead means that you should test your approaches against reality, observe what works, revise what doesn't, and gradually get better at navigating the world as it actually is.
a more usefully worded def of Pragmatism as philosophical school than usually encountered. Vgl [[Filosofische stromingen als gereedschap 20030212105451]] [[Pragmatisme Rorty 20200924110612]]
Original 'Vulcan group' (2011) says they're not the ones who sent a letter taking resp for the Sept and Dec 2025 arson attacks on Berlin energy infra. Prev saw an analysis by a linguist that the December letter wasn't written by native speakers, but seemed translated (not saying what from)
edges
cheating/squaring zooko's triangle
DECENTralized is decent but not enough Need interpersonal verifiably named people \ resource \ co-laboration networks the IndyWeb.
use names that are acnhoed in the petname of the person sharing the resource!
bt
but
open sourcesauce
should be
Open Constructs - that are not only Open Source - but the sources shared are developed by making the secret that makes it wall work Open, so it is Open Sauce - and developed so that it is easy to understand, extend, compelling to use and easy to emulate
Zooko’s Triangle states that we can’t have all three.
Oh Yes We Can
DECENTralized is decent but not enough
use names that are acnhoed in the petname of the person sharing the resource!
We can’t give every example, but here is a range of different things social media platforms do (though this is all an oversimplification).
A major issue with “social media” and trying to apply ethical safeguards to it, is that social media is a vast collection of digital platforms. This vastness makes social media hard to define, and thus it becomes difficult to address issues on a wide scale, for example through law. Additionally, social media varies across different regions, peoples, and languages, and similarly, so do ethics.
Platforms can be minimalist, like Yo, which only lets you say “yo” to people and nothing else. Platforms can also be tailored for specific groups of people, like a social media platforms for low-income blind people in India.
I think it was interesting to put a minimalist platform like Yo together with a platform specifically for low-income blind people. The former looks "simple and easy to use," but the fact that it has fewer features means it can't do much. Specialized platforms, on the other hand, are more complex but really help those who need it most. It made me think: Platform design really depends on who you want to serve.
Meanwhile, 31% of 8-12 year olds have spoken with large language models. 23% have talked to strangers online, while only 44% have physically spoken to a neighbor without their parents. 50% have seen pornography by the time they turn 13.
Kids are independent in the digital world before the physical one.
That flips the historical order. Earlier generations learned:
Now it’s:
This matters because:
**Digital spaces forgive mistakes cheaply
Physical spaces teach consequences viscerally** - “Physical spaces teach consequences viscerally”
In physical space, mistakes hit the nervous system, not just the mind.
Run too fast → you fall. Ignore a curb → you trip. Misjudge a car → you don’t get a second try. Shout too loud → heads turn. Cross a boundary → someone reacts in front of you.
The feedback is:
Immediate
Embodied
Often irreversible
Digital spaces teach what you can get away with. Physical spaces teach what actually happens.
Both are necessary. But they are not interchangeable.
One final note we’d like to make here is that, as we said before, we can use ethics frameworks as tools to help us see into situations. But just because we use an ethics framework to look at a situation doesn’t mean that we will come out with a morally good conclusion.
I feel like this statement in itself is a little contradictory to what this entire chapter is about. We've learned about multiple different kinds of ethical frameworks there are, which in turn means that we've learned that what might be morally right and wrong differs for everyone. Nihilism in itself rejects the existence of morally good conclusions, so how can we say that just because we use an ethical framework to look at a situation, doesn't mean we will come out with a morally good conclusion, when everyone's definition of a "morally good conclusion" differs? I initially agreed with the statement, but upon further processing, I think it'd be much harder to firmly decide a situation to be a morally bad conclusion since the opinion can fluctuate depending on who you ask and where. It doesn't make much of a difference to consider someone's ethical framework if it differs from our own if we're still going to make a definitive conclusion that their situation is morally bad from our own framework.
There is no clear single definition for what counts as social media. John Hartley points out that you could consider almost all of culture as “social media.”
I thought this was a really interesting concept, it's true and it makes sense. All culture, all we digest all our society is and can be considered social media. But when we think about social media, we think of media in the technological sense as an app.
How do you think about the relationship between social media and “real life”?
In contrast to how people are saying "social media isn't real life," I think that people tend to be a more exaggerated version of themselves when engaging online. I think this might be because they don't feel an immediate consequence for poor or extreme behavior alongside the fact that it's very easy to just say whatever you want and click post. It's easy to post impulsive thoughts and not think about how others would react, especially when you're not physically seeing any effects in your real life. So in a way, I think that social media is a part of our real life, but in a much more exaggerated way now that social pressure is generally lightened.
Alicia enjoys her classes, even when she has to get up early after working or studying late the night before. She sometimes gets so excited by something she learns in class that she rushes up to the instructor after class to ask a question. In class discussions, she’s not usually the first to speak out, but by the time another student has given an opinion, she’s had time to organize her thoughts and enjoys arguing her ideas. Nearing the end of her sophomore year and unsure of what to major in given her many interests, she talks things over with one of her favorite instructors, whom she has gotten to know through office visits. The instructor gives her some insights into careers in that field and helps her explore her interests. She takes two more courses with this instructor over the next year, and she’s comfortable in her senior year going to him to ask for a job reference. When she does, she’s surprised and thrilled when he urges her to apply for a high-level paid internship with a company in the field—that happens to be run by a friend of his.
Alicia is a model student and goes to her classes in person all the time and keeps up with the work.
Karen wants to have a good time in college and still do well enough to get a good job in business afterward. Her sorority keeps a file of class notes for her big lecture classes, and from talking to others and reviewing these notes, she’s discovered she can skip almost half of those big classes and still get a B or C on the tests. She stays focused on her grades, and because she has a good memory, she’s able to maintain OK grades. She doesn’t worry about talking to her instructors outside of class because she can always find out what she needs from another student. In her sophomore year, she has a quick conversation with her academic advisor and chooses her major. Those classes are smaller, and she goes to most of them, but she feels she’s pretty much figured out how it works and can usually still get the grade. In her senior year, she starts working on her résumé and asks other students in her major which instructors write the best letters of recommendation. She’s sure her college degree will land her a good job.
Karen is doing good on going to most of her classes but she relies on her sorority to give her notes but still is not good because she can not talk to an instructor.
Carla wants to get through college, and she knows she needs the degree to get a decent job, but she’s just not that into it. She’s never thought of herself as a good student, and that hasn’t changed much in college. She has trouble paying attention in those big lecture classes, which mostly seem pretty boring. She’s pretty sure she can pass all her courses, however, as long as she takes the time to study before tests. It doesn’t bother her to skip classes when she’s studying for a test in a different class or finishing a reading assignment she didn’t get around to earlier. She does make it through her freshman year with a passing grade in every class, even those she didn’t go to very often. Then she fails the midterm exam in her first sophomore class. Depressed, she skips the next couple classes, then feels guilty and goes to the next. It’s even harder to stay awake because now she has no idea what they’re talking about. It’s too late to drop the course, and even a hard night of studying before the final isn’t enough to pass the course. In two other classes, she just barely passes. She has no idea what classes to take next term and is starting to think that maybe she’ll drop out for now.
Carla is setting her self up for failure because she dose not go to class and relies on other for notes and cant ask questions.
Carla wants to get through college, and she knows she needs the degree to get a decent job, but she’s just not that into it. She’s never thought of herself as a good student, and that hasn’t changed much in college. She has trouble paying attention in those big lecture classes, which mostly seem pretty boring. She’s pretty sure she can pass all her courses, however, as long as she takes the time to study before tests. It doesn’t bother her to skip classes when she’s studying for a test in a different class or finishing a reading assignment she didn’t get around to earlier. She does make it through her freshman year with a passing grade in every class, even those she didn’t go to very often. Then she fails the midterm exam in her first sophomore class. Depressed, she skips the next couple classes, then feels guilty and goes to the next. It’s even harder to stay awake because now she has no idea what they’re talking about. It’s too late to drop the course, and even a hard night of studying before the final isn’t enough to pass the course. In two other classes, she just barely passes. She has no idea what classes to take next term and is starting to think that maybe she’ll drop out for now.
I think Carla is trying to do the "quicker way". But in reality it's not the fastest way it is in my opinion is the lazier way of going about things in a productive and efficient way.
Behold this and always love it! It is lela wakan [very sacred], and you must treat it as such. No impure man should ever be allowed to see it, for within this bundle there is a sacred pipe. With this you will, during the winters to come, send your voices to Wakan- Tanka, your Father and Grandfather.”
The warning given by the wakan woman to Chief Standing Hollow Horn shows how sacred and important the pipe is. The pipe is not just an object but a way to speak to Wakan Tanka and to his ancestors. Only pure, honest, and respectful people are allowed to see or use it. This shows the value of truth and careful behavior. The quiet winter setting feels empty and lonely, and the silence is very strong. However, the sacred pipe brings life, comfort, and strength. When the pipe is used, it carries prayers and voices to his father and grandfather, showing that their support is always present, even during times of silence and hardship.
Be generous and resepectful to your parent and siblings by taking the hard decision and absorbing whatever upset follows. It might be a substantial sacrifice to let your parent be angry with you, but an exemplary person would respectfully accept the difficulty in order to secure proper care for their parent. Be respectful of your parent’s authority and don’t intervene and try to take the decision away from them. Instead be generous in helping provide for your parent while your parent makes their own decision
The ethics of Confucianism more relies on the legitimacy (mingfen). Because with mingfen, the action will be legimacy. Therefore, with the mingfen of xiao (filial piety), the intervene from Children will be intervene.
It means that we have even more tools at our disposal for thinking through the ethical considerations packed into a situation, as we learn from those who see things differently.
This sentence resonated with a question I often think about. At times, I regret or even reprimand myself for initially disagreeing with an opinion that I later realize is very reasonable. This passage helps me understand that such reactions do not necessarily reflect a flawed ethical framework, but rather limited exposure to diverse perspectives. It suggests that ethical growth comes from learning to engage with new viewpoints and acquiring additional “tools” for ethical reasoning.
Detailed information on fetal growth, such as that derivedfrom serial ultrasound scans, is difficult to obtain. Epide-miological studies instead use proxy measures, such as birthweight, gestational age, and, less often, birth length, headcircumference, placental weight, or ponderal index (4). Wefocus here on birth weight, which is the most commonlystudied measure of fetal development, but it is important tonote that birth weight, gestational age, and birth length mayreflect different underlying mechanisms with independenteffects on specific mental health outcomes
Fetal growth is challenging to assess directly, so researchers rely on proxy measures such as birth weight, gestational age, birth length, head circumference, placental weight, and the ponderal index. However, these indicators may capture different biological processes and can therefore reflect distinct underlying mechanisms, each with independent associations with specific mental health outcomes.
For example, low birth weight may reflect inadequate fetal nutrition, whereas reduced head circumference may indicate altered brain development; each may be linked to different risks for later mental health conditions. In addition, differences seen can be attributed to genetics instead of maternal health.
The quality of fetal growth and development predicts the riskfor a range of noncommunicable, chronic illnesses. Theseobservations form the basis of the “developmental origins ofhealth and disease” hypothesis, which suggests that the in-trauterine signals that compromise fetal growth also act to“program” tissue differentiation in a manner that predisposesto later illness. Fetal growth also predicts the risk for laterpsychopathology. These findings parallel studies showingthat antenatal maternal emotional well-being likewise pre-dicts the risk for later psychopathology. Taken together, thesefindings form the basis for integrative models of fetal neu-rodevelopment, which propose that antenatal maternaladversity operates through the biological pathways associ-ated with fetal growth to program neurodevelopment. Theauthors review the literature and find little support for suchintegrated models. Maternal anxiety, depression, and stressall influence neurodevelopment but show modest, weak, orno associations with known stress mediators (e.g., gluco-corticoids) or with fetal growth. Rather, compromised fetaldevelopment appears to establish a “meta-plastic” state thatincreases sensitivity to postnatal influences. There also re-main serious concerns that observational studies associatingeither fetal growth or maternal mental health with neuro-developmental outcomes fail to account for underlyinggenetic factors. Finally, while the observed relation betweenfetal growth and adult health has garnered considerableattention, the clinical relevance of these associations remainsto be determined. There are both considerable promise andimportant challenges for future studies of the fetal origins ofmental health
This is a review article where the authors refute the claim that maternal stress affects the neurodevelopment of her fetus.
They argue that the literature doesn't support this model of neurodevelopment and that the evidence supporting this claim is modest, weak, or insignificant.
They argue that compromised fetal development makes the fetus ultra susceptible to post-natal influences. That is, they are put into a 'meta-plastic' state where they are vulnerable to such influences.
They also emphasize a weakness in studies of the relationship between maternal stress and fetal neurodevelopment citing a failure to account for underlying genetic factors that could also potentially explain the abnormal development. In other words, instead of maternal stress affecting neurodevelopment of a fetus, it could instead be caused by some underlying genetic factor.
Word spread, and Justine’s tweet went viral. Twitter users found other recent offensive tweets by Justine about countries she was traveling in. IAC (Justine’s employer) called the tweet “outrageous, offensive” but “Unfortunately, the employee in question is unreachable on an international flight.” Twitter users, now knowing that Justine is on a flight, started the hashtag #hasjustinelanedyet, which started trending on Twitter (including some celebrities tweeting about it).
I'm kind of shocked about this, it's alarming how tweets spread because retweeting is kind of like a fungus that multiplies, once you repost, others do it too, and when people repost on Facebook or Instagram or X now, other people see it. This is the problem in today's world, we don't see the power of us retweeting, and it might not always be good, like what happened here, and that's what people often do not realize. Social media needs to do a better job with the repost, and people should consider not reposting anymore.
This article is more about U.S. policy on Venezuela but contains some interesting points about the administration's economic policy proposals. I think this is also a useful overview of the whole situation.
RR\ID Summary of Reviews: This preprint presents findings from a single-center study suggesting that subclinical HBV infection is frequent in infants born to HBsAg-positive mothers, despite receiving standard prophylaxis (HBIG and vaccination). Using highly sensitive PCR methods, the study detected HBV DNA in immune cells of vaccinated neonates who tested negative for HBsAg, raising concerns about the potential for occult infection. Reviewers acknowledge the novelty of detecting intra-cellular HBV in this context but question the high reported prevalence (over 80%), citing critical methodological concerns including timing of testing, reliance on nested PCR, and absence of key controls. While the findings reignite debate over the definition and relevance of occult HBV, reviewers emphasize the need for cautious interpretation and reiterate that current vaccination strategies remain highly effective at preventing overt chronic infection.
Read directly in RR\ID: https://rrid.mitpress.mit.edu/pub/ded6ilxx/release/2
Reviewer #1: Evidentiary Rating: Potentially Informative
Written Review: The paper demonstrates a high incidence of subclinical HBV infection in infants born to HBsAg-positive mothers despite prophylactic measures (immunoglobulin treatment and vaccination). The neonates did not develop evident HBV infection characterized by HBsAg serum positivity, but HBV-DNA in different immune cells was detected using a highly sensitive PCR method.
The data appear robust, though I would recommend the inclusion of a substantial number of negative controls (blood from newborns of healthy mothers) with experiments conducted in a blinded manner using identical methods. Such negative controls would significantly increase the impact of the findings.
The paper is also excessively long. Identical concepts are repeated multiple times throughout the introduction, results, and discussion sections. Additionally, some results are presented twice.
Several concepts need to be refined or better articulated. For instance, the detection of nucleotide differences in the PreS/S region of HBV isolates from mother and child is discussed as evidence of "creation of HBV isolates in the placenta." Beyond the fact that the term "creation" is particularly ambiguous, the variants detected in newborns could simply reflect HBV evolution in an environment with a less dysfunctional immune system than the mother's—one that shares only half of the mother's HLA class I and class II alleles. This could represent straightforward adaptation to escape a new immune system rather than viral creation in the placenta.
I would also recommend less dramatic language when discussing the consequences of these findings. The authors state: "the documented ability of HBV, as well as traces of WHV [28], to integrate into the host's genome almost immediately after infection and persistence of potentially oncogenic virus-host genome integrations may have serious life-threatening consequences." However, the rate of HCC development in individuals who are HBsAg and HBV-DNA negative but with "occult HBV" remains low. I recommend adding a proper reference reporting the incidence of HCC development in patients without overt liver disease who are HBsAg and HBV-DNA negative.
It is also interesting that the conclusive sentence calls for "the tremendous importance of global HBV vaccination, in particular vaccination of infants and young adults, to avert virus vertical spread and protect future generations", even though the data, if taken literally, show that such prophylactic measures are not effective. I would recommend being more pragmatic in the discussion of the impact of such subclinical infection in the general population and to stress that vaccination still blocks the development of real chronic HBV infection with high levels of HBV replication.
It’s wild to think about how new technologies are changing the way we think about teaching and learning. The digital tools many students have access to both inside and outside the classroom require us all to take a hard look at the way we use these tools in the context of learning experiences. It’s easy to get caught up with the shiniest, brightest, or most attention-grabbing digital device or website, but it is possible to pause, reflect, and prioritize tasks over digital tools in the classroom. Are we putting the learning first?
Though, as others have stated this article is a bit dated, the concepts and ideas can still apply to tech tools of today as many of the apps that my students use started many years prior to this article but have advanced in the times. So this opening statement still rings true.
It is OK if more than oneperson answers the same seeded question.
I am interested to see how this works when it comes to specific papers. Will some students read the same information but interpret it differently? How similar or dissimilar will the questions be? I think the variation of scientific and academic backgrounds will allow for a wide array of perspectives to arise from a singular question.
I switched from VSCode to Zed
[tool.pyright] section in pyproject.toml effectively force Basedpyright’s recommended mode.typeCheckingMode in Zed’s settings.json did not help; the fix was explicitly setting typeCheckingMode = "standard" inside each project’s [tool.pyright] config."disablePullDiagnostics": true in Zed’s Basedpyright initialization options.ty language server, found it good, but stayed with Basedpyright to match CI’s Pyright.settings.json showcasing autosave, disabled inline blame, VSCode keymap, fonts, light theme, and customized Basedpyright LSP options.Note: This response was posted by the corresponding author to Review Commons. The content has not been altered except for formatting.
Learn more at Review Commons
We thank the Reviewers for their positive assessment of the quality and significance of our work, as well as for their insightful comments, which have helped us to further improve the manuscript. We have addressed the majority of the comments in the revised version and, for those that require additional time, we outline below a detailed plan of the experiments we intend to perform.
We agree with Reviewer #2 that a more detailed mechanistic understanding of the drug effects would further strengthen the study, and we are grateful to both reviewers for the constructive experimental suggestions provided to address this point. In particular, we are highly motivated to better define the causal role of C18 sphingolipid alterations in mediating the effects of the drugs, as suggested by Reviewer #2, as well as to investigate the involvement of the retromer complex in the lysosome-to-Golgi connection, as suggested by Reviewer #1.
Below, we provide a point-by-point description of the revisions already incorporated into the manuscript, along with the planned experiments that will address the remaining comments
REVIEWER #1:
VPS13B is a bridge-like lipid transfer protein, the loss or mutation of which is associated with Cohen syndrome (CS) involving Golgi fragmentation. In this study, the authors performed image-based chemical screens to identify compounds capable of rescuing the Golgi morphology in VPS13B-KO HeLa cells. They identified 50 compounds, the majority of which are lysosomotropic compounds or cationic amphiphilic drugs (CADs). Treatment of cells with several of these compounds causes lysosomal lipid storage, as assessed by BMP/LBPA staining, filipin staining, or LipidTOX staining. Interestingly, most LipidTOX puncta colocalized with transferrin receptor-positive compartments but not lysosomes. Similar to lysosomotropic compounds, knocking down NPC1 or SMPD1, mimicking lysosomal storage disease, also substantially rescued Golgi morphology. The authors show that VPS13B-KO cells have reduced C18 sphingolipids, which is reversed by treatment with CADs. Finally, the authors show that two CADs partially rescue neurite outgrowth in neuronal cultures. However, these drugs do not rescue the size of VPS13B KO organoids.
Overall, this is an impressive study identifying CADs as potential therapeutics for CS and suggesting sphingolipid upregulation as a general strategy for CS treatment. The morphological and lipidomics analyses unravel important molecular basis of CS pathology. This study will be of high interest to the field of lipid biology and organelle homeostasis. I have a few comments to help improve the quality of this study.
- The reverse of lipid changes in VPS13B-KO cells by CADs is intriguing. Are CAD-mediated benefits such as Golgi morphology recovery permanent or only transient within 24 hours of treatment? How do the CADs affect the Golgi morphology in WT HeLa cells?
RESPONSE:
We thank the reviewer for this insightful question Indeed, the effects of CADs on Golgi organization are most evident in VPS13B KO cells, where the Golgi apparatus is severely fragmented and becomes more compact upon drug treatment, whereas the effect is much less apparent in wild-type cells. Nevertheless, a careful quantitative analysis of the images (now presented in the new Fig. S7) demonstrates that the impact of these compounds on Golgi morphology is not restricted to KO cells but is likely more general, supporting a link between lysosomal storage and Golgi organization. Although this observation indicates an indirect effect (consistent with the proposed mechanism of action), rather than a direct correction of VPS13B loss, it does not compromise in our opinion their potential beneficial effect for KO cells as shown also from the results obtained in organoid-derived neurons.
Under continuous treatment, azelastine keeps the Golgi in a compact state for 72 hours without any noticeable deleterious effect on the cells (see new Fig. S10) Raloxifene, on the contrary proved to be toxic over the same time period. We believe this difference reflects the mechanism of action of CADs, which progressively accumulate within acidic organelles and may eventually reach a toxic threshold upon prolonged exposure. For this reason, lower drug concentrations administered over longer treatment periods may represent a viable alternative strategy. In this regard, we also refer the reviewer to our response to the comment on brain organoids below.
- Is it surprising that Azelastine-induced lipid storage in transferrin receptor compartments (early and recycling endosomes)? I suggest more controls to examine LipidTOX overlap with Golgi markers or other late endosome/lysosome markers such as LBPA and CD63.
RESPONSE:
We agree with the reviewer that this observation is somewhat unexpected. However, we would like to clarify that we do not intend to suggest that lipid storage occurs primarily in early or recycling endosomes, which would indeed contradict a substantial body of existing evidence. Rather, our data indicate that this particular dye (LipidTOX) labels recycling endosomes, at least in HeLa cells. This finding is consistent with the widely accepted view that lysosomal lipid storage exerts broader effects on intracellular trafficking, not limited to late endosomes/lysosomes. We corrected the text in order to clarify this concept.
LipidTOX was specifically developed to detect drug-induced phospholipidosis, and based on our data, it appears suitable for this purpose. To our knowledge, there is no published information detailing its intracellular localization, which motivated us to perform these control experiments. Unfortunately, the proprietary formulation of this product does not allow informed speculations to explain the observed localization or whether this could refer to the intact molecule or to a catabolite.
As suggested by the reviewer, we plan to perform co-staining with additional markers to further clarify this this point.
- Does the LipidTOX/TFRC overlap suggest potential roles of retrograde transport in supplying sphingolipids to the Golgi? The authors can quickly test if the knockdown of a retromer subunit (VPS35) blocks Azelastine-induced recovery of Golgi morphology.
RESPONSE:
We thank the reviewer for this insightful suggestion. Indeed, the retromer complex represents one of the best-characterized trafficking pathways from the endosomal system to the Golgi, and this relatively straightforward experiment could help to mechanistically clarify our observations. We plan to test whether VPS35 knockdown interferes with the effects of the drugs.
What is the rationale to use 500 nM to 1 uM azelastine and raloxifene for neuronal cultures and organoids? At such concentrations, no obvious changes in Golgi morphology or lipid storage were observed (Fig 4). Also, the lipidomics analysis was performed after 10 uM compound treatment. It might be worth trying dose-response experiments in organoid tests.
RESPONSE:
We thank the reviewer for this question. The rationale about this choice was indeed missing from our previous version of the manuscript. The reason of lowering the concentrations comes indeed from toxicity tests, preliminarily performed over long-term treatment of both WT and VPS13B KO organoids. This information has now been explicitly included in the Results section of the revised manuscript, and the broader implications are also discussed in the Discussion section.
MINOR COMMENTS:
It is important to know whether the authors used TGN or cis-Golgi markers for Golgi morphology analysis. Please label the two channels in Fig. 2C and throughout all figures. In many cases, it is not clear what is stained in the green channel to show the Golgi morphology. It was not even stated in the legend.
RESPONSE:
We now included the antibody staining in all figure legends where it was previously missing.
The authors stated that Recovery of Golgi morphology is dependent on lysosomal lipid storage. However, while the data show positive correlation between the two, no causal relationship is established by the data. It seems true that in all conditions (CADs or genetic knockdown) where lysosomal lipid storage was observed, the authors detect the Recovery of Golgi morphology. However, budesonide did not depend on lysosomal lipid storage to recover the Golgi morphology. Thus, the recovery of Golgi morphology is NOT dependent on lysosomal lipid storage, but inducing lysosomal lipid storage appears sufficient to recover Golgi morphology in VPS13B-KO HeLa cells.
RESPONSE:
We thank the reviewer for this comment and we agree that the previous title of the paragraph could have been misleading. This has been now changed in: “Lysosomal lipid storage mediates the recovery of Golgi morphology” which is probably less prone to ambiguous interpretations.
Obviously, in the previous version of the title we wanted to mean that Golgi recovery is dependent on lipid storage “in the context of CAD treatment” and not as a general statement.
With respect to the cause–effect relationship, we believe that the strongest evidence supporting this link is the observation that genetically induced lipid storage phenocopies the effects of drug treatment. We hope that this conclusion is now sufficiently clear from the revised text.
Each figure needs a title before the detailed legends for specific panels.
RESPONSE:
Titles have now been included to all figure legends.
Fig 8. Y axis labeling is missing.
RESPONSE:
Axes labels have now been included
Does U18666A rescues Golgi morphology in VPS13B-KO cells?
RESPONSE:
We thank the reviewer for this comment. U18666A indeed also corrects Golgi morphology. The result is now included in the new figure S5.
Please do not repeat the result section in discussion. Focus on the most important points.
RESPONSE:
We thank the reviewer for this comment. We shortened the descriptive part of the discussion trying as much as possible to avoid repetitions with the result session and keeping only the more essential information for the flow of the discussion.
Reviewer #1 (Significance (Required)):
This is an impressive study that identifies Cationic Amphiphilic Drugs (CADs) as potential therapeutics for Cohen syndrome (CS) and suggests sphingolipid upregulation as a general strategy for diseases driven by VPS13B loss-of-function. The unbiased approaches, notably the chemical screen and lipidomics, provide novel mechanistic insights into the underlying pathology of CS. This study will be of high interest to researchers in the fields of lipid biology and organelle homeostasis. It will also be highly valuable for clinical pediatricians managing CS patients.
REVIEWER #2:
This manuscript describes a compound screening aimed at identifying molecules that can restore Golgi organization in VPS13B knockout (KO) cells. The authors identify several compounds, most of which are lysosomotropic, and analyze their effects on Golgi morphology and lipid composition using multiple approaches. They report that VPS13B KO cells exhibit a reduction in C18-N-acyl sphingolipids, which can be restored by several of the identified compounds. Furthermore, two of these compounds, azelastine and raloxifene, promote neurite outgrowth in VPS13B KO cortical organoids. These findings are interesting and could potentially contribute to a better understanding of the pathophysiology of Cohen syndrome and the development of therapeutic strategies. However, despite the large number of analyses presented, the study remains largely descriptive, and there is no coherent mechanistic explanation for how these compounds restore Golgi structure in VPS13B KO cells. In addition to the reduction in C18-N-acyl sphingolipids, the KO cells display alterations in several other lipid species (LPC, LPE, PC40:1, PE42:1, TG, etc.), and treatment with the selected compounds induces further lipid accumulations, including cholesterol and BMP/LBPA. The relationship between these diverse lipid changes and the observed Golgi recovery lacks clarity and mechanistic consistency.
MAJOR COMMENTS:
The finding that compounds cannot prevent Golgi fragmentation caused by brefeldin A or nocodazole but can suppress statin-induced fragmentation is intriguing, but the underlying mechanism is not addressed. It is not evident whether this difference results from changes in membrane lipid composition or restoration of Rab/SNARE trafficking. The authors should examine Rab prenylation and SNARE localization by immunofluorescence or Western blotting to support their interpretation.
RESPONSE:
We thank the reviewer for this suggestion and agree that the ability of these compounds to counteract statin-induced Golgi fragmentation is indeed intriguing. The primary reason we did not further explore this aspect is that we evaluated the effects of statins not to be a central focus of the present study. Nevertheless, we fully agree that this observation represents a valuable opportunity to gain additional insight into the mechanism underlying drug-induced Golgi recovery.
To address this point, we plan to analyze Rab prenylation by Western blot and Rab localization by microscopy, focusing on a Golgi-associated Rab protein such as Rab6. In addition, we will employ downstream inhibitors of Rab prenylation, such as 3-PEHPC (an inhibitor of type II protein geranylgeranyltransferase (GGTase-II)), which should allow us to formally distinguish effects related to impaired Rab prenylation from those arising from inhibition of cholesterol biosynthesis.
Although restoration of C18 sphingolipids (SM 36:1, CER 36:1) is observed upon compound treatment, its causal role in Golgi recovery or neurite outgrowth is not established. The authors should test whether blocking the increase of C18 SM/CER prevents the rescue of Golgi or neuronal phenotypes.
RESPONSE:
We sincerely thank the reviewer for this comment. We agree that, based on the current data, a definitive cause–effect relationship between Golgi recovery and the increase in C18 sphingolipids cannot be firmly established, and we acknowledge that a deeper understanding of this issue will require further investigation. Furthermore, we believe that addressing this would not only provide a better mechanistic understanding of the biological processes behind the effect of the drugs but provide a potential avenue for therapeutic intervention. For these reasons, we are strongly motivated to pursue this aspect further.
With respect to the reviewer’s specific suggestion, we agree that preventing the increase in C18 sphingolipids would be an ideal experimental approach. However, the limited understanding of the regulatory mechanisms controlling C18 sphingolipid homeostasis currently precludes a fully informed strategy. In principle, if the observed increase were due to enhanced synthesis, one could envisage blocking it by silencing ceramide synthases with C18 selectivity, such as CERS1. The experiment shown in Fig. 7E (azelastine treatment in the presence of sphingolipid synthesis inhibitors) was designed with this rationale in mind. However, these results suggest that azelastine-induced C18 sphingolipid accumulation is unlikely to result from increased synthesis, and is instead more consistent with reduced degradation, in line with the proposed mechanism of action of CADs.
Based on these considerations, we propose to invert the experimental approach and test whether cellular re-complementation with C18 sphingolipids is sufficient to recapitulate the drug-induced Golgi recovery. We are aware of the technical challenges associated with the targeted delivery of exogenously supplied lipids, particularly given the likelihood that effective rescue would require lipid access to the Golgi apparatus. Based on current knowledge, we anticipate that externally supplied lipids would primarily traffic either to the ER via non-vesicular routes or to endosomes/lysosomes through endocytic uptake. From both locations they could eventually reach to some extent the Golgi. The route from endosomes to Golgi in particular as been intensively studied in the past with the use of fluorescent sphingolipid analogs1,2 and may well work also with native lipids.
Since we are not able to predict in advance which lipid species would be more effective or the optimal delivery strategy, we plan to test re-complementation using C18 sphingomyelin and some of its potential precursors, including C18 ceramide as well as using alternative delivery strategies such as incorporation in liposomes of different formulations and delivery at the plasma membrane with bovine serum albumin or cyclodextrins as carriers.
In Figure 7D, comparisons should include the LM and HM fractions isolated from WT cells.
RESPONSE:
Wild-type control were included in the figure as requested.
The subcellular fractionation experiment should be repeated using AZL and RAL, the compounds used in organoid experiments, rather than TFPZ, to assess whether similar results are obtained. The compounds used differ across experiments, making it difficult to draw consistent conclusions.
RESPONSE:
We thank the reviewer for this comment and apology for some inconsistencies in the selection of the compounds to highlight in the figures which are mostly remnants of the drug prioritization history over the progression of the project. We tried to make it more consistent in the current version.
In the new version of figure 7D, AZL is substituting TFPZ, while TFPZ data were moved to supplementary figure S19.
Golgi morphology in VPS13B KO cells is reported to recover in NPC1 KD and SMPD1 KD cells, but it is not shown whether SM 36:1, CER 36:1, or other lipid levels also increase or change in these conditions. If Golgi morphology recovery occurs via the same mechanism as with compound treatment, a similar lipid pattern should be observed.
RESPONSE:
We thank the reviewer for this question that allowed us to expand our study including new interesting findings. We agree that this is an important point to strengthen the link between CAD and genetic perturbation effects. Given the availability of several published lipidomic datasets modelling LDS in HeLa and in other cell lines, we decided to perform a re-analysis of those to specifically focus on C18 sphingolipids. We found a relative increase of 36:1 upon depletion of LSD genes in all analyzed datasets for NPC1 and SMPD1, but also for more than 15 other LSD genes including NPC2, recapitulating what we find with all the CAD molecules tested in our study. These changes, were not noticed or at least not discussed by most of the authors. This is not surprising since those studies are focused on different biological questions. We believe that these findings, besides reinforcing our hypothesis of a common mechanism between CAD and NPC1/SMPD1 KO, have of general interest for the regulation of C18 sphingolipids, which are among the relative few lipid species with a bona fide specific protein binding partner and proposed to play a crucial role in Golgi traffic.
MINOR POINTS:
The manuscript lacks sufficient information about the compound library used for screening (number and source of compounds, compound type).
RESPONSE:
We apologize if this information was not sufficiently visible in the original version of the manuscript. The data about source, catalog number, formulation and several additional identifiers is included in the File S1. This is now clearly indicated in the methods so that I can be more easily visible to the readers
Fig. 3A: a WT control image is required.
RESPONSE:
A WT control image is now included in the new version of Figure 3.
Fig. 4: include representative images at concentrations higher than 1.25 µM.
RESPONSE:
Representative images are now included for all concentrations higher than 1.25 µM, as requested.
Abbreviations such as BMP/LBPA should be defined when first mentioned.
RESPONSE:
The abbreviation of BMP/LBPA was already defined when first mentioned in the original version of the manuscript
The abbreviation for raloxifene is inconsistent (RLX vs RAL) and should be unified.
RESPONSE:
Raloxifene is now abbreviated as RLX all over the manuscript.
Fig. 5C: the meaning of the green and magenta bars is not explained.
RESPONSE:
Color code for figure 5C has been included.
The definitions and centrifugation parameters for light and heavy membrane fractions should be clearly stated in the Methods.
RESPONSE:
The centrifugation parameters were already defined in the original manuscript. It is not clear to us, which parameter the Referee is referring to. Below is the sentence in the methods section:
“Gradients were centrifuged at 165,000 g for 1.5 h at 4°C with a SW40Ti Swinging-Bucket rotor (Beckman-Coulter). The LM and HM fractions were collected at the 35%-HB and 35%-40.6% interfaces, respectively”
The concentration and incubation times for BFA and nocodazole should be included in the main text or figure legends.
RESPONSE:
Concentrations and incubation times of BFA and nocodazole were already present in the legend of figure 5.
Fig. 8C, D, G, H: y-axes lack labels and must be defined.
RESPONSE:
Axes labels have now been included
There are multiple typographical errors, including "VPS12" instead of "VPS13B", that should be corrected.
RESPONSE:
We corrected this specific mistake as well as others that we could identify after careful reading of the manuscript.
Reviewer #2 (Significance (Required)):
While the dataset is extensive and technically detailed, the manuscript lacks a clear mechanistic explanation connecting lipid changes to Golgi restoration. The choice and comparison of compounds are inconsistent across experiments, and the interpretation remains speculative. Substantial revision and additional experiments are required before the study can be considered for publication.
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
This manuscript describes a compound screening aimed at identifying molecules that can restore Golgi organization in VPS13B knockout (KO) cells. The authors identify several compounds, most of which are lysosomotropic, and analyze their effects on Golgi morphology and lipid composition using multiple approaches. They report that VPS13B KO cells exhibit a reduction in C18-N-acyl sphingolipids, which can be restored by several of the identified compounds. Furthermore, two of these compounds, azelastine and raloxifene, promote neurite outgrowth in VPS13B KO cortical organoids. These findings are interesting and could potentially contribute to a better understanding of the pathophysiology of Cohen syndrome and the development of therapeutic strategies. However, despite the large number of analyses presented, the study remains largely descriptive, and there is no coherent mechanistic explanation for how these compounds restore Golgi structure in VPS13B KO cells. In addition to the reduction in C18-N-acyl sphingolipids, the KO cells display alterations in several other lipid species (LPC, LPE, PC40:1, PE42:1, TG, etc.), and treatment with the selected compounds induces further lipid accumulations, including cholesterol and BMP/LBPA. The relationship between these diverse lipid changes and the observed Golgi recovery lacks clarity and mechanistic consistency.
Major comments
The finding that compounds cannot prevent Golgi fragmentation caused by brefeldin A or nocodazole but can suppress statin-induced fragmentation is intriguing, but the underlying mechanism is not addressed. It is not evident whether this difference results from changes in membrane lipid composition or restoration of Rab/SNARE trafficking. The authors should examine Rab prenylation and SNARE localization by immunofluorescence or Western blotting to support their interpretation.
Although restoration of C18 sphingolipids (SM 36:1, CER 36:1) is observed upon compound treatment, its causal role in Golgi recovery or neurite outgrowth is not established. The authors should test whether blocking the increase of C18 SM/CER prevents the rescue of Golgi or neuronal phenotypes.
In Figure 7D, comparisons should include the LM and HM fractions isolated from WT cells.
The subcellular fractionation experiment should be repeated using AZL and RAL, the compounds used in organoid experiments, rather than TFPZ, to assess whether similar results are obtained. The compounds used differ across experiments, making it difficult to draw consistent conclusions.
Golgi morphology in VPS13B KO cells is reported to recover in NPC1 KD and SMPD1 KD cells, but it is not shown whether SM 36:1, CER 36:1, or other lipid levels also increase or change in these conditions. If Golgi morphology recovery occurs via the same mechanism as with compound treatment, a similar lipid pattern should be observed.
Minor points
The manuscript lacks sufficient information about the compound library used for screening (number and source of compounds, compound type).
Fig. 3A: a WT control image is required. Fig. 4: include representative images at concentrations higher than 1.25 µM. Abbreviations such as BMP/LBPA should be defined when first mentioned. The abbreviation for raloxifene is inconsistent (RLX vs RAL) and should be unified. Fig. 5C: the meaning of the green and magenta bars is not explained. The definitions and centrifugation parameters for light and heavy membrane fractions should be clearly stated in the Methods. The concentration and incubation times for BFA and nocodazole should be included in the main text or figure legends. Fig. 8C, D, G, H: y-axes lack labels and must be defined. There are multiple typographical errors, including "VPS12" instead of "VPS13B", that should be corrected.
While the dataset is extensive and technically detailed, the manuscript lacks a clear mechanistic explanation connecting lipid changes to Golgi restoration. The choice and comparison of compounds are inconsistent across experiments, and the interpretation remains speculative. Substantial revision and additional experiments are required before the study can be considered for publication.
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
VPS13B is a bridge-like lipid transfer protein, the loss or mutation of which is associated with Cohen syndrome (CS) involving Golgi fragmentation. In this study, the authors performed image-based chemical screens to identify compounds capable of rescuing the Golgi morphology in VPS13B-KO HeLa cells. They identified 50 compounds, the majority of which are lysosomotropic compounds or cationic amphiphilic drugs (CADs). Treatment of cells with several of these compounds causes lysosomal lipid storage, as assessed by BMP/LBPA staining, filipin staining, or LipidTOX staining. Interestingly, most LipidTOX puncta colocalized with transferrin receptor-positive compartments but not lysosomes. Similar to lysosomotropic compounds, knocking down NPC1 or SMPD1, mimicking lysosomal storage disease, also substantially rescued Golgi morphology. The authors show that VPS13B-KO cells have reduced C18 sphingolipids, which is reversed by treatment with CADs. Finally, the authors show that two CADs partially rescue neurite outgrowth in neuronal cultures. However, these drugs do not rescue the size of VPS13B KO organoids.
Overall, this is an impressive study identifying CADs as potential therapeutics for CS and suggesting sphingolipid upregulation as a general strategy for CS treatment. The morphological and lipidomics analyses unravel important molecular basis of CS pathology. This study will be of high interest to the field of lipid biology and organelle homeostasis. I have a few comments to help improve the quality of this study.
Minor:
This is an impressive study that identifies Cationic Amphiphilic Drugs (CADs) as potential therapeutics for Cohen syndrome (CS) and suggests sphingolipid upregulation as a general strategy for diseases driven by VPS13B loss-of-function. The unbiased approaches, notably the chemical screen and lipidomics, provide novel mechanistic insights into the underlying pathology of CS. This study will be of high interest to researchers in the fields of lipid biology and organelle homeostasis. It will also be highly valuable for clinical pediatricians managing CS patients.
Mijn Bureau, de Franse variant LaSuite en het Duitse openDesk.
Limited thinking, where you have national 'solutions', good they work on integration, but let's aim for European projects from the get-go too. Mijn Bureau, NL, OpenDesk in D, LaSuite in F.
We are aware that AWS, as the largest player, cannot be exactly matched, but we are confident that we do not have to compromise on security and availability with a European counterpart.
this is the sticking point, and true. You cannot expect a plug and play replacement. Analyse the components, make a new necklace of the different pieces.
Peter Vergote, legal advisor at DNS Belgium: "Our data and processes at AWS are located on European soil, spread across multiple data centres. This is perfectly in line with European legislation such as GDPR and NIS2 .
That is technically true, but not in practice. The 2018 Cloud Act makes all EU reg moot points from the US perspective. That makes any American involvement in your stack by def a breach of NIS2, GDPR and other digital and data related regs.
The migration away from AWS is still in its early stages. The market is currently being surveyed. The transition will begin in 2027 and is expected to be completed in the second half of 2027.
The decision is made, but transition still in early planning phases. To be completed 2027. Market survey ongoing, meaning there's no new contractor in sight as yet.
Since 2017, the system that processes the registrations of .be domain names has been located in AWS's European data centres.
DNS Belgium is on AWS since 2017, in EU data centers but that means nothing, definitely not since 2018
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
The manuscript submitted by Langenbacher et al., entitled " Rtf1-dependent transcriptional pausing regulates cardiogenesis", describes very interesting and highly impactful observations about the function of Rtf-1 in cardiac development. Over the last few years, the Chen lab has published novel insights into the genes involved in cardiac morphogenesis. Here, they used the mouse model, the zebrafish model, cellular assays, single cell transcription, chemical inhibition, and pathway analysis to provide a comprehensive view of Rtf1 in RNAPII (Pol2) transcription pausing during cardiac development. They also conducted knockdown-rescue experiments to dissect the functions of Rtf1 domains.
Strengths:
The most interesting discovery is the connection between Rtf1 and CDK9 in regulating Pol2 pausing as an essential step in normal heart development. The design and execution of these experiments also demonstrate a thorough approach to revealing a previously underappreciated role of Pol2 transcription pausing in cardiac development. This study also highlights the potential amelioration of related cardiac deficiencies using small molecule inhibitors against cyclin dependent kinases, many of which are already clinically approved, while many other specific inhibitors are at various preclinical stages of development for the treatment of other human diseases. Thus, this work is impactful and highly significant.
We thank the reviewer for appreciating our work.
Reviewer #2 (Public Review):
Summary:
Langenbacher at el. examine the requirement of Rtf1, a component of the PAF1C, which regulates transcriptional pausing in cardiac development. The authors first confirm their previous morphant study with newly generated rtf1 mutant alleles, which recapitulate the defects in cardiac progenitor and diUerentiation gene expression observed previously in morphants. They then examine the conservation of Rtf1 in mouse embryos and embryonic stem cell-derived cardiomyocytes. Conditional loss of Rtf1 in mesodermal lineages and depletion in murine ESCs demonstrates a failure to turn on cardiac progenitor and diUerentiation marker genes, supporting conservation of Rtf1 in promoting cardiac development. The authors subsequently employ bulk RNA-seq on flow-sorted hand2:GFP+ cells and multiomic single-cell RNA-seq on whole Rtf1-depleted embryos at the 10-12 stage. These experiments corroborate that genes associated with cardiac and muscle development are lost. Furthermore, the diUerentiation trajectories suggest that the expression of genes associated with cardiac maturation is not initiated. Structure-function analysis supports that the Plus3 domain is necessary for its function in promoting cardiac progenitor formation. ChIP-seq for RNA Pol II on 1012 somite stage embryos suggests that Rtf1 is required for proper promoter pausing. This defect can partially be rescued through use of a pharmacological inhibitor for Cdk9, which inhibits elongation, can partially restore elongation in rtf1 mutants.
Strengths:
Many aspects of the data are strong, which support the basic conclusions of the authors that Rtf1 is required for transcriptional pausing and has a conserved requirement in vertebrate cardiac development. Areas of strength include the genetic data supporting the conserved requirement for Rtf1 in promoting cardiac development, the complementary bulk and single-cell RNA-sequencing approaches providing some insight into the gene expression changes of the cardiac progenitors, the structure-function analysis supporting the requirement of the Plus3 domain, and the pharmacological epistasis combined with the RNA Pol II ChIP-seq, supporting the mechanism implicating Cdk9 in the Rtf1 dependent mechanism of RNA Pol II pausing.
We thank the reviewer for the summary and for recognizing many strengths of our work.
Weaknesses:
While most of the basic conclusions are supported by the data, there are a number of analyses that are confusing as to why they chose to perform the experiments the way they did and some places where the interpretations presently do not support the interpretations. One of the conclusions is that the phenotype aUects the maturation of the cardiomyocytes and they are arresting in an immature state. However, this seems to be mostly derived from picking a few candidates from the single cell data in Fig. 6. If that were the case, wouldn't the expectation be to observe relatively normal expression of earlier marker genes required for specification, such as Nkx2.5 and Gata5/6? The in situ expression analysis from fish and mice (Fig. 2 and Fig. 3) and bulk RNA-seq (Fig. 5) seems to suggest that there are pretty early specification and diUerentiation defects. While some genes associated with cardiac development are not changed, many of these are not specific to cardiomyocyte progenitors and expressed broadly throughout the ALPM. Similarly, it is not clear why a consistent set of cardiac progenitor genes (for instance mef2ca, nkx2.5, and tbx20) was analyzed for all the experiments, in particular with the single cell analysis.
A major conclusion of our study is that Rtf1 deficiency impairs myocardial lineage differentiation from mesoderm, as suggested by the reviewer. Thus, the main goal of this study is to understand how Rtf1 drives cardiac differentiation from the LPM, rather than the maturation of cardiomyocytes. Multiple lines of evidence support this conclusion:
(a) In situ hybridization showed that Rtf1 mutant embryos do not have nkx2.5+ cardiac progenitor cells and subsequently fail to produce cardiomyocytes (Figs. 2, 3).
(b) RT-PCR analysis showed that knockdown of Rtf1 in mouse embryonic stem cells causes a dramatic reduction of cardiac gene expression and production of significantly fewer beating patches (Fig.4).
(c) Bulk RNA sequencing revealed significant downregulation of cardiac lineage genes, including nkx2.5 (Fig. 5).
(d) Single cell RNA sequencing clearly showed that lateral plate mesoderm (LPM) cells are significantly more abundant in Rtf1 morphant,s whereas cardiac progenitors are less abundant (Fig. 6 and Fig.6 Supplement 1-5).
When feasible, we used cardiac lineage restricted markers in our assays. Nkx2.5 and tbx5a are not highlighted in the single cell analysis because their expression in our sc-seq dataset was too low to examine in the clustering/trajectory analysis. In this revised manuscript, we provide violin plots showing the low expression levels of these genes in single cells from Rtf1 deficient embryos (Figure 6 Supplement 5).
The point of the multiomic analysis is confusing. RNA- and ATAC-seq were apparently done at the same time. Yet, the focus of the analysis that is presented is on a small part of the RNA-seq data. This data set could have been more thoroughly analyzed, particularly in light of how chromatin changes may be associated with the transcriptional pausing. This seems to be a lost opportunity. Additionally, how the single cell data is covered in Supplemental Fig. 2 and 3 is confusing. There is no indication of what the diUerent clusters are in the Figure or the legend.
In this study, we performed single cell multiome analysis and used both scRNAseq and scATACseq datasets to generate reliable clustering. The scRNAseq analysis reveals how Rtf1 deficiency impacts cardiac differentiation from mesoderm, which inspired us to investigate the underlying mechanism and led to the discovery of defects in Rtf1-dependent transcriptional pause release.
We agree with the reviewer that deep examination of Rtf1-dependent chromatin changes would provide additional insights into how Rtf1 influences early development and careful examination of the scATACseq dataset is certainly a good future direction.
In this revised manuscript, we have revised Fig.6 Supplement 1 to include the predicted cell types and provide an additional excel file showing the annotation of all 39 clusters (Supplementary Table 2).
While the effect of Rtf1 loss on cardiomyocyte markers is certainly dramatic, it is not clear how well the mutant fish have been analyzed and how specific the eUect is to this population. It is interpreted that the eUects on cardiomyocytes are not due to "transfating" of other cell fates, yet supplemental Fig. 4 shows numerous eUects on potentially adjacent cell populations. Minimally, additional data needs to be provided showing the live fish at these stages and marker analysis to support these statements. In some images, it is not clear the embryos are the same stage (one can see pigmentation in the eyes of controls that is not in the mutants/morphants), causing some concern about developmental delay in the mutants.
Single cell RNA sequencing showed an increased abundance of LPM cells and a reduced abundance of cardiac progenitors in Rtf1 morphants (Fig. 6 and Fig.6 Supplement 1-5). The reclustering of anterior lateral plate mesoderm (ALPM) cells and their derivatives further showed that cells representing undifferentiated ALPM were increased whereas cells representing all three ALPM derivatives were reduced. These findings indicate a defect in ALPM differentiation.
The reviewer questioned whether we examined stage-matched embryos. In our assay, Rtf1 mutant embryos were collected from crosses of Rtf1 heterozygotes. Each clutch from these crosses consists of ¼ embryos showing rtf1 mutant phenotypes and ¾ embryos showing wild type phenotypes which were used as control. Mutants and their wild type siblings were fixed or analyzed at the same time.
The reviewer questioned the specificity of the Rtf1 deficient cardiac phenotype and pointed out that Rtf1 mutant embryos do not have pigment cells around the eye. Rtf1 is a ubiquitously expressed transcriptional regulator. Previous studies in zebrafish have shown that Rtf1 deficiency significantly impacts embryonic development. Rtf1 deficiency causes severe defects in cardiac lineage and neural crest cell development; consequently, Rtf1 deficient embryos do not have cardiomyocytes and pigmentation (Langenbacher et al., 2011, Akanuma et al., 2007, and Jurynec et al., 2019). We now provide an image showing a 2-day-old Rtf1 mutant embryo and their wild type sibling to illustrate the cardiac, neural crest, and somitogenesis defects caused by loss of Rtf1 activity (Fig. 2 Supplement 1).
With respect to the transcriptional pausing defects in the Rtf1 deficient embryos, it is not clear from the data how this eUect relates to the expression of the cardiac markers. This could have been directly analyzed with some additional sequencing, such as PRO-seq, which would provide a direct analysis of transcriptional elongation.
We showed that Rtf1 deficiency results in a nearly genome-wide decrease in promoterproximal pausing and downregulation of cardiac makers. Attenuating transcriptional pause release could restore cardiomyocyte formation in Rtf1 deficient embryos. In this revised manuscript, we provide additional RNAseq data showing that the expression levels of critical cardiac development genes such as nkx2.5, tbx5a, tbx20, mef2ca, mef2cb, ttn.2, and ryr2b are significantly rescued. We agree with the reviewer that further analyses using the PRO-seq approach could provide additional insights, but it is beyond the scope of this manuscript.
Some additional minor issues include the rationale that sequence conservation suggests an important requirement of a gene (line 137), which there are many examples this isn't the case, referencing figures panels out of order in Figs. 4, 7, and 8) as described in the text, and using the morphants for some experiments, such as the rescue, that could have been done in a blinded manner with the mutants.
We have clarified the rationale in this revised manuscript and made the eRort to reference figures in order.
The reviewer commented that rescue experiments “could have been done in a blinded manner with the mutants”. This was indeed how the flavopiridol rescue and cdk9 knockdown experiments were carried out. Embryos from crosses of Rtf1 heterozygotes were collected, fixed after treatment and subjected to in situ hybridization. Embryos were then scored for cardiac phenotype and genotyped (Fig.8 d-g). Morpholino knockdown was used in genomic experiments because our characterization of rtf1 morphants showed that they faithfully recapitulate the rtf1 mutant phenotype during the timeframe of interest (Fig. 2).
Recommendations for the authors:
Reviewer #1 (Recommendations For The Authors):
This reviewer has a few suggestions below, aimed at improving the clarity and impact of the current study. Once these items are addressed, the manuscript should be of interest to the Elife reader.
Item 1. Strengthening the interaction between Rfh1 and CDK9 on Pol2 pausing.
The authors have convincingly shown that the chemical inhibition of CDK9 by flavopiridol can partially rescue the expression of cardiac genes in the zebrafish model. Although flavopiridol is FDA approved and has been a classical inhibitor for the dissection of CDK9 function, it also inhibits related CDKs (such as Flavopiridol (Alvocidib) competes with ATP to inhibit CDKs including CDK1, CDK2, CDK4, CDK6, and CDK9 with IC50 values in the 20-100 nM range) Therefore, this study could be more impactful if the authors can provide evidence on which of these CDKs may be most relevant during Rtf1-dependent cardiogenesis. To determine whether the observed cardiac defect indicates a preferential role for CDK9, or that other CDKs may also be able to provide partial rescue may be clarified using additional, more selective small molecules (e.g., BAY1251152, LDC000067 are commercially available).
The reviewer raised a reasonable concern about the specificity of flavopiridol. We thank the reviewer for the insightful suggestion and share the concern about specificity. To address this question, we have used an orthogonal testing through morpholino inhibition where we directly targeted CDK9 and observed the same level of rescue, supporting a critical role of transcription pausing in cardiogenesis.
Item 2. Differences between CRISPR lines and morphants
Much of the work presented used Rtf1 morphants while the authors have already generated 2 CRISPR lines. What is the diUerence between morphants and mutants? The authors should comment on the similarities and/or differences between using morphants or mutants in their study and whether the same Rtf1- CDK9 connection also occurs in the CRISPR lines.
The morphology of our mutants (rtf1<sup>LA2678</sup> and rtf1<sup>LA2679</sup>) resembles the morphants and the previously reported ENU-induced rtf1<sup>KT641</sup> allele. Extensive in situ hybridization analysis showed that the morphants faithfully recapitulate the mutant phenotypes (Fig.2). We have performed rescue experiments (flavopiridol and CDK9 morpholino) using Rtf1 mutant embryos and found that inhibiting Cdk9 restores cardiomyocyte formation (Fig.8).
Item 3. Discuss the therapeutic relevance of study
The authors have already generated a mouse model of Rtf1 Mesp1-Cre knockout where cardiac muscle development is severely derailed (Fig 3B). Thus, a demonstration of a conserved role for CDK9 inhibitor in rescuing cardiogenesis using mouse cells or the mouse model will provide important information on a conserved pathway function relevant to mammalian heart development. In the Discussion, how this underlying mechanistic role may be useful in the treatment of congenital heart disease should be provided.
Thank you for the insight. We have incorporated your comments in the discussion.
Item 4. Insights into the role of CDK9-Rtf1 in response to stress versus in cardiogenesis.
In the Discussion, the authors commented on the role of additional stress-related stimuli such as heat shock and inflammation that have been linked to CDK9 activity. However, the current ms provides the first, endogenous role of Pol2 pausing in a critical developmental step during normal cardiogenesis. The authors should emphasize the novelty and significance of their work by providing a paragraph on the state of knowledge on the molecular mechanisms governing cardiogenesis, then placing their discovery within this framework. This minor addition will also clarify the significance of this work to the broad readership of eLife.
Thank you for the suggestion. We have incorporated your comments and elaborate on the novelty and significance of our work in the discussion.
Reviewer #2 (Recommendations For The Authors):
(1) It is diUicult to assess what the overt defects are in the embryos at any stages. Images of live images were not included in the supplement. Do these have a small, malformed heart tube later or are the embryos just deteriorating due to broad defects?
The Rtf1 deficient embryos do not produce nkx2.5+ cardiac progenitors. Consequently, we never observed a heart tube or detected cells expressing cardiomyocyte marker genes such as myl7. This finding is consistent with previous reports using rtf1 morphants and rtf<sup>1KT64</sup>, an ENU-induced point mutation allele (Langenbacher et al., 2011 and Akanuma, 2007). In this revised manuscript, we provide a live image of 2-day-old wild type and rtf1<sup>LA2679/LA2679</sup> embryos (Fig. 2 Supplement 1). After two days, rtf1 mutant embryos undergo broad cell death.
(2) Fig. 2, although the in situs are convincing, there is not a quantitative assessment of expression changes for these genes. This could have been done for the bulk or single cell RNA-seq experiments, but was not and these genes weren't not included in the heat maps. A quantitative assessment of these genes would benefit the study.
The top 40 most significantly differentially expressed genes are displayed in the heatmap presented in Fig.5d. The complete differential gene expression analysis results for our hand2 FACS-based comparison of rtf1 morphants and controls is presented in Supplementary Data File 1. In this revised manuscript, we provide a new supplemental figure with violin plots showing the expression levels of genes of interest in our single cell sequencing dataset (Fig.6 Supplement 5).
(3) It doesn't not appear that any statistical tests were used for the comparisons in Fig. 2.
We now provide the statistical data in the legend and Fig.2 b, d, f, h and i.
(4) It's not clear the magnifications and orientations of the embryos in Fig. 3b are the same.
Embryos shown in Fig.3b are at the same magnification. However, because Rtf1 mutant embryos display severe morphological defects, the orientation of mutant embryos was adjusted to examine the cardiac tissue.
(5) The n's for analysis of MLC2v in WT Rtf1 CKO embryos in Fig. 3b are only 1. At least a few more embryos should be analyzed to confirm that the phenotype is consistent.
We have revised the figure and present the number of embryos analyzed and statistics in Fig.3c.
(6) A number of figure panels are referred to out of order in the text. Fig. 4E-G are before Fig. 4C, D, Fig. 7C before 7B, Fig. 8D-I before 8A ,B. In general, it is easier for the reader if the figures panels are presented in the order they are referred to in the text.
Revised as suggested.
(7) While additional genes can be included, it is not clear why the same sets of genes are not examined in the bulk or single-cell RNA-seq as with the in situs or expression was analyzed in embryos. I suggest including the genes like nkx2.5, tbx20, myl7, in all the sequencing analysis.
We used the same set of genes in all analyses when possible. However, the low expression of genes such as nkx2.5 and myl7 in our sc-seq dataset preclude them from the clustering/trajectory analysis. In this revised manuscript, we present violin plots showing their expression in wild type and rtf1 morphants (Fig. 6 Supplement 5).
(8) If a multiomic approach was used, why wasn't its analysis incorporated more into the manuscript? In general, a clearer presentation and deeper analysis of the single cell data would benefit the study. The integration of the RNA and ATAC would benefit the analysis.
As addressed in our response to the reviewer’s public review, both datasets were used in clustering. Examining changes in chromatin accessibility is certainly interesting, but beyond the scope of this study.
(9) Many of the markers analyzed are not cardiac specific or it is not clear they are expressed in cardiac progenitors at the stage of the analysis. Hand2 has broader expression. Additional confirmation of some of the genes through in situ would help the interpretations.
Markers used for the in situ hybridization analysis (myl7, mef2ca, nkx2.5, tbx5a, and tbx20) are known for their critical role in heart development. For sc-seq trajectory analyses, most displayed genes (sema3e, bmp6, ttn.2, mef2cb, tnnt2a, ryr2b, and myh7bb) were identified based on their differential expression along the LPM-cardiac progenitor pseudotime trajectory. Rather than selecting genes based on their cardiac specificity, our goal was to examine the progressive gene expression changes associated with cardiac progenitor formation and compare gene expression of wild type and rtf1 deficient embryos.
(10) Additional labels of the cell clusters are needed for Supplemental Figs. 2 and 3.
The cluster IDs were presented on Supplementary Figures 2 and 3. In this revised version, we added predicted cell types to the UMAP (revised Fig.6 Supplement 1) and provided an excel file with this information (revised Supplementary Table 2).
(11) On lines 101-102, the interpretation from the previous data is that diUerentiation of the LPM requires Rtf1. However, later from the single cell data the interpretation based on the markers is that Rtf1 loss aUects maturation. However, it is not clear this interpretation is correct or what changed from the single cell data. If that were the case, one would expect to see maintenance of more early marks and subsequent loss of maturation markers, which does not appear to the be the case from the presented data.
Our data suggests that cardiac progenitor formation is not accomplished by simultaneously switching on all cardiac marker genes. Our pseudotime trajectory analysis highlights tnnt2a, ryr2b, and myh7bb as genes that increase in expression in a lagged manner compared to mef2cb (Fig. 6). Thus, the abnormal activation of mef2cb without subsequent upregulation of tnnt2a, ryr2b, and myh7bb in rtf1 morphants suggests a requirement for rtf1 in the progressive gene expression changes required for proper cardiac progenitor differentiation. Our single cell experiment focuses on the process of cardiac progenitor differentiation and does not provide insights into cardiomyocyte maturation. We have edited the text to clarify these interpretations.
(12) The interpretation that there is not "transfating" is not supported by the shown data. Analysis of markers in other tissues, again with in situ, to show spatially would benefit the study.
As stated in our response to the reviewer’s public review, we observed a dramatic increase of ALPM cells, but a decrease of ALPM derivatives including the cardiac lineage. We did not observe the expansion of one ALPM-derived subpopulation at the expense of the others. These observations suggest a defect in ALPM differentiation and argue against the notion that the region of the ALPM that would normally give rise to cardiac progenitors is instead differentiating into another cell type.
(13) The rationale that sequence conservation means a gene is important (lines 137-139) is not really true. There are examples a lot of highly conserved genes whose mutants don't have defects.
We have revised the text to avoid confusion.
(14) The data showing that the 8 bp mutations do not aUect the RNA transcript is not shown or at least indicated in Fig. 7. It would seem that this experiment could have been done in the mutant embryos, in which case the experiment would have been semi-blinded as the genotyping would occur after imaging.
The modified Rtf1 wt RNA (Rtf1 wt* in revised Fig. 7) robustly rescued nkx2.5 expression in rtf1 deficient embryos, demonstrating that the 8 bp modifications do not negatively impact the activity of the injected RNA. As stated previously, morpholino knockdown was used in some experiments because our characterization of rtf1 morphants showed that they faithfully recapitulate the rtf1 mutant phenotype during the timeframe of interest.
(15) Using a technique like PRO-seq at the same stage as the ChIP-seq would complement the ChIP-seq and allow a more detailed analysis of the transcriptional pausing on specific genes observed in WT and mutant embryos.
As stated in our response to the reviewer’s public review, we appreciate the suggestion but PRO-seq is beyond the scope of this study.
Reviewer #1 (Public review):
Summary:
Here, the authors are proposing a role for miR-196, a microRNA that has been shown to bind and enhance degradation of mRNA targets in the regulation of cell processes, has a novel role in allowing the emergence of CD19+ cells in cells in which Ebf1, a critical B-cell transcription factor, has been genetically removed.
Strengths:
That over-expression of mR-195 can allow the emergence of CD19+ cells missing Ebf1 is somewhat novel.
Their data does perhaps support to a degree the emergence of a transcriptional network that may bypass the absence of Ebf1, including the FOXO1 transcription factor, but this data is not strong or definitive.
Weaknesses:
It is unclear whether this observation is in fact physiological. When the authors analyse a knockout model of miR-195, there is not much of a change in the B-cell phenotype. Their findings may therefore be an artefact of an overexpression system.
The authors have provided insufficient data to allow a thorough appraisal of the step-wise molecular changes that could account for their observed phenotype.
On review of the resubmitted manuscript, while I note the authors have attempted to address several of my comments, unfortunately, their resubmission is not sufficient to address several of the comments I had previously made.
In particular, in the resubmitted data that includes western blots for PAX5 and ERG in their EBF1-/- model, Supp Fig S3, the bands they show infer that that PAX5 and ERG expression can still be significantly detected in their EBF1-/- early B-cell model. This should not be the case, as no expression of PAX5 or ERG should be seen, as has been shown in prior literature.