2026-ban kapott friss doksit tettük be UG-ba. Ezért nem lett validálva.
10,000 Matching Annotations
- Feb 2026
-
clavis-nxt-user-guide-clavisnxt-granit-dev.apps.okd.dorsum.intra clavis-nxt-user-guide-clavisnxt-granit-dev.apps.okd.dorsum.intra
-
clavis-nxt-user-guide-clavisnxt-erste-dev.apps.okd.dorsum.intra clavis-nxt-user-guide-clavisnxt-erste-dev.apps.okd.dorsum.intra
-
2025-ben kapott friss doksit tettük be UG-ba. Ezért nem lett validálva.
-
-
clavis-nxt-user-guide-clavisnxt-erste-dev.apps.okd.dorsum.intra clavis-nxt-user-guide-clavisnxt-erste-dev.apps.okd.dorsum.intra
-
2025-ben kapott friss doksit tettük be UG-ba. Ezért nem lett validálva.
-
-
mirlo.space mirlo.space
-
Mirlo is a platform a la Bandcamp
-
-
www.biorxiv.org www.biorxiv.org
-
RRID:AB_2313606
DOI: 10.64898/2026.01.26.701850
Resource: (Vector Laboratories Cat# BA-1000, RRID:AB_2313606)
Curator: @scibot
SciCrunch record: RRID:AB_2313606
-
-
bpspubs.onlinelibrary.wiley.com bpspubs.onlinelibrary.wiley.com
-
RRID:SCR_023756
DOI: 10.1111/bph.70331
Resource: SwissTargetPrediction (RRID:SCR_023756)
Curator: @scibot
SciCrunch record: RRID:SCR_023756
-
-
noblogo.org noblogo.org
-
cada vez son menos las sociedades en las que importa la tradición oral
La tradición oral por mucho tiempo ha sido la principal manera de transmitir las historias y el conocimiento. En la época actual, como se menciona en el texto, esta práctica ha disminuido drásticamente con la llegada de la escritura y aún más con la llegada de las nuevas tecnologías. Me parece curioso pensar cómo se transmitía el conocimiento de manera oral. Las personas y la humanidad en general no solo dependían de sus habilidades o inteligencia, sino también de su memoria. El cerebro tiene una capacidad enorme para almacenar datos y procesar información, pero tiene ciertas limitaciones a la hora de reproducir la información o almacenarla más allá del periodo de vida de un individuo. Es por lo anterior que la escritura revolucionó por completo la manera en la que la tecnología avanza, el conocimiento ya no se construía a lo largo de los años, se acumulaba generación tras generación. Personalmente creo que esto es un arma de doble filo ya que se puede perder cierta capacidad cognitiva al no usar con la misma frecuencia o intensidad todas las conexiones neuronales que tenemos. Por otro lado, la tradición oral creo que es algo inherente a la especie humana. Por más tecnología que exista no vamos a dejar de contar historias o transmitir conocimientos de la misma manera, solo que la dinámica cambia un poco.
-
Todo estaría por ahí, en papel o en piedra, listo para consultarse cuando se nos diera la gana.
Esta frase me parece un resumen muy claro de lo que para mí es la inteligencia artificial. A través del texto y sus supervínculos, entiendo la IA como una gran base de datos siempre disponible, lista para ser consultada cuando se nos da la gana. En ese sentido, la inteligencia artificial no filtra la información ni distingue necesariamente entre lo verdadero y lo falso, sino que ofrece lo que tiene a su disposición. No importa quién produjo la información ni de dónde proviene, porque todo está ahí, accesible para ser consultado. Esta idea se complementa con el fragmento que plantea que la inteligencia artificial no es una tecnología utópica ni distópica, sino una tecnología más, con usos, aplicaciones y consecuencias, pero que no transformará a la sociedad de pies a cabeza. Desde esta perspectiva, la IA se parece a algo que alguien escribió alguna vez en una piedra, solo que hoy todos tenemos acceso a esas “piedras”, lo que hace que el conocimiento esté al alcance de cualquiera. Además, el texto me invita a reflexionar sobre mi propia relación con la inteligencia artificial. Me hace pensar que soy yo quien decide cómo usarla y con qué propósito, y que depende de mí si la convierto en una herramienta o en algo de lo que termino dependiendo. Sara Castillo Valderrama
-
-
geo.libretexts.org geo.libretexts.org
-
ay have experienced the impacts of less dense air at higher elevations. If you usually live, work, and recreate at lower elevations, you might find yourself getting short of breath while doing some of those same things at higher elevations. You may even get a headache or feel sluggish. This is because the molecules are further apart, resulting in air that is less dense, or “rarified”. Although you are filling your lungs, you still are not getting as many oxygen molecules in.
question 6
-
-
stylo.ecrituresnumeriques.ca stylo.ecrituresnumeriques.ca
-
es sites sont datés. Par exemple, le site Café, Cafés constituait à la fois une sorte de « teaser » pour annoncer l’exposition, mais faisait aussi le point sur « l’état de la connaissance liée au café »32 (Mireille Jacotin). Or, les connaissances sont datée
Suggestion pour éviter la répétition de « datés » : « ces sites sont datés. [...] Or, les connaissances/les savoir rapportés ne sont plus à jour, car [...] »
-
« Ce processus indique une approche du public par l’institution muséale qui n’est ni seulement communicationnelle (centrée sur la transmission du savoir), ni seulement économique (en termes de clientèles), mais va plutôt dans le sens de sa fonction au sein de l’espace public, pour un public qui va, en ce cas, visiter l’opposition comme on va voir un spectacle »
Il faudrait idéalement accompagner cette citation du numéro de page précis auquel on peut la trouver dans l'ouvrage.
-
-
www.youtube.com www.youtube.com
-
Analyse Approfondie du Conflit : Stratégies, Bénéfices et Psychologie
Résumé Exécutif
Ce document synthétise une analyse approfondie de la nature des disputes, s'éloignant de la perception traditionnelle du conflit comme étant purement négatif.
La thèse centrale est que le conflit, loin d'être un obstacle au bien-être, est un phénomène naturel et un moteur essentiel de développement personnel, relationnel et sociétal.
Son caractère constructif ou destructeur dépend entièrement de la manière dont il est géré.
Les points clés révèlent que la maîtrise du conflit repose sur la régulation émotionnelle, l'application de techniques de communication spécifiques et une volonté de remettre en question ses propres certitudes.
Les bénéfices d'une dispute bien menée sont multiples : elle permet * d'affirmer ses valeurs, de poser des limites, * de renforcer les liens en créant un sentiment d'appartenance et de confiance, * et même de stimuler l'excellence en milieu professionnel.
La gestion efficace des émotions, notamment le stress et la colère, est fondamentale.
Des stratégies comme la reconnaissance de ses émotions, le recadrage de la montée d'adrénaline en énergie positive et des techniques d'ancrage physique sont des outils puissants.
L'analyse met également en lumière l'influence déterminante des expériences de l'enfance sur notre rapport adulte au conflit, soulignant que des schémas de communication peuvent être consciemment modifiés.
Enfin, des méthodes concrètes pour désamorcer les tensions et dialoguer, même avec des interlocuteurs aux opinions radicalement opposées, sont présentées, insistant sur l'importance de présumer la bonne foi et de rechercher des solutions communes.
--------------------------------------------------------------------------------
1. La Perception Évolutive du Conflit
La compréhension psychologique du conflit a radicalement changé.
Si elle a longtemps prôné l'évitement des disputes, considérées comme nuisibles au bien-être, la perspective moderne est tout autre.
• Vision Traditionnelle vs. Moderne :
◦ Anciennement : La psychologie défendait l'idée qu'il fallait "éviter les conflits autant que possible".
Le bonheur était assimilé à l'absence de conflit.
◦ Aujourd'hui : Le conflit est considéré comme "tout à fait naturel".
Ce qui est déterminant n'est pas le conflit en soi, mais "la façon dont on se dispute".
• Le Conflit comme Moteur Social :
◦ Le sociologue Georg Simmel est cité pour affirmer que "le conflit est ce qui fait bouger une société".
◦ L'harmonie et le consensus total mènent à la stagnation : "quand on vient en harmonie et qu'on est tous du même avis, il ne se passe rien, tout se met à l'arrêt".
◦ Éviter systématiquement la confrontation ne résout pas les problèmes sous-jacents, qui continuent de "s'aggraver jusqu'à ce que plus rien ne puisse sauver le couple".
2. Les Multiples Bénéfices d'un Conflit Constructif
Lorsqu'il est abordé de manière saine, le conflit devient une source de force et de croissance à plusieurs niveaux.
• Développement Personnel :
◦ Affirmation de soi : Une dispute est une occasion "d'affirmer ses propres valeurs, poser ses limites et savoir qui on est".
Même sans solution, elle permet de "s'exprimer" et de "formuler son opinion à haute voix".
◦ Connaissance de soi : La confrontation peut mener à une meilleure connaissance de soi-même et des autres.
Andj, musicien de punk hardcore, témoigne qu'une "confrontation violente avec ses parents" lui a permis de mieux se connaître et de les voir sous un autre jour, améliorant considérablement leurs relations.
• Renforcement des Relations :
◦ Création de liens : Le conflit peut paradoxalement créer un "sentiment d'appartenance" lorsqu'on réalise qu'on est finalement d'accord avec l'autre sur certains points.
◦ Signe de confiance : Une première dispute avec une nouvelle connaissance peut "briser la glace".
Andj déclare : "je ne me dispute qu'avec les personnes qui comptent pour moi [...] si j'ai une confrontation avec quelqu'un, ça veut dire que je tiens à cette relation".
◦ Évolution commune : Le plus grand bonheur réside dans le fait que l'autre "n'a pas pris la fuite" et a surmonté l'épreuve ensemble, ce qui "nous a fait évoluer ensemble".
• Efficacité Professionnelle :
◦ En management, des équipes "un peu trop harmonieuses" où la critique constructive est absente n'atteignent qu'un "résultat moyen".
◦ Pour "atteindre l'excellence, on a besoin du conflit".
3. La Psychologie du Conflit : Maîtriser le "Tsunami Émotionnel"
La clé d'une dispute constructive réside dans la capacité à gérer le flux d'émotions intenses qu'elle génère.
• La Nature des Émotions en Conflit :
◦ La dispute est décrite comme un "grand tsunami émotionnel".
◦ Les émotions proviennent de trois sources :
1. L'objet direct de la dispute.
2. Les "émotions coptées" : stress ou frustration accumulés durant la journée.
3. Les émotions de la petite enfance.
• Stratégies de Régulation Émotionnelle :
◦ Reconnaissance et acceptation : La première étape est de reconnaître ses émotions ("oui je suis en colère et c'est légitime").
Le simple fait d'accepter et d'accueillir mentalement le stress "le réduit déjà de 30 %".
◦ Recadrage cognitif : Il est possible de "requalifier ce sentiment" et de voir la montée d'adrénaline comme un "surcroix d'énergie" positif.
Cette technique, enseignée en négociation, peut réduire la tension de 40 % supplémentaires.
◦ Ancrage physique : En cas de confusion mentale ou émotionnelle, il est important de "sentir le sol sous mes pieds" pour retrouver un ancrage.
◦ La pause stratégique : Proposer de "quitter la pièce" lorsque la discussion tourne en rond est "très efficace".
En 3 minutes, "le stress disparaît, la colère s'estompe".
◦ L'expression physique : Le cri, pratiqué par Andj, est présenté comme "une forme puissante d'expression de soi et un outil pour réguler ses émotions".
• Le Rôle Positif de la Colère :
◦ La colère n'est pas systématiquement négative. Hasnain Kazim, écrivain, déclare : "j'aime la colère, je trouve qu'elle a quelque chose d'extrêmement direct.
Elle dit clairement : ça me va, ça ça ne me convient pas".
◦ Il la préfère à l'agressivité passive : "ce que je trouve horrible, c'est quand tu vois bien que tu as vexé une personne [...] et qu'elle ne dit rien".
4. Techniques Pratiques pour une Communication Efficace en Situation de Conflit
Des outils rhétoriques et des approches spécifiques peuvent transformer une querelle en un dialogue productif.
| Technique | Description | Exemple / Citation | | --- | --- | --- | | Éviter les Généralisations | Les mots comme "jamais" ou "toujours" enferment l'autre dans une position inconfortable et ferment le dialogue. Il faut "débarrasser le dialogue de ces mots". | "Il est toujours de mauvaise humeur." | | Utiliser le "Je" | Remplacer le "tu accusateur" par des formulations commençant par "je" pour désamorcer l'agressivité et prendre la responsabilité de sa propre perception. | Au lieu de "Tu ne m'as pas compris", dire "Je me suis mal exprimé". | | Formuler son Point de Vue | Argumenter en commençant par "de mon point de vue" pour éviter de présenter son opinion comme une vérité absolue et agressive. | "De mon point de vue, si je dis à l'autre c'est toi qui est stressé, ça ne va faire qu'attiser le débat." | | Poser des Questions Ouvertes | Utiliser des questions qui commencent par "pourquoi", "comment" ou "qu'est-ce que" pour encourager une réponse développée et maintenir la discussion ouverte. | "Qu'est-ce que tu veux dire exactement ?" | | Utiliser les Questions Fermées | Poser des questions auxquelles on ne peut répondre que par "oui", "non" ou "peut-être" pour "fixer quelque chose qui n'était pas clair". | "Ça, ça te dérange ? - Oui." | | Désamorcer les Attaques Déloyales | Identifier les techniques rhétoriques visant à déstabiliser (ex: ad hominem), puis les neutraliser en recentrant calmement le débat sur le sujet principal. | Attaque : "Ça ne vous dérange pas de passer à la télé avec la même veste qu'avanthière ?" Réponse : "Bon, à part la couleur de la veste \[...\] comment peut-on faire pour avancer sur notre sujet ?" |
5. Le Conflit Idéologique : Dialoguer avec l'Opposition
Le journaliste Hasnain Kazim partage son expérience sur la nécessité et la méthode pour engager le dialogue avec des personnes aux opinions radicalement différentes, notamment politiques.
• Principes Fondamentaux :
◦ Quitter sa zone de confort : Il faut être prêt à "remettre en question ses propres certitudes".
◦ Éviter l'étiquetage : Ne pas immédiatement ranger l'autre dans un camp ("droitard", "réac", "gauchiste") pour ne pas le considérer comme un "ennemi" et couper l'échange.
◦ Présomption de bonne foi : Partir du principe que "l'autre ne me tend pas de piège" et qu'il est "honnête et sincère".
Si ses propos semblent contradictoires, au lieu de le juger "stupide", il faut chercher à comprendre "ce qu'il y a derrière".
◦ Courtoisie et absence de condescendance : Ne pas arriver en "donneur de leçon" ou en "maître d'école".
• Stratégies d'Engagement :
◦ L'importance d'initier le dialogue : Le plus important est "déjà de le faire, tout simplement".
◦ Se fixer des limites : Il est crucial de savoir se protéger et de "riposter" si nécessaire.
On n'est pas obligé de "témoigner de l'empathie à tout le monde".
◦ Les effets à retardement : Même si un débat semble échouer sur le moment, il peut avoir des "effets rétroactifs".
L'interlocuteur peut plus tard réfléchir au courage de la démarche, ce qui peut faire évoluer sa pensée.
6. L'Héritage de l'Enfance et son Impact sur le Conflit
Notre manière de gérer les disputes à l'âge adulte est profondément façonnée par les modèles et les expériences de notre enfance.
• L'Apprentissage par l'Exemple :
◦ La relation à la dispute est "héritière de ce que nous ont montré nos modèles d'identification" (parents, figures d'autorité).
◦ Si un enfant apprend qu'il est aimé lorsqu'il est "passif", "discret" et "d'accord avec tout le monde", il évitera la confrontation à l'âge adulte.
• Exemple d'Andj :
◦ Il a grandi dans une famille où les disputes étaient gérées par des cris et parfois des "agressions physiques".
◦ Ses parents "réprimaient leurs propres besoins", générant une "énorme frustration".
◦ Enfant, il essayait de se "rendre invisible".
◦ La musique punk lui a permis de réaliser qu'il n'était pas seul, en entendant d'autres "hurler cette injustice".
◦ Le processus de guérison a nécessité plusieurs années de confrontations directes avec ses parents, où il a pu "verbaliser tout ce qui [l]'avait blessé", ce qui a finalement permis de rétablir une relation saine.
7. La Résolution : Clore la Dispute de Manière Constructive
Une bonne dispute doit avoir une bonne conclusion pour que ses effets soient bénéfiques.
• Rituels de Réconciliation : Des rituels comme "la bise de réconciliation" ou une poignée de main sont particulièrement efficaces, notamment avec les enfants, pour clore un conflit.
• Règles Familiales : Établir des règles claires, comme celle de ne pas "emporter la dispute au lit", aide à contenir le conflit et à préserver la relation.
• Savoir Ne Pas Disputer : Parfois, la dispute "n'en vaut pas la peine".
Lors de retrouvailles familiales, la préservation des liens peut être "plus importante que le sujet qui vient de surgir".
• Le Piège de l'Empathie pour les Spectateurs : Lors d'une dispute de groupe, les spectateurs doivent se méfier de l'empathie, qui peut les transformer en "partie prenante", leur faisant voir le débat d'un seul côté et transformant les autres en "adversaires".
Tags
Annotators
URL
-
-
-
Met de new competition tool (NCT), ook omschreven in het beroemde Draghi report (pagina 302), is het mogelijk bedrijven bindende aanwijzingen te geven als er effectief geen concurrentie mogelijk is. Als voorbeeld, als het praktisch onmogelijk is om te concurreren met Google of Microsoft voor email en agendabeheer kan je hiermee een bindende aanwijzing geven dat deze bedrijven het mogelijk moeten maken om als nieuwe speler mee te doen in hun ecosysteem. Zo’n verplichting is er nu soms ook al, maar alleen onder hele specifieke omstandigheden. Met de NCT kan het ook onder de algemene voorwaarde dat er van een niet-concurrerende markt sprake is.
ACM krijgt beschikking over 'new competition tool' NCT a la Draghi , bindende aanwijzing nieuwe spelers toe te laten.
-
Geen enkele verwijzing naar de eerdere Nederlandse Digitaliseringsstrategie (NDS), of het daaruit voortvloeiende beloofde Overheidsbrede Cloudbeleid - opmerkelijk
Dit is heel vreemd ja. NDS is er nog maar net. Vgl [[Nathan Ducastel p]] NDS gaat ook over het digihuishouden van de overheid (maar incl decentraal: is dat het verschil?)
-
-
-
Synthèse sur les Systèmes d'Émulation en Milieu Scolaire
Résumé Exécutif
Ce document synthétise les principes, les applications et les conditions d'efficacité des systèmes d'émulation (ou systèmes de renforcement) en contexte scolaire, basés sur l'expertise de Nancy Goudreau, docteure en psychopédagogie.
L'utilisation de ces systèmes vise à instaurer ou à renforcer des comportements spécifiques par le biais d'une motivation externe.
L'idée centrale est que ces outils doivent être employés de manière ciblée, judicieuse et temporaire. Ils sont principalement indiqués pour les élèves présentant des difficultés de comportement importantes et pour qui la motivation intrinsèque est faible.
Une erreur fréquente est de les appliquer à des élèves déjà motivés, ce qui risque de substituer une motivation externe à une motivation autonome.
Le succès d'un système d'émulation repose sur des conditions strictes : le choix d'un renforçateur perçu comme agréable par l'élève, l'application immédiate du renforçateur après le comportement (contiguïté), et un lien de cause à effet clair pour l'élève (contingence).
Il est crucial de planifier le retrait progressif du système dès sa mise en place, en parallèle d'un enseignement explicite des compétences visées.
Le système en lui-même n'enseigne rien ; il ne fait que motiver.
Il existe trois types de systèmes : indépendant (individuel), dépendant (la récompense du groupe dépend d'un individu) et interdépendant (défi collectif).
Le système interdépendant est le plus recommandé pour une classe ordinaire afin de relever un défi de groupe précis.
Finalement, il est essentiel de distinguer le système d'émulation, qui est un contrat "si... alors...", de la récompense spontanée, qui est bien plus efficace pour reconnaître la bonne conduite de la majorité des élèves et demande moins d'énergie à l'enseignant.
Définition et Objectifs du Système d'Émulation
Un système d'émulation, aussi appelé système de renforcement, est une intervention structurée mise en place pour encourager un comportement désiré. Son objectif est double :
1. Faire apparaître un comportement qui est actuellement absent chez un élève.
2. Augmenter la fréquence ou la pertinence d'un comportement déjà présent mais manifesté de façon insuffisante ou dans des contextes inappropriés.
Le mécanisme repose sur l'utilisation d'une motivation extrinsèque (externe) pour amener l'élève à adopter le comportement.
Un "renforçateur" est présenté à l'élève immédiatement après la manifestation du comportement attendu.
Il existe deux types de renforcement :
• Renforcement positif : L'ajout d'un stimulus agréable suite au comportement (ex: obtenir un autocollant, un privilège).
• Renforcement négatif : Le retrait d'un stimulus désagréable suite au comportement (ex: être exempté d'une corvée).
Il est important de noter que le terme "négatif" se réfère au retrait et non à une connotation péjorative.
Principes Fondamentaux et Conditions d'Efficacité
L'efficacité d'un système d'émulation n'est pas automatique.
Elle dépend du respect rigoureux de plusieurs principes fondamentaux et de l'évitement de certaines erreurs courantes.
Quand Utiliser un Système d'Émulation ?
Le recours à un système d'émulation devrait être réservé à des contextes précis :
• Lorsqu'un comportement attendu est totalement absent et qu'il faut initier son apparition.
• Avec des élèves présentant des difficultés de comportement importantes, pour qui la motivation à adopter les conduites attendues est très faible.
• Pour contrecarrer des renforçateurs sociaux qui maintiennent des comportements indésirables (ex: un élève qui fait rire la classe).
Le système vise alors à offrir une réponse positive plus forte, associée au comportement attendu.
Les Erreurs à Éviter
L'utilisation inadéquate des systèmes d'émulation peut être contre-productive. Les erreurs les plus fréquentes sont :
1. Utiliser le système avec des élèves déjà motivés : Appliquer un renforçateur externe à un élève qui agit déjà pour les "bonnes raisons" (motivation autonome) risque de déplacer sa motivation.
L'élève commencera à adopter le comportement non plus par plaisir ou par conviction, mais pour obtenir la récompense.
2. Appliquer un système à toute la classe de manière indifférenciée : Souvent, seuls un ou deux élèves ont réellement besoin d'un tel système.
L'appliquer à tous est une source de gestion lourde pour un bénéfice faible, et il est souvent inefficace pour les élèves qui en ont le plus besoin.
3. Omettre de planifier le retrait du système : Un système d'émulation est une mesure temporaire.
S'il n'y a pas de plan pour le retirer progressivement, l'enseignant et les élèves deviennent "pris dans le système", ce qui mène à un cycle où il faut constamment trouver de nouveaux systèmes pour maintenir la motivation.
Les Clés du Succès
Pour qu'un système soit efficace, plusieurs conditions doivent être réunies :
• Choix du Renforçateur : Le renforçateur doit être perçu comme agréable et motivant du point de vue de l'élève.
Un renforçateur choisi par l'adulte sans consultation peut même avoir un effet punitif (ex: un dîner avec l'enseignante pour un adolescent).
Il est donc primordial de consulter les élèves.
• Principe de Contiguïté : Le délai entre le comportement et l'obtention du renforçateur doit être le plus court possible, surtout avec les jeunes enfants.
Un renforçateur reçu le vendredi pour un comportement du mardi n'a aucun effet, car l'association ne se fait pas.
• Principe de Contingence : L'élève doit clairement et systématiquement associer le comportement spécifique avec la conséquence agréable.
• Enseignement Explicite de la Compétence : C'est un point crucial souvent négligé.
"Ça n'enseigne rien ce système-là".
Le système motive, mais il n'enseigne pas comment gérer sa colère, résoudre un conflit ou composer avec la défaite. Il doit impérativement être accompagné d'un enseignement explicite et d'un accompagnement dans le développement de la compétence visée.
• Planification du Retrait : L'objectif ultime est de développer la motivation autonome de l'élève.
L'adulte doit donc activement l'aider à prendre conscience des avantages et du plaisir associés au nouveau comportement pour que celui-ci se maintienne sans renforçateur externe.
La durée d'utilisation doit être "le plus court possible".
Typologie des Renforçateurs et des Systèmes
Les Types de Renforçateurs
• Renforçateurs Sociaux : Ce sont les renforçateurs les plus puissants pour maintenir durablement les comportements.
Ils incluent les rétroactions positives, les félicitations chaleureuses et sincères, les encouragements, etc.
Ils devraient être utilisés constamment, avec ou sans système formel.
Un système purement "comptable", sans chaleur humaine, est peu efficace.
• Renforçateurs Tangibles : Ils incluent les points, autocollants, privilèges, etc.
Ils sont utiles pour amorcer un changement, mais la transition vers les renforçateurs sociaux puis vers la motivation intrinsèque doit être planifiée.
L'apprentissage de la propreté est une analogie parfaite : on commence avec des renforçateurs tangibles (ex: un bonbon) car l'enfant n'a aucune motivation autonome, puis on transitionne rapidement vers des renforçateurs sociaux (bravos, fierté, appel aux grands-parents) jusqu'à ce que l'enfant devienne autonome.
Les Trois Groupes de Contingence
Il existe trois manières d'organiser un système d'émulation en fonction du groupe.
| Type de Système | Description | Usage Recommandé | Avantages et Inconvénients | | --- | --- | --- | --- | | Groupe Indépendant | Chaque élève travaille pour sa propre récompense ("chacun pour soi"). | Pour un ou deux élèves ayant des difficultés spécifiques, dans le cadre d'un plan d'intervention. | Avantage: Entièrement personnalisé.<br>Inconvénient: "Le moins bon choix" lorsqu'appliqué à toute une classe. Lourd à gérer, inefficace pour les élèves en difficulté et démobilisateur. | | Groupe Dépendant | La récompense de toute la classe dépend de la réussite d'un ou de quelques élèves. | Très efficace pour les élèves avec des troubles du comportement, pour mobiliser le soutien des pairs. | Avantage: Le soutien social des pairs devient un puissant levier d'aide.<br>Inconvénient: Nécessite une bonne préparation de la classe pour éviter de blâmer l'élève en cas d'échec. | | Groupe Interdépendant | La classe travaille collectivement à l'atteinte d'un objectif commun pour une récompense collective ("tous pour un, un pour tous"). | La meilleure approche pour une classe ordinaire afin de relever un défi de groupe ponctuel et précis. | Avantage: Facile à gérer, soutient un climat de coopération, inclut tous les élèves.<br>Inconvénient: Doit cibler un seul comportement précis pour être efficace. |
Conclusions et Recommandations Pratiques
Faut-il les Utiliser au Primaire ?
La réponse est nuancée : uniquement si c'est vraiment nécessaire. La plupart des classes fonctionnent bien sans système d'émulation formel.
• Pour un élève avec des besoins importants, un système indépendant sur mesure et intégré à son plan d'intervention est justifié et peut être efficace.
• Pour un défi de groupe ponctuel (ex: un relâchement dans le calme des déplacements), un système interdépendant peut être une solution efficace et temporaire.
Il doit cibler un seul comportement, et non devenir un "melting pot" de toutes les règles de la classe.
• L'utilisation d'un système indépendant pour tous les élèves d'une classe représente "énormément de temps et d'énergie pour peu de résultats".
La Distinction Cruciale : Système d'Émulation vs. Récompense
Il est fondamental de différencier ces deux concepts :
• Système d'Émulation : Un contrat préétabli. L'élève sait à l'avance que s'il produit le comportement X, il obtiendra la récompense Y. (Ex: "Si vous travaillez bien, je vous donnerai 15 minutes de plus dehors.")
• Récompense : Une reconnaissance spontanée et non annoncée d'un bon comportement. (Ex: "Vous avez tellement fait une belle journée que j'ai envie de vous récompenser, on va aller profiter du soleil 15 minutes.")
Pour la grande majorité des élèves, qui ont déjà une motivation autonome, la récompense spontanée est beaucoup plus efficace pour reconnaître leur bonne conduite, tout en étant infiniment moins énergivore pour l'enseignant que la gestion d'un système formel.
Ces élèves ont avant tout besoin "d'encouragement, de félicitations, de tapes dans le dos".
-
-
www.youtube.com www.youtube.com
-
Document d'Information : Les Fonctions Exécutives chez l'Enfant
Résumé Exécutif
Ce document synthétise les perspectives de Julie Beaulieu, docteure en psychopédagogie, sur les fonctions exécutives chez l'enfant.
Considérées comme le « chef d'orchestre » ou la « tour de contrôle » du cerveau, ces habiletés cognitives de haut niveau, situées dans le lobe frontal, sont fondamentales pour la régulation des pensées, des émotions et des comportements.
Elles permettent à l'individu de s'adapter à son environnement et d'atteindre des objectifs.
Leur développement s'étend sur une longue période, débutant dès les premiers mois de la vie pour atteindre leur plein potentiel vers l'âge de 25 ans, avec des phases de progression accélérée durant la période préscolaire (3-7 ans) et au début de l'adolescence.
Si la maturation cérébrale est un facteur essentiel, l'environnement et la diversité des expériences vécues par l'enfant jouent un rôle crucial pour « aiguiser » ces fonctions.
Quatre composantes principales sont identifiées : la mémoire de travail, l'inhibition, la flexibilité cognitive et la planification.
Ces fonctions, bien que distinctes, sont interdépendantes et fonctionnent en synergie, à l'image d'une « toile d'araignée ».
Elles sont la fondation de nombreuses aptitudes et ont un impact direct sur la réussite éducative et le développement global de l'enfant (social, langagier, moteur).
Le soutien parental est déterminant et s'intègre dans les activités quotidiennes.
Les stratégies efficaces incluent le jeu, les interactions sociales, le questionnement (notamment le « pourquoi »), l'offre de choix, la stimulation de l'autonomie et de la créativité, ainsi que la mise en place d'un environnement sécurisant et positif.
L'objectif n'est pas d'entraîner ces fonctions comme un muscle, mais de les mobiliser à travers des expériences ludiques et variées.
--------------------------------------------------------------------------------
1. Définition et Nature des Fonctions Exécutives
Les fonctions exécutives sont définies comme un ensemble de processus cognitifs de haut niveau localisés dans le lobe frontal du cerveau.
Elles sont essentielles pour la réalisation de tâches complexes et pour l'adaptation de l'individu à son environnement.
Leur rôle principal est de réguler trois aspects fondamentaux du comportement humain :
• Les pensées : Organiser ses idées, raisonner, réfléchir.
• Les comportements : Contrôler ses actions, initier des tâches, s'ajuster.
• Les émotions : Gérer ses réactions émotionnelles.
Ces fonctions sont mobilisées en continu dans la vie quotidienne, dès qu'une tâche n'est pas automatique ou routinière et demande de réfléchir, raisonner, anticiper, planifier ou diriger son attention.
La Métaphore du Chef d'Orchestre
Pour illustrer leur rôle, Julie Beaulieu utilise deux analogies principales :
• La tour de contrôle : Elles permettent de se contrôler et de diriger ses actions.
• Le chef d'orchestre : C'est la métaphore la plus développée.
Les fonctions exécutives agissent comme le chef d'orchestre du cerveau, coordonnant l'ensemble des autres habiletés cognitives (les "instruments") pour produire un comportement harmonieux et adapté à un objectif précis.
Elles dirigent les pensées, les émotions et les comportements pour permettre à l'individu de s'adapter et de répondre efficacement aux exigences de son environnement.
2. Le Développement des Fonctions Exécutives
Le développement des fonctions exécutives est un processus long et progressif, influencé par la maturation biologique et les expériences environnementales.
Chronologie du Développement
• Début : Les composantes des fonctions exécutives commencent à se développer dès les premiers mois de la vie.
• Maturité : Elles atteignent leur plein potentiel au début de l'âge adulte, vers 25 ans.
• Périodes Clés : Deux périodes sont identifiées comme particulièrement propices à une progression importante :
1. L'âge préscolaire (3 à 6-7 ans). 2. Le début de l'adolescence.
Il est crucial de noter que le rythme de développement varie d'un enfant à l'autre, en fonction de facteurs individuels comme la maturation de leur propre cerveau.
Le Rôle Crucial de l'Environnement
La maturation du cerveau est une composante essentielle, mais l'environnement dans lequel l'enfant évolue est un moteur fondamental du développement.
• L'expérience : C'est principalement à travers les expériences vécues que les enfants mobilisent et développent leurs fonctions exécutives.
• L'aiguisage : Plus les fonctions sont mobilisées, plus elles s'« aiguisent », devenant « sophistiquées et spécialistes ».
• La variété : Proposer des contextes et des expériences variés permet de mobiliser un plus large éventail de fonctions exécutives et de favoriser un développement plus complet.
Le soutien offert par l'entourage (parents, éducateurs) est donc primordial.
3. Contextes Favorables au Développement
Certains contextes et situations sont particulièrement efficaces pour mobiliser et renforcer les fonctions exécutives, car ils empêchent l'enfant de recourir à des automatismes.
• Situations nouvelles : Toute situation où l'enfant n'a pas de routine établie l'oblige à s'adapter et donc à mobiliser ses fonctions exécutives.
• Tâches complexes : Les activités qui demandent plusieurs étapes, actions ou une réflexion soutenue.
• Situations inattendues : Les imprévus qui forcent une réévaluation et une adaptation.
• Résolution de problèmes : Chercher des solutions à un conflit ou à une difficulté pratique.
• Le jeu : Un contexte extrêmement favorable, en particulier :
◦ Le jeu symbolique (« faire semblant ») : Se mettre dans la peau d'un personnage.
◦ Les jeux de société : Qui demandent de la stratégie, de la mémoire et de l'anticipation.
• Interactions sociales : Les discussions, les échanges et les conversations demandent une réflexion constante, une anticipation des réponses et une adaptation à l'interlocuteur.
• Découvertes et nature : Explorer de nouveaux environnements.
• Projets à long terme : Les projets qui s'étalent sur plusieurs jours ou semaines et nécessitent de la planification.
• Environnement stimulant et sécurisant : Un climat familial positif, où l'enfant se sent en sécurité, est plus propice à la mobilisation des fonctions exécutives.
Un stress élevé ou des émotions négatives intenses peuvent entraver leur fonctionnement.
4. Les Quatre Composantes Principales des Fonctions Exécutives
L'exposé se concentre sur quatre composantes fondamentales, qui sont interdépendantes et fonctionnent comme une « toile d'araignée ».
| Composante | Description | Exemple Clé | | --- | --- | --- | | Mémoire de travail | Capacité à retenir temporairement et à manipuler l'information pour l'utiliser dans une tâche. Elle fait le pont entre la mémoire sensorielle et la mémoire à long terme, et filtre les informations pertinentes. C'est l'aspect "actif" ou "dynamique" de la mémoire. | Le calcul mental (ex: 52 + 48). Il faut retenir les chiffres et activement les additionner pour trouver le résultat. | | Inhibition | Capacité à résister aux impulsions, aux distractions et aux habitudes pour adopter un comportement plus approprié. C'est le « frein » du cerveau qui permet de prendre un temps de recul avant d'agir et de diriger son attention. | Un jeune enfant à l'épicerie qui s'empêche de faire un commentaire à voix haute sur l'apparence physique d'une personne. | | Flexibilité cognitive | Capacité à se désengager d'une tâche ou d'une perspective pour s'adapter à une nouvelle situation, un nouveau point de vue ou de nouvelles règles. Elle permet de changer de stratégie quand la première ne fonctionne pas. | Un enfant qui accepte d'arrêter son jeu vidéo pour venir souper en famille, se désengageant cognitivement de la première activité pour se réengager dans la seconde. | | Planification | Capacité à anticiper des événements futurs, à se fixer un but et à organiser une séquence d'actions pour l'atteindre. Son développement est plus tardif, car il dépend de l'acquisition de la notion du temps. | Prévoir le trajet pour aller au parc, ou suivre les étapes d'une recette de cuisine. |
5. L'Importance Fondamentale des Fonctions Exécutives
Les fonctions exécutives sont décrites comme des compétences de base, constituant la fondation de multiples aptitudes et habiletés. Leur importance est capitale car :
• Elles soutiennent la réussite éducative à tous les niveaux (primaire, secondaire, et même post-secondaire).
• Elles ont un impact sur l'ensemble des sphères de développement de l'enfant : langagier, social, cognitif et même moteur.
• Elles sont essentielles pour fonctionner efficacement dans la vie de tous les jours et pour le bien-être futur de l'individu.
6. Stratégies de Soutien Parental pour le Développement
Les parents peuvent jouer un rôle actif dans le développement des fonctions exécutives de leur enfant à travers des actions intégrées au quotidien, sans nécessiter de matériel sophistiqué.
Approches Générales
• Développer le langage intérieur : Verbaliser à voix haute ses propres pensées pour modéliser, puis encourager l'enfant à se parler dans sa tête pour retenir des consignes ou se réguler.
• Soutenir les habiletés sociales : Favoriser les interactions sociales positives et variées.
• Offrir un soutien émotionnel : Accueillir les émotions de l'enfant avec empathie pour créer un climat de sécurité.
• Pratiquer des activités sportives et artistiques : Ces activités mobilisent l'ensemble des fonctions exécutives.
• Utiliser l'étayage et le questionnement : Accompagner l'enfant et le questionner sur ce qu'il pense, et surtout pourquoi il le pense, pour l'amener à verbaliser son raisonnement.
• Offrir des choix : Faire un choix implique de prioriser et de renoncer, ce qui mobilise la réflexion.
• Favoriser l'autonomie : Encourager l'enfant à faire les choses par lui-même.
• Tenir compte de ses intérêts : Un contexte agréable et motivant est plus propice à la mobilisation cognitive.
• Proposer des défis : Offrir des défis légèrement supérieurs à ses capacités actuelles, tout en l'accompagnant.
Stratégies Ciblées par Composante
Pour la Mémoire de Travail
• Stratégies de mémorisation : Utiliser des images mentales, la répétition (avec le langage intérieur), le regroupement d'informations (ex: retenir trois légumes), des associations ou des moyens mnémotechniques (chansons, rythmes).
• Repères visuels : Utiliser des listes, des pictogrammes pour les routines afin de se souvenir des étapes.
• Résumer et reformuler : Demander à l'enfant de redire avec ses propres mots ce qu'il a entendu ou lu.
• Réduire la charge cognitive : Éviter de surcharger l'enfant d'informations, surtout dans les moments de transition (ex: le matin avant de partir à l'école).
Pour l'Inhibition
• Prendre un temps de recul : Apprendre à l'enfant à compter jusqu'à trois avant de réagir.
• Communiquer les attentes : Avoir des attentes réalistes et les expliquer clairement à l'enfant.
• Gestion des émotions : Mettre en place des stratégies pour gérer la colère ou la tristesse (respiration, visualisation, relaxation).
• Jeux d'adresse et d'équilibre : Ces jeux demandent de contrôler ses gestes et d'éviter les mouvements impulsifs.
• Jeux de vitesse et de réflexes : Jouer à des jeux où il faut réagir rapidement mais au bon moment (ex: le jeu Dobble).
Pour la Flexibilité Cognitive
• Laisser prendre des décisions : Permettre à l'enfant de prendre des décisions par lui-même.
• Déroger aux routines : Modifier occasionnellement une routine pour l'amener à s'adapter.
• Préparer les transitions : Aviser l'enfant à l'avance d'un changement d'activité pour lui permettre de s'y préparer cognitivement.
• Stimuler la créativité : Encourager les activités créatives, les jeux de rôle et l'improvisation.
• Changer la fin des histoires : Avant de lire la fin d'un livre, demander à l'enfant d'inventer sa propre conclusion.
Pour la Planification
• Utiliser des repères de temps visuels : Calendriers, horaires de la journée ou de la fin de semaine.
• Amener à anticiper : Poser des questions comme : «
- Qu'est-ce que tu penses qu'il va se passer après ? »,
- « De quoi auras-tu besoin pour cette activité ? ».
• Créer des plans : Avant une construction en blocs LEGO, suggérer de dessiner un petit plan.
• Remettre en séquence : Après avoir lu une histoire ou vu un film, demander à l'enfant de raconter ce qui s'est passé au début, au milieu et à la fin.
-
-
www.youtube.com www.youtube.com
-
Synthèse sur la Gestion de la Colère et de la Frustration chez l'Enfant
Résumé Exécutif
Ce document de synthèse présente les stratégies et les concepts clés pour aider les enfants à gérer leur colère et leur frustration, basés sur l'expertise de Madame Ly Massy, docteure en psychologie de l'éducation.
L'approche fondamentale est de considérer la colère non pas comme une émotion négative à supprimer, mais comme une émotion normale, un signal d'alarme indiquant une injustice ou un préjudice.
Elle devient problématique uniquement lorsqu'elle est mal exprimée, trop intense ou injustifiée.
La gestion de ces émotions repose sur le développement de la régulation émotionnelle, une compétence qui s'articule autour de trois habiletés fondamentales :
1. L'identification des émotions et de leurs sources.
2. La modulation de l'intensité émotionnelle pour prévenir l'escalade et retrouver le calme.
3. L'expression adéquate des émotions pour résoudre les situations conflictuelles.
Le rôle des parents est central et s'articule autour de la prévention et de l'enseignement de stratégies concrètes.
La prévention consiste à instaurer un environnement sécurisant par le biais de routines prévisibles et de saines habitudes de vie, notamment une activité physique suffisante et un sommeil adéquat.
Pour l'enseignement, il est crucial d'aider l'enfant à développer un vocabulaire émotionnel, à reconnaître ses propres signaux (physiques, cognitifs, comportementaux) et à maîtriser un éventail de stratégies d'apaisement (techniques physiques, diversion de l'attention).
Le modelage parental — la manière dont les parents gèrent et expriment leurs propres émotions — est l'outil le plus puissant pour guider l'enfant.
--------------------------------------------------------------------------------
1. Comprendre la Colère : Une Émotion Normale
Selon Madame Ly Massy, la perception de la colère comme une émotion purement négative est une erreur. Elle doit être comprise comme un mécanisme de signalisation fondamental.
• Une Fonction d'Alarme : La colère est décrite comme "un petit bouton d'alarme" qui signale qu'une situation est perçue comme injuste, moqueuse, ou qu'elle nuit au développement personnel.
• Quand la Colère Devient Problématique : L'enjeu n'est pas de supprimer la colère, mais de gérer son expression. Elle devient problématique dans trois cas de figure :
1. Expression Inadéquate : L'enfant "explose au lieu de dire tranquillement qu'est-ce qui nous fait vivre l'émotion négative".
2. Intensité Excessive : L'émotion est trop intense et l'enfant ne parvient pas à la contrôler.
3. Justification Inappropriée : La colère est déclenchée par une mauvaise interprétation de la situation (par exemple, percevoir un accident comme un acte intentionnel).
2. La Régulation Émotionnelle : Un Apprentissage en Trois Temps
La régulation émotionnelle est une habileté qui se développe tout au long de la vie, mais dont les bases devraient être acquises au début du primaire. Elle se compose de trois compétences interdépendantes.
| Habileté | Description | | --- | --- | | 1\. Identification des Émotions | Comprendre l'émotion ressentie et ses sources. C'est le prérequis à toute gestion. | | 2\. Modulation des Émotions | Être capable de gérer l'intensité de l'émotion pour éviter une escalade ou pour s'apaiser après une crise et retrouver son calme. | | 3\. Expression des Émotions | Communiquer de manière adéquate ce que l'on ressent et ce qui a causé l'émotion, dans le but de résoudre la situation. |
3. Facteurs de Difficulté dans la Régulation Émotionnelle
Plusieurs caractéristiques individuelles peuvent expliquer pourquoi certains enfants ont plus de difficultés à gérer leurs émotions.
• Hypersensibilité ou Hyperréactivité : Une réaction plus intense aux émotions, aux situations et à l'environnement. Le "moindre petit élément va susciter une réaction négative".
• Seuil de Frustration Bas : L'image d'une "mèche courte" est utilisée. Il en faut très peu pour que l'enfant se sente frustré ou en colère.
• Faible Tolérance au Délai de Gratification : Une difficulté à attendre. Le besoin doit être satisfait immédiatement, sinon une réaction négative s'ensuit.
• Rigidité Mentale : Un besoin que les choses soient faites d'une manière précise ou selon leurs goûts.
Un changement dans la routine peut susciter des émotions négatives.
C'est un trait commun chez les enfants avec un trouble du spectre de l'autisme.
• Perceptions Faussées : Une tendance à interpréter les situations neutres comme hostiles.
L'enfant est en état de vigilance constant, "comme s'il était toujours en vigilance d'avoir peur d'être attaqué".
Cela peut mener à généraliser ou dramatiser les événements.
• Trauma : Un traumatisme peut accentuer tous les facteurs précédents et mener à un "surétiquetage" des émotions négatives comme de la colère, l'enfant cherchant inconsciemment à se sentir moins victime et plus combatif.
4. Stratégies Parentales : Prévention et Anticipation
La première étape pour un parent est de mettre en place des conditions pour "prévenir les pertes de contrôle" plutôt que de simplement réagir aux crises.
4.1. Un Environnement Sécurisant
• Instauration de Routines : Mettre en place des routines prévisibles, surtout pour les moments de la journée identifiés comme difficiles (le matin avant l'école, la période des devoirs, le coucher, la fin de l'effet de la médication).
La prévisibilité rend la gestion des frustrations plus facile pour l'enfant.
4.2. Saines Habitudes de Vie
• Activité Physique : Essentielle pour évacuer les tensions accumulées.
Un enfant qui ne bouge pas assez (par exemple, en s'installant directement pour les devoirs après l'école) aura plus de mal à gérer un événement frustrant.
• Sommeil : Le manque de sommeil est un des "premiers facteurs qui va nuire à la régulation émotionnelle". Les besoins varient selon l'âge :
◦ Début du primaire : Entre 11 et 13 heures. ◦ Début du secondaire : Environ 9 heures.
4.3. Identification des Déclencheurs de Crises
Il est crucial de distinguer deux types de facteurs :
• Facteurs Précipitants (Déclencheurs) : Ce qui initie la crise. Exemples : l'envahissement de l'espace personnel, une blessure d'estime de soi, l'accumulation de stress, la faim.
En identifiant la source, on peut agir en amont (par exemple, prévoir une collation dans la voiture au retour de l'école).
• Facteurs Aggravants : Ce qui intensifie la crise. Exemples : la fatigue de fin de journée, la fin de l'effet d'une médication pour un TDAH.
5. Développer les Compétences de Régulation Émotionnelle
Le parent doit activement enseigner les trois habiletés fondamentales de la régulation, en choisissant un moment où l'enfant est calme.
5.1. Habileté 1 : Identifier les Émotions
• Enrichir le Vocabulaire : Aider l'enfant à nommer ses émotions avec précision (irrité, frustré, furieux) et à décrire ses sensations physiques ("vidé", "submergé").
• Reconnaître les Signaux : L'aider à identifier les signaux précurseurs de la colère :
◦ Signaux Corporels : Externes (tension, rougeur, froncement des sourcils) et internes (cœur qui bat vite, souffle court).
◦ Comportements : Tendance à frapper, à se replier sur soi.
◦ Pensées : Identifier les pensées qui nourrissent la colère ("il l'a fait exprès").
• Outils et Approches :
◦ Refléter : "Tu as l'air déçu, est-ce qu'il s'est passé quelque chose ?".
◦ Utiliser des supports ludiques : Livres, histoires, jeux de mimes, affiches, collages, création d'un "dictionnaire des émotions".
5.2. Habileté 2 : Moduler les Émotions
• Évaluer l'Intensité : Utiliser une échelle simple comme un thermomètre des émotions (vert, jaune, orange, rouge) pour aider l'enfant à prendre conscience de son état et à choisir la bonne stratégie.
• Enseigner Deux Grandes Catégories de Stratégies :
1. Stratégies Physiques : Très efficaces pour réduire la tension physique lors d'émotions intenses.
▪ Respiration profonde : "La plus simple et la plus efficace".
▪ Relaxation : Relaxation musculaire, méditation de pleine conscience.
▪ Stimulation Sensorielle : Objets lourds (couverture, toutou), se bercer, se retirer dans un endroit calme et peu stimulant (tente, garde-robe), manipuler des objets (balles de tension, "putty").
2. Stratégies de Diversion de l'Attention : Idéales pour interrompre les pensées négatives en boucle qui nourrissent la colère.
▪ Activités exigeant de la concentration : Compter à l'envers (de 20 à 1, ou de 200 à 0 par bonds de 4), dessiner des mandalas, jouer aux LEGO, faire une activité artistique.
▪ Objets Visuels : Regarder des objets calmants (spirales liquides, lumières douces).
• Stratégie Cognitive : Les "Pensées Froides"
◦ Aider l'enfant à remplacer les "pensées chaudes" qui attisent la colère par des "pensées froides" qui apaisent.
Exemples : "Il ne l'a pas fait exprès", "Ça va me couler sur le dos comme sur un canard".
5.3. Habileté 3 : Exprimer les Émotions
• Le Bon Moment et le Bon Endroit : Cette étape ne peut se faire que lorsque l'enfant s'est calmé, dans un lieu propice à la discussion.
• Le "Message au Je" : Enseigner à l'enfant à structurer son expression :
1. Je me sens... (nommer l'émotion).
2. Parce que... (expliquer la cause).
3. J'aimerais que... (proposer une solution pour que cela ne se répète pas).
• Le Modelage Parental : "Ici encore plus, le modelage est important".
Les parents doivent eux-mêmes utiliser le "message au je" et parler de leurs propres émotions. C'est en voyant ses parents le faire que l'enfant apprendra le mieux.
6. Messages Clés et Conclusion
Madame Ly Massy conclut avec plusieurs messages fondamentaux pour les parents et les éducateurs :
• La Normalité de la Colère : Il faut cesser de diaboliser cette émotion.
• La Primauté de l'Identification : "Si on n'a pas identifié comment on se sent, on ne sera pas capable de prendre les bons moyens pour se contrôler".
• La Diversité des Stratégies : Il faut enseigner un large éventail de stratégies, car aucune n'est universellement efficace.
• Les Exutoires Sains : Proposer des moyens sains de se défouler quand l'expression verbale directe n'est pas possible (écriture, art, sport intense).
• Le Rôle du Parent : Être un modèle de calme, éviter de "mettre de l'huile sur le feu" et savoir prendre soi-même une grande respiration pour garder son calme face à l'enfant.
-
-
www.youtube.com www.youtube.com
-
Soutenir la Participation Parentale à l'École : Synthèse et Stratégies Clés
Résumé Exécutif
Ce document de synthèse analyse les stratégies et les concepts fondamentaux pour favoriser une participation parentale efficace et constructive dans le milieu scolaire, en se basant sur l'expertise d'Elodie Marion, docteure en administration publique et professeure à l'Université de Montréal.
L'implication parentale est définie comme un ensemble de pratiques à la maison, à l'école et dans la communauté, qui soutiennent le développement de l'enfant.
Son efficacité dépend moins des attentes de l'école envers les parents que des pratiques mises en place par l'école elle-même pour la soutenir.
Trois facteurs principaux influencent l'implication des parents : leur contexte de vie, leur perception d'être invités à participer et leur sentiment de compétence.
Pour surmonter les obstacles et catalyser l'engagement, une approche proactive fondée sur les "Cinq R" est proposée :
- Reconnaissance,
- Respect,
- Rôle,
- Résultats et
- Relation.
Cette approche transforme la dynamique de simple diffusion d'information en une véritable collaboration.
Enfin, la mise en place d'un plan de communication stratégique par les enseignants est essentielle.
Ce plan doit être structuré autour de quatre axes : la fréquence, la bidirectionnalité, les moyens adaptés et la diversité des thèmes abordés.
L'objectif ultime est d'instaurer une coéducation, une responsabilité partagée qui non seulement bénéficie à l'élève, mais brise également l'isolement de l'enseignant et augmente l'efficacité collective.
--------------------------------------------------------------------------------
1. Définition et Modalités de l'Implication Parentale
L'implication parentale englobe toutes les pratiques que les parents mettent en œuvre en lien avec l'éducation de leur enfant.
La recherche la catégorise généralement en trois sphères distinctes, chacune avec des actions concrètes.
1.1. Implication à la Maison
• Rôle Parental : Création d'un environnement familial propice à l'apprentissage et au développement.
Cela inclut la gestion des habitudes de vie (sommeil, alimentation), du stress et le soutien général au bien-être de l'enfant.
• Soutien Scolaire : Actions directement liées à la scolarité de l'enfant, telles que les communications sur l'école, les encouragements, et la transmission d'attentes claires concernant la réussite scolaire.
1.2. Implication à l'École
• Communications : Échanges formels et informels avec le personnel de l'école.
◦ Communications en personne : Rencontres parents-enseignants, réunions avec la direction.
◦ Communications écrites : Messages quotidiens, signatures sur les évaluations (noté comme étant souvent une diffusion d'information plutôt qu'une communication bilatérale).
• Participation à la Prise de Décision : Engagement actif dans les instances de gouvernance de l'école, comme les comités d'école, les comités au centre de services scolaires ou la participation aux rencontres pour l'élaboration de plans d'intervention.
• Volontariat : Implication bénévole dans les activités de l'école.
1.3. Implication dans la Communauté
Bien que mentionnée comme une troisième catégorie par les chercheurs, cette dimension n'est pas détaillée dans la source.
2. Facteurs d'Influence sur l'Implication Parentale
L'engagement des parents n'est pas inné ; il est influencé par un ensemble de facteurs personnels et contextuels. L'école doit les comprendre pour adapter ses propres pratiques.
• Le Contexte de Vie des Familles : Les conditions de vie des parents (statut d'emploi, stress, monoparentalité, statut socio-économique) déterminent leur capacité à s'impliquer.
Par exemple, un parent d'un enfant autiste peut trouver le soutien scolaire à la maison extrêmement difficile, non par manque de volonté, mais en raison du contexte familial.
• La Perception d'Être Invité : L'implication dépend de la manière dont le parent perçoit et reçoit l'invitation de l'école à participer.
Une invitation envoyée n'est pas nécessairement une invitation ressentie comme telle.
• Le Sentiment de Compétence Parentale : Les parents, particulièrement ceux d'enfants en difficulté, peuvent se sentir démunis ou incompétents pour aider leur enfant.
Ce manque de confiance peut être interprété à tort comme un désengagement.
De plus, un sentiment de ne pas être à la hauteur des attentes de l'école peut paralyser leur implication.
• La Reconnaissance de l'Expertise Parentale : Les parents possèdent une connaissance fine et continue de leur enfant. Ils sont "le filon du parcours de son enfant".
Ils souhaitent que cette expertise, acquise au quotidien et souvent documentée, soit reconnue et valorisée par l'école, plutôt que de se sentir dans une relation hiérarchique face à "l'expert" scolaire.
3. Une Approche Stratégique : Le Cadre des "Cinq R" pour Favoriser l'Engagement
Pour passer d'une posture d'attente à une posture de soutien actif, Elodie Marion propose un cadre mnémonique basé sur "cinq R".
Cette approche vise à regarder ses propres pratiques ("se regarder dans le miroir") plutôt que de juger celles des parents ("regarder par la fenêtre").
| Le "R" | Définition | Actions Concrètes | | --- | --- | --- | | Reconnaissance | Valider et reconnaître les efforts et contributions des parents, passés et présents. | \- Mentionner les progrès accomplis les années précédentes.<br>\- Souligner les efforts actuels, même s'ils sont modestes.<br>\- Éviter de se concentrer uniquement sur les nouveaux défis sans reconnaître le travail déjà fait. | | Respect | Tenir compte de l'opinion, de l'expertise, de la culture et du contexte de vie des parents. | \- Pratiquer l'écoute active lors des rencontres.<br>\- Arriver en mode discussion plutôt qu'avec un ordre du jour rigide.<br>\- Adapter les attentes de l'école à la réalité des familles (ex: trouver des alternatives au soutien scolaire à la maison si nécessaire). | | Rôle | Clarifier et définir explicitement le rôle attendu du parent pour éviter les malentendus. | \- Expliciter ce que l'école attend (ex: encourager, questionner) et ce qu'elle n'attend pas (ex: ré-enseigner la matière).<br>\- Co-construire un rôle clair et accepté par le parent.<br>\- Donner des pistes concrètes sur comment le parent peut questionner son enfant sur les apprentissages. | | Résultats | Mettre en évidence les progrès et les objectifs pour que l'engagement soit perçu comme utile. | \- Définir des objectifs clairs pour chaque rencontre.<br>\- Communiquer fréquemment les améliorations observées chez l'enfant suite à l'implication du parent.<br>\- Célébrer les succès pour motiver la poursuite des efforts, au lieu d'attendre les bilans officiels. | | Relation | Bâtir une véritable relation de confiance et un sentiment d'appartenance. | \- Organiser des moments de rencontre informels (ex: fête de la rentrée).<br>\- Concevoir les rencontres pour permettre un réel échange et pas seulement une transmission d'information.<br>\- Intégrer un temps pour la "présentation des parents" lors des réunions de début d'année. |
4. Mettre en Place un Plan de Communication Efficace à l'École
Un plan de communication réfléchi est un outil puissant pour structurer et améliorer les interactions avec les parents tout au long de l'année.
Il doit s'articuler autour de quatre axes.
4.1. Fréquence
• Planification : Établir dès le début de l'année un rythme de communication réaliste pour l'enseignant et pertinent pour les parents.
• Routine : Créer des habitudes de communication (ex: un courriel hebdomadaire) pour éviter les contacts sporadiques ou uniquement déclenchés par des problèmes.
4.2. Bidirectionnalité
• Passer de la diffusion à l'échange : Concevoir les communications pour solliciter l'avis et les informations des parents.
• Poser des questions ouvertes : Utiliser des questions commençant par "comment" ou "pourquoi" pour inviter à des réponses développées, plutôt que des questions fermées (oui/non).
• Expliciter les besoins : Indiquer clairement aux parents les types d'informations que l'enseignant souhaite recevoir (ex: un changement à la maison, une inquiétude de l'enfant).
4.3. Moyens de Communication
• Adaptation : Choisir des moyens de communication adaptés au contexte des parents (ex: langue, niveau de littératie).
• Simplification : Alléger les communications écrites pour en faciliter la compréhension. Éviter les lettres de plusieurs pages.
• Le Piège Technologique : La multiplication des plateformes de communication (portails, applications) augmente souvent la quantité d'informations diffusées sans pour autant améliorer la relation.
Il est recommandé de réduire le nombre de canaux pour se concentrer sur la qualité des échanges.
4.4. Diversité des Thèmes
• Prévoir à l'avance : Planifier la diversité des sujets à aborder durant l'année pour éviter de ne communiquer que sur les difficultés.
• Inciter à l'action : Pour chaque information transmise, se poser la question : "Qu'est-ce que je veux que le parent fasse de cette information ?".
Cela permet de formuler des suggestions concrètes (ex: "Vous pourriez l'encourager en...", "N'hésitez pas à lui demander comment...").
5. Vers la Coéducation : Bénéfices d'une Responsabilité Partagée
L'instauration d'une collaboration solide et d'une relation de confiance entre l'école et la famille mène à une véritable coéducation.
Cette responsabilité partagée génère des bénéfices significatifs pour toutes les parties prenantes.
• Pour l'enseignant :
◦ Briser l'isolement : Le parent devient un allié dans la résolution de problèmes.
◦ Stimuler la créativité : L'échange avec le parent apporte de nouvelles perspectives et idées d'intervention.
◦ Gagner en efficacité : Bien que la collaboration demande du temps initialement, elle permet de comprendre plus vite les situations et de trouver des solutions plus durables, rendant le travail plus efficace à long terme.
• Pour le parent :
◦ Se sent valorisé, compétent et partie prenante de la réussite de son enfant.
• Pour l'élève :
◦ Bénéficie d'un soutien cohérent et aligné entre la maison et l'école, favorisant sa réussite scolaire et éducative.
-
-
www.youtube.com www.youtube.com
-
Soutenir l'Enfant face au Stress Toxique : Synthèse des Idées d'Alexandra Mathurin-Landry
Résumé Exécutif
Ce document de synthèse présente les perspectives d'Alexandra Mathurin-Landry, psychologue, neuropsychologue et professeure spécialisée dans le trauma infantile.
L'analyse distingue trois types de stress — positif, tolérable et toxique — en se concentrant sur les effets dévastateurs du stress toxique, étroitement lié au concept de "trauma complexe".
Le trauma complexe découle d'une exposition répétée et prolongée à des expériences d'adversité (abus, négligence) durant des périodes de développement critiques, souvent perpétrées par les figures de soin, ce qui prive l'enfant de tout effet protecteur.
Les données scientifiques, notamment une méta-analyse récente, révèlent l'ampleur du problème : 60 % des adultes rapportent avoir vécu au moins une expérience d'adversité dans l'enfance, et 16 % en ont vécu quatre ou plus, un seuil qui augmente substantiellement les risques développementaux.
Face à ce constat, les "approches sensibles au trauma" offrent une voie d'intervention prometteuse.
Le modèle "Attachement, Régulation et Compétences" (ARC) est présenté comme un cadre d'intervention concret, de plus en plus utilisé au Québec.
La mise en œuvre de ces approches repose sur une posture adulte de "syntonie" (ajustement empathique), qui consiste à devenir un "détective des émotions" pour comprendre les besoins non comblés derrière les comportements.
Trois stratégies concrètes sont proposées :
1) établir des routines prévisibles et sécurisantes ;
2) répondre aux besoins sous-jacents plutôt que de se focaliser sur les comportements visibles (l'analogie de l'iceberg) ; et
3) soutenir activement la régulation des émotions de l'enfant par la "corégulation", où le calme de l'adulte aide l'enfant à retrouver le sien.
--------------------------------------------------------------------------------
Introduction de l'Experte et du Sujet
L'intervenante, Alexandra Mathurin-Landry, est une experte reconnue dans le domaine du développement de l'enfant.
En tant que professeure agrégée à l'École de travail social et de criminologie, titulaire de la Chaire Richelieu de recherche sur la jeunesse, l'enfance et la famille, psychologue et neuropsychologue, ses travaux se concentrent sur l'amélioration des connaissances sur le trauma et le développement de pratiques innovantes et sensibles aux traumas.
L'objectif de la discussion est d'explorer les différentes formes de stress et d'identifier les moyens de soutenir le développement et la réussite des enfants qui y sont confrontés.
Les Trois Formes de Stress : Un Modèle de Compréhension
Contrairement à une perception monolithique, le stress n'est pas uniforme.
Un modèle distinguant trois formes de stress permet de mieux comprendre leurs impacts respectifs sur le développement de l'enfant.
1. Le Stress Positif
◦ Description : Une forme de stress bénéfique associée à des événements normatifs qui mobilisent la préparation et les apprentissages.
◦ Exemples : Un examen scolaire, une présentation orale.
◦ Impact : N'a pas d'effets négatifs à long terme sur le développement. Il contribue à l'adaptation et à la survie.
2. Le Stress Tolérable
◦ Description : Résulte d'un événement plus exceptionnel, unique et intense. Il a un début et une fin clairs.
◦ Exemple : Un enfant se faisant frapper par une automobile.
◦ Facteur Clé : La présence d'un adulte ou d'une figure de soutien qui offre un "effet protecteur" est cruciale.
Cet adulte croit, soutient et accompagne l'enfant, ce qui permet de mitiger les effets du stress et de le rendre "tolérable".
Le message clé est de ne "pas négliger le rôle qu'on peut avoir protecteur auprès des enfants".
3. Le Stress Toxique
◦ Description : Cette forme de stress est la plus dommageable.
Elle survient dans un contexte où un enfant est exposé de manière répétée et prolongée à des expériences de vie très difficiles.
◦ Exemples : Abus, négligence.
◦ Facteurs Aggravants :
▪ Absence de Protection : L'effet protecteur de l'adulte est absent. Pire, les parents ou figures de soin peuvent être la source du trauma.
▪ Nature du Trauma : Les événements menacent l'intégrité physique et psychologique de l'enfant, affectent son identité et peuvent être associés à un sentiment de trahison.
◦ Impact : A des effets substantiels et à long terme sur tous les domaines du développement de l'enfant, y compris le développement du cerveau.
Du Stress Toxique au Trauma Complexe
Les travaux d'Alexandra Mathurin-Landry portent spécifiquement sur le "trauma complexe", un concept étroitement lié au stress toxique.
• Définition du Trauma Complexe : La situation d'un enfant exposé de façon répétée et prolongée à des expériences d'abus, de négligence ou d'autres problématiques familiales lors de périodes vulnérables du développement (petite enfance, adolescence).
• Chevauchement : Le trauma complexe peut être directement associé au stress toxique en raison de la nature chronique de l'adversité et de l'implication des figures de soin.
• Statut Diagnostique : Il est important de noter que ni le stress toxique ni le trauma complexe ne sont des diagnostics officiels, ce qui complique leur étude et leur quantification précise.
Prévalence de l'Adversité Infantile : Les Données Probantes
En l'absence de diagnostic formel, l'étude de la fréquence des expériences d'adversité (abus, négligence, problématiques familiales) permet d'estimer l'ampleur du phénomène.
Une méta-analyse récente (2023 ou 2024), combinant plus de 200 études et un demi-million d'individus, fournit des données robustes :
| Indicateur | Pourcentage | Interprétation | | --- | --- | --- | | Adultes ayant vécu au moins une forme d'adversité infantile | 60 % | Un chiffre qualifié d'"énorme" et de "trop élevé". | | Adultes ayant vécu quatre formes ou plus d'adversité infantile | 16 % | Un cumul qui augmente "substantiellement les risques" pour le développement. |
Pour rendre ce chiffre concret, 16 % équivaut à quatre enfants dans une classe de 25, dont le parcours de vie est marqué par une adversité significative.
Les Approches Sensibles au Trauma : Une Voie vers la Guérison
Malgré la gravité du problème, il existe des solutions efficaces.
Les "approches sensibles au trauma" sont des interventions qui ont montré des effets positifs significatifs.
• Définition : Des approches systémiques qui modifient la "manière de voir les choses, notre manière de penser, nos manières de faire".
Elles visent à ce que les pratiques, politiques et procédures tiennent compte de la réalité du trauma.
• Objectif : Créer des environnements sécurisants, accueillants et engageants pour les jeunes.
• Portée Universelle : Bien que conçues pour les enfants ayant un vécu traumatique, ces approches sont bénéfiques pour tous les enfants et ne présentent "pas de contre-indication".
Elles sont particulièrement importantes pour ceux qui vivent un stress toxique ou un trauma complexe.
Le Modèle ARC : Attachement, Régulation et Compétences
Le modèle ARC est un cadre d'intervention concret et tangible qui incarne les principes des approches sensibles au trauma.
Il connaît un essor important au Québec, notamment dans les services de protection de la jeunesse et dans les écoles.
• Cible : Il vise à outiller les intervenants, mais aussi les parents (biologiques, d'accueil) et autres "piliers de résilience".
• Objectifs Fondamentaux :
1. Attachement : Aider l'enfant à développer une base relationnelle sécurisante.
2. Régulation : Soutenir l'acquisition de compétences et de stratégies de régulation des émotions.
Un point crucial est que cette acquisition doit être soutenue par l'adulte ; l'enfant ne doit pas être laissé seul.
3. Compétences : Développer diverses compétences chez l'enfant.
La Syntonie : La Clé de Voûte de l'Intervention
Au cœur du modèle ARC se trouve le concept de "syntonie", aussi appelé "ajustement empathique". C'est un élément clé de la posture de l'adulte.
• Description : Une posture axée sur l'accueil, la sensibilité aux besoins et aux émotions de l'enfant, et la capacité à détecter ses signaux verbaux et non verbaux.
• La Métaphore du Détective : L'adulte doit agir comme un "détective des émotions", cherchant à comprendre ce qui se cache derrière un comportement.
Les comportements souvent jugés "perturbateurs" ou "oppositionnels" peuvent en réalité être la manifestation de besoins non comblés (affection, écoute, affirmation de soi) ou d'émotions non régulées.
Trois Stratégies Concrètes Issues du Modèle ARC
Pour traduire ces principes en actions, trois stratégies fondamentales sont proposées :
1. Établir des Routines
◦ Objectif : Sécuriser l'enfant par la prévisibilité. Les enfants ayant un vécu traumatique ont particulièrement besoin de prévisibilité, car leur vie passée en a souvent manqué.
◦ Application : Les routines doivent être individualisées pour répondre aux besoins spécifiques de chaque enfant (ex: besoin de s'activer ou de se reposer le matin).
2. Répondre aux Besoins Sous-Jacents (L'Analogie de l'Iceberg)
◦ Concept : Les comportements visibles ne sont que la pointe de l'iceberg.
La majeure partie, sous la surface de l'eau, est constituée des besoins non comblés, des émotions non régulées et du vécu traumatique.
◦ Intervention : Pour être efficace, l'intervention doit "plonger sous l'eau" et s'adresser aux causes profondes (besoins, émotions) plutôt que de se limiter à la gestion du comportement en surface.
En répondant aux besoins, les comportements problématiques sont susceptibles de diminuer.
3. Soutenir la Régulation des Émotions (La Corégulation)
◦ Principe : Ne pas laisser l'enfant seul face à ses émotions.
La "corégulation" est le processus par lequel "l'état calme de l'adulte va aider l'enfant à retrouver un état calme".
◦ Action : L'adulte doit aider l'enfant à remplir sa "boîte à outils" de stratégies de régulation saines (car il n'en a peut-être pas appris ou en a développé de dangereuses comme l'automutilation).
Plus important encore, l'adulte doit être présent pour "ouvrir sa boîte à outils avec lui" et lui montrer comment utiliser les outils.
-
-
www.youtube.com www.youtube.com
-
Document de Synthèse : Contrer l'Absentéisme au Secondaire
Résumé Exécutif
Ce document synthétise une approche innovante pour la gestion de l'absentéisme dans une école secondaire, développée et présentée par Véronique Sir, directrice d'établissement et candidate au doctorat.
Le projet marque une transition fondamentale d'un modèle punitif, jugé lourd et inefficace, vers un modèle relationnel qui responsabilise et outille les enseignants.
Cette nouvelle stratégie a permis de réduire de 50 % le nombre d'élèves présentant plus de 15 absences non motivées en une seule année scolaire.
Au-delà des chiffres, la retombée la plus significative est l'amélioration notable de la relation entre les enseignants et les élèves, les premiers n'étant plus perçus comme des "polices de la retenue" mais comme des adultes bienveillants et soucieux de la présence de chaque jeune.
La mise en œuvre s'est articulée en cinq étapes clés, incluant une analyse rigoureuse, la création d'un sous-comité stratégique, une approche pilote par "petits pas", une intégration systémique et un partage des connaissances.
Le projet met en lumière l'importance du temps, de l'adhésion des équipes et de la focalisation sur le pouvoir d'agir collectif de l'école plutôt que sur des facteurs externes.
Contexte et Problématique Initiale
À l'arrivée de la nouvelle direction il y a trois ans, deux irritants majeurs étaient palpables et verbalisés par le personnel de l'école :
1. Un manque de cohérence dans l'application du code de vie.
2. Une gestion des absences perçue comme excessivement lourde et inefficace.
Cette dernière tâche était si pesante que la majorité des enseignants souhaitaient s'en dégager.
L'analyse initiale des données a permis de "neutraliser l'effet négatif" des perceptions en démontrant que le problème, bien que réel, ne concernait que deux ou trois élèves par groupe, et non une majorité comme il était parfois ressenti.
Le Projet de Gestion des Absences : Une Approche Relationnelle
Philosophie et Changement de Paradigme
Le cœur du projet est un changement radical de philosophie, passant d'un système répressif à une approche humaine et proactive.
• D'un modèle punitif à un modèle relationnel : L'ancienne méthode, qui consistait à sanctionner l'absence (par exemple, par une retenue), est abandonnée au profit d'une démarche qui cherche à comprendre les causes de l'absence et à outiller l'élève.
Comme le résume Mme Sir : "On est passé d'un modèle punitif à un modèle relationnel et outillé soutenu par des facilitateurs à l'école."
• Le rôle central de l'enseignant : Le projet repose sur l'implication directe des enseignants, qui deviennent les premiers intervenants.
Ils sont responsables des sept premières interventions auprès de leurs élèves tuteurs, incluant deux appels aux parents pour les sensibiliser.
Cette approche s'oppose au réflexe de déléguer cette responsabilité à l'équipe de soutien, reconnaissant qu'une poignée d'intervenants ne peut gérer efficacement les absences de plus de 900 élèves.
La présence des enseignants est donc jugée "essentielle".
Résultats Quantitatifs
Le projet, axé sur une gestion par les résultats, a démontré un impact mesurable et significatif sur la réduction de l'absentéisme chronique non motivé.
| Période | Contexte | Nombre d'élèves avec >15 absences non motivées | | --- | --- | --- | | Juin 2024 | Fin de la phase pilote (3 mois, 3 groupes sur 35) | Environ 120 élèves | | Juin 2025 | Fin de la première année complète (tous les groupes) | Environ 60 élèves | | 31 octobre 2025 | Début de l'année scolaire en cours | 6 élèves |
Ces chiffres représentent une diminution d'environ 50 % des cas d'absentéisme chronique en un an.
Il est noté que le mois de juin tend à augmenter le nombre d'absences, ce qui rend la comparaison encore plus probante.
Le principal fait saillant est que tous les élèves de l'école (clientèle d'environ 950 jeunes) sont désormais connus et suivis, ne permettant à personne de "passer sous la craque".
Les Cinq Étapes de la Mise en Œuvre
Le cheminement réflexif du projet a été structuré en cinq phases distinctes, menées en collaboration avec des chercheurs universitaires.
1. Analyse de la situation : La première étape a consisté à faire émerger des données factuelles pour objectiver les deux irritants majeurs (code de vie et gestion des absences).
2. Création du sous-comité : Considérée comme le "cœur de la démarche", cette étape a impliqué la sélection stratégique de ses membres.
Le comité inclut non seulement des personnes ouvertes au changement, mais aussi des enseignants plus critiques et des membres du personnel encore attachés au modèle punitif.
L'objectif était de créer un espace de réflexion pour confirmer la fin du statu quo et construire une vision commune.
3. Culture des "petits pas" : Pour gérer le changement, le projet a débuté par un pilote limité : trois groupes, trois enseignants volontaires, pendant trois mois.
Ce n'est que la deuxième année que l'approche a été étendue à toute l'école.
Cette phase a été marquée par des "allers-retours constants" et un "droit à l'erreur", permettant d'ajuster les moyens tout en gardant le cap sur la finalité (le modèle relationnel).
4. Veilles et intégration systémique : Cette étape, imbriquée dans les autres, a consisté à ancrer le projet dans toutes les instances de l'école :
◦ Comité projet éducatif : Intégration d'indicateurs sur l'assiduité.
◦ Plan de lutte contre la violence et l'intimidation : Favoriser un climat scolaire sécuritaire.
◦ Assemblées générales : Véhiculer l'importance du projet, en faisant témoigner les "agents facilitateurs".
◦ Rencontres de niveaux : Instaurer un point statutaire toutes les deux semaines pour suivre les élèves absentéistes.
5. Partage à la communauté : La dernière étape consiste à diffuser le projet pour "faire gagner du temps" à d'autres équipes-écoles, évitant ainsi de réinventer des solutions existantes.
Défis, Facteurs de Succès et Recommandations
Défis Rencontrés
• La gestion du temps et des attentes : Les résultats ne sont pas immédiats.
Comprendre les causes profondes de l'absentéisme prend du temps, ce qui peut être un défi dans une culture axée sur les résultats rapides.
• L'adhésion de l'équipe : La deuxième année, lorsque tout le personnel est impliqué, est cruciale et peut voir émerger plus de résistance.
Le sous-comité joue un rôle fondamental pour accueillir ces résistances sans reculer.
• La gestion des cas chroniques : Certains élèves, aux prises avec des enjeux de santé mentale ou de démotivation scolaire importants, résistent aux interventions.
L'implication des professionnels (psychoéducateurs, conseillers d'orientation) est ici fondamentale.
• Le roulement du personnel : L'arrivée de personnel non formé en pédagogie peut rendre la création de liens plus difficile, nécessitant un soutien accru de la part des "agents facilitateurs" internes.
Principale Réussite : L'Amélioration de la Relation Enseignant-Élève
Le gain le plus "magnifique" et le plus positif du projet est l'amélioration de la qualité des relations.
Les enseignants ne sont plus vus comme des agents de sanction. Un enseignant a partagé une anecdote révélatrice :
"Les élèves m'ont dit à plusieurs reprises cette année : 'Cou'donc, avez-vous une vie à part nous regarder à l'école ?'".
Pour l'équipe, cette remarque est une "victoire", car elle signifie que chaque élève sait qu'au moins un adulte se soucie de sa présence.
Erreurs à Éviter
1. Aller trop vite : Le changement culturel et la compréhension des causes profondes de l'absence exigent du temps.
2. Remettre le sort aux parents : Plutôt que de se concentrer sur les motifs d'absence (sur lesquels l'école a peu de contrôle), la discussion doit être réorientée vers le "pouvoir d'agir collectif" à l'interne.
3. Utiliser les données à mauvais escient : Un outil de suivi (Power BI) a été développé pour fournir des données quotidiennes.
La vigilance est de mise pour que ces données servent à comprendre et agir, et non à "masquer artificiellement" les problèmes ou à créer une compétition entre les écoles.
Retombées Stratégiques et Pérennité du Projet
Outre la baisse de l'absentéisme et l'amélioration des relations, le projet a généré plusieurs impacts positifs durables :
• Approche personnalisée : L'école est passée d'une généralisation ("tous les élèves de 4e secondaire s'absentent") à une analyse fine et personnalisée des besoins de chaque élève.
• Standardisation des interventions : Un protocole écrit garantit la qualité et la pérennité des interventions, indépendamment du personnel en place.
• Autonomisation et résilience des équipes : Les enseignants ont développé une autonomie ("empowerment") et une résilience face à la problématique, conscients de leur pouvoir d'agir collectif.
• Préparation à la croissance : La structure mise en place est comparée aux "fondations d'une maison", rendant l'école prête à accueillir une hausse de sa clientèle.
• Pérennité du modèle : Le projet est conçu pour être durable. L'objectif final est de développer une autonomie telle que le projet puisse survivre au départ de la direction actuelle.
Comme le conclut Mme Sir : "demain matin si je pars comme direction d'établissement, le projet va survivre grâce à nos agents facilitateurs qui vont assurer la pérennité du projet."
-
-
www.elcolombiano.com www.elcolombiano.com
-
Si bien el artículo presenta hallazgos interesantes sobre la relación entre el cuidado de los nietos y el mantenimiento de la agudeza mental en personas mayores, su lectura resulta parcial si no se considera el contexto sociocultural y económico en el que fue realizado el estudio. En países como Colombia, muchos abuelos y abuelas continúan trabajando en edades avanzadas debido a la falta de una pensión estable, lo que implica cargas físicas y emocionales distintas a las analizadas en la investigación. Además, no se contemplan otras actividades cognitivas y físicas que también podrían incidir positivamente en la salud mental, como el trabajo, la participación comunitaria o el autocuidado. En el contexto bogotano, marcado por el tráfico, la inseguridad y las dificultades de movilidad, el cuidado de los nietos no siempre es una actividad estimulante, sino que puede convertirse en una fuente adicional de estrés. Y aunque los resultados pueden ser válidos en casos muy específicos, no son fácilmente generalizables al contexto colombiano sin un análisis mucho más situado.
-
el cuidado de los nietos puede mejorar la salud de los abuelos, al mismo tiempo que ralentiza su deterioro cognitivo.
Los niños requieren mucha atención y actividad física, por lo que un adulto mayor debe ejercitarse tanto mental como físicamente durante el cuidado de un menor. Esto hace que los cuidadores deban mantenerse atentos y físicamente aptos para el juego y demás tareas de cuidado. Yo lo veo en mi familia, porque mi mamá cuida a mi sobrino y eso la mantiene alegre y activa no solo a causa de la labor de cuidado como tal, sino por la relación cercana y amorosa que comparte con el niño, que se ha convertido en su principal motivación para estar bien.
-
Una nueva investigación revela que las personas de edades avanzadas que proporcionan cuidado infantil poseen mejores puntuaciones en pruebas de memoria y fluidez mental. Conozca los beneficios de esta práctica.
¿con qué intención se hace el estudio? sería interesante conocer los objetivos y financiamientos de los estudios en estas notas que lo que buscan son vistas. Yo veo una invitación a una población, que ha trabajado durante de más de 40 años, a seguir trabajando, sin remuneración, dado que el detrimento de la capacidad adquisitiva y la precarización de los derechos laborales, hace necesario encender las ventajas de la familia extensa, aunque la legislación favorece la familia nuclear. Todo lo anterior, aportando a la precarización de los cuidados y los cuidadores. Conozco algunos abuelos explotados que verían en esta nota la justificación del maltrato de sus hijos.
-
asistían a los pequeños
"Asistir" es un término muy impreciso. No distingue entre el cuidado recreativo y el cuidado intensivo o forzoso. Con seguridad, para los abuelos un poco de cuidado es beneficioso , ya que permite la socialización y les da un propósito, pero una carga excesiva, como la crianza a tiempo completo, sin duda genera estrés crónico, lo cual es neurotóxico y acelera el deterioro cognitivo. Al no mencionar la frecuencia ideal ni los riesgos del "síndrome de la abuela esclava", el artículo ofrece una visión incompleta y potencialmente dañina de la dinámica familiar.
-
de 67 años
Los abuelos que cuentan con una mejor salud física y cognitiva son, precisamente, aquellos a quienes se les confía el cuidado de los nietos o quienes se sienten capaces de hacerlo. Ahora bien, aquellos que ya presentan un deterioro cognitivo suelen ser apartados de estas responsabilidades en la mayorìa de casos por seguridad. Por tanto, no es necesariamente que los nietos mejoren el cerebro, como afirma el artículo, sino que se requiere un cerebro sano para poder cuidar de ellos.
-
-
www.examtopics.com www.examtopics.com
-
NOTA: Cada respuesta correcta vale un punto.
Asegúrese de que los clientes que procesan eventos marquen la posición dentro de una secuencia de eventos.
Marque la posición del cliente procesador de eventos dentro de la secuencia de eventos de una partición.
-
-
www.youtube.com www.youtube.com
-
Pratiques Punitives en Milieu Scolaire : Analyse, Effets et Recommandations
Résumé Exécutif
Ce document de breffage synthétise les conclusions de l'expert Vincent Bernier, docteur en psychopédagogie, concernant les pratiques punitives en milieu scolaire, et plus particulièrement la suspension.
La recherche, unanime depuis près de 50 ans, démontre que ces pratiques sont non seulement inefficaces, mais aussi profondément néfastes pour les élèves.
Loin de corriger les comportements problématiques, elles les exacerbent et entraînent une cascade d'effets négatifs à court, moyen et long terme, incluant la détérioration du rendement scolaire, le décrochage, l'augmentation des inégalités sociales et des problèmes de santé mentale et physique graves.
Face à ce constat, le document expose des alternatives fondées sur des approches éducatives qui visent à développer les compétences des élèves plutôt qu'à les sanctionner.
Ces alternatives se déclinent en deux catégories : des pratiques concrètes de gestion de classe, comme les conséquences éducatives, et des programmes structurants d'alternative à la suspension. Six recommandations stratégiques sont formulées pour le système éducatif québécois, appelant à un changement de paradigme.
Celles-ci incluent la sensibilisation du personnel, l'interdiction de la suspension externe sans service, et le financement de programmes alternatifs éprouvés, soulignant que l'investissement dans la prévention est socialement et économiquement plus judicieux que la gestion des conséquences coûteuses de l'inaction.
--------------------------------------------------------------------------------
1. Définition et Typologie des Pratiques Punitives
Selon Vincent Bernier, professeur et chercheur à l'Université de Sherbrooke, il est essentiel de distinguer clairement les pratiques punitives des pratiques éducatives.
Les pratiques punitives s'inscrivent dans une logique de répression plutôt que d'éducation.
Leur objectif principal est d'infliger une sanction en réponse à une faute afin de dissuader l'élève de répéter un comportement.
Ces pratiques se situent sur un continuum d'intensité et peuvent être regroupées en trois grandes catégories :
• Les punitions classiques : Ces mesures visent à sanctionner directement un élève.
◦ La copie de lignes ou de textes.
◦ L'assignation de travaux supplémentaires.
◦ Le retrait de privilèges ou de droits.
• Les pratiques d'exclusion : Ces mesures consistent à retirer l'élève d'une situation ou d'un milieu.
◦ La retraite de classe (qui peut être punitive ou éducative selon son application).
◦ Les retenues.
◦ La suspension scolaire (interne ou externe).
• Les punitions physiques : Bien que moins courantes au Québec, elles existent dans certains contextes.
◦ Les châtiments corporels.
◦ La contention physique.
En opposition, les pratiques éducatives visent à aider l'élève à développer ses compétences socio-émotionnelles, son autonomie et sa responsabilité par des mesures d'aide et de soutien.
2. Les Effets Négatifs Documentés des Pratiques Punitives
La littérature scientifique, québécoise et internationale, documente les effets des pratiques punitives depuis les années 1970 et est décrite comme "unanime" sur leurs conséquences négatives.
Impacts Comportementaux et Académiques
Contrairement à l'objectif visé, les pratiques punitives ne règlent pas les problèmes de comportement ; elles les aggravent.
• Escalade des comportements : On observe une détérioration et une aggravation des comportements problématiques chez les élèves exposés à ces pratiques.
• Désengagement scolaire : Les élèves développent des trajectoires d'évitement, une réduction de la motivation à apprendre et un désengagement progressif de l'école, menant souvent à l'absentéisme.
• Baisse du rendement : Une diminution significative du rendement scolaire est fréquemment constatée.
• Décrochage scolaire : La suspension scolaire est fortement corrélée au risque de décrochage.
Conséquences Sociales et Relationnelles
Ces pratiques endommagent le lien entre l'élève et l'école.
• Relations négatives : Elles contribuent au développement de relations conflictuelles et négatives avec le personnel scolaire.
• Sentiment d'exclusion : Les élèves se sentent isolés, mis de côté et exclus, ce qui renforce leur marginalisation.
• Affiliation à des pairs déviants : L'exclusion du milieu scolaire augmente le risque d'affiliation à des groupes de pairs déviants et l'enrôlement potentiel dans des gangs de rue.
Exacerbation des Inégalités
Les pratiques punitives ne sont pas appliquées uniformément et ont pour effet d'aggraver les inégalités sociales, socio-économiques et culturelles existantes.
• Marginalisation des groupes vulnérables : Elles touchent de manière disproportionnée certains groupes, notamment :
◦ Les garçons.
◦ Les élèves ayant des difficultés d'apprentissage.
◦ Les minorités ethniques.
◦ Les élèves issus de milieux défavorisés.
Effets à Long Terme et sur la Santé
Les études longitudinales démontrent que les conséquences de l'exposition à ces pratiques se prolongent bien au-delà de la période scolaire et affectent la santé globale des individus.
• Santé mentale et physique : Une diminution du bien-être général est observée, avec des risques accrus de :
◦ Dépression.
◦ Consommation de drogues.
◦ Automutilation.
◦ Grossesses à risque.
• Judiciarisation : Le risque d'arrestation à l'âge adulte est plus élevé pour les élèves ayant été fréquemment suspendus.
3. Alternatives aux Pratiques Punitives : Une Approche Éducative
Pour remplacer les pratiques punitives, deux grandes catégories d'alternatives sont proposées, toutes deux centrées sur l'éducation et le développement de compétences.
Catégorie 1 : Pratiques Éducatives Concrètes
Ces alternatives peuvent être mises en œuvre quotidiennement par le personnel scolaire pour prévenir et gérer les écarts de conduite.
• Enseignement explicite des comportements attendus.
• Stratégies de gestion de classe proactives (ex: précorrection, proximité).
• Développement de l'autorégulation (ex: autoévaluation, autotraitement).
• Mise en place de conséquences éducatives : Cette approche est particulièrement efficace. Elle se distingue de la punition par sa finalité.
La maxime qui la guide est que "la sanction elle est pas là pour faire mal mais elle est là pour faire sens".
◦ Logique et naturelle : La conséquence découle directement du comportement problématique.
◦ Réparatrice : Elle vise à réparer le tort causé.
◦ Responsabilisante : Elle implique l'élève dans la solution et l'aide à assumer ses responsabilités.
◦ Éducative : Elle est présentée comme une mesure d'aide et une occasion d'apprentissage pour développer des compétences.
Catégorie 2 : Alternatives Structurantes
Ces approches sont plus globales et s'appliquent à l'échelle de l'école ou du centre de services scolaire.
• Programmes d'alternative à la suspension scolaire :
Ces programmes, souvent menés par des organismes externes comme le YMCA, offrent un cadre structuré aux élèves qui auraient autrement été suspendus à la maison sans services.
◦ Fonctionnement :
▪ Matin : Réalisation des travaux scolaires envoyés par l'école.
▪ Après-midi : Participation à des groupes animés pour développer des compétences spécifiques (socio-émotionnelles, résolution de conflits, etc.).
◦ Efficacité prouvée : Les études sur ces programmes démontrent des effets positifs à court et moyen terme, incluant une amélioration des comportements et une diminution du recours futur à la suspension.
4. Recommandations Stratégiques pour le Système Éducatif Québécois
Vincent Bernier formule six recommandations claires pour systématiser le passage d'une approche punitive à une approche éducative au Québec.
| N° | Recommandation | Description | | --- | --- | --- | | 1 | Sensibiliser le personnel scolaire | Il est impératif d'informer l'ensemble du personnel scolaire sur les effets négatifs documentés des pratiques punitives afin de créer une prise de conscience collective. | | 2 | Soutenir le développement professionnel | Offrir de la formation continue pour outiller le personnel avec des pratiques alternatives efficaces et pour déconstruire les croyances erronées sur l'efficacité des punitions. | | 3 | Interdire la suspension externe sans services | Modifier les règlements d'école, les codes de vie et la Loi sur l'instruction publique pour interdire formellement la suspension scolaire à l'externe (à la maison) sans aucune mesure d'aide ou de soutien. | | 4 | Recadrer la suspension comme mesure de dernier recours | Si une suspension est inévitable, elle doit être utilisée de manière constructive : soit à l'interne avec des services et des mesures d'aide, soit à l'externe mais en référant l'élève à un programme d'alternative structuré. L'objectif doit être de "servir plutôt que de sévir". | | 5 | Financer et déployer les programmes alternatifs | Investir dans le déploiement de programmes d'alternative à la suspension, via des organismes communautaires locaux, pour que chaque école au Québec puisse avoir accès à ce type de service. | | 6 | Consigner systématiquement les suspensions | Mettre en place un système provincial pour consigner toutes les suspensions (internes et externes) afin d'obtenir un portrait juste et clair du phénomène (fréquence, durée, profils d'élèves) et ainsi prendre des décisions éclairées. |
5. Conclusion : L'Urgence d'un Virage vers la Prévention
La discussion met en lumière une réalité documentée depuis un demi-siècle : les pratiques punitives sont contre-productives.
L'expert souligne que le Québec doit réfléchir aux services qu'il souhaite offrir à ses jeunes.
Il est démontré que le coût sociétal de l'inaction (gestion de la criminalité, des problèmes de santé mentale, du décrochage) est largement supérieur au coût d'investissement dans des mesures de prévention et des programmes alternatifs.
Le passage d'une culture de la punition à une culture du soutien n'est pas seulement une question de bienveillance, mais une décision stratégique et économique pour l'avenir.
-
-
pesquisa.icea.decea.mil.br pesquisa.icea.decea.mil.br
-
CINDACTA IV (MANAUS - AM)
A imagem sugere uma divisão da FIR, o que só ocorre na FIR-RE com a FIR-AO
-
CINDACTA II (CURITIBA - PR)
A imagem sugere uma divisão da FIR, o que só ocorre na FIR-RE com a FIR-AO
-
-
www.youtube.com www.youtube.com
-
Analyse de la Psychologie des Séparations
Résumé Exécutif
Ce document de synthèse analyse les multiples facettes de la séparation, qu'elle soit amoureuse ou amicale, en se basant sur une combinaison de témoignages personnels et d'analyses psychologiques.
Il en ressort que la séparation est un processus psychobiologique profond, dont la tolérance se construit dès l'enfance à travers les mécanismes d'attachement.
La douleur de la rupture n'est pas purement émotionnelle ; elle se manifeste par des symptômes physiques concrets et des changements hormonaux mesurables, activant dans le cerveau des zones similaires à celles de la douleur physique.
Le processus de deuil est une étape cruciale et individuelle, où le refoulement des émotions peut mener à une "explosion" ou une "implosion".
La guérison passe souvent par la capacité à donner un sens à l'événement, en construisant un récit cohérent de ce qui s'est passé. Des approches thérapeutiques comme l'EMDR peuvent aider à retraiter les souvenirs traumatiques.
Enfin, la séparation, bien que douloureuse, peut se révéler être une opportunité de croissance, permettant une meilleure connaissance de soi, la redéfinition de ses propres limites et une forme de "renaissance" personnelle.
Le soutien social et l'expression des émotions, notamment par l'écriture, sont des aides précieuses tout au long de ce processus.
--------------------------------------------------------------------------------
1. La Nature Fondamentale de la Séparation : Attachement et Anxiété
La séparation est un événement complexe qui puise ses racines dans les fondements psychologiques de l'individu et génère une anxiété profonde.
• L'Importance de l'Attachement Primal : La capacité à gérer une séparation et à tolérer la solitude est directement liée aux expériences d'attachement vécues dès les premiers instants de la vie.
Les relations avec les parents et les amis durant l'enfance déterminent la manière dont un individu entrera en relation et vivra les ruptures futures.
• La Douleur Partagée de la Rupture : Contrairement à une idée reçue, l'acte de quitter une personne peut être aussi difficile et douloureux que d'être quitté.
La décision de rompre est souvent lourde et entraîne des conséquences importantes pour celui qui la prend. Un témoin exprime ce dilemme : "je savais que j'allais lui faire très mal mais je pouvais pas faire autrement".
• Anxiété et Peur de la Solitude : La séparation engendre une forte anxiété, souvent liée à la peur de se retrouver seul et d'affronter les étapes du deuil.
• Perte d'Identité et Confiance en Soi : Pour certains, la relation est si fusionnelle que la séparation entraîne un sentiment de néant.
Une personne témoigne : "à partir du moment où il est parti vraiment et c'est là que j'ai commencé à ressentir que j'étais plus rien quoi comme si ma vie finalement était suspendue à la sienne".
La capacité à se détacher est donc intrinsèquement liée à l'estime de soi et à la conviction de pouvoir exister indépendamment de l'autre.
• Le Poids des Modèles Familiaux : Les schémas parentaux peuvent influencer la perception de la séparation.
Une personne ayant vu sa mère "tout supporter" pour "préserver la famille" a ressenti sa propre rupture comme un échec à maintenir ce modèle, faisant le "deuil de la famille".
2. Les Manifestations Physiques et Neurologiques de la Douleur
La souffrance d'une rupture n'est pas une simple métaphore ; elle s'inscrit dans le corps et le cerveau de manière quantifiable et observable.
• Réactions Organiques Généralisées : L'ensemble de l'organisme réagit à une séparation. Les symptômes physiques courants incluent :
◦ Changements d'appétit (perte de 6 kg en trois mois de grossesse pour une témoin).
◦ Maux de tête et maux d'estomac.
◦ Troubles du sommeil et du rythme cardiaque.
◦ Sensations localisées, comme une "boule au ventre" persistante.
◦ Réactions de stress physique comme la transpiration des mains.
• La Douleur Émotionnelle et la Douleur Physique : Des études montrent que la souffrance émotionnelle d'une rupture active des zones cérébrales très proches de celles activées par une douleur physique intense.
La source cite : "la souffrance qu'on ressent quand quelqu'un qu'on aime n'est plus à nos côtés cette douleur est vraiment très proche de la douleur physique qu'on ressent quand on se fait très mal".
• Changements Hormonaux et État de Manque : La période qui suit immédiatement la rupture est marquée par des bouleversements hormonaux significatifs, créant un état de manque comparable à une addiction.
Le "système de récompense" du cerveau "réclame la personne qui est partie".
| Hormone | Niveau après la rupture | Rôle et Conséquence | | --- | --- | --- | | Dopamine | Très élevé | Associée au système de récompense, son taux élevé explique les pensées obsessionnelles envers l'autre. | | Ocytocine | Baisse | "L'hormone de l'attachement", sa diminution accentue le sentiment de détachement et de perte. | | Sérotonine | Très faible | "L'hormone du contentement", son faible niveau contribue à l'état de tristesse et de nostalgie profonde. |
3. Le Processus de Deuil et les Stratégies de Guérison
La traversée de la séparation est un processus long et individuel, qui nécessite d'affronter la douleur pour pouvoir la surmonter.
• La Nécessité de Vivre ses Émotions : Le refoulement est une stratégie à risque. Il est essentiel de "s'autoriser à vivre son émotion, son chagrin, sa tristesse".
Le refoulement mène à deux issues possibles : "soit au bout d'un moment on explose ou soit on implose en soi".
• La Création d'un Récit Cohérent : Une étape clé de la guérison est la capacité à élaborer un récit, à trouver un sens à ce qui s'est passé.
Se poser les questions "pourquoi ça m'est arrivé ?", "quelles leçons j'en tire ?" et "qu'est-ce que je ferais différemment ?" permet de créer une histoire cohérente qui aide à "surmonter les coups durs".
• Thérapies et Neuroplasticité :
◦ Traumatisme et Système Limbique : Un événement traumatisant comme une rupture peut rester "bloqué dans le cerveau émotionnel" (le système limbique).
◦ La Thérapie EMDR : Cette thérapie utilise des stimulations bilatérales alternées (mouvements oculaires) pour permettre au cerveau de "retraiter" le souvenir traumatique.
Grâce à la neuroplasticité, elle aide à créer de nouvelles connexions synaptiques, permettant de "porter un nouveau regard sur ce qui nous est arrivé et de soulager son vécu émotionnel".
• L'Importance du Soutien Social : L'isolement aggrave la douleur. Rompre l'isolement en se reconnectant à sa "communauté" et à ses amis est crucial.
Le soutien consiste principalement à "être à ses côtés et de l'écouter".
• Autres Mécanismes d'Adaptation :
◦ L'Écriture : Écrire, comme rédiger des lettres (même non envoyées), est un moyen de se "plonger en soi" et de clarifier ses pensées.
◦ Le Temps : Le processus est individuel et ne suit pas un schéma unique. Il est essentiel de "laisser du temps au processus de séparation".
4. La Rupture Amicale : Une Douleur Similaire
La source souligne que la fin d'une amitié profonde peut être aussi dévastatrice qu'une rupture amoureuse, car les mêmes mécanismes d'attachement sont en jeu.
• Une Souffrance Équivalente : La douleur ressentie est décrite comme étant la même que pour une perte amoureuse.
Un témoin décrit son état : "c'était douloureux c'était comme une rupture active", "j'avais une boule dans la gorge parfois j'avais envie de pleurer".
• Le Manque de Communication : Contrairement aux ruptures amoureuses, qui sont souvent officialisées, les amitiés ont tendance à "se défaire lentement".
Le manque de communication directe est souvent au cœur du problème.
"Ce qui nous a manqué avec mon ami cette communiquer", constate un témoin.
L'idéal serait de "rompre avec un ami comme on rond avec un amoureux ou une amoureuse".
• L'Incompréhension et les Questions Sans Réponse : L'absence d'explication claire laisse la personne dans le questionnement et la douleur.
"Je sais pas exactement ce qui s'est passé pour ça il aurait fallu qu'on s'explique".
5. La Séparation comme Catalyseur de Croissance Personnelle
Bien qu'étant une épreuve difficile, la séparation peut paradoxalement devenir une opportunité de transformation et de renouveau.
• Une "Renaissance" : Plusieurs témoignages évoquent un sentiment de renaissance après avoir traversé le deuil.
Une personne décrit cette étape comme "une renaissance de maréchaux", où elle a retrouvé la force et le courage d'agir selon ses propres envies.
• Meilleure Connaissance de Soi : La rupture force à l'introspection.
Elle "peut aussi nous permettre de nous connaître et de savoir qu'est-ce qu'on veut qu'est-ce qu'on ne veut plus pour sa vie future".
• Redéfinition des Limites : C'est l'occasion de définir des limites personnelles qui n'étaient pas respectées.
Une conclusion forte émerge d'un témoignage : "on ne doit pas douter de quelque chose qui concerne son intégrité. [...] je veux plus que mon intégrité soit atteint".
• Vers des Relations Futures Plus Stables :
L'expérience montre que les personnes qui se donnent le temps de vivre pleinement le processus de séparation "parviennent généralement mieux à nouer une relation stable par la suite".
• La Possibilité d'une Nouvelle Relation : Une séparation amoureuse peut évoluer.
En se débarrassant des "restes de relations amoureuses" et en trouvant un "nouveau cadre", il est possible de rester amis.
-
-
test2025.mitkoforevents.cz test2025.mitkoforevents.czKontakt1
-
Souhlasím se zpracováním svých osobních údajů v uvedeném formuláři společností MITKO Sp. z o. o. za účelem vyřízení dotazu. Poskytnutí údajů je dobrovolné, ale nezbytné pro zpracování dotazu. Byl/a jsem informován/a, že mám právo na přístup ke svým údajům, možnost jejich opravy a požadovat zastavení jejich zpracování. Správcem osobních údajů je MITKO Sp. z o.o. se sídlem ve Wodzisławě Śląském, 44-304, ul. 1 Maja 16G.
Mitko Sp.?
-
-
www.biorxiv.org www.biorxiv.org
-
Author response:
The following is the authors’ response to the previous reviews
Reviewer 1
Minor
The main substance of my previous comment I suppose targeted a deeper issue - namely whether such a result is reflecting a resolution to a 'neural prediction' puzzle or a 'perceptual prediction' puzzle. Of course, these results tell us a great deal about a potential resolution for how dampening and sharpening might co-exist in the brain - but in the absence of corresponding perceptual effects (or a lack of correlation between neural and perceptual variables - as outlined in this revision) I do wonder if any claims about implications for perception might need moderation or caveating. To be honest, I don't think the authors *need* to make any more changes along these lines for this paper to be acceptable - it is more an issue they might wish to consider themselves when contextualizing their findings.
Thank you for the thoughtful comment. We have now added a caveat to the relevant section of the discussion to make it clearer that we are discussing neural results, not perceptual results (p.20, lines 378-379).
I am also happy with the changes that the authors have made justifying which claims can and cannot made based on a statistical decoding test against 'chance' in a single condition using t-tests. I was perhaps a little unclear when I spoke about 'comparisons against 0' in my original review, when the key issue (as the authors have intuited!) is about comparisons against 'chance' (where e.g., 0% decoding above chance is the same thing as 'chance'!). The authors are of course correct in the amendment they have made on p.29 to make clear this is a 'fixed effects analysis' - though I still worry this could be a little cryptic for the average reader. I am not suggesting that the authors run more analyses, or revise any conclusions, but I think it would be more transparent if a note was added along the lines of "while the fixed effects approach (one-sample t-test) enables us to establish whether some consistent informative patterns are detectable in these particular subjects, the results from our paired t-tests support inference to the wider population".
This sentence has been added for increased transparency (p. 27, lines 544-547).
Reviewer 3
Major
(1) In the previous round of comments, I noted that: "I am not fully convinced that Figures 3A/B and the associated results support the idea that early learning stages result in dampening and later stages in sharpening. The inference made requires, in my opinion, not only a significant effect in one-time bin and the absence of an effect in other bins. Instead to reliably make this inference one would need a contrast showing a difference in decoding accuracy between bins, or ideally an analysis not contingent on seemingly arbitrary binning of data, but a decrease (or increase) in the slope of the decoding accuracy across trials. Moreover, the decoding analyses seem to be at the edge of SNR, hence making any interpretation that depends on the absence of an effect in some bins yet more problematic and implausible". The authors responded: "we fitted a logarithmic model to quantify the change of the decoding benefit over trials, then found the trial index for which the change of the logarithmic fit was < 0.1%. Given the results of this analysis and to ensure a sufficient number of trials, we focused our further analyses on bins 1-2". However, I do not see how this new analysis addresses the concern that the conclusion highlights differences in decoding performance between bins 1 and 2, yet no contrast between these bins are performed. While I appreciate the addition of the new model, in my current understanding it does not solve the problem I raised. I still believe that if the authors wish to conclude that an effect differs between two bins they must contrast these directly and/or use a different appropriate analysis approach.
Relatedly, the logarithmic model fitting and how it justifies the focus on analysis bin 1-2 needs to be explained better, especially the rationale of the analysis, the choice of parameters (e.g., why logarithmic, why change of logarithmic fit < 0.1% as criterion, etc), and why certain inferences follow from this analysis. Also, the reporting of the associated results seems rather sparse in the current iteration of the manuscript.
We thank the reviewer for this important point. Following your suggestion, we conducted additional post-hoc tests directly comparing the first and second bins. We found significant differences between bins in the invalid trials, but not the valid trials, suggesting that sharpening/dampening effects are condition specific. This is discussed in the manuscript on p.14, lines 268-271; p.15, 280-284; p.20, lines 382-386.
A logarithmic analysis was chosen as learning is usually found to be a nonlinear process; learning effects occur rapidly before stabilising relatively early, as seen in Fig. 2D. This is consistent with other research which found that logarithmic fits efficiently describe learning curves in statistical learning (Kang et al., 2023; Siegelman et al., 2018; Choi et al., 2020). By utilising a change of logarithmic fit at <0.1% as a criterion, it is ensured that virtually zero learning took place after that point, allowing us to focus our analysis on learning effects as they developed and providing a more accurate model of representational change. This is explained in the manuscript on p.13, lines 250-251; p.27-28, lines 557-563.
(2) A critical point the authors raise is that they investigate the buildup of expectations during training. They go on to show that the dampening effect disappears quickly, concluding: "the decoding benefit of invalid predictions [...] disappeared after approximately 15 minutes (or 50 trials per condition)". Maybe the authors can correct me, but my best understanding is as follows: Each bin has 50 trials per condition. The 2:1 condition has 4 leading images, this would mean ~12 trials per leading stimulus, 25% of which are unexpected, so ~9 expected trials per pair. Bin 1 represents the first time the participants see the associations. Therefore, the conclusion is that participants learn the associations so rapidly that ~9 expected trials per pair suffice to not only learn the expectations (in a probabilistic context) but learn them sufficiently well such that they result in a significant decoding difference in that same bin. If so, this would seem surprisingly fast, given that participants learn by means of incidental statistical learning (i.e. they were not informed about the statistical regularities). I acknowledge that we do not know how quickly the dampening/sharpening effects develop, however surprising results should be accompanied with a critical evaluation and exceptionally strong evidence (see point 1). Consider for example the following alternative account to explain these results. Category pairs were fixed across and within participants,i.e. the same leading image categories always predicted the same trailing image categories for all participants. Some category pairings will necessarily result in a larger representational overlap (i.e., visual similarity, etc.) and hence differences in decoding accuracy due to adaptation and related effects. For example, house barn will result in a different decoding performance compared to coffee cup barn, simply due to the larger visual and semantic similarity between house and barn compared to coffee cup and barn. These effects should occur upon first stimulus presentation, independent of statistical learning, and may attenuate over time e.g., due to increasing familiarity with the categories (i.e., an overall attenuation leading to smaller between condition differences) or pairs.
We apologise for the confusion, there are 50 expected trials per bin per condition. The trial breakdown is as follows. Each participant completed 1728 trials, split equally across 3 mappings (two 2:1 maps and one 1:2 map), giving 1152 trials in the 2:1 mapping. Stimuli were expected in 75% of trials (864), leaving 216 per bin, and 54 per leading image in each bin. We have clarified this in the script (p.14, line 267; p.15, line 280). This is in line with similar studies in the field (e.g. Han et al., 2019).
(3) In response to my previous comment, why the authors think their study may have found different results compared to multiple previous studies (e.g. Han et al., 2019; Kumar et al., 2017; Meyer and Olson, 2011), particularly the sharpening to dampening switch, the authors emphasize the use of non-repeated stimuli (no repetition suppression and no familiarity confound) in their design. However, I fail to see how familiarity or RS could account for the absence of
sharpening/dampening inversion in previous studies.
First, if the authors argument is about stimulus novelty and familiarity as described by Feuerriegel et al., 2021, I believe this point does not apply to the cited studies. Feuerriegel et al., 2021 note: "Relative stimulus novelty can be an important confound in situations where expected stimulus identities are presented often within an experiment, but neutral or surprising stimuli are presented only rarely", which indeed is a critical confound. However, none of the studies (Han et al., 2019; Richter et al., 2018; Kumar et al., 2017; Meyer and Olson, 2011) contained this confound, because all stimuli served as expected and unexpected stimuli, with the expectation status solely determined by the preceding cue. Thus, participants were equally familiar with the images across expectation conditions.
Second, for a similar reason the authors argument for RS accounting for the different results does not hold either in my opinion. Again, as Feuerriegel et al. 2021 correctly point out: "Adaptation-related effects can mimic ES when the expected stimuli are a repetition of the last-seen stimulus or have been encountered more recently than stimuli in neutral expectation conditions." However, it is critical to consider the precise design of previous studies. Taking again the example of Han et al., 2019; Kumar et al., 2017; Meyer and Olson, 2011. To my knowledge none of these studies contained manipulations that would result in a more frequent or recent repetition of any specific stimulus in the expected compared to unexpected condition. The crucial manipulation in all these previous studies is not that a single stimulus or stimulus feature (which could be subject to familiarity or RS) determines the expectation status, but rather the transitional probability (i.e. cue-stimulus pairing) of a particular stimulus given the cue. Therefore, unless I am missing something critical, simple RS seems unlikely to differ between expectation condition in the previous studies and hence seems implausible to account for differences in results compared to the current study.
Moreover, studies cited by the authors (e.g. Todorovic & de Lange, 2012) showed that RS and ES are separable in time, again making me wonder how avoiding stimulus repetition should account for the difference in the present study compared to previous ones. I am happy to be corrected in my understanding, but with the currently provided arguments by the authors I do not see how RS and familiarity can account for the discrepancy in results.
The reviewer is correct in that the studies cited (Han et al., 2019; Kumar et al., 2017; Meyer and Olson, 2011) ensure that participants are equally familiar with the images across expectation conditions. Where the present study differs is that participants are not familiar with individual exemplars at all. Han et al., 2019 used a pool of 30 individual images, and subjects underwent exposure sessions lasting two hours each daily for 34 days prior to testing. Kumar et al., 2017 used a pool of 12 images with subjects being exposed to each sequential pair 816 times over the course of the training period. Meyer & Olsen, 2011 used pure tones at five different pitch levels. While familiarity of stimuli across conditions was controlled for in these studies in the sense that familiarity was constant across conditions, novelty was not controlled for. The present study uses a pool of ~3500 images, which are unrepeated across trials.
Feuerriegel et al., 2021 also points out: “There are also effects of adaptation that are dependent on the recent stimulation history extending beyond the last encountered stimulus and long-lag repetition effects that occur when the first and second presentation of a stimulus is separated by tens or even hundreds of intervening images”. Bearing this in mind, and given the very small pool of stimuli being used by Han et al., 2019; Kumar et al., 2017; Meyer and Olson, 2011, it stands to reason that these studies may still have built-in but unaccounted for effects relating to the repetition of exemplars. Thus, our avoidance of those possible confounds, in addition to foregoing any prior training, may elicit differing results. Furthermore, as pointed out by Walsh et al. 2020, methodological heterogeneity (such as subject training) can produce contrasting results as PP makes divergent predictions regarding the properties of prediction error given different permutations of variables such as training, transitional probabilities, and conditional probabilities. In our case, the use of differing methodology was intentional. These issues have been discussed in more detail on p.5, lines 112-115; p.19, lines 368-377; p.20, lines 378-379).
Minor
(1) The authors note in their reply to my previous questions that: "As mentioned above, we opted to target our ERP analyses on Oz due to controversies in the literature regarding univariate effects of ES (Feuerriegel et al., 2021)". This might be a lack of understanding on my side, but how are concerns about the reliability of ES, as outlined by Feuerriegel et al. (2021), an argument for restricting analyses to 1 EEG channel (Oz)? Could one not argue equally well that precisely because of these concerns we should be less selective and instead average across multiple (occipital) channels to improve the reliability of results?
The reviewer is correct in suggesting that a cluster of occipital electrodes may be more reliable than reporting one single electrode. We have amended the analysis to examine electrodes Oz, O1, and O2 (p.9, lines 187-188; p.11, lines 197-201).
(2) The authors provide a github link for the dataset and code. However, I doubt that github is a suitable location to share EEG data (which at present I also cannot find linked in the github repo). Do the authors plan to share the EEG data and if so where?
Thank you for bringing this to my attention. EEG data has now been uploaded at osf.io/x7ydf and linked to the github repository (p.28, lines 569-570).
(3) The figure text could benefit from additional information; e.g. Fig.1C and Fig.3 do not clarify what the asterisk indicates; p < ? with or without multiple comparison correction?
Thank you for pointing out this oversight, the figure texts have been amended (p. 9, line 168; p.16, line 289).
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
Brands and colleagues investigate how temporal adaptation can aid object recognition, and what neural computations may underlie these effects. They employed a previously published experimental paradigm to study how adaptation to temporally constant distractor input facilitates the recognition of a newly appearing target object. Specifically, they studied how this effect is modulated by the contrast of the target object.
They found that adaptation enhances the recognition of high-contrast objects more than that of low-contrast objects. This behavioral effect was mirrored by a larger effect of adaptation on the response to the high-contrast objects in relatively higher visual areas.
To investigate what neural computations can support this interaction, they implement several candidate neural mechanisms in a deep convolutional neural network: additive suppression, divisive suppression, and lateral recurrence. The authors conclude that divisive and additive suppression, which are intrinsic to the neuron, best explain the interaction between contrast and adaptation in the human data. They further show that these mechanisms, and divisive suppression in particular, show increased robustness to spatial shifts of the adaptor stimulus, hinting and potential perceptual benefits.
Strengths:
(1) Overall, this is a well-written paper, supported by thorough analyses and illustrated with clear, well-designed figures that effectively show overall trends as well as data variance. The authors tell a compelling story while responsibly steering away from overreaching conclusions.
(2) What makes this paper stand out is its comprehensive approach to understanding the behavioral benefit of neural adaptation and its mechanistic underpinnings. The authors effectively achieve this through integrating new behavioral and neural data with simulations using neural network models.
(3) The findings convincingly demonstrate that neuronally intrinsic adaptation mechanisms are sufficient to explain the observed interaction between temporal adaptation, contrast, and object recognition. Furthermore, the paper highlights that these intrinsic mechanisms offer superior robustness compared to learned lateral recurrence mechanisms, which, while being more expressive, can also be more brittle.
Weaknesses:
While the results and conclusion are well supported, there were a few major points that need clarification for me.
(1) Divisive normalization
I was confused by the author's classification of divisive normalization as a neuronally intrinsic mechanism, that is, one that operates within a single neuron, independent of interactions with other neurons.
My understanding is that divisive normalization, as originally proposed by Heeger in the early nineties, describes a mechanism where neurons integrate pooled activity from neighboring cells to mutually inhibit one another. In this form, divisive normalization is fundamentally an interneuronal mechanism involving recurrence. Adding to the confusion, the authors highlight in the introduction their interest in divisive normalization for its relation to stimulus contrast, a relation likely linked to neuronal pooling.
However, my reading of the methods section (Equations 6 and 7) suggests the authors implemented only a temporal feedback component, leaving out the pooling across neurons (Equation 5). This distinction should be disambiguated early in the paper. I recommend choosing a less ambiguous term than "divisive normalization". Even "temporal divisive normalization" is still ambiguous, as lateral neuronal interactions are also inherently temporal.
(2) Parietal electrodes
The paper's adapter-specific effects are centered around the P9/P10 electrodes, which the authors identify as "parietal." However, it is unclear to me which part of the cortex drives these electrodes, particularly whether it is actually the parietal cortex. I am no expert in EEG, but based on the topomaps in Figures 4 and 5, it appears that these electrodes cover more posterior occipito-temporal regions rather than truly parietal regions. Given the central role of P9/P10 to the main findings, the paper would be significantly improved for non-EEG readers by clarifying which cortical regions are covered by these electrodes.
(3) Interpretation of non-significant statistical results
In some places, the authors attach relatively strong claims to non-significant statistical results. For example, in Figure 5D, they claim that there is no effect of contrast on occipital electrodes, based on a non-significant p-value. P-values do not quantify evidence for the null hypothesis, so the authors should be careful with such claims. In fact, Figure 5D shows such a clear negative slope, with variance comparable to Figure 5A, that I am surprised that the p-value for the slope of Figure 5D was in fact so large. A similar issue arises in the discussion for Figure 6, where the authors claim that the effect of contrast is adapter-specific. However, this claim is based on the observation that is significant for same-noise trials, but not for different-noise or blank trials. To statistically substantiate the claims that there is an adapter-specific effect, the authors should directly compare the slope for same-noise trials with the slope for different-noise/blank trials.
(4) The match between behavior and models
The authors' claim that models with intrinsic adaptation better match the interaction between contrast and temporal adaptation observed in human behavior is not fully substantiated. This conclusion appears to be based on a qualitative assessment of Figure 8, which, in my view, does not unambiguously rule out an interaction for lateral recurrence. Furthermore, a potential confounding factor is the ceiling effect that limits higher accuracy values. Indeed, conditions where the interaction was not/less (i.e., shorter time sequences and lateral inhibition) are also the conditions where accuracy values are closer to this ceiling, which may mask a potential interaction.
-
-
www.biorxiv.org www.biorxiv.org
-
Note: This response was posted by the corresponding author to Review Commons. The content has not been altered except for formatting.
Learn more at Review Commons
Reply to the reviewers
We sincerely appreciate the feedback, attention to detail and timeliness of the referees for our manuscript. Below, we provide a point-by-point response to all comments from the referees, detailing the changes we have already made, and those that are in progress. Referee's comments will appear in bolded text, while our responses will be unbolded. Any text quoted directly from the manuscript will be italicised and contained within "quotation marks". Additionally, we have grouped all comments into four categories (structural changes, minor text changes, experimental changes, figure changes), comments are numbered 1-n in each of these categories. Please note: this response to reviewer's comments included some images that cannot be embedded in this text-only section.
1. General Statements
We appreciate the overall highly positive and enthusiastic comments from all reviewers, who clearly appreciated the technical difficulty of this study, and noted amongst other things that this study represents" a major contribution to the future advancement of oocyst-sporozoite biology" and the development of the segmentation score for oocysts as a "major advance[ment]". We apologise for the omission of line numbers on the document sent to reviewers, we removed these for the bioRxiv submission without considering that this PDF would be transferred across to Review Commons.
We have responded to all reviewers comments through a variety of text changes, experimental inclusions, or direct query response. Significant changes to the manuscript since initial submission are as follows:
- Refinement of rhoptry biogenesis model: Reviewers requested more detail around the content of the AORs, which we had previously suggested were a vehicle for rhoptry biogenesis as we saw they carried the rhoptry neck protein RON4. To address this, we first attempted to address this using antibodies against rhoptry bulb proteins but were unsuccessful. We then developed a * berghei* line where there rhoptry bulb protein RhopH3 was GFP-tagged. Using this parasite line, we observed that the earliest rhoptry-like structure, which we had previously interpreted as an AOR contained RhopH3. By contrast, RhopH3 was absent from AORs. Reflecting these observations we have renamed this initial structure the 'pre-rhoptry' and suggested a model for rhoptry biogenesis where rhoptry neck cargo are trafficked via the AOR but rhoptry bulb cargo are trafficked by small vesicles that move along the rootlet fibre (previously observed by EM).
- Measurement of rhoptry neck vs bulb: While not directly suggested by the reviewers, we have also included an analysis that estimates the proportion of the sporozoite rhoptry that represents the rhoptry neck. By contrast to merozoites, which we show are overwhelmingly represented by the rhoptry bulb, the vast majority of the sporozoite rhoptry represents the rhoptry neck.
- Measurement of subpellicular microtubules: One reviewer asked if we could measure the length of subpellicular microtubules where we had previously observed that they were longer on one side of the sporozoite than the other. We have now provided absolute and relative (% sporozoite length) length measurements for these subpellicular microtubules and also calculated the proportion of the microtubule that is polyglutamylated.
- More detailed analysis of RON11cKD rhoptries: Multiple comments suggested a more detailed analysis of the rhoptries that were formed/not formed in RON11cKD We have included an updated analysis that shows the relative position of these rhoptries in sporozoites.
2. Point-by-point description of the revisions
Reviewer #1
Minor text changes (Reviewer #1)
-
__Text on page 12 could be condensed to highlight the new data of ron4 staining of the AOR. __
We agree with the reviewer that it is a reasonable suggestion. After obtaining additional data on the contents of the AOR (as described in General Statements #1), this section has been significantly rewritten to highlight these findings. 2.
__Add reference on page 3 after 'disrupted parasites' __
This sentence has been rewritten slightly with some references included and now reads: "Most data on these processes comes from electron microscopy studies 6-8, with relatively few functional reports on gene deleted or disrupted parasites9-11. 3.
__Change 'the basal complex at the leading edge' - this seems counterintuitive __
This change has been made. 4.
__Change 'mechanisms underlying SG are poorly' - what mechanisms? of invasion or infection? __
This was supposed to read "SG invasion" and has now been fixed. 5.
__On page 4: 'handful of proteins' __
This error has been corrected. 6.
__What are the 'three microtubule spindle structures'? __
The three microtubule spindle structures: hemispindle, mitotic spindle, and interpolar spindle are now listed explicitly in the text. 7.
__On page 5: 'little is known' - please describe what is known, also in other stages. At the end of the paper I would like to know what is the key difference to rhoptry function in other stages? __
The following sentence already detailed that we had recently used U-ExM to visualise rhoptry biogenesis in blood-stage parasites, but the following two sentences have been added to provide extra detail on these findings: "In that study, we defined the timing of rhoptry biogenesis showing that it begun prior to cytokinesis and completed approximate coincident with the final round of mitosis. Additionally, we observed that rhoptry duplication and inheritance was coupled with centriolar plaque duplication and nuclear fission." 8.
__change 'rhoptries golgi-derived, made de novo' __
This has been fixed. 9.
__change 'new understand to' __
This change has been made 10.
__'rhoptry malformations' seem to be similar in sporozoites and merozoites. Is that surprising/new? __
We assume this is in reference to mention of "rhoptry malformations" in the abstract. In the RON11 merozoite study (PMID:39292724) the authors noted no gross rhoptry malformations, only that one was not formed/missing. The abstract sentence has been changed to the following to better reflect this nuance: "*We show that stage-specific disruption of RON11 leads to a formation of sporozoites that only contain half the number of rhoptries of controls like in merozoites, however unlike in merozoites the majority of rhoptries appear grossly malformed."
* 11.
__What is known about crossing the basal lamina. Where rhoptries thought to be involved in this process? Or is it proteins on the surface or in other secretory organelles? __
We are unaware of any studies that specifically look at sporozoites crossing the SG basal lamina. A review, although now ~15 years old stated that "No information is available as to how the sporozoites traverse the basal lamina" (PMID:19608457) and we don't know any more information since then. To try and better define our understanding of rhoptry secretion during SG invasion, we have added the following sentence:
"It is currently unclear precisely when during these steps of SG invasion rhoptry proteins are required, but rhoptry secretion is thought to begin before in the haemolymph before SG invasion16." 12.
__On page change/specify: 'wide range of parasite structures' __
The structures observed have been listed: centriolar plaque, rhoptry, apical polar rings, rootlet fibre, basal complex, apicoplast. 13.
__On page 7: is Airyscan2 a particular method or a specific microscope? __
Airyscan2 is a detector setup on Zeiss LSM microscopes, this was already detailed in the materials and methods sections, but figure legends have been clarified to read: "...imaged by an LSM900 microscopy with an Airyscan2 detector". 14.
__how large is RON11? __
RON11 is 112 kDa in * berghei*, as noted in the text. 15.
__There is no causal link between ookinete invasion and oocyst developmental asynchrony __
We have deleted the sentence that implied that ookinete invasion was responsible for oocyst asynchrony. This section now simply states that "Development of each oocyst within a midgut is asynchronous..." 16.
__First sentence of page 24 appears to contradict what is written in results____ I don't understand the first two sentences in the paragraph titled Comparison between Plasmodium spp __
This sentence was worded confusingly, making it appear contradictory when that was not the intention. The sentence has been changed to more clearly support what is written in the discussion and now reads: "Our extensive analysis only found one additional ultrastructural difference between Plasmodium spp."
__On page 25 or before the vast number of electron microscopy studies should be discussed and compared with the authors new data. __
It is not entirely clear which new data should be specifically discussed based on this comment. However, we have added a new paragraph that broadly compares MoTissU-ExM and our findings with other imaging methods previously used on mosquito-stage malaria parasites:
"*Comparison of MoTissU-ExM and other imaging modalities
Prior to the development of MoTissU-ExM, imaging of mosquito-stage malaria parasites in situ had been performed using electron microscopy7,8,11,28, conventional immunofluorescence assays (IFA)10, and live-cell microscopy25. MoTissU-ExM offers significant advantages over electron microscopy techniques, especially volume electron microscopy, in terms of accessibility, throughput, and detection of multiple targets. While we have benchmarked many of our observations against previous electron microscopy studies, the intracellular detail that can be observed by MoTissU-ExM is not as clear as electron microscopy. For example, previous electron microscopy studies have observed Golgi-derived vesicles trafficking along the rootlet fibre8 and distinguished the apical polar rings44; both of which we could not observe using MoTissU-ExM. Compared to conventional IFA, MoTissU-ExM dramatically improves the number and detail of parasite structures/organelles that can be visualised while maintaining the flexibility of target detection. By contrast, it can be difficult or impossible to reliably quantify fluorescence intensity in samples prepared by expansion microscopy, something that is routine for conventional IFA. For studying temporally complex processes, live-cell microscopy is the 'gold-standard' and there are some processes that fundamentally cannot be studied or observed in fixed cells. We attempt to increase the utility of MoTissU-ExM in discerning temporal relationships through the development of the segmentation score but note that this cannot be applied to the majority of oocyst development. Collectively, MoTissU-ExM offers some benefits over these previously applied techniques but does not replace them and instead serves as a novel and complementary tool in studying the cell biology of mosquito-stage malaria parasites.**"
*
__First sentence on page 27: there are many studies on parasite proteins involved in salivary gland invasion that could be mentioned/discussed. __
The sentence in question is "To the best of our knowledge, the ability of sporozoites to cross the basal lamina and accumulate in the SG intercellular space has never previously been reported."
This sentence has now been changed to read as follows: "While numerous studies have characterized proteins whose disruption inhibited SG invasion9,10,15,59-63, to the best of our knowledge the ability of sporozoites to cross the basal lamina and accumulate in the SG intercellular space has never previously been reported ."
__On page 10 I suggest to qualify the statement 'oocyst development has typcially been inferred by'. There seem a few studies that show that size doesn't reflect maturation. __
In our opinion, this statement is already qualified in the following sentence which reads: "Recent studies have shown that while oocysts increase in size initially, their size eventually plateaus (11 days pot infection (dpi) in P. falciparum4)."
__On page 16 the authors state that different rhoptries might have different function. This is an interesting hypothesis/result that could be mentioned in the abstract. __
The abstract already contains the following statement: "...and provide the first evidence that rhoptry pairs are specialised for different invasion events." We see this as an equivalent statement.
Experimental changes (Reviewer #1)
-
On page 19: do the parasites with the RON11 knockout only have the cytoplasmic or only the apical rhoptries?
The answer to this is not completely clear. We have added the following data to Figures 6 and 8 where we quantify the proportion of rhoptries that are either apical or cytoplasmic: In both wildtype parasites and RON11ctrl parasites, oocyst spz rhoptries are roughly 50:50 apical:cytoplasmic (with a small but consistent majority apical), while almost all rhoptries are found at the apical end (>90%) in SG spz. Presumably, after the initial apical rhoptries are 'used up' during SG invasion, the rhoptries that were previously cytoplasmic take their place. In RON11cKD the ratio of apical:cytoplasmic rhoptries is fairly similar to control oocyst spz. In RON11cKD SG spz, the proportion of cytoplasmic rhoptries decreases but not to the same extent as in wildtype or RON11Ctrl. From this, we infer that the two rhoptries that are lost/not made in RON11cKD sporozoites are likely a combination of both the apical and cytoplasmic rhoptries we find in control sporozoites.
__in panel G: Are the dense granules not micronemes? What are the dark lines? Rhoptries?? __
We have labelled all of Figure 1 more clearly to point out that the 'dark lines' are indeed rhoptries. Additionally, we have renamed the 'protein-dense granules' to 'protein-rich granules', as it seems we are suggesting that these structures are dense granules the secretory organelle. At this stage we simply do not know what all of these granules are. The observation that some but not all of these granules contain CSP (Supplementary Figure 2) suggests that they may represent heterogenous structures. It is indeed possible that some are micronemes, however, we think it is unlikely that they are all micronemes for a number of reasons: (1) micronemes are not nearly this protein dense in other Plasmodium lifecycle stages, (2) some of them carry CSP which has not been demonstrated to be micronemal, (3) very few of these granules are present in SG sporozoites, which would be unexpected because microneme secretion is required for hepatocyte invasion.
__Figure 2 seems to add little extra compared to the following figures and could in my view go to the supplement. __
We agree that Figure 2b adds little and so have moved that to Supplementary Figure 2, but think that the relative ease at which it can be distinguished if sporozoites are in the secretory cavity or SG epithelial cell is a key observation because of the difficulty in doing this by conventional IFA.
__On page 8 the authors mention a second layer of CSP but do not further investigate it. It is likely hard to investigate this further but to just let it stand as it is seems unsatisfactory, considering that CSP is the malaria vaccine. What happens if you add anti-CSP antibodies? I would suggest to shorten the opening paragraphs of this paper and to focus on the rhoptries. This could be done be toning down the text on all aspects that are not rhoptries and point to the open question some of the observations such as the CSP layers raise for future studies. __
When writing the manuscript, we were unsure whether to include this data at all as it is a purely incidental finding. We had no intention of investigating CSP specifically, but anti-CSP antibodies were included in most of the salivary gland imaging experiments so we could more easily find sporozoites. Given the tremendous importance of CSP to the field, we figured that these observations were potentially important enough that they should be reported in the literature even though they are not something we have the intention or resources to investigate subsequently. Additionally, after consultation with other microscopists we think there is a reasonable chance that this double-layer effect could be a product of chemical fixation. To account for this, we have qualified the paragraph on CSP with this sentence:
"We cannot determine if there is any functional significance of this second CSP layer and considering that it has not been observed previously it may well represent an artefact of chemical (paraformaldehyde) fixation."
__Maybe include more detail of the differences between species on rhoptry structure into Figure 4. I would encourage to move the Data on rhoptries in Figure S6 to the main text ie to Figure 4. __
We have moved the images of developing rhoptries in * falciparum *(previously Figure S6a and b) into figure 4, which now looks as follows:
Figure S8 (previously S6c) now consists only of the MG spz rhoptry quantification
Manuscript structural changes (Reviewer #1)
-
Abstract: don't focus on technique but on the questions you tried to answer (ie rewrite or delete the 3rd and 4th sentence)
-
'range of cell biology processes' - I understand the paper that the key discovery concerns rhoptry biogenesis and function, so focus on that, all other aspects appear rather peripheral.
-
'Much of this study focuses on the secretory organelles': I would suggest to rewrite the intro to focus solely on those, which yield interesting findings.
-
Page 11: I am tempted to suggest the authors start their study with Figure 3 and add panel A from Figure 2 to it. This leads directly to their nice work on rhoptries. Other features reported in Figures 1 and 2 are comparatively less exciting and could be moved to the supplement or reported in a separate study.____ Page 23: I suggest to delete the first sentence and focus on the functional aspects and the discoveries.
-
__Maybe add a conclusion section rather than a future application section, which reads as if you want to promoted the use of ultrastructure expansion microscopy. To my taste the technological advance is a bit overplayed considering the many applications of this techniques over the last years, especially in parasitology, where it seems widely used. In any case, please delete 'extraordinarily' __
Response to Reviewer#1 manuscript structural changes 1-5: This reviewer considers the findings related to rhoptry biology as the most significant aspect of the study and suggests rewriting the manuscript to emphasize these findings specifically. Doing so might make the key findings easier to interpret. However, in our view, this approach could misrepresent how the study originated and what we see as the most important outcomes. We did not develop MoTissU-ExM specifically to investigate rhoptry biology. Instead, this technique was created independently of any particular biological question, and once established, we asked what questions it could answer, using rhoptry biology as a proof of concept. Given the authors' previous work and available resources, we chose to focus on rhoptry biology. Since this was driven by basic research rather than a specific hypothesis, it's important to acknowledge this in the manuscript. While we agree that the findings related to rhoptry biology are valuable, we believe that highlighting the technique's ability to observe organelles, structures, and phenotypes with unprecedented ease and detail is more important than emphasizing the rhoptry findings alone. For these reasons, we have decided not to restructure the manuscript as suggested.
Reviewer #2
Minor text changes (Reviewer #2)
-
__The 'image Z-depth' value indicated in the figures is ambiguous. It is not clear whether this refers to the distance from the coverslip surface or the starting point of the z-stack image acquisition. A precise definition of this parameter would be beneficial. __
In the legend of Figure 1, the image Z-depth has been clarified as "sum distance of Z-slices in max intensity projection". 2.
__Paragraph 3 of the introduction - line 7, "handful or proteins" should be handful of proteins __
This has been corrected. 3.
__Paragraph 5 of the introduction - line 7, "also able to observed" should be observe __
This has been changed. 4.
__In the final paragraph of the introduction - line 1, "leverage this new understand" should be understanding __
This has been fixed. 5.
__The first paragraph of the discussion summary contains an incomplete sentence on line 7, "PbRON11ctrl-infected SGs." __
This has been removed. 6.
__The second paragraph of the discussion - line 10, "until cytokinesis beings" should be begins __
This mistake has been corrected. 7.
__One minor point that author suggest that oocyst diameter is not appropriate for the development of sporozoite develop. This is not so true as oocyst diameter tells between cell division and cell growth so it is important parameter especially where the proliferation with oocyst does not take place but the growth of oocyst takes place. __
We agree that this was not highlighted enough in the text. The final sentence of the results section about this now reads:
"While diameter is a useful readout for oocyst development in the early stages of its growth, this suggests that diameter is a poor readout for oocyst development once sporozoite formation has begun and highlights the usefulness of the segmentation score as an alternative.", and the final sentence of the discussion section about this now reads "Considering that oocyst size does not plateau until cytokinesis begins4, measuring oocyst diameter may represent a useful biological clock specifically when investigating the early stages of oocyst development." 8.
__How is the apical polarity different to merozoite as some conoid genes are present in ookinete and sporozoite but not in merozoite. __
Our hypothesis is that apical polarity is established by the positioning and attachment of the centriolar plaque to the parasite plasma membrane in both forming merozoites and sporozoites. While the apical polar ring proteins are obviously present at the apical end, and have important functions, we think that they themselves are unlikely to regulate polarity establishment directly. Additionally, it seems that the apical polar rings are visible in forming sporozoites far before the comparable stages of merozoite formation. An important note here is that at this point, this is largely inferences based on observational differences and there is relatively little functional data on proteins that regulate polarity establishment at any stage of the Plasmodium 9.
__Therefore, I think that electron microscopy remains essential for the observation of such ultra-fine structures __
We have added a paragraph in the discussion that provides a more clear comparison between MoTissU-ExM and other imaging modalities previously applied on mosquito-stage parasites (see response to Reviewer#1 (Minor text changes) comment #17). 10.
__The author have not mentioned that sometimes the stage oocyst development is also dependent on the age of mosquito and it vary between different mosquito gut even if the blood feed is done on same day. __
In our opinion this can be inferred through the more general statement that "development of each oocyst within a midgut is asynchronous..."
Figure changes (Reviewer #2)
-
__Fig 3B: stage 2 and 6 does not show the DNA cyan, it would-be good show the sate of DNA at that particular stage, especially at stage 2 when APR is visible. And box the segment in the parent picture whose subset is enlarged below it. __
We completely agree with the reviewer that the stage 2 image would benefit from the addition of a DNA stain. Many of the images in Figure 3b were done on samples that did not have a DNA stain and so in these * yoelii samples we did not find examples of all segmentation scores with the DNA stain. Examples of segmentation score 2 and 6 for P. berghei, and 6 for P. falciparum* can be found with DNA stains in Figure S8. 2.
__For clarity, it would be helpful to add indicators for the centriolar plaques in Figure 1b, as their locations are not immediately obvious. __
The CPs in Figure 1a and 1b have been circled on the NHS ester only panel for clarity. +
__Regarding Figure 1c, the authors state that 'the rootlet fiber is visible'. However, such a structure cannot be confirmed from the provided NHS ester image. Can the authors present a clearer image where the rootlet fibre is more distinct? Furthermore, please provide the basis for identifying this structure as a rootlet fiber based on the NHS ester observation alone. __
The image in Figure 1c has been replaced with one that more clearly shows the rootlet fibre.
Based on electron microscopy studies, the rootlet fibre has been defined as a protein dense structure that connects the centriolar plaque to the apical polar rings (PMID: 17908361). Through NHS ester and tubulin staining, we could identify the apical polar rings and centriolar plaque as sites on the apical end of the parasite and nucleus that microtubules are nucleated from. There is a protein dense fibre that connects these two structures. Based on the fact that the protein density of this structure was previously considered sufficient for its identification by electron microscopy, we consider its visualisation by NHS ester staining sufficient for its identification by U-ExM.
__Fig 1B - could the tubulin image in the hemispindle panel be made brighter? __
The tubulin staining in this panel was not saturated, and so this change has been made.
__Fig 4A - the green text in the first image panel is not visible. Also, the cyan text in the 3rd image in Fig 1A is also difficult to see. There's a few places where this is the case __
We have made all microscopy labels legible at least when printed in A4/Letter size.
__Fig 6A - how do the authors know ron11 expression is reduced by 99%? Did they test this themselves or rely on data from the lab that gifted them the construct? Also please provide mention the number of oocyst and sporozoites were observed. __
The way Figure 6a was previously designed and described was an oversight, that wrongly suggested we had quantified a >99% reduction in *ron11 * The 99% reduction has been removed from Figure 6a and the corresponding part of the figure legend has been rewritten to emphasise that this was previously established:
"(a) Schematic showing previously established Ron11Ctrl and Ron11cKD parasite lines where ron11 expression was reduced by >99%9."
As to the second part of the question, we did not independently test either protein or RNA level expression of RON11, but we were gifted the clonal parasite lines established by Prof. Ishino's lab in PMID: 31247198 not just the genetic constructs.
__Fig 6E - are the data point colours the wrong way round on this graph? Just looking at the graph it looks as though the RON11cKD has more rhoptries than the control which does not match what is said in the text. __
Thank you for pointing out this mistake, the colours have now been corrected.
__Fig S8C, PbRON11 ctrl, pie chart shows 89.7 % spz are present in the secretory cavity while the text shows 100 %, 35/35 __
The text saying 100% (35/35) only considered salivary glands that were infected (ie. Uninfected SGs were removed from the count. The two sentences that report this data have been clarified to reflect this better:
"Of *PbRON11ctrl SGs that were infected (35/39), 100% (35/35) contained sporozoites in the secretory cavity (Figure S8c). Conversely of infected PbRON11cKD SGs (59/82), only 24% (14/59) contained sporozoites within the secretory cavity (Figure S9d)."
*
__Fig S9D shows that RON11 ckd contains 17.1% sporozoites in secretory cavity while the text says 24%. __
Please see the response to Reviewer#2 Figure Changes Comment #8 where this was addressed.
Experimental changes (Reviewer #2)
-
__Why do the congruent rhoptries have similar lengths to each other, while the dimorphic rhoptries have different lengths? Is this morphological difference related to the function of these rhoptries? __
We hypothesise that this morphological difference arises because the congruent rhoptries are 'used' during SG invasion, while the dimorphic rhoptries are utilized during hepatocyte invasion. It is not straightforward to test this functionally at this point, as no protein is known to have differential localization between the two. Additionally, RON11 is likely directly involved in both SG and hepatocyte invasion through a secreted portion of the protein (as seen in RBC invasion). Therefore, RON11cKD sporozoites may have combined defects, meaning we cannot assume any defect is solely due to the absence of two rhoptries. Determining this functionally is of high interest to our research groups and remains an area of ongoing study, but it is beyond the scope of this study. 2.
Would it be possible to show whether RON11 localises to the dimorphic rhoptries, the congruent rhoptries, or both, by using expansion microscopy and a parasite line that expresses RON11 tagged with GFP or a peptide tag?
__ __We do not have access to a parasite line that expresses a tagged copy of RON11, or anti-PbRON11 antibodies. Based on previously published localisation data, however, it seems likely that RON11 localises to both sets of rhoptries. Below are excerpts from Figure 1c of PMID: 31247198, where RON11 (in green) seems to have a more basally-extended localisation in midgut (MG) sporozoites than in salivary gland (SG) sporozoites. From this we infer that in the MG sporozoite you're seeing RON11 in both pairs of rhoptries, but only the one remaining pair in the SG sporozoite.
__The knockdown of RON11 disrupts the rhoptry structure, making the dimorphic and congruent rhoptries indistinguishable. Does this suggest that RON11 is important for the formation of both types of rhoptries? I believe that it would be crucial to confirm whether RON11 localises to all rhoptries or is restricted to specific rhoptries for a more precise discussion of RON11's function. __
Based on our analysis, it does indeed seem that RON11 is important for both types of rhoptries as when RON11 isn't expressed sporozoites still have both apical and cytoplasmic rhoptries (ie. Not just one pair is lost; see Reviewer #1 Experimental changes comment #1).
__The authors state that 64% of RON11cKD SG sporozoites contained no rhoptries at all. Does this mean RON11cKD SG sporozoites used up all rhoptries corresponding to the dimorphic and congruent pairs during SG invasion? If so, this contradicts your claims that sporozoites are 'leaving the dimorphic rhoptries for hepatocyte invasion' and that 'rhoptry pairs are specialized for different invasion events'. If that is not the case, does it mean that RON11cKD sporozoites failed to form the rhoptries corresponding to the dimorphic pair? A more detailed discussion would be needed on this point and, as I mentioned above, on the specific role of RON11 in the formation of each rhoptry pair. __
We do not agree that this constitutes a contradiction; instead, more nuance is needed to fully explain the phenotype. As shown in the new graph added in response to Reviewer#1 Figure changes comment #1 in RON11cKD oocyst sporozoites, 64% of all rhoptries are located at the apical end. Our hypothesis is that these rhoptries are used for SG invasion and, therefore, would not be present in RON11cKD SG sporozoites. Consequently, the fact that 64% of RON11cKD sporozoites lack rhoptries is exactly what we would expect. Essentially, we predict three slightly different 'pathways' for RON11cKD sporozoites: If they had 2 apical rhoptries in the oocyst, we predict they would have zero rhoptries in the SG. If they had 2 cytoplasmic rhoptries in the oocyst, we predict they would have two rhoptries in the SG. If they had one apical and one cytoplasmic rhoptry in the oocyst, we predict they would have one rhoptry in the SG. In any case, we expect the apical rhoptries to be 'used up,' which appears to be supported by the data.
__Out of pure curiosity, is it possible to measure the length and number of subpellicular microtubules in the sporozoites observed in this study using expansion microscopy? __
We have performed an analysis of subpellicular microtubules which is now included as Supplementary Figure 2. We could not always distinguish every SPMT from each other and so have not quantified SPMT number. We have, however, quantified their absolute length on both the 'long side' and 'short side', their relative length (as % sporozoite length) and the degree to which they are polyglutamylated.
A description of this analysis is now found in the results section as follows: "*We quantified the length and degree of polyglutamylation of SPMTs on the 'long side' and 'short side' of the sporozoite (Figure S2). 'Short side' SPMTs were on average 33% shorter (mean = 3.6 µm {plus minus}SD 1.0 µm) than 'long side' SPMTs (mean = 5.3 µm {plus minus}SD 1.5 µm) and extended 17.4% less of the total sporozoite length. While 'short side' SPMTs were significantly shorter, a greater proportion of their length (87.9% {plus minus}SD 11.2%) was polyglutamylated compared to 'long side' SPMTs (69.4% {plus minus}SD 13.8%)." *
Supplementary Figure 2: Analysis of sporozoite subpellicular microtubules. Isolated P. yoelii salivary gland sporozoites were prepared by U-ExM and stained with anti-tubulin (microtubules) and anti-PolyE (polyglutamylated SPMTs) antibodies. SPMTs were defined as being on either the 'long side' (nucleus distant from plasma membrane) or 'short side' (nucleus close to plasma membrane) of the sporozoite as depicted in Figure 1f. (a) SPMT length along with (b) SPMT length as a proportion of sporozoite length were both measured. (c) Additionally, the proportion of the SPMT that was polyglutamylated was measured. Analysis comprises 25 SPMTs (11 long side, 14 short side) from 6 SG sporozoites. ** = p The following section has also been added to the methods to describe this analysis: * "Subpellicular microtubule measurement
- To measure subpellicular microtubule length and polyglutamylation maximum intensity projections were made of sporozoites stained with NHS Ester, anti-tubulin and anti-PolyE antibodies, and SYTOX Deep Red. The side where the nucleus was closest to the parasite plasma membrane was defined as the 'short side', while the side where the nucleus was furthest from the parasite plasma membrane was defined as the 'long side'. Subpellicular microtubules were then measured using a spline contour from the apical end of the sporozoite to the basal-most end of the microtubule with fluorescence intensity across the contour plotted (Zeiss ZEN 3.8). Sporozoite length was defined as the distance from the sporozoite apical polar rings to the basal complex, measuring through the centre of the cytoplasm. The percentage of the subpellicular microtubule that was polyglutamylated was determined by assessing when along the subpellicular microtubule contour the anti-PolyE fluorescence intensity last dropped below a pre-defined threshold."
*
__In addition to the previous point, in the text accompanying Figure 7a, the authors claim that "64% of PbRON11cKD SG sporozoites contained no rhoptries at all, while 9% contained 1 rhoptry and 27% contained 2 rhoptries". Could this data be used to infer which rhoptry pair are missing from the RON11cKD oocyst sporozoites? Can it be inferred that the 64% of salivary gland sporozoites that had no rhoptries in fact had 2 congruent rhoptries in the oocyst sporozoite stage and that these have been discharged already? __
Please see the response to Reviewer #2 Experimental Changes Comment #4.
__Is it possible that the dimorphic rhoptries are simply precursors to the congruent rhoptries? Could it be that after the congruent rhoptries are used for SG invasion, new congruent rhoptries are formed from the dimorphic ones and are then used for the next invasion?____ Would it be possible to investigate this by isolating sporozoites some time after they have invaded the SG and performing expansion microscopy? This would allow you to confirm whether the dimorphic rhoptries truly remain in the same form, or if new congruent rhoptries have been formed, or if there have been any other changes to the morphology of the dimorphic rhoptries. __
In theory, it is possible that the dimorphic rhoptries are precursors to the uniform rhoptries, specifically how the larger one of the two in the dimorphic pair might be a precursor. Maybe the smaller one is, but we have no evidence to suggest that this rhoptry lengthens after SG invasion. We are interested in isolating sporozoites from SGs to add a temporal perspective, but currently, this isn't feasible. When sporozoites are isolated from SGs, they are collected at all stages of invasion. Additionally, we don't know how long each step of SG invasion takes, so a time-based method might not be effective either. We are developing an assay to better determine the timing of events during SG invasion with MoTissU-ExM, but this is beyond the scope of this study.
__In the section titled "Presence of PbRON11cKD sporozoites in the SG intercellular space", the authors state that "the majority of PbRON11cKD-infected mosquitoes contained some sporozoites in their SGs, but these sporozoites were rarely inside either the SG epithelial cell or secretory cavity". - this is suggestive of an invasion defect as the authors suggest. Could the authors collect these sporozoites and see if liver hepatocyte infection can be established by the mutant sporozoites? They previously speculate that the two different types of rhoptries (congruent and dimorphic) may be specific to the two invasion events (salivary gland epithelial cell and liver cell infection). __
It has already been shown that RON11cKD sporozoites fail hepatocyte invasion (PMID: 31247198), even when isolated from the haemolymph and so it seems very unlikely that they would be invasive following SG isolation. As mentioned in the discussion, RON11 in merozoites has a 'dual-function' where it is partially secreted during merozoite invasion in addition to its rhoptry biogenesis functions. Assuming this is also the case in sporozoites, using the RON11cKD parasite line we cannot differentiate these two functions and therefore cannot ascribe invasion defects purely to issues with rhoptry biogenesis. In order to answer this question functionally, we would need to identify a protein that only has roles in rhoptry biogenesis and not invasion directly.
Reviewer #3
Minor text changes (Reviewer #3)
-
__Page 3 last paragraph: ...the molecular mechanisms underlying SG (invasion?) are poorly understood. __
This has been corrected 2.
__The term "APR" does not refer to a tubulin structure per se, but rather to the proteinaceous structure to which tubulin anchors. Are there any specific APR markers that can be used in Figure 1C? If not, I recommend avoiding the use of "APR" in this context. __
The text does not state that the APR is a tubulin structure. Given that it is a proteinaceous structure, we visualise the APRs through protein density (NHS Ester). It has been standard for decades to define APRs by protein density using electron microscopy, and it has previously been sufficient in Plasmodium using expansion microscopy (PMIDs: 41542479, 33705377) so it is unclear why it should not be done so in this study. 3.
__I politely disagree with the bold statements ‚ Little is known about cell biology of sporozoite formation.....from electron microscopy studies now decades old' (p.3, 2nd paragraph); ‚To date, only a handful of (instead of ‚or') proteins have been implicated in SG invasion' (p. 4, 1st paragraph). These claims may overlook existing studies; a more thorough review of the literature is recommended. __
This study includes at least 50 references from papers broadly related to sporozoite biology, covering publications from every decade since the 1970s. The most recent review that discusses salivary gland invasion cites 11 proteins involved in SG invasion. We have replaced "handful" with a more precise term, as it is not the best adjective, but it is hardly an exaggeration.
Figure changes (Reviewer #3)
-
__The hypothesis that Plasmodium utilizes two distinct rhoptry pairs for invading the salivary gland and liver cells is intriguing but remains clearly speculative. Are the "cytoplasmic pair" and "docked pair" composed of the same secretory proteins? Are the paired rhoptries identical? How does the parasite determine which pair to use for salivary gland versus liver cell invasion? Is there any experimental evidence showing that the second pair is activated upon successful liver cell invasion? Without such data this hypothesis seems rather premature. __
We are unaware of any direct protein localisation evidence suggesting that the rhoptry pairs may carry different cargo. However, only a few proteins have been localised in a way that would allow us to determine if they are associated with distinct rhoptry pairs, so this possibility cannot be ruled out either. It seems unlikely that the parasite 'selects' a specific pair, as rhoptries are typically always found at the apical end. What appears more plausible is that the "docked pair" forms first and immediately occupies the apical docking site, preventing the cytoplasmic pair from docking there. Regarding any evidence that the second pair is activated during liver cell invasion, it has been well documented over decades that rhoptries are involved in hepatocyte invasion. If the dimorphic rhoptries are the only ones present in the parasite during hepatocyte invasion, then they must be used for this process. 2.
__The quality of the "Roolet fibre" image is not good and resembles background noise from PolyE staining. Additional or alternative images should be provided to convincingly demonstrate that PolyE staining indeed visualizes the Roolet fibre. It is puzzling that the structure is visible with PolyE staining but not with tubulin staining. __
This is a logical misinterpretation based on the image provided in Figure 1c. Our intention was not to imply that PolyE staining enables us to see the rootlet fibre but that PolyE and tubulin allow us to see the APR to which the rootlet fibre is connected. There is some PolyE staining that likely corresponds to the early SPMTs that in 1c appears to run along the rootlet fibre but this is a product of the max-intensity projection. Please see Reviwer#2 Figure Changes Comment #3 for the updated Figure 1c. 3.
__More arrows should be added to Figures 6b and 6c to guide readers and improve clarity. __
We have added arrows to Figure 6b and 6c which point out what we have defined as normal and aberrant rhoptries more clearly. These panels now look like this: 4.
__Figure 2a zoomed image of P. yoelii infected SG is different than the highligted square. __
We agree that the highlighted square and the zoomed area appear different, but this is due to the differing amounts of light captured by the objectives used in these two panels. The entire SG panel was captured with a 5x objective, while the zoomed panel was captured with a 63x objective. Because of this difference, the plane of focus of the zoomed area is hard to distinguish in the whole SG image. The zoomed image is on the 'top' of the SG (closest to the coverslip), while most of the signal you see in the whole SG image comes from the 'middle' of the SG. To demonstrate this more clearly, we have provided the exact region of interest shown in the 63x image alongside a 5x image and an additional 20x image, all of which are clearly superimposable.__
__ 5.
__Figure 3 legend: "P. yoelii infected midguts harvested on day 15" should be corrected. More general, yes, "...development of each oocyst within a single midgut is asynchronous." but it is still required to provide the dissection days. __
We are unsure what the suggested change here is. We do not know what is wrong with the statement about day 15 post infection, that is when these midguts were dissected. __ Experimental Changes (Reviewer #3)__
-
__The proposed role of AOR in rhoptry biogenesis appears highly speculative. It is unclear how the authors conclude that "AORs carry rhoptry cargo" solely based on the presence of RON4 within the structure. Inclusion of additional markers to characterize the content of AOR and rhoptries will be essential to substantiate the hypothesis that this enigmatic structure supports rhoptry biogenesis. __
It is important to note that the hypothesis that AORs, or rhoptry anlagen, carry rhoptry cargo and serve as vehicles of rhoptry biogenesis was proposed long before this study (PMID: 17908361). In that study, it was assumed that structures now called AORs or rhoptry anlagen were developing rhoptries. Although often visualised by EM and presumed to carry rhoptry cargo (PMID: 33600048, 26565797, 25438048), it was only more recently that AORs became the subject of dedicated investigation (PMID: 31805442), where the authors stated that "...AORs could be immature rhoptr[ies]...". Our observation that AORs contain the rhoptry protein RON4, which is not known to localize to any other organelle, we therefore consider sufficient to conclude that AORs carry rhoptry cargo and are thus vehicles for rhoptry biogenesis. 2.
__The study of RON11 appears to be a continuation of previous work by a collaborator in the same group. However, neither this study nor the previous one adequately addresses the evolutionary context or structural characteristics of RON11. Notably, the presence of an EF-hand motif is an important feature, especially considering the critical role of calcium signaling in parasite stage conversion. Given the absence of a clear ortholog, it would be interesting to know whether other Apicomplexan parasites harbor rhoptry proteins with transmembrane domains and EF-hand motifs, and if these proteins might respond similarly to calcium stimulation. Investigating mutations within the EF-hand domain could provide valuable functional insights into RON11. __
We are unsure what suggests that RON11 lacks a clear orthologue. RON11 is conserved across all apicomplexans and is also present in Vitrella brassicaformis (OrthoMCL orthogroup: OG7_0028843). A phylogenetic comparison of RON11 across apicomplexans has previously been performed (PMID: 31247198), and this study provides a structural prediction of PbRON11 with the dual EF-hand domains annotated (Supplementary Figure 9). 3.
__The study cannot directly confirm that membrane fusion occurs between rhoptries and AORs. __
This is already stated verbatim in the results "Our data cannot directly confirm that membrane fusion occurs between rhoptries and AORs..." 4.
__It is unclear what leads to the formation of the aberrant rhoptries observed in RON11cKD sporozoites. Since mosquitoes were not screened for infection prior to salivary gland dissection, The defect reports and revisited of RON11 knockdown does not aid in interpreting rhoptry pair specialization, as there was no consistent trend as to which rhoptry pair was missing in RON11cKD oocyst sporozoites. The notion that RON11cKD parasites likely have ‚combinatorial defects that effect both rhoptry biogenesis and invasion' poses challenges to understand the molecular role(s) of RON11 on biogenesis versus invasion. Of note, RON11 also plays a role in merozoite invasion. __
We are unclear about the comment or suggestion here, as the claims that RON11cKD does not help interpret rhoptry pair specialization, and that these parasites have combined defects, are both directly stated in the manuscript. 5.
__Do all SG PbRON11cKD sporozoites lose their reduced number of rhoptries during SG invasion as in Figure 7a (no rhoptries)? __
Not all RON11cKD SG sporozoites 'use up' their rhoptries during SG invasion. This is quantified in both Figure 7a and the text, which states: "64% of *PbRON11cKD SG sporozoites contained no rhoptries at all, while 9% contained 1 rhoptry and 27% contained 2 rhoptries."
* 6.
Different mosquito species/strains are used for P. yoelii, P. berghei, and P. falciparum. Does it effect oocyst sizes/stages? Is it ok to compare?
__ __We agree that a direct comparison between for example * yoelii and P. berghei *oocyst size would be inappropriate, however Figure 3c and Supplementary Figure 4 are not direct comparisons between two species, but a summation of all oocysts measured in this study to indicate that the trends we observe transcend parasite/mosquito species differences. Our study was not set up with the experimental power to determine if mosquito host species alter oocyst size. 7.
__While I acknowledge that UExM has significantly advanced resolution capabilities in parasite studies, the value of standard microscopy technique should not be overlooked. Particularly, when discussing the function of RON11, relevant IFA and electron microscopy (EM) images should be included to support claims about RON11's role in rhoptry biogenesis. This would complement the UExM data and substantially strengthen the conclusions. Importantly, UExM can sometimes produce unexpected localization patterns due to the denaturation process, which warrants caution. __
The purpose of this study is not to discredit, undermine, or supersede other imaging techniques. It is simply to use U-ExM to answer biological questions that cannot or have not been answered using other techniques. Please refer to Reviewer # 1 Minor text changes comment#17 to see the new paragraph "Comparison of MoTissU-ExM and other imaging modalities" that addresses this
Both conventional IFA and immunoEM have already been performed on RON11 in sporozoites before (PMID: 31247198). When assessing defects caused by RON11 knockdown, conventional IFA isn't especially helpful because it doesn't allow visualization of individual rhoptries. Thin-section TEM also doesn't provide the whole-cell view needed to draw these kinds of conclusions. Volume EM could likely support these observations, but we don't have access to or expertise in this technique, and we believe it is beyond the scope of this study. It's also important to note that for the defect we observe-missing or abnormal rhoptries-the visualization with NHS ester isn't significantly different from what would be seen with EM-based techniques, where rhoptries are easily identified based on their protein density.
The statement that "UExM can sometimes produce unexpected localisation patterns due to the denaturation process..." is partially correct but lacks important nuance in this context. Based on our extensive experience with U-ExM, there are two main reasons why the localisation of a single protein may look different when comparing U-ExM and traditional IFA images. First, denaturation: in conventional IFAs, antibodies need to recognize conformational epitopes to bind to their target, whereas in U-ExM, antibodies must recognize linear epitopes. This doesn't mean the target protein's localisation changes, only that the antibody's ability to recognize it does. Second, antibody complexes seem unable to freely diffuse out of the gel, which can result in highly fluorescent signals not related to the target protein appearing in the image, as we have previously reported (PMID: 36993603). Importantly, neither of these factors applies to our phenotypic analysis of RON11 knockdown. All phenotypes described are based solely on NHS Ester (total protein) staining, so the considerations about changes in the localisation of individual proteins are not relevant.
-
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Referee #3
Evidence, reproducibility and clarity
Overall, the manuscript is well-written and -structured. However, I would like to raise several major points for consideration:
- While I acknowledge that UExM has significantly advanced resolution capabilities in parasite studies, the value of standard microscopy technique should not be overlooked. Particularly, when discussing the function of RON11, relevant IFA and electron microscopy (EM) images should be included to support claims about RON11's role in rhoptry biogenesis. This would complement the UExM data and substantially strengthen the conclusions. Importantly, UExM can sometimes produce unexpected localization patterns due to the denaturation process, which warrants caution.
- The proposed role of AOR in rhoptry biogenesis appears highly speculative. It is unclear how the authors conclude that "AORs carry rhoptry cargo" solely based on the presence of RON4 within the structure. Inclusion of additional markers to characterize the content of AOR and rhoptries will be essential to substantiate the hypothesis that this enigmatic structure supports rhoptry biogenesis.
- The hypothesis that Plasmodium utilizes two distinct rhoptry pairs for invading the salivary gland and liver cells is intriguing but remains clearly speculative. Are the "cytoplasmic pair" and "docked pair" composed of the same secretory proteins? Are the paired rhoptries identical? How does the parasite determine which pair to use for salivary gland versus liver cell invasion? Is there any experimental evidence showing that the second pair is activated upon successful liver cell invasion? Without such data this hypothesis seems rather premature.
- The study of RON11 appears to be a continuation of previous work by a collaborator in the same group. However, neither this study nor the previous one adequately addresses the evolutionary context or structural characteristics of RON11. Notably, the presence of an EF-hand motif is an important feature, especially considering the critical role of calcium signaling in parasite stage conversion. Given the absence of a clear ortholog, it would be interesting to know whether other Apicomplexan parasites harbor rhoptry proteins with transmembrane domains and EF-hand motifs, and if these proteins might respond similarly to calcium stimulation. Investigating mutations within the EF-hand domain could provide valuable functional insights into RON11.
- The study cannot directly confirm that membrane fusion occurs between rhoptries and AORs.
- It is unclear what leads to the formation of the aberrant rhoptries observed in RON11cKD sporozoites. Since mosquitoes were not screened for infection prior to salivary gland dissection, The defect reports and revisited of RON11 knockdown does not aid in interpreting rhoptry pair specialization, as there was no consistent trend as to which rhoptry pair was missing in RON11cKD oocyst sporozoites. The notion that RON11cKD parasites likely have ‚combinatorial defects that effect both rhoptry biogenesis and invasion' poses challenges to understand the molecular role(s) of RON11 on biogenesis versus invasion. Of note, RON11 also plays a role in merozoite invasion. I like the introduction of a segmentation score to Plasmodium oocyst maturation.
Minor comments:
- The term "APR" does not refer to a tubulin structure per se, but rather to the proteinaceous structure to which tubulin anchors. Are there any specific APR markers that can be used in Figure 1C? If not, I recommend avoiding the use of "APR" in this context.
- The quality of the "Roolet fibre" image is not good and resembles background noise from PolyE staining. Additional or alternative images should be provided to convincingly demonstrate that PolyE staining indeed visualizes the Roolet fibre. It is puzzling that the structure is visible with PolyE staining but not with tubulin staining.
- Figure 2a zoomed image of P. yoelii infected SG is different than the highligted square.
- Figure 3 legend: "P. yoelii infected midguts harvested on day 15" should be corrected. More general, yes, "...development of each oocyst within a single midgut is asynchronous." but it is still required to provide the dissection days.
- More arrows should be added to Figures 6b and 6c to guide readers and improve clarity.
- Do all SG PbRON11cKD sporozoites lose their reduced number of rhoptries during SG invasion as in Figure 7a (no rhoptries)?
- Different mosquito species/strains are used for P. yoelii, P. berghei, and P. falciparum. Does it effect oocyst sizes/stages? Is it ok to compare?
- I politely disagree with the bold statements ‚ Little is known about cell biology of sporozoite formation.....from electron microscopy studies now decades old' (p.3, 2nd paragraph); ‚To date, only a handful of (instead of ‚or') proteins have been implicated in SG invasion' (p. 4, 1st paragraph). These claims may overlook existing studies; a more thorough review of the literature is recommended.
- Page 3 last paragraph: ...the molecular mechanisms underlying SG (invasion?) are poorly understood.
Significance
In this study, the authors explore Ultrastructure Expansion Microscopy (U-ExM) in Plasmodium-infected mosquito tissue with the aim to enhance the visualization of parasite ultrastructure. For this purpose, they revisit sporogony, the maturation of sporozoites inside oocysts, and sporozoite invasion of salivary glands, which has been studied both by cell biological methods and experimental genetics over four decades. They focus their analysis on the biogenesis and function of key secretory organelles, termed rhoptries, which are central to parasite invasion and, again, have been studied extensively.
This study is a follow-up of a previous study by the same authors (Ref. 19). In the former study the authors showed that U-ExM allows to visualize subcellular structures in sporozoites, including the nucleus, rhoptries, Golgi, apical polar rings (APR), and basal complex, as well as midgut-associated oocysts with developing sporozoites. Here, the authors claim a new finding by stating that sporozoites possess two distinct rhoptry pairs. Supposedly, only one pair is utilized during salivary gland invasion. The authors suggest specialization of rhoptries for different cell invasion events. The authors also revisit a RON11 knock-down parasite line, which was previously shown to be deficient in salivary gland invasion, host cell attachment, gliding locomotion, and liver invasion (Ref. 14).
I find it difficult to estimate the significance. Obviously, attention will be limited to Plasmodium researchers only, as this study is descriptive and revisits a well-studied aspect of the Plasmodium life cycle in the Anopheles vector.
-
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Referee #2
Evidence, reproducibility and clarity
The manuscript by Liffner et al have used the modified expansion microscopy as they term Mosquito Tissue Ultrastructure Expansion Microscopy (MoTissU-ExM) to study a cell biology of temporal development of malaria parasite sporozoite biogenesis within mosquito host. They employed three different malaria parasite models Plasmodium yoelii, P.beghei and P falciparum and infected them in mosquito host.
The application of MoTissU-ExM to infected mosquito tissues is a significant technical advance, enabling visualizations previously only achievable with electron microscopy.
The major conclusion and advances are as following
- The establishment of a "segmentation score" as a great tool for staging asynchronous oocyst development.
- The location of Centriolar plaques, rootlet and other structures which are difficult to analyse
- The first detailed timeline for sporozoite rhoptry biogenesis.
- Clear quantification showing that sporozoites possess four rhoptries and utilise two during salivary gland (SG) invasion.
- A characterization of the RON11 knockout phenotype, linking it to defects in rhoptry biogenesis and a specific block in SG epithelial cell invasion. The following points are intended to further strengthen the paper for publication.
Points for Revision
- For clarity, it would be helpful to add indicators for the centriolar plaques in Figure 1b, as their locations are not immediately obvious.
- The 'image Z-depth' value indicated in the figures is ambiguous. It is not clear whether this refers to the distance from the coverslip surface or the starting point of the z-stack image acquisition. A precise definition of this parameter would be beneficial.
- Regarding Figure 1c, the authors state that 'the rootlet fiber is visible'. However, such a structure cannot be confirmed from the provided NHS ester image. Can the authors present a clearer image where the rootlet fibre is more distinct? Furthermore, please provide the basis for identifying this structure as a rootlet fiber based on the NHS ester observation alone.
- Why do the congruent rhoptries have similar lengths to each other, while the dimorphic rhoptries have different lengths? Is this morphological difference related to the function of these rhoptries?
- Would it be possible to show whether RON11 localises to the dimorphic rhoptries, the congruent rhoptries, or both, by using expansion microscopy and a parasite line that expresses RON11 tagged with GFP or a peptide tag?
- The knockdown of RON11 disrupts the rhoptry structure, making the dimorphic and congruent rhoptries indistinguishable. Does this suggest that RON11 is important for the formation of both types of rhoptries? I believe that it would be crucial to confirm whether RON11 localises to all rhoptries or is restricted to specific rhoptries for a more precise discussion of RON11's function.
- The authors state that 64% of RON11cKD SG sporozoites contained no rhoptries at all. Does this mean RON11cKD SG sporozoites used up all rhoptries corresponding to the dimorphic and congruent pairs during SG invasion? If so, this contradicts your claims that sporozoites are 'leaving the dimorphic rhoptries for hepatocyte invasion' and that 'rhoptry pairs are specialized for different invasion events'. If that is not the case, does it mean that RON11cKD sporozoites failed to form the rhoptries corresponding to the dimorphic pair? A more detailed discussion would be needed on this point and, as I mentioned above, on the specific role of RON11 in the formation of each rhoptry pair.
- Out of pure curiosity, is it possible to measure the length and number of subpellicular microtubules in the sporozoites observed in this study using expansion microscopy?
- Is it possible that the dimorphic rhoptries are simply precursors to the congruent rhoptries? Could it be that after the congruent rhoptries are used for SG invasion, new congruent rhoptries are formed from the dimorphic ones and are then used for the next invasion? Would it be possible to investigate this by isolating sporozoites some time after they have invaded the SG and performing expansion microscopy? This would allow you to confirm whether the dimorphic rhoptries truly remain in the same form, or if new congruent rhoptries have been formed, or if there have been any other changes to the morphology of the dimorphic rhoptries.
- In addition to the previous point, in the text accompanying Figure 7a, the authors claim that "64% of PbRON11cKD SG sporozoites contained no rhoptries at all, while 9% contained 1 rhoptry and 27% contained 2 rhoptries". Could this data be used to infer which rhoptry pair are missing from the RON11cKD oocyst sporozoites? Can it be inferred that the 64% of salivary gland sporozoites that had no rhoptries in fact had 2 congruent rhoptries in the oocyst sporozoite stage and that these have been discharged already?
- In the section titled "Presence of PbRON11cKD sporozoites in the SG intercellular space", the authors state that "the majority of PbRON11cKD-infected mosquitoes contained some sporozoites in their SGs, but these sporozoites were rarely inside either the SG epithelial cell or secretory cavity". - this is suggestive of an invasion defect as the authors suggest. Could the authors collect these sporozoites and see if liver hepatocyte infection can be established by the mutant sporozoites? They previously speculate that the two different types of rhoptries (congruent and dimorphic) may be specific to the two invasion events (salivary gland epithelial cell and liver cell infection).
There are a few typing errors in the document:
- Paragraph 3 of the introduction - line 7, "handful or proteins" should be handful of proteins
- Paragraph 5 of the introduction - line 7, "also able to observed" should be observe
- In the final paragraph of the introduction - line 1, "leverage this new understand" should be understanding
- The first paragraph of the discussion summary contains an incomplete sentence on line 7, "PbRON11ctrl-infected SGs."
- The second paragraph of the discussion - line 10, "until cytokinesis beings" should be begins
Some suggestions for figures
Fig 1B - could the tubulin image in the hemispindle panel be made brighter?
Fig 3B: stage 2 and 6 does not show the DNA cyan, it would-be good show the sate of DNA at that particular stage, especially at stage 2 when APR is visible. And box the segment in the parent picture whose subset is enlarged below it.
Fig 4A - the green text in the first image panel is not visible. Also, the cyan text in the 3rd image in Fig 1A is also difficult to see. There's a few places where this is the case
Fig 6A - how do the authors know ron11 expression is reduced by 99%? Did they test this themselves or rely on data from the lab that gifted them the construct? Also please provide mention the number of oocyst and sporozoites were observed.
Fig 6E - are the data point colours the wrong way round on this graph? Just looking at the graph it looks as though the RON11cKD has more rhoptries than the control which does not match what is said in the text.
Fig S8C, PbRON11 ctrl, pie chart shows 89.7 % spz are present in the secretory cavity while the text shows 100 %, 35/35
Fig S9D shows that RON11 ckd contains 17.1% sporozoites in secretory cavity while the text says 24%.
Some point to discuss
1.One minor point that author suggest that oocyst diameter is not appropriate for the development of sporozoite develop. This is not so true as oocyst diameter tells between cell division and cell growth so it is important parameter especially where the proliferation with oocyst does not take place but the growth of oocyst takes place.<br /> 2. The author have not mentioned that sometimes the stage oocyst development is also dependent on the age of mosquito and it vary between different mosquito gut even if the blood feed is done on same day. 3. How is the apical polarity different to merozoite as some conoid genes are present in ookinete and sporozoite but not in merozoite.
Significance
The following aspects are important:
This is novel and more cell biology approach to study the challenging stage of malaria parasite within mosquito. By using MoTissU-ExM, the authors have enabled the three-dimensional observation of ultrastructures of oocyst-sporozoite development that were previously difficult to observe with conventional electron microscopy alone. This includes the developmental process and entire ultrastructure of oocysts and sporozoites, and even the tissue architecture of the mosquito salivary gland and its epithelia cells.
Advances:
By observing sporozoites formation within the oocyst and the overall ultrastructure of the sporozoite with MoTissU-ExM, the authors have provided detailed descriptions of the complete structure and three-dimensional spatial relationships of the rhoptries, rootlet fibre, nucleus, and other organelles. Furthermore, their detailed localisation analysis of sporozoites within the salivary gland is also a great achievement. Considering that such observations were technically and laboriously very difficult with conventional electron microscopy, enabling these analyses with higher efficiency and relatively lower difficulty represents a major contribution to the future advancement of oocyst-sporozoite biology. The development of the 'segmentation score' for sporozoite formation within the oocyst is another major advance. I think this will enable detailed descriptions of structural changes at each developmental stage and of the molecular mechanisms involved in the development of oocysts-sporozoites This has its advantages if antibodies can be used and somewhat reduces the need for immuno-EM. Secondly, in terms of sporozoite rhoptry biology, the Schrevel et al Parasitology 2007 seems to only focus on oocyst sporozoite rhoptries as they say that the sporozoites have 4 rhoptries. This study on the other hand also looks at salivary gland sporozoites and shows that there are potentially important differences between the two - namely the reduction from 4 rhoptries to two. This also leads to further questions about the different types of rhoptries in oocyst sporozoites and whether they're adapted to invasion of different cell types (sal gland epithelial cells or liver hepatocytes)
Limitation
It would be that expansion microscopy alone still has its limits when it comes to observing ultra-fine structures. For example, visualising the small vesicular structures that Schrevel et al. observed in detail with electron microscopy, or seeing ultra-high resolution details such as the fusion of membrane structures and their interactions with structures like the rootlet fibre and microtubules. Therefore, I think that electron microscopy remains essential for the observation of such ultra-fine structures The real impact of this work is mostly cell biologist working with malaria parasite and more in mosquito stages. But the approaches can be applied to any material from any species where temporal dynamics need to be studied with tissue related structures and where UExM can be applied. I am parasite cell biologist working with parasites stages within mosquito vector host.
-
-
www.youtube.com www.youtube.com
-
Violences Institutionnelles : Analyse et Perspectives Juridiques et Pratiques
Synthèse Exécutive
Ce document de synthèse analyse les dimensions multiples des violences institutionnelles, en s'appuyant sur une expertise croisée du droit, des politiques publiques et de la recherche en sciences sociales.
Il ressort que la notion de "violence institutionnelle" est complexe, marquée par une ambiguïté juridique persistante malgré des avancées législatives récentes.
Le terme de "maltraitance institutionnelle" est souvent privilégié pour souligner la relation de pouvoir asymétrique inhérente entre l'institution et l'usager.
Les points critiques à retenir sont les suivants :
1. Une Définition Juridique Incomplète : La loi du 7 février 2022 a introduit dans le Code de l'Action Sociale et des Familles (art. L119-1) une définition de la maltraitance qui englobe l'origine institutionnelle.
Cependant, elle ne définit pas spécifiquement ce que constitue la "maltraitance institutionnelle", laissant une marge d'interprétation et posant des défis en matière de qualification et de traitement.
2. Un Phénomène Peu Quantifié : Il existe une carence significative de données statistiques publiques permettant de mesurer l'ampleur des violences institutionnelles en France.
Les données disponibles indiquent toutefois une forte exposition des professionnels du secteur social et de la santé à la violence, à des niveaux comparables à ceux des forces de l'ordre, ce qui témoigne d'un climat de travail particulièrement difficile.
3. Des Responsabilités Partagées : La lutte contre la maltraitance institutionnelle ne peut se limiter à la sanction des fautes individuelles.
Elle engage des chaînes de responsabilité plurielles et complexes, impliquant les professionnels, les institutions, et plus largement la société dans sa capacité à définir des seuils de tolérance et à protéger les plus vulnérables.
4. L'Importance Cruciale du Soutien Organisationnel : Une étude menée à la Ville de Paris révèle que le bien-être des professionnels du travail social n'est pas corrélé au nombre d'actes de violence subis, mais plutôt à la qualité du soutien organisationnel perçu.
La "détresse morale", liée au manque de marges de manœuvre pour répondre adéquatement aux besoins des usagers, est également un facteur déterminant.
Ces constats identifient le soutien aux équipes et le renforcement de l'autonomie professionnelle comme des leviers d'action stratégiques pour la prévention.
1. Le Cadre Conceptuel et Juridique des Violences Institutionnelles
1.1. Ambiguïtés Sémantiques : Violence vs. Maltraitance
Une distinction fondamentale est établie entre les notions de "violence" et de "maltraitance".
Alors que la violence peut survenir dans n'importe quel contexte, la maltraitance se caractérise par une relation asymétrique de pouvoir ou de dépendance entre l'auteur et la victime.
Dans le contexte institutionnel, la victime se trouve dans une position d'infériorité dont il lui est difficile de s'extraire.
• Perspective de la recherche : La littérature scientifique suggère de privilégier le terme de "maltraitance institutionnelle", car elle implique une relation de pouvoir où la victime est en position d'infériorité, ce qui est particulièrement vrai pour les enfants relevant de l'aide sociale à l'enfance.
• Perspective des personnes concernées :
Le plaidoyer d'ATD Quart Monde ("Stop à la maltraitance institutionnelle", septembre 2024) met en lumière le caractère systémique du phénomène et la forte exposition des personnes en situation de pauvreté.
Une citation issue de ce travail illustre la dépendance de la personne vis-à-vis de l'institution :
La maltraitance institutionnelle peut prendre deux formes :
1. Une réalité factuelle et objectivable : Des actes pouvant constituer des infractions pénales (violences, négligences graves).
2. Une réalité subjective : Le vécu ou le ressenti d'une personne qui s'estime victime, même en l'absence d'infraction pénale caractérisée.
1.2. L'Évolution du Droit et des Politiques Publiques
La reconnaissance des violences institutionnelles dans le droit et les politiques publiques a progressé par à-coups successifs.
| Année | Événement Clé | Contribution | | --- | --- | --- | | 1970 | Opération "pouponnière" lancée par Simone Veil. | Première action ciblée sur les violences institutionnelles envers les enfants, en parallèle des travaux sociologiques d'Erving Goffman sur l'"institution totale". | | Années 2000 | Loi du 2 janvier 2002. | Promotion des droits des usagers pour pallier l'asymétrie de la relation avec l'institution et favoriser l'expression des victimes. | | 2008 | Réforme constitutionnelle. | Création du Défenseur des droits, permettant notamment au Défenseur des enfants de recevoir des réclamations individuelles. | | 2022 | Loi du 7 février 2022. | Première définition légale de la maltraitance dans le secteur social et médico-social. | | 2022 | Loi du 21 mars 2022. | Amélioration de la protection des lanceurs d'alerte, un enjeu connexe à la révélation des dysfonctionnements institutionnels. |
En matière de protection de l'enfance spécifiquement, la terminologie a évolué, passant des "maltraitances" et "mauvais traitements" (loi de 1989) à la notion de "danger" (loi de 2007), pour finalement réintégrer les termes d'"enfant victime de violence" et d'"enfant maltraité" dans les lois de 2016 et 2022.
1.3. La Définition de la Maltraitance par la Loi du 7 février 2022
L'article L119-1 du Code de l'Action Sociale et des Familles (CASF) constitue une avancée majeure. Il définit la maltraitance comme suit :
"La maltraitance [...] vise toute personne en situation de vulnérabilité lorsqu'un geste, une parole, une action ou un défaut d'action compromet ou porte atteinte à son développement, à ses droits, à ses besoins fondamentaux ou à sa santé [...] et que cette atteinte intervient dans une relation de confiance, de dépendance, de soin ou d'accompagnement.
Les situations de maltraitance peuvent être ponctuelles ou durables, intentionnelles ou non.
Leur origine peut être individuelle, collective ou institutionnelle."
Analyse de cette définition :
• Points positifs : Elle est large, reconnaît la vulnérabilité de la personne et la relation de dépendance.
Elle dissocie la maltraitance de l'infraction pénale, permettant de qualifier des situations sans qu'un délit soit nécessairement constitué.
Elle nomme explicitement l'origine "institutionnelle".
• Limites : Le texte ne définit pas ce qu'est la maltraitance institutionnelle en soi.
Par ailleurs, cette approche se heurte à la logique du droit pénal, qui repose sur le principe de la responsabilité personnelle et ne prévoit pas d'infraction spécifique liée au contexte institutionnel ou à la vulnérabilité des publics accompagnés.
2. Quantification et Mesure du Phénomène
2.1. Un Manque de Données Statistiques
Un obstacle majeur à la compréhension et à la lutte contre les violences institutionnelles est l'absence de quantification claire dans la statistique publique.
Les enquêtes nationales (ONPE, INED) fournissent peu d'éléments spécifiques sur ce phénomène, ce qui rend son ampleur difficile à évaluer.
2.2. L'Exposition des Professionnels à la Violence
Malgré le manque de données globales, les chiffres sur la violence subie par les professionnels sont révélateurs du climat dans le secteur social.
• Les données de la fonction publique montrent que les professions intermédiaires de la santé et du travail social sont particulièrement victimes de violence dans l'exercice de leurs fonctions.
• Leur niveau d'exposition à la violence est presque aussi élevé que celui des forces de l'ordre, ce qui souligne l'intensité des tensions et la possible banalisation de la violence dans ce champ.
• Un très faible pourcentage de ces violences fait l'objet d'une plainte et aboutit à une condamnation pénale, ce qui constitue un enjeu majeur pour la reconnaissance des préjudices subis.
3. La Question Centrale de la Responsabilité
3.1. Dépassement de la Responsabilité Individuelle
La Commission nationale de lutte contre les maltraitances souligne que la maltraitance institutionnelle et la responsabilité individuelle ne sont pas exclusives l'une de l'autre.
Il est essentiel de distinguer les comportements individuels déviants des dysfonctionnements collectifs ou systémiques qui engagent la société tout entière.
L'enjeu est de ne pas réduire la maltraitance institutionnelle à une simple somme de fautes professionnelles.
3.2. Des Chaînes de Responsabilité Plurielles
La protection de l'enfant, en particulier, met en jeu des chaînes de responsabilité complexes et entremêlées :
• Responsabilité familiale : Souvent déjà mise à mal dans les situations de protection.
• Responsabilité des professionnels : Directement en contact avec les usagers.
• Responsabilité des institutions : Liée à l'organisation, aux moyens, à la culture interne.
• Responsabilité sociétale : Reflétant les seuils de tolérance collectifs et les dispositifs mis en place pour protéger les plus vulnérables.
De plus, la jurisprudence européenne se montre de plus en plus ferme, ayant déjà condamné la France pour des dysfonctionnements dans son dispositif de protection de l'enfance sur le motif de traitement inhumain et dégradant.
4. Perspectives et Leviers d'Action : L'Étude de la Ville de Paris
En partenariat avec l'Université de Lille, l'Observatoire social de la Ville de Paris a lancé en 2023 une étude sur les violences institutionnelles, axée sur le vécu des professionnels des politiques sociales (protection de l'enfance, autonomie, etc.).
4.1. Principaux Enseignements Préliminaires
1. Forte Exposition, Fort Engagement : L'étude confirme une forte exposition des professionnels à la violence, mais révèle également un niveau d'engagement au travail particulièrement élevé.
2. Le Rôle Clé du Soutien Organisationnel : De manière contre-intuitive, le bien-être au travail des professionnels n'est pas directement corrélé au nombre d'actes de violence subis.
Le facteur le plus déterminant est le soutien organisationnel perçu par les agents.
Un professionnel qui se sent soutenu par son institution vivra mieux son quotidien, même dans un contexte de violence.
3. L'Impact de la "Détresse Morale" : Le second facteur déterminant est la "détresse morale".
Ce concept, issu de travaux canadiens, décrit le sentiment d'impuissance des professionnels qui estiment ne pas avoir les marges de manœuvre ou les moyens nécessaires pour répondre de manière satisfaisante aux besoins des usagers.
4.2. Pistes de Travail Identifiées
Ces résultats, bien que préliminaires, ouvrent des pistes d'action concrètes pour prévenir la maltraitance institutionnelle en agissant sur le climat de travail et le bien-être des professionnels.
Les leviers identifiés sont :
• Renforcer le soutien organisationnel : Mettre en place des dispositifs d'écoute, de reconnaissance et d'appui concrets pour les équipes.
• Améliorer le soutien en équipe : Favoriser la cohésion et l'entraide entre collègues.
• Accroître les marges de manœuvre : Redonner aux professionnels la capacité d'agir de manière adaptée aux situations, réduisant ainsi la détresse morale.
• Travailler sur l'éthique et les valeurs partagées : Consolider une culture professionnelle commune pour guider l'action dans des contextes complexes.
-
-
www.examtopics.com www.examtopics.com
-
NOTA: Cada respuesta correcta vale un punto.
El código configura la duración del bloqueo para la cola.
El último mensaje leído permanece en la cola después de que se ejecuta el código.
La cola de almacenamiento permanece en la cuenta de almacenamiento después de que se ejecuta el código.
-
-
www.kjzz.org www.kjzz.org
-
Conozca a una de las pocas personas
Investigar y redactar una entrevista a una persona de la comunidad que tenga un oficio análogo y/o que sea experto en un saber tradicional o local
-
-
www.google.com www.google.com
-
¿cómo se podrìa convertir la escuela en un laboratorio de experimentaciòn tecnológica que actúe en función de su mejoramiento y bienestar?
Tags
Annotators
URL
-
-
perio.unlp.edu.ar perio.unlp.edu.ar
-
fontanas
tensión entre la incertidumbre humana y la búsqueda tecnológica de una certeza absoluta
-
Chilled by the Present, its gloom and its noise,On waking we sigh for an ancient South,A warm nude age of instinctive poise,A taste of joy in an innocent mouth.At night in our huts we dream of a partIn the balls of the Future: each ritual mazeHas a musical plan, and a musical heartCan faultlessly follow its faultless ways.We envy streams and houses that are sure,But, doubtful, articled to error, weWere never nude and calm as a great door.And never will be faultless like our fountains:We live in freedom by necessity,A mountain people dwelling among mountains.[Helados por el Presente, su pesadumbre y su ruido,al despertar suspiramos por un Sur antiguo,una cálida y desnuda era de instintivo aplomo,en boca inocente, un sabor a gozo.De noche, en nuestros refugios, soñamos tener un huecoen los bailes del Futuro: cada laberinto ritualviene con un plano musical, y un corazón musicala la perfección puede orientarse por tan perfectos recovecos.Envidiamos los arroyos y las casas que son seguros,pero, presa de la duda, aprendices del error,nunca cual puerta grande estuvimos tranquilamente desnudos,y jamás seremos impecables como nuestras fontanas:vivimos en libertad por necesidad,pueblo montaraz que vive entre montañas.]W. H. AUDEN, Sonnets from China, XVIII
Traducción del poema de Auden: Helados por el Presente, su oscuridad y su ruido, Al despertar suspiramos por un antiguo Sur, Una época cálida y desnuda de aplomo instintivo, Un sabor de alegría en una boca inocente. Por la noche, en nuestras chozas soñamos con una parte En los bailes del Futuro: cada laberinto ritual Tiene un plan musical, y un corazón musical Puede seguir sin error sus caminos impecables. Envidiamos arroyos y casas que son seguras, pero, dudosas, articuladas al error, nunca estuvimos desnudas y tranquilas como una gran puerta. Y nunca seremos impecables como nuestras fuentes: vivimos en libertad por necesidad, un pueblo montañés que habita entre montañas. [Helados por el Presente, su pesadumbre y su ruido, al despertar suspiramos por un Sur antiguo, una cálida y desnuda era de instintivo aplomo, en boca inocente, un sabor a gozo. De noche, en nuestros refugios, soñamos tener un hueco en los bailes del Futuro: cada laberinto ritual viene con un plano musical, y un corazón musical a la perfección puede orientarse por tan perfectos recovecos. Envidiamos los arroyos y las casas que son seguros, pero, presa de la duda, aprendices del error, nunca cual puerta grande estuvimos tranquilamente desnudos, y jamás seremos impecables como nuestras fontanas: vivimos en libertad por necesidad, pueblo montaraz que vive entre montañas.] W. H. AUDEN, Sonnets from China, XVIII
-
-
www.examtopics.com www.examtopics.com
-
NOTA: Cada respuesta correcta vale un punto.
Advertir automáticamente sobre posibles problemas de rendimiento y anomalías de fallos en la aplicación web.
Recopilar automáticamente el estado del código fuente y de las variables cuando se produce una excepción en la aplicación web.
Capturar trazas de rendimiento de la aplicación web sin afectar negativamente a los usuarios de la aplicación.
-
-
-
.ca window.navmet.push({key:'Logo',end:+new Date(),begin:window.navmet.tmp}); Delivering to Mississauga L5H Update location window.navmet.tmp=+new Date(); All Select the department you want to search in All Departments Alexa Skills Amazon Devices Amazon Resale Apps & Games Audible Audiobooks Automotive Baby Beauty Books Clothing, Shoes & Jewellery Women's Clothing, Shoes & Jewellery Men's Clothing, Shoes & Jewellery Girl's Clothing, Shoes & Jewellery Boy's Clothing, Shoes & Jewellery Baby Clothing, Shoes and Jewellery Electronics Gift Cards Grocery Handmade Health & Personal Care Home & Kitchen Industrial & Scientific Kindle Store Luggage & Travel Gear Luxury Beauty Movies & TV MP3 Music Music Musical Instruments, Stage & Studio Office Products Patio, Lawn & Garden Pet Supplies Prime Video Smart Home Software Sports & Outdoors Subscribe & Save Tools & Home Improvement Toys & Games Video Games Search Amazon.ca window.navmet.push({key:'Search',end:+new Date(),begin:window.navmet.tmp}); window.navmet.tmp=+new Date(); EN Hello, sign in Account & Lists Returns & Orders 0 Cart window.navmet.push({key:'Tools',end:+new Date(),begin:window.navmet.tmp}); ._cGluL_welcome_u2Ojz{background:-webkit-linear-gradient(#01a4b5,#008296,#383e3e);background:linear-gradient(#01a4b5,#008296,#383e3e);color:#fff;padding:40px;text-align:center}._cGluL_welcome_u2Ojz h1{font-size:4em}._cGluL_welcome_u2Ojz h2{font-size:2.5em}._cGluL_example_2f4_K{padding:40px}._cGluL_contentWrapper_39oOQ{word-wrap:break-word;min-width:0;text-align:center}._cGluL_contentWrapper_39oOQ h1{font-weight:700}#nav-flyout-amazonprime{width:max(200px,min(30vw,310px))!important} ._cGluL_prime-text-primary_1QpJT{color:#0f1111}._cGluL_prime-text-secondary_1EWgx{color:#565959}._cGluL_prime-text-disabled_3eKRp{color:#6f7373}._cGluL_prime-text-inverse_2ZGSJ,._cGluL_prime-text-prime-white_3lz0a{color:#fff}._cGluL_prime-text-link_1xXsF{color:#007185}._cGluL_prime-text-link-dark_1uTFO{color:#3db7cc}._cGluL_prime-text-error_2QaQs,._cGluL_prime-text-sale_PWqdK{color:#cc0c39}._cGluL_prime-text-success_33wHO{color:#067d62}._cGluL_prime-text-prime-blue_3PjwL{color:#0578ff}._cGluL_prime-text-dark-blue_3VdPR{color:#0064f9}._cGluL_prime-text-prime-squid-ink_1hyUs{color:#0f141a}._cGluL_jp-green_2dnsF{color:#05a934}._cGluL_jp-red_d4SU-{color:#e10600} ._cGluL_prime-text-underline_1_XCk{text-decoration:underline!important} ._cGluL_heading-text-background-image-component-vertical-link_14EEU,._cGluL_heading-text-background-image-component-vertical-link_14EEU:hover{color:#0f1111!important;display:block;text-decoration:none!important} ._cGluL_flyout-footer-wrapper_13vTX{-webkit-box-pack:end;-ms-flex-pack:end;-webkit-box-align:center;-ms-flex-align:center;align-items:center;display:-webkit-box;display:-ms-flexbox;display:flex;justify-content:flex-end} Amazon PrimeEnjoy fast, free shipping on millions of items when you start your 30-day free trial.Try Prime FREE Sign inNew customer? Start here.Your ListsCreate a Wish ListWish from Any WebsiteFind a GiftBaby RegistryWedding RegistryDiscover Your StyleExplore ShowroomYour AccountYour AccountYour OrdersKeep shopping forYour RecommendationsYour Subscribe & Save ItemsYour Prime MembershipMemberships & SubscriptionsYour Amazon Credit CardManage Your Content and DevicesYour Music LibraryYour Prime VideoYour Amazon PhotosYour Apps & DevicesCreate your free business account .gc-flyout-section { width: 335px; height: 335px; margin-top: 0px; margin-bottom: 10px; } .gc-flyout-section-stores { margin-top: 20px; margin-bottom: 5px; font-size: 0; } .gc-flyout-store-box { background-color: #F2F5F7; border: #F2F5F7; width: 165px; height: 165px; display: inline-block; vertical-align: middle; } .gc-flyout-section-top-left { margin-right: 5px; margin-bottom: 4px; position: relative; } .gc-flyout-section-top-right { margin-right: 0px; margin-bottom: 4px; position: relative; } .gc-flyout-section-bot-left { margin-right: 5px; margin-top: 5px; position: relative; } .gc-flyout-section-bot-right { margin-right: 0px; margin-top: 5px; position: relative; } .gc-flyout-single-tile { background-color: #F2F5F7; border: #F2F5F7; margin: 0 18px 0 18px; display: inline-block; vertical-align: middle; align-content: center; } .gc-flyout-store-box:hover { background-color: #EBEEF0; transition: background-color 250ms ease-in-out; } .gc-flyout-store-box:active { background-color: #E3E6E8; transition: background-color 250ms ease-in-out; } .gc-flyout-divider-static-links { margin-top: 20px; margin-bottom: 10px; } Explore gift cards Browse all gift cards Change language English - EN français - FR You are shopping on Amazon.ca Change country/region Change language English - EN français - FR You are shopping on Amazon.ca Change country/region window.navmet.tmp=+new Date(); All var hmenu = document.getElementById("nav-hamburger-menu"); hmenu.setAttribute("href", "javascript: void(0)"); window.navHamburgerMetricLogger = function() { if (window.ue && window.ue.count) { var metricName = "Nav:Hmenu:IconClickActionPending"; window.ue.count(metricName, (ue.count(metricName) || 0) + 1); } window.$Nav && $Nav.declare("navHMenuIconClicked",!0); window.$Nav && $Nav.declare("navHMenuIconClickedNotReadyTimeStamp", Date.now()); }; hmenu.addEventListener("click", window.navHamburgerMetricLogger); window.$Nav && $Nav.declare('hamburgerMenuIconAvailableOnLoad', false); window.navmet.push({key:'HamburgerMenuIcon',end:+new Date(),begin:window.navmet.tmp}); window.navmet.tmp=+new Date(); Best Sellers AmazonBasics Prime Electronics
POUR: Operable<br /> The main navigation bar includes multiple links and dropdown menus. If these elements are not fully operable using only a keyboard, users who cannot use a mouse may struggle to navigate the site. Visible focus indicators and logical tab order are important for accessibility.
-
-
taxodium.ink taxodium.ink
-
Starter 不够用,Pro 和 Ultimate 的 AI 功能对我来说又完全没用。
有些月份用不到 300 次总感觉有点亏,比如 2 月回老家过年,肯定用不满 300 次,这时还订阅 Pro 就觉得好像在浪费。
最近发现年付 Starter 的用量是按一整年来计算,就改成了年付。
如果一年里用超过 3600 次,在剩余月份再升级 Pro。
-
-
www.frontiersin.org www.frontiersin.org
-
Argumentamos por la continua relevancia de una definición de "gran carpa" de MR y, de hecho, abogamos por ampliar aún más la carpa al incluir todas las experiencias mediadas por la tecnología bajo el término realidad mixta. Presentamos una nueva taxonomía para describir las experiencias de MR con las dimensiones de alcance del conocimiento del mundo, inmersión y coherencia . La nueva taxonomía se inspiró en la de Milgram y Kishino, la cual, en nuestra opinión, ha sido subestimada en comparación con las demás contribuciones de su artículo original. Si bien nos inspiramos en el trabajo original de Milgram y Kishino y estamos en deuda con ellos, esperamos que este artículo fomente el debate y la investigación en esta área.
Su abordaje es limitado, supeditado al elemento técnico, se erige bajo los estamentos de una industria consolidada y que controla el mercado.
-
n un juego de realidad virtual, es más importante que sea entretenido que inmersivo. En un protocolo de inducción de estrés en realidad virtual, es más importante que sea estresante y controlable que que genere presencia.
Eso es crucial puesto que la narrativa siempre va a ser primaria a la experiencia sensorial.
-
La esquina superior derecha del gráfico requiere un IM alto y un EWK alto juntos, lo que da como resultado un gemelo digital perfecto , que ofrece seguimiento y representación en tiempo real del mundo real.
Revisar fotogrametría
-
la experiencia de un usuario y abordar la tercera limitación identificada en la sección 2, debemos ir más allá de los elementos de realismo co
La experiencia de usuario se limita a lo "Realista", no a lo narrativo ni a lo emocional
-
Además, Slater ha argumentado que la inmersión comprende dos tipos de acciones válidas: acciones sensoriomotoras válidas y acciones efectivas válidas. Estas acciones válidas resultan en cambios en la percepción del usuario del entorno y cambios en el entorno mismo, respectivamente. Argumentamos que tanto la RF como la EPM forman parte de las acciones sensoriomotoras válidas.
No se tiene en cuenta a lo emocional, revisar articulo de: Sora-Domenjó C (2022) Disrupting the “empathy machine”: The power and perils of virtual reality in addressing social issues. Front. Psychol. 13:814565. doi: 10.3389/fpsyg.2022.814565 y Reis, António. (2023). Immersive media and social change: The ‘empathy machine’ is dead, long live ‘emotional geography’!. Anàlisi. 68. 135-154. 10.5565/rev/analisi.3539.
-
os de nuestras dimensiones propuestas se derivan de las tres originales de Milgram y Kishino. Adoptamos directamente la dimensión de Alcance del Conocimiento del Mundo (EWK) , ya que consideramos que captura un componente clave de las experiencias de realidad aumentada y virtualidad aumentada: el grado en que el sistema es consciente de su entorno real y puede responder a los cambios en dicho entorno.
Que sucede en ambientes de arquitectura sin gravedad, o aquellas posibilidades no experienciales en un mundo fisico. o aquellas experiencia que no son de conocimiento sino que se viven a partir de las prácticas culturales propias de los pueblos del sur global.
-
Milgram y Kishino requería que el contenido mostrado (visualmente) fuera una mezcla de real y virtual, mientras que nuestra redefinición propuesta simplemente requiere que la experiencia sensorial general del usuario, la percepción , sea una mezcla de real y virtual. Nuestra respuesta a la crítica de que nuestra definición de MR es demasiado amplia es doble. Primero, como se ilustró anteriormente en esta sección, las muchas definiciones de MR ya eran una fuente de confusión. Segundo, como discutimos en la siguiente sección, la "realidad mixta" no pretende describir completamente un sistema o una experiencia. E
Revisar folk computing
-
or ejemplo, simular una comida mediante la aplicación de las señales visuales, auditivas, hápticas, olfativas y gustativas más sofisticadas puede resultar convincente para un usuario, pero en algún momento probablemente se dará cuenta de que aún no está saciado y, de hecho, podría tener más hambre que al principio. Este conflicto entre la exterocepción y la interocepción muestra cómo las señales contradictorias en una sola percepción pueden generar una experiencia incongruente. Por esta razón, existe una discontinuidad en nuestro continuo revisado, ya que la verdadera realidad virtual solo existe cuando todos los sentidos (exteroceptivos e interoceptivos) son completamente anulados por el contenido generado por computadora.
La simulación como forma de engaño, también esa conexión entre ambientes físicos y digitales y su tensión como formas para crear nuevas formas de percepción.
-
Por ejemplo, la RM se ha definido como una combinación de RA y RV, como sinónimo de RA, como una versión "más potente" de RA, o como la definieron Milgram y Kishino (Speicher et al., 2019 ). En la cultura popular, la distinción entre realidad aumentada y mixta también se ha difuminado, con algunas empresas como Intel.1describiendo la realidad mixta como espacialmente ubicada e interactiva con el mundo real, mientras que la realidad aumentada específicamente no incluye interacción. Microsoft2Define la realidad aumentada como la superposición de gráficos sobre vídeo, como la RA presentada en teléfonos móviles o tabletas, mientras que la realidad mixta requiere una combinación de lo físico y lo virtual. Un ejemplo es el juego RoboRaid de Microsoft HoloLens.3En este juego, los enemigos parecen existir en las paredes y pueden ser ocultados por objetos reales en la habitación real. Si te mueves a otra habitación, la ubicación de los enemigos se adapta a la nueva configuración física. Una abreviatura común es que los sistemas de realidad aumentada (RA)
Definiciones que vienen desde la propia industria, es decir que el lugar donde los autores se situan es precisamente desde esta visión.
-
Nuestro continuo revisado de realidad-virtualidad. Cabe destacar que el Entorno Virtual Externo (tradicionalmente llamado «Realidad Virtual») sigue formando parte de la Realidad Aumentada (RM).
La revisión tiene los mismos problemas del articulo inicial, pero amplia el espectro de manejo de la tecnología. Es preciso buscar el articulo en donde la película Matrix basa su conexión en la base del craneo.
-
La discontinuidad en nuestro continuo revisado hace explícito que existen diferencias reales y sustanciales entre los entornos virtuales externos y los entornos virtuales "similares a Matrix".
Es posible hacer inmersiones espiritu-tecnológicos? Es posible integrar o simular experiencias o practicas ontológicas propias del pueblos del sur global desde estas tecnologías?
-
Continuo realidad-virtualidad de Milgram y Kishino (adaptado de Milgram et al., 1994 ).
Visiones lineales, que son propiamente características de un continuum, pero que es contradictoria en un contexto donde la red, la malla, el hipertexto son considerados lugares de referencia.
-
la propagación del sonido en espacios de varias salas
Panel de sonología, comportamientos del sonido en ambientes físicos, frente a estos comportamientos en la digitalidad.
-
(En la cultura popular, la Pantalla Definitiva se puede ver en las Holocubiertas de la saga Star Trek ).
Establecer relación entre elementos referenciales de la ciencia ficción en este continuum. También hacer critica a esas visiones de futuros que se direccionan a una visión de norte global
-
La experiencia completa del gusto combina sonido, olor, háptica y una sustancia química que imita el gusto natural y estimula las papilas gustativas. El proyecto Food Simulator (Iwata et al., 2004 ) abordó el componente háptico del gusto, y trabajos recientes informan sobre una pantalla de sabor que sintetiza y ofrece sabores que coinciden con los obtenidos con un sensor de sabor (Miyashita, 2020 ).
Revisión de los sentidos en otras practicas culturales. Asimismo, trabajos que han explorado las percepciones y sensaciones en lo digital.
-
como lo propusieron inicialmente Milgram y Kishino, se muestra en la Figura 1. Anclan un extremo con un entorno puramente real, "que consiste únicamente en objetos reales", y el otro, con un entorno puramente virtual, "que consiste únicamente en objetos virtuales"
Unos ambientes puramente reales que son perspectivas que no exploran visiones pluriversales del mundo. Se establece desde una base científica que las creencias, percepciones, practicas de pueblos afro, indigenas, mestizos, entre otros no son realidades considerables.
-
ninguno cuestiona la noción central del continuo RV ni, en general, proponen definiciones alternativas de la realidad mixta.
Precisamente ese es el problema, esa visión positivista hace que se considere este concepto como verdad absoluta
-
realidad virtual" es inalcanzable, y que cualquier forma de realidad mediada por la tecnología es, de hecho, realidad mixta.
Es preciso definir cuales tecnologías abordan porque desde ese momento han habido avances desde las propias experiencias que han ampliado el nivel de percepción.
-
Minority Report, Iron Man y Ready Player One
La primera película la estuvo asesorada por Alex McDowell del Worlbuilding Institute, permitiendo hacer su diseño de producción desde una perspectiva de construcción de universo narrativo.
-
Revisando el continuo realidad-virtualidad de Milgram y Kishino
Esta revisión se hace desde un lugar meramente técnico, anglosajón, purista y que desconoce de forma tajante otras maneras de percepción de la realidad. Es decir, que actualiza una propuesta que se hizo en 1994 pero surge desde ella misma para ampliar su alcance; parte de una realidad tangible para describir su accionar.
Tags
- ontologías
- sonología
- matrix
- linealidad
- realidad virtual
- norte global
- emociones
- realidades extendidas
- puriverso
- Experiencia de usuario
- emotional geography
- malla
- sensaciones
- empathy machine
- realidad mixta
- Crítica
- espiritu-tecnologico
- realidad aumentada
- oligotecnologia
- UX
- Fotogrametría
- comunidades étnicas
- sonido
- worldbuilding
- ciencia ficción
- industria
- narrativa
- folk computing
- tecnologias inmersivas
- Percepción
- conocimiento situado
- hipertexto
- sentidos
- positivismo
- Diseño de producción para el siglo XXI
- red
- Simulación
Annotators
URL
-
-
-
La Maison Perchée : Analyse d'une Communauté de Rétablissement
Résumé Exécutif
Ce document de synthèse analyse les thèmes, les témoignages et la structure de l'association "La Maison Perchée", tels que présentés dans le contexte source.
Fondée en juin 2020 par un groupe de jeunes adultes, dont trois sur quatre sont directement concernés par des troubles psychiques,
La Maison Perchée se positionne comme "la brique manquante du rétablissement".
Son objectif principal est de créer une communauté d'entraide non médicalisée pour les 18-40 ans vivant avec un trouble psychique, en se basant sur le principe fondamental de la pair-aidance.
L'analyse révèle plusieurs axes majeurs :
1. Le Vécu Intime des Troubles : Des récits personnels poignants sur la bipolarité et la schizophrénie illustrent la réalité des symptômes (manie, dépression, psychose, paranoïa), la violence des crises, l'épreuve de l'hospitalisation et la souffrance psychique intense, souvent qualifiée de "handicap invisible".
2. La Philosophie du Rétablissement : Le concept de rétablissement est central. Il ne s'agit pas d'une guérison, mais d'un processus d'apprentissage pour "vivre et non plus survivre" avec une maladie chronique. Ce chemin est jalonné de difficultés et de petites victoires qui doivent être célébrées.
3. La Puissance de la Pair-Aidance : L'entraide entre pairs est le cœur du projet.
Le partage d'un vécu similaire offre des modèles, une compréhension authentique et un soutien que le système de soin traditionnel ne fournit pas toujours.
L'association forme activement ses membres à cette pratique.
4. La Création d'un Espace Physique : Le projet s'est concrétisé par l'ouverture d'un café-lieu de rencontre à Paris.
Cet espace vise à déstigmatiser la santé mentale et à offrir un refuge où les conversations profondes et l'acceptation sont la norme, créant ainsi des liens sociaux essentiels.
En somme, La Maison Perchée émerge comme une initiative vitale, une "pulsion de vie" qui répond à un besoin criant de soutien communautaire, de reconnaissance et d'espoir pour une population souvent marginalisée par la maladie et la stigmatisation.
--------------------------------------------------------------------------------
1. Origine et Mission de La Maison Perchée
1.1. Fondation et Équipe
L'association "La Maison Perchée" a été officiellement fondée en juin 2020. L'équipe fondatrice est composée de quatre jeunes adultes, tous âgés de moins de 30 ans au moment de la création :
• Maxime : Diagnostiqué bipolaire.
• Victoria : Diagnostiquée d'un trouble bipolaire psychotique.
• Lucile : Atteinte de schizophrénie.
• Caroline Mats : Non concernée directement par un trouble psychique.
Cette composition, où trois des quatre fondateurs sont "concernés", incarne la philosophie de l'association.
L'idée est née d'expériences personnelles directes et indirectes avec la maladie psychique, l'hospitalisation et le sentiment d'injustice face au manque de soutien.
Lucile évoque le déclenchement de sa schizophrénie en 2016 et son expérience en tant qu'enfant d'un parent malade. Maxime mentionne l'injustice ressentie à l'hôpital en voyant d'autres patients sans visites, ce qui a été un moteur initial : "je me suis dit il leur faut un endroit".
1.2. Philosophie et Objectifs
La mission de La Maison Perchée repose sur des concepts forts visant à combler un vide dans le parcours de soin.
• "La brique manquante du rétablissement" : L'association se définit comme le "lieu manquant de la psychiatrie", un espace non médicalisé essentiel qui complète le suivi clinique.
• "Apprendre à vivre et non plus survivre" : L'objectif est de dépasser la simple gestion de la maladie pour permettre aux individus de s'épanouir et de mener une vie riche de sens.
C'est une démarche décrite comme une "p***** de pulsion de vie".
• La Pair-Aidance : Le principe de l'aide par les pairs (P.A.I.R.) est central.
Il s'agit de s'appuyer sur "une personne avec un vécu similaire au sien" pour obtenir des conseils, des modèles de rétablissement et une compréhension authentique de ce qu'ils traversent.
• Une Communauté Jeune : Le ciblage d'une tranche d'âge spécifique (18-40 ans) répond à un besoin d'être avec des personnes de son âge, une problématique rencontrée par certains dans des structures de soin plus traditionnelles (C.T.T.P.).
2. Témoignages : Le Vécu des Troubles Psychiques
Le documentaire est jalonné de témoignages directs et crus sur l'expérience de la maladie mentale.
2.1. Le Trouble Bipolaire
• L'expérience de Maxime : Il décrit une phase de manie survenue à 22 ans en Chine, où il avait créé quatre entreprises.
Cette période était caractérisée par un sentiment de toute-puissance ("devant moi un vieux sage"), des délires mystiques ("prières cheloues avec des couleurs") et des comportements obsessionnels ("je compte tout"). Cette phase a été suivie d'une hospitalisation sous contention, puis de 17 mois de profonde dépression et une tentative de suicide.
• L'expérience de Victoria : Diagnostiquée à 20 ans, elle décrit le trouble comme une "alternation assez extrême de l'humeur". La maladie et les effets secondaires du traitement ont été un "parcours du combattant" qui l'a contrainte à interrompre ses études de design pendant un an.
2.2. La Schizophrénie
• Le vécu de Lucile : Elle insiste sur le fait qu'il n'y a pas "une schizophrénie, mais des schizophrénies". Son quotidien est marqué par des épisodes psychotiques avec une "impression de mort imminente" et la peur que les lieux publics "explosent".
Elle décrit une anxiété constante et une "impression paranoïque que les gens parlent sur nous" dans les environnements bruyants. Elle qualifie sa condition de "handicap invisible", car la souffrance n'est pas apparente.
2.3. Crises, Hospitalisation et Souffrance
Les récits des crises psychotiques sont particulièrement intenses.
• Dissociation et mise en danger : Victoria raconte des "bouffées délirantes" où, se sentant "complètement enfermée", elle a couru nue dans Paris et s'est "tabassée contre le sol", animée par un désir de "désexister" et de "s'atomiser". Une "toute petite partie" d'elle restait consciente, rendant l'expérience "terrible".
• L'expérience de l'hôpital : Les séjours en psychiatrie sont décrits comme des périodes d'"incertitude et de solitude".
Les témoignages évoquent l'isolement (se faire livrer des choses "à travers la grille"), le choc de la contention et la froideur du système.
• La réalité de la souffrance : Un point est martelé : "les maladies psy c'est une p***** de souffrance". La douleur psychique est comparée à la douleur physique, avec une échelle de souffrance de 0 à 10.
Le suicide est présenté comme une pensée qui émerge de cette souffrance extrême, non par désir de mort mais par besoin que "ça s'arrête".
3. Le Rétablissement et la Pair-Aidance
3.1. Définir le Rétablissement
Le concept de rétablissement est présenté de manière nuancée, loin de "vendre du rêve".
• Un Processus, pas une Guérison : Il s'agit d'apprendre à mener une vie épanouissante malgré la maladie. C'est un compromis, car "vivre avec une maladie chronique, c'est un compromis".
• Un Chemin Laborieux : Le processus peut être long et difficile ("Est-ce que c'est facile ? Non. Est-ce qu'on en chie par un moment ? Oui.").
• Célébrer les Petites Victoires : Le rétablissement passe par la reconnaissance des progrès, même les plus simples, comme "reprendre les transports" ou "sortir de chez soi".
• Les Marqueurs du Rétablissement : Pour Maxime, être rétabli signifie avoir surmonté les prédictions négatives de ses psychiatres ("vous pourrez plus jamais entreprendre, vous pourrez pas avoir de vie amoureuse") et simplement "être content de se lever le matin".
3.2. Le Pouvoir de la Pair-Aidance
La pair-aidance est le pilier de La Maison Perchée. Son efficacité réside dans le partage d'expériences vécues.
• Compréhension Mutuelle : Elle permet des "échanges de gens qui peuvent réellement comprendre ce que vous vivez".
• Source d'Espoir : Recevoir une carte postale d'un pair en étant hospitalisé est décrit comme un contact qui rend l'hôpital "un peu moins froid, moins impressionnant".
• Formation Active : L'association identifie des membres "plus ou moins rétablis" et les forme à la pair-aidance pour qu'ils puissent à leur tour "donner en retour".
4. La Concrétisation du Projet : Du Virtuel au Physique
4.1. Les Débuts et la Stratégie
La création de l'association a suivi une approche pragmatique et résiliente. Face à l'hésitation initiale, la stratégie de Maxime était directe : "Bon les gars, vous allez chercher trois potes d'HP et on va faire un zoom". Ils ont persisté en créant des événements en ligne, même lorsque personne ne s'inscrivait au début.
4.2. Le Lieu : Un Café Communautaire
Le projet a franchi une étape décisive avec l'ouverture d'un lieu physique.
• Adresse : Avenue de la République, à Paris.
• Concept : Un café ouvert à tous, conçu pour parler de santé mentale et déstigmatiser les troubles psychiques.
• Ambiance : Le lieu favorise des interactions immédiates et profondes. Un témoignage décrit la magie du lieu : "tout le monde se parle de trucs méga deep et méga perso, genre au bout de 1 seconde 30". Il attire un public mixte : membres, proches, et simples passants.
• Financement : L'aménagement du lieu a été en partie financé par l'obtention d'un "appel à projet" de 20 000 €, destinés à l'achat de matériel, dont une machine à café professionnelle estimée à 10 000 €.
4.3. Activités et Communauté
Le lieu est un hub pour diverses activités qui renforcent les liens et le soutien.
• Écriture de cartes : Des ateliers sont organisés pour écrire des mots de soutien à des personnes actuellement hospitalisées.
• Discussions thématiques : Des événements comme les "Tête Perchée" permettent de discuter publiquement de sujets comme la vie professionnelle avec un trouble psychique.
• Création de liens : Le but premier pour beaucoup de membres est simple : "j'ai surtout envie d'avoir des amis". Le lieu offre cet espace de socialisation sécurisant.
5. Thèmes Transversaux et Enjeux
5.1. La Stigmatisation
La lutte contre la stigmatisation est un combat permanent.
• Dans le monde professionnel : La réponse à la question "Comment avez-vous géré le regard d'autrui ?" est "En disant rien pendant longtemps".
• Préjugés du public : Une anecdote relate une rencontre avec une personne exprimant sa peur des "fous" et des "schizophrènes", à qui les fondatrices ont dû répondre : "on vous fait peur ?".
• Action de déstigmatisation : La Maison Perchée combat ces stéréotypes par la prise de parole publique et l'ouverture de son lieu à tous.
5.2. Le Rôle des Proches
La famille et les amis jouent un rôle ambivalent mais souvent crucial.
• Souffrance collatérale : Lucile et Caroline témoignent de la souffrance vécue "en tant que proche", une expérience qui a motivé leur engagement.
• Soutien fondamental : Maxime souligne l'importance du soutien de sa mère, sa tante et sa sœur à sa sortie d'hôpital, le qualifiant de "chance dans sa malchance".
5.3. Identité et Créativité
La maladie est également présentée comme une facette de l'identité, parfois source de force et de créativité.
• Une vision positive : Un membre compare les personnes atteintes de troubles à des "X-Men", possédant "un truc en plus que les autres n'ont pas".
• L'art comme exutoire : Plusieurs membres canalisent leurs expériences à travers l'art : une membre travaille sur un roman graphique de sa vie de 15 ans à aujourd'hui, Lucile lit des poèmes, un autre écrit, chante et filme.
• La citation inspirante : Une phrase partagée lors d'un atelier d'écriture résume cette dualité : "Si tu es tombé si bas, dis-toi que tu peux monter aussi haut".
-
-
Local file Local file
-
Finally, while TIR-Flow signifi-564cantly elevates reasoning ceilings, the iterative na-565ture of System-2 active perception entails a mod-566est trade-off in inference speed compared to static,567single-pass baselines.
speed is mentioned as a limitation, but the paper doesn't include any measures for this.
-
-
www.france.tv www.france.tv
-
Synthèse sur la Santé Mentale : Témoignages et Enjeux
Résumé Exécutif
Ce document de synthèse analyse un ensemble de témoignages et d'expertises sur la santé mentale, en se concentrant sur les expériences vécues par des athlètes de haut niveau et des jeunes.
Il met en lumière le caractère universel des troubles psychiques, qui touchent toutes les strates de la société, indépendamment du succès ou du statut social. Les principaux points à retenir sont les suivants :
1. Universalité du Mal-être : La santé mentale est une composante de la vie de chacun, susceptible de se dégrader. Les témoignages révèlent que même les champions du monde, au sommet de leur carrière, peuvent souffrir de dépression sévère, illustrant la dichotomie fréquente entre la réussite professionnelle et la détresse personnelle.
2. Stigmatisation et Silence : La honte, la peur d'être perçu comme "faible" et la pression sociale, particulièrement dans des environnements compétitifs comme le sport de haut niveau, constituent des obstacles majeurs à l'expression de la souffrance. Beaucoup souffrent en silence, cachant leur mal-être derrière une façade de normalité.
3. Facteurs de Risque Spécifiques :
◦ Pour les Athlètes : La pression de la performance, la gestion difficile de la "redescente" après un grand succès, l'isolement lié aux blessures mal gérées, et la culture du vestiaire où "il n'y a pas de place pour les faibles". ◦ Pour les Jeunes : La pression scolaire, les traumatismes infantiles, l'environnement familial, le harcèlement (scolaire et en ligne), et l'influence des algorithmes des réseaux sociaux qui peuvent créer des "bulles numériques" toxiques.
4. Symptômes et Conséquences : La détresse psychologique se manifeste par un large éventail de symptômes : isolement social, anxiété, crises d'angoisse, troubles du comportement alimentaire (TCA), scarifications, idées suicidaires, et symptômes physiques chroniques. La solitude est décrite comme une force destructrice capable de "tuer une personne".
5. Voies de Guérison : Le déclencheur de la guérison est souvent une rencontre ou la décision de parler. Les solutions passent par la recherche d'une aide professionnelle adaptée (psychologues, coachs mentaux), le soutien de l'entourage, l'expression des émotions (par l'écriture, le sport, l'art) et l'acceptation que la vulnérabilité n'est pas une faiblesse.
6. Initiatives et Prise de Conscience : Des initiatives comme le fonds de dotation "Génération 2018", créé par des footballeurs champions du monde, témoignent d'une volonté croissante de "mettre le sujet sur la table", de déstigmatiser la santé mentale et d'avoir un impact positif sur la jeunesse.
1. Définitions et Perceptions de la Santé Mentale
La santé mentale est décrite comme une composante fondamentale et universelle de l'être humain, souvent comparée à la santé physique. Elle n'est pas un état statique mais un équilibre fluctuant.
• Définitions Personnelles :
◦ "La santé mentale au sens large, on en a tous une, plus ou moins bonne, plus ou moins efficiente." ◦ C'est "comment tu te sens à l'intérieur de soi", le "bien-être du cerveau", et se ressent "dans la tête, dans le cœur, dans le ventre". ◦ Elle est perçue comme un équilibre essentiel : "si on n'a pas ça, ben on a plus envie de rien faire." ◦ Elle inclut la connaissance de sa "propre écologie", l'harmonie entre le mental et le physique.
• Déterminants Multiples : La santé mentale est influencée par une multitude de facteurs : le logement, le travail, l'ambiance sociale, les amis, le sport, et même la météo ou le contexte international. L'état psychologique (bonheur, conflits, questionnements) en est un déterminant central.
• Dégradation Générale : Des études médicales sérieuses indiquent une "aggravation constante sur les 10 dernières années" des troubles anxieux et dépressifs, une tendance accentuée par la crise du Covid-19. La santé mentale des jeunes semble s'être "considérablement dégradée depuis 2019".
2. Le Sport de Haut Niveau : Une Arène de Pression Psychologique
Les athlètes, bien que perçus comme des modèles de force, font face à des défis psychologiques uniques et intenses, souvent cachés derrière l'image du succès.
La Dichotomie entre la Réussite et la Souffrance
Le succès professionnel peut coexister avec une profonde détresse personnelle, créant un décalage difficile à gérer.
• Témoignage d'un Champion du Monde : "Quand tu es champion du monde, ben on voit la coupe et elle est en or quoi. Et on voit que ça et mais derrière il y a beaucoup de choses qui se passent dans la tête des joueurs."
• Expérience Personnelle : Un joueur décrit son arrivée en Espagne à 18 ans : "à la fois, je vivais le rêve au niveau professionnel et à la fois au niveau personnel, j'étais pas bien, j'étais en dépression."
La "Redescente" Post-Victoire et les Blessures
L'après-succès et la gestion des blessures sont des périodes de grande vulnérabilité.
• L'Après-Graal : "Quand tu es arrivé tout en haut, tu as atteint le graal, une redescente après qui n'est pas facile à gérer." Un joueur mentionne qu'Adil Rami a vécu un "espèce de burnout après le mondial".
• L'Impact des Blessures : Une blessure peut signifier la perte de son identité. Un joueur raconte : "j'avais du mal à même le matin me réveiller, c'était dur. Je me demandais ce que j'allais faire parce que pour moi, il y avait que le foot dans la vie."
• Mauvaise Gestion par les Clubs : Un joueur témoigne avoir été mal accompagné, poussé à jouer malgré une blessure pour les besoins de l'équipe, ce qui a aggravé son état physique. Les fuites d'informations confidentielles dans la presse l'ont fait passer pour "un poids pour ce club-là", entraînant un harcèlement de la part des supporters et un profond isolement.
La Culture du Silence et la Peur de la Faiblesse
Le milieu sportif impose une culture où la vulnérabilité est souvent assimilée à une faiblesse inacceptable.
• La Loi du Vestiaire : "Quand tu arrives dans le vestiaire, il y a pas de place pour les faibles. [...] Il faut être meilleur que l'autre pour avoir sa place le weekend."
• Stigmatisation par l'Encadrement : Une joueuse de handball en national raconte avoir été renvoyée par sa coach avec cette justification : "je préfère quelqu'un de moins fort que toi mais de stable mentalement plutôt que toi qui est instable mentalement." Elle ajoute : "je me suis vraiment rendu compte que la santé mentale dans le sport c'était tabou."
• Auto-Censure : Les athlètes eux-mêmes intériorisent cette norme. L'un d'eux déclare : "Le fond du problème, c'est que eux-mêmes se l'interdisent."
3. La Vulnérabilité de la Jeunesse
Les jeunes sont confrontés à une série de pressions et de traumatismes qui peuvent déclencher des crises de santé mentale sévères.
Pression Scolaire et Épuisement
L'environnement scolaire et la pression à la réussite sont des facteurs de stress majeurs.
• Origine des Troubles : Une jeune femme lie sa dépression aux cours : "très vite on m'a mis la pression et je me suis mise la pression. Et donc ça, ça s'est enchaîné à la phobie scolaire et puis après [...] mes TCA."
• Statistiques : Il est noté que "7-8 jeunes sur 10 ont une pression à la réussite plus ou moins forte et plus ils se la mettent forte et plus effectivement ils ont un risque anxio-dépressif qui est élevé."
• Surmenage (Burnout) : Le surmenage n'est pas exclusif au monde du travail. Une jeune femme engagée dans une association décrit un épuisement physique et émotionnel : "Le rythme il baissait pas, il baissait jamais. [...] je pense qu'à un moment mon cerveau il s'est dit stop."
Traumatismes et Environnement Familial
Les expériences vécues dans l'enfance et la dynamique familiale jouent un rôle crucial.
• Traumatisme Précoce : Un jeune homme raconte la mort de son petit frère de deux mois alors qu'il n'avait que 14 ans, et le sentiment de culpabilité qui l'a envahi : "je me suis senti coupable. Je me suis dit c'est de ma faute." Il a gardé ce fardeau pour lui pendant des mois.
• Environnement Familial :
◦ Certaines familles sont "assez réservées" et ont du mal à exprimer leurs problèmes. ◦ Une jeune femme décrit une famille "dans le paraître", où il faut "paraître forte, faut pas montrer ses faiblesses", ce qui l'a empêchée de parler de sa scarification. Sa mère a qualifié ses blessures de "foutaises", provoquant un sentiment de non-compréhension et de repli sur soi. ◦ Un autre jeune, ayant grandi dans un quartier populaire, a endossé très tôt le rôle de "sauveur" familial, portant une pression immense.
Harcèlement et L'Influence des Réseaux Sociaux
Le harcèlement, qu'il soit physique, psychologique ou en ligne, a des conséquences dévastatrices.
• Définition du Harcèlement : "Si quelqu'un dit qu'il est harcelé, il est harcelé. C'est ça la définition du harcèlement."
• Violence Psychologique : Une jeune femme décrit un harcèlement moral au collège : "de m'humilier, de me dire des mots méchants. Il y a aussi du chantage", ce qui a "ruiné" son estime de soi.
• Algorithmes Toxiques : Les réseaux sociaux peuvent enfermer les utilisateurs dans une "bulle numérique".
◦ Un jeune homme raconte comment, en état de dépression, les algorithmes lui proposaient des contenus "qui glamorisaient, qui incitaient au comportement autodommageable". ◦ Une autre témoigne : "j'avais peur d'avoir une maladie grave, bah moi mon algorithme vraiment il y avait que des personnes qui allaient mourir." ◦ Ces contenus, bien que procurant un sentiment paradoxal de "réconfort" et de "compréhension", aggravent le mal-être.
4. Les Chemins vers le Rétablissement
Malgré la profondeur de la souffrance, les témoignages montrent que la guérison est possible, bien que le chemin soit souvent long et difficile.
L'Importance Cruciale de la Parole
Briser le silence est la première étape, et la plus fondamentale, vers la guérison.
• Le Déclic : Souvent, "le déclic, il vient souvent d'une rencontre".
• Parler pour Aller Mieux : "On va mieux grâce à l'échange, à pouvoir parler de sa situation." L'important est de trouver une "oreille attentive", que ce soit un ami, un professionnel, ou un guide spirituel.
• Lignes d'Écoute : L'appel à une ligne d'écoute, même tenue par des étudiants, a été une porte d'entrée pour "déballer son histoire".
La Recherche d'un Soutien Professionnel
Trouver le bon professionnel est un processus parfois ardu mais essentiel.
• La Difficulté de Trouver le Bon Psy : Un jeune homme exprime son blocage : "je suis tombé sur des psy qui qui était pas bien après avoir entendu mon histoire. C'est à dire voir un psy pleurer."
• Persévérance : La recommandation est de ne pas abandonner : "c'est pas parce que tu en as tu es tombé sur trois pas bien que le 4e ou la 4e ou la 5e ou le 5e va pas être bien."
• Terminologie : Dans le sport, il est plus courant de parler de "prépa mental" ou "coach mental" que de "psychologue", car le terme "psy" "fait un peu peur". L'important reste la démarche.
• Médicaments : Les traitements comme les antidépresseurs sont mentionnés comme une aide potentielle, souvent mal vue mais qui, "bien utilisé, ça peut vraiment aider."
Stratégies Personnelles de Guérison
Chaque individu développe ses propres outils pour surmonter les épreuves.
• Faire ce qu'on a pas envie de faire : "Quand tu fais une grosse dépression, tout ce que tu tout ce que tu as pas envie de faire, c'est ce qu'il faut que tu fasses." Cela inclut bien manger, faire du sport, et parler à un psychologue.
• L'Écriture : "J'ai acheté un cahier. J'écrivais ma colère, ma tristesse. Je cherchais à comprendre." Noter les déclencheurs des crises d'angoisse permet de "déconstruire" le processus.
• Activités Passion : L'équitation est citée comme un refuge permettant de "stopper mon cerveau". Retrouver goût à la vie passe par "faire les choses que j'aime", comme le dessin, sortir, manger.
• L'Humour et le Soutien des Pairs : Un joueur raconte comment ses coéquipiers ont désamorcé un titre de presse ("le mal-aimé") en chantant la chanson, transformant une situation douloureuse en un moment de camaraderie.
Le Rôle des Initiatives Collectives
La prise de conscience collective mène à la création de structures de soutien.
• Génération 2018 : Ce fonds de dotation, créé par les joueurs de l'équipe de France de football 2018, a choisi la santé mentale comme thématique pour "laisser une trace, un héritage" et "mettre le sujet sur la table". Leur but est d'aider les autres, mais aussi de "s'aider nous-mêmes".
5. Ressources et Lignes d'Aide
Plusieurs organisations sont mentionnées comme des ressources pour obtenir de l'aide et de l'information :
• E-Enfance 3018 : Lutte contre le harcèlement et les violences numériques. Numéro d'appel : 3018.
• Nightline : Association visant à améliorer la santé mentale des jeunes et étudiants. Site web : nightline.fr.
• Psycom : Propose de l'information fiable et accessible sur la santé mentale. Site web : psycom.org.
-
-
-
La Crise de la Santé Mentale Étudiante : Pressions, Impacts et Initiatives
Synthèse
Ce document de synthèse analyse la crise profonde et multifactorielle de la santé mentale qui affecte la population étudiante. La détresse psychologique, déjà présente avant la pandémie de COVID-19 (touchant 20-25% des étudiants), a été exacerbée par la crise sanitaire, atteignant un pic de 43% en 2021. Cette situation critique est alimentée par une convergence de facteurs de stress : une pression académique intense, la précarité financière, l'isolement social, et une anxiété profonde face à un avenir jugé incertain, marqué par le changement climatique et l'instabilité économique.
Le système de soins en santé mentale s'avère largement insuffisant pour répondre à cette demande croissante, caractérisé par un manque criant de professionnels (un psychologue pour 15 000 étudiants en milieu universitaire) et des listes d'attente prohibitively longues. Dans ce contexte, des initiatives d'aide par les pairs, telles que l'association Nightline, émergent comme une ressource essentielle, offrant un soutien accessible, anonyme et gratuit par des étudiants formés à l'écoute.
Malgré l'ampleur de leur souffrance, les étudiants ne se positionnent pas comme une génération décliniste. Au contraire, ils manifestent une forte volonté d'agir et de s'engager, cherchant à construire un avenir où le bien-être prime sur la réussite matérielle, et affirmant leur capacité à être des acteurs du changement dès maintenant.
--------------------------------------------------------------------------------
1. Un Constat Alarmant : L'ampleur de la détresse psychologique
Les enquêtes épidémiologiques successives révèlent une dégradation continue de la santé mentale des étudiants.
• Avant la crise sanitaire : Les études montraient déjà des taux de détresse psychologique élevés, oscillant entre 20% et 25%. Ce mal-être étudiant, documenté depuis des années, n'a pas été suffisamment pris en compte par les pouvoirs publics, malgré des rapports comme celui de Danièle Viard en 2006.
• L'effet "loupe" du COVID-19 : La pandémie a agi comme un catalyseur et un amplificateur d'une souffrance préexistante. Le taux de détresse psychologique a grimpé à 43% en septembre 2021. Le nombre d'étudiants présentant des symptômes dépressifs a doublé entre l'avant et l'après-COVID, un constat partagé dans plusieurs pays européens.
• Nature de la détresse : Il ne s'agit pas d'une maladie psychiatrique en soi, mais d'un syndrome qui affecte plusieurs sphères de la vie :
◦ Affective et émotionnelle : idées tristes, morosité, irritabilité. ◦ Somatique : troubles du sommeil, fatigue intense, incapacité à accomplir des tâches simples. ◦ Cognitive : baisse de la concentration, de la mémorisation et de la motivation, ce qui impacte directement la capacité à étudier.
2. Les Multiples Facteurs de Stress des Étudiants
La détresse étudiante est le résultat d'une accumulation de facteurs de stress spécifiques à leur condition et à l'époque actuelle.
Pression Académique et Élitiste
Un environnement universitaire extrêmement compétitif est une source majeure de stress.
• Discours élitiste : Certains professeurs instaurent une pression intense dès le début du cursus avec des phrases telles que : "Vous avez choisi un cursus très difficile, il va falloir s'accrocher. Votre voisin de table ne sera pas là dans un an."
• Surcharge de travail : Des étudiants témoignent d'un épuisement total, sacrifiant leur vie sociale et leurs loisirs. Une étudiante déclare : "J'ai arrêté le sport, j'ai arrêté de voir des gens. Je me concentre vraiment sur mes études [...] Les études, c'est toute ma vie."
• Conséquences directes : Cette pression mène à des burnouts, des dépressions et une angoisse constante de l'échec. Un professeur de japonais admet : "On le sait tous les ans, il y a plusieurs suicides. Ils s'en vantent pas mais on le sait quand même."
Précarité et Conditions de Vie
Les difficultés matérielles sont un fardeau quotidien.
• Précarité financière : La gestion du loyer, des courses, des transports et le manque de soutien familial pour certains créent une anxiété permanente.
• Pression familiale : Pour certains, la réussite est une obligation due à l'investissement des parents. Une étudiante aînée de trois sœurs confie : "Je me dis que je n'ai pas le droit à l'échec."
L'Impact Dévastateur de la Crise Sanitaire
La pandémie a eu des effets particulièrement destructeurs sur les jeunes.
• Isolation et perte de socialité : La privation de contacts sociaux a été qualifiée de "cataclysmique" pour une tranche d'âge où la vie relationnelle est fondamentale.
• Traumatisme psychique : Les confinements ont été vécus comme un traumatisme, générant des états de stress post-traumatique, un phénomène inédit à cette échelle. Les peurs (mourir, que les proches meurent, un prochain confinement) ont été des facteurs traumatisants majeurs.
• Stigmatisation : Les étudiants ont été désignés comme un des vecteurs de la circulation virale, ce qui a eu un "effet terrible sur leur santé mentale".
• Conséquences à long terme : Les experts estiment que les conséquences de cette période se feront sentir pendant des décennies, tant sur la santé mentale que sur les parcours de formation et professionnels.
Anxiété face à un Avenir Incertain
Le climat général de la société est perçu comme particulièrement anxiogène.
• Crises multiples : Le changement climatique, l'inflation, la guerre, le chômage et les violences sexistes et sexuelles créent un contexte d'incertitude généralisée.
• Écoanxiété : Il ne s'agit pas d'un trouble, mais d'une peur légitime et saine face à un danger concret et actuel. Un expert précise : "Quand on n'a pas peur face à un danger, c'est pas qu'on est courageux, c'est qu'on est inconscient."
• Perte de prévisibilité : Les structures sociales et les projections (formation menant à un métier, planète viable pour fonder une famille) sont disloquées, ce qui est particulièrement déstabilisant pour des psychismes en construction.
Vulnérabilités Spécifiques
| Catégorie | Description de la Vulnérabilité | | --- | --- | | L'Âge (15-25 ans) | Le cerveau continue sa maturation jusqu'à 25 ans, le rendant particulièrement sensible aux influences environnementales ("plasticité pour le meilleur et pour le pire"). 75% des pathologies psychiatriques (schizophrénie, trouble bipolaire) se déclarent durant cette période, soulignant l'importance de la prévention et de la détection précoce. | | Étudiants Étrangers | Ils subissent une double pression : la pression académique commune, et une pression supplémentaire pour être "excellents" afin d'assurer le renouvellement de leur titre de séjour. À cela s'ajoutent les défis de l'intégration (logement, solitude, choc culturel). |
3. Nightline : Une Réponse par les Pairs
Face à la saturation des services professionnels, l'association étudiante Nightline offre un modèle de soutien alternatif et vital.
Mission et Dispositifs
• Objectif : Améliorer la santé mentale des jeunes, en particulier des étudiants.
• Principe fondamental : L'aide par les pairs. Le service est composé et destiné aux étudiants.
• Actions : L'association informe, déstigmatise, soutient et oriente à travers :
◦ Une ligne d'écoute nocturne (tous les soirs de 21h à 2h30).
◦ Des formations dans les établissements d'enseignement supérieur.
◦ Des actions de terrain et des campagnes sur les réseaux sociaux.
◦ Des actions de plaidoyer pour améliorer les politiques de santé mentale.
Le Rôle des Bénévoles Étudiants
• Profil : Ce ne sont pas des professionnels, mais des étudiants formés à l'écoute active et bienveillante.
• Anonymat : C'est une règle cardinale, valable pour l'appelant comme pour le bénévole. Les bénévoles ne doivent pas révéler leur engagement à leur entourage pour protéger le service et ne pas décourager des proches d'appeler. La seule rupture de l'anonymat est envisagée en cas de risque suicidaire immédiat pour appeler les secours, et l'appelant en est prévenu.
• Soutien des bénévoles : Un cadre strict est mis en place pour les protéger :
◦ Débriefings à chaud entre bénévoles juste après les appels difficiles. ◦ Réunions de partage mensuelles et obligatoires, encadrées par une psychologue, pour extérioriser et analyser les appels marquants.
Un Mouvement International
Le concept de Nightline est né dans les années 1970 au Royaume-Uni. Il s'est depuis étendu en Irlande, en Allemagne, en Suisse, en Autriche et en France. Des projets sont en cours pour une implantation en Belgique, montrant que le problème de la santé mentale étudiante est une préoccupation européenne.
4. Les Défaillances du Système de Soins en Santé Mentale
L'accès aux soins professionnels est un parcours du combattant pour les étudiants en souffrance.
• Pénurie de professionnels : Le ratio dans les services de santé universitaires est de un psychologue pour 15 000 étudiants, soit dix fois moins que la recommandation internationale.
• Listes d'attente "lunaires" : Les délais pour obtenir un rendez-vous dans le secteur public (Centres Médico-Psychologiques) sont de 6 à 18 mois, ce qui est incompatible avec l'urgence d'une détresse psychique. Le système est décrit comme "bouché de chez boucher".
• Coût des soins : Le secteur privé est souvent inaccessible financièrement pour une population précaire, ce qui conduit de nombreux étudiants à renoncer aux soins.
5. Au-delà du Désarroi : L'Engagement et la Résilience Étudiante
Contrairement à une vision parfois décliniste, les témoignages révèlent une jeunesse engagée et tournée vers l'action.
• Rejet du catastrophisme : Les experts notent que les étudiants, malgré leurs difficultés, ne sont pas dans une vision catastrophique du monde. Ils conservent le sentiment d'avoir les moyens d'intervenir sur leur avenir grâce à leur formation et leur vitalité.
• Volonté d'agir maintenant : Le discours dominant est celui de l'action immédiate. Une étudiante déclare : "On est acteur et actrice déjà maintenant [...] on leur dise pas vous ferez plus tard, mais ils peuvent faire maintenant et ils ont envie de faire maintenant."
• Redéfinition de la réussite : Il y a une prise de conscience que le bien-être et un bon cadre de vie sont plus importants qu'un "cadre de vie luxueux" ou un gros salaire. La quête de sens prime sur la seule réussite professionnelle.
-
-
-
L'Anxiété chez les Jeunes : Synthèse et Analyse
Résumé Exécutif
Ce document de synthèse analyse le phénomène croissant de l'anxiété chez les jeunes, un enjeu de plus en plus qualifié d'« épidémie » ou de « maladie de l'heure ».
Les témoignages de jeunes et d'experts révèlent un problème multifactoriel profondément ancré dans les structures sociales, éducatives et familiales contemporaines.
Les principaux moteurs de cette anxiété incluent une pression de performance omniprésente, des horaires surchargés qui éliminent le temps de repos et de jeu, et l'influence des réseaux sociaux qui promeuvent une culture de la perfection irréaliste.
L'analyse scientifique distingue le stress (une réponse à une menace immédiate) de l'anxiété (une menace anticipée), tout en soulignant que le cerveau y réagit de manière identique.
Des facteurs déclencheurs universels — un faible sens de contrôle, l'imprévisibilité, la nouveauté et une menace à l'ego (acronyme SPIN) — sont identifiés comme étant au cœur des expériences anxieuses des jeunes. Parallèlement, l'omniprésence du numérique contribue à une régression de l'intelligence émotionnelle en réduisant les interactions en face à face.
Les experts appellent à un « grand réveil », arguant que l'anxiété juvénile n'est pas un simple problème individuel à traiter, mais un symptôme d'un système qui « use le cerveau » des enfants.
Les solutions résident non seulement dans des stratégies d'adaptation individuelles comme la respiration ou la méditation, mais aussi dans des changements systémiques, notamment une remise en question du modèle éducatif axé sur la performance et la création de conditions plus propices à une croissance saine.
--------------------------------------------------------------------------------
1. Le Phénomène de l'Anxiété : Une "Épidémie" Moderne
L'anxiété chez les jeunes est décrite comme un phénomène majeur et en pleine expansion.
Elle est qualifiée de « maladie de l'heure », succédant à d'autres diagnostics prévalents comme le TDAH il y a quelques années.
Prévalence et Perception
• Consultations en hausse : Un professionnel de la santé mentale note une augmentation significative du phénomène depuis environ six à sept ans, déclarant que l'anxiété représente aujourd'hui la majorité de ses consultations.
• Impression générale : La narratrice, travaillant dans les médias, a constaté à travers diverses statistiques que le problème est bien plus répandu qu'elle ne l'imaginait initialement.
Les jeunes eux-mêmes décrivent l'anxiété comme une expérience quasi universelle dans leur entourage.
Manifestations et Symptômes
Les témoignages des jeunes décrivent un large éventail de manifestations physiques, cognitives et émotionnelles.
| Type de Symptôme | Description | Exemples de Témoignages | | --- | --- | --- | | Physique | Réactions corporelles involontaires déclenchées par la panique ou le stress. | Maux de ventre (Julian, Sopiane), pleurs incontrôlables, hyperventilation, respiration rapide (Magalie). | | Cognitif | Pensées envahissantes et scénarios catastrophiques. | Le "cerveau qui arrête jamais" (Alexandra), se concentrer uniquement sur la panique au point d'en oublier la cause initiale (Sopiane). | | Émotionnel | Sentiment de perte de contrôle, de solitude et de peur. | Peur du jugement et du rejet (Alexandra), impression que l'anxiété "finit plus", difficulté à gérer ses émotions et à montrer sa vulnérabilité (Magalie). | | Comportemental | Réactions externes observables, souvent interprétées différemment par l'entourage. | Colère et agressivité (Julian), comportement très oppositionnel lors des crises (Sopiane), perfectionnisme extrême (arracher une feuille de dessin pour une erreur). |
--------------------------------------------------------------------------------
2. Les Causes Multifactorielles de l'Anxiété Juvénile
L'anxiété des jeunes ne peut être attribuée à une cause unique. Elle émerge de l'intersection de pressions sociales, de l'environnement numérique et de dynamiques scolaires et familiales.
La Pression de la Performance
Un thème central est la quête incessante de la performance dans toutes les sphères de la vie.
• Horaires surchargés : Les jeunes comme Magalie et Marie décrivent des emplois du temps extrêmement chargés, remplis d'études, de tutorat, d'activités parascolaires et de travail, ne leur laissant jamais le temps de se reposer. Marie répond "jamais" à la question "Tu te reposes quand ?".
• Compétition interne et externe : Magalie décrit son besoin de tout faire comme "une compétition contre [elle]-même".
Alexandra exprime un désir de plaire à tout le monde et d'être perçue comme "parfaite".
Cette pression est si normalisée que pleurer à cause des devoirs devient un sujet de conversation banal entre amis.
• Pression sociétale et parentale : Les jeunes ressentent que la performance à l'école, dans les sports et sur les réseaux sociaux est ce qui est valorisé par la société.
Les parents, même avec de bonnes intentions, peuvent amplifier cette pression, comme le reconnaît la narratrice à propos des cours de violon de sa fille Julian.
L'Influence du Monde Numérique et des Réseaux Sociaux
L'environnement numérique joue un rôle significatif dans l'amplification de l'anxiété.
• Culture de la perfection : Les plateformes comme Instagram présentent une vision idéalisée de la vie ("la photo parfaite avec le l'auto parfaite avec la maison parfaite").
Cette exposition constante à la perfection crée un décalage avec la réalité et renforce le sentiment de ne jamais être à la hauteur.
• Régression de l'intelligence émotionnelle : Un spécialiste du non-verbal explique que la diminution des interactions en face à face au profit des communications numériques appauvrit la capacité à décoder les émotions.
Une étude comparant deux camps de vacances (avec et sans téléphones) a montré qu'après une semaine, les jeunes privés de leur appareil étaient devenus plus aptes à juger les émotions des autres.
Le Rôle du Système Scolaire
Le système éducatif est identifié comme un facteur de stress majeur.
• Surcharge d'examens : La chercheuse Sonia Lupien note une augmentation constante du nombre d'examens, qu'elle qualifie scientifiquement de "périodes de stress". L'accumulation de ces déclencheurs de stress épuise les élèves.
• Contagion du stress : Des études montrent qu'un professeur souffrant d'épuisement professionnel peut voir les enfants de sa classe produire davantage d'hormones de stress, car ils "raisonnent avec le professeur".
• Le modèle finlandais comme contre-exemple : La professeure Tina Montreuil décrit le système finlandais, qui minimise les facteurs de stress :
◦ Journées d'école plus courtes (fin vers 14h30-15h).
◦ Pauses déjeuner très longues.
◦ Absence quasi totale de devoirs et d'examens formels.
◦ L'évaluation se fait par des projets, des discussions et des mises en contexte.
◦ Bien que non utopique, ce système génère des niveaux d'anxiété de performance bien plus faibles qu'en Amérique du Nord.
--------------------------------------------------------------------------------
3. Perspectives d'Experts et Compréhension Scientifique
Les experts consultés appellent à un changement de perspective, passant d'une vision individualisée de l'anxiété à une compréhension systémique.
Un Problème Systémique, Pas Individuel
Le professeur Carl la Charité soutient que l'anxiété des enfants est un message adressé à la société.
• Du corps au cerveau : Il établit un parallèle historique : "Il y a [...] un siècle, la façon dont on traitait les enfants, ça nous amenait à user leur corps. [...] Aujourd'hui, en fait, ce qu'on use, c'est leur cerveau."
• Un appel au "grand réveil" : Il considère l'anxiété comme un signal que les méthodes éducatives actuelles ne fonctionnent plus. La solution n'est pas de "réparer" l'enfant, mais de changer les conditions dans lesquelles il grandit.
Démystifier le Stress et l'Anxiété (Approche de Sonia Lupien)
La chercheuse Sonia Lupien propose un cadre scientifique pour comprendre et gérer le stress.
• Stress vs. Anxiété : L'analogie du "mammouth" illustre la différence :
◦ Stress : La réponse biologique à une menace réelle et présente ("le mammouth est devant toi").
◦ Anxiété : La réponse à une menace anticipée ou imaginaire ("le mammouth est dans ta tête").
◦ Pour le cerveau, la production d'hormones de stress est identique dans les deux cas.
• Les quatre déclencheurs du stress (SPIN) : Le cerveau détecte une menace et déclenche une réponse de stress lorsqu'il est confronté à une ou plusieurs de ces situations.
| Acronyme | Caractéristique | Description | | --- | --- | --- | | S | Sens de contrôle faible | Impression de ne pas avoir le contrôle sur la situation. | | P | Personnalité menacée | L'ego ou les compétences sont remis en question. | | I | Imprévisibilité | La situation est nouvelle ou inattendue. | | N | Nouveauté | La situation n'a jamais été rencontrée auparavant. |
• Le danger de la narration "stress toxique" : Sonia Lupien soutient que le discours médiatique et scientifique présentant tout stress comme négatif est contre-productif.
Il peut engendrer une "sensibilité à l'anxiété" ou la "peur d'avoir peur", où une personne panique face aux sensations physiques normales du stress, croyant qu'elles sont dangereuses.
Il est crucial de normaliser la réponse de stress comme étant nécessaire à la survie.
--------------------------------------------------------------------------------
4. Stratégies d'Adaptation et Solutions
Face à ce constat, des solutions émergent à la fois au niveau individuel, familial et sociétal.
Techniques Personnelles et Familiales
• Suivre son intuition : Les jeunes gèrent souvent mieux le stress en suivant leur intuition : jouer, chanter, rire pour dépenser l'énergie accumulée.
• Techniques de régulation simples : Des méthodes scientifiquement prouvées sont souvent les plus simples.
◦ Respiration abdominale : Une "belle grosse respiration avec une belle grosse bédaine" active le système parasympathique et arrête la réponse de stress.
◦ Chanter : A un effet similaire à la respiration profonde.
◦ Méditation : Pratiquée par Magalie pour faire le vide et analyser sa journée.
• Communication et soutien : L'écoute est fondamentale.
Le père de Sopiane a appris qu'au lieu de proposer des solutions, il est plus efficace d'écouter, de valider l'émotion et d'offrir un soutien inconditionnel ("je suis là pour toi").
• Changement de vocabulaire et mantras : Adopter un langage qui normalise l'inconfort sans le dramatiser.
Le mantra utilisé par Judith et Sopiane est un exemple puissant : "C'est vraiment inconfortable en ce moment mais ça va pas durer."
• Prise de conscience parentale : Les parents doivent réfléchir à l'impact de leur propre anxiété de performance sur leurs enfants et ajuster leurs attentes, en se concentrant sur le processus d'apprentissage plutôt que sur le résultat.
Pistes de Réflexion Sociétales
• Créer des conditions de croissance : Selon Carl la Charité, les enfants n'ont pas besoin qu'on "tire sur la fleur pour qu'elle pousse". Ils ont besoin de conditions propices :
◦ Jouer dehors.
◦ Interagir avec d'autres enfants, y compris dans le conflit.
◦ Avoir des adultes qui fixent des limites mais prennent aussi le temps d'écouter.
• Réformer le système éducatif : S'inspirer de modèles moins axés sur la performance et les examens, comme le système finlandais, pour recentrer l'éducation sur le goût d'apprendre.
• Parler ouvertement : La démarche de la narratrice montre que le fait de parler ouvertement de l'anxiété, de poser des questions et de partager des expériences permet de briser l'isolement, de valider les ressentis et d'ouvrir la voie à des solutions collectives.
-
-
www.france.tv www.france.tv
-
Synthèse sur la Crise de la Santé Mentale de la Jeunesse en France
Résumé Exécutif
Le système de santé mentale pour la jeunesse en France est en état de crise profonde, caractérisée par une augmentation exponentielle du mal-être psychique chez les mineurs et un effondrement structurel du système de soin public.
Un jeune sur cinq souffre désormais de symptômes dépressifs sévères, et les passages aux urgences psychiatriques pour mineurs ont triplé en cinq ans.
Cette détresse est aggravée par une pénurie abyssale de moyens : plus de la moitié des lits en pédopsychiatrie ont été supprimés en une décennie, les Centres Médico-Psychologiques (CMP) sont saturés avec des listes d'attente pouvant atteindre deux ans, et le nombre de pédopsychiatres est dramatiquement insuffisant.
Face à ce système public exsangue, les familles et les soignants sont poussés à bout.
Les médecins sont contraints de "trier" les adolescents suicidaires, renvoyant chez eux des cas graves par manque de place, ce qui mène à des situations de non-assistance à personne en danger.
En réponse, des solutions palliatives émergent, dont l'efficacité est souvent contestée.
On observe une sur-médicamentation croissante, avec un doublement des prescriptions d'antidépresseurs en sept ans, parfois sans suivi psychologique adéquat et avec des effets secondaires dangereux.
Simultanément, le secteur privé lucratif se développe, profitant des défaillances du public mais soulevant des questions d'inégalité d'accès aux soins et de déstabilisation de l'hôpital public.
Les initiatives gouvernementales, comme les formations aux premiers secours en santé mentale ou les financements par appels à projets, apparaissent largement insuffisantes pour endiguer une crise systémique qui nécessite une refonte politique et financière massive.
--------------------------------------------------------------------------------
I. Une Crise Sanitaire d'Ampleur Inédite
La santé mentale de la jeunesse française traverse une période de détresse sans précédent.
Les témoignages et les données statistiques dressent le portrait d'une génération en grande souffrance, confrontée à un vide psychologique et un système de soin incapable de répondre à ses besoins.
A. L'Augmentation Alarmante du Mal-être Psychique
Les indicateurs de santé mentale chez les jeunes sont au rouge, témoignant d'une épidémie silencieuse.
• Statistiques Clés :
◦ Un jeune sur cinq en France souffre de symptômes dépressifs sévères.
◦ Le nombre de passages de mineurs aux urgences psychiatriques a triplé au cours des cinq dernières années.
◦ Les tentatives de suicide sont de plus en plus nombreuses et concernent des enfants de plus en plus jeunes (8, 9, 10 ans).
• Témoignages de la Souffrance :
◦ Un jeune décrit sa dépression : "L'envie de mourir me revient. Elle plane au-dessus de moi comme un grand oiseau sombre [...]. Elle prend toute la place. Elle prend ma place."
◦ Un adolescent de 15 ans aux urgences confie avoir des idées suicidaires "en permanence" depuis l'âge de 11 ans.
◦ Une mère de famille résume l'expérience : "Si j'avais à résumer et à imager en un mot la dépression de notre enfant, ce serait enfer."
B. Les Facteurs Contributifs
Plusieurs phénomènes sociaux et conjoncturels sont identifiés comme des catalyseurs de cette crise.
• Facteurs Sociaux : Le harcèlement scolaire, l'impact des réseaux sociaux, les difficultés économiques et sociales des familles, ainsi que l'éco-anxiété sont cités comme des éléments fragilisant les adolescents et les jeunes adultes.
• L'Effet du Covid-19 : La période de la pandémie a exacerbé ce phénomène de mal-être de manière significative.
--------------------------------------------------------------------------------
II. L'Effondrement du Système de Soin Public en Pédopsychiatrie
Le principal paradoxe de cette crise est que, alors que la détresse psychique n'a jamais été aussi forte, l'accès à une prise en charge adéquate et durable n'a jamais été aussi difficile.
Le système public de pédopsychiatrie est en état de délabrement avancé.
A. Une Pénurie Structurelle et Historique
La crise actuelle est le résultat de décennies de décisions politiques et de sous-investissement.
• Origine Politique : Le "virage ambulatoire" initié dans les années 1980, visant à favoriser les soins en dehors de l'hôpital, a conduit à une fermeture excessive de lits.
• Manque de Lits :
◦ Plus de la moitié des lits en pédopsychiatrie ont disparu en seulement 10 ans.
◦ Une dizaine de départements français ne possèdent plus aucun lit de pédopsychiatrie.
◦ En Loire-Atlantique, il n'y a que 14 lits pour plus de 320 000 mineurs.
• Manque de Personnel :
◦ Seulement 600 pédopsychiatres exercent encore dans le service public sur tout le territoire.
◦ Les conditions de travail exténuantes provoquent un épuisement professionnel et des démissions, comme en témoigne un médecin :
"En moins de 6 mois, ce sont deux de mes collègues pédopsychiatres excellents [...] qui sont partis [...] sinon ils allaient y laisser leur peau."
• Surcharge des Structures Ambulatoires : Les Centres Médico-Psychologiques (CMP), devenus le pilier du soin, sont submergés.
Dans certaines régions, les délais d'attente pour un rendez-vous peuvent atteindre deux ans.
B. Conséquences Dévastatrices pour les Patients et les Familles
Le "manque de soin est abyssal", forçant les familles à un véritable "parcours du combattant".
| Problème Systémique | Conséquences Directes | | --- | --- | | Manque de lits et de places | Tri des patients suicidaires : Les médecins doivent choisir qui hospitaliser. Un médecin de l'hôpital Kremlin-Bicêtre explique : "On avait à manager trois. Ben, on prend le plus grave." | | Saturation des urgences | Renvois à domicile de cas critiques : Au CHU de Nantes, 162 enfants ont été renvoyés chez eux en 2024, bien que leur état ait été jugé comme nécessitant une hospitalisation. | | Absence de structures adaptées | Hospitalisations inappropriées : Des mineurs sont placés dans des services de psychiatrie pour adultes, une pratique traumatisante et désormais illégale mais toujours en cours. Une jeune fille témoigne : "J'étais avec des personnes qui criaient la nuit, qui se cognent la tête contre les vitres." | | Désengagement de l'État | Isolement et détresse des familles : Laissés sans solution, les parents vivent dans une peur constante. "On a peur d'ouvrir la porte \[de la chambre de notre enfant\]", confie un père. Certains sont contraints d'arrêter de travailler, s'endettant pour tenter de sauver leur enfant. Ils créent des groupes de soutien sur les réseaux sociaux pour briser l'isolement. |
C. L'Épuisement et la Colère des Soignants
Les professionnels de santé sont en première ligne, confrontés à l'impossibilité d'accomplir leur mission.
• Souffrance Morale : Un psychiatre déclare : "J'ai affaire tous les jours à des collègues qui pleurent régulièrement parce que [...] on fait de la merde au quotidien."
• Sentiment d'Impuissance : La seule réponse souvent possible est : "Nous n'avons rien à vous proposer."
• Actions de Protestation : À Nantes, des soignants ont saisi symboliquement la justice pour "non-assistance à personne en danger".
Des manifestations et des alertes publiques se multiplient depuis deux décennies, sans résultats concrets.
--------------------------------------------------------------------------------
III. Des Solutions Contestées et l'Avènement du Secteur Privé
Face à la défaillance du public, diverses réponses émergent, allant de la sur-médicamentation à des initiatives gouvernementales jugées insuffisantes, jusqu'à une privatisation croissante du secteur.
A. La Sur-médicamentation et ses Dangers
Le manque de temps et de moyens pour des suivis psychologiques pousse à un recours massif aux traitements médicamenteux.
• Explosion des Prescriptions : La prescription d'antidépresseurs chez les jeunes a plus que doublé en seulement 7 ans.
• Une Pratique Douteuse : Des médecins avouent prescrire massivement, à l'encontre de leurs pratiques passées.
"Avant, j'avais l'impression de soigner et maintenant je prescris", affirme une professionnelle.
Ces prescriptions sont souvent faites dès la première consultation, sans accompagnement psychologique, ce qui est contraire aux recommandations officielles.
• Effets Secondaires Graves :
◦ Les antidépresseurs peuvent aggraver les pulsions suicidaires dans les premières semaines de traitement.
Une jeune fille témoigne avoir fait une tentative de suicide avec les médicaments prescrits par un psychiatre qui l'avait laissée sans suivi.
◦ Le cas de Romain, 16 ans, est emblématique. Après la prescription de paroxétine, il a développé des comportements désinhibés et euphoriques avant de se suicider.
• Mise en Cause des Laboratoires : Les parents de Romain ont découvert que les essais cliniques de la paroxétine (laboratoire GSK) sur les enfants avaient été faussés.
GSK a été condamné à une amende de 3 milliards de dollars aux États-Unis pour ces faits.
B. Des Réponses Politiques et Technologiques Insuffisantes
Les autorités tentent de répondre à la crise, mais les mesures mises en place sont critiquées pour leur superficialité.
• "Premiers Secours en Santé Mentale" : Une formation de 14 heures importée d'Australie et massivement déployée.
Une chercheuse ayant analysé 46 études internationales conclut que rien ne prouve l'efficacité de ce programme pour inciter les personnes en souffrance à se faire soigner.
L'argent investi (plus de 12 millions d'euros) serait mieux utilisé ailleurs.
• Le Système des "Appels à Projet" : Le gouvernement finance des projets "innovants" pour une durée limitée (3 ans).
◦ Exemple du CAP d'Amiens : Ce centre d'accueil post-crise a montré des résultats probants (80% des jeunes n'ont pas été ré-hospitalisés dans les 6 mois).
◦ Problème de Pérennité : Malgré son succès, son financement n'a pas été pérennisé, au motif qu'il n'avait pas dépensé tout son budget, illustrant une logique comptable déconnectée des besoins de soin à long terme.
• Thérapies Virtuelles et Chatbots : Une tendance venue des États-Unis, où des start-ups développent des IA pour remplacer les thérapeutes.
Des experts alertent sur le danger de ces outils, qui peuvent créer une dépendance, aggraver l'isolement et ont été reliés à une augmentation des actes suicidaires chez les 10-14 ans.
C. La Montée en Puissance du Privé Lucratif
Le vide laissé par le service public est de plus en plus comblé par des acteurs privés à but lucratif.
• Le Dernier Recours des Familles : Des familles se tournent vers des cliniques privées, souvent à des centaines de kilomètres de chez elles et à des coûts prohibitifs (ex: 210 € par jour).
• Le Cas d'Éméis à Strasbourg : Ce groupe (anciennement Orpea) prévoit de construire une clinique psychiatrique de 80 lits.
◦ Controverse : Le projet est critiqué car il risque de "dépouiller" l'hôpital public de ses professionnels et de ne prendre en charge que les cas les moins complexes, laissant les situations les plus dramatiques au public.
◦ Rôle Ambigu de l'ARS : L'Agence Régionale de Santé a approuvé ce projet privé tout en refusant la création d'un hôpital de jour public au même endroit, soulevant des questions sur l'influence du lobbying.
◦ Interrogations : Le délégué ministériel à la santé mentale s'étonne lui-même d'un tel projet, le jugeant à contre-courant des recommandations qui privilégient les prises en charge ambulatoires.
-
-
www.france.tv www.france.tv
-
Document d'information : Jeunes Sans-Abri à 20 ans
Synthèse Exécutive
Ce document de synthèse analyse les témoignages de plusieurs jeunes sans-abri, mettant en lumière les causes multifactorielles de leur situation, les dures réalités de leur quotidien et leurs sources de résilience.
Les points clés sont les suivants :
• Des parcours de rupture profonde : L'itinérance des jeunes est rarement un choix, mais plutôt la conséquence de traumatismes et de ruptures systémiques.
Les causes principales identifiées sont le rejet familial violent, l'abandon à la majorité par les services de l'Aide Sociale à l'Enfance (ASE), et les parcours migratoires périlleux en tant que demandeurs d'asile.
• Une survie quotidienne brutale : La vie dans la rue est une lutte incessante contre l'épuisement physique et psychologique. Elle est marquée par la mendicité ("la manche"), l'exposition aux éléments, le manque d'hygiène, et un sentiment d'invisibilité et de danger permanent. La précarité expose à la violence, au vol et à une déconnexion progressive avec la réalité.
• La résilience face à l'adversité : Malgré des conditions extrêmes, ces jeunes font preuve d'une forte résilience. Ils s'accrochent à des passions personnelles (musique, sport, jeux), à des relations humaines (amour, amitié) et à des aspirations fortes pour l'avenir, comme la poursuite des études, la fondation d'une famille ou l'obtention d'un logement stable, qui deviennent des moteurs pour s'en sortir.
• Les obstacles systémiques et les lueurs d'espoir : La dépendance aux drogues apparaît comme un facteur aggravant majeur, créant un cercle vicieux difficile à briser seul.
L'accès aux services d'urgence (115) est souvent saturé.
Cependant, l'engagement collectif, la solidarité entre pairs et l'aide des associations spécialisées offrent des perspectives concrètes de réinsertion, menant dans certains cas à des réussites tangibles (obtention de statut, logement, reprise des études).
--------------------------------------------------------------------------------
1. Origines de l'itinérance : Des parcours de rupture
L'entrée dans la rue à un jeune âge est systématiquement liée à des événements traumatisants et à des failles dans les filets de sécurité sociaux et familiaux.
1.1. La rupture familiale
Le rejet et les violences intra-familiales sont une cause directe de l'itinérance pour plusieurs jeunes. L'environnement familial est décrit non pas comme un refuge mais comme la source du problème.
• Le rejet parental dès la naissance : Une jeune femme raconte avoir été un enfant non désiré, un sentiment verbalisé par sa mère et renforcé par l'abandon de son père.
• La fuite comme seule issue : Face à un harcèlement familial constant (rabaissement, dénigrement), la fugue est perçue comme un acte de survie pour éviter une implosion.
1.2. La sortie de l'Aide Sociale à l'Enfance (ASE)
La fin de la prise en charge à 18 ans constitue un point de bascule critique, projetant des jeunes sans soutien familial directement dans la précarité.
• L'abandon institutionnel : Un jeune homme, placé de 2 à 18 ans, décrit sa mise à la rue comme une conséquence directe du refus de son contrat jeune majeur.
1.3. Les parcours migratoires
Pour les jeunes étrangers, l'itinérance est une conséquence directe de leur parcours d'exil et des difficultés rencontrées à leur arrivée en France.
• La fuite de la violence : Un jeune homme brésilien explique avoir fui la favela en raison des menaces des narcotrafiquants et des dettes de drogue de sa famille.
• La désillusion à l'arrivée : Un jeune demandeur d'asile pensait être pris en charge immédiatement, mais a été confronté à la réalité de la rue dès sa première nuit.
2. La réalité quotidienne de la rue : Survie et vulnérabilité
La vie sans-abri est un combat permanent marqué par la précarité matérielle, la détresse psychologique et une exposition constante aux dangers.
2.1. Les difficultés matérielles et physiques
Le quotidien est une lutte pour subvenir aux besoins les plus élémentaires.
• L'absence de ressources : Une jeune femme enceinte décrit son dénuement total.
• La mendicité ("la manche") : C'est une activité épuisante et nécessaire pour se nourrir.
• L'épuisement et l'inconfort : L'exposition aux intempéries et la marche continue mènent à un épuisement extrême.
2.2. L'impact psychologique
La rue a des effets dévastateurs sur la santé mentale.
• Le sentiment d'invisibilité et de solitude : L'isolement est profond, menant à la peur de mourir seul et sans que personne ne s'en aperçoive.
• La confusion entre rêve et réalité : L'épuisement et le stress permanent créent un état de déphasage mental.
• La survie plutôt que la vie : L'existence se réduit à une lutte pour les besoins primaires, effaçant toute notion de vie épanouie.
2.3. Les dangers et l'insécurité
La rue est un environnement hostile où la méfiance est une règle de survie.
• La violence et le vol : Les agressions sont une réalité tangible et le vol est fréquent, même pendant le sommeil.
• La vulnérabilité des femmes : Les femmes seules sont particulièrement exposées aux agressions, notamment la nuit.
• Le squat dans des lieux dangereux : Pour se mettre à l'abri, certains occupent des bâtiments délabrés et dangereux.
3. Mécanismes de résilience et sources d'espoir
Face à cette brutalité, les jeunes développent des stratégies de survie psychologique et s'accrochent à des projets et des relations qui leur donnent la force de continuer.
3.1. Les passions personnelles comme échappatoires
Les activités personnelles permettent de s'évader mentalement de la dureté du quotidien.
• Le Rubik's Cube : Pour une jeune réfugiée, ce jeu fait disparaître les problèmes.
• Le piano : Pour un jeune brésilien, jouer du piano est un lien avec son passé et sa grand-mère, apportant joie et motivation.
• Le roller : Cette activité est synonyme de liberté et permet de combattre la solitude.
3.2. L'importance des relations et de la parentalité
Les liens affectifs sont un puissant soutien. L'arrivée d'un enfant devient un objectif majeur pour s'en sortir.
• L'amour dans la rue : Être en couple est une épreuve mais aussi une force.
• La grossesse comme moteur : Un enfant à naître transforme la lutte pour la survie en un projet de construction d'un avenir stable, motivé par la peur d'un placement.
3.3. La quête de dignité et de normalité
Conserver une part de "vie normale" est essentiel pour le moral.
• L'apparence : Rester bien habillé et propre grâce aux associations est une façon de maintenir son estime de soi.
• La douche : Ce moment est décrit comme une renaissance, un moyen de laver la saleté et de retrouver une part de dignité aux yeux des autres.
4. La dépendance comme facteur aggravant
Pour une des jeunes femmes, la consommation de crack est à la fois une conséquence et une cause de son maintien dans la rue, créant un cycle d'autodestruction.
• La conscience de l'addiction : Elle reconnaît lucidement sa dépendance et le fait qu'elle l'empêche d'avancer.
• L'isolement social : La drogue et la mendicité qui en découle l'ont coupée de toute relation sociale normale.
• La prise de conscience : Elle réalise qu'elle ne peut pas s'en sortir seule et qu'elle doit accepter de l'aide extérieure.
5. L'engagement collectif et la recherche de solutions
Face à l'inertie des institutions, certains jeunes choisissent l'action collective pour faire entendre leur voix.
• La solidarité entre pairs : L'union est vue comme une nécessité pour trouver des solutions.
• L'action politique : Des actions comme des occupations sont menées pour interpeller la mairie et exiger des solutions de logement, considéré comme un droit fondamental.
• L'épreuve des démantèlements : Les expulsions de campements sont vécues comme des traumatismes répétés, aggravant l'épuisement physique et moral.
6. Les perspectives d'avenir : Entre aspiration et précarité
Malgré les obstacles, tous les jeunes expriment des rêves et des projets concrets. La fin du documentaire montre que des évolutions positives sont possibles.
| Individu | Situation Initiale | Aspiration / Projet | Évolution à la fin | | --- | --- | --- | --- | | La jeune étudiante | Vit en campement avec sa famille. | Poursuivre ses études, devenir avocate ou médecin, avoir sa propre chambre. | A obtenu son brevet avec mention, est affectée en lycée général. Vit en hôtel avec sa famille. | | Le jeune demandeur d'asile (africain) | Dort dans le parc de Belleville. | Être considéré comme mineur, aller à l'école, avoir un toit. | Est reconnu mineur, suit un CAP et a obtenu un hébergement. | | Le jeune demandeur d'asile (brésilien) | Squatte un bâtiment incendié. | Travailler déclaré, quitter la précarité. | Travaille au noir, a pu louer une petite chambre, fait des économies. | | Le couple attendant un enfant | Vit dans la rue. | Trouver un logement stable pour accueillir leur enfant et éviter le placement. | Sont toujours à la recherche d'une solution rapide. | | La jeune femme dépendante | Lutte contre son addiction et la vie dans la rue. | Partir faire les saisons avec un camion aménagé. | A pris contact avec une association, initiant une première étape vers la sortie de rue. |
Ces parcours montrent que si la sortie de l'itinérance est un processus long et difficile, l'accès à un statut, un logement, une formation ou un accompagnement social constitue le point de départ indispensable à la reconstruction.
-
-
www.france.tv www.france.tv
-
Document d'information : La mouvance du "mâle alpha"
Synthèse
Ce document d'information synthétise les thèmes, les figures clés et les impacts de la mouvance du "mâle alpha", un phénomène social émanant principalement des réseaux sociaux.
Porté par des influenceurs comme Andrew Tate à l'international et des figures québécoises telles que Julien Bournival, ce mouvement prône un retour aux valeurs traditionnelles et à des rôles de genre strictement définis, où l'homme est le pourvoyeur et le leader, et la femme, plus soumise, se consacre au foyer.
L'idéologie fondamentale repose sur une forme de déterminisme biologique, affirmant que les hommes et les femmes possèdent des caractéristiques innées et distinctes qui les destinent à des rôles différents.
Ce discours trouve un écho particulier auprès de jeunes hommes en quête de repères, attirés par un message mêlant développement personnel (discipline, forme physique, succès entrepreneurial) et une rhétorique de rébellion contre un establishment perçu comme hostile.
Les experts analysent cette mouvance comme une manifestation contemporaine d'un discours antiféministe récurrent, intrinsèquement misogyne, qui exprime une crainte de la perte des privilèges masculins face à l'avancée de l'égalité des genres.
Ce phénomène est étroitement lié à une méfiance généralisée envers les institutions (gouvernement, médias, science), à l'adhésion à des théories du complot concernant une "élite" manipulatrice, et à une convergence avec les idéologies de la droite conservatrice, incluant un retour à la religion chrétienne.
Socialement, cette mouvance contribue à une polarisation idéologique croissante entre les jeunes hommes, qui tendent à devenir plus conservateurs, et les jeunes femmes, de plus en plus progressistes.
Son influence est désormais palpable jusque dans les salles de classe, où des discours rétrogrades et masculinistes refont surface, témoignant de la nécessité d'une vigilance continue face à la remise en question des acquis en matière d'égalité.
--------------------------------------------------------------------------------
1. Définition et idéologie du mouvement "mâle alpha"
Le mouvement "mâle alpha" est défini comme un phénomène émanant d'influenceurs du web et des réseaux sociaux qui prônent un retour à certaines valeurs traditionnelles. Son idéologie repose sur plusieurs piliers fondamentaux.
Principes fondamentaux :
• Rôles de genre traditionnels : L'homme assume le rôle de leader et de pourvoyeur ("provider", "chef à la maison"), tandis que la femme est plus soumise et se consacre au foyer et à la famille ("nurture").
Un influenceur affirme : "La règle c'est que l'homme est un homme puis est masculin puis il doit être le chef à la maison puis c'est lui le provider."
• Force et responsabilité masculine : L'homme "alpha" doit être fort physiquement et mentalement, prendre ses responsabilités, protéger et subvenir aux besoins de sa famille.
• Contrôle dans la relation : Certains discours promeuvent un contrôle sur la partenaire féminine. Un extrait viral stipule : "Quand tu es en couple, tu laisses pas ta blonde sortir d'un club.
Tu laisses pas ta blonde aller dans un festival. Tu laisses pas ta blonde de mettre des photos de ses fesses en Gstring sur Instagram."
Justification par le déterminisme biologique :
• Différences innées : Les partisans soutiennent que les hommes et les femmes sont biologiquement différents, ce qui détermine leurs traits de caractère. L'homme serait naturellement "assertif", "direct" et "fonceur", tandis que la femme serait dotée d'une "sensibilité" et d'une "intuition" supérieures.
• Rejet de l'égalité des compétences : L'idée que les hommes et les femmes sont égaux en aptitudes et en compétences est jugée "complètement ridicule" par Julien Bournival.
• Hypergamie : Un coach en dating du mouvement affirme que les femmes sont biologiquement attirées par des hommes qui leur sont supérieurs en termes de confiance, charisme, salaire, grandeur et force, un concept qu'il nomme "hypergamie".
La "crise de la masculinité" :
• Les influenceurs du mouvement estiment qu'il existe une "crise de la masculinité" causée par une société qui perçoit la masculinité comme "toxique" et tente d' "émasculiner" les hommes.
• Cette perception est partagée par de jeunes hommes qui se sentent attaqués ou dévalorisés. Une jeune femme observe : "À force de se faire dire qu'on est méchant, qu'on est pas bon, qu'on est un problème, mais je pense que leur réaction c'est la colère."
2. Figures clés et leurs discours
Plusieurs influenceurs sont identifiés comme des figures centrales de ce mouvement, chacun avec un style et une portée distincts.
Andrew Tate : La figure de proue internationale
• Profil : Influenceur britanno-américain, ancien champion de kickboxing, décrit comme une "méga star" et l'une des personnes les plus recherchées sur Google. Il a été arrêté en Roumanie pour trafic d'êtres humains, viol et formation d'un gang criminel.
• Message double : Son discours est un mélange de développement personnel (discipline, détermination, prise de responsabilité) et de propos jugés "irrespectueux, misagènes [sic] envers la femme".
• Défense de ses partisans : Ses adeptes, comme Julien Bournival, défendent "l'essence de son message" tout en minimisant ses controverses, les qualifiant de "jokes déplacés" ou d'actes d'un "personnage" destiné à provoquer. Un jeune homme affirme : "si m'aide à faire de l'argent. Je vois pas pourquoi je veux dire c'est une mauvaise personne".
Julien Bournival : Le modèle québécois en Floride
• Profil : Entrepreneur québécois installé en Floride, il se décrit comme faisant partie du "1 % en terme de revenu" et du "1 % en terme de fitness". Il a quitté le Québec, qu'il qualifiait de "République socialiste" durant la pandémie.
• Discours : Il prône un retour aux valeurs traditionnelles, se définit comme un "pourvoyeur" et vit une relation où sa femme s'occupe de la maison et de la famille. Il lie de plus en plus ses valeurs à sa foi chrétienne.
• Activité entrepreneuriale : Il dirige une entreprise (Global) dans le domaine de l'amélioration énergétique, mais utilise ses réunions d'employés comme des séances de "croissance personnelle" où il promeut sa vision du monde, affirmant que les entrepreneurs ont une "responsabilité morale" de bâtir un peuple fort contre les "dirigeants" qui veulent un peuple faible et contrôlable.
Louis Rassico : L'influenceur repenti
• Parcours : Jeune entraîneur québécois, il a été l'une des premières figures "mâle alpha" au Québec. Il admet avoir été influencé par Andrew Tate et avoir copié son style "intense" ("Ferme ta gueule") pour gagner en popularité, ce qui a fonctionné.
• Prise de distance : Il a depuis changé de discours, qualifiant Tate de "manipulateur" et décrivant son propre parcours comme une "déprogrammation" ou une "déradicalisation". Il a réalisé qu'il "perdai[t] contact avec la vraie réalité des choses".
Chloé Roma : La défenseure des droits des hommes
• Position : Canadienne connaissant un grand succès en défendant les droits des hommes. Elle soutient que les hommes sont en crise, manquent de modèles positifs et sont toujours soumis à l'attente d'être "protecteur et pourvoyeur", contrairement aux femmes qui sont maintenant perçues comme capables de multiples rôles.
• Analyse sur Tate : Elle pense que le succès de Tate s'explique par le fait qu'il a touché une audience d'hommes sans figure paternelle ou modèle masculin positif, mais critique le fait que son message renforce les attentes négatives déjà pesantes sur les hommes.
3. Analyse critique et impacts sociétaux
Des experts et des acteurs de la société civile offrent une analyse critique de ce mouvement et de ses conséquences.
Perspective sociologique (Francis Dupuis-Déri) :
• Discours récurrent : La "crise de la masculinité" n'est pas un phénomène nouveau. Des discours similaires existent depuis l'Antiquité romaine et à chaque siècle depuis, quel que soit le contexte politique ou culturel.
• Nature misogyne : Le discours de la crise est "nécessairement misogyne" car il postule que (1) les hommes vont mal, (2) c'est à cause des femmes, et (3) la solution est un retour à une masculinité traditionnelle.
• Réponse à l'égalité : Ce mouvement est une forme d'antiféminisme porté par des hommes qui "ne veulent pas de l'égalité" et voient le progrès des droits des femmes comme une "menace" à leurs privilèges.
• Réfutation du déterminisme : L'idée de rôles biologiquement définis est contredite par l'histoire de l'humanité, qui montre une grande diversité de rôles assumés par les hommes et les femmes. La différence des rôles est avant tout liée à la "socialisation et des éducations différentes".
Impact en milieu scolaire (Véronique Guitras, enseignante) :
• Retour de discours rétrogrades : L'enseignante a constaté un "clash de discours" dans sa classe après un congé de maternité. Des élèves masculins tiennent désormais des propos "conservateurs, traditionnels, masculinistes".
• Exemples concrets : Un élève lui a affirmé que l'aspiration de toutes les femmes est d'être "invité sur un yat à Dubaï", et qu'elles ne sont pas des "bâtisseuses" comme les hommes. Elle décrit ce phénomène comme un retour "60 ans en arrière".
Polarisation idéologique croissante :
• Fossé de genre : Un fossé idéologique se creuse chez les jeunes en Occident : les jeunes femmes deviennent de plus en plus progressistes et féministes, tandis que les jeunes hommes deviennent de plus en plus conservateurs.
• Débat "l'homme ou l'ours" : Ce débat viral illustre la méfiance des femmes envers les hommes.
Une jeune femme explique préférer rencontrer un ours dans la forêt, car "l'ours quand il va m'attaquer, on va pas me demander comment j'étais habillée avant".
Une autre affirme qu'il est "nécessaire pour nous de se méfier de tous les hommes" pour leur propre sécurité.
4. Liens avec le conservatisme et les théories du complot
Le discours "mâle alpha" est intrinsèquement lié à une méfiance envers les institutions et à une adhésion à des idéologies conservatrices et conspirationnistes.
Méfiance envers les institutions :
• Rejet de l'autorité : Il existe une perte de confiance généralisée envers la science, la médecine, le gouvernement et surtout les médias, qualifiés d' "agence de publicité du gouvernement".
Ce phénomène a été "considérablement accéléré" par la pandémie.
• Sentiment d'abandon : Selon l'anthropologue Samuel Viger, ce rejet peut provenir d'un sentiment d'abandon par le système (crises du logement, de la santé, inégalités croissantes), poussant certains individus vers des discours marginaux.
Rhétorique conspirationniste :
• L'élite manipulatrice : Les influenceurs de la mouvance véhiculent l'idée qu'une "élite" satanique contrôle le monde et cherche à affaiblir la population en s'attaquant à la famille traditionnelle, en "brainwashant" les enfants et en promouvant une société de "weak person".
• La posture de rébellion : Adopter les valeurs "mâle alpha" est présenté comme "l'ultime rébellion" contre ce système de contrôle.
Convergence avec la droite et la religion :
• Idéologie de droite : Le mouvement s'aligne sur des valeurs conservatrices. Julien Bournival admire Donald Trump et s'est installé en Floride pour le mode de vie républicain promu par Ron DeSantis.
• Retour à la foi chrétienne : Plusieurs figures du mouvement, dont Bournival, se tournent vers la Bible pour justifier les valeurs traditionnelles.
Le passage biblique sur la soumission de la femme à l'homme (Éphésiens 5:22-33) est cité comme un "code d'éthique". La foi est présentée comme une garantie morale pour la soumission de la femme.
Hostilité envers les minorités de genre :
• Vision rigide des genres : L'existence de personnes transgenres et de drag queens est perçue comme une attaque directe à leur conception "biologisante" et naturelle de l'homme et de la femme.
• Accusations de "grooming" : Les drag queens qui lisent des contes aux enfants sont accusées de "grooming" et de faire partie d'un "agenda satanique".
Cette rhétorique escalade jusqu'à des comparaisons avec la pédophilie : "c'est quoi la prochaine affaire [...] c'est on va accepter les pédophiles".
5. Citations marquantes
| Thème | Citation | Locuteur | | --- | --- | --- | | Idéologie Mâle Alpha | "Chris, allez au gym, arrêtez de faire vos couches de guilleir." | Extrait audio d'influenceur | | Rôles Traditionnels | "La règle c'est que l'homme est un homme puis est masculin puis il doit être le chef à la maison puis c'est lui le provider." | Julien Bournival | | Soumission féminine | "Moi j'aime mieux être dans le shadow, m'occuper de notre maison \[...\] Va à la guerre, va au front, moi je reste derrière." | Partenaire de Julien Bournival | | Critique d'Andrew Tate | "On s'entend que Andw Tate a des propos irrespectueux, misagène envers la femme en général." | Journaliste | | Défense d'Andrew Tate | "L'essence de son message \[...\] c'est respecte-toi, respecte les autres, prends soin de toi. Assure-toi que quand tu dis de quoi, ta parole vaut de quoi." | Julien Bournival | | Impact scolaire | "Je me retrouve devant des jeunes qui ont des discours conservateurs, traditionnels, masculinistes \[...\] A on est revenu 60 ans en arrière." | Véronique Guitras, enseignante | | Analyse sociologique | "Le discours de crise \[...\] il dit les hommes vont mal. Ils vont mal à cause de qui ? Ils vont mal à cause des femmes." | Francis Dupuis-Déri, sociologue | | Polarisation | "L'ours quand il va m'attaquer, on va pas me demander comment j'étais habillée avant." | Jeune femme | | Théorie du complot | "Ceux qui contrôlent le monde sont satanique. Ils contrôlent les gouvernements." | Julien Bournival | | Repentir | "\[Andrew Tate\] est manipulateur clairement. \[...\] Moi-même je me suis fait influencer par lui \[...\] Je me suis déprogrammé." | Louis Rassico, entraîneur | | Vigilance | "J'ai deux filles. J'ai pas envie qu'elle vivent dans un monde inégalitaire. J'ai pas envie qu'elle soit soumise à quiconque. \[...\] il y a rien de gagner pour toujours." | Journaliste |
-
-
-
Synthèse sur le Harcèlement Scolaire : Causes, Conséquences et Stratégies de Prévention
Résumé Exécutif
Ce document de synthèse analyse en profondeur la problématique du harcèlement scolaire en France, en s'appuyant sur des témoignages et des études de cas poignants.
Il met en lumière les conséquences tragiques du harcèlement à travers les suicides de Dina et Lucas, deux adolescents de 13 ans, tout en explorant les mécanismes sous-jacents, les défaillances institutionnelles et les stratégies de prévention émergentes.
Les principaux points à retenir sont les suivants :
1. Les Visages de la Tragédie : Les cas de Dina et Lucas illustrent la violence extrême du harcèlement. Les motifs sont multiples et ciblent la différence :
- l'intelligence perçue,
- l'apparence physique,
- l'exploration de l'identité sexuelle (Dina),
- ou encore l'homosexualité affirmée et la réussite scolaire (Lucas).
Ces agressions répétées, verbales et physiques, créent un sentiment insupportable de honte et d'isolement menant à des actes désespérés.
2. Défaillances Systémiques : Les témoignages révèlent de graves lacunes dans la réponse des institutions scolaires et judiciaires.
Les collèges ont souvent minimisé les faits, tardé à agir ou adopté des approches inefficaces qui ont aggravé la souffrance des victimes.
Le système judiciaire a initialement classé sans suite l'affaire de Dina et annulé en appel la condamnation des harceleurs de Lucas, niant le lien entre le harcèlement et le suicide, et laissant les familles dans un profond désarroi.
3. Un Changement de Paradigme : Face à ces échecs, de nouvelles stratégies proactives sont mises en œuvre.
Le programme national Phare et la méthode de la préoccupation partagée, adaptée par Jean-Pierre Bélon, marquent une rupture avec l'approche punitive traditionnelle.
Cette méthode non-accusatoire vise à responsabiliser les intimidateurs en les impliquant dans la recherche de solutions pour le bien-être de la victime, transformant ainsi leur comportement par l'empathie.
4. Prévention et Reconstruction : La prévention commence dès l'école primaire, avec des programmes axés sur l'intelligence émotionnelle, la communication non-violente ("messages clairs") et le développement de la confiance en soi.
Pour les victimes, la reconstruction est un long processus impliquant des thérapies (comme celles de l'association Marion la main tendue) pour surmonter des traumatismes profonds tels que la phobie scolaire et la perte d'estime de soi, qui peuvent persister des décennies plus tard.
En conclusion, si le harcèlement scolaire a longtemps été banalisé, une prise de conscience sociétale, catalysée par des drames comme celui de Dina, a conduit à une évolution législative et à l'adoption de méthodes préventives prometteuses.
Celles-ci reposent sur une approche systémique, éducative et empathique, visant à transformer le climat scolaire pour garantir la sécurité et le bien-être de chaque élève.
--------------------------------------------------------------------------------
I. Études de Cas : Les Histoires de Dina et Lucas
A. Le Cas de Dina : Une Spirale de Jalousie et d'Homophobie
Dina, 13 ans, est décrite par son frère, Ryan Gontier, comme une jeune fille "en avance sur son temps", intelligente, curieuse de la politique et de l'actualité, ce qui a suscité la jalousie de ses amies.
• Facteurs Déclencheurs : Le harcèlement a commencé en 4ème (2019) lorsque ses amies ont commencé à lui reprocher son intelligence et sa manière de s'exprimer : "tu es toujours en train de nous corriger parce qu'on parle pas bien français par rapport à toi." Le harcèlement s'est intensifié lorsque Dina a commencé à s'intéresser au courant LGBT et à évoquer la possibilité d'aimer aussi bien les filles que les garçons, menant à son exclusion du groupe.
• La Trahison : Un tournant tragique a eu lieu lorsqu'une de ses amies a prétendu être lesbienne pour sortir avec elle.
Après une vingtaine de jours, elle a rompu brutalement en révélant la supercherie : "Mais moi je t'aime pas, c'est pas vrai. Je tiens à inventer cette histoire."
• L'Escalade du Harcèlement : La nouvelle a été propagée dans la cour du collège, déclenchant des insultes homophobes ("guine", "lesbienne") et des violences physiques.
Le harcèlement s'est ensuite étendu à son physique ("trop grosse") et à ses vêtements.
Sa mère témoigne : "Elle pouvait porter n'importe quoi. C'était prétexte à se faire insulter."
• Première Tentative de Suicide : En mars 2021, profondément blessée, Dina fait une première tentative de suicide par ingestion de médicaments.
Hospitalisée, elle explique ne plus vouloir vivre pour ne pas affronter les moqueries à son retour au collège.
• Le Retour Impossible : Son retour au collège est un calvaire. Sa mère raconte recevoir des appels de détresse : "Maman, je suis aux toilettes. Maman, je vais mourir. Maman, il faut venir me chercher."
• Le Suicide : Malgré un changement de lycée en septembre 2021, une rencontre fortuite avec ses anciennes harceleuses à la cantine un mois plus tard a ravivé son traumatisme. "Je pense qu'elle s'est dit ça va recommencer", analyse son frère.
Une semaine après, Dina met fin à ses jours en se pendant dans son armoire.
B. Le Cas de Lucas : La Différence Ciblée
Lucas, 13 ans, s'est suicidé en janvier 2023 après avoir été harcelé en raison de son homosexualité et de sa réussite scolaire au collège Louis Armand de Golbey.
• Affirmation de Soi : Lucas avait fait son coming-out à sa mère, qui l'avait pleinement soutenu : "Sois heureux mon fils parce que je le savais déjà en fait."
Il est décrit comme un très bon élève, assidu et participatif, ce qui aurait provoqué la jalousie d'autres élèves.
• Harcèlement Continu : Dès son arrivée au collège, Lucas subit des insultes sur son physique, sa manière de s'habiller et de parler, et son orientation sexuelle. Sa mère explique : "Ça a été crescendo avec les quatre élèves dont il m'avait donné les noms déjà."
• Un Refuge dans la Danse : Lucas trouvait un exutoire et un espace d'acceptation dans un club de danse hip-hop, basé sur des valeurs de diversité et d'unité.
• L'Acte Final : Le harcèlement s'est poursuivi jusqu'à la veille de sa mort.
Sa mère pense que les insultes du vendredi 6 janvier ont été "la goutte de trop."
Il a été retrouvé pendu par sa petite sœur de 5 ans et son ex-conjoint.
Sa mère interprète son geste : "Je pense que c'était pour couper sa voix en fait. C'était pour dire stop."
II. Défaillances Institutionnelles et Judiciaires
A. La Réponse des Établissements Scolaires
Dans les deux cas, la réponse des institutions scolaires est présentée comme largement insuffisante.
• Pour Dina : Les parents ont rencontré la Conseillère Principale d'Éducation (CPE) à plusieurs reprises.
Au lieu de confronter les harceleuses, la CPE a demandé à Dina d'écrire leurs noms sur un papier, une tâche que l'adolescente, tremblante, n'a pas pu accomplir.
Cette approche a mis le fardeau de la preuve et de la dénonciation sur la victime déjà fragilisée.
• Pour Lucas : Sa mère a signalé les faits et les noms des harceleurs.
Seul un professeur a réagi en envoyant un communiqué à ses collègues, mais sans suite. "Il y a pas eu de convocation des parents, il y a rien eu." Les sanctions n'ont été prises qu'après le drame : les quatre élèves responsables ont été exclus 14 jours après le décès de Lucas. "C'est avant qu'il aurait fallu le protéger."
• Banalisation du Harcèlement : Un expert souligne une tendance historique en France à banaliser les brimades, considérées à tort comme "formatrices".
Or, "on sait aujourd'hui que ces brimades répétées, ces insultes à répétition, elles sont dévastatrices."
B. Les Obstacles Judiciaires
Les familles ont également dû faire face à un système judiciaire qui a, dans un premier temps, nié la gravité des faits.
• Affaire Dina : L'enquête a été initialement classée sans suite par la procureure. La famille a appris cette décision par la presse.
Le procureur a suggéré que Dina s'était suicidée en raison de sa "fragilité psychologique" et que "rien ne s'était passé dans le collège".
La famille a dû se constituer partie civile pour que l'enquête reprenne finalement en novembre 2023, sur la base de chefs d'accusation incluant harcèlement, provocation au suicide et homicide involontaire.
• Affaire Lucas : En juin 2023, les quatre harceleurs ont été reconnus coupables de harcèlement, mais le tribunal a refusé d'établir un lien avec le suicide.
En novembre 2023, la condamnation a été annulée par la cour d'appel de Nancy, qui a estimé que le harcèlement n'était pas prouvé, ni son effet sur la santé mentale de Lucas.
Sa mère a annoncé un pourvoi en cassation. Les harceleurs, se sentant acquittés, continueraient de rire en parlant de Lucas.
L'affaire Dina a cependant eu un impact législatif majeur, conduisant au vote en mars 2022 d'une nouvelle loi créant le délit de harcèlement scolaire, passible de peines allant jusqu'à 10 ans de prison et 150 000 € d'amende en cas de tentative de suicide de la victime.
III. L'Émergence de Nouvelles Stratégies de Prévention
Face à l'échec des approches traditionnelles, des méthodes alternatives axées sur la prévention et l'empathie sont mises en place dans certains établissements, notamment dans le cadre du programme national Phare.
A. La Méthode de la Préoccupation Partagée
Adaptée en France par Jean-Pierre Bélon, cette méthode vise à résoudre les situations de harcèlement sans passer par l'accusation et la sanction, qui se révèlent souvent contre-productives.
• Principe Clé : Au lieu de confronter l'intimidateur, l'équipe éducative le convoque en lui exprimant son inquiétude pour la victime.
L'objectif est de le transformer en acteur de la solution. On lui demande : "qu'est-ce qu'il pourrait mettre en place pour cet élève ?"
• Posture de l'Adulte : L'entretien doit être court, direct et ferme, mais courtois.
L'adulte ne doit pas entrer dans un débat argumentatif ("c'est elle qui a commencé") mais maintenir le cap sur le mal-être de la victime.
L'autorité est incarnée par la certitude que les brimades doivent cesser.
• Efficacité : Cette méthode montre des résultats très positifs.
Elle permet de "casser ce cercle vicieux" et de créer une nouvelle dynamique. Un principal de collège, Philippe Weis, témoigne que grâce à cette approche, de nombreuses situations (46 en avril) sont réglées avant de devenir graves.
B. L'Implication des Élèves : Les Ambassadeurs Anti-Harcèlement
Le programme Phare prévoit la formation d'élèves ambassadeurs pour jouer un rôle de "sentinelle".
• Rôle : Ces élèves sont formés pour détecter les situations de harcèlement, écouter leurs camarades et alerter les adultes.
Ils ne sont pas des "balances" mais des relais de confiance. Comme l'explique un élève : "Nous, on est des enfants et eux aussi et ils se sentent plus peut-être plus en sécurité."
• Motivation : Beaucoup d'ambassadeurs, comme Charlotte, ont eux-mêmes été victimes de harcèlement, ce qui renforce leur engagement. Dans un collège, 30 élèves se sont portés volontaires pour 12 places initialement prévues.
C. La Prévention dès l'École Primaire
La lutte contre le harcèlement commence dès le plus jeune âge par le développement de compétences psychosociales.
• Intelligence Émotionnelle : Une école primaire de la banlieue parisienne a mis en place des outils pour aider les enfants à identifier et nommer leurs émotions (par exemple, un système de "souris" de différentes couleurs à l'entrée de la classe).
• Méthode des "Messages Clairs" : Les élèves de CM2 apprennent à gérer les conflits en exprimant leur ressenti de manière structurée et non-violente, à l'écart des autres.
Un élève dit à un autre : "Quand tu m'as déplacé mon manteau, je me suis senti mal parce que tu fais ça tout le temps. J'aimerais que tu arrêtes s'il te plaît."
• Confiance en Soi : La directrice de l'école insiste sur l'importance de la confiance en soi, qui permet à un enfant de "réagir face à des brimades" et de "solliciter l'adulte" si besoin.
IV. La Reconstruction et les Séquelles à Long Terme
A. Le Traumatisme des Victimes Survivantes
Le harcèlement laisse des cicatrices psychologiques profondes et durables.
• Savana : Harcelée pour ses bonnes notes, elle a développé une phobie scolaire et sociale qui l'a contrainte à arrêter ses activités extrascolaires. "Ils ont complètement gâché ma vie en fait."
Grâce à l'association Marion la main tendue, elle participe à des groupes de parole et des séances de thérapie où elle apprend à "mettre des mots sur ses maux (m-a-u-x)".
• Julien Masquino : Harcelé 20 ans avant Lucas dans le même collège pour son "côté un peu efféminé", son "embonpoint" et son "langage châtié", il a souffert de l'effet de meute.
Le harcèlement a détruit son estime de soi : "Vous vous dites qu'un moment peut-être que si je suis victime c'est que je le mérite."
Il a repris confiance en lui grâce au sport et enseigne aujourd'hui le self-défense à des adolescents.
B. Le Deuil Impossible des Familles
Pour les familles des victimes, le deuil est inextricablement lié à la reconnaissance du harcèlement comme cause du décès.
• Le frère de Dina déclare : "Je pense pas que mes parents et moi on puisse faire notre deuil sans que son harcèlement scolaire soit reconnu comme étant la cause de son suicide."
• La douleur est constante, comme en témoigne le texte lu à la fin du document :
Ton visage, ton sourire me hante, tes câlins, tes mots doux me manquent. [...] J'aimerais tellement savoir si de là-haut tu pouvais me voir, si de là où tu pouvais comprendre que sans toi plus rien n'a de sens.
-
-
www.france.tv www.france.tv
-
Synthèse sur les Auteurs Mineurs de Violences Sexuelles
Résumé Exécutif
Ce document de synthèse analyse les témoignages et expertises concernant des mineurs auteurs de violences sexuelles.
L'analyse révèle une corrélation quasi systématique entre le passage à l'acte et un historique de victimisation durant l'enfance.
Ces jeunes, loin d'être des "monstres", sont souvent des individus au psychisme "cabossé" par des traumatismes précoces, notamment des abus sexuels, des dysfonctionnements familiaux profonds (abandon, négligence, secrets) et une exposition précoce à une pornographie violente.
Le passage à l'acte est mû par des mécanismes psychologiques complexes tels que la répétition traumatique, l'identification à l'agresseur, la vengeance ou une dépendance compulsive.
Face à cette complexité, la prise en charge psychiatrique et psychologique est présentée comme un levier fondamental et efficace.
Elle vise à la fois la responsabilisation de l'auteur et la compréhension des racines de son acte, permettant une évolution positive et un faible taux de récidive.
L'impact sur les familles, qualifiées de "victimes collatérales", est immense, marqué par la culpabilité, la honte et un besoin crucial de soutien.
Enfin, le document souligne un manque criant de moyens dédiés à la santé mentale des jeunes et l'urgence de renforcer la prévention, arguant que prendre en charge les auteurs est une forme essentielle de protection des futures victimes.
--------------------------------------------------------------------------------
I. Le Cycle Victime-Agresseur : Une Thématique Centrale
L'un des constats les plus saillants émergeant des sources est le lien direct entre le statut de victime dans l'enfance et celui d'agresseur à l'adolescence.
La quasi-totalité des parcours examinés débute par une expérience traumatisante de violence sexuelle subie.
• Traumatisme Précoce comme Racine du Comportement :
Un expert psychiatre souligne qu'un jeune homme consommateur de pédopornographie "a subi des violences sexuelles lorsqu'il était lui-même enfant et qui a marqué son cerveau de cette expérience".
Cette expérience, même "fugace", a été "extrêmement choquante" et a "traumatisé" son psychisme.
• Reproduction de Schémas Subis : Un homme, qui a agressé deux mineurs à 14 ans, avait été lui-même violé par son frère aîné dès l'âge de 8 ans.
Il explique que son frère lui a appris ces actes "sous forme de jeu".
Plus tard, son propre passage à l'acte visait à "assouvir la pulsion pour retrouver le bien-être" qu'il associait à la fin de la relation abusive avec son frère, percevant cette dynamique comme "la normalité".
• De la Victimisation à l'Agression au sein de la Fratrie : Un autre témoignage relate un homme abusé par son beau-père dès l'âge de 3 ans, forcé de tourner dans des films pornographiques et d'abuser de sa propre petite sœur.
Il exprime son incapacité à se pardonner, le beau-père lui ayant martelé "Tu vas me ressembler, tu vas me ressembler".
II. Facteurs Déclenchants et Mécanismes Psychologiques
Le passage à l'acte chez les mineurs s'inscrit dans un contexte de vulnérabilités personnelles et de dynamiques familiales complexes, catalysé par des mécanismes psychologiques spécifiques.
A. Traumatismes et Dysfonctionnements Familiaux
Les témoignages mettent en lumière des environnements familiaux profondément insécurisants qui constituent un terreau fertile pour le développement de comportements déviants.
• Rupture des Liens Affectifs : Une jeune femme explique son passage à l'acte sur son petit frère par le divorce de ses parents, son sentiment d'abandon par un père avec qui elle était "fusionnelle" et la dépression de sa mère.
Elle se sentait seule, sans "place", et nourrissait des "envies de mourir".
• Le Poids des Secrets de Famille : Dans un cas, le fils passe à l'acte alors qu'il est révélé plus tard que l'ex-compagnon de la mère était lui-même auteur de violences sexuelles sur d'autres enfants de la famille.
Le passage à l'acte du fils est interprété par une thérapeute comme une manière inconsciente de "faire exploser tout ça".
• La Négligence Parentale : Un agresseur exprime de la "colère envers mes parents qui n'ont rien vu", soulignant que le manque de surveillance et de protection est un facteur aggravant.
L'environnement familial est décrit comme "un système de toutes les sécurités mais aussi de tous les dangers".
B. Passages à l'Acte : Logiques et Motivations
Plusieurs logiques psychologiques distinctes semblent présider au passage à l'acte.
| Mécanisme | Description | Exemples et Citations | | --- | --- | --- | | La Dépendance | Le comportement devient une compulsion, une addiction qu'il faut nourrir pour apaiser une angoisse ou un manque. | Un consommateur de pédopornographie décrit son envie comme une "faim" ou une "soif" qu'il doit "nourrir \[...\] quotidiennement". Il note que "à force de consommer, on se satisfait plus de ce qu'on a en fait et on cherche toujours plus en fait plus de sensation". | | La Vengeance | L'agression est une forme d'agressivité déplacée, dirigée vers une cible de substitution pour punir une injustice perçue. | Une jeune femme ayant agressé son petit frère explique : "C'était de la haine, de l'injustice. Mon petit frère a pris ma place \[...\] C'était une vengeance avec le recul d'aujourd'hui. C'était pour le punir". | | La Répétition Traumatique | L'individu rejoue activement le trauma subi, passant du rôle de victime passive à celui d'agresseur actif, dans une tentative inconsciente de maîtriser l'expérience. | Un homme abusé par son beau-père explique qu'il a reproduit les actes car il a été "démoli psychologiquement". Un autre, abusé par son frère, n'avait "pas la conscience de faire mal" car pour lui, c'était la "normalité". | | Théorie du "Foutu pour Foutu" | L'enfant internalise un sentiment de dévalorisation profonde et agit de manière à confirmer cette mauvaise image de lui-même. | Un psychiatre explique ce mécanisme : "puisque je suis pas bon, autant que je sois vraiment pas bon. Et une fois que j'ai fait quelque chose \[...\] là je mérite qu'on ne s'occupe plus de moi". |
III. La Prise en Charge Thérapeutique : Un Levier Essentiel
Face à la gravité des actes, les experts et les témoignages insistent unanimement sur le rôle crucial du soin et de l'accompagnement psychologique pour prévenir la récidive et permettre la reconstruction.
A. Objectifs et Modalités du Soin
La prise en charge est un processus structuré et multifacette.
• Évaluation et Responsabilisation : La première étape consiste en une "évaluation" pour cerner "le trouble du jeune" et son niveau de "dangerosité".
Le but n'est pas de "minimiser ou le banaliser" l'acte, mais de mettre les jeunes "face à leur responsabilité".
• Approches Thérapeutiques : Le soin est principalement "psychothérapeutique", en groupe ou en individuel, et peut s'accompagner d'une "prise en charge médicamenteuse". L'objectif est de trouver la racine du mal pour "le soigner depuis la racine".
• Briser l'Isolement : Un psychiatre identifie le problème central comme "la question de la solitude et l'isolement". La thérapie offre un espace pour parler et ne plus "rester tout seul".
B. L'Efficacité du Suivi et la Prévention de la Récidive
Les experts se montrent optimistes quant aux perspectives d'évolution positive des mineurs pris en charge.
• Évolution Positive : Une mère témoigne que son fils, après sa condamnation et son suivi, a "littéralement changé", a "pris en maturité" et en "réflexion".
• Faible Taux de Récidive : Un professionnel affirme que "dans les faits, il y a peu de récidives".
Prendre en charge un jeune "tôt" permet une intervention "active, précoce" et de "l'orienter vers le mieux".
• Une Vision Humaniste : La prise en charge repose sur le postulat que "Ce ne sont pas des monstres, ce sont des jeunes qui commettent fait monstrueux".
Un psychiatre affirme : "je ne renoncerai jamais à accompagner un patient dans sa quête de ne plus recommencer".
• Protéger les Futures Victimes : L'argument est clairement posé : "mieux on va prendre en charge les auteurs, moins il y aura de victimes. Donc, on a tout intérêt à travailler ensemble."
IV. L'Impact sur l'Entourage Familial
Les familles des jeunes auteurs sont profondément et durablement affectées, portant un fardeau de douleur, de culpabilité et de questionnements.
• Les Parents, "Victimes Collatérales" : Une mère décrit l'onde de choc de la révélation : "c'est tout votre monde s'écroule".
Les émotions ressenties incluent "colère, de honte, de culpabilité, de jugement". Beaucoup se demandent "qu'est-ce qu'elles ont fait pour en arriver là".
La culpabilité est omniprésente : "si j'avais fait ce qu'il fallait, peut-être ça serait jamais arrivé".
• La Gestion du Secret : La question de la parole est centrale. Un couple en thérapie s'interroge sur le moment et la manière d'expliquer le passé du père (victime et auteur) à leurs filles.
L'enjeu est de transmettre la vérité sans créer de jugement ou de confusion, reconnaissant que "quand un enfant pose une question, il faut lui répondre".
• La Reconstruction Familiale : Malgré le tsunami, la thérapie familiale aide à "analyser la situation et à voir les choses autrement".
Une mère explique comment, après la révélation des secrets, la famille "reconstruit" et que ses "enfants se parlent".
V. Enjeux Sociétaux et Perspectives
Les témoignages convergent vers un constat alarmant sur l'état des ressources allouées à la jeunesse et un appel pressant à l'action.
• Manque de Moyens : Un constat est dressé sur le "manque criant de moyens à la disposition de leur santé mentale, physique et sexuelle" des enfants.
• Urgence de la Prévention : La prévention est jugée "indispensable" mais "pas encore assez présente" dans les mentalités.
Son développement est considéré comme "bénéfique pour tous les mineurs".
• Un Appel à l'Action : La conclusion est sans appel : "La situation est très urgente".
Il est impératif que la société dans son ensemble gagne à "prendre soin de ses enfants même quand ils ont commis des faits aussi graves".
-
-
www.france.tv www.france.tv
-
Synthèse du débat sur la santé mentale et la schizophrénie
Résumé Exécutif
Ce document synthétise les échanges d'un débat entre experts, soignants, représentants d'associations et une personne concernée, centré sur la schizophrénie et les troubles psychiques en France.
Les discussions mettent en lumière plusieurs constats critiques : la stigmatisation profondément ancrée autour des maladies psychiques sévères, qui engendre honte et isolement, reste un obstacle majeur.
Le parcours de soin est un véritable "labyrinthe", caractérisé par un délai de prise en charge moyen d'un an et demi après les premiers symptômes et des diagnostics initiaux souvent complexes et incertains.
Une révolution s'opère cependant dans l'approche des soins, passant d'un modèle paternaliste à une vision centrée sur le "rétablissement" de la personne, où le patient devient l'acteur principal de son projet de vie, soutenu par sa famille qui est désormais considérée comme un partenaire essentiel.
Enfin, le débat souligne les faiblesses structurelles du système psychiatrique français, marqué par un manque de moyens endémique, de fortes inégalités territoriales d'accès aux soins et un besoin crucial de décloisonnement entre les différents acteurs sanitaires, sociaux et professionnels.
L'espoir, porté par les pairs-aidants et une approche plus humaine, émerge comme le moteur fondamental de la reconstruction des parcours de vie.
--------------------------------------------------------------------------------
Introduction
Suite à la diffusion du documentaire "Nous, schizophrènes et combattants", un débat a réuni plusieurs acteurs clés du domaine de la santé mentale pour discuter des défis liés à la schizophrénie et aux troubles psychiques.
Les intervenants ont partagé leurs expertises et expériences sur la détection, la prise en charge et les perspectives de rétablissement, tout en abordant l'état du système psychiatrique en France.
Participants au débat :
• Léopole de Pomier : Patient et pair-aidant à l'association Père et danse Montpellier.
• Alexandre de Conor : Docteur en psychologie, responsable du centre de rétablissement et de réhabilitation Jean Mainviel à Montpellier.
• Étienne Verry : Praticien hospitalier au CHU de Toulouse et au centre expert schizophrénie.
• Michel Comte : Délégué régional de l'Unafam Occitanie (Union nationale de familles et amis de personnes malades et/ou handicapées psychiques).
--------------------------------------------------------------------------------
Thème 1 : La Stigmatisation et le Tabou Persistants
La perception sociale des troubles psychiques, et en particulier de la schizophrénie, reste un obstacle majeur au diagnostic, à la prise en charge et au rétablissement.
• Persistance des clichés : La schizophrénie est encore largement et à tort associée à la "double personnalité" dans les médias et l'imaginaire collectif.
Alexandre de Conor souligne que "c'est encore un mot qui est utilisé pour désigner un changement de comportement brusque".
• Honte et auto-stigmatisation : Les personnes concernées intériorisent cette image négative. Julien, dans le documentaire, se dit "rouge de honte", ce à quoi sa mère répond : "Un diabétique n'a pas honte d'être malade".
Cette stigmatisation a des conséquences concrètes : isolement, difficultés d'accès aux relations amicales, amoureuses, au logement.
• Vulnérabilité accrue : Contrairement à l'image du "fou dangereux", les chiffres montrent que les personnes souffrant de schizophrénie sont plus souvent victimes de personnes malveillantes que auteurs de violences. Alexandre de Conor insiste : "les chiffres nous montrent que c'est des gens qui sont plus victimes".
• Barrières culturelles et religieuses : Léopole de Pomier témoigne de l'influence de certaines croyances qui associent la maladie à une forme de possession : "on pense que c'est des esprits qui nous habitent et tout. Ça, je trouve que c'est une vraie bêtise".
Thème 2 : Le Parcours du Dépistage et du Diagnostic
L'accès aux soins est un processus long et complexe, souvent qualifié de "parcours du combattant" pour les personnes et leurs familles.
• Délai de prise en charge : En France, comme en Europe, la durée moyenne entre l'apparition des premiers symptômes psychotiques et la première prise en charge spécialisée est d'environ un an et demi.
Ce délai est un enjeu majeur sur lequel il est possible d'agir.
• Difficultés du repérage précoce : Les premiers signes apparaissent souvent à l'adolescence et peuvent être confondus avec une "crise d'adolescence un peu violente", retardant ainsi le début d'un accompagnement adapté.
• Un système de soins labyrinthique : Alexandre de Conor décrit le système comme "un labyrinthe en fait entre la psychiatrie, le médico-social".
Cette complexité et le manque de communication entre les structures peuvent décourager les familles et faire perdre un temps précieux.
• La prudence du diagnostic :
◦ Étienne Verry explique qu'un diagnostic posé trop rapidement lors d'un premier épisode psychotique se révèle erroné dans un cas sur deux à cinq ans.
La prudence est donc de mise, en parlant initialement de "premier épisode psychotique" plutôt que de schizophrénie.
◦ Alexandre de Conor ajoute que des diagnostics hâtifs peuvent être dangereux, notamment pour les minorités ethniques ou les populations défavorisées, et mener à des propositions de soins inadaptées (ex: prescription de neuroleptiques non justifiée).
Thème 3 : La Prise en Charge et le Concept de Rétablissement
Une transformation profonde de l'approche des soins est en cours, axée sur la personne, son projet de vie et son rétablissement.
• L'hospitalisation sous contrainte : Souvent redoutée, elle est décrite par la mère de Julien comme "la pire décision de ma vie".
Michel Comte la qualifie d'expérience "violente" et "terrible" pour une famille. Les intervenants s'accordent sur la nécessité de l'éviter au maximum grâce à une intervention précoce et des équipes mobiles, tout en reconnaissant sa nécessité dans certaines situations de perte de contact avec la réalité.
• Du soin à la guérison, au rétablissement : La schizophrénie étant une maladie chronique, on ne parle pas de guérison mais de "rétablissement".
Il s'agit de "vivre une vie satisfaisante et qui a du sens pour la personne malgré cette présence de la maladie" (Alexandre de Conor).
• L'approche centrée sur la personne :
◦ Cette approche constitue une "petite révolution" face à une médecine historiquement "patriarcale et très autoritaire" (Étienne Verry).
◦ Elle implique de ne plus parler des personnes sans leur présence et de partir de leurs projets (emploi, logement) pour proposer un accompagnement.
◦ Exemple concret : Si un jeune veut reprendre un emploi mais est gêné par des voix, on lui propose une aide pour gérer ce symptôme au service de son objectif professionnel.
◦ Cela implique d'accepter une prise de risque accompagnée, par exemple en soutenant un jeune qui souhaite essayer d'arrêter son traitement, car "s'il a décidé de le faire, si on lui dit non, il le fera sans nous" (Alexandre de Conor).
Thème 4 : Le Rôle Central des Familles et des Proches
La famille, longtemps tenue à l'écart ou culpabilisée par la psychiatrie, est aujourd'hui reconnue comme un pilier du rétablissement.
• De la culpabilité à la solution : Michel Comte décrit le parcours des familles, qui passent souvent par le déni, la sidération et un sentiment de culpabilité ("Qu'est-ce que j'ai loupé ?") avant de comprendre qu'elles ne sont "pas le problème" mais "la solution".
• L'impact du soutien familial : Les chiffres confirment que lorsque les familles comprennent la maladie et sont soutenantes, "clairement ça a un impact très fort sur le rétablissement".
• L'intégration des familles aux soins : Des programmes de psychoéducation, comme le programme BREF, sont mis en place pour intégrer les familles, les personnes concernées et les soignants dès le début de la maladie.
• Soutien par les pairs-aidants familiaux : Des associations comme l'Unafam, composées de proches et d'aidants, offrent un espace d'écoute, de partage et de "reconstruction" pour les familles qui se sentent "en miettes".
Thème 5 : Les Défis du Système Psychiatrique en France
Le débat a mis en exergue les failles structurelles du système de santé mentale français, qui freinent l'accès à des soins de qualité pour tous.
• Manque de moyens et de personnel : Le secteur souffre d'un "manque de moyen endémique". Une grande proportion des postes de psychiatres dans les établissements publics ne sont pas pourvus.
• Inégalités territoriales : L'accès aux soins est très inégal. Des départements comme le Gers ou l'Ariège comptent moins de 6 psychiatres pour 100 000 habitants.
Les Centres Médico-Psychologiques (CMP), essentiels dans la psychiatrie de secteur, connaissent des dysfonctionnements majeurs dans certains territoires, avec des délais de rendez-vous de plusieurs mois.
• Nécessité de décloisonnement : Il y a un manque critique de coordination entre la psychiatrie, les autres spécialités médicales (pour les maladies somatiques comme l'obésité liée aux traitements) et les secteurs social et professionnel. La fonction de "case manager" est présentée comme un outil pour orchestrer le parcours du jeune.
• Nouveaux modèles à développer : L'importance de l'approche "aller vers", qui consiste à se déplacer pour rencontrer les jeunes là où ils sont, est soulignée.
Ce modèle est efficace mais coûteux car il requiert d'importantes ressources humaines.
Thème 6 : L'Espoir, la Pair-Aidance et la Force du Collectif
Malgré les difficultés, un message d'espoir traverse l'ensemble du débat, incarné par de nouvelles approches et la valorisation de l'expérience vécue.
• Le pouvoir de l'espoir : Il est crucial de lutter contre la représentation fataliste de la schizophrénie. Étienne Verry rappelle que 20% des patients connaissent un rétablissement complet, parfois même sans traitement, et que le pronostic est donc extrêmement variable. La maladie ne doit pas "tuer l'ambition, le rêve".
• La pair-aidance : Le rôle de Léopole de Pomier est emblématique.
En tant que pair-aidant, il utilise son propre parcours pour aider les autres : "Si j'ai le contrôle sur ma maladie, je peux aider les autres". Cette transmission d'expérience donne de l'espoir et des outils concrets.
• La force du collectif : Les groupes de parole, pour les personnes concernées comme pour les proches, sont des lieux d'enrichissement mutuel où les expériences et les stratégies sont partagées.
• Le combat pour l'inclusion : Le mot "combattant" du titre du documentaire est validé par tous les participants.
C'est un combat pour le rétablissement personnel, mais aussi un combat sociétal pour "abattre les stéréotypes" et pour l'inclusion.
Comme le conclut Étienne Verry, améliorer le pronostic des jeunes est "une affaire de tous. C'est pas juste une affaire de la santé".
-
-
www.france.tv www.france.tv
-
Dossier d'Information : Le Groupe de Protection des Mineurs
Résumé Exécutif
Ce document de synthèse analyse le fonctionnement, les défis et les succès d'un Groupe de protection des mineurs (GPM) au sein de la gendarmerie, basé sur des extraits d'enregistrements.
Il met en lumière une problématique de violences sur mineurs d'une ampleur alarmante sur le territoire du Plateau Picard, avec une fréquence d'un viol tous les six jours et trois nouveaux dossiers par semaine, un constat qui a motivé la création de cette unité spécialisée.
Le groupe a été formé pour rationaliser et professionnaliser le traitement de ces affaires complexes et urgentes, souvent mal gérées par des militaires non formés.
Les résultats sont probants, avec une réduction drastique des délais de traitement des procédures de plusieurs mois à seulement trois ou quatre mois.
La méthodologie du groupe repose sur deux piliers : une prise en charge immédiate et bienveillante des victimes pour éviter les rétractations, et l'application d'un protocole d'audition judiciaire rigoureux pour les enfants, visant à recueillir un témoignage précis et non-induit, essentiel pour caractériser les infractions.
Parallèlement, les interrogatoires des mis en cause (gardes à vue) sont menés avec une stratégie mêlant pression psychologique, analyse de preuves numériques et recherche de rapport humain pour obtenir des aveux.
Le travail des enquêteurs est psychologiquement éprouvant, les confrontant à des "actes de barbarie" et à une misère humaine intense.
Pour y faire face, ils développent une "carapace" et s'appuient sur une cohésion de groupe exceptionnelle, décrite comme une relation amicale fondamentale à leur équilibre et à l'efficacité de l'unité.
Cette solidarité, combinée à une forte conscience de leur mission de protection de l'enfance, constitue le moteur de leur engagement.
--------------------------------------------------------------------------------
I. Contexte et Création du Groupe
A. Un Constat Alarmant sur le Plateau Picard
À son arrivée, un responsable a rapidement constaté que la priorité de la lutte contre la délinquance (cambriolages, stupéfiants) masquait un problème plus profond et plus grave : l'ampleur des violences, en particulier celles commises sur les mineurs.
• Volume des affaires : La compagnie enregistrait environ trois nouveaux dossiers par semaine concernant des violences sur mineurs.
• Gravité des faits : Les statistiques révélaient un viol tous les six jours sur le territoire de la compagnie, une fréquence jugée effrayante.
• Lieux des violences : Ces actes se produisaient dans divers contextes : le milieu familial, les foyers et les écoles.
B. La Nécessité d'une Approche Spécialisée
Avant la création du groupe, le traitement de ces affaires présentait de graves lacunes :
• Manque de formation : Certains militaires ne se sentaient pas capables de traiter ces dossiers sensibles.
• Prise en charge inadaptée : Les victimes n'étaient pas prises en charge correctement, ce qui compromettait le recueil de la parole.
• Lenteur des procédures : Les dossiers "traînaient sur les bureaux", accumulant des mois de retard, ce qui nuisait à l'efficacité de la réponse pénale.
Face à ce constat, la décision a été prise de "rationaliser ce travail" en créant un groupe d'enquêteurs spécialisés et dédiés, afin de concentrer les compétences et de traiter ces affaires complexes en urgence.
C. Résultats et Efficacité
Malgré un scepticisme initial ("Au mois de septembre, le groupe il est mort, il existera pas"), le groupe a rapidement prouvé son efficacité :
• Réduction des délais : Le temps de traitement des procédures a été réduit à une moyenne de trois à quatre mois, contre des retards de plusieurs mois auparavant.
• Professionnalisation : L'unité permet une gestion centralisée et experte des dossiers, garantissant que les enquêtes sont menées par du personnel formé.
II. Le Processus d'Enquête : De la Victime à l'Auteur
A. La Prise en Charge Immédiate des Victimes
L'un des principes cardinaux du groupe est l'intervention rapide auprès des victimes.
• L'urgence de l'écoute : "Le plus important, c'est la prise en compte immédiate des victimes. [...] si on attend des fois peut-être deux jours, c'est suffisant pour qu'elle se rétracte parce que ils vont penser aux conséquences."
• Le rôle proactif : Les enquêteurs se déplacent immédiatement pour rassurer les victimes et leur signifier leur soutien ("maintenant on est là, on va vous aider").
B. L'Audition des Mineurs : Un Exercice Délicat
L'audition d'un enfant victime est une étape cruciale et encadrée par un protocole strict.
• Objectif judiciaire : L'audition vise à recueillir des éléments permettant de "caractériser l'infraction" et d'éviter un "classement sans suite".
Le témoignage de l'enfant est souvent la pièce maîtresse du dossier.
• Le protocole comme guide : Un protocole d'audition filmée est utilisé pour rassurer l'enquêteur et l'enfant.
Il fournit des outils pour ramener l'enfant sur le sujet de la discussion et instaure un climat de confiance.
• Le danger de la suggestion : La principale difficulté est de ne jamais induire les réponses. "Il faut surtout pas suggérer à l'enfant quand on fait les auditions quoi que ce soit en fait."
Des erreurs, comme des questions fermées ou suggestives, peuvent être "préjudiciables" à la procédure.
• La gestion du stress : Le stress de l'enquêteur peut le faire "perdre pied". Il est conseillé de faire des pauses pour se recentrer.
C. La Garde à Vue et l'Interrogatoire des Auteurs
Les enregistrements détaillent une garde à vue de 48 heures, illustrant la stratégie des enquêteurs.
| Étape | Description | | --- | --- | | Interpellation | L'opération est menée à 6h du matin pour créer un effet de surprise. Le dispositif est sécurisé pour parer à un risque de fuite, même si l'individu n'est pas connu comme violent. | | Perquisition | Une fouille minutieuse du domicile est effectuée, avec saisie de tout le matériel informatique et multimédia ("Tout nous intéresse clairement"). | | Exploitation Numérique | Les enquêteurs s'appuient sur la fiabilité de la "preuve numérique" (journaux d'appels, localisation, etc.), car "on laisse toujours une trace". Un logiciel spécialisé extrait toutes les données des appareils saisis. | | Auditions | Les auditions progressent d'un "CV" général à l'abord des faits. La stratégie consiste à alterner des moments de tension et de relâchement ("C'est important dans une garde à vue qu'il y a des moments \[...\] pour souffler, pour réfléchir"). L'enquêteur confronte le suspect à ses contradictions ("vous êtes en train de vous tirer une balle dans le pied comme un con") pour briser le déni. | | Les Aveux | Les aveux sont obtenus progressivement. Le suspect passe de la négation à la reconnaissance de consultations de sites, puis à des aveux partiels sur des attouchements, et enfin à une description précise des faits, incluant une tentative de pénétration. L'enquêteur doit "mouliner" dès que le suspect "ferre le poisson" et est "prêt à parler". | | Gestion Juridique | Le procureur est tenu informé en permanence et autorise la prolongation de la garde à vue pour mener toutes les investigations nécessaires. | | Défèrement | À l'issue des 48 heures, le suspect est déféré devant le substitut du procureur, qui lui notifie les faits reprochés et saisit le juge en vue d'une détention provisoire. |
III. La Dimension Humaine et Psychologique du Travail d'Enquêteur
A. Le Poids Émotionnel des Dossiers
Le quotidien des enquêteurs est marqué par l'exposition à des faits d'une violence extrême.
• L'horreur des récits : "La plupart de la population pourrait pas entendre ce que nous on entend. Il y a des choses horribles. On peut aller jusqu'à des actes de barbarie."
• La nécessité d'une "carapace" : Pour se protéger, les enquêteurs doivent se détacher émotionnellement. "On se crée une carapace, on est obligé. [...] tu rentres chez toi, il faut que tu penses à autre chose."
L'un des gendarmes trouve son équilibre dans le jardinage, une activité qui "soigne le corps" et "l'esprit".
• L'impact persistant : Malgré cette protection, le travail les poursuit parfois. "Ça m'arrive hein des fois le soir d'aller me coucher, de penser aux procédures."
B. La Motivation et le Sens de la Mission
La principale motivation des membres du groupe est un sentiment d'utilité et la conviction de mener une mission essentielle.
• Protéger les enfants : L'objectif premier est clair : "Mon but c'est ça en fait, protéger les enfants."
La "première victoire, c'est quand l'enfant arrive à dire les choses et que on voit à la fin qui est ce soulagement chez l'enfant."
• Un travail sur le long terme : Les enquêteurs ont conscience de l'impact durable de leur action. "Je fais un travail dans le temps, je fais pas un travail dans l'immédiat. Je travaille sur des dizaines d'années. [...] on va le construire en tant qu'adulte."
• Un lien avec les victimes : Un contact direct et un "affect" se développent avec les familles, qui ont leurs numéros de téléphone et peuvent appeler même en dehors des heures de service.
C. L'Importance Cruciale de la Cohésion de Groupe
La solidarité au sein de l'équipe est présentée comme la clé de voûte de leur résilience et de leur succès.
• Plus que des collègues : "On n'est pas que des collègues, on est des amis quoi.
Maintenant, c'est c'est magique quoi."
Ils se décrivent comme "quatre enquêteurs mais aussi quatre copains."
• Un soutien mutuel indispensable : "Si on avait pas ce groupe là, ça serait beaucoup plus compliqué de passer autre chose le soir quand tu rentres chez toi."
Le groupe se relance mutuellement en cas de "fatigue mentale".
• Condition de fonctionnement : "Je pense que s'il y avait pas cette cohésion entre nous quatre, le groupe ne pourrait pas fonctionner comme il fonctionne actuellement."
IV. Citations Clés
Sur la Mission et son Impact
"Le plus important, c'est la prise en compte immédiate des victimes. [...] si on attend [...] c'est suffisant pour qu'elle se rétracte."
"Redonner le sourire à un enfant, l'aider à se reconstruire, c'est ça ça te nourrit en fait."
"Je fais un travail dans le temps, je fais pas un travail dans l'immédiat. Je travaille sur des dizaines d'années."
Sur la Difficulté et la Réalité du Métier
"La plupart de la population pourrait pas entendre ce que nous on entend. Il y a des choses horribles. On peut aller jusqu'à des actes de barbarie."
"On se crée une carapace, on est obligé. [...] tu rentres chez toi, il faut que tu penses à autre chose."
"Il faut surtout pas suggérer à l'enfant quand on fait les auditions quoi que ce soit en fait."
Sur l'Importance du Groupe
"Si on avait pas ce groupe là, ça serait beaucoup plus compliqué de passer autre chose le soir quand tu rentres chez toi."
"On s'appelle groupe d'atteinte aux personnes mais c'est vraiment ça, on est un groupe de quatre enquêteurs mais aussi de quatre quatre copains maintenant."
"Il faut que des groupes comme les nôtres se créent partout en France. C'est super important la prise en charge de la victime."
-
-
www.france.tv www.france.tv
-
Synthèse sur la Prostitution des Mineurs au Sein de l'Aide Sociale à l'Enfance
Résumé Exécutif
Ce document de synthèse analyse une enquête approfondie sur le phénomène systémique de la prostitution des mineurs placés sous la protection de l'Aide Sociale à l'Enfance (ASE) en France.
L'enquête révèle une faille profonde dans le système censé protéger les enfants les plus vulnérables.
Le constat est accablant : une commission d'enquête parlementaire a établi que sur les 20 000 mineurs prostitués en France, 80 % sont issus des dispositifs de l'ASE.
Les foyers de l'ASE, loin d'être des sanctuaires, sont décrits comme des "zones de non-droit" où les jeunes, souvent en fugue, deviennent des proies faciles pour les réseaux d'exploitation sexuelle.
Le phénomène est alimenté par plusieurs facteurs : la précarité matérielle des jeunes (parfois 20€ d'argent de poche par mois), l'impuissance ou l'inaction des équipes éducatives face aux fugues, et des protocoles administratifs qui permettent à l'institution de se déresponsabiliser.
Des témoignages poignants de jeunes filles, dont certaines ont été exploitées dès l'âge de 11 ans, illustrent des parcours de traumatismes répétés, incluant séquestration, violences et proxénétisme au sein même des foyers.
L'enquête met également en lumière l'impunité relative des clients, comme le démontre un procès à Albi où 18 hommes ont été condamnés à de simples amendes pour avoir eu des relations avec une jeune fille de 15 ans.
Face à ce scandale, les responsables politiques des départements, qui ont la tutelle de l'ASE, tendent à minimiser leur responsabilité, invoquant une problématique de société qui les dépasse.
Face à ce tableau sombre, des familles se mobilisent et engagent des poursuites judiciaires contre les départements pour "faute lourde".
En parallèle, une initiative unique, la Maison Gaia près de Lille, démontre qu'une prise en charge spécialisée, sécurisée et bienveillante peut offrir une voie de reconstruction à ces jeunes victimes. Cependant, cette structure reste une exception dans un système en crise profonde.
Analyse Détaillée des Thèmes Centraux
Un Scandale Systémique : L'Échec de l'Aide Sociale à l'Enfance (ASE)
Ampleur et Nature du Phénomène
Le reportage expose une réalité alarmante qualifiée de "pandémie" et de "raz de marée" par les acteurs du secteur.
La prostitution des mineurs placés n'est pas un fait isolé mais un problème structurel et national, touchant des villes comme Paris, Marseille et Albi.
• Statistiques Choc : Une commission d'enquête parlementaire d'avril 2025 révèle que 80 % des 20 000 mineurs prostitués en France sont placés sous la responsabilité de l'ASE.
• Explosion du Phénomène : Le nombre de cas a été multiplié par 10 en 10 ans.
• Des Foyers Ciblés : Les proxénètes connaissent les adresses des foyers et les ciblent délibérément, attendant à l'extérieur que les jeunes sortent ou fuguent.
Une éducatrice à Marseille confirme : "Vous venez 21h, 22h, il y a plein de proxos devant les portes."
Défaillances Structurelles et Institutionnelles
Les foyers, censés être des lieux de protection, sont décrits comme des environnements favorisant l'exploitation.
• Une "Zone de Non-Droit" : Une mère de famille qualifie l'ASE de "zone de non-droit où les jeunes sont livrés à eux-mêmes".
Les éducateurs ne peuvent légalement retenir les adolescents qui souhaitent sortir, facilitant ainsi les fugues et l'emprise des réseaux.
• Précarité Matérielle : Aïana, une ancienne pensionnaire, témoigne avoir reçu 20 € d'argent de poche par mois, la poussant à se prostituer à 13 ans pour acheter des produits de première nécessité (produits d'hygiène, vêtements, nourriture).
• Inaction des Équipes : Si certains éducateurs sont désabusés, d'autres semblent faire preuve d'une passivité coupable.
Une éducatrice aurait dit à une jeune fille se prostituant : "Je sais que tu as des clients ce soir mais rentre pas tard quand même."
• Déresponsabilisation Administrative : L'enquête menée en caméra cachée révèle un protocole bien rodé : lorsqu'une jeune fugue, les éducateurs remplissent une "déclaration de fugue" transmise à la brigade des mineurs.
À partir de là, l'ASE n'est "plus tenue pour responsable du sort de ces jeunes filles".
• Placements Inadaptés : Le cas d'Alice, placée à 16 ans dans un hôtel meublé pendant 6 mois (alors que la loi l'interdit au-delà de 2 mois), montre un placement dans un environnement non sécurisé, au milieu d'adultes et de résidents de passage, où la prostitution a également lieu.
Les Victimes : Parcours de Traumatismes Répétés
Les témoignages des jeunes filles constituent le cœur de l'enquête, révélant la profondeur de la souffrance et la faillite du système.
• Témoignage de la mère d'une fille de 14 ans (Paris) : Après une fugue et des accusations de violence parentale non vérifiées, sa fille est placée.
Les parents découvrent via un détective privé (coût : 35 000 €) qu'elle sort la nuit pour se prostituer. La juge maintient le placement malgré les preuves.
• Aïana (18 ans) : A commencé à se prostituer à 13 ans dans son foyer à Paris par nécessité.
Elle affirme que les éducateurs étaient au courant mais n'offraient aucune alternative.
• Lila (16 ans, Marseille) : Victime d'exploitation dès 11 ans. Placée pour fuir des violences paternelles, elle est forcée de se prostituer par une autre jeune fille du foyer.
Elle passera par 15 foyers différents, se prostituant dans plusieurs d'entre eux.
Elle a contracté des MST (Chlamydia) sans jamais voir un médecin.
• Alice (Essonne) : Placée depuis 6 ans, elle est kidnappée à 14 ans devant son foyer, séquestrée à Toulon et forcée à se prostituer.
Elle affirme avoir alerté ses éducateurs en vain. Après avoir été libérée par la police, elle est replacée dans un autre foyer gangréné par la prostitution, puis dans un hôtel.
Les Auteurs de l'Exploitation : Proxénètes et Clients
L'enquête identifie clairement les deux maillons de la chaîne d'exploitation.
• Le Rôle des Proxénètes :
◦ Proxénétisme Interne : Un schéma récurrent est celui où les pensionnaires plus âgées ("les grandes") deviennent les proxénètes des plus jeunes.
◦ Réseaux Structurés : Les proxénètes externes sont décrits comme "beaucoup plus structurés, beaucoup plus dangereux" et utilisent des méthodes de manipulation et de menace pour contrôler les jeunes filles et leur famille.
◦ Exploitation Financière : Les jeunes filles ne tirent quasiment aucun profit de leur exploitation.
Un éducateur explique le mécanisme : "Finalement tiens, repart avec 20 €."
• Le Profil et l'Impunité des Clients :
◦ Des Profils Variés : Lila témoigne avoir eu des clients de tous âges et de toutes professions : "des policiers comme des pompiers, des ambulanciers comme des avocats".
◦ Le Cas du Tribunal d'Albi : Un procès emblématique a eu lieu en septembre 2025. 18 hommes, clients d'une jeune fille de 15 ans placée en foyer et séquestrée, comparaissaient. Ils ont tous plaidé l'ignorance de sa minorité, bien qu'elle "paraissait mineure".
La justice les a crus, les jugeant devant un tribunal de police et les condamnant à des amendes de 500 à 700 €.
Les Réponses : Déni, Mobilisation et Solutions Alternatives
La Réponse Insuffisante des Autorités
Les responsables politiques et institutionnels, directement mis en cause, peinent à assumer leurs responsabilités.
• Présidents de Départements :
◦ Christophe Ramon (Tarn, PS) : Répond par écrit en parlant de "fléau national" et de "responsabilité collective et partagée".
◦ François Durovray (Essonne, LR) : Affirme que ses services ont "correctement fait leur travail" et que la responsabilité incombe aux "voyous", à la police et à la justice.
Il nie tout problème de moyens financiers (budget de 245 millions d'euros pour l'ASE en Essonne en 2024).
◦ Martine Vassal (Bouches-du-Rhône, LR) : Refuse de commenter l'affaire judiciaire en cours, soulignant la difficulté de l'accompagnement de ces jeunes.
• Le Gouvernement : La ministre de la Santé, Stéphanie Rist, a annulé une interview prévue en réaction au reportage après que son équipe l'ait visionné.
Une Initiative Exemplaire : La Maison Gaia
En contraste avec l'échec généralisé, une structure se distingue.
• Un Refuge Spécialisé : Près de Lille, la Maison Gaia est un foyer unique en France, à l'emplacement secret, qui accueille 10 jeunes filles victimes de prostitution.
• Une Prise en Charge Holistique : L'approche est centrée sur la reconstruction de l'estime de soi (ateliers, repas partagés, soins esthétiques) et la reconnexion à leur corps.
L'objectif est de ne jamais couper le lien, même en cas de fugue, pour permettre un retour.
La Mobilisation des Familles
Face à l'inertie du système, les familles des victimes s'organisent pour obtenir justice.
• Procédures Judiciaires : Les mères d'Alice et de Lila ont décidé d'assigner en justice les départements de l'Essonne et des Bouches-du-Rhône pour "faute lourde".
• Action Collective : Maître Michel Hamas représente 49 familles dans des procédures similaires contre sept départements.
Données Clés et Citations Marquantes
| Donnée | Source / Contexte | | --- | --- | | 80% | Pourcentage des mineurs prostitués en France issus de l'ASE. (Commission d'enquête parlementaire) | | 20 000 | Nombre total estimé de mineurs prostitués en France. | | 10 fois | Multiplication du phénomène de prostitution des mineurs en 10 ans. | | 11 ans | Âge de Lila lors de sa première exploitation sexuelle. | | 20 € / mois | Argent de poche d'Aïana au foyer, la poussant à se prostituer. | | 35 000 € | Somme dépensée par une famille pour un détective privé afin de prouver la prostitution de leur fille. | | 500 - 700 € | Montant des amendes infligées à 18 clients d'une mineure de 15 ans à Albi. | | 245 M€ | Budget de l'ASE pour le département de l'Essonne en 2024. | | 66 M€ | Budget de l'ASE pour le département du Tarn en 2024. |
--------------------------------------------------------------------------------
Citations Marquantes :
• Une mère de famille : "L'ASE, c'est une zone de non-droit en fait. C'est une zone où les jeunes sont livrés à eux-mêmes."
• Une juge des enfants à Marseille : "Le tribunal de Marseille, comme beaucoup de tribunaux en France, connaît une explosion de ce phénomène. (...) Nous sommes devant un raz de marée."
• Alice, victime : "La loi, c'est de la merde. Sortez tous les jeunes de là-bas. (...) Faut arrêter de faire semblant."
• Une éducatrice à Marseille : "Les clients se garent devant, l'éducateur le savait. Ma copine quand elle le faisait, l'éducatrice elle disait 'bon, je sais que tu as des clients ce soir mais rentre pas tard quand même'."
• François Durovray, Président du département de l'Essonne : "Les services du département ont correctement fait leur travail, même si effectivement il y a une situation d'exploitation sexuelle."
• Lila, victime, à propos de l'ASE : "Ils doivent répondre parce que, en soit, ils ont tué nos vies un peu."
-
-
pmc.ncbi.nlm.nih.gov pmc.ncbi.nlm.nih.gov
-
218798
DOI: 10.1002/biot.70179
Resource: None
Curator: @olekpark
SciCrunch record: RRID:Addgene_218798
-
196690
DOI: 10.1002/biot.70179
Resource: None
Curator: @olekpark
SciCrunch record: RRID:Addgene_196690
-
-
human.libretexts.org human.libretexts.org
-
La fuente es demasiado grande en tamaño 16. La fuente es poco convencional. En la información del ensayo en la parte superior izquierda, se omite el título de la clase. La numeración de páginas en la parte superior derecha tiene el apellido del profesor, no el del estudiante. El título “Revisión del ensayo n.º 1” es genérico, no creativo y no despierta el interés del lector. Cuando se presenta la novela, también es necesario mencionar el autor. El título de la novela está entre comillas cuando debería estar en cursiva. El personaje principal se conoce con diferentes apodos informales que no se usan en el texto, y uno está mal escrito. Siempre sea preciso y coherente con los nombres de los personajes y del autor, y asegúrese de que estén bien escritos. Hay un espacio extra después del primer párrafo y el segundo párrafo necesita sangría. Los márgenes del lado izquierdo no miden una pulgada.
In the first page of the essay, there are several MLA formatting errors. The header “Lachmayr 1” is incorrectly placed on the left instead of being right-aligned and ½ inch from the top margin, and it uses the professor’s last name instead of the student’s, which should read “Bell 1.” The date is written in the wrong format, as MLA requires “21 May 2013.” The title is not properly centered, is incorrectly capitalized, and uses improper styling, since MLA titles should be in plain text without bold, italics, or underlining. The abbreviation “n.°” in the title is not MLA style and should be written as “No. 1” or avoided. Additionally, the book title within the paragraph is incorrectly placed in quotation marks instead of italics, and there is an unnecessary space before the closing quotation mark and comma.
-
- Jan 2026
-
www.examtopics.com www.examtopics.com
-
NOTA: Cada selección correcta vale un punto.
¿Qué páginas visitadas por los usuarios se correlacionan con mayor frecuencia con una compra de producto?
¿Cómo afecta el tiempo de carga de la página de visualización del producto a la decisión del usuario de comprar un producto?
¿Qué eventos influyen más en la decisión de un usuario de seguir usando la aplicación?
¿Existen lugares en la aplicación donde los usuarios suelen realizar acciones repetitivas con frecuencia?
-
-
Local file Local file
-
Este articulo surge en el contexto de mis estudios de doctorado en Comunicación, Lenguaje e Información en la Pontificia Universidad Javeriana. Trata sobre el tema del racismo en Colombia y como éste asume muchas formas; desde el mas cotidiano y vulgar, hasta el mas sutil. Fue elaborado para la asignatura de Teorías Contemporáneas de corte decolonial.
-
-
www.elcolombiano.com www.elcolombiano.com
-
: los nietos están involucrados
Considero que es un una creencia muy arraigada en la cultura tener la posibilidad de criar nietos, compartir con ellos y que trasmiten mucha alegría, sobre todo para esa etapa de la vida donde los adultos mayores están mas solos. La pregunta que se puede dejar planteada es Considerando la tendencia de la baja natalidad ¿Como sería el involucramiento de los nietos? o ¿Como mantener esa agudeza mental o en qué otro mecanismo pensar para mantener esa agudeza mental? como el mejor entorno para asegurar una calidad de vida y la longevidad.
-
-
www.examtopics.com www.examtopics.com
-
Para cada una de las siguientes afirmaciones, seleccione Sí si la afirmación es verdadera. De lo contrario, seleccione No.
1.- NO. 2.- NO. 3.- SI. 4:- NO.
-
NOTA: Cada respuesta correcta vale un punto.
A los usuarios no autenticados se les solicita autenticarse con GitHub.
Un archivo inexistente en la carpeta /images/ generará un código de respuesta 404.
Las solicitudes HTTP GET de usuarios autenticados con el rol llamado registeredusers se envían a la carpeta API.
Los usuarios autenticados que no pertenecen al rol registeredusers y los usuarios no autenticados reciben un error HTTP 401 al acceder a la carpeta API.
-
-
www.examtopics.com www.examtopics.com
-
NOTA: Cada respuesta correcta vale un punto.
El rendimiento mínimo del contenedor es de 400 RU/s.
La primera instrucción de consulta es una consulta dentro de una sola partición (in-partition query).
La segunda instrucción de consulta es una consulta entre múltiples particiones (cross-partition query).
-
-
www.examtopics.com www.examtopics.com
-
NOTA: Cada respuesta correcta vale un punto.
Los blobs de tipo block con el prefijo container1/salesorders o container2/inventory que no hayan sido modificados en más de 60 días se mueven al almacenamiento cool.
Los blobs que no hayan sido modificados en 120 días se mueven a la capa de almacenamiento archive.
Los blobs se mueven al almacenamiento cool si no han sido accedidos durante 30 días.
Los blobs se moverán automáticamente de cool a hot si se acceden nuevamente después de haber sido movidos a cool.
Todos los blobs de tipo block con una antigüedad mayor a 730 días se eliminarán.
-
Seleccionar y colocar:
Generar una clave de intercambio de claves (KEK).
Recuperar la clave pública de la clave de intercambio de claves (KEK).
Generar un archivo blob de transferencia de claves usando la herramienta proporcionada por el proveedor del HSM.
Ejecutar el comando az keyvault key import.
-
-
www.youtube.com www.youtube.com
-
Rapport de Synthèse : Crise et Dérives du Secteur Périscolaire et de l’Enseignement Privé
Résumé Exécutif
Une enquête approfondie de l'émission « Cash Investigation » révèle des défaillances systémiques graves au sein du secteur périscolaire et de certains établissements d'enseignement privé sous contrat en France.
Le constat est alarmant : violences physiques et verbales, suspicions d'agressions sexuelles, et manque de surveillance généralisé.
Les points clés à retenir :
• Ampleur nationale : À Paris, 128 animateurs ont été suspendus en 10 ans pour suspicion de violences sexuelles.
Le phénomène touche l'ensemble du territoire, des grandes métropoles aux zones rurales.
• Failles de recrutement : Une infiltration de 27 heures a suffi pour constater des comportements inappropriés (cris, usage de téléphones, gestes déplacés) et un non-respect flagrant des quotas d'encadrement.
• Inertie institutionnelle : Des signalements d'abus restent parfois sans réponse pendant plusieurs années avant qu'une action concrète ne soit entreprise.
• Échec judiciaire : Seules 3 % des plaintes pour violences sexuelles sur mineurs aboutissent à une condamnation, tandis que 73 % sont classées sans suite.
• Manque de contrôle du privé : Les établissements privés sous contrat, financés à 75 % par l'État, échappent à une surveillance rigoureuse malgré des cas de maltraitance institutionnalisée.
--------------------------------------------------------------------------------
I. Analyse du Secteur Périscolaire : Un Système en Souffrance
L'enquête met en lumière une réalité quotidienne brutale dans les accueils périscolaires (cantines, garderies), souvent méconnue des parents qui supposent leurs enfants protégés par l'institution scolaire.
A. Des conditions d'exercice dégradées
L'infiltration d'une journaliste, embauchée sans expérience préalable malgré la possession du BAFA, révèle des dérives immédiates après seulement 27 heures d'observation :
• Comportements inappropriés : Animateurs focalisés sur leurs téléphones portables, hurlements constants contre les enfants, et gestes graves comme une animatrice embrassant des enfants de 4 ans sur la bouche.
• Sous-effectif chronique : Dans certains cas, le nombre d'enfants par animateur est plus du double du quota légal (par exemple, une seule personne pour 23 enfants en cour de récréation).
• Manque de considération : Le métier souffre de bas salaires, d'un recrutement précaire et d'une absence de reconnaissance professionnelle, favorisant un climat d'agressivité.
B. Problématiques de recrutement et de formation
Les municipalités, responsables du périscolaire, font face à des difficultés majeures pour trouver du personnel qualifié.
• Règles floues : Le recrutement se base parfois uniquement sur la "bienveillance et le bon sens", sans formation approfondie.
• Hétérogénéité de gestion : La qualité de l'encadrement dépend directement des choix budgétaires et politiques de chaque mairie.
--------------------------------------------------------------------------------
II. Violences Sexuelles et Impunité
Le document souligne une défaillance critique dans la protection des mineurs face aux prédateurs potentiels au sein des structures d'accueil.
A. Données chiffrées et réalité du terrain
Il n'existe pas de statistiques officielles nationales, forçant les enquêteurs à compiler leurs propres données à partir de la presse quotidienne régionale.
• Suspensions : 128 animateurs suspendus à Paris en une décennie.
• Répartition : Des affaires sont recensées partout en France (Nancy, Marseille, Haute-Savoie, Moselle).
B. Le traitement des signalements
Un problème majeur réside dans la lenteur de réaction des autorités face aux alertes :
• Cas du 15e arrondissement : Un animateur a été interpellé en 2023 pour des jeux et danses à caractère sexuel, alors que des signalements le concernant existaient depuis 2019.
• Culture du silence : Trop souvent, les signalements ne sont pas correctement pris en compte, permettant à des individus suspects de rester au contact des enfants pendant des mois, voire des années.
C. Statistiques Judiciaires (Source : Ciivise)
| Type de procédure | Pourcentage | | --- | --- | | Plaintes classées sans suite | 73 % | | Plaintes aboutissant à une condamnation | 3 % |
Cette difficulté de condamnation s'explique souvent par la situation de "parole contre parole", où la parole de l'enfant n'est pas toujours recueillie dans des conditions optimales.
--------------------------------------------------------------------------------
III. L'Enseignement Privé sous Contrat : Une Zone d'Ombre
L'enquête s'étend aux établissements privés catholiques sous contrat, qui scolarisent près de 2 millions d'élèves en France.
• Financement Public : Ces établissements sont financés à environ 75 % par l'argent public.
• Maltraitance Institutionnalisée : Des témoignages font état de violences physiques (ex: "roustes" en public au self) sur des enfants dès l'âge de 12 ans.
• Absence de contrôle : Malgré l'importance des fonds publics engagés, l'État n'exerce pas de contrôle suffisant sur ces établissements.
Les syndicats dénoncent un silence persistant (omerta) au sein de l'Enseignement catholique.
--------------------------------------------------------------------------------
IV. Recommandations et Solutions
A. Modèles de réussite
L'enquête cite l'exemple d'un village ayant réussi à transformer son périscolaire en augmentant son budget de seulement 8 %. Cette hausse a permis :
• La prise en compte du temps de préparation des ateliers.
• Une formation accrue des animateurs.
• Un environnement où les enfants se sentent en sécurité et épanouis.
B. Vigilance des parents
Face à ces risques, les pédopsychiatres conseillent d'être attentifs aux changements brusques de comportement chez l'enfant :
• Troubles du sommeil (énurésie nocturne/pipi au lit).
• Troubles alimentaires (perte d'appétit).
• Changements d'humeur ou repli sur soi.
Il est préconisé de favoriser des moments de "parole libre" lors de situations calmes (bain, trajet en voiture, lecture du soir) pour permettre à l'enfant d'exprimer un éventuel malaise.
-
-
www.youtube.com www.youtube.com
-
État des Lieux du Secteur Périscolaire à Paris : Dysfonctionnements et Dérives
Résumé Exécutif
Ce document de synthèse expose les défaillances systémiques au sein des structures périscolaires de la Ville de Paris, telles que révélées par une enquête en immersion.
Le constat met en lumière une gestion de crise permanente caractérisée par un recrutement fondé sur la simple disponibilité plutôt que sur les compétences, une absence de formation réelle et des violations récurrentes des taux d'encadrement légaux.
L'analyse révèle un environnement où la sécurité affective et physique des enfants est compromise par des violences verbales, un désinvestissement de certains agents et une impunité institutionnalisée pour les titulaires problématiques via un système de "chaises musicales".
Le manque de contrôle et d'exigence de la hiérarchie favorise l'émergence de dérives graves, allant de la négligence à des signalements d'attouchements sexuels.
--------------------------------------------------------------------------------
1. Un Processus de Recrutement Fondé sur l'Urgence
Le recrutement des animateurs vacataires semble dicté par une nécessité de combler les effectifs plutôt que par une évaluation des aptitudes pédagogiques.
• Critères de sélection minimaux : La mairie exige uniquement d'avoir 18 ans, d'être motivé, d'avoir un casier judiciaire vierge (vérification du FIJAIS) et des vaccins à jour.
• Absence d'évaluation des compétences : Les entretiens ne comportent aucune question sur la gestion des enfants ou l'expérience éducative. L'atout majeur retenu est la disponibilité du candidat.
• Le dogme du "bon sens" : À défaut de directives claires, le recrutement repose sur deux notions vagues : la "bienveillance" et le "bon sens", y compris pour prodiguer des soins en l'absence d'infirmière.
• Profils à risque : Le manque d'exigence conduit au recrutement de profils inadaptés.
Un témoignage interne (Karim, responsable éducatif ville) rapporte l'envoi de remplaçants arrivant en état d'ébriété ou issus de parcours de réinsertion sans évaluation préalable suffisante, malgré des avis de recrutement signalant une instabilité.
2. Déficience de la Formation et de l'Intégration
L'immersion démontre un décalage profond entre les ambitions affichées par la Ville et la réalité du terrain.
• L'inexistence du parcours d'intégration : La formation se résume à un "briefing express" (chronométré à 6 minutes et 42 secondes dans un cas précis) avant une mise en poste immédiate.
• La Charte de l'animateur ignorée : Bien que ce document définisse les missions éducatives et proscrive les comportements ambigus, il n'est souvent ni présenté ni signé par les nouveaux arrivants.
• Improvisation pédagogique : Alors que le temps périscolaire est censé être éducatif, aucune consigne ne soutient les projets pédagogiques.
Les animateurs sont invités à "faire des dessins" ou à improviser des jeux sur leur temps personnel.
3. Conditions de Travail et Non-Respect de la Légalité
La gestion des effectifs se heurte à une pénurie chronique de personnel qualifié, entraînant des entorses régulières à la réglementation.
Taux d'encadrement
| Norme Légale (moins de 6 ans) | Réalité constatée sur le terrain | | --- | --- | | 1 animateur pour 14 enfants | Jusqu'à 1 animateur pour 23 enfants | | Surveillance active requise | Sous-effectifs fréquents (ex: 2 animateurs pour 70 enfants) |
• Précarité et rotation : Les postes de vacataires (payés 15 € brut de l'heure) servent à "boucher les trous" sans continuité éducative, les remplaçants changeant de groupe quotidiennement sans présentation préalable.
• Difficulté de recrutement : La mairie peine à trouver des profils compétents en raison des plannings morcelés et des temps partiels, ne parvenant à compléter ses équipes qu'en période de fin d'études universitaires.
4. Dérives Comportementales et Climat de Violence
Le manque de cadre et de formation génère des comportements abusifs au sein des écoles.
• Violences verbales systématiques : L'usage des cris, des menaces ("vous allez rien manger") et des humiliations est une pratique quotidienne pour obtenir le silence ou le respect des règles à la cantine.
• Négligence et désinvestissement : De nombreux animateurs privilégient l'usage personnel de leur smartphone au détriment de la surveillance active des enfants, en violation directe de la charte professionnelle.
• Absence de limites physiques : L'immersion a révélé des gestes inappropriés, tels que des baisers forcés sur la bouche imposés aux enfants par certains membres du personnel sous couvert d'affection.
5. Défaillances de la Hiérarchie et Impunité
Le système de contrôle interne semble incapable de réguler ou d'écarter les profils dangereux.
• Pilotage à distance : La hiérarchie supérieure se rend rarement sur le terrain (environ trois fois par an), adoptant une posture de "no news good news".
• Le système des "chaises musicales" : Pour les agents titulaires (fonctionnaires) faisant l'objet de signalements pour maltraitance (fessées, secousses par les oreilles), la Ville privilégie le déplacement géographique plutôt que la sanction disciplinaire ou l'exclusion.
Cela permet à des individus problématiques de poursuivre leur carrière en changeant simplement d'établissement.
• Gravité des faits signalés : L'enquête mentionne une plainte pour attouchements sexuels sur une enfant pendant la sieste, ayant entraîné la suspension d'un agent au nom du principe de précaution.
Citations Clés
« Si un enfant se blesse, elle fait comment ? [...] Voilà, c'est votre bon sens. » — Recruteuse de la Ville de Paris.
« On forme sur le tas. [...] Allez, bienvenue à bord du briefing express. » — Responsable Éducatif Ville (REV).
« Si ça te fait de la peine [que les enfants pleurent], c'est pas fait pour toi ce travail. Parce que sinon tu vas te faire bouffer. » — Animatrice à une nouvelle recrue.
« On peut faire une carrière entière en fait en étant déplacé tout le temps, ça il n'y a aucun problème. » — Karim, responsable périscolaire anonyme.
Tags
Annotators
URL
-
-
www.youtube.com www.youtube.com
-
Synthèse Analytique : Les Mécanismes du Contrôle Coercitif et de la Violence Intrafamiliale
Résumé Exécutif
Ce document analyse les dynamiques du contrôle coercitif à travers le prisme des audiences judiciaires et des témoignages d'experts présentés dans l'enquête d'ARTE.
Le contrôle coercitif ne se limite pas à des actes isolés de violence physique, mais constitue un système délibéré de domination visant à aliéner la liberté de la victime. Les points clés identifiés incluent :
• La nature systémique du contrôle : Il s'agit d'une stratégie globale incluant la micro-surveillance, l'isolement social et la dévalorisation psychologique.
• L'arsenal tactique : L'utilisation de technologies (GPS, caméras, accès aux réseaux sociaux) et de pressions économiques pour maintenir une emprise totale.
• La rhétorique de l'agresseur : Une tendance systématique à l'inversion de la culpabilité, à la minimisation des faits et à l'utilisation de prétextes émotionnels pour justifier la violence.
• L'évolution juridique : La nécessité d'intégrer la notion de contrôle coercitif dans le droit pour déconstruire les rapports de domination ancrés historiquement dans le Code civil.
--------------------------------------------------------------------------------
1. Définition et Stratégies du Contrôle Coercitif
Le contrôle coercitif est décrit comme une « arme par excellence » pour soumettre l'autre.
Contrairement à la violence ponctuelle, il s'inscrit dans la durée et l'omniprésence.
Mécanismes de surveillance et de micro-contrôle
L'agresseur cherche à envahir l'espace psychique, intime et professionnel de la victime par divers moyens :
• Surveillance technologique : Installation de traceurs GPS sous les véhicules, utilisation de caméras de surveillance au domicile, et exigence des codes d'accès aux téléphones et réseaux sociaux.
• Intrusion nocturne : Privation de sommeil par la musique forte ou réveils forcés durant la nuit pour obtenir des « aveux » d'infidélité imaginaire.
• Contrôle du corps : Surveillance de la tenue vestimentaire et, dans des cas extrêmes, inspection des sous-vêtements pour déceler des preuves de rapports extra-conjugaux ou de prostitution.
Isolement et dévalorisation
Le contrôle passe par la création d'un désert social autour de la victime :
• Rupture des liens : Interdiction ou limitation des visites à la famille (notamment la mère) et aux amis, sauf en présence de l'agresseur.
• Atteinte à la dignité : Utilisation d'un langage dégradant (« pute », « salope », « moins que rien ») et dénigrement constant des capacités professionnelles ou maternelles.
• Pathologisation de la victime : Traiter la victime d'« hystérique » ou de « folle » pour discréditer sa parole et justifier le contrôle.
--------------------------------------------------------------------------------
2. Le Cycle de la Domination et de la Contrainte
Le passage de la violence verbale à la violence physique et à la séquestration suit une progression souvent prévisible.
La contrainte physique et matérielle
• Séquestration : Fermer la maison à clé pour empêcher la victime de sortir, ou s'enfermer avec elle pour l'empêcher de fuir une dispute.
• Contrôle des besoins vitaux : Interdiction d'accès à la cuisine pour les enfants ou la conjointe, contrôle strict des courses alimentaires (ne rapporter que de l'eau, forcer la victime à consommer des produits périmés).
• Emprise économique : Captation des prestations sociales et reproches systématiques sur la gestion financière, visant à créer une dépendance totale.
Menaces et terrorisme domestique
Le climat de peur est maintenu par des menaces de mort explicites et récurrentes :
• Menaces d'homicide : SMS répétés (« je vais te tuer », « je vais te crever »), mise en joue avec une arme à feu chargée, ou menaces de précipiter la victime d'un pont ou contre un mur sur l'autoroute.
• Chantage au suicide : Utilisation de la menace de se donner la mort pour manipuler la victime et l'empêcher de rompre.
• Violence physique directe : Crachats au visage, strangulation jusqu'à la perte de connaissance, coups de tête et pressions physiques pour « faire taire » la victime.
--------------------------------------------------------------------------------
3. Analyse de la Défense des Agresseurs
L'analyse des audiences révèle des schémas de défense récurrents chez les auteurs de violences, visant à éluder leur responsabilité.
| Tactique de défense | Manifestation constatée dans les sources | | --- | --- | | Inversion de la culpabilité | Affirmer que la victime est « capricieuse », « exigeante » ou qu'elle a « provoqué » l'acte par son comportement ou son infidélité supposée. | | Minimisation | Qualifier un crachat de « simple réaction », ou des menaces de mort de « mots dits sous le coup de la colère ». | | Justification par le trauma personnel | Invoquer le décès d'un proche, une éducation violente ou une surcharge de travail pour excuser le passage à l'acte. | | Déni de la réalité | Contester les faits malgré les preuves matérielles (SMS, rapports de police, expertises médicales). | | Présentation de soi comme victime | Se décrire comme le « véritable lésé » de l'histoire, celui qui a donné tout son amour sans retour. |
--------------------------------------------------------------------------------
4. Impact Traumatique et Conséquences Sociales
La violence ne s'arrête pas à la victime directe ; elle irradie sur l'ensemble du cercle familial.
• Impact sur les enfants : Les enfants sont témoins et parfois cibles des violences.
Ils vivent dans une atmosphère de terreur (« se cacher dans la chambre », « avoir peur de son père »).
L'agresseur peut même les rendre responsables de son propre état émotionnel.
• Le cycle intergénérationnel : Les experts et magistrats soulignent le risque que les enfants reproduisent ces schémas de violence une fois adultes s'ils ne sont pas interrompus.
• La période de séparation : Identifiée comme la phase la plus dangereuse.
C'est souvent au moment où la victime tente de reprendre sa liberté que la violence culmine (harcèlement par SMS, rodéos autour du domicile, usage de traceurs).
--------------------------------------------------------------------------------
5. Perspectives Institutionnelles et Juridiques
Le document souligne le décalage entre la perception de l'agresseur et la norme légale.
• L'héritage historique : Le rappel de l'ancien article 213 du Code civil (1803), qui imposait l'obéissance de la femme à son mari, explique la persistance de structures de domination archaïques dans l'esprit de certains agresseurs.
• Le rôle de la justice : La magistrature, aujourd'hui majoritairement féminine, a pour mission de rappeler la loi et de déconstruire ces rapports de force.
Le droit doit s'immiscer « au cœur des rapports intimes » pour protéger la liberté individuelle.
• Sanctions et obligations : Les condamnations citées incluent des peines de prison (dont certaines sous surveillance électronique), des amendes pour préjudice moral, des interdictions de paraître au domicile et l'obligation de suivre des stages de responsabilisation contre les violences sexistes.
--------------------------------------------------------------------------------
Note finale : Le contrôle coercitif se définit par la volonté de « conserver sous un contrôle permanent, total et absolu » une personne, la réduisant à un objet de propriété plutôt qu'à un sujet de droit.
Sa reconnaissance judiciaire est l'outil essentiel pour briser ce système d'oppression.
Tags
Annotators
URL
-
-
www.biorxiv.org www.biorxiv.org
-
Note: This response was posted by the corresponding author to Review Commons. The content has not been altered except for formatting.
Learn more at Review Commons
Reply to the reviewers
Reviewer #1 Reviewer 1 Point 1- The authors describe cortical neuronal counts across several mammalian species, which is quite impressive, but the information on the methods of counting is lacking: how representative are the data used / shown; how many individuals / brains / sections were used for each species considered? Much more detailed description of the quantifications should be provided to judge the validity of this first conclusion.
Response: We sincerely thank the reviewer for this insightful and constructive suggestion. We agree that the methodological description of our comparative histological analysis, which is the fundamental basis of this study, was insufficient in the original manuscript. Following the reviewer’s advice, we have extensively revised the Materials and Methods section entitled “Nissl staining and neuronal cell number count” (Page 32, Line 15).
Reviewer 1 Point 2- The authors use several markers of cortical neuron identity to confirm their neuron number measurements, but from the data shown in Figure 1D,E it seems that only some markers (Satb2) show species-differences while others do not (CTIP2 / Tbr1). How do the authors explain this discrepancy - does this mean that it is mainly Satb2 neurons that are increased in number? But if so how to explain the relative increase in subcortical projections shown in Figure S7?
Response: We appreciate the reviewer’s insightful comments regarding the marker expression patterns. Upon re-evaluating our data in light of your feedback, we agree that the species differences in deep-layer (DL) markers such as Ctip2 and Tbr1 in the adult stage appear relatively modest compared to the robust differences observed in Satb2 and the projection data shown in Figure S8.
To address this point, we have incorporated a comparison between the adult data (Figure 1) and our findings from P7 (Figure S2). As shown in the revised manuscript, the species differences for all markers are significantly more pronounced at P7 than in the adult. Notably, in the lower layers, rats exhibit a significantly higher number of marker-positive cells across all markers, including those newly added in this revision, compared to mice.
We offer the following interpretation regarding these temporal differences:
- Developmental Relevance: The marker molecules analyzed are well-established regulators of neuronal subtype fate and projection identity during development. Their critical fate-determining functions are primarily exercised during the migration and maturation phases of nascent neurons.
- Postnatal Expression Shifts: Whether these molecules maintain functional roles in the fully matured adult brain remains less certain. It is plausible that marker expression may diminish in certain neuronal populations during late postnatal development, leading to the attenuated species differences observed in adults. Consequently, we believe the strong correlation between P7 quantitative data and projection fate provides a biologically sound validation of our hypothesis.
While we have kept the discussion in the main text concise to maintain focus for the general reader, we have provided comprehensive data in Figure 1 and Figure S2. This ensures that the necessary evidence is readily available for specialists interested in these developmental dynamics.
Reviewer 1 Point 3- The authors focus their study almost exclusively on somatosensory cortex, but can they comment on other areas (motor, visual for instance)? It would be nice to provide additional comparative data on other areas, at least for some of the parameters examined across mouse and rat. Alternatively the authors should be more explicit in the abstract and description of the study that it is limited to a single area.
Response: We sincerely appreciate the reviewer’s insightful comment. As suggested, we have revised the Abstract to explicitly state that our current analysis is focused on the somatosensory cortex. Furthermore, as demonstrated in Figure 1B, we have added a discussion regarding the possibility that the species differences observed in the primary somatosensory cortex may be a general feature shared across the entire cerebral cortex, as follows: “This DL-biased thickening in rats was evident in the primary somatosensory area, but is consistently observed throughout the rostral-caudal cortical regions. (Page 19, Lines 29-31)“
Reviewer 1 Point 4- The authors provide convincing evidence of increased Wnt signaling pathway in the rat. They should show more explicitly how other classical pathways of neurogenic balance / temporal patterning are expressed in their mouse and rat transcriptome data sets. These would include Notch, FGF, BMP, for which all the data should be available to provide meaningful species comparison.
Response: We sincerely thank the reviewer for this insightful suggestion. Following your advice, we have newly included comparative data on key signaling pathways essential for cortical development—namely Wnt, FGF, NOTCH, mTOR, SHH, and BMP—across different species. These results are now presented in Figure S17. Rat progenitors show comparable patterns to other species for FGF, mTOR, and Notch signaling, but elevated Wnt and BMP expression, especially at early stages. A detailed heatmap of raw Wnt pathway gene expression across species is also included in the same supplementary figure. We believe these additions provide a more comprehensive evolutionary perspective and significantly strengthen our findings.
Reviewer 1 Point 5- The alignment of mouse and rat trajectories is very nicely showing a delay at early-mid-corticogenesis. But there is also heterochronic transcriptome at latest stages (end of 5). How can this be interpreted? Does this mean potentially prolonged astrogliogenesis in the rat cortex?
Response: We sincerely appreciate the reviewer’s insightful comment and the meticulous attention given to our data. Regarding the heterochronic shift observed at Day 5, we agree that this point was not sufficiently addressed in the original manuscript.
We would like to clarify the two primary reasons for this omission, which are inherent to the current study’s design:
- Resolution of Stage Alignment at Temporal Extremes: In our developmental stage alignment analysis, corresponding stages are defined by pairs showing the highest transcriptomic similarity within the sampled range. By definition, the precision of this alignment tends to decrease at the earliest and latest time points of a dataset. Since the "true" biological equivalent might lie outside our sampling window, we must be cautious in interpreting shifts at these temporal boundaries.
- Difference in Validation Rigor: Our study prioritized the early stages of deep-layer (DL) neuron production. Consequently, we rigorously defined the onset of neurogenesis in rats (Day 1) using multiple independent methods, including clonal analysis, immunohistochemistry, and gene expression. In contrast, Day 5 was defined simply as five days post-initiation of neurogenesis, without equivalent multi-modal validation. Given that our primary focus is the early phase of neurogenesis, the precision of the transition from late neurogenesis to gliogenesis is relatively lower. For these reasons, we believe that an in-depth discussion of the heterochronic shift at Day 5 might lead to over-interpretation. To reflect this more accurately and avoid misleading the reader, we have revised Figure 6F to de-emphasize the Day 5 shift. In addition, we revised the manuscript as “Importantly, while this analysis identified stage pairs with the highest similarity, the correspondence at the edges of the temporal sampling window is inherently less certain than at the center. Consequently, we focus on the notable reflection point at the center of our dataset. (Page 13, Lines 37-39)”.
We believe these changes more faithfully represent the biological scope of our data while maintaining the scientific integrity of our primary conclusions.
Reviewer 1 Point 6- Figure 7: description implies that module 3 is a subset of module 4, but this is not obvious at all from the panels shown. Please clarify.
Response: We sincerely appreciate the reviewer’s careful reading of our manuscript. As suggested, we have revised Figure 7 to clarify the hierarchical relationship between Module 3 and Module 4, ensuring that their inclusion is now explicitly presented.
Reviewer #2 Reviewer 2 Point 1. The introduction lacks sufficient background and fails to convey the significance of the study. Specifically, why the research was undertaken, what knowledge gap it addresses, and how the findings could be applied. Addressing these questions already in the introduction would enhance the impact of the work and broaden its readership.
Response: We sincerely appreciate the reviewer’s insightful comment on this point. Our study reports evolutionary insights gained through an unconventional approach: a single-cell level comparison between mice and rats. We agree that clarifying the necessity of this specific approach is crucial for the manuscript. Accordingly, we have added the following two points to the Introduction:
- At the end of the first paragraph, we emphasized the current lack of research on the evolutionary adaptation of cortical circuits, despite the established functional importance of evolutionarily conserved circuits. (Page 3, Lines 7-10); “Paradoxically, despite the importance of these variations, research has predominantly focused on the conserved aspects of cortical architecture. Consequently, the degree of evolutionary plasticity inherent in these circuits and the cell-intrinsic mechanisms driving their modification remain profoundly enigmatic.”)
- At the end of the third paragraph, we revised and added text (Page3, Lines 26-27; “This lack of comparative insight represents a significant gap in our understanding of how conserved developmental programs give rise to species-specific brain architectures.”).
Reviewer 2 Point 2. In figure 5 the authors conclude that "differences in cell cycle kinetics and indirect neurogenesis are unlikely to be the primary factors driving the species-specific variation in DL neuron production. Instead, the temporal regulation of progenitor neurogenic competence, which determines the duration of the DL production phase, provides a more plausible explanation for the greater number of DL subtypes observed in rats". It is not clear to this reviewer how the authors come to this conclusion. Authors observe a significant proportion of mitotic cells in rat VZ from day 1, and a higher constant proportion of mitotic progenitors in SVZ rats compared to mouse (Figure 5C). This points to an early difference in mitotic progenitors that may also lead to increased IP numbers, and potentially an increased number in DL cells, even before day 1. In addition, the higher abundance of IPs in the G2/S phase (statistically significant in 4 of the 7 time points) (Figure 5F), would suggest that this difference might play a role in the species-specific variation of DL neuron production. The authors should estimate cell cycle length instead of just measuring proportions to conclude something about cell cycle kinetics. They can then model growth curves to predict the effect caused if there were differences in cell cycle length between equivalent cell types across species.
Response: We sincerely thank the reviewer for their careful reading of our manuscript and for pointing out the overstatements in our original descriptions. We agree that a more nuanced interpretation of the data was necessary. In response to these constructive suggestions, we have made the following revisions:
- Refinement of Descriptions: We have revised the text to more accurately reflect our findings, specifically noting that the increase in RG division on Day 1 and IP proliferation throughout the neurogenic period showed a significant trend. These features are now described more fairly and cautiously in the revised manuscript. (Page 11, Lines 42-46; “Remarkably, while the temporal dynamics of mitotic density were strikingly conserved between the two species, subtle yet discernible species-specific signatures emerged. Specifically, rats exhibited a higher ratio of mitotic cells in the VZ at the onset of neurogenesis, the precise period when DL subtypes are generated in both species. Further assessment of G2/S-phase cells via pulse-EdU labeling (Figure 5D, E) “)
- Inclusion of Time-lapse Imaging Data: The reviewer is correct that measuring the proportions of M and G2/S phases provides only a limited snapshot of cell cycle dynamics. To gain a more precise insight, we performed primary cultures of neural progenitor cells (NPCs) from Day 1 and conducted live-cell time-lapse imaging. This allowed us to directly quantify the cell cycle duration of mouse and rat NPCs (Figure S9A-C).
- Comparative Analysis and Mathematical Modeling: Our new data revealed that the cell cycle lengths of the two species are remarkably similar, with no significant differences observed under these culture conditions. Furthermore, to validate the impact of these findings on overall brain development, we developed a mathematical model based on our experimental data. This model predicts the total number of cells produced over the five-day neurogenic period, providing a more robust theoretical framework for our conclusions (Figure S9D). We believe these additions significantly strengthen the manuscript and address the reviewer's concerns regarding the physiological relevance of our observations.
Reviewer 2 Point 3. In Figure 6 the authors focus only on the mouse and rat datasets. Given the availability of datasets from primates that the author used already for Figure 7, it would give the reader a broader prospective if also these datasets would be integrated in the analysis done for Figure 6, particularly it would be interesting to integrate them in the pseudotime alignment of cortical progenitor. How do human and/or macaque early and late neurogenic phase would compare to mouse and rat in this model?
Response: We sincerely appreciate the reviewer’s insightful suggestion. In accordance with this comment, we have now incorporated pseudotime alignments of cortical progenitors between primates (human, macaque) and rodents (mouse, rat), presented as pairwise gene expression distance matrices with dynamic time warping in Figure S13. These heatmaps illustrate temporal compression or stretching in progenitor gene expression progression across species. Notably, macaque progenitors show no definitive deviations from rodents, whereas human progenitors exhibit distinct protraction relative to rats and even more so to mice. These additions provide a more comprehensive cross-species perspective without altering the study's core conclusions.
Reviewer 2 Point 4. In Figures 6C and 6D, the authors distinguish between cycling and non-cycling NECs and RGCs. Could the authors clarify the rationale behind making this distinction? Could the authors comment on how they interpret the impact of cycling versus non-cycling states on species-specific non-uniform scaling? Do they consider the observed non-linear correspondences to be driven by differences in cell cycle activity?
Response: We are grateful to the reviewer for their insightful observation. We agree that our initial classification of neural progenitor cell (NPC) populations based on proliferation marker expression levels followed a convention used in other studies but was, in the context of this work, unnecessary and potentially misleading. To avoid further confusion and focus on the core biological question, we have re-organized the data by pooling these populations into a single group. Regarding the concern about species differences in cell cycle kinetics, we believe there is no significant divergence between mice and rats that could explain the observed developmental patterns in temporal progression of neurogenesis. This is supported by two lines of evidence:
- Quantitative analysis of pH3-positive cells (Figure 5).
- New time-lapse imaging data of primary cultured NPCs, which shows no substantial difference in cell cycle length between the two species (Figure S9). These results indicate that the species-specific differences in deep-layer (DL) neuron production are not driven by cell division kinetics. Consequently, we conclude that the non-linear developmental progression of NPCs occurs independently of cell cycle regulation.
Reviewer 2 Point 5. For the non-uniform scaling in Figure 6F, the authors identify critical inflection points and mention that "the largest delay in rat progenitors occurring where Day 1 and Day 3 progenitors overlapped". It would be good if the authors could discuss what they think all the inflection points represents. How much can it be explained by the heterogeneity within progenitors per time point? There is a clear higher spread of histograms at days 3 and 5, and the histogram at day 5 almost overlaps with day 1. I wonder if the same conclusion about non-uniform scaling would be detected if the distance matrix was built separately for specific cell types, for example only looking at NECs or RGCs.
Response: We sincerely appreciate the reviewer’s insightful perspective on this point. In alignment with the suggestions from both this reviewer and Reviewer 1 (Point 5), we have updated the manuscript to discuss all identified inflection points. Specifically, we have clarified why our discussion focuses on the correspondence between Mouse D1 and Rat Day 3.
A recognized limitation of our current analytical approach is that it identifies the closest matching expression profiles within the specific timeframes sampled for each species. For stages at the beginning or end of our sampling window, the "true" corresponding stage in the other species may lie outside our sampled range, which naturally limits the strength of any conclusions regarding those boundary points. Consequently, while we can confidently confirm the correspondence between Mouse Day 1 and Rat Day 3—both of which sit centrally within our sampled window—we have intentionally avoided over-interpreting data near the temporal boundaries.
Regarding the cell types analyzed, this specific analysis was conducted exclusively on NECs and RGs (now shown in Figure 6F). Extensive prior research (Susan McConnell lab, Sally Temple lab, Fumio Matsuzaki lab, Dennis Jabaudon lab, and more) has established that the time-dependent mechanisms governing the fate determination of cortical excitatory neuron subtypes are encoded within RGs. Therefore, we focused our investigation on these lineages and did not include other cell types in this study. We believe this focused approach maintains the highest degree of biological relevance for our conclusions.
Reviewer 2 Point 6. The authors conclude that the elevated and prolonged expression of Wnt-ligand genes in rat RGs extend the DL neurogenic window and contribute to rat-specific expansion of deep cortical layer. In order to validate this finding it would be good for the authors to perform a perturbation experiment and reduce Wnt signalling/ Axin 2 levels in rats or depleted the Lmx1a and Lhx2 double-positive population. Response: __We thank the reviewer for this insightful suggestion. We agree that providing direct experimental evidence is crucial to demonstrating that elevated Wnt signaling in RG progenitors drives the production of DL subtype neurons in rats. To address this, we performed a functional intervention on Day 3, a stage when Wnt signaling (indicated by Axin2 expression) is significantly higher in rats than in mice (__Figure 7C, D). By introducing a dominant-negative form of TCF7L2 (dnTCF7L2) to inhibit Wnt signaling specifically in RG progenitors, we tracked the fate of the resulting neurons (Figure 7I, J). Our results showed a clear reduction in the proportion of DL neurons, accompanied by a reciprocal increase in upper-layer (UL) neurons. These findings demonstrate that maintained high levels of Wnt signaling are essential for the prolonged neurogenic capacity for DL neurons in rats. This new data has been incorporated into Figure 7.
Reviewer 2 Point 7. The authors conclude that Wnt signaling is a rat specific effect since they did not observe any clear temporal change in wnt receptors in gyrencephalic species, and only a subset of RG in rats co-express Lmx1a and Lhx2. However, specific Wntligands and receptors (Wnt5a, Fzd and Lrp6) seem to be upregulated in human as well (Fig 7G), non RG cells could act as wnt ligand inducers in other species, and it has not been demonstrated that Lmx1a and Lhx2 are the source for Wntligand production. I wonder if the authors can completely rule out a role for Wnt in the protracted neurogenesis of other species.
Response: We sincerely appreciate the reviewer’s insightful and broad perspective regarding Wnt signaling dynamics across diverse species. In this study, our primary focus was to elucidate the specific mechanisms underlying the differences between mice and rats. Consequently, we did not initially explore Wnt dynamics in other species or their roles in developmental timing in great depth in the original manuscript. We fully acknowledge that lineage-specific adaptations occur at the individual gene level; for instance, Silver and colleagues have reported that human-specific upregulation of Wnt receptor gene FZD8 modulates neural progenitor behavior (Boyd et al., Current Biology 2008, Liu et al., Nature 2025). However, our comparative analysis of five mammalian species—carefully aligned by developmental stage—reveals a distinct global trend. While individual gene variations exist like human FZD8, the expression levels of multiple Wnt-related genes, particularly ligands, are markedly higher in rats than in the other four species.
Following the reviewer’s insightful suggestion, we examined the potential role of Lmx1a in activating Wnt ligand transcription in rat cortical progenitors by analyzing their expression correlation at the single-cell level. Our analysis revealed that several Wnt ligand genes are co-expressed with Lmx1a with a remarkably strong positive correlation. While we have not yet experimentally demonstrated the direct transcriptional activation of Wnt ligands by Lmx1a in these cells, this robust correlation at single-cell resolution strongly suggests that Lmx1a regulates Wnt ligand expression. These new findings are now included in Figure 7 and Figure S16, and the corresponding results section (Page 15, Lines 42-44) has been revised accordingly.
__Reviewer 2 Point 8 __Minor comments: The RNAscope experiment is currently qualitative. Is it the mRNA copy number per cell equal in both species but more cells are positive in rat, or are there differences in number of mRNA molecules as well? It is not indicated if the RNAscopeprobes are the same for mouse and rat.
Response: We sincerely thank the reviewer for this insightful suggestion. Following the comment, we performed RNAscope analysis for Axin2 in both mice and rats and quantified the results (now included in Figure 7D). The new data successfully validate the species differences initially observed in our scRNAseq analysis: specifically, the period of high-level Axin2 expression is significantly extended in rats compared to mice. These findings provide histological evidence that reinforces our conclusions regarding the distinct temporal dynamics between the two species.
Regarding probe design, the Axin2 RNAscope probes target conserved and corresponding sequences between mouse and rat, with species-specific probes optimized for each organism to ensure maximal specificity and sensitivity. We have updated the Methods section ("Fluorescent in situ hybridization with RNAscope") to include these details.
Reviewer #3
Reviewer 3 Point 1. Satb2 is also widely recognized as a deep layer marker. The authors need to perform analysis and quantification in Figs 1 and 4 with other II/III and IV markers such as Cux1 and Rorb.
Response: We thank the reviewer for their insightful comments regarding the marker specificity. We fully agree that while Satb2 is a robust marker for callosal projection identity, its broad distribution across both deep and upper layers limits its utility as a layer-specific marker. As the reviewer suggested, Cux1 (Layers 2/3) and Rorb (Layer 4) are indeed superior markers for defining laminar identity.
To address this, we have incorporated new immunohistochemical data for these markers in both the quantification of somatosensory cortical neurons (Figure S2) and the birth-dating analysis (Figure 4).
Our new findings are as follows:
- Layer Quantification (Figure S2): By utilizing Cux1 and Rorb as more specific upper-layer (UL) markers, we confirmed that there are no significant differences in the number of these neurons between mice and rats.
- Birth-dating Analysis (Figure 4): These markers allowed us to more precisely define the timing of Cux1/Rorb-positive cell generation, revealing subtle but important differences between the two species. While these additions do not alter the fundamental narrative of the original manuscript, they have significantly enhanced the precision and rigor of our analysis. We are grateful to the reviewer for guiding us toward this more robust validation.
Reviewer 3 Point 2. Rats have larger cortices. Therefore, quantification of neurons should also be normalized to cortical thickness in Fig 1E and also represented with individual data points.
Response: We sincerely appreciate the reviewer’s constructive suggestion. We agree that normalizing the number of cortical neurons by thickness provides a more rigorous comparison. Accordingly, we have calculated the neuronal density (cell count per unit thickness) for Tbr1- and Ctip2-positive cells and included these data in Figure S2C. Our analysis confirms that these populations are distributed at a significantly higher density in mice compared to rats.
Furthermore, we have updated the visualization in Figure 1E to display individual data points, ensuring full transparency of the underlying distribution. We believe these revisions, prompted by the reviewer’s insight, have substantially strengthened the clarity and persuasiveness of our manuscript.
Reviewer 3 Point 3. The clonal analysis in Figs 2 and 3 quantifies GFP and RFP and reports these as neurons. However, without using cell-specific markers, it seems the authors cannot exclude that some progeny are also glia derived from a radial glial progeny. I don't expect all experiments to have this but they must have some measures of both populations to address this possibility. This needs to be addressed to build confidence in the conclusion that there is clonal production of neurons.
Related to this, the relationship between position and fate is not always 1 to 1. The data summarized in Fig 2G are based on position and not using subtype markers. They should include assessment of markers as they do in Fig 4.
Response: We sincerely thank the reviewer for this insightful comment. We agree that a clear definition of cell types is essential for the accuracy of clonal analysis.
In this study, we primarily identified neurons based on their distinct morphological characteristics and performed measurements specifically on these cells. To validate this approach, we confirmed that the vast majority of cells identified as neurons were positive for NeuN and cortical excitatory neuron markers, while remaining negative for glial markers such as Olig2 and SOX9. (Notably, at postnatal day 7, most cells in the glial lineage exist as undifferentiated Olig2-positive progenitors). These observations support our conclusion that the cells analyzed based on morphology are indeed cortical excitatory neurons.
As the reviewer rightly pointed out, evaluating cell composition using fate-specific marker expression is the ideal approach. However, our current experimental setup required multiple fluorescence channels for DAPI staining (to assess tissue architecture) and immunostaining for GFP and RFP (to identify labeled clones). Due to these technical constraints regarding available detection channels and host species compatibility, we relied on morphological criteria for the primary analysis.
To address this concern and ensure the reliability of our findings, we performed additional analyses using a subset of samples. By co-staining retrovirally labeled neurons with cell-fate markers, we obtained results consistent with our other data (Figures 1 and 4) regarding laminar position and marker expression. Based on this consistency, we are confident that our classification based on morphology and laminar position does not alter the fundamental conclusions of this study.
Reviewer 3 Point 4. In Fig 5, the authors use PH3 as well as EdU to measure differences in indirect neurogenesis. Using EdU and Tbr2 they report more dividing IPs. However they need to measure this over the total number of Tbr2 cells as it is not normalized to differences in Tbr2 cells between species. Are there total differences in Tbr2+ cells when normalized to DAPI as well? Moreover, little analyses is performed to measure any impact on radial glia. As no striking differences were observed in IPs this leaves the cellular mechanism a bit unclear and begs the impact on radial glia. Measuring PH3+ cells in VZ and SVZ is not cell specific nor does it yield information to support the prolonged neurogenesis.
Response: We sincerely thank the reviewer for this insightful suggestion. We agree that quantifying Tbr2+/EdU+ double-positive cells alone was insufficient to fully capture the IP dynamics. Following the reviewer’s advice, we have now quantified the total population of Tbr2+ cells, normalized to the number of DAPI-stained nuclei. This new analysis reveals that mice and rats exhibit nearly indistinguishable temporal dynamics (Figure S10). When integrated with the original Tbr2+/EdU+ data in Figure 5, these findings suggest that rats maintain a slightly higher IP pool throughout the neurogenic period. This implies that the increased neuronal production in rats is not restricted to a specific phase, but rather occurs consistently across all developmental stages. We believe these additional data significantly strengthen our conclusions.
Reviewer 3 Point 5. The sc-seq is done in rat and compared to published mouse data from corresponding stages. They conclude species specific differences in progenitor gene expression. I am unsure how appropriate this is. Are similar sequencing platforms used? Can they find similar results if using multiple dataset? There are other datasets that may be used to validate these findings beyond DiBella et al.
Response: We sincerely thank the reviewer for this insightful comment. We agree that establishing the validity of our analytical approach is crucial for the reader’s confidence in our findings. To address this, we have explicitly stated in the revised manuscript that both our rat scRNAseq data and the publicly available datasets were generated using consistent experimental platforms. This ensures that the integration process is technically sound.
Revised text (Page 13, Lines 16-18): “After quality control, we integrated these profiles with previously published mouse cortical cell data from corresponding neurogenic stages, which is prepared using the consistent platform with ours (35) (Figure S11).”
Furthermore, to ensure the robustness of our comparative analysis, we have incorporated an additional independent dataset (Ruan et al., PNAS 2021) in addition to the Di Bella et al. Nature 2021 data used in the original manuscript. We confirmed that the results obtained using this second dataset are highly consistent with our initial findings, further validating our conclusions across different studies (Figure S13A).
Reviewer 3 Point 6. Wnt ligand analysis requires validation in situ across developmental stages, to support their conclusions. Ideally they might consider doing some manipulations to provide context to this observation.
Response: We sincerely thank the reviewer for these insightful suggestions. We agree that validating the spatial expression patterns of Wnt ligands and confirming their expression in rat-specific RG, as suggested by our scRNAseq data, is crucial for strengthening our conclusions.
Regarding the expression of Wnt3a, a key ligand in cortical development: although immunohistochemical analysis clearly identified Wnt3a expression in the cortical hem, the expression levels in RG within the cortical area were substantially lower than those in the hem, making definitive visualization challenging. To complement these findings and provide more robust evidence, we performed the following additional experiments:
- Validation of Wnt signaling levels: Using RNAscope-based in situ hybridization for Axin2, we successfully confirmed the elevated Wnt signaling levels in rat-specific RG (Figure 7C, D), consistent with our scRNAseq findings.
- Elucidating strikingly high correlated expressions of Lmx1a and Wnt ligand genes in the rat cortical progenitors in our scRNAseq dataset (Figure S16B).
- Functional analysis: To test the functional significance of this signaling, we inhibited Wnt signaling by electroporating dominant-negative TCF7L2 into rat RG at E15.5. This manipulation resulted in a subtype shift of the generated neurons toward an upper-layer identity (Figure 7I, J). These new results demonstrate that the rat-specific extension of high Wnt signaling levels serves as a fundamental mechanism for the prolonged production of deep-layer (DL) neurons. We are grateful to the reviewer for these suggestions; these additional data have significantly strengthened our core argument that the heterochronic regulation of Wnt signaling states drives the evolution of cortical neuronal composition.
__Reviewer 3 Point 7 __Minor concerns-1
Please separate images in Fig 1D it is very strange to have them all on top of each other.
Response: We sincerely thank the reviewer for this suggestion. As requested, we have provided individual channel images alongside the merged multicolor panels. We agree that this modification significantly enhances the clarity of our data and makes the results much easier to interpret.
__Reviewer 3 Point 8 __Minor concerns-2
Are data in Fig 4E Edu+Tbr1+EdU+? This should be clarified and would be most accurate.
Response: We appreciate the reviewer’s suggestion. We added the label of Y axes of the plots in Figure 4E-K. The procedure of cell count in these analyses are documented in the caption of Figure 4E-K, “Normalized counts of neurons colabeled for EdU and projection-specific markers, relative to the peak of EdU+ and marker+ cells.”.
__Reviewer 3 Point 9 __Minor concerns-3
Fig 4 graphs only have titles without Y axis. Please adjust location of title or repeat for clarity.
Response: We thank the reviewer for this helpful suggestion. To clarify the definition of the Y-axis, we have now added a descriptive label to the axis in the revised figure.
__Reviewer 3 Point 10 __Minor concerns-4
Fig 4A implies cumulative incorporation which I don't think is being performed here. They should clarify this in the figure.
Response: We appreciate the reviewer’s insightful comment. To avoid any potential misunderstanding regarding the additivity of the effect, we have revised the illustration in Figure 4A for greater clarity.
__Reviewer 3 Point 11 __Minor concerns-5
Fig 5 needs labels for the actual stages assayed, as illustrated in Fig 4A.
Response: We thank the reviewer for this helpful suggestion. Following your comment, we have added the developmental stage information (expressed as embryonic days) for both mice and rats in the revised manuscript.
-
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Referee #3
Evidence, reproducibility and clarity
In this study the authors investigate differences between two closely related species, rats and mice, in terms of cortical development and neuronal composition. They first perform comparative analysis of cortical layers which revealed the density and markers of deep layer neurons of rats is disproportionately larger compared to adult mice. They then use retroviruses for lineage analysis from embryonic stages to P7. They find in general that there are temporal differences in when mice and rats produce upper versus deep layer neurons, with the process being protracted in rats. EdU injections were used to report differences in the timing of cortical neuron generation between species and they note no striking differences in IPs. Sc-sequencing of rat cortices at different stages was then used to measure temporal changes in gene expression and compared to published mouse data. They note that rats have sustained Wnt ligand expression in radial glia highlighting that as a potential mechanism of action.
Major concerns 1. Satb2 is also widely recognized as a deep layer marker. The authors need to perform analysis and quantification in Figs 1 and 4 with other II/III and IV markers such as Cux1 and Rorb. 2. Rats have larger cortices. Therefore, quantification of neurons should also be normalized to cortical thickness in Fig 1E and also represented with individual data points. 3. The clonal analysis in Figs 2 and 3 quantifies GFP and RFP and reports these as neurons. However, without using cell-specific markers, it seems the authors cannot exclude that some progeny are also glia derived from a radial glial progney. I don't expect all experiments to have this but they must have some measures of both populations to address this possibility. This needs to be addressed to build confidence in the conclusion that there is clonal production of neurons. Related to this, the relationship between position and fate is not always 1 to 1. The data summarized in Fig 2G are based on position and not using subtype markers. They should include assessment of markers as they do in Fig 4. 4. In Fig 5, the authors use PH3 as well as EdU to measure differences in indirect neurogenesis. Using EdU and Tbr2 they report more dividing IPs. However they need to measure this over the total number of Tbr2 cells as it is not normalized to differences in Tbr2 cells between species. Are there total differences in Tbr2+ cells when normalized to DAPI as well? Moreover, little analyses is performed to measure any impact on radial glia. As no striking differences were observed in IPs this leaves the cellular mechanism a bit unclear and begs the impact on radial glia. Measuring PH3+ cells in VZ and SVZ is not cell specific nor does it yield information to support the prolonged neurogenesis. 5. The sc-seq is done in rat and compared to published mouse data from corresponding stages. They conclude species specific differences in progenitor gene expression. I am unsure how appropriate this is. Are similar sequencing platforms used? Can they find similar results if using multiple dataset? There are other datasets that may be used to validate these findings beyond DiBella et al. 6. Wnt ligand analysis requires validation in situ across developmental stages, to support their conclusions. Ideally they might consider doing some manipulations to provide context to this observation.
Minor concerns 1. Please separate images in Fig 1D it is very strange to have them all on top of each other. 2. Are data in Fig 4E Edu+Tbr1+EdU+? This should be clarified and would be most accurate. 3. Fig 4 graphs only have titles without Y axis. Please adjust location of title or repeat for clarity. 4. Fig 4A implies cumulative incorporation which I don't think is being performed here. They should clarify this in the figure. 5. Fig 5 needs labels for the actual stages assayed, as illustrated in Fig 4A.
Significance
Strengths:
The finding that there are differences in cortical composition between rats and mice and that this is linked to prolonged neurogenesis in rats Use of careful and detailed lineage analysis to define differences in temporal production of neurons Inclusion of single cell sequencing
Limitations:
Largely descriptive Requires additional investigation to support some conclusions about neurons Concerns about inferring too much from single cell sequencing done by the authors but compared to publication
Advance: Finding that there are differences in neurogenesis between closely related species is interesting and provides insight into mechanisms of cortical evolution.
Audience: Evolution, cortical development
Expertise: Cortical development, evolution
-
-
pmc.ncbi.nlm.nih.gov pmc.ncbi.nlm.nih.gov
-
RRID:AB_3717302
DOI: 10.7554/eLife.99438
Resource: None
Curator: @evieth
SciCrunch record: RRID:AB_3717302
Tags
Annotators
URL
-
-
pmc.ncbi.nlm.nih.gov pmc.ncbi.nlm.nih.gov
-
RRID:SCR_016137
DOI: 10.1038/s44319-025-00653-9
Resource: Microsoft Excel (RRID:SCR_016137)
Curator: @scibot
SciCrunch record: RRID:SCR_016137
Tags
Annotators
URL
-
-
www.examtopics.com www.examtopics.com
-
NOTA: Cada respuesta correcta vale un punto.
Los blobs de tipo block con el prefijo transactions moverán automáticamente a almacenamiento cool aquellos blobs que no hayan sido modificados en más de 60 días, y eliminarán los blobs que no hayan sido modificados en 365 días.
Los blobs se moverán al nivel de almacenamiento cool si no han sido accedidos durante 60 días.
La regla de la política moverá a la capa cool las versiones anteriores dentro de un contenedor llamado transactions que tengan 60 días o más, y eliminará las versiones anteriores que tengan 365 días o más.
Los blobs se volverán a mover automáticamente de cool a hot si se acceden nuevamente después de haber sido movidos a cool.
-
NOTA: Cada respuesta correcta vale un punto.
SaveScore funcionará con Cosmos DB.
SaveScore actualizará y reemplazará un registro si ya existe uno con el mismo playerId y gameId.
Los datos del leaderboard del juego se particionarán automáticamente usando gameId.
SaveScore almacenará en la base de datos los valores de los parámetros gameId y playerId.
-
Seleccionar y colocar:
Si ningún servidor edge en el POP tiene la imagen en caché, el POP solicita el archivo al servidor de origen.
Un usuario solicita la imagen desde la URL del CDN. El DNS dirige la solicitud al POP con mejor rendimiento.
Las solicitudes posteriores del archivo pueden dirigirse al mismo POP usando la URL de la imagen del logo del CDN. El servidor edge del POP devuelve el archivo desde la caché si el TTL no ha expirado.
El servidor de origen devuelve la imagen del logo a un servidor edge en el POP. Un servidor edge en el POP almacena la imagen del logo en caché y devuelve la imagen al cliente.
-
-
pmc.ncbi.nlm.nih.gov pmc.ncbi.nlm.nih.gov
-
PDKP (PDKEY01879)
DOI: 10.14348/molcells.2022.5002
Resource: Bloomington Drosophila Stock Center (RRID:SCR_006457)
Curator: @bdscstockkeepers
SciCrunch record: RRID:SCR_006457
Tags
Annotators
URL
-
-
pmc.ncbi.nlm.nih.gov pmc.ncbi.nlm.nih.gov
-
da-Gal4
DOI: 10.18632/aging.204005
Resource: Bloomington Drosophila Stock Center (RRID:SCR_006457)
Curator: @bdscstockkeepers
SciCrunch record: RRID:SCR_006457
Tags
Annotators
URL
-
-
54.241.53.176 54.241.53.176
-
42880
DOI: 10.1073/pnas.2118124119
Resource: RRID:BDSC_42880
Curator: @maulamb
SciCrunch record: RRID:BDSC_42880
-
-
pmc.ncbi.nlm.nih.gov pmc.ncbi.nlm.nih.gov
-
Df(2R)7094
DOI: 10.1038/s41467-022-31253-z
Resource: Bloomington Drosophila Stock Center (RRID:SCR_006457)
Curator: @bdscstockkeepers
SciCrunch record: RRID:SCR_006457
Tags
Annotators
URL
-
-
stylo.ecrituresnumeriques.ca stylo.ecrituresnumeriques.ca
-
musée. » Voir Ferloni et Sitzia (2022).↩︎
Il faudrait intégrer le numéro de page auquel se trouve la citation.
-
-
observatoire.francophonie.org observatoire.francophonie.org
-
5e langue mondiale par le nombre de ses locuteurs, après l’anglais, le mandarin, l’hindi et l’espagnol, la langue française est la seule, avec l’anglais, à être présente sur les 5 continents.
It's really interesting that out of all the languages in the world, French and English are the only languages that are present on the 5 continents. It's important to understand how widespread the language is.
-
angue du foyer et de l’école, transmise par les parents et/ou apprise à l’école, là où elle est langue de scolarisation ; langue officielle (seule ou aux côtés d’autres langues) qui sert pour les démarches administratives, les relations professionnelles, les contacts avec les institutions… langue sociale, quand elle permet de communiquer entre concitoyens de langues maternelles différentes ; langue de communication et de culture, très utilisée dans les médias, les activités culturelles, la littérature
French is used in many social contexts. It is used at home, school, in media, cultural activities, and literature. French is mostly acquired during childhood and learned at school or through parents.
-
Cette francophonie mondiale recouvre des réalités fort différentes et les dynamiques qui la traversent méritent un examen attentif. En effet, les usages de la langue française (en famille, à l’école, au travail, dans les échanges internationaux…), sa présence dans l’environnement sonore et visuel des populations, la fréquence de son emploi, sont très variables selon les régions, voire selon les pays observés.
This website was created by an organization to inform about the globalization of the French language. French is a global language as there are many French speaking countries. French culture varies from country to country as well.
-
Évolution de la population de cinq espaces linguistiques définis selon la langue officielle
This sentence along with this graph shows the progress/growth over time of French speaking countries. This is very informative and quite interesting.
-
-
telecty-docs.vercel.app telecty-docs.vercel.app
-
timeline administrativo asociado a la SIM (e
falta el paso de filtros. no relevar, proponer
-
-
telecty-docs.vercel.app telecty-docs.vercel.app
-
ncola y ejecuta la operación técnica (interno)
cuando lo decimos los cambios en service_profile? sim_profile_change
-
-
telecty-docs.vercel.app telecty-docs.vercel.app
-
Provider Integration Registry (read-only): consulta a provider-service el registro de integraciones habilitadas y capacidades soportadas, para ejecutar la importación.
me parece que esto salio
-
-
-
Analyse de la Tendance « Mamans Ghettossori » : Entre Lutte des Classes et Réalités Éducatives
Résumé Exécutif
L'émergence de la tendance « mamans ghettossori » sur TikTok à la fin de l'année 2024 marque un tournant dans la représentation de la parentalité sur les réseaux sociaux.
Née en réaction au modèle « maman Montessori » — perçu comme bourgeois, permissif et déconnecté — cette tendance met en scène une éducation ancrée dans la réalité des milieux populaires.
Si ce mouvement permet de déculpabiliser de nombreux parents face aux injonctions de la parentalité positive, il soulève également des inquiétudes quant à la validation potentielle des Violences Éducatives Ordinaires (VEO).
Au-delà du simple contenu viral, ce phénomène cristallise une « lutte des classes 2.0 » et une réponse aux accusations de démission parentale, soulignant la nécessité de retrouver des espaces de dialogue neutres et bienveillants, tels que les associations de parents.
--------------------------------------------------------------------------------
1. Genèse et Mécanismes du Phénomène TikTok
La tendance a été initiée par l'influenceuse niçoise Jessica French Riviera le 13 décembre 2024, avant de se propager massivement sous forme de « reels ».
Le format « Ghettossori »
Le modèle type des vidéos repose sur une litanie de phrases commençant par : « Je suis une maman ghettossori, alors bien entendu que... ». Parmi les exemples cités dans les sources :
• Gestion des loisirs : Accepter des soirées pyjamas avec de nombreux copains, pop-corn et chips sans restriction.
• Défense personnelle : Inscrire l'enfant au kickboxing pour qu'il puisse se défendre en cas de conflit (« heja »).
• Rapport à l'autorité : L'enfant craint davantage la réaction de sa mère s'il ne s'est pas défendu que celle de l'école.
• Réalité quotidienne : Commander une pizza par « flemme » de cuisiner tout en le présentant comme un cadeau à l'enfant.
• Compétition : Ne pas laisser gagner l'enfant aux jeux de société (Mario Kart, galette des rois).
La figure de proue Montessori : Armelle
En opposition, la maman Montessori type est représentée par Armelle (@moharmelle).
Ses vidéos, où elle prône une bienveillance absolue (comme attendre des heures dans une voiture qu'un enfant se réveille), cumulent des millions de vues mais suscitent l'incrédulité, voire la moquerie.
Elle est devenue la cible principale des pastiches et le contre-modèle sur lequel s'est construit l'archétype ghettossori.
--------------------------------------------------------------------------------
2. Une Lutte des Classes 2.0 par Réseaux Interposés
Le conflit entre « Montessori » et « Ghettossori » dépasse le cadre pédagogique pour devenir un enjeu sociologique majeur.
• Une fracture socio-économique : En France, le réseau Montessori est majoritairement privé et coûteux (sur 300 établissements, seuls trois sont sous contrat avec l'État en 2022).
Cela associe cette pédagogie aux classes supérieures, créant une opposition naturelle avec les milieux populaires.
• Perception culturelle :
◦ Modèle Montessori : Perçu comme celui des « parents bobos permissifs », prescripteurs d'une morale normative.
◦ Modèle Ghettossori : Revendiqué comme celui de la « vraie vie », valorisant l'imperfection et l'authenticité face aux contraintes du quotidien.
• Effet de halo en Belgique : Bien que les vidéos soient principalement d'origine française, le phénomène touche pleinement la Belgique francophone, les archétypes étant facilement identifiables par le public belge.
--------------------------------------------------------------------------------
3. Enjeux Éducatifs et Risques de Dérapages
L'analyse de la FAPEO souligne une tension entre la déculpabilisation nécessaire et le risque de dérive comportementale.
Le risque de banalisation des VEO
Certaines affirmations dans les vidéos ghettossori inquiètent les spécialistes. Sous couvert d'humour et de sarcasme, certains propos pourraient valider des Violences Éducatives Ordinaires (VEO).
Les journaux (Le Figaro, Le Point, RTL) notent que la frontière entre l'éloge de la « parentalité imparfaite » et la promotion d'une éducation « à la dure » est parfois poreuse.
Une réponse à la « démission parentale »
La tendance est analysée comme une réponse indirecte aux discours politiques et médiatiques stigmatisants sur la prétendue démission des parents des classes populaires.
• C'est un refus de la morale normative qui juge les mères.
• C'est une revendication du droit à l'erreur et une manifestation d'un ras-le-bol face aux injonctions académiques.
--------------------------------------------------------------------------------
4. Dynamiques Sociales et Limites de l'Humour
Un phénomène genré
Il est frappant de noter l'absence quasi totale de « papas Montessori » ou « papas ghettossori ».
L'éducation des jeunes enfants reste un domaine fortement imprégné par le genre, où les mères portent l'essentiel de la charge et du jugement social.
La dérive du « Roasting »
L'humour utilisé sur TikTok s'inscrit dans la tendance actuelle du roasting (mise en boîte). Cependant, l'analyse rappelle que :
• Du rire à l'humiliation, il n'y a qu'un pas.
• La moquerie répétée peut s'apparenter à du harcèlement ou de la maltraitance.
• L'utilisation de « l'écriture en creux » permet de railler un modèle (et la personne qui l'incarne) sans la nommer explicitement, renforçant l'agressivité du sous-texte.
--------------------------------------------------------------------------------
5. Conclusions et Perspectives : Vers un Espace de Dialogue
Pour sortir de la logique clivante des réseaux sociaux et des algorithmes qui favorisent le « drama », la FAPEO propose plusieurs pistes de réflexion :
• Unir plutôt que diviser : Au-delà des méthodes, toutes ces mères partagent le désir d'épanouissement de leurs enfants.
• Valoriser les associations de parents : Ces structures sont présentées comme des lieux idéaux pour un échange de pair à pair, sans jugement, loin de l'agressivité numérique.
• Promouvoir la « finesse » de l'humour : S'inspirer de figures comme Coluche, qui ridiculisait les faits et les travers collectifs sans jamais s'attaquer à l'individu ou tomber dans la vulgarité.
• Adopter un « regard qui écoute » : Selon Max Dorra, il est crucial de libérer l'autre de toute évaluation permanente pour restaurer un lien social authentique.
En définitive, l'opposition entre Montessori et Ghettossori ne doit pas masquer l'essentiel : la nécessité de soutenir tous les parents dans leur réalité quotidienne, en favorisant l'entraide plutôt que le clash médiatique.
-
-
fisher-j.github.io fisher-j.github.io
-
It is important to point out here that the HA and HD “high-density” treatments in this experiment only targeted a residual overstory relative density of 20% (that is 20% of assumed total carrying capacity of the site), and this was selected as an upper limit to support the objective of maintaining conifer growth (Berrill & O’Hara, 2009).
Feels like methods too. Unless this point is specifically needed to explain results...I'm not sure it is needed in the discussion.
-
-
www.biorxiv.org www.biorxiv.org
-
Author response:
We thank all reviewers for their comments. We appreciate the acknowledgement that the paper is important and that results support the major conclusions. We are planning to address the specific concerns as noted by the reviewers in the following way:
Public Reviews:
Reviewer #2 (Public review):
(1) The authors generate a new tool, a Gal4 knock-in of the jam2b locus, to track EGFP-expressing cells over time and follow the developmental trajectory of jam2b-expressing cells. Figure 1 characterizes the line. However, it lacks quantification, e.g., how many etv2-expressing cells also show EGFP expression or the contribution of EGFP-expressing cells to different types of blood vessels. This type of quantification would be useful, as it would also allow for comparison of their findings to their previous data examining the contribution of SVF cells to different types of blood vessels. All the authors state that at 30 hpf, EGFP-expressing cells can be seen in the vasculature (apparently the PCV).
It is not clear why the authors do not use a nuclear marker for both ECs (as they did in their previous publication) and for jam2b-expressing cells. UAS:nEGFP and UAS:NLS-mcherry (e.g. pt424tg) transgenic lines are available. This would circumvent the problem the authors encounter with the strong fluorescence visible in the yolk extension. It would also facilitate quantifying the contribution of jam2b cells to different types of blood vessels.
We agree with the importance of quantification. We had performed quantification of jam2b<sup>Gt(2A-Gal4)</sup>;UAS:GFP contribution to different vascular beds, which was shown in Suppl. Fig. S3. We will clarify this in the revision. We also agree that nuclear GFP or mCherry would help to visualize and quantify cells. Unfortunately, we do not have nuclear UAS:GFP or UAS:mCherry line in our possession, and it will take too long to import it for the standard revision timeline. We are working on the construct, and will attempt to establish the line; therefore we are hoping to clarify these results with the nuclear line in the revised manuscript.
(2) The time-lapse movie in Figure 2 is not very informative, as it just provides a single example of a dividing cell contributing to the PCV. Also, quantifications are needed. As SVF cells appear to expand significantly after their initial specification, it would be informative to know how many cell divisions and which types of blood vessels jam2b-expressing cells contribute to. Can the authors observe cells that give rise to different types of blood vessels? Jam2b expression in LPM cells apparently precedes expression of etv2. Is etv2 needed for maintenance, or do Jam2b-expressing cells contribute to different types of tissues in etv2 mutant embryos? Comparing time-lapse analysis in wildtype and etv2 mutant embryos would address this question.
The time-lapse was meant to serve as an illustration and confirmation of jam2b cell contribution to vasculature. As noted above, Suppl. Fig. S3 provides quantification of jam2b cell contribution to different vascular beds. We had previously performed detailed time-lapse analysis and quantification of SVF cell migration to PCV, SIA and SIV using etv2-2A-Venus line (Metikala et al 2022, Dev Cell), which has some of the same (or similar) information. It is very challenging to obtain this data using jam2b reporter line due to extensive and bright GFP expression in the mesothelial layer over the yolk and yolk extension; for that reason we can only trace some GFP cells but not all of them. Regarding etv2 requirement for jam2b maintenance, we intend to address this question by analyzing jam2b cell contribution in etv2 MO injected embryos, which recapitulates the phenotype in jam2b mutants.
(3) In Figure 3, the authors generate UAS:Cre and UAS:Cre-ERT2 transgenic lines to lineage trace the jam2b-expressing cells. It is again not clear why the authors do not use a responder line containing nuclear-localized fluorescent proteins to circumvent the strong expression of fluorescent proteins in the yolk extension. It is also unclear why the two transgenic lines give very different results regarding the number of cells being labelled. The ERT2 fusions label around 3 cells in the SIA, while the Cre line labels only about 1.5 cells per embryo, with very little contribution of labelled cells to other blood vessels. One would expect the Cre line requiring tamoxifen induction to label fewer cells when compared to the constitutive Cre line. What is the reason for this discrepancy? Are the lines single integration? Is there silencing? This needs to be better characterized, also regarding the reproducibility of the experiments. If the Cre lines were to be multiple copy integrations, outcrossing the line might lead to lower expression levels in future generations.
It is also not clear how the authors conclude from these findings that "SVF cells show major contribution to the SIA and SIV" when only 1.5 or 3 cells of the SIA are labelled, with even fewer cells labelled in other blood vessels. They speculate that this might be due to low recombination efficiency, a question they then set out to answer using photoconversion of etv2:KAEDE expressing cells, an experiment that they also performed in their 2014 and 2022 publications. To check for low recombination efficiency, the authors could examine the expression of Cre mRNA in their transgenic embryos. Do many more jam2b expressing cells express Cre mRNA than they observe in their switch lines? They could also compare their experiments using Cre recombinase with those using EGFP expression in jam2b cells. EGFP is relatively stable, and the time frames the authors analyze are short. As no quantification of EGFP-expressing cells is provided in Figure 1, this comparison is currently not possible. Do these two different approaches answer different questions here?
The reviewer brings up important points, we appreciate that. Unfortunately, we do not have a nuclear switch line in our possession, and it is not possible to obtain it in the normal manuscript revision time line. Regarding UAS:Cre and UAS:CreERT2 lines, they both show rather similar labeling, with most labeled cells present in the SIA. The difference in cell number (1.5 versus 3) is likely due to different levels of Cre expression, which may vary dependent on the integration site. The lines most likely are multi-copy integrations, which can be helpful, as this would result in higher Cre expression. We will address the silencing question by performing in situ hybridization or HCR analysis for Cre or CreERT2 and comparing it with endogenous jam2b expression, as the reviewer suggested. We have noticed that the switch line used, actb2:loxP-BFP-loxP-dsRed, exhibits lower recombination frequency compared to other switch lines (we used it because it was compatible with endothelial fli1:GFP line). We will attempt to answer this question by crossing to other switch lines, which may exhibit higher recombination frequency. In principle, UAS:GFP and switch lines should produce a similar result, except that GFP decays over time and therefore our initial expectation was that switch lines may produce a more accurate result. However, this may not be the case due to low recombination efficiency, which we will attempt to address in the revision.
(4) Concerning the etv2:KAEDE photoconversion experiments: The percentages the authors report for SVF cells' contribution to the SIV and SIA differ from their previous study (Dev Cell, 2022). In that publication, SVF cells contributed 28% to the SIA and 48% to the SIV. In the present study, the numbers are close to 80% for both vessels. The difference is that the previous study analyzed 2dpf old embryos and the new one 4dpf old embryos. Do SVF-derived cells proliferate more than PCV-derived cells, or is there another explanation for this change in percentage contribution?
These numbers refer to different experiments; we apologize for the confusion. As reported earlier in Metikala et 2022, 28% of SVF cells contributed to the SIA and 48% to the SIV by 3 dpf (not 2 dpf; only PCV analysis was done at 2 dpf); SIA and SIV analysis was done based on time-lapse image analysis of etv2-2A-Venus line at 3 dpf, shown in Fig. 3C in Metikala et al. However, this only refers to SVF cell contribution. It does not mean that 28% or 48% cells in SIA or SIV are derived from SVF. The total fraction of SIA and SIV cells that are derived from SVF has not been quantified in the previous study, because that would require accurate tracking of all SVF cells, which is experimentally challenging. Etv2:Kaede experiment is slighly different, because it reports newly formed cells after 24 hpf. It cannot tell if new cells are all derived from SVF cells, although we are not aware of any other source of new endothelial cells at these stages. In the previous study by Metikala et al 2022, we reported ~22 newly formed SIA and ~50 newly formed cells in SIV by 3 dpf (Fig. 1 in Metikala et al 2022), although the entire number of cells was not quantified, therefore the percentage was not known. In the current study, we attempted to estimate the entire percentage of green only Kaede cells, which was close to 80% in both SIA or SIV at 4 dpf. Please note that this estimate was performed in the posterior portion of SIA and SIV that overlies the yolk extension and where SVF cells are observed. We did not quantify cells in the anterior SIV portion, which forms the basket over the yolk.
(5) Single-cell sequencing data: Why do the authors not show jam2b expression in their single-cell sequencing data? They sorted for (presumably) jam2b-expressing cells and hypothesize that jam2b expression in ECs at this time point is important for the generation of intestinal vasculature. Do ECs in cluster 15 express jam2b? Why are no other top marker genes (tal1, etv2, egfl7, npas4l) included in the dot blot in Figure 5b?
We appreciate the suggestion and will include additional marker genes as well as jam2b in the revised version of the manuscript.
(6) Concerns about cell autonomy of mutant phenotypes: The authors need to perform in situ hybridization to characterize jam2a expression. Can it be seen in SVF cells? The double mutants show a clear phenotype in intestinal vessel development; however, it is unclear whether this is due to a cell-autonomous function of jam2a/b within SVF cells. The authors need to address this issue, as jam2b and potentially also jam2a are expressed within the tissue surrounding the forming SVF. For instance, do transplanted mutant cells contribute to the intestinal vasculature to the same extent as wild-type cells do?
jam2a expression has been characterized in the previous studies and it is shown in the Suppl. Fig. S4E. It is primarily enriched in the skeletal muscle. However, our single-cell RNA-seq analysis shows that SVF cells also express jam2a. We will include additional data on jam2a expression in the revised manuscript. We agree that transplation to address cell autonomy is an important experiment, yet there are some practical challenges to it. Jam2a,jam2b mutant phenotype is only partially penetrant, and about 50% reduction in SVF cell number, as well as partial SIA and SIV phenotypes are observed. Only a small number of transplanted cells may contribute to intestinal vasculature, therefore it may be challenging to see the differences, given the partial penetrance. In an attempt to address cell -autonomy question, we will try a different approach. We will overexpress jam2b labeled with 2A-mCherry, and test if it can rescue the mutant phenotype in cell autonomous manner. Overexpression will be done in a mosaic manner, with higher number of cells labeled than in a typical transplantation experiment.
(7) Finally, the authors analyze the phenotypes of hand2 mutants and their impact on the expression of jam2b and etv2. They observe a reduction in jam2b and etv2 expression in SVF cells. However, they do not show the vascular phenotypes of hand2 mutants. Is the formation of the SIA and SIV disturbed? Is hand2 cell autonomously needed in ECs? The authors suggest that hand2 controls SVF development through the regulation of jam2b. However, they also show that jam2b mutants do not have a phenotype on their own. Clearly, hand2, if it were to be required in ECs, regulates other genes important for SVF development. These might then regulate jam2b expression. The clear linear relationship, as the title suggests, is not convincingly shown by the data.
As suggested, we will add the analysis of SIA and SIV in hand2 mutants during the revision process. We could not assess that easily because the line was not maintained in vascular fli1:GFP background. We do not know if hand2 is required cell-autonomously. This is an important question, but it may be answered better in a separate study. Regarding hand2-jam2b axis, it is very clear that jam2b expression in the posterior lateral plate mesoderm is completely lost in hand2 mutants, except for its more anterior domain over the yolk. This does support the idea that hand2 functions upstream of jam2b. However, the relationship may not be necessarily direct. We agree that hand2 may regulate additional genes involved in SVF cell development. We will attempt to clarify this relationship and test if jam2b overexpression may rescue hand2 mutant phenotype.
Reviewer #3 (Public review):
(1) Overall molecular mechanisms of Jam2 function are not fully uncovered in the study. How do the adhesion molecules Jam2a and Jam2b regulate SVF cell formation? Are they responsible for migration, adhesion or fate determination of these structures? The authors should provide a more in-depth study of the jam2a, jam2b mutations and assess the processes affected in these mutants. Combining these mutants with etv2:Kaede can also provide a stronger causative link between their functions and defects in SVF formation.
Our data argue that the initial SVF cell specification (based on etv2 expression) is reduced in jam2a;jam2b mutants. We do not know if the migration or fate determination of the remaining SVF cells is also affected, although this may be more challenging to answer, as there are only few SVF cells remaining. We agree that further mechanistic studies of jam2a,jam2b function are needed. However, we think that this would be better addressed in a separate study. We are currently raising mutants crossed into fli1:Kaede line, which should confirm that there are fewer new cells that emerge after Kaede photoconversion in jam2a,jam2b mutants.
(2) Have the authors tested the specificity of the jam2b knock-in reporter line? This is an important experiment, as many of the conclusions derive from lineage tracing and fluorescence reporting from this knock-in line. One suggestion is to cross the jam2b:GFP or jam2b:Gal4, UAS:GFP line to the generated jam2b mutants, and examine the expression pattern of these lines. Considering that the ISH experiment showed lack of jam2b expression, the reporter line should not be expressed in the jam2b mutants.
We show in Suppl. Fig. 2 that jam2b<sup>Gt(2A-Gal4)</sup>;UAS:GFP knock-in line has similar expression pattern as jam2b mRNA by in situ hybridization, which argues for its specificity. In the revision, we plan to use HCR analysis to confirm than jam2b mRNA is expressed in the same cells as jam2b<sup>Gt(2A-Gal4)</sup>;UAS:GFP, as an additional evidence for its specificity. Unfortunately, it is not feasible to cross jam2b knock-in line into jam2b mutants, as suggested by the reviewer. Because jam2b knock-in line targets the endogenous jam2b genomic locus, which is very close in the genome to jam2b promoter deletion in jam2b mutants, the recombination frequency would be very low, and we would not get double jam2b knock-in and knock-out events in the same chromosome.
(3) The rationale behind the regeneration study is not clear, and the mechanisms underlying the phenotype are not well described. How do the authors explain the phenotype with the impaired regeneration, and what is the significance of this finding as it relates to SVF formation and function?
We apologize for this omission. This experiment was more thouroughly described in our previous study by Metikala et al 2022. In that study we showed that when endothelial cells are ablated by treating with MTZ from 6 to 45 hpf, this results in ablation of all vascular endothelial cells except for SVF cells, because they originate later than other cells. We subsequently showed that these SVF cells can partially form PCV and intestinal vasculature, helping them regenerate, which was confirmed by time-lapse imaging. In the current study, we tested if jam2a; jam2b double mutants show defects in such vascular regeneration. Indeed, regeneration after cell ablation was reduced, which correlated with reduction in SVF cell number. This argues that jam2a/b function is required for SVF cell emergence and vascular recovery after endothelial cell ablation. We will provide better description of this experiment and discuss interpretations in the revised manuscript.
(4) The authors need to include representative images of jam2b>CreERT2 with 4-OH activation at different timepoints in Figure 3.
Yes, thanks for noting this; these images will be included in the revised manuscript.
(5) The etv2:Kaede photoconversion experiment to show that the majority of intestinal vasculature derives after 24 hours needs to be supplemented with additional data on photoconverted post-24-hour-old endothelial cells, with the expectation that the majority of intestinal endothelial cells at 4 days will then be labeled with red Kaede. In addition, there have been data that show the red Kaede protein is not stable past several days in vivo, and 3 days might be sufficient for the removal or degradation of this photoconverted protein. Thus, the statement that intestinal vasculature forms largely by new vasculogenesis might be too strong based on existing data.
It is apparent from Fig. 4B that many other vessels, such as the dorsal aorta and many intersegmental vessels show robust red Kaede expression at 4 dpf, arguing that there is sufficient photoconverted Kaede present at this stage, and its degradation is unlikely to be the reason. However, we are planning to include additional control experiments, as suggested by the reviewer, to make this argument stronger.
(6) To strengthen the claim that hand2 acts upstream of jam2b, the authors can perform combinatorial genetic epistatic analysis and examine whether jam2b mutations worsen hand2 homozygous or heterozygous effects on the SVF. Similarly, overexpressing jam2b might rescue the loss of SVF/etv2 expression in hand2 mutants.
We appreciate this suggestion. Double epistatic analysis, while informative, can be tricky. In this case, we are dealing with jam2a; jam2b redundancy and also the maternal effect. It may take a while considerable effort to generate different combinations of tripple mutant lines (jam2a,jam2b,hand2), and it is unclear whether double or tripple heterozygous embryos will show any defects to clarify their epistatic relationship. Instead, as suggested, we are planning to overexpress jam2b in wild-type and hand2 mutants to address this point.
-
-
www.biorxiv.org www.biorxiv.org
-
Author response:
The following is the authors’ response to the previous reviews
Public Reviews:
Reviewer #1 (Public review):
Summary:
Wu and colleagues aimed to explain previous findings that adolescents, compared to adults, show reduced cooperation following cooperative behaviour from a partner in several social scenarios. The authors analysed behavioural data from adolescents and adults performing a zero-sum Prisoner's Dilemma task and compared a range of social and non-social reinforcement learning models to identify potential algorithmic differences. Their findings suggest that adolescents' lower cooperation is best explained by a reduced learning rate for cooperative outcomes, rather than differences in prior expectations about the cooperativeness of a partner. The authors situate their results within the broader literature, proposing that adolescents' behaviour reflects a stronger preference for self-interest rather than a deficit in mentalising.
Strengths:
The work as a whole suggests that, in line with past work, adolescents prioritise value accumulation, and this can be, in part, explained by algorithmic differences in weighted value learning. The authors situate their work very clearly in past literature, and make it obvious the gap they are testing and trying to explain. The work also includes social contexts that move the field beyond non-social value accumulation in adolescents. The authors compare a series of formal approaches that might explain the results and establish generative and modelcomparison procedures to demonstrate the validity of their winning model and individual parameters. The writing was clear, and the presentation of the results was logical and wellstructured.
We thank the reviewer for recognizing the strengths of our work.
Weaknesses:
(Q1) I also have some concerns about the methods used to fit and approximate parameters of interest. Namely, the use of maximum likelihood versus hierarchical methods to fit models on an individual level, which may reduce some of the outliers noted in the supplement, and also may improve model identifiability.
We thank the reviewer for this suggestion. Following the comment, we added a hierarchical Bayesian estimation. We built a hierarchical model with both group-level (adolescent group and adult group) and individual-level structures for the best-fitting model. Four Markov chains with 4,000 samples each were run, and the model converged well (see Figure supplement 7)
We then analyzed the posterior parameters for adolescents and adults separately. The results were consistent with those from the MLE analysis (see Figure 2—figure supplement 5). These additional results have been included in the Appendix Analysis section (also see Figure supplement 5 and 7). In addition, we have updated the code and provided the link for reference. We appreciate the reviewer’s suggestion, which improved our analysis.
(Q2) There was also little discussion given the structure of the Prisoner's Dilemma, and the strategy of the game (that defection is always dominant), meaning that the preferences of the adolescents cannot necessarily be distinguished from the incentives of the game, i.e. they may seem less cooperative simply because they want to play the dominant strategy, rather than a lower preferences for cooperation if all else was the same.
We thank the reviewer for this comment and agree that adolescents’ lower cooperation may partly reflect a rational response to the incentive structure of the Prisoner’s Dilemma.
However, our computational modeling explicitly addressed this possibility. Model 4 (inequality aversion) captures decisions that are driven purely by self-interest or aversion to unequal outcomes, including a parameter reflecting disutility from advantageous inequality, which represents self-oriented motives. If participants’ behavior were solely guided by the payoff-dominant strategy, this model should have provided the best fit. However, our model comparison showed that Model 5 (social reward) performed better in both adolescents and adults, suggesting that cooperative behavior is better explained by valuing social outcomes beyond payoff structures.
Besides, if adolescents’ lower cooperation is that they strategically respond to the payoff structure by adopting defection as the more rewarding option. Then, adolescents should show reduced cooperation across all rounds. Instead, adolescents and adults behaved similarly when partners defected, but adolescents cooperated less when partners cooperated and showed little increase in cooperation even after consecutive cooperative responses. This pattern suggests that adolescents’ lower cooperation cannot be explained solely by strategic responses to payoff structures but rather reflects a reduced sensitivity to others’ cooperative behavior or weaker social reciprocity motives. We have expanded our Discussion to acknowledge this important point and to clarify how the behavioral and modeling results address the reviewer’s concern.
“Overall, these findings indicate that adolescents’ lower cooperation is unlikely to be driven solely by strategic considerations, but may instead reflect differences in the valuation of others’ cooperation or reduced motivation to reciprocate. Although defection is the payoffdominant strategy in the Prisoner’s Dilemma, the selective pattern of adolescents’ cooperation and the model comparison results indicate that their reduced cooperation cannot be fully explained by strategic incentives, but rather reflects weaker valuation of social reciprocity.”
Appraisal & Discussion:
(Q3) The authors have partially achieved their aims, but I believe the manuscript would benefit from additional methodological clarification, specifically regarding the use of hierarchical model fitting and the inclusion of Bayes Factors, to more robustly support their conclusions. It would also be important to investigate the source of the model confusion observed in two of their models.
We thank the reviewer for this comment. In the revised manuscript, we have clarified the hierarchical Bayesian modeling procedure for the best-fitting model, including the group- and individual-level structure and convergence diagnostics. The hierarchical approach produced results that fully replicated those obtained from the original maximumlikelihood estimation, confirming the robustness of our findings. Please also see the response to Q1.
Regarding the model confusion between the inequality aversion (Model 4) and social reward (Model 5) models in the model recovery analysis, both models’ simulated behaviors were best captured by the baseline model. This pattern arises because neither model includes learning or updating processes. Given that our task involves dynamic, multi-round interactions, models lacking a learning mechanism cannot adequately capture participants’ trial-by-trial adjustments, resulting in similar behavioral patterns that are better explained by the baseline model during model recovery. We have added a clarification of this point to the Results:
“The overlap between Models 4 and 5 likely arises because neither model incorporates a learning mechanism, making them less able to account for trial-by-trial adjustments in this dynamic task.”
(Q4) I am unconvinced by the claim that failures in mentalising have been empirically ruled out, even though I am theoretically inclined to believe that adolescents can mentalise using the same procedures as adults. While reinforcement learning models are useful for identifying biases in learning weights, they do not directly capture formal representations of others' mental states. Greater clarity on this point is needed in the discussion, or a toning down of this language.
We sincerely thank the reviewer for this professional comment. We agree that our prior wording regarding adolescents’ capacity to mentalise was somewhat overgeneralized. Accordingly, we have toned down the language in both the Abstract and the Discussion to better align our statements with what the present study directly tests. Specifically, our revisions focus on adolescents’ and adults’ ability to predict others’ cooperation in social learning. This is consistent with the evidence from our analyses examining adolescents’ and adults’ model-based expectations and self-reported scores on partner cooperativeness (see Figure 4). In the revised Discussion, we state:
“Our results suggest that the lower levels of cooperation observed in adolescents stem from a stronger motive to prioritize self-interest rather than a deficiency in predicting others’ cooperation in social learning”.
(Q5) Additionally, a more detailed discussion of the incentives embedded in the Prisoner's Dilemma task would be valuable. In particular, the authors' interpretation of reduced adolescent cooperativeness might be reconsidered in light of the zero-sum nature of the game, which differs from broader conceptualisations of cooperation in contexts where defection is not structurally incentivised.
We thank the reviewer for this comment and agree that adolescents’ lower cooperation may partly reflect a rational response to the incentive structure of the Prisoner’s Dilemma. However, our behavioral and computational evidence suggests that this pattern cannot be explained solely by strategic responses to payoff structures, but rather reflects a reduced sensitivity to others’ cooperative behavior or weaker social reciprocity motives. We have expanded the Discussion to acknowledge this point and to clarify how both behavioral and modeling results address the reviewer’s concern (see also our response to Q2).
(Q6) Overall, I believe this work has the potential to make a meaningful contribution to the field. Its impact would be strengthened by more rigorous modelling checks and fitting procedures, as well as by framing the findings in terms of the specific game-theoretic context, rather than general cooperation.
We thank the reviewer for the professional comments, which have helped us improve our work.
Reviewer #2 (Public review):
Summary:
This manuscript investigates age-related differences in cooperative behavior by comparing adolescents and adults in a repeated Prisoner's Dilemma Game (rPDG). The authors find that adolescents exhibit lower levels of cooperation than adults. Specifically, adolescents reciprocate partners' cooperation to a lesser degree than adults do. Through computational modeling, they show that this relatively low cooperation rate is not due to impaired expectations or mentalizing deficits, but rather a diminished intrinsic reward for reciprocity. A social reinforcement learning model with asymmetric learning rate best captured these dynamics, revealing age-related differences in how positive and negative outcomes drive behavioral updates. These findings contribute to understanding the developmental trajectory of cooperation and highlight adolescence as a period marked by heightened sensitivity to immediate rewards at the expense of long-term prosocial gains.
Strengths:
(1) Rigid model comparison and parameter recovery procedure.
(2) Conceptually comprehensive model space.
(3) Well-powered samples.
We thank the reviewer for highlighting the strengths of our work.
Weaknesses:
(Q1) A key conceptual distinction between learning from non-human agents (e.g., bandit machines) and human partners is that the latter are typically assumed to possess stable behavioral dispositions or moral traits. When a non-human source abruptly shifts behavior (e.g., from 80% to 20% reward), learners may simply update their expectations. In contrast, a sudden behavioral shift by a previously cooperative human partner can prompt higher-order inferences about the partner's trustworthiness or the integrity of the experimental setup (e.g., whether the partner is truly interactive or human). The authors may consider whether their modeling framework captures such higher-order social inferences. Specifically, trait-based models-such as those explored in Hackel et al. (2015, Nature Neuroscience)-suggest that learners form enduring beliefs about others' moral dispositions, which then modulate trial-bytrial learning. A learner who believes their partner is inherently cooperative may update less in response to a surprising defection, effectively showing a trait-based dampening of learning rate.
We thank the reviewer for this thoughtful comment. We agree that social learning from human partners may involve higher-order inferences beyond simple reinforcement learning from non-human sources. To address this, we had previously included such mechanisms in our behavioral modeling. In Model 7 (Social Reward Model with Influence), we tested a higher-order belief-updating process in which participants’ expectations about their partner’s cooperation were shaped not only by the partner’s previous choices but also by the inferred influence of their own past actions on the partner’s subsequent behavior. In other words, participants could adjust their belief about the partner’s cooperation by considering how their partner’s belief about them might change. Model comparison showed that Model 7 did not outperform the best-fitting model, suggesting that incorporating higher-order influence updates added limited explanatory value in this context. As suggested by the reviewer, we have further clarified this point in the revised manuscript.
Regarding trait-based frameworks, we appreciate the reviewer’s reference to Hackel et al. (2015). That study elegantly demonstrated that learners form relatively stable beliefs about others’ social dispositions, such as generosity, especially when the task structure provides explicit cues for trait inference (e.g., resource allocations and giving proportions). By contrast, our study was not designed to isolate trait learning, but rather to capture how participants update their expectations about a partner’s cooperation over repeated interactions. In this sense, cooperativeness in our framework can be viewed as a trait-like latent belief that evolves as evidence accumulates. Thus, while our model does not include a dedicated trait module that directly modulates learning rates, the belief-updating component of our best-fitting model effectively tracks a dynamic, partner-specific cooperativeness, potentially reflecting a prosocial tendency.
(Q2) This asymmetry in belief updating has been observed in prior work (e.g., Siegel et al., 2018, Nature Human Behaviour) and could be captured using a dynamic or belief-weighted learning rate. Models incorporating such mechanisms (e.g., dynamic learning rate models as in Jian Li et al., 2011, Nature Neuroscience) could better account for flexible adjustments in response to surprising behavior, particularly in the social domain.
We thank the reviewer for the suggestion. Following the comment, we implemented an additional model incorporating a dynamic learning rate based on the magnitude of prediction errors. Specifically, we developed Model 9: Social reward model with Pearce–Hall learning algorithm (dynamic learning rate), in which participants’ beliefs about their partner’s cooperation probability are updated using a Rescorla–Wagner rule with a learning rate dynamically modulated by the Pearce–Hall (PH) Error Learning mechanism. In this framework, the learning rate increases following surprising outcomes (larger prediction errors) and decreases as expectations become more stable (see Appendix Analysis section for details).
The results showed that this dynamic learning rate model did not outperform our bestfitting model in either adolescents or adults (see Figure supplement 6). We greatly appreciate the reviewer’s suggestion, which has strengthened the scope of our analysis. We now have added these analyses to the Appendix Analysis section (also Figure Supplement 6) and expanded the Discussion to acknowledge this modeling extension and further discuss its implications.
(Q3) Second, the developmental interpretation of the observed effects would be strengthened by considering possible non-linear relationships between age and model parameters. For instance, certain cognitive or affective traits relevant to social learning-such as sensitivity to reciprocity or reward updating-may follow non-monotonic trajectories, peaking in late adolescence or early adulthood. Fitting age as a continuous variable, possibly with quadratic or spline terms, may yield more nuanced developmental insights.
We thank the reviewer for this professional comment. In addition to the linear analyses, we further conducted exploratory analyses to examine potential non-linear relationships between age and the model parameters. Specifically, we fit LMMs for each of the four parameters as outcomes (α+, α-, β, and ω). The fixed effects included age, a quadratic age term, and gender, and the random effects included subject-specific random intercepts and random slopes for age and gender. Model comparison using BIC did not indicate improvement for the quadratic models over the linear models for α<sup>+</sup> (ΔBIC<sub>quadratic-linear</sub> = 5.09), α<sup>-</sup>(ΔBIC<sub>quadratic-linear</sub> = 3.04), β (ΔBIC<sub>quadratic-linear</sub> = 3.9), or ω (ΔBIC<sub>quadratic-linear</sub>= 0). Moreover, the quadratic age term was not significant for α<sup>+</sup>, α<sup>−</sup>, or β (all ps > 0.10). For ω, we observed a significant linear age effect (b = 1.41, t = 2.65, p = 0.009) and a significant quadratic age effect (b = −0.03, t = −2.39, p = 0.018; see Author response image 1). This pattern is broadly consistent with the group effect reported in the main text. The shaded area in the figure represents the 95% confidence interval. As shown, the interval widens at older ages (≥ 26 years) due to fewer participants in that range, which limits the robustness of the inferred quadratic effect. In consideration of the limited precision at older ages and the lack of BIC improvement, we did not emphasize the quadratic effect in the revised manuscript and present these results here as exploratory.
Author response image 1.
Linear and quadratic model fits showing the relationship between age and the ω parameter, with 95% confidence intervals.
(Q4) Finally, the two age groups compared - adolescents (high school students) and adults (university students) - differ not only in age but also in sociocultural and economic backgrounds. High school students are likely more homogenous in regional background (e.g., Beijing locals), while university students may be drawn from a broader geographic and socioeconomic pool. Additionally, differences in financial independence, family structure (e.g., single-child status), and social network complexity may systematically affect cooperative behavior and valuation of rewards. Although these factors are difficult to control fully, the authors should more explicitly address the extent to which their findings reflect biological development versus social and contextual influences.
We appreciate this comment. Indeed, adolescents (high school students) and adults (university students) differ not only in age but also in sociocultural and socioeconomic backgrounds. In our study, all participants were recruited from Beijing and surrounding regions, which helps minimize large regional and cultural variability. Moreover, we accounted for individual-level random effects and included participants’ social value orientation (SVO) as an individual difference measure.
Nonetheless, we acknowledge that other contextual factors, such as differences in financial independence, socioeconomic status, and social experience—may also contribute to group differences in cooperative behavior and reward valuation. Although our results are broadly consistent with developmental theories of reward sensitivity and social decisionmaking, sociocultural influences cannot be entirely ruled out. Future work with more demographically matched samples or with socioeconomic and regional variables explicitly controlled will help clarify the relative contributions of biological and contextual factors. Accordingly, we have revised the Discussion to include the following statement:
“Third, although both age groups were recruited from Beijing and nearby regions, minimizing major regional and cultural variation, adolescents and adults may still differ in socioeconomic status, financial independence, and social experience. Such contextual differences could interact with developmental processes in shaping cooperative behavior and reward valuation. Future research with demographically matched samples or explicit measures of socioeconomic background will help disentangle biological from sociocultural influences.”
Reviewer #3 (Public review):
Summary:
Wu and colleagues find that in a repeated Prisoner's Dilemma, adolescents, compared to adults, are less likely to increase their cooperation behavior in response to repeated cooperation from a simulated partner. In contrast, after repeated defection by the partner, both age groups show comparable behavior.
To uncover the mechanisms underlying these patterns, the authors compare eight different models. They report that a social reward learning model, which includes separate learning rates for positive and negative prediction errors, best fits the behavior of both groups. Key parameters in this winning model vary with age: notably, the intrinsic value of cooperating is lower in adolescents. Adults and adolescents also differ in learning rates for positive and negative prediction errors, as well as in the inverse temperature parameter.
Strengths:
The modeling results are compelling in their ability to distinguish between learned expectations and the intrinsic value of cooperation. The authors skillfully compare relevant models to demonstrate which mechanisms drive cooperation behavior in the two age groups.
We thank the reviewer’s recognition of our work’s strengths.
Weaknesses:
(Q1) Some of the claims made are not fully supported by the data:
The central parameter reflecting preference for cooperation is positive in both groups. Thus, framing the results as self-interest versus other-interest may be misleading.
We thank the reviewer for this insightful comment. In the social reward model, the cooperation preference parameter is positive by definition, as defection in the repeated rPDG always yields a +2 monetary advantage regardless of the partner’s action. This positive value represents the additional subjective reward assigned to mutual cooperation (e.g., reciprocity value) that counterbalances the monetary gain from defection. Although the estimated social reward parameter ω was positive, the effective advantage of cooperation is Δ=p×ω−2. Given participants’ inferred beliefs p, Δ was negative for most trials (p×ω<2), indicating that the social reward was insufficient to offset the +2 advantage of defection. Thus, both adolescents and adults valued cooperation positively, but adolescents’ smaller ω and weaker responsiveness to sustained partner cooperation suggest a stronger weighting on immediate monetary payoffs.
In this light, our framing of adolescents as more self-interested derives from their behavioral pattern: even when they recognized sustained partner cooperation and held high expectations of partner cooperation, adolescents showed lower cooperative behavior and reciprocity rewards compared with adults. Whereas adults increased cooperation after two or three consecutive partner cooperations, this pattern was absent among adolescents. We therefore interpret their behavior as relatively more self-interested, reflecting reduced sensitivity to the social reward from mutual cooperation rather than a categorical shift from self-interest to other-interest, as elaborated in the Discussion.
(Q2) It is unclear why the authors assume adolescents and adults have the same expectations about the partner's cooperation, yet simultaneously demonstrate age-related differences in learning about the partner. To support their claim mechanistically, simulations showing that differences in cooperation preference (i.e., the w parameter), rather than differences in learning, drive behavioral differences would be helpful.
We thank the reviewer for raising this important point. In our model, both adolescents and adults updated their beliefs about partner cooperation using an asymmetric reinforcement learning (RL) rule. Although adolescents exhibited a higher positive and a lower negative learning rate than adults, the two groups did not differ significantly in their overall updating of partner cooperation probability (Fig. 4a-b). We then examined the social reward parameter ω, which was significantly smaller in adolescents and determined the intrinsic value of mutual cooperation (i.e., p×ω). This variable differed significantly between groups and closely matched the behavioral pattern.
Following the reviewer’s suggestion, we conducted additional simulations varying one model parameter at a time while holding the others constant. The difference in mean cooperation probability between adults and adolescents served as the index (positive = higher cooperation in adults). As shown in the Author response image 2, decreases in ω most effectively reproduced the observed group difference (shaded area), indicating that age-related differences in cooperation are primarily driven by variation in the social reward parameter ω rather than by others.
Author response image 2.
Simulation results showing how variations in each model parameter affect the group difference in mean cooperation probability (Adults – Adolescents). Based on the bestfitting Model 8 and parameters estimated from all participants, each line represents one parameter (i.e., α+, α-, ω, β) systematically varied within the tested range (α±:0.1–0.9; ω, β:1–9) while other parameters were held constant. Positive values indicate higher cooperation in adults. Smaller ω values most strongly reproduced the observed group difference, suggesting that reduced social reward weighting primarily drives adolescents’ lower cooperation.
(Q3) Two different schedules of 120 trials were used: one with stable partner behavior and one with behavior changing after 20 trials. While results for order effects are reported, the results for the stable vs. changing phases within each schedule are not. Since learning is influenced by reward structure, it is important to test whether key findings hold across both phases.
We thank the reviewer for this thoughtful and professional comment. In our GLMM and LMM analyses, we focused on trial order rather than explicitly including the stable vs. changing phase factor, due to concerns about multicollinearity. In our design, phases occur in specific temporal segments, which introduces strong collinearity with trial order. In multi-round interactions, order effects also capture variance related to phase transitions.
Nonetheless, to directly address this concern, we conducted additional robustness analyses by adding a phase variable (stable vs. changing) to GLMM1, LMM1, and LMM3 alongside the original covariates. Across these specifications, the key findings were replicated (see GLMM<sub>sup</sub>2 and LMM<sub>sup</sub>4–5; Tables 9-11), and the direction and significance of main effects remained unchanged, indicating that our conclusions are robust to phase differences.
(Q4) The division of participants at the legal threshold of 18 years should be more explicitly justified. The age distribution appears continuous rather than clearly split. Providing rationale and including continuous analyses would clarify how groupings were determined.
We thank the reviewer for this thoughtful comment. We divided participants at the legal threshold of 18 years for both conceptual and practical reasons grounded in prior literature and policy. In many countries and regions, 18 marks the age of legal majority and is widely used as the boundary between adolescence and adulthood in behavioral and clinical research. Empirically, prior studies indicate that psychosocial maturity and executive functions approach adult levels around this age, with key cognitive capacities stabilizing in late adolescence (Icenogle et al., 2019; Tervo-Clemmens et al., 2023). We have clarified this rationale in the Introduction section of the revised manuscript.
“Based on legal criteria for majority and prior empirical work, we adopt 18 years as the boundary between adolescence and adulthood (Icenogle et al., 2019; Tervo-Clemmens et al., 2023).”
We fully agree that the underlying age distribution is continuous rather than sharply divided. To address this, we conducted additional analyses treating age as a continuous predictor (see GLMM<sub>sup</sub>1 and LMM<sub>sup</sub>1–3; Tables S1-S4), which generally replicated the patterns observed with the categorical grouping. Nevertheless, given the limited age range of our sample, the generalizability of these findings to fine-grained developmental differences remains constrained. Therefore, our primary analyses continue to focus on the contrast between adolescents and adults, rather than attempting to model a full developmental trajectory.
(Q5) Claims of null effects (e.g., in the abstract: "adults increased their intrinsic reward for reciprocating... a pattern absent in adolescents") should be supported with appropriate statistics, such as Bayesian regression.
We thank the reviewer for highlighting the importance of rigor when interpreting potential null effects. To address this concern, we conducted Bayes factor analyses of the intrinsic reward for reciprocity and reported the corresponding BF10 for all relevant post hoc comparisons. This approach quantifies the relative evidence for the alternative versus the null hypothesis, thereby providing a more direct assessment of null effects. The analysis procedure is now described in the Methods and Materials section:
“Post hoc comparisons were conducted using Bayes factor analyses with MATLAB’s bayesFactor Toolbox (version v3.0, Krekelberg, 2024), with a Cauchy prior scale σ = 0.707.”
(Q6) Once claims are more closely aligned with the data, the study will offer a valuable contribution to the field, given its use of relevant models and a well-established paradigm.
We are grateful for the reviewer’s generous appraisal and insightful comments.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
(1) I commend the authors on a well-structured, clear, and interesting piece of work. I have several questions and recommendations that, if addressed, I believe will strengthen the manuscript.
We thank the reviewer for commending the organization of our paper.
(2) Introduction: - Why use a zero-sum (Prisoner's Dilemma; PD) versus a mixed-motive game (e.g. Trust Task) to study cooperation? In a finite set of rounds, the dominant strategy can be to defect in a PD.
We thank the reviewer for this helpful comment. We agree that both the rationale for using the repeated Prisoner’s Dilemma (rPDG) and the limitations of this framework should be clarified. We chose the rPDG to isolate the core motivational conflict between selfinterest and joint welfare, as its symmetric and simultaneous structure avoids the sequential trust and reputation dependencies/accumulation inherent to asymmetric tasks such as the Trust Game (King-Casas et al., 2005; Rilling et al., 2002).
Although a finitely repeated rPDG theoretically favors defection, extensive prior research shows that cooperation can still emerge in long repeated interactions when players rely on learning and reciprocity rather than backward induction (Rilling et al., 2002; Fareri et al., 2015). Our design employed 120 consecutive rounds, allowing participants to update expectations about partner behavior and to establish stable reciprocity patterns over time. We have added the following clarification to the Introduction:
“The rPDG provides a symmetric and simultaneous framework that isolates the motivational conflict between self-interest and joint welfare, avoiding the sequential trust and reputation dynamics characteristic of asymmetric tasks such as the Trust Game (Rilling et al., 2002; King-Casas et al., 2005)”
(3) Methods:
Did the participants know how long the PD would go on for?
Were the participants informed that the partner was real/simulated?
Were the participants informed that the partner was going to be the same for all rounds?
We thank the reviewer for the meticulous review work, which helped us present the experimental design and reporting details more clearly. the following clarifications: I. Participants were not informed of the total number of rounds in the rPDG. This prevented endgame expectations and avoided distraction from counting rounds, which could introduce additional effects. II. Participants were told that their partner was another human participant in the laboratory. However, the partner’s behavior was predetermined by a computer program. This design enabled tighter experimental control and ensured consistent conditions across age groups, supporting valid comparisons. III. Participants were informed that they would interact with the same partner across all rounds, aligning with the essence of a multiround interaction paradigm and stabilizing partner-related expectations. For transparency, we have clarified these points in the Methods and Materials section:
“Participants were told that their partner was another human participant in the laboratory and that they would interact with the same partner across all rounds. However, in reality, the actions of the partner were predetermined by a computer program. This setup allowed for a clear comparison of the behavioral responses between adolescents and adults. Participants were not informed of the total number of rounds in the rPDG.”
(4) The authors mention that an SVO was also recorded to indicate participant prosociality. Where are the results of this? Did this track game play at all? Could cooperativeness be explained broadly as an SVO preference that penetrated into game-play behaviour?
We thank the reviewer for pointing this out. We agree that individual differences in prosociality may shape cooperative behavior, so we conducted additional analyses incorporating SVO. Specifically, we extended GLMM1 and LMM3 by adding the measured SVO as a fixed effect with random slopes, yielding GLMM<sub>sup</sub>3 and LMM<sub>sup</sub>6 (Tables 12–13). The results showed that higher SVO was associated with greater cooperation, whereas its effect on the reward for reciprocity was not significant. Importantly, the primary findings remained unchanged after controlling for SVO. These results indicate that cooperativeness in our task cannot be explained solely by a broad SVO preference, although a more prosocial orientation was associated with greater cooperation. We have reported these analyses and results in the Appendix Analysis section.
(5) Why was AIC chosen rather an BIC to compare model dominance?
Sorry for the lack of clarification. Both the Akaike Information Criterion (AIC, Akaike, 1974) and Bayesian Information Criterion (BIC, Schwarz, 1978) are informationtheoretic criterions for model comparison, neither of which depends on whether the models to be compared are nested to each other or not (Burnham et al., 2002). We have added the following clarification into the Methods.
“We chose to use the AICc as the metric of goodness-of-fit for model comparison for the following statistical reasons. First, BIC is derived based on the assumption that the “true model” must be one of the models in the limited model set one compares (Burnham et al., 2002; Gelman & Shalizi, 2013), which is unrealistic in our case. In contrast, AIC does not rely on this unrealistic “true model” assumption and instead selects out the model that has the highest predictive power in the model set (Gelman et al., 2014). Second, AIC is also more robust than BIC for finite sample size (Vrieze, 2012).”
(6) I believe the model fitting procedure might benefit from hierarchical estimation, rather than maximum likelihood methods. Adolescents in particular seem to show multiple outliers in a^+ and w^+ at the lower end of the distributions in Figure S2. There are several packages to allow hierarchical estimation and model comparison in MATLAB (which I believe is the language used for this analysis;
see https://journals.plos.org/ploscompbiol/article?id=10.1371/journal.pcbi.1007043).
We thank the reviewer for this helpful comment and for referring us to relevant methodological work (Piray et al., 2019). We have addressed this point by incorporating hierarchical Bayesian estimation, which effectively mitigates outlier effects and improves model identifiability. The results replicated those obtained with MLE fitting and further revealed group-level differences in key parameters. Please see our detailed response to Reviewer#1 Q1 for the full description of this analysis and results.
(7) Results: Model confusion seems to show that the inequality aversion and social reward models were consistently confused with the baseline model. Is this explained or investigated? I could not find an explanation for this.
The apparent overlap between the inequality aversion (Model 4) and social reward (Model 5) models in the recovery analysis likely arises because neither model includes a learning mechanism, making them unable to capture trial-by-trial adjustments in this dynamic task. Consequently, both were best fit by the baseline model. Please see Response to Reviewer #1 Q3 for related discussion.
(8) Figures 3e and 3f show the correlation between asymmetric learning rates and age. It seems that both a^+ and a^- are around 0.35-0.40 for young adolescents, and this becomes more polarised with age. Could it be that with age comes an increasing discernment of positive and negative outcomes on beliefs, and younger ages compress both positive and negative values together? Given the higher stochasticity in younger ages (\beta), it may also be that these values simply represent higher uncertainty over how to act in any given situation within a social context (assuming the differences in groups are true).
We appreciate this insightful interpretation. Indeed, both α+ and α- cluster around 0.35–0.40 in younger adolescents and become increasingly polarized with age, suggesting that sensitivity to positive versus negative feedback is less differentiated early in development and becomes more distinct over time. This interpretation remains tentative and warrants further validation. Based on this comment, we have revised the Discussion to include this developmental interpretation.
We also clarify that in our model β denotes the inverse temperature parameter; higher β reflects greater choice precision and value sensitivity, not higher stochasticity. Accordingly, adolescents showed higher β values, indicating more value-based and less exploratory choices, whereas adults displayed relatively greater exploratory cooperation. These group differences were also replicated using hierarchical Bayesian estimation (see Response to Reviewer #1 Q1). In response to this comment, we have added a statement in the Discussion highlighting this developmental interpretation.
“Together, these findings suggest that the differentiation between positive and negative learning rates changes with age, reflecting more selective feedback sensitivity in development, while higher β values in adolescents indicate greater value sensitivity. This interpretation remains tentative and requires further validation in future research.”
(9) A parameter partial correlation matrix (off-diagonal) would be helpful to understand the relationship between parameters in both adolescents and adults separately. This may provide a good overview of how the model properties may change with age (e.g. a^+'s relation to \beta).
We thank the reviewer for this helpful comment. We fully agree that a parameter partial correlation matrix can further elucidate the relationships among parameters. Accordingly, we conducted a partial correlation analysis and added the visually presented results to the revised manuscript as Figure 2-figure supplement 4.
(10) It would be helpful to have Bayes Factors reported with each statistical tests given that several p-values fall within the 0.01 and 0.10.
We thank the reviewer for this important recommendation. We have conducted Bayes factor analyses and reported BF10 for all relevant post hoc comparisons. We also clarified our analysis in the Methods and Materials section:
“Post hoc comparisons were conducted using Bayes factor analyses with MATLAB’s bayesFactor Toolbox (version v3.0, Krekelberg, 2024), with a Cauchy prior scale σ = 0.707.”
(11) Discussion: I believe the language around ruling out failures in mentalising needs to be toned down. RL models do not enable formal representational differences required to assess mentalising, but they can distinguish biases in value learning, which in itself is interesting. If the authors were to show that more complex 'ToM-like' Bayesian models were beaten by RL models across the board, and this did not differ across adults and adolescents, there would be a stronger case to make this claim. I think the authors either need to include Bayesian models in their comparison, or tone down their language on this point, and/or suggest ways in which this point might be more thoroughly investigated (e.g., using structured models on the same task and running comparisons: https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0087619).
We thank the reviewer for the comments. Please see our response to Reviewer 1 (Appraisal & Discussion section) for details.
Reviewer #2 (Recommendations for the authors):
(1) The authors may want to show the winning model earlier (perhaps near the beginning of the Results section, when model parameters are first mentioned).
We thank the reviewer for this suggestion. We agree that highlighting the winning model early improves clarity. Currently, we have mentioned the winning model before the beginning of the Results section. Specifically, in the penultimate paragraph of the Introduction we state:
“We identified the asymmetric RL learning model as the winning model that best explained the cooperative decisions of both adolescents and adults.”
Reviewer #3 (Recommendations for the authors):
(1) In addition to the points mentioned above, I suggest the following:
Clarify plots by clearly explaining each variable. In particular, the indices 1 vs. 1,2 vs 1,2,3 were not immediately understandable.
We thank the reviewer for this suggestion. We agree that the indices were not immediately clear. We have revised the figure captions (Figure 1 and 4) to explicitly define these terms more clearly:
“The x-axis represents the consistency of the partner’s actions in previous trials (t<sub>−1</sub>: last trial; t<sub>−1,2</sub>: last two trials;<sub>t−1,2,3</sub>: last three trials).”
(2) It's unclear why the index stops at 3. If this isn't the maximum possible number of consecutive cooperation trials, please consider including all relevant data, as adolescents might show a trend similar to adults over more trials.
We thank the reviewer for raising this point. In our exploratory analyses, we also examined longer streaks of consecutive partner cooperation or defection (up to four or five trials). Two empirical considerations led us to set the cutoff at three in the final analyses. First, the influence of partner behavior diminished sharply with temporal distance. In both GLMMs and LMMs, coefficients for earlier partner choices were small and unstable, and their inclusion substantially increased model complexity and multicollinearity. This recency pattern is consistent with learning and decision models emphasizing stronger weighting of recent evidence (Fudenberg & Levine, 2014; Fudenberg & Peysakhovich, 2016). Second, streaks longer than three were rare, especially among some participants, leading to data sparsity and inflated uncertainty. Including these sparse conditions risked biasing group estimates rather than clarifying them. Balancing informativeness and stability, we therefore restricted the index to three consecutive partner choices in the main analyses, which we believe sufficiently capture individuals’ general tendencies in reciprocal cooperation.
(3) The term "reciprocity" may not be necessary. Since it appears to reflect a general preference for cooperation, it may be clearer to refer to the specific behavior or parameter being measured. This would also avoid confusion, especially since adolescents do show negative reciprocity in response to repeated defection.
We thank you for this comment. In our work, we compute the intrinsic reward for reciprocity as p × ω, where p is the partner cooperation expectation and ω is the cooperation preference. In the rPDG, this value framework manifests as a reciprocity-derived reward: sustained mutual cooperation maximizes joint benefits, and the resulting choice pattern reflects a value for reciprocity, contingent on the expected cooperation of the partner. This quantity enters the trade-off between U<sub>cooperation</sub> and U<sub>defection</sub> and captures the participant’s intrinsic reward for reciprocity versus the additional monetary reward payoff of defection. Therefore, we consider the term “reciprocity” an acceptable statement for this construct.
(4) Interpretation of parameters should closely reflect what they specifically measure.
We thank the reviewer for pointing this out. We have refined the relevant interpretations of parameters in the current Results and Discussion sections.
(5) Prior research has shown links between Theory of Mind (ToM) and cooperation (e.g., Martínez-Velázquez et al., 2024). It would be valuable to test whether this also holds in your dataset.
We thank the reviewer for this thoughtful comment. Although we did not directly measure participants’ ToM, our design allowed us to estimate participants’ trial-by-trial inferences (i.e., expectations) about their partner’s cooperation probability. We therefore treat these cooperation expectations as an indirect representation for belief inference, which is related to ToM processes. To test whether this belief-inference component relates to cooperation in our dataset, we further conducted an exploratory analysis (GLMM<sub>sup</sub>4) in which participants’ choices were regressed on their cooperation expectations, group, and the group × cooperation-expectation interaction, controlling for trial number and gender, with random effects. Consistent with the ToM–cooperation link in prior research (MartínezVelázquez et al., 2024), participants’ expectations about their partner’s cooperation significantly predicted their cooperative behavior (Table 14), suggesting that decisions were shaped by social learning about others’ inferred actions. Moreover, the interaction between group and cooperation expectation was not significant, indicating that this inference-driven social learning process likely operates similarly in adolescents and adults. This aligns with our primary modeling results showing that both age groups update beliefs via an asymmetric learning process. We have reported these analyses in the Appendix Analysis section.
(6) More informative table captions would help the reader. Please clarify how variables are coded (e.g., is female = 0 or 1? Is adolescent = 0 or 1?), to avoid the need to search across the manuscript for this information.
We thank the reviewer for raising this point. We have added clear and standardized variable coding in the table notes of all tables to make them more informative and avoid the need to search the paper. We have ensured consistent wording and formatting across all tables.
(7) I hope these comments are helpful and support the authors in further strengthening their manuscript.
We thank the three reviewers for their comments, which have been helpful in strengthening this work.
References
(1) Fudenberg, D., & Levine, D. K. (2014). Recency, consistent learning, and Nash equilibrium. Proceedings of the National Academy of Sciences of the United States of America, 111(Suppl. 3), 10826–10829. https://doi.org/10.1073/pnas.1400987111.
(2) Fudenberg, D., & Peysakhovich, A. (2016). Recency, records, and recaps: Learning and nonequilibrium behavior in a simple decision problem. ACM Transactions on Economics and Computation, 4(4), Article 23, 1–18. https://doi.org/10.1145/2956581
(3) Hackel, L., Doll, B., & Amodio, D. (2015). Instrumental learning of traits versus rewards: Dissociable neural correlates and effects on choice. Nature Neuroscience, 18, 1233– 1235. https://doi.org/10.1038/nn.4080
(4) Icenogle, G., Steinberg, L., Duell, N., Chein, J., Chang, L., Chaudhary, N., Di Giunta, L., Dodge, K. A., Fanti, K. A., Lansford, J. E., Oburu, P., Pastorelli, C., Skinner, A. T.Sorbring, E., Tapanya, S., Uribe Tirado, L. M., Alampay, L. P., Al-Hassan, S. M.,Takash, H. M. S., & Bacchini, D. (2019). Adolescents’ cognitive capacity reaches adult levels prior to their psychosocial maturity: Evidence for a “maturity gap” in a multinational, cross-sectional sample. Law and Human Behavior, 43(1), 69–85. https://doi.org/10.1037/lhb0000315
(5) Krekelberg, B. (2024). Matlab Toolbox for Bayes Factor Analysis (v3.0) [Computer software]. Zenodo. https://doi.org/10.5281/zenodo.13744717
(6) Martínez-Velázquez, E. S., Ponce-Juárez, S. P., Díaz Furlong, A., & Sequeira, H. (2024). Cooperative behavior in adolescents: A contribution of empathy and emotional regulation? Frontiers in Psychology, 15,1342458. https://doi.org/10.3389/fpsyg.2024.1342458
(7) Tervo-Clemmens, B., Calabro, F. J., Parr, A. C., et al. (2023). A canonical trajectory of executive function maturation from adolescence to adulthood. Nature Communications, 14, 6922. https://doi.org/10.1038/s41467-023-42540-8
(8) King-Casas, B., Tomlin, D., Anen, C., Camerer, C. F., Quartz, S. R., & Montague, P. R. (2005). Getting to know you: reputation and trust in a two-person economic exchange. Science, 308(5718), 78-83. https://doi.org/10.1126/science.1108062
(9) Rilling, J. K., Gutman, D. A., Zeh, T. R., Pagnoni, G., Berns, G. S., & Kilts, C. D. (2002).A neural basis for social cooperation. Neuron, 35(2), 395-405. https://doi.org/10.1016/s0896-6273(02)00755-9
(10) Fareri, D. S., Chang, L. J., & Delgado, M. R. (2015). Computational substrates of social value in interpersonal collaboration. Journal of Neuroscience, 35(21), 8170-8180. https://doi.org/10.1523/JNEUROSCI.4775-14.2015
(11) Akaike, H. (2003). A new look at the statistical model identification. IEEE transactions on automatic control, 19(6), 716-723. https://doi.org/10.1109/TAC.1974.1100705
(12) Schwarz, G. (1978). Estimating the dimension of a model. The annals of statistics, 461464. https://doi.org/10.1214/aos/1176344136
(13) Burnham, K. P., & Anderson, D. R. (2002). Model selection and multimodel inference: A practical information-theoretic approach (2nd ed.). Springer.https://doi.org/10.1007/b97636
(14) Gelman, A., & Shalizi, C. R. (2013). Philosophy and the practice of Bayesian statistics. British Journal of Mathematical and Statistical Psychology, 66(1), 8–38. https://doi.org/10.1111/j.2044-8317.2011.02037.x
(15) Gelman, A., Carlin, J. B., Stern, H. S., Dunson, D. B., Vehtari, A., & Rubin, D. B. (2014). Bayesian data analysis (3rd ed.). Chapman and Hall/CRC. https://doi.org/10.1201/b16018
(16) Vrieze, S. I. (2012). Model selection and psychological theory: A discussion of the differences between the Akaike Information Criterion (AIC) and the Bayesian Information Criterion (BIC). Psychological Methods, 17(2), 228–243. https://doi.org/10.1037/a0027127
-
-
www.france.tv www.france.tv
-
www.france.tv www.france.tv
-
Analyse Documentaire : Portrait d'un Père Désemparé et Destin de Jonathan Jensen
Résumé Analytique
Ce document synthétise le récit autobiographique et tragique d'un réalisateur de documentaires hanté par la disparition de son fils, Jonathan Jensen.
À travers le prisme de sa caméra, le père explore une relation marquée par une quête de masculinité héritée de modèles d'exploration classiques (comme Roald Amundsen) et pervertie par les influences numériques contemporaines (telles qu'Andrew Tate).
Jonathan, né en 2002 et décrit comme un enfant d'une énergie débordante, a fini par s'exiler au Brésil pour poursuivre une réussite matérielle rapide, influencé par des "coachs" prônant une vision binaire du monde (gagnants contre perdants).
Ce parcours s'est achevé brutalement par son assassinat à l'âge de 21 ans, commis par son associé et ami d'enfance, Oscar Milander.
Le documentaire met en lumière l'échec d'un père à protéger son fils contre une réalité numérique toxique, malgré ses tentatives répétées de forger son caractère à travers des expériences physiques et des projets cinématographiques.
--------------------------------------------------------------------------------
I. La Dynamique Père-Fils : La Caméra comme Filtre et Lien
La relation entre le narrateur et Jonathan est intrinsèquement liée à la profession du père. Le cinéma documentaire a servi à la fois de pont et de barrière protectrice.
• Le filtre protecteur : Le père admet avoir passé la majeure partie de son temps avec son fils une caméra à la main, l'utilisant comme un « filtre protecteur » entre eux deux.
• L'héritage de l'absence : Le narrateur attribue ses propres lacunes paternelles à l'absence de son propre père, capitaine de navires de croisière qu'il n'a rencontré qu'une seule fois à l'âge de 30 ans.
• La documentation du lien : Des moments clés ont été filmés, notamment un incident où Jonathan a perdu l'usage de ses jambes le lendemain d'une soirée passée seul avec son père, un événement qui a paradoxalement renforcé leur lien malgré l'incompétence paternelle ressentie.
II. Modèles de Masculinité et Quête de Caractère
Le père a tenté d'inculquer à Jonathan des valeurs de force et de discipline, souvent basées sur des figures historiques ou des expériences extrêmes.
L'Influence de Roald Amundsen
Le narrateur voue une obsession à l'explorateur polaire Amundsen, symbole de conquête et de virilité.
• L'expédition manquée : Le père a tenté d'organiser une expédition polaire avec Jonathan pour reproduire (et réussir) une traversée qu'Amundsen avait échouée en 1893.
• Échec de la transmission : Jonathan a rejeté cette épreuve physique, préférant le confort et la socialisation, qualifiant le projet d'« absurde ».
L'Expérience Militaire
Inquiet du manque de discipline de son fils, le père l'a poussé à intégrer un camp militaire pour jeunes.
• Objectif : Enseigner la rigueur et la discipline.
• Résultat : Bien que Jonathan ait semblé enthousiaste au début, le père a rapidement regretté cette décision, percevant une perte de contact avec la nature enfantine de son fils.
III. La Rupture et la Dérive vers la "Matrice" Numérique
À sa majorité, Jonathan s'est radicalement éloigné des valeurs de ses parents pour poursuivre une vie d'entrepreneur numérique, influencé par des idéologies de succès agressif.
• Le départ pour le Brésil : En pleine pandémie de COVID-19, Jonathan et son ami Mons (Oscar Milander) se sont installés au Brésil après que Jonathan a vidé ses comptes d'épargne (30 000 $).
• L'influence d'Andrew Tate : Jonathan a adopté le discours d'Andrew Tate, divisant le monde en "gagnants" et "perdants" et affirmant vouloir « s'échapper de la Matrice ».
• Changement de personnalité : Son petit ami brésilien, Gustavo (un influenceur majeur), a témoigné de la transformation inquiétante de Jonathan sous l'influence de "coachs" psychopathes, passant d'un jeune homme amoureux à une personne froide, homophobe et obsédée par l'argent.
IV. Le Conflit Financier et Idéologique
La tension entre le père et le fils a atteint son paroxysme autour de la gestion de l'argent et de la vision du futur.
| Point de Conflit | Perspective du Père | Perspective de Jonathan | | --- | --- | --- | | Argent | Épargne destinée à sécuriser l'avenir (25 ans). | Capital nécessaire immédiatement pour le business. | | Méthode | Patience, études, vie de classe moyenne. | "Hustle", prise de risque, succès immédiat. | | Vision du monde | Empathie et connexion émotionnelle. | Domination, élimination de la faiblesse. |
Jonathan accusait son père de vouloir le freiner par "peur" et de ne pas comprendre les règles du succès moderne.
V. Le Dénouement Tragique
Malgré un bref retour en Norvège où Jonathan a semblé retrouver une certaine sérénité auprès de son père, il est reparti pour le Brésil, affirmant que son héritage continuerait à travers ses actions.
• Le meurtre : Onze jours après son retour au Brésil, Jonathan Jensen a été retrouvé mort à Balneário Camboriú, poignardé à mort.
• Le coupable : Oscar Milander (Mons), son meilleur ami et associé, a été arrêté alors qu'il fuyait au volant de la Mercedes de la victime.
• Conséquences : Le père décrit une douleur omniprésente, une « pression dans chaque cellule » de son être, réalisant que le rêve de succès de son fils s'est transformé en un cauchemar de "logistique d'horreur".
--------------------------------------------------------------------------------
Citations Clés
« La connexion est rompue. Il n'est plus là. Jonathan, mon beau garçon, est mort. » — Le Père
« Tu ne peux pas te promener et aimer tout le monde, parce que ça finit mal. On a son cercle, comme une famille. » — Jonathan (discutant de l'amour)
« En tant que père, si tu as bien fait ton travail, ta vie continue dans ton fils... Tu crées une meilleure version de toi-même. » — Jonathan (quelques jours avant sa mort)
« Je m'échappe de la Matrice. » — Jonathan
-
-
www.france.tv www.france.tv
-
Éduquons nos fils : Synthèse sur la Déconstruction de la Masculinité et de la Violence
Résumé Exécutif
Ce document analyse les mécanismes de construction de la masculinité et leurs conséquences sur la société française, tels qu'exposés dans les témoignages et analyses du projet "Infrarouge - Éduquons nos fils".
Le constat central est que la violence n'est pas une essence masculine, mais le produit d'un conditionnement éducatif et social profond.
Les points clés sont les suivants :
• Le poids des stéréotypes : Dès l'enfance, les garçons sont incités à refouler leur vulnérabilité, assimilée à une faiblesse, pour incarner le modèle du "mâle alpha" invulnérable.
• Un coût sociétal colossal : La "virilité" coûte environ 100 milliards d'euros par an à l'État français (justice, santé, insécurité routière).
• La masculinité comme facteur de risque : Les hommes représentent 97 % de la population carcérale et sont responsables de 84 % des accidents de la route mortels. Ils sont également les premières victimes de leur propre conditionnement (morts prématurées, conduites à risques).
• L'urgence éducative : La solution réside dans une réinvention de l'éducation des fils, passant par l'autorisation de l'expression émotionnelle, la déconstruction des modèles de domination et une présence paternelle sécurisante.
--------------------------------------------------------------------------------
1. La Construction du "Mâle Alpha" : Un Conditionnement Précoce
Le document met en lumière un système éducatif et social qui formate les garçons selon des codes de virilité hégémonique.
L'injonction à l'invulnérabilité
• Refoulement émotionnel : L'éducation traditionnelle enseigne que "pleurer est une affaire de filles".
Ce blocage des émotions, de l'enfance à l'âge adulte, génère une souffrance psychique et physique (douleurs somatiques, burn-out).
• L'image de la force : Le modèle valorisé est celui de "Superman" : fort, musclé, insensible et protecteur.
Le culte du corps (musculation à outrance) est souvent utilisé comme une armure pour pallier une insécurité intérieure ou une vulnérabilité passée.
• Modèles culturels : Les figures d'identification des années 80 et 90 (Stallone, Schwarzenegger, James Bond) ont ancré l'image de l'homme "sandwich de l'hypervirilité", où la réussite passe par la puissance physique et la domination.
L'influence de l'environnement social
• Le groupe de pairs (la "meute") : La bande de copains joue un rôle de renforcement. La peur d'être exclu ou jugé "faible" pousse les jeunes hommes à la surenchère dans la prise de risque ou la violence.
• Le sexisme ordinaire : La publicité et l'espace public ont longtemps objectivé les femmes, renforçant chez les jeunes garçons une vision prédatrice ou supérieure du masculin.
--------------------------------------------------------------------------------
2. Les Conséquences de la Virilité : Statistiques et Coûts
L'adhésion aux stéréotypes masculins n'est pas seulement un enjeu privé ; elle impacte lourdement la structure socio-économique du pays.
Le coût financier de la virilité (Analyse de Lucile Petavin)
L'historienne Lucile Petavin estime que les comportements antisociaux et violents des hommes coûtent 100 milliards d'euros par an à la France.
| Secteur de dépense | Coût estimé (par an) | | --- | --- | | Justice et forces de l'ordre | 7 milliards € (sur 9 au total) | | Coups et violences volontaires | 18 milliards € (sur 26 au total) | | Insécurité routière | 13,3 milliards € | | Total Global estimé | 100 milliards € |
L'asymétrie de genre dans la criminalité et les risques
• Incarcération : 97 % des personnes détenues sont des hommes.
• Sécurité routière : Les hommes sont responsables de 84 % des accidents mortels.
• Santé : Les hommes ont 2 à 3 fois plus de risques de mourir prématurément (avant 65 ans) à cause de comportements à risques (alcool, tabac, vitesse). À 14 ans, un garçon a déjà 70 % de risques de plus qu'une fille de mourir dans un accident.
--------------------------------------------------------------------------------
3. Les Espaces de Reproduction et de Mutation
La masculinité se joue dans des lieux clés où les stéréotypes sont soit renforcés, soit déconstruits.
L'école et la cour de récréation
L'école est décrite comme un concentré de patriarcat où règnent la compétition et la loi du plus fort.
• Violences scolaires : 71 % des élèves de la 6ème à la terminale ont subi des violences.
• Stéréotypes sportifs : Les activités comme la danse classique ou l'équitation restent perçues comme "féminines", tandis que le foot et la boxe sont vus comme le domaine naturel des garçons. Des initiatives pédagogiques tentent de briser ces barrières par la mixité réelle dans l'effort sportif.
Le danger du numérique et du masculinisme
Les réseaux sociaux (TikTok, Instagram) servent de vecteurs à des discours de haine misogyne extrêmes.
• Contenus toxiques : Des influenceurs militent pour un retour au patriarcat pur et dur, prônant la soumission des femmes et la violence comme outil de respect.
• Absence de modération : Les plateformes sont critiquées pour leur manque de régulation, privilégiant le profit généré par l'engagement (vues/likes) au détriment de la sécurité des mineurs.
--------------------------------------------------------------------------------
n4. Vers une Nouvelle Paternité et Responsabilité
Le changement passe par une redéfinition du rôle de père et une prise de conscience collective des hommes.
Le rôle du père "nouveau"
• Expression de l'affection : Dire "je t'aime" et assurer une sécurité émotionnelle sans menace physique est présenté comme le levier principal pour briser le cycle de la violence.
• Implication domestique : Sortir du cliché "l'homme au barbecue, la femme aux légumes" et s'impliquer dans le soin quotidien des enfants (le "care") permet de normaliser une masculinité attentive et non dominante.
La responsabilité collective (All Men)
Le champion de MMA Cédric Dumbé souligne que si tous les hommes ne sont pas coupables, ils sont tous responsables du climat social.
• Sortir de l'indifférence : Refuser de rire aux blagues sexistes, ne pas relayer de contenus dégradants et soutenir activement les victimes de violences sont des actes de "vrais hommes".
• Légitimation de la vulnérabilité : Accepter sa propre part de féminité et ses émotions est la clé pour une virilité "réglée" qui ne débouche pas sur l'agression.
--------------------------------------------------------------------------------
Conclusion
Le document conclut sur la nécessité d'une transformation profonde de l'éducation.
En autorisant les fils à explorer toute la gamme des émotions humaines et en leur offrant des modèles de force qui ne reposent pas sur la domination, la société peut espérer réduire drastiquement la violence masculine et ses coûts humains et financiers.
Comme l'indique l'un des intervenants : "Tous les hommes ne sont pas violents, mais la violence est masculine. Éduquez vos fils."
-
-
viewer.athenadocs.nl viewer.athenadocs.nl
-
residuele schizofrenie
wat na een episode komt
-
-
shs-cairn-info.ezproxy.u-paris.fr shs-cairn-info.ezproxy.u-paris.fr
-
Fabre souligne la spécificité et l’opportunité “d’un langage fondé sur l’image” pour exprimer/métaphoriser puis symboliser les vécus archaïques
collage
-
-
www.youtube.com www.youtube.com
-
Synthèse du Documentaire : « Didier, moi et les autres... les enfants du silence »
Résumé Exécutif
Ce document synthétise les témoignages et les analyses issus du documentaire de LCP consacré aux victimes masculines de violences sexuelles durant l'enfance.
Le récit s'articule autour de quatre hommes — Nicolas, Didier, Arnaud et Adrien — qui, après des décennies de silence, tentent de reconstruire leur identité fracturée.
Les thèmes centraux incluent le mécanisme de l'amnésie traumatique, l'impact dévastateur du déni familial, et l'utilisation de l'image ou du militantisme comme outils de résilience.
Le document souligne une réalité systémique : 160 000 enfants sont victimes de violences sexuelles chaque année en France, soit un enfant toutes les trois minutes, souvent dans une indifférence ou une impuissance collective.
--------------------------------------------------------------------------------
I. La Mécanique du Silence et du Traumatisme
Le documentaire explore comment le silence devient une stratégie de survie pour l'enfant victime, se transformant en une « expertise » qui dure parfois des décennies.
1. L'Amnésie Traumatique et la Dissociation
L'amnésie traumatique est présentée non comme un oubli volontaire, mais comme une réponse biologique à une horreur indicible.
• Processus biologique : Face à une charge insupportable d'adrénaline et de cortisol sécrétée par l'amygdale, le cerveau « disjoncte » pour protéger l'individu d'une mort potentielle par peur.
• Conséquences : Cette déconnexion empêche la mémoire d'encoder l'événement.
La victime est alors incapable de faire le récit de l'agression, ce qui peut être interprété à tort par l'entourage comme du mensonge ou de l'indifférence.
• La Réviviscence : Le retour des souvenirs est décrit comme un séisme.
Pour Adrien, cela s'est manifesté par des douleurs somatiques (« des fourmis sous la peau ») avant que les images ne se recomposent comme une mosaïque 19 ans plus tard.
2. La Confusion et la Culpabilité
La question du plaisir physiologique durant l'agression est abordée comme un « tabou dans le tabou ».
• Réaction somatique vs Consentement : Les victimes clarifient qu'une réaction physique à une stimulation n'est pas du désir.
• Outil de destruction : Cette confusion est souvent utilisée par l'agresseur pour silencer la victime ou par la victime elle-même pour s'auto-accuser, renforçant la conviction d'avoir été « consentant ».
--------------------------------------------------------------------------------
II. Le Rôle de l'Entourage : Entre Déni et Impuissance
La réaction des parents et de la famille constitue souvent une « double peine » pour la victime.
| Type de Réaction | Description et Citations Clés | | --- | --- | | Le Déni Total | La mère de Nicolas refuse de croire les faits : « Si ce que tu as vécu est vrai, tu me l'aurais dit quand tu avais neuf ans. Donc c'est pas vrai. » | | La Minimisation | La mère de Didier, bien que croyante, incite à l'oubli : « Didier, allez, c'est bon... il faut oublier. » | | L'Incapacité d'Agir | Les parents de Nicolas expliquent leur silence par la prescription légale : « On ne pouvait plus rien faire parce qu'il y a péremption. » | | L'Exclusion | Arnaud relate avoir été exclu des réunions familiales au profit de ses agresseurs (ses cousins) : « Ils préféraient avoir mes cousins agresseurs à la table, plutôt que moi. » |
--------------------------------------------------------------------------------
III. Parcours de Résilience : Transformer la Douleur
Chaque victime développe sa propre méthode pour « recoller les morceaux » de son identité lézardée.
1. La Maîtrise par l'Image (Didier et Nicolas)
Pour Didier et Nicolas, devenir caméraman ou photographe n'est pas un hasard.
C'est une manière de « cadrer le monde » pour y mettre de l'ordre face au chaos intérieur.
• Le Cadre : « Un truc qui borne, qui contient, qui arrête un peu le bordel. »
• Le Négatif et la Lumière : Didier utilise la métaphore photographique : faire émerger la lumière à partir du noir pour « recoller le trou noir » de sa vie.
2. Le Militantisme comme Arme (Arnaud)
Arnaud a transformé sa souffrance en un combat public et tranchant.
• La Voix des Victimes : Il dénonce les institutions (notamment l'Église et l'enseignement catholique) et réclame « Vérité, Justice, Réparation, Prévention ».
• La Grenade Dégoupillée : Nicolas décrit les victimes qui parlent comme des « grenades dégoupillées » que la société préfère éviter car elles sont « radioactives ».
--------------------------------------------------------------------------------
IV. La Quête de Justice et la Confrontation
Le documentaire souligne le besoin de reconnaissance, même lorsque l'action judiciaire est entravée par la prescription.
• La Traque de l'Agresseur : Didier a passé des années à traquer son agresseur, le filmant à son insu comme un « exutoire ».
Son but ultime était qu'il sache qu'une plainte existait, même pour des faits prescrits depuis 50 ans.
• L'Échec de la Confrontation Physique : Une tentative de Didier pour confronter son agresseur se solde par une méprise sur l'identité de l'homme, illustrant l'obsession de « faire cesser le cirque » et le besoin désespéré de conclusion.
• L'Évolution Judiciaire : Le Garde des Sceaux a sollicité les procureurs pour enquêter sur des faits prescrits afin de permettre une forme de reconnaissance officielle, bien que l'accès à ces enquêtes reste difficile pour les victimes.
--------------------------------------------------------------------------------
V. Conclusions et Perspectives
Le traumatisme des violences sexuelles est décrit comme une « effraction à vie ». La guérison ne signifie pas l'effacement du souvenir, mais l'apprentissage d'une vie avec la cicatrice.
• L'Importance de la Parole : Témoigner permet de briser le cycle de l'abandon et d'aider d'autres hommes à sortir du silence.
• Un Enjeu Collectif : Le documentaire conclut que la protection des enfants n'est pas seulement une affaire privée mais un devoir sociétal. Comme le souligne Adrien, il s'agit de sortir du « cadre limitant » du passé pour retrouver une place saine dans la société.
« On ne tourne jamais la page. Je pense qu'on vit avec. » — Didier
Tags
Annotators
URL
-
-
www.youtube.com www.youtube.com
-
Manifeste de l'hostilité misogyne : Analyse de la haine systémique contre les femmes dans l'espace public
Résumé Exécutif
Le présent document synthétise les mécanismes, les acteurs et les conséquences de la recrudescence de la violence misogyne, telle qu'analysée dans le contexte européen et international.
Il apparaît que les femmes occupant des postes à responsabilité ou s'exprimant avec conviction dans la sphère publique — politiciennes, journalistes, activistes — sont les cibles privilégiées d'une haine organisée.
Cette hostilité se manifeste par un cyber-harcèlement d'une violence extrême, allant de l'insulte sexiste aux menaces de viol et de mort.
L'analyse révèle que cette haine n'est pas fortuite : elle est instrumentalisée par des mouvements populistes et d'extrême droite, et théorisée par des groupes masculinistes qui perçoivent l'égalité des genres comme une menace identitaire et sociale.
Le monde numérique, régi par des algorithmes privilégiant l'engagement (souvent généré par la haine) et protégé par un certain vide juridique, sert d'incubateur à une radicalisation pouvant mener au terrorisme (mouvement « Incel »).
Face à l'insuffisance des réponses institutionnelles, des initiatives de soutien émergent, tandis que des voix s'élèvent pour appeler à une déconstruction systémique des stéréotypes de masculinité.
--------------------------------------------------------------------------------
1. Une Violence Ciblée et Systémique
La haine envers les femmes n'est pas le fait d'individus isolés, mais un phénomène structurel visant à exclure les femmes de la sphère publique.
Les cibles privilégiées
Le mépris et l'agressivité visent particulièrement les femmes qui manifestent de l'assurance et occupent des domaines traditionnellement masculins :
• Femmes politiques : En Allemagne (34 % de députées au Bundestag) comme en France (40 % à l'Assemblée nationale), elles subissent des interruptions systématiques et une remise en question de leur légitimité.
• Journalistes sportives : L'exemple de Claudia Neumann illustre l'hostilité rencontrée lorsqu'une femme investit le "pré carré" masculin du football.
• Activistes et Écrivaines : Des figures comme Alice Barbe ou Leila Slimani subissent des campagnes de dénigrement massives en raison de leurs engagements pour les réfugiés ou les droits des femmes.
La nature des agressions
Contrairement aux hommes, les femmes reçoivent des attaques "taillées sur mesure" liées à leur genre :
• Sexualisation et humiliation : Envoi de "dickpics", insultes à caractère sexuel, commentaires sur le physique ou l'âge.
• Menaces extrêmes : Descriptions détaillées de viols, menaces de mutilations génitales ou de mort.
• Invisibilisation : Au sein même des institutions, la parole des femmes est souvent accueillie par un brouhaha ou un désintérêt feint de la part des collègues masculins.
--------------------------------------------------------------------------------
2. Les Racines Idéologiques et l'Instrumentalisation Politique
La misogynie moderne s'appuie sur des discours structurés et une exploitation politique délibérée.
Le mouvement masculiniste
Les masculinistes considèrent que les femmes ont pris le dessus et opprimeraient désormais les hommes.
• Rhétorique de la crise : Ils présentent l'égalité comme une "maladie" ou une "injustice" envers les jeunes hommes.
• Défense du patriarcat : Des auteurs comme Julien Rochedy prônent un retour aux codes virils traditionnels, affirmant que le manque de patriarcat est la cause du féminisme actuel.
L'extrême droite et le populisme
Des partis comme l'AfD en Allemagne ou des figures comme Éric Zemmour en France utilisent le sexisme comme stratégie électorale :
• Provocation délibérée : Utilisation de termes sexistes pour susciter l'émotion et gagner en visibilité médiatique.
• Lien avec d'autres haines : La misogynie s'accompagne souvent d'idéologies racistes et antisémites. Le féminisme est présenté comme un facteur de "décadence civilisationnelle".
• Déstabilisation : Au Parlement, l'entrée de partis d'extrême droite a entraîné une augmentation nette des exclamations et comportements sexistes.
--------------------------------------------------------------------------------
3. L'Espace Numérique : Un "Lieu de Terreur"
Internet et les réseaux sociaux agissent comme des catalyseurs de haine en raison de leur fonctionnement intrinsèque.
La mécanique du cyber-harcèlement
• L'anonymat : Il favorise la levée des inhibitions. Les agresseurs peuvent être des "monsieur tout le monde" (professeurs, ingénieurs, avocats).
• Les algorithmes : La haine et l'incitation à la violence génèrent plus de vues et de clics, ce qui est financièrement profitable pour les plateformes.
• La radicalisation en ligne : Des forums privés permettent à des hommes de s'enfermer dans une vision étriquée du monde, se confortant mutuellement dans leur détestation des femmes.
La menace "Incel" (Célibataires involontaires)
Cette sous-culture numérique représente un danger sécuritaire réel :
• Idéologie : Conviction que les femmes sont des êtres inférieurs et que l'accès à leur corps est un droit fondamental.
• Apologie de la violence : Célébration de terroristes comme Elliot Rodger ou Alec Minassian.
• Passage à l'acte : Plusieurs attentats (Santa Barbara, Toronto, Halle) ont été motivés, totalement ou partiellement, par cette haine des femmes.
--------------------------------------------------------------------------------
4. Conséquences Sociales et Réponses Institutionnelles
L'impact de cette violence dépasse les victimes individuelles pour menacer la démocratie elle-même.
L'impact sur les victimes et la démocratie
• Charge mentale et traumatisme : Les victimes doivent mettre en place des protocoles de sécurité lourds et subissent un stress psychologique intense.
• Le recul de la parole : Une femme sur deux craint d'exprimer son opinion en ligne. Ce retrait forcé crée un déséquilibre dans le débat public, laissant le champ libre aux harceleurs.
• Le sentiment d'isolement : Les militantes se sentent souvent seules face à des réseaux d'agresseurs très bien organisés.
Un cadre juridique et policier défaillant
• Sous-estimation des faits : Les plaintes sont souvent classées sans suite ou banalisées par les forces de l'ordre ("supprimez votre compte").
• Complexité législative : Les plateformes, souvent basées à l'étranger, collaborent peu avec la justice. En France, des tentatives de régulation ont été déclarées inconstitutionnelles au nom de la liberté d'expression.
• Initiatives de soutien : Des structures comme HateAid en Allemagne offrent désormais un accompagnement juridique et psychologique pour pallier les carences de l'État.
--------------------------------------------------------------------------------
5. Perspectives de Lutte et Déconstruction
Pour enrayer ce phénomène, l'analyse suggère des actions à la racine du problème.
| Axe d'intervention | Actions préconisées | | --- | --- | | Éducation | Déconstruire les stéréotypes de genre dès le plus jeune âge et remettre en question la "masculinité toxique". | | Engagement masculin | Impliquer les hommes dans le discours féministe. Comme le souligne Leila Slimani, les hommes ont tout à gagner à un monde moins violent et moins régi par l'obligation de virilité brute. | | Justice | Former la police et le parquet à reconnaître la misogynie comme un délit grave. Créer une jurisprudence forte, à l'instar du procès remporté par Alice Barbe. | | Régulation | Imposer des règles communes au niveau européen pour contraindre les plateformes à transmettre les données des auteurs de messages haineux. |
Conclusion : La lutte contre la misogynie organisée est un enjeu de sécurité publique et de survie démocratique.
Refuser de céder à la peur et briser le silence sont les premiers remparts contre cette tentative de "silencier" les femmes dans l'espace public.
Tags
Annotators
URL
-
-
www.youtube.com www.youtube.com
-
Masculinité en crise : la fin d’Homo Virilus ?
Ce document de synthèse analyse les fondements, l'évolution historique et la remise en question contemporaine de l'archétype de la virilité, désigné sous le terme d'« Homo Virilus ».
À travers un prisme historique, sociologique et personnel, il explore comment ce modèle de domination, longtemps hégémonique, fait face à une crise profonde et à une nécessité de redéfinition.
--------------------------------------------------------------------------------
Résumé exécutif
L’idéal de l'« Homo Virilus » repose sur un triptyque de puissance, de courage et de domination.
Historiquement ancré dans l'héroïsme guerrier, ce modèle a subi des chocs successifs : la déshumanisation par la guerre industrielle en 1914, l'autonomisation des femmes lors des conflits mondiaux, et les révolutions féministes des années 1970.
Aujourd'hui, la masculinité est à la croisée des chemins. D'un côté, une réaction masculiniste et « viriliste » s'intensifie, portée par la manosphère et des discours réactionnaires.
De l'autre, des mouvements de déconstruction émergent, portés par de nouvelles formes de paternité et une distinction claire entre masculinité et domination.
Le coût social de la virilité obligatoire est désormais quantifié (environ 100 milliards d'euros par an en France), soulignant l'urgence d'une transition vers des modèles plus équitables.
--------------------------------------------------------------------------------
1. L'Archétype d'Homo Virilus : Un Idéal de Domination
Le concept d'Homo Virilus n'est pas une donnée biologique, mais une construction culturelle héritée de l'Antiquité gréco-romaine.
• Les piliers de la virilité : Ce modèle exige puissance, courage et domination. Il impose une maîtrise de soi constante, tout en justifiant une violence explosive si nécessaire.
• Une hiérarchie sociale : La vertu virile justifie la domination des femmes et des enfants, mais aussi celle des hommes jugés « non virils ».
• Une fragilité intrinsèque : Contrairement à l'apparence de force, la virilité est décrite comme une « insoutenable fragilité » car elle doit être prouvée et maintenue en permanence. On ne naît pas Homo Virilus, on le devient par des rites de passage, souvent liés au combat.
--------------------------------------------------------------------------------
2. Les Ruptures Historiques : De la Tranchée au Foyer
Le XXe siècle a marqué le début du déclin de l'héroïsme viril traditionnel.
L'épreuve de la Grande Guerre (1914-1918)
• La fin du romantisme militaire : La guerre industrielle, avec son « orage d'acier », a rendu le courage individuel obsolète face aux obus.
• Le traumatisme et la « dévirilisation » : Les survivants sont revenus démembrés ou atteints de blessures invisibles (« l'obusite » ou syndrome post-traumatique). À l'époque, la psychiatrie associait ces troubles nerveux à une forme de féminisation ou de lâcheté.
• Inversion des rôles : Pendant que les hommes étaient au front, les femmes (« munitionnettes », conductrices, postières) ont prouvé leur capacité à faire fonctionner la société, brisant le mythe de leur infériorité physique et morale.
La réaction totalitaire des années 1930
• Le fascisme comme rempart : Face à la fluidité de la République de Weimar, les régimes totalitaires (Hitler, Mussolini) ont prôné un retour à une virilité antique et implacable.
• Idéologie et exclusion : La virilité nazie était indissociable de l'antisémitisme et de l'homophobie. Les homosexuels, perçus comme des « hommes-femmes » incapables de combattre, ont été persécutés et envoyés en camps (marqués du triangle rose).
• Relégation des femmes : Le modèle « Kinder, Kuche, Kirche » (Enfant, Cuisine, Église) visait à supprimer les droits politiques des femmes pour les cantonner au rôle d'épouse-mère du guerrier.
--------------------------------------------------------------------------------
3. Les Révolutions du Corps et du Pouvoir
À partir des années 1960, l'hégémonie virile est attaquée de front par les mouvements sociaux.
• L'ère des luttes LGBT et féministes : La dépénalisation de l'homosexualité (1982 en France) et les victoires législatives (contraception, IVG avec la loi Veil) ont dissocié la sexualité de la reproduction.
• La fin du Pater Familias : Le passage de l'autorité paternelle à l'autorité parentale conjointe a mis fin à la domination juridique absolue du père dans le foyer.
• Le sentiment d'humiliation : Pour les nostalgiques de la domination, ces changements sont vécus comme une dépossession de pouvoir et une « féminisation » de la société.
--------------------------------------------------------------------------------
4. La Virilité comme Performance Sexuelle et Sociale
La puissance masculine reste souvent indexée sur des attributs physiques et des performances quantifiables.
• Le Phallus comme sceptre : La puissance virile est fréquemment réduite à la puissance phallique, créant une obsession pour la taille, la durée et le nombre de conquêtes.
• La culture du vestiaire : Dès l'adolescence, la pression des pairs impose un masque de dureté. La sensibilité est assimilée à une défaillance.
• La domination économique : Au travail, Homo Virilus adopte l'uniforme du cadre ou du manager performant. Les « broligarches » (contraction de brothers et oligarques) utilisent des codes de guerre pour écraser la concurrence.
• La détresse silencieuse : L'échec professionnel est vécu comme une perte de valeur sociale totale. Le tabou de la santé mentale masculine mène à une « épidémie silencieuse » de suicides et de consommations excessives de drogues.
--------------------------------------------------------------------------------
5. Violences et Consentement : L'impact de MeToo
La dénonciation des violences sexistes et sexuelles remet en cause le « droit de cuissage » implicite de l'Homo Virilus.
• De la culture du viol au consentement : Des procès historiques (menés par Gisèle Halimi) à l'affaire Mazan, la société prend conscience que le viol n'est pas le fait de marginaux, mais d'hommes « ordinaires » issus de tous les milieux.
• Prise de conscience masculine : Le mouvement MeToo a forcé certains hommes à réévaluer leurs comportements passés, notamment sur la notion de rapport forcé ou de pression exercée sur les femmes.
• Le coût de la virilité : Les statistiques révèlent que les hommes sont responsables de :
◦ 91 % des tentatives d'homicides. ◦ 99 % des incendies volontaires. ◦ 84 % des accidents de la route mortels. ◦ Coût total : Près de 100 milliards d'euros par an en France.
--------------------------------------------------------------------------------
6. La Montée du Masculinisme et la « Manosphère »
En réaction à ces évolutions, un courant réactionnaire puissant s'organise, notamment sur les réseaux sociaux.
• Le discours du déclin : Des personnalités politiques et médiatiques prédisent l'effondrement de la civilisation face à la « castration » de l'homme blanc.
• L'influence de TikTok : La « manosphère » attire les jeunes hommes avec des conseils de drague et de fitness qui cachent un agenda antiféministe radical. Les femmes y sont souvent désignées comme des « féminazis ».
• Le mouvement Incel : Les « célibataires involontaires » développent une haine profonde des femmes, allant parfois jusqu'à l'apologie du terrorisme ou au passage à l'acte meurtrier (Montréal, Californie).
--------------------------------------------------------------------------------
7. Vers une Déconstruction et de Nouveaux Modèles
Des alternatives à l'Homo Virilus émergent, cherchant à dissocier la masculinité de la domination.
• La paternité active : Des ateliers de préparation à la paternité aident les hommes à s'approprier les gestes du soin et de l'éducation, sortant du rôle binaire « autorité et finance ».
• La masculinité au pluriel : Les sociologues et les personnes trans (comme Léon) soulignent que la testostérone ne crée pas le comportement dominateur ; c'est l'éducation qui façonne la virilité toxique.
• Distinction clé :
Les hommes se trouvent aujourd'hui à la croisée des chemins : perpétuer un stéréotype obsolète et coûteux, ou explorer des formes de masculinité fondées sur l'équité, la sensibilité et le partage des responsabilités.
Tags
Annotators
URL
-
-
learninglab.gitlabpages.inria.fr learninglab.gitlabpages.inria.fr
-
Question 4/6 : reformuler la question
-
Question 3/6 : reformuler la question ? Par exemple : A quels niveaux un robot peut-il généralement surpasser un humain ?
-
Explication Paro : "fins thérapeutiques" => ajouter un s à thérapeutique. Ajouter un "." à la fin de la phrase, et de manière générale à la fin de toutes les phrases d'explication.
-
-
www.youtube.com www.youtube.com
-
Briefing : Sensibilisation au Trouble du Développement Intellectuel (TDI)
Ce document de synthèse analyse les enjeux, les caractéristiques et les modalités d'accompagnement liés au trouble du développement intellectuel (TDI), sur la base des expertises de Fabienne Rudler et Marie-Thé (Job coaches et spécialistes en neurodéveloppement).
Résumé Exécutif
Le trouble du développement intellectuel (TDI), autrefois nommé handicap mental ou déficience intellectuelle, est un trouble neurodéveloppemental (TND) apparaissant durant l'enfance (avant 18 ans).
Il se définit par l'association de déficits des fonctions intellectuelles et de limitations significatives du comportement adaptatif. Contrairement aux troubles "Dys", le TDI impacte l'intelligence de manière globale, bien que les profils soient hétérogènes.
La prise en charge efficace repose sur une distinction cruciale entre le langage expressif (ce que la personne dit) et le langage réceptif (ce qu'elle comprend réellement), ainsi que sur une adaptation de l'environnement face aux difficultés d'abstraction, de repérage temporel et de généralisation des apprentissages.
--------------------------------------------------------------------------------
I. Cartographie et Typologie du Handicap
Il est essentiel de situer le TDI parmi les différentes catégories de handicap pour éviter les confusions diagnostiques.
| Type de Handicap | Caractéristiques Principales | | --- | --- | | Moteur | Limitation de la mobilité (membres) ou troubles d'origine motrice (ex: visuels). | | Sensoriel | Touche la vision et l'auditif (le goût/l'odorat ne sont pas encore reconnus suite au COVID). | | Psychique | Pathologies mentales (psychoses, bipolarité, dépression, TOC). Apparition souvent à l'adolescence ou à l'âge adulte. | | Dégénératif | État qui s'aggrave par paliers (pas de retour en arrière possible). | | Neurodéveloppemental (TND) | Apparaît durant le développement neurologique (0-18 ans). Inclut les troubles "Dys", le TDAH, l'autisme (TSA) et le TDI. |
Distinction entre "Dys" et TDI
• Troubles Spécifiques des Apprentissages (Dys) : L'intelligence est préservée. Le trouble est spécifique à un domaine (lecture, calcul, geste).
• TDI : Les capacités cognitives globales sont impactées. La pédagogie doit être radicalement différente car elle ne peut pas s'appuyer sur le même bagage cognitif.
--------------------------------------------------------------------------------
II. Étiologie et Critères Diagnostiques du TDI
Le TDI est un trouble dynamique. L'origine du trouble est multiple, bien qu'une part importante reste inexpliquée.
Les causes (Étiologie)
• 80 % d'origine prénatale (anténatale) : Aberrations chromosomiques (Trisomie 21), malformations cérébrales, prématurité, infections (rubéole, toxoplasmose) ou intoxications durant la grossesse.
• 10 % d'origine périnatale : Accidents lors de l'accouchement (anoxie cérébrale, hémorragies).
• 10 % d'origine postnatale : Séquelles de méningites, traumatismes crâniens (bébé secoué), noyades ou intoxications (plomb).
• Donnée clé : 40 % à 50 % des TDI restent d'origine indéterminée, ce chiffre montant à 80 % pour les déficiences légères.
Les trois piliers du diagnostic
1. Déficit des fonctions intellectuelles : Évalué par des tests de QI (inférieur à 70).
2. Limitation du comportement adaptatif : Difficultés dans la vie quotidienne (autonomie, relations sociales).
3. Apparition durant la période développementale : Avant l'âge de 18 ans.
--------------------------------------------------------------------------------
III. Analyse des Fonctions Cognitives Impactées
Le TDI se manifeste par des altérations dans plusieurs domaines clés du traitement de l'information.
1. Le repérage Espace-Temps
C'est l'un des domaines les plus lourdement impactés.
• Le Temps : Étant abstrait, il est plus difficile à saisir que l'espace.
Les personnes peuvent savoir lire l'heure (apprentissage mécanique) sans comprendre la notion de durée ou d'anticipation (ex: estimer le temps de trajet ou comprendre qu'un interrupteur s'éteint en secondes et non en minutes).
• L'Espace : Le repérage sur plan est complexe. L'apprentissage est souvent pragmatique (un trajet précis appris par cœur) plutôt que conceptuel (comprendre le réseau de transport).
2. Les fonctions exécutives
• Inhibition : Difficulté à s'empêcher de dire ou de faire quelque chose (impulsivité sociale).
• Flexibilité : Difficulté à passer d'une tâche à une autre ou à gérer l'imprévu.
• Mémoire de travail : Capacité limitée à maintenir une information à court terme pour réaliser une action immédiate.
3. La catégorisation
L'incapacité à regrouper des objets par concepts (ex: fruits et légumes) rend le quotidien coûteux en énergie. Au supermarché, une personne ne sachant pas catégoriser cherchera chaque produit individuellement plutôt que par rayon.
--------------------------------------------------------------------------------
IV. Le Comportement Adaptatif
Aujourd'hui, le degré de sévérité du TDI (léger, moyen, grave, profond) est déterminé par le fonctionnement adaptatif et non plus seulement par le score de QI.
Les habiletés évaluées (échelle Vineland-II)
• Conceptuelles : Lecture, écriture, calcul, gestion de l'argent.
• Pratiques : Tâches ménagères, déplacements, utilisation d'appareils (automates, machine à laver).
• Sociales : Demander de l'aide, éviter les abus, respecter les distances sociales.
La généralisation : un défi majeur
Une personne avec TDI peut apprendre à utiliser une machine à laver spécifique, mais ne saura pas forcément utiliser un autre modèle. Elle ne généralise pas le principe de fonctionnement. L'apprentissage doit être multiplié dans divers contextes.
--------------------------------------------------------------------------------
V. Focus sur la Communication et le Langage
La communication orale est un piège fréquent dans l'accompagnement, car elle repose sur de nombreux codes implicites.
Le paradoxe "Expressif vs Réceptif"
• Le "beau vernis" : Certaines personnes s'expriment très bien (langage expressif) mais ne comprennent pas les mots qu'elles emploient ou ceux des autres (langage réceptif).
• Vigilance : Il est impératif de vérifier systématiquement la compréhension, même si la personne semble éloquente.
Les obstacles linguistiques
• L'abstraction : Les mots ne renvoyant pas à un objet physique (ex: "attendre", "organiser", "ailleurs") sont difficiles à saisir.
• Les doubles sens : Mots phonétiquement identiques mais de sens différents (ex: "glace", "moule", "vase").
• Les petits mots "outils" : Les pronoms (je/tu/moi), les articles et les prépositions peuvent brouiller le message.
• La conditionnalité : Les phrases commençant par "Si..." (ex: "Si tu es malade...") sont souvent incomprises car elles demandent de se projeter dans une situation inexistante. La réponse sera souvent : "Mais je ne suis pas malade".
--------------------------------------------------------------------------------
VI. Recommandations pour l'Accompagnement
Pour favoriser l'inclusion et l'apprentissage des personnes avec TDI, plusieurs leviers doivent être activés :
1. Respecter le temps de latence : Le traitement de l'information est plus lent. Il faut ralentir le débit, poser une seule question à la fois et attendre la réponse sans reformuler trop vite.
2. Passer par le concret : Utiliser des supports visuels, des objets réels et éviter les métaphores ou le second degré.
3. Décomposer les tâches : Ce qui semble être une étape pour une personne ordinaire (ex: traverser la rue) est en réalité une succession de traitements cognitifs complexes (vitesse, distance, durée).
4. Partir de la réponse de la personne : Si une personne répond "à côté", il faut analyser sa logique pour adapter notre message (ex: si elle ne comprend pas "fromage", utiliser le mot "camembert" si c'est sa référence).
5. Anticiper les ruptures : Les changements de trajets, les grèves ou les pannes d'appareils sont des situations de mise en échec si elles n'ont pas été travaillées de manière pragmatique.
Conclusion sur l'autonomie
L'autonomie n'est pas de savoir tout faire seul, mais de savoir quand et comment demander de l'aide.
C'est sur ce levier social et adaptatif que les éducateurs et employeurs peuvent avoir le plus d'impact, car si le QI est fixe, le comportement adaptatif peut toujours progresser par l'expérience.
Tags
Annotators
URL
-
-
learninglab.gitlabpages.inria.fr learninglab.gitlabpages.inria.fr
-
Dan
Il y a écrit "Résumé" sous la vidéo mais vous ne proposez finalement pas de résumé ? uniquement la transcription ?
-
histoire et évolution de la robotique, mouvement et mécanique d’un robot, commande d’un robot, et limites & éthique de la robotique.
A présenter sous forme de liste numérotée ?
-
À la fin de ce cours, vous serez capable de : Acquérir des connaissances théoriques sur la genèse de la robotique,
La formulation du premier objectif ne convient pas :
-
La robotique est un domaine très large et il a fallu faire des choix sur les thèmes abordés.
La robotique étant un domaine très large, il a fallu faire des choix sur les thèmes abordés.
-
-
shs-cairn-info.ezproxy.u-paris.fr shs-cairn-info.ezproxy.u-paris.fr
-
expérience de « terreur agonistique » inélaborée et contre laquelle l’ensemble de l’appareil psychique s’est construit.
pareil que Green quand il dit que l'archaique c'est constitué en défense de ces états & pareil que winnicott pour la crainte de l'effondrement
-
-
www.youtube.com www.youtube.com
-
Guide de Référence : Le Programme Google Ad Grants pour les Associations
Résumé Exécutif
Le programme Google Ad Grants offre aux associations de loi 1901 une enveloppe de publicité gratuite sur le moteur de recherche Google s'élevant à 10 000 dollars par mois.
Malgré son potentiel massif pour accroître la notoriété, recruter des bénévoles ou collecter des fonds, ce programme reste largement sous-exploité en France, avec seulement 2 000 à 3 000 associations actives sur les millions existantes.
Ce document détaille les mécanismes du référencement payant, les critères d'éligibilité technique pour les structures, le processus d'activation en quatre étapes, ainsi que les stratégies optimales pour structurer des campagnes performantes.
Il souligne également les limites du programme, notamment la priorité donnée aux annonceurs payants et la nécessité d'une gestion rigoureuse pour maximiser l'impact du crédit quotidien de 329 dollars.
--------------------------------------------------------------------------------
1. Fondamentaux du Référencement Payant (SEA)
Le programme Google Ad Grants s'inscrit dans le cadre du référencement payant (SEA), qu'il convient de distinguer du référencement naturel (SEO).
Différences Clés : SEA vs SEO
| Caractéristique | Référencement Payant (SEA) | Référencement Naturel (SEO) | | --- | --- | --- | | Position | Haut de la page (résultats sponsorisés) | Sous les annonces sponsorisées | | Délai de résultat | Court terme (immédiat après lancement) | Long terme et incertain | | Coût | Paiement au clic (offert par Ad Grants) | "Gratuit" (nécessite du temps/contenu) | | Contrôle | Choix précis des mots-clés et zones | Dépend de l'algorithme de Google |
Spécificités du Compte Ad Grants
Contrairement à un compte Google Ads classique, le compte Ad Grants présente des particularités :
• Enveloppe virtuelle : Aucun budget réel n'est déboursé par l'association ; Google déduit les frais de l'enveloppe de 10 000 $.
• Hiérarchie de diffusion : Les annonces Ad Grants apparaissent en dessous des annonces payantes des entreprises privées ou des institutions disposant d'un budget marketing.
En cas de forte concurrence (ex: "collecte de dons"), il est parfois impossible de diffuser si les espaces publicitaires sont déjà saturés par des annonceurs payants.
--------------------------------------------------------------------------------
2. Éligibilité et Critères Techniques
Pour bénéficier du programme, une organisation doit remplir des critères statutaires et techniques précis.
Structures Éligibles
• Associations loi 1901.
• Fonds de dotation et fondations reconnues d'utilité publique.
• Exclusions : Les entités gouvernementales, les hôpitaux, les centres de soins et les écoles ne sont pas éligibles directement (sauf via une fondation ou une structure associative dédiée).
Exigences pour le Site Web
Google effectue une vérification manuelle du site Internet lors de la demande. Celui-ci doit présenter :
1. Un nom de domaine propre : Les sites hébergés sur des sous-domaines gratuits (ex: .wix.com, .google.site) sont refusés.
2. Un contenu substantiel : Un minimum de 5 pages est requis.
3. Une clarté institutionnelle : La mission et le statut associatif doivent être mentionnés en page d'accueil, dans une page "À propos" et dans le pied de page (footer).
4. Performance technique : Le site doit être "responsive" (adapté aux mobiles) et avoir une vitesse de chargement satisfaisante (idéalement un score > 50/100 sur PageSpeed Insights).
--------------------------------------------------------------------------------
3. Processus d'Activation en 4 Étapes
Le lancement d'un compte Ad Grants suit un parcours structuré :
1. Création du compte Google pour les associations : Utiliser de préférence une adresse email professionnelle liée au domaine de l'association pour simplifier la validation.
2. Validation de l'identité : Google vérifie le statut juridique de l'association (via le numéro RNA). Cette étape prend généralement 24 heures.
3. Activation de Google Ad Grants : Soumission du site web pour examen des critères de contenu et de performance. Le délai varie de 2 à 14 jours.
4. Configuration finale : Validation du profil de paiement (sans carte bancaire) et accès définitif au compte.
--------------------------------------------------------------------------------
4. Stratégie et Structuration des Campagnes
Une gestion efficace repose sur une structure logique et l'alignement entre l'intention de l'utilisateur et le contenu proposé.
Les 4 Piliers du Succès
• Le Ciblage : Sélection de mots-clés pertinents (volume de recherche > 50/mois) et spécifiques à la cause, en évitant les termes trop génériques ou ultra-concurrentiels.
• Les Annonces : Rédaction de titres percutants (jusqu'à 15 variantes) qui reprennent les mots-clés tapés par l'utilisateur.
• Les Enchères : Utilisation impérative de la stratégie "Maximiser les conversions" pour permettre à l'algorithme de Google d'optimiser la diffusion.
• Le Tracking : Connexion indispensable avec Google Analytics pour mesurer les actions concrètes (dons, inscriptions bénévoles, téléchargements).
Exemple de Structure de Compte (Cas d'un refuge animalier)
• Campagne Adoptions : Groupes d'annonces séparés pour "Adopter un chien" et "Adopter un chat" renvoyant vers les pages respectives du site.
• Campagne Bénévolat : Mots-clés sur le don de temps, le soin aux animaux ou le travail associatif.
• Campagne Marque : Protection du nom de l'association pour apparaître systématiquement en haut lors d'une recherche directe.
--------------------------------------------------------------------------------
5. Outils et Maintenance
Le maintien de la performance nécessite l'usage d'outils complémentaires et une surveillance régulière.
| Outil | Utilité | Niveau de difficulté | | --- | --- | --- | | Google Keyword Planner | Trouver des mots-clés et analyser leur volume/concurrence. | Débutant | | IA (ChatGPT, Gemini) | Aide à la rédaction des titres et descriptions d'annonces. | Débutant | | Google Analytics | Analyser le comportement des visiteurs après le clic. | Intermédiaire | | Google Tag Manager | Installer des marqueurs de conversion précis sans code. | Avancé |
Conseils de Gestion
• Ne jamais supprimer de campagne : Il est préférable de mettre en pause les campagnes inactives pour conserver l'historique et gagner du temps lors de la réactivation.
• Utilisation du budget : Le plafond de 10 000 $ est réparti à hauteur de 329 $ par jour. Les crédits non utilisés un jour donné sont définitivement perdus et ne sont pas reportables.
• Sécurité des accès : Il est crucial de nommer plusieurs administrateurs pour éviter la perte du compte en cas de départ d'un collaborateur ou d'un bénévole.
--------------------------------------------------------------------------------
6. Éthique et Transparence
Bien que les annonces soient financées par Google, elles portent la mention "Sponsorisé".
Cette transparence est renforcée par le Google Ads Transparency Center, qui permet au public de consulter les publicités diffusées par n'importe quelle entité.
Le programme s'inscrit dans la politique de Responsabilité Sociétale des Entreprises (RSE) de Google, agissant comme un don en nature sous forme d'espace publicitaire.
-
-
fisher-j.github.io fisher-j.github.io
-
For Duff & Litter, the largest difference was between the HD and HA treatments. The HD treatment had about 54.4 Mg ha-1, and was about 14.39 Mg ha-1 greater than the HA treatment (p = 0.09). Generally, all treatments were similar, with estimated loading of around 47 Mg ha-1.
as stated above...weird to lump these two measures. Seems like you could find some representative bulk density for duff and litter separately. Graham has this...but it's oddly reported. Or you could use some generic values or representative species from van wagtendonk Sierra Nevada species...not great but would still allow fair comparisons between treatments.
-
Figure 3.2 shows the same model as Figure 3.1, but with an emphasis on treatment comparisons between redwood and tanoak. This shows that we expect on average, 7.88 m2 ha-1 greater redwood basal area than tanoak basal area in the GS treatment (p = 0.1), about 1.69 m2 ha-1 in the LD treatment (p = 0.4), and about 0.6 m2 ha-1 in the HA treatment (p = 0.48). In the HD treatment, we expect to see slightly higher tanoak basal area (p = 0.55).
not sure this adds distinct value since you already provided this information.
-
Table 3.2: Basal area (m2 ha-1) modeled at the vegetation plot level for four harvest treatments and four species classes (n = 16). The asymptotic 95% confidence intervals are based on the normal approximation.
separate table is not needed if data is already presented as figure.
-
Redwood regeneration basal area showed the greatest treatment response. The GS treatment had the greatest basal area of redwood regeneration at 10.12 m2 ha-1, which was 9.28 m2 ha-1 greater than in the HD treatment (p = 0.19). The LD and HD treatments were intermediate. Tanoak regeneration basal area was intermediate between that of redwood and Douglas-fir and other species. The GS and LD treatments had similar responses, as did the HA and HD treatments. The GS treatment resulted in 2.24 m2 ha-1 of tanoak basal area, which was 1.33 m2 ha-1 greater than in the HA treatment (p = 0.18). On average, for Douglas-fir, we expect about 0.17 m2 ha-1 of basal area across treatments. The greatest basal area of Douglas-fir was in the GS treatment which was 0.12 m2 ha-1 greater than in the HA treatment (p = 0.76). The LD, HA, and HD treatments were all comparatively similar. Other species included grand fir, madrone, and California wax-myrtle, of which there was a total of 23, 28, and 16 observations across our 16 macro plots (comprising 64 tree density plots). Generally, each plot had between 0 and 9 observations of other species, except for one macro plot with the LD treatment, which had 16 observations (data not shown). According to predictions made from this model for other species, there was not enough evidence to confirm a statistically significant difference between treatments. On average, we expect about 0.11 m2 ha-1 of basal area across treatments. The greatest basal area of other species was in the HD treatment which was 0.12 m2 ha-1 greater than in the HA treatment (p = 0.26). The GS and LD treatments were intermediate.
might rethink the structure of presenting results. Looks like none of the treatments had differences in basal area for any treatment. I would start with this. Then you can briefly explain some of the trends across species and treatments.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The manuscript by Poh et al. investigated whether dopamine release in the ventral medial striatum integrates information about action selection, controllability of reward pursuit, effort, and reward approach. Rats were implanted with FSCV probes and trained in four Go/No Go task variants:
(1) trials were self-initiated and had two trial types (Go vs. No Go) that were auditorily cued,
(2) trials were cue-initiated and had two trial types (Go vs. No Go) that were auditorily cued,
(3) trials were self-initiated and had three trial types (Go vs. No Go vs. free reward) that were auditorily cued, and effort was increased,
(4) trials were cue-initiated and had three trial types (Go vs. No Go vs. free reward) that were auditorily cued.
The authors report that dopamine levels rose during Go trials and slowly rose in No Go trials, but this pattern did not differ across task variants that modified effort and whether trials were cued or initiated. They also report that dopamine levels rose as rats approached the reward location and were greater in rats that bit the noseport while holding during the No Go response.
Strengths:
(1) Interesting task and variants within the task paradigm that would allow the authors to isolate specific behavioral metrics.
(2) The goal of determining precisely what VMS dopamine signals do is highly significant and would be of interest to many researchers.
Weaknesses:
(1) This Go/No-Go procedure is different from the traditional tasks, and this leads to several problems with interpreting the results:
(a) Go/No Go tasks typically require subjects to refrain from doing any action. In this task, a response is still required for the No Go trials (e.g., continue holding the nosepoke). The problem with this modified design is that failure to withhold a response on No Go trials could be because i) rats could not continue holding the response, as holding responses are difficult for rodents, or ii) rats could not suppress the prepotent go response. This makes interpreting the behavior and the dopamine signal in No Go trials very difficult.
(b) Most Go/No Go tasks bias or overrepresent Go trials so that the Go response is prepotent, and consequently, successful suppression of the Go response is challenging. I didn't see any information in the manuscript about how often each trial type was presented or how the authors ensured that No Go responses (or lack thereof) were reflecting a suppression of the Go response.
(2) The authors observe relatively consistent differences in the DA signal between Go and No Go trials after the action-cue onset. However, the response type was not randomized between trial type, so there is a confound between trial type (Go/No Go) and response (lever/nosepoke). The difference in DA signal may have nothing to do with the cue type, but reflects differences in DA signal elicited by levers vs. nosepokes.
(3) Both Go and No Go trials start with the rat having their nose in the noseport. One cue (Go cue) signals the rat to remove their nose from the noseport and make two lever responses in 5 seconds, whereas the other cue (No Go cue) signals the rat to keep their nose in the noseport for an additional 1.7-1.9 s. The authors state that the time between cue onset and reward delivery was kept the same for all trial types, and Figure 1 suggests this is 2 s, so was reward delivered before rats completed the two lever presses? I would imagine reward was only delivered if rats completed the FR requirement, but again, the descriptions in the text and figures are incongruent.
(4) The manuscript is difficult to understand because key details are not in the main text or are not mentioned at all. I've outlined several points below:
(a) The author's description in the manuscript makes it appear as a discrimination task versus a Go/No Go task. I suggest including more details in the main text that clarify what is required at each step in the task. Additionally, providing clarity regarding what task events the voltammetry traces are aligned to would be very useful.
(b) How many subjects were included in each task variant? The text makes it seem like all rats complete each task variant, but the behavioral data suggest otherwise. Moreover, it appears that some rats did more than one version. Was the order counterbalanced? If not, might this influence the DA signal?
(5) There is a major challenge in their design and interpretation of the dopamine signal. Both trial types (Go and No Go) start with the rat having their nose in the noseport. An auditory cue is presented for 2-3 s signaling to the rat to either leave the noseport and make a lever response (Go trial) or to stay in the noseport (No Go trial). The timing of these actions and/or decisions is entirely independent, so it is not clear to me how the authors would ever align these traces to the exact decision point for each trial type. They attempt to do this with the nose-port exit analysis, but exiting the noseport for a Go trial (a rat needs to make 2 lever presses and then get a reward) versus a No Go trial (a rat needs to go retrieve the reward) is very different and not comparable.
(6) The voltammetry analysis did not appear to test the hypotheses the authors outlined in the intro. All comparisons were done within task variants (DA dynamics in Go vs. No Go trials, aligned to different task events), but there were no comparisons across task variants to determine if the DA signal differed in cued vs self-initiated trials.
(7) Classification of No Go behaviors was interesting, but was not well integrated with the rest of the paper and was underdeveloped. It also raised more questions for me than answers. For example:
(a) Was the behavior classification consistent across rats for all No Go trials? If not, did the DA signal change within subjects between biting vs digging vs calm?
(b) If "biting rats" were not always biting rats on every No Go trial, then is it fair to collapse animals into a single measure (Figure 3C).
(c) Some of the classification groups only had 2 or fewer rats in them, making any statistical comparison and inference difficult.
-
-
shs-cairn-info.ezproxy.u-paris.fr shs-cairn-info.ezproxy.u-paris.fr
-
Je retiendrai trois notions principales : l’interdépendance et les interactions, différentes représentations de l’existence fantasmée du groupe, et le groupe en tant qu’espace transitionnel. Ainsi que nous le verrons, celles-ci interrogent une compréhension des processus inconscients : a) centrés principalement sur « le sujet », dont la vie intrapsychique se structure en fonction de la place réelle et imaginaire qu’il occupe dans les différents groupes auxquels il appartient ; b) centrés sur « le groupe », représentation commune à un ensemble d’individus. Ces notions rendent compte de la fluidité des frontières existant entre l’individu et les groupes, et situent l’espace analytique à l’articulation des limites fluctuantes du Je et du Nous, du dedans et du dehors, du morcellement et de l’unité somatique, et de la pluralité psychique des identités singulière et collective.
L’auteur dit qu’il va s’appuyer sur trois idées clés : 1. l’interdépendance et les interactions → dans un groupe, personne ne fonctionne seul : chacun influence et est influencé. 2. les représentations fantasmées du groupe → le groupe est aussi une construction imaginaire (mère, menace, refuge, etc.), pas seulement un ensemble réel de personnes. 3. le groupe comme espace transitionnel → le groupe est un entre-deux, ni totalement interne ni totalement externe, où peuvent se déposer et se transformer des vécus archaïques.
À partir de là, il distingue deux façons de penser l’inconscient : • soit centré sur le sujet (comment sa vie psychique dépend de sa place dans les groupes), • soit centré sur le groupe (le groupe comme représentation commune).
Ces notions montrent que les frontières entre individu et groupe sont mobiles : le travail analytique se situe précisément dans cet espace intermédiaire, entre Je et Nous, dedans et dehors, morcellement et unité, identité individuelle et collective.
👉 En très court : le groupe est un espace transitionnel où s’entrelacent psychisme individuel et collectif, avec des frontières fluctuantes, permettant l’expression et la transformation de l’archaïque.
-
Avant de traiter de processus primaires tels que l’incorporation, l’introjection et l’identification projective, il est nécessaire de rappeler brièvement certaines notions concernant la conception du « groupe » et de la pratique d’analyse, en les étayant de considérations nouvelles et de formulations plus précises. Ces premières approximations, à partir desquelles j’ai pu développer une pratique et une théorie de l’analyse de groupe, nous conduiront des imagos maternelles archaïques et du groupe comme objet transitionnel aux processus primaires et aux modalités, par lesquels des émotions transférées se substituent à la réalité des personnes présentes dans le groupe, et surdéterminent leurs rapports.
… des imagos maternelles archaïques et du groupe comme objet transitionnel… --> imagos maternelles archaïques = images internes très primitives de la mère (pas la mère réelle). Elles sont : • idéalisées, • persécutrices, • fusionnelles. C’est ce que les patients projettent dans le groupe.
Groupe comme objet transitionnel Le groupe est vécu comme : • ni complètement soi, • ni complètement extérieur. = Comme l’objet transitionnel chez Winnicott. Donc : le groupe devient un support archaïque.
… aux processus primaires et aux modalités par lesquels des émotions transférées se substituent à la réalité des personnes présentes dans le groupe… --> dans le groupe : les affects transférentiels remplacent la perception réelle des autres. On ne voit plus les personnes telles qu’elles sont. On voit : • la mère archaïque, • l’objet persécuteur, • l’objet idéal. Les émotions prennent le dessus sur la réalité.
… et surdéterminent leurs rapports. --> les relations ne sont plus libres. Elles sont dirigées par ces projections archaïques.
-
C’est une introduction à la psychanalyse du lien primaire qui, au-delà de la relation d’objet, est un des éléments fondamentaux du travail de groupe.
!!!
-
-
localhost:3003 localhost:3003
-
GSTIN - NA
Point 1: Newly Added- default value as NA
-
-
pmc.ncbi.nlm.nih.gov pmc.ncbi.nlm.nih.gov
-
RRID: AB_443425
DOI: 10.1038/s41419-025-08362-w
Resource: (Abcam Cat# ab16669, RRID:AB_443425)
Curator: @evieth
SciCrunch record: RRID:AB_443425
Tags
Annotators
URL
-
-
pmc.ncbi.nlm.nih.gov pmc.ncbi.nlm.nih.gov
-
RRID:CVCL_1544
DOI: 10.3390/v18010102
Resource: (KCLB Cat# 90226, RRID:CVCL_1544)
Curator: @scibot
SciCrunch record: RRID:CVCL_1544
Tags
Annotators
URL
-
-
pmc.ncbi.nlm.nih.gov pmc.ncbi.nlm.nih.gov
-
RRID:AB_477043
DOI: 10.1242/dev.204964
Resource: (Sigma-Aldrich Cat# H0412, RRID:AB_477043)
Curator: @scibot
SciCrunch record: RRID:AB_477043
Tags
Annotators
URL
-
-
pmc.ncbi.nlm.nih.gov pmc.ncbi.nlm.nih.gov
-
RRID:AB_312686
DOI: 10.1158/2326-6066.CIR-25-0444
Resource: (BioLegend Cat# 100401, RRID:AB_312686)
Curator: @scibot
SciCrunch record: RRID:AB_312686
Tags
Annotators
URL
-
-
www.sciencedirect.com www.sciencedirect.com
-
RRID:AB_2804516
DOI: 10.1016/j.xcrm.2025.102580
Resource: (Thermo Fisher Scientific Cat# PA5-87933, RRID:AB_2804516)
Curator: @scibot
SciCrunch record: RRID:AB_2804516
-
RRID:AB_2745193
DOI: 10.1016/j.xcrm.2025.102580
Resource: (Thermo Fisher Scientific Cat# MA5-28223, RRID:AB_2745193)
Curator: @scibot
SciCrunch record: RRID:AB_2745193
-
-
www.sciencedirect.com www.sciencedirect.com
-
RRID:AB_2562039
DOI: 10.1016/j.cell.2025.12.043
Resource: (BioLegend Cat# 100237, RRID:AB_2562039)
Curator: @scibot
SciCrunch record: RRID:AB_2562039
-
-
www.biorxiv.org www.biorxiv.org
-
AbstractCancer cells are heterogeneous, each harboring distinct molecular aberrations and are dependent on different genes for their survival and proliferation. While successful targeted therapies have been developed based on driver DNA mutations, many patient tumors lack druggable mutations and have limited treatment options. Here, we hypothesize that new precision oncology targets may be identified through “expression-driven dependency”, whereby cancer cells with high expression of a targeted gene are more vulnerable to the knockout of that gene. We introduce a Bayesian approach, BEACON, to identify such targets by jointly analyzing global transcriptomic and proteomic profiles with genetic dependency data of cancer cell lines across 17 tissue lineages. BEACON identifies known druggable genes, e.g., BCL2, ERBB2, EGFR, ESR1, MYC, while revealing new targets confirmed by both mRNA- and protein-expression driven dependency. Notably, the identified genes show an overall 3.8-fold enrichment for approved drug targets and enrich for druggable oncology targets by 7 to 10-fold. We experimentally validate that the depletion of GRHL2, TP63, and PAX5 effectively reduce tumor cell growth and survival in their dependent cells. Overall, we present the catalog of express-driven dependency targets as a resource for identifying novel therapeutic targets in precision oncology.
This work has been peer reviewed in GigaScience (see https://doi.org/10.1093/gigascience/giag011), which carries out open, named peer-review. These reviews are published under a CC-BY 4.0 license and were as follows:
Reviewer 2
The authors introduce BEACON, a Bayesian correlation approach designed to identify expression-driven dependency in cancer. Their hypothesis suggests that cancer cells with elevated expression of specific genes demonstrate increased vulnerability to the knockout of those same genes, thereby unveiling a promising new category of targets in precision oncology—particularly valuable for targeting cancer cells lacking druggable mutations. BEACON models expression levels and dependency scores as bivariate Gaussians and employs Markov Chain Monte Carlo (MCMC) sampling to estimate the correlation coefficient between them. They then compute p-values followed by rigorous multiple testing correction (BH based FDR correction). A notable strength of their approach lies in the integration of mass spectrometry proteomics data alongside transcriptomic and perturbation screening data, enhancing the robustness of their findings. Their work highlights some key insights: - Gene expression-driven dependency (GED) candidates identified across lineages demonstrate enrichment for "DNA-binding transcription activator activity" and "DNA-binding transcription activator activity, RNA polymerase II-specific" pathways. - The analysis successfully identifies compelling candidates with robust signals in both GED and PED (FERMT2, GRHL2, KLF5, CDK6, and CCND1), which are well-supported by existing drug evidence or established literature - Clustering analyses reveal that cancer cells from pancreas and biliary tract tissues, as well as kidney and urinary tract tissue lineages, exhibit remarkably similar expression-driven dependency profiles. Additionally, lineage-specific genes such as transcription factors, cluster together in a manner consistent with existing literature - Through Fisher's exact test, the authors demonstrate significant enrichments of druggable gene lists from DrugBank with expression-driven dependency patterns at both proteomic and transcriptomic levels - Experimental validation shows that PAX5 is essential for PAX5-high B cell lymphoma cell growth, while TP63 and GRHL2 are essential for LSCC cell growth.
However, I have several principal concerns about the study that should be addressed to demonstrate the robust and superior performance of this proposed approach.
Major Comments:
1.Quantitative benchmarking: While the authors present a valuable contribution, the concept of correlating gene dependency scores to expression has been explored previously through approaches like Project DRIVE (E. Robert McDonald, III et al.) and APSIC (Montazeri et al.). BEACON demonstrates strong correlations across multiple lineages, representing broader scope compared to existing methods that appear more lineage-restricted. However, establishing BEACON's comparative advantages requires more rigorous evaluation. Notably, Project DRIVE—a foundational paper in this field—already identified several BEACON candidates in their "Expression Correlation Analysis Identifies Oncogenes and Lineage-Specific Transcription Factors" section, while APSIC characterized many lineage-specific discoveries as tumor effector genes. BEACON's strength lies in integrating proteomic data with transcriptomic and perturbation screens, enabling identification of additional candidates like PAX5 for hematopoietic and lymphoid tissue. To demonstrate the method's impact, I recommend systematic quantitative benchmarking against existing approaches.
Importantly, BEACON utilizes richer/complementary datasets than previous studies. Disentangling contributions of data richness versus methodological innovation would provide valuable insights into whether enhanced performance stems from improved data availability or genuine method improvements.
Overall for benchmarking, the authors are strongly encouraged to utilize any comprehensive datasets that best demonstrate their method's competitive advantage and are not limited to the specific comparisons recommended above.
2.Correlation method comparisons: Figure S2 shows that BEACON exhibits higher MSE at extremes, and the claimed advantage over Pearson for small sample sizes is difficult to quantify from the current visualization. While the theoretical expectation that BEACON should outperform Pearson in small samples is reasonable, the practical significance remains unclear from these simulations. I recommend demonstrating BEACON's advantage using real data by creating a curated list of established GEDs/PEDs and comparing performance between the two methods. This is particularly important since several of BEACON's hits were previously reported by Project DRIVE using simple Pearson correlations. Alternatively, if BEACON's advantage is indeed significant, please elaborate on the simulation results to better justify this claim with clearer quantitative metrics.
3.Validation experiments: I'm seeking clarification on the validation experiments for TP63 and GRHL2. These candidates were not sensitive to predicted dependency and the authors say that "pan-lineage targets may represent universal vulnerability and their inhibition may lead to undesired off-target effects on other cells". Are the authors positioning them as weaker candidates to illustrate the superiority of lineage-specific predictions like PAX5? Additionally, why were different experimental approaches used—CRISPR for PAX5 versus shRNA for TP63 and GRHL2? For a method aimed at identifying druggable targets, would drug based experiments be more relevant than knockdown approaches to better demonstrate clinical applicability?
Minor comments
- In Figure 4A, the caption refers to the plot as a heatmap, but the visualization appears to be a scatterplot. Please clarify whether the heatmap is missing or modify the caption appropriately. Additionally, I recommend using a different shade of green, as the current color choice makes some gene names difficult to read.
- In Fig S5A, please add a legend for tumor and normal
- For the TP63 and GRHL2 validation experiments, please include results for all four cell lines. The current manuscript is missing HCC15-shTP63, HCC15-shGRHL2, and HARA-shGRHL2 plots.
- How many replicates were the experiments performed on? Is it N= 3 for all experiments?
- Missing some text here - "BEACON offers the unique advantage of utilizing prior distributions that are less susceptible to outliers, especially in multiple lineages where the number of cell lines."
-
-
www.examtopics.com www.examtopics.com
-
NOTA: Cada respuesta correcta vale un punto.
El código crea un arrendamiento (lease) infinito.
El código en la línea 06 siempre crea un nuevo blob.
El bloque finally libera el arrendamiento (lease).
-
-
-
Otros (no son candidaturas)
La mayoría de las candidaturas no superan el margen de error.
-
-
www.examtopics.com www.examtopics.com
-
¿Cuáles son las tres acciones que debe realizar en secuencia? Para responder, mueva las acciones correspondientes de la lista de acciones al área de respuestas y ordénelas correctamente.
azcontainerapp env create
azcontainerapp job create
azcontainerapp job start
-
-
www.examtopics.com www.examtopics.com
-
Necesita implementar la aplicación en Azure.
Azure Container Instance.
Restar policy.
-
-
www.examtopics.com www.examtopics.com
-
Área de interés:
Se crea una alerta de registros que envía un correo electrónico cuando el porcentaje de CPU es superior al 60 % durante cinco minutos.
Se crea una alerta de registros que envía un correo electrónico cuando el número de latidos (heartbeats) de máquinas virtuales en la última hora es menor que cinco.
La alerta de registros está programada para ejecutarse cada dos horas.
-
Seleccionar y colocar:
Export a Resource Manager template (Exportar una plantilla de Azure Resource Manager desde la cuenta de almacenamiento existente)
Modify the template by changing the storage account name and region (Modificar la plantilla para usar un nuevo nombre de cuenta y la región de destino)
Create a new template deployment (Crear una nueva implementación usando la plantilla modificada)
Deploy the template to create a new storage account in the target region (Implementar la plantilla para crear la nueva cuenta de almacenamiento en la región destino)
Use AzCopy to copy the data to the new storage account (Copiar los datos directamente entre cuentas usando AzCopy, servidor-a-servidor)
-
NOTA: Cada selección correcta vale un punto.
1.- Devuelva el estado más reciente del paciente.
2.- Devuelva los datos de supervisión de la salud que no estén retrasados más de una versión.
3.- Después de que el paciente sea dado de alta y se hayan evaluado todos los cargos, recupere los datos de facturación correctos con los cargos finales.
-
NOTA: Cada respuesta correcta vale un punto.
1.- El código registrará la hora en la que el pedido fue procesado desde la cola.
2.- Cuando la función ProcessOrders falla, la función se reintentará hasta cinco veces para un pedido determinado, incluyendo el primer intento.
3.- Cuando hay varios pedidos en la cola, se recuperará un lote de pedidos desde la cola y la función ProcessOrders ejecutará múltiples instancias de forma concurrente para procesar los pedidos.
4.- La función ProcessOrders enviará el pedido como salida a una tabla Orders en Azure Table Storage.
-
-
hypothes.is hypothes.is
-
ando la barra lateral a la de
dEsta parte me indica que deboo
-
-
www.biorxiv.org www.biorxiv.org
-
Note: This response was posted by the corresponding author to Review Commons. The content has not been altered except for formatting.
Learn more at Review Commons
Reply to the reviewers
We thank the reviewers and editors for their careful evaluation of our manuscript and their positive comments on the importance and rigor of the work. Below you will find our point-by-point response to each reviewer's suggestions. We believe that we have addressed (in the response and the revised manuscript) all of the concerns. Please note that in some cases, we have numbered a reviewer's comments for clarity, however beyond this, we have not altered any of the reviewers' text.
Reviewer #1 (Evidence, reproducibility and clarity (Required)):
Lo et al., report a high-throughput functional profiling study on the gene encoding for argininosuccinate synthase (ASS1), done in a yeast experimental system. The study design is robust (see lines 141-143, main text, Methods), whereby "approximately three to four independent transformants of each variant would be isolated and assayed." (lines 140 - 141, main text, Methods). Such a manner of analysis will allow for uncertainty of the functional readout for the tested variants to be accounted for.
This is an outstanding study providing insights on the functional landscape of ASS1. Functionally impaired ASS1 may cause citrullinemia type I, and disease severity varies according to the degree of enzyme impairment (line 30, main text; Abstract). Data from this study forms a valuable resource in allowing for functional interpretation of protein-altering ASS1 variants that could be newly identified from large-scale whole-genome sequencing efforts done in biobanks or national precision medicine programs. I have some suggestions for the Authors to consider:
- The specific function of ASS1 is to condense L-citrulline and L-aspartate to form argininosuccinate. Instead of measuring either depletion of substrate or formation of product, the Authors elected to study 'growth' of the yeast cells. This is a broader phenotype which could be determined by other factors outside of ASS1. Whereas i agree that the experiments were beautifully done, the selection of an indirect phenotype such as ability of the yeast cells to grow could be more vigorously discussed.
We appreciate the reviewer's point regarding the indirect nature of growth as a functional readout. In our system, yeast growth is tightly and specifically coupled to ASS enzymatic activity. The strains used are isogenic and lack the native yeast argininosuccinate synthetase, such that arginine biosynthesis, and therefore yeast replication on minimal medium lacking arginine, depends exclusively on the activity of human ASS1. Under these defined and limiting conditions, growth provides a quantitative proxy for ASS1 function. However, we acknowledge that this assay does not resolve specific molecular mechanisms underlying reduced function, such as altered catalytic activity versus effects on protein stability. We have updated the text to clarify these points.
"While growth is an indirect phenotype relative to direct measurement of substrate turnover or product formation, it is tightly coupled to ASS enzymatic activity in this system and is expected to be impaired by amino acid substitutions that reduce catalytic activity or protein stability. Therefore, growth on minimal medium lacking arginine is a quantitative measure of ASS enzyme function, allowing the impact of ASS1 missense variants to be assessed at scale through a high-throughput growth assay, in a single isogenic strain background, under controlled, defined conditions that limit confounding factors unrelated to ASS1 activity. We expect that the assay will detect reductions in both catalytic activity and protein stability but will not distinguish between these mechanisms."
- One of the key reasons why studies such as this one are valuable is due to the limitations of current variant classification methods that rely on 'conservation' status of amino acid residues to predict which variants might be 'pathogenic' and which variants might be 'likely benign'. However, there are serious limitations, and Figures 2 and 6 in the main text shows this clearly. Specifically, there is an appreciable number of variants that, despite being classified as "ClinVar Pathogenic", were shown by the assay to unlikely be functionally impaired. This should be discussed vigorously. Could these inconsistencies be potentially due to the read out (growth instead of a more direct evaluation of ASS1 function)?
We interpret this discrepancy as reflecting a sensitivity limitation of the growth-based readout rather than a fundamental disagreement between functional effect and clinical annotation. Specifically, we believe that our assay is unable to resolve the very mildest hypomorphic variants from true wild type, i.e., the residual activity of these variants is sufficient to fully support yeast growth under the conditions used. On this basis, we have chosen not to treat wild-type-like growth in our assay as informative for benignity; conversely, reduced growth provides evidence supporting pathogenicity (all clinically validated variants examined in this range are pathogenic).
We have revised the manuscript to clarify this point explicitly and to frame these variants as lying outside the effective resolution limit of the assay rather than representing true false positives. Additional discussion of this limitation and its implications is provided in our responses to Reviewer 2 (points 1 and 4) along with specific changes made to the text.
- Figure 3 is very interesting, showing a continuum of functional readout ranging from 'wild-type' to 'null'. It is very interesting that the Authors used a threshold of less than 0.85 as functionally hypomorphic. What does this mean? It would be very nice if they have data from patients carrying two hypomorphic ASS1 alleles, and correlate their functional readout with severity of clinical presentation. The reader might be curious as to the clinical presentation of individuals carrying, for example, two ASS1 alleles with normalized growth of 0.7 to 0.8.
I hope you will find these suggestions helpful.
We thank the reviewer for this thoughtful comment. Figure 3 indeed illustrates a continuum of functional effects, and we agree that careful interpretation of the thresholds used is important. To clarify the rationale for the hypomorphic threshold, the interpretation of intermediate growth values, and to emphasize that these labels reflect only behavior in the functional assay, we have rewritten the relevant section of the Results:
"The normalized growth scores of the 2,193 variants tested in our functional assay form a clear bimodal distribution (Figure 3), with two distinct peaks corresponding to functional extremes, as is commonly reported in large-scale functional assays of protein function [9, 10]. The smaller peak, centered around the null control (normalized growth = 0), represents variants that fail to support growth in the assay (growth 0.85). Variants with growth values falling between these two peak-based thresholds display partial functional impairment and are classified as functionally hypomorphic (n = 323). Crucially, these classifications are entirely derived from the observed peaks in the distribution of growth values and reflect differences in functional activity under the assay conditions. They do not provide direct evidence for clinical pathogenicity or benignity and should not be used for clinical variant interpretation without proper benchmarking against clinical reference datasets, as implemented below within an OddsPath framework."
We agree with the reviewer that correlating functional measurements with clinical severity in individuals carrying two hypomorphic ASS1 alleles would be highly informative, particularly given that ASS1 deficiency is an autosomal recessive disorder. While mild hypomorphic variants (for example, variants with normalized growth values of 0.7-0.8 in our assay) could plausibly contribute to disease when paired with a complete loss-of-function allele, systematic analysis of combinatorial genotype effects and genotype-phenotype correlations is beyond the scope of the present study, which focuses on the functional effects of individual variants. We view this as an important direction for future work.
Reviewer #1 (Significance (Required)):
This is an outstanding study providing insights on the functional landscape of ASS1. Functionally impaired ASS1 may cause citrullinemia type I, and disease severity varies according to the degree of enzyme impairment (line 30, main text; Abstract). Data from this study forms a valuable resource in allowing for functional interpretation of protein-altering ASS1 variants that could be newly identified from large-scale whole-genome sequencing efforts done in biobanks or national precision medicine programs.
Reviewer #2 (Evidence, reproducibility and clarity (Required)):
In this manuscript, Lo et al characterize the phenotypic effect of ~90% of all possible ASS1 missense mutations using an elegant yeast-based system, and use this dataset to aid the interpretation of clinical ASS1 variants. Overall, the manuscript is well-written and the experimental data are interpretated rigorously. Of particular interest is the identification of pairs of deleterious alleles that rescue ASS1 activity in trans. My comments mainly pertain to the relevance of using a yeast screening methodology to infer functional effects of human ASS1 mutations.
- Since human ASS1 is heterologously expressed in yeast for this mutational screen, direct comparison of native expression levels between human cells and yeast is not possible. Could the expression level of human ASS1 (driven by the pARG1 promoter) in yeast alter the measured fitness defect of each variant? For instance, if ASS1 expression in yeast is sufficiently high to mask modest reductions in catalytic activity, such variants may be misclassified as hypomorphic rather than amorphic. Conversely, if expression is intrinsically low, even mild catalytic impairments could appear deleterious. While it is helpful that the authors used non-human primate SNV data to calibrate their assay, experiments could be performed to directly address this possibility.
The nature of the relationship between yeast growth and availability of functional ASS1 could also influence the interpretation of results from the yeast-based screen. Does yeast growth scale proportionately with ASS1 enzymatic activity?
We completely agree that the expression level of human ASS1 in yeast could influence the measured fitness effects of individual variants. We expect the rank ordering of variants in our growth assay to reflect their relative enzymatic activity (i.e. a monotonic relationship) but acknowledge that the precise mapping between activity and growth is unknown and may include ceiling and floor effects that limit the assay's dynamic range. As the reviewer notes, under high expression conditions moderate loss-of-function variants could appear indistinguishable from wild type (ceiling effect), whereas under lower expression the same variants could behave closer to the null control (floor effect).
In our system, ASS1 is expressed from the pARG1 promoter, chosen under the assumption that the native expression level of ARG1 (the yeast ASS1 ortholog) is appropriately tuned for yeast growth. Crucially, rather than assuming a fixed mapping from assay growth to clinical pathogenicity (given potential nonlinearities in the relationship between ASS function and growth) we benchmark the assay against external data, including known pathogenic and benign variants and non-human primate SNVs, to calibrate thresholds and guide interpretation within an OddsPath framework. This benchmarking indicates that ceiling effects are likely present, with some mild loss-of-function pathogenic variants appearing indistinguishable from wild type in the growth assay. We explicitly account for this by not using high-growth scores as evidence toward benignity. We have made the following changes the manuscript:
"A subset of clinically pathogenic ASS1 variants exhibit near-wild-type growth in our yeast assay. In general, we expect a monotonic relationship between ASS function and yeast growth, but with the potential for floor and ceiling effects that constrain the assay's dynamic range. In this context, we interpret high-growth pathogenic variants as likely causing mild loss of function that cannot be distinguished from wild type in our assay"
"Based on these findings and given that 22/56 pathogenic variants show >85% growth, we conclude that growth above this threshold should not be used as evidence toward benignity."
- It would be helpful to add an additional diagram to Figure 1A explaining how the screen was performed, in particular: when genotype and phenotype were measured, relative to plating on selective vs non-selective media? This is described in "Variant library sequence confirmation" and "Measuring the growth of individual isolates" of the Methods section but could also be distilled into a diagram.
We thank the reviewer for this helpful suggestion. We have updated Figure 1 by adding a new schematic panel (Figure 1C) that distills the experimental workflow into a visual overview. This diagram is intended to complement the detailed descriptions in the Methods and improve clarity for the reader.
- The authors rationalize the biochemical consequences of ASS1 mutations in the context of ASS1 per se - for example, mutations in the active site pocket impair substrate binding and therefore catalytic activity, which is expected. Does ASS1 physically interact with other proteins in human cells, and could these interactions be altered in the presence of specific ASS1 mutations? Such effects may not be captured by performing mutational scanning in yeast.
We are not aware of any specific protein-protein interactions involving ASS that are required for its enzymatic function. However, we agree that ASS could engage in non-essential interactions with other human proteins that might be altered by specific missense variants and that such interactions would not necessarily be captured in a yeast-based assay.
Importantly, our complementation system depends on human ASS providing the essential enzymatic activity required for arginine biosynthesis in yeast. If ASS1 required obligate human-specific protein interactions to function, even the wild-type enzyme would fail to support yeast growth, which is clearly not the case. We therefore conclude that the assay robustly reports on the intrinsic enzymatic activity of ASS, while acknowledging that non-essential human-specific interactions may not be assessed. We have updated the manuscript to reflect this point.
"Importantly, successful functional complementation indicates that ASS enzymatic activity does not depend on any obligate human-specific protein interactions."
- The authors note that only a small number (2/11) of mutations at the ASS1 monomer-monomer interface lead to growth defects in yeast. It would be helpful for the authors to discuss this further.
As discussed in response to the reviewer's comments on the relationship between ASS activity and yeast growth (point 1 above), we expect growth to be a monotonic but nonlinear function of enzymatic activity, with potential ceiling effects at high activity. Under this model, variants causing weak or moderate loss of function may remain indistinguishable from wild type when residual activity is sufficient to support normal growth. We favor this explanation for the observation that only 2/11 interface variants show reduced growth, as many pathogenic interface substitutions are associated with milder disease presentations, consistent with higher residual enzyme function. Consistent with this interpretation, variants affecting the active site, where substitutions are expected to cause large reductions in catalytic activity, are readily detected by the assay.
Although we cannot exclude partial buffering of dimerization defects in yeast, we interpret the reduced sensitivity to interface variants primarily as a general limitation of growth-based assays. Accordingly, our decision not to use growth >85% as evidence toward benignity is conservative relative to approaches that would classify high-growth variants as benign except at the monomer-monomer interface, avoiding reliance on structural subclassification and minimizing the risk of false benign interpretation. Reduced growth, by contrast, provides strong evidence of loss of ASS1 function and pathogenicity, validated under the OddsPath framework.
We have updated the Results and Discussion sections to clarify these points (also see response to the reviewer's point 1).
"A subset of clinically pathogenic ASS1 variants exhibit near-wild-type growth in our yeast assay. In general, we expect a monotonic relationship between ASS function and yeast growth, but with the potential for floor and ceiling effects that constrain the assay's dynamic range. In this context, we interpret high-growth pathogenic variants as likely causing mild loss of function that cannot be distinguished from wild type in our assay. Consistent with this view, many pathogenic variants with high assay growth are located at the monomer-monomer interface rather than the active site, and are associated with milder or later-onset clinical presentations, suggesting partial enzymatic impairment that is clinically relevant in humans but not resolved by the yeast assay."
"Based on these findings and given that 22/56 pathogenic variants show >85% growth, we conclude that growth above this threshold should not be used as evidence toward benignity. Notably, this approach is conservative relative to treating high-growth variants as benign except at the monomer-monomer interface, avoiding reliance on structural subclassification and minimizing the risk of false benign interpretation arising from assay ceiling effects. Conversely, the variants with
Reviewer #2 (Significance (Required)):
This study presents the first comprehensive mutational profiling of human ASS1 and would be of broad interest to clinical geneticists as well as those seeking biochemical insights into the enzymology of ASS1. The authors' use of a yeast system to profile human mutations would be particularly useful for researchers performing deep mutational scans, given that it provides functional insights in a rapid and inexpensive manner.
Reviewer #3 (Evidence, reproducibility and clarity (Required)):
Section 1 - Evidence, reproducibility, and clarity Summary This manuscript presents a comprehensive functional profiling of 2,193 ASS1 missense variants using a yeast complementation assay, providing valuable data for variant interpretation in the rare disease citrullinemia type I. The dataset is extensive, technically sound, and clinically relevant. The demonstration of intragenic complementation in ASS1 is novel and conceptually important. Overall, the study represents a substantial contribution to functional genomics and rare disease variant interpretation.
Major comments 1. This is an exciting paper as it can provide support to clinicians to make actionable decisions when diagnosing infants. I have a few major comments, but I want to emphasize the label of "functionally unimpaired" variants to be misleading. The authors explain that there are several pathogenic ClinVar variants that fall into this category (above the >.85 growth threshold) but I think this category needs a more specific name and I would ask the authors to reiterate the shortcomings of the assay again in the Discussion section.
We thank the reviewer for raising this important point. We agree that the label "functionally unimpaired" could be misleading if interpreted as implying clinical benignity rather than assay behavior. We have therefore clarified that this designation refers strictly to variant behavior in the yeast growth assay and does not imply absence of pathogenicity.
In addition, we have expanded the Discussion to explicitly address the existence of clinically pathogenic variants with high growth scores (>0.85), emphasizing that these likely reflect a ceiling effect of the assay and represent a key limitation for interpretation. This clarification reiterates that high-growth scores should not be used as evidence toward benignity, while reduced growth provides strong functional evidence of pathogenicity. Relevant revisions are described in our responses to Reviewers 1 and 2.
- I think there's an important discussion to be had here, is the assay detecting variants that alter the function of ASS or is it detecting a complete ablation of enzymatic activity? The results might be strengthened with a follow-up experiment that identifies stably expressed ASS1 variants.
We agree with the review that distinguishing between stability and enzyme activity would be valuable information. Unfortunately, we do not currently have the resources to perform this type of large-scale study. We have acknowledged in the text that our assay does not distinguish between enzyme activity and protein stability:
"We expect that the assay will detect reductions in both catalytic activity and protein stability, but will not distinguish between these mechanisms."
At the very least, it would be great to see the authors replicate some of their interesting results from the high-throughput screen by down-selecting to ~12 variants of uncertain significance that could be newly considered pathogenic.
We have included new analysis of all 25 VUS variants falling in the pathogenic range of our assay (Supplemental Table S7). Reclassification under current guidelines (in the absence of our data) shifts six variants to Pathogenic/Likely Pathogenic and 11 more are reclassified to Likely Pathogenic with the application of our functional data as PS3_Supporting. The remaining eight VUS are all reclassified to Likely Pathogenic when inclusion of homozygous PrimateAI-benign variants allows the assay to satisfy full PS3 criteria.
- I would ask the authors to provide more citations of the literature in the introduction of the manuscript. I would be especially interested in knowing more about human ASS being identified as a homolog of yeast ARG1, as they share little sequence similarity (27.5%) at the protein level. That said, I find the yeast complementation assay exciting.
We thank the reviewer for this suggestion. Human ASS and yeast Arg1 catalyze the same biochemical reaction and share approximately 49% amino acid sequence identity. We have revised the Introduction to clarify this relationship and to note explicitly that the Saccharomyces Genome Database (SGD) identifies the human gene encoding argininosuccinate synthase (ASS1) as the ortholog of yeast ARG1. An appropriate citation has been added to support this statement. The protein alignments have been provided as File S2.
"This assay is based on the ability of human ASS to functionally replace (complement) its yeast ortholog (Arg1) in S. cerevisiae (Saccharomyces Genome Database, 2026). Importantly, successful functional complementation indicates that ASS enzymatic activity does not depend on any obligate human-specific protein interactions. At the protein level, human ASS and yeast Arg1 display 49% sequence identity (File S2) and share identical enzymatic roles in converting citrulline and aspartate into argininisuccinate."
- I appreciate the efforts made by the authors to share their work and make this study more reproducible, such as sharing the hASS1 and yASS1 plasmids being shared on NCBI Genbank (Line 121) and publishing the ONT reads on SRA (Line 154). I made a requests for additional data to be shared, such as the custom method/code for codon optimization and a table of Twist variant cassettes that were ordered. I would also love to see these results shared on MaveDB.org.
We thank the reviewer for these suggestions regarding data sharing and reproducibility. As requested, we have provided the custom codon optimization script as File S1 and the amino acid alignment used to perform codon harmonization as File S2. The sequence of the underlying variant cassette is included in the corresponding GenBank entry, and we have clarified this point in the legend of Figure 1. For each amino acid substitution, Twist Bioscience used a yeast-specific codon scheme with a single consistent codon per amino acid; accordingly, the sequence of each variant cassette can be inferred from the base construct and the specified amino acid change. A complete list of variant amino acid substitutions used in this study is provided in Table S3.
- I find this manuscript very exciting as the authors have a compelling assay that identifies pathogenic variants, but I was generally disappointed by the quality and organization of the figures. For example, Figure 4 provides very little insight, but could be dramatically improved with an overlay of the normalized growth score data or highlighting variants surrounding the substrate or ATP interfaces. There are some very interesting aspects of this manuscript that could be shine through with some polished figures.
We thank the reviewer for this feedback and agree that clear and well-organized figures are essential for conveying the key results of the study. In response, we have substantially revised Figure 4 by adding colored overlays showing residue conservation and median normalized growth scores (new panels Figure 4C and 4D), which more directly link structural context to functional outcomes and highlight patterns surrounding the active site and substrate interfaces.
I would also encourage the authors to generate a heatmap of the data represented in Figure 2 (see Fowler and Fields 2014 PMID 25075907, Figure 2), this would be more helpful reference to the readers.
The reviewer also suggested that a heatmap representation, similar to that used in Fowler and Fields (2014), might aid interpretation of the data shown in Figure 2. Because our dataset consists of sparse single-amino acid substitutions rather than a complete mutational scan, such heatmaps are inherently less dense and less effective at conveying patterns than in saturation mutagenesis studies. Nevertheless, to aid readers who may find this visualization useful, we have generated and included a single-nucleotide variant heatmap as Supplemental Figure S1.
My major comments are as follows: 6. Citations needed - especially in the introduction and for establishing that hASS is a homolog of yARG1
We have added the requested citations and clarified the ASS1-ARG1 orthology in the Introduction, as described in our response to point 3 above.
- Generally, the authors do a nice job distinguishing the ASS1 gene from the ASS enzyme, though I found some ambiguities (Line 685). Please double-check the use of each throughout the manuscript.
We have edited the manuscript to ensure consistent and unambiguous use of gene and enzyme nomenclature throughout.
- Generally, I'm confused about what strain was used for integrating all these variants, was is the arg1 knock-out strain from the yeast knockout collection or was it FY4? I think FY4 was used for the preliminary experiments, then the KO collection strain was used for making the variant library but I think this could be made more clear in the text and figures. Lines 226-229 describes introducing the hASS1 and yASS1 sequences into the native ARG1 locus in strain FY4, but the Fig1A image depicts the ASS1 variants going into arg1 KO locus. Fig1A should be moved to Fig2.
We agree that the strain construction steps were not described as clearly as they could have been. We have therefore clarified the strain construction workflow in the Materials & Methods and Results sections, as well as in the Figure 1 legend, to explicitly distinguish preliminary experiments performed in strain FY4 from construction of the variant library in the arg1 knockout background.
As we have also added an additional panel to Figure 1 that schematically explains how the screen was performed (per Reviewer #2's request), we believe that Figure 1A is appropriately placed and should remain in Figure 1.
- Line 303 - "We classify these variants as 'functionally unimpaired'", this is not an accurate description of these variants as Figure 2 highlights 24 pathogenic ClinVar variants that would fall into this category of "functionally unimpaired". The yeast growth assay appears to capture pathogenic variants, but there is likely some nuance of human ASS functionality that is not being assessed here. I would make the language more specific, e.g. "complementary to Arg1" or "growth-compatible".
We agree that the label "functionally unimpaired" could be misinterpreted if read as implying clinical benignity. We have therefore clarified within the manuscript that this designation refers strictly to variant behavior in the yeast growth assay (i.e., wild-type-like growth under assay conditions) and does not imply absence of pathogenicity. We also expanded the Discussion to explicitly address the subset of clinically pathogenic variants with high growth scores (>0.85), consistent with a ceiling effect of the assay and a key limitation for interpretation. See response to reviewer #3 point 1. Relevant revisions are also discussed in our responses to Reviewers #1 and #2.
- Lines 345-355 - It is interesting that there are variants that appear functional at the substrate interfacing sites. Is there anything common across these variants? Are they maintaining the polarity or hydrophobicity of the WT residue? Are any of these variants included in ClinVar or gnomAD? Are pathogenic variants found at any of these sites
Yes. For highly sensitive active-site residues that have few permissible variants, the vast majority of amino acid substitutions that do retain activity preserve key physicochemical properties of the wild-type residue, such as hydrophobicity or charge. We have added this important observation to the manuscript:
"Any variants at these sensitive residues that are permissive for activity in our assay retain hydrophobicity or charged states relative to the original amino acid side chain (Figure 5A & Table S5)."
None of these variants are present in ClinVar. Only L15V and E191D are present in gnomAD (Table S4).
- Lines 423-430 - The OddsPath calculation would seem to rely heavily on the thresholds of .85 for normalized growth. The OddsPath calculation could be bolstered with some additional analysis that emphasizes the robustness to alternative thresholds.
We agree that the sensitivity of the OddsPath calculation to the choice of growth thresholds is an important consideration. In our assay, benign ClinVar variants and non-human primate variants are observed exclusively within the peak centered on wild-type growth, whereas clinically annotated variants falling below this peak are exclusively pathogenic. On this basis, we defined the upper boundary of the assay range interpreted as supporting pathogenicity as the lower boundary of the wild-type-centered peak in the growth distribution (as defined in Figure 3), rather than selecting a cutoff by direct optimization of the OddsPath. This choice reflects the observed concordance, in our dataset, between the onset of measurable functional impairment in the assay and clinical pathogenic annotation. Importantly, in practice the OddsPath value is locally robust to the precise placement of this boundary, remaining invariant across the range 0.82-0.88. Supporting our chosen threshold of 0.85, the lowest-growth benign or primate variant observed has a normalized growth value of 0.88, while the lowest growth observed among variants present as homozygotes in gnomAD was 0.86. We have clarified this rationale and analysis in the revised manuscript.
"Notably, the "Among all nine of the human ASS1 missense variants observed as homozygotes in gnomAD which were tested as amino acid substitutions in our assay, the lowest observed growth value was 0.86 (Ala258Val) consistent with the lower boundary of the PrimateAI variants which was a growth value of 0.87 (Ala81Thr) (Figure 6) and with our use of a 0.85 classification threshold."
"If we treat PrimateAI variants as benign (solely for OddsPath calculation purposes), the OddsPath for growth
- Lines 432-441 - This is an interesting idea to use variants observed in primates, has ACMG weighed in on this? I understand that CTLN1 is an autosomal recessive disorder but I'd still be interested in seeing how the observed ASS1 missense variants in gnomAD perform in your growth assay, possibly a supplemental figure?
To our knowledge, the ACMG/AMP guidelines do not currently address the use of homozygous missense variants observed in non-human primates. We are currently in discussion with two ClinGen working groups to discuss the possibility of formalizing the use of this data source.
We agree that comparison with human population data is also important. Accordingly, total gnomAD allele counts and homozygous counts for all applicable ASS1 missense variants are provided in Table S4, and the growth behavior of ASS1 missense variants observed in the homozygous state in gnomAD is shown in Figure 6. These homozygous variants uniformly exhibit high growth in our assay, consistent with the absence of strong loss-of-function effects. We have updated the manuscript text to clarify these points.
Minor comments 1. Lines 53-59 - This paragraph needs to cite the literature, especially lines 56, 57, and 59 2. Line 61 - no need to repeat "citrullinemia type I", just use the abbreviation as it was introduced in the paragraph above 3. Lines 61-71 - again, this paragraph needs more literature citations 4. Line 62 - change to "results"
The changes suggested in points 1-4 have all been implemented in the revised manuscript.
- Line 74-75 - "RUSP" acronym not needed as it's never used in the manuscript, the same goes for "HHS"
We agree that the acronyms "RUSP" and "HHS" are not reused elsewhere in the manuscript. We have nevertheless retained them at first mention, alongside the expanded names, because these acronyms are commonly used in newborn screening and public health policy contexts and may be more familiar to some readers than the expanded terms. We would be happy to remove the acronyms if preferred.
- Line 86 - "ASS1" I think is referring to the enzyme and should just be "ASS"? If referring to the gene then italicize to "ASS1"
- Lines 91-93 - It would be helpful to mention this is a functional screen in yeast
- Line 101 - It would be helpful to the readers to define SD before using the acronym, consider changing to "minimal synthetic defined (SD) medium" and afterwards can refer to as "SD medium"
- 109-114 - It would be great if you could share your method for designing the codon-harmonized yASS1 gene, consider sharing as a supplemental script or creating a GitHub repository linked to a Zenodo DOI for publication.
The changes suggested in points 6-9 have all been implemented in the revised manuscript. The codon harmonization script has been provided as File S1.
- Lines 135-137 - I think it's helpful to provide a full table of the cassettes ordered from Twist as well as the primers used to amplify them, consider a supplemental table.
Details of Twist cassette and the primer sequences used for amplification have been added to the Materials & Methods.
- Line 138 - "standard methods" is a bit vague, I'm guessing this is a Geitz and Schiestl 2007 LiAc/ssDNA protocol (PMID 17401334)? Also, was ClonNAT used to select for natMX colonies?
The reviewer is correct about which protocol was used, and we have added the citation. We have also clarified that selection was carried out based on resistance to nourseothricin.
- Line 150 - change to "sequence the entire open reading frame, as previously described [4]."
- Line 222-223 - remove "replace" and just use "complement" (and remove the parenthesis)
- Line 249 - It would be great to see a supplemental alignment of the hASS1 and yASS1 sequences.
- Line 261 - spelling "citrullemia" should be corrected to "citrullinemia"
- Line 280 - "using Oxford Nanopore sequencing" is a bit vague, I suggest specifying the equipment used (e.g. Oxford Nanopore Technologies MinION platform) or simplify to "via long-read sequencing (see Materials & Methods)"
The changes suggested in points 12-16 have all been implemented in the revised manuscript. An alignment of the ASS and Arg1 protein sequences has been provided as File S2.
- Line 287-289 - It would be great to see the average number of isolates per variant, as well as a plot of the variant growth estimate vs individual isolate growth.
We agree with the reviewer that conveying measurement precision is important. The number of isolates assayed per variant is provided in Table S4, and we have added explicit mention of this in the text. Because variants were assayed with a mixture of 1, 2, or {greater than or equal to}3 independent isolates, a scatterplot of variant-level growth estimates versus individual isolate measurements would be difficult to interpret and potentially misleading. Instead, we report standard error estimates for each variant in Table S4, derived from the linear model used to estimate growth effects, which more appropriately summarizes measurement uncertainty given the experimental design.
- Lines 324-25 - consider removing the last sentence of this paragraph, it is redundant as the following paragraph starts with the same statement.
We have removed this sentence.
- Lines 327-335 - This is interesting and would benefit from its own subpanel or plot in which the normalized growth score is plotted against variants that are at conserved or diverse residues in human ASS, and see if there's a statistical difference in score between the two groupings.
As suggested by the reviewer, we have added Supplemental Figure 2 (Figure S2) in which the normalized growth score of each variant is plotted against the conservation of the corresponding residue, as measured by ConSurf. The manuscript already includes a statistical analysis of the relationship between residue conservation and functional impact, showing that amorphic variants occur significantly more frequently at highly conserved residues than unimpaired variants do (one-sided Fisher's exact test). We now refer to this new supplemental figure in the relevant Results section.
- Lines 339-341 - As written, it is unclear if aspartate interacts with all of the same residues as citrulline or just Asn123 and Thr119.
- Lines 345-355 - As with my above comment, I find this interesting and would
- Line 353 - add a period to "al" in "Diez-Fernandex et al."
The issues raised in points 20 and 22 have all addressed. Point 21 appears to be truncated.
- Figure 1 a. Remove "Figure" from the subpanels and show just "A" and "B" (as you do for Figure 4) and combine the two images into a single image. Also make this correction to Figure 5 and Figure 8. b. Panel A - I thought the hASS1 and yASS1 were dropped into FY4, not the arg1 KO strain. This needs clarification. c. Panel A - I'm assuming the natMX cassette contains its own promoter, you could use a right-angled arrow to indicate where the promotors are in your construct. d. Panel B - I'm not sure the bar graph is necessary, it would be more helpful to see calculations of the colony size (or growth curves for each strain) and plot the raw values (maybe pixel counts?) for each replicate rather than normalizing to yeast ARG1. I would be great to have a supplemental figure showing all the replicates side-by-side. e. Panel B - Would be helpful to denote the pathogenic and benign ClinVar variants with an icon or colored text.
f. Figure 1 Caption - make "A)" and "B)" bold.
We have implemented the requested changes in Figure 1 with the following exceptions. We have retained panels A and B as separate subfigures because they illustrate distinct experimental concepts. In addition, we respectfully disagree with point (d). The bar graph is intended to provide a clear, high-level comparison of functional complementation by hASS1 versus yASS1 and to illustrate the gross differences in growth between benign and pathogenic proof-of-principle variants. As the bar graph includes error bars for standard deviations, presenting raw colony size measurements or growth curves for individual replicates would substantially complicate the figure without materially improving interpretability for this purpose.
- Figure 2 a. "Shown in magenta are amino acid substitutions corresponding to ClinVar pathogenic, pathogenic/likely pathogenic, and likely pathogenic variants" is repeated in the figure caption. b. "Shown in green are amino acid substitutions corresponding to ClinVar benign and likely benign variants." I don't see any green points. c. Identify the colors used for ASS1 substrate binding residues. d. This plot would benefit from a depiction of the human ASS secondary structure and any protein domains (nucleotide-binding domain, synthase domain, and C-terminal helix from Fig4B)
e. Line 685 675 - "ASS1" is being used in reference to the enzyme, is this correct or should it be "ASS"?
We have made the requested changes to Figure 2. The repeated caption text has been removed, and references to green points have been corrected to orange points to match the figure. The colors used to indicate ASS substrate-binding residues are explicitly described in the figure key. Secondary structure annotations have been added. References to the enzyme have been corrected to "ASS" rather than "ASS1" where appropriate.
- Figure 3 a. Rename the "unimpaired" category as there are several pathogenic ClinVar variants that fall into this category.
To address this point, we have clarified the labeling by adding "in our yeast assay" to the figure legend, making explicit that the "unimpaired" category refers only to wild-type-like behavior under assay conditions and does not imply clinical benignity. See also response to Reviewer #3, Major Comment 1.
- Figure 4 a. List the PDB or AlphaFold accession used for this structure b. Panel A - state which colors are used for to depict each monomer. It is confusing to see several shades of pink/purple used to depict a single monomer in Panel A. c. It is very difficult to make out the aspartate and citrulline substrates in the catalytic binding activity, consider making an inset zooming-in on this domain and displaying a ribbon diagram of the structure rather than the surface. d. Generally, it would be more helpful here to label any particular residues that were identified as pathogenic from your screen, or to overlay average grow scores per residue data onto the structure
We have implemented the requested changes to Figure 4. The relevant PDB/AlphaFold accession is now listed, and the colors used to depict each monomer in Panel A are clarified in the figure legend. An inset focusing on the active site has been added to improve visualization of the citrulline and aspartate substrates. In addition, we have added new panels (Figure 4C and 4D) overlaying pathogenic residues and average growth scores onto the structure to more directly link structural context with functional data.
- Figure 5 a. Line 716 - Insert a page break to place Figure 5 on its own page b. I suggest using a heatmap for this type of plot, as it is very difficult to track which color corresponds to which residue.
c. Fig5A - This plot could be improved by identifying which residue positions interface with which substrate.
We have placed Figure 5 on its own page and added information to the legend identifying which residue positions interface with each substrate. We have retained the active-site variant strip charts raised in point (b), as we believe they effectively illustrate how the distribution of variant effects differs between residues. In addition, we have provided a supplemental heatmap showing variant growth across the entire protein (Figure S1), and individual variant scores for all residues are provided in Table S4.
- Figure 7 a. Line 735 - Insert page break to place figure on a new page
List the PDB accession used for these images. c. For clarity I would mention "human ASS" in the figure title d. State the colors of the substrates e. Panels A and B could be combined into a single panel, making it easier to distinguish the active site and dimerization variants.
f. Could be interesting to get SASA scores for the ClinVar structural variants to determine if they are surface-accessible
We have implemented the requested changes in Figure 7 with the following exceptions. For point (e), there is no single orientation of the structure that allows a clear simultaneous view of both active-site and dimerization variants; accordingly, we have retained panels A and B as separate subfigures to preserve clarity. With respect to point (f), we agree that solvent accessibility analysis could be informative in other contexts. However, such an analysis does not integrate naturally with the functional and assay-based framework of the present study and was therefore not included.
- Figure 8 a. Panel B - overlay a square frame in the larger protein structure that depicts where the below inset is focused, and frame inset image as well.
We have framed the inset image as requested. We did not add a corresponding frame to the full protein structure, as doing so obscured structural details in the region of interest.
Reviewer #3 (Significance (Required)):
Section 2 - Significance This study represents a substantial technical, functional, and translational advance in the interpretation of missense variation in ASS1, a gene of high clinical relevance for the rare disease citrullinemia type I. Its principal strength lies in the generation of an experimentally validated functional atlas of ASS1 missense variants that covers ~90% of all SNV-accessible substitutions. The scale, internal reproducibility, and careful benchmarking of the yeast complementation assay against known pathogenic and benign variants provide a robust foundation for identifying pathogenic ASS1 variants. Particularly strong aspects include the rigorous quality control of variant identities, the quantitative nature of the functional readout, and the thoughtful integration of results into the ACMG/AMP OddsPath framework. The discovery of intragenic complementation between variants affecting distinct structural regions of the enzyme is a notable conceptual and mechanistic contribution. Limitations include the assay's reduced sensitivity to variants impacting oligomerization or subtle folding defects, and the use of yeast as a heterologous system, which may mask disease-relevant mechanisms as several pathogenic ClinVar variants were found to be "functionally unimpaired". Future work extending functional testing to additional cellular contexts or expanding genotype-level combinatorial analyses would further enhance clinical applicability. Relative to prior studies, which have relied on small numbers of patient-derived variants or low-throughput biochemical assays, this work extends the field decisively by delivering a comprehensive, variant-resolved functional map for ASS1. To the best of my current knowledge, this is the first systematic functional screen of ASS1 at this scale and the first direct experimental demonstration that ASS active sites span multiple subunits, enabling intragenic complementation consistent with Crick and Orgel's classic variant sequestration model. As such, the advance is simultaneously technical (high-throughput functional genomics), mechanistic (defining structural contributors to catalysis and epistasis), and clinical (enabling evidence-based reclassification of VUS). I find the use of homozygous non-human primate variants as an orthogonal benign calibration set both creative and controversial, my hope would be that this manuscript will prompt a productive discussion.
-
Note: This preprint has been reviewed by subject experts for Review Commons. Content has not been altered except for formatting.
Learn more at Review Commons
Referee #2
Evidence, reproducibility and clarity
In this manuscript, Lo et al characterize the phenotypic effect of ~90% of all possible ASS1 missense mutations using an elegant yeast-based system, and use this dataset to aid the interpretation of clinical ASS1 variants. Overall, the manuscript is well-written and the experimental data are interpretated rigorously. Of particular interest is the identification of pairs of deleterious alleles that rescue ASS1 activity in trans. My comments mainly pertain to the relevance of using a yeast screening methodology to infer functional effects of human ASS1 mutations.
- Since human ASS1 is heterologously expressed in yeast for this mutational screen, direct comparison of native expression levels between human cells and yeast is not possible. Could the expression level of human ASS1 (driven by the pARG1 promoter) in yeast alter the measured fitness defect of each variant? For instance, if ASS1 expression in yeast is sufficiently high to mask modest reductions in catalytic activity, such variants may be misclassified as hypomorphic rather than amorphic. Conversely, if expression is intrinsically low, even mild catalytic impairments could appear deleterious. While it is helpful that the authors used non-human primate SNV data to calibrate their assay, experiments could be performed to directly address this possibility.
- The nature of the relationship between yeast growth and availability of functional ASS1 could also influence the interpretation of results from the yeast-based screen. Does yeast growth scale proportionately with ASS1 enzymatic activity?
- It would be helpful to add an additional diagram to Figure 1A explaining how the screen was performed, in particular: when genotype and phenotype were measured, relative to plating on selective vs non-selective media? This is described in "Variant library sequence confirmation" and "Measuring the growth of individual isolates" of the Methods section but could also be distilled into a diagram.
- The authors rationalize the biochemical consequences of ASS1 mutations in the context of ASS1 per se - for example, mutations in the active site pocket impair substrate binding and therefore catalytic activity, which is expected. Does ASS1 physically interact with other proteins in human cells, and could these interactions be altered in the presence of specific ASS1 mutations? Such effects may not be captured by performing mutational scanning in yeast.
- The authors note that only a small number (2/11) of mutations at the ASS1 monomer-monomer interface lead to growth defects in yeast. It would be helpful for the authors to discuss this further.
Significance
This study presents the first comprehensive mutational profiling of human ASS1 and would be of broad interest to clinical geneticists as well as those seeking biochemical insights into the enzymology of ASS1. The authors' use of a yeast system to profile human mutations would be particularly useful for researchers performing deep mutational scans, given that it provides functional insights in a rapid and inexpensive manner.
-
-
k51qzi5uqu5dm9g6q2l275rmato8ce47dz8cn4di3nglvgpjqd93p2rubxv5t3.ipns.inbrowser.link k51qzi5uqu5dm9g6q2l275rmato8ce47dz8cn4di3nglvgpjqd93p2rubxv5t3.ipns.inbrowser.link
-
inbrowser.link
as long as the IPNS used get refreshed regularly these links will work
https://k51qzi5uqu5dm9g6q2l275rmato8ce47dz8cn4di3nglvgpjqd93p2rubxv5t3.ipns.inbrowser.link/%F0%9F%A7%8A.%E2%99%96.indy.html
-
-
accessmedicine.mhmedical.com accessmedicine.mhmedical.com
-
a FD
La colinesterasa es una enzima cuya función es:
👉 Degradar (romper) la acetilcolina En la EA ocurre que:
Se pierden neuronas colinérgicas, Como la acetilcolina ya está baja, si la colinesterasa la destruye rápido → empeora el déficit.
-
Las mutaciones causantes de enfermedades en PSEN1, PSEN2 o APP causan EA.
es decir que mutaciones en estos genes pueden causar enfermedades como aumento de β-amiloide anormal, lo que favorece:
Placas amiloides
Daño neuronal
Alzheimer, generalmente de inicio temprano
-
-
docdrop.org docdrop.org
-
I można na całą stronę dać notatki
-
-
tomasurzuam.github.io tomasurzuam.github.io
-
Por último, con el objetivo de ahondar en una caracterización por carreras, este estudio tendrá una segunda sección de resultados con la unidad de análisis centrada en los cursos.
- posibles futuros pasos: mirar hacia pre-pandemia, link con evaluación docente
-
-
tomasurzuam.github.io tomasurzuam.github.io
-
Lista de figuras 4.5: Distribución de notas por carrera según tipo de colegio 2021 - 2024
la primera es de la facultad?
-
-
tomasurzuam.github.io tomasurzuam.github.io
-
Resumen ejecutivo del estudio.
abajo también en un párrafo (o call out) sobre temas de apertura y reproducibilidad del reporte
y también el gráfico de la estructura del repo comentado (o su link al readme. Mejor, que el readme sea idéntico al resumen ejecutivo
-
Estudio de Notas de la Facultad de Ciencias Sociales
pasaste todo x grammarly?
-
-
www.youtube.com www.youtube.com
-
Briefing : L'émancipation de l'Éducation nationale face au monopole de Microsoft
Ce document synthétise les enjeux de la dépendance technologique de l'Éducation nationale française envers Microsoft et l'émergence d'une alternative structurée autour du logiciel libre et de la collaboration enseignante.
Résumé Exécutif
L'Éducation nationale française fait face à une dépendance coûteuse et structurelle vis-à-vis des solutions propriétaires, principalement Microsoft.
Le passage imposé de Windows 10 à Windows 11 illustre cette vulnérabilité : l'obsolescence logicielle pourrait coûter jusqu'à un milliard d'euros à l'échelle nationale pour le renouvellement du parc informatique.
Face à ce constat, une "guérilla" de l'open source s'organise. Portée par la Direction du numérique pour l'éducation (DNE) et des initiatives comme « La Forge », cette dynamique mobilise désormais 10 000 enseignants-développeurs.
L'objectif est de substituer aux licences onéreuses des « communs numériques » (Linux, BigBlueButton, NextCloud), garantissant la souveraineté des données, la pérennité des investissements publics et une pédagogie adaptée aux besoins réels du terrain.
--------------------------------------------------------------------------------
1. Le constat d'une dépendance critique : Le "cas d'école" Microsoft
La relation entre l'institution scolaire et Microsoft est décrite comme une forme d'addiction budgétaire et technique.
Le coût de l'obsolescence imposée
• L'exemple des Hauts-de-France : Suite à une cyberattaque par ransomware, la région a dû envisager la migration vers Windows 11.
Un membre de la DSI a estimé à 100 millions d'euros le coût pour renouveler 30 000 PC incapables de supporter cette mise à jour.
• Extrapolation nationale : Les Hauts-de-France représentant environ 10 % de l'éducation nationale, le coût total pour la mise à jour forcée du parc (300 000 machines) est estimé à 1 milliard d'euros.
• La vente liée : Le monopole s'appuie sur le mécanisme de la vente liée, où le système d'exploitation est pré-installé sans distinction de prix entre le matériel et le logiciel, imposant une solution "clé en main" qui freine l'adoption d'alternatives.
Limites des services propriétaires
• Coûts récurrents : Des dizaines de millions d'euros sont versés chaque année en licences.
• Failles systémiques : La crise du Covid-19 a révélé les carences du système numérique éducatif, notamment sa dépendance à des solutions propriétaires onéreuses et son manque de cohérence globale.
--------------------------------------------------------------------------------
2. La stratégie de rupture par le Logiciel Libre
Face au monopole, des solutions basées sur Linux et l'open source prouvent leur viabilité sur le terrain.
Distributions Linux dédiées à l'éducation
Il existe des alternatives robustes permettant d'adapter l'ordinateur aux besoins pédagogiques :
• PrimTux : Système d'exploitation spécifique pour les écoles primaires.
• ND (Numérique Inclusif, Responsable et Durable) : Distribution destinée au secondaire.
Obstacles et leviers d'adoption
| Obstacle | État des lieux | Perspectives | | --- | --- | --- | | Logiciels métiers | Certains éditeurs (SVT, physique, techno) ne développent que pour Windows. | Pression par la masse : l'augmentation du parc Linux doit forcer les éditeurs à s'adapter. | | Logiciels de vie scolaire | Pronote dispose d'un client Windows complet mais d'une version web dégradée sous Linux. | Nécessité d'une évolution des clients vers des standards interopérables. | | Résilience | En cas d'attaque (ransomware), les systèmes sous Windows ont été paralysés. | Des lycées sous Linux (ex: Lycée Carnot à Bruay-la-Buissière) ont pu proposer leur aide et leurs outils. |
--------------------------------------------------------------------------------
3. « La Forge » : L'industrialisation de l'innovation enseignante
« La Forge » représente un changement de paradigme : passer de l'enseignant "bricoleur" isolé à une communauté structurée de développeurs au sein de l'État.
Un modèle collaboratif massif
• Effectifs : 10 000 enseignants inscrits.
• Volume : 6 500 projets (dépôts de code) enregistrés.
• Fonctionnement : Outil de travail collaboratif (basé sur le modèle GitHub) permettant de fédérer, tester et partager des codes sources et des ressources pédagogiques.
Exemples de projets emblématiques
• MindStory : Alternative open source à Minecraft, permettant aux élèves de collaborer sur des constructions sans dépendre d'un compte Microsoft payant.
• Philo GPT : Interface permettant de dialoguer avec des simulations de grands philosophes.
• Execubot : Outil d'apprentissage de la programmation via un robot virtuel.
• Créa-appli : Outil utilisant l'IA pour aider les profs à générer des prototypes d'applications (HTML/JS) via le "vibe coding" (codage par prompt).
--------------------------------------------------------------------------------
4. Souveraineté, Communs Numériques et Commande Publique
L'enjeu n'est pas seulement technique, il est politique et financier : assurer que l'argent public finance des biens publics.
La notion de "Communs Numériques"
Un commun numérique repose sur trois piliers : une ressource, une communauté et une gouvernance. L'idée est que l'amélioration d'un logiciel par le ministère bénéficie à tous.
Les services souverains déjà déployés
Le ministère opère et héberge ses propres instances de logiciels libres pour s'affranchir des GAFAM :
• BigBlueButton : Alternative à Zoom/Meet pour la visioconférence (participation financière du ministère au développement du projet global).
• Apps.education.fr : Portail regroupant des outils comme Tube (alternative à YouTube basée sur PeerTube) ou NextCloud (alternative à Google Drive).
Critique du modèle traditionnel de commande publique
Par le passé, l'État stimulait les start-ups ("EdTech") via des marchés publics sans exiger la propriété intellectuelle :
1. Les entreprises conservaient le code source et les données.
2. L'État devait payer des abonnements pour continuer à utiliser ce qu'il avait financé.
3. Résultat : Aucune capitalisation sur le long terme.
La nouvelle approche privilégie la pérennité : Investir dans l'open source permet à l'institution de conserver la maîtrise de ses outils, même après la fin d'un contrat avec un prestataire.
--------------------------------------------------------------------------------
Citations Clés
« L'éducation nationale est accro à Microsoft. Chaque année, des dizaines de millions d'euros s'envolent en licences. »
« Le slogan de la forge c'est : "L'union fait la forge". »
« On a oublié que nos profs étaient aussi capables de fabriquer leurs propres ressources... On a passé des marchés avec ces EdTech où on n'exigeait rien en termes de propriété intellectuelle. Les boîtes repartaient avec l'ensemble du code. »
« Un milliard pour faire une mise à jour de système d'exploitation qui était imposée par Microsoft parce que Microsoft a déclaré qu'ils arrêtent le support de Windows 10. »
-
-
tomasurzuam.github.io tomasurzuam.github.io
-
El rendimiento académico ha representado históricamente uno de los elementos constitutivos de la educación superior, siendo el mejor indicador para medir el logro de los aprendizajes de los estudiantes (Garbanzo Vargas, 2013). En este sentido, las calificaciones otorgan un tipo de credencial a quienes cursan una carrera universitaria al demostrar empíricamente si los contenidos impartidos fueron o no aprehendidos.
esto lo incorporaría al párrafo 3 actual, y partiría con el 2 actual
-
-
telecty-docs.vercel.app telecty-docs.vercel.app
-
sesión conocid
attach/dettach. es la ultima sesion attache. snapshot
-
-
telecty-docs.vercel.app telecty-docs.vercel.app
-
ventana temporal.
hoy por sim recorre el mes, si esta guardado no lo toca sino sigo elcon el que sigue) -> analizar con ia como es mejor. de cada icc se pide el consumo por dia de todo el mes, lo ultimos dias del mes se hace pesado. todos los dias se rechequea. generalmente falla el dia de atras. se necesita cada 10 minutos
-
lemetry valida versión, tipo de evento e identificadores míni
la normalizacion gruesa de esto es de provider no?
-
eventos en la cola telemetry-ingest.
varias colas una por proceso
-
Provider ejecuta un proceso batch consultando a la telco por una ventana temporal.
son 4: cdr, sms, location, eventos de red ( pasa algo con el sim:{ "_id": { "$oid": "68c9d39139836c2793527aa5" }, "iccid": "89444611503504960517", "current_country": "Argentina", "current_location": "722310", "current_network": "AMX Argentina S.A.", "domain": "S6a (4G)", "imsi": "204046848359268", "operation": "Location Update", "result": "success", "roaming_profile": "GREENFIVE", "timestamp": { "$date": "2025-09-11T17:18:46.000Z" }, "timestamp_str": "2025-09-11T17:18:46", "synced_at": { "$date": "2025-09-16T18:16:01.575Z" } }
-
ma asíncro
falta el paso de guardar
-
n evento técnico desde la telco vía webhook.
sms recibido y iniciar y terminar sesion hoy se esta tomando con movistar.
-
Provider publica el evento en la cola telemetry-ingest usando un envelope versionado.
tipar la cola para que luego se banque otra cosa que no sea sms
-
-
noblogo.org noblogo.org
-
Bender también argumenta que aún podemos, como sociedad, influenciar el impacto que pueda tener la inteligencia artificial en nuestras vidas.
Depende del factor humano el uso que se le da a la IA. Es distinto querer usar la escritura para evitar memorizar que utilizar la escritura para tener un registro para futuras generaciones. De la misma manera, es distinto usar la inteligencia artificial para resolver una tarea que usarla para brindarnos métodos de aprendizaje. La regulación debe existir, pero, en especial, una autoregulación dentro de la cual no le deleguemos nuestra vida entera a la máquina.
-
-
pavels130.github.io pavels130.github.io
-
Teorema de Gronwal
Añadir a la lista de preliminares con referencia cruzada.
-
-
-
imilar to wild-type animals, the germline rde-1 rescue strainshowed germline defects upon the gld-1 RNAi treatment with around 50% penetrance (Fig. 2A, Table 1). egg-5encodes a pseudo-tyrosine phosphatase in oocytes.
50% penetrance is expected for RNAi, and it confirms that RNAis active in the germline.
-
-
shs-cairn-info.ezproxy.u-paris.fr shs-cairn-info.ezproxy.u-paris.fr
-
Les sujets du groupe co-éprouvent à partir des affects partagés et ont la possibilité d’entrer en résonance affective, fantasmatique, inconsciente, spontanée et immédiate avec les autres psychismes. C’est là que se situe le lien inconscient commun le plus profond entre les membres du groupe.
!!!
-
-
www.biorxiv.org www.biorxiv.org
-
Author response:
The following is the authors’ response to the current reviews.
Public Reviews:
Reviewer #1 (Public review):
Summary:
The manuscript by Yamamoto et al. presents a model by which the four main axes of the limb are required for limb regeneration to occur in the axolotl. A longstanding question in regeneration biology is how existing positional information is used to regenerate the correct missing elements. The limb provides an accessible experimental system by which to study the involvement of the anteroposterior, dorsoventral, and proximodistal axes in the regenerating limb. Extensive experimentation has been performed in this area using grafting experiments. Yamamoto et al. use the accessory limb model and some molecular tools to address this question. There are some interesting observations in the study. In particular, one strength the potent induction of accessory limbs in the dorsal axis with BMP2+Fgf2+Fgf8 is very interesting. Although interesting, the study makes bold claims about determining the molecular basis of DV positional cues, but the experimental evidence is not definitive and does not take into account the previous work on DV patterning in the amniote limb. Also, testing the hypothesis on blastemas after limb amputation would be needed to support the strong claims in the study.
Strengths:
The manuscript presents some novel new phenotypes generated in axolotl limbs due to Wnt signaling. This is generally the first example in which Wnt signaling has provided a gain of function in the axolotl limb model. They also present a potent way of inducing limb patterning in the dorsal axis by the addition of just beads loaded with Bmp2+Fgf8+Fgf2.
Comments on revised version:
Re-evaluation: The authors have significantly improved the manuscript and their conclusions reflect the current state of knowledge in DV patterning of tetrapod limbs. My only point of consideration is their claim of mesenchymal and epithelial expression of Wnt10b and the finding that Fgf2 and Wnt10b are lowly expressed. It is based upon the failed ISH, but this doesn't mean they aren't expressed. In interpreting the Li et al. scRNAseq dataset, conclusions depend heavily on how one analyzes and interprets it. The 7DPA sample shows a very low representation of epithelial cells compared to other time points, but this is likely a technical issue. Even the epithelial marker, Krt17, and the CT/fibroblast marker show some expression elsewhere. If other time points are included in the analysis, Wnt10b, would be interpreted as relatively highly expressed almost exclusively in the epithelium. By selecting the 7dpa timepoint, which may or may not represent the MB stage as it wasn't shown in the paper, the conclusions may be based upon incomplete data. I don't expect the authors to do more work, but it is worth mentioning this possibility. The authors have considered and made efforts to resolve previous concerns.
We are grateful for the constructive comments. As Reviewer #1 suggested, we noted that clearer expression patterns of Wnt10b and Fgf2 may be detectable in scRNA-seq analyses at other stages, and we also clarified that low-level signals of epithelial and CT/fibroblast markers outside their expected clusters may reflect technical bias in the Discussion section. In addition, we agree with the reviewer’s point that our unsuccessful ISH experiments and the low abundance detected by RT-qPCR do not demonstrate absence of expression, and that conclusions from reanalyzing the Li et al. scRNA-seq dataset can depend strongly on analytical choices; therefore, while we focused on the 7 dpa sample because our RT-qPCR data suggested that Wnt10b and Fgf2 may be most enriched around the MB stage (the original study refers to 7 dpa as MB), we explicitly acknowledged that analyzing a single time point—especially one with a low representation of epithelial cells—may yield incomplete or stage-biased interpretations, and that inclusion of additional datasets could reveal clearer and potentially different expression patterns in the Discussion section. We also tempered our wording regarding the inferred cellular sources to avoid over-interpretation based on the current data in the Results section.
Reviewer #2 (Public review):
Summary:
This study explores how signals from all sides of a developing limb, front/back and top/bottom, work together to guide the regrowth of a fully patterned limb in axolotls, a type of salamander known for its impressive ability to regenerate limbs. Using a model called the Accessory Limb Model (ALM), the researchers created early staged limb regenerates (called blastemas) with cells from different sides of the limb. They discovered that successful limb regrowth only happens when the blastema contains cells from both the top (dorsal) and bottom (ventral) of the limb. They also found that a key gene involved in front/back limb patterning, called Shh (Sonic hedgehog), is only turned on when cells from both the dorsal and ventral sides come into contact. The study identified two important molecules, Wnt10B and FGF2, that help activate Shh when dorsal and ventral cells interact. Finally, the authors propose a new model that explains how cells from all four sides of a limb, dorsal, ventral, anterior (front), and posterior (back), contribute at both the cellular and molecular level to rebuilding a properly structured limb during regeneration.
Strengths:
The techniques used in this study, like delicate surgeries, tissue grafting, and implanting tiny beads soaked with growth factors, are extremely difficult, and only a few research groups in the world can do them successfully. These methods are essential for answering important questions about how animals like axolotls regenerate limbs with the correct structure and orientation. To understand how cells from different sides of the limb communicate during regeneration, the researchers used a technique called in situ hybridization, which lets them see where specific genes are active in the developing limb. They clearly showed that the gene Shh, which helps pattern the front and back of the limb, only turns on when cells from both the top (dorsal) and bottom (ventral) sides are present and interacting. The team also took a broad, unbiased approach to figure out which signaling molecules are unique to dorsal and ventral limb cells. They tested these molecules individually and discovered which could substitute for actual dorsal and ventral cells, providing the same necessary signals for proper limb development. Overall, this study makes a major contribution to our understanding of how complex signals guide limb regeneration, showing how different regions of the limb work together at both the cellular and molecular levels to rebuild a fully patterned structure.
Weaknesses:
Because the expressional analyses are performed on thin sections of regenerating tissue, in the original manuscript, they provided only a limited view of the gene expression patterns in their experiments, opening the possibility that they could be missing some expression in other regions of the blastema. Additionally, the quantification method of the expressional phenotypes in most of the experiments did not appear to be based on a rigorous methodology. The authors' inclusion of an alternate expression analysis, qRT-PCR, on the entire blastema helped validate that the authors are not missing something in the revised manuscript.
Overall, the number of replicates per sample group in the original manuscript was quite low (sometimes as low as 3), which was especially risky with challenging techniques like the ones the authors employ. The authors have improved the rigor of the experiment in the revised manuscript by increasing the number of replicates. The authors have not performed a power analysis to calculate the number of animals used in each experiment that is sufficient to identify possible statistical differences between groups. However, the authors have indicated that there was not sufficient preliminary data to appropriately make these quantifications.
Likewise, in the original manuscript, the authors used an AI-generated algorithm to quantify symmetry on the dorsal/ventral axis, and my concern was that this approach doesn't appear to account for possible biases due to tissue sectioning angles. They also seem to arbitrarily pick locations in each sample group to compare symmetry measurements. There are other methods, which include using specific muscle groups and nerve bundles as dorsal/ventral landmarks, that would more clearly show differences in symmetry. The authors have now sufficiently addressed this concern by including transverse sections of the limbs annd have explained the limitations of using a landmark-based approach in their quantification strategy.
We are grateful for the careful evaluation of the technical rigor and quantification. We have benefited from the reviewer’s earlier feedback, which guided revisions that improved the manuscript’s rigor and presentation.
Reviewer #3 (Public review):
Summary:
After salamander limb amputation, the cross-section of the stump has two major axes: anterior-posterior and dorsal-ventral. Cells from all axial positions (anterior, posterior, dorsal, ventral) are necessary for regeneration, yet the molecular basis for this requirement has remained unknown. To address this gap, Yamamoto et al. took advantage of the ALM assay, in which defined positional identities can be combined on demand and their effects assessed through the outgrowth of an ectopic limb. They propose a compelling model in which dorsal and ventral cells communicate by secreting Wnt10b and Fgf2 ligands respectively, with this interaction inducing Shh expression in posterior cells. Shh was previously shown to induce limb outgrowth in collaboration with anterior Fgf8 (PMID: 27120163). Thus, this study completes a concept in which four secreted signals from four axial positions interact for limb patterning. Notably, this work firmly places dorsal-ventral interactions upstream of anterior-posterior, which is striking for a field that has been focussed on anterior-posterior communication. The ligands identified (Wnt10b, Fgf2) are different to those implicated in dorsal-ventral patterning in the non-regenerative mouse and chick models. The strength of this study is in the context of ALM/ectopic limb engineering. Although the authors attempt to assay the expression of Wnt10b and Fgf2 during limb regeneration after amputation, they were unable to pinpoint the precise expression domains of these genes beyond 'dorsal' and 'ventral' blastema. Given that experimental perturbations were not performed in regenerating limbs - almost exclusively under ALM conditions - this author finds the title "Dorsoventral-mediated Shh induction is required for axolotl limb regeneration" a little misleading.
Strengths:
(1) The ALM and use of GFP grafts for lineage tracing (Figures 1-3) take full advantage of the salamander model's unique ability to outgrow patterned limbs under defined conditions. As far as I am aware, the ALM has not been combined with precise grafts that assay 2 axial positions at once, as performed in Figure 3. The number of ALMs performed in this study deserves special mention, considering the challenging surgery involved.
(2) The authors identify that posterior Shh is not expressed unless both dorsal and ventral cells are present. This echoes previous work in mouse limb development models (AER/ectoderm-mesoderm interaction) but this link between axes was not known in salamanders. The authors elegantly reconstitute dorsal-ventral communication by grafting, finding that this is sufficient to trigger Shh expression (Figure 3 - although see also section on Weaknesses).
(3) Impressively, the authors discovered two molecules sufficient to substitute dorsal or ventral cells through electroporation into dorsal- or ventral- depleted ALMs (Figure 5). These molecules did not change the positional identity of target cells. The same group previously identified the ventral factor (Fgf2) to be a nerve-derived factor essential for regeneration. In Figure 6, the authors demonstrate that nerve-derived factors, including Fgf2, are alone sufficient to grow out ectopic limbs from a dorsal wound. Limb induction with a 3-factor cocktail without supplementing with other cells is conceptually important for regenerative engineering.
(4) The writing style and presentation of results is very clear.
Overall appraisal:
This is a logical and well-executed study that creatively uses the axolotl model to advance an important framework for understanding limb patterning. The relevance of the mechanisms to normal limb regeneration are not yet substantiated, in the opinion of this reviewer. Additionally, Wnt10b and Fgf2 should be considered molecules sufficient to substitute dorsal and ventral identity (solely in terms of inducing Shh expression). It is not yet clear whether these molecules are truly necessary (loss of function would address this).
Comments on revisions:
Congratulations - I still find this an elegant and easy-to-read study with significant implications for the field! Linking your mechanisms to normal limb regeneration (i.e. regenerating blastema, not ALM), as well as characterising the cell populations involved, will be interesting directions for the future.
We are grateful for the constructive comments. To mitigate the concerns raised by Reviewer #3, we cited a previous study suggesting that ALM was used as the alternative experimental system for studying limb regeneration (Nacu et al., 2016, Nature, PMID: 27120163; Satoh et al., 2007, Developmental Biology, PMID: 17959163) in the Introduction section. We are confident that our ALM-based data provide a reasonable basis for understanding limb regeneration. We agree that there are important remaining questions—such as which cell populations express Wnt10b and Fgf2 and how endogenous WNT10B and FGF2 signals induce Shh expression in normal regeneration—which should be investigated in future studies to deepen our understanding of limb regeneration.
The following is the authors’ response to the original reviews.
Recommendations for the authors:
Reviewing Editor Comments:
The authors should be commended for addressing this gap - how cues from the DV axis interact with the AP axis during limb regeneration. Overall, the concept presented in this manuscript is extremely interesting and could be of high value to the field. However, the manuscript in its current form is lacking a few important data and resolution to fully support their conclusions, and the following needs to be addressed before publication:
(1) ISH data on Wnt10b and FGF2 from various regeneration time points are essential to derive the conclusion. Preferably multiplex ISH of Wnt10b/Fgf2/Shh or at least canonical ISH on serial sections to demonstrate their expression in dermis/epidermis and order of gene expression i.e. Shh is only expressed after expression of Wnt10b/FGF2. It would certainly help if this can also be shown in regular blastema.
We are grateful for the constructive suggestion on assessing Wnt10b and Fgf2 expression during regular regeneration, and we agree that clarifying their expression patterns in regular blastemas is important for strengthening the conclusions of our study. Because we cannot currently ensure sufficient sensitivity with multiplex FISH in our laboratory—partly due to high background—, we conducted conventional ISH on serial sections of regular blastemas at several time points (Fig. S5A). However, the expression patterns of Wnt10b and Fgf2 were not clear. To complement the ISH results, we performed RT-qPCR on microdissected dorsal and ventral halves of regular blastemas at the MB stage (Fig. S5B). We found that Wnt10b and Fgf2 were expressed at significantly higher levels in the dorsal and ventral halves, respectively, compared to the opposite half. This dorsal/ventral biased expression of Wnt10b/Fgf2 is consistent with our RNA-seq data. We further quantified expression levels of Wnt10b, Fgf2, and Shh across stages (intact, EB, MB, LB, and ED) and found that Wnt10b and Fgf2 peaked at the MB stage, whereas Shh peaked at the LB stage—consistent with the editor’s request regarding the order of gene expression (Fig. S5C). This temporal offset in upregulation supports our model. These results are now included in the revised manuscript (Line 294‒306).
To identify the cell types expressing Wnt10b or Fgf2, we analyzed published single-cell RNA-seq data (7 dpa blastema (MB), Li et al., 2021). As a result, Fgf2 expression was observed in the mesenchymal cluster, whereas Wnt10b expression was observed in both mesenchymal and epithelial clusters (Fig. S6). However, because only a small fraction of cells expressed Wnt10b, the principal cellular source of WNT10B protein remains unclear. The apparent low abundance likely contributes to the weak ISH signals and reflects current technical limitations. In addition, Wnt10b and Fgf2 expression did not follow Lmx1b expression (Fig. S6J, K), and Wnt10b and Fgf2 themselves were not exclusive (Fig. S6L). These results are now included in the revised manuscript (Line 307‒321). Together with the RT-qPCR data (Fig. S5B), these results suggest that Wnt10b and Fgf2 are not exclusively confined to purely dorsal or ventral cells at the single-cell level, even though they show dorsoventral bias when assessed in bulk tissue. These results suggest that Wnt10b/Fgf2 expression is not restricted to dorsal/ventral cells but mediated by dorsal/ventral cells, and co-existence of both signals should provide a permissive environment for Shh induction. Defining the precise spatial patterns of Wnt10b and Fgf2 in regular regeneration will therefore be an important goal for future work.
(2) Validation of the absence of gene expression via qRT PCR in the given sample will increase the rigor, as suggested by reviewers.
We thank for this important suggestion and agree that validation by qRT-PCR increases the rigor of our study. Accordingly, we performed RT-qPCR on AntBL, PostBL, DorBL, and VentBL to corroborate the ISH results. The results are now included in Fig. 2. We also verified by RT-qPCR that Shh expression following electroporation and the quantitative results are now provided in Fig. 5.
(3) Please increase n for experiments where necessary and mention n values in the figures.
We thank for this helpful comment and agree on the importance of providing sufficient sample sizes. Accordingly, we increased the n for the relevant experiments and have indicated the n values in the corresponding figure legends.
(4) Most comments by all three reviewers are constructive and largely focus on improving the tone and language of the manuscript, and I expect that the authors should take care of them.
We thank the reviewers for their constructive feedback on the tone and language of the manuscript. We have carefully revised the text according to each comment, and we hope these modifications have improved both clarity and readability.
In addition, in revising the manuscript we also refined the conceptual framework. Our new analysis of Wnt10b and Fgf2 expression during normal regeneration suggests that these genes are not expressed in a strictly dorsal- or ventral-specific manner at the single-cell level. When these observations are considered together with (i) the RNA-seq comparison of dorsally and ventrally induced ALM blastemas, (ii) RT-qPCR of microdissected dorsal and ventral halves of regenerating blastemas, and (iii) the functional electroporation experiments, our interpretation is that Wnt10b and Fgf2 act as dorsal- and ventral-mediated signals, respectively: their production is regulated by dorsal or ventral cells, and the presence of both signals is required to induce Shh expression. Given those, we now think our conclusion might be explained without using the confusing term, “positional cue”. Because the distinction between “positional cue” and “positional information” could be confusing as noted by the reviewers, we rewrote our manuscript without using “positional cue.”
Reviewer #1 (Recommendations for the authors):
(1) Line 61: More explanation for what a double-half limb means is needed.
We thank the reviewer for this suggestion. We have revised the manuscript (Line 73‒76). Specifically, we now explain that a double-dorsal limb, for example, is a chimeric limb generated by excising the ventral half and replacing it with a dorsal half from the contralateral limb while preserving the anteroposterior orientation.
(2) Line 63-65: "Such blastemas form hypomorphic, spike-like structures or fail to regenerate entirely." This statement does not represent the breadth of work on the APDV axis in limb regeneration. The cited Bryant 1976 reference tested only double-posterior and double-anterior newt limbs, demonstrating the importance of disposition along the AP axis, not DV. Others have shown that the regeneration of double-half limbs depends upon the age of the animal and the length of time between the grafting of double-half limbs and amputation. Also, some double-dorsal or double-ventral limbs will regenerate complete AP axes with symmetrical DV duplications (Burton, Holder, and Jesani, 1986). Also, sometimes half dorsal stylopods regenerate half dorsal and half ventral, or regenerate only half ventral, suggesting there are no inductive cues across the DV axis as there are along the AP axis. Considering this is the basis of the study under question, more is needed to convince that the DV axis is necessary for the generation of the AP axis.
We thank the reviewer for this detailed and constructive comment. We acknowledge that previous studies have reported a range of outcomes for double-half limbs. For example, Burton et al. (1986) described regeneration defects in double-dorsal (DD) and double-ventral (VV) limbs, although limb patterning did occur in some cases (Burton et al., 1986, Table 1). As the reviewer notes, regenerative outcomes depend on variables such as animal age and the interval between construction of the double-half limb and amputation, sometimes called the effect of healing time (Tank and Holder, 1978). Moreover, variability has been reported not only in DD/VV limbs but also in double-anterior (AA) and double-posterior (PP) limbs (e.g., Bryant, 1976; Bryant and Baca, 1978; Burton et al., 1986). In the revised manuscript, we have therefore modified the statement to avoid over-generalization and to emphasize that regeneration can be incomplete under these conditions (Line 76‒82). Importantly, in order to provide the additional evidence requested and to directly re-evaluate whether dorsal and ventral cells are required for limb patterning, we performed the ALM experiments shown in Fig. 1. The ALM system allows us to assess this question in a binary manner (regeneration vs. non-regeneration), thereby strengthening the rationale for our conclusions regarding the necessity of the APDV orientations. We also revised a sentence at the beginning of the Results section to emphasize this point (Line 139‒140).
(3) Line 71: These findings suggest that specific signals from all four positional domains must be integrated for successful limb patterning, such that the absence of any one of them leads to failure." I was under the impression that half posterior limbs can grow all elements, but half anterior can only grow anterior elements.
We thank the reviewer for this helpful clarification. As summarized by Stocum, half-limb experiments show that while some digit formation can occur, limb patterning remains incomplete in both anterior-half and posterior-half limbs in some cases (Stocum, 2017). We see this point as closely related to the broader question of whether proper limb patterning requires the integration of signals from all four positional domains. As noted in our response above, our ALM experiments in Fig. 1 were designed to test this point directly, and our data support the interpretation that cells from all four orientations are necessary for correct limb patterning.
(4) Line 79-81: This is stated later in lines 98-105. I suggest expanding here or removing it here.
We thank the reviewer for this suggestion. In the original version, lines 79–81 introduced our use of the terms “positional cue” and “positional information,” and this content partially overlapped with what later appeared in lines 98–105. In the revised manuscript, we have substantially rewritten this section (Line 82‒84), including the sentences corresponding to lines 79–81 in the original version, to remove the term “positional cue,” as explained in our response to the Editor’s comment (4); our revision reflects new analyses indicating that Wnt10b and Fgf2 appear not be strictly restricted to dorsal or ventral cell populations, and we now describe these factors as dorsal- or ventral-mediated signals that act across dorsoventral domains to induce Shh expression. Accordingly, we no longer maintain the original use of “positional cue” and “positional information.”
(5) Line 92 - 93: "Similarly, an ALM blastema can be induced in a position-specific manner along the limb axes. In this case, the induced ALM blastema will lack cells from the opposite side." This sentence is difficult to follow. Isn't it the same thing stated in lines 88-90?
We thank the reviewer for this comment. We revised the sentence to improve readability and to avoid redundancy with original Lines 88–90 (Line 104‒106).
(6) Line 107: I think the appropriate reference is McCusker et al., 2014 (Position-specific induction of ectopic limbs in non-regenerating blastemas on axolotl forelimbs), although Vieira et al., 2019 can be included here. In addition, Ludolph et al 1990 should be cited.
We thank the reviewer for this suggestion. We have added McCusker et al. (2014) and Ludolph et al. (1990) as references in the revised manuscript (Line 120‒121).
(7) Line 107-109: A missing point is how the ventral information is established in the amniote limb. From what I remember, it is the expression of Engrailed 1, which inhibits the ventral expression of Wnt7a, and hence Lmx1b. This would suggest that there is no secreted ventral cue. This is a relatively large omission in the manuscript.
We thank the reviewer for this comment. We agree that ventral fate in amniotes is specified by En1 in the ventral ectoderm, which represses Wnt7a and thereby prevents induction of Lmx1b; accordingly, a secreted ventral morphogen analogous to dorsal Wnt7a has not been established. We added this point to the revised Introduction (Line 61‒64).
By contrast, in axolotl limb regeneration, our previous work on Lmx1b expression suggests that DV identities reflect the original positional identity rather than being re-specified during regeneration (Yamamoto et al., 2022). Within this framework, our original use of the term “ventral positional cue” does not imply a ventral patterning morphogen in the amniote sense; rather, it denotes downstream signals induced by cells bearing ventral identity that are required for the blastema to form a patterned limb. This interpretation is consistent with classic studies on double-half chimeras and ectopic contacts between opposite regions (Iten & Bryant, 1975; Bryant & Iten, 1976; Maden, 1980; Stocum, 1982) as well as with our ALM data (Fig. 1). For this reason, we intentionally used the term “positional cues” to refer to signals provided by cells bearing ventral identity, which can be considered separable from the DV patterning mechanism itself, in the original text. As explained in our response to the Editor’s comment (4), we describe these signals as “signals mediated by dorsal/ventral cells,” rather than “positional cues” in the revised manuscript.
The necessity of dorsal- and ventral-mediated signals is supported by classic studies on the double-half experiment. In the non-regenerating cases, structural patterns along the anteroposterior axis appear to be lost even though both anterior and posterior cells should, in principle, be present in a blastema induced from a double-dorsal or double-ventral limbs. In limb development of amniotes, Wnt7a/Lmx1b or En-1 mutants show that limbs can exhibit anteroposterior patterning even when tissues are dorsalized or ventralized—that is, in the relative absence of ventral or dorsal cells, respectively (Riddle et al., 1995; Chen et al., 1998; Loomis et al., 1996). Taken together, axolotl limb regeneration, in which the presence of both dorsal and ventral cells plays a role in anteroposterior patterning, should differ from other model organisms. It is reasonable to predict the dorsal- and ventral-mediated signals in axolotl limb regeneration. We included this point in the revised manuscript (Line 82‒89). However, there is no evidence that these signals are secreted molecules. For this reason, we have carefully used the term “dorsal-/ventral-mediated signals” in the Introduction without implying secretion.
(8) Introduction - In general, the argument is a bit misleading. It is written as if it is known that a ventral cue is necessary, but the evidence from other animal models is lacking, from what I know. I may be wrong, but further argument would strengthen the reasoning for the study.
We thank the reviewer for this thoughtful comment. We agree that it should not read as if it is known that a ventral cue is necessary. In the revised Introduction, we have addressed this in several ways. First, as described in our response to comment (7), we now explicitly note that in amniote limb development ventral identity is specified by En1-mediated repression of Wnt7a, and that a secreted ventral morphogen equivalent to dorsal Wnt7a has not been established. Second, we removed the term “positional cue” and no longer present “ventral positional cue” as a defined entity. Instead, we use mechanistic phrasing such as “signals mediated by ventral cells” and “signals mediated by dorsal cells,” which does not assume that such signals are secreted morphogens or universally conserved. Third, we have reframed the role of dorsal- and ventral-mediated signals as a working hypothesis specific to axolotl limb regeneration, rather than as a general conclusion across model systems.
(9) Line 129: Remove "As mentioned before".
We thank the reviewer for this suggestion. We have removed the phrase “As mentioned before” in the revised manuscript (Line 143).
(10) Figure 1: Are Lmx1, Fgf8, and Shh mutually exclusive? Multiplexed FISH would provide this information, and is a relatively important question considering the strong claims in the study.
We thank the reviewer for raising this important point. As noted in our response to the editor’s comment, we cannot currently ensure sufficiently high detection sensitivity with multiplex FISH in our laboratory. However, based on previous reports (Nacu et al., 2016), Fgf8 and Shh should be mutually exclusive. In contrast, with respect to Lmx1b, our analysis suggests that its expression is not mutually exclusive with either Fgf8 or Shh, at least their expression domains. To confirm this, we analyzed the published scRNA-seq data and the results were added to the supplemental figure 6. Fgf8 and Shh were expressed in both Lmx1b-positive and Lmx1b-negative cells (Fig. S6H, I), but Fgf8 and Shh themselves were mutually exclusive (Fig. S6M). This point is now included in the revised manuscript (Line 314‒317).
(11) Results section and Figure 2: More evidence is needed for the lack of Shh expression ISH in tissue sections. Demonstrating the absence of something needs some qPCR or other validation to make such a claim.
We thank the reviewer for this suggestion. We performed qRT-PCR on ALM blastemas to complement the ISH data (Fig. 2).
(12) Line 179: I think they are likely leucistic d/d animals and not wild-type animals based upon the images.
We thank the reviewer for this observation. In the revised manuscript, we have corrected the description to “leucistic animals” (Line 194).
(13) Line 183-186: I'm a bit confused about this interpretation. If Shh turns on in just a posterior blastema, wouldn't it turn on in a grafted posterior tissue into a dorsal or ventral region? Isn't this independent of environment, meaning Shh turns on if the cells are posterior, regardless of environment?
Our interpretation is that only posterior-derived cells possess the competency to express Shh. In other words, whether a cell is capable of expressing Shh depends on its original positional identity (Iwata et al., 2020), but whether it actually expresses Shh depends on the environment in which the cell is placed. The results of Fig. 3E and G indicate that Shh activation is dependent on environment and that the posterior identity is not sufficient to activate Shh expression. We have revised the manuscript to emphasize this distinction more clearly (Line 198‒203).
(14) Figure 4: Do the limbs have an elbow, or is it just a hand?
We thank the reviewer for this thoughtful question. From the appearance, an elbow-like structure can occasionally be seen; however, we did not examine the skeletal pattern in detail because all regenerated limbs used for this analysis were sectioned for the purpose of symmetry evaluation, and we therefore cannot state this conclusively. While this is indeed an important point, analyzing proximodistal patterning would require a very large number of additional experiments, which falls outside the main focus of the present study. For this reason, and also to minimize animal use in accordance with ethical considerations, we did not pursue further experiments here. In response to this point, we have added a description of the skeletal morphology of ectopic limbs induced by BMP2+FGF2+FGF8 bead implantation (Fig. 6). In these experiments, multiple ectopic limbs were induced along the same host limb. In most cases, these ectopic limbs did not show fusion with the proximal host skeleton, similar to standard ALM-induced limbs, although in one case we observed fusion at the stylopod level. We now note this observation in the revised manuscript (Line 347‒354).
We regard the relationship between APDV positional information and proximodistal patterning as an important subject for future investigation.
(15) Line 203 - 237: I appreciate the symmetry score to estimate the DV axis. Are there landmarks that would better suggest a double-dorsal or double-ventral phenotype, like was done in the original double-half limb papers?
We thank the reviewer for this thoughtful comment. In most cases, the limbs induced by the ALM exhibit abnormal and highly variable morphologies compared to normal limbs, making it difficult to apply consistent morphological landmarks as used in the original double-half limb studies. For this reason, we focused our analysis on “morphological symmetry” as a quantitative measure of DV axis patterning, and we have added this explanation to the manuscript (Line 232‒235). Additionally, we provided transverse sections along the proximodistal axis as supplemental figures (Figs. S2 and S4). In addition to reporting the symmetry score, we have explicitly stated in the text that symmetry was also assessed by visual inspection of these sections.
(16) Line 245-247: The experiment was done using bulk sequencing, so both the epithelium and mesenchyme were included in the sample. The posterior (Shh) and anterior (Fgf8) patterning cues are mesenchymally expressed. In amniotes, the dorsal cue has been thought to be Wnt7a from the epithelium. Can ISH, FISH, or previous scRNAseq data be used to identify genes expressed in the mesenchyme versus epithelium? This is very important if the authors want to make the claim for defining "The molecular basis of the dorsal and ventral positional cues" as was stated by the authors.
We thank the reviewer for highlighting this important point. As the reviewer notes, our bulk RNA-seq data do not distinguish between epithelial and mesenchymal expression domains. As noted in our response to the editor’s comment, we performed ISH and qPCR on regular blastemas. However, these approaches did not provide definitive information regarding the specific cell types expressing Wnt10b and Fgf2. To complement this, we re-analyzed publicly available single-cell RNA-seq data (from Li et al., 2021). As a results, Fgf2 was expressed mainly by the mesenchymal cells, and Wnt10b expression was observed in both mesenchymal and epithelial cells. These results are now included in the revised manuscript (Line 294‒321) and in supplemental figures (Fig. S6, S7).
(17) Was engrailed 1, lmx1b, or Wnt7a differentially expressed along the DV axis, suggesting similar signaling between? Are these expressed in mesenchyme? Previous work suggests Wnt7a is expressed throughout the mesenchyme, but publicly available scRNAseq suggests that it is expressed in the epithelium.
We thank the reviewer for this important comment. As noted, the reported expression patterns of DV-related genes are not consistent across studies, which likely reflects the technical difficulty of detecting these genes with high sensitivity. In our own experiments, expression of DV markers other than Lmx1b has been very weak or unclear by ISH. Whether these genes are expressed in the epithelium or mesenchyme also appears to vary depending on the detection method used. In our RNA-seq dataset, Wnt7a expression was detected at very low levels and showed no significant difference along the DV axis, while En1 expression was nearly absent. We have clarified these results in the revised manuscript (Line 437‒441). Our reanalysis of the published scRNA-seq likewise detected Wnt7a in only a very small fraction of cells. Accordingly, we consider it premature to reach a definitive conclusion—such as whether Wnt7a is broadly mesenchymal or restricted to epithelium—as suggested in prior reports. We also note that whether Wnt7a is epithelial or mesenchymal does not affect the conclusions or arguments of the present study. Although the roles of Wnt7a and En1 in axolotl DV patterning are certainly important, we feel that drawing a definitive conclusion on this issue lies beyond the scope of the present study, and we have therefore limited our description to a straightforward presentation of the data.
(18) Line 247-249: The sentence suggests that all the ligands were tried. This should be included in the supplemental data.
We thank the reviewer for this clarification. In fact, we tested only Wnt4, Wnt10b, Fgf2, Fgf7, and Tgfb2, and all of these results are presented in the figures. To avoid misunderstanding, we have revised the text to explicitly state that our analysis focused on these five genes (Line 272‒274).
(19) Line 249: An n =3 seems low and qPCR would be a more sensitive means of measuring gene induction compared to ISH. The ISH would confirm the qPCR results. Figure 5C is also not the most convincing image of Shh induction without support from a secondary method.
We have increased the sample size for these experiments (Line 277‒280). In addition, to complement the ISH results, we confirmed Shh induction by qPCR following electroporation of Wnt10b and Fgf2 (Fig. 5D, E). In addition, because Shh signal in the Wnt10b-electroporated VentBL images was particularly weak and difficult to discern, we replaced that panel with a representative example in which Shh signal is more clearly visible. These data are now included in the revised manuscript (Line 280‒282).
(20) Line 253: It is confusing why Wnt10b, but not Wnt4 would work? As far as I know, both are canonical Wnt ligands. Was Wnt7a identified as expressed in the RNAseq, but not dorsally localized? Would electroporation of Wnt7a do the same thing as Wnt10b and hence have the same dorsalizing patterning mechanisms as amniotes?
We thank the reviewer for raising this challenging but important question. Wnt10b was identified directly from our bulk RNA-seq analysis, as was Wnt4. The difference in the ability of Wnt10b and Wnt4 to induce Shh expression in VentBL may reflect differences in how these ligands activate downstream WNT signaling programs. WNT10B is a potent activator of the canonical WNT/β-catenin pathway (Bennett et al., 2005), although WNT10B has also been reported to trigger a β-catenin–independent pathway (Lin et al., 2021). By contrast, WNT4 can signal through both canonical and non-canonical (β-catenin–independent) pathways, and the balance between these outputs is known to depend on cellular context (Li et al., 2013; Li et al., 2019). Consistent with a requirement for canonical WNT signaling, we found that pharmacological activation of canonical WNT signaling with BIO (a GSK3 inhibitor) was also sufficient to induce Shh expression in VentBL. However, despite this, it is still unclear why Wnt10b, but not Wnt4, was able to induce Shh under our experimental conditions. One possible explanation is that different WNT ligands can engage the same receptors (e.g., Frizzled/LRP6) yet can drive distinct downstream transcriptional programs (This may depend on the state of the responding cells, as Voss et al. predicted), resulting in ligand-specific outputs (Voss et al., 2025). This point is now included in the revised discussion section (Line 402‒412). At present, we cannot distinguish between these possibilities experimentally, and we therefore refrain from making a stronger mechanistic claim.
With respect to Wnt7a, we detected Wnt7a expression at very low levels, and without a clear dorsoventral bias, in our RNA-seq analysis of ALM blastemas (we describe this point in Line 437‒440). This is consistent with previous work suggesting that axolotl Wnt7a is not restricted to the dorsal region in regeneration. Because of this low and unbiased expression, and because our data already implicated Wnt10b as a dorsal-mediated signal that can act across dorsoventral domains to permit Shh induction, we did not prioritize Wnt7a electroporation in the present study. We therefore cannot conclude whether Wnt7a would behave similarly to Wnt10b in this context.
Importantly, these uncertainties about ligand-specific mechanisms do not alter our main conclusion. Our data support the idea that a dorsal-mediated WNT signal (represented here by WNT10B and canonical WNT activation) and a ventral-mediated FGF signal (FGF2) must act together to permit Shh induction, and that the coexistence of these dorsal- and ventral-mediated signals is required for patterned limb formation in axolotl limb regeneration.
(21) Is canonical Wnt signaling induced after electroporation of Wnt10b or Wnt4? qPCR of Lef1 and axin is the most common way of showing this.
We thank the reviewer for this helpful suggestion. In addition to examining Shh expression, we also assessed canonical WNT signaling by qPCR analysis of Axin2 and Lef1 following Wnt10b electroporation. The data is now included in Fig. 5.
(22) Line 255-256: qPCR was presented for Figure 5D, but ISH was used for everything else. Is there a technical reason that just qPCR was used for the bead experiments?
We thank the reviewer for this helpful comment. In the original submission, our goal was to test whether treatment with commercial FGF2 protein or BIO could reproduce the results obtained by electroporation. In the revised manuscript, to avoid confusion between distinct experimental aims, we removed the FGF2–bead data from this section and instead used RT-qPCR to quantitatively corroborate Shh induction after electroporation (Fig. 5D–E). RT-qPCR provided a sensitive, whole-blastema readout and allowed a paired design (left limb: factor; right limb: GFP control) that increased statistical power while minimizing animal use. To address the reviewer’s point more directly, we additionally performed ISH for the BIO treatment and now include those results in Supplementary Figure 3 (Line 287‒288).
(23) Line 261-263: The authors did not show where Wnt10B or Fgf2 is expressed in the limb as claimed. The RNAseq was bulk, so ISH of these genes is needed to make this claim. Where are Wnt10b and Fgf2 expressed in the amputated limb? Do they show a dorsal (Wnt10b) and ventral (Fgf2) expression pattern?
We thank the reviewer for raising this important point. As noted in our response to the editor’s comment, we performed ISH on serial sections of regular blastemas at several time points (Fig. S5A). However, the expression patterns of Wnt10b and Fgf2 along the dorsoventral axis were not clear. To complement the ISH results, we performed RT-qPCR on microdissected dorsal and ventral halves of regular blastemas at the MB stage (Fig. S5B). We found that Wnt10b and Fgf2 were expressed at significantly higher levels in the dorsal and ventral halves, respectively, compared to the opposite half. This dorsal/ventral biased expression of Wnt10b/Fgf2 is consistent with our RNA-seq data. To identify the cell types expressing Wnt10b or Fgf2, we analyzed published single-cell RNA-seq data (7 dpa blastema (MB), Li et al., 2021). As a result, Fgf2 expression was observed in the mesenchymal cluster, whereas Wnt10b expression was observed in both mesenchymal and epithelial clusters (Fig. S6). However, because only a small fraction of cells expressed Wnt10b, the principal cellular source of WNT10B protein remains unclear. The apparent low abundance likely contributes to the weak ISH signals and reflects current technical limitations. In addition, Wnt10b and Fgf2 expression did not follow Lmx1b expression (Fig. S6J, K), and Wnt10b and Fgf2 themselves were not exclusive (Fig. S6L). Together with the RT-qPCR data (Fig. S5B), these results suggest that Wnt10b and Fgf2 are not exclusively confined to purely dorsal or ventral cells at the single-cell level, even though they show dorsoventral bias when assessed in bulk tissue, suggesting that Wnt10b/Fgf2 expression is not dorsal-/ventral-specific but mediated by dorsal/ventral cells. Defining the precise spatial patterns of Wnt10b and Fgf2 in regular regeneration will therefore be an important goal for future work. These points are now included in the revised manuscript (Line 485‒501).
(24) Line 266-288: The formation of multiple limbs is impressive. Do these new limbs correspond to the PD location they are generated?
We thank the reviewer for this interesting question. Interestingly, from our observations, there does appear to be a tendency for the induced limbs to vary in length depending on their PD location. The skeletal patterns of the induced multiple limbs are now included in Fig. 6. However, as noted earlier, the supernumerary limbs exhibit highly variable morphologies, and a rigorous analysis of PD correlation would require a large number of induced limbs. Since this lies outside the main focus of the present study, we have not pursued this point further in the manuscript.
(25) Line 288: The minimal requirement for claiming the molecular basis for DV signaling was identified is to ISH or multiplexed FISH for Wnt10b and Fgf2 in amputated limb blastemas to show they are expressed in the mesenchyme or epithelium and are dorsally and ventrally expressed, respectively. In addition, the current understanding of DV patterning through Wnt7a, Lmx1b, and En1 shown not to be important in this model.
We thank the reviewer for this comment and fully agree with the point raised. We would like to clarify that we are not claiming to have identified the molecular basis of DV patterning. As the reviewer notes, molecules such as Lmx1b, Wnt7a, and En1 are well identified in other animal models as key regulators of DV positional identity. There is no doubt that these molecules play central roles in DV patterning. However, in axolotl limb regeneration, clear DV-specific expression has not been demonstrated for these genes except for Lmx1b. Therefore, further studies will be required to elucidate the molecular basis of DV patterning in axolotls.
Our focus here is more limited: we aim to identify the molecular basis for the mechanisms in which positional domain-mediated signals (FGF8, SHH, WNT10B, and FGF2) regulate the limb patterning process, rather than the molecular basis of DV patterning. In fact, our results on Wnt10b and Fgf2 suggest that these genes did not affect dorsoventral identities.
We recognize that this distinction was not sufficiently clear in the original text, and we have revised the manuscript to describe DV patterning mechanisms in other animals and clarify that the dorsal- and ventral-mediated signals are distinct from DV patterning (Line 444‒450). At least, we avoid claiming that the molecular basis for DV signaling was identified.
(26) Line 335: References are needed for this statement. From what I found, Wnt4 can be canonical or non-canonical.
We thank the reviewer for this helpful comment. We have revised the manuscript (Line 404‒407). We added these citations at the relevant location and adjusted nearby wording to avoid implying pathway exclusivity, in alignment with our response to comment (20).
(27) Line 337-338: The authors cannot claim "that canonical, but not non-canonical, WNT signaling contributes to Shh induction" as this was not thoroughly tested is based upon the negative result that Wnt4 electroporation did not induce Shh expression.
We thank the reviewer for this important clarification. We agree that our data do not allow us to conclude that non-canonical WNT signaling in general does not contribute to Shh induction. Accordingly, we have removed the phrase “but not non-canonical” and revised the text to emphasize that, within the scope of our experiments, Shh induction was not observed following Wnt4 electroporation, whereas it was observed with Wnt10b.
(28) Line 345: In order to claim "WNT10B via the canonical WNT pathway...appears to regulate Shh expression" needs at least qPCR to show WNT10B induces canonical signaling.
We thank the reviewer for this comment. As noted in our response to comment (21), we also assessed canonical WNT signaling by qPCR analysis of Axin2 and Lef1 following Wnt10b electroporation (Line 282‒285).
(29) Lines 361-372: A few studies have been performed on DV patterning of the mouse digit regeneration in regards to Lmx1b and En1. It may be good to discuss how the current study aligns with these findings.
We appreciate the reviewer’s suggestion. As the reviewer refers, several studies have been performed on dorsoventral (DV) patterning in mouse digit tip regeneration in relation to Lmx1b and En1 (e.g., Johnson et al., 2022; Castilla-Ibeas et al., 2023). In the present study, however, our main conclusion is different in the scope of studies on mouse digit tip regeneration. We show that, in the axolotl, pre-existing dorsal and ventral identities (as reflected by dorsally derived and ventrally derived cells in the ALM blastema) are required together to induce Shh expression, and that this Shh induction in turn supports anteroposterior interaction at the limb level. This mechanism—dorsal-mediated and ventral-mediated signals acting in combination to permit Shh expression—does not have a clear direct counterpart in the mouse digit tip literature. Moreover, even with respect to Lmx1b, the two systems behave differently. In mouse digit tip regeneration, loss of Lmx1b during regeneration does not grossly affect DV morphology of the regenerate (Johnson et al., 2022). By contrast, in our axolotl ALM system, the presence or absence of Lmx1b-positive dorsal tissue correlates with the final dorsoventral organization of the induced limb-like structures (e.g., production of double-dorsal or double-ventral symmetric structures in the absence of appropriate dorsoventral contact). Thus, the role of dorsoventral identity in our model is directly tied to patterned limb outgrowth at the whole-limb scale, whereas in the mouse digit tip it has been reported primarily in the context of digit tip regrowth and bone regeneration competence, not robust DV repatterning (Johnson et al., 2022).
For these reasons, we believe that an extended discussion of mouse digit tip regeneration would risk implying a mechanistic equivalence between axolotl limb regeneration and mouse digit tip regeneration that is not supported by current data. Because the regenerative contexts differ, and because Lmx1b does not appear to re-establish DV patterning in the mouse regenerates (Johnson et al., 2022), we have chosen not to include an explicit discussion of mouse digit tip regeneration in the main text.
(30) Line 408-433: Although I appreciate generating a model, this section takes some liberties to tell a narrative that is not entirely supported by previous literature or this study. For example, lines 415-416 state "Wnt10b and Fgf2 are expressed at higher levels in dorsal and the ventral blastemal cells, respectively" which were not shown in the study or other studies.
We thank the reviewer for this important comment. We agree that the original model based on RNA-seq data overstated the evidence. To address this point experimentally, we examined Wnt10b and Fgf2 expression in regular blastemas (Supplemental Figure 5 and 6). Accordingly, our model is now framed as an inductive mechanism for Shh expression—supported by results in ALM (WNT10B in VentBL; FGF2 in DorBL) and by DV-biased expression. Concretely, the sentence previously paraphrased as “Wnt10b and Fgf2 are expressed at higher levels in dorsal and ventral blastemal cells, respectively” has been replaced with wording that (i) avoids single-cell DV specificity and (ii) emphasizes dorsal-/ventral-mediated regulation and the requirement for both signals to allow Shh induction (Line 510‒511).
Reviewer #2 (Recommendations for the authors):
(1) Introduction:
The authors' definitions of positional cues vs positional information are a little hard to follow, and do not appear to be completely accurate. From my understanding of what the authors explain, "positional information" is defined as a signal that generates positional identities in the regenerating tissue. This is a somewhat different definition than what I previously understood, which is the intrinsic (likely epigenetic) cellular identity associated with specific positional coordinates. On the other hand, the authors define "positional cues" as signals that help organize the cells according to the different axes, but don't actually generate positional identities in the regenerating cells. The authors provide two examples: Wnt7a as an example of positional information, and FGF8 as a positional cue. I think that coording to the authors definitions, FGF8 (and probobly Shh) are bone fide positional cues, since both signals work together to organize the regenerating limb cells - yet do not generate positional identities, because ectopic limbs formed from blastemas where these pathways have been activated do not regenerate (Nacu et al 2016). However, I am not sure Wnt7a constitutes an example of a "positional information" signal, since as far as I know, it has not been shown to generate stable dorsal limb identities (that remain after the signal has stopped) - at least yet. If it has, the authors should cite the paper that showed this. I think that some sort of diagram to help define these visually will be really helpful, especially to people who do not study regenerative patterning.
We thank the reviewer for this thoughtful comment. We now agree with the reviewer that our use of “positional cue” and “positional information” may have been confusing. In the revision—and as noted in our response to the Editor’s comment (4)—we have removed the term “positional cue” and no longer attempt to contrast it with “positional information.” Instead, we adopt phrasing that reflects our data and hypothesis: during limb patterning, dorsal-mediated signals act on ventral cells and ventral-mediated signals act on dorsal cells to induce Shh expression. This wording avoids implying that these signals specify dorsoventral identity.
Regarding WNT7A, we agree it has not been shown to generate a stable dorsal identity after signal withdrawal. In the revised Introduction we therefore describe WNT7A in amniote limb development as an extracellular regulator that induces Lmx1b in dorsal mesenchyme (with En1 repressing Wnt7a ventrally), rather than labeling it as “positional information” in a strict, identity-imprinting sense. We highlight this contrast because, in our axolotl experiments, WNT10B and FGF2 did not alter Lmx1b expression or dorsal–ventral limb characteristics when overexpressed, consistent with the idea that they act downstream of DV identity to enable Shh induction, not to establish DV identity.
(2) Results:
It would be helpful if the number of replicates per sample group were reported in the figure legends.
We thank the reviewer for this suggestion. In accordance with the comment, we have added the number of replicates (n) for each sample group in the figure legends.
Figure 2 shows ISH for A/P and D/V transcripts in different-positioned blastemas without tissue grafts. The images show interesting patterns, including the lack of Shh expression in all blastemas except in posterior-located blastemas, and localization of the dorsal transcript (Lmx1b) to the dorsal half of A or P located blastemas. My only concern about this data is that the expression patterns are described in only a small part of the ectopic blastema (how representative is it?) and the diagrams infer that these expression patterns are reflective of the entire blastema, which can't be determined by the limited field of view. It is okay if the expression patterns are not present in the entire blastema -in fact, that might be an important observation in terms of who is generating (and might be receiving) these signals.
We thank the reviewer for this insightful comment. Because Fgf8 and Shh expression was detectable only in a limited subset of cells, the original submission included only high-magnification images. In response to the reviewer’s valid concern about representativeness, we have now added low-magnification overviews of the entire blastema as a supplemental figure (Fig. S1) and clarified in the figure legend that these expression patterns can be focal rather than pan-blastemal (Line 795‒796).
In Figure 3, they look at all of these expression patterns in the grafted blastemas, showing that Shh expression is only visible when both D and V cells are present in the blastema. My only concern about this data is that the number of replicates is very low (some groups having only an N=3), and it is unclear how many sections the authors visualized for each replicate. This is especially important for the sample groups where they report no Shh expression -I agree that it is not observable in the single example sections they provide, but it is uncertain what is happening in other regions of the blastema.
We thank the reviewer for this important comment. To increase the reliability of the results, we have increased the number of biological replicates in groups where n was previously low. For all samples, we collected serial sections spanning the entire blastema. For blastemas in which Shh expression was observed, we present representative sections showing the signal. For blastemas without detectable Shh expression, we selected a section from the central region that contains GFP-positive cells for the Figure. To make these points explicit, we have added the following clarification to the Fig. 3 legend (Line 811‒815).
Figure 4: Shh overexpression in A/P/D/V blastemas - expression induces ectopic limbs in A/D/V locations. They analyzed the symmetry of these regenerates (assuming that Do and V located blastemas will exhibit D/V symmetry because they only contain cells from one side of that axis. I am a little concerned about how the symmetry assay is performed, since oblique sections through the digits could look asymmetric, while they are actually symmetric. It is also unclear how the angle of the boxes that the symmetry scores were based on was decided - I imagine that the score would change depending on the angle. It also appears that the authors picked different digits to perform this analysis on the different sample groups. I also admit that the logic of classification scheme that the authors used AI to perform their symmetry scoring analysis (both in Figures 4 and 5) is elusive to me. I think it would have been more informative if the authors leveraged the structural landmarks, like the localization of specific muscle groups. (If this experiment were performed in WT animals, the authors could have used pigment cell localization)... or generate more proximal sections to look at landmarks in the zeugopod.
We thank the reviewer for these detailed comments regarding the symmetry analysis. Because reliance on a computed symmetry score alone could raise the concerns noted by the reviewer, we now provide transverse sections along the proximodistal axis as supplemental figures (Figs. S2 and S4). These include levels corresponding to the distal end of the zeugopod and the proximal end of the autopod. In addition to reporting the symmetry score, we have explicitly stated in the text that symmetry was also assessed by visual inspection of these sections.
As also noted in our response to Reviewer #1 (comment 15), ALM-induced limbs frequently exhibit abnormal and highly variable morphologies, which makes it difficult to use consistent anatomical landmarks such as particular digits or muscle groups. For this reason, we focused our analysis on morphological symmetry rather than landmark-based metrics, and we emphasize this rationale in the revised text (Line 232‒235).
Regarding the use of bounding boxes, this procedure was chosen to minimize the effects of curvature or fixation-induced distortion. For each section, the box angle was adjusted so that the outer contour (epidermal surface) was aligned symmetrically; this procedure was applied uniformly across all conditions to avoid bias. We analyzed multiple biological replicates in each group, which helps mitigate potential artifacts due to oblique sectioning. To further reduce bias, we increased the number of fields included in the analysis to n = 24 per group in the revised version.
In addition, staining intensity varied among samples, such that a region identified as “muscle” in one sample could be assigned differently in another if classification were based solely on color. To avoid this problem, we used a machine-learning classifier trained separately for each sample, allowing us to group the same tissues consistently within that sample irrespective of intensity differences. In the context of ALM-induced limbs, where stable anatomical landmarks are not available, we consider this strategy the most appropriate. We have added this rationale to the revised manuscript for clarity (Line 239‒247).
Figure 5: The number of replicates in sample groups is relatively low and is quite variable between groups (ranging between 3 and 7 replicates). Zoom in to visualize Shh expression is small relative to the blastema, and it is difficult to discern why the authors positioned the window where they did, and how they maintained consistency among their different sample groups. In the examples of positive Shh expression - the signal is low and hard to see. Validating these expression patterns using some sort of quantitative transcriptional assay (like qRTPCR) would increase the rigor of this experiment ... especially given that they will be able to analyze gene expression in the entire blastema as opposed to sections that might not capture localized expression.
We thank the reviewer for this important comment. To increase the rigor of these experiments, we have increased the number of biological replicates in groups where n was previously low. In addition, because Shh signal in the Wnt10b-electroporated VentBL images was particularly weak and difficult to discern, we replaced that panel with a representative example in which Shh signal is more clearly visible. We also validated the Shh expression for Wnt10b–electroporated VentBL and Fgf2–electroporated DorBL by RT-qPCR, which assesses gene expression across the entire blastema. These results are now included in Fig. 5 and Line 280‒282. Finally, we clarified in the figure legend how the “window” for imaging was chosen: for samples with detectable Shh expression, the window was placed in the region where the signal was observed; for conditions without detectable Shh expression, the window was positioned in a comparable region containing GFP-positive cells (Line 836‒839). These revisions are included in the revised manuscript.
Figure 6: They treat dorsal and ventral wounds with gelatin beads soaked in a combination of BMP2+FGF8 (nerve factors) and FGF2 proposed ventral factor). Remarkably, they observe ectopic limb expression in only dorsal wounds, further supporting the idea that FGF2 provides the "ventral" signal. They show examples of this impressive phenotype on limbs with multiple ectopic structures that formed along the Pr/Di axis. Including images of tubulin staining (as they have in Figures 1 and 2) to ensure that the blastemas (or final regenerates) are devoid of nerves. The authors' whole-mount skeletal staining which shows fusion of the ectopic humerus with the host humerus, is a phenotype associated with deep wounding, which could provide an opportunity for more cellular contribution from different limb axes.
We thank the reviewer for these constructive comments. As noted in the prior study, when beads are used to induce blastemas without surgical nerve orientation, fine nerve ingrowth can still occur (Makanae et al., 2014), and the induced blastemas are not completely devoid of nerves. While it is still uncertain whether these recruited nerves are functional after blastema induction, it is an important point, and we added sentences about this in the revised manuscript (Line 341‒345).
Regarding the skeletal phenotype, despite careful implantation to avoid injuring deep tissues, bead-induced ectopic limbs on the dorsal side occasionally displayed fusion of the stylopod with the host humerus—a phenotype associated with deep wounding, as the reviewer notes. This observation suggests that contributions from a broader cellular population cannot be excluded. However, because fusion was observed in only 1 of 16 induced limbs analyzed, and because ectopic limbs induced at the forearm (zeugopod) level did not exhibit such fusion (n=1/6 for stylopod-level inductions; n=0/10 for zeugopod-level inductions), we believe that our main conclusion remains valid. Because fusion is not a typical outcome, we now present representative non-fusion cases—including zeugopod-origin examples—in the figure (Fig. 6L1, L2), and we report the fusion incidence explicitly in the text (Line 350‒354). We also note in the revised manuscript that stylopod fusion can occur in a minority of cases (Line 347‒349).
Figure 7 nicely summarizes their findings and model for patterning.
We thank the reviewer for this positive comment.
The table is cut off in the PDF, so it cannot be evaluated at this time.
In our copy of the PDF, the table appears in full, so this may have been a formatting issue. We have carefully checked the file and ensured that the table is completely included in the revised submission.
There is a supplemental figure that doesn't seem to be referenced in the text.
The supplemental figure (Fig. S1 of the original manuscript) is referenced in the text, but it may have been overlooked. To improve clarity, we have expanded the description in the manuscript so that the supplemental figure is more clearly referenced (Line 285‒291).
(3) Materials and Methods:
No power analysis was performed to calculate sample group sizes. The authors have used these experimental techniques in the past and could have easily used past data to inform these calculations.
We thank the reviewer for this important comment. We did not include a power analysis in the manuscript because this was the first time we compared Shh and other gene expression levels among ALM blastemas of different positional origins using RT-qPCR in our experimental system. As we did not have prior knowledge of the expected variability under these specific conditions, it was difficult to predetermine appropriate sample sizes.
Reviewer #3 (Recommendations for the authors):
General:
Congratulations - I found this an elegant and easy-to-read study with significant implications for the field! If possible, I would urge you to consider adding some more characterisation of Wnt10b and Fgf2- which cell types are they expressed in? If you can link your mechanisms to normal limb regeneration too (i.e., regenerating blastema, not ALM), this would significantly elevate the interest in your study.
We sincerely thank the reviewer for these encouraging comments. As also noted in our response to the editor’s comment, we have analyzed the expression patterns of Wnt10b and Fgf2 in regular blastemas (Line 294‒306). Although clear specific expression patterns along dorsoventral axis were not detected by ISH, likely due to technical limitations of sensitivity, RT-qPCR revealed significantly higher expression levels of Wnt10b in the dorsal half and Fgf2 in the ventral half of a regular blastema (Fig. S5). In addition, we analyzed published single-cell RNA-seq data (7 dpa blastema, Li et al., 2021) (Line 307‒321). As a result, Fgf2 expression was observed in the mesenchymal clusters, whereasWnt10b expression was observed in both mesenchymal and epithelial clusters (Fig. S6). However, because only a small fraction of cells expressed Wnt10b, the principal cellular source of WNT10B protein remains unclear. Therefore, defining the precise spatial patterns of Wnt10b and Fgf2 in regular regeneration will be an important goal for future work.
Data availability:
I assume that the RNA-sequencing data will be deposited at a public repository.
RNA-seq FASTQ files have been deposited in the DNA Data Bank of Japan (DDBJ; https://www.ddbj.nig.ac.jp/) under BioProject accession PRJDB38065. We have added a Data availability section to the revised manuscript.
References
Castilla-Ibeas, A., Zdral, S., Oberg, K. C., & Ros, M. A. (2024). The limb dorsoventral axis: Lmx1b’s role in development, pathology, evolution, and regeneration. Developmental Dynamics, 253(9), 798–814. https://doi.org/10.1002/dvdy.695
Johnson, G. L., Glasser, M. B., Charles, J. F., Duryea, J., & Lehoczky, J. A. (2022). En1 and Lmx1b do not recapitulate embryonic dorsal-ventral limb patterning functions during mouse digit tip regeneration. Cell Reports, 41(8), 111701. https://doi.org/10.1016/j.celrep.2022.111701
Stocum, D. (2017). Mechanisms of urodele limb regeneration. Regeneration, 4. https://doi.org/10.1002/reg2.92
Tank, P. W., & Holder, N. (1978). The effect of healing time on the proximodistal organization of double-half forelimb regenerates in the axolotl, Ambystoma mexicanum. Developmental Biology, 66(1), 72–85. https://doi.org/10.1016/0012-1606(78)90274-9
-
-
milenio-nudos.github.io milenio-nudos.github.iopaper1
-
This study
faltan dos cosas esenciales de toda intro acá: 1) la relevancia, 2) EL ARGUMENTOOOO
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #3 (Public review):
Summary:
The manuscript explores the dependence of ABC transporter activity on membrane curvature. The underlying concept being analysed here is whether membrane mechanics can regulate the conformation of the protein and thereby its activity.
Strengths:
The protein of choice here is BmrA, a bacterial transmembrane ABC transporter. This protein was previously found to exhibit two states: open conformation with Nucleotide Binding domains (NBDs) separated from each other and an ATP-bound closed conformation with dimerised NBDs. The protein was purified and reconstituted into liposomes of varying diameters, largely categorised as Small vesicles (SV) and Large vesicles (LV). The authors find that the activity of the protein is reduced with the changing curvature of the membrane vesicles used to make the proteoliposomes. This could be modulated by making vesicles at different temperatures, LV at high and SV at lower temperature (4 {degree sign}C), following which they perform biochemical measurement of activity or smFRET experiments at HT or RT. They use well-characterized single-molecule FRET-based measurements to assess the change in conformation of the protein during the ATPase cycle. They find that a significant fraction of the protein is in an open (inactive) conformation in vesicles of higher curvature (SVs) at a given temperature. The authors develop a simple yet elegant theoretical model based on the energy of protein configuration states and their coupling to membrane energetics (bending rigidity) and curvature to explain these findings. The model provides a parameter-free fit that predicts the open/closed state distributions as well as the ATPase activity differences between SV and LV. Using experimentally determined values of the protein conicity, the authors to extract reasonable values of membrane rigidity, consistent with available literature.
The data and theoretical model together convincingly support the claim that membrane mechanics via local curvature modulation may bias membrane protein conformation states and thereby modify the activity of membrane proteins. This is an important and general conclusion that the authors also elaborate on in their discussion.
Weaknesses:
The authors say that the protein activity is irreversibly inhibited by orthovanadate, but 50% of the proteins are still in open conformation, while being accessible to the analogue (Table 2). It is unclear what this means in the context of activity vs. conformation.
The difference in the fraction of proteins in closed conformation is quite similar between LV and SV treated with AMP-PNP at 20 {degree sign}C (Figure 2B), and it is not clear if the difference is significant. The presence of a much higher FRET tail in the plots of smFRET experiment in SVs at 20 {degree sign}C or 33 {degree sign}C in the apo conformation of the protein (Figure 3A-B) is cause of some concern since one would not expect BmrA to access the closed states more frequently in the Apo conformation especially when incorporated in the SV. This is because the subtraction of the higher fraction of closed states in the Apo conformation contributes directly to enhancing the bias between the closed states in SV versus LV membrane bilayers.
-
Author response:
Global answer about the ATP analogs (concerns the 3 reviewers)
We use ATP-Vanadate essentially for detecting the FRET efficiency for the closed state. But these data are not included in our theoretical model. Thus, even if the comments of the reviewers on the observation of a non-negligible fraction of proteins in the open state in the presence of ATP-vanadate are justified, this has no consequence on our conclusions on the effect of curvature on BmrA on the conformational changes with ATP or AMP-PNP.
We agree with the comments of the reviewers that the binding of vanadate is not irreversible, but the reported lifetime of the closed state is very long compared to our experimental conditions (see (Urbatsch et al. JBC (1995)) on PgP).
Nevertheless, we will perform new experiments independent of ATP analogs using the E504A BmrA mutant. It has been shown structurally and enzymatically to bind and not hydrolyze ATP and to be 100% in a closed conformation at 5 mM ATP (A. Gobet et al., Nat. Commun. 16, 1745 (2025)). It will clear up all doubts about our experiments.
We will also add new references:
I. L. Urbatsch, B. Sankaran, J. Weber, A. E. Senior, J. Biol. Chem. 270, 19383 (1995)
T. Baukrowitz, T.-C. Hwang, A. C. Nairn, D. C. Gadsby, Neuron 12, 473 (1994)
A. Gobet et al., Nat. Commun. 16, 1745 (2025)
Y. Liu, M. Liao, Sci. Adv. 11, eadv9721 (2025) (on the effect of vanadate and temperature on a plant ABC)
Public Reviews:
Reviewer #1 (Public review):
(1) An important aspect of this paper is the difference in mechanism between inhibitors AMP-PNP (a substrate analog) and vanadate (together with ADP, forms a transition state analog inhibitor). The mechanisms and inhibitory constants/binding affinities of these inhibitors are not very well-supported in the current form of the manuscript, either through citations or through experiments. Related to this, the interpretation of the different curvature response of BmrA in the presence of vanadate vs AMPPNP is not very clear.
See the global answer about ATP-analogs (above)
(2) Overall, the energetic contribution of the membrane curvature is subtle (less than a kT), so while the principles seem generalizable among membrane proteins, whether these principles impact transport or cell physiology remains to be established.
This is correct that the effect is limited to high curvature in the case of BmrA. Our theoretical model allows predictions for different protein parameters. The effect is particularly dependent on the protein size and on protein conicity, which can vary over a wide range. We show that larger proteins, such as piezo 1 are in principle expected to display a much stronger curvature dependence than BmrA. But testing our predictions on other proteins and on their physiological function is indeed an exciting perspective but beyond the objective of the current manuscript.
Reviewer #2 (Public review):
(1) Although this study may be considered as a purely biophysical investigation of the sensitivity of an ABC transporter to mechanical perturbation of the membrane, the impact would be strengthened if a physiological rationale for this mode of regulation were discussed. Many factors, including temperature, pH, ionic strength, or membrane potential, are likely to affect flux through the transport cycle to some extent, without justifying describing BmrA as a sensor for changes in any of these. Indeed, a much stronger dependence on temperature than on membrane curvature was measured. It is not clear what radii of curvature BmrA would normally be exposed to, and whether this range of curvatures corresponds to the range at which modulation of transport activity could occur. Similarly, it is not clear what biological condition would involve a substantial change to membrane curvature or tension that would necessitate altered BmrA activity.
Reviewers 1 and 2 both stressed that we showed that activity and conformational changes are mechanosensitive, not that the function of the protein is to be a mechanosensor. This will be corrected.
Regarding the physiological relevance of the mechanosensitivity of BmrA, we have addressed this point in the manuscript (bottom of page 10 and top of page 11). This discussion was positively appreciated by Reviewer #3. We stress that we have used BmrA as a model system, but considering our results and the theoretical model, we can predict the parameters that are relevant for future studies on the sensitivity of other transmembrane proteins to membrane mechanical properties. And, as stated by the reviewer, "mechanosensitivity of proteins is an understudied phenomenon".
(2) The size distributions of vesicles were estimated by cryoEM. However, grid blotting leaves a very thin layer of vitreous ice that could sterically exclude large vesicles, leading to a systematic underestimation of the vesicle size distribution.
We used Lacey carbon grids with large mesh size ranges for our cryoEM images, and we blot on the backside, precisely to measure the largest size range accessible to cryoEM. In our hands, this was not the case when using Quantifoil or C-Flat grids with uniform hole sizes and a large fraction of carbon where the vesicles adhere. With our grids, we are able to image vesicles from 20 to 200 nm diameter and the precision on the diameter is high, but the statistics might not be as good as with DLS or other diffusion-based methods. DLS is an indirect method (as compared to cryoEM) to measure vesicle size distribution, that may overestimate the fraction of large objects and underestimate the small ones. We will perform DLS experiments for comparison purpose.
(3) The relative difference in ATP turnover rates for BmrA in small versus large vesicles is modest (~2-fold) and could arise from different success rates of functional reconstitution with the different protocols.
The ATPase activity is sensitive to several parameters. We thus carefully characterized our reconstituted samples, including ATPase activity, yield of incorporation and orientation of proteins that are often reported. In addition, we showed by cryo-EM the unilamellarity of the proteoliposomes and their stability during the experiments, which were never reported. The ATPase activity of our samples reconstituted in liposomes at 20 ° and at 4°C are high, among the highest reported for BmrA, and less sensitive to errors as compared to the low activities in micelles of detergent.
We would also like to stress that with our protocol, we have prepared the same batch of lipid/protein mixture that we have split it 2 for the reconstitution at 4°C and 20°C conversely. Both preparations contain the same amount of detergent. The only difference is that we include more BioBeads for the preparation at 4°C to account for the difference of absorption of the detergent on the beads at low temperature (D. Lévy, A. Bluzat, M. Seigneuret, J.L. Rigaud Biochim. Biophys. Acta. 179 (1990)), but we also showed that the proteins do not adsorb on the BioBeads (J.-L. Rigaud, B. Pitard, D. Levy, Biochim. Biophys. Acta 1231, 223 (1995)). In addition, the activity of the protein at 37°C is high and comparable to those reported in the literature (E. Steinfels et al., Biochemistry 43, 7491 (2004)., W. Mi et al., Nature 549, 233 (2017).), which speaks for a good functional reconstitution. Finally, our results are consistent between the smFRET where we have only one protein maximum per vesicle and the activity measurements where the amount of protein is higher.
We also performed reconstitution from molar LPR= 1:13600 to 1:1700 and found the same activity per protein, confirming that the proteins are functional, independently of their surface fraction. We will add these data in the revision.
Altogether, these data suggest that we correctly estimate the rate of functional reconstitution in our experiments.
Nevertheless, we will design additional experiments to further compare the activity of the proteins before and after reconstitution.
(4) The conformational state of the NBDs of BmrA was measured by smFRET imaging. Several aspects of these investigations could be improved or clarified. Firstly, the inclusion and exclusion criteria for individual molecules should be more quantitatively described in the methods. Secondly, errors were estimated by bootstrapping. Given the small differences in state occupancies between conditions, true replicates and statistical tests would better establish confidence in their significance. Thirdly, it is concerning that very few convincing dynamic transitions between states were observed. This may in part be due to fast photobleaching compared to the rate of isomerization, but this could be overcome by reducing the imaging frequency and illumination power. Alternatively, several labs have established the ability to exchange solution during imaging to thereby monitor the change in FRET distribution as a ligand is delivered or removed. Visualizing dynamic and reversible responses to ligands would greatly bolster confidence in the condition-dependent changes in FRET distributions. Such pre-steady state experiments would also allow direct comparison of the kinetics of isomerization from the inward-facing to the outward-facing conformation on delivery of ATP between small and large vesicles.
(a) We will better detail the inclusion and exclusion criteria.
(b) For the smFRET, we have performed N=3 true replicates. We will add statistical tests on our graphs.
(c) We will detail more how we have optimized our illumination protocol, considering the signal to noise ratio and the photobleaching. Practically, we cannot add ATP to our sealed observation chamber on our TIRF system to detect dynamical changes on our immobilized liposomes. The experiment suggested by the reviewer would imply to build a flow chamber to exchange the medium around immobilized liposomes, compatible with TIRF microscopy. This is an excellent idea, which has been achieved only recently (S. N. Lefebvre, M. Nijland, I. Maslov, D. J. Slotboom, Nat. Commun. 16, 4448 (2025)). It will require a full new study to optimize both the flow chamber and the dyes to track the smFRET changes over long periods of time.
Nevertheless, we would like to stress that our objective is not to study the dynamics of the conformational changes, and that we expect it to be slow for BmrA, even at 33°C.
(5) A key observation is that BmrA was more prone to isomerize ATP- or AMP-PNP-dependently to the outward-facing conformations in large vesicles. Surprisingly, the same was not observed with vanadate-trapping, although the sensitivity of state occupancy to membrane curvature would be predicted to be greatest when state occupancies of both inward- and outward-facing states are close to 50%. It is argued that this was due to irreversibility of vanadate-trapping, but both vanadate and AMP-PNP should work fully reversibly on ABC transporters (see e.g. PMID: 7512348 for vanadate). Further, if trapping were fully irreversible, a quantitative shift to the outward-facing condition would be predicted.
See the global answer about ATP-analogs (above)
Reviewer #3 (Public review):
(1) The authors say that the protein activity is irreversibly inhibited by orthovanadate, but 50% of the proteins are still in open conformation, while being accessible to the analogue (Table 2). It is unclear what this means in the context of activity vs. conformation.
See the global answer about ATP-analogs (above)
(2) The difference in the fraction of proteins in closed conformation is quite similar between LV and SV treated with AMP-PNP at 20 {degree sign}C (Figure 2B), and it is not clear if the difference is significant. The presence of a much higher FRET tail in the plots of smFRET experiment in SVs at 20 {degree sign}C or 33 {degree sign}C in the apo conformation of the protein (Figure 3A-B) is cause of some concern since one would not expect BmrA to access the closed states more frequently in the Apo conformation especially when incorporated in the SV. This is because the subtraction of the higher fraction of closed states in the Apo conformation contributes directly to enhancing the bias between the closed states in SV versus LV membrane bilayers.
We have consistently observed, both at 20°C and at 33°C, a fraction of proteins with a high FRET signal in our measurements, higher in SV (about 15% and 17%) than in LV (about 10% and 6%). We have quantified the fraction of proteins with NBDs facing inside the liposomes (page 5), 20% in LV and 23.85% in SV. Considering the inverted curvature of the membrane, this orientation could favor the closed conformation, even in the absence of ATP, more for SV than LV. The fraction with inverted orientation could explain our higher fraction of high FRET signal in SV.
Moreover, for part of it, it can be due to a fraction of proteins with a non-specific labeling that would produce a higher FRET signal. We will add data with Cys-less mutants showing that less than 4% are labeled.
-
-
www.biorxiv.org www.biorxiv.org
-
Author response:
The following is the authors’ response to the previous reviews
Public Reviews:
Reviewer #3 (Public review):
To summarize: The authors' overfilling hypothesis depends crucially on the premise that the very quickly reverting paired-pulse depression seen after unusually short rest intervals of << 50 ms is caused by depletion of release sites whereas Dobrunz and Stevens (1997) concluded that the cause was some other mechanism that does not involve depletion on. The authors now include experiments where switching extracellular Ca2+ from 1.2 to 2.5 mM increases synaptic strength on average, but not by as much as at other synapse types. They contend that the result supports the depletion on hypothesis. I didn't agree because the model used to generate the hypothesis had no room for any increase at all, and because a more granular analysis revealed a mixed population with a subset where: (a) synaptic strength increased by as much as at standard synapses; and yet (b) the quickly reverting depression for the subset was the same as the overall population.
The authors raise the possibility of additional experiments, and I do think this could clarify things if they pre-treat with EGTA as I recommended initially. They've already shown they can do this routinely, and it would allow them to elegantly distinguish between pv and pocc explanations for both the increases in synaptic strength and the decreases in the paired pulse ratio upon switching Ca2+ to 2.5 mM. Plus/minus EGTA pre-treatment trials could be interleaved and done blind with minimal additional effort.
Showing reversibility would be a great addition too, because, in our experience, this does not always happen in whole-cell recordings in ex-vivo tissue even when electrical properties do not change. If the goal is to show that L2/3 synapses are less sensitive to changes in Ca2+ compared to other synapse types - which is interesting but a bit off point - then I would additionally include a positive control, done by the same person with the same equipment, at one of those other synapse types using the same kind of presynaptic stimulation (i.e. ChRs).
Specific points (quotations are from the Authors' rebuttal)
(1) Regarding the Author response image 1, I was instead suggesting a plot of PPR in 1.2 mM Ca2+ versus the relative increase in synaptic strength in 2.5 versus in 1.2 mM. This continues to seem relevant.
Complying with your suggestion, we studied the effects of external [Ca<sup>2+</sup>] ([Ca<sup>2+</sup>]<sub>o</sub>) after pre-incubating the slice in aCSF containing 50 μM EGTA-AM, and added the results as Figure 3—figure supplement 3C-D. Elevation of ([Ca<sup>2+</sup>]<sub>o</sub>) from 1.3 to 2.5 mM produced no significant change in either baseline EPSC amplitude or PPR, supporting that the p<sub>v</sub> is already saturated at 1.3 mM [Ca<sup>2+</sup>]<sub>o</sub> and implying that the modest Ca<sup>2+</sup> dependence of baseline EPSCs and PPR in the absence of EGTA (Figure 3—figure supplement 3A-B) is mediated by the change in baseline vesicular occupancy of release sites (p<sub>occ</sub>) rather than fusion probability of docked vesicles (p<sub>v</sub>).
We found some correlation of high Ca<sup>2+</sup>-induced relative increase in synaptic strength with the PPR at low Ca<sup>2+</sup> (Author response image 1-A). But this correlation was abolished by pre-incubating the slices in EGTA-AM too (Author response image 1-B). It should be noted that high PPR does not always mean low p<sub>v</sub>. For example, when the replenishment is equal between high and low baseline p<sub>occ</sub> synapses, the PPR would be higher at low p<sub>occ</sub> synapses than that at high p<sub>occ</sub> synapses, even if p<sub>v</sub> is close to unity. Therefore, high baseline release probability (Pr), whatever it is attributed to high p<sub>v</sub> or high p<sub>occ</sub>, can result in low PPR, considering that Pr = p<sub>occ</sub> x p<sub>v</sub>.
As we have already mentioned in our previous letter, the relationship of PPR with refilling rate is complicated and can be bidirectional, whereas an increase in p<sub>v</sub> always results in a reduction of PPR. For example, PPR can be reduced by both a decrease and an increase in the refilling rate (Figure 2— figure supplement 1 and Lin et al., 2025). Therefore, the PPR analysis alone is insufficient to differentiate the contributions of p<sub>v</sub> and p<sub>occ</sub> Thanks to your suggestion, we could resolve this ambiguity by the EGTA-AM pre-incubation study (Figure 3—figure supplement 3C-D).
Author response image 1.
Plot of PPR at low [Ca<sup>2+</sup>]<sub>o</sub> (1.3 mM) as a function of the baseline EPSC at high [Ca<sup>2+</sup>]<sub>o</sub> (2.5 mM) normalized to that at low [Ca<sup>2+</sup>]<sub>o</sub> measured at recurrent excitatory synapses in L2/3 of the prelimbic cortex under the conditions without EGTA-AM (A) and after pre-incubating the slices in EGTA-AM (50 μM) (B)
(2) "Could you explain in detail why two-fold increase implies pv < 0.2?"
(a) start with power((2.5/(1 + (2.5/K1) + 1/2.97)),4) = 2<sup>*</sup>power((1.3/(1 + (1.3/K1) + 1/2.97)),4);
(b) solve for K1 (this turns out to be 0.48);
(c) then implement the premise that pv -> 1.0 when Ca2+ is high by calculating Max = power((C/(1 + (C/K1) + 1/2.97)),4) where C is [Ca] -> infinity.
(d) pv when [Ca] = 1.3. mM must then be power((1.3/(1 + (1.3/K1) + 1/2.97)),4)/Max, which is <0.2. Note that modern updates of Dodge and Rahamimoff typically include a parameter that prevents pv from approaching 1.0; this is the gamma parameter in the versions from Neher group.
Thank you very much for your kind explanation. This interpretation, however, based on the premise that pv is not saturated at low[Ca<sup>2+</sup>]<sub>o</sub>, and that Pr = p<sub>v</sub>. In the present study, however, we presented multiple convergent lines of evidence supporting that p<sub>v</sub> is already saturated at 1.3 mM [Ca<sup>2+</sup>]<sub>o</sub> as follows: (1) little effect of EGTA-AM on the baseline EPSCs (Figure 2—figure supplement 1); (2) high double failure rates (Figure 3—figure supplement 2); (3) little effect of high [Ca<sup>2+</sup>]<sub>o</sub> on baseline EPSC (Figure 3—figure supplement 3). Therefore, our results suggest that the classical Dodge-Rahamimoff fourth-power relationship can not be applied to estimate p<sub>v</sub> at the L2/3 recurrent excitatory synapses.
(3) "If so, we can not understand why depletion-dependent PPD should lead to PPF." When PPD is caused by depletion and pv < 0.2, the number of occupied release sites should not be decreased by more than one-filth at the second stimulus so, without facilitation, PPR should be > 0.8. The EGTA results then indicate there should be strong facilitation, driving PPR to something like 1.2 with conservative assumptions. And yet, a value of < 0.4 is measured, which is a large miss.
As mentioned above, the framework used for inferring that p<sub>v</sub> < 0.2, the Dodge-Rahamimoff equation, is not applicable to our experimental system. Consequently, the subsequent deduction— that depletion-dependent PPD should logically lead to PPF—is based on a model that does not compatible with aforementioned multiple convergent lines of evidence, which supports high p<sub>v</sub> rather than the low p<sub>v</sub> facilitation model.
(4) Despite the authors' suggestion to the contrary, I continue to think there is a substantial chance that Ca2+-channel inactivation is the mechanism underlying the very quickly reverting paired-pulse depression. However, this is only one example of a non-depletion mechanism among many, with the main point being that any non-depletion mechanism would undercut the reasoning for overfilling. And, this is what Dobrunz and Stevens claimed to show; that the mechanism - whatever it is - does not involve depletion. The most effective way to address this would be affirmative experiments showing that the quickly reverting depression is caused by depletion after all. Attempting to prove that Ca2+channel inactivation does not occur does not seem like a worthwhile strategy because it would not address the many other possibilities.
We have systematically ruled out alternative possibilities that may underlie the strong PPD observed at our synapses and demonstrated that it arises from high p<sub>v</sub>-induced vesicle depletion through multiple independent lines of evidence. First, we excluded (1) AMPAR desensitization or saturation (Figure 1—figure supplement 5), (2) Ca<sup>2+</sup> channel inactivation (Figure 2—figure supplement 2), (3) channelrhodopsin inactivation (Figure 1—figure supplement 2), (4) artificial bouton stimulation (Figure 1—figure supplement 4), and (5) transient vesicle undocking (Figure 5; addressed in our previous rebuttal). Second, EGTA-AM experiments (Figure 2, Figure 2—figure supplement 1) revealed that release sites are tightly coupled to Ca<sup>2+</sup> channels, and that EGTA further exacerbates PPD. Third, we validated high baseline p<sub>v</sub> through analysis of double failure rates (Figure 3—figure supplement 2). Fourth, the minimal increase in baseline EPSCs upon elevation of external [Ca<sup>2+</sup>] (Figure 3—figure supplement 3) further supports that baseline p<sub>v</sub> is already saturated at low [Ca<sup>2+</sup>]<sub>o</sub>. Additionally, to further validate our hypothesis, we performed the specific experiment suggested by the reviewer. We have now added EGTA pre-incubation experiments (Figure 3—figure supplement 3C-D) and have revised the manuscript. Specifically, when slices were pre-incubated with 50 μM EGTA-AM, elevation of extracellular [Ca<sup>2+</sup>] from 1.3 to 2.5 mM produced no significant change in either baseline EPSC amplitude or PPR, strongly supporting that the high [Ca<sup>2+</sup>]<sub>o</sub> effects in the absence of EGTA are primarily mediated by changes in p<sub>occ</sub> rather than p<sub>v</sub>
(5) True that Kusick et al. observed morphological re-docking, but then vesicles would have to re-prime and Mahfooz et al. (2016) showed that re-priming would have to be slower than 110 ms (at least during heavy use at calyx of Held).
As previously discussed, Kusick et al. (2020) demonstrated that the transient destabilization of the docked vesicle pool recovers very rapidly within 14 ms after stimulation. This implies that any posts stimulation undocking events are likely recovered before the 20 ms ISI used in our PPR experiments. Consequently, transient undocking/re-docking events are unlikely to significantly influence the PPR measured at this interval. Furthermore, regarding the slow re-priming kinetics (>100 ms) reported by Mahfooz et al. (2016) and Kusick et al., (2020), our 20 ms ISI effectively falls into a me window that avoids the potential confounds of both processes: it is long enough for the rapid morphological recovery (~14 ms) of docked vesicles to occur, yet too short for the slow re-priming process to make a substantial contribution. Furthermore, Vevea et al. (2021) showed that post-stimulus undocking is facilitated in synaptotagmin-7 (Syt7) knockout synapses. In our study, however, Syt7 knockdown did not affect PPR at 20 ms ISI, suggesting that the undocking process described in Kusick et al. (2020) is not a major contributor to the PPD observed at 20 ms intervals in our experiments. Therefore, we conclude that the 20 ms ISI used in our experiments falls within a me window that is influenced neither by the rapid undocking (<14 ms) reported nor by the slow re-priming process (>100 ms).
-
-
shs-cairn-info.ezproxy.u-paris.fr shs-cairn-info.ezproxy.u-paris.fr
-
Par ailleurs, considérer que les expériences précoces sont au cœur de la souffrance humaine, et de toutes ses formes de détresse, suppose de bien penser l’infantile comme toujours actuel, et non pas comme seulement convoqué par des effets d’après-coup, ou comme une seule relique du passé.
=/ que penser que dans l'après coup
-
On peut même dire que c’est à ces aspects que s’adresse prioritairement le soin psychique, car ce sont les souffrances dans ces zones de la subjectivité qui, lorsqu’elles font retour, sont les plus douloureuses et harcelantes.
!!!!
-
-
www.biorxiv.org www.biorxiv.org
-
Author response:
The following is the authors’ response to the current reviews.
Reviewer #4 (Public review):
Summary:
The authors demonstrate a computational rational design approach for developing RNA aptamers with improved binding to the Receptor Binding Domain (RBD) of the SARS-CoV-2 spike protein. They demonstrate the ability of their approach to improve binding affinity using a previously identified RNA aptamer, RBD-PB6-Ta, which binds to the RBD. They also computationally estimate the binding energies of various RNA aptamers with the RBD and compare against RBD binding energies for a few neutralizing antibodies from the literature. Finally, experimental binding affinities are estimated by electrophoretic mobility shift assays (EMSA) for various RNA aptamers and a single commercially available neutralizing antibody to support the conclusions from computational studies on binding. The authors conclude that their computational framework, CAAMO, can provide reliable structure predictions and effectively support rational design of improved affinity for RNA aptamers towards target proteins. Additionally, they claim that their approach achieved design of high affinity RNA aptamer variants that bind to the RBD as well or better than a commercially available neutralizing antibody.
Strengths:
The thorough computational approaches employed in the study provide solid evidence of the value of their approach for computational design of high affinity RNA aptamers. The theoretical analysis using Free Energy Perturbation (FEP) to estimate relative binding energies supports the claimed improvement of affinity for RNA aptamers and provides valuable insight into the binding model for the tested RNA aptamers in comparison to previously studied neutralizing antibodies. The multimodal structure prediction in the early stages of the presented CAAMO framework, combined with the demonstrated outcome of improved affinity using the structural predictions as a starting point for rational design, provide moderate confidence in the structure predictions.
We thank the reviewer for this accurate summary and for recognizing the strength of our integrated computational–experimental workflow in improving aptamer affinity.
Weaknesses:
The experimental characterization of RBD affinities for the antibody and RNA aptamers in this study present serious concerns regarding the methods used and the data presented in the manuscript, which call into question the major conclusions regarding affinity towards the RBD for their aptamers compared to antibodies. The claim that structural predictions from CAAMO are reasonable is rational, but this claim would be significantly strengthened by experimental validation of the structure (i.e. by chemical footprinting or solving the RBD-aptamer complex structure).
The conclusions in this work are somewhat supported by the data, but there are significant issues with experimental methods that limit the strength of the study's conclusions.
(1) The EMSA experiments have a number of flaws that limit their interpretability. The uncropped electrophoresis images, which should include molecular size markers and/or positive and negative controls for bound and unbound complex components to support interpretation of mobility shifts, are not presented. In fact, a spliced image can be seen for Figure 4E, which limits interpretation without the full uncropped image.
Thank you for your valuable comments and careful review.
In response to your suggestion, we will provide all uncropped electrophoresis raw images corresponding to the results in the main figures and supplementary figures (Figure 2F, 3D, 3E, 4E, S9A and S10 of the original manuscript) in the revised version. Regarding the spliced image in Figure 4E, the uncropped raw gel image clearly shows that the two C23U samples were run on an adjacent lane of the same gel due to the total number of samples exceeding the well capacity of a single lane. All samples were electrophoresed and signal-detected under identical experimental conditions in one single experiment, ensuring the validity of direct signal intensity comparison across all samples. These complete uncropped raw images will be supplemented in the revised manuscript as Figure S12 (also see Author response image 1).
Author response image 1.
Uncropped electrophoresis images corresponding to Figures 2F, 3D, 3E, 4E, S9A and S10 of the original manuscript.
Additionally, he volumes of EMSA mixtures are not presented when a mass is stated (i.e. for the methods used to create Figure 3D), which leaves the reader without the critical parameter, molar concentration, and therefore leaves in question the claim that the tested antibody is high affinity under the tested conditions.
Thank you for your valuable comment on this oversight.
For the EMSA assay in Figure 3D, the reaction mixture (10 μL total volume) contained 3 μg of RBD protein and 3 μg of antibody (40592-R001), either individually or in combination, with incubation at room temperature for 20 minutes. Based on the molecular weights (35 kDa for RBD and 150 kDa for the IgG antibody), the corresponding molar concentrations in the mixture were calculated as 8.57 μM for RBD and 2 μM for the antibody. To ensure consistency, clarity and provide the critical molar concentration parameter, we will revise the legend of Figure 3D, replacing the mass values with the calculated molar concentrations as you suggested in the revised manuscript.
Additionally, protein should be visualized in all gels as a control to ensure that lack of shifts is not due to absence/aggregation/degradation of the RBD protein. In the case of Figure 3E, for example, it can be seen that there are degradation products included in the RBD-only lane, introducing a reasonable doubt that the lack of a shift in RNA tests (i.e. Figure 2F) is conclusively due to a lack of binding.
We sincerely appreciate your careful evaluation of our work, which helps us further clarify the experimental details and data reliability.
First, we would like to clarify the nature of the gel electrophoresis in Figure 3E: the RBD protein was separated by native-PAGE rather than denaturing SDS-PAGE. The RBD protein used in all experiments was purchased from HUABIO (Cat. No. HA210064) with guaranteed quality, and its integrity and purity were independently verified in our laboratory via denaturing SDS-PAGE (see Author response image 2), which showed a single, intact band without any degradation products. The ladder-like bands observed in the RBD-only lane of the native-PAGE gel are not a result of protein degradation. Instead, they arise from two well-characterized properties of recombinant SARS-CoV-2 Spike RBD protein expressed in human cells: intrinsic conformational heterogeneity (the RBD domain exists in multiple dynamic conformations due to its structural flexibility) (Cai et al., Science, 2020; Wrapp et al., Science, 2020) and heterogeneity in N-glycosylation modification (variable glycosylation patterns at the conserved N-glycosylation sites of RBD) (Casalino et al., ACS Cent. Sci., 2020; Ives et al., eLife, 2024), both of which could cause distinct migration bands in native-PAGE under non-denaturing conditions.
Second, to ensure the reliability of the RNA-binding results, the EMSA experiments for determining the binding affinity (K<sub>d</sub>) of RBD to Ta, Tc and Ta variants were performed with three independent biological replicates (the original manuscript includes all replicate data in Figure 2F and S9). Consistent results were obtained across all replicates, which effectively rules out false-negative outcomes caused by accidental absence or loss of functional RBD protein in the reaction system. In addition, our gel images (Figure 2F and S9 in the original manuscript) and uncropped raw images of all EMSA gels (see Author response image 1) show no significant signal accumulation in the sample wells, confirming the absence of RBD protein aggregation in the binding reactions—an issue that would otherwise interfere with RNA-protein interaction and band shift detection.
New results for RBD analysis by denaturing SDS-PAGE, along with the associated discussion, will be added to the revised manuscript as Figure S10 (also see Author response image 2).
Author response image 2.
SDS-PAGE analysis of the SARS-CoV-2 Spike RBD protein, neutralizing antibody (40592-R001) and BSA reference. This gel validates the high purity and structural integrity of the commercially sourced RBD protein and neutralizing antibody used in this study.
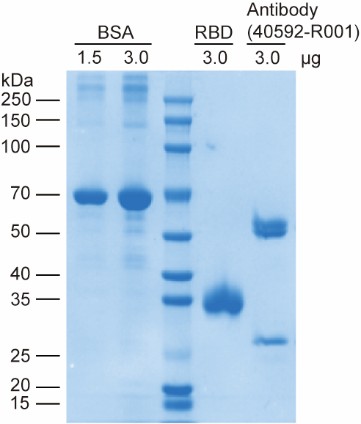
References
Cai, Y. et al. Distinct conformational states of SARS-CoV-2 spike proteins. Science 369, 1586-1592 (2020).
Casalino, L. et al. Beyond shielding: the roles of glycans in the SARS-CoV-2 spike protein. ACS Cent. Sci. 6, 1722-1734 (2020).
Ives, C.M. et al. Role of N343 glycosylation on the SARS-CoV-2 S RBD structure and co-receptor binding across variants of concern. eLife 13, RP95708 (2024).
Wrapp, D. et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 367, 1260-1263 (2020).
Finally, there is no control for nonspecific binding, such as BSA or another non-target protein, which fails to eliminate the possibility of nonspecific interactions between their designed aptamers and proteins in general. A nonspecific binding control should be included in all EMSA experiments.
Thank you for this constructive comment.
Following your recommendation, we are currently supplementing the EMSA assays with BSA as a non-target protein control to rigorously exclude potential non-specific binding between our designed aptamers (Ta and Ta variants) and exogenous proteins. These additional experiments are designed to directly assess whether the aptamers exhibit unintended interactions with unrelated proteins and to further validate the protein specificity of the RBD–aptamer interaction observed in our study.
The resulting nonspecific binding control data will be formally incorporated into the revised manuscript as Figure S11, and the corresponding Results and Discussion sections will be updated accordingly to reflect this critical validation once the experiments are completed.
(2) The evidence supporting claims of better binding to RBD by the aptamer compared to the commercial antibody is flawed at best. The commercial antibody product page indicates an affinity in low nanomolar range, whereas the fitted values they found for the aptamers in their study are orders of magnitude higher at tens of micromolar. Moreover, the methods section is lacking in the details required to appropriately interpret the competitive binding experiments. With a relatively short 20-minute equilibration time, the order of when the aptamer is added versus the antibody makes a difference in which is apparently bound. The issue with this becomes apparent with the lack of internal consistency in the presented results, namely in comparing Fig 3E (which shows no interference of Ta binding with 5uM antibody) and Fig 5D (which shows interference of Ta binding with 0.67-1.67uM antibody). The discrepancy between these figures calls into question the methods used, and it necessitates more details regarding experimental methods used in this manuscript.
Thank you for your insightful comments, which have helped us refine the rigor of our study. We address each of your concerns in detail below:
First, we agree with your observation that the commercial neutralizing antibody (Sino Biological, Cat# 40592-R001) is reported to bind Spike RBD with low nanomolar affinity on its product page. However, this discrepancy in affinity values (nanomolar vs. micromolar) stems from the use of distinct analytical methods. The product page affinity was determined via the Octet RED System, a technique analogous to Surface Plasmon Resonance (SPR) that offers high sensitivity for kinetic and affinity measurements. In contrast, our study employed EMSA, a method primarily optimized for semi-quantitative assessment of binding interactions. The inherent differences in sensitivity and principle between these two techniques—with Octet RED System enabling real-time monitoring of biomolecular interactions and EMSA relying on gel separation—account for the observed variation in affinity values.
Second, regarding the competitive binding experiments, we appreciate your note on the critical role of reagent addition order and equilibration time. To eliminate potential biases from sequential addition, we clarify that Cy3-labeled RNAs, RBD proteins, and the neutralizing antibody were added simultaneously to the reaction system. We will revise the Methods section in the revised manuscript to provide a detailed protocol for the EMSA experiments, to ensure full reproducibility and appropriate interpretation of the results.
Third, we acknowledge and apologize for a critical error in the figure legends of Figure 3E: the concentrations reported (5 μM aptamer and antibody 40592-R001) refer to stock solutions, not the final concentrations in the EMSA reaction mixture. The correct final concentrations are 0.5 μM for aptamer Ta, and 0.5 μM for the antibody. This correction resolves the apparent inconsistency between Figure 3E and Figure 5D, as the final antibody concentration in Figure 3E is now consistent with the concentration range used in Figure 5D. We will update the figure legends for Figure 3E and revise the Methods section to explicitly distinguish between stock and final reaction concentrations, ensuring clarity and internal consistency of the results.
We sincerely thank you for highlighting these issues, which will prompt important revisions to improve the clarity, accuracy, and rigor of our manuscript.
(3) The utility of the approach for increasing affinity of RNA aptamers for their targets is well supported through computational and experimental techniques demonstrating relative improvements in binding affinity for their G34C variant compared to the starting Ta aptamer. While the EMSA experiments do have significant flaws, the observations of relative relationships in equilibrium binding affinities among the tested aptamer variants can be interpreted with reasonable confidence, given that they were all performed in a consistent manner.
We sincerely appreciate your valuable concerns and constructive feedback, which have greatly facilitated the improvement of our manuscript. Regarding the flaws of the EMSA experiments you pointed out, we have provided a detailed response to clarify the related issues and supplemented necessary experimental details to enhance the rigor and reproducibility of our work (see corresponding response above). It is worth noting that EMSA remains a classic and widely used technique for studying biomolecular interactions, and its reliability in qualitative and semi-quantitative analysis of binding events has been well recognized in the field. Furthermore, we fully agree with and are grateful for your view that, since all tested aptamer variants were analyzed using a consistent experimental protocol, the observations on the relative relationships of their equilibrium binding affinities can be interpreted with reasonable confidence. This recognition reinforces the validity of the relative affinity improvements we observed for the G34C variant compared to the parental Ta aptamer, which is a key finding of our study.
(4) The claim that the structure of the RBD-Aptamer complex predicted by the CAAMO pipeline is reliable is tenuous. The success of their rational design approach based on the structure predicted by several ensemble approaches supports the interpretation of the predicted structure as reasonable, however, no experimental validation is undertaken to assess the accuracy of the structure. This is not a main focus of the manuscript, given the applied nature of the study to identify Ta variants with improved binding affinity, however the structural accuracy claim is not strongly supported without experimental validation (i.e. chemical footprinting methods).
We thank the reviewer for this comment and agree that experimental validation would be required to establish the structural accuracy of the predicted RBD–aptamer complex. We note, however, that the primary aim of this study is not structural determination, but the development of a general computational framework for aptamer affinity maturation. In most practical applications, experimentally resolved structures of aptamer–protein complexes are unavailable. Accordingly, CAAMO is designed to operate under such conditions, using computationally generated binding models as working hypotheses to guide rational optimization rather than as definitive structural descriptions. In this context, the predicted structure is evaluated by its utility for affinity improvement, rather than by direct structural validation. We will revise the manuscript accordingly to further clarify this scope.
(5) Throughout the manuscript, the phrasing of "all tested antibodies" was used, despite there being only one tested antibody in experimental methods and three distinct antibodies in computational methods. While this concern is focused on specific language, the major conclusion that their designed aptamers are as good or better than neutralizing antibodies in general is weakened by only testing only three antibodies through computational binding measurements and a fourth single antibody for experimental testing. The contact residue mapping furthermore lacks clarity in the number of structures that were used, with a vague description of structures from the PDB including no accession numbers provided nor how many distinct antibodies were included for contact residue mapping.
We thank the reviewer for this important comment regarding language precision, experimental scope, and clarity of the antibody dataset used in this study. We agree that the phrase “all tested antibodies” was imprecise and could lead to overgeneralization. We will carefully revise the manuscript to use more accurate and explicit wording throughout, clearly distinguishing between experimentally tested antibodies, computationally analyzed antibodies, and antibody structures used for large-scale contact analysis.
Specifically, the experimental comparison in this study was performed using one commercially available SARS-CoV-2 neutralizing antibody, whereas free energy–based computational analyses were conducted on three representative neutralizing antibodies with available structural data. We will revise the manuscript to explicitly state these distinctions and avoid general statements referring to neutralizing antibodies as a class.
Importantly, the residue-level contact frequency analysis was not based solely on these individual antibodies. Instead, this analysis leveraged a comprehensive set of experimentally resolved SARS-CoV-2 RBD–antibody complex structures curated from the Coronavirus Antibody Database (CoV-AbDab), a publicly available and actively maintained resource developed by the Oxford Protein Informatics Group. CoV-AbDab aggregates all published coronavirus-binding antibodies with associated PDB structures and provides a systematic and unbiased structural foundation for antibody–RBD interaction analysis. All available high-resolution RBD–antibody complex structures indexed in CoV-AbDab at the time of analysis were included to compute contact residue frequencies across the structural ensemble. We will explicitly state this data source, clarify the number and nature of structures used, and add the appropriate citation (Raybould et al., Bioinformatics, 2021, doi: 10.1093/bioinformatics/btaa739).
Finally, we will revise the conclusions to avoid claims that extend beyond the scope of the data. The comparison between aptamers and antibodies is now framed in terms of representative antibodies and consensus interaction patterns derived from a large structural ensemble, rather than as a general statement about all neutralizing antibodies. These revisions will improve the clarity, rigor, and reproducibility of the manuscript, while preserving the core conclusion that the CAAMO framework enables effective structure-guided affinity maturation of RNA aptamers.
Overall, the manuscript by Yang et al presents a valuable tool for rational design of improved RNA aptamer binding affinity toward target proteins, which the authors call CAAMO. Notably, the method is not intended for de novo design, but rather as a tool for improving aptamers that have been selected for binding affinity by other methods such as SELEX. While there are significant issues in the conclusions made from experiments in this manuscript, the relative relationships of observed affinities within this study provide solid evidence that the CAAMO framework provides a valuable tool for researchers seeking to use rational design approaches for RNA aptamer affinity maturation.
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
Summary:
In this study, the authors attempt to devise general rules for aptamer design based on structure and sequence features. The main system they are testing is an aptamer targeting a viral sequence.
Strengths:
The method combines a series of well-established protocols, including docking, MD, and a lot of system-specific knowledge, to design several new versions of the Ta aptamer with improved binding affinity.
We thank the reviewer for this accurate summary and for recognizing the strength of our integrated computational–experimental workflow in improving aptamer affinity.
Weaknesses:
The approach requires a lot of existing knowledge and, importantly, an already known aptamer, which presumably was found with SELEX. In addition, although the aptamer may have a stronger binding affinity, it is not clear if any of it has any additional useful properties such as stability, etc.
Thanks for these critical comments.
(1) On the reliance on a known aptamer: We agree that our CAAMO framework is designed as a post-SELEX optimization platform rather than a tool for de novo discovery. Its primary utility lies in rationally enhancing the affinity of existing aptamers that may not yet be sequence-optimal, thereby complementing experimental technologies such as SELEX. The following has been added to “Introduction” of the revised manuscript. (Page 5, line 108 in the revised manuscript)
‘Rather than serving as a de novo aptamer discovery tool, CAAMO is designed as a post-SELEX optimization platform that rationally improves the binding capability of existing aptamers.’
(2) On stability and developability: We also appreciate the reviewer’s important reminder that affinity alone is not sufficient for therapeutic development. We acknowledge that the present study has focused mainly on affinity optimization, and properties such as nuclease resistance, structural stability, and overall developability were not evaluated. The following has been added to “Discussion and conclusion” of the revised manuscript. (Page 25, line 595 in the revised manuscript)
‘While the present study primarily focused on affinity optimization, we acknowledge that other key developability traits—such as nuclease resistance, structural and thermodynamic stability, and in vivo persistence—are equally critical for advancing aptamers toward therapeutic applications. These properties were not evaluated here but will be systematically addressed in future iterations of the CAAMO framework to enable comprehensive optimization of aptamer candidates.’
Reviewer #2 (Public review):
Summary:
This manuscript proposes a workflow for discovering and optimizing RNA aptamers, with application in the optimization of a SARS-CoV-2 RBD. The authors took a previously identified RNA aptamer, computationally docked it into one specific RBD structure, and searched for variants with higher predicted affinity. The variants were subsequently tested for RBD binding using gel retardation assays and competition with antibodies, and one was found to be a stronger binder by about three-fold than the founding aptamer.
Overall, this would be an interesting study if it were performed with truly high-affinity aptamers, and specificity was shown for RBD or several RBD variants.
Strengths:
The computational workflow appears to mostly correctly find stronger binders, though not de novo binders.
We thank the reviewer for the clear summary and for acknowledging that our workflow effectively prioritizes stronger binders.
Weaknesses:
(1) Antibody competition assays are reported with RBD at 40 µM, aptamer at 5 µM, and a titration of antibody between 0 and 1.2 µg. This approach does not make sense. The antibody concentration should be reported in µM. An estimation of the concentration is 0-8 pmol (from 0-1.2 µg), but that's not a concentration, so it is unknown whether enough antibody molecules were present to saturate all RBD molecules, let alone whether they could have displaced all aptamers.
Thanks for your insightful comment. We have calculated that 0–1.2 µg antibody corresponds to a final concentration range of 0–1.6 µM (see Author response image 1). In practice, 1.2 µg was the maximum amount of commercial antibody that could be added under the conditions of our assay. In the revised manuscript, all antibody amounts previously reported in µg have been converted to their corresponding molar concentrations in Fig. 1F and Fig. 5D. In addition, the exact antibody concentrations used in the EMSA assays are now explicitly stated in the Materials and Methods section under “EMSA experiments.” The following has been added to “EMSA experiments” of the revised manuscript. (Page 30 in the revised manuscript)
‘For competitive binding experiments, 40 μM of RBP proteins, 5 μM of annealed Cy3-labelled RNAs and increasing concentrations of SARS-CoV-2 neutralizing antibody 40592-R001 (0–1.67 μM) were mixed in the EMSA buffer and incubated at room temperature for 20 min.’
Author response image 1.
Estimation of antibody concentration. Assuming a molecular weight of 150 kDa, dissolving 1.2 µg of antibody in a 5 µL reaction volume results in a final concentration of 1.6 µM.

As shown in Figure 5D, the purpose of the antibody–aptamer competition assay was not to achieve full saturation but rather to compare the relative competitive binding of the optimized aptamer (Ta<sup>G34C</sup>) versus the parental aptamer (Ta). Molecular interactions at this scale represent a dynamic equilibrium of binding and dissociation. While the antibody concentration may not have been sufficient to saturate all available RBD molecules, the experimental results clearly reveal the competitive binding behavior that distinguishes the two aptamers. Specifically, two consistent trends emerged:
(1) Across all antibody concentrations, the free RNA band for Ta was stronger than that of Ta<sup>G34C</sup>, while the RBD–RNA complex band of the latter was significantly stronger, indicating that Ta<sup>G34C</sup> bound more strongly to RBD.
(2) For Ta, increasing antibody concentration progressively reduced the RBD–RNA complex band, consistent with antibody displacing the aptamer. In contrast, for Ta<sup>G34C</sup>, the RBD–RNA complex band remained largely unchanged across all tested antibody concentrations, suggesting that the antibody was insufficient to displace Ta<sup>G34C</sup> from the complex.
Together, these observations support the conclusion that Ta<sup>G34C</sup> exhibits markedly stronger binding to RBD than the parental Ta aptamer, in line with the predictions and objectives of our CAAMO optimization framework.
(2) These are not by any means high-affinity aptamers. The starting sequence has an estimated (not measured, since the titration is incomplete) K<sub>d</sub> of 110 µM. That's really the same as non-specific binding for an interaction between an RNA and a protein. This makes the title of the manuscript misleading. No high-affinity aptamer is presented in this study. If the docking truly presented a bound conformation of an aptamer to a protein, a sub-micromolar K<sub>d</sub> would be expected, based on the number of interactions that they make.
In fact, our starting sequence (Ta) is a high-affinity aptamer, and then the optimized sequences (such as Ta<sup>G34C</sup>) with enhanced affinity are undoubtedly also high-affinity aptamers. See descriptions below:
(1) Origin and prior characterization of Ta. The starting aptamer Ta (referred to as RBD-PB6-Ta in the original publication by Valero et al., PNAS 2021, doi:10.1073/pnas.2112942118) was selected through multiple positive rounds of SELEX against SARS-CoV-2 RBD, together with counter-selection steps to eliminate non-specific binders. In that study, Ta was reported to bind RBD with an IC₅₀ of ~200 nM as measured by biolayer interferometry (BLI), supporting its high affinity and specificity. The following has been added to “Introduction” of the revised manuscript. (Page 4 in the revised manuscript)
‘This aptamer was originally identified through SELEX and subsequently validated using surface plasmon resonance (SPR) and biolayer interferometry (BLI), which confirmed its high affinity (sub-nanomolar) and high specificity toward the RBD. Therefore, Ta provides a well-characterized and biologically relevant starting point for structure-based optimization.’
(2) Methodological differences between EMSA and BLI measurements. We acknowledge that the discrepancy between our obtained binding affinity (K<sub>d</sub> = 110 µM) and the previously reported one (IC<sub>50</sub> ~ 200 nM) for the same Ta sequence arises primarily from methodological and experimental differences between EMSA and BLI. Namely, different experimental measurement methods can yield varied binding affinity values. While EMSA may have relatively low measurement precision, its relatively simple procedures were the primary reason for its selection in this study. Particularly, our framework (CAAMO) is designed not as a tool for absolute affinity determination, but as a post-SELEX optimization platform that prioritizes relative changes in binding affinity under a consistent experimental setup. Thus, the central aim of our work is to demonstrate that CAAMO can reliably identify variants, such as Ta<sup>G34C</sup>, that bind more strongly than the parental sequence under identical assay conditions. The following has been added to “Discussion and conclusion” of the revised manuscript. (Page 24 in the revised manuscript)
‘Although the absolute K<sub>d</sub> values determined by EMSA cannot be directly compared with surface-based methods such as SPR or BLI, the relative affinity trends remain highly consistent. While EMSA provides semi-quantitative affinity estimates, the close agreement between experimental EMSA trends and FEP-calculated ΔΔG values supports the robustness of the relative affinity changes reported here. In future studies, additional orthogonal biophysical techniques (e.g., filter-binding, SPR, or BLI) will be employed to further validate and refine the protein–aptamer interaction models.’
(3) Evidence of specific binding in our assays. We emphasize that the binding observed in our EMSA experiments reflects genuine aptamer–protein interactions. As shown in Figure 2G, a control RNA (Tc) exhibited no detectable binding to RBD, whereas Ta produced a clear binding curve, confirming that the interaction is specific rather than non-specific.
(3) The binding energies estimated from calculations and those obtained from the gel-shift experiments are vastly different, as calculated from the K<sub>d</sub> measurements, making them useless for comparison, except for estimating relative affinities.
Author Reply: We thank the reviewer for raising this important point. CAAMO was developed as a post-SELEX optimization tool with the explicit goal of predicting relative affinity changes (ΔΔG) rather than absolute binding free energies (ΔG). Empirically, CAAMO correctly predicted the direction of affinity change for 5 out of 6 designed variants (e.g., ΔΔG < 0 indicates enhanced binding free energy relative to WT); such predictive power for relative ranking is highly valuable for prioritizing candidates for experimental testing. Our prior work on RNA–protein interactions likewise supports the reliability of relative affinity predictions (see: Nat Commun 2023, doi:10.1038/s41467-023-39410-8). The following has been added to “Discussion and conclusion” of the revised manuscript. (Page 24 in the revised manuscript)
‘While EMSA provides semi-quantitative affinity estimates, the close agreement between experimental EMSA trends and FEP-calculated ΔΔG values supports the robustness of the relative affinity changes reported here.’
Recommendations for the Authors:
Reviewer #1 (Recommendations for the authors)
(1) Overall, the paper is well-written and, in the opinion of this reviewer, could remain as it is.
We thank the reviewer for the positive evaluation and supportive comments regarding our manuscript. We are grateful for the endorsement of its quality and suitability for publication.
Reviewer #2 (Recommendations for the authors)
(1) All molecules present in experiments need to be reported with their final concentrations (not µg).
We thank the reviewer for raising this important point. In the revised manuscript, all antibody amounts previously reported in µg have been converted to their corresponding molar concentrations in Fig. 1F and Fig. 5D. In addition, the exact antibody concentrations used in the EMSA assays are now explicitly stated in the Materials and Methods section under “EMSA experiments.” The following has been added to “EMSA experiments” of the revised manuscript. (Page 30 in the revised manuscript)
‘For competitive binding experiments, 40 μM of RBP proteins, 5 μM of annealed Cy3-labelled RNAs and increasing concentrations of SARS-CoV-2 neutralizing antibody 40592-R001 (0–1.67 μM) were mixed in the EMSA buffer and incubated at room temperature for 20 min.’
(2) An independent K<sub>d</sub> measurement, for example, using a filter binding assay, would greatly strengthen the results.
We thank the reviewer for this constructive suggestion and agree that an orthogonal biophysical measurement (e.g., a filter-binding assay, SPR or BLI) would further strengthen confidence in the reported dissociation constants. Unfortunately, all available SARS-CoV-2 RBD protein used in this study has been fully consumed and, due to current supply limitations, we were unable to perform new orthogonal binding experiments for the revised manuscript. We regret this limitation and have documented it in the Discussion as an item for future work.
Importantly, although we could not perform a new filter-binding experiment at this stage, we have multiple independent lines of evidence that support the reliability of the EMSA-derived affinity trends reported in the manuscript:
(1) Rigorous EMSA design and reproducibility. All EMSA binding curves reported in the manuscript (e.g., Figs. 2F–G, 4E–F, 5A and Fig. S9) are derived from three independent biological replicates and include standard deviations; the measured binding curves show good reproducibility across replicates.
(2) Appropriate positive and negative controls. Our gel assays include clear internal controls. The literature-reported strong binder Ta forms a distinct aptamer–RBD complex band under our conditions, whereas the negative-control aptamer Tc shows no detectable binding under identical conditions (see Fig. 2F). These controls demonstrate that the EMSA system discriminates specific from non-binding sequences with high sensitivity.
(3) Orthogonal computational validation (FEP) that agrees with experiment. The central strength of the CAAMO framework is the integration of rigorous physics-based calculations with experiments. We performed FEP calculations for the selected single-nucleotide mutations and computed ΔΔG values for each mutant. The direction and rank order of binding changes predicted by FEP are in good agreement with the EMSA measurements: five of six FEP-predicted improved mutants (Ta<sup>G34C</sup>, Ta<sup>G34U</sup>, Ta<sup>G34A</sup>, Ta<sup>C23A</sup>, Ta<sup>C23U</sup>) were experimentally confirmed to have stronger apparent affinity than wild-type Ta (see Fig. 4D–F, Table S2), yielding a success rate of 83%. The concordance between an independent, rigorous computational method and our experimental measurements provides strong mutual validation.
(4) Independent competitive binding experiments. We additionally performed competitive EMSA assays against a commercial neutralizing monoclonal antibody (40592-R001). These competition experiments show that Ta<sup>G34C</sup>–RBD complexes are resistant to antibody displacement under conditions that partially displace the wild-type Ta–RBD complex (see Fig. 5D). This result provides an independent, functionally relevant line of evidence that Ta<sup>G34C</sup> binds RBD with substantially higher affinity and specificity than WT Ta under our assay conditions.
Given these multiple, independent lines of validation (rigorous EMSA replicates and controls, FEP agreement, and antibody competition assays), we are confident that the relative affinity improvements reported in the manuscript are robust, even though the absolute K<sub>d</sub> values measured by EMSA are not directly comparable to surface-based methods (EMSA typically reports larger apparent K<sub>d</sub> values than SPR/BLI due to methodological differences). The following has been added to “Discussion and conclusion” of the revised manuscript. (Page 24 in the revised manuscript)
‘Although the absolute K<sub>d</sub> values determined by EMSA cannot be directly compared with surface-based methods such as SPR or BLI, the relative affinity trends remain highly consistent. While EMSA provides semi-quantitative affinity estimates, the close agreement between experimental EMSA trends and FEP-calculated ΔΔG values supports the robustness of the relative affinity changes reported here. In future studies, additional orthogonal biophysical techniques (e.g., filter-binding, SPR, or BLI) will be employed to further validate and refine the protein–aptamer interaction models.’
(3) The project would really benefit from a different aptamer-target system. Starting with a 100 µM aptamer is really not adequate.
We thank the reviewer for this important suggestion and for highlighting the value of testing the CAAMO framework in additional aptamer–target systems.
First, we wish to clarify the rationale for selecting the Ta–RBD system as the proof-of-concept. The Ta aptamer is not an arbitrary or weak binder: it was originally identified by independent SELEX experiments and subsequently validated by rigorous biophysical assays (SPR and BLI) (see: Proc. Natl. Acad. Sci. 2021, doi: 10.1073/pnas.2112942118). That study confirmed that Ta exhibits high-affinity and high-specificity binding to the SARS-CoV-2 RBD, which is why it serves as a well-characterized and biologically relevant system for method validation and optimization. We have added a brief clarification to the “Introduction” to emphasize these points. The following has been added to “Introduction” of the revised manuscript. (Page 4 in the revised manuscript)
‘This aptamer was originally identified through SELEX and subsequently validated using surface plasmon resonance (SPR) and biolayer interferometry (BLI), which confirmed its high affinity and high specificity toward the RBD. Therefore, Ta provides a well-characterized and biologically relevant starting point for structure-based optimization.’
Second, we agree that apparent discrepancies in absolute K<sub>d</sub> values can arise from different experimental platforms. Surface-based methods (SPR/BLI) and gel-shift assays (EMSA) have distinct measurement principles; EMSA yields semi-quantitative, solution-phase, apparent K<sub>d</sub> values that are not directly comparable in absolute magnitude to surface-based measurements. Crucially, however, our study focuses on relative affinity change. EMSA is well suited for parallel, comparative measurements across multiple variants when all samples are assayed under identical conditions, and thus provides a reliable readout for ranking and validating designed mutations. We have added a short statement in the “Discussion and conclusion”. The following has been added to “Discussion and conclusion” of the revised manuscript. (Page 24 in the revised manuscript)
‘Although the absolute K<sub>d</sub> values determined by EMSA cannot be directly compared with surface-based methods such as SPR or BLI, the relative affinity trends remain highly consistent. While EMSA provides semi-quantitative affinity estimates, the close agreement between experimental EMSA trends and FEP-calculated ΔΔG values supports the robustness of the relative affinity changes reported here. In future studies, additional orthogonal biophysical techniques (e.g., filter-binding, SPR, or BLI) will be employed to further validate and refine the protein–aptamer interaction models.’
Third, and importantly, CAAMO is inherently generalizable. In addition to the Ta–RBD application presented here, we have already begun applying CAAMO to other aptamer–target systems. In particular, we have successfully deployed the framework in preliminary optimization studies of RNA aptamers targeting the epidermal growth factor receptor (EGFR) (see: Gastroenterology 2021, doi: 10.1053/j.gastro.2021.05.055) (see Author response image 2). These preliminary results support the transferability of the CAAMO pipeline beyond the SARS-CoV-2 RBD system. We have added a short statement in the “Discussion and conclusion”. The following has been added to “Discussion and conclusion” of the revised manuscript. (Page 259 in the revised manuscript)
‘In addition to the Ta–RBD system, the CAAMO framework itself is inherently generalizable. More work is currently underway to apply CAAMO to optimize aptamers targeting other therapeutically relevant proteins, such as the epidermal growth factor receptor (EGFR) [45], in order to further explore its potential for broader aptamer engineering.’
Author response image 2.
Overview of the predicted binding model of the EGFR–aptamer complex generated using the CAAMO framework.
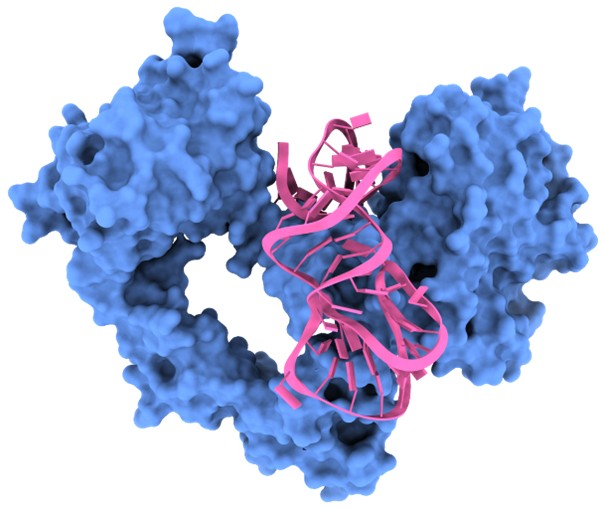
(4) Several RBD variants should be tested, as well as other proteins, for specificity. At such weak affinities, it is likely that these are non-specific binders.
We thank the reviewer for this important concern. Below we clarify the basis for selecting Ta and its engineered variants, summarize the experimental controls that address specificity, and present the extensive in silico variant analysis we performed to assess sensitivity and breadth of binding.
(1) Origin and validation of Ta. As noted in our response to “Comment (3)”, the Ta aptamer was not chosen arbitrarily. Ta was identified by independent SELEX with both positive and negative selection and subsequently validated using surface-based biophysical assays (SPR and BLI), which reported low-nanomolar affinity and high specificity for the SARS-CoV-2 RBD. Thus, Ta is a well-characterized, experimentally validated starting lead for method development and optimization.
(2) Experimental specificity controls. We appreciate the concern that weak apparent affinities can reflect non-specific binding. As noted in our response to “Comment (2)”, we applied multiple experimental controls that argue against non-specificity: (i) a literature-reported weak binder (Tc) was used as a negative control and produced no detectable complex under identical EMSA conditions (see Figs. 2F–G), demonstrating the assay’s ability to discriminate non-binders from specific binders; (ii) competitive EMSA assays with a commercial neutralizing monoclonal antibody (40592-R001) show that both Ta and Ta<sup>G34C</sup> engage the same or overlapping RBD site as the antibody, and that Ta<sup>G34C</sup> is substantially more resistant to antibody displacement than WT Ta (see Figs. 3D–E, 5D). Together, these wet-lab controls support that the observed aptamer-RBD bands reflect specific interactions rather than general, non-specific adsorption.
(3) Variant and specificity analysis by rigorous FEP calculations. To address the reviewer’s request to evaluate variant sensitivity, we performed extensive free energy perturbation combined with Hamiltonian replica-exchange molecular dynamics (FEP/HREX) for improved convergence efficiency and increased simulation time to estimate relative binding free energy changes (ΔΔG) of both WT Ta and the optimized Ta<sup>G34C</sup> against a panel of RBD variants. Results are provided in Tables S4 and S5. Representative findings include: For WT Ta versus early lineages, FEP reproduces the experimentally observed trends: Alpha (B.1.1.7; N501Y) yields ΔΔG<sub>FEP</sub> = −0.42 ± 0.07 kcal/mol (ΔΔG<sub>exp</sub> = −0.24), while Beta (B.1.351; K417N/E484K/N501Y) gives ΔΔG<sub>FEP</sub> = 0.64 ± 0.25 kcal/mol (ΔΔG<sub>exp</sub> = 0.36) (see Table S4). The agreement between the computational and experimental results supports the fidelity of our computational model for variant assessment. For the engineered Ta<sup>G34C</sup>, calculations across a broad panel of variants indicate that Ta<sup>G34C</sup> retains or improves binding (ΔΔG < 0) for the majority of tested variants, including Alpha, Beta, Gamma and many Omicron sublineages. Notable examples: BA.1 (ΔΔG = −3.00 ± 0.52 kcal/mol), BA.2 (ΔΔG = −2.54 ± 0.60 kcal/mol), BA.2.75 (ΔΔG = −5.03 ± 0.81 kcal/mol), XBB (ΔΔG = −3.13 ± 0.73 kcal/mol) and XBB.1.5 (ΔΔG = −2.28 ± 0.96 kcal/mol). A minority of other Omicron sublineages (e.g., BA.4 and BA.5) show modest positive ΔΔG values (2.11 ± 0.67 and 2.27 ± 0.68 kcal/mol, respectively), indicating a predicted reduction in affinity for those specific backgrounds. Overall, these data indicate that the designed Ta<sup>G34C</sup> aptamer can maintain its binding ability with most SARS-CoV-2 variants, showing potential for broad-spectrum antiviral activity (see Table S5). The following has been added to “Results” of the revised manuscript. (Page 22 in the revised manuscript)
‘2.6 Binding performance of Ta and Ta<sup>G34C</sup> against SARS-CoV-2 RBD variants
To further evaluate the binding performance and specificity of the designed aptamer Ta<sup>G34C</sup> toward various SARS-CoV-2 variants [39], we conducted extensive free energy perturbation combined with Hamiltonian replica-exchange molecular dynamics (FEP/HREX) [40–42] for both the wild-type aptamer Ta and the optimized Ta<sup>G34C</sup> against a series of RBD mutants. The representative variants include the early Alpha (B.1.1.7) and Beta (B.1.351) lineages, as well as a panel of Omicron sublineages (BA.1–BA.5, BA.2.75, BQ.1, XBB, XBB.1.5, EG.5.1, HK.3, JN.1, and KP.3) carrying multiple mutations within the RBD region (residues 333–527). For each variant, mutations within 5 Å of the bound aptamer were included in the FEP to accurately estimate the relative binding free energy change (ΔΔG).
For the wild-type Ta aptamer, the FEP-predicted binding affinities toward the Alpha and Beta RBD variants were consistent with the previous experimental results, further validating the reliability of our model (see Table S4). Specifically, Ta maintained comparable or slightly enhanced binding to the Alpha variant and showed only marginally reduced affinity for the Beta variant.
In contrast, the optimized aptamer Ta<sup>G34C</sup> exhibited markedly improved and broad-spectrum binding capability toward most tested variants (see Table S5). For early variants such as Alpha, Beta, and Gamma, Ta<sup>G34C</sup> maintained enhanced affinities (ΔΔG < 0). Notably, for multiple Omicron sublineages—including BA.1, BA.2, BA.2.12.1, BA.2.75, XBB, XBB.1.5, XBB.1.16, XBB.1.9, XBB.2.3, EG.5.1, XBB.1.5.70, HK.3, BA.2.86, JN.1 and JN.1.11.1—the calculated binding free energy changes ranged from −1.89 to −7.58 kcal/mol relative to the wild-type RBD, indicating substantially stronger interactions despite the accumulation of multiple mutations at the aptamer–RBD interface. Only in a few other Omicron sublineages, such as BA.4, BA.5, and KP.3, a slight reduction in binding affinity was observed (ΔΔG > 0).
These computational findings demonstrate that the Ta<sup>G34C</sup> aptamer not only preserves high affinity for the RBD but also exhibits improved tolerance to the extensive mutational landscape of SARS-CoV-2. Collectively, our results suggest that Ta<sup>G34C</sup> holds promise as a high-affinity and potentially cross-variant aptamer candidate for targeting diverse SARS-CoV-2 spike protein variants, showing potential for broad-spectrum antiviral activity.’
The following has been added to “Materials and Methods” of the revised manuscript. (Page 29 in the revised manuscript)
‘4.7 FEP/HREX
To evaluate the binding sensitivity of the optimized aptamer Ta<sup>G34C</sup> toward SARS-CoV-2 RBD variants, we employed free energy perturbation combined with Hamiltonian replica-exchange molecular dynamics (FEP/HREX) simulations for enhanced sampling efficiency and improved convergence. The relative binding free energy changes (ΔΔG) upon RBD mutations were estimated as:
ΔΔ𝐺 = Δ𝐺<sub>bound</sub> − Δ𝐺<sub>free</sub>
where ΔG<sub>bound</sub> and ΔG<sub>free</sub> represent the RBD mutations-induced free energy changes in the complexed and unbound states, respectively. All simulations were performed using GROMACS 2021.5 with the Amber ff14SB force field. For each mutation, dual-topology structures were generated in a pmx-like manner, and 32 λ-windows (0.0, 0.01, 0.02, 0.03, 0.06, 0.09, 0.12, 0.16, 0.20, 0.24, 0.28, 0.32, 0.36, 0.40, 0.44, 0.48, 0.52, 0.56, 0.60, 0.64, 0.68, 0.72, 0.76, 0.80, 0.84, 0.88, 0.91, 0.94, 0.97, 0.98, 0.99, 1.0) were distributed uniformly between 0.0 and 1.0. To ensure sufficient sampling, each window was simulated for 5 ns, with five independent replicas initiated from distinct velocity seeds. Replica exchange between adjacent λ states was attempted every 1 ps to enhance phase-space overlap and sampling convergence. The van der Waals and electrostatic transformations were performed simultaneously, employing a soft-core potential (α = 0.3) to avoid singularities. For each RBD variant system, this setup resulted in an accumulated simulation time of approximately 1600 ns (5 ns × 32 windows × 5 replicas × 2 states). The Gromacs bar analysis tool was used to estimate the binding free energy changes.’
Tables S4 and S5 have been added to Supplementary Information of the revised manuscript.
-
-
www.france.tv www.france.tv
-
Synthèse : "Braver l'ombre, nos vies après le harcèlement"
Synthèse Exécutive
Ce document analyse les dynamiques complexes du harcèlement scolaire en s'appuyant sur les témoignages croisés d'Anna, une jeune femme victime de harcèlement sévère, et de Zacharie, un ancien harceleur.
Il explore les motivations profondes derrière les actes de harcèlement, notamment la quête de statut social et la peur d'être soi-même une victime, souvent déclenchés par la perception d'une "différence" chez la cible.
L'analyse met en lumière les conséquences dévastatrices et durables pour la victime, incluant des traumatismes psychologiques profonds, l'anxiété chronique, des troubles du comportement et une perte d'estime de soi.
Parallèlement, elle retrace le parcours de l'auteur, de la recherche d'une réputation à une prise de conscience tardive, suivie d'un besoin de réparation.
Le rôle crucial, et souvent défaillant, des adultes et des institutions scolaires est examiné, soulignant leur incapacité à identifier, comprendre et gérer adéquatement les situations.
Enfin, le document se conclut sur les thèmes de la reconstruction personnelle et du potentiel de la justice restaurative, incarnée par la rencontre entre Anna et Zacharie, comme voie vers la guérison et la compréhension mutuelle.
Profils Centraux : La Victime et l'Auteur
Anna : Le Parcours de la Victime
Anna est décrite comme une jeune fille initialement « très enjouée » et « curieuse », mais marquée dès l'enfance par une anxiété profonde.
Cette anxiété, ressentie dès le CM1, la plaçait déjà en décalage avec ses pairs. Son parcours illustre l'escalade et les effets à long terme du harcèlement.
• Nature du harcèlement : Le harcèlement commence par des moqueries dès la 6ème, s'intensifie brutalement en 4ème pour devenir quotidien et violent. Il prend plusieurs formes :
◦ Psychologique : Moqueries constantes sur son apparence (« castor », « cheval »), sa dentition, son style vestimentaire et son comportement. Insultes quotidiennes : « tu es vraiment moche », « tu pues », « va te pendre », « crève ».
◦ Social : Isolement complet, y compris de la part de ses amis qui, par peur, se joignent aux harceleurs.
◦ Physique : Elle reçoit un coup de coude violent dans le dos, la faisant s'effondrer en larmes.
◦ Institutionnel : Une professeure en 3ème participe activement à son humiliation publique, se moquant de ses problèmes d'anxiété et de sommeil devant toute la classe, provoquant un sentiment d'humiliation constant et une perte totale de repères.
• Conséquences : Les répercussions du harcèlement sur Anna sont profondes et multidimensionnelles.
◦ Traumatismes : Elle souffre de traumatismes si sévères qu'elle n'a « zéro souvenir » de son année de 5ème. Elle développe une phobie scolaire, décrivant l'entrée au collège comme une « angoisse qui me prenait tellement fort ».
◦ Santé mentale et physique : Elle développe des troubles anxieux majeurs, des crises de panique, des troubles du sommeil, des troubles alimentaires et recourt à l'automutilation (« me gratter, à me couper volontairement ou à me brûler ») pour gérer un « surplus d'émotion ».
Elle exprime avoir perdu toute raison de vivre.
◦ Séquelles durables : Adulte, elle vit avec une peur constante du jugement, interprétant le moindre rire dans la rue comme une moquerie dirigée contre elle.
Elle lutte pour reconstruire son estime de soi et apprend à « s'apprécier sur des petits points ».
Zacharie : La Trajectoire de l'Auteur
Zacharie, surnommé « le malaimé » dans son enfance, était un enfant « turbulent » cherchant à se faire remarquer.
Son parcours révèle comment la quête d'une place dans la hiérarchie sociale peut mener au harcèlement.
• Motivations : Ses actions sont motivées par un désir profond d'exister et d'appartenir aux « classes dominantes de la cour de récréation ».
Il explique : « Je voulais pas être quelqu'un qui se faisait dominer, je voulais être quelqu'un qui était reconnu ».
Le harcèlement devient un outil pour se forger une réputation de « mauvais garçon » et s'assurer qu'on ne l'« embête pas ».
Il admet que le comportement d'un professeur en CM1, qui utilisait l'humiliation, a contribué à normaliser ce type de violence.
• Actes de harcèlement : Il commence à harceler activement à la fin du CM1.
◦ Ciblage : Il identifie des cibles faciles, des élèves affichant des « normes déviantes », une « forme de fragilité » ou une « marginalité ».
◦ Escalade de la violence : Ses actes vont des brimades verbales à la violence physique.
Un événement particulièrement grave le voit projeter un camarade dans une poubelle en béton, menant à une convocation au commissariat.
En 5ème, il agresse un autre élève en lui frottant le visage avec une feuille urticante à laquelle il est allergique, ce qui entraîne une intervention des pompiers et son exclusion temporaire.
• Prise de conscience et réparation :
◦ Conséquences pour lui : Son comportement entraîne une chute drastique de ses résultats scolaires (de 14 à 7 de moyenne), son exclusion et un sentiment d'abandon.
◦ Le déclic : Sa prise de conscience s'opère en plusieurs temps : une remarque malheureuse à son petit frère qui lui répond qu'il se suiciderait s'il était homosexuel ; l'apprentissage qu'une de ses victimes souhaitait se venger physiquement ; et ses études en sociologie qui lui permettent de comprendre les mécanismes sociaux à l'œuvre.
◦ Démarche de réparation : Il décide de recontacter ses anciennes victimes pour présenter ses excuses, une démarche qu'il juge essentielle.
Pour lui, témoigner est une manière de « mettre mon expérience à contribution pour faire en sorte que on identifie vraiment les causes du harcèlement ».
Thèmes et Dynamiques Clés du Harcèlement
La "Différence" comme Facteur de Risque
La notion de "différence" est identifiée comme un catalyseur central du harcèlement.
• Pour Anna, son anxiété, son introversion et son côté « artiste » la singularisent et en font une cible.
Ses parents lui disent initialement : « tu es un peu différent, un peu bizarre, mets-toi un peu dans les normes et ça ira mieux ».
• Pour Zacharie, les victimes sont choisies parce qu'elles « affichent des normes déviantes » ou une « fragilité ».
Il explique que la différence, qu'elle soit due à l'introversion ou l'extraversion, est un « vrai facteur de risque ».
• Lui-même n'était « pas dans le moule », son comportement turbulent le distinguant et le poussant à trouver une autre manière d'exister.
Le Rôle Crucial et Ambivalent des Adultes
Le témoignage met en évidence l'impact déterminant, positif comme négatif, des adultes.
• L'institution Scolaire : L'école apparaît souvent comme une institution défaillante.
◦ Manque de réaction : Des professeurs sont témoins de moqueries répétées mais « ne disaient rien du tout ».
◦ Participation active : Le cas de la professeure d'Anna en 3ème, qui l'humilie publiquement, est l'exemple le plus extrême de la faillite de l'adulte protecteur.
◦ Réponses inadaptées : Les sanctions contre Zacharie (convocation à la police, exclusion) ne sont pas accompagnées d'une démarche pédagogique pour l'amener à réfléchir à ses actes.
Une conseillère d'orientation déclare même à sa mère : « Votre fils madame Zacharie, il est foutu ».
◦ Interventions positives : À l'inverse, certaines figures comme la professeure principale d'Anna (qui finit par comprendre), la CPE et l'infirmière, ou la nouvelle équipe enseignante de Zacharie au lycée, montrent qu'un soutien bienveillant peut radicalement changer une trajectoire.
• Les Parents : Les parents sont souvent démunis.
◦ Incompréhension initiale : Les parents d'Anna ne mesurent pas la gravité de la situation au début, ce qu'elle a ressenti comme un manque d'écoute.
◦ Déni : La mère de Zacharie a du mal à accepter la réalité des faits, parlant de « chamaillerie » et se demandant pourquoi les professeurs n'ont pas « mis de mots » sur la situation.
◦ Soutien et inquiétude : Une fois la situation comprise, les parents d'Anna deviennent un soutien indéfectible, bien que leur vie soit « beaucoup impactée ». Ils expriment leur inquiétude constante et leur sentiment d'impuissance.
Les Conséquences à Long Terme
Les effets du harcèlement s'inscrivent durablement dans la vie des individus concernés.
| Impact Psychologique | Impact Physique et Comportemental | Impact Social et Relationnel | | --- | --- | --- | | Anxiété chronique et crises d'angoisse | Troubles du sommeil | Isolement et perte des amis | | Traumatismes profonds et amnésie partielle | Troubles du comportement alimentaire | Difficulté à demander de l'aide et à faire confiance | | Perte d'estime de soi (« je vaux quelque chose ») | Automutilation (scarification, brûlures) | Perte de confiance en soi et peur du jugement | | Idées suicidaires (« plus de raison de vivre ») | Phobie scolaire | Difficulté à se reconstruire une identité positive |
La Reconstruction et la Justice Restaurative
Chemins de Guérison Individuels
• Pour Anna : La reconstruction passe par un suivi thérapeutique, le soutien familial, et la redécouverte de passions comme les réseaux sociaux (Musical.ly) qui lui ont permis de reprendre confiance.
Son projet de devenir infirmière est directement lié à son vécu : « je veux être utile dans la vie des gens ».
• Pour Zacharie : Son changement de collège et son orientation en bac professionnel marquent une rupture.
Il trouve une manière positive d'exister en devenant un bon élève et en aidant ses camarades.
Ses études en sociologie lui fournissent les outils intellectuels pour analyser son passé. Il choisit de vivre dans un camping-car, symbole d'une quête de « liberté ».
La Rencontre : Dialogue et Reconnaissance
La rencontre organisée entre Anna et Zacharie incarne une démarche de justice restaurative.
• Attentes : Anna espère que cette rencontre lui apportera des réponses et l'aidera à « faire le deuil de la période de mon harcèlement ». Zacharie y voit l'opportunité de montrer les « deux côtés d'une même histoire ».
• Déroulement : L'échange leur permet de confronter leurs expériences et leurs ressentis. Zacharie explique ses motivations, tandis qu'Anna décrit la violence de ce qu'elle a subi.
• Impact : La rencontre est un moment de reconnaissance mutuelle. Anna conclut que Zacharie n'est pas « une mauvaise personne parce que tu regrettes ce que tu as fait ».
Pour Zacharie, présenter ses excuses et montrer qu'il a changé est « beaucoup plus valorisant » que le harcèlement lui-même. Léa, une autre de ses victimes, confirme que sa démarche l'a aidée à « clore un chapitre ».
Messages et Conclusions
Les témoignages convergent vers plusieurs messages forts.
• Pour les victimes : Il est crucial de parler et de ne pas s'isoler dans la souffrance. Anna insiste : « c'est pas vous les coupables ». Le harcèlement est une épreuve qui « suit toute sa vie » si elle n'est pas stoppée.
• Pour les auteurs : Zacharie affirme que le harcèlement n'est « pas cool », « pas stylé » et qu'il ne rend pas « plus intéressant ». Il souligne que reconnaître ses torts est une étape difficile mais nécessaire et positive.
• Pour la société : Les témoignages appellent à une plus grande attention et à une prise au sérieux du phénomène.
Ils rappellent que le harcèlement prend racine dans une souffrance, y compris celle de l'auteur qui ne trouve pas sa place.
Le dialogue et la reconnaissance du mal causé sont présentés comme des outils puissants pour la réparation et la prévention.
-
-
www.biorxiv.org www.biorxiv.org
-
Reviewer #1 (Public review):
Summary:
The temporal regulation of neuronal specification and its molecular mechanisms are important problems in developmental neurobiology. This study focuses on Kenyon cells (KCs), which form the mushroom body in Drosophila melanogaster, in order to address this issue. Building on previous findings, the authors examine the role of the transcription factor Eip93F in the development of late-born KCs. The authors revealed that Eip93F controls the activity of flies at night through the expression of the calcium channel Ca-α1T. Thus, the study clarifies the molecular machinery that controls temporal neuronal specification and animal behavior.
Strengths:
The convincing results are based on state-of-the-art molecular genetics, imaging, and behavioral analysis.
-
Reviewer #2 (Public review):
Summary:
Understanding the mechanisms of neural specification is a central question in neurobiology. In Drosophila, the mushroom body (MB), which is the associative learning region in the brain, consists of three major cell types: γ, α'/β' and α/β kenyon cells. These classes can be further subdivided into seven subtypes, together comprising ~2000 KCs per hemi-brain. Remarkably, all of these neurons are derived from just four neuroblasts in each hemisphere. Therefore, a lot of endeavours are put to understand how the neuron is specified in the fly MB.
Over the past decade, studies have revealed that MB neuroblasts employ a temporal patterning mechanism, producing distinct neuronal types at different developmental stages. Temporal identity is conveyed through transcription factor expression in KCs. High levels of Chinmo, a BTB-zinc finger transcription factor, promote γ-cell fate (Zhu et al., Cell, 2006). Reduced Chinmo levels trigger expression of mamo, a zinc finger transcription factor that specifies α'/β' identity (Liu et al., eLife, 2019). However, the specification of α/β neurons remains poorly understood. Some evidence suggests that microRNAs regulate the transition from α'/β' to α/β fate (Wu et al., Dev Cell, 2012; Kucherenko et al., EMBO J, 2012). One hypothesis even proposes that α/β represents a "default" state of MB neurons, which could explain the difficulty in identifying dedicated regulators.
The study by Chung et al. challenges this hypothesis. By leveraging previously published RNA-seq datasets (Shih et al., G3, 2019), they systematically screened BAC transgenic lines to selectively label MB subtypes. Using these tools, they analyzed the consequences of manipulating E93 expression and found that E93 is required for α/β specification. Furthermore, loss of E93 impairs MB-dependent behaviors, highlighting its functional importance.
Strengths:
The authors conducted a thorough analysis of E93 manipulation phenotypes using LexA tools generated from the Janelia Farm and Bloomington collections. They demonstrated that E93 knockdown reduces expression of Ca-α1T, a calcium channel gene identified as an α/β marker. Supporting this conclusion, one LexA line driven by a DNA fragment near EcR (R44E04) showed consistent results. Conversely, overexpression of E93 in γ and α'/β' Kenyon cells led to downregulation of their respective subtype markers.
Another notable strength is the authors' effort to dissect the genetic epistasis between E93 and previously known regulators. Through MARCM and reporter analyses, they showed that Chinmo and Mamo suppress E93, while E93 itself suppresses mamo. This work establishes a compelling molecular model for the regulatory network underlying MB cell-type specification.
Weaknesses:
The interpretation of E93's role in neuronal specification requires caution. Typically, two criteria are used to establish whether a gene directs neuronal identity:
(1) gene manipulation shifts the neuronal transcriptome from one subtype to another, and
(2) gene manipulation alters axonal projection patterns.
The results presented here only partially satisfy the first criterion. Although markers are affected, it remains possible that the reporter lines and subtype markers used are direct transcriptional targets of E93 in α/β neurons, rather than reflecting broader fate changes. Future studies using transcriptomics would provide a more comprehensive assessment of neuronal identity following E93 perturbation.
With respect to the second criterion, the evidence is also incomplete. While reporter patterns were altered, the overall morphology of the α/β lobes appeared largely intact after E93 knockdown. Overexpression of E93 in γ neurons produced a small subset of cells with α/β-like projections, but this effect warrants deeper characterization before firm conclusions can be drawn.
Overall, this study has nicely shown that E93 can regulate α/β neural identities. Further studies on the regulatory network will help to better understand the mechanism of neurogenesis in mushroom body.
-
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
Summary:
The temporal regulation of neuronal specification and its molecular mechanisms are important problems in developmental neurobiology. This study focuses on Kenyon cells (KCs), which form the mushroom body in Drosophila melanogaster, in order to address this issue. Building on previous findings, the authors examine the role of the transcription factor Eip93F in the development of late-born KCs. The authors revealed that Eip93F controls the activity of flies at night through the expression of the calcium channel Ca-α1T. Thus, the study clarifies the molecular machinery that controls temporal neuronal specification and animal behavior.
Strengths:
The convincing results are based on state-of-the-art molecular genetics, imaging, and behavioral analysis.
Weaknesses:
Temporal mechanisms of neuronal specification are found in many nervous systems. However, the relationship between the temporal mechanisms identified in this study and those in other systems remains unclear.
We have discussed the temporal mechanisms between different nervous systems at the beginning of the Discussion section.
Reviewer #2 (Public review):
Summary:
Understanding the mechanisms of neural specification is a central question in neurobiology. In Drosophila, the mushroom body (MB), which is the associative learning region in the brain, consists of three major cell types: γ, α'/β', and α/β kenyon cells. These classes can be further subdivided into seven subtypes, together comprising ~2000 KCs per hemi-brain. Remarkably, all of these neurons are derived from just four neuroblasts in each hemisphere. Therefore, a lot of endeavors are put into understanding how the neuron is specified in the fly MB.
Over the past decade, studies have revealed that MB neuroblasts employ a temporal patterning mechanism, producing distinct neuronal types at different developmental stages. Temporal identity is conveyed through transcription factor expression in KCs. High levels of Chinmo, a BTB-zinc finger transcription factor, promote γ-cell fate (Zhu et al., Cell, 2006). Reduced Chinmo levels trigger expression of mamo, a zinc finger transcription factor that specifies α'/β' identity (Liu et al., eLife, 2019). However, the specification of α/β neurons remains poorly understood. Some evidence suggests that microRNAs regulate the transition from α'/β' to α/β fate (Wu et al., Dev Cell, 2012; Kucherenko et al., EMBO J, 2012). One hypothesis even proposes that α/β represents a "default" state of MB neurons, which could explain the difficulty in identifying dedicated regulators.
The study by Chung et al. challenges this hypothesis. By leveraging previously published RNA-seq datasets (Shih et al., G3, 2019), they systematically screened BAC transgenic lines to selectively label MB subtypes. Using these tools, they analyzed the consequences of manipulating E93 expression and found that E93 is required for α/β specification. Furthermore, loss of E93 impairs MB-dependent behaviors, highlighting its functional importance.
Strengths:
The authors conducted a thorough analysis of E93 manipulation phenotypes using LexA tools generated from the Janelia Farm and Bloomington collections. They demonstrated that E93 knockdown reduces expression of Ca-α1T, a calcium channel gene identified as an α/β marker. Supporting this conclusion, one LexA line driven by a DNA fragment near EcR (R44E04) showed consistent results. Conversely, overexpression of E93 in γ and α'/β' Kenyon cells led to downregulation of their respective subtype markers.
Another notable strength is the authors' effort to dissect the genetic epistasis between E93 and previously known regulators. Through MARCM and reporter analyses, they showed that Chinmo and Mamo suppress E93, while E93 itself suppresses Mamo. This work establishes a compelling molecular model for the regulatory network underlying MB cell-type specification.
Weaknesses:
The interpretation of E93's role in neuronal specification requires caution. Typically, two criteria are used to establish whether a gene directs neuronal identity:
(1) gene manipulation shifts the neuronal transcriptome from one subtype to another, and
(2) gene manipulation alters axonal projection patterns.
The results presented here only partially satisfy the first criterion. Although markers are affected, it remains possible that the reporter lines and subtype markers used are direct transcriptional targets of E93 in α/β neurons, rather than reflecting broader fate changes. Future studies using single-cell transcriptomics would provide a more comprehensive assessment of neuronal identity following E93 perturbation.
We do plan conduct multi-omics experiments to provide a more comprehensive assessment of neuronal identity upon loss-of-function of E93. However, omics results take time to be conducted and analyzed, so the result will be summarized in a future manuscript.
With respect to the second criterion, the evidence is also incomplete. While reporter patterns were altered, the overall morphology of the α/β lobes appeared largely intact after E93 knockdown. Overexpression of E93 in γ neurons produced a small subset of cells with α/β-like projections, but this effect warrants deeper characterization before firm conclusions can be drawn. While the results might be an intrinsic nature of KC types in flies, the interpretation of the reader of the data should be more careful, and the authors should also mention this in their main text.
We have toned down our description on the effect of E93 (especially in the loss-offunction) in specifying the α/β-specific cell identity and discussed whether unidentified regulators would work together with E93 in α/β neural fate specification.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
(1) Changes in nighttime activity in flies upon knocking down Ca_α1T and Eip93F are interesting (Fig. 2C). However, examining the morphological changes in the mushroom body under these conditions would be essential.
We did not find the morphological change of mushroom body lobes by examining with the Fas2 staining (shown in Figure S8D).
(2) Temporal mechanisms of neuronal specification have been identified in various nervous systems, including the embryonic central nervous system (CNS), the optic lobe of Drosophila, and the nervous systems of other organisms. The Discussion section should address the relationship between the temporal mechanisms identified in this study and those identified in other systems.
We have discussed the temporal mechanisms between different nervous systems at the beginning of the Discussion section.
(3) Eip93F is an Ecdysone-induced protein. In the Discussion section, the authors should discuss the relationship between the ecdysone signal and the roles of Eip93F.
We have added the discussion on the relationship between the ecdysone signal and the roles of Eip93F.
Reviewer #2 (Recommendations for the authors):
(1) The behavioral effect of Ca-α1T knockdown is pretty interesting. But how the downregulation of Ca-α1T in the mushroom body can affect locomotion is puzzling. Even though the mushroom body is known to suppress locomotion (Matin et al., Learn Mem, 1998), the real results are opposite. Can authors give further explanation in the discussion? Also, the behavioral experiments are hard to interpret, given that Figure 2C(1) and Figure 2C(3) as a control, also vary a lot. Since the behavioral experiments don't affect the main conclusion of the paper, I would suggest removing that part or adding more explanation in the discussion.
First, we have discussed the puzzling part on the MB influence in locomotion between the previous study using tetanus toxin light chain (TeNT-Ln) and our Ca-α1T knockdown result. It is possible that the different effect is derived from TeNT-Ln’s function in MB axons and Ca-α1T’s function in MB dendrites. Secondly, we have re-conducted the behavioral results using a new α/β driver (13F02-AD/70F05-DBD) to replace our initial behavioral results (using c739-GAL4, which would cause the abnormal wing when drives E93 RNAi expression; see S8C(2) Fig). Current results (now in Fig 2I) are more consistent in control groups.
(2) In the manuscript, the authors use "subtype" to describe γKC, α'/β'KC and α/βKC in the fly MB. However, in most of the literature, people use "main types" to summarize these three types, and "subtype" is mostly about the difference in γd, γm, α'/β'ap, α'/β'm, α/βp, α/βs and α/βc KC (Shih et al., G3, 2019). Replacing "subtypes" with "main types" will help to increase the clarity.
We have replaced "KC subtypes" with "main KC types" or just “KC types”.
(3) The authors have identified a lot of new markers for the KC cell types, and some of them are used in this manuscript. It will be helpful if they can have a figure to summarize the markers they used in this study and what cell types they labeled.
We have summarized expression patterns of these markers in Supplemental table 1.
(4) In the method, the authors mentioned that only females were selected for analysis of Ca-α1T-GFSTF. Could the authors explain the reasons in more detail?
Since homozygous Ca-α1T-GFSTF female flies and hemizygous Ca-α1T-GFSTF male are a bit sick and hard to collect, we therefore used heterozygous Ca-α1T-GFSTF female in our experiments. I have added this description in the Materials and Methods section.
(5) Figure S1: The legend of magenta fluorescence is missing. Please add which protein is shown in magenta.
We have added the legend of magenta fluorescence, which is Trio.
(6) The detailed genotypes of Figure 2C and Figure S7 are missing in Supplementary Table 1. Please include that, so that readers can know the genetic background.
We have added genotypes of Figure 2I (previously Figure 2C) and Figure S8 (previously as Figure S7) in Supplementary Table 2.
(7) Figure 2D-G: It will be helpful if the authors can outline the lobe (γ, α'/β', and α/β) in the figure, which will help readers to understand the images.
We have outlined α, α', β, β' and γ lobes in Figure 2C-F (previously as Figure 2D-G).
-
-
-
Analyse d'une Rencontre sur le Harcèlement Scolaire
Résumé Exécutif
Ce document de synthèse analyse le dialogue entre un ancien harceleur, aujourd'hui sociologue, et une ancienne victime de harcèlement, étudiante infirmière de 22 ans.
Leur échange met en lumière les mécanismes complexes du harcèlement scolaire, explorant à la fois ses origines et ses conséquences durables.
Du côté du harceleur, l'acte est présenté comme une stratégie de survie sociale, une quête de statut et d'existence au sein de la hiérarchie de la cour de récréation, souvent initiée par la normalisation de comportements dégradants par des figures d'autorité (un professeur).
Malgré une conscience sous-jacente du mal infligé, des mécanismes de justification et le déni de l'entourage parental empêchent une remise en question immédiate.
Pour la victime, l'expérience est définie par la violence psychologique et l'isolement.
Ciblée pour sa "différence" (timidité, style), elle subit des agressions verbales quotidiennes qui engendrent une souffrance profonde, exacerbée par l'incompréhension de ses parents et l'absence de soutien de ses pairs.
Les répercussions sont profondes et durables, se manifestant par une phobie scolaire, une anxiété chronique, une peur persistante du jugement et une difficulté à demander de l'aide à l'âge adulte.
La rencontre souligne l'importance de la reconnaissance et du dialogue.
Elle conclut sur un double message : un appel à la reconstruction et à l'affirmation de soi pour les victimes, et une mise en garde pour les harceleurs sur l'inutilité et les conséquences négatives de leurs actes, en les encourageant à verbaliser leur propre souffrance.
Contexte de la Rencontre
La discussion rassemble deux individus aux parcours de vie marqués par le harcèlement scolaire, mais de côtés opposés du spectre.
• L'ancien harceleur : Un homme au parcours scolaire qualifié de "chaotique".
Bon élève en primaire, il a connu une "descente abyssale" au collège avant de se réorienter plusieurs fois (bac pro commerce, licence pro en communication, master en sciences politiques et sociologie). Il a commencé à harceler dès le CM2.
• L'ancienne victime : Une femme de 22 ans, étudiante en première année de soins infirmiers.
Passionnée de cinéma, elle se décrit comme une enfant "renfermée" et très émotive, ce qui a fait d'elle une cible. Le harcèlement à son encontre s'est intensifié en classe de 4ème.
La Genèse du Harcèlement : Perspective du Harceleur
L'analyse de son propre comportement révèle plusieurs facteurs déclencheurs et de maintien du harcèlement.
• L'influence d'une figure d'autorité : Il cite l'exemple d'un professeur de CM1 qui avait mis en place des "dispositifs d'humiliation" (surnoms dégradants, moqueries au tableau).
Ce comportement a été perçu comme une normalisation de la violence verbale, qu'il a ensuite reproduite.
• Une stratégie d'existence sociale : Le harcèlement est décrit comme "une manière d'exister, de trouver une place, de grimper dans la hiérarchie de la cour de récrée".
Il admet que ce comportement, ainsi que les bêtises en général, lui permettait d'obtenir une reconnaissance de ses pairs.
• La cible : Son premier acte de harcèlement en CM2 visait un élève qui "déviait des normes de genre".
Les brimades initiales portaient sur sa "féminité apparente" avant de s'intensifier jusqu'à la violence physique.
• La conscience de l'acte et l'auto-justification : Il affirme que "toute personne qui harcèle se rend compte de son acte".
Cependant, cette conscience est neutralisée par des justifications : "soit il a cherché, soit bah c'est lui ou moi".
Il explique qu'il ne prenait "pas particulièrement de plaisir" à le faire.
• L'inefficacité des sanctions initiales : Une plainte du directeur de son école primaire a mené à une enquête et une convocation par la police.
Si l'expérience a été "marquante", il admet qu'elle ne l'a "pas suffisamment marqué" pour qu'il arrête ses agissements.
• Le déni parental : Lorsque, plus tard, il a qualifié ses propres actions de "harcèlement" auprès de sa mère, celle-ci a nié la gravité des faits, parlant de "chamaillerie" et affirmant : "tu n'étais pas un garçon comme ça".
L'Expérience de la Victime : Violence et Isolement
Le témoignage de l'ancienne victime met en exergue la violence de l'expérience et le sentiment d'abandon qui l'accompagne.
• La "différence" comme déclencheur : Elle identifie sa nature "renfermée" et sa forte émotivité (pleurer lorsqu'elle était interrogée) comme les "fragilités" exploitées par les harceleurs.
• La nature des agressions : Le harcèlement a débuté en 4ème, initié par une élève populaire.
Les attaques étaient quotidiennes et verbales, portant sur son physique et son style vestimentaire, avec des insultes telles que "tu es un castor", "regarde le cheval", "tu es vraiment débile", "tu es une merde".
• L'incompréhension familiale : Lorsqu'elle en a parlé à ses parents, leur réaction a été de lui conseiller de s'adapter : "oui bah tu es un peu différent un peu bizarre, mets-toi un peu dans les normes et ça ira mieux".
Elle souligne l'absurdité de demander à une victime de changer.
• L'absence de soutien des pairs : Elle explique que ses "amis" de l'époque pratiquaient un "harcèlement passif" ou qu'elle n'en avait pas réellement.
Ce manque de soutien a été un facteur aggravant, car sa souffrance n'a pas été prise en compte.
Conséquences à Long Terme et Reconstruction
Les deux intervenants discutent des impacts durables du harcèlement sur leur vie respective.
• Pour la victime : un traumatisme durable
◦ Santé mentale : Elle a développé une phobie scolaire, des angoisses et une anxiété persistante.
Elle vivait dans la peur constante du lendemain au collège.
◦ Impacts sociaux : L'expérience l'a marquée durablement. Elle explique : "quand quelqu'un rigole dans la rue, pour moi il se moque de moi". Elle vit avec une "peur constante du jugement" et une incapacité à demander de l'aide.
◦ Orientation professionnelle : Son vécu a conditionné son choix de devenir infirmière, motivée par le désir "d'être utile dans la vie des gens" et de s'assurer que d'autres puissent être entendus.
◦ Reconstruction : Elle a transformé sa "différence" en force, appréciant aujourd'hui son style particulier.
• Pour le harceleur : une prise de conscience tardive
◦ La reconnaissance de la souffrance de l'autre : Il comprend que ses actes ont engendré un "traumatisme" qui est "resté ancré" chez la victime.
◦ L'analyse de ses propres motivations : Il théorise que le harcèlement provient également "d'une forme de souffrance", de questionnements et d'un sentiment de ne pas trouver sa place.
◦ L'importance de la remise en question : Il insiste sur la nécessité pour les harceleurs de "questionner ses comportements" et de reconnaître le mal causé, même si c'est une étape difficile.
Citations Clés
Le tableau suivant présente des citations marquantes qui illustrent les thèmes principaux de la discussion.
| Citation | Intervenant | Thème Illustré | | --- | --- | --- | | "J'ai harcelé parce que c'était une manière de d'exister, de trouver une place, de grimper dans la hiérarchie de la cour de récrée." | L'ancien harceleur | La motivation sociale du harcèlement | | "On m'a dit c'est 'oui bah tu es un peu différent un peu bizarre, mets-toi un peu dans les normes et ça ira mieux.'" | L'ancienne victime | L'incompréhension de l'entourage et la culpabilisation de la victime | | "Je pense que les personnes qui m'ont fait subir ça, je pense qu'elles ont oublié en fait. Elles se sont même pas rendu compte de la violence que encore maintenant ça m'impacte." | L'ancienne victime | L'asymétrie de l'impact et la persistance du traumatisme | | "Ma mère m'a dit 'bah non c'est de la chamaillerie, s'est pas passé comme ça, tu étais pas un garçon comme ça.'" | L'ancien harceleur | Le déni parental face au comportement de son enfant | | "Le harcèlement ça vient aussi d'une forme de souffrance \[...\] d'élèves qui trouvent pas leur place." | L'ancien harceleur | L'analyse de l'origine du comportement du harceleur | | "Tu peux te reconstruire après ça et tu peux avancer dans ta vie, tu peux faire quelque chose de ta vie." | L'ancienne victime | Le message d'espoir et de résilience | | "C'est pas cool de harceler. C'est pas stylé et ça va pas te rendre plus intéressant, ça va pas t'aider à avancer dans ta vie." | L'ancien harceleur | Le message de dissuasion adressé aux harceleurs potentiels |
-
-
www.biorxiv.org www.biorxiv.org
-
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public review):
Monziani and Ulitsky present a large and exhaustive study on the lncRNA EPB41L4A-AS1 using a variety of genomic methods. They uncover a rather complex picture of an RNA transcript that appears to act via diverse pathways to regulate the expression of large numbers of genes, including many snoRNAs. The activity of EPB41L4A-AS1 seems to be intimately linked with the protein SUB1, via both direct physical interactions and direct/indirect of SUB1 mRNA expression.
The study is characterised by thoughtful, innovative, integrative genomic analysis. It is shown that EPB41L4A-AS1 interacts with SUB1 protein and that this may lead to extensive changes in SUB1's other RNA partners. Disruption of EPB41L4A-AS1 leads to widespread changes in non-polyA RNA expression, as well as local cis changes. At the clinical level, it is possible that EPB41L4A-AS1 plays disease-relevant roles, although these seem to be somewhat contradictory with evidence supporting both oncogenic and tumour suppressive activities.
A couple of issues could be better addressed here. Firstly, the copy number of EPB41L4A-AS1 is an important missing piece of the puzzle. It is apparently highly expressed in the FISH experiments. To get an understanding of how EPB41L4A-AS1 regulates SUB1, an abundant protein, we need to know the relative stoichiometry of these two factors. Secondly, while many of the experiments use two independent Gapmers for EPB41L4A-AS1 knockdown, the RNA-sequencing experiments apparently use just one, with one negative control (?). Evidence is emerging that Gapmers produce extensive off-target gene expression effects in cells, potentially exceeding the amount of on-target changes arising through the intended target gene. Therefore, it is important to estimate this through the use of multiple targeting and non-targeting ASOs, if one is to get a true picture of EPB41L4A-AS1 target genes. In this Reviewer's opinion, this casts some doubt over the interpretation of RNA-seq experiments until that work is done. Nonetheless, the Authors have designed thorough experiments, including overexpression rescue constructs, to quite confidently assess the role of EPB41L4A-AS1 in snoRNA expression.
It is possible that EPB41L4A-AS1 plays roles in cancer, either as an oncogene or a tumour suppressor. However, it will in the future be important to extend these observations to a greater variety of cell contexts.
This work is valuable in providing an extensive and thorough analysis of the global mechanisms of an important regulatory lncRNA and highlights the complexity of such mechanisms via cis and trans regulation and extensive protein interactions.
Reviewer #2 (Public review):
Summary:
In this manuscript, Monziani et al. identified long noncoding RNAs (lncRNAs) that act in cis and are coregulated with their target genes located in close genomic proximity. The authors mined the GeneHancer database, and this analysis led to the identification of four lncRNA-target pairs. The authors decided to focus on lncRNA EPB41L4A-AS1.
They thoroughly characterised this lncRNA, demonstrating that it is located in the cytoplasm and the nuclei, and that its expression is altered in response to different stimuli. Furthermore, the authors showed that EPB41L4A-AS1 regulates EPB41L4A transcription, leading to a mild reduction in EPB41L4A protein levels. This was not recapitulated with siRNA-mediated depletion of EPB41L4AAS1. RNA-seq in EPB41L4A-AS1-depleted cells with single LNA revealed 2364 DEGs linked to pathways including the cell cycle, cell adhesion, and inflammatory response. To understand the mechanism of action of EPB41L4A-AS1, the authors mined the ENCODE eCLIP data and identified SUB1 as an lncRNA interactor. The authors also found that the loss of EPB41L4A-AS1 and SUB1 leads to the accumulation of snoRNAs, and that SUB1 localisation changes upon the loss of EPB41L4A-AS1. Finally, the authors showed that EPB41L4A-AS1 deficiency did not change the steady-state levels of SNORA13 nor RNA modification driven by this RNA. The phenotype associated with the loss of EPB41L4A-AS1 is linked to increased invasion and EMT gene signature.
Overall, this is an interesting and nicely done study on the versatile role of EPB41L4A-AS1 and the multifaceted interplay between SUB1 and this lncRNA, but some conclusions and claims need to be supported with additional experiments. My primary concerns are using a single LNA gapmer for critical experiments, increased invasion, and nucleolar distribution of SUB1- in EPB41L4A-AS1-depleted cells. These experiments need to be validated with orthogonal methods.
Strengths:
The authors used complementary tools to dissect the complex role of lncRNA EPB41L4A-AS1 in regulating EPB41L4A, which is highly commendable. There are few papers in the literature on lncRNAs at this standard. They employed LNA gapmers, siRNAs, CRISPRi/a, and exogenous overexpression of EPB41L4A-AS1 to demonstrate that the transcription of EPB41L4A-AS1 acts in cis to promote the expression of EPB41L4A by ensuring spatial proximity between the TAD boundary and the EPB41L4A promoter. At the same time, this lncRNA binds to SUB1 and regulates snoRNA expression and nucleolar biology. Overall, the manuscript is easy to read, and the figures are well presented. The methods are sound, and the expected standards are met.
Weaknesses:
The authors should clarify how many lncRNA-target pairs were included in the initial computational screen for cis-acting lncRNAs and why MCF7 was chosen as the cell line of choice. Most of the data uses a single LNA gapmer targeting EPB41L4A-AS1 lncRNA (eg, Fig. 2c, 3B, and RNA-seq), and the critical experiments should be using at least 2 LNA gapmers. The specificity of SUB1 CUT&RUN is lacking, as well as direct binding of SUB1 to lncRNA EPB41L4A-AS1, which should be confirmed by CLIP qPCR in MCF7 cells. Finally, the role of EPB41L4A-AS1 in SUB1 distribution (Figure 5) and cell invasion (Figure 8) needs to be complemented with additional experiments, which should finally demonstrate the role of this lncRNA in nucleolus and cancer-associated pathways. The use of MCF7 as a single cancer cell line is not ideal.
Reviewer #3 (Public review):
Summary:
In this paper, the authors made some interesting observations that EPB41L4A-AS1 lncRNA can regulate the transcription of both the nearby coding gene and genes on other chromosomes. They started by computationally examining lncRNA-gene pairs by analyzing co-expression, chromatin features of enhancers, TF binding, HiC connectome, and eQTLs. They then zoomed in on four pairs of lncRNA-gene pairs and used LNA antisense oligonucleotides to knock down these lncRNAs. This revealed EPB41L4A-AS1 as the only one that can regulate the expression of its cis-gene target EPB41L4A. By RNA-FISH, the authors found this lncRNA to be located in all three parts of a cell: chromatin, nucleoplasm, and cytoplasm. RNA-seq after LNA knockdown of EPB41L4A-AS1 showed that this increased >1100 genes and decreased >1250 genes, including both nearby genes and genes on other chromosomes. They later found that EPB41L4A-AS1 may interact with SUB1 protein (an RNA-binding protein) to impact the target genes of SUB1. EPB41L4A-AS1 knockdown reduced the mRNA level of SUB1 and altered the nuclear location of SUB1. Later, the authors observed that EPB41L4A-AS1 knockdown caused an increase of snRNAs and snoRNAs, likely via disrupted SUB1 function. In the last part of the paper, the authors conducted rescue experiments that suggested that the full-length, intron- and SNORA13-containing EPB41L4A-AS1 is required to partially rescue snoRNA expression. They also conducted SLAM-Seq and showed that the increased abundance of snoRNAs is primarily due to their hosts' increased transcription and stability. They end with data showing that EPB41L4A-AS1 knockdown reduced MCF7 cell proliferation but increased its migration, suggesting a link to breast cancer progression and/or metastasis.
Strengths:
Overall, the paper is well-written, and the results are presented with good technical rigor and appropriate interpretation. The observation that a complex lncRNA EPB41L4A-AS1 regulates both cis and trans target genes, if fully proven, is interesting and important.
Weaknesses:
The paper is a bit disjointed as it started from cis and trans gene regulation, but later it switched to a partially relevant topic of snoRNA metabolism via SUB1. The paper did not follow up on the interesting observation that there are many potential trans target genes affected by EPB41L4A-AS1 knockdown and there was limited study of the mechanisms as to how these trans genes (including SUB1 or NPM1 genes themselves) are affected by EPB41L4A-AS1 knockdown. There are discrepancies in the results upon EPB41L4A-AS1 knockdown by LNA versus by CRISPR activation, or by plasmid overexpression of this lncRNA.
Recommendations for the authors:
Reviewer #1 (Recommendations for the authors):
(1) Copy number:
Perhaps I missed it, but it seems that no attempt is made to estimate the number of copies of EPB41L4A-AS1 transcripts per cell. This should be possible given RNAseq and FISH. At least an order of magnitude estimate. This is important for shedding light on the later observations that EPB41L4A-AS1 may interact with SUB1 protein and regulate the expression of thousands of mRNAs.
We thank the reviewer for the insightful suggestion. We agree that an estimate of EPB41L4A-AS1 copy number might further strengthen the hypotheses presented in the manuscript. Therefore, we analyzed the smFISH images and calculated the copy number per cell of this lncRNA, as well as that of GAPDH as a comparison.
Because segmenting MCF-7 cells proved to be difficult due to the extent of the cell-cell contacts they establish, we imaged multiple (n = 14) fields of view, extracted the number of EPB41L4A-AS1/GAPDH molecules in each field and divided them by the number of cells (as assessed by DAPI staining, 589 cells in total). We detected an average of 33.37 ± 3.95 EPB41L4A-AS1 molecules per cell, in contrast to 418.27 ± 61.79 GAPDH molecules. As a comparison, within the same qPCR experiment the average of the Ct values of these two RNAs is about 22.3 and 17.5, the FPKMs in the polyA+ RNA-seq are ~ 2479.4 and 35.6, and the FPKMs in the rRNA-depleted RNA-seq are ~ 3549.9 and 19.3, respectively. Thus, our estimates of the EPB41L4A-AS1 copy number in MCF-7 cells fits well into these observations.
The question whether an average of ~35 molecules per cell is sufficient to affect the expression of thousands of genes is somewhat more difficult to ascertain. As discussed below, it is unlikely that all the genes dysregulated following the KD of EPB41L4A-AS1 are all direct targets of this lncRNA, and indeed SUB1 depletion affects an order of magnitude fewer genes. It has been shown that lncRNAs can affect the behavior of interacting RNAs and proteins in a substoichiometric fashion (Unfried & Ulitsky, 2022), but whether this applies to EPB41L4A-AS1 remains to be addressed in future studies. Nonetheless, this copy number appears to be sufficient for a trans-acting functions for this lncRNA, on top of its cis-regulatory role in regulating EPB41L4A. We added this information in the text as follows:
“Using single-molecule fluorescence in-situ hybridization (smFISH) and subcellular fractionation we found that EPB41L4A-AS1 is expressed at an average of 33.37 ± 3.95 molecule per cell, and displays both nuclear and cytoplasmic localization in MCF-7 cells (Fig. 1D), with a minor fraction associated with chromatin as well (Fig. 1E).”
We have updated the methods section as well:
“To visualize the subcellular localization of EPB41L4A-AS1 in vivo, we performed single-molecule fluorescence in situ hybridization (smFISH) using HCR™ amplifiers. Probe sets (n = 30 unique probes) targeting EPB41L4A-AS1 and GAPDH (positive control) were designed and ordered from Molecular Instruments. We followed the Multiplexed HCR v3.0 protocol with minor modifications. MCF-7 cells were plated in 8-well chambers (Ibidi) and cultured O/N as described above. The next day, cells were fixed with cold 4% PFA in 1X PBS for 10 minutes at RT and then permeabilized O/N in 70% ethanol at -20°C. Following permeabilization, cells were washed twice with 2X SSC buffer and incubated at 37°C for 30 minutes in hybridization buffer (HB). The HB was then replaced with a probe solution containing 1.2 pmol of EPB41L4A-AS1 probes and 0.6 pmol of GAPDH probes in HB. The slides were incubated O/N at 37°C. To remove excess probes, the slides were washed four times with probe wash buffer at 37°C for 5 minutes each, followed by two washes with 5X SSCT at RT for 5 minutes. The samples were then pre-amplified in amplification buffer for 30 minutes at RT and subsequently incubated O/N in the dark at RT in amplification buffer supplemented with 18 pmol of the appropriate hairpins. Finally, excess hairpins were removed by washing the slides five times in 5X SSCT at RT. The slides were mounted with ProLong™ Glass Antifade Mountant (Invitrogen), cured O/N in the dark at RT, and imaged using a Nikon CSU-W1 spinning disk confocal microscope. In order to estimate the RNA copy number, we imaged multiple distinct fields, extracted the number of EPB41L4A-AS1/GAPDH molecules in each field using the “Find Maxima” tool in ImageJ/Fiji, and divided them by the number of cells (as assessed by DAPI staining).”
(2) Gapmer results:
Again, it is quite unclear how many and which Gapmer is used in the genomics experiments, particularly the RNA-seq. In our recent experiments, we find very extensive off-target mRNA changes arising from Gapmer treatment. For this reason, it is advisable to use both multiple control and multiple targeting Gapmers, so as to identify truly target-dependent expression changes. While I acknowledge and commend the latter rescue experiments, and experiments using multiple Gapmers, I'd like to get clarification about how many and which Gapmers were used for RNAseq, and the authors' opinion on the need for additional work here.
We agree with the Reviewer that GapmeRs are prone to off-target and unwanted effects (Lai et al., 2020; Lee & Mendell, 2020; Maranon & Wilusz, 2020). Early in our experiments, we found out that LNA1 triggers a non-specific CDKN1A/p21 activation (Fig. S5A-C), and thus, we have initially performed some experiments such as RNA-seq with only LNA2.
Nonetheless, other experiments were performed using both GapmeRs, such as multiple RT-qPCRs, UMI-4C, SUB1 and NPM1 imaging, and the in vitro assays, among others, and consistent results were obtained with both LNAs.
To accommodate the request by this and the other reviewers, we have now performed another round of polyA+ RNA-seq following EPB41L4A-AS1 knockdown using LNA1 or LNA2, as well as the previously used and an additional control GapmeR. The FPKMs of the control samples are highly-correlated both within replicates and between GapmeRs (Fig. S6A). More importantly, the fold-changes to control are highly correlated between the two on-target GapmeRs LNA1 and LNA2, regardless of the GapmeR used for normalization (Fig. S6B), thus showing that the bulk of the response is shared and likely the direct result of the reduction in the levels of EPB41L4A-AS1. Notably, key targets NPM1 and MTREX (see discussion, Fig. S12A-C and comments to Reviewer 3) were found to be downregulated by both LNAs (Fig. S6C).
However, we acknowledge that some of the dysregulated genes are observed only when using one GapmeR and not the other, likely due to a combination of indirect, secondary and non-specific effects, and as such it is difficult to infer the direct response. Supporting this, LNA2 yielded a total of 1,069 DEGs (617 up and 452 down) and LNA1 2,493 DEGs (1,328 up and 1,287 down), with the latter triggering a stronger response most likely as a result of the previously mentioned CDKN1A/p21 induction. Overall, 45.1% of the upregulated genes following LNA2 transfection were shared with LNA1, in contrast to only the 24.3% of the downregulated ones.
We have now included these results in the Results section (see below) and in Supplementary Figure (Fig. S6).
“Most of the consequences of the depletion of EPB41L4A-AS1 are thus not directly explained by changes in EPB41L4A levels. An additional trans-acting function for EPB41L4A-AS1 would therefore be consistent with its high expression levels compared to most lncRNAs detected in MCF-7 (Fig. S5G). To strengthen these findings, we have transfected MCF-7 cells with LNA1 and a second control GapmeR (NT2), as well as the previous one (NT1) and LNA2, and sequenced the polyadenylated RNA fraction as before. Notably, the expression levels (in FPKMs) of the replicates of both control samples are highly correlated with each other (Fig. S6A), and the global transcriptomic changes triggered by the two EPB41L4A-AS1-targeting LNAs are largely concordant (Fig. S6B and S6C). Because of this concordance and the cleaner (i.e., no CDKN1A upregulation) readout in LNA2-transfected cells, we focused mainly on these cells for subsequent analyses.”
(3) Figure 1E:
Can the authors comment on the unusual (for a protein-coding mRNA) localisation of EPB41L4A, with a high degree of chromatin enrichment?
We acknowledge that mRNAs from protein-coding genes displaying nuclear and chromatin localizations are quite unusual. The nuclear and chromatin localization of some mRNAs are often due to their low expression, length, time that it takes to be transcribed, repetitive elements and strong secondary structures (Bahar Halpern et al., 2015; Didiot et al., 2018; Lubelsky & Ulitsky, 2018; Ly et al., 2022).
We now briefly mention this in the text:
“In contrast, both EPB41L4A and SNORA13 were mostly found in the chromatin fraction (Fig. 1E), the former possibly due to the length of its pre-mRNA (>250 kb), which would require substantial time to transcribe (Bahar Halpern et al., 2015; Didiot et al., 2018; Lubelsky & Ulitsky, 2018; Ly et al., 2022).”
Supporting our results, analysis of the ENCODE MCF-7 RNA-seq data of the cytoplasmic, nuclear and total cell fractions indeed shows a nuclear enrichment of the EPB41L4A mRNA (Author response image 1), in line with what we observed in Fig. 1E by RT-qPCR.
Author response image 1.
The EPB41L4A transcript is nuclear-enriched in the MCF-7 ENCODE subcellular RNA-seq dataset. Scatterplot of gene length versus cytoplasm/nucleus ratio (as computed by DESeq2) in MCF-7 cells. Each dot represents an unique gene, color-coded reflecting if their DESeq2 adjusted p-value < 0.05 and absolute log<sub>2</sub>FC > .41 (33% enrichment or depletion).GAPDH and MALAT1 are shown as representative cytoplasmic and nuclear transcripts, respectively. Data from ENCODE.
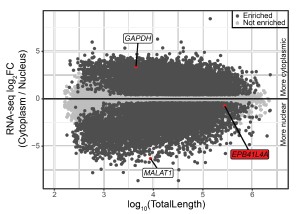
(4) Annotation and termini of EPB41L4A-AS1:
The latest Gencode v47 annotations imply an overlap of the sense and antisense, different from that shown in Figure 1C. The 3' UTR of EPB41L4A is shown to extensively overlap EPB41L4A-AS1. This could shed light on the apparent regulation of the former by the latter that is relevant for this paper. I'd suggest that the authors update their figure of the EPB41L4A-AS1 locus organisation with much more detail, particularly evidence for the true polyA site of both genes. What is more, the authors might consider performing RACE experiments for both RNAs in their cells to definitely establish whether these transcripts contain complementary sequence that could cause their Watson-Crick hybridisation, or whether their two genes might interfere with each other via some kind of polymerase collision.
We thank the reviewer for pointing this out. Also in previous GENCODE annotations, multiple isoforms were reported with some overlapping the 3’ UTR of EPB41L4A. In the EPB41L4A-AS1 locus image (Fig. 1C), we report at the bottom the different transcripts isoforms currently annotated, and a schematics of the one that is clearly the most abundant in MCF-7 cells based on RNA-seq read coverage. This is supported by both the polyA(+) and ribo(-) RNA-seq data, which are strand-specific, as shown in the figure.
We now also examined the ENCODE/CSHL MCF-7 RNA-seq data from whole cell, cytoplasm and nucleus fractions, as well as 3P-seq data (Jan et al., 2011) (unpublished data from human cell lines), reported in Author response image 2. All these data support the predominant use of the proximal polyA site in human cell lines. This shorter isoform does not overlap EPB41L4A.
Author response image 2.
Most EPB41L4A-AS1 transcripts end before the 3’ end of EPB41L4A. UCSC genome browser view showing tracks from 3P-seq data in different cell lines and neural crest (top, with numbers representing the read counts, i.e. how many times that 3’ end has been detected), and stranded ENCODE subcellular RNA-seq (bottom).
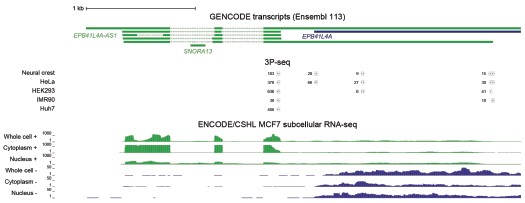
Based on these data, the large majority of cellular transcripts of EPB41L4A-AS1 terminate at the earlier polyA site and don’t overlap with EPB41L4A. There is a small fraction that appears to be restricted to the nucleus that terminates later at the annotated isoform. 3' RACE experiments are not expected to provide substantially different information beyond what is already available.
(5) Figure 3C:
There is an apparent correlation between log2FC upon EPB41L4A-AS1 knockdown, and the number of clip sites for SUB1. However, I expect that the clip signal correlates strongly with the mRNA expression level, and that log2FC may also correlate with the same. Therefore, the authors would be advised to more exhaustively check that there really is a genuine relationship between log2FC and clip sites, after removing any possible confounders of overall expression level.
As the reviewer suggested, there is a correlation between the baseline expression level and the strength of SUB1 binding in the eCLIP data. To address this issue, we built expression-matched controls for each group of SUB1 interactors and checked the fold-changes following EPB41L4A-AS1 KD, similarly to what we have done in Fig. 3C. The results are presented, and are now part of Supplementary Figure 7 (Fig. S7C).
Based on this analysis, while there is a tendency of increased expression with increased SUB1 binding, when controlling for expression levels the effect of down-regulation of SUB1-bound RNAs upon lncRNA knockdown remains, suggesting that it is not merely a confounding effect. We have updated the text as follows:
“We hypothesized that loss of EPB41L4A-AS1 might affect SUB1, either via the reduction in its expression or by affecting its functions. We stratified SUB1 eCLIP targets into confidence intervals, based on the number, strength and confidence of the reported binding sites. Indeed, eCLIP targets of SUB1 (from HepG2 cells profiled by ENCODE) were significantly downregulated following EPB41L4A-AS1 KD in MCF-7, with more confident targets experiencing stronger downregulation (Fig. 3C). Importantly, this still holds true when controlling for gene expression levels (Fig. S7C), suggesting that this negative trend is not due to differences in their baseline expression.”
(6) The relation to cancer seems somewhat contradictory, maybe I'm missing something. Could the authors more clearly state which evidence is consistent with either an Oncogene or a Tumour Suppressive function, and discuss this briefly in the Discussion? It is not a problem if the data are contradictory, however, it should be discussed more clearly.
We acknowledge this apparent contradiction. Cancer cells are characterized by a multitude of hallmarks depending on the cancer type and stage, including high proliferation rates and enhanced invasive capabilities. The notion that cells with reduced EPB41L4A-AS1 levels exhibit lower proliferation, yet increased invasion is compatible with a function as an oncogene. Cells undergoing EMT may reduce or even completely halt proliferation/cell division, until they revert back to an epithelial state (Brabletz et al., 2018; Dongre & Weinberg, 2019). Notably, downregulated genes following EPB41L4A-AS1 KD are enriched in GO terms related to cell proliferation and cell cycle progression (Fig. 2I), whereas those upregulated are enriched for terms linked to EMT processes. Thus, while we cannot rule out a potential function as tumor suppressor gene, our data fit better the notion that EPB41L4A-AS1 promotes invasion, and thus, primarily functions as an oncogene. We now address this in point in the discussion:
“The notion that cells with reduced EPB41L4A-AS1 levels exhibit lower proliferation (Fig. 8C), yet increased invasion (Fig. 8A and 8B) is compatible with a function as an oncogene by promoting EMT (Fig. 8D and 8E). Cells undergoing this process may reduce or even completely halt proliferation/cell division, until they revert back to an epithelial state (Brabletz et al., 2018; Dongre & Weinberg, 2019). Notably, downregulated genes following EPB41L4A-AS1 KD are enriched in GO terms related to cell proliferation and cell cycle progression (Fig. 2I), whereas those upregulated for terms linked to EMT processes. Thus, while we cannot rule out a potential function as tumor suppressor gene, our data better fits the idea that this lncRNA promotes invasion, and thus, primarily functions as an oncogene.”
Reviewer #2 (Recommendations for the authors):
Below are major and minor points to be addressed. We hope the authors find them useful.
(1) Figure 1:
Where are LNA gapmers located within the EPB41L4A-AS1 gene? Are they targeting exons or introns of the EPB41L4A-AS1? Please clarify or include in the figure.
We now report the location of the two GapmeRs in Fig. 1C. LNA1 targets the intronic region between SNORA13 and exon 2, and LNA2 the terminal part of exon 1.
(2) Figure 2B:
Why is a single LNA gapmer used for EPB41L4A Western? In addition, are the qPCR data in Figure 2B the same as in Figure 1B? Please clarify.
The Western Blot was performed after transfecting the cells with either LNA1 or LNA2. We now have replaced Fig. 2C with the full Western Blot image, in order to show both LNAs. With respect to the qPCRs in Fig. 1B and 2B, they represent the results from two independent experiments.
(3) Figure 2F:
2364 DEGs for a single LNA is a lot of deregulated genes in RNA-seq data. How do the authors explain such a big number in DEGs? Is that because this LNA was intronic? Additional LNA gapmer would minimise the "real" lncRNA target and any potential off-target effect.
We agree with the Reviewer that GapmeRs are prone to off-target and unwanted effects (Lai et al.,2020; Lee & Mendell, 2020; Maranon & Wilusz, 2020). Early in our experiments, we found out that LNA1 triggers a non-specific CDKN1A/p21 activation (Fig. S5A-C), and thus, we have initially performed some experiments such as RNA-seq with only LNA2.
Nonetheless, other experiments were performed using both GapmeRs, such as multiple RT-qPCRs, UMI-4C, SUB1 and NPM1 imaging, and the in vitro assays, among others, and consistent results were obtained with both LNAs.
To accommodate the request by this and the other reviewers, we have now performed another round of polyA+ RNA-seq following EPB41L4A-AS1 knockdown using LNA1 or LNA2, as well as the previously used and an additional control GapmeR. The FPKMs of the control samples are highly-correlated both within replicates and between GapmeRs (Fig. S6A). More importantly, the fold-changes to control are highly correlated between the two on-target GapmeRs LNA1 and LNA2, regardless of the GapmeR used for normalization (Fig. S6B), thus showing that despite significant GapmeR-specific effects, the bulk of the response is shared and likely the direct result of the reduction in the levels of EPB41L4A-AS1. Notably, key targets NPM1 and MTREX (see discussion, Fig. S12A-C and comments to Reviewer 3) were found to be downregulated by both LNAs (Fig. S6C).
However, we acknowledge that some of the dysregulated genes are observed only when using one GapmeR and not the other, likely due to a combination of indirect, secondary and non-specific effects, and as such it is difficult to infer the direct response. Supporting this, LNA2 yielded a total of 1,069 DEGs (617 up and 452 down) and LNA1 2,493 DEGs (1,328 up and 1,287 down), with the latter triggering a stronger response most likely as a result of the previously mentioned CDKN1A/p21 induction. Overall, 45.1% of the upregulated genes following LNA2 transfection were shared with LNA1, in contrast to only the 24.3% of the downregulated ones.
We have now included these results in the Results section (see below) and in Supplementary Figure (Fig. S6).
“Most of the consequences of the depletion of EPB41L4A-AS1 are thus not directly explained by changes in EPB41L4A levels. An additional trans-acting function for EPB41L4A-AS1 would therefore be consistent with its high expression levels compared to most lncRNAs detected in MCF-7 (Fig. S5G). To strengthen these findings, we have transfected MCF-7 cells with LNA1 and a second control GapmeR (NT2), as well as the previous one (NT1) and LNA2, and sequenced the polyadenylated RNA fraction as before. Notably, the expression levels (in FPKMs) of the replicates of both control samples are highly correlated with each other (Fig. S6A), and the global transcriptomic changes triggered by the two EPB41L4A-AS1-targeting LNAs are largely concordant (Fig. S6B and S6C). Because of this concordance and the cleaner (i.e., no CDKN1A upregulation) readout in LNA2-transfected cells, we focused mainly on these cells for subsequent analyses.”
(4) Figure 3B: Does downregulation of SUB1 and NPM1 reflect at the protein level with both LNA gapmers? The authors should show a heatmap and metagene profile for SUB1 CUT & RUN. How did the author know that SUB1 binding is specific, since CUT & RUN was not performed in SUB1-depleted cells?
As requested by both Reviewer #2 and #3, we have performed WB for SUB1, NPM1 and FBL following EPB41L4A-AS1 KD with two targeting (LNA1 and LNA2) and the previous control GapmeRs. Interestingly, we did not detect any significant downregulation of either proteins (Author response image 3), although this might be the result of the high variability observed in the control samples. Moreover, the short timeframe in which the experiments have been conducted━that is, transient transfections for 3 days━might not be sufficient time for the existing proteins to be degraded, and thus, the downregulation is more evident at the RNA (Fig. 3B and Supplementary Figure 6C) rather than protein level.
Author response image 3.
EPB41L4A-AS1 KD has only marginal effects on the levels of nucleolar proteins. (A) Western Blots for the indicated proteins after the transfection for 3 days of the control and targeting GapmeRs. (B) Quantification of the protein levels from (A). All experiments were performed in n=3 biological replicates, with the error bars in the barplots representing the standard deviation. ns - P>0.05; * - P<0.05; ** - P<0.01; *** - P<0.001 (two-sided Student’s t-test).
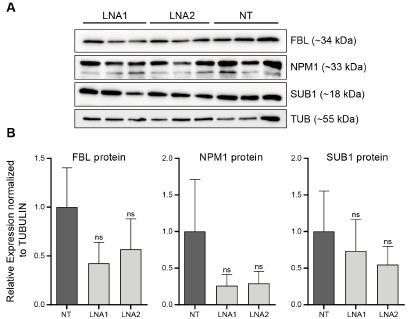
Following the suggestion by the Reviewer, we now show both the SUB1 CUT&RUN metagene profile (previously available as Fig. 3F) and the heatmap (now Fig. 3G) around the TSS of all genes, stratified by their expression level. Both graphs are reported.
We show that the antibody signal is responsive to SUB1 depletion via siRNAs in both WB (Fig. S8F) and IF (Fig. 5E) experiments. As mentioned below, this and the absence of non-specific signals makes us confident in the CUT&RUN data. Performing CUT&RUN in SUB1 depleted cells would be difficult to interpret as perturbations are typically not complete, and so the remaining protein can still bind the same regions. Since there isn’t a clear way to add spike-ins to CUT&RUN experiments, it is very difficult to show specificity of binding by CUT&RUN in siRNA-knockdown cells.
(5) Figure 3D: The MW for the depicted proteins are lacking. Why is there no SUB1 protein in the input? Please clarify. Since the authors used siRNA to deplete SUB1, it would be good to know if the antibody is specific in their CUT & RUN (see above)
We apologize for the lack of the MW in Fig. 3D. As shown in Fig. S8F, SUB1 is ~18 kDa and the antibody signal is responsive to SUB1 depletion via siRNAs in both WB (Fig. S8F) and IF (Fig. 5E) experiments. Thus, given its 1) established specificity in those two settings and 2) the lack of generalized signal at most open chromatin regions, which is typical of nonspecific CUT&RUN experiments, we are confident in the specificity of the CUT&RUN results.
We now mention the MW of SUB1 in Fig. 3D as well and we provide in Author response image 4 the full SUB1 WB picture, enhancing the contrast to highlight the bands. We agree that the SUB1 band in the input is weak, likely reflecting the low abundance in that fraction and the detection difficulty due to its low MW (see Fig. S8F).
Author response image 4.
Western blot for SUB1 following RIP using either a SUB1 or IgG antibody. IN - input, SN - supernatant/unbound, B - bound.
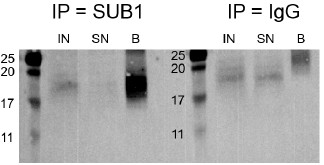
(6) Supplementary Figure 6C:
The validation of lncRNA EPB41L4A-AS1 binding to SUB1 should be confirmed by CLIP qPCR, since native RIP can lead to reassociation of RNA-protein interactions (PMID: 15388877). Additionally, the eclip data presented in Figure 3a were from a different cell line and not MCF7.
We acknowledge that the SUB1 eCLIP data was generated in a different cell line, as we mentioned in the text:
“Indeed, eCLIP targets of SUB1 (from HepG2 cells profiled by ENCODE) were significantly downregulated following EPB41L4A-AS1 KD in MCF-7, with more confident targets experiencing stronger downregulation (Fig. 3C). Importantly, this still holds true when controlling for gene expression levels (Fig. S7C), suggesting that this negative trend is not due to differences in their baseline expression. To obtain SUB1-associated transcripts in MCF-7 cells; we performed a native RNA immunoprecipitation followed by sequencing of polyA+ RNAs (RIP-seq) (Fig. 3D, S7D and S7E).”
Because of this, we resorted to native RIP, in order to get binding information in our experimental system. As we show independent evidence for binding using both eCLIP and RIP, and the substantial challenge in establishing the CLIP method, which has not been successfully used in our group, we respectfully argue that further validations are out of scope of this study. We nonetheless agree that several genes which are nominally significantly enriched in our RIP data are likely not direct targets of SUB1, especially given that it is difficult to assign the perfect threshold that discriminates between bound and unbound RNAs.
We now additionally mention this at the beginning of the paragraph as well:
“In order to identify potential factors that might be associated with EPB41L4A-AS1, we inspected protein-RNA binding data from the ENCODE eCLIP dataset(Van Nostrand et al., 2020). The exons of the EPB41L4A-AS1 lncRNA were densely and strongly bound by SUB1 (also known as PC4) in both HepG2 and K562 cells (Fig. 3A).”
(7) Figure 3G:
Can the authors distinguish whether loss of EPB41L4A-AS1 affects SUB1 chromatin binding or its activity as RBP? Please discuss.
Distinguishing between altered SUB1 chromatin and RNA binding is challenging, as this protein likely does not interact directly with chromatin and exhibits rather promiscuous RNA binding properties (Ray et al., 2023). In particular, SUB1 (also known as PC4) interacts with and regulates the activity of all three RNA polymerases, and was reported to be involved in transcription initiation and elongation, response to DNA damage, chromatin condensation (Conesa & Acker, 2010; Das et al., 2006; Garavís & Calvo, 2017; Hou et al., 2022) and telomere maintenance (Dubois et al., 2025; Salgado et al., 2024).
Based on our data, genes whose promoters are occupied by SUB1 display marginal, yet highly significant changes in their steady-state expression levels upon lncRNA perturbations. We also show that upon EPB41L4A-AS1 KD, SUB1 acquires a stronger nucleolar localization (Fig. 5A), which likely affects its RNA interactome as well. However, further elucidating these activities would require performing RIP-seq and CUT&RUN in lncRNA-depleted cells, which we argue is out of the scope of the current study. We note that KD of SUB1 with siRNAs have milder effects than that of EPB41L4A-AS1 (Fig. S8G), suggesting that additional players and effects shape the observed changes. Therefore, it is highly likely that the loss of this lncRNA affects both SUB1 chromatin binding profile and RNA binding activity, with the latter likely resulting in the increased snoRNAs abundance.
(8) Figure 4: Can the authors show that a specific class of snorna is affected upon depletion of SUB1 and EPB41L4A-AS1? Can they further classify the effect of their depletion on H/ACA box snoRNAs, C/D box snoRNAs, and scaRNAs?
Such potential distinct effect on the different classes of snoRNAs was considered, and the results are available in Fig. S8B and S8H (boxplots, after EPB41L4A-AS1 and SUB1 depletion), as well as Fig. 4F and S9F (scatterplots between EPB41L4A-AS1 and SUB1 depletion, and EPB41L4A-AS1 and GAS5 depletion, respectively). We see no preferential effect on one group of snoRNAs or the other.
(9) Figure 5: From the representative images, it looks to me that LNA 2 targeting EPB41L4A-AS1 has a bigger effect on nucleolar staining of SUB1. To claim that EPB41L4A-AS1 depletion "shifts SUB1 to a stronger nucleolar distribution", the authors need to perform IF staining for SUB1 and Fibrillarin, a known nucleolar marker. Also, how does this data fit with their qPCR data shown in Figure 3B? It is instrumental for the authors to demonstrate by IF or Western blotting that SUB1 levels decrease in one fraction and increase specifically in the nucleolus. They could perform Western blot for SUB1 and Fibrillarin in EPB41L4A-AS1-depleted cells and isolate cytoplasmic, nuclear, and nucleolar fractions.This experiment will strengthen their finding. The scale bar is missing for all the images in Figure 5. The authors should also show magnified images of a single representative cell at 100x.
We apologize for the confusion regarding the scale bars. As mentioned here and elsewhere, the scale bars are present in the top-left image of each panel only, in order to avoid overcrowding the panel. All the images are already at 100X, with the exception of Fig. 5E (IF for SUB1 upon siSUB1 transfection) which is 60X in order to better show the lack of signal. We however acknowledge that the images are sometimes confusing, due to the PNG features once imported into the document. In any case, in the submission we have also provided the original images in high-quality PDF and .ai formats. The suggested experiment would require establishing a nucleolar fractionation protocol which we currently don’t have available and we argue that it is out of scope of the current study.
(10) Additionally, is rRNA synthesis affected in SUB1- and EPB41L4A-AS1-depleted cells? The authors could quantify newly synthesised rRNA levels in the nucleoli, which would also strengthen their findings about the role of this lncRNA in nucleolar biology.
We acknowledge that there are many aspects of the role of EPB41L4A-AS1 in nucleolar biology that remain to be explored, as well as in nucleolar biology itself, but given the extensive experimental data we already provide in this and other subjects, we respectfully suggest that this experiment is out of scope of the current work. We note that a recent study has shown that SUB1 is required for Pol I-mediated rDNA transcription in the nucleolus (Kaypee et al., 2025). In the presence of nucleolar SUB1, rDNA transcription proceeds as expected, but when SUB1 is depleted or its nucleolar localization is affected—by either sodium butyrate treatment or inhibition of KAT5-mediated phosphorylation at its lysine 35 (K35)—the levels of the 47S pre-rRNA are significantly reduced. In our settings, SUB1 enriches into the nucleolus following EPB41L4A-AS1 KD; thus, we might expect to see a slightly increased rDNA transcription or no effect at all, given that SUB1 localizes in the nucleolus in baseline conditions as well. We now mention this novel role of SUB1 both in the results and discussion.
“SUB1 interacts with all three RNA polymerases and was reported to be involved in transcription initiation and elongation, response to DNA damage, chromatin condensation(Conesa & Acker, 2010; Das et al., 2006; Garavís & Calvo, 2017; Hou et al., 2022), telomere maintenance(Dubois et al., 2025; Salgado et al., 2024) and rDNA transcription(Kaypee et al., 2025). SUB1 normally localizes throughout the nucleus in various cell lines, yet staining experiments show a moderate enrichment for the nucleolus (source: Human Protein Atlas; https://www.proteinatlas.org/ENSG00000113387-SUB1/subcellular)(Kaypee et al., 2025).”
“Several features of the response to EPB41L4A-AS1 resemble nucleolar stress, including altered distribution of NPM1(Potapova et al., 2023; Yang et al., 2016). SUB1 was shown to be involved in many nuclear processes, including transcription(Conesa & Acker, 2010), DNA damage response(Mortusewicz et al., 2008; Yu et al., 2016), telomere maintenance(Dubois et al., 2025), and nucleolar processes including rRNA biogenesis(Kaypee et al., 2025; Tafforeau et al., 2013). Our results suggest a complex and multi-faceted relationship between EPB41L4A-AS1 and SUB1, as SUB1 mRNA levels are reduced by the transient (72 hours) KD of the lncRNA (Fig. 3B), the distribution of the protein in the nucleus is altered (Fig. 5A and 5C), while the protein itself is the most prominent binder of the mature EPB41L4A-AS1 in ENCODE eCLIP data (Fig. 3A). The most striking connection between EPB41L4A-AS1 and SUB1 is the similar phenotype triggered by their loss (Fig. 4). We note that a recent study has shown that SUB1 is required for Pol I-mediated rDNA transcription in the nucleolus(Kaypee et al., 2025). In the presence of nucleolar SUB1, rDNA transcription proceeds as expected, but when SUB1 is depleted or its nucleolar localization is affected—by either sodium butyrate treatment or inhibition of KAT5-mediated phosphorylation at its lysine 35 (K35)—the levels of the 47S pre-rRNA are significantly reduced. In our settings, SUB1 enriches into the nucleolus following EPB41L4A-AS1 KD; thus, we might expect to see a slightly increased rDNA transcription or no effect at all, given that SUB1 localizes in the nucleolus in baseline conditions as well. It is however difficult to determine which of the connections between these two genes is the most functionally relevant and which may be indirect and/or feedback interactions. For example, it is possible that EPB41L4A-AS1 primarily acts as a transcriptional regulator of SUB1 mRNA, or that its RNA product is required for proper stability and/or localization of the SUB1 protein, or that EPB41L4A-AS1 acts as a scaffold for the formation of protein-protein interactions of SUB1.”
(11) Figure 8: The scratch assay alone cannot be used as a measure of increased invasion, and this phenotype must be confirmed with a transwell invasion or migration assay. Thus, I highly recommend that the authors conduct this experiment using the Boyden chamber. Do the authors see upregulation of N-cadherin, Vimentin, and downregulation of E-cadherin in their RNA-seq?
We agree with the reviewer that those phenotypes are complex and normally require multiple in vitro, as well as in vivo assays to be thoroughly characterized. However, we respectfully consider those as out of scope of the current work, which is more focused on RNA biology and the molecular characterization and functions of EPB41L4A-AS1.
Nevertheless, in Fig. 8D we show that the canonical EMT signature (taken from MSigDB) is upregulated in cells with reduced expression of EPB41L4A-AS1. Notably, EMT has been found to not possess an unique gene expression program, but it rather involves distinct and partially overlapping gene signatures (Youssef et al., 2024). In Fig. 8D, the most upregulated gene is TIMP3, a matrix metallopeptidase inhibitor linked to a particular EMT signature that is less invasive and more profibrotic (EMT-T2, (Youssef et al., 2024)). Interestingly, we observed a strong upregulation of other genes linked to EMT-T2, such as TIMP1, FOSB, SOX9, JUNB, JUN and KLF4, whereas MPP genes (linked to EMT-T1, which is highly proteolytic and invasive) are generally downregulated or not expressed. With regards to N- and E-cadherin, the first does not pass our cutoff to be considered expressed, and the latter is not significantly changing. Vimentin is also not significantly dysregulated. All these examples are reported, which were added as Fig. 8E:
The text has also been updated accordingly:
“These findings suggest that proper EPB41L4A-AS1 expression is required for cellular proliferation, whereas its deficiency results in the onset of more aggressive and migratory behavior, likely linked to the increase of the gene signature of epithelial to mesenchymal transition (EMT) (Fig. 8D). Because EMT is not characterized by a unique gene expression program and rather involves distinct and partially overlapping gene signatures (Youssef et al., 2024), we checked the expression level of marker genes linked to different types of EMTs (Fig. 8E). The most upregulated gene in Fig. 8D is TIMP3, a matrix metallopeptidase inhibitor linked to a particular EMT signature that is less invasive and more profibrotic (EMT-T2) (Youssef et al., 2024). Interestingly, we observed a stark upregulation of other genes linked to EMT-T2, such as TIMP1, FOSB, SOX9, JUNB, JUN and KLF4, whereas MPP genes (linked to EMT-T1, which is highly proteolytic and invasive) are generally downregulated or not expressed. This suggests that the downregulation of EPB41L4A-AS1 is primarily linked to a specific EMT program (EMT-T2), and future studies aimed at uncovering the exact mechanisms and relevance will shed light upon a possible therapeutic potential of this lncRNA.”
(12) Minor points:
(a) What could be the explanation for why only the EPB41L4A-AS1 locus has an effect on the neighbouring gene?
There might be multiple reasons why EPB41L4A-AS1 is able to modulate the expression of the neighboring genes. First, it is expressed from a TAD boundary exhibiting physical contacts with several genes in the two flanking TADs (Fig. 1F and 2A), placing it in the right spot to regulate their expression. Second, it is highly expressed when compared to most of the genes nearby, with transcription having been linked to the establishment and maintenance of TAD boundaries (Costea et al., 2023). Accordingly, the (partial) depletion of EPB41L4A-AS1 via GapmeRs transfection slightly reduces the contacts between the lncRNA and EPB41L4A loci (Fig. 2E and S4J), although this effect could also be determined by a premature transcription termination triggered by the GapmeRs.
There are a multitude of mechanisms by which lncRNAs with regulatory functions modulate the expression of one or more target genes in cis (Gil & Ulitsky, 2020), and our data do not unequivocally point to one of them. Distinguishing between these possibilities is a major challenge in the field and would be difficult to address in the context of this one study. It could be that the processive RNA polymerases at the EPB41L4A-AS1 locus are recruited to the neighboring loci, facilitated by the close proximity in the 3D space. It could also be possible that chromatin remodeling factors are recruited by the nascent RNA, and then promote and/or sustain the opening of chromatin at the target site. The latter possibility is intriguing, as this mechanism is proposed to be widespread among lncRNAs (Gil & Ulitsky, 2020; Oo et al., 2025) and we observed a significant reduction of H3K27ac levels at the EPB41L4A promoter region (Fig. 2D). Future studies combining chromatin profiling (e.g., CUT&RUN and ATAC-seq) and RNA pulldown experiments will shed light upon the exact mechanisms by which this lncRNA regulates the expression of target genes in cis and its interacting partners.
(b) The scale bar is missing on all the images in the Supplementary Figures as well.
The scale bars are present in the top-left figure of each panel. We acknowledge that due to the export as PNG, some figures (including those with microscopy images) display abnormal font sizes and aspect ratio. All images were created using consistent fonts, sizes and ratio, and are provided as high-quality PDF in the current submission.
(13) Methods:
The authors should double-check if they used sirn and LNA gapmers at 25 and 50um concentrations, as that is a huge dose. Most papers used these reagents in the range of 5-50nM maximum.
We apologize for the typo, the text has been fixed. We performed the experiments at 25 and 50nM, respectively, as suggested by the manufacturer’s protocol.
(14) Discussion:
Which cell lines were used in reference 27 (Cheng et al., 2024 Cell) to study the role of SNORA13? It may be useful to include this in the discussion.
We already mentioned the cell system in the discussion, and now we edited to include the specific cell line that was used:
“A recent study found that SNORA13 negatively regulates ribosome biogenesis in TERT-immortalized human fibroblasts (BJ-HRAS<Sup>G12V</sup>), by decreasing the incorporation of RPL23 into the maturing 60S ribosomal subunits, eventually triggering p53-mediated cellular senescence(Cheng et al., 2024).”
Reviewer #3 (Recommendations for the authors):
Major comments on weaknesses:
(1) The paper is quite disjointed:
(a) Figures1/2 studied the cis- and potential trans target genes altered by EPB41L4A-AS1 knockdown. They also showed some data about EPB41L4A-AS1 overlaps a strong chromatin boundary.
(b) Figures3/4/5 studied the role of SUB1 - as it is altered by EPB41L4A-AS1 knockdown - in affecting genes and snoRNAs, which may partially underlie the gene/snoRNA changes after EPB41L4A-AS1 knockdown.
(c) Figure 6 showed that EPB41L4A-AS1 knockdown did not directly affect SNORA13, the snoRNA located in the intron of EPB41L4A-AS1. Thus, the upregulation of many snoRNAs is not due to SNORA13.
(d) Figure 7 studied whether the changes of cis genes or snoRNAs are due to transcriptional stability.
(e) Figure 8 studied cellular phenotypes after EPB41L4A-AS1 knockdown.
These points are overly spread out and this dilutes the central theme of these results, which this Reviewer considered to be on cis or trans gene regulation by this lncRNA.The title of the paper implies EPB41L4A-AS1 knockdown affected trans target genes, but the paper did not focus on studying cis or trans effects, except briefly mentioning that many genes were changed in Figure 2. The many changes of snoRNAs are suggested to be partially explained by SUB1, but SUB1 itself is affected (>50%, Figure 3B) by EPB41L4A-AS1 knockdown, so it is unclear if these are mostly secondary changes due to SUB1 reduction. Given the current content of the paper, the authors do not have sufficient evidence to support that the changes of trans genes are due to direct effects or indirect effects. And so they are encouraged to revise their title to be more on snoRNA regulation, as this area took the majority of the efforts in this paper.
We respectfully disagree with the reviewer. We show that the effect on the proximal genes are cis-acting, as they are not rescued by exogenous expression, whereas the majority of the changes observed in the RNA-seq datasets appear to be indirect, and the snoRNA changes, that indeed might be indirect and not necessarily involve direct interaction partners of the lncRNA, such as SUB1, appear to be trans-regulated, as they can be rescued partially by exogenous expression of the lncRNA. We also show that KD of the main cis-regulated gene, EPB41L4A, results in a much milder transcriptional response, further solidifying the contribution of trans-acting effects. While we agree that the snoRNA effects are interesting, we do not consider them to be the main result, as they are accompanied by many additional changes in gene expression, and changes in the subnuclear distribution of the key nucleolar proteins, so it is difficult for us to claim that EPB41L4A-AS1 is specifically relevant to the snoRNAs rather than to the more broad nucleolar biology. Therefore, we prefer not to mention snoRNAs specifically in the title.
(2) EPB41L4A-AS1 knockdown caused ~2,364 gene changes. This is a very large amount of change on par with some transcriptional factors. It thus needs more scrutiny. First, on Page 9, second paragraph, the authors used|log2Fold-change| >0.41 to select differential genes, which is an unusual cutoff. What is the rationale? Often |log2Fold-change| >1 is more common. How many replicates are used? To examine how many gene changes are likely direct target genes, can the authors show how many of the cist-genes that are changed by EPB41L4A-AS1 knockdown have direct chromatin contacts with EPB41L4A-AS1 in HiC data? Is there any correlation between HiC contact with their fold changes? Without a clear explanation of cis target genes as direct target genes, it is more difficult to establish whether any trans target genes are directly affected by EPB41L4A-AS1 knockdown.
A |log<sub>2</sub>Fold-change| >0.41 equals a change of 33% or more, which together with an adjusted P < 0.05 is a threshold that has been used in the past. All RNA-seq experiments have been performed in triplicates, in line with the standards in the field. While it is possible that the EPB41L4A-AS1 establishes multiple contacts in trans—a process that has been observed in at least another lncRNA, namely Firre but involving its mature RNA product—we do believe this to be less likely that the alternative, namely that the > 2,000 DEGs are predominantly result from secondary changes rather than genes directly regulated by EPB41L4A-AS1 contacts.
In any case, we have inspected our UMI-4C data to identify other genes exhibiting higher contact frequencies than background levels, and thus, potentially regulated in cis. To this end, we calculated the UMI-4C coverage in a 10kb window centered around the TSS of the genes located on chromosome 5, which we subsequently normalized based on the distance from EPB41L4A-AS1, in order to account for the intrinsic higher DNA recovery the closer to the target DNA sequence. However, in our UMI-4C experiment we have employed baits targeting three different genes—EPB41L4A-AS1, EPB41L4A and STARD4—and therefore such approach assumes that the lncRNA locus has the most regulatory features in this region. As expected, we detected a strong negative correlation between the normalized coverage and the distance from the EPB41L4A-AS1 locus (⍴ = -0.51, p-value < 2.2e-16), and the genes in the two neighboring TADs exhibited the strongest association with the bait region (Author response image 5). The genes that we see are down-regulated in the adjacent TADs, namely NREP, MCC and MAN2A1 (Fig. 2F) show substantially higher contacts than background with the EPB41L4A-AS1 gene, thus potentially constituting additional cis-regulated targets of this lncRNA. We note that both SUB1 and NPM1 are located on chromosome 5 as well, albeit at distances exceeding 75 and 50 Mb, respectively, and they do not exhibit any striking association with the lncRNA locus.
Author response image 5.
UMI-4C coverage over the TSS of the genes located on chromosome 5. (A) Correlation between the normalized UMI-4C coverage over the TSS (± 5kb) of chromosome 5 genes and the absolute distance (in megabases, Mb) from EPB41L4A-AS1. (B) Same as in (A), but with the x axis showing the relative distance from EPB41L4A-AS1. In both cases, the genes in the two flanking TADs are colored in red and their names are reported.
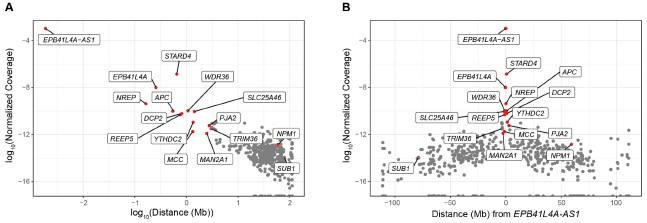
To increase the confidence in our RNA-seq data, we have now performed another round of polyA+ RNA-seq following EPB41L4A-AS1 knockdown using LNA1 or LNA2, as well as the previously used and an additional control GapmeR. The FPKMs of the control samples are highly-correlated both within replicates and between GapmeRs (Fig. S6A). More importantly, the fold-changes to control are highly correlated between the two on-target GapmeRs LNA1 and LNA2, regardless of the GapmeR used for normalization (Fig. S6B), thus showing that despite significant GapmeR-specific effects, the bulk of the response is shared and likely the direct result of the reduction in the levels of EPB41L4A-AS1. Notably, key targets NPM1 and MTREX (see discussion, Fig. S12A-C and comments to Reviewer 3) were found to be downregulated by both LNAs (Fig. S6C).
However, we acknowledge that some of the dysregulated genes are observed only when using one GapmeR and not the other, likely due to a combination of indirect, secondary and non-specific effects, and as such it is difficult without short time-course experiments (Much et al., 2024) to infer the direct response. Supporting this, LNA2 yielded a total of 1,069 DEGs (617 up and 452 down) and LNA1 2,493 DEGs (1,328 up and 1,287 down), with the latter triggering a stronger response most likely as a result of the previously mentioned CDKN1A/p21 induction. Overall, 45.1% of the upregulated genes following LNA2 transfection were shared with LNA1, in contrast to only the 24.3% of the downregulated ones.
We have now included these results in the Results section (see below) and in Supplementary Figure (Fig. S6).
“Most of the consequences of the depletion of EPB41L4A-AS1 are thus not directly explained by changes in EPB41L4A levels. An additional trans-acting function for EPB41L4A-AS1 would therefore be consistent with its high expression levels compared to most lncRNAs detected in MCF-7 (Fig. S5G). To strengthen these findings, we have transfected MCF-7 cells with LNA1 and a second control GapmeR (NT2), as well as the previous one (NT1) and LNA2, and sequenced the polyadenylated RNA fraction as before. Notably, the expression levels (in FPKMs) of the replicates of both control samples are highly correlated with each other (Fig. S6A), and the global transcriptomic changes triggered by the two EPB41L4A-AS1-targeting LNAs are largely concordant (Fig. S6B and S6C). Because of this concordance and the cleaner (i.e., no CDKN1A upregulation) readout in LNA2-transfected cells, we focused mainly on these cells for subsequent analyses.”
Figure 3B, SUB1 mRNA is reduced >half by EPB41L4A-AS1 KD. How much did SUB1 protein reduce after EPB41L4A-AS1 KD? Similarly, how much is the NPM1 protein reduced? If these two important proteins were affected by EPB41L4A-AS1 KD simultaneously, it is important to exclude how many of the 2,364 genes that changed after EPB41L4A-AS1 KD are due to the protein changes of these two key proteins. For SUB1, Figures S7E,F,G provided some answers. But NPM1 KD is also needed to fully understand such. Related to this, there are many other proteins perhaps changed in addition to SUB1 and NPM1, this renders it concerning how many of the EPB41L4A-AS1 KD-induced changes are directly caused by this RNA. In addition to the suggested study of cist targets, the alternative mechanism needs to be fully discussed in the paper as it remains difficult to fully conclude direct versus indirect effect due to such changes of key proteins or ncRNAs (such as snoRNAs or histone mRNAs).
As requested by both Reviewer #2 and #3, we have performed WB for SUB1, NPM1 and FBL following EPB41L4A-AS1 KD with two targeting (LNA1 and LNA2) and the previous control GapmeRs. Interestingly, we did not detect any significant downregulation of either proteins (Author response image 3), although this might be the result of the high variability observed in the control samples. Moreover, the short timeframe in which the experiments have been conducted━that is, transient transfections for 3 days━might not be sufficient time for the existing proteins to be degraded, and thus, the downregulation is more evident at the RNA (Fig. 3B and Supplementary Figure 6C) rather than protein level.
We acknowledge that many proteins might change simultaneously, and to pinpoint which ones act upstream of the plethora of indirect changes is extremely challenging when considering such large-scale changes in gene expression. In the case of SUB1 and NPM1━which were prioritized for their predicted binding to the lncRNA (Fig. 3A)━we show that the depletion of the former affects the latter in a similar way than that of the lncRNA (Fig. 5F). Moreover, snoRNAs changes are also similarly affected (as the reviewer pointed out, Fig. 4F), suggesting that at least this phenomenon is predominantly mediated by SUB1. Other effects might also be indirect consequences of cellular responses, such as the decrease in histone mRNAs (Fig. 4A) that might reflect the decrease in cellular replication (Fig. 8C) and cell cycle genes (Fig. 2I) (although a link between SUB1 and histone mRNA expression has been described (Brzek et al., 2018)).
Supporting the notion that additional proteins might be involved in driving the observed phenotypes, one of the genes that most consistently was affected by EPB41L4A-AS1 KD with GapmeRs is MTREX (also known as MTR4), that becomes downregulated at both the RNA and protein levels (now presented in the main text as Supplementary Figure 12). MTREX it’s part of the NEXT and PAXT complexes (Contreras et al., 2023), that target several short-lived RNAs for degradation, and the depletion of either MTREX or other complex members leads to the upregulation of such RNAs, that include PROMPTs, uaRNAs and eRNAs, among others. Given the lack in our understanding in snoRNA biogenesis from introns in mammalian systems(Monziani & Ulitsky, 2023), it is tempting to hypothesize a role for MTREX-containing complexes in trimming and degrading those introns and release the mature snoRNAs.
We updated the discussion section to include these observations:
“Beyond its site of transcription, EPB41L4A-AS1 associates with SUB1, an abundant protein linked to various functions, and these two players are required for proper distribution of various nuclear proteins. Their dysregulation results in large-scale changes in gene expression, including up-regulation of snoRNA expression, mostly through increased transcription of their hosts, and possibly through a somewhat impaired snoRNA processing and/or stability. To further hinder our efforts in discerning between these two possibilities, the exact molecular pathways involved in snoRNAs biogenesis, maturation and decay are still not completely understood. One of the genes that most consistently was affected by EPB41L4A-AS1 KD with GapmeRs is MTREX (also known as MTR4), that becomes downregulated at both the RNA and protein levels (Fig. S12A-C). Interestingly, MTREX it is part of the NEXT and PAXT complexes(Contreras et al., 2023), that target several short-lived RNAs for degradation, and the depletion of either MTREX or other complex members leads to the upregulation of such RNAs, that include PROMPTs, uaRNAs and eRNAs, among others. It is therefore tempting to hypothesize a role for MTREX-containing complexes in trimming and degrading those introns, and releasing the mature snoRNAs. Future studies specifically aimed at uncovering novel players in mammalian snoRNA biology will both conclusively elucidate whether MTREX is indeed involved in these processes.”
With regards to the changes in gene expression between the two LNAs, we provide a more detailed answer above and to the other reviewers as well.
(3) A Strong discrepancy of results by different approaches of knockdown or overexpression:
(a) CRISPRa versus LNA knockdown: Figure S4 - CRISPRa of EPB41L4A-AS1 did not affect EPB41L4A expression (Figure S4B). The authors should discuss how to interpret this result. Did CRISPRa not work to increase the nuclear/chromatin portion of EPB41L4A-AS1? Did CRISPRa of EPB41L4A-AS1 affect the gene in the upstream, the STARD4? Did CRISPRa of EPB41L4A-AS1 also affect chromatin interactions between EPB41L4A-AS1 and the EPB41L4A gene? If so, this may argue that chromatin interaction is not necessary for cis-gene regulation.
There are indeed several possible explanations, the most parsimonious is that since the lncRNA is already very highly transcribed, the relatively modest effect of additional transcription mediated by CRISPRa is not sufficient to elicit a measurable effect. For this reason, we did not check by UMI-4C the contact frequency between the lncRNA and EPB41L4A upon CRISPRa.
CRISPRa augments transcription at target loci, and thus, the nuclear and chromatin retention of EPB41L4A-AS1 are not expected to be affected. We did not check the expression of STARD4, because we focused on EPB41L4A which appears to be the main target locus according to Hi-C (Fig. 2A), UMI-4C (Fig. 2E and S4J) and GeneHancer (Fig. S1).
We already provide extensive evidence of a cis-regulation of EPB41L4A-AS1 over EPB41L4A, and show that EPB41L4A is lowly-expressed and likely has a limited role in our experimental settings. Thus, we respectfully propose that an in-deep exploration of the mechanism of action of this regulatory axis is out of scope of the current study, that instead focused more on the global effects of EPB41L4A-AS1 perturbation.
(b) Related to this, while CRISPRa alone did not show an effect, upon LNA knockdown of EPB41L4A-AS1, CRISPRa of EPB41L4A-AS1 can increase EPB41L4A expression. It is perplexing as to why, upon LNA treatment, CRISPRa will show an effect (Figure S4H)? Actually, Figures S4H and I are very confusing in the way they are currently presented. They will benefit from being separated into two panels (H into 2 and I into two). And for Ectopic expression, please show controls by empty vector versus EPB41L4A-AS1, and for CRISPRa, please show sgRNA pool versus sgRNA control.
The results are consistent with the parsimonious assumption mentioned above that the high transcription of the lncRNA at baseline is sufficient for maximal positive regulation of EPB41L4A, and that upon KD, the reduced transcription and/or RNA levels are no longer at saturating levels, and so CRISPRa can have an effect. We now mention this interpretation in the text:
“Levels of EPB41L4A were not affected by increased expression of EPB41L4A-AS1 from the endogenous locus by CRISPR activation (CRISPRa), nor by its exogenous expression from a plasmid (Fig. S4B and S4C). The former suggests that endogenous levels of EPB41L4A-AS1—that are far greater than those of EPB41L4A—are sufficient to sustain the maximal expression of this target gene in MCF7 cells.”
We apologize for the confusion regarding the control used in the rescue experiments in Fig. S4H and S4I. The “-” in the Ectopic overexpression and CRISPRa correspond to the Empty Vector and sgControl, respectively, and not the absence of any vector. We changed the text in the figure legends:
“(H) Changes in EPB41L4A-AS1 expression after rescuing EPB41L4A-AS1 with an ectopic plasmid or CRISPRa following its KD with GapmeRs. In both panels (Ectopic OE and CRISPRa) the “-” samples represent those transfected with the Empty Vector or sgControl. Asterisks indicate significance relative to the –/– control (transfected with both the control GapmeR and vector). (I) Same as in (H), but for changes in EPB41L4A expression.”
(c) siRNA versus LNA knockdown: Figure S3A showed that siRNA KD of EPB41L4A-AS1 does not affect EPB41L4A expression. How to understand this data versus LNA?
As explained in the text, siRNA-mediated KD presumably affects mostly the cytoplasmic pool of EPB41L4A-AS1 and not the nuclear one, which we assume explains the different effects of the two perturbations, as observed for other lncRNAs (e.g., (Ntini et al., 2018)). However, we acknowledge that we do not know what aspect of the nuclear RNA biology is relevant, let it be the nascent EPB41L4A-AS1 transcription, premature transcriptional termination or even the nuclear pool of this lncRNA, and this can be elucidated further in future studies.
(d) EPB41L4A-AS1 OE versus LNA knockdown: Figure 6F showed that EPB41L4A-AS1 OE caused reduction of EPB41L4A mRNA, particularly at 24hr. How to interpret that both LNA KD and OE of EPB41L4A-AS1 reduce the expression of EPB41L4A mRNA?
We do not believe that the OE of EPB41L4A-AS1, and in particular the one elicited by an ectopic plasmid affects EPB41L4A RNA levels. In the experiment in Fig. 6F, EPB41L4A relative expression at 24h is ~0.65 (please note the log<sub>2</sub> scale in the graph), which is significant as reported. However, throughout this study (and as shown in Fig. S4C for the ectopic and Fig. S4B for the CRISPRa overexpression, respectively), we observed no such behavior, suggesting that the effect reported in Fig. 6F is the result of either that particular setting, and unlikely to reflect a general phenomenon.
(e) Did any of the effects on snoRNAs or trans target genes after EPB41L4A-AS1 knockdown still appear by CRISPRa?
As mentioned above, we did a limited number of experiments after CRISPRa, prompted by the fact that endogenous levels of EPB41L4A-AS1 are already high enough to sustain its functions. Pushing the expression even higher will likely result in no or artifactual effects, which is why we respectfully propose such experiments are not essential in this current work, which instead mostly relies on loss-of-function experiments.
For issue 3, extensive data repetition using all these methods may be unrealistic, but key data discrepancy needs to be fully discussed and interpreted.
Other comments on weakness:
(1) This manuscript will benefit from having line numbers so comments from Reviewers can be made more specifically.
We added line numbers as suggested by the reviewer.
(2) Figure 2G, to distinguish if any effects of EPB41L4A-AS1 come from the cytoplasmic or nuclear portion of EPB41L4A-AS1, an siRNA KD RNA-seq will help to filter out the genes affected by EPB41L4A-AS1 in the cytoplasm, as siRNA likely mainly acts in the cytoplasm.
This experiment would be difficult to interpret as while the siRNAs mostly deplete the cytoplasmic pool of their target, they can have some effects in the nucleus as well (e.g., (Sarshad et al., 2018)) and so siRNAs knockdown will not necessarily report strictly on the cytoplasmic functions.
(3) Figure 2H, LNA knockdown of EPB41L4A should check the protein level reduction, is it similar to the change caused by knockdown of EPB41L4A-AS1?
As suggested by reviewer #2, we have now replaced the EPB41L4A Western Blot that now shows the results with both LNA1 and LNA2. Please note that the previous Fig. 2C was a subset of this, i.e., we have previously cropped the results obtained with LNA1. Unfortunately, we did not have sufficient antibody to check for EPB41L4A protein reduction following LNA KD of EPB41L4A in a timely manner.
(4) There are two LNA Gapmers used by the paper to knock down EPB41L4A-AS1, but some figures used LNA1, some used LNA2, preventing a consistent interpretation of the results. For example, in Figures 2A-D, LNA2 was used. But in Figures 2E-H, LNA1 was used. How consistent are the two in changing histone H3K27ac (like in Figure 2D) versus gene expression in RNA-seq? The changes in chromatin interaction appear to be weaker by LNA2 (Figure S4J) versus LNA1 (Figure 2E).
As explained above and in response to Reviewer #1, we now provide more RNA-seq data for LNA1 and LNA2. We note that besides the unwanted and/or off-target effects, these two GapmeRs might be not equally effective in knocking down EPB41L4A-AS1, which could explain why LNA1 seems to have a stronger effect on chromatin than LNA2. Nonetheless, when we have employed both we have obtained similar and consistent results (e.g., Fig. 5A-D and 8A-C), suggesting that these and the other effects are indeed on target effects due to EPB41L4A-AS1 depletion.
(5) It will be helpful if the authors provide information on how long they conducted EPB41L4A-AS1 knockdown for most experiments to help discern direct or indirect effects.
The length of all perturbations was indicated in the Methods section, and we now mention them also in the Results. Unless specified otherwise, they were carried out for 72 hours. We agree with the reviewer that having time course experiments can have added value, but due to the extensive effort that these will require, we suggest that they are out of scope of the current study.
(6) In Figures 1C and F, the authors showed results about EPB41L4A-AS1 overlapping a strong chromatin boundary. But these are not mentioned anymore in the later part of the paper. Does this imply any mechanism? Does EPB41L4A-AS1 knockdown or OE, or CRISPRa affect the expression of genes near the other interacting site, STARD4? Do genes located in the two adjacent TADs change more strongly as compared to other genes far away?
We discuss this point in the Discussion section:
“At the site of its own transcription, which overlaps a strong TAD boundary, EPB41L4A-AS1 is required to maintain expression of several adjacent genes, regulated at the level of transcription. Strikingly, the promoter of EPB41L4A-AS1 ranks in the 99.8th percentile of the strongest TAD boundaries in human H1 embryonic stem cells(Open2C et al., 2024; Salnikov et al., 2024). It features several CTCF binding sites (Fig. 2A), and in MCF-7 cells, we demonstrate that it blocks the propagation of the 4C signal between the two flanking TADSs (Fig. 1F). Future studies will help elucidate how EPB41L4A-AS1 transcription and/or the RNA product regulate this boundary. So far, we found that EPB41L4A-AS1 did not affect CTCF binding to the boundary, and while some peaks in the vicinity of EPB41L4A-AS1 were significantly affected by its loss, they did not appear to be found near genes that were dysregulated by its KD (Fig. S11C). We also found that KD of EPB41L4A-AS1—which depletes the RNA product, but may also affect the nascent RNA transcription(Lai et al., 2020; Lee & Mendell, 2020)—reduces the spatial contacts between the TAD boundary and the EPB41L4A promoter (Fig. 2E). Further elucidation of the exact functional entity needed for the cis-acting regulation will require detailed genetic perturbations of the locus, that are difficult to carry out in the polypoid MCF-7 cells, without affecting other functional elements of this locus or cell survival as we were unable to generate deletion clones despite several attempts.”
As mentioned in the text (pasted below) and in Fig. 2F, most genes in the two flanking TADs become downregulated following EPB41L4A-AS1 KD. While STARD4 – which was chosen because it had spatial contacts above background with EPB41L4A-AS1 – did not reach statistical significance, others did and are highlighted. Those included NREP, which we also discuss:
“Consistently with the RT-qPCR data, KD of EPB41L4A-AS1 reduced EPB41L4A expression, and also reduced expression of several, but not all other genes in the TADs flanking the lncRNA (Fig. 2F).Based on these data, EPB41L4A-AS1 is a significant cis-acting activator according to TransCistor (Dhaka et al., 2024) (P=0.005 using the digital mode). The cis-regulated genes reduced by EPB41L4A-AS1 KD included NREP, a gene important for brain development, whose homolog was downregulated by genetic manipulations of regions homologous to the lncRNA locus in mice(Salnikov et al., 2024). Depletion of EPB41L4A-AS1 thus affects several genes in its vicinity.”
(7) Related to the description of SUB1 regulation of genes are DNA and RNA levels: "Of these genes, transcripts of only 56 genes were also bound by SUB1 at the RNA level, suggesting largely distinct sets of genes targeted by SUB1 at both the DNA and the RNA levels." SUB1 binding to chromatin by Cut&Run only indicates that it is close to DNA/chromatin, and this interaction with chromatin may still likely be mediated by RNAs. The authors used SUB1 binding sites in eCLIP-seq to suggest whether it acts via RNAs, but these binding sites are often from highly expressed gene mRNAs/exons. Standard analysis may not have examined low-abundance RNAs close to the gene promoters, such as promoter antisense RNAs. The authors can examine whether, for the promoters with cut&run peaks of SUB1, SUB1 eCLIP-seq shows binding to the low-abundance nascent RNAs near these promoters.
In response to a related comment by Reviewer 1, we now show that when considering expression level–matched control genes, knockdown of EPB41L4A-AS1 still significantly affects expression of SUB1 targets over controls. The results are presented in Supplementary Figure 7 (Fig. S7C).
Based on this analysis, while there is a tendency of increased expression with increased SUB1 binding, when controlling for expression levels the effect of down-regulation of SUB1-bound RNAs upon lncRNA knockdown remains, suggesting that it is not merely a confounding effect. We have updated the text as follows:
“We hypothesized that loss of EPB41L4A-AS1 might affect SUB1, either via the reduction in its expression or by affecting its functions. We stratified SUB1 eCLIP targets into confidence intervals, based on the number, strength and confidence of the reported binding sites. Indeed, eCLIP targets of SUB1 (from HepG2 cells profiled by ENCODE) were significantly downregulated following. EPB41L4A-AS1 KD in MCF-7, with more confident targets experiencing stronger downregulation (Fig. 3C). Importantly, this still holds true when controlling for gene expression levels (Fig. S7C), suggesting that this negative trend is not due to differences in their baseline expression.”
(8) Figure 8, the cellular phenotype is interesting. As EPB41L4A-AS1 is quite widely expressed, did it affect the phenotypes similarly in other breast cancer cells? MCF7 is not a particularly relevant metastasis model. Can a similar phenotype be seen in commonly used metastatic cell models such as MDA-MB-231?
We agree that further expanding the models in which EPB41L4A-AS1 affects cellular proliferation, migration and any other relevant phenotype is of potential interest before considering targeting this lncRNA as a therapeutic approach. However, given that 1) others have already identified similar phenotypes upon the modulation of EPB41L4A-AS1 in a variety of different systems (see Results and Discussion), and 2) we were most interested in the molecular consequences following the loss of this lncRNA, we respectfully suggest that these experiments are out of scope of the current study.
References
Bahar Halpern, K., Caspi, I., Lemze, D., Levy, M., Landen, S., Elinav, E., Ulitsky, I., & Itzkovitz, S. (2015). Nuclear Retention of mRNA in Mammalian Tissues. Cell Reports, 13(12), 2653–2662.
Brabletz, T., Kalluri, R., Nieto, M. A., & Weinberg, R. A. (2018). EMT in cancer. Nature Reviews. Cancer, 18(2), 128–134.
Brzek, A., Cichocka, M., Dolata, J., Juzwa, W., Schümperli, D., & Raczynska, K. D. (2018). Positive cofactor 4 (PC4) contributes to the regulation of replication-dependent canonical histone gene expression. BMC Molecular Biology, 19(1), 9.
Cheng, Y., Wang, S., Zhang, H., Lee, J.-S., Ni, C., Guo, J., Chen, E., Wang, S., Acharya, A., Chang, T.-C., Buszczak, M., Zhu, H., & Mendell, J. T. (2024). A non-canonical role for a small nucleolar RNA in ribosome biogenesis and senescence. Cell, 187(17), 4770–4789.e23.
Conesa, C., & Acker, J. (2010). Sub1/PC4 a chromatin associated protein with multiple functions in transcription. RNA Biology, 7(3), 287–290.
Contreras, X., Depierre, D., Akkawi, C., Srbic, M., Helsmoortel, M., Nogaret, M., LeHars, M., Salifou, K., Heurteau, A., Cuvier, O., & Kiernan, R. (2023). PAPγ associates with PAXT nuclear exosome to control the abundance of PROMPT ncRNAs. Nature Communications, 14(1), 6745.
Costea, J., Schoeberl, U. E., Malzl, D., von der Linde, M., Fitz, J., Gupta, A., Makharova, M., Goloborodko, A., & Pavri, R. (2023). A de novo transcription-dependent TAD boundary underpins critical multiway interactions during antibody class switch recombination. Molecular Cell, 83(5), 681–697.e7.
Das, C., Hizume, K., Batta, K., Kumar, B. R. P., Gadad, S. S., Ganguly, S., Lorain, S., Verreault, A., Sadhale, P. P., Takeyasu, K., & Kundu, T. K. (2006). Transcriptional coactivator PC4, a chromatin-associated protein, induces chromatin condensation. Molecular and Cellular Biology, 26(22), 8303–8315.
Dhaka, B., Zimmerli, M., Hanhart, D., Moser, M. B., Guillen-Ramirez, H., Mishra, S., Esposito, R., Polidori, T., Widmer, M., García-Pérez, R., Julio, M. K., Pervouchine, D., Melé, M., Chouvardas, P., & Johnson, R. (2024). Functional identification of cis-regulatory long noncoding RNAs at controlled false discovery rates. Nucleic Acids Research, 52(6), 2821–2835.
Didiot, M.-C., Ferguson, C. M., Ly, S., Coles, A. H., Smith, A. O., Bicknell, A. A., Hall, L. M., Sapp, E., Echeverria, D., Pai, A. A., DiFiglia, M., Moore, M. J., Hayward, L. J., Aronin, N., & Khvorova, A. (2018). Nuclear Localization of Huntingtin mRNA Is Specific to Cells of Neuronal Origin. Cell Reports, 24(10), 2553–2560.e5.
Dongre, A., & Weinberg, R. A. (2019). New insights into the mechanisms of epithelial-mesenchymal transition and implications for cancer. Nature Reviews. Molecular Cell Biology, 20(2), 69–84.
Dubois, J.-C., Bonnell, E., Filion, A., Frion, J., Zimmer, S., Riaz Khan, M., Teplitz, G. M., Casimir, L., Méthot, É., Marois, I., Idrissou, M., Jacques, P.-É., Wellinger, R. J., & Maréchal, A. (2025). The single-stranded DNA-binding factor SUB1/PC4 alleviates replication stress at telomeres and is a vulnerability of ALT cancer cells. Proceedings of the National Academy of Sciences of the United States of America, 122(2), e2419712122.
Garavís, M., & Calvo, O. (2017). Sub1/PC4, a multifaceted factor: from transcription to genome stability. Current Genetics, 63(6), 1023–1035.
Gil, N., & Ulitsky, I. (2020). Regulation of gene expression by cis-acting long non-coding RNAs. Nature Reviews. Genetics, 21(2), 102–117.
Hou, Y., Gan, T., Fang, T., Zhao, Y., Luo, Q., Liu, X., Qi, L., Zhang, Y., Jia, F., Han, J., Li, S., Wang, S., & Wang, F. (2022). G-quadruplex inducer/stabilizer pyridostatin targets SUB1 to promote cytotoxicity of a transplatinum complex. Nucleic Acids Research, 50(6), 3070–3082.
Jan, C. H., Friedman, R. C., Ruby, J. G., & Bartel, D. P. (2011). Formation, regulation and evolution of Caenorhabditis elegans 3’UTRs. Nature, 469(7328), 97–101.
Kaypee, S., Ochiai, K., Shima, H., Matsumoto, M., Alam, M., Ikura, T., Kundu, T. K., & Igarashi, K. (2025). Positive coactivator PC4 shows dynamic nucleolar distribution required for rDNA transcription and protein synthesis. Cell Communication and Signaling : CCS, 23(1), 283.
Lai, F., Damle, S. S., Ling, K. K., & Rigo, F. (2020). Directed RNase H Cleavage of Nascent Transcripts Causes Transcription Termination. Molecular Cell, 77(5), 1032–1043.e4.
Lee, J.-S., & Mendell, J. T. (2020). Antisense-Mediated Transcript Knockdown Triggers Premature Transcription Termination. Molecular Cell, 77(5), 1044–1054.e3.
Lubelsky, Y., & Ulitsky, I. (2018). Sequences enriched in Alu repeats drive nuclear localization of long RNAs in human cells. Nature, 555(7694), 107–111.
Ly, S., Didiot, M.-C., Ferguson, C. M., Coles, A. H., Miller, R., Chase, K., Echeverria, D., Wang, F., Sadri-Vakili, G., Aronin, N., & Khvorova, A. (2022). Mutant huntingtin messenger RNA forms neuronal nuclear clusters in rodent and human brains. Brain Communications, 4(6), fcac248.
Maranon, D. G., & Wilusz, J. (2020). Mind the Gapmer: Implications of Co-transcriptional Cleavage by Antisense Oligonucleotides. Molecular Cell, 77(5), 932–933.
Monziani, A., & Ulitsky, I. (2023). Noncoding snoRNA host genes are a distinct subclass of long noncoding RNAs. Trends in Genetics : TIG, 39(12), 908–923.
Mortusewicz, O., Roth, W., Li, N., Cardoso, M. C., Meisterernst, M., & Leonhardt, H. (2008). Recruitment of RNA polymerase II cofactor PC4 to DNA damage sites. The Journal of Cell Biology, 183(5), 769–776.
Much, C., Lasda, E. L., Pereira, I. T., Vallery, T. K., Ramirez, D., Lewandowski, J. P., Dowell, R. D., Smallegan, M. J., & Rinn, J. L. (2024). The temporal dynamics of lncRNA Firre-mediated epigenetic and transcriptional regulation. Nature Communications, 15(1), 6821.
Ntini, E., Louloupi, A., Liz, J., Muino, J. M., Marsico, A., & Ørom, U. A. V. (2018). Long ncRNA A-ROD activates its target gene DKK1 at its release from chromatin. Nature Communications, 9(1), 1636.
Oo, J. A., Warwick, T., Pálfi, K., Lam, F., McNicoll, F., Prieto-Garcia, C., Günther, S., Cao, C., Zhou, YGavrilov, A. A., Razin, S. V., Cabrera-Orefice, A., Wittig, I., Pullamsetti, S. S., Kurian, L., Gilsbach, R., Schulz, M. H., Dikic, I., Müller-McNicoll, M., … Leisegang, M. S. (2025). Long non-coding RNAs direct the SWI/SNF complex to cell type-specific enhancers. Nature Communications, 16(1), 131.
Open2C, Abdennur, N., Abraham, S., Fudenberg, G., Flyamer, I. M., Galitsyna, A. A., Goloborodko, A., Imakaev, M., Oksuz, B. A., Venev, S. V., & Xiao, Y. (2024). Cooltools: Enabling high-resolution Hi-C analysis in Python. PLoS Computational Biology, 20(5), e1012067.
Potapova, T. A., Unruh, J. R., Conkright-Fincham, J., Banks, C. A. S., Florens, L., Schneider, D. A., & Gerton, J. L. (2023). Distinct states of nucleolar stress induced by anticancer drugs. https://doi.org/10.7554/eLife.88799.
Ray, D., Laverty, K. U., Jolma, A., Nie, K., Samson, R., Pour, S. E., Tam, C. L., von Krosigk, N., Nabeel-Shah, S., Albu, M., Zheng, H., Perron, G., Lee, H., Najafabadi, H., Blencowe, B., Greenblatt, J., Morris, Q., & Hughes, T. R. (2023). RNA-binding proteins that lack canonical RNA-binding domains are rarely sequence-specific. Scientific Reports, 13(1), 5238.
Salgado, S., Abreu, P. L., Moleirinho, B., Guedes, D. S., Larcombe, L., & Azzalin, C. M. (2024). Human PC4 supports telomere stability and viability in cells utilizing the alternative lengthening of telomeres mechanism. EMBO Reports, 25(12), 5294–5315.
Salnikov, P., Korablev, A., Serova, I., Belokopytova, P., Yan, A., Stepanchuk, Y., Tikhomirov, S., & Fishman, V. (2024). Structural variants in the Epb41l4a locus: TAD disruption and Nrep gene misregulation as hypothetical drivers of neurodevelopmental outcomes. Scientific Reports, 14(1), 5288.
Sarshad, A. A., Juan, A. H., Muler, A. I. C., Anastasakis, D. G., Wang, X., Genzor, P., Feng, X., Tsai, P.-F., Sun, H.-W., Haase, A. D., Sartorelli, V., & Hafner, M. (2018). Argonaute-miRNA Complexes Silence Target mRNAs in the Nucleus of Mammalian Stem Cells. Molecular Cell, 71(6), 1040–1050.e8.
Tafforeau, L., Zorbas, C., Langhendries, J.-L., Mullineux, S.-T., Stamatopoulou, V., Mullier, R., Wacheul, L., & Lafontaine, D. L. J. (2013). The complexity of human ribosome biogenesis revealed by systematic nucleolar screening of Pre-rRNA processing factors. Molecular Cell, 51(4), 539–551.
Unfried, J. P., & Ulitsky, I. (2022). Substoichiometric action of long noncoding RNAs. Nature Cell Biology, 24(5), 608–615.
Van Nostrand, E. L., Freese, P., Pratt, G. A., Wang, X., Wei, X., Xiao, R., Blue, S. M., Chen, J.-Y.,Cody, N. A. L., Dominguez, D., Olson, S., Sundararaman, B., Zhan, L., Bazile, C., Bouvrette, L. P. B., Bergalet, J., Duff, M. O., Garcia, K. E., Gelboin-Burkhart, C., … Yeo, G. W. (2020). A large-scale binding and functional map of human RNA-binding proteins. Nature, 583(7818), 711–719.
Yang, K., Wang, M., Zhao, Y., Sun, X., Yang, Y., Li, X., Zhou, A., Chu, H., Zhou, H., Xu, J., Wu, M., Yang, J., & Yi, J. (2016). A redox mechanism underlying nucleolar stress sensing by nucleophosmin. Nature Communications, 7, 13599.
Youssef, K. K., Narwade, N., Arcas, A., Marquez-Galera, A., Jiménez-Castaño, R., Lopez-Blau, C., Fazilaty, H., García-Gutierrez, D., Cano, A., Galcerán, J., Moreno-Bueno, G., Lopez-Atalaya, J. P., & Nieto, M. A. (2024). Two distinct epithelial-to-mesenchymal transition programs control invasion and inflammation in segregated tumor cell populations. Nature Cancer, 5(11), 1660–1680.
Yu, L., Ma, H., Ji, X., & Volkert, M. R. (2016). The Sub1 nuclear protein protects DNA from oxidative damage. Molecular and Cellular Biochemistry, 412(1-2), 165–171.
-
-
www.youtube.com www.youtube.com
-
Pédocriminalité: dans le piège des loverboys | Reportage | ARTE Regards
186 676 vues 25 sept. 2025 #reportage #prostitution #arte Reportage disponible jusqu'au 27/08/2026
En France, près de 20 000 adolescentes seraient sous l’emprise de proxénètes parfois à peine plus âgés qu’elles.
Derrière une apparence séduisante et beaucoup de tchatche, ces jeunes hommes appelés parfois "loverboys" utilisent la manipulation amoureuse comme arme de contrôle.
La méthode des "loverboys" est redoutable : séduire, isoler, détruire pour mieux asservir.
Ce reportage raconte comment, de Paris à Maastricht, trois jeunes filles se sont retrouvées dans les griffes de ces proxénètes qui après les avoir séduites, les ont vendues sur Internet.
Bao, contrainte à la prostitution pendant cinq ans, livre un témoignage bouleversant.
Jennifer Pailhé, mère de l’une des victimes, raconte son combat acharné pour sauver sa fille et faire tomber son agresseur.
Chemelle Jongen, ancienne victime devenue lanceuse d’alerte, lutte aujourd’hui pour sensibiliser le public aux mécanismes de l’emprise et aux ravages causés par ce système.
Tous les pays européens sont confrontés à la prostitution des mineurs. Mais chaque pays a sa propre législation et ses méthodes.
Reportage (France, 2025, 31mn)
prostitution #reportage #arte
-
-
www.youtube.com www.youtube.com
-
PEUR BLEUE - Anatomie des violences conjugales et du parcours de sortie
31 275 vues 4 oct. 2023 Ce film a été produit par le Conseil Départemental d'Accès au Droit des Hautes-Pyrénées.
Basé sur des témoignages de victimes et de professionnels, ce film a pour objectif de rendre compte du phénomène des violences et de ses différentes formes, qui peuvent ou non se cumuler.
La victime, dépossédée de sa capacité d'action par les effets de l'emprise et du psycho-trauma, peut cependant trouver des voies de protection et bénéficier d'un accompagnement personnalisé dans un lieu spécialisé.
Retrouvez nous sur Instagram : / cdad65000 <br /> Illustrations faites par Ysar : / __ysar<br /> Ce film a été mis en image et monté par Yannick Chaumeil : / @associationlarrache-temps1871
-
-
www.youtube.com www.youtube.com
-
Synthèse de la Conférence sur le Traitement de l'Information Sociale et les Violences Sexuelles
Résumé Exécutif
Ce document synthétise la conférence plénière de Massil Ben Bouriche sur le traitement de l'information sociale et les violences sexuelles, présentée lors des 20e journées du Groupe de Réflexion en Psychopathologie Cognitive (Grepaco).
La présentation met en lumière l'ampleur des violences sexuelles comme un problème de santé publique majeur, soulignant que les données officielles sous-estiment considérablement le phénomène en raison d'un taux d'attrition judiciaire massif (seule une plainte sur dix aboutit à une condamnation).
L'argument central de la conférence est que les modèles explicatifs dominants de l'agression sexuelle, développés principalement à partir d'échantillons d'auteurs judiciarisés, ont une portée limitée et ne peuvent rendre compte de la majorité des violences commises par des personnes jamais identifiées par les autorités.
Pour dépasser cette limite, une approche basée sur les modèles de traitement de l'information sociale (TIS) est proposée.
Ces modèles analysent les violences sexuelles comme le résultat d'une séquence d'opérations mentales (encodage, interprétation, etc.), fortement influencées par des "structures de connaissance" (schémas, mythes sur le viol) issues des expériences individuelles et des normes socioculturelles.
Les recherches empiriques présentées démontrent que les difficultés cognitives ne sont pas généralisées mais spécifiques.
Par exemple, les hommes ne présentent pas un déficit global dans la reconnaissance des intentions sexuelles, mais une difficulté particulière à identifier une absence d'intérêt.
De plus, l'effet de l'alcool sur la perception du consentement n'est significatif que chez les individus qui adhèrent déjà fortement aux mythes du viol.
Ces constats plaident pour des interventions préventives ciblant les structures de connaissance (stéréotypes de genre, mythes) dès le plus jeune âge, une stratégie dont l'efficacité est soutenue par plusieurs méta-analyses.
--------------------------------------------------------------------------------
1. Contexte de la Conférence : 20e Journées du Grepaco
La conférence s'est déroulée dans le cadre des 20e journées du Groupe de Réflexion en Psychopathologie Cognitive (Grepaco) à Lyon, le 15 mai.
L'événement, centré sur les interactions sociales et le rôle de la cognition, a été organisé avec le soutien de plusieurs entités, notamment :
• Le centre d'excellence iMIND : Un centre labellisé en 2020 dans le cadre de la stratégie nationale pour les troubles du neurodéveloppement.
Sa mission est de faire le lien entre la recherche, les usagers et la clinique, en se concentrant spécifiquement sur les problématiques des adultes, souvent "les grands oubliés" des stratégies nationales.
Ses axes directeurs incluent l'inclusion, la déstigmatisation, l'innovation dans la formation et une recherche translationnelle et participative.
• Le laboratoire EMC, l'Université Lyon 1, le Centre Hospitalier Le Vinatier et d'autres sponsors.
Joël Billieux, au nom du comité scientifique du Grepaco, a rappelé l'histoire du groupe, qui a évolué de rencontres informelles à un congrès scientifique plus structuré, tout en insistant sur la volonté de maintenir une plateforme pour les ateliers et les projets collaboratifs. Un appel a été lancé pour trouver des lieux d'organisation pour les éditions futures (2026-2028).
2. La Conférence de Massil Ben Bouriche
Massil Ben Bouriche, maître de conférences en psychologie et justice à l'Université de Lille, a présenté une conférence plénière intitulée "Information sociale, violences sexuelles et comportements violents".
2.1. L'Ampleur et la Nature des Violences Sexuelles : Un Problème de Santé Publique
La présentation a débuté par un rappel contextuel sur l'ampleur et la nature des violences sexuelles, en s'appuyant sur des enquêtes de victimation et de perpétration plutôt que sur des données officielles jugées peu fiables.
Statistiques Clés :
| Type de Donnée | Source / Étude | Chiffres Marquants | | --- | --- | --- | | Victimation (Femmes) | Données internationales convergentes | Au moins 1 femme sur 5 à 1 sur 3 sera victime de violence sexuelle au cours de sa vie. | | Victimation (Hommes) | Étude de Briding (USA) | 2 % des hommes sont victimes de viol au cours de leur vie. | | Victimation (Mineurs) | Publication The Lancet (Mai 2024) | 1 femme sur 5 et 1 homme sur 7 sont victimes avant l'âge de 18 ans. | | Perpétration | Données internationales | 20 à 40 % des personnes reconnaissent avoir commis au moins un fait de violence sexuelle non judiciarisé depuis l'âge de 14 ans. | | Perpétration (Étudiants) | Études françaises (Thèse M. Escargel) | En moyenne, 40 % des étudiants et 18 % des étudiantes rapportent avoir commis un fait de violence sexuelle. |
Stratégies de Perpétrations : Il est souligné que, contrairement aux représentations communes, la force physique ou l'usage d'une arme restent des stratégies peu fréquentes.
La majorité des violences sexuelles résultent de :
• Manipulation et pressions verbales.
• Intoxication de la victime (alcool ou drogues).
2.2. L'Attrition du Système Judiciaire et la Limite des Modèles Explicatifs
Un point crucial de la conférence est l'écart massif entre le nombre de violences commises et les condamnations.
• Le "Rapport de 1/10" : La littérature scientifique estime que seule 1 situation de violence sexuelle sur 10 donne lieu à un dépôt de plainte, et que seule 1 plainte sur 10 aboutit à une condamnation.
• Données Françaises (2012-2021) : Un rapport de l'Institut des politiques publiques (2024) révèle que 86 % des plaintes pour violences sexuelles sont classées sans suite.
Parmi les 14 % de suspects finalement jugés, seuls 13 % sont reconnus coupables.
Cette réalité a une implication majeure : la grande majorité des auteurs de violences sexuelles ne sont jamais identifiés par les autorités.
Par conséquent, les modèles théoriques dominants en psychocriminologie, élaborés quasi exclusivement à partir de l'étude d'auteurs condamnés et incarcérés, ne renseignent que sur une minorité très spécifique et probablement non représentative.
Il existe un risque de postuler à tort qu'une catégorie juridique (l'infraction sexuelle) correspond à un ensemble homogène de processus psychologiques.
2.3. Le Rôle de la Cognition Sociale
Dans l'étude des violences sexuelles, la cognition sociale est un élément central. Quatre composantes sont principalement étudiées :
1. L'empathie et/ou la théorie de l'esprit.
2. La reconnaissance des émotions.
3. La régulation émotionnelle et l'autorégulation.
4. Les distorsions cognitives : Croyances qui servent à rationaliser, minimiser ou justifier l'agression (ex: les mythes sur le viol).
Cependant, les recherches offrent un portrait nuancé, supportant "partiellement et parfois très partiellement" le rôle de ces cognitions.
2.4. Le Cadre Théorique du Traitement de l'Information Sociale (TIS)
Pour mieux comprendre les mécanismes sous-jacents, les modèles de TIS (ex: Crick & Dodge) sont proposés comme un cadre de référence pertinent.
Ces modèles décrivent tout comportement social comme le résultat d'une séquence de six étapes mentales :
1. Encodage des indices (internes et externes).
2. Interprétation des indices (attribution d'intentions).
3. Clarification des objectifs.
4. Génération de réponses alternatives.
5. Évaluation et choix d'une réponse.
6. Mise en œuvre comportementale.
Un élément central de ces modèles est le rôle des structures de connaissance (schémas, théories implicites, mythes), qui sont des contenus cognitifs enracinés en mémoire. Issues des expériences de vie et des normes socioculturelles, elles influencent chaque étape du traitement de l'information, notamment via des biais attentionnels et interprétatifs.
2.5. Résultats des Recherches et Études Empiriques
La conférence a présenté plusieurs résultats de recherche menées au-delà des seules populations judiciarisées.
• Régulation émotionnelle : La suppression expressive apparaît comme un facteur de risque, tandis que la réévaluation cognitive serait un mécanisme de protection.
• Empathie : Contrairement à une idée reçue, l'empathie n'est généralement pas associée aux violences sexuelles en tant que facteur de risque, mais plutôt comme un facteur de protection (effet tampon ou "buffering").
• Structures de connaissance : L'adhésion aux mythes du viol (croyances erronées sur les victimes, les auteurs et les violences) est un facteur clé.
Une méta-analyse récente confirme une relation d'effet modérée, stable depuis 30 ans, et qui semble se renforcer avec l'âge si les croyances ne sont pas déconstruites.
• Perception des intentions sexuelles : Une étude utilisant des stimuli vidéo a montré que les hommes ne présentent pas un déficit général, mais une difficulté spécifique à reconnaître une absence d'intérêt.
Le taux de reconnaissance de cette intention n'est pas statistiquement différent du hasard.
• Effet de l'alcool : Une étude expérimentale a démontré que la consommation d'alcool (jusqu'à 1g/L) dégrade la capacité à percevoir l'absence de consentement uniquement chez les individus qui adhèrent déjà fortement aux mythes du viol.
L'alcool n'a aucun effet chez ceux qui n'adhèrent pas à ces mythes, remettant en cause son utilisation comme excuse.
2.6. Implications et Perspectives
Les résultats présentés ont des implications importantes pour la prévention et la prise en charge.
1. Nécessité d'une approche globale : Il est crucial d'étendre l'étude de la cognition sociale aux populations non judiciarisées pour comprendre les mécanismes à l'œuvre dans la majorité des cas.
2. Cibler les structures de connaissance : Les programmes de prévention primaire et secondaire doivent se concentrer sur la déconstruction des structures de connaissance (stéréotypes de genre, mythes sur le viol).
Ces programmes sont efficaces : une méta-analyse montre une réduction des violences sexuelles de 17 % chez les adolescents.
3. Prévention précoce : Le travail peut commencer très tôt, avant même de parler de sexualité, en agissant sur les stéréotypes de genre dès l'école primaire.
4. Évaluation multidimensionnelle : La prise en charge des auteurs doit reposer sur une évaluation fine et multidimensionnelle de la cognition sociale.
Le programme BOAT, porté par le CHU de Montpellier, est cité comme une initiative ambitieuse adoptant une approche populationnelle pour articuler prévention des violences et promotion de la santé sexuelle.
3. Synthèse de la Session de Questions-Réponses
• Différence entre "crime" et "violence" : Le terme "violence sexuelle" est utilisé dans la recherche pour inclure les comportements non judiciarisés.
L'étude des profils psychopathologiques des victimes est également un champ de recherche actif.
• Différences entre auteurs judiciarisés et non judiciarisés :
L'hypothèse d'une différence se base sur le fait que les cas poursuivis sont souvent ceux où les preuves sont plus fortes ou les faits perçus comme plus graves, ce qui pourrait correspondre à des caractéristiques criminologiques et des profils psychologiques distincts.
• Biais de désirabilité sociale : C'est une limite reconnue des études basées sur des questionnaires autorapportés.
Ce biais est contrôlé statistiquement, mais la limite demeure. Cependant, le fait que 20 à 40 % des personnes rapportent des actes de perpétration malgré ce biais est en soi significatif.
• Différence entre "mythe du viol" et "culture du viol" : Les deux concepts ont un recouvrement conceptuel très fort. "Mythe du viol" est un terme historiquement utilisé dans le champ de la psychologie, tandis que "culture du viol" est davantage employé en sociologie.
-