But model quality is table stakes at this point
meaning it is a no-brainer to start with a good model
But model quality is table stakes at this point
meaning it is a no-brainer to start with a good model
Not all Instructors will expect formal emails, but it’s important to remember that your instructor is not your friend
I think that even if some Instructors don't expect formal emails, respect and professionalism should always be maintained.
but first let me tell ye, if ye should lead her into 1320a fool's paradise, as they say, it were a very gross kind of behavior, as they say: for the gentlewoman is young; and, therefore, if you should deal double with her, truly it were an ill thing to be offered to any gentlewoman, and very weak dealing.
Nurse protects Juliet, warning Romeo not to trick her into a "fool's paradise."
that Petrarch flowed in: Laura to his lady was but a 1200kitchen-wench; marry, she had a better love to be-rhyme her; Dido a dowdy; Cleopatra a gipsy; Helen and Hero hildings and harlots; Thisbe a grey eye or so, but not to the purpose. Signior
Allusion: Says Romeo's love outdoes Petrarch's famous love for Laura, and other famous beauties.
young men's love then lies Not truly in their hearts, but in their eyes.
Critique: Friar says Romeo loves based on looks, not deep feeling.
And yet no further than a wanton's bird;
Simile: She compares him to a spoiled child's pet bird on a string. Wants him close but free.
But old folks, many feign as they were dead; 1390Unwieldy, slow, heavy and pale as lead.
Simile/Character: Juliet impatiently stereotypes the old as slow and dull, unlike her youthful passion.
Nurse. Well, you have made a simple choice; you know not 1415how to choose a man: Romeo! no, not he; though his face be better than any man's, yet his leg excels all men's; and for a hand, and a foot, and a body, though they be not to be talked on, yet they are past compare: he is not the flower of courtesy, 1420but, I'll warrant him, as gentle as a lamb. Go thy ways, wench; serve God. What, have you dined at home?
Comic Delay: The Nurse praises Romeo's body parts (face, leg) but won't give the news, frustrating Juliet.
Nurse. Then hie you hence to Friar Laurence' cell; There stays a husband to make you a wife: Now comes the wanton blood up in your cheeks, They'll be in scarlet straight at any news. Hie you to church; I must another way, 1450To fetch a ladder, by the which your love Must climb a bird's nest soon when it is dark: I am the drudge and toil in your delight, But you shall bear the burden soon at night.
Plot/Imagery: She'll get a ladder for Romeo to climb to Juliet's "bird's nest" (room) after the wedding.
Call me but love, and I'll be new baptized;
Religious Metaphor: He'll renounce his name and be reborn through her love.
Her vestal livery is but sick and green
Symbolism: The pale green of virginity is sickly. He urges her to "cast it off."
Mercutio. Nay, I'll conjure too. 805Romeo! humours! madman! passion! lover! Appear thou in the likeness of a sigh: Speak but one rhyme, and I am satisfied; Cry but 'Ay me!' pronounce but 'love' and 'dove;' Speak to my gossip Venus one fair word, 810One nick-name for her purblind son and heir, Young Adam Cupid, he that shot so trim, When King Cophetua loved the beggar-maid! He heareth not, he stirreth not, he moveth not; The ape is dead, and I must conjure him. 815I conjure thee by Rosaline's bright eyes, By her high forehead and her scarlet lip, By her fine foot, straight leg and quivering thigh And the demesnes that there adjacent lie, That in thy likeness thou appear to us!
Mockery: Mercutio humorously calls for Romeo using love clichés and Rosaline's body parts. Shows he doesn't understand true love.
L'Implication Affective des Enseignants : Synthèse des Recherches de Maël Virat
Ce document de synthèse analyse les travaux de Maël Virat sur l'implication affective des enseignants et son impact sur les élèves.
La thèse centrale est que la relation affective enseignant-élève, loin d'être un simple supplément à la pédagogie, est un moteur fondamental de l'apprentissage et du développement de l'élève.
Cette dynamique s'ancre dans la théorie de l'attachement, où la sécurité affective fournie par l'enseignant libère les capacités d'exploration de l'élève.
Les points clés sont les suivants :
1. Sécurité et Exploration : La relation enseignant-élève est gouvernée par la même dynamique "sécurité-exploration" que celle observée entre un parent et son enfant.
Un enseignant perçu comme une "base de sécurité" permet à l'élève, notamment celui de style d'attachement anxieux, de persévérer face aux difficultés scolaires.
2. L'Engagement comme Médiateur : Des méta-analyses à grande échelle confirment le lien entre la qualité de la relation affective et la réussite scolaire.
Cet effet est principalement médiatisé par l'engagement de l'élève : une relation sécurisante favorise la motivation et l'implication, qui à leur tour améliorent les résultats.
3. L'Amour Compassionnel : Pour caractériser l'implication affective de l'enseignant, Maël Virat propose le concept d'« amour compassionnel ».
Il s'agit d'un sentiment altruiste, centré sur le bien-être de l'autre, qui se distingue de l'amour romantique ou amical.
Cet amour se manifeste par l'attention, le soutien comportemental et une sensibilité émotionnelle aux réussites et aux difficultés de l'élève.
4. Les Facteurs d'Influence : L'implication de l'enseignant n'est pas un trait de personnalité immuable mais dépend fortement du contexte. Les facteurs déterminants incluent :
◦ Le soutien institutionnel : Le soutien perçu de la part des collègues et de la hiérarchie est directement corrélé à la capacité de l'enseignant à s'investir affectivement auprès de ses élèves.
◦ Les croyances professionnelles : L'intention d'un enseignant de fournir un soutien émotionnel est principalement prédite par son attitude (les bénéfices qu'il en retire personnellement en termes de plaisir au travail et de relations), son sentiment de contrôle (se sentir formé, avoir le temps, considérer que cela fait partie de son rôle) et, dans une moindre mesure, par les normes sociales perçues.
◦ Le contexte systémique : La taille de l'établissement, la culture professionnelle, et la formation initiale jouent un rôle crucial dans la facilitation ou l'inhibition de ces relations.
En conclusion, améliorer l'engagement et la réussite des élèves passe par la reconnaissance et la valorisation du rôle affectif des enseignants.
Cela nécessite des interventions qui ne se limitent pas à l'individu, mais qui agissent sur le système : la formation, la culture d'établissement et le soutien offert aux professionnels de l'éducation.
--------------------------------------------------------------------------------
Maël Virat, chercheur en psychologie, concentre une partie significative de ses travaux sur la relation enseignant-élève, bien que ses recherches s'étendent également aux besoins sociaux des adolescents et au vécu des professionnels du travail social, notamment dans la protection de l'enfance.
Ses travaux mobilisent la théorie de l'attachement comme cadre théorique principal pour analyser les dynamiques relationnelles en milieu scolaire.
Il est membre d'un groupe de recherche francophone (FREE) qui s'intéresse à la manière de prendre en compte la dimension relationnelle dans la formation, initiale et continue, des enseignants.
La théorie de l'attachement, développée par John Bowlby, établit un lien fondamental entre la sécurité affective et le comportement d'exploration.
• Les Expériences de Harlow : Les travaux de Harry Harlow avec des bébés singes ont démontré que le besoin de sécurité affective est primordial.
Privés de leur mère mais en présence de substituts maternels (l'un en fil de fer nourrissant, l'autre en tissu doux), les singes privilégiaient le contact réconfortant.
Ce manque de sécurité affective réduisait significativement leurs comportements exploratoires dans un nouvel environnement.
• Une Théorie pour toute la Vie : Cette dynamique n'est pas limitée à la petite enfance.
Une étude sur des couples mariés a montré que lorsqu'un homme était confronté à une tâche impossible (résoudre des puzzles insolubles), la présence de sa partenaire agissant comme une base de sécurité (encouragements, attention, absence d'interférence) augmentait sa persistance dans la tâche.
La figure d'attachement principale à l'âge adulte est souvent le partenaire amoureux, suivi par la mère.
Plusieurs études expérimentales transposent cette dynamique à la relation enseignant-élève, démontrant que l'enseignant peut fonctionner comme une "base de sécurité" qui favorise l'apprentissage.
Étude 1 : Soutien Émotionnel et Comportements Exploratoires
Une étude basée sur l'observation de duos enseignant-élève a établi une chaîne causale claire :
1. Soutien de l'enseignant : Plus l'enseignant manifeste de comportements de soutien émotionnel (temps d'attention, regards, encouragements).
2. Sécurité de l'élève : Plus l'élève montre des signes de sécurité affective (détente, absence de stress, concentration).
3. Exploration : Et plus il met en œuvre des comportements exploratoires (persistance face à la difficulté, concentration accrue).
Étude 2 : L'Amorçage Subliminal par la Photo de l'Enseignant
Des chercheurs allemands et autrichiens ont mené une expérience où des élèves devaient résoudre des tests psychotechniques.
• Protocole : Avant chaque test, la photo de leur enseignant était projetée de manière subliminale (20 à 40 millisecondes), un temps trop court pour une perception consciente.
Pour le groupe contrôle, une image brouillée ayant les mêmes propriétés lumineuses était utilisée.
• Condition : Au préalable, les enseignants avaient évalué la qualité de leur relation avec chaque élève via une échelle mesurant la proximité et la chaleur, un outil fortement corrélé aux mesures d'attachement.
• Résultats : La présentation subliminale de la photo de l'enseignant améliorait les performances des élèves uniquement lorsque l'enseignant avait décrit sa relation avec cet élève comme étant chaleureuse, affective et sécurisante.
Étude 3 : La Persistance des Adolescents face à l'Échec
Une étude menée en Israël par Mario Mikuliner, spécialiste de l'attachement, a examiné la persistance scolaire chez des adolescents.
| Variable mesurée | Méthode | | --- | --- | | Style d'attachement de l'élève | Questionnaire évaluant le niveau de sécurité ou d'anxiété dans les relations. | | Perception de l'enseignant comme "base de sécurité" | Questionnaire demandant aux élèves s'ils perçoivent leur professeur principal comme disponible, accueillant et non rejetant. | | Condition expérimentale (3 semaines plus tard) | Groupe expérimental : Exercice de visualisation demandant à l'élève de penser intensément à son professeur principal. <br> Groupe contrôle : Exercice de visualisation demandant de penser à un voisin neutre. | | Mesure de la persistance | Tâche d'association de mots contenant 4 items impossibles à résoudre. La persistance est mesurée par le temps passé sur ces items impossibles avant d'abandonner, comparativement au temps de réponse moyen de l'élève. |
Résultats principaux :
• Dans le groupe contrôle (pensée neutre), les élèves au style d'attachement anxieux montrent une persistance significativement plus faible que les autres.
• Dans le groupe expérimental, le fait de penser à un enseignant perçu comme une base de sécurité compense totalement le déficit de persistance des élèves anxieux. Leur performance devient indiscernable de celle des élèves sécures.
Conclusion de cette partie : Ces travaux démontrent expérimentalement que la perception d'un enseignant comme une figure sécurisante a un effet direct et mesurable sur les capacités cognitives et la persévérance des élèves, en particulier pour ceux qui sont les plus vulnérables sur le plan affectif.
Une méta-analyse majeure réalisée par Débora Roorda, portant sur 189 études et un total de près de 250 000 élèves du primaire et du secondaire, confirme l'importance de la relation affective.
• Lien avec la réussite et l'engagement : Il existe un lien statistique modéré mais robuste et constant entre la qualité de la relation affective enseignant-élève et à la fois l'engagement scolaire et la réussite scolaire.
• Le rôle médiateur de l'engagement : Le principal mécanisme par lequel la relation affective influence la réussite est l'engagement. Une relation positive renforce la motivation et l'implication de l'élève dans les tâches scolaires.
• Ordre de grandeur de l'effet : La relation positive avec les enseignants peut expliquer environ 10% de la variance de l'engagement des élèves.
Dans le domaine de la psychologie, où il est rare d'expliquer plus de 50% d'un phénomène complexe, ce chiffre est considéré comme important.
Une enquête menée par Maël Virat auprès de collégiens via le questionnaire "Who To ?" (Vers qui te tournes-tu en cas de problème ?) apporte des nuances importantes.
• Diversité des figures d'attachement : Si les enseignants sont fréquemment cités comme personnes ressources, les assistants d'éducation (AED) apparaissent également comme des figures sécurisantes majeures.
• Un constat préoccupant : Dans un premier échantillon, 50% des élèves n'ont nommé aucune personne au sein de leur établissement vers qui se tourner.
• Corrélation : Le nombre de personnes sécurisantes citées par un élève est positivement corrélé à sa motivation, son engagement scolaire et son sentiment d'appartenance à l'école.
Face à l'abondance de littérature sur les effets de la relation, Maël Virat a orienté ses recherches sur une question moins explorée : qu'est-ce que l'implication affective du côté de l'enseignant ?
Son postulat est qu'un élève ne peut se sentir en sécurité affective avec une personne qui n'est pas elle-même impliquée affectivement.
Après avoir écarté des concepts jugés inadéquats :
• La bienveillance : Trop général, pouvant s'appliquer à un voisin dans un train et pas nécessairement doté d'une dimension affective spécifique à la relation pédagogique.
• L'empathie : Décrit davantage une compétence cognitive et émotionnelle mobilisable dans divers contextes (y compris la vente) qu'un engagement relationnel durable.
Il s'est arrêté sur le concept d'amour compassionnel.
Définition de l'Amour Compassionnel : C'est une forme d'amour altruiste, centrée sur le bien et le développement de l'autre.
Dans la théorie de l'attachement, c'est le sentiment éprouvé par la figure de soin (le caregiver) en réponse à l'attachement de l'enfant. Il se construit dans la durée et ne disparaît pas avec la fin de la relation.
Cet amour se compose de trois dimensions :
1. Cognitive : Une attention soutenue à l'autre, des efforts pour comprendre sa perspective.
2. Comportementale : Des actes concrets d'aide, de soutien et de dévouement.
3. Affective : Une sensibilité à l'état de l'autre, se traduisant par :
◦ Des émotions positives (plaisir au contact de l'élève, joie face à ses réussites).
◦ Des émotions négatives (tristesse, peine, lorsque l'élève est en difficulté).
◦ Note : Des études par questionnaire montrent que les enseignants reconnaissent plus facilement les émotions positives que les négatives, possiblement en raison de normes professionnelles.
Une hypothèse centrale est que de nombreuses actions perçues comme purement pédagogiques par l'enseignant sont interprétées par l'élève comme des signes d'implication affective.
Une étude sur des élèves de 4ème en mathématiques a testé cette hypothèse :
• Variable indépendante : La perception par les élèves du "climat de classe" (structure de but), soit centré sur la maîtrise (chacun progresse à son rythme), soit sur la performance (comparaison et classement entre élèves).
• Variable médiatrice : La perception par l'élève de l'amour compassionnel de son enseignant de mathématiques à son égard.
• Variable dépendante : L'engagement affectif de l'élève pour les mathématiques ("j'aime les maths").
Résultat : Un climat de classe centré sur la maîtrise est positivement lié à l'engagement de l'élève parce qu'il est interprété par ce dernier comme un signe que l'enseignant se soucie de lui et l'aime (amour compassionnel).
L'efficacité du choix pédagogique passe par sa signification affective.
L'amour compassionnel n'est pas une émotion arbitraire ("l'amour ne se commande pas"). Il peut être cultivé et dépend fortement de facteurs contextuels et personnels.
| Catégorie de Facteurs | Exemples | | --- | --- | | Facteurs Externes | Taille de l'école et de la classe (plus c'est petit, meilleures sont les relations), type de management du chef d'établissement, culture d'établissement valorisant les relations. | | Facteurs liés à l'Élève | Compétences sociales et scolaires, sexe (très léger effet en faveur des filles). Le facteur le plus puissant est la présence de problèmes de comportement. | | Facteurs liés à l'Enseignant | Quantité et qualité de la formation, état de stress, compétences émotionnelles et sociales, style d'attachement (les enseignants "sécures" ont des relations légèrement meilleures), sentiment d'efficacité, croyances sur leur rôle. |
Une étude montre que plus les enseignants déclarent recevoir de soutien de la part de leurs collègues, plus ils rapportent ressentir de l'amour compassionnel pour leurs élèves.
Cela s'explique par le fait que le système de caregiving (prendre soin) de l'enseignant est d'autant plus actif que son propre système d'attachement est sécurisé par son environnement professionnel.
Une étude récente basée sur la théorie du comportement planifié a cherché à identifier les croyances spécifiques qui prédisent l'intention d'un enseignant de s'impliquer dans le soutien émotionnel.
Le modèle testé explique 68% de la variance de cette intention, un score très élevé.
Voici les croyances les plus déterminantes, qui constituent des cibles d'action pour la formation :
1. L'Attitude (ce que l'enseignant pense du soutien émotionnel) L'intention est plus forte quand l'enseignant croit que le soutien émotionnel est bénéfique... pour lui-même.
• Il améliore ses relations avec les élèves.
• Il augmente son plaisir au travail.
• Il renforce son sentiment d'utilité. (Argumenter sur les seuls bénéfices pour l'élève serait donc moins efficace pour motiver les enseignants).
2. Le Contrôle Comportemental Perçu (se sentir capable) L'intention est plus forte quand l'enseignant :
• Pense que le soutien émotionnel fait partie intégrante de son travail (et n'est pas "en plus").
• Pense qu'il a suffisamment de temps pour cela.
• Se sent formé à cette dimension du métier.
3. Les Normes Sociales (ce qui est attendu, ce que font les autres)
Cet aspect a un effet moins fort.
L'intention est plus forte quand l'enseignant croit que ses collègues investis et compétents fournissent ce type de soutien, et non que seuls ceux qui "ne veulent pas en faire plus" s'en abstiennent.
La recherche de Maël Virat démontre que l'implication affective de l'enseignant est un pilier de la réussite et du bien-être de l'élève, avec des effets qui s'étendent bien au-delà des apprentissages scolaires (bien-être, symptômes dépressifs, rapport à l'autorité).
Cette implication, conceptualisée comme de l'amour compassionnel, n'est pas une simple inclination personnelle mais le résultat d'un écosystème complexe.
Pour la favoriser, il est essentiel d'agir à plusieurs niveaux :
• La formation : Intégrer la dimension relationnelle comme une compétence professionnelle à part entière.
• La culture d'établissement : Promouvoir une culture qui valorise les relations et reconnaît le soutien émotionnel comme partie intégrante du rôle enseignant.
• Le soutien aux professionnels : Assurer que les enseignants eux-mêmes se sentent soutenus par leurs pairs et leur hiérarchie, afin qu'ils puissent à leur tour devenir une base de sécurité pour leurs élèves.
Reviewer #3 (Public review):
This study investigates the characteristics of the autofluorescence signal excited by 740 nm 2-photon excitation, in the range of 420-500 nm, across the Drosophila brain. The fluorescence lifetime (FL) appears bi-exponential, with a short 0.4 ns time constant followed by a longer decay. The lifetime decay and the resulting parameter fits vary across the brain. The resulting maps reveal anatomical landmarks, which simultaneous imaging of genetically encoded fluorescent proteins help identify. Past work has shown that the autofluorescence decay time course reflects the balance of the redox enzyme NAD(P)H vs. its protein bound form. The ratio of free to bound NADPH is thought to indicate relative glycolysis vs. oxidative phosphorylation, and thus shifts in the free-to-bound ratio may indicate shifts in metabolic pathways. The basics of this measure have been demonstrated in other organisms, and this study is the first to use the FLIM module of the STELLARIS 8 FALCON microscope from Leica to measure autofluorescence lifetime in the brain of the fly. Methods include registering brains of different flies to a common template and masking out anatomical regions of interest using fluorescence proteins.
The analysis relies on fitting a FL decay model with two free parameters, f_free and T_bound. F_free is the fraction of the normalized curve contributed by a decaying exponential with a time constant 0.4 ns, thought to represent the FL of free NADPH or NADH, which apparently cannot be distinguished. T_bound is the time constant of the second exponential, with scalar amplitude = (1-f_free). The T_bound fit is thought to represent the decay time constant of protein bound NADPH, but can differ depending on the protein. The study shows that across the brain, T_bound can range from 0 to >5 ns, whereas f_free can range from 0.5 to 0.9 ns (Figure 1a). The paper beautifully lays out the analysis pipeline, providing a valuable resource. The full range of fits are reported, including maximum likelihood quality parameters, and can be benchmarks for future studies.
The authors measure properties of NADPH related autofluorescence of Kenyon Cells (KCs) of the fly mushroom body. The somata and calyx of mushroom bodies have a longer average tau_bound than other regions (Figure 1e); the f_free fit is higher for the calyx (input synapses) region than for KC somata; and the average across flies of average f_free fits in alpha/beta KC somata decreases slightly following paired presentation of odor and shock, compared to unpaired presentation of the same stimuli. Though the change is slight, no comparable change is detected in gamma KCs, suggesting that distributions of f_free derived from FL may be sensitive enough to measure changes in metabolic pathways following conditioning.
FLIM as a method is not yet widely prevalent in fly neuroscience, but recent demonstrations of its potential are likely to increase its use. Future efforts will benefit from the description of the properties of the autofluorescence signal to evaluate how autofluorescence may impact measures of FL of genetically engineered indicators.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
Summary:
The authors present a novel usage of fluorescence lifetime imaging microscopy (FLIM) to measure NAD(P)H autofluorescence in the Drosophila brain, as a proxy for cellular metabolic/redox states. This new method relies on the fact that both NADH and NADPH are autofluorescent, with a different excitation lifetime depending on whether they are free (indicating glycolysis) or protein-bound (indicating oxidative phosphorylation). The authors successfully use this method in Drosophila to measure changes in metabolic activity across different areas of the fly brain, with a particular focus on the main center for associative memory: the mushroom body.
Strengths:
The authors have made a commendable effort to explain the technical aspects of the method in accessible language. This clarity will benefit both non-experts seeking to understand the methodology and researchers interested in applying FLIM to Drosophila in other contexts.
Weaknesses:
(1) Despite being statistically significant, the learning-induced change in f-free in α/β Kenyon cells is minimal (a decrease from 0.76 to 0.73, with a high variability). The authors should provide justification for why they believe this small effect represents a meaningful shift in neuronal metabolic state.
We agree with the reviewer that the observed f_free shift averaged per individual, while statistically significant, is small. However, to our knowledge, this is the first study to investigate a physiological (i.e., not pharmacologically induced) variation in neuronal metabolism using FLIM. As such, there are no established expectations regarding the amplitude of the effect. In the revised manuscript, we have included an additional experiment involving the knockdown of ALAT in α/β Kenyon cells, which further supports our findings. We have also expanded the discussion to expose two potential reasons why this effect may appear modest.
(2) The lack of experiments examining the effects of long-term memory (after spaced or massed conditioning) seems like a missed opportunity. Such experiments could likely reveal more drastic changes in the metabolic profiles of KCs, as a consequence of memory consolidation processes.
We agree with the reviewer that investigating the effects of long-term memory on metabolism represent a valuable future path of investigation. An intrinsic caveat of autofluorescence measurement, however, is to identify the cellular origin of the observed changes. To this respect, long-term memory formation is not an ideal case study as its essential feature is expected to be a metabolic activation localized to Kenyon cells’ axons in the mushroom body vertical lobes (as shown in Comyn et al., 2024), where many different neuron subtypes send intricate processes. This is why we chose to first focus on middle-term memory, where changes at the level of the cell bodies could be expected from our previous work (Rabah et al., 2022). But our pioneer exploration of the applicability of NAD(P)H FLIM to brain metabolism monitoring in vivo now paves the way to extending it to the effect of other forms of memory.
(3) The discussion is mostly just a summary of the findings. It would be useful if the authors could discuss potential future applications of their method and new research questions that it could help address.
The discussion has been expanded by adding interpretations of the findings and remaining challenges.
Reviewer #2 (Public review):
This manuscript presents a compelling application of NAD(P)H fluorescence lifetime imaging (FLIM) to study metabolic activity in the Drosophila brain. The authors reveal regional differences in oxidative and glycolytic metabolism, with a particular focus on the mushroom body, a key structure involved in associative learning and memory. In particular, they identify metabolic shifts in α/β Kenyon cells following classical conditioning, consistent with their established role in energy-demanding middle- and long-term memories.
These results highlight the potential of label-free FLIM for in-vivo neural circuit studies, providing a powerful complement to genetically encoded sensors. This study is well-conducted and employs rigorous analysis, including careful curve fitting and well-designed controls, to ensure the robustness of its findings. It should serve as a valuable technical reference for researchers interested in using FLIM to study neural metabolism in vivo. Overall, this work represents an important step in the application of FLIM to study the interactions between metabolic processes, neural activity, and cognitive function.
Reviewer #3 (Public review):
This study investigates the characteristics of the autofluorescence signal excited by 740 nm 2-photon excitation, in the range of 420-500 nm, across the Drosophila brain. The fluorescence lifetime (FL) appears bi-exponential, with a short 0.4 ns time constant followed by a longer decay. The lifetime decay and the resulting parameter fits vary across the brain. The resulting maps reveal anatomical landmarks, which simultaneous imaging of genetically encoded fluorescent proteins helps to identify. Past work has shown that the autofluorescence decay time course reflects the balance of the redox enzyme NAD(P)H vs. its protein-bound form. The ratio of free-to-bound NADPH is thought to indicate relative glycolysis vs. oxidative phosphorylation, and thus shifts in the free-to-bound ratio may indicate shifts in metabolic pathways. The basics of this measure have been demonstrated in other organisms, and this study is the first to use the FLIM module of the STELLARIS 8 FALCON microscope from Leica to measure autofluorescence lifetime in the brain of the fly. Methods include registering the brains of different flies to a common template and masking out anatomical regions of interest using fluorescence proteins.
The analysis relies on fitting an FL decay model with two free parameters, f_free and t_bound. F_free is the fraction of the normalized curve contributed by a decaying exponential with a time constant of 0.4 ns, thought to represent the FL of free NADPH or NADH, which apparently cannot be distinguished. T_bound is the time constant of the second exponential, with scalar amplitude = (1-f_free). The T_bound fit is thought to represent the decay time constant of protein-bound NADPH but can differ depending on the protein. The study shows that across the brain, T_bound can range from 0 to >5 ns, whereas f_free can range from 0.5 to 0.9 (Figure 1a). These methods appear to be solid, the full range of fits are reported, including maximum likelihood quality parameters, and can be benchmarks for future studies.
The authors measure the properties of NADPH-related autofluorescence of Kenyon Cells(KCs) of the fly mushroom body. The results from the three main figures are:
(1) Somata and calyx of mushroom bodies have a longer average tau_bound than other regions (Figure 1e);
(2) The f_free fit is higher for the calyx (input synapses) region than for KC somata (Figure 2b);
(3) The average across flies of average f_free fits in alpha/beta KC somata decreases from 0.734 to 0.718. Based on the first two findings, an accurate title would be "Autofluorecense lifetime imaging reveals regional differences in NADPH state in Drosophila mushroom bodies."
The third finding is the basis for the title of the paper and the support for this claim is unconvincing. First, the difference in alpha/beta f_free (p-value of 4.98E-2) is small compared to the measured difference in f_free between somas and calyces. It's smaller even than the difference in average soma f_free across datasets (Figure 2b vs c). The metric is also quite derived; first, the model is fit to each (binned) voxel, then the distribution across voxels is averaged and then averaged across flies. If the voxel distributions of f_free are similar to those shown in Supplementary Figure 2, then the actual f_free fits could range between 0.6-0.8. A more convincing statistical test might be to compare the distributions across voxels between alpha/beta vs alpha'/beta' vs. gamma KCs, perhaps with bootstrapping and including appropriate controls for multiple comparisons.
The difference observed is indeed modest relative to the variability of f_free measurements in other contexts. The fact that the difference observed between the somata region and the calyx is larger is not necessarily surprising. Indeed, these areas have different anatomical compositions that may result in different basal metabolic profiles. This is suggested by Figure 1b which shows that the cortex and neuropile have different metabolic signatures. Differences in average f_free values in the somata region can indeed be observed between naive and conditioned flies. However, all comparisons in the article were performed between groups of flies imaged within the same experimental batches, ensuring that external factors were largely controlled for. This absence of control makes it difficult to extract meaningful information from the comparison between naive and conditioned flies.
We agree with the reviewer that the choice of the metric was indeed not well justified in the first manuscript. In the new manuscript, we have tried to illustrate the reasons for this choice with the example of the comparison of f_free in alpha/beta neurons between unpaired and paired conditioning (Dataset 8). First, the idea of averaging across voxels is supported by the fact that the distributions of decay parameters within a single image are predominantly unimodal. Examples for Dataset 8 are now provided in the new Sup. Figure 14. Second, an interpretable comparison between multiple groups of distributions is, to our knowledge, not straightforward to implement. It is now discussed in Supplementary information. To measure interpretable differences in the shapes of the distributions we computed the first three moments of distributions of f_free for Dataset 8 and compared the values obtained between conditions (see Supplementary information and new Sup. Figure 15). Third, averaging across individuals allows to give each experimental subject the same weight in the comparisons.
I recommend the authors address two concerns. First, what degree of fluctuation in autofluorescence decay can we expect over time, e.g. over circadian cycles? That would be helpful in evaluating the magnitude of changes following conditioning. And second, if the authors think that metabolism shifts to OXPHOS over glycolosis, are there further genetic manipulations they could make? They test LDH knockdown in gamma KCs, why not knock it down in alpha/beta neurons? The prediction might be that if it prevents the shift to OXPHOS, the shift in f_free distribution in alpha/beta KCs would be attenuated. The extensive library of genetic reagents is an advantage of working with flies, but it comes with a higher standard for corroborating claims.
In the present study, we used control groups to account for broad fluctuations induced by external factors such as the circadian cycle. We agree with the reviewer that a detailed characterization of circadian variations in the decay parameters would be valuable for assessing the magnitude of conditioning-induced shifts. We have integrated this relevant suggestion in the Discussion. Conducting such an investigation lies unfortunately beyond the scope and means of the current project.
In line with the suggestion of the reviewer, we have included a new experiment to test the influence of the knockdown of ALAT on the conditioning-induced shift measured in alpha/beta neurons. This choice is motivated in the new manuscript. The obtained result shows that no shift is detected in the mutant flies, in accordance with our hypothesis.
FLIM as a method is not yet widely prevalent in fly neuroscience, but recent demonstrations of its potential are likely to increase its use. Future efforts will benefit from the description of the properties of the autofluorescence signal to evaluate how autofluorescence may impact measures of FL of genetically engineered indicators.
Recommendations for the authors
Reviewer #1 (Recommendations for the authors):
(1) Y axes in Figures 1e, 2c, 3b,c are misleading. They must start at 0.
Although we agree that making the Y axes start at 0 is preferable, in our case it makes it difficult to observe the dispersion of the data at the same time (your next suggestion). To make it clearer to the reader that the axes do not start at 0, a broken Y-axis is now displayed in every concerned figure.
(2) These same plots should have individual data points represented, for increased clarity and transparency.
Individual data points were added on all boxplots.
Reviewer #2 (Recommendations for the authors):
I am evaluating this paper as a fly neuroscientist with experience in neurophysiology, including calcium imaging. I have little experience with FLIM but anticipate its use growing as more microscopes and killer apps are developed. From this perspective, I value the opportunity to dig into FLIM and try to understand this autofluorescence signal. I think the effort to show each piece of the analysis pipeline is valuable. The figures are quite beautiful and easy to follow. My main suggestion is to consider moving some of the supplemental data to the main figures. eLife allows unlimited figures, moving key pieces of the pipeline to the main figures would make for smoother reading and emphasize the technical care taken in this study.
We thank the reviewer for their feedback. Following their advice we have moved panels from the supplementary figures to the main text (see new Figure 2).
Unfortunately, the scientific questions and biological data do not rise to the typical standard in the field to support the claims in the title, "In vivo autofluorescence lifetime imaging of the Drosophila brain captures metabolic shifts associated with memory formation". The authors also clearly state what the next steps are: "hypothesis-driven approaches that rely on metabolite-specific sensors" (Intro). The advantage of fly neuroscience is the extensive library of genetic reagents that enable perturbations. The key manipulation in this study is the electric shock conditioning paradigm that subtly shifts the distribution of a parameter fit to an exponential decay in the somas of alpha/beta KCs vs others. This feels like an initial finding that deserves follow-up; but is it a large enough result to motivate a future student to pick this project up? The larger effect appears to be the gradients in f_free across KCs overall (Figure 2b). How does this change with conditioning?
We acknowledge that the observed metabolic shift is modest relative to the variability of f_free and agree that additional corroborating experiments would further strengthen this result. Nevertheless, we believe it remains a valid and valuable finding that will be of interest to researchers in the field. The reviewer is right in pointing out that the gradient across KCs is higher in magnitude, however, the fact that this technique can also report experience-dependent changes, in addition to innate heterogeneities across different cell types, is a major incentive for people who could be interested in applying NAD(P)H FLIM in the future. For this reason, we consider it appropriate to retain mention of the memory-induced shift in the title, while making it less assertive and adding a reference to the structural heterogeneities of f_free revealed in the study. We have also rephrased the abstract to adopt a more cautious tone and expanded the discussion to clarify why a low-magnitude shift in f_free can still carry biological significance in this context. Finally, we have added the results of a new set of data involving the knockdown of ALAT in Kenyon cells, to further support the relevance of our observation relative to memory formation, despite its small magnitude. We believe that these elements together form a good basis for future investigations and that the manuscript merits publication in its present form.
Together, I would recommend reshaping the paper as a methods paper that asks the question, what are the spatial properties of NADPH FL across the brain? The importance of this question is clear in the context of other work on energy metabolism in the MBs. 2P FLIM will likely always have to account for autofluorescence, so this will be of interest. The careful technical work that is the strength of the manuscript could be featured, and whether conditioning shifts f_free could be a curio that might entice future work.
By transferring panels of the supplementary figures to the main text (see new Figure 2) as suggested by Reviewer 2, we have reinforced the methodological part of the manuscript. For the reasons explained above, we however still mention the ‘biological’ findings in the title and abstract.
Minor recommendations on science:
Figure 2C. Plotting either individual data points or distributions would be more convincing.
Individual data points were added on all boxplots.
There are a few mentions of glia. What are the authors' expectations for metabolic pathways in glia vs. neurons? Are glia expected to use one more than the other? The work by Rabah suggests it should be different and perhaps complementary to neurons. Can a glial marker be used in addition to KC markers? This seems crucial to being able to distinguish metabolic changes in KC somata from those in glia.
Drosophila cortex glia are thought to play a similar role as astrocytes in vertebrates (see Introduction). In that perspective, we expect cortex glia to display a higher level of glycolysis than neurons. The work by Rabah et al. is coherent with this hypothesis. Reviewer 2 is right in pointing out that using a glial marker would be interesting. However, current technical limitations make such experiments challenging. These limitations are now exposed in the discussion.
The question of whether KC somata positions are stereotyped can probably be answered in other ways as well. For example, the KCs are in the FAFB connectomic data set and the hemibrain. How do the somata positions compare?
The reviewer’s suggestion is indeed interesting. However, the FAFB and hemibrain connectomic datasets are based on only two individual flies, which probably limits their suitability for assessing the stereotypy of KC subtype distributions. In addition, aligning our data with the FAFB dataset would represent substantial additional work.
The free parameter tau_bound is mysterious if it can be influenced by the identity of the protein. Are there candidate NADPH binding partners that have a spatial distribution in confocal images that could explain the difference between somas and calyx?
There are indeed dozens of NADH- or NADPH-binding proteins. For this reason, in all studies implementing exponential fitting of metabolic FLIM data, tau_bound is considered a complex combination of the contributions from many different proteins. In addition, one should keep in mind that the number of cell types contributing to the autofluorescence signal in the mushroom body calyx (Kenyon cells, astrocyte-like and ensheathing glia, APL neurons, olfactory projection neurons, dopamine neurons) is much higher than in the somas (only Kenyon cells and cortex glia). This could also participate in the observed difference. Hence, focusing on intracellular heterogeneities of potential NAD(P)H binding partners seems premature at that stage.
The phrase "noticeable but not statistically significant" is misleading.
We agree with the reviewer and have removed “noticeable but” from the sentence in the new version of the manuscript.
Minor recommendations on presentation:
The Introduction can be streamlined.
We agree that some parts of the Introduction can seem a bit long for experts of a particular field. However, we think that this level of detail makes the article easily accessible for neuroscientists working on Drosophila and other animal models but not necessarily with FLIM, as well as for experts in energy metabolism that may be familiar with FLIM but not with Drosophila neuroscience.
Reviewer #2 (Public review):
Summary:
In this paper, the authors report on a case-control study in which participants with chronic pain (TMD) were compared to controls on performance of a three-option learning task. The authors find no difference in task behavior, but fit a model to this behavior and suggest that differences in the model-derived metrics (specifically, change in learning rate/estimated volatility/model estimated uncertainty) reveal a relevant between-group effect. They report a mediation effect suggesting that group differences on self-report apathy may be partially mediated by this uncertainty adaptation result.
Strengths:
The role of sensitivity to uncertainty in pathological states is an interesting question and is the focus of a reasonable amount of research at present. This paper provides a useful assessment of these processes in people with chronic pain.
Weaknesses:
(1) The interpretation of the model in the absence of any apparent behavioral effect is not convincing. The model is quite complex with a number of free parameters (what these parameters are is not well explained in the methods, although they seem to be presented in the supplement). These parameters are fitted to participant choice behavior - that is, they explain some sort of group difference in this choice behavior. The authors haven't been able to demonstrate what this difference is. The graphs of learning rate per group (Figure 2) suggest that the control group has a higher initial learning rate and a lower later learning rate. If this were actually the case, you would expect to see it reflected in the choice data (the control group should show higher lose-shift behavior earlier on, with this then declining over time, and the TMD group should show no change). This behavior is not apparent. The absence of a clear effect on behavior suggests that the model results are more likely to be spurious.
(2) As far as I could see, the actual parameters of the model are not reported. The results (Figure 2) illustrate the trial-level model estimated uncertainty/learning rate, etc, but these differ because the fitted model parameters differ. The graphs look like there are substantial differences in v0 (which was not well recovered), but presumably lambda, at least, also differs. The mean(SD) group values for these parameters should be reported, as should the correlations between them (it looks very much like they will be correlated).
(3) The task used seems ill-suited to measuring the reported process. The authors report the performance of a restless bandit task and find an effect on uncertainty adaptation. The task does not manipulate uncertainty (there are no periods of high/low uncertainty) and so the only adaptation that occurs in the task is the change from what appears to be the participants' prior beliefs about uncertainty (which appear to be very different between groups - i.e. the lines in Figure 2a,b,c are very different at trial 0). If the authors are interested in measuring adaptation to uncertainty, it would clearly be more useful to present participants with periods of higher or lower uncertainty.
(4) The main factor driving the better fit of the authors' preferred model over listed alternatives seems to be the inclusion of an additive uncertainty term in the softmax-this differentiates the chosen model from the other two Kalman filter-based models that perform less well. But a similar term is not included in the RW models-given the uncertainty of a binary outcome can be estimated as p(1-p), and the RW models are estimating p, this would seem relatively straightforward to do. It would be useful to know if the factor that actually drives better model fit is indeed in the decision stage (rather than the learning stage).
Reviewer #3 (Public review):
This paper applies a computational model to behavior in a probabilistic operant reward learning task (a 3-armed bandit) to uncover differences between individuals with temporomandibular disorder (TMD) compared with healthy controls. Integrating computational principles and models into pain research is an important direction, and the findings here suggest that TMD is associated with subtle changes in how uncertainty is represented over time as individuals learn to make choices that maximize reward. There are a number of strengths, including the comparison of a volatile Kalman filter (vKF) model to some standard base models (Rescorla Wagner with 1 or 2 learning rates) and parameter recovery analyses suggesting that the combination of task and vKF model may be able to capture some properties of learning and decision-making under uncertainty that may be altered in those suffering from chronic pain-related conditions.
I've focused my comments in four areas: (1) Questions about the patient population, (2) Questions about what the findings here mean in terms of underlying cognitive/motivational processes, (3) Questions about the broader implications for understanding individuals with TMD and other chronic pain-related disorders, and (4) Technical questions about the models and results.
(1) Patient population
This is a computational modelling study, so it is light on characterization of the population, but the patient characteristics could matter. The paper suggests they were hospitalized, but this is not a condition that requires hospitalization per se. It would be helpful to connect and compare the patient characteristics with large-scale studies of TMD, such as the OPPERA study led by Maixner, Fillingim, and Slade.
(2) What cognitive/motivational processes are altered in TMD
The study finds a pattern of alterations in TMD patients that seems clear in Figure 2. Healthy controls (HC) start the task with high estimates of volatility, uncertainty, and learning rate, which drop over the course of the task session. This is consistent with a learner that is initially uncertain about the structure of the environment (i.e., which options are rewarded and how the contingencies change over time) but learns that there is a fixed or slowly changing mean and stationary variance. The TMD patients start off with much lower volatility, uncertainty, and learning rate - which are actually all near 0 - and they remain stable over the course of learning. This is consistent with a learner who believes they know the structure of the environment and ignores new information.
What is surprising is that this pattern of changes over time was found in spite of null group differences in a number of aspects of performance: (1) stay rate, (2) switch rate, (3) win-stay/lose-switch behaviors, (4) overall performance (corrected for chance level), (5) response times, (6) autocorrelation, (7) correlations between participants' choice probability and each option's average reward rate, (7) choice consistency (though how operationalized is not described?), (8) win-stay-lose-shift patterns over time. I'm curious about how the patterns in Figure 2 would emerge if standard aspects of performance are essentially similar across groups (though the study cannot provide evidence in favor of the null). It will be important to replicate these patterns in larger, independent samples with preregistered analyses.
The authors believe that this pattern of findings reveals that TMD patients "maintain a chronically heightened sensitivity to environmental changes" and relate the findings to predictive processing, a hallmark of which (in its simplest form) is precision-weighted updating of priors. They also state that the findings are not related to reduced overall attentiveness or failure to understand the task, but describe them as deficits or impairments in calibrating uncertainty.
The pattern of differences could, in fact, result from differences in prior beliefs, conceptualization of the task, or learning. Unpacking these will be important steps for future work, along with direct measures of priors, cognitive processes during learning, and precision-weighted updating.
(3) Implications for understanding chronic pain
If the findings and conclusions of the paper are correct, individuals with TMD and perhaps other pain-related disorders may have fundamental alterations in the ways in which they make decisions about even simple monetary rewards. The broader questions for the field concern (1) how generalizable such alterations are across tasks, (2) how generalizable they are across patient groups and, conversely, how specific they are to TMD or chronic pain, (3) whether they are the result of neurological dysfunction, as opposed to (e.g.) adaptive strategies or assumptions about the environment/task structure.
It will be important to understand which features of patients' and/or controls' cognition are driving the changes. For example, could the performance differences observed here be attributable to a reduced or altered understanding of the task instructions, more uncertainty about the rules of the game, different assumptions about environments (i.e., that they are more volatile/uncertain or less so), or reduced attention or interest in optimizing performance? Are the controls OVERconfident in their understanding of the environment?
This set of questions will not be easy to answer and will be the work of many groups for many years to come. It is a judgment call how far any one paper must go to address them, but my view is that it is a collaborative effort. Start with a finding, replicate it across labs, take the replicable phenomena and work to unpack the underlying questions. The field must determine whether it is this particular task with this model that produces case-control differences (and why), or whether the findings generalize broadly. Would we see the same findings for monetary losses, sounds, and social rewards? Tasks with painful stimuli instead of rewards?
Another set of questions concerns the space of computational models tested, and whether their parameters are identifiable. An alteration in estimated volatility or learning rate, for example, can come from multiple sources. In one model, it might appear as a learning rate change and in another as a confirmation bias. It would be interesting in this regard to compare the "mechanisms" (parameters) of other models used in pain neuroscience, e.g., models by Seymour, Mancini, Jepma, Petzschner, Smith, Chen, and others (just to name a few).
One immediate next step here could be to formally compare the performance of both patients and controls to normatively optimal models of performance (e.g., Bayes optimal models under different assumptions). This could also help us understand whether the differences in patients reflect deficits and what further experiments we would need to pin that down.<br /> In addition, the volatility parameter in the computational model correlated with apathy. This is interesting. Is there a way to distinguish apathy as a particular clinical characteristic and feature of TMD from apathy in the sense of general disinterest in optimal performance that may characterize many groups?
If we know this, what actionable steps does it lead us to take? Could we take steps to reduce apathy and thus help TMD patients better calibrate to environmental uncertainty in their lives? Or take steps to recalibrate uncertainty (i.e., increase uncertainty adaptation), with benefits on apathy? A hallmark of a finding that the field can build off of is the questions it raises.
(4) Technical questions about the models and results
Clarification of some technical points would help interpret the paper and findings further:
(a) Was the reward probability truly random? Was the random walk different for each person, or constrained?
(b) When were self-report measures administered, and how?
(c) Pain assessments: What types of pain? Was a body map assessed? Widespreadness? Pain at the time of the test, or pain in general?
(d) Parameter recovery: As you point out, r = 0.47 seems very low for recovery of the true quantity, but this depends on noise levels and on how the parameter space is sampled. Is this noise-free recovery, and is it robust to noise? Are the examples of true parameters drawn from the space of participants, or do they otherwise systematically sample the space of true parameters?
(e) What are the covariances across parameter estimates and resultant confusability of parameter estimates (e.g., confusion matrix)?
(f) It would be helpful to have a direct statistical comparison of controls and TMD on model parameter estimates.
(g) Null statistical findings on differences in correlations should not be interpreted as a lack of a true effect. Bayes Factors could help, but an analysis of them will show that hundreds of people are needed before it is possible to say there are no differences with reasonable certainty. Some journals enforce rules around the kinds of language used to describe null statistical findings, and I think it would be helpful to adopt them more broadly.
(h) What is normatively optimal in this task? Are TMD patients less so, or not? The paper states "aberrant precision (uncertainty) weighting and misestimation of environmental volatility". But: are they misestimates?
(i) It's not clear how well the choice of prior variance for all parameters (6.25) is informed by previous research, as sensible values may be task- and context-dependent. Are the main findings robust to how priors are specified in the HBI model?
Follow-up post by Brennan Kenneth Brown after a previous one urging people to their own web sites. The response was large, but w many questions on how to discover more, and 'find the others'
You probably don't need Oh My Zsh
He made you for a highway to my bed; But I, a maid, die maiden-widowed. Come, cords, come, nurse; I'll to my wedding-bed; 1860And death, not Romeo, take my maidenhead!
Symbolism: The ropes were for their wedding night. Now they’re useless.
Some word there was, worser than Tybalt's death, That murder'd me: I would forget it fain; But, O, it presses to my memory, Like damned guilty deeds to sinners' minds: 1835'Tybalt is dead, and Romeo—banished;' That 'banished,' that one word 'banished,' Hath slain ten thousand Tybalts. Tybalt's death Was woe enough, if it had ended there:
Theme: Exile is a living death. She’d rather have Romeo dead than gone.
But, wherefore, villain, didst thou kill my cousin? That villain cousin would have kill'd my husband: 1825Back, foolish tears, back to your native spring; Your tributary drops belong to woe, Which you, mistaking, offer up to joy. My husband lives, that Tybalt would have slain; And Tybalt's dead, that would have slain my husband:
She realizes Tybalt would have killed Romeo. This comforts her.
Dove-feather'd raven!
Oxymoron: A bird of peace (dove) that’s actually a dark bird (raven). Romeo seemed good but did evil.
This torture should be roar'd in dismal hell. Hath Romeo slain himself? say thou but 'I,' And that bare vowel 'I' shall poison more Than the death-darting eye of cockatrice: I am not I, if there be such an I; 1770Or those eyes shut, that make thee answer 'I.' If he be slain, say 'I'; or if not, no: Brief sounds determine of my weal or woe.
Juliet thinks Romeo killed himself. She’s frantic and heartbroken.
Nurse. Ah, well-a-day! he's dead, he's dead, he's dead!
Dramatic Irony: Nurse says “he’s dead” but doesn’t say who. Juliet thinks it’s Romeo.
Prince Escalus. And for that offence 1705Immediately we do exile him hence:
Plot: Prince exiles Romeo instead of death. This is “mercy” but will cause more problems.
Prince Escalus. And for that offence 1705Immediately we do exile him hence:
Plot: Prince exiles Romeo instead of death. This is “mercy” but will cause more problems.
Benvolio. Tybalt, here slain, whom Romeo's hand did slay; Romeo that spoke him fair, bade him bethink 1670How nice the quarrel was, and urged withal Your high displeasure: all this uttered With gentle breath, calm look, knees humbly bow'd, Could not take truce with the unruly spleen Of Tybalt deaf to peace, but that he tilts 1675With piercing steel at bold Mercutio's breast, Who all as hot, turns deadly point to point, And, with a martial scorn, with one hand beats Cold death aside, and with the other sends It back to Tybalt, whose dexterity, 1680Retorts it: Romeo he cries aloud, 'Hold, friends! friends, part!' and, swifter than his tongue, His agile arm beats down their fatal points, And 'twixt them rushes; underneath whose arm 1685An envious thrust from Tybalt hit the life Of stout Mercutio, and then Tybalt fled; But by and by comes back to Romeo, Who had but newly entertain'd revenge, And to 't they go like lightning, for, ere I 1690Could draw to part them, was stout Tybalt slain. And, as he fell, did Romeo turn and fly. This is the truth, or let Benvolio die.
Plot: Benvolio tells the truth about the fight, defending Romeo.
Mercutio. No, 'tis not so deep as a well, nor so wide as a church-door; but 'tis enough,'twill serve: ask for me to-morrow, and you shall find me a grave man.
Even dying, Mercutio makes puns (“grave man”). Shows his wit and bravery.
Mercutio. Good king of cats, nothing but one of your nine lives; that I mean to make bold withal, and as you shall use me hereafter, drybeat the rest of the eight. Will you pluck your sword out of his pitcher by the ears? make haste, lest mine be about your 1580ears ere it be out.
Mercutio calls Tybalt “king of cats” (mockery) and challenges him to fight.
Mercutio. And but one word with one of us? couple it with something; make it a word and a blow.
Wordplay: “A word and a blow” means talk then fight. Mercutio is looking for a fight.
Mercutio. Nay, an there were two such, we should have none shortly, for one would kill the other. Thou! why, thou wilt quarrel with a man that hath a hair more, 1515or a hair less, in his beard, than thou hast: thou wilt quarrel with a man for cracking nuts, having no other reason but because thou hast hazel eyes: what eye but such an eye would spy out such a quarrel? Thy head is as fun of quarrels as an egg is full of 1520meat, and yet thy head hath been beaten as addle as an egg for quarrelling: thou hast quarrelled with a man for coughing in the street, because he hath wakened thy dog that hath lain asleep in the sun: didst thou not fall out with a tailor for wearing 1525his new doublet before Easter? with another, for tying his new shoes with old riband? and yet thou wilt tutor me from quarrelling!
Mercutio’s exaggeration. Hyperbole: Mercutio claims Benvolio would fight over anything, even a cough or someone’s shoes. This is funny exaggeration.
Reviewer #1 (Public review):
Summary:
This manuscript by Wang et al. reports the potential involvement of an asymmetric neurocircuit in the sympathetic control of liver glucose metabolism.
Strengths:
The concept that the contralateral brain-liver neurocircuit preferentially regulates each liver lobe may be interesting.
Weaknesses:
However, the experimental evidence presented did not support the study's central conclusion.
(1) Pseudorabies virus (PRV) tracing experiment:<br /> The liver not only possesses sympathetic innervations but also vagal sensory innervations. The experimental setup failed to distinguish whether the PRV-labeling of LPGi (Lateral Paragigantocellular Nucleus) is derived from sympathetic or vagal sensory inputs to the liver.
(2) Impact on pancreas:<br /> The celiac ganglia not only provide sympathetic innervations to the liver but also to the pancreas, the central endocrine organ for glucose metabolism. The chemogenetic manipulation of LPGi failed to consider a direct impact on the secretion of insulin and glucagon from the pancreas.
(3) Neuroanatomy of the brain-liver neurocircuit:<br /> The current study and its conclusion are based on a speculative brain-liver sympathetic circuit without the necessary anatomical information downstream of LPGi.
(4) Local manipulation of the celiac ganglia:<br /> The left and right ganglia of mice are not separate from each other but rather anatomically connected. The claim that the local injection of AAV in the left or right ganglion without affecting the other side is against this basic anatomical feature.
Reviewer #2 (Public review):
Summary:
The manuscript by Wang and colleagues aims to determine whether the left and right LPGi differentially regulate hepatic glucose metabolism and to reveal decussation of hepatic sympathetic nerves.
The authors used tissue clearing to identify sympathetic fibers in the liver lobes, then injected PRV into the hepatic lobes. Five days post-injection, PRV-labeled neurons in the LPGi were identified. The results indicated contralateral dominance of premotor neurons and partial innervation of more than one lobe. Then the authors activated each side of the LPGi, resulting in a greater increase in blood glucose levels after right-sided activation than after left-sided activation, as well as changes in protein expression in the liver lobes. These data suggested modulation of HGP (hepatic glucose production) in a lobe-specific manner. Chemical denervation of a particular lobe did not affect glucose levels due to compensation by the other lobes. In addition, nerve bundles decussate in the hepatic portal region.
Strengths:
The manuscript is timely and relevant. It is important to understand the sympathetic regulation of the liver and the contribution of each lobe to hepatic glucose production. The authors use state-of-the-art methodology.
Weaknesses:
(1) The wording/terminology used in the manuscript is misleading, and it is not used in the proper context. For instance, the goal of the study is "to investigate whether cerebral hemispheres differentially regulate hepatic glucose metabolism..." (see abstract); however, the authors focus on the brainstem (a single structure without hemispheres). Similarly, symmetric is not the best word for the projections.
(2) Sparse labeling of liver-related neurons was shown in the LPGi (Figure 1). It would be ideal to have lower magnification images to show the area. Higher quality images would be necessary, as it is difficult to identify brainstem areas. The low number of labeled neurons in the LPGi after five days of inoculation is surprising. Previous findings showed extensive labeling in the ventral brainstem at four days post-inoculation (Desmoulins et al., 2025). Unfortunately, it is not possible to compare the injection paradigm/methods because the PRV inoculation is missing from the methods section. If the PRV is different from the previously published viral tracers, time-dependent studies to determine the order of neurons and the time course of infection would be necessary.
(3) Not all LPGi cells are liver-related. Was the entire LPGi population stimulated, or was it done in a cell-type-specific manner? What was the strain, sex, and age of the mice? What was the rationale for using the particular viral constructs?
(4) The authors should consider the effect of stimulation of double-labeled neurons (innervating more than one lobe) and potential confounding effects regarding other physiological functions.
(5) The authors state that "central projections directly descend along the sympathetic chain to the celiac-superior mesenteric ganglia". What they mean is unclear. Do the authors refer to pre-ganglionic neurons or premotor neurons? How does it fit with the previous literature?
(6) How was the chemical denervation completed for the individual lobes?
(7) The Western Blot images look like they are from different blots, but there are no details provided regarding protein amount (loading) or housekeeping. What was the reason to switch beta-actin and alpha-tubulin? In Figures 3F -G, the GS expression is not a good representative image. Were chemiluminescence or fluorescence antibodies used? Were the membranes reused?
(8) Key references using PRV for liver innervation studies are missing (Stanley et al, 2010 [PMID: 20351287]; Torres et al., 2021 [PMID: 34231420]; Desmoulins et al., 2025 [PMID: 39647176]).
Reviewer #4 (Public review):
Summary:
The studies here are highly informative in terms of anatomical tracing and sympathetic nerve function in the liver related to glucose levels, but given that they are performed in a single species, it is challenging to translated them to humans, or to determine whether these neural circuits are evolutionarily conserved. Dual-labeling anatomical studies are elegant, and the addition of chemogenetic and optogenetic studies is mechanistically informative. Denervation studies lack appropriate controls, and the role of sensory innervation in the liver is overlooked.
Specific Weaknesses - Major:
(1) The species name should be included in the title.
(2) Tyrosine hydroxylase was used to mark sympathetic fibers in the liver, but this marker also hits a portion of sensory fibers that need to be ruled out in whole-mount imaging data
(3) Chemogenetic and optogenetic data demonstrating hyperglycemia should be described in the context of prior work demonstrating liver nerve involvement in these processes. There is only a brief mention in the Discussion currently, but comparing methods and observations would be helpful.
(4) Sympathetic denervation with 6-OHDA can drive compensatory increases to tissue sensory innervation, and this should be measured in the liver denervation studies to implicate potential crosstalk, especially given the increase in LPGi cFOS that may be due to afferent nerve activity. Compensatory sympathetic drive may not be the only culprit, though it is clearly assumed to be. The sensory or parasympathetic/vagal innervation of the liver is altogether ignored in this paper and could be better described in general.
Author response:
Public Reviews:
Reviewer #1 (Public review):
Summary:
This manuscript by Wang et al. reports the potential involvement of an asymmetric neurocircuit in the sympathetic control of liver glucose metabolism.
Strengths:
The concept that the contralateral brain-liver neurocircuit preferentially regulates each liver lobe may be interesting.
Weaknesses:
However, the experimental evidence presented did not support the study's central conclusion.
We sincerely thank the reviewer for recognizing the conceptual novelty of our work and for constructive comments aimed at enhancing its rigor and clarity. In response, we will carry out targeted experiments to address the points raised, including: (i) further characterization of LPGi projections to vagal and sympathetic circuits; (ii) evaluation of potential pancreatic involvement; and (ii) validation of the specificity of chemogenetic activation within the proposed circuit. We anticipate completing the revised version within 8 weeks.
(1) Pseudorabies virus (PRV) tracing experiment:
The liver not only possesses sympathetic innervations but also vagal sensory innervations. The experimental setup failed to distinguish whether the PRV-labeling of LPGi (Lateral Paragigantocellular Nucleus) is derived from sympathetic or vagal sensory inputs to the liver.
Thank you for raising this important point. We fully agree that the liver receives both sympathetic and vagal sensory innervation, and we acknowledge that PRV-based tracing alone does not definitively distinguish between these two pathways. This represents a limitation of the original experimental design.
Based on established anatomical literature as well as our experimental observations, vagal sensory neuron cell bodies reside in the nodose ganglion (NG), and their central projections terminate predominantly in the nucleus of the solitary tract (NTS) (Nature. 2023;623(7986):387-396; Curr Biol. 2020;30(20):3986-3998.e5.), which is located in the dorsomedial medulla. In contrast, the LPGi, together with other sympathetic-related nuclei, is predominantly distributed in the ventral medulla (Cell Metab. 2025;37(11):2264-2279.e10; Nat Commun. 2022;13(1):5079.).
To directly assess the contribution of vagal sensory pathways, we will perform an additional PRV tracing experiment using two groups of mice: one with bilateral nodose ganglion (NG) removal and a sham-operated control group. Identical PRV injections will be delivered to the liver in both groups, and PRV labeling in the LPGi will be quantitatively compared. Preservation of LPGi labeling following NG ablation would indicate that PRV transmission occurs primarily via sympathetic, rather than vagal sensory, pathways. These data will be incorporated into the revised manuscript and are expected to be completed within 3 weeks.
(2) Impact on pancreas:
The celiac ganglia not only provide sympathetic innervations to the liver but also to the pancreas, the central endocrine organ for glucose metabolism. The chemogenetic manipulation of LPGi failed to consider a direct impact on the secretion of insulin and glucagon from the pancreas.
Thank you for this important comment. We agree that the celiac ganglia (CG) provide sympathetic innervation not only to the liver but also to the pancreas, which plays a central role in glucose homeostasis through the secretion of both insulin and glucagon. Therefore, the potential pancreatic implications associated with LPGi chemogenetic manipulation worth careful consideration.
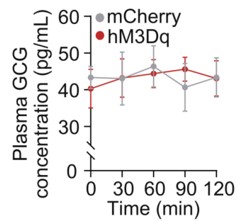
To address this concern, we examined circulating glucagon levels following chemogenetic manipulation of the LPGi. As shown in the Supplementary Figure below, plasma glucagon (GCG) concentrations were not significantly altered at 30, 60, 90, or 120 minutes compared with control mice (n = 6), indicating that LPGi manipulation does not measurably affect glucagon secretion under our experimental conditions.
We acknowledge that insulin secretion was not assessed in the study, which represents an important limitation given the pancreatic innervation of the CG. To further strengthen our interpretation, we are performing additional experiments in newly prepared mice to measure circulating insulin levels following LPGi manipulation. These data together with Author response image 1 below will be included in the revised manuscript upon completion.
Author response image 1.
Plasma concentrations of GCG in mice following LPGi GABAergic neurons activation.
(3) Neuroanatomy of the brain-liver neurocircuit:<br /> The current study and its conclusion are based on a speculative brain-liver sympathetic circuit without the necessary anatomical information downstream of LPGi.
Thank you for raising this important point. A clear anatomical definition of the downstream pathways linking the brain to the liver is essential for interpreting the proposed brain-liver sympathetic circuit.
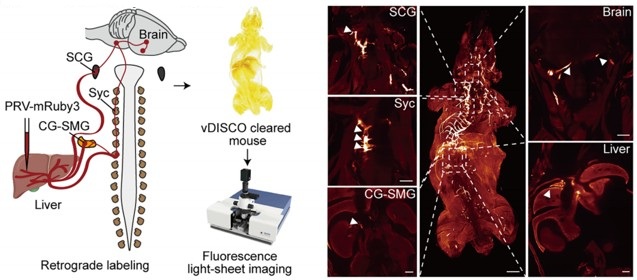
However, the present study (Figure 4A) provides direct anatomical evidence supporting the organization of the brain–liver sympathetic neurocircuit. These observations are consistent with our recent detailed characterization of the brain-liver sympathetic circuit published in Cell Metabolism (Cell Metab. 2025;37(11):2264–2279), LPGi GABAergic neurons inhibit GABAergic neurons in the caudal ventrolateral medulla (CVLM). Disinhibition of CVLM reduces GABAergic suppression of rostral ventrolateral medulla (RVLM) neurons, which are key excitatory drivers of sympathetic tone. RVLM neurons project to sympathetic preganglionic neurons in the sympathetic chain (Syc). These neurons synapse with postganglionic sympathetic neurons in ganglia such as the celiac-superior mesenteric ganglion (CG-SMG). Postganglionic sympathetic fibers then innervate the liver, releasing NE to activate hepatic β<sub>2</sub>-adrenergic receptors and stimulate HGP.
Together, these data establish a coherent anatomical basis for the proposed brain-liver sympathetic pathway and clarify the downstream organization relevant to the functional experiments presented here.
Author response image 2.
Tracing scheme (Left) and whole-mount imaging (Right) of PRV-labeled brain-liver neurocircuit. Scale bars, 3,000 (whole mount) or 1,000 (optical sections) μm.
(4) Local manipulation of the celiac ganglia:<br /> The left and right ganglia of mice are not separate from each other but rather anatomically connected. The claim that the local injection of AAV in the left or right ganglion without affecting the other side is against this basic anatomical feature.
Thank you for raising this important anatomical point. We fully acknowledge that the left and right celiac ganglia (CG) in mice are interconnected, and that unilateral viral injection could theoretically affect the contralateral side. The celiac–superior mesenteric ganglion (CG-SMG) complex serves as a major sympathetic hub that regulates visceral organ functions. Recent transcriptomic, anatomical, and functional studies have revealed that the CG-SMG is not a homogeneous structure but is composed of molecularly and functionally distinct neuronal populations. These populations exhibit specialized projection patterns and regulate different aspects of gastrointestinal physiology, supporting a model of modular sympathetic control. (Nature. 2025 Jan;637(8047):895-902). Therefore, we were aware of this phenomenon during the initial stages of these experiments.
To minimize unintended spread to the contralateral CG, we took two complementary approaches.
First, we optimized the injection strategy by using an extremely small injection volume (100 nL per site), with a very slow infusion rate (50 nL/min), and fine glass micropipettes. With these refinements, contralateral viral spread was rarely observed.
Second, and importantly, all animals included in the final analyses were subjected to post hoc anatomical verification. After completion of the experiments, CG were collected, sectioned, and examined for viral expression. As shown in Supplementary Figure 5F, only mice in which viral expression was strictly confined to the targeted CG, with no detectable infection in the contralateral ganglion, were included in the presented data.
Together, these measures ensure that the reported effects are attributable to local manipulation of the intended CG. We will ensure that the Methods section more explicitly details these technical precautions and that the legend for Figure S5F clearly states its role in validating injection specificity.
Reviewer #2 (Public review):
Summary:
The manuscript by Wang and colleagues aims to determine whether the left and right LPGi differentially regulate hepatic glucose metabolism and to reveal decussation of hepatic sympathetic nerves.
The authors used tissue clearing to identify sympathetic fibers in the liver lobes, then injected PRV into the hepatic lobes. Five days post-injection, PRV-labeled neurons in the LPGi were identified. The results indicated contralateral dominance of premotor neurons and partial innervation of more than one lobe. Then the authors activated each side of the LPGi, resulting in a greater increase in blood glucose levels after right-sided activation than after left-sided activation, as well as changes in protein expression in the liver lobes. These data suggested modulation of HGP (hepatic glucose production) in a lobe-specific manner. Chemical denervation of a particular lobe did not affect glucose levels due to compensation by the other lobes. In addition, nerve bundles decussate in the hepatic portal region.
We thank the reviewer for the thorough and constructive evaluation of our manuscript. In direct response, we will undertake comprehensive revisions to enhance the rigor and clarity of the study, including: (i) correcting ambiguous or misleading terminology pertaining to anatomical resolution and sympathetic circuit organization; (ii) expanding the Methods section with complete experimental details, improved image presentation, and explicit justification of our viral and genetic approaches; and (iii) strengthening data interpretation by addressing issues related to sparse PRV labeling, projection heterogeneity, and the functional implications of double-labeled neurons. All revisions are expected to be completed within 8 weeks.
Strengths:
The manuscript is timely and relevant. It is important to understand the sympathetic regulation of the liver and the contribution of each lobe to hepatic glucose production. The authors use state-of-the-art methodology.
Weaknesses:
(1) The wording/terminology used in the manuscript is misleading, and it is not used in the proper context. For instance, the goal of the study is "to investigate whether cerebral hemispheres differentially regulate hepatic glucose metabolism..." (see abstract); however, the authors focus on the brainstem (a single structure without hemispheres). Similarly, symmetric is not the best word for the projections.
We thank the reviewer for raising these critical points regarding terminology and conceptual framing. We acknowledge that certain phrases in our original manuscript may have been overly broad or ambiguous, particularly in describing the scope of sympathetic heterogeneity and the specificity of neural projections. Due to practical constraints and the scope of our study, our investigation is focused on the brainstem, which represents the final common pathway for these lateralized commands. We acknowledge that terms referring to the cerebral hemispheres do not accurately describe our study.
We are revising the manuscript to ensure accurate and consistent terminology and will submit the revised version with these corrections.
(2) Sparse labeling of liver-related neurons was shown in the LPGi (Figure 1). It would be ideal to have lower magnification images to show the area. Higher quality images would be necessary, as it is difficult to identify brainstem areas. The low number of labeled neurons in the LPGi after five days of inoculation is surprising. Previous findings showed extensive labeling in the ventral brainstem at four days post-inoculation (Desmoulins et al., 2025). Unfortunately, it is not possible to compare the injection paradigm/methods because the PRV inoculation is missing from the methods section. If the PRV is different from the previously published viral tracers, time-dependent studies to determine the order of neurons and the time course of infection would be necessary.
We sincerely thank the reviewer for these detailed and constructive comments regarding the PRV tracing experiments. We fully agree that careful presentation and interpretation of the anatomical data are essential for ensuring rigor and transparency. We address each point in detail below.
(1) Image magnification and anatomical context of LPGi labeling
We agree that the original images did not sufficiently convey the broader anatomical context of the LPGi. In the revised manuscript, we will replace the original panels in Figure 1 with new images that include lower-magnification overviews of the brainstem, alongside higher-magnification views of the LPGi. These images clearly delineate the LPGi with respect to established anatomical landmarks and atlas boundaries. Image contrast and resolution will also be optimized to allow unambiguous identification of PRV-labeled neurons and surrounding structures.
(2) Sparse LPGi labeling at 5 days post-injection and methodological details
We apologize for the omission of the detailed PRV injection protocol in the original Methods section. We deliberately used small-volume, focal injections (1 µL per liver lobe) to minimize viral spread and to restrict labeling to circuits specifically connected to the targeted hepatic region. Under these conditions, early-stage or intermediate-order upstream nuclei such as the LPGi are expected to exhibit relatively sparse labeling compared to more proximal autonomic nuclei. This information will add, including the PRV strain, viral titer, injection volume, precise injection coordinates, and surgical procedures.
(3) Not all LPGi cells are liver-related. Was the entire LPGi population stimulated, or was it done in a cell-type-specific manner? What was the strain, sex, and age of the mice? What was the rationale for using the particular viral constructs?
We thank the reviewer for this insightful and important question. We agree that not all neurons within the LPGi are liver-related, and we apologize that our rationale was not clearly articulated in the original manuscript.
(1) Our decision to target GABAergic neurons in the LPGi using Gad1-Cre mice was based on prior experimental evidence rather than an assumption about the entire LPGi population. In our previous study (Cell Metab. 2025;37(11):2264-2279.e10), we performed single-cell RNA sequencing on retrogradely labeled LPGi neurons following liver tracing. These analyses revealed that the majority of liver-projecting LPGi neurons are GABAergic in nature. Based on these findings, we chose to selectively manipulate GABAergic neurons in the LPGi rather than the entire LPGi neuronal population, in order to achieve greater cellular specificity and to minimize potential confounding effects arising from heterogeneous neuron types within this region. We regret that this rationale was not clearly described in the original submission and have now revised the manuscript to explicitly state this reasoning.
(2) In addition, we apologize for the omission of mouse strain, sex, and age information in the Methods section. These details will be fully added.
(3) We selected AAV-based viral vectors, specifically the AAV9 serotype, due to their well-established efficiency in transducing neurons in the brainstem, relatively low toxicity, and widespread use in circuit-level chemogenetic and optogenetic studies. When combined with Cre-dependent viral constructs in Gad1-Cre mice, this approach enabled selective and reliable manipulation of LPGi GABAergic neurons.
(4) The authors should consider the effect of stimulation of double-labeled neurons (innervating more than one lobe) and potential confounding effects regarding other physiological functions.
We thank the reviewer for raising this important point. We agree that neurons innervating more than one liver lobe could, in principle, introduce potential confounding effects and may reflect higher-order integrative autonomic neurons.
This consideration is consistent with a key finding of the cited study: the celiac-superior mesenteric ganglion (CG-SMG) contains molecularly distinct sympathetic neuron populations (e.g., RXFP1<sup>+</sup> vs. SHOX2<sup>+</sup>) that exhibit complementary organ projections and separate, non‑overlapping functions. Specifically, RXFP1<sup>+</sup> neurons innervate secretory organs (pancreas, bile duct) to regulate secretion, while SHOX2<sup>+</sup> neurons innervate the gastrointestinal tract to control motility. This functional segregation supports the concept of specialized autonomic modules rather than a uniform,“fight or flight”response, reinforcing the need for careful interpretation of circuit-specific manipulations. (Nature. 2025;637(8047):895-902; Neuron. Published online December 10, 2025).
In our PRV tracing experiments, the proportion of double-labeled neurons was relatively small, suggesting that the majority of labeled LPGi neurons preferentially associate with individual hepatic lobes. Nevertheless, we recognize that activation of this minority population could contribute to broader physiological effects beyond strictly lobe-specific regulation. We acknowledge that the absence of single-cell-level resolution in the current study limits our ability to further dissect the functional heterogeneity of these projection-defined neurons, and we will explicitly state this as a limitation in the revised manuscript. We will explicitly acknowledge this possibility in the revised manuscript and included it as a limitation of the current study. We thank the reviewer for highlighting this important conceptual consideration.
(5) The authors state that "central projections directly descend along the sympathetic chain to the celiac-superior mesenteric ganglia". What they mean is unclear. Do the authors refer to pre-ganglionic neurons or premotor neurons? How does it fit with the previous literature?
We thank the reviewer for pointing out this imprecise wording. We agree that the original phrasing was anatomically inaccurate and potentially confusing. The pathways we intended to describe involve brainstem premotor neurons that project to sympathetic preganglionic neurons in the spinal cord. These preganglionic neurons then innervate neurons in the celiac–superior mesenteric ganglia, which in turn provide postganglionic input to the liver.
We are revising the manuscript to clearly distinguish premotor from preganglionic neurons and to describe this pathway in a manner consistent with the established organization of sympathetic autonomic circuits reported in the previous literature. The revised wording will explicitly reflect this hierarchical relay structure.
(6) How was the chemical denervation completed for the individual lobes?
We thank the reviewer for raising this important methodological concern. We agree that potential diffusion of 6-OHDA is a critical issue when performing lobe-specific chemical denervation, and we apologize that our original description did not sufficiently clarify how this was controlled.
In the revised Methods section, we will provide a detailed description of the denervation procedure, including the injection volume and concentration of 6-OHDA, as well as the physical separation and isolation of individual hepatic lobes during application to minimize diffusion to adjacent tissue.
To directly assess the specificity of the chemical denervation, we included immunofluorescence and Western blot analyses demonstrating a selective reduction of sympathetic markers in the targeted lobe, with minimal effects on non-targeted lobes. These results support the effectiveness and relative spatial confinement of the 6-OHDA treatment under our experimental conditions.
We thank the reviewer for highlighting this point, which has helped us improve both the clarity and rigor of the manuscript.
(7) The Western Blot images look like they are from different blots, but there are no details provided regarding protein amount (loading) or housekeeping. What was the reason to switch beta-actin and alpha-tubulin? In Figures 3F -G, the GS expression is not a good representative image. Were chemiluminescence or fluorescence antibodies used? Were the membranes reused?
We thank the reviewer for this careful and detailed evaluation of the Western blot data. We apologize that insufficient methodological detail was provided in the original submission.
(1) We would like to clarify that the protein bands shown within each panel were derived from the same membrane. To improve transparency, we will provide full, uncropped images of the corresponding membranes in the supplementary materials. In addition, detailed information regarding protein loading amounts, gel conditions, and housekeeping controls will be added to the Methods section.
(2) The use of different loading controls (β-actin or α-tubulin) reflects a technical consideration rather than an experimental inconsistency. In our experiments, the molecular weight of the TH (62kDa) was too close to α-tubulin (55kDa), and β-actin (42kDa) was therefore used to avoid band overlap and to ensure accurate quantification.
(3) Regarding the GS signal shown in Figures 3F–G, we agree that the original representative image was suboptimal. This appears to be related to antibody performance rather than sample quality. To address this, we are repeating the GS Western blot using a newly validated antibody. The original tissue samples had been aliquoted and stored at −80 °C, allowing reliable re-analysis. This work will be done in 8 weeks.
(4) All Western blot experiments were detected using chemiluminescence, and membrane stripping and reprobing procedures are now explicitly described in the Methods section.
We thank the reviewer for highlighting these issues, which significantly improve the rigor and clarity of our data presentation.
(8) Key references using PRV for liver innervation studies are missing (Stanley et al, 2010 [PMID: 20351287]; Torres et al., 2021 [PMID: 34231420]; Desmoulins et al., 2025 [PMID: 39647176]).
We thank the reviewer for pointing out these important and highly relevant references that were inadvertently omitted in our initial submission. The studies by Stanley et al. (Proc Natl Acad Sci U S A, 2010), Torres et al. (Am J Physiol Regul Integr Comp Physiol, 2021), and Desmoulins et al. (Auton Neurosci, 2025) represent key PRV-based retrograde tracing work that has mapped central neural circuits innervating the liver and thus provide essential context for our anatomical analyses.
We agree that inclusion of these studies is necessary to properly situate our findings within the existing literature. Accordingly, we will incorporate citations to these references in the revised manuscript and discuss their relationship to our results.
Reviewer #3 (Public review):
Summary:
This study found a lobe-specific, lateralized control of hepatic glucose metabolism by the brain and provides anatomical evidence for sympathetic crossover at the porta hepatis. The findings are particularly insightful to the researchers in the field of liver metabolism, regeneration, and tumors.
Strengths:
Increasing evidence suggests spatial heterogeneity of the liver across many aspects of metabolism and regenerative capacity. The current study has provided interesting findings: neuronal innervation of the liver also shows anatomical differences across lobes. The findings could be particularly useful for understanding liver pathophysiology and treatment, such as metabolic interventions or transplantation.
Weaknesses:
Inclusion of detailed method and Discussion:
We sincerely thank the reviewer for the positive and constructive feedback, which will significantly enhance both the methodological rigor and the broader biological interpretation of our study. In direct response, we will revise the Discussion to elaborate on the potential physiological advantages of a lateralized and lobe-specific pattern of liver innervation. Furthermore, we will expand the Methods section to include a comprehensive description of the quantitative analysis applied to PRV-labeled neurons. Together, these revisions will strengthen the manuscript’s clarity, depth, and relevance to researchers in hepatic metabolism, regeneration, and disease. We expect to complete all updates within 8 weeks.
(1) The quantitative results of PRV-labeled neurons are presented, and please include the specific quantitative methods.
We thank the reviewer for this helpful suggestion. We will add a detailed description of the quantitative methods used to analyze PRV-labeled neurons in the revised Methods section. This includes information on the counting criteria, the brain regions analyzed, how the regions of interest were delineated, and the normalization procedures applied to obtain the reported neuron counts.
(2) The Discussion can be expanded to include potential biological advantages of this complex lateralized innervation pattern.
We appreciate the reviewer’s suggestion. We will expand the Discussion to include a paragraph addressing the potential biological significance of lateralized liver innervation. We highlight that this asymmetric organization could allow for more precise, lobe-specific regulation of hepatic metabolism, enable integration of distinct physiological signals, and potentially provide robustness against perturbations. These points will discuss in the revised manuscript.
Reviewer #4 (Public review):
Summary:
The studies here are highly informative in terms of anatomical tracing and sympathetic nerve function in the liver related to glucose levels, but given that they are performed in a single species, it is challenging to translated them to humans, or to determine whether these neural circuits are evolutionarily conserved. Dual-labeling anatomical studies are elegant, and the addition of chemogenetic and optogenetic studies is mechanistically informative. Denervation studies lack appropriate controls, and the role of sensory innervation in the liver is overlooked.
We sincerely appreciate the reviewer's thoughtful evaluation and fully agree that findings derived from a single-species model must be interpreted with caution in relation to human physiology. In direct response, we will revise the manuscript to explicitly clarify that all experimental data were obtained in mice and to provide a discussion of the limitations regarding direct extrapolation to humans. Concurrently, we will expand the Discussion section by integrating our findings with recent human and translational studies, including a multicenter clinical trial demonstrating that catheter-based endovascular denervation of the celiac and hepatic arteries significantly improved glycemic control in patients with poorly controlled type 2 diabetes, without major adverse events (Signal Transduct Target Ther. 2025;10(1):371). While our current work focuses on defining the anatomical organization and functional asymmetry of this circuit in mice, the clinical findings suggest that the core principles, sympathetic control of hepatic glucose metabolism via CG-liver pathways, may be conserved and of translational relevance. Additionally, we will clarify the interpretation of tyrosine hydroxylase labeling and expand the discussion of hepatic sensory and parasympathetic innervation, acknowledging their important roles in liver–brain communication and identifying them as key directions for future research. Collectively, these revisions will provide a more balanced, clinically informed, and rigorous framework for interpreting our findings, and we aim to complete all updates within 8 weeks.
Specific Weaknesses - Major:
(1) The species name should be included in the title.
We thank the reviewer for this suggestion. We agree that the species should be clearly indicated. The findings presented in this study were obtained in mice using tissue clearing and whole-organ imaging approaches. Due to technical limitations, these observations are currently limited to the mouse strain. We will update the title and clarified the species used throughout the manuscript.
(2) Tyrosine hydroxylase was used to mark sympathetic fibers in the liver, but this marker also hits a portion of sensory fibers that need to be ruled out in whole-mount imaging data
We thank the reviewer for pointing this out. We acknowledge that tyrosine hydroxylase (TH) labels not only sympathetic fibers but also a subset of sensory fibers. We will add a limitation of this point in the revised manuscript. In addition, ongoing experiments using retrograde PRV labeling from the liver, combined with sectioning, are being used to distinguish sympathetic fibers from vagal and dorsal root ganglion–derived sensory fibers. These data will be included in a forthcoming update of the manuscript and are expected to be completed in approximately 6 weeks.
(3) Chemogenetic and optogenetic data demonstrating hyperglycemia should be described in the context of prior work demonstrating liver nerve involvement in these processes. There is only a brief mention in the Discussion currently, but comparing methods and observations would be helpful.
We thank the reviewer for this suggestion. Previous studies largely relied on electrical stimulation to modulate liver innervation, which provides relatively coarse control of neural activity (Eur J Biochem. 1992;207(2):399-411). By contrast, our use of chemogenetic and optogenetic approaches allows selective, cell-type–specific manipulation of LPGi neurons. We will revise the Discussion to place our functional data in the context of prior work, highlighting how these more precise approaches improve understanding of the contribution of liver-innervating neurons to hyperglycemia.
(4) Sympathetic denervation with 6-OHDA can drive compensatory increases to tissue sensory innervation, and this should be measured in the liver denervation studies to implicate potential crosstalk, especially given the increase in LPGi cFOS that may be due to afferent nerve activity. Compensatory sympathetic drive may not be the only culprit, though it is clearly assumed to be. The sensory or parasympathetic/vagal innervation of the liver is altogether ignored in this paper and could be better described in general.
We thank the reviewer for this insightful comment and agree that chemical sympathetic denervation with 6-OHDA may induce compensatory changes in non-sympathetic hepatic inputs, including sensory and parasympathetic (vagal) innervation. As the reviewer correctly points out, increased LPGi cFOS activity may reflect afferent nerve engagement rather than solely compensatory sympathetic drive.
More broadly, we agree that the central nervous system functions as an integrated homeostatic network that continuously processes diverse afferent signals, including hepatic sensory and vagal inputs, as well as other interoceptive cues. From this perspective, the LPGi cFOS changes observed in our study likely represent one component of a complex integrative response rather than evidence for a single dominant pathway.
We acknowledge that the present study did not directly assess hepatic sensory or parasympathetic innervation, which represents a limitation in scope. In the revised manuscript, we will expand the Discussion to explicitly note this limitation and provide a more balanced consideration of potential crosstalk among sympathetic, sensory, and parasympathetic pathways in shaping LPGi activity following hepatic denervation.
Recommendations for the authors:
Reviewer #2 (Recommendations for the authors):
Although the findings are interesting, this reviewer has major concerns about the experimental design, methodology, results, and interpretation of the data. Experimental details are lacking, including basic information (age, sex, strain of mice, procedures, magnification, etc.).
We thank the reviewer for this important recommendation. We agree that comprehensive reporting of experimental details is essential for rigor and reproducibility.
In the revised manuscript, we will add complete information regarding mouse strain, sex, age, and sample size for each experiment. In addition, detailed descriptions of surgical procedures, viral constructs, injection parameters, imaging magnification, and analysis methods have been incorporated into the Methods section.
These revisions ensure that all experiments are described with sufficient technical detail and clarity to allow accurate interpretation and replication of our findings.
Reviewer #3 (Recommendations for the authors):
Addressing a few questions might help:
(1) The study found that liver-associated LPGi neurons are predominantly GABAergic. It would be informative to molecularly characterize the PRV-traced, liver-projecting LPGi neurons to determine their neurochemical phenotypes.
We thank the reviewer for this insightful suggestion. We agree that molecular characterization of liver-projecting LPGi neurons is important for understanding their functional identity.
This issue has been addressed in detail in our recent study (Cell Metab. 2025;37(11):2264-2279.e10), in which we performed single-cell RNA sequencing on retrogradely traced LPGi neurons connected to the liver. These analyses demonstrated that the majority of liver-projecting LPGi neurons are GABAergic, with a defined transcriptional profile distinct from neighboring non–liver-related populations.
Based on these findings, the current study selectively targets GABAergic LPGi neurons using Gad1-Cre mice. We are now explicitly referencing and summarizing these molecular results in the revised manuscript to clarify the neurochemical identity of the PRV-traced LPGi neurons.
(2) Is it possible to do a local microinjection of a sodium channel blocker (e.g., lidocaine) or an adrenergic receptor antagonist into the porta hepatis? That would potentially provide additional evidence for the porta hepatis as the functional crossover point.
We appreciate the reviewer’s thoughtful suggestion. While pharmacological blockade at the porta hepatis could modulate local neural activity, the proposed approach may not fully capture the distinction between ipsilateral and contralateral inputs, and may not conclusively establish neural crossover at this particular site.
In our view, the anatomical evidence provided by whole-mount tissue clearing, dual-labeled tracing, and direct visualization of decussating nerve bundles at the porta hepatis offers a more definitive demonstration of sympathetic crossover. Pharmacological blockade would affect both crossed and uncrossed fibers simultaneously and therefore would not specifically resolve the anatomical organization of this decussation.
Nevertheless, we agree that functional interrogation of the porta hepatis represents an interesting direction for future work, and we will now acknowledge this possibility in the Discussion.
(3) It is possible to investigate the effects of unilateral LPGi manipulation or ablation of one side of CG/SMG on liver metabolism, such as hyperglycemia?
We thank the reviewer for this important suggestion. We agree that unilateral ablation or silencing of the CG-SMG could provide additional insight into lateralized sympathetic control of liver metabolism.
However, precise and selective ablation of one side of the CG-SMG through 6-OHDA without affecting the contralateral ganglion or adjacent autonomic structures remains technically challenging, particularly given the anatomical connectivity between the two sides. We are currently optimizing approaches to achieve reliable unilateral manipulation.
If successful within the revision timeframe, we will include these experiments and corresponding metabolic analyses in the revised manuscript. If not, we will explicitly discuss this experimental limitation and the predicted metabolic consequences of unilateral CG-SMG ablation as an important direction for future studies. This work will be done in 6 weeks.
Reviewer #4 (Recommendations for the authors):
In the abstract and elsewhere, the use of the term 'sympathetic release' is unclear - do you mean release of nerve products, such as the neurotransmitter norepinephrine? This should be more clearly defined.
We thank the reviewer for pointing out this ambiguity. We agree that the term “sympathetic release” was imprecise. In the revised manuscript, we will explicitly refer to the release of sympathetic neurotransmitters, primarily norepinephrine, from postganglionic sympathetic fibers.
We will revise the wording throughout the manuscript to ensure accurate and consistent terminology and to avoid potential confusion regarding the underlying neurobiological mechanisms.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews
Reviewer #1 (Public review):
Summary:
In this paper, the authors investigate the effects of Miro1 on VSMC biology after injury. Using conditional knockout animals, they provide the important observation that Miro1 is required for neointima formation. They also confirm that Miro1 is expressed in human coronary arteries. Specifically, in conditions of coronary diseases, it is localized in both media and neointima, and, in atherosclerotic plaque, Miro1 is expressed in proliferating cells.
However, the role of Miro1 in VSMC in CV diseases is poorly studied, and the data available are limited; therefore, the authors decided to deepen this aspect. The evidence that Miro-/- VSMCs show impaired proliferation and an arrest in S phase is solid and further sustained by restoring Miro1 to control levels, normalizing proliferation. Miro1 also affects mitochondrial distribution, which is strikingly changed after Miro1 deletion. Both effects are associated with impaired energy metabolism due to the ability of Miro1 to participate in MICOS/MIB complex assembly, influencing mitochondrial cristae folding. Interestingly, the authors also show the interaction of Miro1 with NDUFA9, globally affecting super complex 2 assembly and complex I activity.
Finally, these important findings also apply to human cells and can be partially replicated using a pharmacological approach, proposing Miro1 as a target for vasoproliferative diseases.
Strengths:
The discovery of Miro1 relevance in neointima information is compelling, as well as the evidence in VSMC that MIRO1 loss impairs mitochondrial cristae formation, expanding observations previously obtained in embryonic fibroblasts.
The identification of MIRO1 interaction with NDUFA9 is novel and adds value to this paper. Similarly, the findings that VSMC proliferation requires mitochondrial ATP support the new idea that these cells do not rely mostly on glycolysis.
Weaknesses:
(1) Figure 3:
I appreciate the system used to assess mitochondrial distribution; however, I believe that time-lapse microscopy to evaluate mitochondrial movements in real time should be mandatory. The experimental timing is compatible with time-lapse imaging, and these experiments will provide a quantitative estimation of the distance travelled by mitochondria and the fraction of mitochondria that change position over time. I also suggest evaluating mitochondrial shape in control and MIRO1-/- VSMC to assess whether MIRO1 absence could impact mitochondrial morphology, altering fission/fusion machinery, since mitochondrial shape could differently influence the mobility.
Mitochondrial motility experiments. WT and Miro1-/- VSMCs were transiently transfected with mito-ds-red and untargeted GFP adenoviruses to fluorescently label mitochondria and cytosol, respectively. Live-cell fluorescence confocal microscopy was used to acquire mitochondrial images at one-minute intervals over a 25-30-minute period. WT cells exhibited dynamic reorganization of the mitochondrial network, whereas Miro1-/- VSMCs displayed minimal mitochondrial movement, characterized only by limited oscillatory behavior without network remodeling (Supplemental Video 1).
Mitochondrial shape (form factor) was assessed by confocal microscopy in WT and Miro1-/- VSMCs. Analysis of the mitochondrial form factor (defined as the ratio of mitochondrial length to width) during cell cycle progression revealed morphological changes in wild type (WT) cells, characterized by an increase in form factor. In contrast, Miro1-/- cells exhibited no significant alterations in mitochondrial morphology (Figure 3- Figure supplement 1B).
(2) Figure 6:
The evidence of MIRO1 ablation on cristae remodeling is solid; however, considering that the mechanism proposed to explain the finding is the modulation of MICOS/MIB complex, as shown in Figure 6D, I suggest performing EM analysis in each condition. In my mind, Miro1 KK and Miro1 TM should lead to different cristae phenotypes according to the different impact on MICOS/MIB complex assembly. Especially, Miro1 TM should mimic Miro1 -/- condition, while Miro1 KK should drive a less severe phenotype. This would supply a good correlation between Miro1, MICOS/MIB complex formation and cristae folding.
I also suggest performing supercomplex assembly and complex I activity with each plasmid to correlate MICOS/MIB complex assembly with the respiratory chain efficiency.
Complex I activity assays revealed that overexpression of MIRO1-WT fully restored enzymatic activity in MIRO1-/- cells, whereas MIRO1-KK provided partial rescue. In contrast, a MIRO1 mutant lacking the transmembrane domain failed to restore activity and resembled the Miro1-/- phenotype (Figure 6- Figure supplement 2).
The Complex I activity in each Miro1 mutant correlated with the degree of MICOS/MIB complex assembly in pulldown assays, implying a functional link between Miro1 and mitochondrial cristae organization.
Moreover, an in-gel Complex V activity assay was performed to evaluate the enzymatic activity of mitochondrial ATP synthase in a native gel following electrophoresis. To normalize the activity signal, a Blue Native PAGE of the same samples was probed for the ATP5F1 subunit. A modest, yet statistically significant reduction in Complex V activity was observed in Miro1-/- cells (Figure 6- Figure supplement 1).
(3) I noticed that none of the in vitro findings have been validated in an in vivo model. I believe this represents a significant gap that would be valuable to address. In your animal model, it should not be too complex to analyze mitochondria by electron microscopy to assess cristae morphology. Additionally, supercomplex assembly and complex I activity could be evaluated in tissue homogenates to corroborate the in vitro observations.
We appreciate the reviewer’s comment. However, our currently available samples have been processed by light microscopy and are therefore not suitable for embedding for light for electron microscopy.
(4) I find the results presented in Figure S7 somewhat unclear. The authors employ a pharmacological strategy to reduce Miro1 and validate the findings previously obtained with the genetic knockout model. They report increased mitophagy and a reduction in mitochondrial mass. However, in my opinion, these changes alone could significantly impact cellular metabolism. A lower number of mitochondria would naturally result in decreased ATP production and reduced mitochondrial respiration. This, in turn, weakens the proposed direct link between Miro1 deletion and impaired metabolic function or altered electron transport chain (ETC) activity. I believe this section would benefit from additional experiments and a more in-depth discussion.
We initially conducted experiments using the MIRO1 reducer to explore the translational potential of our findings. These experiments aimed to provide a foundation in vivo studies. However, despite multiple attempts, we were unable to demonstrate a significant effect of MIRO1reducer, delivered via a Pluronic gel, on the mitochondria of the vascular wall. Of note, he role of MIRO1 in mitophagy has been well-established in several studies (for example, PMID: 34152608), which show that genetic deletion of Miro1 delays the translocation of the E3 ubiquitin ligase Parkin onto damaged mitochondria, thereby reducing mitochondrial clearance in fibroblasts and cultured neurons. Furthermore, loss of Miro1 in the hippocampus and cortex increases mitofusin levels with the appearance of hyperfused mitochondria and activation of the integrated stress response. Thus, MIRO1 deletion in genetic models does not result in a substantial reduction of mitochondria but causes hyperfused mitochondria. The rationale for developing the MIRO1 reducer stems from genetic forms of Parkinson’s disease, where Miro1 is retained in PD cells but degraded in healthy cells following mitochondrial depolarization (PMID: 31564441). Thus, the degradation of mutant MIRO1 by the reducer does not phenocopy the effects of genetic MIRO1 depletion. Thus, we believe the data with the reducer demonstrate that MIRO1 can be acutely targeted in vitro, but the mechanism of action (as the reviewer points out, the reduction of mitochondrial mass may lead to decreased ATP levels, potentially reducing cell proliferation) differs from that of chronic genetic deletion. In fact, we observe somewhat increased mitochondrial length in MIRO1-/- cells. We acknowledge that this is complex and have revised the paragraph to clarify the use of the MIRO1 reducer.
Reviewer #2 (Public review):
Summary:
This study identifies the outer mitochondrial GTPase MIRO1 as a central regulator of vascular smooth muscle cell (VSMC) proliferation and neointima formation after carotid injury in vivo and PDGF-stimulation ex vivo. Using smooth muscle-specific knockout male mice, complementary in vitro murine and human VSMC cell models, and analyses of mitochondrial positioning, cristae architecture, and respirometry, the authors provide solid evidence that MIRO1 couples mitochondrial motility with ATP production to meet the energetic demands of the G1/S cell cycle transition. However, a component of the metabolic analyses is suboptimal and would benefit from more robust methodologies. The work is valuable because it links mitochondrial dynamics to vascular remodeling and suggests MIRO1 as a therapeutic target for vasoproliferative diseases, although whether pharmacological targeting of MIRO1 in vivo can effectively reduce neointima after carotid injury has not been explored. This paper will be of interest to those working on VSMCs and mitochondrial biology.
Strengths:
The strength of the study lies in its comprehensive approach, assessing the role of MIRO1 in VSMC proliferation in vivo, ex vivo, and importantly in human cells. The subject provides mechanistic links between MIRO1-mediated regulation of mitochondrial mobility and optimal respiratory chain function to cell cycle progression and proliferation. Finally, the findings are potentially clinically relevant given the presence of MIRO1 in human atherosclerotic plaques and the available small molecule MIRO1.
Weaknesses:
(1) There is a consistent lack of reporting across figure legends, including group sizes, n numbers, how many independent experiments were performed, or whether the data is mean +/- SD or SEM, etc. This needs to be corrected.
These data were added in the revised manuscript.
(2) The in vivo carotid injury experiments are in male mice fed a high-fat diet; this should be explicitly stated in the abstract, as it's unclear if there are any sex- or diet-dependent differences. Is VSMC proliferation/neointima formation different in chow-fed mice after carotid injury?
This is an important point, and we appreciate the feedback. In this model, the transgene is located on the Y chromosome. As a result, only male mice can be studied. However, in our previous experiments, we have not observed any sex-dependent changes in neointimal formation. Additionally, please note that smooth muscle cell proliferation in neointimal formation is enhanced in models of cholesterol-fed mice on a high-fat diet.
(3) The main body of the methods section is thin, and it's unclear why the majority of the methods are in the supplemental file. The authors should consider moving these to the main article, especially in an online-only journal.
We thank the reviewer for this suggestion. We moved the methods to the main manuscript.
(4) Certain metabolic analyses are suboptimal, including ATP concentration and Complex I activity measurements. The measurement of ATP/ADP and ATP/AMP ratios for energy charge status (luminometer or mass spectrometry), while high-resolution respirometry (Oroboros) to determine mitochondrial complex I activity in permeabilized VSMCs would be more informative.
ATP/ADP and ATP/AMP ratios were assessed on samples from WT and Miro1-/- VSMCs using an ATP/ADP/AMP Assay Kit (Cat#: A-125) purchased from Biomedical Research Service, University at Buffalo, New York). Miro1-/- samples exhibited reduced ATP levels accompanied by elevated concentrations of ADP and AMP. As a result, both ATP/ADP and ATP/AMP ratios were significantly lower in MIRO1-/- cells compared to WT, indicating impaired cellular energy homeostasis (Figure 5B, C).
(5) The statement that 'mitochondrial mobility is not required for optimal ATP production' is poorly supported. XF Seahorse analysis should be performed with nocodazole and also following MIRO1 reconstitution +/- EF hands.
To evaluate the metabolic effects of Nocodazole, we conducted Seahorse metabolic assays on vascular smooth muscle cells with various conditions (VSMCs). We used WT VSMCs, Miro1-/- VSMCs, and Miro1-/- VSMCs that expressed either MIRO1-WT, KK, or ΔTM mutants.Our results demonstrate that Nocodazole exposure did not compromise mitochondrial respiratory activity. However, Miro1-/- VSMCs displayed a trend toward reduced basal and maximal mitochondrial respiration when compared to WT cells. This deficit was only partially corrected by the expression of the MIRO1-KK mutant. In contrast, reintroducing MIRO1-WT through adenoviral delivery fully restored mitochondrial respiration to normal levels (Figure 5- Figure supplement 1).
(6) The authors should consider moving MIRO1 small molecule data into the main figures. A lot of value would be added to the study if the authors could demonstrate that therapeutic targeting of MIRO1 could prevent neointima formation in vivo.
We appreciate the reviewer's comment and attempted the suggested in vivo experiments using the commercially available Miro1 reducer. For these experiments, we used a pluronic gel to deliver the reducer to the adventitial area surrounding the carotid artery. Despite numerous attempts to optimize the experimental conditions, we were unable to reliably detect a significant effect of the reducer on mitochondria in the vascular wall.
Reviewer #3 (Public review):
Summary:
This study addresses the role of MIRO1 in vascular smooth muscle cell proliferation, proposing a link between MIRO1 loss and altered growth due to disrupted mitochondrial dynamics and function. While the findings are potentially useful for understanding the importance of mitochondrial positioning and function in this specific cell type within health and disease contexts, the evidence presented appears incomplete, with key bioenergetic and mechanistic claims lacking adequate support.
Strengths:
(1)The study focuses on an important regulatory protein, MIRO1, and its role in vascular smooth muscle cell (VSMC) proliferation, a relatively underexplored context.
(2) It explores the link between smooth muscle cell growth, mitochondrial dynamics, and bioenergetics, which is a potentially significant area for both basic and translational biology.
(3) The use of both in vivo and in vitro systems provides a potentially useful experimental framework to interrogate MIRO1 function in this context.
Weaknesses:
(1) The central claim that MIRO1 loss impairs mitochondrial bioenergetics is not convincingly demonstrated, with only modest changes in respiratory parameters and no direct evidence of functional respiratory chain deficiency.
(2) The proposed link between MIRO1 and respiratory supercomplex assembly or function is speculative, lacking mechanistic detail and supported by incomplete or inconsistent biochemical data.
(3) Key mitochondrial assays are either insufficiently controlled or poorly interpreted, undermining the strength of the conclusions regarding oxidative phosphorylation.
(4) The study does not adequately assess mitochondrial content or biogenesis, which could confound interpretations of changes in respiratory activity.
(5) Overall, the evidence for a direct impact of MIRO1 on mitochondrial respiratory function in the experimental setting is weak, and the conclusions overreach the data.
Recommendations for the authors:
Reviewer #3 (Recommendations for the authors):
(1) Throughout the manuscript, the authors incorrectly use "mobility" to describe the active transport of mitochondria. The appropriate term is "mitochondrial motility," which refers to active, motor-driven movement. "Mobility" implies passive diffusion and is not scientifically accurate in this context.
(2) "Super complex" should be consistently written as "supercomplex," in line with accepted mitochondrial biology terminology.
We thank the reviewer for this comment and revised the text accordingly.
(3) A significant limitation of the in vivo model is the mild phenotype observed, which is expected from an inducible knockout system. The authors should clarify whether a constitutive, tissue-specific knockout was considered and, if not, whether embryonic lethality or another limitation prevented its generation.
This genetic model was originally developed by Dr. Janet Shaw at the University of Utah. In the original publication, Miro1 was constitutively knocked out in neurons. Germline inactivation of Miro1 was achieved by crossing mice harboring the Miro1F allele with a mouse line expressing Cre recombinase under the control of the hypoxanthine-guanine phosphoribosyltransferase (HPRT) promoter. Mating Miro1+/− mice resulted in Miro1−/− animals, which were cyanotic and died shortly after birth. Due to this outcome, we opted to develop an inducible, smooth muscle-specific model. Additionally, we considered testing whether the acute use of an inhibitor or a knockdown system targeting Miro1 could be evaluated as a potential therapeutic approach.
(4) In Figure 1A and S1A, the authors use Western blotting to validate the knockout in the aorta and IHC in carotid arteries. The choice of different methods does not seem justified, and qPCR data are shown only for the aorta. IHC appears to be suboptimal for assessing MIRO1 levels in vascular tissue due to high autofluorescence, and IHC in Figure S1A is merely qualitative, with no quantification provided.
We present complementary approaches to validate the deletion of Miro1. For Western blot analysis, we used the aorta because it provides more material for analysis. The autofluorescence observed via immunofluorescence is characteristic of elastin fibers within the media layer, making our results typical for this technique. As shown in Figure 1- Figure supplement 1, our data demonstrate a significant decrease, if not a complete knockout, of the target protein specifically in smooth muscle cells.
(5) In Figure 1G, the bottom left panel (magnification) shows a lower green signal than the top left panel, suggesting these may have been collected with different signal intensity. This raises concerns about image consistency and representation.
Top images in Figure 1G are taken at magnification 63x. Bottom images were made at magnification 20x. The intensity is different between the two magnifications, but similar between genotypes.
(6) In Figure S3, the sampling is uncontrolled: the healthy subject and the patient differ markedly in age. The claim of colocalization is not substantiated with any quantitative analysis.
As outlined in the Methods section, our heart samples were obtained from LVAD patients or explanted hearts from transplant recipients. Due to the limited availability of such samples, there is indeed a difference in age between the healthy subject and the patient. While we acknowledge this limitation, the scarcity of samples made it challenging to control for age. Additionally, we determined that performing a quantitative analysis of colocalization would not yield robust or meaningful data given the constraints of our sample size and variability.
(7) Figure S4A lacks statistical analysis, which is necessary for interpreting the data shown.
This appears to be a misunderstanding. In this manuscript, we do present statistically significant differences and focus on those that are biologically meaningful. Specifically, we highlight differences between PDGF treatment versus no treatment within the same genotype, as well as differences between the two genotypes under the same treatment condition (control or PDGF treatment). In this particular case, there is only a statistical difference between WT+PDGF and SM-Miro1-/, but since this is not a meaningful comparison, it is not shown. Please note that this approach applies to all figures in the manuscript. Including all comparisons—whether statistically significant or not, and whether biologically meaningful or not—may appear rigorous but in our opinion, ultimately detracts from the main message of this paper.
(8) The authors state, "given the generally poor proliferation of VSMCs from SM-MIRO1-/- mice, in later experiments we used VSMCs from MIRO1fl/fl mice and infected them with adenovirus expressing cre." This is not convincing, especially since in vivo cre efficiency is generally lower than in vitro. Moreover, the methods indicate that "VSMCs from littermate controls were subjected to the same procedure with empty vector control adenovirus," yet in Figure 2A, the control appears to be MIRO1fl/fl VSMCs transduced with Ad-EV. The logic and consistency of the controls used need clarification.
For the initial experiments, cells were explanted from SM-MIRO1-/- mice (Figure 2- Figure supplement 1). In these mice, Cre recombination had occurred in vivo, and the cells exhibited very poor growth. In fact, their growth was so limited that we decided not to pursue this experimental approach after three independent experiments.
For subsequent experiments, cells were explanted from Miro1fl/fl mice and passaged several times, which allowed us to generate the number of cells required for the experiments (Figure 2B). Once sufficient Miro1fl/fl cells were obtained, they were treated with adenovirus expressing Cre, as described in the Methods section. Control cells were treated with an empty vector adenovirus. To clarify, the control cells are Miro1fl/fl cells infected with an empty vector adenovirus, while the MIRO1-/- cells are Miro1fl/fl cells infected with adenovirus expressing Cre. The statement that “littermate controls were used” is incorrect as in fact, Miro1fl/fl cells from the same preparation were either infected with an empty vector adenovirus, or with adenovirus expressing Cre. As mentioned, the knockdown was confirmed by Western blotting.
(9) Figure 2C shows a growth delay in MIRO1-/- cells. Have the authors performed additional time points to determine when these cells return to G1 and quantify the duration of the lag?
This is an excellent suggestion. So far, we have not performed this experiment.
(10) In the 24 h time point of Figure 2C, MIRO1-/- cells appear to be cycling, yet no cyclin E signal is detected. How do the authors explain this inconsistency? Additionally, in Figure 2H, the quantification of cyclin E is unreliable, given that lanes 3 and 4 show no detectable signal.
We agree with the reviewer—the inconsistency is driven by the exposure of the immunoblot presented. We revisited the data, reviewed the quantification, and performed an additional experiment. We are now presenting an exposure that demonstrates levels of cyclin E (Figure 2G).
(11) In Figure 3D, the authors present mitochondrial probability map vs. distance from center curves. How was the "center" defined in this analysis? Were radial distances normalized across cells (e.g., to the cell radius or maximum extent)? If not, variation in cell and/or nucleus size or shape could significantly affect the resulting profiles. No statistical analysis is provided for this assessment, which undermines its quantitative value. Furthermore, the rationale behind the use of mito95 values is not clearly explained.
The center refers to the center of the microchip's Y-shaped pattern, to which each cell is attached. Since all Y-shapes on the chip are identical in size, normalization is not required. The size of the optimal Y-shapes was tested as recommended by CYTOO. For further context, please refer to the papers by the Kittler group.
Additionally, a graph demonstrating the percentage of mitochondria localized at specific distances can be produced for any given distance. Notably, the further from the center of the chip, the more pronounced the differences become.
(12) The authors apply a 72 h oligomycin treatment to assess proliferation and a 16 h treatment to measure ATP levels. This discrepancy in experimental design is not justified in the manuscript. The length of treatment directly impacts the interpretation of the data in Figures 4C, 4D, and 4E, and needs to be addressed.
Thank you for this comment. We have performed additional experiments to align these time points. In the revised manuscript, we now present proliferation and ATP production measured at the same time point (Figure 4A, B for proliferation and ATP levels).
(13) The manuscript repeatedly suggests that MIRO1 loss causes a defect in mitochondrial ATP production, yet no direct demonstration of a bioenergetic defect is provided. The claim relies on a modest decrease in supercomplex species (of undefined composition) and a mild reduction in complex I activity that does not support a substantial OXPHOS defect. Notably, the respirometry data in Figure 5I do not align with the BN-PAGE results in Figure 6I. There is increasing evidence that respiratory chain supercomplexes do not confer a catalytic advantage. The authors should directly assess the enzymatic activities of all respiratory complexes. Reported complex I activity in MIRO1-/- cells appears rotenone-like (virtually zero, figure 3K) or ~30% residual (Figure 3L), suggesting a near-total loss of functional complex I, which is not reflected in the BN-PAGE. Additionally, complex I activity has not been normalized to a mitochondrial reference, such as citrate synthase.
Given that we work in primary cells and are limited by the number of cells we can generate, we concentrated on ETC1 and 5 and performed experiments in cells after expression of MIRO1 WT and MIRO1 mutants (Figure 6- Figure supplement 1). Please note that the addition of Rotenone abolishes the slope of NADH consumptions (Figure 6- Figure supplement 2F).
While the ETC1 activity is measured in Fig. 6K, the blue native gel shown in Figure 6I is performed without substrate and thus, indicative of protein complex abundance rather than complex activity.
In additional experiments, we normalized the activity to citrate synthase as requested.
(14) In the methods section, the complex I activity assay is incorrectly described: complex I is a NADH dehydrogenase, so the assay measures NADH oxidation, not NADPH.
We thank the reviewer for his comment and revised the manuscript accordingly.
(15) The authors have not assessed mitochondrial mass, which is a critical omission. Differences in mitochondrial biogenesis or content could underlie several observed phenotypes and should be controlled for.
A qPCR assay was used to assess mitochondrial DNA copy number in WT and Miro1-/- VSMCs. We determined the abundance of COX1 and MT-RNR1 DNA as mitochondrial gene targets and NDUFV DNA as the nuclear reference gene. While the results in Miro1-/- cells were highly variable, no statistically significant reduction of copy numbers was detected (Figure 3- Figure supplement 1B).
(16) Complex IV signal is missing in Figure 6I. Its omission is not acknowledged or explained.
Thank you for this comment. We believe this is due to a technical issue. Complex IV can be challenging to detect consistently, as its visibility is highly dependent on sample preparation conditions. In this specific case, we suspect that the buffer used during the isolation process may have influenced the detection of Complex IV.
(17) Figure 6D does not appear representative of the quantifications shown. C-MYC signal is visibly reduced in the mutant, consistent with the lower levels of interactors such as Sam50 and NDUFA9. Additionally, the SDHA band is aligned at the bottom of the blot box. The list of antibodies used, and their catalog number is missing, or it was not provided to the reviewers. It seems plausible that the authors used a cocktail antibody set (e.g., Abcam ab110412), which includes anti-NDUFA9. This would contradict the claim of reduced complex I and SC levels, as the steady-state levels of NDUFA9 appear unchanged.
We acknowledge that the expression of the myc-MIRO1 mutant is lower compared to myc-MIRO1 WT or myc-MIRO1 KK. Achieving identical expression levels when overexpressing multiple MIRO1 constructs is challenging. We agree that the lower expression of this mutant contributes to a reduced pull-down. Our quantification shows a reduction in association, although it is not statistically significant.
A list of the antibodies was provided in the Methods section.
We would like to clarify that we did not use an antibody cocktail in our experiments.
(18) The title of Figure 6, "Loss of Miro1 leads to dysregulation of ETC activity under growth conditions," is vague. The term "dysregulation" should be replaced with a more specific mechanistic descriptor-what specific regulatory defect is meant?
We thank the reviewer for this suggestion and rephrased the title.
(19) In the results text for Figure 6, the authors state: "These data demonstrate that MIRO1 associates with MIB/MICOS and that this interaction promotes the formation of mitochondrial super complexes and the activity of ETC complex I." This conclusion is speculative and not mechanistically supported by the data presented.
We appreciate the reviewer's feedback. We have revised the text to clarify the relationship between MIRO1, MIB/MICOS, supercomplex formation, and ETC activity. The updated text now states: "These data demonstrate that MIRO1 associates with MIB/MICOS. Additionally, MIRO1 promotes the formation of mitochondrial supercomplexes and enhances the activity of ETC complex I.”
(20) In Figure 7A, it is unclear what the 3x siControl/siMiro1 pairs represent-are these different cell lines or technical replicates of the same line? No loading control is shown. If changes in mitochondrial protein abundance are being evaluated, using COX4 as a loading control is inappropriate. The uneven COX4 signal across samples further complicates interpretation
Please note that we used primary cells, not cell lines. The three siControl/siMiro1 pairs represent independent cell isolations, each transfected with either siControl or. siMIRO1 mRNA. While the possibility of a difference in mitochondrial mass is an interesting question, the primary objective of this experiment is to demonstrate that the technique effectively results in the knockdown of Miro1, which is exclusively localized to mitochondria and not present in the cytosol. As such, we believe that Cox4 serves as a reasonable loading control. Although Miro1 knockdown may lead to a reduction in mitochondrial mass, the focus of this experiment is not to assess mitochondrial mass but to confirm the reduction in Miro1 protein levels on mitochondria. We also performed anti-VDAC immunoblots on the same membranes as alternative loading control (Author response image 1).
Author response image 1.
(21) Figure 7G is difficult to interpret. Why did the authors choose to use a sensor-based method instead of the chemiluminescent assay to measure ATP in these samples?
Both methods were employed to assess ATP levels in human samples. ATP measurements obtained with luminescent assay are provided.
Loops
The use of loops and conditionals in social media bots highlights a major ethical challenge regarding scale and context. While a human can manually block a few trolls, a bot using a loop can automatically block hundreds of users in seconds based on a simple conditional like an iphone or android tag. This demonstrates how automation doesn't just make tasks easier but changes the power dynamics of online interaction by allowing a single user to exert massive influence without manual effort.
Joint Public Review:
Quite obviously, the brain encodes "time", as we are able to tell if something happened before or after something else. How this is done, however, remains essentially not understood. In the context of Working Memory tasks, many experiments have shown that the neural activity during the retention period "encodes" time, besides the stimulus to be remembered; that is, the time elapsed from stimulus presentation can be reliably inferred from the recordings, even if time per se is not important for the task. This implies 'mixed selectivity', in the weak sense of neural activity varying with both stimulus identity and time elapsed (since presentation).
In this paper, the authors investigate the implications of a specific form of such mixed selectivity, that is, conjunctive coding of what (stimulus) and when (time) at the single-neuron level, on the resulting dynamics of the population activity when 'viewed' through linear dimensionality-reduction techniques, essentially Principal Component Analysis (PCA). The theoretical/modeling results presented provide a useful guide to the interpretation of the experimental results; in particular, with respect to what can, or cannot, be rightfully inferred from those experimental results (using PCA-like techniques). The results are essentially theoretical in nature; there are, however, some conclusions that require a more precise justification, in my opinion. More generally, as the authors themselves discuss in the paper, it is not clear how to generalize this coding scheme to more complicated, but behaviorally and cognitively relevant, situations, such as multi-item WM or WM for sequences.
(1) It is unclear to me how the conjunctive code that the authors use (i.e., Equation (3)) is constrained by the theoretical desiderata (i.e., compositionality) they list, or whether it is simply an ansatz, partly motivated by theoretical considerations and experimental observations.
The "what" part: What the authors mean by "relationships" between stimuli is never clearly defined. From their argument (and from Figure 1b), it would seem that what they mean is "angles" between population vectors for all pairs of stimuli. If this is so, then the effect of the passing time can only amount to a uniform rescaling of the components of the population vector (i.e., it must be a similarity transformation; rotations are excluded, if the linear-decoder vectors are to be time-independent); the scaling factor, then, must be a strictly monotonous function of time (increasing or decreasing), if one is to decode time. In other words, the "when" receptive fields must be the same for all neurons.
The "when" part: The condition, \tau_3=\tau_1+\tau_2, does not appear to be used at all. In fact, it is unclear (to me at least) whether the model, as it is formulated, is able to represent time intervals between stimuli.
(2) For the specific case considered, i.e., conjunctive coding, it would seem that one should be able to analytically work out the demixed PCA (see Kobak et al., 2016). More generally, it seems interesting to compare the results of the PCA and the demixed PCA in this specific case, even just using synthetic data.
(3) In the Section "Dimensionality of neural trajectories...", there is some claim about how the dimensionality of the population activity goes up with the observation window T, backed up by numerical results that somehow mimic the results of Cueva et al. (2020) on experimental data. Is this a result that can be formally derived? Related to this point, it would be useful to provide a little more justification for Equation (17). Naively, one would think that the correlation matrix of the temporal component is always full-rank nominally, but that one can get excellent low-rank approximations (depending on T, following your argument).
eLife Assessment
The authors provide a scholarly review of intracranial research into the neural correlates of consciousness (NCCs). To our knowledge, this is the first such review, and it therefore may become a must-read for anyone working in the field of consciousness research. It is not so persuasive that intracranial recordings are better suited to identifying pure NCCs than other methods, which appears a problem instead solved through novel paradigms and better-developed theories - but this no doubt reflects an in-depth, timely, and insightful contribution to the literature.
Reviewer #1 (Public review):
Summary
In this review paper, the authors describe the concept of neural correlates of consciousness (NCC) and explain how noninvasive neuroimaging methods fall short of being able to properly characterise an unconfounded NCC. They argue that intracranial research is a means to address this gap and provide a review of many intracranial neuroimaging studies that have sought to answer questions regarding the neural basis of perceptual consciousness.
Strengths
The authors have provided an in-depth, timely, and scholarly contribution to the study of NCCs. First and foremost, the review surveys a vast array of literature. The authors synthesise findings such that a coherent narrative of what invasive electrophysiology studies have revealed about the neural basis of consciousness can be easily grasped by the reader. The review is also, to the best of my knowledge, the first review to specifically target intracranial approaches to consciousness and to describe their results in a single article. This is a credit to the authors, as it becomes ever harder to apply strict tests to theories of consciousness using methods such as fMRI and M/EEG it is important to have informative resources describing the results of human intracranial research so that theorists will have to constrain their theories further in accordance with such data. As far as the authors were aiming to provide a complete and coherent overview of intracranial approaches to the study of NCCs, I believe they have achieved their aim.
Weaknesses
Overall, I feel positive about this paper. However, there are a couple of aspects to the manuscript that I think could be improved.
(1) Distinguishing NCCs from their prerequisites or consequences
This section in the introduction was particularly confusing to me. Namely, in this section, the authors' aim is to explain how intracranial recordings can help distinguish 'pure' NCCs from their antecedents and consequences. However, the authors almost exclusively describe different tasks (e.g., no-report tasks) that have been used to help solve this problem, rather than elaborating on how intracranial recordings may resolve this issue. The authors claim that no-report designs rely on null findings, and invasive recordings can be more sensitive to smaller effects, which can help in such cases. However, this motivation pertains to the previous sub-section (limits of noninvasive methods), since it is primarily concerned with the lack of temporal and spatial resolution of fMRI and M/EEG. It is not, in and of itself, a means to distinguish NCCs from their confounds.
As such, in its current formulation, I do not find the argument that intracranial recordings are better suited to identifying pure NCCs (i.e. separating them from pre- or post-processing) convincing. To me, this is a problem solved through novel paradigms and better-developed theories. As it stands, the paper justifies my position by highlighting task developments that help to distinguish NCCs from prerequisites and consequences, rather than giving a novel argument as to why intracranial recordings outperform noninvasive methods beyond the reasons they explained in the previous section. Again, this position is justified when, from lines 505-506, the authors describe how none of the reported single-cell studies were able to dissociate NCCs from post-perceptual processing. As such, it seems as if, even with intracranial recording, NCCs and their confounds cannot be disentangled without appropriate tasks.
The section 'Towards Better Behavioural Paradigms' is a clear attempt to address these issues and, as such, I am sure the authors share the same concerns as I am raising. Still, I remain unconvinced that the distinguishing of NCCs from pre-/post- processing is a fair motivation for using intracranial over noninvasive measures.
(2) Drawing misleading conclusions from certain studies
There are passages of the manuscript where the authors draw conclusions from studies that are not necessarily warranted by the studies they cite. For instance:
Lines 265 - 271: "The results of these two studies revealed a complex pattern: on the one hand, HGA in the lateral occipitotemporal cortex and the ventral visual cortex correlated with stimulus strength. On the other hand, it also correlated with another factor that does not appear to play a role in visibility (repetition suppression), and did not correlate with a non-sensory factor that affects visibility reports (prior exposure). These results suggest that activity in occipitotemporal cortex regions reflecting higher-order visual processing may be a precursor to the NCC but not an NCC proper."
It's possible to imagine a theory that would predict HGA could correlate with stimulus strength and repetition suppression, or that it would not correlate with prior exposure (e.g. prior exposure could impact response bias without affecting subjective visibility itself). The authors describe this exact ambiguity in interpretation later in the article (line 664), but in its current form, at least in line 270 (when the study is most extensively discussed), the manuscript heavily implies that HGA is not an NCC proper. This generates a false impression that intracranial recordings have conclusively determined that occipitotemporal HGA is not a pure NCC, which is certainly a premature conclusion.
Line 243: "Altogether, these early human intracranial studies indicate that early-latency visual processing steps, reflected in broadband and low gamma activity, occur irrespective of whether a stimulus is consciously perceived or not. They also identified a candidate NCC: later (>200 ms) activity in the occipitotemporal region responsible for higher-order visual processing."
The authors claim in this section that later (>200ms) activity in occipitotemporal regions may be a candidate for an NCC. However, the Fisch et al. (2009) study they describe in support of this conclusion found that early (~150ms) activity could dissociate conscious and unconscious processing. This would suggest that it is early processing that lays claim to perceptual consciousness. The authors explicitly describe the Fisch et al results as showing evidence for early markers of consciousness (line 240: '...exhibited an early...response following recognized vs unrecognised stimuli.) Yet only a few lines later they use this to support the conclusion that a candidate NCC is 'later (>200ms) activity in the occipitotemporal region' (line 245). As such, I am not sure what conclusion the authors want me to make from these studies.
This problem is repeated in lines 386-387: "Altogether, studies that investigated the cortical correlates of visual consciousness point to a role of neural responses starting ~250 ms after stimulus onset in the non-primary visual cortex and prefrontal cortex."
This seems to be directly in conflict with the Fisch et al results, which show that correlates of consciousness can begin ~100ms earlier than the authors state in this passage.
(3) Justifying single-neuron cortical correlates of consciousness
The purpose of the present manuscript is to highlight why and how intracortical measures of neural activity can help reveal the neural correlates of perceptual consciousness. As such, in the section 'Single-neuron cortical correlates of perceptual consciousness', I think the paper is lacking an argument as to why single-neuron research is useful when searching for the NCC. Most theories of consciousness are based around circuit or system-level analyses (e.g., global ignition, recurrent feedback, prefrontal indexing, etc.) and usually do not make predictions about single cells. Without any elaboration or argument as to why single-cell research is necessary for a science of consciousness, the research described in this section, although excellent and valuable in its own right, seems out of place in the broader discussion of NCCs. A particularly strong interpretation here could be that intracranial recordings mislead researchers into studying single cells simply because it is the finest level of analysis, rather than because it offers helpful insight into the NCCs.
(4) No mention of combined fMRI-EEG research
A minor point, but I was surprised that the authors did not mention any combined fMRI-EEG research when they were discussing the limits of noninvasive recordings. Intracortical recordings are one way to surpass the spatial and temporal resolution limits of M/EEG and fMRI respectively, but studies that combine fMRI and EEG are also an alternative means to solve this problem: by combining the spatial resolution of fMRI with the temporal resolution of EEG, researchers can - in theory - compare when and where certain activity patterns (be they univariate ERPs or multivariate patterns) arise. The authors do cite one paper (Dellert et al., 2021 JNeuro) that used this kind of setup, but they discuss it only with respect to the task and ignore the recording method. The argument for using intracranial recordings is weaker for not mentioning a viable, noninvasive alternative that resolves the same issues.
Reviewer #2 (Public review):
Summary:
In this work, the authors review the study of the neural correlates of consciousness (NCCs). They discuss several of the difficulties that researchers must face when studying NCCs, and argue that several of these difficulties can be alleviated by using intracranial recordings in humans.
They describe what constitutes an NCC, and the difficulties to distinguish between an NCC proper from the prerequisites and consequences of conscious processing.
They also describe the two main types of experimental designs used to study NCCs. These are the contrastive approach (with its report and non-report variants), and the supraliminal approach, each with its own merits and pitfalls.
They discuss the limitations of non-invasive methods, such as fMRI, EEG and MEG, as well as the limitations of the use of invasive recordings in non-human animals.
After setting the stage in this way, the authors provide an extensive review of the knowledge acquired by using invasive recordings in humans. This included population-level measurements in vision and in other sensory modalities, as well as single-neuron level studies. The authors also discuss studies of subcortical NCCs.
The second half of this work discusses the theoretical insights gained through the use of intracranial recordings, as well as their limitations, and a perspective for future work.
Strengths:
This work offers an impressive review, which will serve as a useful reference document, both for newcomers to the study of NCC and for experienced researchers. The inclusion of non-visual and subcortical NCCs is of particular merit, as these have been understudied.
Besides serving as a review, this work includes a perspective, exploring several directions to pursue for the progress of the field.
Weaknesses:
The intention of the authors is to argue how some of the problems faced when studying NCCs are alleviated by the use of intracranial recordings in humans. But in some cases, the link between the problems related to the study of NCCs and the advantages of intracranial recordings over non-invasive methods is not clear.
For example, the authors explain the difficulties in distinguishing between true NCCs from their prerequisites and consequences. This constitutes a difficult conceptual problems that plague all recording techniques. The authors don't provide a convincing explanation of how intracranial recordings offer advantages over EEG or MEG when dealing with these problems.
For example, the authors explain how the use of non-report designs to rule out post-perceptual processing relies on null results, which, according to them, are harder to interpret given the low resolution of non-invasive methods. But the interpretation of null results is actually more complicated in the case of intracranial recordings. As the coverage achieved by the electrodes is sparse, if a null result is attested, it remains possible that a true effect was present in a nearby patch of cortex out of coverage.
The authors argue that the spatial resolution of intracranial recordings is better than that of EEG and MEG. While this is technically true (especially compared to EEG), the true spatial scale of the NCCs is unknown. If NCCs' span is in the mm range, then the additional spatial resolution of intracranial recordings might not be an advantage.
Another factor that should be taken into consideration when assessing the spatial resolution of intracranial recordings is that while the listening zone of individual intracranial contacts is small, coverage is sparse and defined by clinical criteria (something that the authors discuss). In practice, the activity recorded by contacts is usually attributed to anatomically defined ROIs with a scale in the cm range. Given the sparse and uneven (across regions and patients) coverage afforded by intracranial recordings, the advantage of intracranial recordings in terms of spatial resolution is overstated.
Appraisal of whether the authors achieved their aims:
In this work, the authors have gathered an impressive review and have discussed several important problems in the field of study of NCCs, as well as provided a perspective on how the field could move forward.
What is less clear is how the use of intracranial recordings per se holds potential to overcome problems such as the distinction between true NCCs and the prerequisites and consequences of conscious processing.
Discussion of the likely impact of the work on the field:
This work has the potential of becoming a must-read for anyone working in the field of consciousness research.
on International Women’s Day, the bot automatically finds when any of those companies make an official tweet celebrating International Women’s Day and it quote tweets it with the pay gap at that company:
This is a fascinating idea. I think this is a distillation of the ideal of that fact-checking feature that Meta tried and then abandoned, or the community notes feature on Twitter currently. The ability to add context in real time to an issue which is politically multi-faceted (such as corporations both paying lip service to international women's day while not eradicating the structures of misogyny and patriarchy in their own companies) is something social media ought to be doing, but doesn't. I wonder what factors led to more social media companies not attempting to add or push these features to their platforms. Is it profit? Is it their own political beliefs? Or is it simply not feasible?
But Kurt Skelton was an actual human (in spite of the well done video claiming he was fake). He was just trolling his audience. Professor Casey Fiesler [c16] talked about it on her TikTok channel:
This actually got me. When I watched the first video and it was "revealed" that the creator was "not real," I fully believed it because I've definitely been tricked by AI videos before. After watching the second video by Professor Casey Fiesler, I was genuinely shocked, both because I fell for it so fast and because I felt double tricked. I might've believed it as fast as I did because I've never seen any of Kurt Skelton's videos before, so it was easy for me to imagine that he might've just been curated by another creator. Maybe had I seen that creator's videos prior to that video, I would've been more skeptical since I would've seen a longer history of the creator and his personality. Definitely a shocker to me either way though!
“human computers”
I think this concept of a human computer is very thought provoking in a myriad of ways. My own research pertains to how identity is being reshaped along the boundaries of recommendation algorithms, and I feel like this idea of a human computer has a lot of thematic overlap. I know that the first "computers" were people who manually did computation as their job, but to what extent did their status as essentially beings of code shape their lives? Did they (or do they, in the case of modern human computers) see the world differently than I? What do they notice? Does living so in tune with the digital shape their philosophical framework? I feel like the fact that so much of culture and my own time is mediated by retention-based algorithms drastically shapes my ability to imagine another world different from my own. Does the same hold true for human computers?
Reviewer #1 (Public review):
Summary:
In this study, the authors trained rats on a "figure 8" go/no-go odor discrimination task. Six odor cues (3 rewarded and 3 non-rewarded) were presented in a fixed temporal order and arranged into two alternating sequences that partially overlap (Sequence #1: 5⁺-0⁻-1⁻-2⁺; Sequence #2: 3⁺-0⁻-1⁻-4⁺) --forming an abstract figure-8 structure of looping odor cues.
This task is particularly well-suited for probing representations of hidden states, defined here as the animal's position within the task structure beyond superficial sensory features. Although the task can be solved without explicit sequence tracking, it affords the opportunity to generalize across functionally equivalent trials (or "positions") in different sequences, allowing the authors to examine how OFC representations collapse across latent task structure.
Rats were first trained to criterion on the task and then underwent 15 days of self-administration of either intravenous cocaine (3 h/day) or sucrose. Following self-administration, electrodes were implanted in lateral OFC, and single-unit activity was recorded while rats performed the figure-8 task.
Across a series of complementary analyses, the authors report several notable findings. In control animals, lOFC neurons exhibit representational compression across corresponding positions in the two sequences. This compression is observed not only in trial/positions involving overlapping odor (e.g., Position 3 = odor 1 in sequence 1 vs sequence 2), but also in trials/positions involving distinct, sequence-specific odors (e.g., Position 4: odor 2 vs odor 4) --indicating generalization across functionally equivalent task states. Ensemble decoding confirms that sequence identity is weakly decodable at these positions, consistent with the idea that OFC representations collapse incidental differences in sensory information into a common latent or hidden state representation. In contrast, cocaine-experienced rats show persistently stronger differentiation between sequences, including at overlapping odor positions.
Strengths:
Elegant behavioral design that affords the detection of hidden-state representations.
Sophisticated and complementary analytical approaches (single-unit activity, population decoding, and tensor component analysis).
Weaknesses:
The number of subjects is small --can't fully rule out idiosyncratic, animal-specific effects.
Comments
(1) Emergence of sequence-dependent OFC representations across learning.
A conceptual point that would benefit from further discussion concerns the emergence of sequence-dependent OFC activity at overlapping positions (e.g., position P3, odor 1). This implies knowledge of the broader task structure. Such representations are presumably absent early in learning, before rats have learned the sequence structure. While recordings were conducted only after rats were well trained, it would be informative if the authors could comment on how they envision these representations developing over learning. For example, does sequence differentiation initially emerge as animals learn the overall task structure, followed by progressive compression once animals learn that certain states are functionally equivalent? Clarifying this learning-stage interpretation would strengthen the theoretical framing of the results.
(2) Reference to the 24-odor position task
The reference to the previously published 24-odor position task is not well integrated into the current manuscript. Given that this task has already been published and is not central to the main analyses presented here, the authors may wish to a) better motivate its relevance to the current study or b) consider removing this supplemental figure entirely to maintain focus.
(3) Missing behavioral comparison
Line 117: the authors state that absolute differences between sequences differ between cocaine and sucrose groups across all three behavioral measures. However, Figure 1 includes only two corresponding comparisons (Fig. 1I-J). Please add the third measure (% correct) to Figure 1, and arrange these panels in an order consistent with Figure 1F-H (% correct, reaction time, poke latency).
(4) Description of the TCA component
Line 220: authors wrote that the first TCA component exhibits low amplitude at positions P1 and P4 and high amplitude at positions P2 and P3. However, Figure 3 appears to show the opposite pattern (higher magnitude at P1 and P4 and lower magnitude at P2 and P3). Please check and clarify this apparent discrepancy. Alternatively, a clearer explanation of how to interpret the temporal dynamics and scaling of this component in the figure would help readers correctly understand the result.
(5) Sucrose control<br /> Sucrose self-administration is a reasonable control for instrumental experience and reward exposure, but it means that this group also acquired an additional task involving the same reinforcer. This experience may itself influence OFC representations and could contribute to the generalization observed in control animals. A brief discussion of this possibility would help contextualize the interpretation of cocaine-related effects.
(6) Acknowledge low N
The number of rats per group is relatively low. Although the effects appear consistent across animals within each group, this sample size does not fully rule out idiosyncratic, animal-specific effects. This limitation should be explicitly acknowledged in the manuscript.
(7) Figure 3E-F: The task positions here are ordered differently (P1, P4, P2, P3) than elsewhere in the paper. Please reorder them to match the rest of the paper.
Reviewer #2 (Public review):
In the current study, the authors use an odor-guided sequence learning task described as a "figure 8" task to probe neuronal differences in latent state encoding within the orbitofrontal cortex after cocaine (n = 3) vs sucrose (n = 3) self-administration. The task uses six unique odors which are divided into two sequences that run in series. For both sequences, the 2nd and 3rd odors are the same and predict reward is not available at the reward port. The 1st and 4th odors are unique, and are followed by reward. Animals are well-trained before undergoing electrode implant and catheterization, and then retrained for two weeks prior to recording. The hypothesis under test is that cocaine-experienced animals will be less able to use the latent task structure to perform the task, and instead encode information about each unique sequence that is largely irrelevant. Behaviorally, both cocaine and sucrose-experienced rats show high levels of accuracy on task, with some group differences noted. When comparing reaction times and poke latencies between sequences, more variability was observed in the cocaine-treated group, implying animals treated these sequences somewhat differently. Analyses done at the single unit and ensemble level suggests that cocaine self-administration had increased the encoding of sequence-specific information, but decreased generalization across sequences. For example, the ability to decode odor position and sequence from neuronal firing in cocaine-treated animals was greater than controls. This pattern resembles that observed within the OFC of animals that had fewer training sessions. The authors then conducted tensor component analysis (TCA) to enable a more "hypothesis agnostic" evaluation of their data.
Overall, the paper is well written and the authors do a good job of explaining quite complicated analyses so that the reader can follow their reasoning. I have the following comments.
While well-written, the introduction mainly summarises the experimental design and results, rather than providing a summary of relevant literature that informed the experimental design. More details regarding the published effects of cocaine self-administration on OFC firing, and on tests of behavioral flexibility across species, would ground the paper more thoroughly in the literature and explain the need for the current experiment.
For Fig 1F, it is hard to see the magnitude of the group difference with the graph showing 0-100%- can the y axis be adjusted to make this difference more obvious? It looks like the cocaine-treated animals were more accurate at P3- is that right?<br /> The concluding section is quite brief. The authors suggest that the failure to generalize across sequences observed in the current study could explain why people who are addicted to cocaine do not use information learned e.g. in classrooms or treatment programs to curtail their drug use. They do not acknowledge the limitations of their study e.g. use of male rats exclusively, or discuss alternative explanations of their data.
Is it a problem that neuronal encoding of the "positions" i.e. the specific odors was at or near chance throughout in controls? Could they be using a simpler strategy based on the fact that two successive trials are rewarded, then two successive trials are not rewarded, such that the odors are irrelevant?
When looking at the RT and poke latency graphs, it seems the cocaine-experienced rats were faster to respond to rewarded odors, and also faster to poke after P3. Does this mean they were more motivated by the reward?
Reviewer #2 (Public review):
Summary:
This study provides evidence for the integration-segregation theory of an attentional effect, widely cited as inhibition of return (IOR), from a neuroimaging perspective, and explores neural interactions between IOR and cognitive conflict, showing that conflict processing is potentially modulated by attentional orienting.
Strengths:
The integration-segregation theory was examined in a sophisticated experimental task that also accounted for cognitive conflict processing, which is phenomenologically related to IOR but "non-spatial" by nature. This study was carefully designed and executed. The behavioral and neuroimaging data were carefully analyzed and largely well presented.
Weaknesses:
The rationale for the experimental design was not clearly explained in the manuscript; more specifically, why the current ER-fMRI study would disentangle integration and segregation processes was not explained. The introduction of "cognitive conflict" into the present study was not well reasoned for a non-expert reader to follow.
The presentation of the results can be further improved, especially the neuroimaging results. For instance, Figure 4 is challenging to interpret. If "deactivation" (or a reduction in activation) is regarded as a neural signature of IOR, this should be clearly stated in the manuscript.
Reviewer #3 (Public review):
Summary:
This study aims to provide the first direct neuroimaging evidence relevant to the integration-segregation theory of exogenous attention - a framework that has shaped behavioral research for more than two decades but has lacked clear neural validation. By combining an inhibition-of-return (IOR) paradigm with a modified Stroop task in an optimized event-related fMRI design, the authors examine how attentional integration and segregation processes are implemented at the neural level and how these processes interact with semantic and response conflicts. The central goal is to map the distinct neural substrates associated with integration and segregation and to clarify how IOR influences conflict processing in the brain.
Strengths:
The study is well-motivated, addressing a theoretically important gap in the attention literature by directly testing a long-standing behavioral framework with neuroimaging methods. The experimental approach is creative: integrating IOR with a Stroop manipulation expands the theoretical relevance of the paradigm, and the use of a genetic-algorithm-optimized fMRI design ensures high efficiency. Methodologically, the study is sound, with rigorous preprocessing, appropriate modeling, and analyses that converge across multiple contrasts. The results are theoretically coherent, demonstrating plausible dissociations between integration-related activity in the fronto-parietal attention network (FEF, IPS, TPJ, dACC) and segregation-related activity in medial temporal regions (PHG, STG). The findings advance the field by supplying much-needed neural evidence for the integration-segregation framework and by clarifying how IOR modulates conflict processing.
Weaknesses:
Some interpretive aspects would benefit from clarification, particularly regarding the dual roles ascribed to dACC activation and the circumstances under which PHG and STG are treated as a single versus separate functional clusters. Reporting conventions are occasionally inconsistent (e.g., statistical formatting, abbreviation definitions), which may hinder readability. More detailed reporting of sample characteristics, exclusion criteria, and data-quality metrics-especially regarding the global-variance threshold-would improve transparency and reproducibility. Finally, some limitations of the study, including potential constraints on generalization, are not explicitly acknowledged and should be articulated to provide a more balanced interpretation.
Reviewer #1 (Public review):
Summary:
Fahdan et al. present a study investigating the molecular programs underlying axon initial growth and regrowth in Drosophila mushroom body (MB) neurons. The authors leverage the fact that different Kenyon cell (KC) subtypes undergo distinct axonal events on the same developmental timeline: γ KCs prune and then regrow their axons during early pupation, whereas α/β KCs extend their axons for the first time during the same pupal period. Using bulk Smart-seq2 RNA sequencing across six developmental time points, the authors identify genes enriched during γ KC regrowth and α/β KC initial outgrowth, and subsequently perform an RNAi screen to determine which candidates are functionally required for these processes.
Among these, they focus on Pmvk, a key enzyme in the mevalonate pathway. Both RNAi knockdown and a CRISPR-generated mutant produce strong γ KC regrowth defects. Knockdown of other mevalonate pathway components (Hmgcr, Mvk) partially recapitulates this phenotype. The authors propose that Pmvk promotes axonal regrowth through effects on the TOR pathway.
Overall, this work identifies new molecular players in developmental axon remodeling and provides intriguing evidence connecting Pmvk to γ KC regrowth.
While the Pmvk knockdown and loss-of-function data are compelling, the evidence that the mevalonate pathway broadly regulates γ KC axon regrowth is less clear. RNAi knockdown of enzymes upstream of Pmvk (Hmgcr, Mvk) produces only mild phenotypes, and knockdown of several downstream enzymes produces no phenotype. The authors attribute this discrepancy to the possibility of weak RNAi constructs, which is plausible but not fully demonstrated. It would be helpful for the authors to discuss alternative explanations, including non-canonical roles for Pmvk that may not require the full pathway, and clarify the extent to which the current data support the conclusion that the mevalonate pathway, rather than Pmvk specifically, is a core regulator of regrowth.
It is not clear from the Methods whether γ KCs and α/β KCs were sorted from the same brains using orthogonal binary expression systems (e.g., Gal4 > reporter 1 and LexA > reporter 2), or isolated separately from different fly lines. If the latter, differences in genetic background, staging, or batch effects could influence transcriptional comparisons. This should be explicitly clarified in the Methods, and any associated limitations discussed in the manuscript.
The authors have made important findings that contribute to our understanding of axon growth and regrowth. As written, some major claims are only partially supported, but these issues can be addressed through reframing and clarification. In particular, the manuscript would benefit from (1) a more cautious interpretation of the mevalonate pathway's role, potentially considering Pmvk non-canonical functions, and (2) addressing methodological ambiguities in the transcriptomic analysis.
Reviewer #2 (Public review):
Fahdan et al. set out to build upon their previous work outlining the genes involved in axon growth, targeting two axon growth states: initial growth and regrowth. They outline a debate in the field that axon regrowth (For instance, after injury or in the peripheral nervous system) is different from initial axon growth, for which the authors have previously demonstrated distinct mechanisms. The authors set out to directly compare the transcriptomes of initial axon growth and regrowth, specifically within the same neuronal environment and developmental time point. To this end, the authors used the well-characterized genetic tools available in Drosophila melanogaster (the fruit fly) to build a valuable dataset of genes involved at different time points in axon growth (alpha/beta Mushroom Body Kenyon cells) and regrowth (gamma Mushroom Body Kenyon cells). The authors then focus on genes that are upregulated during both initial axon growth and axon regrowth. Then, using this subset of genes, they screen for axonal growth and regrowth deficits by knocking down 300 of these genes. 12 genes are found to be phenotypically involved in both axon growth and regrowth based on RNAi gene-targeted knockdown in the Mushroom Body. Of these 12 genes, the authors focus on one gene, Pmvk, which is part of the mevalonate pathway. They then highlight other genes in this pathway. But these genes primarily affect axon regrowth, not initial axon growth, implicating metabolic pathways in axon regrowth. This comprehensive RNA-seq dataset will be a valuable resource for the field of axon growth and regrowth, as well as for other researchers studying the Mushroom Body.
Strengths:
This paper contains many strengths, including the in-depth sequencing of overlapping developmental time points during the alpha/beta KCs' initial axon growth and gamma KCs' regrowth. This produces a rich dataset of differentially expressed genes across different time points in either cell population during development. In addition, the authors characterized expression patterns at developmental time points for 30 Gal4 lines previously identified as alpha/beta KC-expressing. This is very helpful for Drosophila
Mushroom Body researchers because the authors not only characterized alpha/beta expression but also alpha'/beta' expression, gamma expression, and non-MB expression. The authors comprehensively walked through identifying differentially expressed genes during alpha/beta axon growth, identifying a subset of overlapping upregulated genes between cell types, then systematically characterized whether knockdown of a subset of these genes produced an axonal growth defect, and finally selected 1 of 3 cell-autonomous genes important for gamma KCs regrowth to further study.
The authors utilized the developing Mushroom Body in Drosophila melanogaster, which happens to have new neurons developing axons and neurons that have undergone pruning and are regrowing neurons at the same developmental time. They are also in the same part of the brain (the Mushroom Body) and, in theory, since the authors implicate a metabolic pathway, they will have similar metabolic growth conditions.
Identifying Pmvk and two other components of the mevalonate pathway in axon regrowth opens up novel avenues for future studies on the role this metabolic pathway may have in axon growth. The authors of this paper are also very upfront about their negative results, allowing researchers to avoid running redundant experiments and truly build on this work.
Weaknesses:
While the dataset produced in this study is a strength, certain aspects make it more challenging to interpret. For instance, the authors state that roughly equal numbers of males and females are used for sequencing, and this vagueness, coupled with only taking a subset of the GFP-labeled neurons during FACs sorting, can introduce confounds into the dataset. This may hold true in imaging studies as well, in which males and females were used interchangeably.
Additionally, a rationale is needed to explain why random numbers of 1-7 were assigned to zero-expressing genes in the DESeq analysis. This does not seem to conform to the usual way this analysis is normally performed. This can alter how genes across the dataset are normalized and requires further explanation.
The display and discussion of the data set do not always align with the authors' stated goal of having a comprehensive description of the genes that dynamically change during axon<br /> growth and regrowth. Displaying more information about genes differentially expressed in the alpha/beta KCs, or any information about the genes diƯerentially expressed in the gamma KCs when using the same criteria as the alpha/beta KCs, or the 676 overlapping upregulated genes, would significantly add to this paper. The authors previously performed a similar study across developmental time points for gamma KCs, and it is not clear whether any overlapping genes were identified. Also, more information on the genes consisting of PC1 and PC3 when showing the PCA analysis would be helpful. Within the text, there is a discussion of why certain genes or gene groups were omitted or selected, such as clusters 1 and 2, and then some of their subgroups based on expected genes. There is also some discussion of omitted gene groups, but this is not complete across the different clusters, nor is there a discussion of why PC2 was not selected or of which genes might exhibit greater variability than cell type. The authors would make a stronger case for the genes they pursued if they showed that groups of genes already known to be involved in axon growth clustered within the selected groups. Since we do not see the gene lists, this is unclear and adds to the sometimes arbitrary nature of the author's choices about what to pursue in this paper. A larger set of descriptors, such as gene lists and Gene Ontology analysis beyond what is shown, would be very helpful in putting the results in context and determining whether this is a resource beneficial to others.
While the Pmvk story is interesting, the authors appear to make some arbitrary decisions in what is shown or pursued in this paper. Visually, CadN and Twr appear to be more severe axon regrowth phenotypes, where the peduncle appears intact, and axons are not regrowing in Figures 3 N and O. In contrast, Pmvk visually appears to lose neurons in Figure 3 M. With a change of the Gal4 driver (Figure 4), Pmvk now produces a gamma axon regrowth phenotype similar to CadN and Twr in Figure 3. This diƯerence in the use of Gal4 for characterizing axonal phenotypes is not discussed, making some interpretations more challenging due to diƯerences in Gal4 expression strength. For instance, the sequencing work was done with a diƯerent Gal4 MB expressing line than the characterization of gene knockdowns. Further characterization of the Pmvk was performed in the same Gal4 lines as the sequencing (Figure 4), suggesting a potential diƯerence in Gal4 strength that may play a role in their rescue experiments if they are using a slightly weaker Gal4 for gamma lobe expression. A broader discussion of this may make the selection of Pmvk less arbitrary if the phenotype is similar to those of CadN and Twr. Along the lines of the sometimes arbitrary nature of the genes chosen to pursue further, the authors state that they selected genes that showed differential expression at any time point. As they refine their list of genes to pursue further, they seem to prioritize genes that change at 18-21 APF. This appears to be the early period for axon growth in alpha/beta KCs and gamma KCs, based on Figure 1. A stronger case might be made at longer time points when the axon is growing or regrowing.
The paper would benefit from scaling back the claim that the mevalonate pathway is involved. The authors identified only a subset of genes from the mevalonate pathway, all immediately upstream of Pmvk, with no effect on downstream genes. Along these lines, the paper would benefit from a discussion of non-canonical PmvK signaling.
While the ability to take neurons at the same developmental time and from the same brain region is a strength, they are still 2 different types of neurons. Although gamma neuron axon growth occurs very early in development, it would be interesting to know whether the same genes are involved in their initial growth. A caveat to the author's conclusion is that these are 2 different cell types, and they might use different genetic programs or use overlapping ones at other times. The authors did not show that gamma KCs use these genes in their initial axon growth.
There is no way in which police can maintain dignity in seizing and destroying a donkey on whose flank a political message has been inscribed.
INFO 103 Winter 2026. I think this is absolutely right. It reminds me of the rapid development of AI in recent years. One point that people are extremely worried about is that if Ai really becomes so powerful that even humans cannot control it, and if it invays media platforms like a hacker and then changes the direction of public opinion, this would be a very terrifying thing. But at this point, how should we convict?
In this example, some clever protesters have made a donkey perform the act of protest: walking through the streets displaying a political message. But, since the donkey does not understand the act of protest it is performing, it can’t be rightly punished for protesting. The protesters have managed to separate the intention of protest (the political message inscribed on the donkey) and the act of protest (the donkey wandering through the streets). This allows the protesters to remain anonymous and the donkey unaware of it’s political mission.
This example shows how separating intention from action can be used to avoid responsibility. This can be applied to bots, since people who deploy them can hide behind the automated actions while denying responsibility for what the bot produces.
Antagonistic bots can also be used as a form of political pushback that may be ethically justifiable. For example, the “Gender Pay Gap Bot” bot on Twitter is connected to a database on gender pay gaps for companies in the UK. Then on International Women’s Day, the bot automatically finds when any of those companies make an official tweet celebrating International Women’s Day and it quote tweets it with the pay gap at that company:
It is "confrontational", but it has a social justice purpose - to use automation to counter the "pseudo-equality propaganda" of corporate marketing and bring the real structural problem (the wage gap) to the public. This example shows that some antagonistic bots can instead become tools for monitoring power.
Bots might have significant limits on how helpful they are, such as tech support bots you might have had frustrating experiences with on various websites. 3.2.2. Antagonistic bots:# On the other hand, some bots are made with the intention of harming, countering, or deceiving others.
The "bot" itself is not good or bad, but depends on what it is designed for and how the rules of the platform constrain it. For example, friendly bots (automatic captioning, vaccine progress, red panda images) essentially improve the efficiency of information acquisition and enhance the user experience; antagonistic bots (spam, fake fans, astroturfing), however, can create false public opinion and make people think that "many people support/oppose a certain opinion", which directly affects public judgment
We also would like to point out that there are fake bots as well, that is real people pretending their work is the result of a Bot. For example, TikTok user Curt Skelton posted a video claiming that he was actually an AI-generated / deepfake character:
As someone who's majoring in a creative field, I find it both incredibly interesting and concerning just how advanced AI is getting, and where this rapid innovation will take us in just a few years. It's so jarring to be watching a video on Tiktok or Instagram and fully believe it to be completely real, just to feel the need to dissect the video to see if it's really real. I can't begin to imagine how the job industry will change due to AI, but with innovation there (hopefully) comes opportunity.**
3.2.3. Corrupted bots# As a final example, we wanted to tell you about Microsoft Tay a bot that got corrupted. In 2016, Microsft launched a Twitter bot that was intended to learn to speak from other Twitter users and have conversations. Twitter users quickly started tweeting racist comments at Tay, which Tay learned from and started tweeting out within one day. Read more about what went wrong from Vice How to Make a Bot That Isn’t Racist 3.2.4. Registered vs. Unregistered bots# Most social media platforms provide an official way to connect a bot to their platform (called an Application Programming Interface, or API). This lets the social media platform track these registered bots and provide certain capabilities and limits to the bots (like a rate limit on how often the bot can post). But when some people want to get around these limits, they can make bots that don’t use this official API, but instead, open the website or app and then have a program perform clicks and scrolls the way a human might. These are much harder for social media platforms to track, and they normally ban accounts doing this if they are able to figure out that is what is happening. 3.2.5. Fake Bots# We also would like to point out that there are fake bots as well, that is real people pretending their work is the result of a Bot. For example, TikTok user Curt Skelton posted a video claiming that he was actually an AI-generated / deepfake character:
This passage uses three levels to remind us that "robots" themselves do not equate to intelligence or objectivity. Tay's "contamination" illustrates that machine learning-based conversational robots absorb biases from the platform as "language norms"—when training data comes from an environment full of provocation and racism, the system becomes an amplifier of prejudice; the problem is not just a technical failure, but a governance failure of treating a "public platform" as a safe training ground. Next, the "registered vs. unregistered bots" reveal the cat-and-mouse game of platform regulation and countermeasures: API restrictions act as rules and guardrails, while simulated clicks bypassing APIs disguise automation as "human," making it harder for platforms to track, demonstrating that visibility and controllability are themselves forms of power. Finally, the "fake bots" point to another form of deception: humans pretending to be AI to gain traffic, a sense of mystery, or immunity from responsibility—this blurs the line of "authenticity" and reminds us that in the attention economy, technological identity can also be used for performance and marketing.
On the other hand, some bots are made with the intention of harming, countering, or deceiving others. For example, people use bots to spam advertisements at people. You can use bots as a way of buying fake followers, or making fake crowds that appear to support a cause (called Astroturfing). As one example, in 2016, Rian Johnson, who was in the middle of directing Star Wars: The Last Jedi, got bombarded by tweets that all originated in Russia (likely making at least some use of bots). “I’ve gotten a rush of tweets – coordinated tweets. Like, somewhere else on the internet there’s like a group on the internet saying, ‘Okay, everyone tweet Rian Johnson.’ All from Russian accounts, and all begging me not to kill Admiral Hux in this movie.” From: https://www.imdb.com/video/vi3962091545 (start at 7:49) After the Star Wars: Last Jedi was released, there was a significant online backlash. When a researcher looked into it: [Morten] Bay found that 50.9% of people tweeting negatively about “The Last Jedi” were “politically motivated or not even human,” with a number of these users appearing to be Russian trolls. The overall backlash against the film wasn’t even that great, with only 21.9% of tweets analyzed about the movie being negative in the first place. https://www.indiewire.com/2018/10/star-wars-last-jedi-backlash-study-russian-trolls-rian-johnson-1202008645/ Antagonistic bots can also be used as a form of political pushback that may be ethically justifiable. For example, the “Gender Pay Gap Bot” bot on Twitter is connected to a database on gender pay gaps for companies in the UK. Then on International Women’s Day, the bot automatically finds when any of those companies make an official tweet celebrating International Women’s Day and it quote tweets it with the pay gap at that company:
This passage shifts the discussion of "bots" from neutral tools back into the context of power and manipulation: they can not only automate the dissemination of information but also automate the creation of "false impressions of public opinion" (follower boosting, astroturfing) and targeted harassment (the coordinated attack on Rian Johnson). More notably, the research mentions that a large number of negative tweets were "politically motivated or non-human," meaning that the anger, ridicule, and boycotts we see online may not be a natural aggregation of "genuine public opinion," but rather an emotional landscape that is organized, amplified, and fabricated. Finally, the "Gender Pay Gap Bot" provides a counterexample: this "adversarial" automation can be used for public accountability—by forcibly juxtaposing corporate holiday statements with structural data (wage gaps), it forces people to see the reality obscured by public relations language. The key is not whether "bots are good or bad," but who uses them and whose perceptions and interests they are used to shape.
As a final example, we wanted to tell you about Microsoft Tay a bot that got corrupted. In 2016, Microsft launched a Twitter bot that was intended to learn to speak from other Twitter users and have conversations. Twitter users quickly started tweeting racist comments at Tay, which Tay learned from and started tweeting out within one day. Read more about what went wrong from Vice How to Make a Bot That Isn’t Racist
The discussion of bots influencing public opinion raises important ethical questions about power and accountability. Even if a bot spreads accurate information, the scale and speed of automation can still distort public discourse. This suggests that ethical evaluation of bots should consider not only content accuracy but also their impact on human decision-making and democratic processes.
The discussion of bots influencing public opinion raises important ethical questions about power and accountability. Even if a bot spreads accurate information, the scale and speed of automation can still distort public discourse. This suggests that ethical evaluation of bots should consider not only content accuracy but also their impact on human decision-making and democratic processes.
ethically justifiable
To me I find it problematic in practice for there to be a distinction between ethical and non-ethical use of antagonistic bots. Everybody has their own worldview and values. To define some of these values as ethical on social media is to impose them on everyone. Maybe this would be okay if there was a democratic way for this. But there isn't. These bots are made to "get a rise out of people" or stir emotions. Subjecting people to that through automated bots under the guise of ethics I disagree with
Some bots are intended to be helpful, using automation to make tasks easier for others or to provide information, such as:
I was already familiar with harmful bots, but it was interesting to learn about friendly bots! I learned more about their abilities while reading this section, which I was previously unaware of.
The truth is, your biological sex isn’t carved in stone, but a living system with the potential for change.
Actual research shows that sex is anything but binary
Sun’s article argues that biological sex cannot be reduced to a simple binary and that scientific research contradicts the “basic biology” often cited to oppose transgender identities.
Nearly everyone in middle school biology learned that if you’ve got XX chromosomes, you’re a female; if you’ve got XY, you’re a male
(4. Audience) This text eludes to who is reading the text/meant to read the text. It is likely that not actual middle schoolers are reading this, but many people in the audience probably remember learning about sex like this. Maybe the audience is those with conventional ideas of sex similar to this and those who have been possible swayed by the bad actors listed prior (trolls and politics). In other words, people uneducated on the issue
Let’s just take the most famous example of sexual dimorphism in the brain: the sexually dimorphic nucleus of the preoptic area (sdnPOA). This tiny brain area with a disproportionately sized name is slightly larger in males than in females. But it’s unclear if that size difference indicates distinctly wired sdnPOAs in males versus females, or if—as with the bipotential primordium—the same wiring is functionally weighted toward opposite ends of a spectrum. Throw in the observation that the sdnPOA in gay men is closer to that of straight females than straight males, and the idea of “the male brain” falls apart.
This small paragraph displays that the text is meant for college students and also individuals familiar with biology, and can understand scientific reasoning from research. We also know because most likely, those who are subscribing to the SCI AM are generally interested in or have a background in STEM/science
Actual research shows that sex is anything but binary
The popular belief that your sex arises only from your chromosomal makeup is wrong.The truth is, your biological sex isn’t carved in stone, but a living system with the potential for change.
The popular belief that your sex arises only from your chromosomal makeup is wrong.The truth is, your biological sex isn’t carved in stone, but a living system with the potential for change.
In other words, there is no such thing as “the male brain” or “the female brain.” This is not to say that there are no observable differences. Certain brain characteristics can be sexually dimorphic: observable average differences across males and females. But like biological sex, pointing to “brain sex” as the explanation for these differences is wrong and hinders scientific research.
The popular belief that your sex arises only from your chromosomal makeup is wrong.The truth is, your biological sex isn’t carved in stone, but a living system with the potential for change.
In other words, there is no such thing as “the male brain” or “the female brain.” This is not to say that there are no observable differences. Certain brain characteristics can be sexually dimorphic: observable average differences across males and females. But like biological sex, pointing to “brain sex” as the explanation for these differences is wrong and hinders scientific research.
Genre: The author writes in a scientific commentary style. Throughout the article they are constantly not just summarizing the research, but giving commentary and critiquing scientific errors. This type of genre generally aims to challenge misinformation and persuade readers as a call to action.
Note that sometimes people use “bots” to mean inauthentically run accounts, such as those run by actual humans, but are paid to post things like advertisements or political content. We will not consider those to be bots, since they aren’t run by a computer. Though we might consider these to be run by “human computers” who are following the instructions given to them, such as in a click farm:
This paragraph, I mean, it's very important that people know what bots do, they misconceptualize them, but in this day of ChatGPT, Microsoft Copilot, Google Gemini, DuckAI. These bots though are actually interesting in a sense how that not all accounts called ‘bots’ are truly automated. I think this distinction is important because it changes how we understand online content, whether it’s influenced by algorithms or by campaigns.
Fig. 3.1 A photo that is likely from a click-farm, where a human computer is paid to do actions through multiple accounts, such as like a post or rate an app. For our purposes here, we consider this a type of automation, but we are not considering this a “bot,” since it is not using (electrical) computer programming.
This is my first time seeing a click-farm, and I usually associate these accounts with bot accounts because they perform similar things. Both are used to boost inauthentic engagement for posts, advertisements, or other content.
Note that sometimes people use “bots” to mean inauthentically run accounts, such as those run by actual humans, but are paid to post things like advertisements or political content. We will not consider those to be bots, since they aren’t run by a computer. Though we might consider these to be run by “human computers” who are following the instructions given to them, such as in a click farm:
I like this clarification because “bot” gets thrown around so loosely online. It makes sense to draw the line at whether an account is actually run by software versus a human, even if that human is being paid and tightly scripted. Calling click-farm workers “human computers” is kind of unsettling, but it does a good job of showing how inauthentic behavior isn’t always automated in a technical sense.
his approach concludes that most of the works that were found atNag Hammadi are not Gnostic because they lack the Gnostic myth evenif some include certain of its characters or motifs in otherwise quite differ-ent stories.
But certainly there's some reason for these parallels, yes?
Thispicture of Christian life may not match conditions of the middle of thesecond century, but instead may suit better the more developed Chris-tian churches of the third century, long after Irenaeus.
This is a very tenuous premise.
hose the name Rebeccafor Pocahontas with the hope that, like the biblical Rebecca, the newconvert would favor her white children (like Jacob, whose skin waslight) over her "red" children (referring to Esau, who was characterizedas red).
This is a thing that a lot of people do currently as well. They choose a different name to fit in to the different environment, people, and language. This shows that Pocahontas was not as happy as described in the movies, but tried to fit in and adjust to her situation. This was particularly interesting to me as this happens a lot today as well.
The Hebrew word Sakal is often translated as "prosperity" or "success," but the literal meaning is "Tactical Insight" or "Circumspection."
📝 Intel Expansion: The word Sakal (שָׂכַל) implies looking at the mechanics of a situation. It is the opposite of "blind faith."
Cross-Reference: Ephesians 5:15 (KJV) uses the same concept: "See then that ye walk circumspectly, not as fools, but as wise." To walk circumspectly means to look around (circum) and inspect (spect). It is a scan of the environment.
That is not peace; that is denial.
[NEURO-LINK] In psychology, this is called "Avoidance Coping." It feels like safety, but it actually spikes cortisol levels over time because the threat remains unresolved in your subconscious processing. True Shalom disarms the threat; it doesn't ignore it.
To this I would simply say that the burden of proof is ontheir side: we’ve been at this ‘study of religion’ thing for a while now, and still, we have nosingular definition of ‘Religion’ which the majority of scholars can agree on. If it is the case thatall of our various attempts to impose one abstract concept on disparate phenomena constantlycome up short, then isn’t it perhaps more likely that the abstract concept which we are trying toemploy is precisely not a derivation from the phenomena, but rather an imposition on them? Thehistorical perspective, on the other hand, shows that religion changes, and that this is because wechange with it.
There are some good ideas raised here, but the argument, as is, is unpersuasive.
that one can abstract ‘Religion’ from‘religions’ in some meaningful sense
But this isn't the only way to define religion, and it would actually be circular to do so. This seems, again, like a category error, or a strawman at best.
such a concrete thing as religion
But I don't think even most perennialists would argue this. Religion is an abstract thing by definition, it would seem, but does that make it any less "real," any less a "thing"?
but it didn’t have secularists—which is to say it didn’t havereligion either.
Again...very debatable, but also, I don't think the argument follows anyway.
any, perhaps even most or all,people who practice a religion today have some understanding of themselves as doing thisbroader, second order thing, ‘religion’, and this would seem perfectly natural to us
Maybe, but ofc the Evangelical "relationship, not religion" people.
Even a distinctionsuch as that between Christianity and ‘Heathenism’, for example, could have been made withoutrecourse to the general category of religion
But what about Christianity vs. atheism or "superstition"?
But is it not the case that in our average understanding religion and science are so oftenopposed, or if not outright opposed, then somehow at odds?
"Average understanding"? Maybe to a post-Enlightenment European, but even many of those see no opposition.
Does the fact that it is a bot change how you feel about its actions?
Not really. I think in a lot of controversial cases involving bot behavior are often not excused because at the end of the day, someone programmed it. Maybe it's because I don't know exactly how bots work, but I feel like a bot's actions are limited to what its programmer allows it to do, so if a bot is able to do something (such as post racist comments, as explained in an earlier section of this chapter), then something in its code allowed it to process racist information and express it. In the case from earlier this chapter, it probably should've been considered that if the bot is using other twitter users' tweets toward the company and that information is public, there probably should've been safeguards to detect racial remarks and block it from the bot's vocabulary and processing. It could be argued that that scenario slipped from their minds, but I think that anything that will be released to the public should be tested and be given feedback from a smaller community before released to the mass public. Surely someone would've thought that the bot could pick up racist tweets.
hey also emerge within specific social, political, andmaterial conditions that shape what can be thought, said, and done. An emphasis on intelligibilitymay therefore leave insufficient room for analyzing how religious forms are produced
But can't these elements simply be integrated into an intelligible account?
I argue that an assumption of coherence is methodologically necessary forunderstanding how practitioners experience religious objects as meaningful, but that this must betreated as a heuristic tool rather than as a description of lived religious reality.
Agreed.
interpretive reconstruction
But this strikes me as very "external." What is the "internal" sense that I am supposed to get?
The Aché, a foraging group living in the subtropical rainforest in Paraguay, eat 33 different kinds of mammals, more than 15 species of fish, the adult forms of 5 insects, 10 types of larvae, and at least 14 kinds of honey. This is in addition to finding and collecting 40 species of plants.[5] The !Kung foragers, who live in the Kalahari Desert in southern Africa, treasure the mongongo nut, which is tasty, high in protein, and abundant for most of the year, but they also hunt giraffes, six species of antelope, and many kinds of smaller game like porcupine.[6]
I once would have said this was disgusting, but I have grown beyond that
Many of the foods regularly eaten by foragers, such as insects and worms, would not necessarily be considered edible by many people in the United States.
Unless we were starving and we'd be forced to, but hey, I wonder what they taste like
Imagine if we could bring world-class healthcare, education, finance and creative services to anyone for almost zero cost. People able to afford these services don’t doomscroll looking for more stuff. They goal seek to flourish more.
Not only is production at a low cost but distribution is now at the point and available hyper targeted.
biosocial
Although this likely pertains to more contemporary times, I think this is a great reminder that biology, although often seen solely as physical, can also be molded through social and cultural constructs. Thus, in interpreting skeletal remains, it is important to consider what biology may reveal about the social environment the individual inhabited during life. In deep time, this obviously may be difficult, but I think it serves to demonstrate that great thought must be devoted when attempting to assign individual anatomy to larger populations.
perperson
Perhaps it is mentioned elsewhere, but what does this mean by 'per person?' Does this refer to every maxilla/mandible that had articulated teeth? I feel like teeth would fall out fairly easily within a burial context, though I may be wrong. If the teeth are disarticulated, it seems it would be rather difficult to assign them to a particular person before analysis was conducted on them.
control the story
I think this relates to one of the reading questions: "The commitment on the part of the HWN to allow this biological material to be retained is beneficial to researchers, but is it beneficial to them?" While I do think the research was beneficial for the Huron-Wendat as it revealed scientific data regarding their past, one has to question how much authority the group actually had in 'controlling their story.' Though sometimes individuals from Indigenous groups may be involved in the research, often times it will be individuals outside of these groups running the tests in labs and publishing papers. In this sense, though the Huron-Wendat did play an important role in this research, one has to question how much authority they actually have in 'controlling their story' within a scientific context. I don't know if there is a simple, practical solution to this, however.
Dear authors, congratulations on the beautiful work. - This is clearly a very important work to define the ON-DSGC subtype and investigate the developmental regulation of the subtype using genetic and functional approaches. I have a minor critique - For Figure 6, I suggest that your revised/peer-reviewed paper should cite the original publication (PMID: 35524141) in both text/method on mWGA-mCherry as (mWmC+) instead of saying a generic (WGA+), as your staining was RFP (the mWmC fusion ) but not WGA-protein. The fusion protein (but not the original plant lectin from Yoshihara 1999) made visualization robust and quantifiable.
A goal, on the other hand, is something that determines a process, yet lies in the future.
The goal as something that is not present, but which the system works towards is an example of an absential.
16.Evolution and immortality Cybernetic integration of humans must preserve the creative core of human individual, because it is the engine of evolution. And it must make it immortal, because for the purpose of evolution there is no sense in killing humans. In natural selection, the source of change is the mutation of the gene; nature creates by experimenting on genes and seeing what kind of a body they produce. Therefore, nature has to destroy older creations in order to make room for the newer ones. The mortality of multicellular organisms is an evolutionary necessity. At the present new stage of evolution, the evolution of human-made culture, the human brain is the source of creativity, not an object of experimentation. Its loss in death is unjustifiable; it is an evolutionary absurdity. The immortality of human beings is on the agenda of Cosmic Evolution. 17.Evolution of the human person The future immortality of the human person does not imply its frozen constancy. We can understand the situation by analogy with the preceding level of organization. Genes are controllers of biological evolution and they are immortal, as they should be. They do not stay unchanged, however, but undergo mutations, so that human chromosomes are a far cry from the chromosomes of primitive viruses. Cybernetically immortal human persons may mutate and evolve in interaction with other members of the super-being, while possibly reproducing themselves in different materials. Those human persons who will evolve from us may be as different from us as we are different from viruses. But the defining principle of the human person will probably stay fixed, as did the defining principle of the gene.
CEStoicism does not desire the immortality of the individual, to which points 16 and 17 aspire. The evolution of culture/worldview/p-individual can occur with or without the indefinite continuation of individuals. Instead, creative immortality and cultural P-individual immortality suffice.
old religious systems are slowly but surely losing their influence.
CEStoicism is unsure if older religious systems, and the idea of metaphysical immortality, are losing their influence. However, we do agree that other forms of immortality are gaining appeal, including the immortality of stable conceptual systems (SCSs).
Continuing constructive evolution is a possibility but not a necessity. Acts of will can contribute to evolution or counter it. Because of the natureof evolution, there is a fundamental difference between constructive and destructive contributions.
Living parts of existence that continue to survive) have periods of constructive evolution in the K-phase of their adaptive cycle, and also demonstrate resilience both within one cycle and throughout many cycles in a panarchy.
Thoracic cavity
A further break down of the thoracic cavity to include the pleural cavity, pericardial cavity, and superior mediastinum terms would be helpful here. The information is a part of the diagram, but the terms are not found in the descriptive text.
distinguishes us from the animals
We respect the specific definition of 'thought' defined by Principia Cybernetica, but we feel the real distinction of the human animal from other animals is humans' extensive use of fine-grained signs. We also stress that self-consciousness, tool making, imagination, planning, play, sense of humor and esthetic feelings are likely abilities/activities shared by some other animals, although maybe not to the same fine-grained ability as humans. We further stress that human are still animals.
an additional mechanism which controls the behavior and production of the S-subsystems
Specifically, this mechanism is our nomic and entailment net.
In some situations the control mechanisms may come from a place external of the systems under control. For example, from a farmer (C) that controls many of the conditions/inputs of the plants (Sn) on the farm (S').
But in the situation of a true democracy (S'), the control mechanism (perhaps better called a coordination mechanism) comes from the people themselves (Sn). This coordination mechanism (C) is composed of a good-faith process where the people (Sn) create and edit the rules they agree to live under.
selection
See https://cestoicism.neocities.org/selection
Also see Rachel Burnham's review (http://rachelburnham.blogspot.com/2019/07/review-of-curation-power-of-selection.html) of Michael Bhaskar's book "Curation. The power of selection in a world of excess" describes curation in terms of selection:
[Bhaskar] describes curation as 'using acts of selection and arrangement (but also refining, reducing, displaying, simplifying, presenting and explaining) to add value. ... He sees expert selection as at the start of good curation and quotes from Maria Popova, curator of the highly thought of 'Brain Pickings': "The art of curation isn't about the individual pieces of content, but about how these pieces fit together, what story they tell by being placed next to each other, and what statement the context they create makes about the culture and the world at large. This is, she argues, a process of 'pattern recognition'. Seeing how things fit together, understanding connections (which multiply in a networked environment), but then also, crucially, creating new ones by recombining them, is a massive part of curation."
Under these conditions, ironically, the OLP might for the time being remain ‘a democratic procedure for regulatory issues only’. However, the passage to bicameralism is real, and intergovernmental carve outs and parliamentary conservatism are part and parcel of the deepening of co-decision. The OLP compels Member States to ‘manage’ the EP in areas of sensitive issues where they would rather just ignore it. Likewise, it exposes MEPs to the growing public expectations of civil society organizations. EU bicameralism is growing out of the straitjacket of intergovernmental relations—even in sensitive cases of market regulation.
Basically, all this resistence from Member States i in fact (as author argues) EVIDENCE OF CO - DECISION AND INTEGRATION WORKING - > necssary hurdles to overcome -> successful negortaions processes.
OLP makes Member States "manage" the EP in areas where BEFORE LISBON they would have ignored it (selective exits? Less possible after Lisbon. Good queston idk)
OLP is theoretically now ONLY for regulatory issues but that IS changing (ad evidenced)
In conc -> trilogues need to be more transparent, EP needs better procedures for it. Need to connect Member States and citizens more -> therefore need more open dialogue.
"Under these conditions, ironically, the OLP might for the time being remain ‘a democratic procedure for regulatory issues only’. However, the passage to bicameralism is real, and intergovernmental carve outs and parliamentary conservatism are part and parcel of the deepening of co-decision. The OLP compels Member States to ‘manage’ the EP in areas of sensitive issues where they would rather just ignore it. Likewise, it exposes MEPs to the growing public expectations of civil society organizations. EU bicameralism is growing out of the straitjacket of intergovernmental relations—even in sensitive cases of market regulation."
All in all, the democratic effects of the co-decision and OLP procedures seem to trickle down in everyday law-making through a sedimental process where democratic norms play a certain, but not uncontested, role.
Anyway, existence of trilogues (they are regulated -> EP has reporting requirements on them, but still controversial) is evidence of DEMOCRATIZATION of co-decision process -> i.e., not those macro treaty level things, but in DAILY PRACTICE -> in other words, Democratic effects of the OLP and co-decision being codified are influence on EVERY DAY actings of legislative bodies and this is EVIDENCED by the existence of trilogues.
Trilogue "socialization" is the "realist" interpretation of co-decision.
"the trilogues themselves have generated their own norms and expectations as part and parcel of the broader process of socialization between the Commission, EP, and Council participants. It has been argued that trilogue socialization has promoted a more ‘realist’ interpretation of co-decision, which has moderated the reformist ambitions of the EP by linking its credibility to expectations of ‘responsible’ legislator (Ripoll Servent 2012). All in all, the democratic effects of the co-decision and OLP procedures seem to trickle down in everyday law-making through a sedimental process where democratic norms play a certain, but not uncontested, role."
a political entity which had evolved institutions and procedures well beyond those of an international organization, but remained fundamentally messy.
IN OTHER WORDS -> looking at how this helped bring the EU out of an "international organization" and into some sort of actual democratic / legislative body -> elevates it from simply regulatory body to government - BEFORE Maastricht, scholars tried to find out what the new "POLITCAL ORDER" was now that the EC was more unified -> again, conclude that the nascent EU/EC is WAY MORE POWERFUL THAN ANYTHING ELSE on an international scale, but remains "messy." - Part of this mess was fact that there was no OVERARCHING POLICY OR GOAL -> simply "general principles or tendencies" - Again, many hope at this point (again, like 1990 before EU treaty of 1992) for greater transnational demoicratic institutions, not just (appointed?) regulatory body.
hereditary
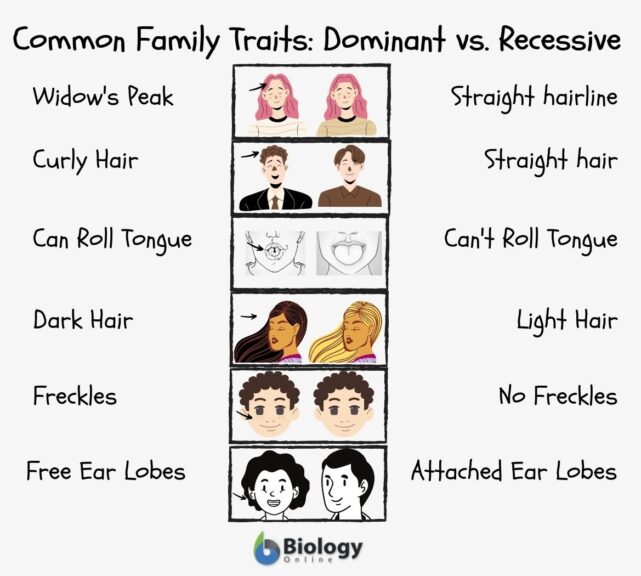
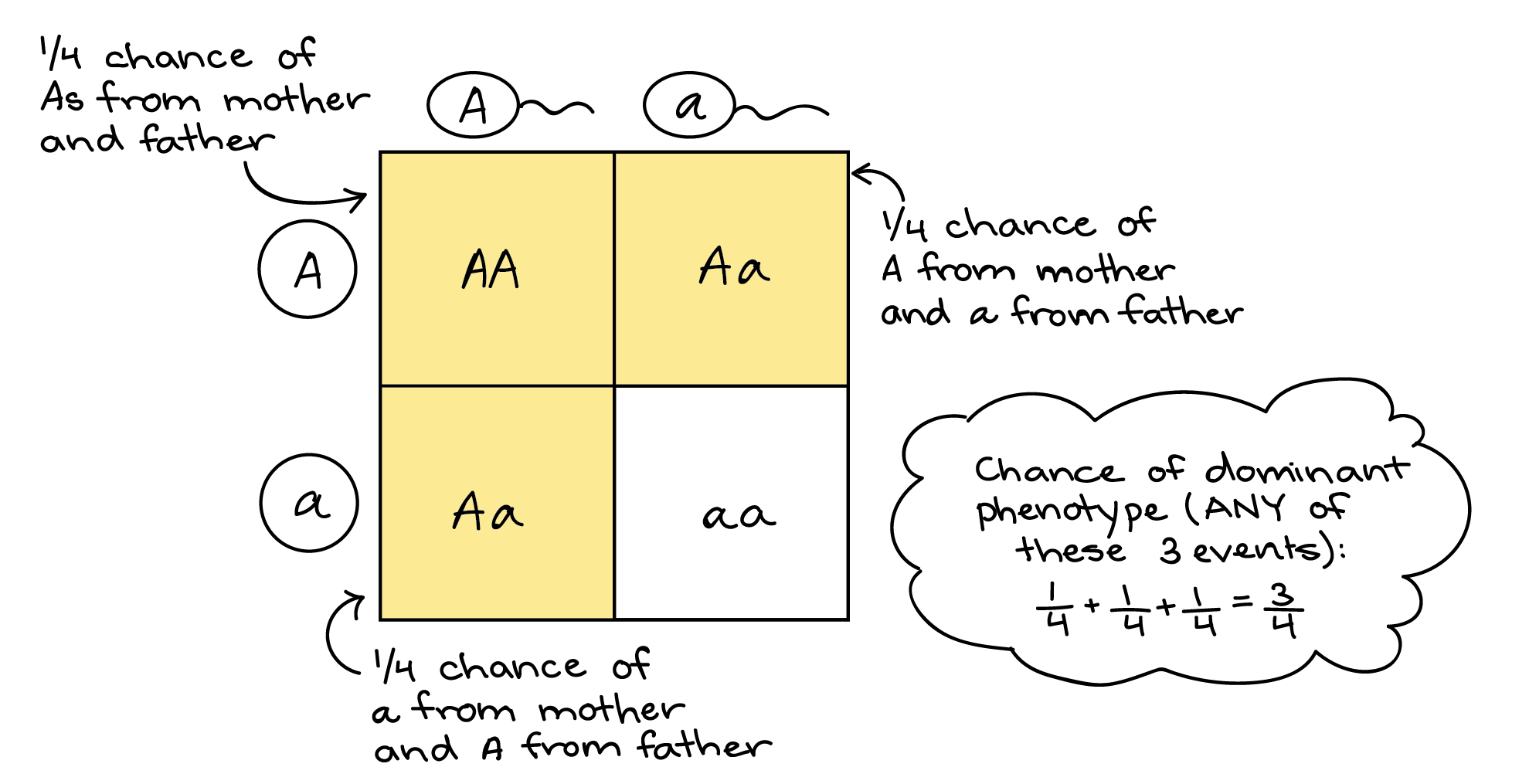

Hereditary means passed from parents to offspring through genes. If something is hereditary, it can be inherited biologically because it is encoded in DNA.
Each person usually inherits two versions of a gene (alleles):
one from the mother
These alleles influence:
traits (eye color, hair texture)
Important:
遗传的(hereditary) 指 通过基因从父母传给子女的性状或特征,本质上与 DNA 有关。
等位基因共同决定:
外貌或生理性状
遗传 ≠ 一定会发生
有些人携带基因,但并不表现症状
看到 hereditary,可以立刻联想到这条逻辑链: DNA → 基因 → 等位基因 → 遗传方式 → 性状 / 疾病
如果你愿意,我可以下一步把它做成 中英对照生物词卡(flashcards),或直接嵌入你之前的 Unit 1 DNA HTML 教学页面。
alleles

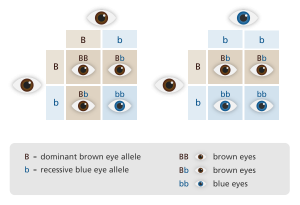


Alleles are different versions of the same gene. They control variations of a trait, such as eye color or flower color.
Allele = one version of a gene
Each gene can have two or more alleles, but an individual organism usually carries two alleles per gene (one from each parent).
📌 This is why offspring show traits from both parents.
Trait: Seed color
Possible allele combinations:
These combinations affect the trait that appears.
Dominant allele
Recessive allele
📌 Example:
| Term | Meaning | Example | | --------- | ------------------ | ---------- | | Allele | Version of a gene | A or a | | Genotype | Allele combination | Aa | | Phenotype | Physical trait | Brown eyes |
Alleles determine the genotype, which determines the phenotype.
Punnett squares:
📌 Example: Parents: Aa × Aa
Alleles:
Alleles are different versions of the same gene that determine variations in traits.
等位基因(alleles)是指同一基因的不同版本,决定同一性状的不同表现。
等位基因 = 同一基因的不同形式
性状:豌豆高度
组合可能是:
📌 Tt → 显性性状 📌 tt → 隐性性状
等位基因是控制同一性状的不同基因形式。
如果你愿意,我可以把 alleles → genotype → phenotype → Punnett squares 整理成 Science 10 中英对照闪卡或互动练习,直接用于复习或教学。
等位基因(allele) EN: Different versions of the same gene that may produce different forms of a trait. Example: For pea flower colour, one allele codes for purple and another for white. 中文:位于同源染色体相同位置、控制同一性状的基因的不同形式。 例子:例如花色基因可以有紫花等位基因和白花等位基因。
The blog won't save us. But it's one of the tools we'll need if we're going to save ourselves.
Blogs as self care
Diderot understood that the container shapes the contents. The Encyclopédie was a collection of facts, yes, but more fundamentally it was an argument about how knowledge should be organized. Cross-references between entries were themselves a form of commentary, connecting ideas that authorities wanted kept separate.
Posits the original encyclopedia was both a k organising attempt and a network of references between ideas, thus bringing them together
But it also produced actual intellectual communities. Remember those?People wrote long responses to each other's posts, those responses generated further responses, and you could follow the thread of an argument across multiple sites and weeks of discussion. The format rewarded careful thinking because careful thinking was legible in a way that it simply isn't on platforms designed for rapid-fire engagement.
describing the distributed conversations that were important to me for blogging too. Not sure we were as exalted though.
When people talk about the Enlightenment as if it were an intellectual garden party where everyone sipped wine and agreed about reason, they're missing the part where producing and distributing ideas was (in fact) dangerous and thankless work
Enlightenment was not a salon, but an era where coming up with ideas and spreading them carried risk.
Some good pointers to [[Brian Eno c]] work and thinking, to follow up.
Also good anecdote from one of those links on Rem Koolhaas notion of n:: premature sheen Making things look nice early takes away from thinking about other points of quality. Jeremy applies it to AI too, the premature sheen generate awe, but not quality output.
"I think of Cognitive Debt as ‘where we have the answers, but not the thinking that went into producing those answers”. It is a phenomenal largely (but not exclusively) fuelled by the deployment of LLMs at scale. Answers are now much, much cheaper to come by.
Additionally, I am most interested in exploring Cognitive Debt not from an individual perspective, but from a group one. It is critical to thinking through the implications of using these technologies inside an organisation, or between an organisation and its employees, a government and its citizens, and so on and so forth."
n:: cognitive debt - [ ] return
This requirement is the biggest difference between skills and other previous attempts at expanding the abilities of LLMs, such as MCP and ChatGPT Plugins. It’s a significant dependency, but it’s somewhat bewildering how much new capability it unlocks.
需要依赖本地环境(文件系统、工具)来拓展 agent 的能力,是 skills 和 mcp 的巨大不同。
The /compact command summarizes your entire conversation history while preserving the key information Claude has learned. This is ideal when: Claude has gained valuable knowledge about your project You want to continue with related tasks The conversation has become long but contains important context
用 /compact 命令来总结当前的上下文,让 claude 记住当前得到的知识
Segregation is not simply physical separation; it is an attempt to deny and prevent association with another group. Denying association with another group is another way of denying that group’s basic humanity. In that sense, segregation is not just spatial projects but ontological.
I imagine that the author means to imply that the minority group's humanity is the one being denied. What if there are 2 groups of almost equal presence but there is also conflict between them, would segregation work then?
In another study, images of persons identified by varying social groupings triggered different responses in the brain when observed under an MRI.45Lasana T. Harris and Susan T. Fiske, “Dehumanizing the Lowest of the Low: Neuroimaging Responses to Extreme Out-Groups,” Psychological Science 17, no. 10 (2006): 847–53. jQuery("#footnote_plugin_tooltip_2620_45").tooltip({ tip: "#footnote_plugin_tooltip_text_2620_45", tipClass: "footnote_tooltip", effect: "fade", fadeOutSpeed: 100, predelay: 400, position: "top right", relative: true, offset: [10, 10] }); Persons belonging to these especially marginalized outgroups did not even trigger recognition at a neural level as being human, as if they were animals or objects.
It is a very interesting discovery but also I must show my fascination that we have the technology to reach these conclusions.
Through talk, tales, stories, gossip, anecdotes, pronouncements, news accounts, orations, sermons, preachments, and the like, definitions are presented and feelings expressed…If the interaction becomes increasingly circular and reinforcing, devoid of serious inner opposition, such currents grow, fuse, and become strengthened. It is through such a process that a collective image of a subordinate group is formed, and a sense of group position is set
I agree that it is difficult to resist the urge to naturally group with those you find similar especially when it is placed in the environment stated here so I am starting to believe that there is nothing wrong with the formation of the group itself but rather how the group chooses to interact with what they consider outsiders.
Studies since the 1950s demonstrate the tendency of people to identify with whom they are grouped, no matter how arbitrary or even silly the group boundaries may be, and to judge members of their own group as superior. Studies dividing students into completely fabricated groups lead to consistently different perceptions of in-group and out-group members.
To me, this is enough to draw the conclusion that humans are biologically predisposed to partake in othering. This however is a truth that I do not want to accept and will settle for accepting that humans are predisposed to group with those they find similar but are not predisposed to treat those outside of the group negatively.
demagoguery is not an inevitable feature of political life in periods of geopolitical change or economic turmoil. It is a strategy dependent upon the choices of political actors.
I strongly agree that there would not be an issue (politically) to begin with if the politicians didn't view othering as a weapon to begin with but since there is no way to guarantee the morality of a politician I find it beat to take away the weapon entirely by educating people
The idea of stoking anxiety, resentment, or fear of the “other” is not a new electoral strategy in American politics. Appeals to nativism, racism, and xenophobia are evident in almost every period of American history.
I do not find it at all surprising that Americans have found a way to leverage hatred, ignorance and confusion to their advantage in winning elections but in a way it is not just the fault of the person using their ignorance for their own gain but especially the fault of the ignorant part that refuses to educate itself and do its own research.
this article investigates the forces that contribute to othering
While I am sure that many many conditions apply to othering and many factors can influence its commonality and severity, I believe that othering will always exist as long as people are different. And since it is impossible to have 8 billion of the same person in our world othering will always exist. We can only suppress and mitigate but I dont believe it possible to eliminate it unfortunately.
“Othering” is a term that not only encompasses the many expressions of prejudice on the basis of group identities, but we argue that it provides a clarifying frame that reveals a set of common processes and conditions that propagate group-based inequality and marginality.
I feel like this comment has helped me differentiate between othering and bias better since that was a question that was posed during class.
You accept that the work will wait, but the human moment will expire. You are not a processor; you are a partner. (Cause).
This results in an immediate spike of Cortisol and Adrenaline, narrowing your visual and emotional field—a "biological hijack." To counter this, you must engage the Vagus Nerve. By pressing your feet flat and extending your exhale, you signal the parasympathetic nervous system to downregulate the alarm, shifting the energy from "Defence" back to "Social Engagement." You are effectively clearing the cache of your emotional processor to make room for a new, higher-priority input.
The "Physics of the Kingdom" dictates that Love (Agape) is the primary vector. In 1 Corinthians 13, Paul describes Love as not being "easily provoked" (paroxynetai—literally, "not sparked into a sharp edge"). When you snap, you have allowed your internal friction to create a spark that severs the "cord" of connection. By hitting the "Save State," you are aligning with the Sabbath Logic: the world (and the code) is sustained by God, allowing you the freedom to stop, turn, and witness the Image Dei standing in your doorway.
The Physical Save (5 Seconds): Physically press Ctrl + S (or Command + S). This tactile movement tells your brain: "The data is safe; the PFC can let go."
The Grounding (10 Seconds): Press both feet into the floor. Feel the weight. Exhale slowly through pursed lips as if blowing out a candle.
The Pivot (5 Seconds): Physically rotate your chair or your torso 180 degrees away from the screen.
The Identification: Look at the person and internally label them: "Image of God. Priority One."
The Opening: Say: "I've saved my place. I'm listening."
Antenatalmaternal symptoms of anxiety or depression are associatedwith more difficult/reactive infant temperament (17–19),independent of postnatal maternal mental health (18, 19)
Symptoms of anxiety and depression in mothers during pregnancy are associated with difficult/reactive infant temperament, but not when they occur after pregnancy. That is, mental health of the mother during pregnancy affects the child but not mental health after. So, some sort of transmission from mother to child during pregnancy affects infant temperament instead of geneticss?
It was a balmy night in Deerfield Beach, Florida. The conference was packed with philosophers, sociologists, and programmers, all intent on examining the latest developments in consciousness and artificial intelligence. Papers had been presented, models dissected, scenarios examined. I had brought my camera along, without any clear idea of what I meant to photograph. But seeing Sophia there sparked an idea. Portrait photography is usually about connecting with other human beings and trying to capture their essence, presenting whatever it is that makes them beautiful and unique. What if I were to photograph Sophia—a humanoid robot developed by Hanson Robotics—and then, in a separate session, the philosopher David Chalmers, a prominent theorist of consciousness, and reflect on the experience? What might I learn from those encounters that I had not already gleaned from the analytical papers and philosophical discussions?
will this be seen
It seems fitting to follow up the expectations for the first year with a list of common challenges that college students encounter along the way to a degree.
Annotation 2: Reading about imposter syndrome really stood out to me because it’s something a lot of students probably experience but don’t talk about. I have experienced it myself, especially going back to school after many years. Knowing that feeling unsure or overwhelmed is normal makes college feel less intimidating. It reminded me to be patient with myself while adjusting and to not give up when things get hard.
Public memory is produced from a political discussion that involvesnot so much specific economic or moral problems but rather funda-mental issues about the entire existence of a society: its organization,| structure of power, and the very meaning of its past and present.
The struggle with memory can be political because it is how we understand the power of politics in the past.
he stories I keep close to me, day by day, are those that yield more and more of their spirit in time.
Hence, why its important to ask but not always necessary to solve.
t remains for me one of the most intensely vital stories in my experience, not only because it is a supernal example of the warrior idea—an adventure story in the best sense—but because it is a story about story, about the efficacy of language and the power of words. One does not come to the end of such a story.
Details the story's thematic contents, and the meanings it conveys.
But most of all, enshittification is the result of anticircumvention law's ban on interoperability.
Core premise: enshittification is caused by anticircumvention bc you've nowhere to go. This is a deepening of his adversarial interoperability concept
But what if European firms want to go on taking advantage of anticircumvention laws? Well, there's good news there, too. "Good news," because the EU firms that rely on anticircumvention are engaged in the sleaziest, most disgusting frauds imaginable.
ah, right on time. On to EU firms counting on anticircumvention
It used to be that countries that depended on USAID had to worry about losing food, medical and cash supports if they pissed off America. But Trump killed USAID, so now that's a dead letter.
The erosion of all soft-power of the US is feeding into this
But crises precipitate change. Remember when another mad emperor – Vladimir Putin – invaded Ukraine, and Europe experienced a dire energy shortage? In three short years, the continent's solar uptake skyrocketed. The EU went from being 15 years behind in its energy transition, to ten years ahead of schedule.
True? Did we see acceleration in solar since early 2022, shortening the timeline with 10 yrs? What was the sched anyway?
When I was walking the picket line in Hollywood during the writer's strike, a writer told me that you prompt an AI the same way a studio boss gives shitty notes to a writer's room: "Make me ET, but make it about a dog, and give it a love interest, and a car-chase in the third act.
great quote
The thing is, software is not an asset, it's a liability. The capabilities that running software delivers – automation, production, analysis and administration – those are assets. But the software itself? That's a liability. Brittle, fragile, forever breaking down as the software upstream of it, downstream of it, and adjacent to it is updated or swapped out, revealing defects and deficiencies in systems that may have performed well for years.
software is a liability. Dutch equiv of this phrase? The assets are its impact : automation, production, analysis, admin
But there's one post-American system that's easy to imagine. The project to rip out all the cloud connected, backdoored, untrustworthy black boxes that power our institutions, our medical implants, our vehicles and our tractors; and replace it with collectively maintained, open, free, trustworthy, auditable code. This project is the only one that benefits from economies of scale, rather than being paralyzed by exponential crises of scale. That's because any open, free tool adopted by any public institution – like the Eurostack services – can be audited, localized, pen-tested, debugged and improved by institutions in every other country.
digital transition is possible because it scales through spreading. You don't have to solve exponential scale first.
But Eurostack is heading for a crisis. It's great to build open, locally hosted, auditable, trustworthy services that replicate the useful features of Big Tech, but you also need to build the adversarial interoperability tools that allow for mass exporting of millions of documents, the sensitive data-structures and edit histories.
Applaudes Eurostack but signals problem: you need to build adversarial interoperability to export all that is locked in current silos, and that is illegal as it takes anticircumvention. Not sure if that is true at scale though
This is exactly the kind of infrastructural risk that we were warned of if we let Chinese companies like Huawei supply our critical telecoms equipment. Virtually every government ministry, every major corporation, every small business and every household in the world have locked themselves into a US-based, cloud-based service.
Warning of Chinese intrusion but not seeing the US one etc
When the EU hit Apple with an enforcement order under the Digital Markets Act, Apple responded by offering to allow third party app stores, but it would only allow those stores to sell apps that Apple had approved of.
delaying tactic by Apple
As Jeff Bezos said to the publishers: "Your margin is my opportunity." With these guys, it's always "disruption for thee, but not for me." When they do it to us, that's progress. When we do it to them, it's piracy, and every pirate wants to be an admiral. Well, screw that. Move fast and break Tim Cook's things. Move fast and break kings!
n:: move fast and break bigtech
But there's a third possible response to tariffs, one that's just sitting there, begging to be tried: what about repealing anticircumvention law
if anticircumvention came to pass to avoid tariffs, why not abolish it as retaliation
The alternative was tariffs. Well, I don't know if you've heard, but we've got tariffs now!
US exported anticircumvention in their trade deals and won it to avoid tariffs. Doesn't say how that worked for the EU
every other country in the world has passed a law just like this in the years since. Here in the EU, it came in through Article 6 of the 2001 EU Copyright Directive.
art 6 of Copyright and Information Society Directive 2001/29
Note 2001/29 has been amended by Directive on Copyright in the Digital Single Market 2019/790, but I don't think in this aspect.
Today's links The Post-American Internet: My speech from Hamburg's Chaos Communications Congress. Hey look at this: Delights to delectate. Object permanence: Error code 451; Public email address Mansplaining Lolita; NSA backdoor in Juniper Networks; Don't bug out; Nurses whose shitty boss is a shitty app. Upcoming appearances: Where to find me. Recent appearances: Where I've been. Latest books: You keep readin' em, I'll keep writin' 'em. Upcoming books: Like I said, I'll keep writin' 'em. Colophon: All the rest. The Post-American Internet (permalink) On December 28th, I delivered a speech entitled "A post-American, enshittification-resistant internet" for 39C3, the 39th Chaos Communications Congress in Hamburg, Germany. This is the transcript of that speech. Video Playerhttps://archive.org/download/doctorow-39c3/39c3-1421-eng-A_post-American_enshittification-resistant_internet.mp400:0000:0001:01:12Use Up/Down Arrow keys to increase or decrease volume. Many of you know that I'm an activist with the Electronic Frontier Foundation – EFF. I'm about to start my 25th year there. I know that I'm hardly unbiased, but as far as I'm concerned, there's no group anywhere on Earth that does the work of defending our digital rights better than EFF. I'm an activist there, and for the past quarter-century, I've been embroiled in something I call "The War on General Purpose Computing." If you were at 28C3, 14 years ago, you may have heard me give a talk with that title. Those are the trenches I've been in since my very first day on the job at EFF, when I flew to Los Angeles to crash the inaugural meeting of something called the "Broadcast Protection Discussion Group," an unholy alliance of tech companies, media companies, broadcasters and cable operators. They'd gathered because this lavishly corrupt American congressman, Billy Tauzin, had promised them a new regulation – a rule banning the manufacture and sale of digital computers, unless they had been backdoored to specifications set by that group, specifications for technical measures to block computers from performing operations that were dispreferred by these companies' shareholders. That rule was called "the Broadcast Flag," and it actually passed through the American telecoms regulator, the Federal Communications Commission. So we sued the FCC in federal court, and overturned the rule. We won that skirmish, but friends, I have bad news, news that will not surprise you. Despite wins like that one, we have been losing the war on the general purpose computer for the past 25 years. Which is why I've come to Hamburg today. Because, after decades of throwing myself against a locked door, the door that leads to a new, good internet, one that delivers both the technological self-determination of the old, good internet, and the ease of use of Web 2.0 that let our normie friends join the party, that door has been unlocked. Today, it is open a crack. It's open a crack! And here's the weirdest part: Donald Trump is the guy who's unlocked that door. Oh, he didn't do it on purpose! But, thanks to Trump's incontinent belligerence, we are on the cusp of a "Post-American Internet," a new digital nervous system for the 21st century. An internet that we can build without worrying about America's demands and priorities. Now, don't get me wrong, I'm not happy about Trump or his policies. But as my friend Joey DaVilla likes to say "When life gives you SARS, you make sarsaparilla." The only thing worse than experiencing all the terror that Trump has unleashed on America and the world would be going through all that and not salvaging anything out of the wreckage. That's what I want to talk to you about today: the post-American Internet we can wrest from Trump's chaos. A post-American Internet that is possible because Trump has mobilized new coalition partners to join the fight on our side. In politics, coalitions are everything. Any time you see a group of people suddenly succeeding at a goal they have been failing to achieve, it's a sure bet that they've found some coalition partners, new allies who don't want all the same thing as the original forces, but want enough of the same things to fight on their side. That's where Trump came from: a coalition of billionaires, white nationalists, Christian bigots, authoritarians, conspiratorialists, imperialists, and self-described "libertarians" who've got such a scorching case of low-tax brain worms that they'd vote for Mussolini if he'd promise to lower their taxes by a nickel. And what's got me so excited is that we've got a new coalition in the War on General Purpose Computers: a coalition that includes the digital rights activists who've been on the lines for decades, but also people who want to turn America's Big Tech trillions into billions for their own economy, and national security hawks who are quite rightly worried about digital sovereignty. My thesis here is that this is an unstoppable coalition. Which is good news! For the first time in decades, victory is in our grasp.
Sees the original fight by digital rights activists now joined by geopolitical economics and international cybersec. Thinks this combi will win out
Capulet. O brother Montague, give me thy hand: This is my daughter's jointure, for no more Can I demand. Montague. But I can give thee more: For I will raise her statue in pure gold; 3275That while Verona by that name is known, There shall no figure at such rate be set As that of true and faithful Juliet.
Theme of Resolution. The two families finally make peace. The Capulets and Montagues end their feud. They'll build gold statues of Romeo and Juliet.
Friar Laurence. I will be brief, for my short date of breath Is not so long as is a tedious tale. 3205Romeo, there dead, was husband to that Juliet; And she, there dead, that Romeo's faithful wife: I married them; and their stol'n marriage-day Was Tybalt's dooms-day, whose untimely death Banish'd the new-made bridegroom from the city, 3210For whom, and not for Tybalt, Juliet pined. You, to remove that siege of grief from her, Betroth'd and would have married her perforce To County Paris: then comes she to me, And, with wild looks, bid me devise some mean 3215To rid her from this second marriage, Or in my cell there would she kill herself. Then gave I her, so tutor'd by my art, A sleeping potion; which so took effect As I intended, for it wrought on her 3220The form of death: meantime I writ to Romeo, That he should hither come as this dire night, To help to take her from her borrow'd grave, Being the time the potion's force should cease. But he which bore my letter, Friar John, 3225Was stay'd by accident, and yesternight Return'd my letter back. Then all alone At the prefixed hour of her waking, Came I to take her from her kindred's vault; Meaning to keep her closely at my cell, 3230Till I conveniently could send to Romeo: But when I came, some minute ere the time Of her awaking, here untimely lay The noble Paris and true Romeo dead. She wakes; and I entreated her come forth, 3235And bear this work of heaven with patience: But then a noise did scare me from the tomb; And she, too desperate, would not go with me, But, as it seems, did violence on herself. All this I know; and to the marriage
Plot Summary: Friar explains everything: the marriage, the potion plan, the failed letter.
Friar Laurence. I am the greatest, able to do least, Yet most suspected, as the time and place Doth make against me of this direful murder; 3200And here I stand, both to impeach and purge Myself condemned and myself excused.
Friar admits he's involved but will explain.
Give me the light: upon thy life, I charge thee, Whate'er thou hear'st or seest, stand all aloof, And do not interrupt me in my course. Why I descend into this bed of death, 2965Is partly to behold my lady's face; But chiefly to take thence from her dead finger A precious ring, a ring that I must use In dear employment: therefore hence, be gone: But if thou, jealous, dost return to pry 2970In what I further shall intend to do, By heaven, I will tear thee joint by joint And strew this hungry churchyard with thy limbs: The time and my intents are savage-wild, More fierce and more inexorable far 2975Than empty tigers or the roaring sea.
Romeo threatens Balthasar. Romeo is desperate and violent. He threatens to tear Balthasar apart if he follows.
Paris. Give me thy torch, boy: hence, and stand aloof: Yet put it out, for I would not be seen. 2935Under yond yew-trees lay thee all along, Holding thine ear close to the hollow ground; So shall no foot upon the churchyard tread, Being loose, unfirm, with digging up of graves, But thou shalt hear it: whistle then to me, 2940As signal that thou hear'st something approach. Give me those flowers. Do as I bid thee, go.
Paris at the tomb: Paris is mourning Juliet. He's being secretive.
The letter was not nice but full of charge Of dear import, and the neglecting it May do much danger. Friar John, go hence; 2920Get me an iron crow, and bring it straight Unto my cell.
Plot: The important letter. The letter explained Juliet wasn't really dead.
Apothecary. My poverty, but not my will, consents.
The apothecary only agrees because he's poor, not because he wants to since the law of Mantua forbids selling poison.
Romeo. Art thou so bare and full of wretchedness, And fear'st to die? famine is in thy cheeks, 2880Need and oppression starveth in thine eyes, Contempt and beggary hangs upon thy back; The world is not thy friend nor the world's law; The world affords no law to make thee rich; Then be not poor, but break it, and take this.
Romeo convinces the apothecary with the following argument. He says, since apothecary is so poor and starving, he should break the law for money.
Apothecary. Such mortal drugs I have; but Mantua's law Is death to any he that utters them.
Setting and the law then: Selling poison is punishable by death in Mantua.
'An if a man did need a poison now, Whose sale is present death in Mantua, 2860Here lives a caitiff wretch would sell it him.' O, this same thought did but forerun my need;
Plot: Romeo remembers a poor man who sells poison, which is illegal in Mantua.
[[Cory Doctorow p]] oped in Guardian, on US tech policy and enshittification. Points out that the US basically reneged on a deal (no tariffs if you allow our tech, but now we have the tariffs) so we can renege our part of it (circumvention laws). This aimed at UK audience, points out that that circumvention is inside a EU reg (art 6 of Copyright and Information Society Directive 2001) so it would be relatively easy after brexit to ditch the thing
Note 2001/29 has been amended by Directive on Copyright in the Digital Single Market 2019/790, but I don't think in this aspect.
The FEC critique the “supply-side” approach to growth, both asineffective in providing these elusive “good jobs” and in assum-ing aggregate GDP will “will lift all boats”. The FEC’s goal is toreorient social and economic policy away from a GDP-centred,individualised jobs-and-wages growth to a focus on guaranteedcollective liveability.It is disposable household income, rather than wages perse, that should provide a key metric.
Watch out with displacing definitions and invisibilising the most vulnerable. Who would get to say what accounts as "disposable"? What is "needed" insofar as to be counted only as undisposable: Shelter, food, education? I think I get, though, that all these basic undisposable needs, shall be given to everyone, collectively. But this is already in place as a right, just not enforced whatsoever...
How, then, do we reassert culture’s role in public policy?Many who seek to give culture a distinct function have beentempted to add it as a measure or priority alongside the “eco-nomic”. Advocacy such as the “fourth pillar” adds culture tothe “triple bottom line” of economic, social, and environmen-tal impact. But simply adding “culture” leaves “economy”unexamined, a “black box”.6 Like many such attempts, it isconstantly surprised when the real bottom line turns out to be“economy” after all. 7 In this sense, we need to challenge what“economy” actually entails.
"Economy" reeks of instrumentalism. Granted, it is needed for efficiency, to "save" lives, to "care" for the largest amount, but most of the time, it's not used in this particular efficiency way.
Clear headed explanation of DSA wrt Grok and X. Otoh in the case of Grok, I think the AI Act has a role to play, which is directly and more immediately connected to market access of Grok. It only takes into account the DSA, and the AIR isn't fully applicable yet, but still I think the entire framework needs to be taken into account. Incl the GDPR.
The Override Protocol You cannot out-argue the loop; you must overwrite it with Source Code.
This is the pivot. Everything changes when we spot the lie. If you're stuck here, use the counter-measure - it seems to easy to fix it, but it works!
Some of the buildings are new, so they don't have any employees in them yet, but we need to find some information about them regardless.
this is an example of growing independently:
new building can be without employees
You might see queries with these joins written as LEFT OUTER JOIN, RIGHT OUTER JOIN, or FULL OUTER JOIN, but the OUTER keyword is really kept for SQL-92 compatibility and these queries are simply equivalent to LEFT JOIN, RIGHT JOIN, and FULL JOIN respectively.
outer 关键字是和 SQL-92 兼容,等同于不写
When joining table A to table B, a LEFT JOIN simply includes rows from A regardless of whether a matching row is found in B. The RIGHT JOIN is the same, but reversed, keeping rows in B regardless of whether a match is found in A. Finally, a FULL JOIN simply means that rows from both tables are kept, regardless of whether a matching row exists in the other table.
left join right join full join
You might see queries where the INNER JOIN is written simply as a JOIN. These two are equivalent, but we will continue to refer to these joins as inner-joins because they make the query easier to read once you start using other types of joins,
inner join is join
When an ORDER BY clause is specified, each row is sorted alpha-numerically based on the specified column's value. In some databases, you can also specify a collation to better sort data containing international text.
by default it's sorted alpha-numerically
some db provide collation for better sorting for i18n text, but what?
_ Used anywhere in a string to match a single character (only with LIKE or NOT LIKE) col_name LIKE "AN_" (matches "AND", but not "AN")
_ is for one character used with like, not like
Windows XP vs Vista vs 7 vs 8.1 vs 10 vs 11 | Speed Test
Based on the "polish," "speed," and "stability" on older hardware as tested in the video:
Where bloody Tybalt, yet but green in earth,
Fear: Tybalt's fresh corpse will be in the tomb with her.
The horrible conceit of death and night, Together with the terror of the place,— 2590As in a vault, an ancient receptacle, Where, for these many hundred years, the bones Of all my buried ancestors are packed: Where bloody Tybalt, yet but green in earth, Lies festering in his shroud; where, as they say, 2595At some hours in the night spirits resort;— Alack, alack, is it not like that I, So early waking, what with loathsome smells, And shrieks like mandrakes' torn out of the earth, That living mortals, hearing them, run mad:— 2600O, if I wake, shall I not be distraught, Environed with all these hideous fears? And madly play with my forefather's joints? And pluck the mangled Tybalt from his shroud? And, in this rage, with some great kinsman's bone, 2605As with a club, dash out my desperate brains?
Gothic Horror: She imagines Tybalt's rotting body, ghosts, bones, and going mad.
Juliet. Ay, those attires are best: but, gentle nurse, I pray thee, leave me to myself to-night, 2550For I have need of many orisons To move the heavens to smile upon my state, Which, well thou know'st, is cross, and full of sin.
Plot: Juliet needs to be alone to take the potion. She lies about praying.
When presently through all thy veins shall run A cold and drowsy humour, for no pulse Shall keep his native progress, but surcease: No warmth, no breath, shall testify thou livest; The roses in thy lips and cheeks shall fade 2465To paly ashes, thy eyes' windows fall, Like death, when he shuts up the day of life; Each part, deprived of supple government, Shall, stiff and stark and cold, appear like death:
Description of the effects of the portion: She'll look dead: cold, no pulse, pale, stiff.
Things that, to hear them told, have made me tremble;
Juliet admits these ideas scare her, but she'd still do them.
Paris. God shield I should disturb devotion! Juliet, on Thursday early will I rouse ye: Till then, adieu; and keep this holy kiss.
Action: Paris kisses Juliet. She doesn't want it but must accept it.
Paris. Poor soul, thy face is much abused with tears.
Paris notices her crying face but misses her real pain.
Juliet. What must be shall be.
Fatalism: Juliet sounds resigned, but she's secretly planning to stop it.
Evolution tunes functional sub-state interconversion to boost enzyme function
This work offers a compelling view of the emergence of ceftazidime resistance in OXA-48 through conformational dynamics, framing adaptation as selection acting on rare, pre-existing catalytic behavior rather than the invention of new chemistry. A key observation is that adaptive mutations amplify an early, stochastic phase of ceftazidime turnover without changing steady-state k<sub>cat</sub>, supporting a phenotype-first view of enzyme evolution in which weak, probabilistic activity precedes genetic stabilization. The kinetic trade-offs observed across substrates are consistent with specialization within a constrained conformational landscape, and the ensemble-based interpretation provides a clear framework for linking dynamics, adaptation, and constraint.
A central implication of this framing is that productive sub-states must exist before selection acts, with adaptation increasing access to rare activity already present in the ensemble. Do you view the conformational ensemble that gives rise to these weak activities as fixed primarily by protein sequence, with the cellular environment acting mainly as a selector? Or could cellular conditions associated with antibiotic stress (such as altered proteostasis, macromolecular crowding, or chaperone engagement) transiently shift sub-state populations in vivo, changing both which weak activities are expressed and how strongly they translate into fitness early in adaptation? In this view, environmental context would shape not just the visibility of rare catalytic events, but their selective weight. Further, could such stress-associated conditions also influence which mutations are sampled or retained, for example by coupling altered protein dynamics to stress-induced mutagenesis or to differential tolerance of destabilizing variants?
Finally, you frame adaptation as tuning the rates of interconversion between pre-existing functional sub-states. In that view, do conformational dynamics simply provide a permissive background on which selection acts, or do they meaningfully bias which adaptive solutions are most accessible in the first place? More concretely, how would you distinguish a system where many evolutionary outcomes are equally reachable from one where the structure of the conformational ensemble makes certain trajectories more likely under specific substrates or stresses?
built their notebooks as simple web pages. The interface is missing Mathematica’s Steve Jobsian polish, and its sophistication. But by latching itself to the web, IPython got what is essentially free labor: Any time Google, Apple, or a random programmer open-sourced a new plotting tool, or published better code for rendering math, the improvement would get rolled into IPython. “It has paid off handsomely,” Pérez said.
Algo similar es lo que quiero capitalizar con Cardumem y luego portar a Grafoscopio, pues, como lo ha mostrado la experiencia con este último, las interfaces en Spec, el toolkit gráfico de Pharo, si bien brindan algunas cosas que las interfaces web no tienen, adolecen del basto ecosistema de ésta última y mantienen los documentos y la computación aisladas dentro de la imagen.
La web, por el contrario, es casi ubicua en términos de las tecnologías ya instaladas y así no se cuente con una conexión a internet en el equipo de cómputo, si este tiene una interfaz gráfica, muy seguramente contará con un naveador web. Y ahora que los sistemas hipermedia, hacen posible programar la web desde cualquier lenguaje (HOWL: Hypermedia On Whatever you Like), se puede aprovechar tanto lo que sabemos de los lenguajes/entornos que nos gustan (Pharo o Lua) como del amplio sistema de la web. Antes de 2023, que se popularizaron los sistemas hipermedia, teníamos que elegir entre lo uno y lo otro. Y yo deselegí activamente la web, debido al adefesio de JavaScript y lo engorroso del CSS. Hoy, las condiciones son bien distintas.
Out of the box, Python is a much less powerful language than the Wolfram Language that powers Mathematica. But where Mathematica gets its powers from an army of Wolfram Research programmers, Python’s bare-bones core is supplemented by a massive library of extra features—for processing images, making music, building AIs, analyzing language, graphing data sets—built by a community of open-source contributors working for free. Python became a de facto standard for scientific computing because open-source developers like Pérez happened to build useful tools for it; and open-source developers have flocked to Python because it happens to be the de facto standard for scientific computing. Programming-language communities, like any social network, thrive—or die—on the strength of these feedback loops.
A potential evolutionary explanation for the derived male cVA reduction in D. santomea was lacking until our unexpected discovery of a second sexual valence switch (and third pheromonal change in total), this time in the behavioral response to cVA by females. cVA promotes mating receptivity in D. melanogaster females (89, 130, 131) but has the opposite effect in D. santomea. The switch may be related to interspecific sexual dynamics between D. santomea and D. yakuba at their geographic boundary in São Tomé. D. yakuba males court D. santomea females vigorously (120, 121) and rare (sterile) F1 hybrid males can be collected within the sympatric zone (116–118). Thus, we speculate that the D. santomea female cVA valence switch arose due to ongoing selection pressure to maintain reproductive isolation from D. yakuba (122). If this explanation is correct, it would imply that D. santomea females use the high level of cVA on D. yakuba males as a sex- and species-identifying cue to ward off unwanted advances from heterospecific suitors (though we cannot exclude that D. yakuba courtship song is also used by D. santomea females for species discrimination) (61–63). This female aversion to cVA may have led in turn to reduced cVA production by D. santomea males under sexual selection pressure (140).However, two other evolutionary scenarios are also possible. D. santomea males may have initially reduced cVA abundance due to neutral drift or to pheromonally distinguish themselves from D. yakuba males (141), which in turn may have pressured D. santomea females to develop cVA aversion by sexual selection operating in the reverse direction. Alternatively, D. santomea female cVA aversion and male cVA reduction may have evolved simultaneously, potentially in genetic linkage (142). In any of these scenarios, the ultimate outcome would still be an elegant pheromonal mechanism which maintains efficient intraspecific mating in D. santomea while minimizing unfit hybridization with D. yakuba. Interestingly, a similar sexual reversal from cVA promoting to inhibiting female receptivity has been reported in one other drosophilid, the agricultural pest D. suzukii (143), whose males show fully eliminated cVA production (144). The evolutionary circumstances and selection pressures leading to D. suzukii cVA changes are unclear, however, as are potential consequences for intermale social behaviors.
Providing three independently compelling hypotheses for the evolutionary basis of this behavior is very much appreciated. Observation of these mating behaviors in their natural context (São Tomé) would both strengthen these hypotheses and may provide crucial ethological data to distinguish between them.
shown
it would be interesting to estimate the stoichiometry of each protein via the gel (obviously not the most quantitative, but you could get an estimate). Does the size of the complex off of the SEC suggest a 1:1:1 relationship? That would be a nice check that your are purifying what you think you are.
eluate
This could also be an issue of stoichiometry. If the interaction is strong, but the yield of expressed DDB1 for some reason was low, this could also be the case. It would be interesting to assay the amount of each DCAF bound to DDB1 versus free.
The treatment of serum-deprived wild-type NIH3T3 cells with MG132 resulted in an increased PCD (Fig. 3B) supporting our hypothesis. To test whether MG132 treatment affects PCD due to the inhibition of the ciliary proteasome or due to the inhibition of the overall cellular proteasomal activity, we treated serum-deprived Rpgrip1l-/-NIH3T3 cells with MG132. We found that the percentage of CC3-positive Rpgrip1l-/- NIH3T3 cells was not increased by the treatment with MG132
Curious what the ciliation levels are in MG132 treated serum starved cells in these experiments. There is a difference between ciliation percentage in the RPGRIP1l mutants vs wild type as in Figure 1C which also show a different percentage of CC3+ PCD in figure 1D but perhaps no difference in CC3+ PCD between the mutant and wild-type (RPGRIP1l +/+ w MG132 and RPGRIP1l -/- w MG132) in the presence of MG132 (though these are not shown on the same graph and perhaps not done in the same experiment?)
Free-swimming flagellated cells were also107observed, but couldn’t be identified further using optical microscopy
Very cool work! In Figure 3A, both algae from Clade A and Clade B appear to have flagella. Is it known whether symbiont motility/flagellar beating impacts host development? For instance, does it affect gastrocoel roof plate function during left-right asymmetry establishment? This would be interesting to explore, especially given the potential for both clades to colonize eggs in the same populations.
I can see why you can't just copy and paste the entire paper but I feel like you should be able to use previous findings, especially if you mention the paper you previously wrote.
I never thought this, but now I understand why
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Sheidaei et al., report how chromosomes are brought to positions that facilitate kinetochore-microtubule interactions during mitosis. The study focusses on an important early step of the highly orchestrated chromosome segregation process. Studying kinetochore capture during early prophase is extremely difficult due to kinetochore crowding but the team has taken up the challenge by classifying the types of kinetochore movements, carefully marking kinetochore positions in early mitosis and linking these to map their fate/next-positions over time. The work is an excellent addition to the field as most of the literature has thus far focussed on tracking kinetochore in slightly later stages of mitosis. The authors show that the PANEM facilitates chromosome positioning towards the interior of the newly forming spindle, which in turn facilitates chromosome congression - in the absence of PANEM chromosomes end up in unfavourable locations, and they fail to form proper kinetochore-microtubule interactions. The work highlights the perinuclear actomyosin network in early mitosis (PANEM) as a key spatial and temporal element of chromosome congression which precedes the segregation process.
Major points
1) The complexity of tracking has been managed by classifying kinetochore movements into 4 categories, considering motions towards or away from the spindle mid-plane. While this is a very creative solution in most cases, there may be some difficult phases that involve movement in both directions or no dominant direction (eg Phase3-like). It is unclear if all kinetochores go through phase1, 2, 3 and 4 in a sequential or a few deviate from this pattern. A comment on this would be helpful. Also, it may be interesting to compare those that deviate from the sequence, and ask how they recover in the presence and absence of azBB.
2) Would peripheral kinetochore close to poles behave differently compared to peripheral kinetochore close to the midplane (figure S4) ?In figure 3D, are they separated? If not, would it look different?
3) Uncongressed polar chromosomes (eg., CENPE inhibited cells) are known to promote tumbling of the spindle. In figure 5B with polar chromosomes, it will be helpful to indicate how the authors decouple spindle pole movements from individual kinetochore movements.
4) The work has high quality manual tracking of objects in early mitosis- if this would be made available to the field, it can help build AI models for tracking. The authors could consider depositing the tracking data and increasing the impact of their work.
Minor points
The current work builds upon their previous work, in which the authors demonstrated that an actomyosin network forms on the cytoplasmic side of the nuclear envelope during prophase. This work explains how the network facilitates chromosome capture and congression by tracking motions of individual kinetochores during early mitosis. The findings can be broadly useful for cell division and the cytoskeletal fields.
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
In this manuscript, Sheidaei et al. reported on their study of chromosome congression during the early stages of mitotic spindle assembly. Building on their previous study (ref. #15, Booth et al., Elife, 2019), they focused on the exact role of the actin-myosin-based contraction of the nuclear envelope. First, they addressed a technical issue from their previous study, finding a way to specifically impair the actomyosin contraction of the nuclear membrane without affecting the contraction of the plasma membrane. This allowed them to study the former more specifically. They then tracked individual kinetochores to reveal which were affected by nuclear membrane contraction and at what stage of displacement towards the metaphase plate. The investigation is rigorous, with all the necessary controls performed. The images are of high quality. The analyses are accurate and supported by convincing quantifications. In summary, they found that peripheral chromosomes, which are close to the nuclear membrane, are more influenced by nuclear membrane contraction than internal chromosomes. They discovered that nuclear membrane contraction primarily contributes to the initial displacement of peripheral chromosomes by moving them towards the microtubules. The microtubules then become the sole contributors to their motion towards the pole and subsequently the midplane. This step is particularly critical for the outermost chromosomes, which are located behind the spindle pole and are most likely to be missegregated.
While the conclusions are somewhat intuitive and could be considered incremental with regard to previous works, they are solid and improve our understanding of mitotic fidelity. The authors had already reported the overall role of nuclear membrane contraction in reducing chromosome missegregation in their previous study, as mentioned fairly and transparently in the text. However, the reason for this is now described in more detail with solid quantification. Overall, this is good-quality work which does not drastically change our understanding of chromosome congression, but contributes to improving it. Personally, I am surprised by the impact of such a small contraction (of around one micron) on the proper capture of chromosomes and wonder whether the signalling associated with the contraction has a local impact on microtubule dynamics. However, investigating this point is clearly beyond the scope of this study, which can be published as it is.
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Summary
Sheidaei and colleagues report a novel and potentially important role for an early mitotic actomyosin-based mechanism, PANEM contraction, in promoting timely congression of chromosomes located at the nuclear periphery, particularly those in polar positions. The manuscript will interest researchers studying cell division, cytoskeletal dynamics, and motor proteins. Although some data overlap with the group's prior work, the authors extend those findings by optimizing key perturbations and performing more detailed analyses of chromosome movements, which together provide a clearer mechanistic explanation. The study also builds naturally on recent ideas from other groups about how chromosome positioning influences both early and later mitotic movements.
In its current form, however, the manuscript is not acceptable for publication. It suffers from major organizational problems, an overcrowded and confusing Results section and figures, and a lack of essential experimental controls and contextual discussion. These deficiencies make it difficult to evaluate the data and the authors' conclusions. A substantial structural revision is required to improve clarity and persuasiveness. In addition, several key control experiments and more conceptual context are needed to establish the specificity and relevance of PANEM relative to other microtubule- and actin-based mitotic mechanisms. Testing PANEM in additional cell lines or contexts would also strengthen the claim. I therefore recommend Major Revision, addressing the structural, conceptual, and experimental issues detailed below.
Major Comments
The Results section is overly dense, lacks clear structure, and includes descriptive content that belongs in the Methods. Many figure panels should be moved to Supplementary Materials. A substantial reorganization is required to transform the manuscript into a focused, "Reports"-type article. - Move methodological and descriptive details (e.g., especially from the second Results subheading and Figure 2) to the Methods or Supplementary Materials. - Remove repetitive statements that simply restate that later phenotypes arise as consequences of delayed Phase 1 (applicable to subheadings 3 onward). - Figure 4I: This panel is currently unclear and should be drastically simplified. I recommend to reorganize figures as follows: - Figure I: Keep as single figure but simplify. Figure 1D and 1E could be combined, move unnormalized SCV to supplementary materials. Same goes for 1F. - New Figure 2: Combine current Figures 2A, 3A, 3C, 3D, 4C, 4F, and 4H to illustrate how PANEM contraction facilitates initial interactions of peripheral chromosomes with spindle microtubules which increases speed of congression initiation. - New Figure 3: Combine current Figures 5A, 5C, 5D, 5F, 6B, 6C, and lower panels of 4H to show how PANEM contraction repositions polar chromosomes and reduces chromosome volume in early mitosis to enable rapid initiation of congression. - New Figure 4: Combine Figures 7A, 7B, 7D, 7E, 7F, expanded Supplementary Figure S7, and new data to demonstrate that PANEM actively pushes peripheral chromosomes inward which is important for efficient chromosome congression in diverse cellular contexts. 2. Specificity and redundancy of actin perturbation
To establish the specificity and relevance of PANEM, the authors should include or discuss appropriate controls:
- Apply global actin inhibitors (e.g., cytochalasin D, latrunculin A) to disrupt the entire actin cytoskeleton. These perturbations strongly affect mitotic rounding and cytokinesis but only modestly influence early chromosome movements, as reported previously (Lancaster et al., 2013; Dewey et al., 2017; Koprivec et al., 2025). The minimal effect of global inhibition must be addressed when proposing a localized actomyosin mechanism. Comment if the apparent differences in this approach and one that the authors were using arises due to different cell types.
- Clarify why spindle-associated actin, especially near centrosomes, as reported in prior studies using human cultured cells (Kita et al., 2019; Plessner et al., 2019; Aquino-Perez et al., 2024), was not observed in this study. The Myosin-10 and actin were also observed close to centrosomes during mitosis in X.laevis mitotic spindles (Woolner et al., 2008). Possible explanations include differences in fixation, probe selection, imaging methods, or cell type. Note that some actin probes (e.g., phalloidin) poorly penetrate internal actin, and certain antibodies require harsh extraction protocols. Comment on possibility that interference with a pool of Myo10 at the centrosomes is important for effects on congression.
To strengthen the conclusions and broaden the study beyond the group's previous work, PANEM function should be tested in additional contexts (some may be considered optional but important for broader impact): - Test PANEM function in at least one additional cell line that displays PANEM to rule out cell-line-specific effects. - Examine higher-ploidy or binucleated cells to determine whether multiple PANEM contractions are coordinated and if PANEM contraction contributes more in cells of higher ploidies or specific nuclear morphologies. - Investigate dependency on nuclear shape or lamina stiffness; test whether PANEM force transmission requires a rigid nuclear remnant. - Analyze PANEM's contribution under mild microtubule perturbations that are known to induce congression problems (e.g., low-dose nocodazole). - Evaluate PANEM contraction role in unsynchronized U2OS cells, where centrosome separation can occur before NEBD in a subset of cells (Koprivec et al., 2025), and in other cell types with variable spindle elongation timing. - Quantify not only the percentage of affected cells after azBB but also the number of chromosomes per cell with congression defects in the current and future experiments. 4. Conceptual integration in Introduction and Discussion The manuscript should better situate its findings within the context of early mitotic chromosome movements: - Clearly state in the Introduction and elaborate in the Discussion that initiation of congression is coupled to biorientation (Vukušić & Tolić, 2025). This provides essential context for how PANEM-mediated nuclear volume reduction supports efficient congression of polar chromosomes. - Explain that PANEM is most critical for polar chromosomes because their peripheral positions are unfavorable for rapid biorientation (Barišić et al., 2014; Vukušić & Tolić, 2025). - Discuss how cell lines lacking PANEM (e.g., HeLa and others) nonetheless achieve efficient congression, and what alternative mechanisms compensate in the absence of PANEM. For example, it is well established that cells congress chromosomes after monastrol or nocodazole washout, which essentially bypasses the contribution of PANEM contraction.
Minor Comments
These issues are more easily addressable but will significantly improve clarity and presentation.
Introduction
Results (by subheading)
Discussion
Advance
This study's main strength is its novel and potentially important demonstration that contraction of PANEM, a peripheral actomyosin network that operates contracts early mitosis, contributes to the timely initiation of chromosome congression, especially for polar chromosomes. While PANEM itself was previously described by this group, this manuscript provides new mechanistic evidence, improved perturbations, and detailed chromosome tracking. To my knowledge, no prior studies have mechanistically connected this contraction to polar chromosome congression in this level of detail. The work complements dominant microtubule-centric models of chromosome congression and introduces actomyosin-based forces as a cooperating system during very early mitosis. However, the impact of the study is currently limited by major organizational issues, insufficient controls, and incomplete contextualization within existing literature. Addressing these issues will substantially improve clarity and credibility.
Audience
Primary audience of this study will be researchers working in cell division, mitosis, cytoskeleton dynamics, and motor proteins. The findings may interest also the wider cell biology community, particularly those studying chromosome segregation fidelity, spindle mechanics, and cytoskeletal crosstalk. If validated and clarified, the concept of PANEM could be integrated into textbooks and models of chromosome congression and could inform studies on mitotic errors and cancer cell mechanics.
Expertise
My expertise lies in kinetochore-microtubule interactions, spindle mechanics, chromosome congression, and mitotic signaling pathways.
This makes questions like “how fast is WebAssembly” a bit hard to answer. You don’t ask how fast algebraic notation is—it’s not a very sensible question. Taken in the context of something like JavaScript, the language is only as fast as the engine running it. JavaScript the language has no speed, but you can benchmark JS engines like V8, SpiderMonkey, and JavaScriptCore. You can benchmark the IO libraries of JS runtimes like Bun, Deno, and Node. What people actually mean is “how useful are the constructs of this language to efficient mappings of modern hardware” and “what is the current landscape of systems taking advantage of these constructs”.
Romeo. I fear, too early: for my mind misgives Some consequence yet hanging in the stars Shall bitterly begin his fearful date 610With this night's revels and expire the term Of a despised life closed in my breast By some vile forfeit of untimely death. But He, that hath the steerage of my course, Direct my sail! On, lusty gentlemen.
Foreshadowing: Romeo has a bad feeling, hence predicts something bad will start tonight and lead to his death.
Romeo. Give me a torch: I am not for this ambling; Being but heavy, I will bear the light.
Romeo is too sad to dance. "Ambling" means dancing.
Juliet. I'll look to like, if looking liking move: But no more deep will I endart mine eye Than your consent gives strength to make it fly.
Juliet is obedient but not enthusiastic. She'll look but not promise anything.
Nurse. Even or odd, of all days in the year, Come Lammas-eve at night shall she be fourteen. Susan and she—God rest all Christian souls!— Were of an age: well, Susan is with God; She was too good for me: but, as I said, 405On Lammas-eve at night shall she be fourteen; That shall she, marry; I remember it well. 'Tis since the earthquake now eleven years; And she was wean'd,—I never shall forget it,— Of all the days of the year, upon that day: 410For I had then laid wormwood to my dug, Sitting in the sun under the dove-house wall; My lord and you were then at Mantua:— Nay, I do bear a brain:—but, as I said, When it did taste the wormwood on the nipple 415Of my dug and felt it bitter, pretty fool, To see it tetchy and fall out with the dug! Shake quoth the dove-house: 'twas no need, I trow, To bid me trudge: And since that time it is eleven years; 420For then she could stand alone; nay, by the rood, She could have run and waddled all about; For even the day before, she broke her brow: And then my husband—God be with his soul! A' was a merry man—took up the child: 425'Yea,' quoth he, 'dost thou fall upon thy face? Thou wilt fall backward when thou hast more wit; Wilt thou not, Jule?' and, by my holidame, The pretty wretch left crying and said 'Ay.' To see, now, how a jest shall come about! 430I warrant, an I should live a thousand years, I never should forget it: 'Wilt thou not, Jule?' quoth he; And, pretty fool, it stinted and said 'Ay.'
The Nurse is talkative, funny, and loves telling stories about Juliet's childhood.
his pencil, and the painter with his nets; but I am sent to find those persons whose names are here 315writ, and can never find what names the writing person hath here writ. I must to the learned.—In good time.
Plot Device: This illiteracy causes the servant to ask Romeo for help, leading Romeo to the party.
But woo her, gentle Paris, get her heart,
Capulet is somehow progressive. He wants Juliet to consent to marriage.
Capulet. But saying o'er what I have said before: My child is yet a stranger in the world; She hath not seen the change of fourteen years, Let two more summers wither in their pride, 280Ere we may think her ripe to be a bride.
Capulet is protective. He thinks Juliet is too young to marry at 14.