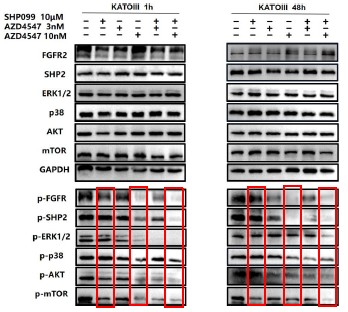
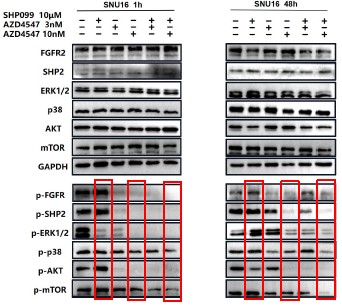
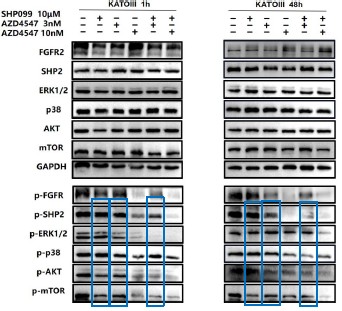
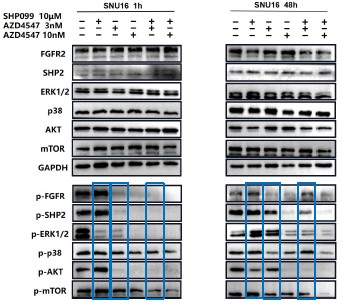
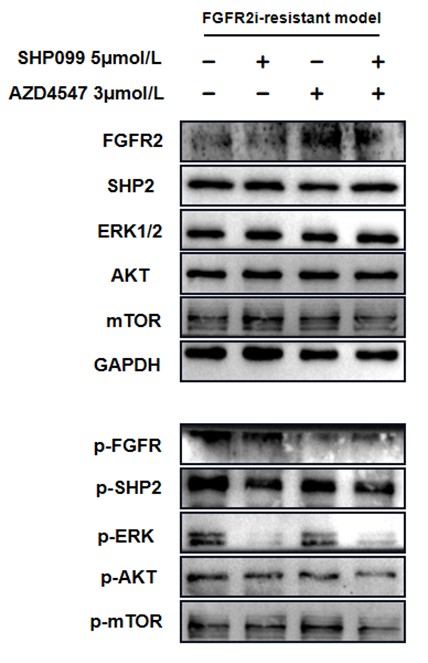
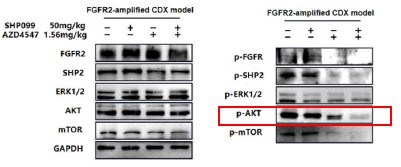
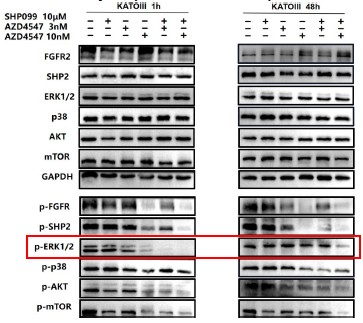
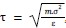1. General Statements [optional]
*We thank the reviewers for their insightful and constructive comments, which have substantially strengthened the manuscript. We have addressed all concerns and replaced the previous non-quantitative RNA-seq analysis with a new analysis that allowed for quantitative assessment. We were encouraged to find that the revised analysis not only confirmed our original observations but also reinforced and extended our conclusions. *
2. Point-by-point description of the revisions
Reviewer #1
Significance
Comment 1: At its current stage, this work represents a robust resource for molecular parasitology research programs, paving the way for mechanistic studies on multilayered gene expression control and it would benefit from experimental evidence for some of the claims concerning the in silico regulatory networks. Terms like "regulons", "recursive feedback loop" are employed without solid confirmation or extensive literature support. In my view, the most relevant contribution of this study is centered in the direct association between proteasome-dependent degradation and Leishmania differentiation.
__Response: __We thank the reviewer to acknowledge the impact of our work as a robust resource for further mechanistic studies. We agree that the new concepts emerging from our multilayered analysis should be experimentally assessed. However, given the scope of our analysis (i.e. a complete systems-level analysis of bona fide, hamster-isolated L. donovani amastigotes and derived promastigotes) and the amount of data presented in the current manuscript, such functional genetic analysis will merit an independent, in-depth investigation. The current version has been very much toned down and modified to emphasize the impact of our work as a powerful new resource for downstream functional analyses.
Evidence, reproducibility and clarity
Comment 1: The narrative becomes somewhat diffuse with the shift to putative multilevel regulatory networks, which would benefit from further experimental validation.
Response: We agree with the reviewer and toned down the general discussion while suggesting putative multilevel regulatory networks for follow-up, mechanistic analyses. We now emphasize those networks for which evidence in trypanosomatids and other organisms has been published. Experimental validation of some of these regulatory networks is outside the scope of our manuscript and will be pursued as part of independent investigations.
Major issues
Comment 1: Fig.1D suggests a significant portion of the SNPs are exclusive, with a frequency of zero in one of the two stages. Were only the heterozygous and minor alleles plotted in Fig.1D, since frequencies close to 1 are barely observed? Is the same true in Sup Fig. S2B? Why do chrs 4 and 33 show unusual patterns in S2B?
__Response: __We thank the reviewer for this observation. The SNPs exclusive to either one or the other stage are likely the result of the 10% cutoff we use for this kind of analysis (eliminating SNPs that lack sufficient support, i.e. less than 10 reads). Due to bottle neck events (such as in vitro culture or stage differentiation), many low frequency SNPs are either 'lost' (filtered out) or 'gained' (passing the 10% cutoff) between the ama and pro samples. All SNPs above 10% were plotted. The absence of SNPs at 100% is one of the hallmarks of the Ld1S L. donovani strain we are using. Instead, these parasites show a majority of SNPs at a frequency of around 50%, which is likely a sign of a previous hybridization event. Chr 4 and chr 33 show a very low SNP density, most likely as they went through a transient monosomy at one moment of their evolutionary history, causing loss of heterozygosity. We now explain these facts in the figure legend.
Comment 2: Chr26 revealed a striking contrasting gene coverage between H-1 and the other two samples. While a peak is observed for H-1 in the middle of this chr, the other two show a decrease in coverage. Is there any correlation with the transcriptomic/proteomic findings?
Response: This analysis is based on normalized median read depth, taking somy variations into account. This is now more clearly specified in the figure legend. We do not see any significant expression changes that would correlate with the observed (minor) read depth changes. As indicated in the legend, we do not consider such small fluctuations (less than +/- 1,5 fold) as significant. The reversal of the signal for chr 26 sample H1 eludes us (but again, these fluctuations are minor and not observed at mRNA level).
Comment 3: The term "regulon" is used somewhat loosely in many parts of the text. Evidence of co-transcriptomic patterns alone does not necessarily demonstrate control by a common regulator (e.g., RNA-binding protein), and therefore does not fulfill the strict definition of a regulon. It should be clear whether the authors are highlighting potential multiple inferred regulons within a list of genes or not. Maybe functional/ gene module/cluster would be more appropriate terms.
Response: We thank the reviewer for this important comment. We replaced 'regulon' throughout the manuscript by 'co-regulated, functional gene clusters' (or similar).
Comment 4: It is unclear whether the findings in Fig.3E are based on previous analysis of stage-specific rRNA modifications or inferred from the pre-snoRNA transcriptomic data in the current work or something else. I struggle to find the significance of presenting this here.
__Response: __We thank the reviewer for this comment. Yes, these data show stage-specific rRNA modifications based on previous analyses that mapped stage-specific differences of pseudouridine (Y) (Rajan et al., Cell Reports 2023, DOI: 10.1016/j.celrep.2024.114203) and 2'-O-modifications (Rajan et al., Nature Com, in revision) by various RNA-seq analyses and cryoEM. This figure has been modified in the revised version to consider the identification of stage-regulated snoRNAs in our new and statistically robust RNA-seq analysis. These data are shown to further support the existence of stage-regulated ribosomes that may control mRNA translatability, as suggested by the enriched GO terms 'ribosome biogenesis', 'rRNA processing' and 'RNA methylation' shown in Figure 2. We better integrated these analyses by moving the panels from Figure 3 to Figure 2.
Comment 5: The protein turnover analysis is missing the critical confirmation of the expected lactacystin activity on the proteasome in both ama and pro. A straightforward experiment would be an anti-polyUb western blotting using a low concentration SDS-PAGE or a proteasome activity assay on total extracts.
Response: We thank the reviewer for this comment and have now included an anti-polyUb Western blot analysis (see Fig S7).
Comment 6: The viability tests upon lactacystin treatment need a positive control for the PI and the YoPro staining (i.e., permeabilized or heat-killed promastigotes).
Response: This control is now included in Fig S7 and we have added the corresponding description to the text.
Comment 7: I found that the section on regulatory networks was somewhat speculative and less focused. Several of the associated conclusions are, in some parts, overstated, such as in "uncovered a similar recursive feedback loop" (line 566) or "unprecedented insight into the regulatory landscape" (line 643). It would be important to provide some form of direct evidence supporting a functional connection between phosphorylation/ubiquitination, ribosome biogenesis/proteins and gene expression regulation.
Response: We agree with the reviewer and have considerably toned down our statements. Functional analyses to investigate and validate some of the shown network interactions are planned for the near future and will be published separately.
Minor issues
1) The ordinal transition words "First,"/"Second," are used too frequently in explanatory sections. I noted six instances. I suggest replacing or rephrasing some to improve flow.
Response: Rectified, thanks for pointing this out.
2) Ln 168: Unformatted citations were given for the Python packages used in the study.
Response: Rectified, thanks for pointing this out.
3) Fig.1D: "SNP frequency" is the preferred term in English.
Response: Corrected.
4) Fig.2A: not sure what "counts}1" mean.
__Response: __This figure has been replaced.
5) Ln 685: "Transcripts with FC 0.01 are represented by black dots" -> This sentence is inaccurate. The intended wording might be: "Transcripts with FC 0.01 are represented by black dots"
Response: We thank the reviewer and corrected accordingly.
6) Ln 698: Same as ln 685 mentioned above.
Response: We thank the reviewer and corrected accordingly.
7) Fig.2B and elsewhere: The legend key for the GO term enrichment is a bit confusing. It seems like the color scales represent the adj. p-values, but the legend keys read "Cluster efficiency" and "Enrichment score", while those values are actually represented by each bar length. Does light blue correspond to a max value of 0.05 in one scale, and dark blue to a max value of 10-7 in the other scale?
Response: This was corrected in the figure and the legends were updated accordingly.
8) Sup Figure S3A and S4A: The hierarchical clustering dendrograms are barely visible in the heatmaps.
Response: Thanks for the comment. Figure S3 was removed and replaced by a hierarchical clustering and a PCA plot.
9) S3A Legend: The following sentence sounds a bit awkward: "Rows and columns have been re-ordered thanks to a hierarchical clustering". I suggest switching "thanks to a hierarchical clustering" to "based on hierarchical clustering".
Response: This figure was removed and the legend modified.
10) Fig.5D: The font size everywhere except the legend key is too small. In addition, on the left panel, gene product names are given as a column, while on the right, the names are shown below the GeneIDs. Consistency would make it clearer.
Response: Thank you, this is now rectified. To ensue readability, we reduced the number of shown protein kinase examples.
Reviewer #2
Evidence, reproducibility and clarity
Comment 1: In the absence of riboprofiling the authors return to the RNA-seq to assess the levels of pre-Sno RNA (the role of the could be more explicitly stated).
Response: We thank the reviewer for this comment. We moved the snoRNA analysis from Fig 3 to Fig 2 (see also the similar comment of reviewer 1), which better integrates and justifies this analysis. Based on the new and statistically robust RNA-seq analysis, the volcano plot showing differential snoRNA expression and possible ribosome modification has been adjusted (Figures 2C and D).
__Comment 2: __The authors provide a clear and comprehensive description of the data at each stage of the results and this in woven together in the discussion allowing hypotheses to be formed on the potential regulatory and signalling pathways that control the differentiation of amastigotes to promastigotes. Given the amount and breadth of data presented the authors are able to present a high-level assessment of the processes that form feedback loops and/or intersectional signalling, but specific examples are not picked out for deeper validation or exploration.
__Response: __We thank the reviewer to acknowledge the amount and breadth of data presented. As indicated above (see responses to reviewer 1), mechanistic studies will be conducted in the near future to validate some of the regulatory interactions. These will be subject of separate publications. As noted above (response to reviewer 1), we toned down the general discussion, suggest follow-up mechanistic analyses and emphasize those networks for which evidence in trypanosomatids and other organisms has been published.
__
__
Major comments:
Are the claims and the conclusions supported by the data or do they require additional experiments or analyses to support them?
Comment 1: As I have understood it from the description in the text, and in Data Table 4, the RNA-seq element of the work has only been conducted using two replicates. If this is the case, it would substantially undermine the RNA-seq and the inferences drawn from it. Minimum replicates required for inferential analysis is 3 bio-replicates and potentially up to 6 or 12. It may be necessary for the authors to repeat this for the RNA-seq to carry enough weight to support their arguments. (PMID: 27022035)
Response: We agree with the reviewer and conducted a new RNA-seq analysis with 4 independent biological replicates of spleen-purified amastigotes and derived promastigotes. Given the robustness of the stage-specific transcriptome, and the legal constrains associated with the use of animals, we chose to limit the number of replicates to the necessary. We thank the reviewer for this important comment, and the new data not only confirm the previous one (providing a high level of robustness to our data) but allowed us to increase the number of identified stage-regulated snoRNAs, thus further supporting a possible role of ribosome modification in Leishmania stage development.
Comment 2: There are several examples that are given as reciprocal or recursive signalling pathways, but these are not followed up with independent, orthogonal techniques. I think the paper currently forms a great resource to pursue these interesting signalling interactions and is certainly more than just a catalogue of modifications, but to take it to the next level ideally a novel signalling interaction would be demonstrated using an orthogonal approach. Perhaps the regulation of the ribosomes could have been explored further (same teams recently published related work on this). Or perhaps more interestingly, a novel target(s) from the ubiquitinated protein kinases could have been explored further; for example making precision mutants that lack the ubiquitination or phosphorylation sites - does this abrogate differentiation?
Response: We agree with the reviewer that the paper currently forms a great resource. In-depth molecular analysis investigating key signaling pathways and regulatory interactions are outside the scope of the current multilevel systems analysis but will be pursued in independent investigations.
Comment 3: I found the use of lactacystin a bit curious as there are more potent and specific inhibitors of Leishmania proteasomes e.g. LXE-408. This could be clarified in the write-up (See below).
__Response: __We thank the reviewer for this comment. We opted for the highly specific and irreversible proteasome inhibitor lactacystin that has been previously applied to study the Leishmania proteasome (PMID: 15234661) rather than the typanosomatid-specific drug candidate LXE408 as the strong cytotoxic effect of the latter makes it difficult to distinguish between direct effects on protein turnover and secondary effects resulting from cell death, limiting its utility for dissecting proteasome function in living parasites. We have added this information in the Results section.
Comment 4: If it is the case that only 2 replicates of the RNA-Seq have been performed it really is not the accepted level of replication for the field. Most studies use a minimum of 3 bioreplicates and even a minimum of 6 is recommended by independent assessment of DESeq2.
__Response: __See response to comment 1 above.
Comment 5: As far as I could see, the cell viability assay does not include a positive control that shows it is capable of detecting cytotoxic effects of inhibitors. Add treatment showing that it can differentiate cytostatic vs cytotoxic compound.
__Response: __This control has now been added to Fig S7.
If you have constructive further reaching suggestions that could significantly improve the study but would open new lines of investigations, please label them as "OPTIONAL".
Are the suggested experiments realistic in terms of time and resources? It would help if you could add an estimated time investment for substantial experiments.
Comment 6: It is realistic for the authors to validate the cell viability assay. If the RNA-seq needs to be repeated then this would be a substantial involvement.
Response: Redoing the RNA-seq analysis was entirely feasible and very much improved the robustness of our results.
Are the data and the methods presented in such a way that they can be reproduced?
Comment 7: All the methods are written to a good level of detail. The sample prep, acquisition and data analysis of the protein mass spectrometry contained a high level of detail in a supplemental section. The authors should be more explicit about the amount of replication at each stage, as in parts of the manuscript this was quite unclear.
Response: We thank the reviewer for this comment and explicitly state the number of replicates in Methods, Results and Figure legends for all analyses. The number of replicates for each analysis is further shown in the overview Figure S1.
Are the experiments adequately replicated and statistical analysis adequate?
Comment 8: Unless I have misunderstood the manuscript, I believe the RNA-seq dataset is underpowered according to the number of replicates the authors report in the text.
Response: See response to comment 1 above.
Comment 9: Looking at Figure 1 and S1 and Data Table 4 to show the sample workflow I was surprised to see that the RNA-seq only used 2 replicates. The authors do show concordance between the individual biological replicates, but I would consider that only having 2 is problematic here, especially given the importance placed on the mRNA levels and linkage in this study. This would constitute a major weakness of the study, given that it is the basis for a crucial comparison between the RNA and protein levels.
Response: We agree and have repeated the RNAseq analysis using four independent biological replicates - see response to comment 1.
Comment 10: It also wasn't clear to me how many replicates were performed at each condition for the lactacystin treatment experiment - can the authors please state this clearly in the text, it looks like 4 replicates from Figure S1 and Data Table 8.
Response: Indeed, we did 4 replicates. This is now clarified in Methods, Results and Figure legends and shown in Figure S1.
Comment 11: Four replicates are used for the phosphoproteomics data set, which is probably ok, but other researchers have used a minimum of 5 in phosphoproteomics experiments to deal with the high level of variability that can often be observed with low abundance proteins & modifications. The method for the phosphoproteomics analysis suggests that a detection of a phosphosite in 1 sample (also with a localisation probability of >0.75) was required for then using missing value imputation of other samples. This seems like a low threshold for inclusion of that phosphosite for further relative quantitative analysis. For example, Geoghegan et al (2022) (PMID: 36437406) used a much more stringent threshold of greater than or equal to 2 missing values from 5 replicates as an exclusion criteria for detected phoshopeptides. Please correct me if I misunderstood the data processing, but as it stands the imputation of so many missing values (potentially 3 of 4 per sample category) could be reducing the quality of this analysis.
Response: We thank the reviewer for this remark and for highlighting best practices in phosphoproteomics data analysis. Unlike other studies that use cultured parasites and thus have access to unlimited amounts, our study employs bona fide amastigotes isolated from infected hamster spleens. In France, the use of animals is tightly controlled and only the minimal number of animals to obtain statistically significant results is tolerated (and necessary to obtain permission to conduct animal experiments).
Regarding the number of biological replicates, we would like to emphasize that the use of four biological replicates is fully acceptable and used in quantitative proteomics and phosphoproteomics, particularly when combined with high-quality LC-MS/MS data and stringent peptide-level filtering. While some studies indeed employ five or more replicates, this is not a strict requirement, and many high-impact phosphoproteomics studies have successfully relied on four replicates when experimental quality and depth are high. In the present study, we adopted a discovery-oriented approach, aimed at detecting as many confidently identified phosphopeptides as possible. The consistency between replicates, combined with the depth of coverage and signal quality, indicates that four replicates are adequate for both the global proteome and the phosphoproteome in this context. Importantly, the quality of the MS data in this study is supported by (i) a high number of confidently identified peptides and phosphopeptides (identification FDR0.75), and (iii) reproducible quantitative profiles across replicates. Notably, most of the identified phosphopeptides are quantified in at least two replicates within a given condition (between 73.2% and 83.4% of all the identified phosphopeptides among replicates of the same condition).
Regarding missing value imputation, we appreciate that our initial description may have been unclear and we have revised the Methods to avoid misunderstanding. Phosphosites were only considered if detected with high confidence (identification FDR0.75) in at least one replicate. This criterion was chosen to retain biologically relevant, low-abundance phosphosites, which are more difficult to identify and are often stochastically sampled in phosphoproteomics datasets. For statistical analyses, missing values within a given condition were imputed with a well-established algorithm (MLE) only when at least one observed value was present in that condition. Notably, they were replaced by values in the neighborhood of the observed intensities, rather than by globally low, noise-like values.
We agree that more stringent exclusion rules, such as those used by Geoghegan et al. (2022), are appropriate in some contexts. However, there is no universally accepted standard for missingness thresholds in phosphoproteomics, and different strategies reflect trade-offs between sensitivity and stringency. In our discovery-oriented approach, we deliberately prioritized biological coverage while maintaining data quality. Our main conclusions are supported by coherent biological patterns, rather than by isolated phosphosite measurements.
Comment 12: For the metabolomics analysis it looks like 2 amastigote samples were compared against 4 promastigote samples. Why not triplicates of each?
Response: We thank the reviewer for noticing this point. It is an error in the figure file (Sup figure S1). Four biological replicates of splenic amastigotes were prepared (H130-1, H130-2, H133-1 and H133-2). Amastigotes from 2 biological replicates (H131-1 and H131-2) were seeded for differentiation into promastigotes in 4 flasks (2 per biological replicate) that were collected at passage 2. We have updated the figure file accordingly.
Minor comments:
__
__Specific experimental issues that are easily addressable.
Are prior studies referenced appropriately?
*
*Comment 1: Yes
Are the text and figures clear and accurate?
*
*Comment 2: The write up is clear, with the data presented coherently for each method. The analyses that link everything together are well discussed. The figures are mostly clear (see below) and are well described in the legends. There is good use of graphics to explain the experimental designs and sample names - although it is unclear if technical replicates are defined in these figures.
Response: We thank the reviewer for these positive comments. We now included the information on replicates in the overview figure (Figure S1).
Do you have suggestions that would help the authors improve the presentation of their data and conclusions?
Comment 3: As I have understood it, the authors have calculated the "phosphostoichiometry" using the ratio of change in the phosphopeptide to the ratio of the change in total protein level changes. This is detailed in the supplemental method (see below). Whilst this has normalised the data, it has not resulted in an occupancy or stoichiometry measurement, which are measured between 0-1 (0% to 100%). The normalisation has probably been sufficient and useful for this analysis, but this section needs to be re-worded to be more precise about what the authors are doing and presenting. These concepts are nicely reviewed by Muneer, Chen & Chen 2025 (PMID: 39696887) who reference seminal papers on determination of phosphopeptide occupancy - and may be a good place to start. An alternative phrase should be used to describe the ratio of ratios calculated here, not phosphostoichiometry.
Response: We thank the reviewer for this insightful comment and fully agree with the conceptual distinction raised. The reviewer is correct that the approach used in this study does not measure absolute phosphosite occupancy or stoichiometry, which would indeed require dedicated experimental strategies and would yield values bounded between 0 and 1 (0-100%). Instead, we calculated a normalized phosphorylation change, defined as the ratio of the change in phosphopeptide abundance relative to the change in the corresponding total protein abundance (a ratio-of-ratios approach - see doi :10.1007/978-1-0716-1967-4_12), and we tested whether this normalized phosphorylation change differed significantly from zero. This normalization approach is comparable to those previously published in the « Experimental Design and Statistical Analysis of the Proteome and the Phosphoproteome » section of the following paper (DOI: 10.1016/j.mcpro.2022.100428).
Our intention was to account for protein-level regulation and thereby better isolate changes in phosphorylation dynamics. While this normalization is informative and appropriate for the biological questions addressed here, we agree that the term "phosphostoichiometry" is imprecise and not correct in this context.
In response, we (i) replaced the term "phosphostoichiometry" throughout the manuscript with a more accurate description, such as "normalized phosphorylation level", or "relative phosphorylation change normalized to protein abundance", and (ii) revised the corresponding Methods and Results text to clearly state that absolute occupancy was not measured.
This rewording will improve conceptual accuracy without altering the validity or interpretation of the results.
Comment 4: From the authors methods describing the ratio comparison approach:
"Another statistical test was performed in a second step: a contrasted t-test was performed to compare the variation in abundance of each modified peptide to the one of its parent unmodified protein using the limma R package {Ritchie, 2015; Smyth, 2005}. This second test allows determining whether the fold-change of a phosphorylated peptide between two conditions is significantly different from the one of its parent and unmodified protein (paragraph 3.9 in Giai Gianetto et al 2023). An adaptive Benjamini-Hochberg procedure was applied on the resulting p-values thanks to the adjust.p function of R package cp4p {Giai Gianetto, 2016} using the Pounds et al {Pounds, 2006} method to control the False Discovery Rate level."
Response: The references have been formatted.
Comment 5: Several aspects of the figures that contain STRING networks are quite useful, particularly the way colour around the circle of each node to denote different molecular functions/biological processes. However, some have descended into "hairball" plots that convey little useful information that would be equally conveyed in a table, for example. Added to this, the points on the figure are identified by gene IDs which, while clear and incontrovertible, are lacking human readability. I suggest that protein name could be included here too.
Response: We thank the reviewer for this comment but for readability we opted to keep the figure as is. We now refer to Tables 8, 9, and 12 that allow the reader to link gene IDs to protein name and annotation (if available).
Comment 6: It is also not clear what STRING data is being plotted here, what are the edges indicating - physical interactions proven in Leishmania, or inferred interactions mapped on from other organisms? Perhaps as supplemental data provide the Cytoscape network files so readers can explore the networks themselves?
Response: We thank the reviewer for this comment. While the STRING plugin in Cytoscape enables integrated network-based analyses, it represents protein-protein associations as a single edge per protein pair derived from the combined confidence score. Consequently, the specific contribution of individual evidence channels (e.g. experimental evidence, curated databases, co-expression, or text mining) cannot be disentangled within this framework. However, this representation was considered appropriate for the present study, which focused on global network topology and functional enrichment rather than on the interpretation of individual interaction types. The information on stringency has been added to the Methods section and the Figure legends (adding the information on confidence score cutoff).
We decided not to submit the Cytoscape files as they were generated with previous versions of Cytoscape and the STRING plugin. Based on the differential abundance data shown in the tables it will be very easy to recreate these networks with the new versions for any follow up study.
Comment 7: The title of columns in table S10 panel A are written in French, which will be ok for many people particularly those familiar with proteomics software outputs, but everything else is in English so perhaps those titles could be made consistent.
__Response: __We apologize and have translated the text in English.
Comment 8: I would suggest that the authors provide a table that has all the gene IDs of the Ld1S2D strain and the orthologs for at least one other species that is in TriTrypDB. This would make it easy to interrogate the data and make it a more useful resource for the community who work on different strains and species of Leishmania. Although this data is available it is a supplemental material file in a previous paper (Bussotti et al PNAS 2021) and not easy to find.
Response: We thank the reviewer for this very useful suggestion and have added this table (Table S13).
Comment 9: Figure 5b - from the legend it is not clear where the confidence values were derived in this analysis, although this is explained in the supplemental method. Perhaps the legend can be a bit clearer.
Response: We have the following statement to the legend: 'Confidence values were derived as described in Supplementary Methods'.
Comment 10: Can the authors discuss why lactacystin was used? While this is a commonly used proteasome inhibitor in mammalian cells there is concern that it can inhibit other proteases. At the concentrations (10 µM) the authors used there are off-target effects in Leishmania, certainly the inhibition of a carboxypeptidase (PMID: 35910377) and potentially cathepsins as is observed in other systems (PMID: 9175783). There is a specific inhibitor of the Leishmania proteasome LXE-408 (PMID: 32667203), which comes closer to fulfilling the SGC criteria (PMID: 26196764) for a chemical probe - why not use this. Does lactacystin inhibit a different aspect of proteasome activity compared to LXE-408?
Response: We have add the following justification to the results section (see also response above to comment 3 for reviewer 2): We chose the highly specific and irreversible proteasome inhibitor lactacystin over the typanosomatid-specific, reversible drug candidate LXE408 as the latter's potent cytotoxicity can confound direct effects on protein turnover with secondary consequences of cell death, limiting its utility for dissecting proteasome function in living parasites.
Comment 11: The application of lactacystin is changing the abundance of a multitude of proteins but no precision follow up is done to identify if those proteins are necessary and/or sufficient from driving/blocking differentiation. This could be tested using precision edited lines that are unable to be ubiquitinated? There is a lack of direct evidence that the proteins protected from degradation by lactacystin are ubiquitinated? Perhaps some of these could be tagged and IP'd then probed for ubiquitin signal. Di-Gly proteomics to reveal ubiquitinated proteins? These suggestions should be considered as OPTIONAL experiments in the relevant section above.
Response: We very much appreciate these very interesting suggestions, which we will be considered for ongoing follow-up studies.
Comment 12: In the data availability RNA-seq section the text for the GEO link is : (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc= GSE227637) but the embedded link takes me to (https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE165615) which is data for another, different study. Also, the link to the GEO site for the DNA seq isn't working and manual searches with the archive number (BioProject PRJNA1231373 ) does not appear to find anything. The IDs for the mass spec data PRIDE/ProteomeXchange don't seem to bring up available datasets: PXD035697 and PXD035698
Response: The links have now been rectified and validated. For those data that are still under quarantine, here is the login information: To access the data:
DNAseq data: https://dataview.ncbi.nlm.nih.gov/object/PRJNA1231373?reviewer=6qt24dd7f475838rbqfn228d0
RNAseq data:
https://www.ebi.ac.uk/biostudies/ArrayExpress/studies/E-MTAB-16528?key=65367b55-d77f-4c06-b4bd-bc10f2dc0b14
Proteomic data: http://www.ebi.ac.uk/pride
__Username: __reviewer_pxd035698@ebi.ac.uk
__Password: __gOIcRx0g
Phosphoproteomic data: http://www.ebi.ac.uk/pride
__Username: __reviewer_pxd035697@ebi.ac.uk
__Password: __7GWtBmvx
Significance
Provide contextual information to readers (editors and researchers) about the novelty of the study, its value for the field and the communities that might be interested.
The following aspects are important:
*
General assessment: provide a summary of the strengths and limitations of the study. What are the strongest and most important aspects? What aspects of the study should be improved or could be developed?*
Strengths:
Comment 1: The molecular pathways that regulate Leishmania life-stage transitions are still poorly understood, with many approaches exploring single proteins/RNAs etc in a reductionist manner. This paper takes a systems-scale approach and does a good job of integrating the disparate -omics datasets to generate hypotheses of the intersections of regulatory proteins that are associated with life-cycle progression.
Response: We thank the reviewer for this positive assessment of our work.
Comment 2: The differentiation step studied is from amastigote to promastigote. I am not aware that this has been studied before using phosphoproteomics. The use of the hamster derived amastigotes is a major strength. While a difficult/less common model, the use of hamsters permits the extraction of parasites that are host adapted and represent "normal", host-adapted Leishmania ploidy, the promastigote experiments are performed at a low passage number. This is a strength or the work as it reduces the interference of the biological plasticity of Leishmania when it is cultured outside the host.
Response: We thank the reviewer for the acknowledgment of our relevant hamster system, for which we face many challenges (financial, ethical, administrative as protocols need to be approved by the French government).
Limitations:
__
__Comment 1: Potential lack of appropriate replication (see above).
Response: See response to comment 1.
Comment 2: Lack of follow up/validation of a novel signalling interaction identified from the systems-wide approach. There is a lack of assessment of whether a single signalling cascade is driving the differentiation or these are all parallel, requisite pathways. The authors state the differentiation is not driven by a single master regulator, but I am not sure there is adequate evidence to rule this in or out.
Response: See response to comment 2 above.
Advance: compare the study to the closest related results in the literature or highlight results reported for the first time to your knowledge; does the study extend the knowledge in the field and in which way? Describe the nature of the advance and the resulting insights (for example: conceptual, technical, clinical, mechanistic, functional,...).
Comment 3: The study applies well established techniques without any particular technical step-change. The application of large-scale multi-omics techniques and integrated comparisons of the different experimental workflows allow a synthesis of data that is a step forward from that existing in the previous Leishmania literature. It allows the generation of new hypotheses about specific regulatory pathways and crosstalk that potentially drive, or are at least active, during amastigote>promastigote differentiation.
Response: We thank the reviewer for these positive comments.
*Audience: describe the type of audience ("specialized", "broad", "basic research", "translational/clinical", etc...) that will be interested or influenced by this research; how will this research be used by others; will it be of interest beyond the specific field?
*
This manuscript will have primary interest to those researchers studying the molecular and cell biology of Leishmania and other kinetoplastid parasites. The approaches used are quite standard (so not so interesting in terms of methods development etc.) and given the specific quirks of Leishmania biology it may not be that relevant to those working more broadly in parasites from different clades/phyla, or those working on opisthokont systems- yeast, humans etc. Other Leishmania focused groups will surely cherry-pick interesting hits from this dataset to advance their studies, so this dataset will form a valuable reference point for hypothesis generation.
Response: We thank the reviewer for this assessment and agree that our data sets will be very valuable for us and other teams to generate hypotheses for follow-up studies.
Please define your field of expertise with a few keywords to help the authors contextualize your point of view. Indicate if there are any parts of the paper that you do not have sufficient expertise to evaluate.
Relevant expertise: Trypanosoma & Leishmania molecular & cell biology, RNA-seq, proteomics, transcriptional/epigenetic regulation, protein kinases - some experience of UPS system.
I have not provided comment on the metabolomics as it is outside my core expertise. However, I can see it was performed at one of the leading parasitology metabolomics labs.
Response: We thank the reviewer for sharing expertise, investing time and intelligence in the assessment of our manuscript, and the highly constructive criticisms provided.
Reviewer #3 (Evidence, reproducibility and clarity (Required)):
__Summary:
__The study presents a comprehensive multi-omics investigation of Leishmania differentiation, combining genomic, transcriptomic, proteomic, phospho-proteomic and metabolomic data. The authors aim to uncover mechanisms of post-transcriptional and post-translational regulation that drive the stage-specific biology of L. donovani. The authors provide a detailed characterization of transcriptomic, proteomic, and phospho-proteomic changes between life stages, and dissect the relative contributions of mRNA abundance and protein degradation to stage-specific protein expression. Notably, the study is accompanied by comprehensive supplementary materials for each molecular layer and provides public access to both raw and processed data, enhancing transparency and reproducibility. While the data are rich and compelling, several mechanistic interpretations (e.g., "feedback loops," "recursive networks," "signaling cascades") are overstated. Similarly, the classification of gene sets as "regulons" is not adequately supported, as no common regulatory factor has been identified and only a single condition change (amastigote to promastigote) was assessed.
__Response: __We thank the reviewer for these comments and have corrected the manuscript to eliminate all unjustified mechanistic interpretations.
Major Comments:
Comment 1:__ Across several sections (incl abstract, L559-565, L589-599, L600-L603, L610-612, L613-614, L625, L643-645, L650-652), the manuscript describes "recursive or self-controlling networks", "signaling cascades", "self-regulating", and "recursive feedback loops" - involving protein kinases, phosphatases, and translational regulators. While the data convincingly demonstrate stage-specific changes in phosphorylation and abundance changes in key molecules, the language used implies causal, direct and directional regulatory relationships that have not been experimentally validated.
Response: __We agree with the reviewer and have corrected the text, replacing all expressions that may allude to causal or directional relationships by more neutral expressions such as 'co-expression'. __
Comment 2: Co-expression and shared function alone do not define a regulon (L363, and several other places in the manuscript). A regulon also requires the gene set to be regulated by the same factor, for which there is no evidence here. Regulons can be derived from transcriptomic experiments, but then they need to show the same transcriptional behavior across many biological conditions, while here just 1 condition change is evaluated. Therefore, this analysis is conventional GO enrichment analysis and should not be overinterpreted into regulons.
__Response: __We agree with the reviewer and have replaced 'regulon' with 'co-regulated gene clusters' (or similar).
Comment 3: LFQ intensity of 0 (e.g., L389): An LFQ intensity of 0 does not necessarily indicate that a protein is absent, but rather that it was not detected. This can occur for several reasons: (1) true biological absence in one condition, (2) low abundance below the detection threshold, or (3) stochastic missingness due to random dropout in mass spectrometry. While the authors state that adjusted p-values for the 1534 proteins exclusively detected in either amastigotes or promastigotes are below 0.01, I could not find corresponding p-values for these proteins in Table 8 ('Global_Proteomic'). An appropriate statistical method designed to handle this type of missingness should be used. In this context, I also find the following statement unclear: "identified over 4000 proteins at each stage in at least 3 out of 4 biological replicates, representing 3521 differentially expressed proteins (adjusted p-value Response: We fully agree with the reviewer, an LFQ intensity of 0 may results from various reasons. We realize that our wording may have been ambiguous. For clarity, we have modified the original text to: 'Label-free quantitative proteomic analysis of 4 replicates of amastigotes and derived promastigotes identified over 4000 proteins, including 1987 differentially expressed proteins (adjusted p-value<br />
Comment 4: L412 - Figure 3B: The figure shows proteins with infinite fold changes, which result from division by zero due to LFQ intensity values of zero in one of the compared conditions. As previously noted, interpreting LFQ zero values as true absence of expression is problematic, since these zeros can arise from several technical reasons - such as proteins being just below the detection threshold or due to stochastic dropout during MS analysis. Therefore, the calculated fold changes for these proteins are likely highly overestimated. This concern is visually supported by the large gap on the y-axis (even in log scale) between these "infinite" fold changes and the rest of the data. Moreover, given Leishmania's model of constitutive gene expression, it seems biologically implausible that all these proteins would be completely absent in one stage. This issue applies not only to Figure 3B, but also to the analyses presented in Figures 4D and 4E.
Response: __We thank the reviewer for this comment. To clarify this section, we modified the text as follows: 'Only expression changes were considered that either showed statistically significant differential abundance at both RNA and protein levels (p <br />
__Minor Comments:
Methods
L132: Typo: "A according" should be "according."
__Response: __The 'A' refers to RNase A. We added a comma for clarification (...RNase A, according to...)
L158: How exactly were somy levels calculated? Please specify the method used, as I could not find a clear description in the referenced manuscript.
__Response: __We thank the reviewer for this comment. Aside the already quite detailed description in Methods and the reference there to the paper describing the pipeline, we now added a link to the description of the karyotype module of the giptools package (https://gip.readthedocs.io/en/latest/giptools/karyotype.html). There the following explanation can be found: "The karyotype module aims at comparing the chromosome sequencing coverage distributions of multiple samples. This module is useful when trying to detect chromosome ploidy differences in different isolates. For each sample the module loads the GIP files with the bin sequencing coverage (.covPerBin.gz files) and normalizes the meancoverage values by the median coverage of all bins. The bin scores are then converted to somy scores which are then used for producing plots and statistics." The description then goes into further detail.
L158: Chromosome 36 is not consistently disomic, as stated. It has been observed in other somy states (e.g., Negreira et al. 2023, EMBO Reports, Figure 1), even if such occurrences are rare in the studied context. Normalizing by chr36 remains a reasonable choice, but it would be helpful to confirm that the majority of chromosomes appear disomic post-normalization to support the assumption that chr36 is disomic in this dataset as well.
__Response: __We thank the reviewer for this comment. Unlike the paper cited above (using long-term cultured promastigotes), our analysis uses promastigote parasites from early culture adaptation (p2) that were freshly derived from splenic amastigotes known to be disomic (and confirmed here), which represents an internal control validating our analysis.
L163: Suggestion: Cite the GIP pipeline here rather than delaying the reference until L173.
Response: corrected
L188: "Controlled" may be a miswording. Consider replacing with "confirmed" or "validated."
Response: corrected to 'validated'
L214: Please specify which statistical test was used to assess differential expression at the protein level. L227: Similarly, clarify which statistical test was applied for determining differential expression in the phospho-proteomics data.
Response: As noted in the Methods section, a limma t-test was applied to determine proteins/phosphoproteins with a significant difference in abundance while imposing a minimal fold change of 2 between the conditions to conclude that they are differentially abundant {Ritchie, 2015; Smyth, 2005}.
__Results __
L337-339: The interpretation here is too speculative. Phrases like "suggesting" and "likely" are too strong given the evidence presented. Alternative explanations, such as mosaic variation combined with early-stage selective pressure in the culture environment, should be considered.
Response: We thank the reviewers for these suggestions and have reformulated into: 'In the absence of convergent selection, it is impossible to distinguish if these gene CNVs provide some strain-specific advantage or are merely the result of random genetic drift.'
L340: The "undulating pattern" mentioned is somewhat subjective. To support this interpretation, consider adding a moving average (or similar) line to Figure 3A, which would more clearly highlight this trend across the data points.
Response: These lines have been added to Figure 1C (not 3A).
L356: It may be more accurate to say "control of individual gene expression," since Leishmania does have promoters - the key distinction is that initiation does not occur on a gene-by-gene basis.
Response: corrected
L403-405: The statement "this is because these metabolites comprise a glycosomal succinate shunt..." should be rephrased as a hypothesis rather than a definitive explanation, as this causal link has not been experimentally validated.
Response: Thank you for the comment - we followed your advice.
L407: Replace "confirming" with "matching" to avoid overstating the agreement with previous observations.
Response: corrected
L408: Replace "correlated" with "matched" for more accurate interpretation of results.
Response: corrected
L433: It is unclear how differential RNA modifications were detected. Please specify which biological material was used, the number of replicates per life stage, and how statistical evaluation of differential modifications was performed.
Response: This figure has now been updated using our statistically robust RNA-seq analysis conducted for the revision. See comments above.
L436: This conclusion appears incomplete. While the manuscript mentions transcript-regulated proteins, it should also note that other proteins showed discordant mRNA/protein patterns. A more balanced conclusion would mention both the matching and non-matching subsets.
Response: We thank the reviewer for this comment and have made the necessary adjustments to better balance this conclusion.
L441: The phrase "poor correlation" overgeneralizes and lacks nuance. Earlier sections of the manuscript describe hundreds of genes where mRNA and protein levels correlate well, suggesting that mRNA turnover plays a key regulatory role. Please rephrase this sentence to clarify that poor correlation applies only to a subset of the data.
Response: This has been corrected to 'The discrepancies we observed in a sub-set of genes between....'.
L454: The claim that "epitranscriptomic regulation and stage-adapted ribosomes are key processes" should be supported with references. If this builds on previously published work, please cite it accordingly.
Response: corrected
L457: Proteasomal degradation is a well-established mechanism in Leishmania. These findings are interesting but should be presented in the context of existing literature (e.g. Silva-Jardim et al.2014, [PMID: 15234661]) rather than as entirely novel.
Response: corrected
L459: The authors shoumd add a microscopy image of promastigotes treated with lactacystin. This would provide insight into whether treatment affects morphology, as is known in T. cruzi (see Dias et al., 2008). It would be particularly informative if Leishmania behaves differently.
Response: We added this information to Figure S7.
L472 + L481: Table 9 shows several significant GO terms not discussed in the manuscript. Please clarify how the subset presented in the text was selected.
Response: We added this information to the text ('some of the most significantly enrichment terms included ...').
L482: The argument that a single master regulator can be excluded is unclear. Could the authors please elaborate on the reasoning or data supporting this conclusion?
Response: This statement was too speculative and has been removed. Instead, we added 'Thus, Leishmania differentiation correlates with the expression of complex signaling networks that are established in a stage-specific manner'.
L494: The term "unexpected" may not be appropriate here, as protein degradation is a well-established regulatory mechanism in trypanosomatids. Consider omitting this term to better reflect the field's current understanding.
Response: We deleted the term as suggested and reformulated to '....our results confirm the important role of protein degradation....'.
L543: The term "feedback loop" should be used more cautiously. The current data are correlative, and no interventional experiments are provided to support a causal regulatory loop between proteasomal activity and protein kinases. As such, this remains a hypothesis rather than a confirmed mechanism.
Response: We fully agree and have toned down the entire manuscript, referring to feedback loops only as a hypothesis and not as a fact emerging from our datasets, which set the stage for future functional analyses.
__Discussion __
L555: As noted in L494, reconsider using the word "unexpected."
Response: removed
L589: The data do not fully support the presence of stage-specific ribosomes. Rather, they suggest differential ribosomal function through changes in abundance and regulation. Please consider rephrasing.
Response: We thank the reviewer for this comment and have follow the advice reformulating the sentence according to the suggestion.
L657-658: The discussion of post-transcriptional and post-translational regulation of gene dosage effects would benefit from citing additional literature beyond the authors' own work. E.g. the study by Cuypers et al. (PMID: 36149920) offers a relevant and comprehensive analysis covering 4 'omic layers.
Response: We apologize for this omission and now describe and cite this publication in the Results section when concluding the results shown in Figure 1.
L659-664: The reference to deep learning for biomarker discovery appears speculative and loosely connected to the current findings. As no such methods were applied in the study, and the manuscript does not clarify what types of biomarkers are intended, this statement could be seen as aspirational rather than evidence-based. Consider either omitting or elaborating with clear justification.
Response: We agree and have deleted this section.
L690 + L705 (Figure 2): The phrase "main GO terms" is vague. Please clarify the criteria for selecting the GO terms shown - were they chosen based on adjusted p-value, enrichment score, or another metric? Additionally, define "cluster efficiency," explaining how it was calculated and what it represents.
Response: Corrected to 'some of the most significantly enriched GO terms'.
Signed: Bart Cuypers, PhD
**Referee cross-commenting**
Overall, I think the other reviewers' comments are fair. They seem to align particularly on the following points:
1) Reviewers agree that this is a comprehensive body of work with original contributions to the field of Leishmania/trypanosomatid molecular biology, and that it will serve as a valuable reference for hypothesis generation.
2) Several reviewers raise concerns about overinterpretation of the data, particularly regarding regulatory networks, regulons, and master regulators. The interpretation and large parts of the discussion are considered too speculative without additional functional validation.
3) There are comments about the incorrect statistical treatment of missing values in the proteomics experiments, which affects confidence in some of the conclusions.
4) While the correlation between the two RNA-Seq replicates is high, the decision to include only two biological replicates is seen as unfortunate and not ideal for statistical robustness.
5) The use of lactacystin should be more clearly motivated, and its limitations discussed in the context of the experiments.
Even though I did not remark on the last two points (4 and 5) in my own review, I agree with them.
Response: We thank the reviewer for this cross-comparison, which served us as guide to revise our manuscript. We believe that we have responded to all these concerns.
Reviewer #3 (Significance (Required)):
This study provides a rich, integrative multi-omics dataset that advances our understanding of stage-specific adaptation in the transcriptionally unique parasite Leishmania. By dissecting the relative contributions of mRNA abundance and protein turnover to final protein levels across life stages, the authors offer valuable insights into post-transcriptional and post-translational regulation. The work represents a resource-driven yet conceptually informative contribution to the field, with comprehensive supplementary materials and transparent data sharing standing out as additional strengths.
However, the mechanistic insights proposed are speculative in several places and require more cautious language. The study is most impactful as a resource and descriptive atlas, initiating hypotheses for future validation. The broad scientific community working on Leishmania, trypanosomatids, and post-transcriptional regulation in eukaryotes would benefit from this work.
Response: We thank the reviewer for this positive assessment and have modified the manuscript to further emphasize its strength as an important resource to incite mechanistic follow-up studies.
Field of reviewer expertise: multi-omics integration, bioinformatics, molecular parasitology, transcriptomics, proteomics, metabolomics, Leishmania, Trypanosoma.
Reviewer #4 (Evidence, reproducibility and clarity (Required)):
__
__Summary:
This study investigates the regulatory mechanisms underlying stage differentiation in Leishmania donovani, a parasitic protist. Pesher et al., aim to address the central question of how these parasites establish and maintain distinct life cycle stages in mostly the absence of transcriptional control. The authors employed a five-layered systems-level analysis comparing hamster-derived amastigotes and their in vitro-derived promastigotes. From those parasites, they performed a genomic, transcriptomic, proteomic, metabolomic and phosphoproteomic analysis to reveal the changes the parasites undertook between the two life stages.
The main conclusion stated by the authors are:
- The stage differentiation in vitro is largely independent of major changes in gene dosage or karyotype.
-
RNA-seq analysis identified substantial stage-specific differences in transcript abundance, forming distinct regulons with shared functional annotations. Amastigotes showed enrichment in transcripts related to amastins and ribosome biogenesis, while promastigotes exhibited enrichment in transcripts associated with ciliary cell motility, oxidative phosphorylation, and post-transcriptional regulation itself.
-
Quantitative phosphoproteome analysis revealed a significant increase in global protein phosphorylation in promastigotes. Normalizing phosphorylation changes against protein abundance identified numerous stage-specific phosphoproteins and phosphosites, indicating that differential phosphorylation also plays a crucial role in establishing stage-specific biological networks. The study identified recursive feedback loops (where components of a pathway regulate themselves) in post-transcriptional regulation, protein translation (potentially involving stage-specific ribosomes), and protein kinase activity. Reciprocal feedback loops (where components of different pathways cross-regulate each other) were observed between kinases and phosphatases, kinases and the translation machinery, and crucially, between kinases and the proteasomal system, with proteasomal inhibition disrupting promastigote differentiation.
Response: We thank the reviewer for the time and implication dedicated to our manuscript.
Comments:
Further details are organised by order of apparition in the text:
Comment 1: Material and Methods: while the authors are indicating some key parameters, providing the codes and scripts they used throughout the manuscript would improve reproducibility.
Response: We thank the reviewer for this comment and added the URL for the codes to the data availability section.
Comment 2: Why only 2 biological replicates for RNA while the others layers have 3 or 4?
__Response: __We agree with the other reviewers and have repeated this analysis to have statistically more robust results.
Comment 3: Is the slight but reproducible increase in median coverage observed for chr 1, 2, 3, 4, 6 and 20 stable on longer culture derived promastigotes and sandfly derived promastigotes ?
Response: No, as published in Barja et al Nature EcolEvol 2017 (PMID: 29109466) and Bussotti et al PNAS 2023 (PMID: 36848551), these minor fluctuations are not predicting subsequent aneuploidies in long-term culture nor in sand fly-derived promastigotes. This information has been added to the text.
Comment 4: Is this change of ploidy a culture adaptation representation rather than a life cycle event as the authors discuss later on? (This is probably an optional request that would be nice to include, if the authors have performed the sequencing of such parasites. Otherwise, it should be mentioned in the discussion).
__Response: __Yes, this is a well-known culture adaptation phenomenon, on which we have published extensively. We added this conclusion and the references to the text.
Comment 5: L333 "Likewise, stage differentiation was not associated with any major gene copy number variation (Figure 1C, Table 2)". The authors are looking here at steady differentiated stages rather than differentiation itself. "Likewise, stage differentiation was.." would be more appropriate.
__Response: __We corrected this sentence to 'Likewise, differentiation of promastigotes was not associated with any major gene copy number variation at early passage 2'.
Comment 6: L349-355: have the mRNA presenting change in abundance between stages been normalised by their relative DNA abundance ? Said otherwise, can the wave patterns observed at the genome level explain the respective mRNA level ? Can the authors plot in a similar way the enrichment scores in regards to the position on the genome and can the authors indicate if there is a positional enrichment in addition to the functional one they observe ? This may affect the conclusion in L356-358.
Response: As noted above, we did not see any significant read depth changes at DNA level when comparing amastigotes and promastigotes. Thus there is no need to normalize the RNA-seq results to DNA read depth. Furthermore, in our comparative transcriptomics analysis, we only consider 2-fold or higher changes in mRNA abundance (which is far beyond the non-significant read depth change we have observed on DNA level). Manual inspection of the enrichment scores with respect to position did not reveal any significant signal (other than revealing some over-represented tandem gene arrays where all gene copies share the same location and GO term).
Comment 8: L415 "stage-specific expression changes correlate between protein and RNA levels, suggesting that the abundance of these proteins is mainly regulated by mRNA turn-over". Overstatement. Correlation does not suggest causation. "suggesting that the abundance of these proteins could be regulated by mRNA turn-over" would be more appropriate.
Response: We thank the reviewer for this comment and have corrected the statement accordingly.
Comment 9: Figure 3B, could the authors clarify what are the "unique genes" that are on the infinite quadrants? It seems these proteins are identified in one stage and not the other. This implies that the corresponding missing values are missing non-at random (MNAR). Rather than removing those proteins containing NMAR from the differential expression analysis, the authors should probably impute those missing values. Methods of imputation of NMAR and MAR can be found in the literature. Indeed, the level of expression in one stage of those proteins is now missing, while it could strongly affect the conclusions the authors are drawing in figure 4E regarding the proteins targeted for degradation and rescued in presence of the proteasome inhibitor.
Response: We thank the reviewer for this important comment. However, we would like to clarify several key points regarding the treatment of proteins identified in only one condition.
First, the reviewer assumes that proteins identified in one stage but not the other are necessarily missing not-at-random (MNAR). However, this cannot be definitively established, as these missing values could equally be missing completely at random (MCAR). Without additional information, categorizing them specifically as MNAR may be an oversimplification. More importantly, we have concerns about the reliability of imputation methods in this specific context. Algorithms designed to impute MNAR values (such as QRILC) replace absent data using random sampling from arbitrary probability distributions, typically assuming low intensity values. However, when no intensity value has been detected or quantified for a protein in a given condition, imputing an arbitrary low value raises significant concerns about data interpretation. Such imputed values would not reflect actual measurements but rather statistical assumptions that could introduce bias into downstream analyses. For instance, imputed values could lead to the conclusion that a protein is not differentially abundant, when in reality it is detected in one condition but completely absent in the other. In our view, there are two biologically plausible scenarios: either these proteins are expressed at levels below our detection threshold, or they are genuinely absent (or present at negligible levels) in the corresponding stage. Rather than introducing potentially misleading imputed values, we chose to treat these as genuine stage-specific differences (presence/absence), which results in infinite fold-changes in Figure 3B. Critically, our approach is strongly supported by independent validation through RNA-seq data, which corroborates the differential presence/absence patterns observed at the protein level. Furthermore, our enrichment analyses reveal significant over-representation of specific biological terms among these stage-specific proteins, providing biological coherence to these findings. These converging lines of evidence (proteomics, transcriptomics, and functional enrichment) strengthen our confidence that these represent biologically meaningful differences rather than technical artifacts.Therefore, we believe our conservative approach of treating these as genuine presence/absence differences, validated by orthogonal data, is more appropriate than introducing imputed values based on arbitrary statistical assumptions.To clarify this section, we modified the text as follows: 'Only expression changes were considered that either showed statistically significant differential abundance at both RNA and protein levels (p Comment 10: L430-435 "These data fit with the GO [...] the ribosome translational activity (34)." This discussion feels out of place and context. It is too speculative and with little support by the data presented at this stage of the manuscript. It should be removed as Figure 3E or could be placed in the discussion and supplementary information.
Response: We agree with the reviewer. In response to a comment from reviewer 1, we have moved both panels to Figure 2, which much better integrates these data.
Comment 10: The authors present an elegant way to show stage specific degradation through the comparison of stage specific proteasome blockages that show rescue in ama of proteins present in pro and vice versa. L494 "reveal an unexpected but substantial" the term unexpected is inappropriate, as several studies have shown in kinetoplastids the essential role of protein turnover through degradation / autophagy during differentiation. Furthermore the conclusions may be strongly affected by the level of expression of the proteins in the infinite quadrants as we discussed above, and should be revised accordingly.
Response: We rephrased the conclusion to 'In conclusion, our results confirm the important role of protein degradation in regulating the L. donovani amastigote and promastigote proteomes and identify protein kinases as key targets of stage-specific proteasomal activities.' Please see the response to comment 9 regarding the unique proteins.
Comment 11: L518 "These data reveal a surprising level of stage-specific phosphorylation in promastigotes, which may reflect their increased biosynthetic and proliferative activities compared to amastigotes." Overstatement. Could also be due to culture adaptation - What is the overlap of stage-specific phosphorylations with previous published datasets in other species of Leishmania? Looking at such comparisons could help to decipher the role of culture adaptation response, species specificity and true differentiation conserved mechanisms.
Response: We agree with the reviewer and have toned this statement down by adding the statement '....or simply be a consequence of culture adaptation'.
Comment 12: The discussion is extremely speculative. While some speculation at this stage is acceptable, claiming direct link and feedback without further validation is probably far too stretched. For example, the changes of phosphorylation observed on particular sets of proteins, such as phosphatase and DUBs, need to be validated for their respective change of protein activity in the direction that fits the model of the authors. Those discussions should be toned down.
Response: We agree with the reviewer and have strongly toned down the entire discussion, emphasizing the hypothesis-building character of our results, which provide a novel framework for future experimental analyses.
Comment 13: A couple of typos:
-
In the phosphoproteome analysis section, "...0,2 % DCA..." should be "...0.2 % DCA..." (use a decimal point).
-
L225 "...peptide match was disable." should be "...peptide match was disabled."
Response: both corrected
__Reviewer #4 (Significance (Required)): __
While there is not too much novelty around the emphasis of gene expression at post-translational level in kinetoplastid organisms, the scale of the work presented here, looking at 5 layers of potential regulations, is. Therefore, this study represents a substantial amount of work and provides interesting and comprehensive datasets useful for the parasitology community.
Response: We thank the reviewer for this positive statement.
Several potential concerns regarding the biological meaning of the findings were identified. These include the limitations of in vitro systems promastigote differentiation potentially limiting the conclusions, the challenge of inferring causality from correlative "omics" data, and the complexities of functional interpretation of changes in phosphorylation and metabolite levels. The proposed feedback loops and functional roles of specific molecules would require further experimental validation to confirm their biological relevance in the natural life cycle of Leishmania, but that would probably fall out of the scope of this manuscript.
Response: We agree with the reviewer and have modified pour manuscript throughout to remove any causal relationships. Indeed, this work is setting the stage for future investigations on dissecting some of the suggested regulatory mechanisms.
Area of expertise of the reviewers: Kinetoplastid, Differentiation, Signalling, Omics